冰岛硫化叶菌引发酶突变体CHI-ΔX菌株及其构建方法与应用
本发明属于微生物菌株改造。
背景技术:
1、古菌-真核类型的引发酶作为古菌dna复制起始过程中合成引物的关键酶,由催化小亚基pris和非催化大亚基pril构成。硫化叶菌引发酶非催化亚基pril的c末端(pril-ctd)含有4个半胱氨酸(cys)残基可以形成铁硫簇。先前研究发现,缺失pril-ctd后,古菌引发酶在体外具有引物合成活性(lao-sirieix,et al.2005);而真核生物引发酶缺失该区域后,不再具有引物合成能力(klinge,et al.2007;weiner,et al.2007)。2015年本实验室研究发现硫化叶菌中还存在另外一个非催化亚基prix可以与prisl存在相互作用且能在生理温度(75℃)下明显提高prisl的引发活性。系统发育学分析显示prix的c末端(prix-ctd,该末端不含铁硫簇)与真核生物的pril的c末端存在相似性(liu,et al.2015)。holzer等人的进一步研究表明含有野生型pris以及由pril的n端结构域和prix的c端部分组成的融合蛋白prisln-xc与野生型引发酶异源三聚体prislx一样可以在体外高效地合成引物。因此,体外引物的合成不依赖于fe-s簇(holzer,et al.2017)。因此,古菌引发酶的铁硫簇所起的作用应该不同于真核生物。对于古菌引发酶铁硫簇的研究可能进一步揭示古菌引发酶的引物合成机制及其生理功能。
技术实现思路
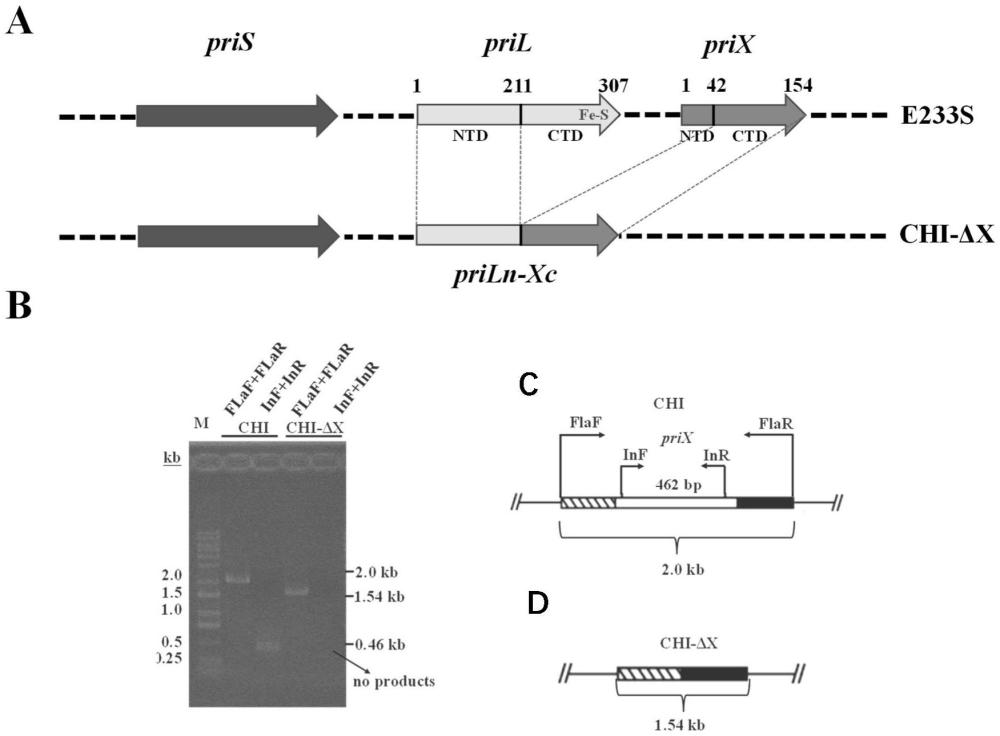
1、有鉴于此,本发明构建了一株冰岛硫化叶菌引发酶突变体chi-δx菌株。该菌株含有pril的n末端(pril-ntd)和prix-ctd融合在一起的嵌合蛋白priln-xc的dna片段;所述priln-xc的基因序列如序列表seq id no.4所示。
2、同时,还提供了该突变体chi-δx菌株的构建方法,将极端嗜酸嗜热古菌冰岛硫化叶菌(sulfolobus islandicus)菌株中的prix-ctd取代pril-ctd的同时并敲除了编码prix的基因后获得;所述prix-ctd的基因序列如序列表seq id no.3所示;所述pril-ctd的基因序列如序列表seq id no.2所示;所述prix的基因序列如序列表seq id no.5所示。
3、本发明是利用古菌ia型crispr遗传操作系统在极端嗜酸嗜热古菌冰岛硫化叶菌e233s菌株(亲本株)中获得了一个不含铁硫簇的引发酶)嵌合突变体菌株,该菌株在90℃以及低铁培养条件下比亲本菌株生长更好;体外生化活性实验也表明嵌合突变体蛋白比野生型蛋白更耐高温和低铁。因此该突变体菌株可能更具某些抗胁迫潜力,可进一步作为嗜热古菌的底盘细胞使用。
技术特征:
1.冰岛硫化叶菌引发酶突变体chi-δx菌株,其与出发菌株相比,含有嵌合蛋白priln-xc的dna片段并敲除prix;
2.权利要求1所述冰岛硫化叶菌引发酶突变体chi-δx菌株的构建方法,其特征在于,
3.微生物菌剂,其特征在于,包括权利要求1所述的冰岛硫化叶菌引发酶突变体chi-δx菌株。
4.化合物,其特征在于,包括权利要求1所述的冰岛硫化叶菌引发酶突变体chi-δx菌株。
5.权利要求1所述的冰岛硫化叶菌引发酶突变体chi-δx菌株在制备耐高温菌剂中的应用,其特征在于,所述高温为90℃。
6.权利要求1所述的冰岛硫化叶菌引发酶突变体chi-δx菌株在制备低铁菌剂中的应用。
技术总结
本发明属于微生物菌株改造技术领域。本发明提供了冰岛硫化叶菌引发酶突变体CHI‑ΔX菌株,其与出发菌株相比,含有嵌合蛋白PriLn‑Xc的DNA片段并敲除priX;所述PriLn‑Xc的基因序列如序列表SEQ ID NO.4所示;所述priX的基因序列如序列表SEQ ID NO.5所示;所述出发菌株为冰岛硫化叶菌E233S。同时还提供了该突变体CHI‑ΔX菌株的构建方法,将极端嗜酸嗜热古菌冰岛硫化叶菌(Sulfolobus islandicus)E233S菌株中的PriX‑CTD取代PriL‑CTD同时敲除PriX基因后获得;所述PriX‑CTD的基因序列如序列表SEQ ID NO.3所示;所述PriL‑CTD的基因序列如序列表SEQ ID NO.2所示;所述PriX的基因序列如序列表SEQ ID NO.5所示。本发明提供的突变体CHI‑ΔX菌株能在90℃以及低铁培养条件下比亲本菌株生长更好;体外生化活性实验也表明嵌合突变体蛋白比野生型蛋白更耐高温和低铁。
技术研发人员:黄力,熊代江
受保护的技术使用者:中国科学院微生物研究所
技术研发日:
技术公布日:2024/8/26
- 还没有人留言评论。精彩留言会获得点赞!