双特异性抗原结合构建体的制作方法
相关申请本申请要求于2017年12月22日提交的美国临时专利申请号62/609,523的权益,该美国临时专利申请的全部内容以引用的方式并入本文。序列表本申请含有序列表,该序列表已经以ascii格式以电子方式递交并且在此以引用的方式整体并入本文。所述于2018年12月21日创建的ascii副本被命名为607772_agx5-031_st25.txt并且具有59,420字节的大小。本发明涉及多特异性,例如双特异性抗原结合构建体,其包含常规抗体和vhh抗体以改善与蛋白复合物的结合。所述抗原结合构建体能够靶向蛋白质复合物内至少两种不同的抗原。本发明还涉及多特异性抗原结合构建体,其包含常规抗体和vhh抗体以改善通过例如尺寸排阻色谱法(sec)进行的纯化。
背景技术:
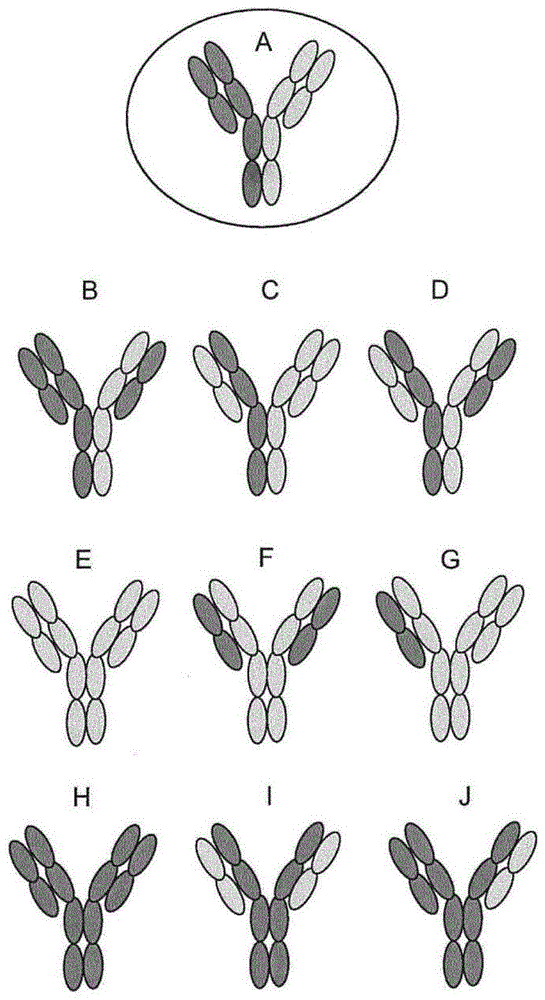
:包括二价抗体在内的天然存在的抗体对特定靶抗原上的特定表位表现出免疫反应性。顾名思义,多特异性抗体是被工程化以识别并且结合可能位于所关注的不同靶抗原上的多于一个表位的抗体。天然存在的常规抗体通常包括免疫球蛋白重链和轻链的组合,其中分子的抗原结合特性由重链和轻链的可变区或结构域(即分别为vh结构域和vl结构域)决定。更具体地,常规抗体的抗原结合位点通常包括由vh结构域和vl结构域中的每一个内的三个互补决定区(cdr)贡献的残基。多特异性抗体与天然存在的抗体的不同之处通常在于它们包含多于一个vh-vl结构域配对以使得它们可以识别并且结合多于一个表位。在商业上,这些抗体由于它们结合多于一种靶抗原的能力而是非常重要的。然而,由于并入同一抗体分子中的不同重链与轻链之间的错配,因此在多特异性抗体的制造和分离方面存在显著的困难。这些错配可能会导致单特异性抗体或具有非功能性或无成效性抗原结合位点的抗体的无意产生,从而降低了所关注的多特异性抗体的产率。图1图示了在对两个不同的表位表现出免疫反应性的双特异性抗体的制备中可能出现的困难。如所示的双特异性抗体(a)包括两条不同的重链和两条不同的轻链。然而,只有这四条免疫球蛋白链的正确配对才能产生具有所期望的结合特性,即对两种靶抗原具有特异性的抗体。实际上,由所示的四条重链和轻链的混合物可以形成九种其它潜在的组合,从而产生二价单特异性抗体(e和h)、单价单特异性抗体(b、c、g和j)和非结合抗体(d、f和i)。多特异性抗体分子越复杂,即抗体意图结合的表位或抗原越多,则该问题会变得越严重。已经进行了各种尝试以通过解决不正确的链配对的问题来改善多特异性抗体的制备。几种方法已经侧重于将抗体工程化以促进vh-vl结构域之间的正确配对。例如,美国专利申请号2010/0254989a1描述了双特异性cmet-erbb1抗体的构建,其中单个抗体的vh和vl通过glyser接头遗传融合。一种替代方法使用大鼠-小鼠四源杂交瘤产生双特异性抗体,其中小鼠和大鼠抗体主要形成原始的vh-vl配对并且所述双特异性抗体由大鼠和小鼠fc组成(lindhofer等,jimmunol.(1995)155:1246-1252)。对于包括fc结构域的双特异性抗体,研究人员还已经侧重于将突变引入到重链的恒定区中以促进fc部分的正确异二聚化。在klein等(mabs(2012)4:6,1-11)中综述了几种这样的技术,该文献的内容以引用的方式整体并入本文。这些技术包括“杵入臼”(knobs-into-holes,kih)方法,其涉及将大体积的残基引入到抗体重链之一的ch3结构域之一中。该大体积的残基适合于配对的重链的另一个ch3结构域中的互补“臼状结构”,从而促进重链的正确配对。研究人员还已经试图解决实现重链和轻链对的正确缔合的问题。一种方法使用crossmab原理(如klein等中所综述),其涉及重链与轻链之间的结构域交换以促进正确配对的形成。其它方法已经试图将重链和轻链的配对的vh-vl结构域或配对的ch1-cl结构域之间的界面工程化改造以增加重链与它的同源轻链之间的亲和力(lewis等,naturebiotechnology(2014)32:191-198)。诸如上述那些的需要广泛的抗体工程化的技术已经取得了一些成功;然而,带有特定突变的抗体的制备可能是劳动密集型的,并且可能产生在人类中具有高免疫原性和/或遭受效应子功能的丧失的抗体。制备具有正确的抗原特异性的多特异性抗体制剂的替代方法是富集具有正确的重链-轻链配对的抗体的方法的开发。例如,spiess等(naturebiotechnology(2013)31:753-758)描述了一种用于从表达两种不同的半抗体的细菌共培养物中制备met-egfr双特异性抗体的方法。还已经描述了其中使双特异性抗体的重链中的至少一条的恒定区突变以改变它对亲和剂,例如蛋白a的结合亲和力的方法。这允许基于利用两条重链与亲和剂的差异结合的纯化技术来分离正确配对的重链异二聚体(参见美国专利申请公开号2010/0331527和wo2013/136186)。基于差异结合来选择正确的重链异二聚化的方法的局限性在于它们不选择具有正确的重链-轻链配对的抗体,因此这些技术通常应用于具有共有或共同的轻链的多特异性抗体。国际专利申请号pct/ep2012/071866(wo2013/064701)使用用于基于使用抗独特型结合剂,特别是抗独特型抗体分离多特异性抗体的方法解决了不正确链配对的问题。将所述抗独特型结合剂用于两步选择法中,其中使用第一试剂捕捉具有对第一抗原具有特异性的vh-vl结构域配对的抗体,随后使用第二试剂捕捉还具有对第二抗原具有特异性的第二vh-vl结构域配对的抗体。这种方法的缺点在于用于分离抗体的抗独特型结合剂必须对所制备的每一种多特异性抗体具有特异性,这取决于它的抗原结合谱。因此,尽管pct/ep2012/071866中描述的方法的原理一般适用于任何多特异性抗体的分离,但是必须根据要分离的多特异性抗体的特定vh-vl结构域配对来产生试剂,即抗独特型结合剂。技术实现要素:本发明通过提供多特异性抗原结合构建体而改进了现有技术,所述多特异性抗原结合构建体具有至少一个常规fab结合区和一个单结构域抗体(vhh)结合区。这种形式与异二聚化方法的组合迫使产生一种双特异性抗体构型。所使用的异二聚化方法迫使fab的重链区与完整的仅重链vhh结合。由于vhh链不与轻链缔合,因此fab部分的轻链区将仅与它的相应重链缔合。本发明通过提供所期望的多特异性抗原结合构建体,例如双特异性抗体而进一步改进了现有技术,所述多特异性抗原结合构建体可以基于尺寸从不期望的抗体纯化。所述所期望的多特异性抗原结合构建体将具有约112kda的尺寸,而不期望的抗体将具有约150kda和/或75kda的尺寸。在第一方面,本发明提供了一种双特异性抗原结合构建体,其包含:(a)结合第一靶抗原的单结构域抗体(vhh)结合区,其中所述单结构域抗体结合区与第一iggfc结构域多肽可操作地连接;和(b)结合第二靶抗原的常规igg抗体的fab部分,其中所述fab部分与第二iggfc结构域多肽可操作地连接;其中所述第一iggfc结构域多肽和第二iggfc结构域多肽二聚化而形成所述双特异性抗原结合构建体。在一个实施方案中,fc结构域二聚化通过杵入臼相互作用、fab臂交换(fae)、静电转向相互作用或疏水性相互作用而发生。在一个实施方案中,所述第一iggfc结构域多肽含有杵状结构取代,并且所述第二iggfc结构域多肽含有臼状结构取代。在一个实施方案中,所述杵状结构取代选自由精氨酸(r)、苯丙氨酸(f)、酪氨酸(y)、色氨酸(w)和其任何组合组成的组中。在一个实施方案中,所述臼状结构取代选自由丙氨酸(a)、天冬酰胺(n)、天冬氨酸(d)、甘氨酸(g)、丝氨酸(s)、苏氨酸(t)、缬氨酸(v)和其任何组合组成的组中。在一个实施方案中,所述双特异性抗原结合构建体具有在约100kda至约120kda的范围的分子量。在一个实施方案中,所期望的双特异性抗原结合构建体是约112kda。在另一个方面,本发明提供了一种纯化双特异性抗原结合构建体的方法,所述方法包括以下步骤:(a)提供包含具有不同尺寸的抗原结合构建体的混合抗原结合构建体组合物;和(b)基于尺寸分离所述混合抗原结合构建体组合物;其中所述所期望的双特异性抗原结合构建体包含(i)结合第一靶抗原的单结构域抗体(vhh)结合区,其中所述单结构域抗体结合区与第一iggfc结构域多肽可操作地连接;和(ii)结合第二靶抗原的常规igg抗体的fab部分,其中所述fab部分与第二iggfc结构域多肽可操作地连接;其中所述第一iggfc结构域多肽和第二iggfc结构域多肽二聚化而形成所述双特异性抗原结合构建体。在一个实施方案中,所述基于尺寸的分离包括尺寸排阻色谱法。在一个实施方案中,最初通过蛋白a、蛋白g、蛋白l或ch1选择性色谱法纯化所述混合抗原结合构建体组合物。在另一个方面,本发明提供了一种确定其它抗原结合构建体的混合物内所期望的双特异性抗原结合构建体的量的方法,所述方法包括以下步骤:(a)提供包含具有不同尺寸的抗原结合构建体的混合抗原结合构建体组合物;和(b)基于尺寸分离所述混合抗原结合构建体组合物;其中所述所期望的抗原结合构建体包含(i)结合第一靶抗原的单结构域抗体(vhh)结合区,其中所述单结构域抗体结合区与第一iggfc结构域多肽可操作地连接;和(ii)结合第二靶抗原的常规igg抗体的fab部分,其中所述fab部分与第二iggfc结构域多肽可操作地连接;其中所述第一iggfc结构域多肽和第二iggfc结构域多肽二聚化而形成所述双特异性抗原结合构建体。在一个实施方案中,所述基于尺寸的分离包括凝胶电泳。在一个实施方案中,所述所期望的双特异性抗原结合构建体是约100kda至约120kda。在一个实施方案中,所述所期望的双特异性抗原结合构建体是约112kda。在一个实施方案中,所述其它抗原结合构建体是约75kda或约150kda。附图说明图1示出了可以由具有两条不同的重链和两条不同的轻链的双特异性抗体的制备而产生的重链和轻链组合的十种不同的配对((a)-(j))。图2a-图2c描绘了抗原结合构建体的示意图。图2a描绘了常规抗体的示意图。图2b描绘了vhh结构域的示意图。图2c描绘了本发明的fab结构域/vhh双特异性抗体的示意图。图3描绘了在两个fc部分中具有杵入臼fc突变和fc失活无糖基化n297a突变的双特异性抗体的抗原结合构建体示意图。常规的fab部分含有臼状结构突变并且vhh部分含有杵状结构t366w突变。图4描绘了代表性的igg2fc区的氨基酸序列比对。序列如下:migg2a臼状结构:seqidno:32;migg2a杵状结构:seqidno:33;mfc融合igg2a臼状结构:seqidno:34;mfc融合igg2a杵状结构:seqidno:35;和pupex36-migg2.p9:seqidno:36。图5a-图5b描绘了抗原结合构建体的考马斯亮蓝染色的sds-page。示出了未还原样品和还原样品的染色凝胶。图5a描绘了针对靶a和靶b的抗原结合构建体的sds-page。图5b描绘了针对靶c和靶d的抗原结合构建体的sds-page。图6a-图6c分别描绘了抗原结合构建体vhh2h3-mfc_臼状结构+4r36b7migg_杵状结构、vhh3h2-mfc_臼状结构+4r36b7migg_杵状结构和仅vhh3h2-mfc_臼状结构的尺寸排阻色谱(sec)数据。图7a-图7d描绘了如通过biacore(spr)所测量的抗原结合构建体对(图7a)固定的靶抗原b、(图7b)固定的靶a、(图7c)pbs和(图7d)同种型对照的结合亲和力。图8a-图8c描绘了抗原结合构建体与(图8a)靶a、(图8b)靶b和(图8c)靶a-靶b复合物的结合动力学。具体实施方式a.定义“抗体”或“免疫球蛋白”:如本文所用的术语“免疫球蛋白”包括具有两条重链和两条轻链的组合的多肽,无论它是否具有任何相关的特异性免疫反应性。“抗体”指的是这样的组装体,其对所关注的抗原(例如人类抗原)具有显著的已知的特异性免疫反应活性。对特定人类抗原的“特异性”不排除与该抗原的物种同源物的交叉反应性。抗体和免疫球蛋白包含轻链和重链,而在它们之间有或没有链间共价键。脊椎动物系统中的基本免疫球蛋白结构被相对充分了解。通用术语“免疫球蛋白”包含可以在生物化学上区分的五种不同类别的抗体。所有五类抗体均落入本发明的范围内,以下论述将大体上涉及igg类别的免疫球蛋白分子。关于igg,免疫球蛋白包含具有约23,000道尔顿的分子量的两条相同的轻链多肽链和具有53,000-70,000的分子量的两条相同的重链。这四条链通过二硫键连接成“y”构型,其中轻链从“y”的口部开始并且继续通过可变区包围重链。抗体的轻链被分类为κ或λ(κ、λ)。每一个重链类别可以与κ轻链或λ轻链结合。一般来说,在免疫球蛋白由杂交瘤、b细胞或遗传工程化的宿主细胞产生时,轻链和重链彼此共价键合,并且两条重链的“尾区”部分通过共价二硫键或非共价键彼此键合。在重链中,氨基酸序列从y构型的分叉末端处的n末端延伸到每一条链的尽头处的c末端。本领域技术人员将了解的是,重链被分类为γ、μ、α、δ或ε,在它们当中存在一些亚类(例如γ1-γ4)。这条链的性质决定了抗体的“类别”分别是igg、igm、iga、igg或ige。免疫球蛋白亚类(同种型),例如igg1、igg2、igg3、igg4、iga1等被充分表征并且已知赋予功能特化。鉴于本公开,这些类别和同种型中的每一种的修饰型式对于本领域技术人员来说是容易辨别的并且因此,落入本发明的范围内。如上文所示,抗体的可变区允许抗体选择性地识别并且特异性地结合抗原上的表位。也就是说,抗体的vl结构域和vh结构域组合形成限定三维抗原结合位点的可变区。该四级抗体结构形成了存在于y的每个臂的末端处的抗原结合位点。更具体地,抗原结合位点是由vh链和vl链中的每一个上的三个互补决定区(cdr)限定的。“分离的抗体”:如本文所用的“分离的抗体”是已经从它的天然环境的组分分离和/或回收的抗体。它的天然环境的污染物组分是会干扰抗体的诊断用途或治疗用途的物质,并且可以包括酶、激素和其它蛋白质或非蛋白质组分。分离的抗体包括原位处于重组细胞内的抗体,这是因为抗体的天然环境中的至少一种组分将不会存在。然而,通常,将通过至少一个纯化步骤来制备分离的抗体。“亲和变体”:如本文所用的术语“亲和变体”指的是变体抗体,所述变体抗体与参考抗体相比在氨基酸序列上表现出一个或多个变化,其中所述亲和变体与参考抗体相比表现出改变的对蛋白质的亲和力。通常,与参考抗体相比,亲和变体将表现出提高的对蛋白质的亲和力。所述提高可以是更低的kd、更快的解离速率或与所述蛋白质的非人类同源物的交叉反应模式的改变。与参考抗体相比,亲和变体通常在cdr中的氨基酸序列上表现出一个或多个变化。这样的取代可以引起存在于cdr中的给定位置处的原始氨基酸被不同的氨基酸残基置换,所述不同的氨基酸残基可以是天然存在的氨基酸残基或非天然存在的氨基酸残基。所述氨基酸取代可以是保守的或非保守的。“结合位点”:如本文所用的术语“结合位点”包含多肽的区域,其负责选择性地结合所关注的靶抗原(例如人类抗原)。结合结构域包含至少一个结合位点。示例性结合结构域包括抗体可变结构域。本发明的抗体分子可以包含多个(例如两个、三个或四个)结合位点。“骆驼科动物来源的”:在某些优选的实施方案中,本发明的抗原结合构建体包含源自于通过对骆驼科动物进行主动免疫接种而产生的骆驼科动物常规抗体的框架氨基酸序列和/或cdr氨基酸序列。然而,包含骆驼科动物来源的氨基酸序列的本发明的抗体可以被工程化以包含源自于人类氨基酸序列(即人类抗体)或其它非骆驼科动物哺乳动物物种的框架和/或恒定区序列。例如,人类或非人类灵长类动物框架区、重链部分和/或铰链部分可以被包括在主题抗体中。在一个实施方案中,一个或多个非骆驼科动物氨基酸可以存在于“骆驼科动物来源的”抗体的框架区中,例如骆驼科动物框架氨基酸序列可以包含一个或多个氨基酸突变,其中存在相应的人类或非人类灵长类动物氨基酸残基。此外,骆驼科动物来源的vh结构域和vl结构域或其人源化变体可以与人类抗体的恒定结构域连接以产生嵌合分子。“保守氨基酸取代”:“保守氨基酸取代”是其中氨基酸残基被具有相似侧链的氨基酸残基置换的氨基酸取代。具有相似侧链的氨基酸残基的家族在本领域中已经被定义,包括碱性侧链(例如赖氨酸、精氨酸、组氨酸)、酸性侧链(例如天冬氨酸、谷氨酸)、不带电荷的极性侧链(例如甘氨酸、天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸、酪氨酸、半胱氨酸)、非极性侧链(例如丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、甲硫氨酸、色氨酸)、β-支化侧链(例如苏氨酸、缬氨酸、异亮氨酸)和芳族侧链(例如酪氨酸、苯丙氨酸、色氨酸、组氨酸)。因此,免疫球蛋白多肽中的非必需氨基酸残基可以被来自同一侧链家族的另一个氨基酸残基置换。“表位”:术语“表位”指的是位于与抗体或抗体片段结合的肽或蛋白质上的氨基酸的特定排列。表位通常由诸如氨基酸或糖侧链的分子的化学活性表面分组组成,并且具有特定的三维结构特征和特定的电荷特征。表位可以是线性的,即涉及与单个氨基酸序列的结合,或可以是构象的,即涉及与抗原的各个区域中可能不一定连续的两个或更多个氨基酸序列的结合。“重链部分”:如本文所用的术语“重链部分”包括源自于免疫球蛋白重链的恒定结构域的氨基酸序列。包含重链部分的多肽包含以下各项中的至少一个:ch1结构域、铰链结构域(例如上铰链区、中铰链区和/或下铰链区)、ch2结构域、ch3结构域或其变体或片段。在一个实施方案中,本发明的结合分子可以包含免疫球蛋白重链的fc部分(例如铰链部分、ch2结构域和ch3结构域)。在另一个实施方案中,本发明的结合分子缺少恒定结构域的至少一部分(例如ch2结构域的全部或一部分)。在某些实施方案中,恒定结构域中的至少一个并且优选地全部源自于人类免疫球蛋白重链。例如,在一个优选的实施方案中,重链部分包含完全人类铰链结构域。在其它优选的实施方案中,重链部分包含完全人类fc部分(例如来自人类免疫球蛋白的铰链、ch2和ch3结构域序列)。在某些实施方案中,重链部分的组成恒定结构域来自不同的免疫球蛋白分子。例如,多肽的重链部分可以包含源自于igg1分子的ch2结构域和源自于igg3或igg4分子的铰链区。在其它实施方案中,恒定结构域是嵌合结构域,其包含不同免疫球蛋白分子的部分。例如,铰链可以包含来自igg1分子的第一部分和来自igg3分子或igg4分子的第二部分。如上文所述,本领域的普通技术人员将了解的是,重链部分的恒定结构域可以被修饰以使得它们在氨基酸序列上不同于天然存在的(野生型)免疫球蛋白分子。也就是说,本文公开的本发明的多肽可以包含对重链恒定结构域(ch1、铰链、ch2或ch3)中的一个或多个和/或轻链恒定结构域(cl)的改变或修饰。示例性修饰包括一个或多个结构域中一个或多个氨基酸的添加、缺失或取代。“可变区”或“可变结构域”:术语“可变”指的是以下事实,即可变结构域vh和vl的某些部分在抗体之间在序列上广泛不同并且用于每一种特定抗体对它的靶抗原的结合和特异性。然而,可变性在抗体的整个可变结构域中不是均匀分布的。它集中在vl结构域和vh结构域中的每一个中被称作“高变环”的三个区段中,它们形成抗原结合位点的一部分。vλ轻链结构域的第一高变环、第二高变环和第三高变环在本文中被称为l1(λ)、l2(λ)和l3(λ)并且可以被定义为包含vl结构域中的残基24-33(l1(λ),由9个、10个或11个氨基酸残基组成)、残基49-53(l2(λ),由3个残基组成)和残基90-96(l3(λ),由5个残基组成)(morea等,methods20:267-279(2000))。vκ轻链结构域的第一高变环、第二高变环和第三高变环在本文中被称为l1(κ)、l2(κ)和l3(κ)并且可以被定义为包含vl结构域中的残基25-33(l1(κ),由6个、7个、8个、11个、12个或13个残基组成)、残基49-53(l2(κ),由3个残基组成)和残基90-97(l3(κ),由6个残基组成)(morea等,methods20:267-279(2000))。vh结构域的第一高变环、第二高变环和第三高变环在本文中被称为h1、h2和h3并且可以被定义为包含vh结构域中的残基25-33(h1,由7个、8个或9个残基组成)、残基52-56(h2,由3个或4个残基组成)和残基91-105(h3,长度高度可变)(morea等,methods20:267-279(2000))。除非另外指明,否则术语l1、l2和l3分别指的是vl结构域的第一高变环、第二高变环和第三高变环,并且涵盖从vκ同种型和vλ同种型中获得的高变环。术语h1、h2和h3分别指的是vh结构域的第一高变环、第二高变环和第三高变环,并且涵盖从已知重链同种型中的任一种中获得的高变环,所述已知重链同种型包括γ、ε、δ、α或μ。高变环l1、l2、l3、h1、h2和h3可以各自构成如下文所定义的“互补决定区”或“cdr”的一部分。术语“高变环”和“互补决定区”不是严格同义的,这是因为高变环(hv)是基于结构定义的,而互补决定区(cdr)是基于序列可变性定义的(kabat等,sequencesofproteinsofimmunologicalinterest(有免疫学意义的蛋白质序列),第5版,马里兰州贝塞斯达国家卫生研究院的公共卫生服务部(publichealthservice,nationalinstitutesofhealth,bethesda,md.),1983)并且在一些vh结构域和vl结构域中hv和cdr的限制可能是不同的。vl结构域和vh结构域的cdr通常可以被定义为包含以下氨基酸:轻链可变结构域中的残基24-34(cdrl1)、残基50-56(cdrl2)和残基89-97(cdrl3);和重链可变结构域中的残基31-35或31-35b(cdrh1)、残基50-65(cdrh2)和残基95-102(cdrh3)(kabat等,sequencesofproteinsofimmunologicalinterest(有免疫学意义的蛋白质序列),第5版,马里兰州贝塞斯达国家卫生研究院的公共卫生服务部(1991))。因此,hv可以被包含在相应的cdr内并且除非另外指明,否则在本文中提到vh结构域和vl结构域的“高变环”时,应当被解释为还涵盖相应的cdr,反之亦然。可变结构域的更高度保守的部分被称作框架区(fr),如下文所定义。天然重链和轻链的可变结构域各自包含四个fr(分别是fr1、fr2、fr3和fr4),它们主要采用β-折叠构型,通过三个高变环连接。每一条链中的高变环通过fr紧密靠近地保持在一起,并且连同来自另一条链的高变环一起,有助于形成抗体的抗原结合位点。抗体的结构分析揭示了由互补决定区形成的结合位点的序列与形状之间的关系(chothia等,j.mol.biol.227:799-817(1992));tramontano等,j.mol.biol,215:175-182(1990))。尽管它们具有高度的序列可变性,但是这六个环中的五个仅采用了一小部分的主链构象,被称作“规范结构”。这些构象首先由环的长度决定,其次由环中和框架区中某些位置处关键残基的存在决定,这些关键残基通过它们的堆积、氢键键合或呈现不寻常的主链构象的能力决定构象。“cdr”:如本文所用的术语“cdr”或“互补决定区”意指在重链多肽和轻链多肽的可变区内都存在的非连续抗原结合位点。这些特定区域已经由kabat等,j.biol.chem.252,6609-6616(1977)和kabat等,sequencesofproteinofimmunologicalinterest(有免疫学意义的蛋白质序列)(1991)以及chothia等,j.mol.biol.196:901-917(1987)和maccallum等,j.mol.biol.262:732-745(1996)描述,其中定义包括在彼此比较时氨基酸残基的重叠或子集。示出了构成如由上文所引用的参考文献中的每一篇所定义的cdr的氨基酸残基以进行比较。优选的是,术语“cdr”是如由kabat基于序列比较所定义的cdr。表1:cdr定义1残基编号遵循kabat等(同上)的命名法。2残基编号遵循chothia等(同上)的命名法。3残基编号遵循maccallum等(同上)的命名法。根据kabat等给出的vh结构域的一般编号对vhh结构域的氨基酸残基进行编号。应当指出的是,如本领域中对于vh结构域和vhh结构域所公知的那样,每一个cdr中氨基酸残基的总数可以变化并且可能不对应于由kabat编号所指示的氨基酸残基的总数(也就是说,根据kabat编号的一个或多个位置可能不存在于实际序列中,或实际序列可能含有比kabat编号所允许的数量更多的氨基酸残基)。这意味着,一般,根据kabat的编号可能对应于或可能不对应于实际序列中氨基酸残基的实际编号。关于vhh结构域编号的细节描述于美国专利号8,703,131中,该美国专利以引用的方式并入本文。“框架区”:如本文所用的术语“框架区”或“fr区”包括作为可变区的一部分,但是不是cdr的一部分的氨基酸残基(例如使用cdr的kabat定义)。因此,可变区框架具有约100个-120个氨基酸的长度,但是仅包括cdr之外的那些氨基酸。对于重链可变区的具体实例和如由kabat等所定义的cdr,框架区1对应于包含氨基酸1-30的可变区的结构域;框架区2对应于包含氨基酸36-49的可变区的结构域;框架区3对应于包含氨基酸66-94的可变区的结构域;并且框架区4对应于从氨基酸103到可变区的末端的可变区的结构域。轻链的框架区类似地由轻链可变区cdr中的每一个隔开。类似地,使用chothia等或mccallum等的cdr定义,框架区边界由如上文所述的对应的cdr末端隔开。在优选的实施方案中,cdr如由kabat所定义。在天然存在的抗体中,在每一个单体抗体上存在的六个cdr是短的非连续的氨基酸序列,它们在抗体在水性环境中呈现它的三维构型时特异性定位以形成抗原结合位点。重链可变结构域和轻链可变结构域的其余部分在氨基酸序列上显示出更小的分子间可变性并且被称作框架区。框架区主要采用β-折叠构象并且cdr形成环,这些环连接β-折叠结构并且在一些情况下,形成β-折叠结构的一部分。因此,这些框架区用以形成通过链间非共价相互作用使得六个cdr以正确的取向定位的支架。由定位的cdr形成的抗原结合位点限定了与免疫反应性抗原上的表位互补的表面。该互补表面促进了抗体与免疫反应性抗原表位的非共价结合。cdr的位置可以容易地由本领域的普通技术人员鉴定出。“铰链区”:如本文所用的术语“铰链区”包括重链分子的将ch1结构域与ch2结构域连接的部分。该铰链区包含约25个残基并且是柔性的,从而允许两个n末端抗原结合区独立地移动。铰链区可以被细分成三个不同的结构域:上铰链结构域、中铰链结构域和下铰链结构域(roux等,j.immunol.161:4083(1998))。包含“完全人类”铰链区的抗体可以含有下表2中所示的铰链区序列之一。表2:人类铰链序列“片段”:术语“片段”指的是抗体或抗体链的一部分或部分,其包含比完整或完全的抗体或抗体链更少的氨基酸残基。术语“抗原结合片段”指的是免疫球蛋白或抗体的多肽片段,其结合抗原或与完整抗体(即与作为它们的来源的完整抗体)竞争结合抗原(即特异性结合人类抗原)。如本文所用的术语抗体分子的“片段”包括抗体的抗原结合片段,例如抗体轻链(vl)、抗体重链(vh)、单链抗体(scfv)、f(ab')2片段、fab片段、fd片段、fv片段和单结构域抗体片段(dab)。片段可以例如经由对完整或完全抗体或抗体链进行化学或酶处理或通过重组手段获得。“特异性”:术语“特异性”指的是与给定的靶抗原,例如人类靶抗原特异性结合(例如免疫反应)的能力。抗体或其抗原结合片段可以是单特异性的并且含有特异性结合靶标的一个或多个结合位点,或抗体或其抗原结合片段可以是多特异性的并且含有特异性结合相同或不同的靶标的两个或更多个结合位点。在某些实施方案中,本发明的抗原结合构建体对多于一种靶抗原具有特异性。在某些实施方案中,本发明的抗原结合构建体对两种靶抗原具有特异性。在某些实施方案中,本发明的抗原结合构建体对多于一种人类靶抗原具有特异性。在某些实施方案中,本发明的抗原结合构建体对两种人类靶抗原具有特异性。例如,在一个实施方案中,本发明的双特异性结合分子与靶抗原a(靶a)和靶抗原b(靶b)结合。作为另一个实例,在一个实施方案中,本发明的双特异性结合分子与靶抗原c(靶c)和靶抗原d(靶d)结合。“合成”:如本文所用的关于多肽的术语“合成”包括包含非天然存在的氨基酸序列的多肽。例如,非天然存在的多肽,其是天然存在的多肽的修饰形式(例如包含突变,如添加、取代或缺失)或包含第一氨基酸序列(其可能是或可能不是天然存在的),所述第一氨基酸序列与在自然界中不与它天然连接的第二氨基酸序列(其可能是或可能不是天然存在的)连接成线性氨基酸序列。“工程化”:如本文所用的术语“工程化”包括通过合成手段(例如通过重组技术、体外肽合成、通过肽的酶促或化学偶联或这些技术的某种组合)操纵核酸或多肽分子。优选的是,本发明的抗原结合构建体是工程化的,包括例如人源化和/或嵌合抗体,和已经被工程化以改善一种或多种特性,如抗原结合、稳定性/半衰期或效应子功能的抗体。“fc区”:如本文所用的术语“fc区”指的是天然免疫球蛋白的由它的两条重链的fc结构域形成的部分。天然fc区是同二聚体。如本文所用的术语“变体fc区”指的是相对于天然fc区具有一个或多个改变的fc区。可以通过氨基酸取代、添加和/或缺失、另外的部分的连接和/或天然聚糖的改变来改变fc区。所述术语涵盖了其中组成fc结构域中的每一个不同的fc区。异二聚体fc区的实例包括但不限于使用如例如美国专利号8,216,805中所述的“杵入臼”技术制备的fc区。所述术语还涵盖其中组成fc结构域由接头部分连接在一起的单链fc区,如例如美国专利申请公开号2009/0252729a1和us2011/0081345a1中所述。除非另有说明,否则所有抗体恒定区编号均指的是eu编号方案,如edelman等,proc.natl.acad.sci.usa63(1):78-85(1969)中所述。“人源化取代”:如本文所用的术语“人源化取代”指的是氨基酸取代,其中存在于抗体(例如骆驼科动物来源的抗体)的vh结构域或vl结构域中的特定位置处的氨基酸残基被存在于参考人类vh结构域或vl结构域中的等同位置处的氨基酸残基置换。参考人类vh结构域或vl结构域可以是由人类种系编码的vh结构域或vl结构域。可以在本文定义的抗体的框架区和/或cdr中进行人源化取代。“人源化抗体或变体”:如本文所用的术语“人源化抗体”或“人源化变体”指的是与参考抗体相比含有一个或多个“人源化取代”的变体抗体,其中所述参考抗体的一部分(例如vh结构域和/或vl结构域或其含有至少一个cdr的部分)具有源自于非人类物种的氨基酸,并且“人源化取代”发生在源自于非人类物种的氨基酸序列内。“仅重链抗体”或“vhh抗体”:如本文所用的术语“仅重链抗体”或“vhh抗体”指的是仅由骆驼科的物种产生的第二类型的抗体,所述物种包括骆驼、美洲驼、羊驼。仅重链抗体由两条重链构成并且不含轻链。每一条重链在n末端处均具有可变结构域,并且这些可变结构域被称为“vhh”结构域以将它们与常规的异四聚体抗体的重链的可变结构域(即上述vh结构域)区分开。“修饰的抗体”:如本文所用的术语“修饰的抗体”包括抗体的合成形式,其被改变以使得它们不是天然存在的,例如包含至少两个重链部分,但是不包含两条完整重链的抗体(如结构域缺失的抗体或微型抗体);抗体的多特异性形式(例如双特异性、三特异性等),其被改变以与两种或更多种不同的抗原或单个抗原上的不同表位结合;与scfv分子连接的重链分子等。scfv分子是本领域已知的并且描述于例如美国专利号5,892,019中。此外,术语“修饰的抗体”包括抗体的多价形式(例如三价、四价等抗体,其与相同抗原的三个或更多个拷贝结合)。在另一个实施方案中,本发明的修饰的抗体是融合蛋白,其包含缺少ch2结构域的至少一个重链部分并且包含多肽的结合结构域,所述多肽包含受体配体对的一个成员的结合部分。术语“修饰的抗体”在本文中也可以用于指的是抗体的氨基酸序列变体。本领域的普通技术人员将了解的是,可以修饰抗体以产生变体抗体,所述变体抗体与作为它的来源的抗体相比在氨基酸序列上不同。例如,可以进行核苷酸取代或氨基酸取代,从而在“非必需”氨基酸残基处引起保守取代或变化(例如在cdr和/或框架残基中)。氨基酸取代可以包括一个或多个氨基酸被天然存在或非天然的氨基酸置换。“靶a”:如本文所用的术语“靶a”或“靶抗原a”指的是靶抗原,本发明的抗体的含有常规fab的部分对所述靶抗原具有结合特异性。“靶b”:如本文所用的术语“靶b”或“靶抗原b”指的是靶抗原,本发明的抗体的含有vhh的部分对所述靶抗原具有结合特异性。靶a和靶b可以一起形成复合物。在某些实施方案中,对靶a和靶b具有特异性的本发明的双特异性抗体也可以结合靶a和靶b的复合物。“靶c”:如本文所用的术语“靶c”或“靶抗原c”指的是靶抗原,本发明的抗体的含有常规fab的部分对所述靶抗原具有结合特异性。“靶d”:如本文所用的术语“靶d”或“靶抗原d”指的是靶抗原,本发明的抗体的含有vhh的部分对所述靶抗原具有结合特异性。“抗原结合构建体”:如本文所用的术语“抗原结合构建体”包含与iggfc结构域部分融合的单结构域抗体(vhh)结合区;常规igg抗体的重链-fc结构域部分;和所述常规igg抗体的轻链部分。vhh结合区是由单个单体可变抗体结构域组成的抗体片段。它选择性地结合特定的第一抗原。常规igg抗体的重链-fc结构域部分包括常规igg重链的可变结构域和恒定结构域。常规igg抗体的轻链部分包括常规igg轻链的可变结构域和恒定结构域。常规igg抗体的重链和轻链部分选择性地结合特定的第二抗原。b.多特异性抗原结合构建体本发明的多特异性抗原结合构建体的一个组分是常规抗体或其抗原结合片段的重链和轻链,其中术语“常规抗体”在本文中用于描述含有根据图1中所示的“y”构型排列的免疫球蛋白重链和轻链的异四聚体抗体。这样的常规抗体可以源自于任何合适的物种,包括但不限于美洲驼、小鼠、大鼠、兔、山羊、仓鼠、鸡、猴或人类来源的抗体。在某些示例性实施方案中,所述常规抗体是如例如美国专利号us8,524,231中所描述和要求保护的simple抗体,该美国专利的全部内容以引用的方式并入本文。simple抗体可以包含重链可变结构域(vh)和轻链可变结构域(vl),其中所述vh结构域和/或vl结构域或其一个或多个互补决定区(cdr)源自于骆驼科的动物,即源自于通过对骆驼科动物(例如美洲驼)进行免疫接种而产生的常规抗体。表现出高人类同源性或具有至少一个骆驼科动物来源的cdr序列、vh结构域和/或vl结构域的抗体或抗原结合片段可以是来自骆驼科动物常规抗体的vh结构域或vl结构域的人源化或种系化变体,其中术语“人源化”和“种系化”如本文其它地方所定义。在某些实施方案中,常规抗体抗原结合区可以被称为“fab”(抗原结合片段)。所述fab包含来自重链和轻链中的每一个的一个恒定结构域和一个可变结构域。可变重链和轻链含有负责抗原结合的cdr。常规抗体的fab部分见于图2a和图2c的示意图中。本发明的多特异性抗原结合构建体的另一个组分包含vhh抗体或纳米抗体的vhh结构域或重链。作为骆驼科动物来源的重链抗体的vhh抗体由两条重链构成并且没有轻链(hamers-casterman等,nature.1993;363;446-8)。vhh抗体的每一条重链在n末端都具有可变结构域,并且这些可变结构域在本领域中被称为“vhh”结构域以将它们与常规抗体的重链的可变结构域(即vh结构域)区分开。类似于常规抗体,所述分子的vhh结构域赋予抗原结合特异性,并且因此,vhh抗体或片段,如分离的vhh结构域适合作为本公开的抗原结合构建体的组分。对于本公开的多特异性抗原结合构建体,可以通过用包含对应的选择性靶抗原的多肽对宿主物种进行主动免疫接种而产生或获得对所述抗原具有特异性的常规异四聚体抗体或vhh抗体。为了产生常规抗体,宿主物种可以选自以下各项中的任一种:小鼠、大鼠、兔、山羊、仓鼠、鸡、猴或骆驼科的物种。为了产生vhh抗体,可以用包括对应抗原的多肽对来自骆驼科的任何物种(包括美洲驼物种)进行免疫接种。本发明的示例性抗原结合构建体示于下文中。表3:靶a/靶b抗体的抗原结合构建体序列。根据kabat编号的hcdr1-hcdr3和lcdr1-lcdr3是粗体的。表4:靶c/靶d抗体的抗原结合构建体序列。c.vhh和常规抗体的二聚化本发明克服了传统的双特异性抗体形式存在的链错配的问题。例如,传统的双特异性抗体链配对可能会产生多达十种不同的抗体种类(图1)。在十种不同的可能的选择方案下,所期望的双特异性抗体的产率一般较低并且难以从其它抗体的群体中分离所期望的双特异性抗体。在一些情况下,可以通过使用迫使形成某些所期望的构型的异二聚化方法来部分克服这个问题。例如,下文所述的异二聚化方法迫使发生特定的fc结构域相互作用,从而产生四种可能的构型之一(图1(a)-图1(d))。然而,令人遗憾的是,这些异二聚化方法仍然允许轻链的随机配对并且阻止了具有单一构型的双特异性抗体的均一产生。为了克服本领域中的这种缺陷,本发明的多特异性抗原结合构建体包含结合第一靶抗原的单个vhh结合区,其中所述vhh结合区与第一iggfc结构域多肽可操作地连接;和结合第二靶抗原的常规igg抗体的fab部分,其中所述fab部分与第二iggfc结构域多肽可操作地连接,并且其中所述第一iggfc结构域多肽和第二iggfc结构域多肽二聚化而形成双特异性抗原结合构建体(图2c)。所述fab的轻链部分只能与fab的重链部分配对。因此,这确保了仅获得所期望的双特异性构型。在本发明的一个方面,所述抗原结合构建体的两个fc结构域通过杵入臼配对而异二聚化。这种二聚化技术利用了被工程化到ch3结构域的界面中的隆起(“杵状结构”)和空腔(“臼状结构”)。在第一ch3结构域或第二ch3结构域的界面处存在适当定位和尺寸设定的杵状结构或臼状结构的情况下,仅需要在相邻的界面处分别对相应的臼状结构或杵状结构进行工程化,从而促进并且增强ch3/ch3结构域界面中的fc结构域配对。与vhh融合的iggfc结构域具有杵状结构,并且常规抗体的iggfc结构域具有被设计成容纳所述杵状结构的臼状结构,或反之亦然。“杵状结构”指的是从第一fc结构域的ch3部分的界面突出的至少一个氨基酸侧链,通常是较大的侧链。所述突出产生“杵状结构”,其与第二fc结构域的ch3部分中的“臼状结构”互补并且被“臼状结构”容纳。“臼状结构”是从第二fc结构域的ch3部分的界面内缩的至少一个氨基酸侧链,通常是较小的侧链。该技术描述于例如美国专利号5,821,333;ridgway等,proteinengineering9:617-621(1996);和carterp.,j.immunol.methods248:7-15(2001)中。可以用作杵状结构的示例性氨基酸残基包括精氨酸(r)、苯丙氨酸(f)、酪氨酸(y)和色氨酸(w)。ch3结构域中存在的氨基酸残基可以被杵状结构氨基酸残基置换或取代。要被取代的优选的氨基酸可以包括具有小侧链的任何氨基酸,如丙氨酸(a)、天冬酰胺(n)、天冬氨酸(d)、甘氨酸(g)、丝氨酸(s)、苏氨酸(t)或缬氨酸(v)。可以用作臼状结构的示例性氨基酸残基包括丙氨酸(a)、丝氨酸(s)、苏氨酸(t)和缬氨酸(v)。ch3结构域中存在的氨基酸残基可以被臼状结构氨基酸残基置换或取代。要被取代的优选的氨基酸可以包括具有大或大体积的侧链的任何氨基酸,如精氨酸(r)、苯丙氨酸(f)、酪氨酸(y)或色氨酸(w)。ch3结构域优选地源自于人类igg1抗体。ch3结构域的示例性氨基酸取代包括t366y、t366w、f405a、f405w、y407t、y407a、y407v、t394s和其组合。优选的示例性组合是用于第一ch3结构域上的杵状结构突变的t366y或t366w和用于第二ch3结构域上的臼状结构突变的y407t或y407v。在本发明的某些实施方案中,所述抗原结合构建体的两个fc结构域通过fab臂交换(fae)而异二聚化。具有p228s铰链突变的人类igg1可以含有f405l或k409rch3结构域突变。将这两种抗体与还原剂混合会引起fae。这种技术描述于美国专利号9,212,230和labrijna.f.,procnatlacadsciusa110(13):5145-5150(2013)中。在本发明的某些实施方案中,所述抗原结合构建体的两个fc结构域通过静电转向效应而异二聚化。该二聚化技术利用静电转向来促进和增强ch3/ch3结构域界面中的fc结构域配对。相对于同二聚化(相同电荷配对),改变两个ch3结构域之间的电荷互补性有利于促进异二聚化(相反电荷配对)。在这种方法中,静电排斥力阻止同二聚化。示例性的氨基酸残基取代可以包括第一ch3结构域中的k409d、k392d和/或k370d,以及第二ch3结构域中的d399k、e356k和/或e357k。该技术描述于美国专利申请公开号2014/0154254a1和gunasekarank.,jbiolchem285(25):19637-19646(2010)中。在本发明的某些实施方案中,所述抗原结合构建体的两个fc结构域通过疏水性相互作用效应而异二聚化。该二聚化技术利用疏水性相互作用而非静电相互作用来促进和增强ch3/ch3结构域界面中的fc结构域配对。示例性的氨基酸残基取代可以包括第一ch3结构域中的k409w、k360e、q347e、y349s和/或s354c,以及第二ch3结构域中的d399v、f405t、q347r、e357w和/或y349c。在第一ch3结构域与第二ch3结构域之间的优选的氨基酸残基取代对包括k409w:d399v、k409w:f405t、k360e:q347r、y349s:e357w和s354c:y349c。该技术描述于美国专利申请公开号2015/0307628a1中。在本发明的某些实施方案中,可以通过使用亮氨酸拉链融合体来介导异二聚化。与抗体链的每一个ch3结构域的c末端融合的亮氨酸拉链结构域迫使进行异二聚化。该技术描述于wranikb.,jbiolchem287(52):43331-43339(2012)中。在本发明的一个方面,可以通过使用链交换工程结构域(seed)体来介导异二聚化。源自于igg和iga形式的ch3结构域迫使进行异二聚化。该技术描述于mudam.,proteineng.des.sel.24(5):447-454(2011)中。除非另有说明,否则本文所用的所有抗体恒定区编号均对应于eu编号方案,如edelman等,proc.natl.acad.sci.usa63(1):78-85(1969)中所述。重链和/或轻链的异二聚化以及不对称抗体的产生和纯化的另外的方法是本领域已知的。参见例如kleinc.,mabs4(6):653-663(2012)和美国专利号9,499,634,这些文献中的每一篇以引用的方式并入本文。d.fc区的基于非异二聚化的修饰本发明的抗体分子可以在重链和/或轻链的恒定区内,特别是在fc区内具有一个或多个氨基酸取代、插入或缺失。氨基酸取代可以引起取代的氨基酸被不同的天然存在的氨基酸或非天然或修饰的氨基酸置换。还容许其它结构修饰,例如像糖基化模式的变化(例如通过n-连接或o-连接的糖基化位点的添加或缺失)。可以在fc区内修饰本发明的双特异性抗原结合构建体以增加对新生儿fc受体(fcrn)的结合亲和力。增加的结合亲和力可以是在酸性ph值(例如约ph5.5至约ph6.0)下可测量的。增加的结合亲和力也可以是在中性ph值(例如约ph6.9至约ph7.4)下可测量的。“增加的结合亲和力”意指相对于未修饰的fc区,对fcrn的结合亲和力增加。通常,未修饰的fc区将具有人类igg1、igg2、igg3或igg4的野生型氨基酸序列。在这样的实施方案中,将相对于野生型igg1、igg2、igg3或igg4对fcrn的结合亲和力来测量具有修饰的fc区的抗体分子的增加的fcrn结合亲和力。已经报道了几种fc取代会增加fcrn结合并且从而改善抗体药物代谢动力学。这样的取代在例如zalevsky等(2010)nat.biotechnol.28(2):157-9;hinton等(2006)jimmunol.176:346-356;yeung等(2009)jimmunol.182:7663-7671;prestalg.(2008)curr.opin.immunol.20:460-470;和vaccaro等(2005)nat.biotechnol.23(10):1283-88中有所报道,这些文献的内容以引用的方式整体并入本文。在某些实施方案中,本文所述的双特异性抗原结合构建体的抗体分子中的一种或多种包含修饰的人类iggfc结构域,其包含修饰,所述修饰包含氨基酸取代h433k和n434f或由氨基酸取代h433k和n434f组成,其中fc结构域编号是根据eu编号。在另一个特定的实施方案中,本文所述的组合的抗体分子中的一种或多种包含修饰的人类iggfc结构域,其包含修饰,所述修饰包含氨基酸取代m252y、s254t、t256e、h433k和n434f或由氨基酸取代m252y、s254t、t256e、h433k和n434f组成,其中fc结构域编号是根据eu编号。所述双特异性抗原结合构建体也可以被修饰以形成免疫缀合物,所述免疫缀合物包含与细胞毒性剂缀合的抗体,所述细胞毒性剂诸如化学治疗剂、毒素(例如细菌、真菌、植物或动物来源的酶活性毒素或其片段)或放射性同位素(即放射性缀合物)。fc区也可以被工程化以延长半衰期,如chan和carter(2010)naturereviews:immunology10:301-316所述,该文献以引用的方式并入本文。在某些实施方案中,fc区可以被工程化以使得它缺乏效应子功能,例如缺乏引导抗体依赖性细胞毒性(adcc)或补体依赖性细胞毒性(cdc)的能力。这样的工程化的无效应子的fc区在本文中被称为“fc失活”。在某些实施方案中,本发明的双特异性抗原结合构建体可以具有源自于具有降低的效应子功能的天然存在的igg同种型(例如igg4)的fc区。可以进一步修饰源自于igg4的fc区以增加治疗效用,例如通过引入使体内igg4分子之间的臂交换减到最低限度的修饰。可以修饰源自于igg4的fc区以包括s228p取代。在某些实施方案中,本发明的双特异性抗原结合构建体在糖基化方面被修饰。例如,可以制备无糖基化的抗体(即所述抗体缺乏糖基化)。可以改变糖基化以例如增加抗体对靶抗原的亲和力。这样的碳水化合物修饰可以通过例如改变抗体序列内一个或多个糖基化位点来实现。例如,可以进行一个或多个氨基酸取代,其引起一个或多个可变区框架糖基化位点的消除,从而消除该位点处的糖基化。这样的无糖基化可以增加抗体对抗原的亲和力。在某些实施方案中,可以对fc区进行工程化以减少或消除糖基化。例如,可以使ch2结构域中的n297位点突变以减少或消除糖基化,其中fc结构域编号是根据eu编号。在某些实施方案中,本发明的双特异性抗原结合构建体包含一个或多个n297a突变。已经证实n297a突变会降低抗体依赖性细胞毒性(adcc)。参见例如borrok等(2012)acschembiol.7(9):1596-1602。e.编码多特异性抗原结合构建体的多核苷酸本发明还提供了编码本发明的多特异性抗原结合构建体的多核苷酸分子,以及表达载体,所述表达载体含有与调节序列可操作地连接的编码本发明的多特异性抗原结合构建体的核苷酸序列,所述调节序列允许所述多特异性抗原结合构建体多肽在宿主细胞或无细胞表达系统中表达。本发明还提供了含有这些表达载体的宿主细胞或无细胞表达系统。编码本发明的多特异性抗原结合构建体的多核苷酸分子包括例如重组dna分子。如本文所用的术语“核酸”、“多核苷酸”和“多核苷酸分子”是可互换的并且指的是任何dna分子或rna分子,无论是单链还是双链,并且如果是单链,则也指它的互补序列的分子。在论述核酸分子时,特定核酸分子的序列或结构在本文中可以根据在5'到3'方向上提供序列的常规惯例来描述。在本发明的一些实施方案中,核酸或多核苷酸是“分离的”。该术语在应用于核酸分子时指的是这样的核酸分子,在作为它的来源的生物体的天然存在的基因组中所述核酸分子与它直接相邻的其它组分(包括序列)分离。例如,“分离的核酸”可以包含插入载体(如质粒或病毒载体)中或整合到原核或真核细胞或非人类宿主生物体的基因组dna中的dna分子。当应用于rna时,术语“分离的多核苷酸”主要指的是由如上文所定义的分离的dna分子编码的rna分子。可选地,所述术语可以指的是这样的rna分子,所述rna分子已经从在它的天然状态下(即在细胞或组织中)原本与它缔合的其它核酸纯化/分离。分离的多核苷酸(dna或rna)可以进一步代表通过生物学手段或合成手段直接制备并且与在它的制备期间存在的其它组分分离的分子。为了重组产生根据本发明的多特异性抗原结合构建体,可以制备(使用标准分子生物学技术)编码各种构建体组分的重组多核苷酸并且将其插入可复制的载体中以在所选择的宿主细胞或无细胞表达系统中表达。合适的宿主细胞可以是原核生物、酵母或高等真核生物细胞,特别是哺乳动物细胞。有用的哺乳动物宿主细胞系的实例是由sv40转化的猴肾cv1细胞系(cos-7,atcccrl1651);人类胚肾细胞系(被亚克隆以在悬浮培养中生长的293或293细胞,graham等,j.gen.virol.36:59(1977));幼仓鼠肾细胞(bhk,atccccl10);中国仓鼠卵巢细胞/-dhfr(cho,urlaub等,proc.natl.acad.sci.usa77:4216(1980));小鼠塞尔托利氏细胞(tm4,mather,biol.reprod.23:243-251(1980));小鼠骨髓瘤细胞sp2/0-ag14(atcccrl1581;atcccrl8287)或ns0(hpa培养物保藏号85110503);猴肾细胞(cv1atccccl70);非洲绿猴肾细胞(vero-76,atcccrl-1587);人类宫颈癌细胞(hela,atccccl2);犬肾细胞(mdck,atccccl34);布法罗大鼠肝细胞(brl3a,atcccrl1442);人类肺细胞(w138,atccccl75);人类肝细胞(hepg2,hb8065);小鼠乳腺肿瘤(mmt060562,atccccl51);tri细胞(mather等,annalsn.y.acad.sci.383:44-68(1982));mrc5细胞;fs4细胞;和人类肝细胞瘤细胞系(hepg2)以及dsm公司的perc-6细胞系。适用于这些宿主细胞中的每一种中的表达载体也是本领域公知的。应当指出的是,术语“宿主细胞”一般指培养的细胞系。已经引入了编码根据本发明的抗原结合多肽的表达载体的整个人类明确地被排除在“宿主细胞”的定义之外。f.多特异性抗原结合构建体的治疗效用如本文所用的术语“治疗(treat)”、“治疗(treating)”或“治疗(treatment)”意指减慢、中断、阻止、控制、停止症状、病症、疾患或疾病、降低其严重程度,但是不一定涉及所有疾病相关的症状、疾患或病症的完全消除。对于人类治疗用途,本文所述的多特异性抗原结合构建体可以“有效量”向需要治疗的人类受试者施用。术语“有效量”指的是足以实现所期望的结果的药剂的量或剂量。在某些实施方案中,术语“有效量”指的是在向受试者单剂量或多剂量施用后,在疾病的治疗方面提供治疗功效的药剂的剂量。在某些实施方案中,术语“有效量”指的是在向人类患者单剂量或多剂量施用后,在疾病的治疗方面提供治疗功效的药剂的剂量。例如,术语“有效量”可以指的是在向患有疾病的人类患者单剂量或多剂量施用后,在疾病的治疗方面提供治疗功效的本发明的多特异性抗原结合构建体的量或剂量。所述多特异性抗原结合构建体的治疗有效量可以包括在每单次剂量每kg体重约0.1mg至每kg体重约20mg的范围的量。在某些实施方案中,所述多特异性抗原结合构建体的治疗有效量可以包括在每单次剂量每kg体重约1mg至每kg体重约10mg的范围的量。用于任何单个患者的治疗有效量可以取决于诸如患者的年龄和总体状况以及要治疗的疾病的性质和严重程度的因素。用于任何单个患者的治疗有效量可以由医疗保健专业人员,通过例如监测抗原结合构建体对生物标志物(如肿瘤组织中靶抗原的细胞表面表达)或症状(如肿瘤消退)等的作用来确定。在任何给定时间点施用的抗原结合构建体的量可以变化以使在治疗过程中施用最佳量的抗原结合构建体,无论是单独使用还是与任何其它治疗剂组合使用。还考虑将本文所述的抗原结合构建体或包含这样的构建体的药物组合物与任何其它治疗联合施用,作为联合疗法。g.药物组合物本发明的范围包括药物组合物,所述药物组合物含有本发明的抗原结合构建体,用一种或多种药学上可接受的载体或赋形剂配制。用于配制用于人类治疗用途的诸如单克隆抗体的抗原结合构建体的技术是本领域公知的并且例如在wang等,jpharmsci96:1-26(2007)中进行了综述。所述药物组合物可以被配制用于通过任何合适的施用途径施用,包括但不限于静脉内(i.v.)、腹膜内(i.p.)、肌内(i.m.)、肿瘤内、口服、其它肠内、皮下(s.c.)和肺部。h.多特异性抗原结合构建体的纯化本发明的范围包括纯化本发明的多特异性抗原结合构建体的方法。纯化方法可以包括基于物理化学分级分离的方法。这样的方法包括尺寸排阻色谱法(sec)、硫酸铵沉淀、离子交换色谱法(iec)和固定化金属螯合色谱法(imac)。纯化方法可以包括亲和纯化。这样的方法依赖于蛋白a、蛋白g和/或蛋白l作为抗原结合构建体的保守区域的亲和配体。亲和配体可以与树脂缀合以促进纯化。另外的亲和纯化方法可以依赖于与抗体的iggch1结构域的特异性结合。这是ch1选择性色谱法。可以通过sec纯化本公开的多特异性结合构建体。本公开的多特异性结合构建体的尺寸预期是约112kda,其包含各自与fc结构域可操作地连接的vhh结合区和常规igg抗体(例如simple抗体)的fab部分。通过各自与fc结构域可操作地连接的两个fab部分的二聚化而形成的常规igg抗体预期是约150kda。通过各自与fc结构域可操作地连接的两个vhh部分的二聚化而形成的vhh抗体预期是约75kda。鉴于所期望的多特异性结合构建体(约112kda)与两种可能的不期望的替代结合构建体(约150kda和/或75kda)之间的尺寸差异,本领域技术人员将容易理解的是,所期望的多特异性抗原结合构建体可以容易地通过sec去除不期望的结合构建体来纯化。无论所得的多特异性抗原结合构建体是否已经通过交联而稳定化,在一些实施方案中,本发明的方法可以包括纯化多特异性抗原结合构建体的进一步的步骤。含有多特异性抗原结合构建体的混合物可以使用标准色谱技术来纯化,诸如(但不限于)标准蛋白a色谱法、蛋白g、蛋白l、阳离子/阴离子交换色谱法、尺寸排阻色谱法、疏水性相互作用色谱法、亲硫色谱法或使用被设计成捕捉igg分子的配体(蛋白a模拟物、美洲驼vhh配体等)。可选地,可以使用标准技术使多特异性抗原结合构建体沉淀,如盐诱导的沉淀(硫酸铵)、添加有机溶剂(dmso、乙醇)、改变ph值或非离子型聚合物(聚乙二醇)。在另一种背景下,可以将多特异性抗原结合构建体应用于使用允许浓缩多特异性抗原结合构建体的膜的过滤技术。可能需要所有这些技术的组合来将多特异性抗原结合构建体纯化至完全均质,这是因为某些混合物可能仍然含有亲本单特异性igg分子以及多特异性抗原结合构建体。然后可能需要另外的纯化步骤来将多特异性抗原结合构建体与亲本单特异性igg分子分离。这可以通过使用针对第一结合特异性的亲和柱进行结合和洗脱,继而使用针对第二结合特异性的亲和柱进行结合和洗脱来进行纯化而完成。(纯化的)多特异性抗原结合构建体的量、质量和纯度可以使用常规的生物化学技术来分析,如吸光度测量、hp-sec、sds-page、非变性page和rp-hplc。可以将多特异性抗原结合构建体与亲本igg分子区分开的另外的技术包括但不限于ief、cief、ciex和质谱(esi、maldi),从而允许基于电荷和/或质量非常准确地分离和检测分子。双特异性抗原结合构建体的双重结合特异性可以使用多种不同的结合测定形式中的任一种,使用例如elisa、ria、表面等离子体共振(spr)、生物层干涉测量法、delfia、fret、ecl、gyros和alfascreen来评估。纯化方法描述于美国专利号9,212,230中。以引用的方式并入在上述说明中和整个以下实施例中引用了各种专利、公开的专利申请和出版物,它们中的每一篇以引用的方式整体并入本文。实施例将参考以下非限制性实施例进一步理解本发明。实施例1:抗靶a/抗靶b抗原结合构建体的产生和纯化抗靶a是对靶a具有结合特异性的常规抗体。抗靶a能够中和鼠类cd4+脾细胞中靶a的激活。抗靶b是对靶b具有结合特异性的vhh。使用杵入臼(kih)技术产生了用于结合靶a、靶b或靶a-靶b复合物的抗靶a和抗靶b的双特异性抗原结合构建体。将抗靶bvhh与migg2a_臼状结构融合。将抗靶a-vh克隆成migg2a_杵状结构,并且将抗靶a-vl克隆成clvλ。为了产生抗靶a/抗靶b抗原结合构建体,每100ml使用20μg以下构建体中的每一种转染10×100ml的293e细胞:mfc融合igg2a杵状结构_抗靶b-vhh、migg2a臼状结构_抗靶a_vh和clvλ_抗靶a。通过kpni-noti克隆到pupe(pcdna3.1样载体)中来产生migg2a臼状结构、migg2a杵状结构、mfc融合igg2a臼状结构和mfc融合igg2a杵状结构构建体。将抗靶a_vh进行bsmbi克隆到pupe-migg2a臼状结构载体中。将抗靶b-vhh进行bsmbi克隆到pupe-mfc融合igg2a杵状结构载体中。将样品经过蛋白a珠粒纯化。将结合的抗原结合构建体洗脱并且使用sds-page分析。图5a示出了用考马斯蓝染色的sds-page凝胶,说明了以112kda的正确尺寸纯化抗原结合构建体。请注意,与样品相比,分子量标记物迁移得更快。与完整igg相比,抗原结合构建体具有更低的条带。在正常情况下,错配将引起150kda的完整igg、75kda的vhh-fc融合体和约112kda的双特异性形式的形成。然而,图5a示出了仅产生了112kda的产物,这表明了优先产生所期望的双特异性形式。在还原条件下,对于两条重链看到双重条带。请注意,mfc融合igg2a杵状结构_抗靶b-vhh不含ch1结构域并且没有轻链,从而产生约37kda的产物。migg2a臼状结构_抗靶a-vh的重链预期是约50kda。抗靶a的轻链是约25kda。因此,本发明的双特异性形式确保了仅获得正确的双特异性抗体。特别是,migg2a臼状结构_抗靶a_vh构建体仅与mfc融合igg2a杵状结构_抗靶b-vhh构建体配对。剩余的轻链clvλ_抗靶a构建体只能与抗靶a_vh链配对,这是因为轻链不能与vhh链配对。对于本领域技术人员来说将显而易见的是,本发明不限于上述的kih异二聚化方法。任何适当的异二聚化方法,如本文其它地方所述的方法均可以用于上述双特异性抗体形式。实施例2:抗靶a/抗靶b抗原结合构建体的表征在biacore(spr)上分析实施例1的抗原结合构建体与靶a和靶b的结合特征。所述构建体能够捕捉靶a和靶b这两者。(将图7a和图7b与图7c和图7d的pbs和同种型对照进行比较)。与对靶a或靶b的亲和力相比,所述抗原结合构建体对靶a-靶b复合物的亲和力高得多,这表明了二价结合(图8a-图8c和表5)。请注意,biacore的检测限度是+/-1e-12,因此图8中所示的亲和力是高估值。表5:通过biacore测量抗原结合构建体对靶a-靶b复合物、靶a和靶b的亲和力。ka(1/ms)kd(1/s)kd(m)靶a-靶b复合物5.15e+054.22e-088.20e-14靶a2.16e+055.05e-042.34e-09靶b4.86e+051.42e-042.93e-10有可能的是,图8a-图8c和表5中所示的抗原结合构建体的二价相互作用是抗原结合构建体与一个复合物的靶b和相邻复合物的靶a结合的结果,而不是与同一复合物分子内的靶b和靶a结合的结果。为了确定抗原结合构建体是否在同一复合物分子内结合,将biacorecm5芯片用抗原结合构建体包被。使靶a-靶b复合物、靶a和靶b结合芯片(表6)。对靶b的亲和力类似于在固定靶b并且使抗原结合构建体结合时所显示的亲和力。靶a以相对高的亲和力与固定的抗原结合构建体结合,这是因为靶a是二聚体。这允许靶a与抗原结合构建体包被的芯片发生二价相互作用。复合物对固定的抗原结合构建体的亲和力明显高于靶b对固定的抗原结合构建体的亲和力的事实表明了抗原结合构建体在一个复合物分子内结合。表6:通过biacore对靶a-靶b复合物、靶a和靶b的亲和力测量。kd(m)靶a-靶b复合物1.25e-12靶a8.04e-11靶b1.14e-10实施例3:靶c/靶d抗原结合构建体的产生和纯化vhh2h3和vhh3h2是对靶d具有结合特异性的不同的vhh抗体。4r36b7是对靶c具有结合特异性的常规抗体。使用杵入臼(kih)技术产生用于结合靶c和靶d的vhh2h3或vhh3h2和4r36b7的双特异性抗原结合构建体。将vhh2h3和vhh3h2与migg2a_臼状结构融合。将4r36b7-vh克隆成migg2a_杵状结构,并且将4r36b7-vl克隆成clvλ。在293e细胞中产生靶c/靶d抗原结合构建体。将样品经过蛋白a珠粒纯化。将结合的抗原结合构建体洗脱并且使用sds-page分析。图5b示出了用考马斯蓝染色的sds-page凝胶,说明了以约120kda的正确尺寸纯化抗原结合构建体。请注意,与样品相比,分子量标记物迁移得更快。与完整igg相比,抗原结合构建体具有更低的条带。在正常情况下,错配将引起150kda的完整igg、75kda的vhh-fc融合体和约120kda的双特异性形式的形成。然而,图5b示出了仅产生120kda的产物,这表明了优先产生所期望的双特异性形式。因此,本发明的双特异性形式确保了仅获得正确的双特异性抗体。特别是,4r36b7migg2afc失活_杵状结构构建体仅与vhh2h3-mfc失活_臼状结构或vhh3h2-mfc失活_臼状结构构建体配对。剩余的轻链clvλ_4r36b7构建体只能与4r36b7migg2afc失活重链配对,这是因为轻链不能与vhh链配对。对于本领域技术人员来说将显而易见的是,本发明不限于上述的kih异二聚化方法。任何适当的异二聚化方法,如本文其它地方所述的方法均可以用于上述双特异性抗体形式。实施例4:抗原结合构建体的尺寸排阻色谱法(sec)所使用的色谱系统是agilent1260infinityii,其配备有四元泵、自动进样器、冷藏自动进样器(6℃)、在线脱气器和dad检测器。柱是watersxbridgebehsec(3.5μm,7.8×300mm;沃特世公司(waters),目录号176003596),其与watersxbridgebehsec预保护柱(3.5μm,7.8×30mm;沃特世公司,目录号176003591)联接。首先用流动相将柱平衡10倍柱体积(cv),并且不将它保持在恒温隔室中。在使用pbs作为流动相(西格玛公司(sigma),目录号d8537)的同时,将操作流速设置为1毫升/分钟,持续30分钟。将检测器同时设置为波长280nm和214nm(参考波长为360nm,截止波长为100nm)。在已知具有3.5%聚集体的高聚集对照样品(hac)的存在下,在通道214nm上跟踪聚集监测。一起获取在该过程中多次监测的样品作为qc样品。使用chemstationopenlabcdc软件(安捷伦科技公司(agilent))进行数据采集。图6a-图6c和下表7示出了靶c/靶d双特异性抗体的sec分析的结果。表7:靶c/靶d抗体的sec结果。实施例5:靶c/靶d抗原结合构建体的表征使用biacore3000测量靶c/靶d结合构建体的亲和力。使用如所推荐的nhs化学方法,将cm5芯片用约500ru的靶c或靶d包被。将一定浓度范围的抗体添加到芯片中(9种浓度,从10μg/ml开始)。所使用的分子量如下:migg是约150kda;migg-vhh-fc是约120kda;vhh-fc二聚体是约80kda。通过使用biaevaluation软件将动力学参数与1:1朗缪尔结合模型拟合来确定亲和力。结果示于下表8中。发现两个抗体臂均具有功能性。表8:通过biacore对靶c和靶d的亲和力测量。“nb”意指无结合。序列表<110>阿根思公司<120>双特异性抗原结合构建体<130>607772:agx5-031<150>us62/609,523<151>2017-12-22<160>36<170>patentin3.5版<210>1<211>10<212>prt<213>智人<400>1gluprolyssercysasplysthrhisthr1510<210>2<211>5<212>prt<213>智人<400>2cysproprocyspro15<210>3<211>8<212>prt<213>智人<400>3alaprogluleuleuglyglypro15<210>4<211>10<212>prt<213>智人<400>4cyscysvalglucysproproprocyspro1510<210>5<211>7<212>prt<213>智人<400>5alaproprovalalaglypro15<210>6<211>12<212>prt<213>智人<400>6gluleulysthrproleuglyaspthrthrhisthr1510<210>7<211>50<212>prt<213>智人<400>7cysproargcysprogluprolyssercysaspthrproproprocys151015proargcysprogluprolyssercysaspthrproproprocyspro202530argcysprogluprolyssercysaspthrproproprocysproarg354045cyspro50<210>8<211>7<212>prt<213>智人<400>8gluserlystyrglypropro15<210>9<211>5<212>prt<213>智人<400>9cysprosercyspro15<210>10<211>8<212>prt<213>智人<400>10alaproglupheleuglyglypro15<210>11<211>373<212>dna<213>人工序列<220><223>合成多核苷酸<400>11gagttgcaggtggtggagtctgggggaggattggtgcaggctggggcctctctgagactc60tcctgtgtagcctctggacgcaccttcagtagttattccatggcctggttccgccaggct120ccagggaaggagcgtgagtttgtcgcgacggttagtaattggaatgattacatcacaacc180tatgcagactccgtgaagggccgattcaccatctccagagacaatgccaaaaacacggtg240tctctgcaaatgaacggcctgaaacctgaggacacggccgtttattactgtgcagcgcgg300accggggctcctagggtcacttccggacagtatgactactggggccaggggacccaggtc360accgtgtcctcag373<210>12<211>124<212>prt<213>人工序列<220><223>合成多肽<400>12gluleuglnvalvalgluserglyglyglyleuvalglnalaglyala151015serleuargleusercysvalalaserglyargthrphesersertyr202530sermetalatrppheargglnalaproglylysgluargglupheval354045alathrvalserasntrpasnasptyrilethrthrtyralaaspser505560vallysglyargphethrileserargaspasnalalysasnthrval65707580serleuglnmetasnglyleulysprogluaspthralavaltyrtyr859095cysalaalaargthrglyalaproargvalthrserglyglntyrasp100105110tyrtrpglyglnglythrglnvalthrvalserser115120<210>13<211>361<212>dna<213>人工序列<220><223>合成多核苷酸<400>13gagttgcagctggtggagtctgggggaggcttggtgcagcctggggggtctcggagactc60tcctgtgctgcctctgggttcacctacagtgactactggatgtattgggtccgtcagcct120ccagggaagggactcgagtgggtctcaaatattaattctgaaggtggtagcacagcctat180gcagactccgtgaagggccgattcaccatctccagagacaacgccaagaacacgctgtat240ctgcaaatgaacagtctgaaatctgaggacacggccgtatactactgtgtaagagcatta300agcagtggtcaatggtacccggcctactggggccaggggacccaggtcaccgtgtcctcc360g361<210>14<211>120<212>prt<213>人工序列<220><223>合成多肽<400>14gluleuglnleuvalgluserglyglyglyleuvalglnproglygly151015serargargleusercysalaalaserglyphethrtyrserasptyr202530trpmettyrtrpvalargglnproproglylysglyleuglutrpval354045serasnileasnsergluglyglyserthralatyralaaspserval505560lysglyargphethrileserargaspasnalalysasnthrleutyr65707580leuglnmetasnserleulyssergluaspthralavaltyrtyrcys859095valargalaleuserserglyglntrptyrproalatyrtrpglygln100105110glythrglnvalthrvalserser115120<210>15<211>796<212>dna<213>人工序列<220><223>合成多核苷酸<400>15ctgtcgtctcactctgagttgcagctggtggagtctgggggaggcttggtgcagcctggg60gggtctcggagactctcctgtgctgcctctgggttcacctacagtgactactggatgtat120tgggtccgtcagcctccagggaagggactcgagtgggtctcaaatattaattctgaaggt180ggtagcacagcctatgcagactccgtgaagggccgattcaccatctccagagacaacgcc240aagaacacgctgtatctgcaaatgaacagtctgaaatctgaggacacggccgtatactac300tgtgtaagagcattaagcagtggtcaatggtacccggcctactggggccaggggacccag360gtcaccgtgtcctccgcctccgcggccgcaagcggtggaggcggttcaggcggaggtgga420tctggcggtggcggaagtgcacaggcagggctgactcagctgccctccgtgtctggatcc480ccaggccagaagatcaccatctcctgcactggaagcagcagcaacatcagggttggttat540aatgttcagcggttccagcacctcccaggaacacccccccaactgctcatctatggtaac600agcaatcaagcttcgggggtcccagaccgcttctctggctccaagtctggcagctcggcc660tccctgaccatcactgggctccaggctgaggacgaggctgactattactgtgaatgctat720gacagcggcctcagtgggcctgtgttcggcggagggaccaagctgaccgtcctcggtgag780cctgagacgacacgcc796<210>16<211>260<212>prt<213>人工序列<220><223>合成多肽<400>16gluleuglnleuvalgluserglyglyglyleuvalglnproglygly151015serargargleusercysalaalaserglyphethrtyrserasptyr202530trpmettyrtrpvalargglnproproglylysglyleuglutrpval354045serasnileasnsergluglyglyserthralatyralaaspserval505560lysglyargphethrileserargaspasnalalysasnthrleutyr65707580leuglnmetasnserleulyssergluaspthralavaltyrtyrcys859095valargalaleuserserglyglntrptyrproalatyrtrpglygln100105110glythrglnvalthrvalserseralaseralaalaalaserglygly115120125glyglyserglyglyglyglyserglyglyglyglyseralaglnala130135140glyleuthrglnleuproservalserglyserproglyglnlysile145150155160thrilesercysthrglyserserserasnileargvalglytyrasn165170175valglnargpheglnhisleuproglythrproproglnleuleuile180185190tyrglyasnserasnglnalaserglyvalproaspargphesergly195200205serlysserglyserseralaserleuthrilethrglyleuglnala210215220gluaspglualaasptyrtyrcysglucystyraspserglyleuser225230235240glyprovalpheglyglyglythrlysleuthrvalleuglyglupro245250255gluthrthrarg260<210>17<211>1104<212>dna<213>人工序列<220><223>合成多核苷酸<400>17cgtttaaacggtaccgccgccaccatgggctggtcctgcatcatcctgtttctggtggcc60accgccacaggcgtccactctggagacgccaatccttcactcgaattccgtctcgctaaa120acaacagccccatcggtctatccactggcccctgtgtgtggagatacaactggctcctcg180gtgactctaggatgcctggtcaagggttatttccctgagccagtgaccttgacctggaac240tctggatccctgtccagtggtgtgcacaccttcccagctgtcctgcagtctgacctctac300accctcagcagctcagtgactgtaacctcgagcacctggcccagccagtccatcacctgc360aatgtggcccacccggcaagcagcaccaaggtggacaagaaaattgagcccagagggccc420acaatcaagccctgtcctccatgcaaatgcccagcacctaacctcttgggtggaccagac480gtcttcatcttccctccaaagatcaaggatgtactcatgatctccctgagccccatagtc540acatgtgtggtggtggatgtgagcgaggatgacccagatgtccagatcagctggtttgtg600aacaacgtggaagtacacacagctcagacacaaacccatagagaggattaccagagtact660ctccgggtggtcagtgccctccccatccagcaccaggactggatgagtggcaaggagttc720aaatgcaaggtcaacaacaaagacctcccagcgcccgaagagagaaccatctcaaaaccc780aaagggtcagtaagagctccacaggtatatgtcttgcctccaccagaagaagagatgact840aagaaacaggtcactctgtcctgcgctgtcacagacttcatgcctgaagacatttacgtg900gagtggaccaacaacgggaaaacagagctaaactacaagaacactgaaccagtcctggac960tctgatggttcttacttcatggtgagcaagctgagagtggaaaagaagaactgggtggaa1020agaaatagctactcctgttcagtggtccacgagggtctgcacaatcaccacacgactaag1080agcttctcccggactccgggtaaa1104<210>18<211>368<212>prt<213>人工序列<220><223>合成多肽<400>18argleuasnglythralaalathrmetglytrpsercysileileleu151015pheleuvalalathralathrglyvalhisserglyaspalaasnpro202530serleuglupheargleualalysthrthralaproservaltyrpro354045leualaprovalcysglyaspthrthrglyserservalthrleugly505560cysleuvallysglytyrpheprogluprovalthrleuthrtrpasn65707580serglyserleuserserglyvalhisthrpheproalavalleugln859095seraspleutyrthrleuserserservalthrvalthrserserthr100105110trpproserglnserilethrcysasnvalalahisproalaserser115120125thrlysvalasplyslysilegluproargglyprothrilelyspro130135140cysproprocyslyscysproalaproasnleuleuglyglyproasp145150155160valpheilepheproprolysilelysaspvalleumetileserleu165170175serproilevalthrcysvalvalvalaspvalsergluaspasppro180185190aspvalglnilesertrpphevalasnasnvalgluvalhisthrala195200205glnthrglnthrhisarggluasptyrglnserthrleuargvalval210215220seralaleuproileglnhisglnasptrpmetserglylysgluphe225230235240lyscyslysvalasnasnlysaspleuproalaproglugluargthr245250255ileserlysprolysglyservalargalaproglnvaltyrvalleu260265270proproproglugluglumetthrlyslysglnvalthrleusercys275280285alavalthraspphemetprogluaspiletyrvalglutrpthrasn290295300asnglylysthrgluleuasntyrlysasnthrgluprovalleuasp305310315320seraspglysertyrphemetvalserlysleuargvalglulyslys325330335asntrpvalgluargasnsertyrsercysservalvalhisglugly340345350leuhisasnhishisthrthrlysserpheserargthrproglylys355360365<210>19<211>1104<212>dna<213>人工序列<220><223>合成多核苷酸<400>19cgtttaaacggtaccgccgccaccatgggctggtcctgcatcatcctgtttctggtggcc60accgccacaggcgtccactctggagacgccaatccttcactcgaattccgtctcgctaaa120acaacagccccatcggtctatccactggcccctgtgtgtggagatacaactggctcctcg180gtgactctaggatgcctggtcaagggttatttccctgagccagtgaccttgacctggaac240tctggatccctgtccagtggtgtgcacaccttcccagctgtcctgcagtctgacctctac300accctcagcagctcagtgactgtaacctcgagcacctggcccagccagtccatcacctgc360aatgtggcccacccggcaagcagcaccaaggtggacaagaaaattgagcccagagggccc420acaatcaagccctgtcctccatgcaaatgcccagcacctaacctcttgggtggaccagac480gtcttcatcttccctccaaagatcaaggatgtactcatgatctccctgagccccatagtc540acatgtgtggtggtggatgtgagcgaggatgacccagatgtccagatcagctggtttgtg600aacaacgtggaagtacacacagctcagacacaaacccatagagaggattacaacagtact660ctccgggtggtcagtgccctccccatccagcaccaggactggatgagtggcaaggagttc720aaatgcaaggtcaacaacaaagacctcccagcgcccgaagagagaaccatctcaaaaccc780aaagggtcagtaagagctccacaggtatatgtcttgcctccaccagaagaagagatgact840aagaaacaggtcactctgtggtgcatggtcacagacttcatgcctgaagacatttacgtg900gagtggaccaacaacgggaaaacagagctaaactacaagaacactgaaccagtcctggac960tctgatggttcttacttcatgtacagcaagctgagagtggaaaagaagaactgggtggaa1020agaaatagctactcctgttcagtggtccacgagggtctgcacaatcaccacacgactaag1080agcttctcccggactccgggtaaa1104<210>20<211>368<212>prt<213>人工序列<220><223>合成多肽<400>20argleuasnglythralaalathrmetglytrpsercysileileleu151015pheleuvalalathralathrglyvalhisserglyaspalaasnpro202530serleuglupheargleualalysthrthralaproservaltyrpro354045leualaprovalcysglyaspthrthrglyserservalthrleugly505560cysleuvallysglytyrpheprogluprovalthrleuthrtrpasn65707580serglyserleuserserglyvalhisthrpheproalavalleugln859095seraspleutyrthrleuserserservalthrvalthrserserthr100105110trpproserglnserilethrcysasnvalalahisproalaserser115120125thrlysvalasplyslysilegluproargglyprothrilelyspro130135140cysproprocyslyscysproalaproasnleuleuglyglyproasp145150155160valpheilepheproprolysilelysaspvalleumetileserleu165170175serproilevalthrcysvalvalvalaspvalsergluaspasppro180185190aspvalglnilesertrpphevalasnasnvalgluvalhisthrala195200205glnthrglnthrhisarggluasptyrasnserthrleuargvalval210215220seralaleuproileglnhisglnasptrpmetserglylysgluphe225230235240lyscyslysvalasnasnlysaspleuproalaproglugluargthr245250255ileserlysprolysglyservalargalaproglnvaltyrvalleu260265270proproproglugluglumetthrlyslysglnvalthrleutrpcys275280285metvalthraspphemetprogluaspiletyrvalglutrpthrasn290295300asnglylysthrgluleuasntyrlysasnthrgluprovalleuasp305310315320seraspglysertyrphemettyrserlysleuargvalglulyslys325330335asntrpvalgluargasnsertyrsercysservalvalhisglugly340345350leuhisasnhishisthrthrlysserpheserargthrproglylys355360365<210>21<211>813<212>dna<213>人工序列<220><223>合成多核苷酸<400>21cgtttaaacggtaccgccgccaccatgggctggtcctgcatcatcctgtttctggtggcc60accgccacaggcgtccactctggagacgccaatccttcactcgaattccgtctcgagccc120agagggcccacaatcaagccctgtcctccatgcaaatgcccagcacctaacctcttgggt180ggaccagacgtcttcatcttccctccaaagatcaaggatgtactcatgatctccctgagc240cccatagtcacatgtgtggtggtggatgtgagcgaggatgacccagatgtccagatcagc300tggtttgtgaacaacgtggaagtacacacagctcagacacaaacccatagagaggattac360aacagtactctccgggtggtcagtgccctccccatccagcaccaggactggatgagtggc420aaggagttcaaatgcaaggtcaacaacaaagacctcccagcgcccgaagagagaaccatc480tcaaaacccaaagggtcagtaagagctccacaggtatatgtcttgcctccaccagaagaa540gagatgactaagaaacaggtcactctgtcctgcgctgtcacagacttcatgcctgaagac600atttacgtggagtggaccaacaacgggaaaacagagctaaactacaagaacactgaacca660gtcctggactctgatggttcttacttcatggtgagcaagctgagagtggaaaagaagaac720tgggtggaaagaaatagctactcctgttcagtggtccacgagggtctgcacaatcaccac780acgactaagagcttctcccggactccgggtaaa813<210>22<211>271<212>prt<213>人工序列<220><223>合成多肽<400>22argleuasnglythralaalathrmetglytrpsercysileileleu151015pheleuvalalathralathrglyvalhisserglyaspalaasnpro202530serleuglupheargleugluproargglyprothrilelysprocys354045proprocyslyscysproalaproasnleuleuglyglyproaspval505560pheilepheproprolysilelysaspvalleumetileserleuser65707580proilevalthrcysvalvalvalaspvalsergluaspaspproasp859095valglnilesertrpphevalasnasnvalgluvalhisthralagln100105110thrglnthrhisarggluasptyrasnserthrleuargvalvalser115120125alaleuproileglnhisglnasptrpmetserglylysgluphelys130135140cyslysvalasnasnlysaspleuproalaproglugluargthrile145150155160serlysprolysglyservalargalaproglnvaltyrvalleupro165170175proproglugluglumetthrlyslysglnvalthrleusercysala180185190valthraspphemetprogluaspiletyrvalglutrpthrasnasn195200205glylysthrgluleuasntyrlysasnthrgluprovalleuaspser210215220aspglysertyrphemetvalserlysleuargvalglulyslysasn225230235240trpvalgluargasnsertyrsercysservalvalhisgluglyleu245250255hisasnhishisthrthrlysserpheserargthrproglylys260265270<210>23<211>813<212>dna<213>人工序列<220><223>合成多核苷酸<400>23cgtttaaacggtaccgccgccaccatgggctggtcctgcatcatcctgtttctggtggcc60accgccacaggcgtccactctggagacgccaatccttcactcgaattccgtctcgagccc120agagggcccacaatcaagccctgtcctccatgcaaatgcccagcacctaacctcttgggt180ggaccagacgtcttcatcttccctccaaagatcaaggatgtactcatgatctccctgagc240cccatagtcacatgtgtggtggtggatgtgagcgaggatgacccagatgtccagatcagc300tggtttgtgaacaacgtggaagtacacacagctcagacacaaacccatagagaggattac360aacagtactctccgggtggtcagtgccctccccatccagcaccaggactggatgagtggc420aaggagttcaaatgcaaggtcaacaacaaagacctcccagcgcccgaagagagaaccatc480tcaaaacccaaagggtcagtaagagctccacaggtatatgtcttgcctccaccagaagaa540gagatgactaagaaacaggtcactctgtggtgcatggtcacagacttcatgcctgaagac600atttacgtggagtggaccaacaacgggaaaacagagctaaactacaagaacactgaacca660gtcctggactctgatggttcttacttcatgtacagcaagctgagagtggaaaagaagaac720tgggtggaaagaaatagctactcctgttcagtggtccacgagggtctgcacaatcaccac780acgactaagagcttctcccggactccgggtaaa813<210>24<211>271<212>prt<213>人工序列<220><223>合成多肽<400>24argleuasnglythralaalathrmetglytrpsercysileileleu151015pheleuvalalathralathrglyvalhisserglyaspalaasnpro202530serleuglupheargleugluproargglyprothrilelysprocys354045proprocyslyscysproalaproasnleuleuglyglyproaspval505560pheilepheproprolysilelysaspvalleumetileserleuser65707580proilevalthrcysvalvalvalaspvalsergluaspaspproasp859095valglnilesertrpphevalasnasnvalgluvalhisthralagln100105110thrglnthrhisarggluasptyrasnserthrleuargvalvalser115120125alaleuproileglnhisglnasptrpmetserglylysgluphelys130135140cyslysvalasnasnlysaspleuproalaproglugluargthrile145150155160serlysprolysglyservalargalaproglnvaltyrvalleupro165170175proproglugluglumetthrlyslysglnvalthrleutrpcysmet180185190valthraspphemetprogluaspiletyrvalglutrpthrasnasn195200205glylysthrgluleuasntyrlysasnthrgluprovalleuaspser210215220aspglysertyrphemettyrserlysleuargvalglulyslysasn225230235240trpvalgluargasnsertyrsercysservalvalhisgluglyleu245250255hisasnhishisthrthrlysserpheserargthrproglylys260265270<210>25<211>991<212>dna<213>人工序列<220><223>合成多核苷酸<400>25agtttaaacggtaccgccgccaccatgggctggtcctgcatcatcctgtttctggtggcc60accgccacaggcgtccactctggagacgcctccttaacactcgaattccgtctcgctaaa120acaacagccccatcggtctatccactggcccctgtgtgtggagatacaactggctcctcg180gtgactctaggatgcctggtcaagggttatttccctgagccagtgaccttgacctggaac240tctggatccctgtccagtggtgtgcacaccttcccagctgtcctgcagtctgacctctac300accctcagcagctcagtgactgtaacctcgagcacctggcccagccagtccatcacctgc360aatgtggcccacccggcaagcagcaccaaggtggacaagaaaattgagcccagagggccc420acaatcaagccctgtcctccatgcaaatgcccagcacctaacctcttgggtggaccatcc480gtcttcatcttccctccaaagatcaaggatgtactcatgatctccctgagccccatagtc540acatgtgtggtggtggatgtgagcgaggatgacccagatgtccagatcagctggtttgtg600aacaacgtggaagtacacacagctcagacacaaacccatagagaggattacaacagtact660ctccgggtggtcagtgccctccccatccagcaccaggactggatgagtggcaaggagttc720aaatgcaaggtcaacaacaaagacctcccagcgcccatcgagagaaccatctcaaaaccc780aaagggtcagtaagagctccacaggtatatgtcttgcctccaccagaagaagagatgact840aagaaacaggtcactctgacctgcatggtcacagacttcatgcctgaagacatttacgtg900gagtggaccaacaacgggaaaacagagctaaactacaagaacactgaaccagtcctggac960tctgatggttcttacttcatgtacagcaagc991<210>26<211>330<212>prt<213>人工序列<220><223>合成多肽<400>26serleuasnglythralaalathrmetglytrpsercysileileleu151015pheleuvalalathralathrglyvalhisserglyaspalaserleu202530thrleuglupheargleualalysthrthralaproservaltyrpro354045leualaprovalcysglyaspthrthrglyserservalthrleugly505560cysleuvallysglytyrpheprogluprovalthrleuthrtrpasn65707580serglyserleuserserglyvalhisthrpheproalavalleugln859095seraspleutyrthrleuserserservalthrvalthrserserthr100105110trpproserglnserilethrcysasnvalalahisproalaserser115120125thrlysvalasplyslysilegluproargglyprothrilelyspro130135140cysproprocyslyscysproalaproasnleuleuglyglyproser145150155160valpheilepheproprolysilelysaspvalleumetileserleu165170175serproilevalthrcysvalvalvalaspvalsergluaspasppro180185190aspvalglnilesertrpphevalasnasnvalgluvalhisthrala195200205glnthrglnthrhisarggluasptyrasnserthrleuargvalval210215220seralaleuproileglnhisglnasptrpmetserglylysgluphe225230235240lyscyslysvalasnasnlysaspleuproalaproilegluargthr245250255ileserlysprolysglyservalargalaproglnvaltyrvalleu260265270proproproglugluglumetthrlyslysglnvalthrleuthrcys275280285metvalthraspphemetprogluaspiletyrvalglutrpthrasn290295300asnglylysthrgluleuasntyrlysasnthrgluprovalleuasp305310315320seraspglysertyrphemettyrserlys325330<210>27<211>450<212>prt<213>人工序列<220><223>合成多肽<400>27gluleuglnleuvalgluserglyglyglyleuvalglnproglygly151015serargargleusercysalaalaserglyphethrtyrserasptyr202530trpmettyrtrpvalargglnproproglylysglyleuglutrpval354045serasnileasnsergluglyglyserthralatyralaaspserval505560lysglyargphethrileserargaspasnalalysasnthrleutyr65707580leuglnmetasnserleulyssergluaspthralavaltyrtyrcys859095valargalaleuserserglyglntrptyrproalatyrtrpglygln100105110glythrglnvalthrvalserseralalysthrthralaproserval115120125tyrproleualaprovalcysglyaspthrthrglyserservalthr130135140leuglycysleuvallysglytyrpheprogluprovalthrleuthr145150155160trpasnserglyserleuserserglyvalhisthrpheproalaval165170175leuglnseraspleutyrthrleuserserservalthrvalthrser180185190serthrtrpproserglnserilethrcysasnvalalahisproala195200205serserthrlysvalasplyslysilegluproargglyprothrile210215220lysprocysproprocyslyscysproalaproasnleuleuglygly225230235240proaspvalpheilepheproprolysilelysaspvalleumetile245250255serleuserproilevalthrcysvalvalvalaspvalsergluasp260265270aspproaspvalglnilesertrpphevalasnasnvalgluvalhis275280285thralaglnthrglnthrhisarggluasptyrglnserthrleuarg290295300valvalseralaleuproileglnhisglnasptrpmetserglylys305310315320gluphelyscyslysvalasnasnlysaspleuproalaprogluglu325330335argthrileserlysprolysglyservalargalaproglnvaltyr340345350valleuproproproglugluglumetthrlyslysglnvalthrleu355360365sercysalavalthraspphemetprogluaspiletyrvalglutrp370375380thrasnasnglylysthrgluleuasntyrlysasnthrgluproval385390395400leuaspseraspglysertyrphemetvalserlysleuargvalglu405410415lyslysasntrpvalgluargasnsertyrsercysservalvalhis420425430gluglyleuhisasnhishisthrthrlysserpheserargthrpro435440445glylys450<210>28<211>357<212>prt<213>人工序列<220><223>合成多肽<400>28gluleuglnvalvalgluserglyglyglyleuvalglnalaglyala151015serleuargleusercysvalalaserglyargthrphesersertyr202530sermetalatrppheargglnalaproglylysgluargglupheval354045alathrvalserasntrpasnasptyrilethrthrtyralaaspser505560vallysglyargphethrileserargaspasnalalysasnthrval65707580serleuglnmetasnglyleulysprogluaspthralavaltyrtyr859095cysalaalaargthrglyalaproargvalthrserglyglntyrasp100105110tyrtrpglyglnglythrglnvalthrvalsersergluproarggly115120125prothrilelysprocysproprocyslyscysproalaproasnleu130135140leuglyglyproaspvalpheilepheproprolysilelysaspval145150155160leumetileserleuserproilevalthrcysvalvalvalaspval165170175sergluaspaspproaspvalglnilesertrpphevalasnasnval180185190gluvalhisthralaglnthrglnthrhisarggluasptyrasnser195200205thrleuargvalvalseralaleuproileglnhisglnasptrpmet210215220serglylysgluphelyscyslysvalasnasnlysaspleuproala225230235240proglugluargthrileserlysprolysglyservalargalapro245250255glnvaltyrvalleuproproproglugluglumetthrlyslysgln260265270valthrleutrpcysmetvalthraspphemetprogluaspiletyr275280285valglutrpthrasnasnglylysthrgluleuasntyrlysasnthr290295300gluprovalleuaspseraspglysertyrphemettyrserlysleu305310315320argvalglulyslysasntrpvalgluargasnsertyrsercysser325330335valvalhisgluglyleuhisasnhishisthrthrlysserpheser340345350argthrproglylys355<210>29<211>242<212>prt<213>人工序列<220><223>合成多肽<400>29glyglyglyglyserglyglyglyglysergluproargglyprothr151015ilelysprocysproprocyslyscysproalaproasnleuleugly202530glyproservalpheilepheproprolysilelysaspvalleumet354045ileserleuserproilevalthrcysvalvalvalaspvalserglu505560aspaspproaspvalglnilesertrpphevalasnasnvalgluval65707580histhralaglnthrglnthrhisarggluasptyralaserthrleu859095argvalvalseralaleuproileglnhisglnasptrpmetsergly100105110lysgluphelyscyslysvalasnasnlysaspleuproalaproile115120125gluargthrileserlysprolysglyservalargalaproglnval130135140tyrvalleuproproproglugluglumetthrlyslysglnvalthr145150155160leutrpcysmetvalthraspphemetprogluaspiletyrvalglu165170175trpthrasnasnglylysthrgluleuasntyrlysasnthrglupro180185190valleuaspseraspglysertyrphemettyrserlysleuargval195200205glulyslysasntrpvalgluargasnsertyrsercysservalval210215220hisgluglyleuhisasnhishisthrthrlysserpheserargthr225230235240progly<210>30<211>242<212>prt<213>人工序列<220><223>合成多肽<400>30glyglyglyglyserglyglyglyglysergluproargglyprothr151015ilelysprocysproprocyslyscysproalaproasnleuleugly202530glyproservalpheilepheproprolysilelysaspvalleumet354045ileserleuserproilevalthrcysvalvalvalaspvalserglu505560aspaspproaspvalglnilesertrpphevalasnasnvalgluval65707580histhralaglnthrglnthrhisarggluasptyralaserthrleu859095argvalvalseralaleuproileglnhisglnasptrpmetsergly100105110lysgluphelyscyslysvalasnasnlysaspleuproalaproile115120125gluargthrileserlysprolysglyservalargalaproglnval130135140tyrvalleuproproproglugluglumetthrlyslysglnvalthr145150155160leutrpcysmetvalthraspphemetprogluaspiletyrvalglu165170175trpthrasnasnglylysthrgluleuasntyrlysasnthrglupro180185190valleuaspseraspglysertyrphemettyrserlysleuargval195200205glulyslysasntrpvalgluargasnsertyrsercysservalval210215220hisgluglyleuhisasnhishisthrthrlysserpheserargthr225230235240progly<210>31<211>330<212>prt<213>人工序列<220><223>合成多肽<400>31alalysthrthralaproservaltyrproleualaprovalcysgly151015aspthrthrglyserservalthrleuglycysleuvallysglytyr202530pheprogluprovalthrleuthrtrpasnserglyserleuserser354045glyvalhisthrpheproalavalleuglnseraspleutyrthrleu505560serserservalthrvalthrserserthrtrpproserglnserile65707580thrcysasnvalalahisproalaserserthrlysvalasplyslys859095ilegluproargglyprothrilelysprocysproprocyslyscys100105110proalaproasnleuleuglyglyproservalpheilephepropro115120125lysilelysaspvalleumetileserleuserproilevalthrcys130135140valvalvalaspvalsergluaspaspproaspvalglnilesertrp145150155160phevalasnasnvalgluvalhisthralaglnthrglnthrhisarg165170175gluasptyralaserthrleuargvalvalseralaleuproilegln180185190hisglnasptrpmetserglylysgluphelyscyslysvalasnasn195200205lysaspleuproalaproilegluargthrileserlysprolysgly210215220servalargalaproglnvaltyrvalleuproproproglugluglu225230235240metthrlyslysglnvalthrleusercysalavalthraspphemet245250255progluaspiletyrvalglutrpthrasnasnglylysthrgluleu260265270asntyrlysasnthrgluprovalleuaspseraspglysertyrphe275280285metvalserlysleuargvalglulyslysasntrpvalgluargasn290295300sertyrsercysservalvalhisgluglyleuhisasnhishisthr305310315320thrlysserpheserargthrproglylys325330<210>32<211>373<212>prt<213>人工序列<220><223>合成多肽<400>32argleuasnglythralaalathrmetglytrpsercysileileleu151015pheleuvalalathralathrglyvalhisserglyaspalaasnpro202530serleuglupheargleualalysthrthralaproservaltyrpro354045leualaprovalcysglyaspthrthrglyserservalthrleugly505560cysleuvallysglytyrpheprogluprovalthrleuthrtrpasn65707580serglyserleuserserglyvalhisthrpheproalavalleugln859095seraspleutyrthrleuserserservalthrvalthrserserthr100105110trpproserglnserilethrcysasnvalalahisproalaserser115120125thrlysvalasplyslysilegluproargglyprothrilelyspro130135140cysproprocyslyscysproalaproasnleuleuglyglyproasp145150155160valpheilepheproprolysilelysaspvalleumetileserleu165170175serproilevalthrcysvalvalvalaspvalsergluaspasppro180185190aspvalglnilesertrpphevalasnasnvalgluvalhisthrala195200205glnthrglnthrhisarggluasptyrglnserthrleuargvalval210215220seralaleuproileglnhisglnasptrpmetserglylysgluphe225230235240lyscyslysvalasnasnlysaspleuproalaproglugluargthr245250255ileserlysprolysglyservalargalaproglnvaltyrvalleu260265270proproproglugluglumetthrlyslysglnvalthrleusercys275280285alavalthraspphemetprogluaspiletyrvalglutrpthrasn290295300asnglylysthrgluleuasntyrlysasnthrgluprovalleuasp305310315320seraspglysertyrphemetvalserlysleuargvalglulyslys325330335asntrpvalgluargasnsertyrsercysservalvalhisglugly340345350leuhisasnhishisthrthrlysserpheserargthrproglylys355360365serargalaalaala370<210>33<211>373<212>prt<213>人工序列<220><223>合成多肽<400>33argleuasnglythralaalathrmetglytrpsercysileileleu151015pheleuvalalathralathrglyvalhisserglyaspalaasnpro202530serleuglupheargleualalysthrthralaproservaltyrpro354045leualaprovalcysglyaspthrthrglyserservalthrleugly505560cysleuvallysglytyrpheprogluprovalthrleuthrtrpasn65707580serglyserleuserserglyvalhisthrpheproalavalleugln859095seraspleutyrthrleuserserservalthrvalthrserserthr100105110trpproserglnserilethrcysasnvalalahisproalaserser115120125thrlysvalasplyslysilegluproargglyprothrilelyspro130135140cysproprocyslyscysproalaproasnleuleuglyglyproasp145150155160valpheilepheproprolysilelysaspvalleumetileserleu165170175serproilevalthrcysvalvalvalaspvalsergluaspasppro180185190aspvalglnilesertrpphevalasnasnvalgluvalhisthrala195200205glnthrglnthrhisarggluasptyrasnserthrleuargvalval210215220seralaleuproileglnhisglnasptrpmetserglylysgluphe225230235240lyscyslysvalasnasnlysaspleuproalaproglugluargthr245250255ileserlysprolysglyservalargalaproglnvaltyrvalleu260265270proproproglugluglumetthrlyslysglnvalthrleutrpcys275280285metvalthraspphemetprogluaspiletyrvalglutrpthrasn290295300asnglylysthrgluleuasntyrlysasnthrgluprovalleuasp305310315320seraspglysertyrphemettyrserlysleuargvalglulyslys325330335asntrpvalgluargasnsertyrsercysservalvalhisglugly340345350leuhisasnhishisthrthrlysserpheserargthrproglylys355360365serargalaalaala370<210>34<211>276<212>prt<213>人工序列<220><223>合成多肽<400>34argleuasnglythralaalathrmetglytrpsercysileileleu151015pheleuvalalathralathrglyvalhisserglyaspalaasnpro202530serleuglupheargleugluproargglyprothrilelysprocys354045proprocyslyscysproalaproasnleuleuglyglyproaspval505560pheilepheproprolysilelysaspvalleumetileserleuser65707580proilevalthrcysvalvalvalaspvalsergluaspaspproasp859095valglnilesertrpphevalasnasnvalgluvalhisthralagln100105110thrglnthrhisarggluasptyrasnserthrleuargvalvalser115120125alaleuproileglnhisglnasptrpmetserglylysgluphelys130135140cyslysvalasnasnlysaspleuproalaproglugluargthrile145150155160serlysprolysglyservalargalaproglnvaltyrvalleupro165170175proproglugluglumetthrlyslysglnvalthrleusercysala180185190valthraspphemetprogluaspiletyrvalglutrpthrasnasn195200205glylysthrgluleuasntyrlysasnthrgluprovalleuaspser210215220aspglysertyrphemetvalserlysleuargvalglulyslysasn225230235240trpvalgluargasnsertyrsercysservalvalhisgluglyleu245250255hisasnhishisthrthrlysserpheserargthrproglylysser260265270argalaalaala275<210>35<211>276<212>prt<213>人工序列<220><223>合成多肽<400>35argleuasnglythralaalathrmetglytrpsercysileileleu151015pheleuvalalathralathrglyvalhisserglyaspalaasnpro202530serleuglupheargleugluproargglyprothrilelysprocys354045proprocyslyscysproalaproasnleuleuglyglyproaspval505560pheilepheproprolysilelysaspvalleumetileserleuser65707580proilevalthrcysvalvalvalaspvalsergluaspaspproasp859095valglnilesertrpphevalasnasnvalgluvalhisthralagln100105110thrglnthrhisarggluasptyrasnserthrleuargvalvalser115120125alaleuproileglnhisglnasptrpmetserglylysgluphelys130135140cyslysvalasnasnlysaspleuproalaproglugluargthrile145150155160serlysprolysglyservalargalaproglnvaltyrvalleupro165170175proproglugluglumetthrlyslysglnvalthrleutrpcysmet180185190valthraspphemetprogluaspiletyrvalglutrpthrasnasn195200205glylysthrgluleuasntyrlysasnthrgluprovalleuaspser210215220aspglysertyrphemettyrserlysleuargvalglulyslysasn225230235240trpvalgluargasnsertyrsercysservalvalhisgluglyleu245250255hisasnhishisthrthrlysserpheserargthrproglylysser260265270argalaalaala275<210>36<211>330<212>prt<213>人工序列<220><223>合成多肽<400>36serleuasnglythralaalathrmetglytrpsercysileileleu151015pheleuvalalathralathrglyvalhisserglyaspalaserleu202530thrleuglupheargleualalysthrthralaproservaltyrpro354045leualaprovalcysglyaspthrthrglyserservalthrleugly505560cysleuvallysglytyrpheprogluprovalthrleuthrtrpasn65707580serglyserleuserserglyvalhisthrpheproalavalleugln859095seraspleutyrthrleuserserservalthrvalthrserserthr100105110trpproserglnserilethrcysasnvalalahisproalaserser115120125thrlysvalasplyslysilegluproargglyprothrilelyspro130135140cysproprocyslyscysproalaproasnleuleuglyglyproser145150155160valpheilepheproprolysilelysaspvalleumetileserleu165170175serproilevalthrcysvalvalvalaspvalsergluaspasppro180185190aspvalglnilesertrpphevalasnasnvalgluvalhisthrala195200205glnthrglnthrhisarggluasptyrasnserthrleuargvalval210215220seralaleuproileglnhisglnasptrpmetserglylysgluphe225230235240lyscyslysvalasnasnlysaspleuproalaproilegluargthr245250255ileserlysprolysglyservalargalaproglnvaltyrvalleu260265270proproproglugluglumetthrlyslysglnvalthrleuthrcys275280285metvalthraspphemetprogluaspiletyrvalglutrpthrasn290295300asnglylysthrgluleuasntyrlysasnthrgluprovalleuasp305310315320seraspglysertyrphemettyrserlys325330当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1