开环异落叶松脂素二葡糖苷(SDG)和相关化合物用于防护辐射和化学损害的用途的制作方法
政府利益
本发明是在由国立卫生研究院授予的资助号r01(ca133470),1p30es013508-02,rc1ai081251和5-p30-ca-016520-34s2的政府支持下进行。政府在本发明中享有一定的权利。
发明领域
本文提供了使用断裂开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其立体异构体、其代谢物及其类似物,用于辐射防护和辐射缓解以及用于化学预防,如避免致癌物诱导的肺癌和间皮瘤或避免次氯酸离子的组合物和方法。
发明背景
电离辐射在生物机体中产生大范围的有害影响。人类因为职业危害暴露于辐射,在诊断和治疗放射照相程序期间,当使用电子设备时,来自核事故的背景辐射,在空中和太空旅行期间,以及来自长时间暴露于太阳(例如,日光浴者或户外工人)。暴露于自然辐射可能以许多形式出现:自然资源如空气、水和土壤在与自然发生的辐射放射物质(放射性核素)接触时可能受到污染;氡是一种这样常见的自然辐射源。当前的全球发展还将恐怖主义定为危险的手段,通过这些手段可能使大量的人暴露于致命量的辐射。因此,鉴别可以在暴露于辐射之前和期间(即,辐射防护剂)以及作为放射性暴露后的治疗(即,辐射缓解剂)给予的试剂是非常重要的。
此外,在美国,肺癌是癌症死亡率的主要原因。尽管存在新颖的靶向治疗剂,改进的分期和外科技术,以及增加针对局部晚期肺癌的伴随化放疗的利用,但是总体死亡率仅有最少降低(坦(tan)和斯皮瓦克(spivack)(2009)肺癌(lungcancer)65:129-137)。癌症化学预防已被定义为使用具有特定天然或合成试剂的饮食和药理学干预,这些天然或合成试剂被设计来预防,抑制或逆转恶性肿瘤发展之前的致癌作用过程(洪(hong)&斯伯恩(sporn),(1997),科学(science)278:1073-1077)。用于肺癌化学预防的一种策略集中于通过解毒ii期酶的上调来调节烟草、环境和其他致癌物的代谢和分布的试剂的使用。已知许多合成和天然发生的化合物诱导ii期酶的表达。已有许多报道支持nrf2/are调节的ii期酶诱导是降低对致癌物的易感性的高度有效策略的想法。我们有数据表明亚麻籽(fs)及其主要木脂素sdg,二者都来自天然材料的富集和合成衍生,是化学致癌物诱导的肺癌的小鼠模型中有效的肺癌化学预防试剂。
大约85%的肺癌是由吸烟引起的。烟草烟雾中的主要肺致癌物是多环芳烃,以苯并[a]芘(bap)为代表。在开发出更好的治疗之前,减少死亡的最佳希望是通过筛查、戒烟或化学预防进行预防。化学预防剂必须在大量暴露但相对健康的受试者中长时间给予。它们必须是安全、无毒、适口,并且理想地,是负担得起的。已经在肺癌中研究了许多化学预防剂,但没有一种符合这些标准。最有希望的方法之一是上调ii期抗氧化剂和解毒酶。不幸的是,迄今为止在患者中测试的ii期酶激活剂,如奥替普拉或莱菔硫烷(sulforophane),已经证明是不可接受的毒性(彭迪亚拉(pendyala)等人,(2001),癌症流行病学、生物标志物和预防(cancerepidemiolbiomarkersprev)10:269-272)。因此,迫切需要安全,无毒的化学预防剂,其有效地防止由烟草烟雾中的肺致癌物诱导的氧化应激和dna损伤。
另一种众所周知的环境致癌物是石棉,其是指商业上用于绝缘的一组天然发生的水合纤维状硅酸盐纤维。现在已经在两种动物模型和患者中清楚地建立,石棉纤维吸入可导致肿瘤性疾病,如恶性间皮瘤(mm)和肺癌(卡蓬(carbone)&杨(yang)(2012),临床癌症研究(clincancerres)18:598-604;内里(neri)等人(2012),抗癌研究(anticancerres)32:1005-1013),以及肺纤维化。mm是由胸膜和腹膜的间皮细胞产生的高度侵袭性癌症,中位存活期为约1年(斯特曼(sterman)等人,(2005),临床癌症研究(clincancerres)11,7444-7453;斯特曼等人,(1999),胸(chest)116,504-520;贝纳德(benard)等人,(1999),核医学杂志(jnuclmed)40,1241-1245)。除了在非常早期的疾病中的手术,目前的治疗不是有疗效的(斯特曼&阿尔贝达(albelda)(2005),呼吸病学(respirology)10,266-283)。目前,mm在美国每年导致约3,000例死亡,并且在西欧每年导致额外5,000例死亡。
虽然石棉的使用在许多西方国家受到限制,但在世界许多国家中仍然使用石棉,并且据估计2008年开采的石棉超过200万吨(苏尔维(survey),b.g.(2010),世界矿产(worldmineralproduction)2004-08.诺丁汉,英国,英国地质调查所(britishgeologicalsurvey))。因此,第三世界(特别是印度)的mm病例可能会大幅增加,在那里使用石棉的人数增加了,而且很少采取预防措施。然而,即使在发达世界,重要的风险仍然存在。这些包括许多类型的职业,这些职业使工人暴露于预先存在的石棉(即,水管工、管道工、从事绝缘工作的工人,绝缘层去除等)以及超级石棉危险废物场所。还有环境和居家暴露。例如,在采矿或石棉工厂关闭的地区,mm的风险增加。
石棉和癌症之间的关系的一个主要问题是吸入的石棉纤维可以在肺中持续很长时间,导致连续损伤,即使患者从暴露中移除。由于这种长的潜伏期(通常长达30-50年),过去暴露的个体在整个生命中仍然存在增加的mm和其他癌症的风险。
癌症的化学预防目的在于预防、阻止或逆转致癌作用的起始阶段或肿瘤细胞进展为癌症。虽然这个定义听起来很简单,但是很难找到有效的化学预防剂。首先,致癌物诱导癌症的机制通常涉及多种机制,使得功效具有挑战性并且需要具有多种活性的试剂。其次,由于该试剂将用于预防大量健康的但处于危险的个体中的少量肿瘤,因此其必须是非常无毒,良好耐受且可负担的。
因此,鉴别可以在暴露于致癌物或其他有害化学试剂(即,化学预防剂)(如,化学战剂,氯和次氯酸离子和其他有害毒物)之前,期间和之后给予的试剂也是非常重要的。
技术实现要素:
在一方面,本发明涉及用于在对其有需要的受试者中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免受辐射损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、降解物或其代谢物。向所述受试者给予涵盖在暴露于损伤性辐射暴露之前,期间和之后的给予。时间之前,期间和之后可以是秒、分钟、小时、天、周、月或甚至年。生物活性成分涵盖开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于治疗或预防已经或将要暴露于辐射的受试者中的辐射损伤的方法,该方法包括:向所述受试者给予有效量的至少一种生物活性成分,其中所述生物活性成分包括开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于在对其有需要的受试者中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免受意外辐射暴露所导致的辐射损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、降解物或其代谢物。
在另一方面,本发明涉及用于保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞或组织免受导致衰老的辐射损伤的方法。
在另一方面,本发明涉及用于保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞或组织免受由暴露于天然和合成的化学致癌物质和毒物产生的损害的方法。
在另一方面,本发明涉及用于在对其有需要的受试者中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免受针对癌症治疗的放射疗法所导致的辐射损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、降解物或其代谢物。
在另一方面,本发明涉及用于在对其有需要的受试者中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免受辐射损伤的方法,该方法包括:向所述受试者给予有效量的至少一种生物活性成分,其中所述生物活性成分包括开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。在一些实施例中,辐射损伤由意外的辐射暴露引起。在一些实施例中,辐射损伤由用于癌症(例如,肺癌)治疗的放射疗法引起。
在另一方面,本发明涉及用于在对其有需要的受试者中预防对生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织的辐射诱导的损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、或其代谢物。
在另一方面,本发明涉及用于在对其有需要的受试者中预防对生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织的辐射诱导的损伤的方法,该方法包括:向所述受试者给予有效量的至少一种生物活性成分,其中所述生物活性成分包括开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于在细胞中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免受辐射损伤的方法,该方法包括:将所述细胞与有效量的至少一种生物活性成分接触,其中所述生物活性成分包括开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于在对其有需要的受试者中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免受致癌物损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、降解物或其代谢物。向所述受试者给予涵盖在暴露于天然和合成的化学致癌物和毒物的损伤性暴露之前,期间和之后的给予。时间之前,期间和之后可以是秒、分钟、小时、天、周、月或甚至年。生物活性成分涵盖开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于在对其有需要的受试者中保护生物分子免受由于意外暴露于天然和合成的化学致癌物和毒物引起的致癌物损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、降解物或其代谢物。
在另一方面,本发明涉及用于保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞或组织免受来自天然和合成的导致肺癌或间皮瘤的化学致癌物和毒物的损害的方法。
在另一方面,本发明涉及用于保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞或组织免于致癌物损伤的方法,该方法包括:将所述生物分子、细胞或组织与有效量的生物活性成分接触。与所述生物分子、细胞或组织接触涵盖在暴露于天然和合成化学致癌物和毒物的损伤性暴露之前,期间和之后的接触。时间之前,期间和之后可以是秒、分钟、小时、天、周、月或甚至年。生物活性成分涵盖开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于在已经或将要暴露于一种或多种致癌物的受试者中治疗或预防致癌物诱导的损伤,恶性转化或癌症发展避免致癌物诱导癌症的方法,该方法包括:向所述受试者给予有效量的至少一种生物活性成分,其中所述生物活性成分包括开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及保护暴露于一种或多种致癌物的受试者免受致癌物诱导的癌症的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分或其代谢物。
在另一方面,本发明涉及用于在对其有需要的受试者中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免由次氯酸根离子造成的损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、降解物或其代谢物。向所述受试者给予涵盖在暴露于天然和合成的化学致癌物和毒物的损伤性暴露之前,期间和之后的给予。时间之前,期间和之后可以是秒、分钟、小时、天、周、月或甚至年。生物活性成分涵盖开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于治疗或预防已经或将要暴露于次氯酸根离子的受试者中的次氯酸根离子诱导的损伤的方法,该方法包括:向所述受试者给予有效量的至少一种生物活性成分,其中所述生物活性成分包括开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞或组织免于由次氯酸根离子造成的损伤的方法,该方法包括:将暴露于或将暴露于次氯酸根离子的所述生物分子、细胞或组织与有效量的生物活性成分接触。与所述生物分子、细胞或组织接触涵盖在暴露于天然和合成化学致癌物和毒物的损伤性暴露之前,期间和之后的接触。时间之前,期间和之后可以是秒、分钟、小时、天、周、月或甚至年。生物活性成分涵盖开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于前述方法之一的组合物。
除非另外定义,在此所使用的全部技术术语和科学术语具有与本发明所属领域之内的普通技术人员通常所理解的相同的意思。本发明的其他特征与优点从以下详细的说明实例以及图中将会变得清楚。然而,应当理解,这种详细说明和这些具体实例虽然指示了本发明的优选的实施例,但它们仅仅是通过说明的方式给出的,因为从这种详细说明在本发明的精神和范围内的不同变化和修改对于本领域的那些普通技术人员而言将变得很清楚。还预期,在适当时,本发明的任何实施例可以与本发明的一个或多个其他实施例组合,即使在本发明的不同方面下描述实施例。
附图简要说明
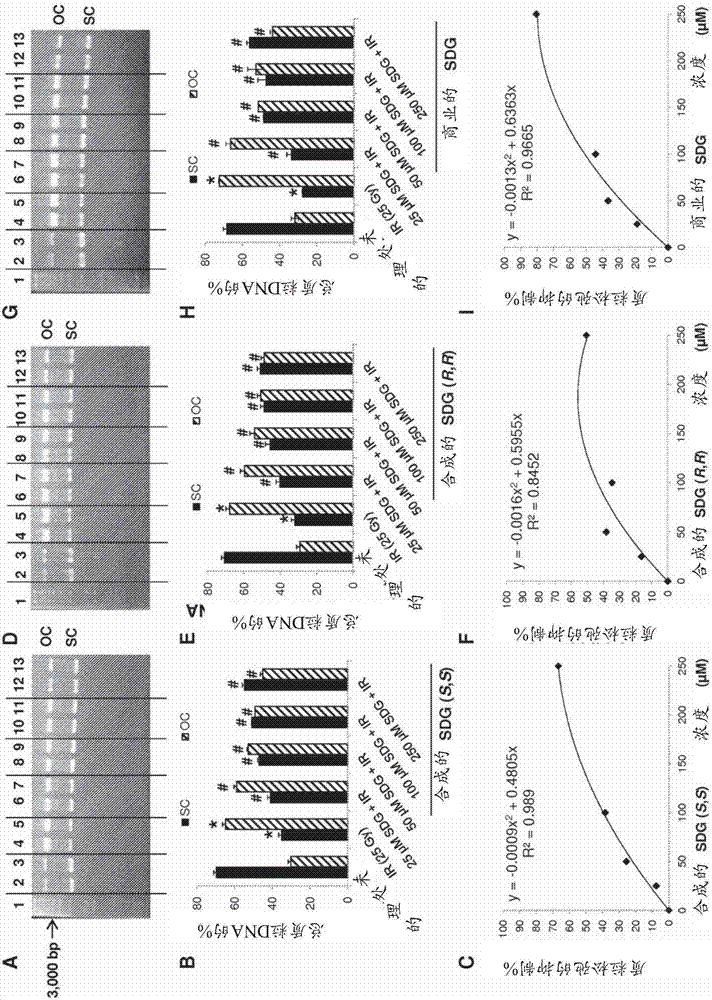
图1:增加剂量的γ辐射对质粒(pbr322)dna松弛的影响。超级螺旋(sc)代表紧凑形式。开放螺旋(oc)形式代表质粒的松弛形式或损伤形式。(a)-超级螺旋(sc)形式是下突出带(在3,000bp处),而开放螺旋(oc)形式是上突出带。泳道1-1kbdna标准梯,泳道2和3-未处理的质粒dna,泳道4和5-暴露于10gy的质粒dna,泳道6和7-暴露于25gy的质粒dna,泳道8和9-暴露于50gy的质粒dna。(b)-sc和oc形式表示为总质粒dna的百分比。针对每种条件,所有样品一式两份运行。数据表示为平均值±标准偏差。p<0.05被认为是有意义的。*和**分别显示与未处理的sc和oc形式相比的显著差异。
图2:增加浓度的合成sdg(s,s)、sdg(r,r)和商业sdg对γ辐射诱导质粒(pbr322)dna松弛的影响。将所有样品暴露于25戈瑞的γ射线剂量。sdg浓度为25、50、100和250μm。在图(a)、(d)、和(g)中-显示了在25、50、100和250μmsdg(s,s)、sdg(r,r)和sdg(商业)存在下暴露于25gy后的质粒dna的代表性琼脂糖凝胶扫描。泳道1-1kbdna标准梯,泳道2和3-未处理的质粒dna,泳道4和5-25μm,泳道6和7-50μm,泳道8和9-100μm,泳道10和11-250μmsdg。在图(b)、(e)和(h)中sc和oc形式表示为总质粒dna的百分比。针对每种条件,所有样品一式两份运行。数据表示为平均值±标准偏差。p<0.05被认为是有意义的。*和#分别显示与未处理的sc和oc形式相比的显著差异。**和##显示与暴露于25gy不含sdg的样品相比的显著差异。在图(c)、(f)和(i)中,显示了质粒dna松弛的sdg依赖性抑制。从曲线下呈现的二次方程式确定ec50值。
图3:增加剂量的γ辐射对小牛胸腺dna片段化的影响。暴露于γ辐射的dna产生小分子量的片段,其移动比更高分子量dna更快。确定低分子量dna片段(<6,000bp)的密度,与高分子量dna(>6,000bp)相比,反映了辐射诱导损伤的程度。(a)泳道1-1kbdna标准梯,泳道2和3-未处理的小牛胸腺dna,泳道4和5-暴露于25gy的dna,泳道6和7-暴露于50gy的dna。(b)-高和低分子量dna形式表示为总dna的百分比。针对每种条件,所有样品一式两份运行。数据表示为平均值±标准偏差。p<0.05被认为是有意义的。*分别显示与未处理形式相比的显著差异。
图4:增加浓度的合成sdg(s,s)、sdg(r,r)和商业sdg对γ辐射诱导的小牛胸腺dna片段化的影响。将所有样品暴露于50戈瑞的γ射线剂量。sdg浓度为25、50、100和250μm。在图(a)、(c)、和(e)中显示了在25、50、100和250μmsdg(s,s)、sdg(r,r)和sdg(商业)存在下暴露于50gy后的小牛胸腺dna的代表性琼脂糖凝胶扫描。泳道1-1kbdna标准梯,泳道2和3-未处理的dna,泳道4和5-25μm,泳道6和7-50μm,泳道8和9-100μm,泳道10和11-250μmsdg。在图(b)、(d)和(f)中高和低分子量dna形式表示为总dna的百分比。针对每种条件,所有样品一式两份运行。数据表示为平均值±标准偏差。p<0.05被认为是有意义的。*显示与未处理的dna相比的显著差异。#显示与暴露于50gy不含sdg的样品相比的显著差异。
图5:非常低浓度的合成sdg(s,s),sdg(r,r)和商业sdg对γ辐射诱导的小牛胸腺dna片段化的影响。将所有样品暴露于50gy的γ辐射剂量。sdg浓度为0.5、1.0、5.0和10μm。在图(a)、(c)、和(e)中显示了在0.5、1.0、5.0和10μmsdg(s,s)、sdg(r,r)和sdg(商业)存在下暴露于50gy后的小牛胸腺dna的代表性琼脂糖凝胶扫描。泳道1-1kbdna标准梯,泳道2和3-未处理的dna,泳道4和5-0.5μm,泳道6和7-1.0μm,泳道8和9-5.0μm,泳道10和11-10μmsdg。在图(b)、(d)和(f)中高和低分子量dna形式表示为总dna的百分比。针对每种条件,所有样品一式两份运行。数据表示为平均值±标准偏差。p<0.05被认为是有意义的。*显示与未处理的dna相比的显著差异。#显示与暴露于50gy不含sdg的样品相比的显著差异。
图6:sdg、seco、ed和el对γ辐射诱导的小牛胸腺dna片段化的影响。将所有样品暴露于50gy的γ辐射剂量。以10μm浓度使用sdg、seco、ed和el。(a)显示在10μmsdg、seco、ed和el存在下暴露于50gy后的小牛胸腺dna的代表性琼脂糖凝胶扫描。泳道1-1kbdna标准梯,泳道2和3-未处理的dna,泳道4、5和6-ir50gy,泳道7和8-sdg,泳道9和10-seco,泳道11和12-ed,泳道13和14-el。(b)高和低分子量dna形式表示为总dna的百分比。针对每种条件,所有样品一式两份运行。数据表示为平均值±标准偏差。p<0.05被认为是有意义的。*显示与未处理的dna相比的显著差异。#显示与暴露于单独的50gy的样品相比的显著差异。
图7:sdg治疗对小鼠原代肺细胞的辐射剂量应答的影响。(a)上皮细胞;(b)内皮细胞和(c)wt成纤维细胞。在γ辐射(0、2、4、6、8gy)之前用不同浓度的sdg处理细胞6小时并孵育。在第12-14天计数所有可见菌落,并将存活部分针对对照值进行标准化。数据表示为平均值±sem。**p≤0.001*p≤0.01、#p≤0.05用于辐射的细胞与50μmsdg预处理的辐射细胞。
图8:使用碱性彗星测定评估肺细胞中辐射诱导的dna单链断裂(ssb)。(a)2-gyγ辐射的原代肺细胞(上皮,内皮和wt成纤维细胞)中dna损伤的动力学评估;对每次处理计数至少100-150个细胞。通过计算每个细胞的“尾力矩”(尾部dna量乘以尾部长度的乘积)来评估dna损伤。*p≤0.001针对非辐射的对照物与其相应的被辐射细胞;(b)sdg(50μm)处理(辐射前0、2、4、6小时)对辐射的原代肺细胞的影响。数据表示为平均值±sem。*p≤0.001、#p≤0.01用于辐射的细胞与sdg预处理的辐射细胞。
插入物:原代肺上皮细胞的代表性荧光显微照片。将细胞用sdg(50μm)预处理并暴露于g-辐射(2gy),包埋在琼脂糖中,裂解,并且进行电泳(0.66v/cm,25min),用sybr绿进行染色并在荧光显微镜下进行可视化并且此后检查彗尾形成。(a)对照细胞,(b)sdg(50μm),(c)暴露30分钟后的ir(2gy),(d)用sdg(50μm,6h)预处理并且辐射的细胞。
图9:荧光评估γ-h2ax病灶在辐射的鼠原代肺细胞(即,上皮细胞,内皮细胞和wt成纤维细胞)中的诱导。将细胞用sdg(50μm)处理6小时,并进行γ-辐射(2gy)。以所需的时间间隔,将细胞在4%多聚甲醛中固定,洗涤,用γ-h2ax抗体探测,并使用dapi对细胞核进行复染。在荧光显微镜下观察细胞。对每个视野计数总细胞(蓝色)γ-h2ax阳性细胞(绿色),并计算γ-h2ax阳性细胞的百分比。对每次处理计数至少500个细胞,并进行两次实验。数据表示为平均值±sem。*p≤0.05,**p≤0.005针对辐射的细胞与sdg预处理的辐射细胞。
图10:小鼠肺肺上皮细胞中γ-h2ax病灶(绿色)的免疫荧光可视化的代表性组。将细胞用sdg预处理6h,进行γ-辐射(2gy)并再孵育30min。将细胞在4%多聚甲醛中固定,并用γ-h2ax抗体探测。用dapi(蓝色)对dna进行复染色。使用荧光显微镜获取图像。
图11:在被辐射的鼠原代肺细胞中γ-h2ax病灶的诱导的流式细胞术(facs)确认。将原发性肺细胞(即,上皮细胞(a),内皮细胞(b)和wt成纤维细胞(c))用sdg(50μm)处理6h并且进行γ辐射(2gy)。以所需的时间间隔,处理细胞用于facs分析。使用summit软件定量数据并表示为平均值±sem。*p≤0.05,**p≤0.01针对辐射的细胞与sdg预处理的辐射细胞。
图12:原发性肺细胞中辐射诱导的细胞凋亡死亡的评估。sdg(50mm)预处理(6h)对辐射的原发性肺细胞(即,上皮细胞(a),内皮细胞(b)和wt成纤维细胞(c))的影响的定量评估。将细胞固定,用dapi染色,并在荧光显微镜下观察用于形态学分析。对于每次处理,从5个不同的视野计数至少500个细胞,并计算凋亡细胞的百分比。进行两次实验。数据表示为平均值±sem。*p≤0.05,**p≤0.005针对辐射的细胞与sdg预处理的辐射细胞。
图13:评估sdg治疗对肺上皮细胞凋亡调节剂的影响。在2gy暴露之前,用sdg(50μm)处理鼠原代肺细胞(上皮细胞)6小时。在辐射后6、24和48小时收获细胞。以期望的时间间隔从上皮细胞分离总rna,并通过针对bax和bcl-2基因表达(a和b)的定量实时rt-pcr分析进行评估。一式三份进行分析,并将基因表达归一化为18s核糖体rna。通过蛋白质印迹分析评估bax和bcl-2蛋白水平;代表性图像(c)和密度测定法分析,其中归一化为β-肌动蛋白。(d和e)。数据表示为平均值±sem。*p≤0.05,**p≤0.01针对与sdg处理的细胞或sdg+ir处理的细胞相比的ir暴露的细胞。
图14:sdg对放射诱导的活性半胱氨酸蛋白酶-3和切割的parp水平的增加的作用。在2gy暴露之前,用sdg(50μm)处理鼠原代肺细胞(上皮细胞)6小时。在辐射后6、24和48小时收获细胞。通过蛋白质印迹分析评估切割的半胱氨酸蛋白酶-3和切割的parp蛋白水平。(a)代表图像和(b和c)密度测定法分析,其中归一化为β-肌动蛋白。一式两份进行分析,并且数据表示为平均值±sem。*p≤0.05,**p≤0.01针对与sdg处理的细胞相比的ir暴露的细胞。
图15:sdg清除次氯酸离子。图15a显示apf和hpf荧光的clo-依赖性增加。图15b显示了由sdg清除clo-。图15c显示了合成的sdg非对映异构体sdg(s,s)和sdg(r,r)的清除效果。所有样品重复运行。数据表示为平均值±标准误差。p<0.05被认为是有意义的。*显示与未处理的对照相比的显著差异。
图16:sdg清除γ-辐射诱导的次氯酸盐的生成。图16a和b显示γ-辐射诱导apf和hpf荧光的增加。图16c和d显示了在具有apf或hpf的磷酸盐缓冲盐水(pbs)中增加剂量的辐射下sdg对次氯酸盐的产生的影响(参见图15图注)。图16e、f和g显示γ-辐射诱导的牛磺酸的氯化。图16e显示了牛磺酸的次氯酸盐依赖性氯化。图16f显示牛磺酸氯胺作为所有实验条件的吸光度。图16g显示了在如图16f中所示的各种条件下的次氯酸盐浓度。对于图16a-e,所有样品都重复运行,而对于图16f和g,所有样品一式四份运行。数据表示为平均值±标准误差。p<0.05被认为是有意义的。*显示与未处理的对照相比的显著差异。
图17:次氯酸盐诱导的小牛胸腺dna损伤。图17a和c显示了暴露于hocl后小牛胸腺dna的代表性琼脂糖凝胶扫描。图17b和d显示高和低分子量dna片段为总dna的百分比。图17e和f显示了sdg对质粒dna的次氯酸盐诱导的损伤的影响。图17e显示了暴露于hocl后质粒dna的代表性琼脂糖凝胶。图17f显示sc和oc形式为总质粒dna的百分比。针对图17a,泳道1-1kbdna标准梯,泳道2和3-未处理的dna,泳道4和5-0.1mm,泳道6和7-0.2mm,泳道8和9-0.4mm,泳道10和11-0.5mm以及泳道12和13-0.6mmclo-。针对图17c,泳道1-1kbdna标准梯,泳道2和3-未处理的dna,泳道4和0.5mmhocl,泳道6和7-0.5mmhocl+sdg(com)1μm,泳道8和9-0.5mmhocl+sdg(s,s)1μm,泳道10-11-0.5mmhocl+sdg(r,r)1μm,泳道12-13-0.5mmhocl+槲皮素1μm,以及泳道14和15-0.5mmhocl+水飞蓟宾1μm。对于图17e,泳道1-1kbdna标准梯,泳道2和3-未处理的质粒dna,泳道4和4.5mmhocl,泳道6和7-4.5mmhocl+sdg25μm。针对每种条件,所有样品一式两份运行。数据表示为平均值±标准误差。p<0.05被认为是有意义的。*和#显示与未处理的dna相比的显著差异。
图18:sdg(前处理和后处理)对2-氨基嘌呤(2-ap)的次氯酸盐诱导的修饰的影响。图18a显示所有条件的代表性光谱。图18b显示在如图18a中所示的不同条件下在374nm下的荧光。图18c显示了sdg的保护%。针对每种条件,所有样品一式两份运行。数据表示为平均值±标准误差。p<0.05被认为是有意义的。*和#分别显示与未处理的2-ap对照和处理的相比的显著差异。
图19:sdg防止2-氨基嘌呤(2-ap)的γ-辐射诱导的修饰。图19a显示所有条件的代表性光谱。图19b显示在374nm下的荧光。针对每种条件,所有样品一式两份运行。数据表示为平均值±标准误差。p<0.05被认为是有意义的。*和#分别显示与未处理的2-ap对照和处理的相比的显著差异。
图20:sdg作用在dna保护免于核碱基氯化中建议机制。
图21:通过亚麻籽及其木脂素的化学预防机制。sdg通过抑制多步致癌作用过程减轻了由烟草和其他环境致癌物造成的肺部肿瘤发生。我们提供证据表明,在两个动物模型中,木脂素sdg具有通过调节nrf2调节的阶段ii解毒通路和可能其他机制的化学预防活性。我们进一步提供数据支持sdg的保护作用由直接ros清除和/或间接抗氧化/抗炎性能以及降低致癌物毒性和dna损伤来介导。
图22:sdg减少由苯并-α-芘在细胞中诱导的氧化性dna损伤。将sdg(10μm)添加至暴露于25μm烟草和环境致癌物苯并-α-芘(bap)的人上皮细胞(a549)中,并使用质谱法检测对dna的氧化损伤,如8-氧代-7,8-二氢鸟嘌呤(8-氧代-dguo)的存在指示的。sdg在致癌物暴露后3和6小时减少dna损伤。
图23:在细胞中ros诱导的致癌物苯并-α-芘。鼠上皮细胞暴露于bap诱导破坏性活性氧簇(ros),如通过氧化还原敏感性荧光染料检测的。早在暴露于致癌物后2小时,荧光强度的强烈增加表明细胞中ros的产生。
图24:sdg防止ros产生免受致癌物暴露。将小鼠上皮细胞暴露于10或20μmbap和增加浓度的sdg(0、0.1、0.5、1、5μmsdg),并在2小时后检测ros(如图23中适当确定的)。sdg清除有害ros至可忽略的水平。
图25:sdg防止暴露于bap的人上皮细胞中的基因毒性应激。将细胞暴露于强致癌物如bap,诱导基因毒性应激,如通过增加的p53蛋白水平所指示的。这通过在5、10、25和50μm浓度下sdg的存在而减轻剂量依赖性。
图26:sdg防止暴露于bap的人上皮细胞中的氧化性dna损伤。将细胞暴露于强致癌物如bap,诱导氧化性dna损伤,如通过增加的γ-h2ax(双链dna断裂的标志物)的水平所指示的。这通过在5、10、25、50和100μm浓度下的存在而减轻剂量依赖性。
图27:sdg防止暴露于bap的人上皮细胞中的dna加合物形成。将细胞暴露于强致癌物,如bap,诱导dna加合物的形成。dna加合物是与致癌物共价连接的dna片段,并与恶性肿瘤的发展直接相关。dna加合物水平通过单独或组合给予sdg或其代谢物ed和el的存在而降低。
图28:化学致癌物诱导的肺肿瘤的小鼠模型。给小鼠(a/j菌株)以1mg/kg剂量经腹膜内注射4次烟草和环境致癌物bap(每周一次)。小鼠在暴露时用亚麻籽或木脂素饮食开始。在暴露后在不同时间评估小鼠,以确定肿瘤负荷,小鼠重量和总体健康状况。
图29:亚麻籽降低小鼠中的肿瘤负荷:暴露于致癌物的鼠肺的总病理学状况。bap暴露和饮食亚麻籽给予后若干个月,鼠肺的代表性临床图像。
图30:亚麻籽降低小鼠中的肿瘤负荷:暴露于致癌物的鼠肺的组织病理学状况。bap暴露和饮食亚麻籽给予后若干个月,鼠肺的代表性h&e应变肺切片。来自喂食对照饮食(上图)或亚麻籽(下图)的小鼠的箭头指示的结节在亚麻籽喂养的小鼠中显示更小。每个小图代表不同的动物。
图31:亚麻籽降低小鼠中的肿瘤负荷:肿瘤负荷的定量评估。使用针对整体肿瘤面积(a)和结节大小(b)的图像分析软件进行形态学评估组织学鼠肺切片。在喂养亚麻籽饮食的小鼠中,肿瘤占据的肺面积显著减少(p<0.03)。类似地,存在更小肿瘤结节尺寸的趋势。
图32:亚麻籽降低小鼠中的肿瘤负荷:肿瘤负荷的定量评估。使用针对每个肺的肿瘤结节的总数(a)和侵入肺的肿瘤%(b)的图像分析软件进行形态学评估组织学鼠肺切片。每个肺存在更少的肿瘤结节的趋势(a)并且在亚麻籽补充剂下更少的肿瘤侵入的趋势(b)
图33:亚麻籽补充剂防止防止来自由bap诱导的肺癌的耗散效应。在bap暴露后纵向测量动物重量200天。饲喂亚麻籽饮食的小鼠暴露于bap下,显示比当对照饮食时暴露于bap的小鼠显示更高的重量。
图34:实例5的实验方案。
图35:在每天摄取亚麻籽(fs)和亚麻籽木质素组分(flc)补充剂的饮食后4天,在血液中可检测到哺乳动物木脂素代谢物。具体地,可以使用液相色谱,串联质谱(lc/ms/ms)检测肠二醇(ed)和肠内酯(el)。饮食被设计为递送可比较的木脂素水平,反映在2种饮食中可检测的木脂素代谢物水平。
图36:在石棉暴露之前给予的亚麻籽(fs)和亚麻籽木质素组分(flc),由经腹膜内青石棉石棉注射诱导的钝性腹部炎症,如通过巨噬细胞(mf)、嗜中性粒细胞(pmn)和淋巴细胞(ly)的数量所证实的。具体来说,fs和flc显著减少腹部中的巨噬细胞流入。*p<0.05
图37:小鼠在fs和flc饮食上开始并在24小时后暴露于石棉。根据图34中的实验方案,暴露后3天,使用elisa确定血浆(b,d)和腹部(a,c)中的细胞因子水平(tnfα和il-1β)。这两种饮食表明防止腹部中的促炎细胞因子的分泌和由石棉暴露诱导的全身循环的趋势。
图38:在饮食(a)的情况下,炎性细胞也趋于更低,同时由400和800mg青石棉石棉诱导的tnfα(b)和il-1β(c)细胞因子水平由石棉暴露后1天饮食中添加的flc显著钝化(*p<0.05)。*p表示与暴露于石棉的对照饮食相比的显著性。
图39:在小鼠中口服灌胃可变sdg浓聚物后sdg和代谢物的血浆浓度。
图40:在小鼠中口服灌胃可变剂量的sdg后肺中的抗氧化剂酶基因表达水平。
图41:在小鼠口服灌胃100mg/kgsdg后,血浆(a)和肺组织(b)中sdg水平的动力学,和aoe基因表达的相应水平的(c)。
图42a-b:实例7的实验方案。
图43:实例7的临床研究设计。
图44:蛋白质印迹显示,喂食10%fs增加小鼠鼻上皮中的ho-1和nqo-1。
图45:40gfs饮食后人颊上皮细胞中ho-1基因表达的动力学。(*p<0.05,从0天)。
图46:在一个fs的患者中尿液isop水平的动力学。
图47:fs的正常和肺移植患者中尿8-氧代-dguo水平的动力学。
图48:假设sdg或亚麻籽饮食减少石棉诱导的ros/炎症。
图49:细胞的石棉暴露以检测炎性细胞因子分泌和亚硝化/氧化应激的实验计划(示意图):
图50:sdg在体外通过人类间皮细胞钝化石棉诱导的ros分泌。
图51:培养物raw巨噬细胞中石棉诱导的氧化应激(ros释放)的评估:石棉诱导的ros在石棉暴露后不久产生,并且在观察期间持续。
图52:在暴露于石棉后几小时向巨噬细胞给予sdg降低氧化应激。
图53:在暴露于石棉后几小时向巨噬细胞给予sdg降低亚硝化应激。
图54:在暴露于石棉后几小时向巨噬细胞给予sdg减少炎症细胞因子分泌(il-1β)。
图55:在暴露于石棉后几小时向巨噬细胞给予sdg减少炎症细胞因子分泌(tnf-α)。
图56:使用两种小鼠模型在石棉诱导的间皮瘤中测试sdg:使用在石棉暴露后遗传倾向形成间皮瘤的小鼠的至少2个模型,我们将:评估亚麻籽和sdg对小鼠中单剂量石棉的急性效应;测试亚麻籽和sdg是否抑制肿瘤在加速,石棉诱导mm的遗传模型中的发展。
图57:向暴露于石棉的mextag小鼠给予富含sdg(35%sdg)的flc饮食减少炎症。
图58:nf2小鼠的石棉暴露实验计划和亚麻籽/sdg木脂素配制品评估:
图59:在石棉暴露后nf2小鼠中腹部炎症的动力学:炎症细胞流入3天达到峰值并在石棉暴露后9天逐渐减少。因此,选择3天作为在所有后续实验中评估炎症的时间点。
图60:亚麻籽及其富含sdg的木脂素组分钝化的石棉诱导的炎症(年轻小鼠):在饮食中添加fs或flc时,总白细胞(a)减少,虽然不显著。然而,当观察细胞差异,并且特别是巨噬细胞水平时,水平显著通过亚麻籽和sdg-木脂素饮食(b)减弱。
图61:亚麻籽及其富含sdg的木脂素组分钝化的石棉诱导的炎症(老龄小鼠):与仅300,000细胞/ml(高出10倍)相比,通过呈现约3,000,000wbc/ml的腹部灌洗液,暴露于腹部石棉的老龄小鼠(a)对石棉更敏感。结果表明,炎症细胞嗜中性粒细胞(b)和巨噬细胞(c)在老龄小鼠中二者都显著高于年轻小鼠。
图62:富含sdg(在饮食配制品中给出)的亚麻籽木脂素提取物使老年小鼠中的石棉炎症钝化:在第0天向雄性nf2(129sv)(+/-)小鼠注射(腹膜内)400μg石棉。小鼠在石棉暴露前一周(第-7天)在测试饮食(0%fs或10%flc)上开始,并在石棉暴露后第3天处死。用5ml1×pbs进行腹部灌洗(al)(将1ml腹部灌洗液离心,并且冷冻上清液)。在-80°下冷冻收集血浆。在灌洗液中评估细胞,并且显示通过富含sdg的饮食,总wbc和嗜中性粒细胞、巨噬细胞和嗜酸性粒细胞都显著减少。
图63:富含sdg(在饮食配制品中给出)的亚麻籽木脂素提取物使老年小鼠中的石棉炎性细胞因子分泌和亚硝化应激钝化:在第0天向雄性nf2(129sv)(+/-)小鼠注射(腹膜内)400μg石棉。小鼠在石棉暴露前一周(第7天)在测试饮食(0%fs或10%flc)上开始,并在石棉暴露后第3天处死。用5ml1×pbs进行腹部灌洗(al)(将1ml腹部灌洗液离心,并且冷冻上清液)。在-80°下冷冻收集血浆。细胞因子il1β和tnfα以及亚硝酸盐的水平被富含sdg的饮食显著钝化。
发明详细说明
在以下详细说明中,陈述了众多具体细节,以便提供对本发明的彻底理解。然而,本领域的技术人员将理解的是,可以在没有这些具体细节的情况下实施本发明。在其他情况下,没有详细说明的熟知方法、程序和组分,以便不使本发明难于理解。
本文提供了使用亚麻籽、其生物活性成分或其代谢物用于辐射防护和化学预防的治疗和预防方法。在示例性实施例中,生物活性成分包括开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其异构体(包括立体异构体)或其组合。
本申请的发明人已经发现亚麻籽、其生物活性成分和/或其降解物或代谢物有效地保护生物分子、细胞和组织免受辐射损伤、次氯酸离子诱导的损伤、致癌物诱导的损伤和恶性肿瘤。因此,诸位本发明人已经发现亚麻籽、其生物活性成分、或其代谢物可用于保护生物分子、细胞和组织免受辐射损伤,次氯酸离子诱导的损伤、致癌物损伤和癌症发展。
根据本文提供的方法需要辐射防护或放射减轻的受试者是将要或已经暴露于潜在有害量的辐射的受试者。应当理解,这种暴露可以是单次暴露、周期性暴露、零星暴露或持续暴露于辐射。还应当理解,这种辐射暴露包括意外暴露、偶然或有意暴露。
根据本发明的方法可能需要辐射防护或放射减轻的受试者的实例包括但不限于:暴露于作为治疗方案的一部分的辐射的患者(例如,需要放射疗法的癌症患者),暴露于辐射以诊断疾病或病症的受试者(例如,接受牙齿的或骨的x射线的受试者,接受pet扫描、ct扫描等的患者)。根据本发明的方法可能需要辐射防护或辐射减轻的受试者的实例还包括:那些由于其职业或生活方式选择暴露于辐射的那些(例如,飞机机组或其他频繁的航空旅行者,以及甚至空间旅行者,他们暴露于高于平均辐射水平;实验室技术人员和其他工人;或通过使用电子设备所暴露的那些)或者暴露于氡的累积(例如,在住处或矿场中的累积)的那些或暴露于来自太阳的自然辐射的户外工人或日光浴者。根据本发明的方法可能需要的辐射防护的其他受试者包括意外暴露于辐射的那些受试者,如泄漏或溢出(例如,核反应堆泄漏或事故或实验室溢出)。还考虑到由于战争或恐怖主义导致核弹头爆炸而暴露于辐射的那些。涵盖的其他受试者是暴露于分散放射性物质的常规爆炸物的恐怖分子的爆炸的那些。
根据本文提供的方法需要化学预防的受试者是将会,或已经暴露于潜在有害量的致癌物或其他毒物的受试者。应当理解,这种暴露可以是单次暴露、周期性暴露,零星暴露或持续暴露于若干种合成或天然发生的致癌物或其他毒物(如,化学战剂)中的一种或组合。还应当理解,这种暴露包括意外暴露、偶然或有意暴露。还应当理解,这种暴露可以是直接暴露或间接暴露。例如,间接暴露于次氯酸离子可以是直接暴露于电离辐射的结果。
根据本发明的方法可能需要化学预防的受试者的实例包括但不限于:由于其职业或生活方式选择而可能暴露于致癌物或其他毒物的那些(例如,石油工业的工人;暴露于机动车废气颗粒的收费站值班人员;实验室技术人员和其他工人)。根据本发明的方法可能需要化学预防的其他受试者包括意外暴露于致癌物的那些,如饮用水或空气中的致癌物(石棉,多环芳烃)的泄漏或溢出。还预期由于习惯(吸烟者)而暴露于致癌物的那些。涵盖的其他受试者是暴露于恐怖分子的分散致癌物和其他癌症促进材料(如化学战剂)行为的那些。
在一方面,本发明涉及用于在对其有需要的受试者中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免受辐射损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、降解物或其代谢物。向所述受试者给予涵盖在暴露于损伤性辐射暴露之前,期间和之后的给予。时间之前,期间和之后可以是秒、分钟、小时、天、周、月或甚至年。生物活性成分涵盖开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于治疗或预防已经或将要暴露于辐射的受试者中的辐射损伤的方法,该方法包括:向所述受试者给予有效量的至少一种生物活性成分,其中所述生物活性成分包括开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于在对其有需要的受试者中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免受意外辐射暴露所导致的辐射损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、降解物或其代谢物。
在另一方面,本发明涉及用于保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞或组织免受导致衰老的辐射损伤的方法。
在另一方面,本发明涉及用于保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞或组织免受由暴露于天然和合成的化学致癌物质和毒物(包括化学战剂)产生的损害的方法。
在另一方面,本发明涉及用于在对其有需要的受试者中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免受针对癌症治疗的放射疗法所导致的辐射损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、降解物或其代谢物。
在另一方面,本发明涉及用于在对其有需要的受试者中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免受辐射损伤的方法,该方法包括:向所述受试者给予有效量的至少一种生物活性成分,其中所述生物活性成分包括开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。在一些实施例中,辐射损伤由意外的辐射暴露引起。在一些实施例中,辐射损伤由用于癌症(例如,肺癌)治疗的放射疗法引起。
在另一方面,本发明涉及用于在对其有需要的受试者中预防对生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织的辐射诱导的损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、或其代谢物。
在另一方面,本发明涉及用于在对其有需要的受试者中预防对生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织的辐射诱导的损伤的方法,该方法包括:向所述受试者给予有效量的至少一种生物活性成分,其中所述生物活性成分包括开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于在细胞中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免受辐射损伤的方法,该方法包括:将所述细胞与有效量的至少一种生物活性成分接触,其中所述生物活性成分包括开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于在对其有需要的受试者中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免受致癌物损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、降解物或其代谢物。向所述受试者给予涵盖在暴露于天然和合成的化学致癌物和毒物的损伤性暴露之前,期间和之后的给予。时间之前,期间和之后可以是秒、分钟、小时、天、周、月或甚至年。生物活性成分涵盖开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于在对其有需要的受试者中保护生物分子免受由于意外暴露于天然和合成的化学致癌物和毒物引起的致癌物损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、降解物或其代谢物。
在另一方面,本发明涉及用于保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞或组织免受来自天然和合成的导致肺癌或间皮瘤的化学致癌物和毒物的损害的方法。
在另一方面,本发明涉及用于保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞或组织免于致癌物损伤的方法,该方法包括:将所述生物分子、细胞或组织与有效量的生物活性成分接触。与所述生物分子、细胞或组织接触涵盖在暴露于天然和合成化学致癌物和毒物的损伤性暴露之前,期间和之后的接触。时间之前,期间和之后可以是秒、分钟、小时、天、周、月或甚至年。生物活性成分涵盖开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于在已经或将要暴露于一种或多种致癌物的受试者中治疗或预防致癌物诱导的损伤,恶性转化或癌症发展避免致癌物诱导癌症的方法,该方法包括:向所述受试者给予有效量的至少一种生物活性成分,其中所述生物活性成分包括开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及保护暴露于一种或多种致癌物的受试者免受致癌物诱导的癌症的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分或其代谢物。
在另一方面,本发明涉及用于在对其有需要的受试者中保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞、或组织免由次氯酸根离子造成的损伤的方法,该方法包括:向所述受试者给予有效量的亚麻籽、其生物活性成分、降解物或其代谢物。向所述受试者给予涵盖在暴露于天然和合成的化学致癌物和毒物的损伤性暴露之前,期间和之后的给予。时间之前,期间和之后可以是秒、分钟、小时、天、周、月或甚至年。生物活性成分涵盖开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于治疗或预防已经或将要暴露于次氯酸根离子的受试者中的次氯酸根离子诱导的损伤的方法,该方法包括:向所述受试者给予有效量的至少一种生物活性成分,其中所述生物活性成分包括开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于保护生物分子(如,遗传物质如核酸、蛋白质或脂质)、细胞或组织免于由次氯酸根离子造成的损伤的方法,该方法包括:将暴露于或将暴露于次氯酸根离子的所述生物分子、细胞或组织与有效量的生物活性成分接触。与所述生物分子、细胞或组织接触涵盖在暴露于天然和合成化学致癌物和毒物的损伤性暴露之前,期间和之后的接触。时间之前,期间和之后可以是秒、分钟、小时、天、周、月或甚至年。生物活性成分涵盖开环异落叶松脂素二葡糖苷(sdg)、开环异落叶松脂素(seco)、肠二醇(ed)、肠内酯(el)、其类似物、其立体异构体或其组合。
在另一方面,本发明涉及用于前述方法之一的组合物。
亚麻籽、其生物活性成分及其代谢物是本领域已知的,并描述于美国专利公开号2010/0239696;2011/0300247;和2014/0308379;和在国际专利公开号wo2014/200964中,其各自通过引用以其整体并入本文。
在亚麻籽中发现的主要木脂素是2,3-双(3-甲氧基-4-羟基苄基)丁烷-1,4-二醇(开环异落叶松脂素或seco),其作为共轭物开环异落叶松脂素二葡糖苷(sdg)以其天然状态存储在植物中。sdg在人肠中代谢为肠二醇(ed)和肠内酯(el)。肠醇和肠内酯的合成类似物是已知的(参见,例如,埃克隆(eklund)等人,有机通讯(org.lett.),2003,5:491)。
“降解物”是分子(如,sdg)分解成更小分子的产物。
“代谢物”是通过代谢或通过代谢过程产生的物质。例如,sdg的代谢物是el或ed。
本领域技术人员应当理解,代谢物可以是天然代谢物的化学合成的等价物。
“类似物”是其结构与另一种化合物的结构相关的化合物。类似物可以是合成类似物。
“成分”或“组分”是混合物或化合物中的元素或组分。
“产物”是由化学反应产生的物质。
“提取物”是包含材料(例如,来自亚麻籽)的活性成分或浓缩精华的制品。
“药物组合物”是指有效量的活性成分,例如,(s,s)-sdg(r,r)-sdg、内消旋-sdg、sdg、seco、el、ed及其类似物,与可接受的载体或稀释剂一起。“治疗有效量”是指对给定条件和给予方案提供治疗效果的量。
本文所述的组合物可以包括“治疗有效量”。“治疗有效量”是指在必需的剂量和时间段内有效达到所希望治疗结果的量。治疗有效量可以根据因素(如个体的疾病状态、年龄、性别和重量)以及组合物在个体中引起所希望应答的能力等因素而变化。治疗有效量也是治疗有益效果超过分子的毒性或有害作用的量。
如在此所用,短语“药学上可接受”是指在合理医学判断的范围内适用于与人类和动物的组织接触而无过度毒性、刺激、过敏应答或其他问题或并发症,与合理益处/风险比相称的那些化合物、材料、组合物、载体和/或剂型。
“药学上可接受的赋形剂”意指在制备药物组合物中有用的一种赋形剂,该组合物通常是安全、无毒的并且不是生物学上或其他方面不希望的,并且包括对于兽用连同人类药用是可接受的一种赋形剂。如本文所用的“药学上可接受的赋形剂”包括一种和多于一种这样的赋形剂。
药物组合物可以通过本领域技术人员已知的任何合适的方法,如口服、经肠胃外、经粘膜、经皮、经肌内、经静脉内、经皮内、经皮下、经膜内、经颅内、经阴道内、经肿瘤内或经颊向受试者给予。也可以通过将活性成分包埋在合适的聚合物中来使用控释,然后可以将其经皮下、经肿瘤内、经颊、作为贴剂在皮肤上、或经阴道内插入。该包括用活性成分涂覆医疗装置。
在一些实施例中,药物组合物经口服给予,并因此配制成适于口服给予的形式,即作为固体或液体配制品。合适的固体口服配制品包括片剂、胶囊剂、丸剂、颗粒剂、小丸剂等。合适的液体口服配制品包括溶液,悬浮液,分散液,乳液,油等。在一些实施例中,将活性成分配制到胶囊剂中。根据该实施例,除了活性化合物和惰性载体或稀释剂外,除了其他赋形剂以及明胶胶囊外,本发明的组合物包括干燥剂。
在一些实施例中,通过经静脉内,经动脉内或经肌内注射液体配制品来给予药物组合物。在一些实施例中,药物组合物是配制用于口服给予的液体制品。在一些实施例中,药物组合物是配制用于阴道内给予的液体制品。合适的液体配制品包括溶液,悬浮液,分散液,乳液,油等。在一些实施例中,药物组合物经静脉内给予,并因此配制成适于静脉内给予的形式。在另一个实施例中,该药物组合物经动脉内给予,并因此被配制为适于动脉内给予的形式。在一些实施例中,药物组合物经肌内给予并因此配制成适于肌内给予的形式。在一些实施例中,药物组合物经颊内给予并因此配制成适于经颊给予的形式。
在一些实施例中,药物组合物局部给予于体表,并且因此配制成适合局部给予的形式。合适的局部配制品包括凝胶、软膏、乳膏、洗剂、滴剂、控释聚合物等。对于局部给予,制备亚麻籽、其生物活性成分或其代谢物,并将其作为溶液,悬浮液或乳液在具有或不具有药物载体的生理学上可接受的稀释剂中施用。
在一些实施例中,本文提供的药物组合物是控释组合物,即亚麻籽、其生物活性成分或其代谢物在给予后一段时间内释放的组合物。控释或缓释组合物包括在亲脂性储库(例如,脂肪酸、蜡、油)中的配制品。在其他实施例中,组合物是立即释放组合物,即所有亚麻籽、其生物活性成分或其代谢物在给予后立即释放的组合物。
在一些实施例中,用于本文提供的方法的组合物以治疗剂量每天一次进行给予。在一些实施例中,组合物每两天一次,每周两次,每周一次,或每两周一次进行给予。
用于提取和纯化sdg的技术是本领域已知的并且描述于美国专利5,705,618中,其通过引用并入本文。用于合成sdg、其立体异构体和类似物的技术描述于米什拉(mishra)op等人,生物有机化学与医药化学通讯(bioorganic&medicinalchemistryletters)2013,(19):5325-5328和国际专利公开号wo2014/200964中,其内容以其整体通过引用在此结合。如本领域已知的,用于本文提供的方法中的生物活性组分还可以直接化学合成为哺乳动物的易于代谢的形式,肠二醇(ed)或肠内酯(el)。
(s,s)-sdg(r,r)-sdg、(s,r)-sdg(r,s)-sdg、内消旋-sdg、seco、el、ed或其类似物可以0.1ng/kg至500mg/kg的剂量给予。
用(s,s)-sdg(r,r)-sdg、(s,r)-sdg(r,s)-sdg、内消旋-sdg、sdg、seco、el、ed或其类似物的治疗是单次给予至若干天、若干个月、若干年或无限期。
如本文所用,“治疗”可以指治疗性治疗或防预性或预防性措施,其中目的是预防或减轻本文所述的靶向病理病症或障碍,或两者。因此,用于本文提供的方法的组合物可以在暴露于,例如,辐射、致癌物、毒物或次氯酸离子之前给予于受试者。在一些情况下,用于本文提供的方法的组合物可以在暴露后给予给受试者。因此,治疗本文所述的病症可以指预防,抑制或遏制受试者的病症。
此外,如本文所使用的,术语“治疗(treat或treatment)”是指治疗性治疗以及防预性或预防性措施,其中目的是预防或减缓(减轻)与疾病或病症相关的不期望的生理变化。有益的或期望的临床结果包括但不限于:症状的缓解,疾病或病症的程度的减轻,疾病或病症的稳定(即,疾病或病症不恶化的情况),延迟或减缓疾病或病症的进展,疾病或病症的改善或缓解,以及疾病或病症的缓解(无论部分或全部),无论是可检测的还是不可检测的。“治疗”还可以指相较于未接受治疗时的预期存活期,延长存活期。需要治疗的那些包括已经暴露于,例如,辐射、致癌物、毒物或次氯酸离子的那些,以及倾向于暴露的那些或预期暴露的那些。
在一些实施例中,需要治疗的受试者和本文所述的方法和组合物可包括但不限于:患有肺部疾病和障碍(如哮喘、癌症、copd和间皮瘤)的受试者。在一些实施例中,合适的受试者可以包括患有与衰老相关的疾病和病症的受试者,如心血管障碍和病症,皮肤下垂和中枢神经系统(cns)疾病(例如,阿尔茨海默尔痴呆)。在一些实施例中,合适的受试者可以包括皮肤障碍和病症(例如,牛皮癣),以及患有化妆品皮肤病症(例如,皱纹和老年斑)的受试者。在一些实施例中,合适的受试者可以包括患有胃肠道障碍和病症(如ibd和慢性病)的受试者。在一些实施例中,合适的受试者可以包括患有心血管障碍和病症的受试者。在一些实施例中,合适的受试者可以包括患有黑色素瘤的受试者。在一些实施例中,合适的受试者可以包括患有眼部疾病(如黄斑变性)的受试者。在一些实施例中,合适的受试者可以包括患有癌症(如乳腺癌、前列腺癌和子宫癌)的受试者。在一些实施例中,合适的受试者包括患有认知缺损和其他认知障碍的受试者。
术语“受试者”包括哺乳动物,例如,人、伴侣动物(例如,狗、猫、鸟等)、农场动物(例如,牛、绵羊、猪、马、家禽等)、以及实验室动物(例如,大鼠、小鼠、豚鼠、鸟等)。除了人以外,受试者可以包括狗、猫、猪、牛、绵羊、山羊、马、水牛、鸵鸟、豚鼠、大鼠、小鼠、鸟类(例如,长尾小鹦鹉)和其他野生,驯养或商业上有用的动物(例如,鸡,鹅,火鸡,鱼)。术语“受试者”不排除在所有方面都是正常的个体。术语“受试者”包括但不限于需要疗法或易感于病症或其后遗症的人。
术语“约”或“大约”意指由本领域普通技术人员确定的特定值的可接受误差范围内,其将部分地取决于如何测量或确定该值,即,测量系统的局限性。
提出以下实例,以便更充分地说明本发明的优选实施例。然而,它们绝不应被解释为限制本发明的广泛范围。
实例
实例1
合成的(s,s)和(r,r)-开环异落叶松脂素二葡糖苷(sdg)保护裸质粒和基因组dna免受γ辐射损伤
作为原子核蜕变的结果,产生三种类型的辐射,即具有正电荷的阿尔法(α),具有负电荷的贝它(β)和不带电荷的伽马(γ)。在γ-辐射的情况下,电磁波具有非常小的波长(<0.005nm),并且因此具有能够电离分子和原子的高能量。在生物系统或溶液中,电离辐射通过水辐射分解产生羟基自由基(.oh)。这些羟基自由基(.oh)是电离辐射诱导的对细胞组分(包括脂质,蛋白质和基因组dna)的损伤的主要来源。由γ辐射产生的羟基自由基(.oh)在dna中导致单链和双链断裂。(.oh)自由基通过从脱氧核糖和嘌呤以及嘧啶碱基提取h原子或添加到碱基的双键上来损伤dna。这些反应导致dna链断裂。
具有抗氧化和自由基清除性能的化合物可以作为辐射防护剂并且防止辐射诱导的dna损伤。由于从自然资源分离开环异落叶松脂素二葡糖苷(sdg)的复杂的提取、纯化和富集方法,该自然资源与高成本、变异性和难以产生大量sdg来使得临床前和床前测试可行相关,因此sdg是化学合成的。使用天然化合物香草醛和葡萄糖,成功地合成了sdg:sdg(s,s)和sdg(r,r)的两种对映异构体(它们的结构如下所示)(米什拉(mishra)等人,生物有机化学与医药化学通讯(bioorganic&medicinalchemistryletters)2013,(19):5325)。
除了其他以外,在克里斯托费杜-索罗米杜(christofidou-solomidou)等人的许多研究中已经显示sdg是有效的抗氧化剂和有效的自由基清除剂。重要的是,在最近的研究中,合成的sdg对映异构体已显示具有强的抗氧化和自由基清除特性(米什拉等人,生物有机化学与医药化学通讯(bioorganic&medicinalchemistryletters)2013,(19):5325-5328)。在本实例中,当合成的sdg对映异构体(s,s)-sdg和(r,r)-sdg与商业sdg相比时,调研并评估了其辐射防护性能。使用质粒dna松弛测定,通过在将质粒暴露于γ-辐射后确定sdg防止超级螺旋至开放螺旋质粒dna(pbr322)转化的能力,以及通过在将dna暴露于γ-辐射后评估基因组dna片段化的抑制,来评估三种化合物的辐射防护特征。sdg由肠道细菌代谢产生开环异落叶松脂素(seco)、肠二醇(ed)和肠内酯(el)。因此,还评估了sdg的这些代谢物对基因组dna的γ-辐射诱导的片段化的影响。
材料和方法
化学品
质粒dna(pbr322)、溴化乙锭、超纯10xtae缓冲液和1kbplusdna梯购自英杰公司(invitrogen)(生命技术公司(lifetechnologies),卡尔斯拜德,加利福利亚州)。琼脂糖(超纯)和小牛胸腺dna购自西格玛奥德里奇公司(sigma-aldrich)(圣路易斯,密苏里州)。开环异落叶松脂素二葡糖苷(sdg)(商业)、开环异落叶松脂素(seco)、肠二醇(ed)、和肠内酯(el)购自美国中草药公司(欧文,加利福尼亚州)。
开环异落叶松脂素二葡糖苷的合成(sdg)
如米什拉(mishra)等人,生物有机化学与医药化学通讯(bioorganic&medicinalchemistryletters)2013,(19):5325-5328中所述,来合成合成的sdg(r,r)和sdg(s,s)立体异构体。
质粒dna和小牛胸腺dna暴露于γ-辐射
将具有或不具有不同浓度的sdg(r,r)、sdg(s,s)和sdg(商业)的质粒dna(pbr322)或小牛胸腺dna样品用mark1铯(cs-137)幅射器(jl谢泼德(shepherd),圣费尔南多,加利福尼亚州)以1.7gy/min的剂量率在磷酸盐缓冲的生理盐水(ph7.4)(pbs)中暴露于γ-辐射。
辐射诱导的质粒dna松弛的确定
使用质粒dna(pbr322)(生命技术公司,卡尔斯拜德,加利福利亚州)确定测试化合物对辐射诱导的链断裂和超级螺旋(sc)至开放螺旋(oc)转化的影响。将pbs(ph7.4)中的质粒dna(500ng)与各种浓度(25-250μm)的sdg(r,r)、sdg(s,s)和sdg(商业)混合,并在pbs中在25gy下进行辐射。在辐射暴露后30min,将样品与负载染料混合,并在100v下在tae缓冲液(ph,8.3)中进行琼脂糖(1%)凝胶电泳。将凝胶用溴化乙锭(0.5μg/ml)染色40min,洗涤20min,并且然后在uv透射仪(伯乐公司,赫库斯,加利福尼亚州)上观察。扫描捕获的凝胶图像,并且通过gel-doc图像分析程序确定开放螺旋(oc)和超级螺旋(sc)质粒dna条带的密度。sc和oc质粒dna的密度表示为总密度(oc+sc)的%。
辐射诱导的dna片段化的确定
使用小牛胸腺dna(西格玛,圣路易斯,密苏里州)确定测试化合物对dna中辐射诱导的链断裂的影响。将pbs(ph7.4)中的dna(500ng)与各种浓度(25-250μm)的sdg(r,r)、sdg(s,s)和sdg(商业)混合,并在50gy下进行辐射持续30min。在从0.5-10μm范围内的不同浓度下进行第二系列实验。将样品与负载染料混合,并在100v下在tae缓冲液(ph,8.3)中进行琼脂糖(1%)凝胶电泳。将凝胶用溴化乙锭(0.5μg/ml)染色40min,洗涤20min,并且然后在uv透射仪(伯乐公司,赫库斯,加利福尼亚州)上观察。扫描捕获的凝胶图像,并且通过gel-pro图像分析程序(mediacybernetics,银泉,马里兰州)确定小牛胸腺dna片段的密度。小牛胸腺dna的低分子量(<6,000bp)和高分子量(>6,000bp)片段的密度表达为总密度的%(低分子量+高分子量)。
数据分析
获得的数据表示为平均值±标准偏差。使用邦弗朗尼校正用statview程序,使数据经受单向方差分析(anova)与事后比较。p值≤0.05被认为是有意义的。
结果
使用质粒dna(pbr322)确定合成sdg(r,r)、sdg(s,s)和sdg(商业)的辐射防护潜力。在本研究中使用的辐射防护测定基于以下原理:暴露于γ-辐射后的质粒dna比未暴露的质粒dna移动更慢。这仅仅是由于超级螺旋质粒dna由于其紧凑的大小而在琼脂糖凝胶中移动更快的事实。相比之下,质粒dna中辐射诱导的缺口解开超级螺旋,导致相对更大尺寸的质粒,其以更慢的速率在凝胶中移动。因此,与超级螺旋的质粒dna相比,确定开放螺旋的密度反映了辐射诱导的损伤的程度。
辐射导致剂量依赖性sc至ocdna质粒转化
为了选择引起显著dna损伤但允许测试辐射减轻剂的治疗窗的辐射剂量,将质粒dna暴露于10、25和50gy的γ辐射。图1a中呈现的结果显示,存在质粒dna的oc形式的辐射剂量依赖性增加以及质粒dna的sc形式的辐射剂量依赖性降低。sc和oc的分布(图1b)显示sc的%分别在0、10、25和50gy下从68.73%±2.54%降低至50.91%±2.31%、38.37%±3.73%和35.66%±4.24%(p<0.05)。同时,oc的的%分别在0、10、25和50gy下从31.26%±2.50%增加至49.08%±2.31%、61.62%±3.73%和67.33%±4.24%(p<0.05)。基于这些初始实验,选择25gy的辐射剂量(在该辐射剂量下实现相当大且清楚可见的损害)用于随后的实验以确定不同sdg的辐射防护特征。
使用质粒dna松弛测定的合成sdg的辐射防护活性
将质粒dna暴露于所选剂量的25gyγ辐射(参见图1),并测定各种浓度(25-250μm)的每种sdg试剂(合成和商业)对dna损伤的抑制%(sc至oc形成)。
在25、50、100和250μmsdg(s,s)存在下暴露于25gy后的质粒dna的代表性凝胶印迹显示于图2a中,并且半定量光密度分析示于图2b中,同时与对照相比的抑制%示于图2c中。有趣的是,在增加浓度的sdg(s,s)(25、50、100和250μm)存在下,sc形式的比例增加并且oc形式的密度显著(p<0.05)和剂量依赖性降低。使用%抑制图(图2c),可以确定每种试剂的ec50值(即,在25gy下防止50%质粒松弛所需的有效浓度(ec)),其对于sdg(s,s)是141.77μm。该用于防止质粒dna松弛的值与清除dpph自由基的ec50值可比较。这些结果证明了合成的sdg(s,s)对映异构体的辐射防护特征。针对sdg(r,r)对映异构体(图2d-f)和sdg(商业的)(图2,g-i)显示了相似的结果,ec50分别为127.96μm和98.38μm。用于防止质粒dna松弛的这些值与清除dpph自由基的相应ec50值可比较。这些结果证明了合成的和可商购的天然sdg的辐射防护特征。
辐射导致从高到低分子量片段的剂量依赖性dna片段化
辐射诱导dna片段化的增加,如图3a中的dna凝胶所示。基于大小,小牛胸腺dna片段分为两组:高分子量(>6,000bp)尺寸和低分子量(<6,000bp)尺寸。高和低分子量片段的分布(图3b)显示高分子量dna的%在25和50gy下分别从88.16%±0.50%降至67.82%±7.89%和34.94%±4.45%(p<0.05)。同时,低分子量片段的比例在25和50gy下分别从11.83%±0.50%增加至32.17%±7.89%和65.05%±4.45%(p<0.05)。结果(图3b)显示高分子量dna的显著降低和低分子量dna片段的显著增加,表明在50gy下对dna的损伤。基于这些初始实验,选择50gy的辐射剂量(观察到清楚的可证实的小牛胸腺dna片段化),用于确定不同sdg的辐射防护特征的以下实验。
使用小牛胸腺dna片段化分析的合成sdg的辐射防护活性
使用如以上所描述的小牛胸腺dna的辐射诱导的片段化确定合成sdg(r,r)、sdg(s,s)和sdg(商业)的辐射防护潜力。
高sdg浓度(25-250μm):图4a显示在25、50、100和250μmsdg(s,s)存在下暴露于50gy后的小牛胸腺dna的代表性dna凝胶。在增加浓度的sdg(s,s)(25、50、100和250μm)的存在下,在辐射暴露后,高分子量dna形式的比例显著增加(p<0.05),而低分子量片段减少。图4b中呈现了在各种浓度的sdg(s,s)下,高和低分子量大小dna形式的分布。这些结果表明使用小牛胸腺基因组dna我们的合成sdg(s,s)对映异构体的辐射防护特征。类似地,图4c-d和4e-f中呈现的结果分别显示合成sdg(r,r)和sdg(商业)的辐射防护性能。这些结果表明使用小牛胸腺基因组dna的合成sdg(r,r)和(s,s)对映异构体的辐射防护特征。
为了进一步确定dna保护中sdg的下限,进行了测试从0.5-10μm范围的低浓度的所有3种sdg的一系列dna片段化实验。
低sdg浓度(0.5-10μm):与其抗氧化剂和自由基清除活性的ec50值相比,在低浓度的sdg(s,s)、sdg(r,r)和sdg(商业)下进行的试验的结果呈现于图5中。类似于更高的sdg浓度,在本节中使用小牛胸腺dna片段化测定法呈现的这些结果证明,即使在低浓度下,合成的sdg(s,s)和sdg(r,r)对映异构体也具有强的辐射防护特征。
使用小牛胸腺dna片段化测定sdg代谢物的辐射防护活性
确定sdg代谢物seco、ed和el的辐射防护潜力,并与如上所述的使用小牛胸腺dna的辐射诱导的片段化的sdg进行比较。基于以上作为中值有效剂量示出的以前的发现,选择每种测试剂的10μm的浓度。结果示于图6中。数据证明sdg及其代谢物seco、ed、el在其防辐射性能方面是等效的。
讨论
本实例的结果表明,合成的sdg(s,s)和sdg(r,r)对映异构体具有强的辐射防护性能。使用质粒dna(pbr322)测定的这些对映异构体的辐射防护潜力,随其浓度的增加而增加。这些合成的sdg(s,s)和sdg(r,r)对映异构体以浓度依赖性方式防止对质粒dna的辐射诱导的损伤。sdg的合成异构体的辐射防护潜力与商业sdg可比较。合成对映异构体sdg(s,s)和sdg(r,r)也防止了小牛胸腺基因组dna的放射诱导的dna片段化。在测试的最低浓度下,sdg(s,s)和sdg(r,r)完全防止辐射诱导的小牛胸腺dna的低分子量片段的产生,证明合成的sdg(s,s)和sdg(r,r)对映异构体的辐射防护特征。使用低浓度的sdg(s,s)、sdg(r,r)和sdg(商业)的结果表明,与其抗氧化剂和自由基清除活性的ec50值相比,用于保护小牛胸腺dna免受γ辐射损伤所需的浓度低得多。重要的是,sdg、seco、ed和el的哺乳动物木脂素代谢物显示出同样有效的dna保护性能。
黄酮类具有强烈的抗氧化剂活性;具体地,这样的多酚具有自由基清除活性,并且已知是比维生素e和c更有效的体外抗氧化剂。还已知具有抗氧化剂性能的饮食和药用植物预防与氧化应激相关的许多人类疾病,并且是有用的辐射防护剂。抗氧化剂,包括维生素和矿物质,在辐射暴露后多年抑制切尔诺贝利工人中的致染色体畸变因子的水平。我们一直使用胸部辐射损伤的小鼠模型调研全谷物饮食亚麻籽、富含木脂素多酚的谷物以及富含sdg的亚麻籽木脂素配制品在辐射诱导损伤中的作用。我们已经表明,当在辐射暴露之前和之后给予时,亚麻籽改善小鼠中辐射诱导的炎症和氧化应激。我们还证明,喂养仅包含亚麻籽的木脂素组分的饮食的辐射小鼠,该饮食富含木脂素双酚sdg,也显示出显著改进的血液动力学测量和存活,同时还改进肺部炎症和氧化组织损伤。这些研究表明亚麻籽通过木脂素sdg的作用针对体内放射诱导组织损伤具有保护性。
活性氧簇(ros)如超氧化物阴离子(o2-)、羟基自由基(.oh)、和过氧化氢的产生的增加导致在各种实验和病理条件下的组织损伤。活性氧簇通过细胞膜脂质、蛋白质和基因组dna的氧化修饰导致细胞损伤。许多研究已经显示,提取的、纯化的或合成的亚麻籽sdg是体外以及体内的有效抗氧化剂。因此,sdg作为抗氧化剂在各种实验和疾病条件下(包括在经历放射疗法的患者中的辐射诱导的组织损伤)具有治疗潜力。
多酚通常在植物中作为糖苷存在并具有抗氧化剂性能。黄酮类作为抗氧化剂,干扰在活性氧簇的产生中涉及的酶活性,淬灭自由基,螯合过渡金属,并使它们在芬顿反应中无氧化还原活性。开环异落叶松脂素(sdg)是亚麻籽中的主要木脂素,并已被证明是一种有效的体外以及体内的抗氧化剂。为了探讨亚麻籽木脂素开环异落叶松脂素(sdg)的治疗潜力,使用香草醛作为前体分子通过化学反应合成sdg,并通过评估以下各项确定合成sdg(r,r)和sdg(s,s)的抗氧化剂性能:其还原能力,金属螯合潜力,和对羟基自由基、过氧化自由基和dpph自由基的自由基清除活性。在本实例中,我们通过评估合成的sdg(r,r)、sdg(s,s)对映异构体和可商购的sdg(作为对照)用于防止γ-辐射诱导的质粒dna(pbr322)和小牛胸腺dna的损伤的潜力,来调研其辐射防护特征。通过质粒dna的开放螺旋形式的增加和质粒dna的超级螺旋形式的减少来评估对质粒dna的辐射诱导的损伤。通过确定dna片段化的水平来评估对基因组dna的辐射诱导的损伤。在该实例中,我们已经检查了合成sdg(r,r)、sdg(s,s)和商业sdg在无细胞系统中对辐射诱导的dna损伤的功效。
之前已经证明了sdg分子的抗氧化剂性能。我们已经表明,天然的,可商购的sdg在暴露于辐射的细胞中具有有效的自由基清除性能。研究了这些合成的sdg(r,r)和sdg(s,s)对映异构体的抗氧化剂和自由基清除特征,并且已经证明具有强的还原能力,高的金属离子螯合潜力,和针对羟基自由基、过氧化自由基和dpph自由基的高自由基清除活性。合成sdg(r,r)和sdg(s,s)的这些特征表明这些分子显示出调节细胞氧化还原状态,降低金属离子浓度和清除氧自由基的很强的潜力。合成的sdg对映异构体的这些特征表明其通过作用于和防止自由基反应的引发、蔓延以及终止的所有三个步骤而起作用的能力,表明这些潜在的机制可能是造成体内sdg(s,s)和sdg(r,r)对映异构体的辐射防护特征的原因。
所进行的一个观察是,通过sdg的基因组dna的最大辐射防护已经在约5.0μm浓度下实现,该浓度远低于其自由基清除和抗氧化剂作用的ec50值。因此,sdg作为抗氧化剂和自由基清除剂也可以作为dna辐射防护剂和辐射缓解剂。
总之,在本实例中,证明合成的sdg(s,s)和sdg(r,r)对映异构体具有强辐射防护特征。使用质粒dna(pbr322)和小牛胸腺dna确定这些对映异构体的辐射防护潜力。合成的sdg(s,s)和sdg(r,r)对映异构体以浓度依赖性方式防止对质粒dna的辐射诱导的损伤。合成对映异构体sdg(s,s)和sdg(r,r)也防止了小牛胸腺基因组dna的放射诱导的片段化。在5μm的浓度下,sdg(s,s)和sdg(r,r)完全防止辐射诱导的小牛胸腺dna的低分子量片段的产生,证明这些对映异构体具有强的辐射防护特征。
实例2
木质素开环异落叶松脂素二葡糖苷(sdg)在肺细胞中的辐射防护性能
细胞的辐射损伤由活性氧簇(ros)的产生引发。ros对细胞机制造成的损伤的谱包括脂质过氧化,dna-蛋白交联,碱基修饰,加合物形成和单链和双链断裂(dnassb和dsb)。这些修饰已经涉及辐射诱导的细胞凋亡和细胞死亡。由于细胞dna损伤是辐射诱导的细胞死亡中已知的决定因素,因此已经做出了显著的努力来鉴定和开发可以通过干扰自由基反应或通过调节辐射诱导的细胞凋亡来保护dna免受辐射损伤的试剂。
通过在暴露时系统中抗氧化剂的存在可以实现防止辐射诱导的基因毒性。由于抗氧化剂可以是干扰自由基链反应的ros清除剂,因此通过补充剂抗氧化剂可以保护细胞dna免受辐射诱导的氧化应激。已经研究了许多合成和天然抗氧化剂化合物的辐射防护功效。然而,它们中的大多数在其有效浓度下表现出固有的毒性和副作用,或者具有短的保质期和低的生物利用度。因此,寻找有效和无毒的辐射防护剂已导致对饮食抗氧化剂和营养制品的调查调研。
我们评估了饮食亚麻籽(fs)补充剂在氧化性肺损伤(如高氧,酸性吸入损伤和缺血/再灌注损伤)的临床前鼠模型中的保护作用。我们确定fs的保护作用可能部分是由于其增强肺组织中的抗氧化剂酶表达的能力。重要的是,饮食fs改善了在暴露之前以及暴露后当给予时胸部辐射的不利影响。在这些研究中,饮食亚麻籽减少辐射诱导氧化肺组织损伤,减少肺部炎症和预防肺纤维化。
早先的报告表明,亚麻籽的多样化作用可能归因于其木脂素,其木脂素已被证明具有抗氧化剂,抗炎和抗致癌作用。开环异落叶松脂素二葡糖苷(sdg)是突出的fs木脂素(约干重的1%),这可能有助于fs颗粒的有益的健康影响。sdg在肠中通过肠道细菌代谢为哺乳动物木脂素,即,肠二醇(ed)和肠内酯(el)。显示出sdg有益于治疗许多疾病(如动脉粥样硬化和糖尿病)的临床前模型。sdg也被报道在动物模型中发挥心脏保护作用。fs木脂素对多种癌症类型具有保护作用,如在阿道夫(adolphe)等人在近期关于sdg的健康影响的综述(英国营养学杂志(brjnutr)2010,103:929)中所总结的,并且报道其减少动物中的黑素瘤转移。
此外,sdg的抗氧化剂和自由基清除性能已有文献记载,这是至关重要的,因为化合物的自由基清除能力可直接与其辐射防护功效相关。在我们对肺内皮细胞的研究中,当细胞暴露于γ-辐射时,sdg显示出自由基清除性能,而全部亚麻籽木脂素组分(flc)富含sdg,介导了小鼠中的辐射防护和辐射缓解。然而,尚未建立sdg的辐射防护性能的表征。
进行这项研究,以确定fs木脂素sdg的辐射防护能力,并探讨负责其作用的可能机制。本研究的第一个目的是评估sdg对原代鼠肺细胞中,特别是上皮,内皮细胞和成纤维细胞中辐射诱导的克隆性死亡的作用。由于辐射诱导的细胞生殖死亡与细胞dna损伤直接相关,因此我们通过使用碱性彗星测定(ssb)和γ-h2ax病灶(dsb)的形成评估了sdg是否可以保护细胞免受辐射诱导的dna链断裂。此外,我们检查了sdg预处理在预防小鼠原代肺细胞避免ir诱导的细胞死亡中的影响。许多研究已经证明促细胞凋亡蛋白bax(bcl-2相关x蛋白)在ir诱导的细胞死亡中的作用。我们还分析了sdg对bax及其拮抗剂bcl-2(b细胞白血病/淋巴瘤2)mrna表达的直接作用,以确定sdg保护的机制是否涉及细胞凋亡的这些关键调节剂的比例的变化。我们的研究结果确定木质素sdg,fs中的有效的生物活性成分,介导肺细胞中的放射防护,从而提供对fs的辐射防护作用的新颖的见解。
材料和方法
试剂
开环异落叶松脂素二葡糖苷(sdg)是可商购的(美国中草药公司,加利福尼亚州)。彗星测定试剂盒购自trevigen公司(盖瑟斯堡,马里兰州)。p-组蛋白h2ax(兔mab)购自细胞信号技术有限公司(cellsignalingtechnology,inc.)(丹弗斯(danvers),马萨诸塞州)。磷酸盐缓冲盐水(pbs),牛血清白蛋白(bsa),具有l-谷氨酰胺、葡萄糖1g/l、不含碳酸氢钠的杜氏改良培养基(dmem),hepes缓冲液,胰蛋白酶,牛血清白蛋白(bsa),乙二胺四乙酸(edta),4,6-二脒基-2-苯基吲哚(dapi),胎牛血清(fbs),胶原酶,triton-x100和分散酶购自西格玛奥德里奇公司,圣路易斯,密苏里州,美国。
细胞系
从c57/bl6小鼠分离成纤维细胞和内皮细胞。对于成纤维细胞分离,收获小鼠肺,切碎,并用分散酶(2mg/ml)孵育45分钟。将这些片铺板,并如前所述培养成纤维细胞,并在通道3和10之间使用。如前所述从小鼠肺分离肺微血管内皮细胞(pmvec)。简言之,用胶原酶处理新鲜收获的小鼠肺,然后通过粘附到用mab包被的磁珠来分离细胞到血小板内皮细胞粘附分子(pecam)上。上皮细胞(c10)细胞最初来源于正常的balb/c小鼠肺外植体,并且是非致瘤性的,接触抑制的,并且在传代早期具有肺泡2型细胞特征。
克隆存活
将指数生长的细胞作为单细胞铺板并孵育过夜。在辐射(2、4、6和8gy)前6h用各种剂量的木脂素sdg(10-50μm)处理细胞。基于当摄取10%亚麻籽时在血液循环中达到的生理水平范围内的动物研究,来选择木脂素剂量。以1.7gy/min的剂量率用mark1铯(cs-137)辐射器(jl谢泼德,圣费尔南多,加利福尼亚州)辐射细胞。辐射后10至15天对菌落进行染色并计数,并且计算存活部分。
彗星分析
培养指数生长的细胞,并在辐射(2gy)之前以不同的时间间隔用sdg(50μm)进行处理。按照制造商的说明书(trevigen公司,盖瑟斯堡,马里兰州)处理细胞用于彗星测定。简言之,将细胞(1x105个细胞/ml,在pbs中)与
γ-h2ax的免疫染色和流式细胞术。
针对γ-h2ax的免疫染色,将细胞铺在玻璃盖玻片(5,000个细胞/盖玻片)上,用50μmsdg预处理(6h)并进行辐射(2gy)。以所需的时间间隔,固定细胞(4%多聚甲醛),洗涤并用pbst(pbs+包含5%山羊血清,1%bsa的0.1%tritonx-100)进行封闭。将细胞用γ-h2ax第一抗体(1:200)在4℃下孵育过夜,接着用pbst(3x5min)进行洗涤并与第二抗体(alexa
针对facs分析,将细胞进行胰酶消化并用pbs进行洗涤。然后将细胞固定(fix/perm缓冲液,ebioscience公司)45分钟,并且此后使用渗透清洗缓冲液(biolegend公司,美国)洗涤。将细胞重悬于与alexa
凋亡细胞的形态学检测
凋亡细胞的形态学特征在于细胞核和细胞质的缩合,膜起泡,细胞收缩以及细胞核dna的分解,首先在大区段中,并且随后在核小体片段中,并且最后形成良好封闭的细胞凋亡小体。从用于微核检测的载玻片经显微镜确定经历细胞凋亡的细胞的百分比(细胞遗传损伤)。对于每个实验(实验进行两次)计数至少500个细胞,并如下确定凋亡细胞百分比:
%细胞死亡=na/ntx100,
其中na是具有细胞凋亡体的细胞的数量,并且nt是所分析的细胞的总数。
通过定量实时pcr(qpcr)的基因表达分析
使用由应用生物系统公司(appliedbiosystems),生命技术公司(卡尔斯拜德,加利福利亚州)提供的
简言之,将细胞用sdg(50μm,6hrs)进行预处理并辐射(2gy)。使用rneasyplus迷你试剂盒(凯杰有限公司,巴伦西亚,加利福尼亚州)分离总rna,并使用nanodrop2000(赛默飞世尔科技公司(thermofisherscientific),沃尔瑟姆,马萨诸塞州)定量。然后使用由应用生物系统公司,生命技术公司(卡尔斯拜德,加利福利亚州)提供的高容量rna至cdna试剂盒在
通过蛋白质印迹法的细胞凋亡检测
通过使用免疫印迹观察到的bax(细胞凋亡启动子),bcl-2(细胞凋亡抑制剂),切割的半胱氨酸蛋白酶-3和切割的聚(腺苷二磷酸-核糖)聚合酶(parp)的水平在小鼠肺上皮细胞中确定细胞凋亡。简言之,在包含蛋白酶抑制剂的pbs中裂解细胞。然后使用10孔sds12%nupage凝胶(英杰公司,卡尔斯巴德,加利福尼亚州)进行细胞裂解物的免疫印迹分析。在200v下进行电泳1小时。在25伏下转移至polyscreenpv转移膜(珀金埃尔默生命科学公司(perkinelmerlifesciences),波士顿,马萨诸塞州)进行1小时。将膜在磷酸盐缓冲盐水中的5%脱脂奶粉中封闭过夜。然后弃去脱脂奶粉,并且用第一抗体孵育该膜。使用制造商推荐的稀释液(细胞信号传导技术公司,丹弗斯,马萨诸塞州),使用针对bax和bcl-2的兔抗小鼠单克隆抗体,以及兔抗小鼠切割的半胱氨酸蛋白酶-3(asp175),单克隆抗体和兔多克隆抗切割的parp(214/215)切割位点特异性抗体,来检测bax、bcl-2、切割的半胱氨酸蛋白酶-3、和切割的parp的蛋白质水平。将膜洗涤5次,并且然后在与辣根过氧化物酶共轭的第二抗体中在室温下孵育45分钟。使用westernlightingchemiluminescencereagentplus(珀金埃尔默生命科学公司,波士顿,马萨诸塞州)显影膜,并且通过特异性条带(bax为20kda,bcl-2为26kda,切割的半胱氨酸蛋白酶-3为17/19kda,并且切割的parp为89kda)的光密度扫描进行定量,该特异性条带使用通过特异性第二抗体(西格玛,圣路易斯,密苏里州)检测的β-肌动蛋白表达水平调整负载。
统计
结果表示为平均值±sem。使用kaleidagraph软件(4.0)制备克隆生成测定的存活曲线。使用单向方差分析(anova)确定组间的统计学差异。当发现统计学显著差异(p<0.05)时,使用bonferoni/dunn测试(statview4.0)进行个体比较。
结果
sdg治疗增加了辐射的原代肺细胞的集落形成能力
克隆生存测定法已广泛用于在暴露于环境和药物致癌物质、电离辐射等之后在细胞经历任何基因毒性应激后确定细胞生殖死亡。在该实例中,评估了sdg(10-50μm)预处理对原发性肺细胞(上皮细胞,内皮细胞和成纤维细胞)的克隆形成中辐射诱导的减少的影响。结果显示,与它们各自的未处理的对照细胞(100%)相比,单独的sdg(10-50μm)不引起对所有三种细胞类型的集落形成能力的任何不利影响(图1a-1c)。
辐射治疗以剂量依赖性方式显著(p≤0.01)降低了上皮和内皮细胞的集落形成能力。当在辐射前用sdg处理细胞时,在所有处理组中存活的部分显著增强(图7a,7b)。在50μmsdg预处理的辐射组中观察到对在成纤维细胞中克隆形成的辐射诱导的损失的最大保护(图7c)。因此,我们选择这种特定浓度的sdg用于我们的进一步研究。
sdg防止在所辐射的原发性肺细胞中形成dnassb
我们首先进行了一项研究,以确定在放射生物相关剂量2gy后在所有细胞类型(内皮、上皮、成纤维细胞)中dna损伤的动力学。如所预期的,与其相应的未经辐射的对照细胞相比,在所有细胞类型中辐射暴露诱导了显著的dna损伤,如增加的尾力矩所证明的。dna损伤的程度在辐射后30分钟最大。dna损伤的程度随着辐射后长达60分钟的时间而降低,并且在辐射后2小时进一步急剧下降。因此,我们选择30分钟时间间隔进行与放射诱导的dna损伤相关的进一步研究(图8a)。
与其未辐射的对照细胞相比,辐射暴露(2gy)导致彗星尾长显著增加。在辐射前以各种时间间隔(0h、2h、4h、6h和24h)处理细胞。在所有时间间隔用sdg(50μm)预处理细胞显著抑制所有三种类型的肺细胞中的辐射诱导的彗星尾长。然而,在辐射前6hsdg处理时观察到在所有细胞类型中对dna的辐射诱导尾力矩的最大保护(图8b)。描述在辐射(在sdg的存在和/或不存在)肺上皮细胞中彗尾形成的代表性荧光显微照片已经在插入物,图8b中显示。
sdg治疗废除了鼠原代肺细胞中γ-h2ax病灶的形成
为了进一步测试我们的假设,sdg是否可以保护免受氧化性dna损伤,我们还评估了sdg对诱导γ-h2ax病灶的作用。使用标准显微镜生成的图像分析(图9,10)和流式细胞术(图11)方法评估sdg预处理对氧化性dna损伤的影响,其由辐射后鼠原代肺细胞中的γ-h2ax形成证实。荧光显微镜分析的结果显示,辐射(2gy)暴露导致所有三种细胞类型中γ-h2ax病灶的形成的显著增加(图9)。病灶/细胞数量基本上增加15分钟,在辐射后30分钟达到峰值(针对上皮、内皮和成纤维细胞的分别为46.7%±0.5、33.6%±3.2和30.0%±1.4的γ-h2ax阳性细胞),而数字在暴露的1小时内显著降低,但仍显著高于未经辐射的对照细胞。与它们各自的未经辐射的对照细胞(针对所有细胞类型p<0.005)相比,所有值都显著更高(图9)。
sdg预处理(ir前6小时)显著降低了γ-h2ax的诱导,因为在经辐射的上皮细胞、内皮细胞和成纤维细胞中,γ-h2ax阳性细胞数量分别降低到22.7%±2.17、21.92%±2.88和22.1%±1.9(针对上皮,p<0.005,并且针对内皮和成纤维细胞,p<0.05)。sdg保护细胞免受γ-h2ax病灶形成的能力似乎独立于细胞类型,因为sdg的预处理保护所有三种类型的肺细胞免受辐射诱导的dna链断裂。图10描述了原代肺上皮细胞中γ-h2ax阳性细胞的显微镜分析的代表性荧光显微照片。
使用流式细胞术进一步证实了辐射暴露后sdg对钝化γ-h2ax阳性细胞的诱导的保护作用。如所预期的,在辐射后观察到在γ-h2ax阳性细胞的诱导中的类似模式;然而,在所有细胞类型中,通过sdg预处理显著消除了γ-h2ax阳性细胞的数量(图11)。
sdg治疗防止原代肺细胞免受ir诱导的细胞凋亡死亡
为了研究sdg在细胞凋亡方面的细胞保护作用,用dapi染色细胞核用于通过显微镜的目测并计数。显微镜分析证实,对照细胞具有完整的染色质(凋亡细胞约4%-5%),而辐射暴露显著(p≤0.05)以时间和剂量依赖性方式增加凋亡细胞的百分比。在24h时,在上皮和内皮细胞中分别观察到凋亡细胞的百分比为13.9%±1.08和15.1%±1.95。sdg预处理(50μm)明显(p≤0.05)反映了ir诱导的凋亡细胞百分比增加(在上皮和内皮细胞中分别为9.9%±1.08和10.71%±1.45凋亡细胞),如图12(a和b)中所示。发现成纤维细胞最敏感,因为辐射暴露导致在24和48hr凋亡细胞分别显著提高36.4%和41.8%。重要的是,如上皮和内皮细胞所示,用sdg预处理也显著降低了肺成纤维细胞中的细胞凋亡程度(图12c)。
sdg治疗修饰鼠原发性肺上皮细胞中细胞凋亡调节子的表达
为了进一步阐明sdg在钝化dna损伤和细胞死亡中的辐射防护作用,我们假设sdg可以改变促细胞凋亡和抗细胞凋亡调节子蛋白的比例。因此,我们测试了在辐射存在或不存在下sdg治疗肺细胞是否会改变bax和bcl-2的基因表达,以及这些变化是否将转化为蛋白质水平的变化。为此,用sdg(50μm)处理肺上皮、内皮和成纤维细胞,并通过qpcr在ir后6、24和48小时评估酶mrna水平(图13a和13b)。我们观察到2gy导致在ir后24和48小时促细胞凋亡baxmrna水平增加约11倍,其通过sdg预处理显著(p<0.05)抑制。可替代地,抗细胞凋亡bcl-2mrna水平不由于辐射暴露而改变,然而在ir处理后24和48小时,在sdg预处理的细胞中分别增加了6.6和3.5倍。当通过蛋白质印迹分析bax和bcl-2mrna水平的变化的蛋白质验证时,我们发现bax和bcl-2蛋白水平的随后增加趋向于在sdg预处理的细胞中降低的水平(图13c描绘了代表性印迹,并且13d和13e显示了带密度定量)。为了进一步调研通过sdg观察到的辐射防护背后的潜在机制,我们评估了sdg在改变细胞凋亡信号级联:执行子切割的半胱氨酸蛋白酶-3和切割的parp中涉及的关键蛋白水平中的作用。总的来说,切割的半胱氨酸蛋白酶-3和切割的parp的水平在经辐射的细胞中在ir后6、24和48小时显著增加,并且趋向于在sdg预处理的细胞中降低的水平(图14a描述了代表性印迹,并且14b和14c显示了带密度定量)。
讨论
在这个实例中,我们证明fs木脂素sdg可以保护鼠原代肺细胞免受辐射诱导的氧化性dna损伤和细胞凋亡死亡。我们观察到sdg预处理不仅改善ir诱导的细胞毒性,如通过克隆形成生存测量的,而且还减少了dna链断裂(dsbs和ssbs)的诱导和肺细胞中的细胞死亡。重要的是,通过sdg治疗还改变了参与细胞凋亡调节的基因的表达。这些研究结果表明sdg可能作为无毒的辐射防护剂在辐射情况下和改进放射治疗的治疗指数中是有用的。
ir诱导的细胞死亡是细胞放射敏感性的经典标志物,其特征在于克隆形成生存的丧失。类似地,我们还注意到所有三种鼠肺细胞中的克隆形成的辐射剂量依赖性损失,其通过sdg预处理显著减弱(图7)。细胞dna是细胞中ir诱导的损伤的主要靶标。在暴露期间产生的ros诱导细胞dna的改变的阵列,其范围从突变、碱基损伤、交联、ssb和dsb。dna中的损伤不能被代替,并且因此必须修复,并且如果其保持未修复,该细胞可以诉诸诱导细胞凋亡或坏死。因此,靶细胞dna的保护赋予针对基因毒性损害的第一道防线。ros产生在暴露的几秒内发生,并且在辐射后持续几分钟。因此,早期辐射应答(如立即dna损伤)在辐射暴露后几分钟内发生。类似地,在受辐射细胞的时间依赖性动力学研究中,我们还观察到dna链断裂的程度,如彗星尾长所证明的,在暴露后30分钟内达到其最大水平,并随后减少(图8a)。
彗星测定是用于在细胞水平下检测dna损伤/修复的敏感技术,并且已广泛用于研究dna链断裂。多酚通过减少ros介导的氧化性dna损伤具有保护正常组织或细胞免受辐射的损伤作用的能力。在本研究中,通过使用标准碱性彗星测定,我们还评估了sdg对ir诱导的ssb在鼠原代肺细胞中的作用。我们的结果与其他报告一致,这些其他报告显示针对其他多酚(如,白杨素和表儿茶酸)免受辐射诱导的dnassb的类似的保护作用。
尽管ssb容易被细胞修复,但是dsb对于细胞更难以修复,并且更可能导致诱变,因此dsb主要代表致死性细胞事件。h2ax分子对于通过辐射引入的每个dna双链断裂沿着兆碱基长染色质结构域变为磷酸化的(γ-h2ax),并且γ-h2ax损失或去磷酸化与dna修复在时间上相关。我们在这里报告,在所有三种类型测试的细胞中,sdg保护细胞dna避免红外诱导dsb。我们的结果与一些其他研究一致,这些其他研究也表明多酚像白藜芦醇和绿茶儿茶素保护细胞免受ir诱导dna链断裂。然而,在辐射不存在下,sdg在这些细胞二者之一中都没有发挥任何毒性作用。由于氧化性dna损伤被认为是许多癌症的前体,所以通过作为抗氧化剂的sdg的这种损伤的减少可以导致降低的癌症风险。
众所周知,ir诱导的细胞和组织中细胞凋亡部分是由于细胞中dna损伤的诱导。虽然若干种机制(o6-甲基鸟嘌呤、碱基n-烷基化、庞大的dna加合物,dna交联)参与dna损伤依赖性细胞凋亡,但是dna双链断裂在诱导凋亡性细胞死亡中起主要作用。在这项研究中,用sdg预处理细胞保护避免肺细胞中辐射诱导细胞凋亡并且抑制彗星尾长和γ-h2ax病灶形成。因此,sdg对辐射诱导的凋亡性细胞死亡的保护作用可以归因于其减少细胞氧化应激的能力、离子稳态恢复及其防止dna损伤的能力。这些发现与在我们的缺血再灌注和辐射诱导的组织损伤的鼠研究中的饮食整粒亚麻籽的抗细胞凋亡性能一致,并且表明sdg作为亚麻籽中可能的生物活性成分,介导组织中的保护性,抗凋亡作用。
因为ir降低了细胞环境中抗氧化剂酶的水平,所以通过抗氧化剂对细胞的预处理可能进一步干扰ros,并且从而降低ros与生物分子相互作用的风险。在前面的实例中,我们早先报道,用包含亚麻籽木脂素复合物的饮食预饲喂动物增加小鼠肺中保护性ii期抗氧化剂酶的水平。这些作用的假设机制涉及激活抗氧化剂应答元件(are)介导的转录诱导。此外,nrf2还启动负责保护免受氧化应激介导的细胞毒性的各种抗氧化剂酶的从头合成。一些其他报告还表明多酚像姜黄素和egcg对氧化应激发挥保护作用,这与它们诱导ii期抗氧化剂酶的能力有关。
从结果显而易见,sdg减少鼠原代肺细胞中的细胞死亡,这是由于其抗氧化剂性能和对dna的保护作用。我们的发现证实了许(hseu)等人(食品和化学毒理学(foodchemtoxicol)2012,50:1245)的结果,其报道了鞣花酸,在石榴中发现的多酚,通过ho-1和nrf-2基因的上调来保护人角质形成细胞免受uva诱导的氧化应激和细胞凋亡。我们的研究提供了新颖的证据,sdg在抗击ir诱导的氧化损伤和凋亡细胞死亡的辐射防护作用可能是通过直接诱导抗氧化剂防御系统。
总之,这些结果证实亚麻籽中的亚麻籽木脂素sdg具有有效的辐射防护性能,其可能有助于在动物研究中观察到全谷物的保护作用。
实例3
开环异落叶松脂素二葡糖苷(sdg)清除次氯酸离子:sdg保护基因组dna免受辐射的新颖机制
次氯酸(hocl),一种有效的氧化剂,是由嗜中性粒细胞通过活化的髓过氧化物酶产生,该髓过氧化物酶催化生理存在的氯离子和过氧化氢(h2o2)之间的反应。活化的嗜中性粒细胞产生h2o2和超氧化物阴离子o2·-。在生理ph下,存在hocl和次氯酸根离子(clo-)的混合物。hocl通过氧化损伤杀死微生物。然而,已知过量的生产会对组织造成损害。次氯酸盐修饰腺嘌呤核苷酸导致氯胺的形成,这似乎是嗜中性粒细胞介导的毒性的主要机制。
hocl及其共轭碱clo-已显示出氧化氨基酸、肽、蛋白质和脂质,并氯化细胞dna和rna中的碱基。hocl/clo-的反应导致嘌呤和嘧啶核苷酸在鸟嘌呤和胸腺嘧啶的环内-nh基团以及鸟嘌呤、腺嘌呤和胞嘧啶衍生物的环外nh2基团上的修饰,导致氯胺(如(rnhcl)和rr’ncl)的形成。发现主要修饰的碱基是在skm-1细胞的dna和rna中的5-氯胞嘧啶、8-氯腺嘌呤和8-氯鸟嘌呤。
众所周知,γ-辐射能够电离原子和分子。在生物系统中或在溶液中,电离辐射产生羟基自由基(·oh),其被认为是电离辐射诱导的对细胞组分(包括脂质、蛋白质和dna)的损伤的来源。然而,这些高度不稳定的自由基可被cl-离子清除,这些cl-离子以非常高的浓度存在于生理介质中。这导致产生含活性氯的中间体,其中相对稳定的clo-是辐射衍生的毒物。在含氯化物的溶液中,clo-和氧化性质的其他活性氯衍生物形成为辐射分解的产物;它们可以有助于抑制有机体的生理功能。因此,我们建议辐射诱导的dna或蛋白质损伤部分地通过辐射生成的clo-来介导。
化学合成的sdg的两种非对映异构体最近已被证明在其抗氧化剂,自由基清除和dna保护性能方面是等效的。本研究使用高度新颖和特异性荧光探针评估在避免生物盐水溶液中γ-辐射诱导产生的clo-的dna辐射防护中的sdg。次氯酸盐特异性3’-(p-氨基苯基)荧光素(apf)和羟基自由基-敏感的3’-(p-羟苯基)荧光素(hpf)提供用于确定上述活性氧簇(ros)的更大的特异性和再现性。
材料与方法
化学品
ros指示探针apf和hpf、质粒dna(pbr322)和1kb加dna梯来自英杰公司(生命技术公司,卡尔斯拜德,加利福利亚州)。
确定次氯酸盐
在次氯酸盐存在下或在γ-辐射暴露后,在490nm/515nm的激发/发射下测量pbs中的ros探针apf和hpf的荧光。数据表示为相对荧光单位(rfu)。
通过确定牛磺酸氯胺γ-辐射诱导产生次氯酸盐
使用tmb测定法确定牛磺酸的氯化。数据表示为牛磺酸氯胺(吸光度)以及clo-浓度(μm)。
次氯酸盐诱导对小牛胸腺和质粒dna的损伤
在37℃下,用次氯酸盐孵育小牛胸腺或质粒dna2hr。使dna样品进行琼脂糖(1%)凝胶电泳并且进行分析。
次氯酸盐诱导的2-氨基嘌呤(2-ap)氯化的确定
将pbs中的2-ap暴露于次氯酸盐,并且在360-390nm之间记录荧光光谱,在374nm下具有发射最大值。计算2-ap的变化%。
数据的统计分析
获得的数据表示为平均值±标准偏差。使用邦弗朗尼校正用statview程序,使数据经受单向方差分析(anova)与事后比较。p值≤0.05被认为是有意义的。
结果
在这项研究中,我们调研sdg清除辐射诱导的clo-的能力,作为在生理溶液中dna保护免受辐射暴露的潜在机制。
sdg清除次氯酸离子
使用次氯酸钠评估所选择的荧光探针的特异性。图15a示出了随着clo-浓度(1-4μm)的增加apf荧光强度的线性增加。重要的是,hpf荧光强度仅略有增加,表明apf荧光主要是clo-依赖性的。选择clo-剂量在后续实验的该范围内。然后评估sdg(可商购的)通过sdg清除clo-的能力。事实上,sdg剂量依赖性地降低clo-(图15b)。最后,我们评估了合成的sdg非对映异构体sdg(s,s)和sdg(r,r)的clo-清除效应。在0.5μm下,sdg(r,r)和sdg(s,s)和sdg(商业)清除clo-(图15c),具有与类似于水飞蓟宾(一种已建立的clo-清除剂)的可比较的效力。
sdg清除γ-辐射诱导的次氯酸盐
辐射剂量依赖性产生clo-,如由apf的荧光强度的增加证明的。具体地,50gyγ-辐射诱导的apf和hpf(图16a),其通过sdg剂量依赖性地降低(图16a和b)。apf荧光(1,816.30)的降低的初始斜率比hpf(695.75)高,表明sdg对clo-的选择性清除效应(插入物,图16a)。与hpf相比,sdg对apf的影响明显更显著(图16b)。sdg钝化来自辐射暴露(25和50gy)的clo-生成,通过增加的apf(图16c)和hpf(图16d)荧光(p<0.05)显示。apf/hpf的比率通过sdg降低,表明clo-的选择性清除。
当氯化牛磺酸时,次氯酸盐辐射剂量依赖性增加
为了建立辐射诱导clo-氯化生物分子中的-nh基团,我们评估了放射诱导的牛磺酸氯化。结果(图16e)显示在生理盐水中,γ-辐射在50、100和200gy下显著增加了牛磺酸氯胺形成,表明γ-辐射诱导clo-产生,其依赖于氯化物浓度(图16f)。结果提供了强有力的证据,辐射诱导clo-在生理溶液中,能够损害生物分子。
sdg保护次氯酸盐诱导对小牛胸腺和质粒dna的损伤
我们确定clo-是否诱导对基因组(图17a-d)和质粒(图17e-f)dna的损伤。实际上,clo-诱导剂量依赖性dna片段化(图17a,b),同时低分子量片段增加。通过所有sdg钝化对暴露于0.5mm次氯酸盐1.0μm的基因组dna的损伤,至与槲皮素,已知的抗氧化剂和水飞蓟宾,clo-清除剂可比较的水平(图17c,d)。类似地,通过sdg钝化由clo-对质粒dna的损伤(图17e,f)。具体来说,与在琼脂糖凝胶电泳中具有不同迁移模式的、损坏的、开放螺旋的dna相比,我们评估超级螺旋(sc)质粒dna的数量与oc(18.6%±9.4%)相比,以(25μm)存在的sdg减少了clo-诱导的对质粒dna的损伤,并保持大部分(81.3%±9.4%)dna处于超级螺旋形式。
sdg对次氯酸盐诱导2-ap氯化的保护作用
为了确定clo-对dna的损伤是否通过碱基修饰发生,我们评估了clo-诱导的2-ap(一种嘌呤的荧光类似物)的氯化作用。以与通过辐射暴露(图15a,16a,17a)产生的那些浓度的可比较浓度(10μm)给出的次氯酸盐降低了2-ap荧光,该2-ap荧光通过用sdg预处理(60秒)来防止(图18a和b)。最重要的是(图18b和c),如果在暴露于clo-后+15、+30、+60、+120、+180或+300秒处添加,用sdg后处理导致从次氯酸盐诱导的2-ap修饰显著恢复。这些结果证实sdg对次氯酸盐诱导的嘌呤碱基修饰的核碱基保护特征。
sdg对2-ap的γ-辐射诱导的氯化的保护作用
为了确定辐射是否诱导核碱基氯化,我们使用与上述相同的系统,即2-ap荧光(图19a)。事实上,暴露于γ-辐射导致荧光强度的剂量依赖性降低(类似于在clo-存在下的观察(图18a),其通过sdg(图19a和b)来防止)。这些结果证明γ-辐射诱导核碱基的氯化并且建立了sdg针对嘌呤碱基的这种辐射诱导修饰的保护性能。
sdg作用在预防或减轻通过辐射引起的核碱基氯化中的建议机制
关于n-氯化核碱基的恢复或预防的机制,我们建议或通过富电子芳环的双电子还原n-cl分子或单电子还原,导致形成游离的n-根,其进而从苯酚sdg部分中的-oh基团捕获氢原子。方案1(图20)表明sdg保护作用的建议机制。我们目前的研究结果提供通过清除次氯酸离子并且保护免受辐射诱导的dna损伤的sdg的辐射防护作用的新机制的证据。
讨论
这项研究的结果提供以下各项的证据:i)sdg,一种已知的木脂素抗氧化剂和自由基清除剂,通过化学方法以及通过辐射在生理溶液中产生的脱毒次氯酸离子;ii)sdg保护基因组dna以及来自次氯酸盐诱导的损伤的质粒dna;iii)sdg防御(从次氯酸盐诱导的dna损伤中保护或恢复)的机制涉及清除次氯酸盐和再生来自氯胺(-ncl)的核碱基上的氨基(-nh);iv)暴露于γ-辐射导致牛磺酸氯胺形成的增加;v)暴露于γ-辐射导致通过sdg防止的嘌呤碱的增加的氯化;vi)当在暴露之前或之后添加时,sdg作用同样有效地保护dna免受次氯酸盐诱导的损伤,即它可以作为核碱基氯化的保护剂和/或缓和剂;vii)与商业的sdg、水飞蓟宾和槲皮素(天然抗氧化剂黄酮类)相比,合成的sdg(s,s)和sdg(r,r)非对映异构体在清除次氯酸离子并且防止次氯酸盐诱导的dna损伤方面同样有效。这些结果证实sdg对于次氯酸盐诱导的核碱基修饰的保护和减轻性能。
使用胸部辐射损伤的小鼠模型,我们建立了全谷物饮食亚麻籽,富含木脂素多酚的谷物以及富含sdg的亚麻籽木脂素配制品的组织辐射防护作用。这些研究强调木质素sdg对辐射诱导的体内组织损伤的辐射防护和辐射减轻性能。提取、纯化或合成亚麻籽sdg是体外以及体内有效的抗氧化剂。为了探索sdg的治疗潜力,我们通过新颖的化学反应合成sdg,并通过评估其还原能力,金属螯合潜力,和针对·oh自由基、过氧化自由基和dpph自由基的自由基清除活性来确定合成的sdg(r,r)和sdg(s,s)的抗氧化剂性能。我们还通过评估其预防γ-辐射诱导对质粒dna(pbr322)和小牛胸腺dna的损伤的潜力,证明了合成的sdg(r,r)、sdg(s,s)非对映异构体的辐射防护特征。
我们还表明,通过sdg的基因组dna的最大辐射防护已经在约5.0μm浓度下实现,该浓度低于其自由基清除和抗氧化剂作用的ec50值,典型地在130-200μm的范围内。有趣的是,在本研究中确定的sdg在清除辐射诱导的次氯酸盐中的最大有效性在0.5至5μm内。这表明sdg的dna保护作用部分是由于清除了损伤性clo-。显示sdg对次氯酸盐诱导的2-ap修饰的保护作用的结果表明,sdg通过防止次氯酸盐诱导的对核碱基的损伤来保护dna。
hocl由活化的嗜中性粒细胞的髓过氧化物酶使用由nadph氧化酶和氯离子作为底物产生的过氧化氢产生。hocl可以氯化并氧化核碱基。由于hocl可氯化核碱基,因此这可能引起基因毒性。氯化核苷已经被鉴定并与炎症和癌症相关。
可能存在若干种潜在的机制,通过这些机制sdg可以保护dna免受辐射诱导的损伤:1)通过清除引起核碱基的氯化和氧化的次氯酸根离子;2)通过清除通过与氯离子反应产生次氯酸盐的·oh自由基。我们观察到在氯离子存在下在磷酸盐缓冲盐水(pbs)中以及在单独的盐水(初步实验)中,·oh的产生急剧下降。考虑到氯离子在体内生理介质中的高浓度,次氯酸盐产生的显著性和次氯酸盐诱导的对包括dna的细胞组分的损伤将是辐射损伤的主要机制;3)通过与dna碱基对结合,作为若干黄酮类如lutiolin、山奈酚和槲皮素;4)通过阻断相似嘌呤和嘧啶碱基,特别是c5、c6和c8上以及在脱氧核糖位点处质子的抽取或·oh的添加。这些机制已被提出用于保护免受自由基诱导的dna损伤;5)最后,通过还原形成的氯胺,从而再生核酸中的内部和外部氨基基团。因此,sdg作为次氯酸离子的清除剂以及作为抗氧化剂和自由基清除剂以及来自次氯酸盐诱导的氯化作用的核碱基的保护剂可以充当dna辐射防护剂和辐射缓解剂。
次氯酸盐以ph依赖的量作为次氯酸盐阴离子(clo-),次氯酸(hclo)和游离氯(cl2)的混合物存在于溶液中。在生理ph下,clo-和hclo是主要分子。与强的单电子氧化剂如·oh相比,次氯酸盐是一种双电子氧化剂,比·oh更少反应性并且更具选择性。次氯酸盐可氯化富含电子的芳环和nh-化合物。它氧化伯醇和仲醇以及苄基亚甲基基团和叔次甲基基团和酚。上述反应的第一步是氯化,然后水解/hcl消除。sdg包含所有上述反应位点,除了使sdg分子成为强效次氯酸盐清除剂的氨基。在方案1(图20)中,我们提出了通过sdg使用核苷碱基的1-电子或2-电子还原的dna保护的新颖机制。
总之,我们已经证明sdg清除次氯酸(clo-)离子并防止辐射诱导的次氯酸盐和dna损伤。由于已知次氯酸离子通过氯化/氧化修饰dna碱基,并且然后随后导致dna损伤,因此我们的发现显示sdg可用作与癌症治疗的放射治疗或意外暴露于辐射相关的正常组织损伤的辐射防护剂。
实例4
亚麻籽及其木脂素保护免受苯并-α-芘的影响
亚麻籽、sdg和木脂素衍生物通过抑制多步致癌作用过程来减轻由烟草和其他环境致癌物造成的肺部肿瘤发生(图21)。在该实例中,提供了实验,表明在动物模型中,木脂素sdg具有通过调节nrf2调节的阶段ii解毒通路和可能其他机制的化学预防活性。sdg的保护作用由直接ros清除和/或间接抗氧化/抗炎性能以及降低致癌物毒性和dna损伤来介导。
鼠上皮细胞暴露于烟草和环境致癌物苯并-α-芘(bap)诱导破坏性活性氧簇(ros),如通过氧化还原敏感性荧光染料检测的(图23)。早在暴露于致癌物后2小时,荧光强度的强烈增加表明细胞中ros的产生。
为了证明sdg降低细胞中致癌物诱导的氧化性dna损伤,将sdg(10μm)添加至暴露于25μm的bap的人上皮细胞(a549)中,并使用质谱法检测对dna的氧化损伤,如8-氧代-7,8-二氢鸟嘌呤(8-氧代-dguo)的存在指示的。sdg在致癌物暴露后3和6小时减少dna损伤(图22)。同样,为了证明sdg防止ros从致癌物暴露中产生,将小鼠上皮细胞暴露于10或20μmbap和增加浓度的sdg(0、0.1、0.5、1、5μmsdg),并在2小时后检测ros。sdg清除有害ros至可忽略的水平(图24)。
类似地,sdg防止暴露于bap的人上皮细胞中的基因毒性应激。将细胞暴露于强致癌物如bap,诱导基因毒性应激,如通过增加的p53蛋白水平所指示的(图25)。这通过在5、10、25和50μm浓度下sdg的存在而减轻剂量依赖性。sdg还防止暴露于bap的人上皮细胞中的氧化性dna损伤。将细胞暴露于强致癌物如bap,诱导氧化性dna损伤,如通过增加的γ-h2ax(双链dna断裂的标志物)的水平所指示的(图26)。这通过在5、10、25和100μm浓度下sdg的存在而减轻剂量依赖性。此外,sdg防止暴露于bap的人上皮细胞中的dna加合物形成。将细胞暴露于强致癌物如bap诱导dna加合物的形成(图27)。dna加合物是与致癌物共价连接的dna片段,并与恶性肿瘤的发展直接相关。dna加合物水平通过单独或组合给予sdg或其代谢物ed和el的存在而降低。
亚麻籽及其木脂素在化学致癌物诱导的肺肿瘤的小鼠模型中提供保护。给小鼠(a/j菌株)以1mg/kg剂量经腹膜内注射4次烟草和环境致癌物bap(每周一次)(图28)。小鼠在暴露时用亚麻籽或木脂素饮食开始。在暴露后在不同时间评估小鼠,以确定肿瘤负荷,小鼠重量和总体健康状况。
亚麻籽降低小鼠中的肿瘤负荷:
图29呈现了bap暴露和饮食亚麻籽给予后若干个月,鼠肺的代表性临床图像。图30呈现了bap暴露和饮食亚麻籽给予后若干个月,鼠肺的代表性h&e应变肺切片。来自喂食对照饮食(上图)或亚麻籽(下图)的小鼠的箭头指示的结节在亚麻籽喂养的小鼠中显示更小。每个小图代表不同的动物。
使用针对整体肿瘤面积和结节大小(图31a和b)的图像分析软件进行形态学评估组织学鼠肺切片。在喂养亚麻籽饮食的小鼠中,肿瘤占据的肺面积显著减少(p<0.03)。类似地,存在更小肿瘤结节尺寸的趋势。针对每个肺的肿瘤结节的总数(图32a)和侵袭肺的肿瘤%(图32b),还将组织学鼠肺切片进行了形态学评估。每个肺存在更少的肿瘤结节的趋势(a)并且在亚麻籽补充剂下更少的肿瘤侵入的趋势(b)。
最后,亚麻籽补充剂显示防止来自由bap诱导的肺癌的耗散效应。在bap暴露后纵向测量动物重量200天。饲喂亚麻籽饮食的小鼠暴露于bap下,显示比当对照饮食时暴露于bap的小鼠显示更高的重量(图33)。
实例5
亚麻籽及其木脂素保护细胞和组织免受石棉诱导的损伤
介绍
恶性间皮瘤(mm)是一种毁灭性的,疼痛的和致死型的癌症,没有疗法和治疗的现实机会。mm的发展与暴露至石棉纤维直接相关。最近的研究表明,石棉诱导的癌症的发病机理是由于通过持续石棉纤维造成的慢性炎症和氧化组织损伤。全粒亚麻籽(fs)具有已知的抗氧化剂,抗炎和癌症化学预防性能。在该实例中,在nf2+/mut;cdkn2a+/mut小鼠中测试了fs及其富含在饮食中给出的木脂素开环异落叶松脂素二葡糖苷(sdg)的木脂素组分(flc)防止急性石棉诱导的炎症和炎症细胞因子释放的能力。
材料和方法
小鼠饮食和石棉暴露
在啮齿动物食物中给出了fs和其富含木脂素开环异落叶松脂素二葡糖苷(sdg)的木脂素组分(flc)。将小鼠(nf2+/mut;cdkn2a+/mut)在单次ip推注400mg青石棉石棉后一天(+1)或前一天(-1)置于10%fs或10%flc补充剂饮食上,并且评估3天后腹部炎症和促炎细胞因子释放(参见图34)。选择nf2小鼠品系,因为当暴露于石棉时其形成mm的加速形式。
组织收获和分析
使用液相色谱,串联质谱(lc/ms/ms),评估亚麻籽木脂素代谢物(如哺乳动物木脂素肠内酯(el)和肠二醇(ed))的系统水平(即,血浆),以确保fs被该小鼠品系的肠道菌群有效代谢,并且水平与其他小鼠模型中的那些可比较。
使用pbs进行腹部灌洗,并使用细胞离心涂片分析确定巨噬细胞(mf)和嗜中性粒细胞(pmn)的水平。
讨论
使用液相色谱,串联质谱(lc/ms/ms),评估亚麻籽木脂素代谢物(如哺乳动物木脂素肠内酯(el)和肠二醇(ed))的系统水平(即,血浆),以确保fs被该小鼠品系的肠道菌群有效代谢,并且水平与其他小鼠模型中的那些可比较(图37)。巨噬细胞(mf)和嗜中性粒细胞(pmn)的腹部灌洗水平表明fs和flc都钝化由石棉诱导的急性腹部炎症(图36和38)。此外,促炎细胞因子tnf-α和il-1β的水平也通过饮食剂降低(图37和38)。
结论:这些发现表明亚麻籽及其木脂素组分的化学预防性能延伸到保护免受石棉诱导的组织和细胞损伤。亚麻籽因此可以用作恶性间皮瘤的化学预防中的饮食剂。
实例6
体内sdg给予的优化剂量和动力学
在本实例中,在通过口服灌胃以水溶性形式给予sdg的小鼠中,呈现了关于药物代谢动力学、生物利用度、和剂量应答的数据。
剂量研究:向小鼠口服给予5、25、50和100mg/kg体重的sdg剂量(n=5/剂量),并且4小时后收集组织用于分析。通过液相色谱和质谱分析sdg及其生物活性代谢物ed、el和seco的血浆浓度,并以ng/ml表示(图39),而nrf2调节的抗氧化剂酶(aoe)(sdg及其代谢物的细胞内靶标)的基因表达水平,使用qrt-pcr从相同动物的肺组织中进行确定(图40)。ho1、nqo1和gst的基因表达水平增加2-3倍,以少至5mgsdg/kg,并且相对于基线达到平均6倍的增加,以100mg/kgsdg。基因水平由肺组织中蛋白质水平的更低但仍显著增加支持。
药代动力学研究:在选择100mg/kg剂量作为诱导显著的aoe表达,同时允许在循环中检测sdg的剂量后,进行药代动力学研究以确定靶组织(即,肺)中的生物利用度和生物效应。在15min、30min、和1、2、4、6、8、12小时收集血液样品以及组织(肺、脑、肝、肾、脾)。如上在血浆和肺中确定sdg水平(图41)。单次剂量100mgsdg/kg被良好吸收,并且在循环中在给予后长达6小时(图41a)和冲洗的、无血的肺组织中持续4小时可检测到完整的sdg(图41b)。sdg的血浆和肺组织浓度分别在给予后30分钟达到0.8和12.6μm的水平。值得注意的是,代表性的aoe基因表达水平(ho-1、nqo1、gst)的强烈诱导显著升高(p<0.05)超过基线(图41c)。观察到的基因表达增加与通过蛋白质印迹确定的蛋白质水平的增加相关(未显示)。重要的是,以相同剂量给予每日两次的sdg持续7天显示无不耐受性也无毒性。这些数据表明,通过这种可溶性sdg形式可以获得显著的生物利用度和效率。因此,大大促进了sdg在人类治疗中的用途,并且不太可能发生毒性风险。因此,50-100mg/kg(每天1或2mgsdg)的剂量足以在靶组织中诱导预期的保护作用,并且可用于进一步研究。
实例7
在nrf2缺陷小鼠中bap诱导的肺肿瘤发生中的sdg对致癌作用的起始或促进阶段的影响。
来自肺损伤模型的数据显示,fs和fs衍生的sdg在饮食中可以上调肺中的nrf2和nrf2激活的基因和蛋白质。在本实例中,使用nrf2敲除小鼠,测试了这种途径的活化是sdg的化学预防效应的重要机制的假说,并且确定sdg在致癌作用的起始和促进阶段是否具有活性。
sdg在nrf2缺陷小鼠中的作用。
在这些实验中,将野生型a/j小鼠中在bap诱导的肺癌模型中口服给予的合成sdg的作用与在a/j背景(目前保持在我们的动物设施处的集群)上回交的nrf2缺陷小鼠进行比较。简言之,对一组野生型(wt)小鼠和一组敲除(ko)小鼠喂食对照饮食。一组wt小鼠和一组ko小鼠通过其饮用水给予sdg,以达到50和100mg/kg的每日sdg消耗(以测试在抑制肺癌发展中剂量应答关系),如图42a中所示。sdg给药研究和动力学确定sdg应当给予持续至少几天,因为这被确定为饮食在组织中达到稳定状态所需的最小时间,随后4次每周注射i.p.给予的1mg/小鼠苯并[a]芘(bap)。口服(口服灌胃或在饮用水中)50和100mg/kgsdg均可诱导肺中的期酶表达(参见图41)。在4、6和9个月处死小鼠(参见图42b),并且收获肺组织,用于:a)肿瘤负荷的组织学评估和通过图像分析进行定量,b)nrf2调节的aoe表达的蛋白质印迹检测和氧化应激,c)在小鼠组织和尿中的8-氧代-7,8-二氢-2’-脱氧鸟苷(8-氧代dguo)水平,和d)dna加合物形成。使用以下各项评估小鼠的亚组的肺炎症:a)支气管肺泡灌洗,和b)使用分别针对cd11b/ly6g、cd11b/f4/80、和抗-cd3的抗体对炎性细胞(如嗜中性粒细胞、活化的巨噬细胞和t细胞)的facs分析。
统计:为了达到统计显著性每组需要的并且建议20只小鼠/组的小鼠的最小数量。
分析:如上所述(图29和31),在获得50和100mg/kgsdg的wt小鼠中预期肿瘤的数量和大小的减少,然而在100mg/kg剂量的情况下,结果更深刻(更少的肿瘤)。在与对照饮食的wt小鼠相比时,由于丧失了使bap解毒的能力,预期在获得对照(无药物)的nrf2ko小鼠中更高数量的肿瘤。然而,预期在接受sdg的nfr2ko小鼠中没有肿瘤减少(与ko小鼠没有药物相比),因为sdg的主要作用是通过nrf2激活介导的。在nrf2/sdg小鼠的肿瘤负荷中观察到的任何减少都归因于也有贡献的其他机制,如sdg的直接自由基清除效应,其应通过测量氧化应激的标志物来检测。
sdg和nrf2激活在致癌作用的起始和/或促进阶段中的作用。
预期上述数据将提供关于在bap的起始和促进阶段期间的sdg给予的明确机械的数据。还感兴趣的是确定在刚刚起始阶段(图42a,计划b)期间或在刚刚促进阶段期间(图42a,计划c)的sdg补充剂的功效。因此,使用这些不同的进食计划在野生型a/j小鼠中进行此图解中概述的实验。
分析:在促进或起始期间给予sdg时,预期肿瘤数量和大小减少。如上所述在nrf2ko小鼠中重复研究,以定义ii期酶上调对该活性的贡献。最近已报道nrf2在致癌作用期间具有两个作用,其中一个在肿瘤起始期间是预防的,并且第二个是促进恶性进展。这些发现表明使用nrf2预防肺癌的恶性进展,而nrf2激活剂更适合于肺癌预防。预期sdg作为化学预防剂是有效的。尽管nrf2在sdg的抗癌作用中起作用,但是可能还涉及其他机制或甚至更重要,如全身流体(血液)或肺组织中mirna的调节。
向吸烟者给予sdg补充剂对氧化应激的全身和肺特异性生物标志物的影响。
设计了具有遗传和生化终点的sdg的口服给予的临床研究,这些终点是遗传和生化的。
所选人群:第一组将由从未吸烟的正常健康志愿者组成。该组允许评估感兴趣的基因和氧化性生物标志物的真实基线水平(可能低),以便评估在“非刺激”环境中由sdg补充剂诱导的变化。第二组受试者将由目前主动的吸烟者组成。选择该组是因为:i)它们处于活动性癌症初始的高风险中并且是化学预防试验中的潜在受试者,并且ii)先前已经显示吸烟诱导在呼吸上皮中的其自身基因组改变,以及活性氧化应激。该组允许吸烟者中的基因组和氧化应激标志物的作图,并且以确定在已经“活化的”环境中由sdg诱导的什么基因组改变。该组还允许在该高风险群体中通过sdg确定升高的氧化应激标志物的减少。选择交叉设计,其中每个患者还将摄入在明胶胶囊剂中给予的安慰剂对照和sdg补充剂(参见图43)。
剂量:选择商业配制品(brevailtm)以避免在不同批次的原或研磨的亚麻籽观的情况下察到的sdg含量和生物利用度的显著变异性。可替代地,将使用合成的sdg如米什拉(mishra)等人,生物有机化学与医药化学通讯(bioorganic&medicinalchemistryletters)2013,(19):5325所述的合成的sdg。药理研究表明,每天给药包含50mg的sdg的这种配制品产生与病例对照研究中癌症发病率的降低相关的最高五分位数中发现的enl水平类似的enl水平(中值,63nmol/l)。
临床研究:该研究将是一个单中心,随机,双盲,两期交叉试验。将存在两个3周的治疗期,其间穿插有一个月的清除期(图43)。交叉设计的原理是受试者中的治疗的比较从比较中去除任何“主体效应”,并提高研究的效率和效能。将二十(20)名健康志愿者(10名吸烟者和10名终身不吸烟者)随机化来摄取以下各项,按次序,或者是每天50mgsdg(一个胶囊剂brevailtm或合成sdg)持续3周,随后1个月的清除期并且然后安慰剂(由相同提供者(木质素研究,圣迭哥,加利福尼亚州)提供的没有sdg的胶囊剂)再持续3周,或者是安慰剂持续3周,一个月的清除,并且然后50mgsdg持续3周。在基线和第2、3、7、9和10周获得口腔上皮(通过口腔拭子获得)、呼气冷凝液、尿和血液。主动吸烟状态将通过尿液可替宁测定来确认。该研究将由佩恩(penn)机构审查委员会批准,并且所有参与者将提供书面知情同意书。将在基线处给予包括综合吸烟史和环境烟草暴露(ets)的详细问卷。参加者将每周参加,用于评估副作用,遵守测试剂摄取,收集间歇的烟草和药物使用的数据以及样品收集。
统计原理/效能分析:感兴趣的主要比较是尿氧化应激的变化,如通过尿8-氧代-7,8-二氢-2’-脱氧鸟苷(8-氧代-dguo),呼气冷凝液的变化(探索性),以及在sdg治疗后研究受试者的颊上皮中基因表达水平的变化所测量的。预期20名患者为这项探索性研究提供合理的效能。由于本研究的性能,校正不适用于多重比较。本研究被设计来获得sdg补充剂的效果的评估。还要注意,存在有限的效能来测试遗留效应,但4周的清除期应避免这些关注点。
口服sdg摄入后颊拭子上皮细胞中基因表达变化的评估
在这些实验中,使用颊粘膜上皮细胞作为支气管上皮组织的替代组织。这类细胞比需要侵入性支气管镜检查的支气管上皮细胞更容易获得(即,通过使用简单的颊擦拭)。这种方法在人类中的有效性由以下证据支持,颊上皮可以作为基因阵列研究中气道支气管组织的替代组织。
这种方法进一步使用小鼠中所摄入的fs进行验证。在对照或10%fs饮食喂养小鼠三周后,收获一定范围的组织,并通过免疫印迹分析两种代表性nrf-2诱导蛋白(ho-1和nqo1)的表达。观察到肺和肝脏中这些标志物的显著增加(数据未显示)。重要的是,观察到两种蛋白质的鼻上皮的明显可检测的增加(图44),因此验证鼻咽组织也可以用作肺的替代物。在人类中也有一些可行性数据支持这种方法。对2名正常志愿者给予整粒fs(每日40g)持续一周。取颊拭子,成功提取rna,并且制备cdna。测量nrf2依赖性基因ho-1的相对表达水平。如图45中所示,ho-1mrna表达在一个受试者中增加了34%,在第二个中增加了63%(p<0.05)。这些数据表明这种方法是可行的,并且fs和sdg促进正常健康志愿者中的nrf2依赖基因。
在试验中,在基线和在纤维对照和fs饮食的2和4周后,在对照和吸烟受试者中取颊拭子。如上产生mrna和cdna,并且使经受rt-pcr以评估代表性抗氧化剂和ii期药物代谢酶的相对表达水平,这些酶包括nqo-1、ho-1和谷胱甘肽-s-转移酶(gst)。除了是nrf2诱导酶的“经典”代表之外,所选择的酶也可以发挥重要的机械作用。nqo1在烟草烟雾中存在的促致癌物的解毒和活化中起双重作用。nqo1的变体基因型与日本受试者中肺癌风险降低显著相关。在遗传研究中,ho-1也被认为是针对香烟烟雾中的氧化剂和芳香烃的细胞保护剂。因此,ho-1也代表了用于监测亚麻籽效应的重要生物标志物。许多研究已经显示gst多态性,吸烟和肺癌之间的关系。重要的是,肺细胞中苯并[a]芘衍生的dna加合物通过由细胞色素p-450-和醛酮还原酶介导的代谢产生的反应性中间体的解毒通过肺特异性gst来调节。由于苯并[a]芘是一种重要的肺致癌物,并且它存在于烟草烟雾中,在吸烟者中gst被监测为亚麻籽的作用的生物标志物。
数据分析:使源自5个正常的非吸烟志愿者的颊拭子的大量cdna库用作所有分析的“基线”比较器。使用β-肌动蛋白和gapdh将来自所有试验样品的cdna标准化至该库。一旦标准化,将每个研究样品与基线比较器进行比较,并计算百分比或倍数变化。主要分析是比较在从不吸烟者和当前吸烟者中sdg摄取的3周之前和之后的基因表达的变化(使用配对t检验)。还将这些值与其安慰剂对照样品中的值进行比较。另外的分析将所有时间点的表达变化与sdg进行比较,包括冲洗值(重复测量anova),以及在基线和sdg和安慰剂之后非吸烟者和吸烟者之间的比较。比较2与3周数据和3周与冲洗数据以建立应答的动力学。
确定同一患者群体中通过sdg的氧化应激的降低
虽然难以测量人类受试者的氧化应激,但最成熟的方法之一是测量isop的尿水平。已经开发了用于这种标志物的可靠和可重复的测定。数据获自等待肺移植的患者,该患者摄取40gfs长达13周。发现在fs时,尿isop稳定下降,表明氧化应激的全身降低(一些isop亚类降低至fs前水平的26%)。代表性患者的数据显示在图46中。bap,香烟烟雾中的致癌物,引起活性氧簇介导的dna链断裂和8-氧代-7,8-二氢-2'-脱氧鸟苷(8-氧代-dguo)形成。可以使用lc-mrm/ms在生物样品(尿、肝、肺)中检测8-氧代-dguo。来自2名人类志愿者的数据表明,每日消耗40gfs持续3周诱导8-氧代-dguo减少的趋势,在同样饮食的肺移植患者(n=5)中也观察到趋势(图47)。
试验:在试验中,在对照和吸烟受试者中在多个时间点采集尿液样品。以盲法方式测量尿液的isop和8-氧代-dguo的水平。
用于评估肺氧化应激的呼气冷凝液:考虑到在这些受试者中进行侵入性测试像支气管镜检查是不可行的,使用一种非侵入性样品收集技术,呼气冷凝液(ebc)用于调研肺氧化剂应激。在呼气中发现许多物质,这些物质在通过冷却(即,冷凝)可获得的液体中是可检测的。这种方法的优点是它作为实时分析和评估肺中氧化应激生物标志物的非侵入性采样方法是非侵入性的并且是方便的。氧化应激的生物标志物包括:h2o2、异前列烷(isop)、丙二醛、4-羟基-2-壬烯醛、抗氧化剂、谷胱甘肽和亚硝化应激标志物。ebc中的异前列烷水平被评估为用于研究fs对肺氧化应激的作用的有用的非侵入性方法。从对照受试者和吸烟者收集的ebc来测量8-isop。非吸烟者的水平一致低-接近测试的灵敏度(1.2pg/ml+/-0.6,n=8),表明ebc在非吸烟者中没有用。虽然只有四个吸烟者测量水平,有趣的是,这些受试者中的两个具有高得多的水平:16pg/ml和6.5pg/ml。因此,在那些具有高基线水平的吸烟者中,ebc用于跟踪肺特异性氧化应激。
数据分析。确定每个受试者的基线值,并且使用配对t检验评估在治疗结束时(3周)观察到的变化。另外的分析将所有时间点的表达变化与sdg进行比较,包括冲洗值(重复测量anova),以及在基线和sdg之后非吸烟者和吸烟者之间的比较。我们可以比较2与3周数据和3周与冲洗数据以建立应答的动力学。
亚麻籽木脂素(sdg)的化学预防性能
在这些实验中,测试了类似于萝卜硫素的sdg可以降低异生物质的毒性并且解毒致癌物(如bap)的假说。首先,评估sdg对支气管上皮细胞的体外作用。针对下述所有sdg剂量进行细胞毒性试验,如mtt测定。第二,评估在bap小鼠模型中口服给予的sdg。
评估在正常小鼠和人原发性肺上皮细胞中sdg介导的bap解毒作用和nrf2介导的ii期酶表达的诱导。
当由上皮细胞代谢激活时,bap(10或20μm)以剂量和时间依赖性方式诱导ros(图23)。针对小鼠研究,使用永生化的c10小鼠支气管细胞系(来源于balb/c小鼠),假定这些细胞是非致瘤性的,接触抑制的,并且包含所有被研究的nrf2调节机构。使用a549细胞进行类似的研究(用于模拟人类1型肺泡上皮细胞的转化的但高度分化的肺癌细胞系-参见图22-23中的数据)并且重要的是原代人支气管上皮细胞。
为了评估直接的ros拦截,将一系列临床相关的sdg浓缩物(0.1-10μm)与10μmb[a]p同时加入,并且测量代表直接抗氧化活性的h2dcf荧光的减少。液相色谱/多反应监测质谱(lc/mrm-ms)也用于确定gssg:gsh比率。bap还可以快速诱导dna加合物形成,该形成可以使用lc-mrm/ms进行测量(图22)。由bap引起的bap诱导的氧化性dna损伤可以通过8-氧代-dguo形成,使用标准comet图像分析的单个细胞的尾力矩(数据未显示),或用于检测γh2ax的免疫印迹的密度计量分析(数据未显示)来测量。为了评估dna加合物和氧化变化,将一些列sdg浓缩物与10μmb[a]p同时添加,并且测量这些参数。此外,进行mtt细胞毒性测定以评估在致癌物不存在和存在下对这些细胞的直接sdg效应(fs和flc只能在体内测试)。接下来,测试纯化的sdg上调支气管上皮细胞中的ii期酶的能力。具体地,测量用木脂素sdg(0.1-10μm)处理孵育0、1、2、4、24、48和72小时后的gst-ya和nqo-1信息和蛋白的诱导。这些酶被选择作为ii期系统的特征代表性are调节酶。进行rt-pcr和免疫印迹分析。在鉴定ii期酶上调的时间过程后,用sdg预处理上皮细胞,并在最大ii期诱导的时间点添加bap。使用上述测定评估氧化应激,dna损伤和细胞死亡。
数据分析,预期结果和其他研究:第一组研究定义了sdg的直接抗氧化剂作用。在sdg自身消失(因此没有直接的抗氧化剂效应存在)的稍后时间点进行的第二组研究,定义了sdg由于其ii期酶诱导效应的影响。进行氧化应激和炎症的另外的测量以确定通过sdg血浆氧化应激测量(血浆丙二醛)和促炎应激标志物(如il-6、il-1α、il1β、tnf-α、c反应蛋白)的减少。
在a/j小鼠-化学致癌物诱导肺肿瘤发生的鼠模型中,sdg的化学预防性能。
在上述相同的研究设计之后,在a/j小鼠肺中研究口服给予的sdg对bap诱导的致癌作用的进展的影响(图42a,计划a)。将sdg补充的小鼠与对照直接比较。进行的后续研究包括:仅在起始阶段使用sdg的研究(图42a,计划b)和仅在促进阶段使用sdg的研究(图22a,计划c)。
统计:动物研究通常包含多于20只小鼠的两组(例如,对照与50mg/kgsdg),并且使用anova或其他合适的线性模型进行分析。所有计算假设为双侧检验,α水平为0.05,并且检验力至少为80%。
表1:将在3个实验计划(参见图42a:计划a、b、c)上放置的3个测试组的小鼠在4、6和9个月(3个时间点)处死,即,3×3×3×20个小鼠=540个小鼠。
实例8
sdg对石棉诱导的致癌作用的影响。
sdg或亚麻籽饮食被假设为减少石棉诱导的ros/炎症,导致:1)ros,2)减少细胞因子;3)减少hmgb1;4)更少的肿瘤发生病灶;和5)更少的肿瘤。下划线假说是减少的炎症和氧化应激将导致细胞的恶性转化减少和更少的肿瘤负荷(参见图48)。
为了测试亚麻籽木脂素sdg将减少石棉诱导的炎症以及氧化和亚硝化应激的假设,将小鼠腹膜巨噬细胞暴露于不同浓度的青石棉石棉纤维(10、20、30、和40ng/cm2)。在石棉暴露(3、4、6和8小时)后的可变时间处,将细胞用sdg(50μm)孵育,并且24小时后收集上清液以检测炎症细胞因子分泌和亚硝化/氧化应激(图49)。
sdg在体外通过人类间皮细胞钝化石棉诱导的ros分泌(图50)。检测细胞中石棉诱导的ros的实验计划:(人间皮细胞)hm细胞或小鼠巨噬细胞在20%dmem中接种24小时;在hbss中添加20μmdcf持续1小时;用包含h2o2(100μm),石棉(10μg青石棉/cm2)和/或sdg(50μm)的1%dmem替代上清液;并且在开始石棉暴露后监测荧光水平长达36小时。结果表明,石棉和h2o2诱导高水平的ros,然而在介质中添加sdg显著降低ros水平至基线水平(图50)。
评估培养物raw巨噬细胞中的石棉诱导的氧化应激(ros释放)。将细胞用ros敏感染料h2dcfda处理30min,并且然后将它们暴露于载体至40μg/cm2石棉纤维或4um次氯酸盐溶液,并用分光光度法测量荧光强度持续90、150分钟和27小时。石棉诱导的ros在石棉暴露后不久产生,并在观察期间持续(图51)。
在暴露于石棉后几小时向巨噬细胞给予sdg降低氧化应激(图52)。雌性c57/bl6小鼠注射2ml巯基乙酸盐,并且3天后收获腹膜巨噬细胞。将2百万个细胞/孔接种在6孔板中并暴露于20μg/cm2石棉。在石棉暴露后3、4、6或8小时用25或50μmsdg(合成sdg)处理细胞。在石棉暴露后24小时收获细胞。对冷冻上清液样品进行分析。结果显示,丙二醛,脂质过氧化的标志物,随着时间的推移随着石棉暴露而增加(图52a)。在将25和50μmsdg添加稻细胞中的情况下,mda的水平显著降低(p<0.05)(图52b)。
暴露于石棉后若干小时向巨噬细胞给予的sdg减少亚硝酸应激(图53)。为了评估在石棉暴露后细胞中的钝化亚硝化应激中的sdg,向雌性c57/bl6小鼠注射2ml巯基乙酸盐并且在3天后收获的腹膜巨噬细胞。将2百万个细胞/孔接种在6孔板中并暴露于20μg/cm2石棉。在石棉暴露后3、4、6或8小时用25或50μmsdg(合成sdg)处理细胞。在石棉暴露后24小时收获细胞。对冷冻上清液样品进行分析。结果显示,亚硝酸盐,亚硝化应激的标志物,随着时间的推移随着石棉暴露而增加(图53a)。在将25和50μmsdg添加稻细胞中的情况下,mda的水平显著降低(p<0.05)(图53b)。
暴露于石棉后若干小时向巨噬细胞给予的sdg减少炎症细胞因子分泌(il-1β)(图54)。为了评估在石棉暴露后细胞中钝性炎症细胞因子分泌(il-1β)中的sdg,向雌性c57/bl6小鼠注射2ml巯基乙酸盐和在3天后收获的腹膜巨噬细胞。将2百万个细胞/孔接种在6孔板中并暴露于20μg/cm2石棉。在石棉暴露后3、4、6或8小时用25或50μmsdg(合成sdg)处理细胞。在石棉暴露后24小时收获细胞。对新鲜上清液样品进行分析。结果显示il-1β,促炎细胞因子,随着时间的推移随着石棉暴露而增加(图54a)。在将25和50μmsdg添加稻细胞中的情况下,il1β的水平显著降低(p<0.05)(图54b)。
暴露于石棉后若干小时向巨噬细胞给予的sdg减少炎症细胞因子分泌(tnf-α)(图55)。为了评估在石棉暴露后细胞中钝性炎症细胞因子分泌(tnf-α)中的sdg,向雌性c57/bl6小鼠注射2ml巯基乙酸盐和在3天后收获的腹膜巨噬细胞。将2百万个细胞/孔接种在6孔板中并暴露于20μg/cm2石棉。在石棉暴露后3、4、6或8小时用25或50μmsdg(合成sdg)处理细胞。在石棉暴露后24小时收获细胞。对新鲜上清液样品进行分析。结果显示tnf-α,促炎细胞因子,随着时间的推移随着石棉暴露而增加(图55a)。在将25和50μmsdg添加稻细胞中的情况下,tnf-α的水平显著降低(p<0.05)(图55b)。
简言之,本实例中的体外实验证明(1)sdg阻断人类间皮细胞和小鼠raw巨噬细胞中的石棉诱导的ros;(2)sdg阻断通过暴露于石棉的小鼠腹膜巨噬细胞的炎性细胞因子分泌;和(3)sdg阻断暴露于石棉的小鼠腹膜巨噬细胞中的氧化(脂质过氧化)和亚硝化应激(亚硝酸盐水平)。这些体外实验支持体内实验以确定sdg在钝性慢性炎症中的有用性并且最终确定由于石棉暴露导致的恶性肿瘤。
因此,使用两种小鼠模型在石棉诱导的间皮瘤中测试sdg,其中该小鼠在石棉暴露后遗传倾向形成间皮瘤。使用这些模型,评估亚麻籽和sdg对单一剂量的石棉在小鼠中的急性效应,以及测试亚麻籽和sdg是否在这些加速的石棉诱导mm模型中抑制肿瘤的发展(图56)。
sdg富集的亚麻籽木脂素饮食(flc)钝化mextag小鼠中的石棉诱导的腹部炎症(图57)。简言之,向6只雄性mextag小鼠(10-12周龄)ip注射400μg的青石棉石棉,体积为0.5ml。在石棉注射之前,将半数小鼠置于富含sdg(35%sdg)的flc饮食上持续两周-其他小鼠在标准饮食上。3天后,用5ml的pbs灌洗小鼠,并且进行灌洗用于sups和细胞计数。进行细胞离心涂片并针对差异,计数3-5个独立的视野-计算总细胞的%。与0%fs饲喂的小鼠相比,喂食10%flc的小鼠在腹部灌洗液wbc中具有26%的减少(p=0.014)(图57)。
在7周龄、雄性、暴露于石棉的nf2(129sv)(+/-)小鼠中进行急性期研究,以评估在饮食中给予的亚麻籽和木脂素sdg配制品的作用。nf2小鼠的石棉暴露的实验计划和亚麻籽/sdg木脂素配制品评估示于图58中。简言之,在第0天通过腹膜内注射将雄性nf2(129sv)(+/-)暴露于400μg石棉。小鼠在石棉暴露前24小时(第-1天)在测试饮食(0%fs、10%fs、10%flc;n=2只小鼠/组)上开始,并在石棉暴露后第3天处死。使用5ml的1×pbs进行腹部灌洗(al)。炎症细胞流入在3天达到峰值,并在石棉暴露后9天逐渐减少。因此,选择3天作为在所有后续实验中评估炎症的时间点(图59)。
亚麻籽及其富含sdg的木脂素成分钝化石棉诱导的炎症(年轻小鼠)(图60)。在饮食中fs或flc添加的情况下,总白细胞(图60a)降低,虽然不显著。然而,当观察细胞差异,并且特别是巨噬细胞水平时,水平显著通过亚麻籽和sdg-木脂素饮食减弱(图60b)。
亚麻籽及其富含sdg的木脂素成分钝化石棉诱导的炎症(老年小鼠)(图61)。与仅300,000个细胞/ml(高出10倍)相比,通过呈现约3,000,000wbc/ml的腹部灌洗液,暴露于腹部石棉的老龄小鼠(图61a)对石棉更敏感。结果表明,炎症细胞嗜中性粒细胞(图61b)和巨噬细胞(图61c)在老龄小鼠中二者都显著高于年轻小鼠。
图62:富含sdg的亚麻籽木脂素提取物(在饮食配制品中给出)使老年小鼠中石棉炎症钝化(图62)。在第0天用400μg石棉注射(经腹膜内)雄性nf2(129sv)(+/-)小鼠。小鼠在石棉暴露前一周(第-7天)在测试饮食(0%fs或10%flc)上开始,并在石棉暴露后第3天处死。用5ml1×pbs进行腹部灌洗(al)(将1ml腹部灌洗液离心,并且冷冻上清液)。在-80°下冷冻收集血浆。在灌洗液中评估细胞,并且显示通过富含sdg的饮食,总wbc和嗜中性粒细胞、巨噬细胞和嗜酸性粒细胞都显著减少。
富含sdg的亚麻籽木脂素提取物(在饮食配制品中给出)使老年小鼠中石棉炎性细胞因子分泌和亚硝化应激钝化(图63)。在第0天用400μg石棉注射(经腹膜内)雄性nf2(129sv)(+/-)小鼠。小鼠在石棉暴露前一周(第7天)在测试饮食(0%fs或10%flc)上开始,并在石棉暴露后第3天处死。用5ml1×pbs进行腹部灌洗(al)(将1ml腹部灌洗液离心,并且冷冻上清液)。在-80°下冷冻收集血浆。细胞因子il1β和tnfα以及亚硝酸盐的水平被富含sdg的饮食显著钝化。
简言之,这些用nf2小鼠的体内实验证明:(1)饲喂富含sdg的饮食的小鼠具有显著减少的腹部炎症,如通过腹部灌洗液wbc所确定;(2)sdg富集的饮食减少腹部灌洗液中嗜中性粒细胞的数量;(3)在饲喂sdg富集的小鼠中,促炎细胞因子、il-1β和tnfα的水平降低;并且(4)富含sdg的饮食喂养的小鼠具有更低水平的腹部灌洗液亚硝酸盐,表示减少由暴露于石棉纤维诱导的亚硝化应激。
这些实例中的这些研究证明sdg的化学预防活性。进行更大规模的生物标志物研究以评估颊上皮中的ii期酶的诱导和通过sdg的氧化应激减少。在暴露于致癌物的受试者(例如,前体或当前吸烟者)中,在口服sdg每天给予后,进行另外的测量,测量氧化应激和炎症,并且包括血浆氧化应激测量(血浆丙二醛)和促炎应激标志物(如il-6、il-1α、il1β、tnf-α、c-反应蛋白、f2-异前列烷)。
尽管本文已举例说明并描述了本发明的某些特征,但是本领域普通技术人员会想到许多修改、替换、改变和等同。因此,应当理解,所附权利要求旨在覆盖落入本发明的真实精神内的所有这些修改和改变。
- 该技术已申请专利。仅供学习研究,如用于商业用途,请联系技术所有人。
- 技术研发人员:迈尔波·赫里斯托费多-所罗米多
- 技术所有人:宾夕法尼亚大学董事会
- 我是此专利的发明人
- 该领域下的技术专家
- 如您需求助技术专家,请点此查看客服电话进行咨询。
- 1、司老师:1.制浆造纸 2.植物资源精细化工与化学 3.生物质精炼 4.天然产物化学
- 2、薛老师:1.CRISPR-Cas系统 2.基因编辑 3.基因修复 4.天然产物合成 5.单分子技术开发与应用
- 3、戴老师:1.天然药物(中药)合成生物学研究 2.酵母生物学与工程化研究
- 4、孟老师:1. 基于糖类的抗肿瘤药物的合成和活性评价及糖类疫苗的研制 2.功能糖类的化学酶法合成及构效关系研究 3.多糖及仿生材料功能的开发及应用
- 5、满老师:1.天然产品的提取分离与活性研究 2.天然产物活性与安全性评价 3.中药组方配伍机制研究
- 如您是高校老师,可以点此联系我们加入专家库。
- 还没有人留言评论。精彩留言会获得点赞!