一种黄籽甘蓝型油菜的获取方法与流程
本发明属于农业技术领域,特别是甘蓝型油菜黄籽新种质资源的获取技术和鉴定的方法。
背景技术:
黄籽油菜具有相对含油量高、菜籽油品质好、菜籽粕营养价值高等优点,选育黄籽油菜已成为当前油菜育种的主要目标之一,成为提高种子含油量、改进油菜品质的一条重要途径。白菜型油菜和芥菜型油菜均有不少黄籽品种,且黄籽性状稳定。而甘蓝型油菜不存在天然黄籽种质,黄色种皮基因资源匮乏。目前培育出的黄籽甘蓝型油菜均来自芸苔属内种间杂交,还未见通过属间杂交获得黄籽甘蓝型油菜的报道,且现有获得的属内起源的甘蓝型油菜黄籽性状存在籽色不纯,遗传不稳定的问题。白芥属白芥种与甘蓝型油菜同属十字花科,其种皮颜色为纯黄色,且遗传稳定。因此,引进白芥属黄籽基因,对拓宽甘蓝型油菜黄籽性状遗传基础,选育籽色较纯、遗传稳定的黄籽甘蓝型油菜将具有非常重要的意义。
技术实现要素:
本发明目的在于提供一种种子含油量高、黄色种皮遗传稳定的黄籽甘蓝型油菜的获取方法。
本发明技术方案包括以下步骤:
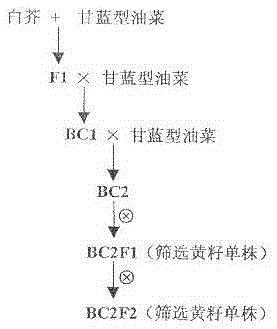
1)通过芸苔属甘蓝型油菜与白芥属白芥野生种原生质体融合,获得属间杂种F1;
2)将属间杂种F1与亲本芸苔属甘蓝型油菜回交,获得BC1;
3)将BC1与亲本芸苔属甘蓝型油菜再回交,获得BC2;
4)从BC2自交后代中筛选获得褐黄籽单株,获得BC2F1;
5)将BC2F1进一步自交,获得具有白芥黄籽表现型的易位系BC2F2,即黄籽甘蓝型油菜种质资源。
通常黄籽甘蓝型油菜种质资源的创建选自同属的不同种进行杂交,而本发明杂交的两个亲本来自不同属,该远缘杂交可以实现不同种间、属间甚至亲缘关系更远的物种间的杂交突破种属界限,扩大遗传变异,从而创造新的变异类型或新物种。但随着亲缘关系差距的加大,不同种属的物种杂交常表现杂交不亲和性。原生质体融合技术可避免受精作用中种的特异性配子识别反应,打破远缘杂交中的有性不亲和界限。通过该技术可克服远缘杂交不亲和性,将十字花科不同属优良性状导入现有甘蓝型油菜栽培品种中,实现优良基因的属间转移。
本发明以白芥属白芥野生种与甘蓝型油菜经原生质体融合获得属间杂种F1,并用亲本甘蓝型油菜回交至BC2,从BC2自交后代中筛选获得褐黄籽单株,该单株进一步自交,筛选获得6个遗传稳定、不同程度黄籽表现型株系,经细胞学和分子生物学鉴定,这6个黄籽材料均为带有白芥黄籽基因的易位系材料。使用丹麦FOSS公司TR-3700近红外分析仪测定黄籽株系的含油量,表明本发明获取的黄籽单株含油量明显高于白芥属白芥型,较芸苔属甘蓝型提高1.7-6%。
由于通过远缘杂交技术获得的杂种及其回交后代中附加有大量外源植物染色体,遗传不稳定,难以在育种直接应用,因此有必要通过细胞学和分子生物学的方法对获得的后代进行鉴定,以筛选遗传稳定的易位系,为育种应用奠定基础。原位杂交(GISH)和分子标记鉴定已成为外源遗传物质鉴定的有效方法。GISH技术利用核酸序列间的同源互补,已被广泛用于分析植物基因组结构、确定外源DNA的插入位点及定量分析、鉴定外源染色体片断、分析植物杂交品种染色体组成来源以及近缘种属或品系的比较研究。分子标记技术(包括RFLP、RAPD、SSR、AFLP等)可在DNA水平上迅速、有效的反映生物个体之间的差异,因而可应用于异染色体系鉴定及辅助育种。
本发明针对分析该黄籽甘蓝型油菜来源的特殊性,对杂种后代减数分裂进行GISH分析,发现甘蓝型油菜的染色体在回/自交各个世代中均能正常配对和分离,部分白芥染色体与甘蓝型油菜染色体形成三价体,可能发生异源染色体重组;BC2F2黄籽材料染色体数目与正常甘蓝型油菜相同,无杂交信号,排除该材料是异附加系或置换系的可能性。微卫星核心序列引物33.6扩增结果证明,在黄籽后代基因组中存在白芥基因组特异序列。类黄酮基因引物TT2-2获得一致结果,即黄籽后代中具有白芥DNA序列;同时获得黄籽特异性分子标记Sa1,为分子标记辅助育种奠定了基础。结合细胞学鉴定结果,本发明可以确认获得的黄籽后代株系是带有白芥DNA序列的易位系。
附图说明
图1为本发明技术方案的流程图。
图2为本发明获得的6个白芥属来源的甘蓝型油菜黄籽株系种子照片图。
具体实施方式
一、获取黄籽甘蓝型油菜种质资源的步骤,如图1所示:
1、白芥属白芥野生种与芸苔属甘蓝型油菜的原生质体融合及植株再生:
将白芥属白芥野生种种子和芸苔属甘蓝型油菜种子分别经70%(v/v)酒精消毒1min、2%(v/v)次氯酸钠消毒15min,无菌水冲洗3遍后,接种于MS培养基,于25℃光照条件下培养16h。
分别取3~4周龄叶片,分别分离出白芥属白芥型油菜原生质体和芸苔属甘蓝型油菜原生质体。
将白芥属白芥型油菜原生质体和芸苔属甘蓝型油菜原生质体以1∶2体积比混合至终浓度≥5×105个/mL后,进行电融合。
将电融合的原始质体于CR培养基中25℃暗培养,2~3周后形成的愈伤组织转至固体培养基(MS+10g/l蔗糖+20g/l甘露醇+250mg/l甘酪素水解物+6g/l琼脂+0.5g/l NAA+0.5g/l 6-BA+1g/l TDZ),25℃16h光照条件下培养。4周后转至分化培养基(MS培养基+20g/l蔗糖+250mg/l甘酪素水解物+10g/l琼脂+10g/l 6-BA+0.04g/l GA3)。3~4周后生根后转到无激素的MS培养基上生长,最终获得7棵属间杂种F1。
2、体细胞杂种后代的黄籽筛选:
将获得的属间杂种F1与芸苔属甘蓝型油菜回交至BC2。
从BC2自交后代中筛选褐黄籽单株,即得BC2F1。
将BC2F1进一步自交,筛选获得6个遗传稳定、不同程度黄籽表现型株系BC2F2。
上述黄籽株系BC2F2继续自交并无分离,相关基因已纯合并稳定遗传。
3、黄籽甘蓝型油菜的细胞学鉴定:
本发明按试剂盒(Catalogue No.976776,Roche,德国)标记白芥基因组为探针,对杂种后代减数分裂和根尖细胞染色体制片进行基因组原位杂交(GISH),使用Olympus BX51荧光显微镜观察染色体和杂交信号,对于分散较好的细胞在不同的滤光片进行拍照,用Image-Pro Plus Version 5.0软件进行图片合成和处理。结果发现甘蓝型油菜的染色体在回/自交各个世代中均能正常配对和分离,而白芥染色体多以单价体形式存在,部分白芥染色体会与甘蓝型油菜染色体形成三价体,可能发生异源染色体重组,为易位系的产生奠定基础。BC2F2黄籽材料根尖细胞和减数分裂细胞的GISH鉴定显示,获得的黄籽材料染色体数目与正常甘蓝型油菜相同,无杂交信号,排除该材料是异附加系或置换系的可能性。
4、黄籽甘蓝型油菜的分子标记鉴定:
本发明参考Somers(2002)的研究结果,利用微卫星核心序列33.6引物(AGGGCTGGAGG)对黄籽后代进行PCR鉴定,扩增产物于9%聚丙烯酰胺凝胶电泳上检测,结果表明黄籽后代的基因组中存在与白芥相同的特异序列(300bp)。同时根据拟南芥类黄酮合成相关基因设计引物对黄籽后代进行PCR鉴定,其中TT2-2引物(正向序列为:5′-CAGAATCTATTCTCAACACAACAC-3′,反向序列为:5′-TCTAGATAWTAYTWGTAATTAGAAG-3′)的扩增结果也表明:黄籽后代中存在白芥特异片段。将PCR扩得的白芥特异条带经TIANgel MidiPurification Kit(QIAGEN)进行切胶回收、纯化,连接pMD19-TVector(TaKaRa),热激转化大肠杆菌DH5α,进行Amp抗性和X-gal/IPTG蓝白斑筛选。选白色菌落进行菌落PCR,结果为阳性的菌落进行LB液体培养,碱裂解法提取质粒。以质粒为模板进行PCR检测。选取含阳性质粒的菌液送上海生工生物工程技术服务有限公司测序。根据TT2-2扩增特异产物测序结果设计的引物Sa1:
正向序列为5′-GAAGTAGTAGGTGGAAAGC-3′,
反向序列为5′-AAGGCAGTAAGGATGTAA-3′
在白芥和黄籽后代中扩增的特异产物具有一致性。
5、黄籽甘蓝型油菜的品质分析:
如图2所示,白芥属白芥野生种(图2中标记:白芥)具有黄籽性状,籽粒颜色淡黄色。
回交亲本芸苔属甘蓝型油菜(图2中标记:扬油6号)籽粒颜色为褐色。
在回交后代BC2F2中筛选出多个具有黄籽或趋向黄籽性状的株系,籽色颜色从红偏黄到鲜黄各个梯度均有如图2中标记D244-18、D244-52、D244-6、D255-3、D246-5、D211-1。
本发明使用丹麦FOSS公司TR-3700近红外分析仪测定黄籽株系的含油量,表明黄籽单株含油量明显提高,黄籽株系D244-18、D244-52、D244-6、D255-3、D246-5、D211-1的含油量分别为43.32%、43.35%、47.60%、42.94%、42.79%、44.57%。而扬油6号和白芥的含油量分别为41.63%和39.04%。结果表明:本发明获取的黄籽材料含油量较芸苔属甘蓝型油菜提高1.7-6%。
- 还没有人留言评论。精彩留言会获得点赞!