预防和治疗组织器官纤维化的药物及其用途的制作方法
本发明涉及一种预防和治疗组织器官纤维化的方法,包括给药受试者有效量的纤溶酶原。
背景技术:
纤维化是一种病理变化,表现为成纤维细胞激活增殖、组织器官内纤维结缔组织增多,实质细胞减少,持续进展可致组织、器官结构破坏和功能丧失。重要脏器的纤维化严重影响患者生存质量,甚至危及生命。在全世界范围内,组织纤维化是许多疾病致残、致死的主要原因,据美国有关统计资料证明,该国因各种疾病而致死的患者中,约45%可以归于组织纤维增生疾病。
纤维化疾病包括累及多系统的疾病,如系统性硬化症、多灶性纤维化、硬皮病、肾源性的多系统纤维化,也包括器官组织特异性疾病,如皮肤、心、肺、肝、肾纤维化等。不同纤维化疾病的病因各不相同,例如组织器官的损伤、感染、免疫反应、慢性炎症等,但其共同的特征是细胞外基质(ECM)在组织中的过度沉积和器官组织重构[1-3]。
心脏纤维化出现于多种心脏疾病的发生与发展过程中,如病毒性心肌炎、心肌梗死和高血压心脏病等。过度的心脏纤维化导致心脏功能受损,是多种疾病走向心衰的主要原因,其中抑制和逆转心肌纤维化己成为治疗心血管疾病的重要环节。
肝纤维化是指由各种致病因子所致肝内结缔组织异常增生,肝内弥漫性细胞外基质过度沉淀的病理过程。多种因素均可引起肝纤维化,如病毒感染、炎症反应、氧化应激和酗酒等。肝纤维化的病理特点为汇管区和肝小叶内有大量纤维组织增生和沉积,但尚未形成小叶内间隔,肝硬化则有假小叶形成,中心静脉区和汇管区出现间隔,肝的正常结构遭到破坏,肝纤维化进一步发展即为肝硬化。我国的慢性肝病中以病毒性肝炎为主,慢性病毒性肝炎的肝组织纤维化与肝内炎症、坏死、病毒复制等有关,而且在早期是可逆的。为此将抗病毒治疗,调整机体免疫功能及保护肝细胞等疗法与抗纤维化结合起来,是防止肝纤维化的积极措施。
肺纤维化疾病包括特发性肺纤维化、结节病、过敏性肺炎、尘肺、药物和放射线导致的纤维化,以及与胶原血管病有关的致纤维化肺泡炎等病因各异、范围广泛的疾病谱。其主要病理特点包括肺组织间充质细胞增殖、细胞外基质增生沉积及肺实质的重构等画。目前主要采用抗炎、抗氧化、抗成纤维细胞增殖和胶原沉积及肺移植等措施治疗肺纤维化。
肾纤维化表现为细胞外基质和不适当结缔组织在肾聚集,导致肾结构改变和功能受损的病理过程,也是几乎所有肾疾病进展到终末期肾衰的共同通路。肾纤维化过程涉及炎症反应,固有细胞和免疫细胞的凋亡以及多种调控纤维化因子失衡等,故可通过抗炎症、抗凋亡和针对纤维化因子治疗等途径抗御肾纤维化。
组织器官的慢性病变常伴有纤维化,例如肺的慢性炎症,慢性病变,伴有肺纤维化。肝纤维化也是这样,像乙肝、丙肝、酒精肝、脂肪肝、血吸虫病等都伴有早期肝纤维化。肝的代偿功能很强,纤维化病变一直潜伏在各种慢性肝病里,一般发展为肝硬化时才被发现,实际上肝硬化已经是肝纤维化的严重阶段。再像慢性肾炎、肾小球炎、肾小管炎等都有肾纤维化;心血管、脑血管、下肢血管硬化、或变窄、或阻塞情况下都有血管纤维化。
皮肤纤维化形成瘢痕组织。瘢痕组织是肉芽组织经改建成熟形成的老化阶段的纤维结缔组织。创伤等情况下,成纤维细胞分裂、增殖,向受损部位迁移,产生细胞外基质,形成瘢痕组织,修复创伤。
瘢痕的形成是肉芽组织逐渐纤维化的过程。此时网状纤维及胶原纤维越来越多,网状纤维胶原化,胶原纤维变粗,与此同时纤维母细胞越来越少,少量剩下者转变为纤维细胞;间质中液体逐渐被吸收,中性粒细胞、巨噬细胞、淋巴细胞和浆细胞先后消失;毛细血管闭合、退化、消失,留下很少的小动脉及小静脉。这样,肉芽组织乃转变成主要由胶原纤维组成的血管稀少的瘢痕组织,肉眼呈白色,质地坚韧。
由于瘢痕坚韧又缺乏弹性,加上瘢痕收缩可引起器官变形及功能障碍,所以发生在关节附近和重要脏器的瘢痕,常引起关节痉挛或活动受限,如在消化道、泌尿道等腔室器官则引起管腔狭窄,在关节附近则引起运动障碍。发生在器官之间或器官与体腔壁之间的瘢痕性粘连,常不同程度地影响其功能。如器官内广泛损伤后发生广泛纤维化、玻璃样变,则导致器官硬化。
系统性硬化症(SSc)也称为硬皮病,是一种以局限性或弥漫性皮肤增厚和纤维化为特征的全身性自身免疫病。病变特点为皮肤纤维增生及血管洋葱皮样改变,最终导致皮肤硬化、血管缺血。本病临床上以局限性或弥漫性皮肤增厚和纤维化为特征,除皮肤受累外,它也可影响内脏(心、肺和消化道等器官)。
动脉粥样硬化常常导致组织、器官缺血损伤,继而引发组织器官的纤维化病变。动脉粥样硬化是一种慢性的、渐进性动脉疾病,发病时动脉中沉积的脂肪部分或全部堵塞血流。当原本光滑、坚实的动脉内膜变粗糙、增厚,并被脂肪、纤维蛋白、钙和细胞碎屑堵塞时,便出现动脉粥样硬化。动脉粥样硬化是动脉内膜的慢性炎症性增生病变,导致大、中型动脉管腔狭窄或闭塞,引起相应器官组织缺血损伤、纤维化,甚至坏死。
动脉粥样硬化与糖尿病关系密切,表现为糖尿病患者出现动脉粥样硬化的时间早、程度重和预后差,而动脉粥样硬化又是糖尿病患者的主要死亡原因。临床发现糖尿病患者的冠状动脉血管病理改变的特点主要是病变累及的血管较多、冠状动脉狭窄严重,病变更加弥漫严重,其机制多认为是血糖代谢异常引起动脉粥样硬化,随着更进一步深入的研究,更多的结果表明,糖尿病引起动脉粥样硬化并非单一因素所致,而是通过多种途径以及较为复杂的机制来诱发和促进动脉粥样硬化的发生及发展,例如巨噬细胞极化、巨噬细胞移动抑制因子途径、糖基化终产物途径、清道夫受体上调、胰岛素抵抗、泛素-蛋白酶体系统激活、血小板源性生长因子激活途径等[4]。
目前,各种原因导致的纤维化疾病发病人群众多,患者往往有多个器官组织受累,尚缺乏有效的治疗方法,社会和经济负担较重。本发明研究发现纤溶酶原可以改善组织器官的纤维化,改善组织器官功能,从而为组织器官纤维化及其相关病症的预防和治疗开辟了一个全新的领域。
发明概述
本发明涉及下述各项:
1.一种预防和/或治疗受试者组织器官胶原蛋白沉积或纤维化及其相关病症的方法,包括给药受试者有效量的纤溶酶原,其中所述受试者易患组织器官纤维化、有组织器官纤维化倾向或罹患其它疾病并伴有组织器官纤维化。
2.项1的方法,其中所述组织器官胶原蛋白沉积或纤维化包括皮肤纤维化、血管纤维化、心脏纤维化、肺纤维化、肝纤维化、肾纤维化。
3.项1或2的方法,其中所述组织器官胶原蛋白沉积或纤维化包括感染、炎症、超敏反应、肿瘤、组织缺血、组织器官淤血、化学物质、辐射或环境污染导致的损伤引发的或伴随的。
4.项3的方法,其中所述组织器官胶原蛋白沉积或纤维化包括细菌、病毒或寄生虫感染引起的组织器官病变导致的组织器官胶原蛋白沉积或纤维化。
5.项4的方法,其中所述组织器官胶原蛋白沉积或纤维化包括结核杆菌感染导致的肺纤维化。
6.项4的方法,其中所述组织器官胶原蛋白沉积或纤维化为乙型肝炎病毒、丙型肝炎病毒或戊型肝炎病毒感染导致的肝脏纤维化。
7.项4的方法,其中所述组织器官胶原蛋白沉积或纤维化为血吸虫感染导致的肝纤维化。
8.项3的方法,其中所述组织器官胶原蛋白沉积或纤维化为无菌性炎症或自身免疫反应导致的。
9.项8的方法,其中所述组织器官胶原蛋白沉积或纤维化为慢性肾小球肾炎、肾盂肾炎、肾病综合征、肾功能不全、尿毒症导致的肾脏纤维化。
9.项3的方法,其中所述组织器官胶原蛋白沉积或纤维化为癌症导致组织器官损伤所致的。
10.项9的方法,所述组织器官胶原蛋白沉积或纤维化为肺癌导致的肺纤维化、肝癌导致的肝纤维化或肾脏癌症导致的肾脏纤维化。
11.项3的方法,其中所述组织器官胶原蛋白沉积或纤维化为慢性缺血性组织损伤导致的。
12.项11的方法,其中所述组织器官胶原蛋白沉积或纤维化为冠状动脉粥样硬化、冠心病导致的心脏缺血性纤维化。
13.项11的方法,其中所述组织器官胶原蛋白沉积或纤维化为慢性缺血性肾损伤导致的肾脏纤维化。
14.项3的方法,其中所述组织器官胶原蛋白沉积或纤维化为心血管疾病导致的组织器官淤血导致的。
15.项14的方法,其中所述组织器官胶原蛋白沉积或纤维化为肝淤血或肺淤血。
16.项3的方法,其中所述组织器官胶原蛋白沉积或纤维化为药物导致的。
17.项16的方法,其中所述组织器官胶原蛋白沉积或纤维化为药物性肝纤维化或肾纤维化。
18.项3的方法,其中所述组织器官胶原蛋白沉积或纤维化为吸入性化学物质或环境污染物导致的肺纤维化。
19.项8的方法,其中所述组织器官胶原蛋白沉积或纤维化为诸如系统性红斑狼疮、系统性硬化症、强直性脊柱炎全身性免疫性疾病导致的。
20.项8的方法,其中所述组织器官纤维化为特发性肺纤维化。
21.项1-20任一项的方法,其中所述组织器官纤维化相关病症包括组织器官因纤维化病变导致的功能减弱、障碍或丧失而引发的病症。
22.项21的方法,其中所述组织器官纤维化相关病症包括动脉粥样硬化、冠心病、心绞痛、心肌梗死、心律失常、脑缺血、脑梗塞、肾功能不全、尿毒症、肝功能障碍、肝硬化、肝昏迷、呼吸困难、肺气肿、肺心病、肺纤维化、强直性脊柱炎。
23.根据项1-22任一项的方法,其中所述纤溶酶原可与一种或多种其它药物或治疗手段联合施用。
24.根据项23的方法,其中所述其它药物包括:降血脂药物、抗血小板药物、降血压药物、扩张血管药物、降血糖药物、抗凝血药物、溶血栓药物,保肝药物,抗纤维化药物、抗心律失常药物,强心药物,利尿药物、抗肿瘤药物、放化疗药物、炎症调节药物、免疫调节药物、抗病毒药物、抗生素。
25.项1-24任一项的方法,其中所述纤溶酶原与序列2、6、8、10或 12具有至少75%、80%、85%、90%、95%、96%、97%、98%或99%的序列同一性,并且仍然具有纤溶酶原活性。
26.项1-25任一项的方法,所述纤溶酶原是在序列2、6、8、10或12 的基础上,添加、删除和/或取代1-100、1-90、1-80、1-70、1-60、1-50、 1-45、1-40、1-35、1-30、1-25、1-20、1-15、1-10、1-5、1-4、1-3、1-2、1 个氨基酸,并且仍然具有纤溶酶原活性的蛋白质。
27.项1-26任一项的方法,所述纤溶酶原是包含纤溶酶原活性片段、并且仍然具有纤溶酶原活性的蛋白质。
28.项1-27任一项的方法,所述纤溶酶原选自Glu-纤溶酶原、Lys-纤溶酶原、小纤溶酶原、微纤溶酶原、delta-纤溶酶原或它们的保留纤溶酶原活性的变体。
29.项1-28任一项的方法,所述纤溶酶原为天然或合成的人纤溶酶原、或其仍然保留纤溶酶原活性的变体或片段。
30.项1-28任一项的方法,所述纤溶酶原为来自灵长类动物或啮齿类动物的人纤溶酶原直向同系物或其仍然保留纤溶酶原活性的变体或片段。
31.项1-30任一项的方法,所述纤溶酶原的氨基酸如序列2、6、8、10 或12所示。
32.项1-31任一项的方法,其中所述纤溶酶原是人天然纤溶酶原。
33.项1-32任一项的方法,其中所述受试者是人。
34.项1-33任一项的方法,其中所述受试者缺乏或缺失纤溶酶原。
35.项1-34任一项的方法,所述缺乏或缺失是先天的、继发的和/或局部的。
36.一种用于项1-35任一项的方法的纤溶酶原。
37.一种药物组合物,其包含药学上可接受的载剂和用于项1-35中任一项所述方法的纤溶酶原。
38.一种预防性或治疗性试剂盒,其包含:(i)用于项1-35中任一项所述方法的纤溶酶原和(ii)用于递送所述纤溶酶原至所述受试者的构件(means)。
39.根据项38所述的试剂盒,其中所述构件为注射器或小瓶。
40.项38或39的试剂盒,其还包含标签或使用说明书,该标签或使用说明书指示将所述纤溶酶原投予所述受试者以实施项1-35中任一项所述方法。
41.一种制品,其包含:
含有标签的容器;和
包含(i)用于项1-35中任一项所述方法的纤溶酶原或包含纤溶酶原的药物组合物,其中所述标签指示将所述纤溶酶原或组合物投予所述受试者以实施项1-35中任一项所述方法。
42.项38-40中任一项的试剂盒或项41的制品,还包含另外的一个或多个构件或容器,该构件或容器中含有其他药物。
43.项42的试剂盒或制品,其中所述其他药物选自下组:降血脂药物、抗血小板药物、降血压药物、扩张血管药物、降血糖药物、抗凝血药物、溶血栓药物,保肝药物,抗纤维化药物、抗心律失常药物,强心药物,利尿药物、抗肿瘤药物、放化疗药物、炎症调节药物、免疫调节药物、抗病毒药物、抗生素。
一方面,本发明涉及一种预防和/或治疗受试者组织器官胶原蛋白沉积或纤维化及其相关病症的方法,包括给药受试者有效量的纤溶酶原,其中所述受试者易患组织器官纤维化、有组织器官纤维化倾向或罹患其它疾病并伴有组织器官纤维化。本发明还涉及纤溶酶原用于预防和/或治疗受试者组织器官胶原蛋白沉积或纤维化及其相关病症的用途。本发明还涉及纤溶酶原用于制备预防和/或治疗受试者组织器官胶原蛋白沉积或纤维化及其相关病症的药物的用途。此外,本发明还涉及用于预防和/或治疗受试者组织器官胶原蛋白沉积或纤维化及其相关病症的纤溶酶原。在一些实施方案中,所述组织器官胶原蛋白沉积或纤维化包括皮肤纤维化、血管纤维化、心脏纤维化、肺纤维化、肝纤维化、肾纤维化。在另一些实施方案中,所述组织器官胶原蛋白沉积或纤维化包括感染、炎症、超敏反应、肿瘤、组织缺血、组织器官淤血、化学物质、辐射或环境污染导致的损伤引发的或伴随的。具体地,所述组织器官胶原蛋白沉积或纤维化包括细菌、病毒或寄生虫感染引起的组织器官病变导致的组织器官胶原蛋白沉积或纤维化,其中,所述组织器官胶原蛋白沉积或纤维化包括结核杆菌感染导致的肺纤维化、乙型肝炎病毒、丙型肝炎病毒或戊型肝炎病毒感染导致的肝脏纤维化、血吸虫感染导致的肝纤维化。在一些实施方案中,所述组织器官胶原蛋白沉积或纤维化为无菌性炎症或自身免疫反应导致的。具体地,所述组织器官胶原蛋白沉积或纤维化为慢性肾小球肾炎、肾盂肾炎、肾病综合征、肾功能不全、尿毒症导致的肾脏纤维化。在另一些实施方案中,所述组织器官胶原蛋白沉积或纤维化为癌症导致组织器官损伤所致的。具体地,所述组织器官胶原蛋白沉积或纤维化为肺癌导致的肺纤维化、肝癌导致的肝纤维化或肾脏癌症导致的肾脏纤维化。在另一些实施方案中,所述组织器官胶原蛋白沉积或纤维化为慢性缺血性组织损伤导致的。具体地,所述组织器官胶原蛋白沉积或纤维化为冠状动脉粥样硬化、冠心病导致的心脏缺血性纤维化和/或慢性缺血性肾损伤导致的肾脏纤维化。在另一些实施方案中,所述组织器官胶原蛋白沉积或纤维化为心血管疾病导致的组织器官淤血导致的。具体地,所述组织器官胶原蛋白沉积或纤维化为肝淤血或肺淤血。在一些实施方案中,所述组织器官胶原蛋白沉积或纤维化为药物导致的。具体地,所述组织器官胶原蛋白沉积或纤维化为药物性肝纤维化或肾纤维化。在一些实施方案中,所述组织器官胶原蛋白沉积或纤维化为吸入性化学物质或环境污染物导致的肺纤维化。在上述实施方案中,所述组织器官胶原蛋白沉积或纤维化为诸如系统性红斑狼疮、系统性硬化症、强直性脊柱炎全身性免疫性疾病导致的。在一些实施方案中,所述组织器官纤维化为特发性肺纤维化。
在上述实施方案中,所述组织器官纤维化相关病症包括组织器官因纤维化病变导致的功能减弱、障碍或丧失而引发的病症。具体地,所述组织器官纤维化相关病症包括动脉粥样硬化、冠心病、心绞痛、心肌梗死、心律失常、脑缺血、脑梗塞、肾功能不全、尿毒症、肝功能障碍、肝硬化、肝昏迷、呼吸困难、肺气肿、肺心病、肺纤维化、强直性脊柱炎。
在上述实施方案中,所述纤溶酶原可与一种或多种其它药物或治疗方法联用。具体地,所述纤溶酶原可与一种或多种选自如下的药物联用:降血脂药物、抗血小板药物、降血压药物、扩张血管药物、降血糖药物、抗凝血药物、溶血栓药物,保肝药物,抗纤维化药物、抗心律失常药物,强心药物,利尿药物、抗肿瘤药物、放化疗药物、炎症调节药物、免疫调节药物、抗病毒药物、抗生素。
在上述实施方案中,所述纤溶酶原与序列2、6、8、10或12具有至少 75%、80%、85%、90%、95%、96%、97%、98%或99%的序列同一性,并且仍然具有纤溶酶原活性。
在上述实施方案中,所述纤溶酶原的氨基酸如序列2、6、8、10或12 所示。在一些实施方案中,所述纤溶酶原是在序列2、6、8、10或12的基础上,添加、删除和/或取代1-100、1-90、1-80、1-70、1-60、1-50、1- 45、1-40、1-35、1-30、1-25、1-20、1-15、1-10、1-5、1-4、1-3、1-2、1 个氨基酸,并且仍然具有纤溶酶原活性的蛋白质。
在上述实施方案中,所述纤溶酶原是包含纤溶酶原活性片段、并且仍然具有纤溶酶原活性的蛋白质。具体地,所述纤溶酶原选自Glu-纤溶酶原、Lys-纤溶酶原、小纤溶酶原、微纤溶酶原、delta-纤溶酶原或它们的保留纤溶酶原活性的变体。
在上述实施方案中,纤溶酶原为天然或合成的人纤溶酶原、或其仍然保留纤溶酶原活性的变体或片段。在一些实施方案中,所述纤溶酶原为来自灵长类动物或啮齿类动物的人纤溶酶原直向同系物或其仍然保留纤溶酶原活性的变体或片段。例如,来自灵长类动物或啮齿类动物的纤维蛋白溶酶原直向同系物,例如来自大猩猩,恒河猴、鼠、牛、马,狗的纤维蛋白溶酶原直向同系物。最优选,本发明的纤维蛋白溶酶原的氨基酸序列如序列2、6、8、10或12所示。
在上述实施方案中,所述受试者是人。在一些实施方案中,其中所述受试者缺乏或缺失纤溶酶原。具体地,所述缺乏或缺失是先天的、继发的和/或局部的。
在一个实施方案中,所述纤维蛋白溶酶原通过全身或局部给药,优选通过以下途径施用:表面、静脉内、肌内、皮下、吸入、椎管内、局部注射、关节内注射或通过直肠。在一个实施方案中,所述局部给药为直接向骨质疏松区域给药,例如通过敷料,导管等方式来进行。
在一个实施方案中,所述纤溶酶原与适当的多肽载体或稳定剂组合施用。在一个实施方案中,所述纤溶酶原以每天0.0001-2000mg/kg、0.001- 800mg/kg、0.01-600mg/kg、0.1-400mg/kg、1-200mg/kg、1-100mg/kg、10- 100mg/kg(以每公斤体重计算)或0.0001-2000mg/cm2、0.001-800 mg/cm2、0.01-600mg/cm2、0.1-400mg/cm2、1-200mg/cm2、1-100 mg/cm2、10-100mg/cm2(以每平方厘米体表面积计算)的剂量施用,优选至少重复一次,优选至少每天施用。在局部施用的情况下,上述剂量还可以根据情况进一步调整。一方面,本发明涉及一种药物组合物,其包含药学上可接受的载剂和用于本发明所述方法的纤溶酶原。
另一方面,本发明涉及一种预防性或治疗性试剂盒,其包含:(i)用于本发明所述方法的纤溶酶原和(ii)用于递送所述纤溶酶原至所述受试者的构件(means),具体地,所述构件为注射器或小瓶。在一些实施方案中,所述试剂盒还包含标签或使用说明书,该标签或使用说明书指示将所述纤溶酶原投予所述受试者以实施本发明所述的方法。
另一方面,本发明还涉及一种制品,其包含:含有标签的容器;和(i) 用于本发明所述方法的纤溶酶原或包含纤溶酶原的药物组合物,其中所述标签指示将所述纤溶酶原或组合物投予所述受试者以实施本发明所述方法。
在上述实施方案中,所述试剂盒或制品还包含另外的一个或多个构件或容器,该构件或容器中含有其他药物。在一些实施方案中,所述其他药物选自下组:降血脂药物、抗血小板药物、降血压药物、扩张血管药物、降血糖药物、抗凝血药物、溶血栓药物,保肝药物,抗纤维化药物、抗心律失常药物,强心药物,利尿药物、抗肿瘤药物、放化疗药物、炎症调节药物、免疫调节药物、抗病毒药物、抗生素。
定义
“纤维化”是肺、肝、肾、血管、腹膜、胰腺、皮肤等组织、器官受到炎症、感染、免疫反应、缺血、化学物质、辐射等各种原因导致持续损伤后,成纤维细胞激活增殖、组织器官内纤维结缔组织增多,实质细胞减少,组织、器官结构破坏和功能丧失的病变。该术语可与“纤维化病变”互换使用。该纤维化病变术语涵盖各种原因导致的心脏纤维化、肺纤维化、肝纤维化、肾纤维化、血管纤维化、皮肤纤维化等组织器官纤维化病变,还包括各种疾病发生、发展过程中伴随产生的心脏纤维化、肺纤维化、肝纤维化、肾纤维化、血管纤维化、皮肤纤维化等组织器官纤维化病变。
组织、器官发生纤维化病变后,其正常结构发生改变,相应的功能减弱或丧失,由此导致的相关病症称为“组织器官纤维化相关病症”。
“心脏纤维化”是指各种原因(例如炎症、感染、免疫反应、缺血、化学物质、辐射)导致的或伴随的心脏组织损伤或各种原因导致的心脏疾病的发生与发展过程中出现的纤维化病变。心脏纤维化病变导致心脏功能受损,由此导致的相关病症称为“心脏纤维化相关病症”,包括但不限于心脏功能受损引起的各器官、组织缺血的症状和病症,例如冠心病、心绞痛、心肌梗死、心律失常、脑缺血、呼吸困难、肾功能不全等。
“肝纤维化”是指由各种原因(例如炎症、感染(例如病毒感染)、免疫反应、缺血、化学物质、辐射、氧化应激和酗酒)导致的或伴随的肝内结缔组织异常增生,肝内弥漫性细胞外基质过度沉淀、肝的正常结构遭到破坏的病理变化(病变)。肝纤维化进一步发展即为肝硬化,也涵盖在本发明“肝纤维化”术语的范围内。肝脏纤维化病变导致肝脏功能受损,由此导致的相关病症称为“肝脏纤维化相关病症”。
“肺纤维化”是指各种原因(例如炎症、感染、免疫反应、缺血、化学物质、辐射)导致的或伴随的肺组织间充质细胞增殖、细胞外基质增生沉积及肺实质的重构而造成的病理过程。肺脏纤维化病变导致肺脏功能受损,由此导致的相关病症称为“肺脏纤维化相关病症”。
“肾脏纤维化”是指各种原因(例如炎症、感染、免疫反应、缺血、化学物质、辐射)导致的或伴随的结缔组织在肾脏的异常聚集,导致肾脏结构改变和功能受损的病理过程。肾脏纤维化病变是几乎所有肾脏疾病进展到后期的共同通路。
肾脏纤维化病变导致肾脏功能受损,由此导致的相关病症称为“肾脏纤维化相关病症”,例如,肾脏功能不全,肾衰,尿毒症等。
组织器官的慢性病变常伴有纤维化,例如肺的慢性炎症,慢性病变,伴有肺纤维化。肝纤维化也是如此,例如乙肝、丙肝、酒精肝、脂肪肝、血吸虫病等都伴有早期肝纤维化。再像慢性肾炎、肾小球炎、肾小管炎等都有肾纤维化;心血管、脑血管、下肢血管硬化、或变窄、或阻塞情况下都有血管纤维化。本发明“纤维化”或“纤维化病变”术语涵盖机体各组织器官的慢性病变常伴有的纤维化病变。
“系统性硬化症”或称“硬皮病”是一种以局限性或弥漫性皮肤增厚和纤维化为特征的全身性自身免疫病。病变特点为皮肤纤维增生及血管洋葱皮样改变,最终导致皮肤硬化、血管缺血。本病临床上以局限性或弥漫性皮肤增厚和纤维化为特征,除皮肤受累外,它也可影响内脏(心、肺和消化道等器官)。
“动脉粥样硬化”是一种慢性的、渐进性动脉疾病,发病时动脉中沉积的脂肪部分或全部堵塞血流。动脉粥样硬化是个渐进的过程。当血液中的脂类浓度大大增加时,便会沿着动脉壁形成脂肪条纹。这些条纹会导致脂肪和胆固醇沉积,这些沉淀依附在原本平滑的动脉内膜上,从而形成小结。这些小结下面继而长出纤维化的瘢痕组织,导致钙沉积。沉积的钙逐渐演变为无法除去的白垩状坚硬薄膜(称为动脉粥样斑)。当与体内的某个组织、器官相连的动脉被堵塞后,因该组织、器官中的动脉受阻而引起的组织、器官的缺血损伤可导致该组织、器官的纤维化病变,例如心脏、肺脏、肝脏、肾脏、血管、腹膜、胰腺、皮肤的纤维化。
糖尿病常伴有动脉粥样硬化的发生,其机制多认为是血糖代谢异常引起的动脉粥样硬化,随着更进一步深入的研究,更多的结果表明,糖尿病引起动脉粥样硬化并非单一因素所致,而是通过多种途径以及较为复杂的机制来诱发和促进动脉粥样硬化的发生及发展[4]。糖尿病以及其伴随的动脉粥样硬化可导致组织、器官损伤和纤维化,例如心脏、肺脏、肝脏、肾脏、血管、腹膜、胰腺、皮肤等组织、器官的纤维化。
发明详述
纤溶酶是纤溶酶原激活系统(PA系统)的关键组分。它是一种广谱的蛋白酶,能够水解细胞外基质(ECM)的几个组分,包括纤维蛋白、明胶、纤连蛋白、层粘连蛋白和蛋白聚糖[5]。此外,纤溶酶能将一些金属蛋白酶前体(pro-MMPs)激活形成具有活性的金属蛋白酶(MMPs)。因此纤溶酶被认为是胞外蛋白水解作用的一个重要的上游调节物[6,7]。纤溶酶是由纤溶酶原通过两种生理性的PAs:组织型纤溶酶原激活剂(tPA)或尿激酶型纤溶酶原激活剂(uPA)蛋白水解形成的。由于纤溶酶原在血浆和其他体液中相对水平较高,传统上认为PA系统的调节主要通过PAs的合成和活性水平实现。PA系统组分的合成受不同因素严格调节,如激素、生长因子和细胞因子。此外,还存在纤溶酶和PAs的特定生理抑制剂。纤溶酶的主要抑制剂是α2-抗纤溶酶(α2-antiplasmin)。PAs的活性同时被uPA和tPA的纤溶酶原激活剂抑制剂-1(PAI-1)抑制以及主要抑制uPA的溶酶原激活剂抑制剂-2(PAI-2)调节。某些细胞表面具有直接水解活性的uPA特异性细胞表面受体(uPAR)[8,9]。
纤溶酶原是一个单链糖蛋白,由791个氨基酸组成,分子量约为92 kDa[10,11]。纤溶酶原主要在肝脏合成,大量存在于胞外液中。血浆中纤溶酶原含量约为2μM。因此纤溶酶原是组织和体液中蛋白质水解活性的一个巨大的潜在来源[12,13]。纤溶酶原存在两种分子形式:谷氨酸-纤溶酶原(Glu- plasminogen)和赖氨酸-纤溶酶原(Lys-plasminogen)。天然分泌和未裂解形式的纤溶酶原具有一个氨基末端(N-末端)谷氨酸,因此被称为谷氨酸-纤溶酶原。然而,在纤溶酶存在时,谷氨酸-纤溶酶原在Lys76-Lys77处水解成为赖氨酸-纤溶酶原。与谷氨酸-纤溶酶原相比,赖氨酸-纤溶酶原与纤维蛋白具有更高的亲和力,并可以更高的速率被PAs激活。这两种形式的纤溶酶原的Arg560-Val561肽键可被uPA或tPA切割,导致二硫键连接的双链蛋白酶纤溶酶的形成[14]。纤溶酶原的氨基末端部分包含五个同源三环,即所谓的kringles,羧基末端部分包含蛋白酶结构域。一些kringles含有介导纤溶酶原与纤维蛋白及其抑制剂α2-AP特异性相互作用的赖氨酸结合位点。最新发现一个纤溶酶原为38kDa的片段,其中包括kringles1-4,是血管生成的有效抑制剂。这个片段被命名为血管抑素,可通过几个蛋白酶水解纤溶酶原产生。
纤溶酶的主要底物是纤维蛋白,纤维蛋白的溶解是预防病理性血栓形成的关键[15]。纤溶酶还具有对ECM几个组分的底物特异性,包括层粘连蛋白、纤连蛋白、蛋白聚糖和明胶,表明纤溶酶在ECM重建中也起着重要作用[11,16,17]。间接地,纤溶酶还可以通过转化某些蛋白酶前体为活性蛋白酶来降解ECM的其他组分,包括MMP-1,MMP-2,MMP-3和MMP-9。因此,有人提出,纤溶酶可能是细胞外蛋白水解的一个重要的上游调节器[18]。此外,纤溶酶具有激活某些潜在形式的生长因子的能力[19-21]。在体外,纤溶酶还能水解补体系统的组分并释放趋化补体片段。
“纤溶酶”是存在于血液中的一种非常重要的酶,能将纤维蛋白凝块水解为纤维蛋白降解产物和D-二聚体。
“纤溶酶原”是纤溶酶的酶原形式,根据swiss prot中的序列,按含有信号肽的天然人源纤溶酶原氨基酸序列(序列4)计算由810个氨基酸组成,分子量约为90kD,主要在肝脏中合成并能够在血液中循环的糖蛋白,编码该氨基酸序列的cDNA序列如序列3所示。全长的纤溶酶原包含七个结构域:位于C末端的丝氨酸蛋白酶结构域、N末端的Pan Apple(PAp)结构域以及5个Kringle结构域(Kringle1-5)。参照swiss prot中的序列,其信号肽包括残基Met1-Gly19,PAp包括残基Glu20-Val98,Kringle1包括残基 Cys103-Cys181,Kringle2包括残基Glu184-Cys262,Kringle3包括残基 Cys275-Cys352,Kringle4包括残基Cys377-Cys454,Kringle5包括残基 Cys481-Cys560。根据NCBI数据,丝氨酸蛋白酶域包括残基Val581- Arg804。
Glu-纤溶酶原是天然全长的纤溶酶原,由791个氨基酸组成(不含有19 个氨基酸的信号肽),编码该序列的cDNA序列如序列1所示,其氨基酸序列如序列2所示。在体内,还存在一种是从Glu-纤溶酶原的第76-77位氨基酸处水解从而形成的Lys-纤溶酶原,如序列6所示,编码该氨基酸序列的cDNA序列如序列5所示。Delta-纤溶酶原(δ-plasminogen)是全长纤溶酶原缺失了Kringle2-Kringle5结构的片段,仅含有Kringle1和丝氨酸蛋白酶域[22,23],有文献报道了delta-纤溶酶原的氨基酸序列(序列8)[23],编码该氨基酸序列的cDNA序列如序列7。小纤溶酶原(Mini-plasminogen)由 Kringle5和丝氨酸蛋白酶域组成,有文献报道其包括残基Val443-Asn791 (以不含有信号肽的Glu-纤溶酶原序列的Glu残基为起始氨基酸)[24],其氨基酸序列如序列10所示,编码该氨基酸序列的cDNA序列如序列9所示。而微纤溶酶原(Micro-plasminogen)仅含有丝氨酸蛋白酶结构域,有文献报道其氨基酸序列包括残基Ala543-Asn791(以不含有信号肽的Glu-纤溶酶原序列的Glu残基为起始氨基酸)[25],也有专利文献CN102154253A 报道其序列包括残基Lys531-Asn791(以不含有信号肽的Glu-纤溶酶原序列的Glu残基为起始氨基酸),本专利序列参考专利文献CN102154253A,其氨基酸序列如序列12所示,编码该氨基酸序列的cDNA序列如序列11 所示。
本发明的“纤溶酶”与“纤维蛋白溶酶”、“纤维蛋白溶解酶”可互换使用,含义相同;“纤溶酶原”与“纤维蛋白溶酶原”、“纤维蛋白溶解酶原”可互换使用,含义相同。
在本申请中,所述纤溶酶原“缺乏”的含义为受试者体内纤溶酶原的含量或活性比正常人低,低至足以影响所述受试者的正常生理功能;所述纤溶酶原“缺失”的含义为受试者体内纤溶酶原的含量或活性显著低于正常人,甚至活性或表达极微,只有通过外源提供才能维持正常生理功能。
本领域技术人员可以理解,本发明纤溶酶原的所有技术方案适用于纤溶酶,因此,本发明描述的技术方案涵盖了纤溶酶原和纤溶酶。
在循环过程中,纤溶酶原采用封闭的非活性构象,但当结合至血栓或细胞表面时,在纤溶酶原激活剂(plasminogen activator,PA)的介导下,其转变为呈开放性构象的活性纤溶酶。具有活性的纤溶酶可进一步将纤维蛋白凝块水解为纤维蛋白降解产物和D-二聚体,进而溶解血栓。其中纤溶酶原的PAp结构域包含维持纤溶酶原处于非活性封闭构象的重要决定簇,而 KR结构域则能够与存在于受体和底物上的赖氨酸残基结合。已知多种能够作为纤溶酶原激活剂的酶,包括:组织纤溶酶原激活剂(tPA)、尿激酶纤溶酶原激活剂(uPA)、激肽释放酶和凝血因子XII(哈格曼因子)等。
“纤溶酶原活性片段”是指在纤溶酶原蛋白中,能够与底物中的靶序列结合并发挥蛋白水解功能的活性片段。本发明涉及纤溶酶原的技术方案涵盖了用纤溶酶原活性片段代替纤溶酶原的技术方案。本发明所述的纤溶酶原活性片段为包含纤溶酶原的丝氨酸蛋白酶域的蛋白质,优选,本发明所述的纤溶酶原活性片段包含序列14、与序列14具有至少80%、90%、 95%、96%、97%、98%、99%同源性的氨基酸序列的蛋白质。因此,本发明所述的纤溶酶原包括含有该纤溶酶原活性片段、并且仍然保持该纤溶酶原活性的蛋白。
目前,对于血液中纤溶酶原及其活性测定方法包括:对组织纤溶酶原激活剂活性的检测(t-PAA)、血浆组织纤溶酶原激活剂抗原的检测(t- PAAg)、对血浆组织纤溶酶原活性的检测(plgA)、血浆组织纤溶酶原抗原的检测(plgAg)、血浆组织纤溶酶原激活剂抑制物活性的检测、血浆组织纤溶酶原激活剂抑制物抗原的检测、血浆纤维蛋白溶酶-抗纤维蛋白溶酶复合物检测(PAP)。其中最常用的检测方法为发色底物法:向受检血浆中加链激酶 (SK)和发色底物,受检血浆中的PLG在SK的作用下,转变成PLM,后者作用于发色底物,随后用分光光度计测定,吸光度增加与纤溶酶原活性成正比。此外也可采用免疫化学法、凝胶电泳、免疫比浊法、放射免疫扩散法等对血液中的纤溶酶原活性进行测定。
“直系同源物或直系同系物(ortholog)”指不同物种之间的同源物,既包括蛋白同源物也包括DNA同源物,也称为直向同源物、垂直同源物。其具体指不同物种中由同一祖先基因进化而来的蛋白或基因。本发明的纤溶酶原包括人的天然纤溶酶原,还包括来源于不同物种的、具有纤溶酶原活性的纤溶酶原直系同源物或直系同系物。
“保守取代变体”是指其中一个给定的氨基酸残基改变但不改变蛋白质或酶的整体构象和功能,这包括但不限于以相似特性(如酸性,碱性,疏水性,等)的氨基酸取代亲本蛋白质中氨基酸序列中的氨基酸。具有类似性质的氨基酸是众所周知的。例如,精氨酸、组氨酸和赖氨酸是亲水性的碱性氨基酸并可以互换。同样,异亮氨酸是疏水氨基酸,则可被亮氨酸,蛋氨酸或缬氨酸替换。因此,相似功能的两个蛋白或氨基酸序列的相似性可能会不同。例如,基于MEGALIGN算法的70%至99%的相似度(同一性)。“保守取代变体”还包括通过BLAST或FASTA算法确定具有60%以上的氨基酸同一性的多肽或酶,若能达75%以上更好,最好能达85%以上,甚至达90%以上为最佳,并且与天然或亲本蛋白质或酶相比具有相同或基本相似的性质或功能。
“分离的”纤溶酶原是指从其天然环境分离和/或回收的纤溶酶原蛋白。在一些实施方案中,所述纤溶酶原会纯化(1)至大于90%、大于95%、或大于98%的纯度(按重量计),如通过Lowry法所确定的,例如超过99%(按重量计),(2)至足以通过使用旋转杯序列分析仪获得N端或内部氨基酸序列的至少15个残基的程度,或(3)至同质性,该同质性是通过使用考马斯蓝或银染在还原性或非还原性条件下的十二烷基硫酸钠-聚丙烯酰胺凝胶电泳 (SDS-PAGE)确定的。分离的纤溶酶原也包括通过生物工程技术从重组细胞制备,并通过至少一个纯化步骤分离的纤溶酶原。
术语“多肽”、“肽”和“蛋白质”在本文中可互换使用,指任何长度的氨基酸的聚合形式,其可以包括遗传编码的和非遗传编码的氨基酸,化学或生物化学修饰的或衍生化的氨基酸,和具有经修饰的肽主链的多肽。该术语包括融合蛋白,包括但不限于具有异源氨基酸序列的融合蛋白,具有异源和同源前导序列(具有或没有N端甲硫氨酸残基)的融合物;等等。
关于参照多肽序列的“氨基酸序列同一性百分数(%)”定义为在必要时引入缺口以实现最大百分比序列同一性后,且不将任何保守替代视为序列同一性的一部分时,候选序列中与参照多肽序列中的氨基酸残基相同的氨基酸残基的百分率。为测定百分比氨基酸序列同一性目的的对比可以以本领域技术范围内的多种方式实现,例如使用公众可得到的计算机软件,诸如BLAST、BLAST-2、ALIGN或Megalign(DNASTAR)软件。本领域技术人员能决定用于比对序列的适宜参数,包括对所比较序列全长实现最大对比需要的任何算法。然而,为了本发明的目的,氨基酸序列同一性百分数值是使用序列比较计算机程序ALIGN-2产生的。
在采用ALIGN-2来比较氨基酸序列的情况中,给定氨基酸序列A相对于给定氨基酸序列B的%氨基酸序列同一性(或者可表述为具有或包含相对于、与、或针对给定氨基酸序列B的某一%氨基酸序列同一性的给定氨基酸序列A)如下计算:
分数X/Y乘100
其中X是由序列比对程序ALIGN-2在该程序的A和B比对中评分为相同匹配的氨基酸残基的数目,且其中Y是B中的氨基酸残基的总数。应当领会,在氨基酸序列A的长度与氨基酸序列B的长度不相等的情况下, A相对于B的%氨基酸序列同一性会不等于B相对于A的%氨基酸序列同一性。除非另有明确说明,本文中使用的所有%氨基酸序列同一性值都是依照上一段所述,使用ALIGN-2计算机程序获得的。
如本文中使用的,术语“治疗”和“处理”指获得期望的药理和/或生理效果。所述效果可以是完全或部分预防疾病或其症状,和/或部分或完全治愈疾病和/或其症状,并且包括:(a)预防疾病在受试者体内发生,所述受试者可以具有疾病的素因,但是尚未诊断为具有疾病;(b)抑制疾病,即阻滞其形成;和(c)减轻疾病和/或其症状,即引起疾病和/或其症状消退。
术语“个体”、“受试者”和“患者”在本文中可互换使用,指哺乳动物,包括但不限于鼠(大鼠、小鼠)、非人灵长类、人、犬、猫、有蹄动物(例如马、牛、绵羊、猪、山羊)等。
“治疗有效量”或“有效量”指在对哺乳动物或其它受试者施用以治疗疾病时足以实现对疾病的所述预防和/或治疗的纤溶酶原的量。“治疗有效量”会根据所使用的纤溶酶原、要治疗的受试者的疾病和/或其症状的严重程度以及年龄、体重等而变化。
本发明纤溶酶原的制备
纤溶酶原可以从自然界分离并纯化用于进一步的治疗用途,也可以通过标准的化学肽合成技术来合成。当通过化学合成多肽时,可以经液相或固相进行合成。固相多肽合成(SPPS)(其中将序列的C末端氨基酸附接于不溶性支持物,接着序贯添加序列中剩余的氨基酸)是适合纤溶酶原化学合成的方法。各种形式的SPPS,诸如Fmoc和Boc可用于合成纤溶酶原。用于固相合成的技术描述于Barany和Solid-Phase Peptide Synthesis;第3-284页于The Peptides:Analysis,Synthesis,Biology.第2卷:Special Methods in Peptide Synthesis,Part A.,Merrifield,等J.Am.Chem.Soc.,85:2149-2156 (1963);Stewart等,Solid Phase Peptide Synthesis,2nd ed.Pierce Chem.Co., Rockford,Ill.(1984);和Ganesan A.2006Mini Rev.Med Chem.6:3-10和 Camarero JA等2005Protein Pept Lett.12:723-8中。简言之,用其上构建有肽链的功能性单元处理小的不溶性多孔珠。在偶联/去保护的重复循环后,将附接的固相游离N末端胺与单个受N保护的氨基酸单元偶联。然后,将此单元去保护,露出可以与别的氨基酸附接的新的N末端胺。肽保持固定在固相上,之后将其切掉。
可以使用标准重组方法来生产本发明的纤溶酶原。例如,将编码纤溶酶原的核酸插入表达载体中,使其与表达载体中的调控序列可操作连接。表达调控序列包括但不限于启动子(例如天然关联的或异源的启动子)、信号序列、增强子元件、和转录终止序列。表达调控可以是载体中的真核启动子系统,所述载体能够转化或转染真核宿主细胞(例如COS或CHO细胞)。一旦将载体掺入合适的宿主中,在适合于核苷酸序列的高水平表达及纤溶酶原的收集和纯化的条件下维持宿主。
合适的表达载体通常在宿主生物体中作为附加体或作为宿主染色体 DNA的整合部分复制。通常,表达载体含有选择标志物(例如氨苄青霉素抗性、潮霉素抗性、四环素抗性、卡那霉素抗性或新霉素抗性)以有助于对外源用期望的DNA序列转化的那些细胞进行检测。
大肠杆菌(Escherichia coli)是可以用于克隆主题抗体编码多核苷酸的原核宿主细胞的例子。适合于使用的其它微生物宿主包括杆菌,诸如枯草芽孢杆菌(Bacillus subtilis)和其他肠杆菌科(enterobacteriaceae),诸如沙门氏菌属(Salmonella)、沙雷氏菌属(Serratia)、和各种假单胞菌属(Pseudomonas)物种。在这些原核宿主中,也可以生成表达载体,其通常会含有与宿主细胞相容的表达控制序列(例如复制起点)。另外,会存在许多公知的启动子,诸如乳糖启动子系统,色氨酸(trp)启动子系统,beta-内酰胺酶启动子系统,或来自噬菌体λ的启动子系统。启动子通常会控制表达,任选在操纵基因序列的情况中,并且具有核糖体结合位点序列等,以启动并完成转录和翻译。
其他微生物,诸如酵母也可用于表达。酵母(例如酿酒酵母(S. cerevisiae))和毕赤酵母(Pichia)是合适的酵母宿主细胞的例子,其中合适的载体根据需要具有表达控制序列(例如启动子)、复制起点、终止序列等。典型的启动子包含3-磷酸甘油酸激酶和其它糖分解酶。诱导型酵母启动于特别包括来自醇脱氢酶、异细胞色素C、和负责麦芽糖和半乳糖利用的酶的启动子。
在微生物外,哺乳动物细胞(例如在体外细胞培养物中培养的哺乳动物细胞)也可以用于表达并生成本发明的抗-Tau抗体(例如编码主题抗-Tau抗体的多核苷酸)。参见Winnacker,From Genes to Clones,VCH Publishers, N.Y.,N.Y.(1987)。合适的哺乳动物宿主细胞包括CHO细胞系、各种Cos 细胞系、HeLa细胞、骨髓瘤细胞系、和经转化的B细胞或杂交瘤。用于这些细胞的表达载体可以包含表达控制序列,如复制起点,启动子和增强子 (Queen等,Immunol.Rev.89:49(1986)),以及必需的加工信息位点,诸如核糖体结合位点,RNA剪接位点,多聚腺苷酸化位点,和转录终止子序列。合适的表达控制序列的例子是白免疫球蛋白基因、SV40、腺病毒、牛乳头瘤病毒、巨细胞病毒等衍生的启动子。参见Co等,J.Immunol.148:1149 (1992)。
一旦合成(化学或重组方式),可以依照本领域的标准规程,包括硫酸铵沉淀,亲和柱,柱层析,高效液相层析(HPLC),凝胶电泳等来纯化本发明所述的纤溶酶原。该纤溶酶原是基本上纯的,例如至少约80%至85%纯的,至少约85%至90%纯的,至少约90%至95%纯的,或98%至99%纯的或更纯的,例如不含污染物,所述污染物如细胞碎片,除主题抗体以外的大分子,等等。
药物配制剂
可以通过将具有所需纯度的纤溶酶原与可选的药用载体,赋形剂,或稳定剂(Remington's Pharmaceutical Sciences,16版,Osol,A.ed.(1980))混合形成冻干制剂或水溶液制备治疗配制剂。可接受的载体、赋形剂、稳定剂在所用剂量及浓度下对受者无毒性,并包括缓冲剂例如磷酸盐,柠檬酸盐及其它有机酸;抗氧化剂包括抗坏血酸和蛋氨酸;防腐剂(例如十八烷基二甲基苄基氯化铵;氯化己烷双胺;氯化苄烷铵(benzalkonium chloride),苯索氯铵;酚、丁醇或苯甲醇;烷基对羟基苯甲酸酯如甲基或丙基对羟基苯甲酸酯;邻苯二酚;间苯二酚;环己醇;3-戊醇;间甲酚);低分子量多肽 (少于约10个残基);蛋白质如血清白蛋白,明胶或免疫球蛋白;亲水聚合物如聚乙烯吡咯烷酮;氨基酸如甘氨酸,谷氨酰胺、天冬酰胺、组氨酸、精氨酸或赖氨酸;单糖,二糖及其它碳水化合物包括葡萄糖、甘露糖、或糊精;螯合剂如EDTA;糖类如蔗糖、甘露醇、岩藻糖或山梨醇;成盐反离子如钠;金属复合物(例如锌-蛋白复合物);和/或非离子表面活性剂,例如TWEENTM,PLURONICSTM或聚乙二醇(PEG)。优选冻干的抗-VEGF 抗体配制剂在WO 97/04801中描述,其包含在本文中作为参考。
本发明的配制剂也可含有需治疗的具体病症所需的一种以上的活性化合物,优选活性互补并且相互之间没有副作用的那些。例如,抗高血压的药物,抗心律失常的药物,治疗糖尿病的药物等。
本发明的纤溶酶原可包裹在通过诸如凝聚技术或界面聚合而制备的微胶囊中,例如,可置入在胶质药物传送系统(例如,脂质体,白蛋白微球,微乳剂,纳米颗粒和纳米胶囊)中或置入粗滴乳状液中的羟甲基纤维素或凝胶-微胶囊和聚-(甲基丙烯酸甲酯)微胶囊中。这些技术公开于Remington's Pharmaceutical Sciences 16th edition,Osol,A.Ed.(1980)。
用于体内给药的本发明的纤溶酶原必需是无菌的。这可以通过在冷冻干燥和重新配制之前或之后通过除菌滤膜过滤而轻易实现。
本发明的纤溶酶原可制备缓释制剂。缓释制剂的适当实例包括具有一定形状且含有糖蛋白的固体疏水聚合物半通透基质,例如膜或微胶囊。缓释基质实例包括聚酯、水凝胶(如聚(2-羟基乙基-异丁烯酸酯)(Langer等,J. Biomed.Mater.Res.,15:167-277(1981);Langer,Chem.Tech.,12:98- 105(1982))或聚(乙烯醇),聚交酯(美国专利3773919,EP 58,481),L-谷氨酸与γ乙基-L-谷氨酸的共聚物(Sidman,等,Biopolymers 22:547(1983)),不可降解的乙烯-乙烯乙酸酯(ethylene-vinyl acetate)(Langer,等,出处同上),或可降解的乳酸-羟基乙酸共聚物如Lupron DepotTM(由乳酸-羟基乙酸共聚物和亮氨酰脯氨酸(leuprolide)乙酸酯组成的可注射的微球体),以及聚D-(-)- 3-羟丁酸。聚合物如乙烯-乙酸乙烯酯和乳酸-羟基乙酸能持续释放分子100 天以上,而一些水凝胶释放蛋白的时间却较短。可以根据相关机理来设计使蛋白稳定的合理策略。例如,如果发现凝聚的机理是通过硫代二硫键互换而形成分子间S-S键,则可通过修饰巯基残基、从酸性溶液中冻干、控制湿度、采用合适的添加剂、和开发特定的聚合物基质组合物来实现稳定。
给药和剂量
可以通过不同方式,例如通过静脉内,腹膜内,皮下,颅内,鞘内,动脉内(例如经由颈动脉),肌内来实现本发明药物组合物的施用。
用于胃肠外施用的制备物包括无菌水性或非水性溶液、悬浮液和乳剂。非水性溶剂的例子是丙二醇、聚乙二醇、植物油如橄榄油,和可注射有机酯,如油酸乙酯。水性载体包括水、醇性/水性溶液、乳剂或悬浮液,包括盐水和缓冲介质。胃肠外媒介物包含氯化钠溶液、林格氏右旋糖、右旋糖和氯化钠、或固定油。静脉内媒介物包含液体和营养补充物、电解质补充物,等等。也可以存在防腐剂和其他添加剂,诸如例如,抗微生物剂、抗氧化剂、螯合剂、和惰性气体,等等。
医务人员会基于各种临床因素确定剂量方案。如医学领域中公知的,任一患者的剂量取决于多种因素,包括患者的体型、体表面积、年龄、要施用的具体化合物、性别、施用次数和路径、总体健康、和同时施用的其它药物。本发明包含纤溶酶原的药物组合物的剂量范围可以例如为例如每天约0.0001至2000mg/kg,或约0.001至500mg/kg(例如0.02mg/kg,0.25 mg/kg,0.5mg/kg,0.75mg/kg,10mg/kg,50mg/kg等等)受试者体重。例如,剂量可以是1mg/kg体重或50mg/kg体重或在1-50mg/kg的范围,或至少1mg/kg。高于或低于此例示性范围的剂量也涵盖在内,特别是考虑到上述的因素。上述范围中的中间剂量也包含在本发明的范围内。受试者可以每天、隔天、每周或根据通过经验分析确定的任何其它日程表施用此类剂量。例示性的剂量日程表包括连续几天1-10mg/kg。在本发明的药物施用过程中需要实时评估治疗效果和安全性。
制品或药盒
本发明的一个实施方案涉及一种制品或药盒,其包含可用于治疗由糖尿病引起的心血管病及其相关病症的本发明纤溶酶原或纤溶酶。所述制品优选包括一个容器,标签或包装插页。适当的容器有瓶子,小瓶,注射器等。容器可由各种材料如玻璃或塑料制成。所述容器含有组合物,所述组合物可有效治疗本发明的疾病或病症并具有无菌入口(例如所述容器可为静脉内溶液包或小瓶,其含有可被皮下注射针穿透的塞子的)。所述组合物中至少一种活性剂为纤溶酶原/纤溶酶。所述容器上或所附的标签说明所述组合物用于治疗本发明所述由糖尿病引起的心血管病及其相关病症。所述制品可进一步包含含有可药用缓冲液的第二容器,诸如磷酸盐缓冲的盐水,林格氏溶液以及葡萄糖溶液。其可进一步包含从商业和使用者角度来看所需的其它物质,包括其它缓冲液,稀释剂,过滤物,针和注射器。此外,所述制品包含带有使用说明的包装插页,包括例如指示所述组合物的使用者将纤溶酶原组合物以及治疗伴随的疾病的其它药物给药患者。
附图简述
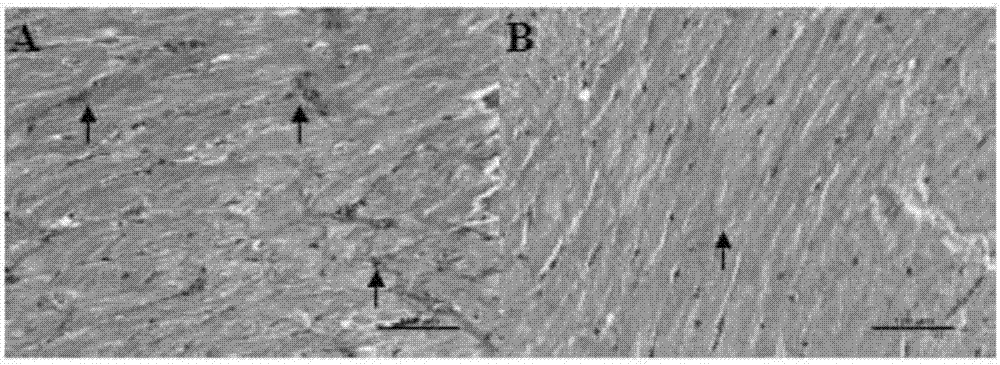
图1博莱霉素诱导的系统性硬化模型小鼠给予纤溶酶原21天后心脏天狼星红染色代表性图片。A为给溶媒PBS对照组,B为给纤溶酶原组。研究发现,在博莱霉素诱导的系统性硬化症小鼠模型中,给溶媒PBS对照组心脏胶原蛋白沉积(箭头标识)程度比给纤溶酶原组高。说明纤溶酶原能有效降低博莱霉素诱导的心脏纤维化。
图2 24-25周龄糖尿病小鼠给予纤溶酶原31天后心脏masson染色观察结果。A为给溶媒PBS对照组,B为给纤溶酶原组。结果显示,给溶媒 PBS对照组心肌纤维之间可见着蓝色的增生的胶原纤维(箭头标识),呈轻度心肌纤维化;给纤溶酶原组心肌纤维之间可见少量的浅蓝色的增生的胶原纤维,较之于对照组,心肌纤维化明显减轻。说明纤溶酶原能够改善糖尿病小鼠心脏的纤维化。
图3 17-18周龄糖尿病小鼠给予纤溶酶原35天后心脏天狼星红染色代表性图片。A为给溶媒PBS对照组,B为给纤溶酶原组。结果显示,给纤溶酶原组小鼠胶原纤维的沉积(箭头标识)明显少于给溶媒PBS对照组。说明纤溶酶原能够减少相对年轻(17-18周龄)糖尿病小鼠心脏的纤维化。
图4 26-27周龄糖尿病小鼠给予纤溶酶原35天后心脏天狼星红染色代表性图片。A为给溶媒PBS对照组,B为给纤溶酶原组。结果显示,给纤溶酶原组小鼠胶原蛋白的沉积(箭头标识)明显少于给溶媒PBS对照组。说明纤溶酶原能减弱相对年老(26-27周龄)糖尿病小鼠心脏的纤维化。
图5 ApoE动脉粥样硬化模型小鼠给予纤溶酶原30天后心脏天狼星红染色代表性图片。A为给溶媒PBS对照组,B为给纤溶酶原组。结果显示,给纤溶酶原组胶原的沉积(箭头标识)明显少于给溶媒PBS对照组,说明纤溶酶原能减轻ApoE动脉粥样硬化模型小鼠心脏纤维化。
图6 C57高血脂模型小鼠给予纤溶酶原30天后心脏天狼星红染色代表性图片。A为给溶媒PBS对照组,B为给纤溶酶原组。结果显示,给纤溶酶原组胶原的沉积(箭头标识)明显少于给溶媒PBS对照组,说明纤溶酶原能减轻高血脂模型小鼠心脏纤维化。
图7 24-25周龄糖尿病小鼠给予纤溶酶原31天后胰岛天狼星红染色观察结果。A为给溶媒PBS对照组,B为给纤溶酶原组,C为定量分析结果。结果显示,给纤溶酶原组小鼠胰岛胶原沉积(箭头标识)明显少于给溶媒PBS对照组,且统计差异显著(*表示P<0.05)。说明纤溶酶原能改善糖尿病所致的胰岛损伤和纤维化。
图8 ApoE动脉粥样硬化模型小鼠给予纤溶酶原30天后主动脉窦天狼星红染色代表性图片。A、C为给溶媒PBS对照组,B、D为给纤溶酶原组。结果显示给纤溶酶原组胶原蛋白沉积(箭头标识)的面积明显小于给溶媒PBS对照组,说明纤溶酶原能够消减动脉粥硬化模型小鼠主动脉窦纤维化水平。
图9给予纤溶酶原14天后四氯化碳诱导的肝纤维化模型小鼠肝脏天狼星红染色代表性图片。A为空白对照组,B为给溶媒PBS对照组,C为给纤溶酶原组。结果显示,给纤溶酶原组胶原蛋白沉积明显少于给溶媒PBS 对照组,且与空白对照小鼠胶原蛋白沉积水平接近。说明纤溶酶原能减少肝脏胶原蛋白的沉积,改善肝纤维化模型小鼠肝脏纤维化。
图10 16周高脂血症模型小鼠给予纤溶酶原30天后主动脉窦天狼星红染色代表性图片。A、C为给溶媒PBS对照组,B、D为给纤溶酶原组。结果显示,给纤溶酶原组主动脉窦管壁内膜胶原蛋白沉积(箭头标识)的面积明显小于给溶媒PBS对照组,说明纤溶酶原能够消减高脂血症模型小鼠主动脉窦管壁内膜纤维化水平。
图11博莱霉素诱导的系统性硬化模型小鼠给予纤溶酶原21天后皮肤天狼星红染色代表性图片。A为空白对照组,B为给溶媒PBS对照组,C 为给纤溶酶原组,D为PLG活性受损组。结果显示,在博莱霉素诱导的系统性硬化症小鼠模型中,给溶媒PBS组和PLG活性受损组,真皮上部胶原纤维束明显增多,胶原纤维粗大,排列致密,真皮层增厚;给纤溶酶原组真皮层中成纤维细胞明显少于给溶媒PBS组,且皮肤真皮层厚度接近正常水平。
图12博莱霉素诱导的系统性硬化模型小鼠给予纤溶酶原21天后肺天狼星红染色代表性图片。A为给溶媒PBS对照组,B为给纤溶酶原组,C 为定量分析结果。结果显示,在博莱霉素诱导的系统性硬化症小鼠模型中,给溶媒PBS组小鼠肺纤维化(箭头标识)程度比给纤溶酶原组高;给纤溶酶原组小鼠肺部肺泡壁形态接近正常,炎症水平细胞明显减少,纤维化程度明显低于给溶媒PBS组,且统计差异显著(*表示P<0.05)。
图13博莱霉素诱导的系统性硬化模型小鼠给予纤溶酶原21天后肾脏天狼星红染色代表性图片。A为给溶媒PBS对照组,B为给纤溶酶原组。结果显示,在博莱霉素诱导的系统性硬化症小鼠模型中,给溶媒PBS对照组肾脏胶原纤维化(箭头标识)程度比给纤溶酶原组高。说明纤溶酶原能有效降低博莱霉素诱导的肾脏纤维化。
图14 24-25周龄糖尿病小鼠给予纤溶酶原31天后肾脏IV型胶原免疫染色观察结果。A为给溶媒PBS对照组,B为给纤溶酶原组。结果显示,给纤溶酶原组IV胶原阳性着色(箭头标识)明显多于给溶媒PBS对照组,说明纤溶酶原能改善糖尿病小鼠肾脏的纤维化。
图15 26周龄糖尿病小鼠给予纤溶酶原35天后肾脏masson染色观察结果。A为给溶媒PBS对照组,B为给纤溶酶原组。结果显示,给溶媒PBS 对照组,肾小球系膜增生,系膜基质增多,肾间质轻度纤维化(箭头标识),增生的纤维化呈蓝色。给纤溶酶原组肾小球系膜细胞及基质明显比对照组少,肾间质纤维化明显减少。说明纤溶酶原能够改善糖尿病小鼠肾脏的纤维化病变。
图16顺铂肾纤维化模型小鼠给予纤溶酶原7天后肾脏IV型胶原免疫染色观察结果。A为给溶媒PBS对照组,B为给纤溶酶原组。结果显示,给溶媒PBS对照组肾脏IV型胶原阳性表达(箭头标识)明显高于给纤溶酶原组。说明纤溶酶原能改善顺铂肾纤维化模型小鼠肾脏的纤维化。
图17嘌呤诱导的慢性肾损伤模型小鼠给予纤溶酶原10天后肾脏天狼星红染色观察结果。A为给溶媒PBS对照组,B为给纤溶酶原组,C为 PLG活性受损组,D为定量分析结果。给纤溶酶原组胶原的沉积(箭头标识)明显少于给溶媒PBS对照组和PLG活性受损组,且给纤溶酶原组与 PLG活性受损组定量分析统计差异显著(*表示P<0.05)。说明纤溶酶原能减轻慢性肾损伤所致的肾脏纤维化,促进肾损伤的修复。
图18给予纤溶酶原30天后3%胆固醇高脂血症模型小鼠肾脏天狼星红染色观察结果。A为空白对照组,B为给溶媒PBS对照组,C为给纤溶酶原组,D为定量分析结果。结果显示,给纤溶酶原组肾脏胶原蛋白沉积 (箭头标识)明显少于给溶媒PBS对照组,且统计差异显著;给纤溶酶原组纤维化基本恢复到正常水平。说明纤溶酶原能有效的减少3%胆固醇高脂血症模型小鼠肾脏纤维化。
图19给予纤溶酶原28天四氯化碳的诱导肝脏纤维化模型小鼠肝脏天狼星红染色观察结果。A为空白对照组,B为给溶媒PBS对照组,C为给纤溶酶原组,D为定量分析结果。结果显示,给纤溶酶原组胶原蛋白沉积 (箭头标识)明显少于给溶媒PBS对照组,且统计差异显著(*表示 P<0.05);与给溶媒PBS对照组相比,给纤溶酶原组胶原沉积水平更加接近空白对照小鼠。说明纤溶酶原能减少肝纤维化模型小鼠肝脏胶原蛋白的沉积,改善肝脏纤维化。
实施例
实施例1纤溶酶原降低系统性硬化症小鼠心脏纤维化
取12周龄C57雄鼠10只,随机分为两组,给溶媒PBS对照组和给纤溶酶原组各5只。实验开始当天记为第0天并称重分组,第1天开始造模给药,按0.1mg/0.1ml/只/天皮下注射博莱霉素诱导系统性硬化症[26],并开始给纤溶酶原或PBS,连续给药21天。给纤溶酶原组小鼠按1mg/0.1ml/只/ 天尾静脉注射纤溶酶原,给溶媒PBS对照组尾静脉注射相同体积的PBS,在第22天处死小鼠并取心脏在4%多聚甲醛固定液中固定24小时。固定后的心脏经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为 3μm,切片脱蜡至水后水洗1次,以0.1%天狼星红饱和苦味酸染色30分钟后,流水冲洗2min,苏木素染色1分钟,流水冲洗,1%盐酸酒精分化,氨水返蓝,流水冲洗,烘干后中性树胶封片,在200倍光学显微镜下观察。
研究发现,在博莱霉素诱导的系统性硬化症小鼠模型中,镜下观察到给溶媒PBS对照组(图1A)心脏胶原蛋白沉积比给纤溶酶原组(图1B) 高。说明纤溶酶原能有效降低博莱霉素诱导的心脏纤维化。
实施例2纤溶酶原改善24-25周龄糖尿病小鼠心脏纤维化
24-25周龄db/db雄鼠10只,随机分为两组,给溶媒PBS对照组和给纤溶酶原组,每组各5只。实验开始当天记为第0天并称重分组,第1天开始给纤溶酶原或PBS,连续给药31天。给纤溶酶原组小鼠按2mg/0.2ml/只/ 天尾静脉注射纤溶酶原,给溶媒PBS对照组给予相同体积的PBS。给纤溶酶原31天后处死小鼠并取心脏组织在4%多聚甲醛固定液中固定24小时。固定后的心脏组织经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为4μm,切片脱蜡复水后放入重铬酸钾溶液过夜。铁苏木素染3到5 分钟,流水稍洗。1%盐酸酒精分化,氨水处理1秒,水洗。丽春红酸性品红液染8分钟,水中快速漂洗。1%磷钼酸水溶液处理约2分钟,苯胺蓝液复染6分钟。1%冰醋酸漂洗大概1分钟左右。无水乙醇脱水二甲苯透明后封片,切片在200倍光学显微镜下观察。
糖尿病最常见的并发症是结缔组织过度累积(病理性纤维化),心肌间质纤维化可能是糖尿病心肌病变的特征性病理改变[28-29]。
Masson染色可以显示组织的纤维化。结果显示,给溶媒PBS对照组 (图2A)心肌纤维之间可见着蓝色的增生的胶原纤维(箭头标识),呈轻度心肌纤维化;给纤溶酶原组(图2B)心肌纤维之间可见少量的浅蓝色的增生的胶原纤维,较之于对照组,心肌纤维化明显减轻。说明纤溶酶原能改善糖尿病小鼠心脏的纤维化。
实施例3纤溶酶原降低17-18周龄糖尿病小鼠心脏胶原蛋白沉积
17-18周龄db/db雄鼠8只,随机分为两组,给溶媒PBS对照组和给纤溶酶原组,每组各4只。实验开始当天记为第0天并称重分组,第1天开始给纤溶酶原或PBS,连续给药35天。给纤溶酶原组小鼠按2mg/0.2ml/只/ 天尾静脉注射纤溶酶原,给溶媒PBS对照组给予相同体积的PBS。给纤溶酶原35天后处死小鼠并取心脏组织在4%多聚甲醛固定液中固定24小时。固定后的心脏经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为3μm,切片脱蜡至水后水洗1次,以0.1%天狼星红饱和苦味酸染色30 分钟后,流水冲洗2min,苏木素染色1分钟,流水冲洗,1%盐酸酒精分化,氨水返蓝,流水冲洗,烘干后中性树胶封片,在200倍光学显微镜下观察。
结果显示,给纤溶酶原组(图3B)小鼠胶原纤维的沉积(箭头标识) 明显少于给溶媒PBS对照组(图3A)。说明纤溶酶原能降低心脏组织胶原蛋白的沉积,提示纤溶酶原有望通过降低心脏组织胶原蛋白的沉积,减少相对年轻(17-18周龄)糖尿病小鼠心脏组织纤维化。
实施例4纤溶酶原降低26-27周龄糖尿病小鼠心脏胶原蛋白沉积
26-27周龄db/db雄鼠9只,随机分为两组,给溶媒PBS对照组5只和给纤溶酶原组4只。实验开始当天记为第0天并称重分组,第1天开始给纤溶酶原或PBS,连续给药35天。给纤溶酶原组小鼠按2mg/0.2ml/只/天尾静脉注射纤溶酶原,给溶媒PBS对照组给予相同体积的PBS。给纤溶酶原 35天后处死小鼠并取心脏组织在4%多聚甲醛固定液中固定24小时。固定后的心脏经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为 3μm,切片脱蜡至水后水洗1次,以0.1%天狼星红染色60分钟后,流水冲洗,苏木素染色1分钟,流水冲洗,1%盐酸酒精和氨水分化返蓝,流水冲洗,烘干后封片,在200倍光学显微镜下观察。
结果显示,给纤溶酶原组(图4B)小鼠胶原纤维的沉积(箭头标识) 明显少于给溶媒PBS对照组(图4A)。说明纤溶酶原能降低心脏组织胶原蛋白的沉积,提示纤溶酶原有望通过降低心脏组织胶原蛋白的沉积,减少相对年老(26-27周龄)糖尿病小鼠心脏组织纤维化。
实施例5纤溶酶原改善ApoE动脉粥样硬化小鼠心脏纤维化水平
6周龄ApoE雄性小鼠13只饲喂高脂高胆固醇饲料(南通特洛菲, TP2031)16周,诱导动脉粥样硬化[31,32]。在给药前三天每只小鼠取血 50μL,检测总胆固醇浓度,并根据检测结果将小鼠随机分为两组,给溶媒 PBS对照组7只以及给纤溶酶原组6只。开始给药定为第1天,给纤溶酶原组小鼠尾静脉注射人源纤溶酶原1mg/0.1ml/只/天,给溶媒PBS对照组尾静脉注射同体积的PBS。给药30天,期间一直饲喂高脂高胆固醇饲料。于第31天处死小鼠,取心脏于4%多聚甲醛固定24-48小时。固定后的组织经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为3μm,切片脱蜡复水后水洗1次,以0.1%天狼星红饱和苦味酸染色30分钟后,流水冲洗2min,苏木素染色1分钟,流水冲洗,1%盐酸酒精分化,氨水返蓝,流水冲洗,烘干后中性树胶封片,在200倍光学显微镜下观察。
结果显示,给纤溶酶原组(图5B)胶原的沉积(箭头标识)明显少于给溶媒PBS对照组(图5A),提示纤溶酶原能通过减少ApoE动脉粥样硬化模型小鼠心脏组织的胶原蛋白沉积,阻止和降低动脉粥样硬化所致的心脏纤维化。
实施例6纤溶酶原降低高血脂模型小鼠心脏纤维化
6周龄C57雄性小鼠11只饲喂高脂高胆固醇饲料(南通特洛菲, TP2031)16周,诱导高血脂[33,34]。在给药前三天每只小鼠取血50μL,检测总胆固醇浓度,并据其将小鼠随机分为两组,给溶媒PBS对照组6只以及给纤溶酶原组5只。开始给药记为第1天,给纤溶酶原组小鼠尾静脉注射人源纤溶酶原1mg/0.1ml/只/天,给溶媒PBS对照组尾静脉注射同体积的 PBS。给药30天,期间一直饲喂高脂高胆固醇饲料。于第31天处死小鼠,取心脏组织于4%多聚甲醛固定24-48小时。固定后的组织经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为3μm,切片脱蜡复水后水洗1次,以0.1%天狼星红饱和苦味酸染色30分钟后,流水冲洗2min,苏木素染色1分钟,流水冲洗,1%盐酸酒精分化,氨水返蓝,流水冲洗,烘干后中性树胶封片,在200倍光学显微镜下观察。
结果显示,给纤溶酶原组(图6B)胶原沉积(箭头标识)明显少于给溶媒PBS对照组(图6A),提示纤溶酶原能通过减少高血脂模型小鼠心脏组织胶原蛋白沉积,从而阻止和降低高血脂所致的心脏纤维化。
实施例7纤溶酶原减少糖尿病小鼠胰岛胶原沉积
24-25周龄db/db雄性小鼠16只,随机分为两组,给纤溶酶原组10 只,给溶媒PBS对照组6只。给纤溶酶原组尾静脉注射人源纤溶酶原 2mg/0.2ml/只/天,给溶媒PBS对照组尾静脉注射同体积的PBS。实验开始当天记为第0天称重分组,第1天开始给纤溶酶原或PBS,连续给药31 天。在第32天处死小鼠,取胰脏在4%多聚甲醛中固定。固定后的胰脏组织经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为3μm,切片脱蜡至水后水洗1次,以0.1%天狼星红染色60分钟后,流水冲洗,苏木素染色1分钟,流水冲洗,1%盐酸酒精和氨水分化返蓝,流水冲洗,烘干后封片,切片在200倍光学显微镜下观察。
结果显示,给纤溶酶原组小鼠(图7B)胰岛胶原沉积(箭头标识)明显低于给溶媒PBS对照组(图7A),且统计差异显著(图7C)。说明纤溶酶原能显著减轻糖尿病小鼠胰腺组织中胶原蛋白的沉积,阻止和减轻胰腺的损伤和纤维化。
实施例8纤溶酶原改善ApoE动脉粥样硬化小鼠主动脉窦纤维化
6周龄雄性ApoE小鼠13只饲喂高脂高胆固醇饲料(南通特洛菲,TP2031)16周以诱导动脉粥样硬化模型[31,32]。成模后的小鼠在给药前三天每只取血50μl以检测总胆固醇(T-CHO)含量,并根据T-CHO含量随机分为两组,给溶媒PBS对照组7只,给纤溶酶原组6只。开始给药定为第 1天,给纤溶酶原组小鼠尾静脉注射人源纤溶酶原1mg/0.1mL/只/天,给溶媒PBS对照组尾静脉注射同体积的PBS。给药30天,并于第31天处死小鼠,取心脏于4%多聚甲醛固定24-48小时,分别于15%、30%蔗糖中4℃过夜沉底,OCT包埋,冰冻切片厚度8μm,以0.1%天狼星红饱和苦味酸染色30分钟后,流水冲洗2min,苏木素染色1分钟,流水冲洗,1%盐酸酒精分化,氨水返蓝,流水冲洗,烘干后中性树胶封片,切片在40(8A、 8B)、200倍(8C、8D)光学显微镜下观察。
结果显示,给纤溶酶原组(图8B、D)主动脉窦管壁内膜胶原蛋白沉积(箭头标识)的面积明显小于给溶媒PBS对照组(图8A、C),说明纤溶酶原能够消减动脉粥硬化模型小鼠主动脉窦纤维化水平。
实施例9纤溶酶原改善四氯化碳诱导的肝脏纤维化
9周龄C57雌性小鼠15只,随机分为三组,空白对照组、给溶媒PBS 对照组和给纤溶酶原组,每组各5只。给溶媒PBS对照组和给纤溶酶原组小鼠按照1mL/kg体重腹腔注射四氯化碳,每周三次,连续注射两周,建立肝纤维化模型[36,37],空白对照小鼠按照模型小鼠注射方式注射相应体积的玉米油。四氯化碳需用玉米油稀释,四氯化碳与玉米油稀释比例为1:3。模型建立后开始给药,开始给药当天记为第1天,给纤溶酶原组小鼠按照1mg/0.1mL/只/天尾静脉注射人源纤溶酶原,给溶媒PBS对照组尾静脉注射同体积的PBS,空白对照组不做注射处理,连续给药14天。第15天处死小鼠取肝脏,于4%多聚甲醛中固定24小时。固定后的肝脏经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为3μm,切片脱蜡至水后水洗1次,以0.1%天狼星红染色60分钟后,流水冲洗,苏木素染色1分钟,流水冲洗,1%盐酸酒精和氨水分化返蓝,流水冲洗,烘干后封片,切片在200倍光学显微镜下观察。
结果显示,给纤溶酶原组(图9C)胶原蛋白沉积明显少于给溶媒PBS 对照组(图9B),且给纤溶酶原组比给PBS组胶原蛋白沉积水平更接近空白对照小鼠(图9A)。说明纤溶酶原能减少肝脏胶原蛋白的沉积,改善肝纤维化模型小鼠肝脏纤维化。
实施例10纤溶酶原降低16周高脂血症模型小鼠主动脉窦纤维化
6周龄雄性C57小鼠11只饲喂高脂高胆固醇饲料(南通特洛菲,货号 TP2031)16周以诱导高脂血症模型[30,31],此模型定为16周高脂血症模型。成模后的小鼠继续饲喂高胆固醇饲料。在给药前三天每只取血50μl以检测总胆固醇(T-CHO)含量,并根据T-CHO含量随机分为两组,给溶媒PBS 对照组6只,给纤溶酶原组5只。开始给药记为第1天,给纤溶酶原组尾静脉注射人源纤溶酶原1mg/0.1ml/只/天,给溶媒PBS对照组尾静脉注射同体积的PBS。给药30天,于第31天处死小鼠,取材心脏于4%多聚甲醛固定24-48小时。固定后的组织经酒精梯度脱水和二甲苯透明后进行石蜡包埋。主动脉窦切片厚度为3μm,切片脱蜡复水后水洗1次,以0.1%天狼星红饱和苦味酸染色30分钟后,流水冲洗2min,苏木素染色1分钟,流水冲洗,1%盐酸酒精分化,氨水返蓝,流水冲洗,烘干后中性树胶封片,在40 (10A、10B)、200倍(10C、10D)光学显微镜下观察。
结果显示,给纤溶酶原组(图10B、10D)主动脉窦管壁内膜胶原蛋白沉积(箭头标识)的面积明显小于给溶媒PBS对照组(图10A、10C),说明纤溶酶原能够消减高脂血症模型小鼠主动脉窦纤维化水平。
实施例11纤溶酶原降低系统性硬化症小鼠皮肤纤维化
取12周龄C57雄鼠15只,随机分为三组,空白对照组、给溶媒PBS (PBS为磷酸缓冲盐溶液(Phosphate Buffer Saline),本文中为纤溶酶原的溶媒。)对照组和给纤溶酶原组,每组各5只,并取13周龄PLG活性受损小鼠5只。实验开始当天记为第0天称重分组,第二天开始造模给药,给溶媒PBS对照组、给纤溶酶原组以及PLG活性受损小鼠按0.1mg/0.1ml/只/天皮下注射博莱霉素诱导系统性硬化症[26]。空白对照组按0.1ml/只/天皮下注射生理盐水,同时开始给纤溶酶原或PBS并记为第1天,连续造模给药21 天。给纤溶酶原组小鼠按1mg/0.1ml/只/天尾静脉注射纤溶酶原,给溶媒 PBS对照组给予相同体积的PBS,正常小鼠组和PLG活性受损小鼠不做给药处理。在第22天处死小鼠并取背部皮肤组织在4%多聚甲醛固定液中固定24小时。固定的皮肤组织经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为3μm,切片脱蜡至水后水洗1次,以0.1%天狼星红饱和苦味酸染色30分钟后,流水冲洗2min,苏木素染色1分钟,流水冲洗, 1%盐酸酒精分化,氨水返蓝,流水冲洗,烘干后中性树胶封片,在100倍光学显微镜下观察。
天狼星红染色可使胶原持久染色,作为病理切片特殊染色方法,天狼星红染色可以特异显示胶原组织。
结果显示,在博莱霉素诱导的系统性硬化症小鼠模型中,显微镜下观察到给溶媒PBS组(图11B)和PLG活性受损组小鼠(图11D)真皮上部胶原纤维束明显增多,胶原纤维粗大,排列致密,真皮层增厚;给纤溶酶原组(图11C),真皮层中成纤维细胞明显少于给溶媒PBS对照组,皮肤真皮层厚度基本接近正常水平(图11A)。说明纤溶酶原能有效降低博莱霉素诱导的皮肤纤维化。
实施例12纤溶酶原降低系统性硬化症小鼠肺纤维化
取12周龄C57雄鼠17只,随机分为两组,给溶媒PBS对照组11只和给纤溶酶原组6只。实验开始当天记为第0天并称重分组,第1天开始造模给药,两组小鼠按0.1mg/0.1ml/只/天皮下注射博莱霉素诱导系统性硬化症[26],并开始给纤溶酶原或PBS,连续造模给药21天。给纤溶酶原组小鼠按1mg/0.1ml/只/天尾静脉注射纤溶酶原,给溶媒PBS对照组以相同方式给予相同体积的PBS。第22天处死小鼠并取肺组织在4%多聚甲醛固定液中固定24小时。固定后的肺组织经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为3μm,切片脱蜡至水后水洗1次,以0.1%天狼星红饱和苦味酸染色30分钟后,流水冲洗2min,苏木素染色1分钟,流水冲洗, 1%盐酸酒精分化,氨水返蓝,流水冲洗,烘干后中性树胶封片,在200倍光学显微镜下观察。
研究发现,在博莱霉素诱导的系统性硬化症小鼠模型中,镜下观察给溶媒PBS组(图12A)胶原纤维化(箭头标识)程度比给纤溶酶原组(图 12B)高;给纤溶酶原组小鼠肺部肺泡壁形态接近正常水平,炎症细胞明显减少,纤维化程度明显低于给溶媒PBS组,且统计差异显著(图12C)。说明纤溶酶原可有效降低博莱霉素诱导的系统性硬化症小鼠肺组织纤维化。
实施例13纤溶酶原降低系统性硬化症小鼠肾脏纤维化
取12周龄C57雄鼠10只以,随机分为两组,给溶媒PBS对照组和给纤溶酶原组各5只。实验开始当天记为第0天并称重分组,第1天开始造模给药,所有小鼠按0.1mg/0.1ml/只/天皮下注射博莱霉素诱导系统性硬化症,并开始给纤溶酶原或PBS,连续造模给药21天。给纤溶酶原组小鼠按 1mg/0.1ml/只/天尾静脉注射纤溶酶原,给溶媒PBS对照组尾静脉注射相同体积的PBS。在第22天处死小鼠并取肾脏在4%多聚甲醛固定液中固定24 小时。固定后的肾脏经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为3μm,切片脱蜡至水后水洗1次,以0.1%天狼星红饱和苦味酸染色30分钟后,流水冲洗2min,苏木素染色1分钟,流水冲洗,1%盐酸酒精分化,氨水返蓝,流水冲洗,烘干后中性树胶封片,在200倍光学显微镜下观察。
结果显示,在博莱霉素诱导的系统性硬化症小鼠模型中,给溶媒PBS 对照组(图13A)肾脏胶原纤维化(箭头标识)程度比给纤溶酶原组(图 13B)高。说明纤溶酶原能有效降低博莱霉素诱导的肾脏纤维化。
实施例14纤溶酶原降低糖尿病小鼠肾脏胶原蛋白沉积
24-25周龄db/db雄鼠10只,随机分为两组,给溶媒PBS对照组和给纤溶酶原组,每组各5只。实验开始当天记为第0天并称重分组,第1天开始给纤溶酶原或PBS,连续给药31天。给纤溶酶原组小鼠按2mg/0.2ml/只/ 天尾静脉注射纤溶酶原,给溶媒PBS对照组给予相同体积的PBS。给纤溶酶原31天后处死小鼠并取肾脏组织在4%多聚甲醛固定液中固定24小时。固定后的肾脏组织经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为4μm,切片脱蜡复水后水洗1次。以3%双氧水孵育15分钟, 0.01MPBS洗2次,每次5分钟。10%正常羊血清(Vector laboratories, Inc.,USA)封闭1小时,时间到后甩去血清,用PAP笔圈出组织。针对IV 胶原的兔抗小鼠多克隆抗体(Abcam)4℃孵育过夜,TBS洗2次,每次5 分钟。山羊抗兔IgG(HRP)抗体(Abcam)二抗室温孵育1小时,TBS洗2 次。按DAB试剂盒(Vector laboratories,Inc.,USA)显色,水洗3次后苏木素复染30秒,流水冲洗5分钟,梯度酒精脱水,二甲苯透明透明,中性树胶封片,切片在200倍光学显微镜下观察。
糖尿病肾病是糖尿病慢性并发症,肾小球硬化以及肾间质纤维化是其典型的病理改变[27]。
结果显示,给纤溶酶原组(图14B)IV胶原阳性着色明显多于给溶媒 PBS对照组(图14A),说明纤溶酶原能降低肾脏组织胶原蛋白的沉积 (箭头标识),提示纤溶酶原有望通过降低肾脏组织胶原蛋白的沉积,阻止糖尿病所致的肾脏组织纤维化。
实施例15纤溶酶原改善糖尿病小鼠肾脏纤维化
26周龄db/db雄鼠10只,随机分为两组,给溶媒PBS对照组和给纤溶酶原组,每组各5只。实验开始当天记为第0天并称重分组,第1天开始给纤溶酶原或PBS,连续给药35天。给纤溶酶原组小鼠按2mg/0.2ml/只/天尾静脉注射纤溶酶原,给溶媒PBS对照组给予相同体积的PBS。第36天处死小鼠并取肾脏组织在4%多聚甲醛固定液中固定24小时。固定后的肾脏组织经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为 4μm,切片脱蜡复水后放入重铬酸钾溶液过夜。铁苏木素染3到5分钟,流水稍洗。1%盐酸酒精分化,氨水处理1秒,水洗。丽春红酸性品红液染8分钟,水中快速漂洗。1%磷钼酸水溶液处理约2分钟,苯胺蓝液复染6分钟。1%冰醋酸漂洗1分钟左右。无水乙醇脱水二甲苯透明后封片,切片在 200倍光学显微镜下观察。
Masson染色可以显示组织的纤维化。结果显示,给溶媒PBS对照组 (图15A)肾小球系膜增生,系膜基质增多,肾间质轻度纤维化(箭头标识),增生的纤维化呈蓝色。给纤溶酶原组(图15B)肾小球系膜细胞及基质明显比对照组少,肾间质纤维化明显减少。说明纤溶酶原能改善糖尿病小鼠肾脏的纤维化。
实施例16纤溶酶原降低顺铂肾脏纤维化模型小鼠肾脏的纤维化
8-9周龄健康的雄性C57小鼠10只,随机分为两组,给溶媒PBS对照组和给纤溶酶原组,每组各5只。分组完成后,按10mg/Kg体重单次腹腔注射顺铂,建立肾脏纤维化模型[30]。建立模型后,给纤溶酶原组 1mg/0.1ml/只/天经尾静脉注射给予纤溶酶原,给溶媒PBS对照组给予相同体积的PBS。实验开始当天记为第0天称体重并分组,第1天开始腹腔注射顺铂造模,造模后3小时内给予纤溶酶原或溶媒PBS,给药周期7天。第8天处死小鼠,取肾脏在4%多聚甲醛固定液中固定24小时。固定后的肾脏组织经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为 5μm,切片脱蜡复水后水洗1次。柠檬酸修复30分钟,室温冷却10分钟后水轻柔冲洗。以3%双氧水孵育15分钟,用PAP笔圈出组织。10%的羊血清(Vector laboratories,Inc.,USA)封闭1小时;时间到后,弃除羊血清液。兔抗小鼠IV胶原抗体(Abcam)4℃孵育过夜,TBS洗2次,每次5 分钟。山羊抗兔IgG(HRP)抗体(Abcam)二抗室温孵育1小时,TBS洗2 次,每次5分钟。按DAB试剂盒(Vector laboratories,Inc.,USA)显色,水洗3次后苏木素复染30秒,流水返蓝5分钟,然后TBS洗1次。梯度脱水透明并封片,切片在200倍光学显微镜下观察。
顺铂是临床上应用广泛、疗效可靠的广谱抗肿瘤药物,但具有严重的肾毒性,主要是肾小管及肾间质损伤,最终发展为肾纤维化[30]。该实验结果显示,给溶媒PBS对照组(图16A)肾脏IV型胶原阳性表达(箭头标识)明显高于给溶纤溶酶原组(图16B)。说明纤溶酶原可改善顺铂肾脏纤维化模型小鼠肾脏的纤维化。
实施例17纤溶酶原修复慢性肾衰竭模型肾脏的纤维化
8-9周龄的PLG活性正常雄性小鼠12只以及PLG活性受损雄性小鼠6 只,PLG活性正常小鼠随机分为两组,给纤溶酶原组和给溶媒PBS对照组各6只。三组小鼠每天饲喂0.25%嘌呤饲料(南通特洛菲),建立慢性肾衰竭模型[35]。造模当天记为第1天,同时开始给药。给纤溶酶原组按1mg/0.1ml/ 只/天给予纤溶酶原,给溶媒PBS对照组以相同方式给予相同体积的PBS,连续造模给药10天,PLG活性受损小鼠不做处理。第11天处死小鼠取肾脏于4%多聚甲醛中固定24小时。固定后的肾脏经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为3μm,切片脱蜡至水后水洗1次,以0.1%天狼星红染色60分钟后,流水冲洗,苏木素染色1分钟,流水冲洗,1%盐酸酒精和氨水分化返蓝,流水冲洗,烘干后封片,切片在200倍光学显微镜下观察。
结果显示,给纤溶酶原组(图17B)胶原的沉积(箭头标识)要明显少于给溶媒PBS对照组(图17A)和PLG活性受损组(图17C),且给纤溶酶原组与PLG活性受损组统计差异显著(P=0.018)(图17D)。说明纤溶酶原能显著减轻慢性肾损伤动物肾脏组织中胶原蛋白的沉积,阻止和减轻慢性肾损伤所致的肾脏纤维化。
实施例18纤溶酶原降低3%胆固醇高脂血症模型小鼠肾脏纤维化
9周龄雄性C57小鼠16只饲喂3%胆固醇高脂饲料(南通特洛菲)4周,诱导高脂血症[30,31],此模型定为3%胆固醇高脂血症模型,成模后的小鼠继续饲喂3%胆固醇高脂饲料。另取相同周龄的雄性C57小鼠5只作为空白对照组,实验期间饲喂普通维持饲料。在给药前三天每只小鼠取血50μL,检测总胆固醇,模型小鼠根据总胆固醇浓度和体重随机分为两组,给纤溶酶原组和给溶媒PBS对照组,每组各8只。开始给药记为第1天,给纤溶酶原组小鼠尾静脉注射人源纤溶酶原1mg/0.1ml/只/天,给溶媒PBS对照组尾静脉注射同体积的PBS。在第30天给药后小鼠给药30天,于第31天处死小鼠,取材肾脏于4%多聚甲醛固定24-48小时。固定后的组织经酒精梯度脱水和二甲苯透明后进行石蜡包埋。切片厚度为3μm,切片脱蜡复水后水洗1次,以0.1%天狼星红饱和苦味酸染色30分钟后,流水冲洗2min,苏木素染色1分钟,流水冲洗,1%盐酸酒精分化,氨水返蓝,流水冲洗,烘干后中性树胶封片,在200倍光学显微镜下观察。
结果显示,给纤溶酶原组(图18C)肾脏胶原蛋白沉积(箭头标识) 明显少于给溶媒PBS对照组(图18B),且统计差异显著(图18D);给纤溶酶原组纤维化基本恢复到正常水平(图18A)。说明纤溶酶原能有效的减少3%胆固醇高脂血症模型小鼠肾脏纤维化。
实施例19纤溶酶原减少四氯化碳诱导肝脏纤维化过程肝脏胶原蛋白沉积
7-8周龄C57雌性小鼠20只,随机分为三组,空白对照组5只、给溶媒PBS对照组7只和给纤溶酶原组8只。给溶媒PBS对照组和给纤溶酶原组小鼠按照1mL/kg体重腹腔注射四氯化碳,每周三次,连续注射四周,建立肝纤维化模型[36,37],空白对照小鼠腹腔注射相应体积的玉米油。四氯化碳需用玉米油稀释,四氯化碳与玉米油稀释比例为1:3。造模当天开始给药,记为第1天,给纤溶酶原组小鼠尾静脉注射人源纤溶酶原1mg/0.1mL/ 只/天,给溶媒PBS对照组尾静脉注射同体积的PBS,空白对照组不做注射处理,连续给药28天。第29天处死小鼠取肝脏于4%多聚甲醛中固定24 小时。固定后的肝脏经酒精梯度脱水和二甲苯透明后进行石蜡包埋。组织切片厚度为3μm,切片脱蜡至水后水洗1次,以0.1%天狼星红染色60分钟后,流水冲洗,苏木素染色1分钟,流水冲洗,1%盐酸酒精和氨水分化返蓝,流水冲洗,烘干后封片,切片在200倍光学显微镜下观察。
结果显示,给纤溶酶原组(图19C)胶原蛋白沉积明显少于给溶媒 PBS对照组(图19B),且统计差异显著(图19D);与给溶媒PBS对照组相比,给纤溶酶原组胶原沉积(箭头标识)水平更加接近空白对照小鼠 (图19A)。说明纤溶酶原能减少肝纤维化模型小鼠肝脏胶原蛋白的沉积,改善肝脏纤维化。
参考文献
[1]Denton CP,Black CM,Abraham DJ.Mechanisms and consequences of fibosis in systemic sclerosis[J].Nat Clin Pract Rheumatol,2006,2(3):134-144.
[2]Wilson MS,Wynn TA.Pulmonary fibrosis:pathogenesis,etiology and regulation[J]. Mucosal Immunol,2009,2(2):103-121.
[3]Liu Y.Renal fibrosis:new insights into the pathogenesis and therapeutics[J].Kidney Int, 2006,69:213-217.
[4]Alexander,C.M.and Werb,Z.(1989).Proteinases and extracellular matrix remodeling. Curr.Opin.Cell Biol.1,974-982.
[5]Alexander CM and Werb,Z.(1991).Extracellular matrix degradation.In Cell Biology of Extracellular Matrix,Hay ED,ed.(New York:Plenum Press),pp.255-302
[6]Werb,Z.,Mainardi,C.L.,Vater,C.A.,and Harris,E.D.,Jr.(1977).Endogenous activiation of latent collagenase by rheumatoid synovial cells.Evidence for a role of plasminogen activator.N.Engl.J.Med.296,1017-1023.
[7]He,C.S.,Wilhelm,S.M.,Pentland,A.P.,Marmer,B.L.,Grant,G.A.,Eisen,A.Z.,and Goldberg,G.I.(1989).Tissue cooperation in a proteolytic cascade activating human interstitial collagenase.Proc.Natl.Acad.Sci.U.S.A 86,2632-2636
[8]Stoppelli,M.P.,Corti,A.,Soffientini,A.,Cassani,G.,Blasi,F.,and Assoian,R.K.(1985). Differentiation-enhanced binding of the amino-terminal fragment of human urokinase plasminogen activator to a specific receptor on U937 monocytes.Proc.Natl.Acad.Sci.U.S.A 82,4939-4943.
[9]Vassalli,J.D.,Baccino,D.,and Belin,D.(1985).A cellular binding site for the Mr 55,000 form of the human plasminogen activator,urokinase.J.Cell Biol.100,86-92.
[10]Wiman,B.and Wallen,P.(1975).Structural relationship between"glutamic acid"and "lysine"forms of human plasminogen and their interaction with the NH2-terminal activation peptide as studied by affinity chromatography.Eur.J.Biochem.50,489-494.
[11]Saksela,O.and Rifkin,D.B.(1988).Cell-associated plasminogen activation:regulation and physiological functions.Annu.Rev.Cell Biol.4,93-126
[12]Raum,D.,Marcus,D.,Alper,C.A.,Levey,R.,Taylor,P.D.,and Starzl,T.E.(1980). Synthesis of human plasminogen by the liver.Science 208,1036-1037
[13]Wallén P(1980).Biochemistry of plasminogen.In Fibrinolysis,Kline DL and Reddy KKN,eds.(Florida:CRC
[14]Sottrup-Jensen,L.,Zajdel,M.,Claeys,H.,Petersen,T.E.,and Magnusson,S.(1975). Amino-acid sequence of activation cleavage site in plasminogen:homology with"pro"part of prothrombin.Proc.Natl.Acad.Sci.U.S.A 72,2577-2581.
[15]Collen,D.and Lijnen,H.R.(1991).Basic and clinical aspects of fibrinolysis and thrombolysis.Blood 78,3114-3124.
[16]Alexander,C.M.and Werb,Z.(1989).Proteinases and extracellular matrix remodeling. Curr.Opin.Cell Biol.1,974-982.
[17]Mignatti,P.and Rifkin,D.B.(1993).Biology and biochemistry of proteinases in tumor invasion.Physiol Rev.73,161-195.
[18]Collen,D.(2001).Ham-Wasserman lecture:role of the plasminogen system in fibrin- homeostasis and tissue remodeling.Hematology.(Am.Soc.Hematol.Educ.Program.)1-9.
[19]Rifkin,D.B.,Moscatelli,D.,Bizik,J.,Quarto,N.,Blei,F.,Dennis,P.,Flaumenhaft,R., and Mignatti,P.(1990).Growth factor control of extracellular proteolysis.Cell Differ.Dev.32, 313-318.
[20]Andreasen,P.A.,Kjoller,L.,Christensen,L.,and Duffy,M.J.(1997).The urokinase-type plasminogen activator system in cancer metastasis:a review.Int.J.Cancer 72,1-22.
[21]Rifkin,D.B.,Mazzieri,R.,Munger,J.S.,Noguera,I.,and Sung,J.(1999).Proteolytic control of growth factor availability.APMIS 107,80-85.
[22]Marder V J,Novokhatny V.Direct fibrinolytic agents:biochemical attributes,preclinical foundation and clinical potential[J].Journal of Thrombosis and Haemostasis,2010,8(3):433- 444.
[23]Hunt J A,Petteway Jr S R,Scuderi P,et al.Simplified recombinant plasmin:production and fu-nctional comparison of a novel thrombolytic molecule with plasma-derived plasmin[J]. Thromb Haemost,2008,100(3):413-419.
[24]Sottrup-Jensen L,Claeys H,Zajdel M,et al.The primary structure of human plasminogen: Isolation of two lysine-binding fragments and one“mini”-plasminogen(MW,38,000)by elastase-catalyzed-specific limited proteolysis[J].Progress in chemical fibrinolysis and thrombolysis,1978,3:191-209
[25]Nagai N,Demarsin E,Van Hoef B,et al.Recombinant human microplasmin:production and potential therapeutic properties[J].Journal of Thrombosis and Haemostasis,2003,1(2): 307-313.
[26]Yosuke Kanno,En Shu,Hiroyuki Kanoh et al.The Antifibrotic Effect of a2AP Neutralization in Systemic Sclerosis Dermal Fibroblasts and Mouse Models of Systemic Sclerosis.J Invest Dermatol.2016 Apr;136(4):762-9.
[27]Donnelly SM,Zhou XP,Huang JT et al.Prevention of early glomerulopathy with tolrestat in the streptozotocin-induced diabetic rat.Biochem Cell Biol.1996;74(3):355-62.
[28]Ashish Aneja,W.H.Wilson Tang,Sameer Bansilal et al.Diabetic Cardiomyopathy: Insights into Pathogenesis,Diagnostic Challenges,and Therapeutic Options.Am J Med.2008 Sep;121(9):748-57.
[29]SamuelCS1,HewitsonTD,ZhangYetal.Relaxin ameliorates fibrosis in experimental diabet c cardiomyopathy.Endocrinology.2008 Jul;149(7):3286-93.
[30]Yutaka Nakashima,Andrew S.Plump,Elaine W.Raines et al.Arterioscler Thromb.1994 Jan;14(1):133-40.
[31]Yutaka Nakashima,Andrew S.Plump,Elaine W.Raines et al.Arterioscler Thromb.1994 Jan;14(1):133-40.
[32]Yvonne Nitschke,Gabriele Weissen-Plenz,Robert Terkeltaub et al.Npp1 promotes atherosclerosis in ApoE knockout mice.J.Cell.Mol.Med.Vol 15,No 11,2011 pp.2273-2283
[33]Dominika Nackiewicz,Paromita Dey,Barbara Szczerba et al.Inhibitor of differentiation 3, a transcription factor regulates hyperlipidemia associated kidney disease.Nephron Exp Nephrol.2014;126(3):141–147.
[34]Ming Gu1,Yu Zhang.,Shengjie Fan et al.Extracts of Rhizoma Polygonati Odorati Prevent High-Fat Diet-Induced Metabolic Disorders in C57BL/6 Mice.PLoS ONE 8(11): e81724.
[35]Cristhiane Favero Aguiar,Cristiane Naffah-de-Souza,Angela Castoldi et al. Administration ofα-Galactosylceramide Improves Adenine-Induced Renal Injury.Mol Med.2015 Jun 18;21:553-62.
[36]Mark A.Barnes,Megan R.McMullen,Sanjoy Roychowdhury et al.Macrophage migration inhibitory factor is required for recruitment of scar-associated macrophages during liver fibrosis.J Leukoc Biol.2015 Jan;97(1):161-9.
[37]Takayoshi Yamaza,Fatima Safira Alatas,Ratih Yuniartha et al.In vivo hepatogenic capacity and therapeutic potential of stem cells from human exfoliated deciduous teeth in liver fibrosis in mice.Stem Cell Res Ther.2015 Sep 10;6:171.
序列表
<110> 深圳瑞健生命科学研究院有限公司
<120> 一种预防和治疗组织器官纤维化的方法
<130> PDK03583
<160> 14
<170> PatentIn version 3.5
<210> 1
<211> 2376
<212> DNA
<213> 不含有信号肽的天然纤溶酶原(Glu-PLG,Glu-纤维蛋白溶酶原)核酸序列
<400> 1
gagcctctgg atgactatgt gaatacccag ggggcttcac tgttcagtgt cactaagaag 60
cagctgggag caggaagtat agaagaatgt gcagcaaaat gtgaggagga cgaagaattc 120
acctgcaggg cattccaata tcacagtaaa gagcaacaat gtgtgataat ggctgaaaac 180
aggaagtcct ccataatcat taggatgaga gatgtagttt tatttgaaaa gaaagtgtat 240
ctctcagagt gcaagactgg gaatggaaag aactacagag ggacgatgtc caaaacaaaa 300
aatggcatca cctgtcaaaa atggagttcc acttctcccc acagacctag attctcacct 360
gctacacacc cctcagaggg actggaggag aactactgca ggaatccaga caacgatccg 420
caggggccct ggtgctatac tactgatcca gaaaagagat atgactactg cgacattctt 480
gagtgtgaag aggaatgtat gcattgcagt ggagaaaact atgacggcaa aatttccaag 540
accatgtctg gactggaatg ccaggcctgg gactctcaga gcccacacgc tcatggatac 600
attccttcca aatttccaaa caagaacctg aagaagaatt actgtcgtaa ccccgatagg 660
gagctgcggc cttggtgttt caccaccgac cccaacaagc gctgggaact ttgtgacatc 720
ccccgctgca caacacctcc accatcttct ggtcccacct accagtgtct gaagggaaca 780
ggtgaaaact atcgcgggaa tgtggctgtt accgtgtccg ggcacacctg tcagcactgg 840
agtgcacaga cccctcacac acataacagg acaccagaaa acttcccctg caaaaatttg 900
gatgaaaact actgccgcaa tcctgacgga aaaagggccc catggtgcca tacaaccaac 960
agccaagtgc ggtgggagta ctgtaagata ccgtcctgtg actcctcccc agtatccacg 1020
gaacaattgg ctcccacagc accacctgag ctaacccctg tggtccagga ctgctaccat 1080
ggtgatggac agagctaccg aggcacatcc tccaccacca ccacaggaaa gaagtgtcag 1140
tcttggtcat ctatgacacc acaccggcac cagaagaccc cagaaaacta cccaaatgct 1200
ggcctgacaa tgaactactg caggaatcca gatgccgata aaggcccctg gtgttttacc 1260
acagacccca gcgtcaggtg ggagtactgc aacctgaaaa aatgctcagg aacagaagcg 1320
agtgttgtag cacctccgcc tgttgtcctg cttccagatg tagagactcc ttccgaagaa 1380
gactgtatgt ttgggaatgg gaaaggatac cgaggcaaga gggcgaccac tgttactggg 1440
acgccatgcc aggactgggc tgcccaggag ccccatagac acagcatttt cactccagag 1500
acaaatccac gggcgggtct ggaaaaaaat tactgccgta accctgatgg tgatgtaggt 1560
ggtccctggt gctacacgac aaatccaaga aaactttacg actactgtga tgtccctcag 1620
tgtgcggccc cttcatttga ttgtgggaag cctcaagtgg agccgaagaa atgtcctgga 1680
agggttgtag gggggtgtgt ggcccaccca cattcctggc cctggcaagt cagtcttaga 1740
acaaggtttg gaatgcactt ctgtggaggc accttgatat ccccagagtg ggtgttgact 1800
gctgcccact gcttggagaa gtccccaagg ccttcatcct acaaggtcat cctgggtgca 1860
caccaagaag tgaatctcga accgcatgtt caggaaatag aagtgtctag gctgttcttg 1920
gagcccacac gaaaagatat tgccttgcta aagctaagca gtcctgccgt catcactgac 1980
aaagtaatcc cagcttgtct gccatcccca aattatgtgg tcgctgaccg gaccgaatgt 2040
ttcatcactg gctggggaga aacccaaggt acttttggag ctggccttct caaggaagcc 2100
cagctccctg tgattgagaa taaagtgtgc aatcgctatg agtttctgaa tggaagagtc 2160
caatccaccg aactctgtgc tgggcatttg gccggaggca ctgacagttg ccagggtgac 2220
agtggaggtc ctctggtttg cttcgagaag gacaaataca ttttacaagg agtcacttct 2280
tggggtcttg gctgtgcacg ccccaataag cctggtgtct atgttcgtgt ttcaaggttt 2340
gttacttgga ttgagggagt gatgagaaat aattaa 2376
<210> 2
<211> 791
<212> PRT
<213> 不含有信号肽的天然纤溶酶原(Glu-PLG,Glu-纤维蛋白溶酶原)氨基酸序列
<400> 2
Glu Pro Leu Asp Asp Tyr Val Asn Thr Gln Gly Ala Ser Leu Phe Ser
1 5 10 15
Val Thr Lys Lys Gln Leu Gly Ala Gly Ser Ile Glu Glu Cys Ala Ala
20 25 30
Lys Cys Glu Glu Asp Glu Glu Phe Thr Cys Arg Ala Phe Gln Tyr His
35 40 45
Ser Lys Glu Gln Gln Cys Val Ile Met Ala Glu Asn Arg Lys Ser Ser
50 55 60
Ile Ile Ile Arg Met Arg Asp Val Val Leu Phe Glu Lys Lys Val Tyr
65 70 75 80
Leu Ser Glu Cys Lys Thr Gly Asn Gly Lys Asn Tyr Arg Gly Thr Met
85 90 95
Ser Lys Thr Lys Asn Gly Ile Thr Cys Gln Lys Trp Ser Ser Thr Ser
100 105 110
Pro His Arg Pro Arg Phe Ser Pro Ala Thr His Pro Ser Glu Gly Leu
115 120 125
Glu Glu Asn Tyr Cys Arg Asn Pro Asp Asn Asp Pro Gln Gly Pro Trp
130 135 140
Cys Tyr Thr Thr Asp Pro Glu Lys Arg Tyr Asp Tyr Cys Asp Ile Leu
145 150 155 160
Glu Cys Glu Glu Glu Cys Met His Cys Ser Gly Glu Asn Tyr Asp Gly
165 170 175
Lys Ile Ser Lys Thr Met Ser Gly Leu Glu Cys Gln Ala Trp Asp Ser
180 185 190
Gln Ser Pro His Ala His Gly Tyr Ile Pro Ser Lys Phe Pro Asn Lys
195 200 205
Asn Leu Lys Lys Asn Tyr Cys Arg Asn Pro Asp Arg Glu Leu Arg Pro
210 215 220
Trp Cys Phe Thr Thr Asp Pro Asn Lys Arg Trp Glu Leu Cys Asp Ile
225 230 235 240
Pro Arg Cys Thr Thr Pro Pro Pro Ser Ser Gly Pro Thr Tyr Gln Cys
245 250 255
Leu Lys Gly Thr Gly Glu Asn Tyr Arg Gly Asn Val Ala Val Thr Val
260 265 270
Ser Gly His Thr Cys Gln His Trp Ser Ala Gln Thr Pro His Thr His
275 280 285
Asn Arg Thr Pro Glu Asn Phe Pro Cys Lys Asn Leu Asp Glu Asn Tyr
290 295 300
Cys Arg Asn Pro Asp Gly Lys Arg Ala Pro Trp Cys His Thr Thr Asn
305 310 315 320
Ser Gln Val Arg Trp Glu Tyr Cys Lys Ile Pro Ser Cys Asp Ser Ser
325 330 335
Pro Val Ser Thr Glu Gln Leu Ala Pro Thr Ala Pro Pro Glu Leu Thr
340 345 350
Pro Val Val Gln Asp Cys Tyr His Gly Asp Gly Gln Ser Tyr Arg Gly
355 360 365
Thr Ser Ser Thr Thr Thr Thr Gly Lys Lys Cys Gln Ser Trp Ser Ser
370 375 380
Met Thr Pro His Arg His Gln Lys Thr Pro Glu Asn Tyr Pro Asn Ala
385 390 395 400
Gly Leu Thr Met Asn Tyr Cys Arg Asn Pro Asp Ala Asp Lys Gly Pro
405 410 415
Trp Cys Phe Thr Thr Asp Pro Ser Val Arg Trp Glu Tyr Cys Asn Leu
420 425 430
Lys Lys Cys Ser Gly Thr Glu Ala Ser Val Val Ala Pro Pro Pro Val
435 440 445
Val Leu Leu Pro Asp Val Glu Thr Pro Ser Glu Glu Asp Cys Met Phe
450 455 460
Gly Asn Gly Lys Gly Tyr Arg Gly Lys Arg Ala Thr Thr Val Thr Gly
465 470 475 480
Thr Pro Cys Gln Asp Trp Ala Ala Gln Glu Pro His Arg His Ser Ile
485 490 495
Phe Thr Pro Glu Thr Asn Pro Arg Ala Gly Leu Glu Lys Asn Tyr Cys
500 505 510
Arg Asn Pro Asp Gly Asp Val Gly Gly Pro Trp Cys Tyr Thr Thr Asn
515 520 525
Pro Arg Lys Leu Tyr Asp Tyr Cys Asp Val Pro Gln Cys Ala Ala Pro
530 535 540
Ser Phe Asp Cys Gly Lys Pro Gln Val Glu Pro Lys Lys Cys Pro Gly
545 550 555 560
Arg Val Val Gly Gly Cys Val Ala His Pro His Ser Trp Pro Trp Gln
565 570 575
Val Ser Leu Arg Thr Arg Phe Gly Met His Phe Cys Gly Gly Thr Leu
580 585 590
Ile Ser Pro Glu Trp Val Leu Thr Ala Ala His Cys Leu Glu Lys Ser
595 600 605
Pro Arg Pro Ser Ser Tyr Lys Val Ile Leu Gly Ala His Gln Glu Val
610 615 620
Asn Leu Glu Pro His Val Gln Glu Ile Glu Val Ser Arg Leu Phe Leu
625 630 635 640
Glu Pro Thr Arg Lys Asp Ile Ala Leu Leu Lys Leu Ser Ser Pro Ala
645 650 655
Val Ile Thr Asp Lys Val Ile Pro Ala Cys Leu Pro Ser Pro Asn Tyr
660 665 670
Val Val Ala Asp Arg Thr Glu Cys Phe Ile Thr Gly Trp Gly Glu Thr
675 680 685
Gln Gly Thr Phe Gly Ala Gly Leu Leu Lys Glu Ala Gln Leu Pro Val
690 695 700
Ile Glu Asn Lys Val Cys Asn Arg Tyr Glu Phe Leu Asn Gly Arg Val
705 710 715 720
Gln Ser Thr Glu Leu Cys Ala Gly His Leu Ala Gly Gly Thr Asp Ser
725 730 735
Cys Gln Gly Asp Ser Gly Gly Pro Leu Val Cys Phe Glu Lys Asp Lys
740 745 750
Tyr Ile Leu Gln Gly Val Thr Ser Trp Gly Leu Gly Cys Ala Arg Pro
755 760 765
Asn Lys Pro Gly Val Tyr Val Arg Val Ser Arg Phe Val Thr Trp Ile
770 775 780
Glu Gly Val Met Arg Asn Asn
785 790
<210> 3
<211> 2433
<212> DNA
<213> 含有信号肽的天然纤溶酶原(来源于swiss prot)的核酸序列
<400> 3
atggaacata aggaagtggt tcttctactt cttttatttc tgaaatcagg tcaaggagag 60
cctctggatg actatgtgaa tacccagggg gcttcactgt tcagtgtcac taagaagcag 120
ctgggagcag gaagtataga agaatgtgca gcaaaatgtg aggaggacga agaattcacc 180
tgcagggcat tccaatatca cagtaaagag caacaatgtg tgataatggc tgaaaacagg 240
aagtcctcca taatcattag gatgagagat gtagttttat ttgaaaagaa agtgtatctc 300
tcagagtgca agactgggaa tggaaagaac tacagaggga cgatgtccaa aacaaaaaat 360
ggcatcacct gtcaaaaatg gagttccact tctccccaca gacctagatt ctcacctgct 420
acacacccct cagagggact ggaggagaac tactgcagga atccagacaa cgatccgcag 480
gggccctggt gctatactac tgatccagaa aagagatatg actactgcga cattcttgag 540
tgtgaagagg aatgtatgca ttgcagtgga gaaaactatg acggcaaaat ttccaagacc 600
atgtctggac tggaatgcca ggcctgggac tctcagagcc cacacgctca tggatacatt 660
ccttccaaat ttccaaacaa gaacctgaag aagaattact gtcgtaaccc cgatagggag 720
ctgcggcctt ggtgtttcac caccgacccc aacaagcgct gggaactttg tgacatcccc 780
cgctgcacaa cacctccacc atcttctggt cccacctacc agtgtctgaa gggaacaggt 840
gaaaactatc gcgggaatgt ggctgttacc gtgtccgggc acacctgtca gcactggagt 900
gcacagaccc ctcacacaca taacaggaca ccagaaaact tcccctgcaa aaatttggat 960
gaaaactact gccgcaatcc tgacggaaaa agggccccat ggtgccatac aaccaacagc 1020
caagtgcggt gggagtactg taagataccg tcctgtgact cctccccagt atccacggaa 1080
caattggctc ccacagcacc acctgagcta acccctgtgg tccaggactg ctaccatggt 1140
gatggacaga gctaccgagg cacatcctcc accaccacca caggaaagaa gtgtcagtct 1200
tggtcatcta tgacaccaca ccggcaccag aagaccccag aaaactaccc aaatgctggc 1260
ctgacaatga actactgcag gaatccagat gccgataaag gcccctggtg ttttaccaca 1320
gaccccagcg tcaggtggga gtactgcaac ctgaaaaaat gctcaggaac agaagcgagt 1380
gttgtagcac ctccgcctgt tgtcctgctt ccagatgtag agactccttc cgaagaagac 1440
tgtatgtttg ggaatgggaa aggataccga ggcaagaggg cgaccactgt tactgggacg 1500
ccatgccagg actgggctgc ccaggagccc catagacaca gcattttcac tccagagaca 1560
aatccacggg cgggtctgga aaaaaattac tgccgtaacc ctgatggtga tgtaggtggt 1620
ccctggtgct acacgacaaa tccaagaaaa ctttacgact actgtgatgt ccctcagtgt 1680
gcggcccctt catttgattg tgggaagcct caagtggagc cgaagaaatg tcctggaagg 1740
gttgtagggg ggtgtgtggc ccacccacat tcctggccct ggcaagtcag tcttagaaca 1800
aggtttggaa tgcacttctg tggaggcacc ttgatatccc cagagtgggt gttgactgct 1860
gcccactgct tggagaagtc cccaaggcct tcatcctaca aggtcatcct gggtgcacac 1920
caagaagtga atctcgaacc gcatgttcag gaaatagaag tgtctaggct gttcttggag 1980
cccacacgaa aagatattgc cttgctaaag ctaagcagtc ctgccgtcat cactgacaaa 2040
gtaatcccag cttgtctgcc atccccaaat tatgtggtcg ctgaccggac cgaatgtttc 2100
atcactggct ggggagaaac ccaaggtact tttggagctg gccttctcaa ggaagcccag 2160
ctccctgtga ttgagaataa agtgtgcaat cgctatgagt ttctgaatgg aagagtccaa 2220
tccaccgaac tctgtgctgg gcatttggcc ggaggcactg acagttgcca gggtgacagt 2280
ggaggtcctc tggtttgctt cgagaaggac aaatacattt tacaaggagt cacttcttgg 2340
ggtcttggct gtgcacgccc caataagcct ggtgtctatg ttcgtgtttc aaggtttgtt 2400
acttggattg agggagtgat gagaaataat taa 2433
<210> 4
<211> 810
<212> PRT
<213> 含有信号肽的天然纤溶酶原(来源于swiss prot)的氨基酸序列
<400> 4
Met Glu His Lys Glu Val Val Leu Leu Leu Leu Leu Phe Leu Lys Ser
1 5 10 15
Gly Gln Gly Glu Pro Leu Asp Asp Tyr Val Asn Thr Gln Gly Ala Ser
20 25 30
Leu Phe Ser Val Thr Lys Lys Gln Leu Gly Ala Gly Ser Ile Glu Glu
35 40 45
Cys Ala Ala Lys Cys Glu Glu Asp Glu Glu Phe Thr Cys Arg Ala Phe
50 55 60
Gln Tyr His Ser Lys Glu Gln Gln Cys Val Ile Met Ala Glu Asn Arg
65 70 75 80
Lys Ser Ser Ile Ile Ile Arg Met Arg Asp Val Val Leu Phe Glu Lys
85 90 95
Lys Val Tyr Leu Ser Glu Cys Lys Thr Gly Asn Gly Lys Asn Tyr Arg
100 105 110
Gly Thr Met Ser Lys Thr Lys Asn Gly Ile Thr Cys Gln Lys Trp Ser
115 120 125
Ser Thr Ser Pro His Arg Pro Arg Phe Ser Pro Ala Thr His Pro Ser
130 135 140
Glu Gly Leu Glu Glu Asn Tyr Cys Arg Asn Pro Asp Asn Asp Pro Gln
145 150 155 160
Gly Pro Trp Cys Tyr Thr Thr Asp Pro Glu Lys Arg Tyr Asp Tyr Cys
165 170 175
Asp Ile Leu Glu Cys Glu Glu Glu Cys Met His Cys Ser Gly Glu Asn
180 185 190
Tyr Asp Gly Lys Ile Ser Lys Thr Met Ser Gly Leu Glu Cys Gln Ala
195 200 205
Trp Asp Ser Gln Ser Pro His Ala His Gly Tyr Ile Pro Ser Lys Phe
210 215 220
Pro Asn Lys Asn Leu Lys Lys Asn Tyr Cys Arg Asn Pro Asp Arg Glu
225 230 235 240
Leu Arg Pro Trp Cys Phe Thr Thr Asp Pro Asn Lys Arg Trp Glu Leu
245 250 255
Cys Asp Ile Pro Arg Cys Thr Thr Pro Pro Pro Ser Ser Gly Pro Thr
260 265 270
Tyr Gln Cys Leu Lys Gly Thr Gly Glu Asn Tyr Arg Gly Asn Val Ala
275 280 285
Val Thr Val Ser Gly His Thr Cys Gln His Trp Ser Ala Gln Thr Pro
290 295 300
His Thr His Asn Arg Thr Pro Glu Asn Phe Pro Cys Lys Asn Leu Asp
305 310 315 320
Glu Asn Tyr Cys Arg Asn Pro Asp Gly Lys Arg Ala Pro Trp Cys His
325 330 335
Thr Thr Asn Ser Gln Val Arg Trp Glu Tyr Cys Lys Ile Pro Ser Cys
340 345 350
Asp Ser Ser Pro Val Ser Thr Glu Gln Leu Ala Pro Thr Ala Pro Pro
355 360 365
Glu Leu Thr Pro Val Val Gln Asp Cys Tyr His Gly Asp Gly Gln Ser
370 375 380
Tyr Arg Gly Thr Ser Ser Thr Thr Thr Thr Gly Lys Lys Cys Gln Ser
385 390 395 400
Trp Ser Ser Met Thr Pro His Arg His Gln Lys Thr Pro Glu Asn Tyr
405 410 415
Pro Asn Ala Gly Leu Thr Met Asn Tyr Cys Arg Asn Pro Asp Ala Asp
420 425 430
Lys Gly Pro Trp Cys Phe Thr Thr Asp Pro Ser Val Arg Trp Glu Tyr
435 440 445
Cys Asn Leu Lys Lys Cys Ser Gly Thr Glu Ala Ser Val Val Ala Pro
450 455 460
Pro Pro Val Val Leu Leu Pro Asp Val Glu Thr Pro Ser Glu Glu Asp
465 470 475 480
Cys Met Phe Gly Asn Gly Lys Gly Tyr Arg Gly Lys Arg Ala Thr Thr
485 490 495
Val Thr Gly Thr Pro Cys Gln Asp Trp Ala Ala Gln Glu Pro His Arg
500 505 510
His Ser Ile Phe Thr Pro Glu Thr Asn Pro Arg Ala Gly Leu Glu Lys
515 520 525
Asn Tyr Cys Arg Asn Pro Asp Gly Asp Val Gly Gly Pro Trp Cys Tyr
530 535 540
Thr Thr Asn Pro Arg Lys Leu Tyr Asp Tyr Cys Asp Val Pro Gln Cys
545 550 555 560
Ala Ala Pro Ser Phe Asp Cys Gly Lys Pro Gln Val Glu Pro Lys Lys
565 570 575
Cys Pro Gly Arg Val Val Gly Gly Cys Val Ala His Pro His Ser Trp
580 585 590
Pro Trp Gln Val Ser Leu Arg Thr Arg Phe Gly Met His Phe Cys Gly
595 600 605
Gly Thr Leu Ile Ser Pro Glu Trp Val Leu Thr Ala Ala His Cys Leu
610 615 620
Glu Lys Ser Pro Arg Pro Ser Ser Tyr Lys Val Ile Leu Gly Ala His
625 630 635 640
Gln Glu Val Asn Leu Glu Pro His Val Gln Glu Ile Glu Val Ser Arg
645 650 655
Leu Phe Leu Glu Pro Thr Arg Lys Asp Ile Ala Leu Leu Lys Leu Ser
660 665 670
Ser Pro Ala Val Ile Thr Asp Lys Val Ile Pro Ala Cys Leu Pro Ser
675 680 685
Pro Asn Tyr Val Val Ala Asp Arg Thr Glu Cys Phe Ile Thr Gly Trp
690 695 700
Gly Glu Thr Gln Gly Thr Phe Gly Ala Gly Leu Leu Lys Glu Ala Gln
705 710 715 720
Leu Pro Val Ile Glu Asn Lys Val Cys Asn Arg Tyr Glu Phe Leu Asn
725 730 735
Gly Arg Val Gln Ser Thr Glu Leu Cys Ala Gly His Leu Ala Gly Gly
740 745 750
Thr Asp Ser Cys Gln Gly Asp Ser Gly Gly Pro Leu Val Cys Phe Glu
755 760 765
Lys Asp Lys Tyr Ile Leu Gln Gly Val Thr Ser Trp Gly Leu Gly Cys
770 775 780
Ala Arg Pro Asn Lys Pro Gly Val Tyr Val Arg Val Ser Arg Phe Val
785 790 795 800
Thr Trp Ile Glu Gly Val Met Arg Asn Asn
805 810
<210> 5
<211> 2145
<212> DNA
<213> LYS77-PLG(Lys-纤溶酶原)核酸序列
<400> 5
aaagtgtatc tctcagagtg caagactggg aatggaaaga actacagagg gacgatgtcc 60
aaaacaaaaa atggcatcac ctgtcaaaaa tggagttcca cttctcccca cagacctaga 120
ttctcacctg ctacacaccc ctcagaggga ctggaggaga actactgcag gaatccagac 180
aacgatccgc aggggccctg gtgctatact actgatccag aaaagagata tgactactgc 240
gacattcttg agtgtgaaga ggaatgtatg cattgcagtg gagaaaacta tgacggcaaa 300
atttccaaga ccatgtctgg actggaatgc caggcctggg actctcagag cccacacgct 360
catggataca ttccttccaa atttccaaac aagaacctga agaagaatta ctgtcgtaac 420
cccgataggg agctgcggcc ttggtgtttc accaccgacc ccaacaagcg ctgggaactt 480
tgtgacatcc cccgctgcac aacacctcca ccatcttctg gtcccaccta ccagtgtctg 540
aagggaacag gtgaaaacta tcgcgggaat gtggctgtta ccgtgtccgg gcacacctgt 600
cagcactgga gtgcacagac ccctcacaca cataacagga caccagaaaa cttcccctgc 660
aaaaatttgg atgaaaacta ctgccgcaat cctgacggaa aaagggcccc atggtgccat 720
acaaccaaca gccaagtgcg gtgggagtac tgtaagatac cgtcctgtga ctcctcccca 780
gtatccacgg aacaattggc tcccacagca ccacctgagc taacccctgt ggtccaggac 840
tgctaccatg gtgatggaca gagctaccga ggcacatcct ccaccaccac cacaggaaag 900
aagtgtcagt cttggtcatc tatgacacca caccggcacc agaagacccc agaaaactac 960
ccaaatgctg gcctgacaat gaactactgc aggaatccag atgccgataa aggcccctgg 1020
tgttttacca cagaccccag cgtcaggtgg gagtactgca acctgaaaaa atgctcagga 1080
acagaagcga gtgttgtagc acctccgcct gttgtcctgc ttccagatgt agagactcct 1140
tccgaagaag actgtatgtt tgggaatggg aaaggatacc gaggcaagag ggcgaccact 1200
gttactggga cgccatgcca ggactgggct gcccaggagc cccatagaca cagcattttc 1260
actccagaga caaatccacg ggcgggtctg gaaaaaaatt actgccgtaa ccctgatggt 1320
gatgtaggtg gtccctggtg ctacacgaca aatccaagaa aactttacga ctactgtgat 1380
gtccctcagt gtgcggcccc ttcatttgat tgtgggaagc ctcaagtgga gccgaagaaa 1440
tgtcctggaa gggttgtagg ggggtgtgtg gcccacccac attcctggcc ctggcaagtc 1500
agtcttagaa caaggtttgg aatgcacttc tgtggaggca ccttgatatc cccagagtgg 1560
gtgttgactg ctgcccactg cttggagaag tccccaaggc cttcatccta caaggtcatc 1620
ctgggtgcac accaagaagt gaatctcgaa ccgcatgttc aggaaataga agtgtctagg 1680
ctgttcttgg agcccacacg aaaagatatt gccttgctaa agctaagcag tcctgccgtc 1740
atcactgaca aagtaatccc agcttgtctg ccatccccaa attatgtggt cgctgaccgg 1800
accgaatgtt tcatcactgg ctggggagaa acccaaggta cttttggagc tggccttctc 1860
aaggaagccc agctccctgt gattgagaat aaagtgtgca atcgctatga gtttctgaat 1920
ggaagagtcc aatccaccga actctgtgct gggcatttgg ccggaggcac tgacagttgc 1980
cagggtgaca gtggaggtcc tctggtttgc ttcgagaagg acaaatacat tttacaagga 2040
gtcacttctt ggggtcttgg ctgtgcacgc cccaataagc ctggtgtcta tgttcgtgtt 2100
tcaaggtttg ttacttggat tgagggagtg atgagaaata attaa 2145
<210> 6
<211> 714
<212> PRT
<213> LYS77-PLG(Lys-纤溶酶原)氨基酸序列
<400> 6
Lys Val Tyr Leu Ser Glu Cys Lys Thr Gly Asn Gly Lys Asn Tyr Arg
1 5 10 15
Gly Thr Met Ser Lys Thr Lys Asn Gly Ile Thr Cys Gln Lys Trp Ser
20 25 30
Ser Thr Ser Pro His Arg Pro Arg Phe Ser Pro Ala Thr His Pro Ser
35 40 45
Glu Gly Leu Glu Glu Asn Tyr Cys Arg Asn Pro Asp Asn Asp Pro Gln
50 55 60
Gly Pro Trp Cys Tyr Thr Thr Asp Pro Glu Lys Arg Tyr Asp Tyr Cys
65 70 75 80
Asp Ile Leu Glu Cys Glu Glu Glu Cys Met His Cys Ser Gly Glu Asn
85 90 95
Tyr Asp Gly Lys Ile Ser Lys Thr Met Ser Gly Leu Glu Cys Gln Ala
100 105 110
Trp Asp Ser Gln Ser Pro His Ala His Gly Tyr Ile Pro Ser Lys Phe
115 120 125
Pro Asn Lys Asn Leu Lys Lys Asn Tyr Cys Arg Asn Pro Asp Arg Glu
130 135 140
Leu Arg Pro Trp Cys Phe Thr Thr Asp Pro Asn Lys Arg Trp Glu Leu
145 150 155 160
Cys Asp Ile Pro Arg Cys Thr Thr Pro Pro Pro Ser Ser Gly Pro Thr
165 170 175
Tyr Gln Cys Leu Lys Gly Thr Gly Glu Asn Tyr Arg Gly Asn Val Ala
180 185 190
Val Thr Val Ser Gly His Thr Cys Gln His Trp Ser Ala Gln Thr Pro
195 200 205
His Thr His Asn Arg Thr Pro Glu Asn Phe Pro Cys Lys Asn Leu Asp
210 215 220
Glu Asn Tyr Cys Arg Asn Pro Asp Gly Lys Arg Ala Pro Trp Cys His
225 230 235 240
Thr Thr Asn Ser Gln Val Arg Trp Glu Tyr Cys Lys Ile Pro Ser Cys
245 250 255
Asp Ser Ser Pro Val Ser Thr Glu Gln Leu Ala Pro Thr Ala Pro Pro
260 265 270
Glu Leu Thr Pro Val Val Gln Asp Cys Tyr His Gly Asp Gly Gln Ser
275 280 285
Tyr Arg Gly Thr Ser Ser Thr Thr Thr Thr Gly Lys Lys Cys Gln Ser
290 295 300
Trp Ser Ser Met Thr Pro His Arg His Gln Lys Thr Pro Glu Asn Tyr
305 310 315 320
Pro Asn Ala Gly Leu Thr Met Asn Tyr Cys Arg Asn Pro Asp Ala Asp
325 330 335
Lys Gly Pro Trp Cys Phe Thr Thr Asp Pro Ser Val Arg Trp Glu Tyr
340 345 350
Cys Asn Leu Lys Lys Cys Ser Gly Thr Glu Ala Ser Val Val Ala Pro
355 360 365
Pro Pro Val Val Leu Leu Pro Asp Val Glu Thr Pro Ser Glu Glu Asp
370 375 380
Cys Met Phe Gly Asn Gly Lys Gly Tyr Arg Gly Lys Arg Ala Thr Thr
385 390 395 400
Val Thr Gly Thr Pro Cys Gln Asp Trp Ala Ala Gln Glu Pro His Arg
405 410 415
His Ser Ile Phe Thr Pro Glu Thr Asn Pro Arg Ala Gly Leu Glu Lys
420 425 430
Asn Tyr Cys Arg Asn Pro Asp Gly Asp Val Gly Gly Pro Trp Cys Tyr
435 440 445
Thr Thr Asn Pro Arg Lys Leu Tyr Asp Tyr Cys Asp Val Pro Gln Cys
450 455 460
Ala Ala Pro Ser Phe Asp Cys Gly Lys Pro Gln Val Glu Pro Lys Lys
465 470 475 480
Cys Pro Gly Arg Val Val Gly Gly Cys Val Ala His Pro His Ser Trp
485 490 495
Pro Trp Gln Val Ser Leu Arg Thr Arg Phe Gly Met His Phe Cys Gly
500 505 510
Gly Thr Leu Ile Ser Pro Glu Trp Val Leu Thr Ala Ala His Cys Leu
515 520 525
Glu Lys Ser Pro Arg Pro Ser Ser Tyr Lys Val Ile Leu Gly Ala His
530 535 540
Gln Glu Val Asn Leu Glu Pro His Val Gln Glu Ile Glu Val Ser Arg
545 550 555 560
Leu Phe Leu Glu Pro Thr Arg Lys Asp Ile Ala Leu Leu Lys Leu Ser
565 570 575
Ser Pro Ala Val Ile Thr Asp Lys Val Ile Pro Ala Cys Leu Pro Ser
580 585 590
Pro Asn Tyr Val Val Ala Asp Arg Thr Glu Cys Phe Ile Thr Gly Trp
595 600 605
Gly Glu Thr Gln Gly Thr Phe Gly Ala Gly Leu Leu Lys Glu Ala Gln
610 615 620
Leu Pro Val Ile Glu Asn Lys Val Cys Asn Arg Tyr Glu Phe Leu Asn
625 630 635 640
Gly Arg Val Gln Ser Thr Glu Leu Cys Ala Gly His Leu Ala Gly Gly
645 650 655
Thr Asp Ser Cys Gln Gly Asp Ser Gly Gly Pro Leu Val Cys Phe Glu
660 665 670
Lys Asp Lys Tyr Ile Leu Gln Gly Val Thr Ser Trp Gly Leu Gly Cys
675 680 685
Ala Arg Pro Asn Lys Pro Gly Val Tyr Val Arg Val Ser Arg Phe Val
690 695 700
Thr Trp Ile Glu Gly Val Met Arg Asn Asn
705 710
<210> 7
<211> 1245
<212> DNA
<213> delta-plg(delta-纤溶酶原)核酸序列
<400> 7
gagcctctgg atgactatgt gaatacccag ggggcttcac tgttcagtgt cactaagaag 60
cagctgggag caggaagtat agaagaatgt gcagcaaaat gtgaggagga cgaagaattc 120
acctgcaggg cattccaata tcacagtaaa gagcaacaat gtgtgataat ggctgaaaac 180
aggaagtcct ccataatcat taggatgaga gatgtagttt tatttgaaaa gaaagtgtat 240
ctctcagagt gcaagactgg gaatggaaag aactacagag ggacgatgtc caaaacaaaa 300
aatggcatca cctgtcaaaa atggagttcc acttctcccc acagacctag attctcacct 360
gctacacacc cctcagaggg actggaggag aactactgca ggaatccaga caacgatccg 420
caggggccct ggtgctatac tactgatcca gaaaagagat atgactactg cgacattctt 480
gagtgtgaag aggcggcccc ttcatttgat tgtgggaagc ctcaagtgga gccgaagaaa 540
tgtcctggaa gggttgtagg ggggtgtgtg gcccacccac attcctggcc ctggcaagtc 600
agtcttagaa caaggtttgg aatgcacttc tgtggaggca ccttgatatc cccagagtgg 660
gtgttgactg ctgcccactg cttggagaag tccccaaggc cttcatccta caaggtcatc 720
ctgggtgcac accaagaagt gaatctcgaa ccgcatgttc aggaaataga agtgtctagg 780
ctgttcttgg agcccacacg aaaagatatt gccttgctaa agctaagcag tcctgccgtc 840
atcactgaca aagtaatccc agcttgtctg ccatccccaa attatgtggt cgctgaccgg 900
accgaatgtt tcatcactgg ctggggagaa acccaaggta cttttggagc tggccttctc 960
aaggaagccc agctccctgt gattgagaat aaagtgtgca atcgctatga gtttctgaat 1020
ggaagagtcc aatccaccga actctgtgct gggcatttgg ccggaggcac tgacagttgc 1080
cagggtgaca gtggaggtcc tctggtttgc ttcgagaagg acaaatacat tttacaagga 1140
gtcacttctt ggggtcttgg ctgtgcacgc cccaataagc ctggtgtcta tgttcgtgtt 1200
tcaaggtttg ttacttggat tgagggagtg atgagaaata attaa 1245
<210> 8
<211> 414
<212> PRT
<213> delta-plg(delta-纤溶酶原)氨基酸序列
<400> 8
Glu Pro Leu Asp Asp Tyr Val Asn Thr Gln Gly Ala Ser Leu Phe Ser
1 5 10 15
Val Thr Lys Lys Gln Leu Gly Ala Gly Ser Ile Glu Glu Cys Ala Ala
20 25 30
Lys Cys Glu Glu Asp Glu Glu Phe Thr Cys Arg Ala Phe Gln Tyr His
35 40 45
Ser Lys Glu Gln Gln Cys Val Ile Met Ala Glu Asn Arg Lys Ser Ser
50 55 60
Ile Ile Ile Arg Met Arg Asp Val Val Leu Phe Glu Lys Lys Val Tyr
65 70 75 80
Leu Ser Glu Cys Lys Thr Gly Asn Gly Lys Asn Tyr Arg Gly Thr Met
85 90 95
Ser Lys Thr Lys Asn Gly Ile Thr Cys Gln Lys Trp Ser Ser Thr Ser
100 105 110
Pro His Arg Pro Arg Phe Ser Pro Ala Thr His Pro Ser Glu Gly Leu
115 120 125
Glu Glu Asn Tyr Cys Arg Asn Pro Asp Asn Asp Pro Gln Gly Pro Trp
130 135 140
Cys Tyr Thr Thr Asp Pro Glu Lys Arg Tyr Asp Tyr Cys Asp Ile Leu
145 150 155 160
Glu Cys Glu Glu Ala Ala Pro Ser Phe Asp Cys Gly Lys Pro Gln Val
165 170 175
Glu Pro Lys Lys Cys Pro Gly Arg Val Val Gly Gly Cys Val Ala His
180 185 190
Pro His Ser Trp Pro Trp Gln Val Ser Leu Arg Thr Arg Phe Gly Met
195 200 205
His Phe Cys Gly Gly Thr Leu Ile Ser Pro Glu Trp Val Leu Thr Ala
210 215 220
Ala His Cys Leu Glu Lys Ser Pro Arg Pro Ser Ser Tyr Lys Val Ile
225 230 235 240
Leu Gly Ala His Gln Glu Val Asn Leu Glu Pro His Val Gln Glu Ile
245 250 255
Glu Val Ser Arg Leu Phe Leu Glu Pro Thr Arg Lys Asp Ile Ala Leu
260 265 270
Leu Lys Leu Ser Ser Pro Ala Val Ile Thr Asp Lys Val Ile Pro Ala
275 280 285
Cys Leu Pro Ser Pro Asn Tyr Val Val Ala Asp Arg Thr Glu Cys Phe
290 295 300
Ile Thr Gly Trp Gly Glu Thr Gln Gly Thr Phe Gly Ala Gly Leu Leu
305 310 315 320
Lys Glu Ala Gln Leu Pro Val Ile Glu Asn Lys Val Cys Asn Arg Tyr
325 330 335
Glu Phe Leu Asn Gly Arg Val Gln Ser Thr Glu Leu Cys Ala Gly His
340 345 350
Leu Ala Gly Gly Thr Asp Ser Cys Gln Gly Asp Ser Gly Gly Pro Leu
355 360 365
Val Cys Phe Glu Lys Asp Lys Tyr Ile Leu Gln Gly Val Thr Ser Trp
370 375 380
Gly Leu Gly Cys Ala Arg Pro Asn Lys Pro Gly Val Tyr Val Arg Val
385 390 395 400
Ser Arg Phe Val Thr Trp Ile Glu Gly Val Met Arg Asn Asn
405 410
<210> 9
<211> 1104
<212> DNA
<213> Mini-plg(小纤维蛋白溶酶原)核酸序列
<400> 9
gtcaggtggg agtactgcaa cctgaaaaaa tgctcaggaa cagaagcgag tgttgtagca 60
cctccgcctg ttgtcctgct tccagatgta gagactcctt ccgaagaaga ctgtatgttt 120
gggaatggga aaggataccg aggcaagagg gcgaccactg ttactgggac gccatgccag 180
gactgggctg cccaggagcc ccatagacac agcattttca ctccagagac aaatccacgg 240
gcgggtctgg aaaaaaatta ctgccgtaac cctgatggtg atgtaggtgg tccctggtgc 300
tacacgacaa atccaagaaa actttacgac tactgtgatg tccctcagtg tgcggcccct 360
tcatttgatt gtgggaagcc tcaagtggag ccgaagaaat gtcctggaag ggttgtaggg 420
gggtgtgtgg cccacccaca ttcctggccc tggcaagtca gtcttagaac aaggtttgga 480
atgcacttct gtggaggcac cttgatatcc ccagagtggg tgttgactgc tgcccactgc 540
ttggagaagt ccccaaggcc ttcatcctac aaggtcatcc tgggtgcaca ccaagaagtg 600
aatctcgaac cgcatgttca ggaaatagaa gtgtctaggc tgttcttgga gcccacacga 660
aaagatattg ccttgctaaa gctaagcagt cctgccgtca tcactgacaa agtaatccca 720
gcttgtctgc catccccaaa ttatgtggtc gctgaccgga ccgaatgttt catcactggc 780
tggggagaaa cccaaggtac ttttggagct ggccttctca aggaagccca gctccctgtg 840
attgagaata aagtgtgcaa tcgctatgag tttctgaatg gaagagtcca atccaccgaa 900
ctctgtgctg ggcatttggc cggaggcact gacagttgcc agggtgacag tggaggtcct 960
ctggtttgct tcgagaagga caaatacatt ttacaaggag tcacttcttg gggtcttggc 1020
tgtgcacgcc ccaataagcc tggtgtctat gttcgtgttt caaggtttgt tacttggatt 1080
gagggagtga tgagaaataa ttaa 1104
<210> 10
<211> 367
<212> PRT
<213> Mini-plg(小纤维蛋白溶酶原)氨基酸序列
<400> 10
Val Arg Trp Glu Tyr Cys Asn Leu Lys Lys Cys Ser Gly Thr Glu Ala
1 5 10 15
Ser Val Val Ala Pro Pro Pro Val Val Leu Leu Pro Asp Val Glu Thr
20 25 30
Pro Ser Glu Glu Asp Cys Met Phe Gly Asn Gly Lys Gly Tyr Arg Gly
35 40 45
Lys Arg Ala Thr Thr Val Thr Gly Thr Pro Cys Gln Asp Trp Ala Ala
50 55 60
Gln Glu Pro His Arg His Ser Ile Phe Thr Pro Glu Thr Asn Pro Arg
65 70 75 80
Ala Gly Leu Glu Lys Asn Tyr Cys Arg Asn Pro Asp Gly Asp Val Gly
85 90 95
Gly Pro Trp Cys Tyr Thr Thr Asn Pro Arg Lys Leu Tyr Asp Tyr Cys
100 105 110
Asp Val Pro Gln Cys Ala Ala Pro Ser Phe Asp Cys Gly Lys Pro Gln
115 120 125
Val Glu Pro Lys Lys Cys Pro Gly Arg Val Val Gly Gly Cys Val Ala
130 135 140
His Pro His Ser Trp Pro Trp Gln Val Ser Leu Arg Thr Arg Phe Gly
145 150 155 160
Met His Phe Cys Gly Gly Thr Leu Ile Ser Pro Glu Trp Val Leu Thr
165 170 175
Ala Ala His Cys Leu Glu Lys Ser Pro Arg Pro Ser Ser Tyr Lys Val
180 185 190
Ile Leu Gly Ala His Gln Glu Val Asn Leu Glu Pro His Val Gln Glu
195 200 205
Ile Glu Val Ser Arg Leu Phe Leu Glu Pro Thr Arg Lys Asp Ile Ala
210 215 220
Leu Leu Lys Leu Ser Ser Pro Ala Val Ile Thr Asp Lys Val Ile Pro
225 230 235 240
Ala Cys Leu Pro Ser Pro Asn Tyr Val Val Ala Asp Arg Thr Glu Cys
245 250 255
Phe Ile Thr Gly Trp Gly Glu Thr Gln Gly Thr Phe Gly Ala Gly Leu
260 265 270
Leu Lys Glu Ala Gln Leu Pro Val Ile Glu Asn Lys Val Cys Asn Arg
275 280 285
Tyr Glu Phe Leu Asn Gly Arg Val Gln Ser Thr Glu Leu Cys Ala Gly
290 295 300
His Leu Ala Gly Gly Thr Asp Ser Cys Gln Gly Asp Ser Gly Gly Pro
305 310 315 320
Leu Val Cys Phe Glu Lys Asp Lys Tyr Ile Leu Gln Gly Val Thr Ser
325 330 335
Trp Gly Leu Gly Cys Ala Arg Pro Asn Lys Pro Gly Val Tyr Val Arg
340 345 350
Val Ser Arg Phe Val Thr Trp Ile Glu Gly Val Met Arg Asn Asn
355 360 365
<210> 11
<211> 750
<212> DNA
<213> Micro-plg(微纤维蛋白溶酶原)核酸序列
<400> 11
gccccttcat ttgattgtgg gaagcctcaa gtggagccga agaaatgtcc tggaagggtt 60
gtaggggggt gtgtggccca cccacattcc tggccctggc aagtcagtct tagaacaagg 120
tttggaatgc acttctgtgg aggcaccttg atatccccag agtgggtgtt gactgctgcc 180
cactgcttgg agaagtcccc aaggccttca tcctacaagg tcatcctggg tgcacaccaa 240
gaagtgaatc tcgaaccgca tgttcaggaa atagaagtgt ctaggctgtt cttggagccc 300
acacgaaaag atattgcctt gctaaagcta agcagtcctg ccgtcatcac tgacaaagta 360
atcccagctt gtctgccatc cccaaattat gtggtcgctg accggaccga atgtttcatc 420
actggctggg gagaaaccca aggtactttt ggagctggcc ttctcaagga agcccagctc 480
cctgtgattg agaataaagt gtgcaatcgc tatgagtttc tgaatggaag agtccaatcc 540
accgaactct gtgctgggca tttggccgga ggcactgaca gttgccaggg tgacagtgga 600
ggtcctctgg tttgcttcga gaaggacaaa tacattttac aaggagtcac ttcttggggt 660
cttggctgtg cacgccccaa taagcctggt gtctatgttc gtgtttcaag gtttgttact 720
tggattgagg gagtgatgag aaataattaa 750
<210> 12
<211> 249
<212> PRT
<213> Micro-plg(微纤维蛋白溶酶原)氨基酸序列
<400> 12
Ala Pro Ser Phe Asp Cys Gly Lys Pro Gln Val Glu Pro Lys Lys Cys
1 5 10 15
Pro Gly Arg Val Val Gly Gly Cys Val Ala His Pro His Ser Trp Pro
20 25 30
Trp Gln Val Ser Leu Arg Thr Arg Phe Gly Met His Phe Cys Gly Gly
35 40 45
Thr Leu Ile Ser Pro Glu Trp Val Leu Thr Ala Ala His Cys Leu Glu
50 55 60
Lys Ser Pro Arg Pro Ser Ser Tyr Lys Val Ile Leu Gly Ala His Gln
65 70 75 80
Glu Val Asn Leu Glu Pro His Val Gln Glu Ile Glu Val Ser Arg Leu
85 90 95
Phe Leu Glu Pro Thr Arg Lys Asp Ile Ala Leu Leu Lys Leu Ser Ser
100 105 110
Pro Ala Val Ile Thr Asp Lys Val Ile Pro Ala Cys Leu Pro Ser Pro
115 120 125
Asn Tyr Val Val Ala Asp Arg Thr Glu Cys Phe Ile Thr Gly Trp Gly
130 135 140
Glu Thr Gln Gly Thr Phe Gly Ala Gly Leu Leu Lys Glu Ala Gln Leu
145 150 155 160
Pro Val Ile Glu Asn Lys Val Cys Asn Arg Tyr Glu Phe Leu Asn Gly
165 170 175
Arg Val Gln Ser Thr Glu Leu Cys Ala Gly His Leu Ala Gly Gly Thr
180 185 190
Asp Ser Cys Gln Gly Asp Ser Gly Gly Pro Leu Val Cys Phe Glu Lys
195 200 205
Asp Lys Tyr Ile Leu Gln Gly Val Thr Ser Trp Gly Leu Gly Cys Ala
210 215 220
Arg Pro Asn Lys Pro Gly Val Tyr Val Arg Val Ser Arg Phe Val Thr
225 230 235 240
Trp Ile Glu Gly Val Met Arg Asn Asn
245
<210> 13
<211> 684
<212> DNA
<213> 丝氨酸蛋白酶(结构)域的核酸序列
<400> 13
gttgtagggg ggtgtgtggc ccacccacat tcctggccct ggcaagtcag tcttagaaca 60
aggtttggaa tgcacttctg tggaggcacc ttgatatccc cagagtgggt gttgactgct 120
gcccactgct tggagaagtc cccaaggcct tcatcctaca aggtcatcct gggtgcacac 180
caagaagtga atctcgaacc gcatgttcag gaaatagaag tgtctaggct gttcttggag 240
cccacacgaa aagatattgc cttgctaaag ctaagcagtc ctgccgtcat cactgacaaa 300
gtaatcccag cttgtctgcc atccccaaat tatgtggtcg ctgaccggac cgaatgtttc 360
atcactggct ggggagaaac ccaaggtact tttggagctg gccttctcaa ggaagcccag 420
ctccctgtga ttgagaataa agtgtgcaat cgctatgagt ttctgaatgg aagagtccaa 480
tccaccgaac tctgtgctgg gcatttggcc ggaggcactg acagttgcca gggtgacagt 540
ggaggtcctc tggtttgctt cgagaaggac aaatacattt tacaaggagt cacttcttgg 600
ggtcttggct gtgcacgccc caataagcct ggtgtctatg ttcgtgtttc aaggtttgtt 660
acttggattg agggagtgat gaga 684
<210> 14
<211> 228
<212> PRT
<213> 丝氨酸蛋白酶(结构)域的氨基酸序列
<400> 14
Val Val Gly Gly Cys Val Ala His Pro His Ser Trp Pro Trp Gln Val
1 5 10 15
Ser Leu Arg Thr Arg Phe Gly Met His Phe Cys Gly Gly Thr Leu Ile
20 25 30
Ser Pro Glu Trp Val Leu Thr Ala Ala His Cys Leu Glu Lys Ser Pro
35 40 45
Arg Pro Ser Ser Tyr Lys Val Ile Leu Gly Ala His Gln Glu Val Asn
50 55 60
Leu Glu Pro His Val Gln Glu Ile Glu Val Ser Arg Leu Phe Leu Glu
65 70 75 80
Pro Thr Arg Lys Asp Ile Ala Leu Leu Lys Leu Ser Ser Pro Ala Val
85 90 95
Ile Thr Asp Lys Val Ile Pro Ala Cys Leu Pro Ser Pro Asn Tyr Val
100 105 110
Val Ala Asp Arg Thr Glu Cys Phe Ile Thr Gly Trp Gly Glu Thr Gln
115 120 125
Gly Thr Phe Gly Ala Gly Leu Leu Lys Glu Ala Gln Leu Pro Val Ile
130 135 140
Glu Asn Lys Val Cys Asn Arg Tyr Glu Phe Leu Asn Gly Arg Val Gln
145 150 155 160
Ser Thr Glu Leu Cys Ala Gly His Leu Ala Gly Gly Thr Asp Ser Cys
165 170 175
Gln Gly Asp Ser Gly Gly Pro Leu Val Cys Phe Glu Lys Asp Lys Tyr
180 185 190
Ile Leu Gln Gly Val Thr Ser Trp Gly Leu Gly Cys Ala Arg Pro Asn
195 200 205
Lys Pro Gly Val Tyr Val Arg Val Ser Arg Phe Val Thr Trp Ile Glu
210 215 220
Gly Val Met Arg
225
- 还没有人留言评论。精彩留言会获得点赞!