一种利用CsXTH04基因沉默以提高柑橘对溃疡病抗性的方法与流程
本发明涉及分子生物学领域,具体涉及一种利用csxth04基因沉默以提高柑橘对溃疡病抗性的方法。
背景技术:
柑橘是我国南方重要的果树作物,因其具有较高的食用价值和药用价值而受到人们的喜爱。柑橘是全球重要经济作物,主要种植在非洲(16.60%),亚洲(52.90%)、美洲(24.50%)、欧洲和大洋洲(6%)(魏方林等,2008)。然而柑橘产业发展却饱受各种病害的困扰,其中柑橘溃疡病(citrusbacterialcanker,cbc)是阻碍柑橘产业发展最为严重的病害之一(胡军华等,2015)。我国广东、广西、福建、湖南等柑橘主产区受柑橘溃疡病侵害较为严重。全世界有30多个国家和地区将柑橘溃疡病列为重要的植物检疫对象。柑橘溃疡病是由黄单胞杆菌柑橘致病变种(xanthomonascitrisubsp.citri,xcc)引起,起源于印度、爪哇等地(pitinoetal,2015)。溃疡病菌主要侵染柑橘叶片、枝梢、果实等,其中苗木、幼树受害较为严重。病树会出现落叶、枯梢、树势衰弱以及落果等现象,严重的影响了柑橘的产量和品质。病树的叶片发病初期表层出现浓黄色小圆斑,后期病变成淡褐色或褐色,最终形成火山口状灰白色凸起(何秀玲等,2007)。
目前为控制柑橘溃疡病危害通常采取化学防治为主,生物防治为辅的综合防治策略。由于以上防治措施对环境不友好,且需要投入大量的人力、物力,亟待通过培育抗病新品种来减少溃疡病带来的损失(陈力等,2008;朱雪梅等,2017)。在抗病育种方面一代代柑橘人进行了长期杂交育种,但由于杂交育种周期长使得育种效率低。随着分子生物学的兴起,人们开始从抗病基因工程方向对病原菌本身和植物的抗病防卫反应进行研究。基因工程手段也在抗溃疡病研究中进行了尝试,获得了一些对溃疡病有抗性的转基因材料(贾瑞瑞等,2017;段敏杰等,2016)。陈善春等获得了对柑橘溃疡病有抗性的转柞蚕抗菌肽d基因的锦橙、新会橙、脐橙植株(陈善春等,1996)。外源基因nls,chit42,xa21和ptha在转移至冰糖橙,椪柑和甜橙后对柑橘溃疡病具有抗性(mendesetal,2010;yangetal,2011)。贾瑞瑞的研究发现csbzip40是响应柑橘溃疡病侵染的一个重要的转录因子,并且猜测是通过sa途径影响柑橘品种的抗性(贾瑞瑞等,2017)。cslob1基因作为溃疡病病原基因ptha的靶蛋白,被认为是是柑橘溃疡菌的感病因子,使得植物对溃疡病更为敏感(lietal,2014)。fuqua等发现由n-酰基高丝氨酸内酯(ahl)可作为诱导细菌群体感应系统的信号分子,并且参与到各种病原体的发病机制中(fuquaetal,1994;withersetal,2001),之后他们将细菌中的水解酶转基因进入植物,发现植物对溃疡病的抵抗力更强(zanetal,2011)。近年来,出现了使用基因敲除技术来防御溃疡病病原菌的方法,彭爱红等对柑橘溃疡病感病基因cslob1启动子进行crispr/cas9靶向敲除从而获得了对柑橘溃疡病抗性提高的植株(pengetal,2017)。
细胞壁是围绕细胞膜外的结构,它是纤维素,果胶多糖,酶和多种结构蛋白的动态构成的(yokoyamaetal,2001)。在生物和非生物胁迫(例如细菌病原体和盐度)下,细胞壁经历许多生理学,分子机械变化以及细胞形态变化,代谢变化和非蛋白质组分变化(cosgroveetal,1997)。木葡聚糖是使相邻的纤维素微纤维与氢键交联的组分,其赋予植物细胞壁强度。细胞壁生物过程中涉及大量酶,如扩展蛋白(exps),1,4-内切葡聚糖酶(egases)和木葡聚糖内转葡糖基酶/水解酶(xyloglucanendotransglucosylase/hydrolase,xth)等(roseetal,2002;maldonado-mendozaetal,2005;xuanetal,2016)。这些酶通过一级或二级壁松弛机制调节细胞壁的形状,在细胞壁构建,降解,延伸,切割木葡聚糖和重新连接木葡聚糖链的过程中起重要作用。xth属于糖苷水解酶gh16家族的亚家族,xth涉及生理过程,例如次生维管组织的形成,植物组织的伸长等(yokoyamaetal,2001)。最近,xth被赋予了涉及植物对胁迫响应的调节的功能。例如,铝过量暴露导致根中的atxth31显着下调表达(yangetal,2011)。过量表达caxth3可明显提高转基因拟南芥对盐和干旱的耐受性(choetal,2006)。xth在植物抗胁迫育种中有重要的潜力。
技术实现要素:
本发明为提高柑橘对溃疡病的抗性提供一种新的选择,公开一种改良柑橘溃疡病抗性的分子育种方法。本发明公开一种利用csxth04基因沉默以提高柑橘对溃疡病抗性的方法,该方法能够显著提高柑橘对溃疡病的抗性。
本发明通过下述技术方案实现:
一种利用csxth04基因沉默以提高柑橘对溃疡病抗性的方法,包括以下步骤:
(1)克隆柑橘csxth04基因片段,然后进行干扰表达载体构建;
(2)干扰表达载体转化柑橘,得到转基因植株。
其中,步骤(1)中,克隆csxth04基因片段所采用的引物为rnai-csxth04-f和rnai-csxth04-r,分别具有如seqidno:1和seqidno:2所示核苷酸序列。
本发明中,csxth04(cds)具有如seqidno:12所示核苷酸序列。
进一步的,步骤(1)中,柑橘csxth04基因片段的克隆方法为:提取柑橘总rna,然后反转录为cdna,最后采用高保真酶pcr克隆csxth04基因片段。
进一步的,步骤(1)中,干扰表达载体构建方法为:根据csxth04编码序列设计上游分别含有xbaⅰ、swaⅰ酶切位点和保护碱基的特异引物,下游分别含有bamhⅰ、ascⅰ酶切位点和保护碱基的特异引物,以柑橘cdna为模板进行pcr扩增得到干扰片段,片段序列如seqidno:3所示。将产物回收后分两组,第一组用swaⅰ和ascⅰ双酶切,第二组用bamhⅰ和xbaⅰ双酶切,酶切回收的两组片段同时连接到pucr干扰载体的相应部位上,得到中间载体pucr-csxth04-rnai进行测序验证,然后将测序正确的产物和plgne超量表达载体用kpnⅰ和salⅰ分别双酶切,将含有干扰片段的酶切产物连接到plgne表达载体上,构建出最终的干扰表达载体plgne-csxth04-rnai。
进一步的,步骤(1)中,plgne载体带有camv35s启动子和gus编码基因,35s启动子为花菜花叶病毒启动子,gus用作转基因植株检测,分别具有seqidno:4和seqidno:5所示的核苷酸序列。
进一步的,步骤(2)中,干扰表达载体转化柑橘的方法为:干扰表达载体通过电击法转化农杆菌,再用农杆菌介导转化柑橘外植体,遗传转化后的外植体细胞再经离体培养、染色鉴定、嫁接后得到转基因植株。
进一步的,步骤(2)得到转基因植株后,对转基因植株进行抗性评价,判定csxth04基因沉默与柑橘溃疡病的相关性。
进一步的,对转基因植株进行抗性评价前,通过pcr验证转基因植株,采用的引物为id-csxth04-f和id-csxth04-r,分别具有如seqidno:6和seqidno:7所示核苷酸序列。
进一步的,pcr验证后,用qrt-pcr再次验证转基因植株,采用的引物为rt-csxth04-f和rt-csxth04-r,分别具有如seqidno:8和seqidno:9所示核苷酸序列。qrt-pcr内参为柑橘actin基因,采用引物为rt-csactin-f和rt-csactin-r,分别具有如seqidno:10和seqidno:11所示核苷酸序列。
本发明与现有技术相比,具有如下的优点和有益效果:
本发明为一种基于csxth04基因沉默以提高柑橘对溃疡病抗性的方法,通过克隆柑橘csxth04基因片段,进行干扰表达载体构建,然后转化柑橘,得到的转基因植株csxth04的表达量显著下调至现有柑橘的25%,溃疡病发病程度可降低至现有柑橘的35%,因而得出沉默csxth04基因可一定程度上提高植株对溃疡病的抗性。
附图说明
此处所说明的附图用来提供对本发明实施例的进一步理解,构成本申请的一部分,并不构成对本发明实施例的限定。在附图中:
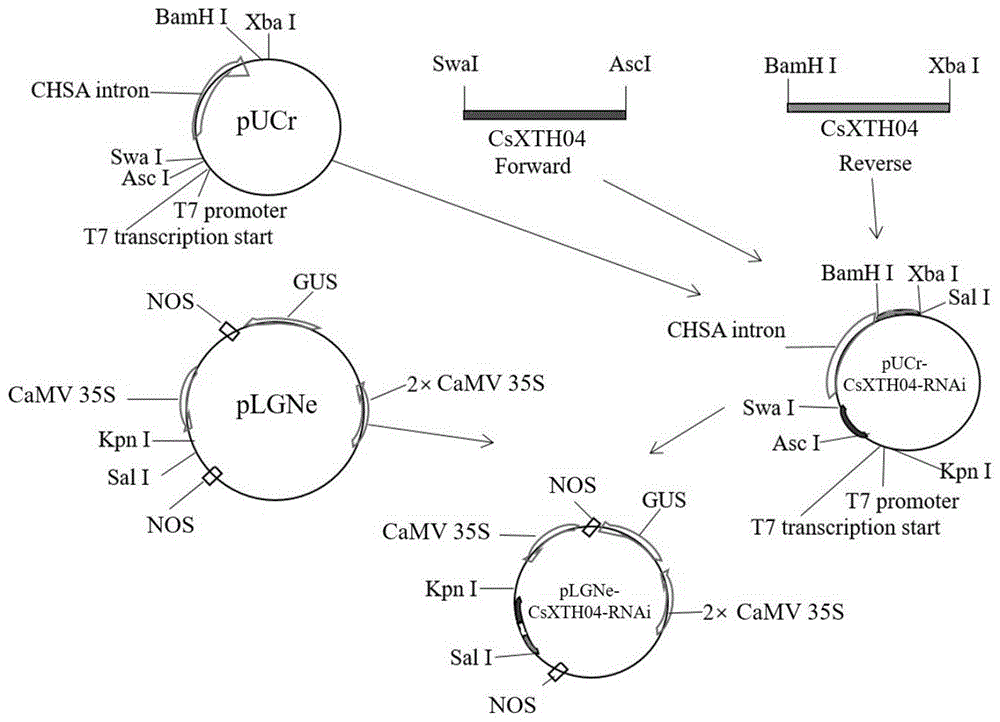
图1为本发明干扰表达载体的构建流程图:gus,β-葡萄糖酸苷酶基因;camv35s,来源于花椰菜花叶病毒的植物组成性启动子;nos,冠瘿碱合成酶基因终止子;载体plgne具有camv35s启动子调控下的gus基因,便于在植物遗传转化过程中对转化子进行gus染色筛选;t7promoter,质粒在大肠杆菌内启动转录使用的启动子;t7transcriptionstart,t7启动子启动转录的起点。
图2为本发明转基因植株的pcr验证图:r1、r2、r3分别代表3个转基因植株;wt代表野生型对照植株,(下同)。
图3为本发明转基因植株的gus染色验证图。
图4为本发明转基因植株的表型。
图5为本发明转基因植株中csxth04的相对表达量:**表示同野生型比较差异极显著(p=0.01),(下同)。
图6为本发明转基因植株叶片接种溃疡病菌十天后的发病情况。
图7为本发明转基因植株接种溃疡病菌10天后的病斑面积统计。
图8为本发明转基因植株接种溃疡病菌10天后的发病程度统计。
具体实施方式
为使本发明的目的、技术方案和优点更加清楚明白,下面结合实施例和附图,对本发明作进一步的详细说明,本发明的示意性实施方式及其说明仅用于解释本发明,并不作为对本发明的限定。
经发明人研究发现:csxth04在溃疡病易感柑橘品种中受溃疡病菌侵染后明显上调表达,说明这个基因是一个潜在的柑橘溃疡病感病基因,所以对其进行沉默,或许可以增加柑橘对溃疡病的抗性。基于此,本发明公开了这种利用csxth04基因沉默以提高柑橘对溃疡病抗性的方法。
本发明实施例以晚锦橙为试验对象,在实际应用中,该方法也能够用于提高其他柑橘品种对溃疡病的抗性。
实施例1
克隆csxth04基因片段
1.rna提取及cdna合成
选取柑橘(晚锦橙)叶片0.1g用easyspin植物rna快速提取试剂盒(艾德莱,cat:rn09)提取叶片总rna,用非变性琼脂糖凝胶电泳验证rna质量,用浓度计测定其浓度。使用recombinantdnasei合成cdna(宝生物),cdna于-20℃保存备用。
2.csxth04基因片段的pcr扩增
使用引物rnai-csxth04-f和rnai-csxth04-r从柑橘cdna中扩增获得csxth04片段,片段长度为216bp。pcr试剂盒采用primestarmastermix(宝生物)。
扩增体系:10xpcrmix:2.5μl;引物rnai-csxth04-f(5μmol/l):1μl;引物rnai-csxth04-r(5μmol/l):1μl;cdna约60ng;加ddh2o至25μl。
扩增程序:94℃,5min;94℃,30s,56℃,30s,72℃,1.5min,35个循环;72℃延伸10min。
3.dna片段回收
紫外灯下,用洁净的刀片切下含有目的片段的琼脂糖凝胶块。回收方法参照试剂盒的使用说明进行(艾德莱),回收片段在浓度测试仪上进行定量。
实施例2
构建csxth04干扰表达载体并转化农杆菌
载体构建流程图如图1,所有限制性内切酶购自(thermo)公司,按照使用说明操作。
具体操作如下:实施例1中获得的csxth04基因片段的pcr产物回收后分两组,第一组用swaⅰ和ascⅰ双酶切,第二组用bamhⅰ和xbaⅰ双酶切,酶切回收的两组片段同时连接到pucr干扰载体的相应部位上,得到中间载体pucr-csxth04-rnai进行测序验证,然后将测序正确的产物和plgne超量表达载体用kpnⅰ和salⅰ分别双酶切,将含有干扰片段的酶切产物连接到plgne表达载体上,构建出最终的干扰表达载体plgne-csxth04-rnai。质粒提取采用试剂盒(艾德莱)。
用电激法将构建的超量表达载体质粒导入根癌农杆菌eha105。预先取冻存的eha105农杆菌感受态细胞(50μl),于冰上融化。加入2μl所构建的超量表达载体的质粒于感受态细胞中,吹打混匀后,冰上放置5min。将混合液转入事先吹干的电击杯底(注意避免产生气泡),并将电击杯放入卡槽调整到正确位点。将电击装置调节为“agr”档,按下电击按钮,检查电击数据确保电击成功。加入1mllb液体培养基至电击杯中,移液枪吹打混匀,移至无菌离心管中,260r/min,28℃摇床振荡培养40min。10000r/min将菌液离心1min,弃上清液(剩约100μl重悬菌体),重悬后用移液枪打到lk固体培养基上(表达载体含有卡那抗性),涂布均匀,28℃倒置暗培养2d。待菌斑长出后,挑取单菌落至lk液体培养基中,恒温摇床上(28℃)振荡过夜,菌液用于进行pcr验证。
实施例3
遗传转化柑橘(晚锦橙)
1.柑橘实生苗上胚轴的获得
取新鲜柑橘(晚锦橙)洗净,用70%酒精表面消毒,在无菌的条件下取出种子,剥掉种皮,接种在种子萌发培养基上萌发,28℃下暗培养2周,然后在16h光照/8h黑暗的光周期下培养1周。无菌条件下取萌发幼苗上胚轴切成1cm左右的茎段,用于根癌农杆菌的遗传转化。
2.根癌农杆菌的制备
用于转染的农杆菌菌液(含csxth04干扰表达载体)加入30%的无菌甘油保存于-70℃的超低温培养箱中。转染前,在含50mg/l卡那霉素的lk固体培养基上划线培养。挑农杆菌单菌落,接种于25ml含有相同抗生素的lk液体培养基中,28℃震荡培养过夜。次日,测浓度后将菌液稀释成od值0.1的菌液进行二摇,3h后,待菌液处于对数生长期(od值为0.5左右)时,于5000r/min离心10min,弃上清,用ph5.4的ms液体培养基重悬后用于转染。
3.柑橘上胚轴茎段的转化
将切成1cm左右的柑橘(晚锦橙)上胚轴茎段在农杆菌中浸泡13min,期间轻微晃动。取出茎段后将表面的菌液吸干;将茎段转移到共培养培养基中,26℃按培养2d。
4.转化子的筛选
共培养完成后,将上胚轴转移到筛选培养基中,28℃暗培养7d,外植体在28℃、16h光照/8h黑暗培养,每两周继代一次。
5.转化子的成苗培养
待幼苗长到1cm以上时,将其切下后嫁接到无菌试管晚锦橙苗,在成苗培养基中进行培养;待幼苗长到5cm左右时将其嫁接到枳实生苗上,在温室中进行培养。
实施例4
转基因植株验证
1.外源基因整合的pcr检测
待长到足够大的时候取初筛得到的植株叶片100mg,使用艾德莱公司试剂盒(cat:dn15)提取基因组dna,pcr检测csxth04干扰片段的整合。pcr反应条件:94℃3min;94℃30s,58℃30s,72℃30s,30次循环;72℃10min。检测引物为id-csxth04-f和id-csxth04-r。pcr结果见图2,阳性植株可以得到1394bp的扩增片段对照植株无扩增。经验证得到3个超量表达转基因植株。
2.转基因植株gus染色鉴定
将转基因植株叶片切成叶圆片(直径7mm),进行gus组织化学染色,如图3所示,阳性植株叶圆片边缘显蓝色,野生型植株叶圆片为白色。
3.转基因植株表型观察
观察分析3株转基因植株表型,发现除了发育快慢稍有差异,其他外观上看并无明显差异。由于本发明得到的转基因植株是嫁接得来的,受到与砧木亲和差异或者伤口愈合等因素的影响,且目前发育时间较短,因此本发明认为其发育方面的不是很显著的差异完全可忽略,认为表型与野生型对照无变化,干扰表达csxth04基因并未对植株的表型和发育产生直接的明显的变化。
实施例5
csxth04基因表达的qrt-pcr分析
提取柑橘叶片,使用easyspin植物rna快速提取试剂盒(艾德莱,catno:rn09)提取叶片总rna,用非变性琼脂糖凝胶电泳验证rna质量,用浓度计测定其浓度。使用recombinantdnasei合成cdna(宝生物)。目的基因的检测引物为rt-csxth04-f和rt-csxth04-r;内参基因actin的检测引物为rt-csactin-f和rt-csactin-r。
反应体积20μl,反应条件:95℃3min,94℃10s;56℃10s,72℃10s,40次循环;72℃10min。实验重复三次。采用2-△△ct法计算转基因植株中csxth04基因的相对表达量:定义水处理的样本为参照因子,即其csxth04的表达水平为1,然后计算转基因柑橘中相对参照因子基因表达的倍数2-△△ct,为其相对表达量。检测结果见图5。结果显示,csxth04基因在转基因植株中相比野生型植株被较大程度沉默,表达量最高下调至现有柑橘的25%。
实施例6
转基因植株的抗性评价
采集成熟叶片清洗后用75%的酒精消毒后置于超纯水中冲洗,置于超净台;以叶脉为中心进行针刺,六针为一组,每侧两组;用移液器点样溃疡病菌液,每针孔点样1μl(1x105cfu/ml)。然后将柑橘叶片的叶柄用浸湿的脱脂棉包裹,石蜡带密封于培养皿,于28℃恒温光照培养箱中培养(16h光照/8h黑暗)。对照组用lb代替溃疡病菌菌液,其他操作保持一致。叶片点菌后培养10天拍照,用imagejv1.47软件统计病斑大小(lesionsizes,ls,mm2)。根据病情指数公式计算发病程度(diseaseindex,di)。按照病斑面积将病情分为0-7共8级,以字母r表示病斑面积,0级(r≤0.25mm2),1级(0.25mm2<r≤0.5mm2),2级(0.5mm2<r≤0.75mm2),3级(0.75mm2<r≤1mm2),4级(1.0mm2<r≤1.25mm2),5级(1.25mm2<r≤1.5mm2),6级(1.5mm2<r≤1.75mm2),7级(r>1.75mm2);根据公式计算发病程度:di=100xσ【各级病斑数x相应级数值】/(病斑总数x最大级数)。
接种溃疡病菌10天后,转基因植株叶片上病斑大小与野生型柑橘叶片上存在大小差异(图6)。对柑橘叶片上的病斑大小和病情指数进行统计后,发现转基因植株叶片上的病斑面积(图7)和发病程度(图8)均小于野生型柑橘叶片,尤其是r3,病情指数仅为野生型柑橘叶片的35%。
由此可见,csxth04的沉默可一定程度上降低溃疡病的病斑面积,减轻溃疡病的发病程度。
根癌农杆转化所用培养基:
种子萌发培养基:ms+30g/l蔗糖+2.5g/lgelrite,ph5.8。
共培养培养基:ms+2mg/lba+0.5mg/liaa+1mg/l2,4–d+100μmolas+30g/l蔗糖+2.5g/lgelrite,ph5.8。
筛选培养基:ms+2mg/lba+0.5mg/liaa+500mg/lcef+50mg/lkan+30g/l蔗糖+2.5g/lgelrite,ph5.8。
成苗培养基:ms+30g/l蔗糖,ph5.8。
以上所述的具体实施方式,对本发明的目的、技术方案和有益效果进行了进一步详细说明,所应理解的是,以上所述仅为本发明的具体实施方式而已,并不用于限定本发明的保护范围,凡在本发明的精神和原则之内,所做的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
序列表
<110>西南大学
<120>一种利用csxth04基因沉默以提高柑橘对溃疡病抗性的方法
<160>12
<170>siposequencelisting1.0
<210>1
<211>38
<212>dna
<213>人工序列(1)
<400>1
gctctagaggcgcgccttcatagcctcctacaaggggt38
<210>2
<211>39
<212>dna
<213>人工序列(2)
<400>2
cgggatccatttaaatgcattctggtgaaggcgtaggga39
<210>3
<211>216
<212>dna
<213>人工序列(3)
<400>3
gtcatagcctcctacaaggggttccacgttgatgggtgtgaggtatctgtgaacgccaag60
ttctgtgagacacagggcaagagatggtgggatcagaaggcgttccaagaccttgacgcc120
taccagtacaggagactcagatgggttcgccagaaatacaccatctataactactgcaac180
gatcgtgcccgcttccctacgccttcaccagaatgc216
<210>4
<211>345
<212>dna
<213>人工序列(4)
<400>4
gtcctctccaaatgaaatgaacttccttatatagaggaagggtcttgcgaaggatagtgg60
gattgtgcgtcatcccttacgtcagtggagatatcacatcaatccacttgctttgaagac120
gtggttggaacgtcttctttttccacgatgctcctcgtgggtgggggtccatctttggga180
ccactgtcggcagaggcatcttcaacgatggcctttcctttatcgcaatgatggcatttg240
taggagccaccttccttttccactatcttcacaataaagtgacagatagctgggcaatgg300
aatccgaggaggtttccggatattaccctttgttgaaaagtctca345
<210>5
<211>1812
<212>dna
<213>人工序列(5)
<400>5
atgttacgtcctgtagaaaccccaacccgtgaaatcaaaaaactcgacggcctgtgggca60
ttcagtctggatcgcgaaaactgtggaattgatcagcgttggtgggaaagcgcgttacaa120
gaaagccgggcaattgctgtgccaggcagttttaacgatcagttcgccgatgcagatatt180
cgtaattatgcgggcaacgtctggtatcagcgcgaagtctttataccgaaaggttgggca240
ggccagcgtatcgtgctgcgtttcgatgcggtcactcattacggcaaagtgtgggtcaat300
aatcaggaagtgatggagcatcagggcggctatacgccatttgaagccgatgtcacgccg360
tatgttattgccgggaaaagtgtacgtatcaccgtttgtgtgaacaacgaactgaactgg420
cagactatcccgccgggaatggtgattaccgacgaaaacggcaagaaaaagcagtcttac480
ttccatgatttctttaactatgccggaatccatcgcagcgtaatgctctacaccacgccg540
aacacctgggtggacgatatcaccgtggtgacgcatgtcgcgcaagactgtaaccacgcg600
tctgttgactggcaggtggtggccaatggtgatgtcagcgttgaactgcgtgatgcggat660
caacaggtggttgcaactggacaaggcactagcgggactttgcaagtggtgaatccgcac720
ctctggcaaccgggtgaaggttatctctatgaactgtgcgtcacagccaaaagccagaca780
gagtgtgatatctacccgcttcgcgtcggcatccggtcagtggcagtgaagggcgaacag840
ttcctgattaaccacaaaccgttctactttactggctttggtcgtcatgaagatgcggac900
ttgcgtggcaaaggattcgataacgtgctgatggtgcacgaccacgcattaatggactgg960
attggggccaactcctaccgtacctcgcattacccttacgctgaagagatgctcgactgg1020
gcagatgaacatggcatcgtggtgattgatgaaactgctgctgtcggctttaacctctct1080
ttaggcattggtttcgaagcgggcaacaagccgaaagaactgtacagcgaagaggcagtc1140
aacggggaaactcagcaagcgcacttacaggcgattaaagagctgatagcgcgtgacaaa1200
aaccacccaagcgtggtgatgtggagtattgccaacgaaccggatacccgtccgcaaggt1260
gcacgggaatatttcgcgccactggcggaagcaacgcgtaaactcgacccgacgcgtccg1320
atcacctgcgtcaatgtaatgttctgcgacgctcacaccgataccatcagcgatctcttt1380
gatgtgctgtgcctgaaccgttattacggatggtatgtccaaagcggcgatttggaaacg1440
gcagagaaggtactggaaaaagaacttctggcctggcaggagaaactgcatcagccgatt1500
atcatcaccgaatacggcgtggatacgttagccgggctgcactcaatgtacaccgacatg1560
tggagtgaagagtatcagtgtgcatggctggatatgtatcaccgcgtctttgatcgcgtc1620
agcgccgtcgtcggtgaacaggtatggaatttcgccgattttgcgacctcgcaaggcata1680
ttgcgcgttggcggtaacaagaaagggatcttcactcgcgaccgcaaaccgaagtcggcg1740
gcttttctgctgcaaaaacgctggactggcatgaacttcggtgaaaaaccgcagcaggga1800
ggcaaacaatga1812
<210>6
<211>28
<212>dna
<213>人工序列(6)
<400>6
cttcaccagaatgcatttaaatgtgtaa28
<210>7
<211>28
<212>dna
<213>人工序列(7)
<400>7
ggtcttacgggatccaaatacctgcaaa28
<210>8
<211>20
<212>dna
<213>人工序列(8)
<400>8
tcagaaggcgttccaagacc20
<210>9
<211>20
<212>dna
<213>人工序列(9)
<400>9
ctggtgaaggcgtagggaag20
<210>10
<211>18
<212>dna
<213>人工序列(10)
<400>10
catccctcagcaccttcc18
<210>11
<211>19
<212>dna
<213>人工序列(11)
<400>11
ccaaccttagcacttctcc19
<210>12
<211>882
<212>dna
<213>人工序列(12)
<400>12
atggcttcttttctatggactctttgcctgagtgctctgtttttggcttctggagttata60
agtgccccaccaaagaggcctatagatgtaccatttggccgaaattatgtgccaacatgg120
gcatttgatcacatcaaatactttaatggaggttctgagattcagcttgtcctggataaa180
tacacagggactggcttccaatcaaaaggctcttatctgtttggtcacttcagcatgcac240
ataaagatggttcctggtgattctgccggaactgtgactgctttttacttatcttctcaa300
aattcggaacatgatgagatagactttgagttcttggggaacagaaccgggcagccatac360
attttgcagacaaatgtgtacactggaggaaagggagacaaagagcagagaatttatctt420
tggtttgatccaaccaaagagtttcacacttactcagttctctggaacatgtatcagatt480
gttttcttagtggacgatgtgccgattagggtgttcaagaacagcaaagatttgggagtg540
aggtttccattcaaccaacccatgaagatatactcaagcctgtggaatgcagatgactgg600
gcaacgaggggcggtttggagaagacggactggtcgaaggctcctttcatagcctcctac660
aaggggttccacgttgatgggtgtgaggtatctgtgaacgccaagttctgtgagacacag720
ggcaagagatggtgggatcagaaggcgttccaagaccttgacgcctaccagtacaggaga780
ctcagatgggttcgccagaaatacaccatctataactactgcaacgatcgtgcccgcttc840
cctacgccttcaccagaatgcaaacgagaccgtgacatttaa882
- 还没有人留言评论。精彩留言会获得点赞!