一种提高玉米高温胁迫下光合电子传递的方法与流程
本发明涉及生物科学技术领域,特别是涉及一种提高玉米高温胁迫下光合电子传递的方法。
背景技术:
近年来,温室效应引起的异常高温天气的频繁发生已成为制约作物生长发育的一个非常重要的环境因素。在世界三大农作物中,玉米产量最高。由于气温每升高1℃,作物产量可减少17%(lobell等人,2003年),因此,高温对玉米生产的不利影响日益受到关注。
光合作用极易受到高温的影响,因为高温不仅损害光合色素、光系统、电子传递而且还影响碳同化(ashraf等人,2013年)。高等植物中存在两种形式的光合电子传递:线性电子传递:水→psii→pq→cytb6f→pc→psi→fd→fnr→nadp+和环式电子传递(cef):psi→fd→pq→cytb6f→pc→psi。高温会降低光合效率,产生光抑制,从而导致过剩光能的产生,这主要发生在psii、pq和psi中(mathur等人,2014;song等人,2014)。psii对温度非常敏感。高温胁迫可降低其供体侧和受体侧的活性,破坏其反应中心(yan等人,2013)。此外,高温胁迫还导致类囊体膜重组、d1和d2蛋白结构变化以及qa和qb电子载体之间氧化还原平衡的改变,从而造成qa-qb电子传递的抑制(cao和govindjee,1990)。在光合反应中,pq库的氧化还原条件也受到调控。高温胁迫可导致其过度还原,增加类囊体中的△ph水平(mathur等人,2014)。此外,伴随着质子进入类囊体腔的pqh2氧化,被称为线性电子传递中的限速步骤,因为其氧化速率(约1-20ms)比其他过程(约1ms)要慢得多(yamori和shikanai,2016)。psi活性比psii稳定(havaux,1993)。中等程度的高温可以增加psi活性、类囊体的跨膜质子梯度以及环psi的cef(bukhovetal.,1999;bukhovetal.,2000)。c4植物光合作用所需的额外atp由环psi的cef提供(havaux等人,2005)。此外,高温还显著改变了电子传输链中每个组分的氧化还原平衡,从而推动电子传递到psi(bukhov等人,2000)。
因此,需要提供能够促进高温胁迫下玉米中的光合电子传递的方法。
技术实现要素:
本发明的目的是提供一种提高玉米高温胁迫下光合电子传递的方法,以解决上述现有技术存在的问题。
为实现上述目的,本发明提供了如下方案:
本发明提供一种提高玉米高温胁迫下光合电子传递的方法,玉米幼苗期添加外源海藻糖。
进一步地,外源海藻糖提高高温胁迫下玉米全链、psii以及psi的光合电子传递链活性。
进一步地,所述外源海藻糖的浓度为0.5mmol/l。
发明人提供外源添加海藻糖研究了玉米高温胁迫下的光合电子传递活性。添加外源海藻糖的植株在正常条件下生长正常,在高温胁迫条件下生长优于对照,且能促进植株早熟,缩短生长周期。
本发明公开了以下技术效果:
本发明在高温胁迫(42℃)和室温(25℃)条件下培育海藻糖预处理的玉米幼苗。然后测量了光合电子传递活性、叶绿素荧光值和p700吸收、快速叶绿素荧光诱导曲线、pq大小和p700+初始还原速率等参数,提供了一种海藻糖对玉米幼苗叶片光合电子传递的作用,为进一步阐明海藻糖介导的玉米耐热性调控机制提供了理论依据。
附图说明
为了更清楚地说明本发明实施例或现有技术中的技术方案,下面将对实施例中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图仅仅是本发明的一些实施例,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他的附图。
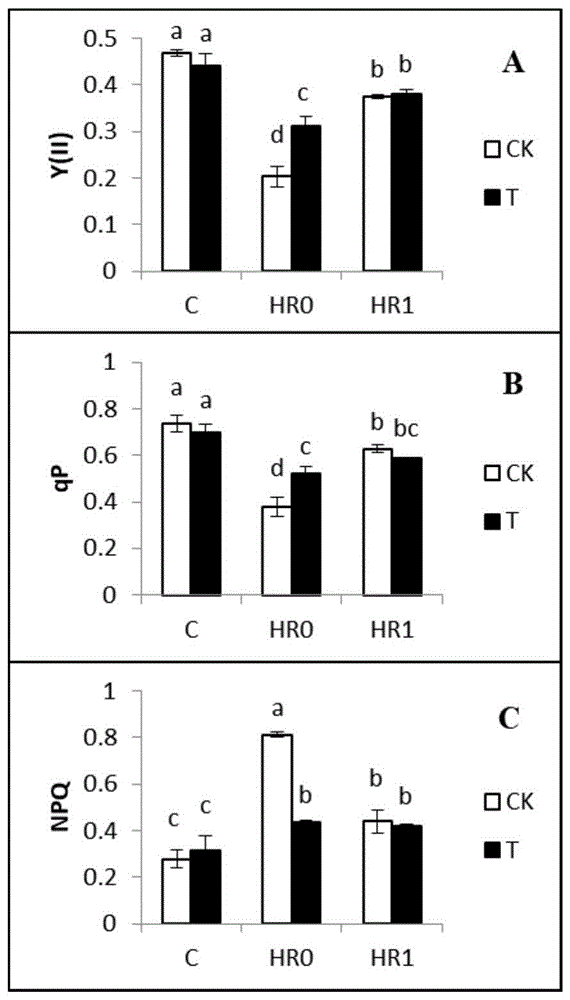
图1为外源海藻糖对不同处理组玉米幼苗叶片y(ii)、qp和npq的影响,其中,(a)y(ii):光系统ii的实际光化学效率;(b)qp:光化学淬灭;(c)npq:非光化学淬灭。ck:对照组;t:海藻糖预处理组;c:未进行高温处理;hr0:高温处理1天;hr1:高温胁迫后,在常温下恢复1天;不同字母代表显著性差异(p<0.05);
图2为外源海藻糖对不同处理组玉米幼苗叶片y(i)、y(nd)和y(na)的影响,其中,(a)y(i):光系统i的实际光化学效率;(b)y(nd):光系统i由于供体侧限制引起的非光化学能量耗散的量子产量;(c)y(na):光系统i由于受体侧限制引起的非光化学能量耗散的量子产量。ck:对照组;t:海藻糖预处理组;c:未进行高温处理;hr0:高温处理1天;hr1:高温胁迫后,在常温下恢复1天;不同字母代表显著性差异(p<0.05);
图3为外源海藻糖对不同处理组玉米幼苗叶片快速叶绿素荧光诱导动力学曲线的影响,其中,ck:对照组;t:海藻糖预处理组;h0:未进行高温处理;h:高温处理1天;hr:高温胁迫后,在常温下恢复1天;不同字母代表显著性差异(p<0.05);
图4为外源海藻糖对不同处理组玉米幼苗叶片pq库大小的影响,其中,ck:对照组;t:海藻糖预处理组;c:未进行高温处理;hr0:高温处理1天;hr1:高温胁迫后,在常温下恢复1天;不同字母代表显著性差异(p<0.05);
图5为外源海藻糖对不同处理组玉米幼苗叶片p700+初始还原速率的影响,其中,ck:对照组;t:海藻糖预处理组;c:未进行高温处理;hr0:高温处理1天;hr1:高温胁迫后,在常温下恢复1天;不同字母代表显著性差异(p<0.05)。
具体实施方式
现详细说明本发明的多种示例性实施方式,该详细说明不应认为是对本发明的限制,而应理解为是对本发明的某些方面、特性和实施方案的更详细的描述。
应理解本发明中所述的术语仅仅是为描述特别的实施方式,并非用于限制本发明。另外,对于本发明中的数值范围,应理解为还具体公开了该范围的上限和下限之间的每个中间值。在任何陈述值或陈述范围内的中间值以及任何其他陈述值或在所述范围内的中间值之间的每个较小的范围也包括在本发明内。这些较小范围的上限和下限可独立地包括或排除在范围内。
除非另有说明,否则本文使用的所有技术和科学术语具有本发明所述领域的常规技术人员通常理解的相同含义。虽然本发明仅描述了优选的方法和材料,但是在本发明的实施或测试中也可以使用与本文所述相似或等同的任何方法和材料。本说明书中提到的所有文献通过引用并入,用以公开和描述与所述文献相关的方法和/或材料。在与任何并入的文献冲突时,以本说明书的内容为准。
在不背离本发明的范围或精神的情况下,可对本发明说明书的具体实施方式做多种改进和变化,这对本领域技术人员而言是显而易见的。由本发明的说明书得到的其他实施方式对技术人员而言是显而易见的。本申请说明书和实施例仅是示例性的。
为验证本发明的有益效果,进行以下实验:
1材料与方法
1.1材料培养与处理
在温室条件下[温度25℃、光周期13h(800μmolm-2s-1ppfd)、相对湿度70%]使用1/2hoagland营养液对“申科糯1号”玉米(zea-maysl.)进行为期14天的培养。等玉米幼苗第三片叶片完全展开后,用0.5mm海藻糖(用1/2的hoagland溶液配制)处理幼苗3d(用不加海藻糖的1/2hoagland溶液作为对照)。随后,对玉米植株进行42℃高温胁迫1d,然后移入室温恢复1d,用各处理的第三片全展叶测定生理参数,重复3-5次。
1.2类囊体膜的分离
根据rintamaki等人(1996)的研究方法稍作修改分离类囊体膜。玉米叶片用含有300mm蔗糖、5mm氯化镁、1mm乙二胺四乙酸钠和10mm氟化钠的50mmhepes-naoh(ph7.5)预冷的分离缓冲液均质。匀浆通过三层尼龙布过滤,并在4℃、1500g离心力下提取4min。用含有10mmhepes-naoh(ph7.5)、5mmmgcl2、5mm蔗糖的溶液冲洗颗粒,将10mmnaf在3000g下离心3min,将类囊体颗粒再悬浮于10mmhepes-naoh(ph7.5)、100mm蔗糖、10mmmgcl2和5mmnacl中,低温保存备用。叶绿素浓度测量采用porro等人的方法(1989年)。
1.3光合电子传递链活性的测定
采用zhang和liu(2016)的方法,使用液相氧电极(hansatech,chlorolab2,英国)测定玉米幼苗叶片的电子传递链活性。反应基质:50mmtricine-koh,50mmkcl,5mmmgcl2,ph7.6含20-50μgchl/ml叶绿素。反应介质:全链(h2o→mv):50μmmv,5mmnh4cl,2mmnan3。全链(dpc→mv):5mmnh4cl,0.5mmdpc,2mmnan3。psii(h2o→p-bq):5mmnh4cl,4mmk3fe(cn)6,1mmp-bq。psi(dcpip→mv):50μmmv,5mmnh4cl,2mmvc,2mmnan3,50μmdcmu。
1.4叶绿素荧光参数和p700参数的同步测定
将样品暗适应20-30min,采用脉冲振幅调制叶绿素荧光仪(dual-pam-100,walz,德国)测定不同处理组叶片的叶绿素荧光参数和p700参数。使用dual-pam软件对诱导曲线进行参数分析。
f0:暗适应下最小叶绿素荧光强度;fm:暗适应下最大叶绿素荧光强度;f0’:光下最小叶绿素荧光强度;fm’:光下最大叶绿素荧光强度;fs:光下稳态荧光强度。光系统ii的最大光化学效率:fv/fm=(fm-fo)/fm;光系统ii的实际光化学效率:y(ii)=(fm’-fs)/fm’;光化学淬灭:qp=(fm’-fs)/(fm’-fo’);非光化学淬灭:npq=fm/fm’-1。
p:光下p700信号的强度;pm:氧化型p700的最大值;pm’:光下氧化型p700的最大值。光系统i的化学量子产量:y(i)=(pm’-p)/pm;光系统i供体侧限制:y(nd)=p/pm;光系统i受体侧限制:y(na)=(pm-pm’)/pm。
1.5快速叶绿素荧光诱导动力学曲线的测定
利用便携式植物效率分析仪(pocketpea,hansatech,uk)测定不同处理组玉米幼苗叶片(暗适应20-30min)的ojip曲线,参考strasser等人(2000)的方法,记录如下原始数据。f1(o相):50μs时的荧光强度;f2(k相):100μs时的荧光强度;f3(j相):300μs时的荧光强度;f4(i相):2ms时的荧光强度;f5(p相):30ms时的荧光强度。对原始曲线进行标准化处理,ojip曲线标准化公式:(ft-fmin)/(fmax-fmin)。
1.6pq库大小的测定
依据schreiber(1988)的方法,采用脉冲振幅调制叶绿素荧光仪(dual-pam-100,walz,德国)测定不同处理组玉米幼苗叶片p700的氧化还原状态。在远红光照射的情况下,分别发射一次单周转闪光(st,50ms,pq库被氧化)和一次多周转闪光(mt,50ms,pq库被完全还原)来获取p700的信号,pq库大小=mt的面积/st的面积(schreiberetal.,1989;savitchetal.,2001)。
1.7p700+暗还原速率的测定
依据bukhov和egorova(2005)的方法,采用脉冲振幅调制叶绿素荧光仪(dual-pam-100,walz,德国)测定不同处理组玉米幼苗叶片p700+的暗还原速率。关掉远红光后,p700+暗还原曲线下降的斜率能够表征cef的速率,是研究cef的重要方法(schreiberandklughammer,2008)。
1.8统计分析
所有数据都使用spss21.0(spss,chicago,usa)进行了统计学分析,依据tukey检验对每个独立样本进行了单因素方差分析(anova),显著性水平设置为p≤0.05(显著)和p≤0.01(极显著)。所有实验均进行至少3次重复,所有数据均以平均值±标准差呈现。
2结果
2.1高温胁迫下外源海藻糖对光合电子传递链活性的影响
首先,本实验分析了外源海藻糖对不同处理组玉米幼苗叶片光合电子传递链活性的影响。表1的实验结果表明,高温胁迫显著降低了全链和psii光合电子传递链的活性,却提高了psi光合电子传递链的活性。psi的光合电子传递链活性的提高是由于高温胁迫下psi周围cef活性的急速提升。同时,也发现在高温胁迫条件下相较于未外源添加海藻糖组,外源添加海藻糖显著提高了全链、psii以及psi的光合电子传递链活性。因此结果表明,外源添加海藻糖可以有效缓解高温胁迫对玉米幼苗光合电子传递链活性的抑制,进而提高光合作用中的光反应效率,提高玉米的耐热性。
表1外源海藻糖对不同处理组玉米幼苗叶片光合电子传递链活性的影响
ck:对照组;tre:海藻糖预处理组;h0:未进行高温处理;h:高温处理1天;hr:高温胁迫后,在常温下恢复1天。不同字母代表显著性差异(p<0.05)。
2.2高温胁迫下外源海藻糖对叶绿素荧光和p700参数的影响
为了探究高温胁迫下外源海藻糖具体保护了光合电子传递链上的哪些受损位点,本发明测定了玉米幼苗叶片中叶绿素荧光和p700的参数。叶绿素荧光参数可以反映psii的活性;p700的参数可以反映psi的活性。高温胁迫显著降低了y(ii)和qp,却显著提升了npq(图1a,b,c)。另外,在高温胁迫条件下相较于对照组,外源添加海藻糖显著提高了玉米幼苗叶片中的y(ii)和qp(图1a,b)。综上,可以得出,外源添加海藻糖不是通过提升高温胁迫条件下玉米幼苗的热耗散能力而是通过提高其光化学效率,即加快光合电子传递的速率,进而提高y(ii)来起到保护光合电子传递链活性的作用。
高温胁迫条件下,y(i)显著降低,而y(nd)和y(na)显著升高,表明高温严重增强对psi供体侧和受体侧的限制,进而降低了psi的实际光化学效率(图2a,b,c)。此外,本发明还发现高温胁迫阶段相较于对照组,外源添加海藻糖降低了y(nd),提高了y(i),而对y(na)没有影响(图2a,b,c)。这表明外源添加海藻糖可以通过缓解高温胁迫对psi供体侧的抑制来显著提高psi的实际光化学效率。
2.3高温胁迫下外源海藻糖对ojip曲线的影响
为了进一步确定海藻糖预处理究竟加快了通过psii的哪段光合电子传递速率,本发明分析了不同处理组玉米幼苗叶片中的ojip曲线。在高温胁迫和后期常温恢复阶段,k相和j相的峰值振幅明显高于常温组(图3)。在高温胁迫和后期常温恢复阶段相比于对照组,海藻糖预处理显著降低了k相和j相的峰值振幅(图3),这表明海藻糖预处理可以有效提高光合电子由pheo到qa再到qb的传递。因此,在高温胁迫及其后续恢复过程中,海藻糖预处理组中psii的实际光化学效率更高。
2.4高温胁迫下外源海藻糖对pq库的影响
pq库介于光系统ii与光系统i之间,是光合电子传递失衡的感应器。高温胁迫显著降低了pq库的大小(图4)。换言之,在高温胁迫阶段,还原态的pq增多,氧化态的pq减少。相比于对照组,海藻糖预处理可以有效增大高温胁迫下pq库的库容(图4)。这表明,海藻糖预处理可以有效缓解高温胁迫引起的pq库的过度还原,进而促进玉米幼苗叶片中两个光系统之间的光合电子传递。
2.5高温胁迫下外源海藻糖对p700+初始还原速率的影响
为了探究海藻糖预处理对cef速率的影响,本发明测定了p700+的初始还原速率,它可以代表psi周围cef的速率。高温胁迫提高了幼苗叶片中cef的速率(图5),进而提高了光系统i的实际光化学效率。高温胁迫阶段相比于对照组,海藻糖预处理显著提高了cef的速率(图5)。
以上所述的实施例仅是对本发明的优选方式进行描述,并非对本发明的范围进行限定,在不脱离本发明设计精神的前提下,本领域普通技术人员对本发明的技术方案做出的各种变形和改进,均应落入本发明权利要求书确定的保护范围内。
- 还没有人留言评论。精彩留言会获得点赞!