从乙酰‑COA产生烯烃的重组微生物的制作方法
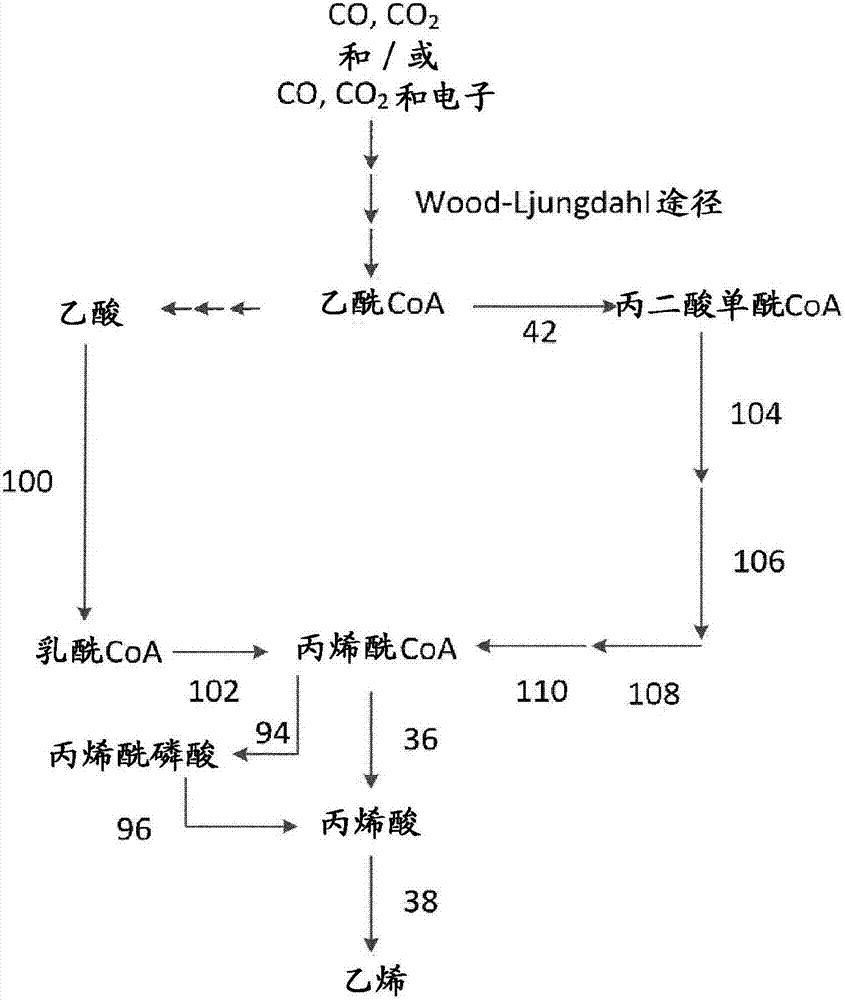
相关申请的交叉参考本申请要求2014年9月3日提交uspto并正式分配系列号us62/045,083的申请“遗传工程微生物和将气态c1-碳源和/或气态c1-碳源和电子转化为烯烃的方法(geneticallyengineeredmicroorganismandprocessforconvertinggaseousc1-carbonsourcesand/orgaseousc1-carbonsourcesandelectronsintoalkenes)”的权益和优先权。本申请还要求2014年9月9日提交uspto并正式分配系列号us62/047,827的申请“遗传工程微生物和将气态c1-碳源和/或气态c1-碳源和电子转化为1,3-丁二烯的方法(geneticallyengineeredmicroorganismandprocessforconvertinggaseousc1-carbonsourcesand/orgaseousc1-carbonsourcesandelectronsinto1,3-butadiene)”的权益和优先权。本申请还要求2014年9月26日提交欧洲专利局并正式分配系列号ep14186574的申请“遗传工程微生物和将气态c1-碳源和/或气态c1-碳源和电子转化为2-甲基-1,3-丁二烯和1,3-丁二烯的方法(geneticallyengineeredmicroorganismandprocessforconvertinggaseousc1-carbonsourcesand/orgaseousc1-carbonsourcesandelectronsinto2-methyl-1,3-butadieneand1,3-butadiene)”的权益和优先权。本申请还要求2014年9月26日提交欧洲专利局并正式分配系列号ep14186690的申请“遗传工程微生物和将气态c1-碳源和/或气态c1-碳源和电子转化为烯烃的方法(geneticallyengineeredmicroorganismandprocessforconvertinggaseousc1-carbonsourcesand/orgaseousc1-carbonsourcesandelectronsintoalkenes)”的权益和优先权。这些申请us62/045,083、us62/047,827、ep14186574和ep14186690的内容在此为了所有目的以其整体引入作为参考,包括所有表格、附图和权利要求,以及依照pct细则4.18包括并入不包含在本文中和在pct细则20.5(a)中提到的说明书、权利要求或附图的任何要素或部分。序列表本申请包括序列表。提供重组微生物。还提供其中使用该重组微生物的方法。该方法是使c1碳源转化为主链具有1至6个碳原子的一种或多种烷烃和/或烯烃的方法。
背景技术:
:提供以下背景讨论仅为了辅助读者理解下文中公开的微生物、方法和用途,不承认描述或构成现有技术。全球经济对能量和大宗化学品的需求增加对烃成本施加了越来越大的压力。许多产业(包括化学和塑料产业)重度依赖可作为其产品原料的化石烃源。烷烃是汽油、柴油和航空煤油的主要组分。烯烃通常通过裂化见于原油中的烷烃来产生。化石燃料的快速耗竭成为鉴定用于产生烷烃的替代来源的驱动力。多种物种天然产生长链烷烃。但是,对于链长小于约10个原子的烷烃和烯烃,仅已知甲烷的天然生物合成途径。这类短链烷烃和烯烃不仅用作燃料,在产生大宗化学品中也很重要。它们例如用作塑料、聚酯、溶剂、燃料、粘合剂或油漆的构件。例如丙烯是端烯烃,其用于制备聚乙烯、聚丙烯、α烯烃、苯乙烯、聚酯、丙烯酸、乙二醇防冻液、聚氯乙烯(pvc)、环氧丙烷、酮醇和异丙醇。丙烯通常衍生自从获自裂化和其他提炼方法的烃混合物分馏。乙烯也作为原材料,例如作为用于产生聚氨基甲酸酯、橡胶制品、实验化学品、工业流体和防冻化合物的单体。它还用于产生燃料和燃料添加剂。1,3-丁二烯是无色气体,其在-4.5℃冷凝为液体。丁二烯是用萃取蒸馏法从乙烯和丙烯产生的裂化副产物之一粗c4流衍生。丁二烯的最大单一用途是用于产生丁苯橡胶(sbr),sbr转而主要用于制造汽车轮胎。sbr还用于粘合剂、密封剂、涂料,及用于橡胶制品如鞋底。聚丁二烯也用于轮胎,且可以在丙烯腈-丁二烯-苯乙烯(abs)的产生中用作中间体。abs广泛用于诸如电话、电脑外罩和其他器具。由丁二烯制成的其他聚合物包括:用于例如地毯背衬和粘合剂的丁苯胶乳;用于软管、燃料管、密封垫片、手套和鞋的丁腈橡胶;用于从沥青改性剂(路面和屋顶建造应用)至粘合剂、鞋和玩具的许多终端用途的苯乙烯-丁二烯嵌段共聚物。由丁二烯制成的化学中间体包括分别用于制造尼龙和氯丁橡胶的己二腈和氯丁二烯。目前每年生产约一千万吨丁二烯。全世界1,3-丁二烯的销售额超过100亿美元。在寻找更可持续的化学生产方法中,从替代和/或可再生原料制造丁二烯的能力是主要进步。2-甲基-1,3-丁二烯或异戊二烯是无色挥发性液体,其由许多植物、动物和细菌天然产生。2-甲基-1,3-丁二烯作为用于合成多种合成聚合物尤其是合成高弹体的原材料。目前可得且为产生合成高弹体所需的大多数2-甲基-1,3-丁二烯是通过热裂化石油或油产生的。高弹体或橡胶不仅在轮胎生产中需要,在橡胶产业中也需要,它们在橡胶产业中用于多种消费品,如医用手套、配件、橡皮筋、鞋、体育用品等。合成橡胶主要基于丁二烯聚合物,丁二烯聚合物作为副产物从乙烯和丙烯生产获得。虽然2-甲基-1,3-丁二烯可以通过分馏石油获得,但石油的纯化昂贵且耗时,与所需付出的努力相比,所获得的2-甲基-1,3-丁二烯产率低。此外,2-甲基-1,3-丁二烯的供给有风险,因为天然橡胶的可用性吃紧,且其价格上涨。另外,由于更深刻的生态意识,对来自烯烃裂化的粗c5原料流的接受度下降。因此,目前,尝试了用于产生2-甲基-1,3-丁二烯的更经济的方法,尤其是基于生物的方法。虽然2-甲基-1,3-丁二烯的生物来源(即某些植物)可这样获得,但迄今未开发出经济上和实际上可行的从植物产生和回收2-甲基-1,3-丁二烯的商业化方法。已进行了从用细菌合成来产生异戊二烯的尝试(美国专利申请us2014/0234926、us2013/0323820,国际专利申请wo2014/065271、wo2013/180584、wo2013/181647),就产率、纯度和稳定性而言,所使用的菌株并不符合工业生产的要求。使合成气转化为液体燃料或化学品的可用技术包括化学催化法,如fischer-tropsch法,以及用于合成甲醇或其他混合醇的方法和生物气体发酵法。fischer-tropsch法从将近100年前就在使用,依赖基于金属的无机催化剂来使合成气转化为更长链的烃。通过天然生物化学代谢过程固定碳的生物系统是已知的。已开发了藻类系统来通过光合反应以及间接依赖光合作用的饲喂糖的异养反应产生烃。已遗传改造细菌细胞来在异养发酵系统中将糖原料加工为有用的烃。已公开了直接或间接产生例如烷烃的生物途径(us2011/0269204、us2011/0091952、us2011/0269204、wo2012/058606、us2012/0329119、us2013/0224808、us2013/0316425、wo2013/148348、us2014/0039143、wo2014/047209)。但是,这些公开的方法具有使它们尤其对大规模工业方法而言无效或不适宜的局限。几篇出版物(lee,c.c.等,science(2010)329,5992,642;yang,z.-y等,journalofbiologicalchemistry(2011)286,22,19417-19421)显示,纯化的钒以及钼固氮酶能够在体外催化co或co2或cn转化形成烃。发明概述提供重组微生物。该重组微生物通常能够从c1碳化合物尤其是一氧化碳和/或二氧化碳产生主链具有2至5个碳原子的一种或多种烯烃。该重组微生物还能够从诸如一氧化碳、二氧化碳和/或氰化物的c1碳产生主链具有1至6个碳原子的一种或多种烷烃和/或烯烃。在一些实施方案中,该重组微生物可以能够从一氧化碳或二氧化碳产生主链具有2或3个碳原子的一种或多种烷烃和/或烯烃。在一些实施方案中,该重组微生物可以能够从一氧化碳或二氧化碳产生主链具有4或5个碳原子的一种或多种烷烃和/或烯烃。在一些实施方案中,该重组微生物不能进行光合作用。在一些实施方案中,该重组微生物具有允许该重组微生物从co或co2和/或氰化物进行乙酰coa的生物合成的内源酶。本文所述的细胞包含用于生物合成烷烃和/或烯烃的人工的和有效的途径。在某些实施方案中,各生物合成基因稳定整合入宿主染色体。还提供可以借助其来以足以满足稳健的商业化方法的需求的高产率和纯度产生2种烷烃和/或烯烃的方法。这些方法非常清洁和有效(就节约成本和时间而言),是工业上可行的产生目的烷烃和/或烯烃的方式。根据第一方面,提供重组微生物。该重组微生物包含异源核酸序列,该异源核酸序列编码催化乙酰coa转化为主链具有1至5个碳原子的一种或多种烷烃和/或烯烃的一种或多种酶。该异源核酸序列包括编码催化乙酰coa转化为主链具有1至5个碳原子的一种或多种烷烃和/或烯烃的一种或多种酶的编码序列或编码序列的组合。各编码序列与转录启动子有效连接。在一些实施方案中,该重组微生物包含编码使乙酰coa转化为主链具有2至5个碳原子的一种或多种烯烃的一种或多种酶的异源核酸序列。在一些实施方案中,该重组微生物还包含使c1碳源转化为乙酰coa的一种或多种内源酶。在一些实施方案中,该c1碳源是一氧化碳、二氧化碳和氰化物中的至少一种。在一些实施方案中,该重组微生物不包含编码催化乙酰coa转化为主链具有1至8个碳原子的一种或多种烷烃和/或烯烃的一种或多种酶的至少一种编码序列或编码序列的组合。在该重组微生物包含编码催化乙酰coa转化为主链具有1至8个碳原子的一种或多种烷烃和/或烯烃的一种或多种酶的一种以上编码序列或一种以上编码序列的组合的实施方案中,该重组微生物可以不包含该编码序列中的至少一种。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码固氮酶ec1.18.6.1或ec1.19.6.1的编码序列。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码催化乙酰coa转化为巴豆酰coa和进一步催化巴豆酰coa转化为烯烃的一种或多种酶的一种或多种编码序列。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码催化乙酰coa转化为3-甲基巴豆酰coa和进一步催化3-甲基巴豆酰coa转化为烯烃的一种或多种酶的一种或多种编码序列。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码催化乙酰coa转化为丙酰coa和进一步催化丙酰coa转化为烯烃的一种或多种酶的一种或多种编码序列。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码催化乙酰coa转化为丙烯酰coa和进一步催化丙烯酰coa转化为烯烃的一种或多种酶的一种或多种编码序列。根据第一方面的重组微生物的一些实施方案,该催化乙酰coa转化为巴豆酰coa的一种或多种酶包括:乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);乙酰乙酰coa还原酶(ec1.1.1.36);和烯酰coa水合酶2(ec4.2.1.119)。根据第一方面的重组微生物的一些实施方案,该催化乙酰coa转化为巴豆酰coa的一种或多种酶包括:乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);和3-羟基丁酰coa脱水酶。根据第一方面的重组微生物的一些实施方案,该催化乙酰coa转化为巴豆酰coa的一种或多种酶包括:4-氨基丁酸氨基转移酶(ec2.6.1.19);谷氨酸脱氢酶(ec1.4.1.2);2-羟基戊二酸脱氢酶(ec1.1.99.2);戊烯二酸coa转移酶(ec2.8.3.12);2-羟基戊二酰coa脱水酶(ec4.2.1.-);和戊烯二酸单酰coa脱羧酶(ec4.1.1.70)。根据第一方面的重组微生物的一些实施方案,该催化乙酰coa转化为巴豆酰coa的一种或多种酶包括:4-氨基丁酸氨基转移酶(ec2.6.1.19);4-羟基丁酸脱氢酶(ec1.1.1.61);4-羟基丁酸coa转移酶(ec2.8.3.-);乙烯乙酰coaδ异构酶(ec4.2.1.120);和4-羟基丁酰coa脱水酶(ec5.3.3.3)。根据第一方面的重组微生物的一些实施方案,该催化乙酰coa转化为3-甲基巴豆酰coa的一种或多种酶包括:乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10);和3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18)。该催化乙酰coa转化为3-甲基巴豆酰coa的一种或多种酶也可以包括3-甲基巴豆酰coa羧化酶(ec6.4.1.4)。在一些实施方案中,该催化乙酰coa转化为3-甲基巴豆酰coa的一种或多种酶可以包括:乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10);3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18);和3-甲基巴豆酰coa羧化酶(ec6.4.1.4)。在一些实施方案中,该催化乙酰coa转化为3-甲基巴豆酰coa的一种或多种酶包括:乙酰coac-乙酰转移酶(ec2.3.1.9);乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);3-羟基-3-甲基戊二酰-coa合酶(ec2.3.3.10);3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18);和3-甲基巴豆酰coa羧化酶(ec6.4.1.4)。根据第一方面的重组微生物的一些实施方案,该催化乙酰coa转化为丙酰coa的一种或多种酶包括:乙酰coa羧化酶(ec6.4.1.2);丙二酸单酰(malonyl)coa还原酶(ec1.2.1.75);丙二酰(malonyl)coa还原酶(ec1.1.1.298);丙酰coa合酶(ec6.2.1.36);丙酰coa合酶/丙烯酰coa合酶(ec4.2.1.116);和丙酰coa合酶(ec1.3.1.84)。根据第一方面的重组微生物的一些实施方案,该催化乙酰coa转化为丙酰coa的一种或多种酶包括:乙酰coa羧化酶(ec6.4.1.2)和甲基丙二酸单酰coa脱羧酶(ec4.1.1.41)。该催化乙酰coa转化为丙酰coa的一种或多种酶可以例如包括:乙酰coa羧化酶(ec6.4.1.2);甲基丙二酸单酰coa变位酶(ec5.4.99.2);和甲基丙二酸单酰coa脱羧酶(ec4.1.1.41)。根据第一方面的重组微生物的一些实施方案,该催化乙酰coa转化为丙酰coa的一种或多种酶包括:2-氧代戊二酸/丁酸铁氧还蛋白氧化还原酶(ec1.2.7.3)和/或丙酮酸/2-氧代丁酸铁氧还蛋白氧化还原酶(ec1.2.7.1)。根据第一方面的重组微生物的一些实施方案,该催化乙酰coa转化为丙酰coa的一种或多种酶包括:乳酸coa转移酶(ec2.8.3.-);乳酰coa脱水酶(ec4.2.1.54);和丙酰coa合酶(ec1.3.1.84)。根据第一方面的重组微生物的一些实施方案,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括:乙酰coa羧化酶(ec6.4.1.2);丙二酸单酰coa还原酶(ec1.2.1.75);丙二酰coa还原酶(ec1.1.1.298);丙酰coa合酶(ec6.2.1.36);和丙酰coa合酶/丙烯酰coa合酶(ec4.2.1.116)。在一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括乳酸coa转移酶(ec2.8.3.-);和乳酰coa脱水酶(ec4.2.1.54)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括酰基coa硫酯酶(ec3.1.2.-)和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:反式-2-烯酰coa还原酶(ec1.1.1.36);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:反式-2-烯酰coa还原酶(ec1.1.1.36);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:醛脱氢酶(ec1.2.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);和牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:醛脱氢酶(ec1.2.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)和/或十一异戊烯醇(undecaprenol)激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:醛铁氧还蛋白氧化还原酶(ec1.2.7.5);酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:醛铁氧还蛋白氧化还原酶(ec1.2.7.5);酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);醛铁氧还蛋白氧化还原酶(ec1.2.7.5);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);醛铁氧还蛋白氧化还原酶(ec1.2.7.5);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:苯基丙烯酸脱羧酶(ec4.1.1.-);和酰基coa硫酯酶(ec3.1.2.-)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:苯基丙烯酸脱羧酶(ec4.1.1.-);磷酸丁酰转移酶(ec2.3.1.19);和丁酸激酶(ec2.7.2.7)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:苯基丙烯酸脱羧酶(ec4.1.1.-);酰基coa硫酯酶(ec3.1.2.-);磷酸丁酰转移酶(ec2.3.1.19);和丁酸激酶(ec2.7.2.7)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:醛铁氧还蛋白氧化还原酶(ec1.2.7.5);酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:醛铁氧还蛋白氧化还原酶(ec1.2.7.5);酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);醛铁氧还蛋白氧化还原酶(ec1.2.7.5);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化巴豆酰coa转化为烯烃的一种或多种酶包括:磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);醛铁氧还蛋白氧化还原酶(ec1.2.7.5);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。在一些实施方案中,该催化丙酰coa转化为烯烃的一种或多种酶包括:乙酰coac-乙酰转移酶(ec2.3.1.9);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该催化丙酰coa转化为烯烃的一种或多种酶包括:乙酰coac-乙酰转移酶(ec2.3.1.9);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该催化丙酰coa转化为烯烃的一种或多种酶包括:乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该催化丙酰coa转化为烯烃的一种或多种酶包括:乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该催化丙烯酰coa转化为烯烃的一种或多种酶包括:酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该催化丙烯酰coa转化为烯烃的一种或多种酶包括:磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);乳酸coa转移酶(ec2.8.3.-);乳酰coa脱水酶(ec4.2.1.54);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:4-氨基丁酸氨基转移酶(ec2.6.1.19);谷氨酸脱氢酶(ec1.4.1.2);2-羟基戊二酸脱氢酶(ec1.1.99.2);戊烯二酸coa转移酶(ec2.8.3.12);2-羟基戊二酰coa脱水酶(ec4.2.1.-);戊烯二酸单酰coa脱羧酶(ec4.1.1.70);酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:4-氨基丁酸氨基转移酶(ec2.6.1.19);4-羟基丁酸脱氢酶(ec1.1.1.61);4-羟基丁酸coa转移酶(ec2.8.3.-);乙烯乙酰coaδ异构酶(ec4.2.1.120);4-羟基丁酰coa脱水酶(ec5.3.3.3);酰基coa硫酯酶(ec3.1.2.-);醛/醇脱氢酶(ec1.1.1.-和/或ec1.2.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:4-氨基丁酸氨基转移酶(ec2.6.1.19);谷氨酸脱氢酶(ec1.4.1.2);2-羟基戊二酸脱氢酶(ec1.1.99.2);戊烯二酸coa转移酶(ec2.8.3.12);2-羟基戊二酰coa脱水酶(ec4.2.1.-);戊烯二酸单酰coa脱羧酶(ec4.1.1.70);4-羟基丁酸脱氢酶(ec1.1.1.61);4-羟基丁酸coa转移酶(ec2.8.3.-);乙烯乙酰coaδ异构酶(ec4.2.1.120);4-羟基丁酰coa脱水酶(ec5.3.3.3);酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:4-氨基丁酸氨基转移酶(ec2.6.1.19);4-羟基丁酸脱氢酶(ec1.1.1.61);4-羟基丁酸coa转移酶(ec2.8.3.-);乙烯乙酰coaδ异构酶(ec4.2.1.120);戊烯二酸单酰coa脱羧酶(ec4.1.1.70);4-羟基丁酰coa脱水酶(ec5.3.3.3);反式-2-烯酰coa还原酶(ec1.1.1.36);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coac-乙酰转移酶(ec2.3.1.9)、3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10)、3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18)、3-甲基巴豆酰coa羧化酶(ec6.4.1.4)、醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coac-乙酰转移酶(ec2.3.1.9)、3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10)、3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18)、3-甲基巴豆酰coa羧化酶(ec6.4.1.4)、醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、和/或牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、和/或羟乙基噻唑激酶(ec2.7.1.50)、和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2)、乙酰乙酰coa合酶(ec2.3.1.194)、3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10)、3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18)、3-甲基巴豆酰coa羧化酶(ec6.4.1.4)、醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2)、乙酰乙酰coa合酶(ec2.3.1.194)、3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10)、3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18)、3-甲基巴豆酰coa羧化酶(ec6.4.1.4)、醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、和/或牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、和/或羟乙基噻唑激酶(ec2.7.1.50)、和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coac-乙酰转移酶(ec2.3.1.9)、3-羟基丁酰coa脱氢酶(ec1.1.1.157)、3-羟基丁酰coa脱水酶(ec4.2.1.55)、醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coac-乙酰转移酶(ec2.3.1.9)、3-羟基丁酰coa脱氢酶(ec1.1.1.157)、3-羟基丁酰coa脱水酶(ec4.2.1.55)、醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、和/或牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、和/或羟乙基噻唑激酶(ec2.7.1.50)、和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2)、乙酰乙酰coa合酶(ec2.3.1.194)、乙酰乙酰coa还原酶(ec1.1.1.36)、烯酰coa水合酶2(ec4.2.1.119)、醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2)、乙酰乙酰coa合酶(ec2.3.1.194)、乙酰乙酰coa还原酶(ec1.1.1.36)、烯酰coa水合酶2(ec4.2.1.119)、醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);法尼醇激酶(ec2.7.1.-)、和/或牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、和/或羟乙基噻唑激酶(ec2.7.1.50)、和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10);3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18);3-甲基戊烯二酸单酰coa脱羧酶;醛/醇脱氢酶(ec1.1.1.-或ec1.2.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10);3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18);3-甲基戊烯二酸单酰coa脱羧酶;醛/醇脱氢酶(ec1.1.1.-或ec1.2.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10);3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18);3-甲基戊烯二酸单酰coa脱羧酶;法尼醇激酶(ec2.7.1.-)、和/或牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、和/或羟乙基噻唑激酶(ec2.7.1.50)、和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10);3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18);3-甲基戊烯二酸单酰coa脱羧酶;法尼醇激酶(ec2.7.1.-)、和/或牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、和/或羟乙基噻唑激酶(ec2.7.1.50)、和/或十一异戊烯醇激酶(ec2.7.1.66);及2-甲基-1,3-丁二烯合酶(ec4.2.3.27)和/或甲基丁烯醇合酶。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);乙酰乙酰coa还原酶(ec1.1.1.36);酰基coa水合酶2(ec4.2.1.119);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10);3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18);3-甲基戊烯二酸单酰coa脱羧酶;酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10);3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18);3-甲基戊烯二酸单酰coa脱羧酶;酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);反式-2-烯酰coa还原酶(ec1.1.1.36);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);反式-2-烯酰coa还原酶(ec1.1.1.36);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);反式-2-烯酰coa还原酶(ec1.1.1.36);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);反式-2-烯酰coa还原酶(ec1.1.1.36);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:4-氨基丁酸氨基转移酶(ec2.6.1.19);4-羟基丁酸脱氢酶(ec1.1.1.61);4-羟基丁酸coa转移酶(ec2.8.3.-);乙烯乙酰coaδ异构酶(ec4.2.1.120);4-羟基丁酰coa脱水酶(ec5.3.3.3);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:4-氨基丁酸氨基转移酶(ec2.6.1.19);谷氨酸脱氢酶(ec1.4.1.2);2-羟基戊二酸脱氢酶(ec1.1.99.2);戊烯二酸coa转移酶(ec2.8.3.12);2-羟基戊二酰coa脱水酶(ec4.2.1.-);戊烯二酸单酰coa脱羧酶(ec4.1.1.70);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:2-氧代戊二酸/丁酸铁氧还蛋白氧化还原酶(ec1.2.7.3);丙酮酸/2-氧代丁酸铁氧还蛋白氧化还原酶(ec1.2.7.1);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:2-氧代戊二酸/丁酸铁氧还蛋白氧化还原酶(ec1.2.7.3);丙酮酸/2-氧代丁酸铁氧还蛋白氧化还原酶(ec1.2.7.1);乙酰coac-乙酰转移酶(ec2.3.1.9);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:2-氧代戊二酸/丁酸铁氧还蛋白氧化还原酶(ec1.2.7.3);丙酮酸/2-氧代丁酸铁氧还蛋白氧化还原酶(ec1.2.7.1);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:2-氧代戊二酸/丁酸铁氧还蛋白氧化还原酶(ec1.2.7.3);丙酮酸/2-氧代丁酸铁氧还蛋白氧化还原酶(ec1.2.7.1);乙酰coac-乙酰转移酶(ec2.3.1.9);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:2-氧代戊二酸/丁酸铁氧还蛋白氧化还原酶(ec1.2.7.3);丙酮酸/2-氧代丁酸铁氧还蛋白氧化还原酶(ec1.2.7.1);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:2-氧代戊二酸/丁酸铁氧还蛋白氧化还原酶(ec1.2.7.3);丙酮酸/2-氧代丁酸铁氧还蛋白氧化还原酶(ec1.2.7.1);乙酰coac-乙酰转移酶(ec2.3.1.9);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乳酸coa转移酶(ec2.8.3.-);乳酰coa脱水酶(ec4.2.1.54);lcdab;丙酰coa合酶(ec1.3.1.84);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乳酸coa转移酶(ec2.8.3.-);乳酰coa脱水酶(ec4.2.1.54);lcdab;丙酰coa合酶(ec1.3.1.84);乙酰coac-乙酰转移酶(ec2.3.1.9);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乳酸coa转移酶(ec2.8.3.-);乳酰coa脱水酶(ec4.2.1.54);lcdab;丙酰coa合酶(ec1.3.1.84);乙酰coac-乙酰转移酶(ec2.3.1.9);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乳酸coa转移酶(ec2.8.3.-);乳酰coa脱水酶(ec4.2.1.54);lcdab;丙酰coa合酶(ec1.3.1.84);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乳酸coa转移酶(ec2.8.3.-);乳酰coa脱水酶(ec4.2.1.54);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);丙二酸单酰coa还原酶(ec1.2.1.75);丙二酰coa还原酶(ec1.1.1.298);丙酰coa合酶(ec6.2.1.36);丙酰coa合酶/丙烯酰coa合酶(ec4.2.1.116);丙酰coa合酶(ec1.3.1.84);乙酰coac-乙酰转移酶(ec2.3.1.9);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);丙二酸单酰coa还原酶(ec1.2.1.75);丙二酰coa还原酶(ec1.1.1.298);丙酰coa合酶(ec6.2.1.36);丙酰coa合酶/丙烯酰coa合酶(ec4.2.1.116);丙酰coa合酶(ec1.3.1.84);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);丙二酸单酰coa还原酶(ec1.2.1.75);丙二酰coa还原酶(ec1.1.1.298);丙酰coa合酶(ec6.2.1.36);丙酰coa合酶/丙烯酰coa合酶(ec4.2.1.116);丙酰coa合酶(ec1.3.1.84);乙酰coac-乙酰转移酶(ec2.3.1.9);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);丙二酸单酰coa还原酶(ec1.2.1.75);丙二酰coa还原酶(ec1.1.1.298);丙酰coa合酶(ec6.2.1.36);丙酰coa合酶/丙烯酰coa合酶(ec4.2.1.116);丙酰coa合酶(ec1.3.1.84);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);丙二酸单酰coa还原酶(形成丙二酸半醛)(ec1.2.1.75);3-羟基丙酸脱氢酶(ec1.1.1.298);3-羟基丙酰coa合酶(ec6.2.1.36);羟基丙酰coa脱水酶(ec4.2.1.116);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第一方面的重组微生物的一些实施方案,该异源核酸序列包含编码以下酶的编码序列:乙酰乙酰辅酶a合酶硫解酶(ec2.3.1.9);磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10);3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18);3-甲基戊烯二酸单酰coa脱羧酶;酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛铁氧还蛋白氧化还原酶(ec1.2.7.5);牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127);法尼醇激酶(ec2.7.1.-);和甲基丁烯醇合酶。在一些实施方案中,本文公开的微生物是厌氧微生物。该微生物可以是厌氧细胞。根据第一方面的重组微生物的一些实施方案,使一氧化碳和二氧化碳之一或二者转化为乙酰coa的内源酶是wood-ljungdahl途径的酶。在一些实施方案中,该重组微生物包含编码wood-ljungdahlpathway途径的所有酶的内源酶的序列。该重组微生物可以表达催化一氧化碳和二氧化碳之一或二者转化为乙酰coa的wood-ljungdahl途径的内源酶。在一些实施方案中,该重组微生物包含编码wood-ljungdahl途径的所有酶的内源酶的序列。该重组微生物可以表达催化一氧化碳和二氧化碳之一或二者转化为乙酰coa的wood-ljungdahl途径酶的内源酶。在一些实施方案中,该重组微生物包含编码wood-ljungdahl途径的所有酶的内源酶的序列。该重组微生物可以表达wood-ljungdahl途径的所有酶。在一些实施方案中,该重组微生物为羧基营养(caroxyotrop)。在一些实施方案中,该重组微生物是产乙酸菌。根据第一方面的重组微生物的一些实施方案,所有编码序列都与相同的常见转录启动子有效连接。根据一些实施方案,该异源基因序列包含在异源核酸分子中或该微生物的染色体中。根据一些实施方案,该微生物属于梭菌属(clostridium)。在一些实施方案中,该微生物可以是杨氏梭菌(clostridiumljungdahlii)。该微生物也可以是clostridiumautoethanogenum。在一些实施方案中,该微生物也可以是clostridiumcarboxidivorans。在一些实施方案中,该微生物也可以是醋酸梭菌(clostridiumaceticum)。在一些实施方案中,该微生物也可以是clostridiumdrakei。在一些实施方案中,该微生物也可以是粪味梭菌(clostridiumscatologenes)。在一些实施方案中,该微生物也可以是clostridiumragsdalei。在一些实施方案中,该微生物也可以是蚁酸醋酸梭菌(clostridiumformicoaceticum)。在一些实施方案中,该微生物也可以是大梭菌(clostridiummagnum)。在一些实施方案中,该微生物也可以是球形梭菌(clostridiumcoccoides)。在一些实施方案中,该微生物也可以是艰难梭菌(clostridiumdifficile)。在一些实施方案中,该微生物也可以是clostridiumformicaceticum。在一些实施方案中,该微生物也可以是clostridiumfervidus。在一些实施方案中,该微生物也可以是热纤梭菌(clostridiumthermocellum)。根据一些实施方案,第一方面的重组微生物属于穆尔氏菌属(moorella)。在一些实施方案中,该微生物可以是热醋穆尔氏菌(moorellathermoacetica)。在一些实施方案中,该微生物可以是moorellathermauto-trophica。在一些实施方案中,该微生物可以是moorellamulderi。在一些实施方案中,该微生物可以是moorellastamsii。在一些实施方案中,该微生物可以是甘油穆尔氏菌(moorellaglycerini)。在一些实施方案中,该微生物属于醋杆菌属(acetobacterium)。在一些实施方案中,该微生物可以是伍氏醋杆菌(acetobacteriumwoodii)。在一些实施方案中,该微生物可以是acetobacteriumbakii。在一些实施方案中,该微生物可以是甲醇醋杆菌(acetobacteriumcarbinolicum)。在一些实施方案中,该微生物可以是acetobacteriumdehalogenans。在一些实施方案中,该微生物可以是苹果酸醋杆菌(acetobacteriummalicum)。在一些实施方案中,该微生物可以是acetobacteriumpaludosum。在一些实施方案中,该微生物可以是acetobacteriumfimetarium。在一些实施方案中,该微生物可以是acetobacteriumtundrae。在一些实施方案中,该微生物可以是威氏醋杆菌(acetobacteriumwieringae)。在一些实施方案中,该微生物可以是acetobacteriumpsammolithicum。根据一些实施方案,第一方面的重组微生物属于鼠孢菌属(sporomusa)。在一些实施方案中,该微生物可以是卵形鼠孢菌(sporomusaovata)。在一些实施方案中,该微生物可以是sporomusasilvacetica。在一些实施方案中,该微生物可以是球形鼠孢菌(sporomusasphaeroides)。在一些实施方案中,该微生物可以是白蚁鼠孢菌(sporomusatermitida)。在一些实施方案中,该微生物可以是食酸鼠孢菌(sporomusaacidovorans)。在一些实施方案中,该微生物可以是sporomusaaerivorans。在一些实施方案中,该微生物可以是sporomusapaucivoransacidovorans。根据一些实施方案,该微生物属于blautia。在一些实施方案中,该微生物可以是blautiaproducta。在一些实施方案中,该微生物可以是blautiawexlerae。在一些实施方案中,该微生物可以是blautiahydrogenotrophica。在一些实施方案中,该微生物可以是blautiahansenii。根据一些实施方案,该微生物属于carboxydocella。在一些实施方案中,该微生物可以是carboxydocellasporoproducens。在一些实施方案中,该微生物可以是carboxydocellaferrireducens。在一些实施方案中,该微生物可以是carboxydocellathermautotrophica。根据一些实施方案,该微生物属于carboxydocella。在一些实施方案中,该微生物可以是carboxydocellasporoproducens。根据一些实施方案,该微生物属于氧化碳嗜热菌属(carboxydothermus)。在一些实施方案中,该微生物可以是生氢氧化碳嗜热菌(carboxydothermushydrogenoformans)。在一些实施方案中,该微生物可以是carboxydothermusferrireducens。在一些实施方案中,该微生物可以是carboxydothermuspertinax。在一些实施方案中,该微生物可以是carboxydothermusislandicus。在一些实施方案中,该微生物可以是carboxydothermussiderophilus。根据一些实施方案,该微生物属于脱硫肠状菌属(desulfotomaculum)。在一些实施方案中,该微生物可以是desulfotomaculumcarboxydivorans。在一些实施方案中,该微生物可以是desulfotomaculumgibsoniae。在一些实施方案中,该微生物可以是库氏脱硫肠状菌(desulfotomaculumkuznetsovii)。在一些实施方案中,该微生物可以是desulfotomaculumthermobenzoicumsubsp.thermosyntrophicum。在一些实施方案中,该微生物可以是醋酸氧化脱硫肠状菌(desulfotomaculumacetoxidans)。在一些实施方案中,该微生物可以是desulfotomaculumalcoholivorax。在一些实施方案中,该微生物可以是desulfotomaculumcarboxydivorans。在一些实施方案中,该微生物可以是desulfotomaculumsapomandens。在一些实施方案中,该微生物可以是desulfotomaculumthermocisternum。根据一些实施方案,第一方面的重组微生物属于脱硫弧菌属(desulfovibrio)。在一些实施方案中,该微生物可以是普通脱硫弧菌(desulfovibriovulgaris)。在一些实施方案中,该微生物可以是desulfovibrioalaskensis。在一些实施方案中,该微生物可以是脱硫脱硫弧菌(desulfovibriodesulfuricans)。在一些实施方案中,该微生物可以是普通脱硫弧菌。根据一些实施方案,该微生物属于分枝杆菌属(mycobacterium)。在一些实施方案中,该微生物可以是耻垢分枝杆菌(mycobacteriumsmegmatis)。在一些实施方案中,该微生物可以是戈氏分枝杆菌(mycobacteriumgordonae)。在一些实施方案中,该微生物可以是结核分枝杆菌(mycobacteriumtuberculosis)。根据一些实施方案,该微生物属于瘤胃球菌属(ruminococcus)。在一些实施方案中,该微生物可以是ruminococcushydrogenotrophicus。在一些实施方案中,该微生物可以是ruminococcusschinkii。在一些实施方案中,该微生物可以是产生瘤胃球菌(ruminococcusproductus)。根据一些实施方案,第一方面的重组微生物是以下的物种:食甲基丁酸杆菌(butyribacteriummethylotrophicum)、alkalibaculumbacchii、aecetoariaerobiumriotera、黏液真杆菌(eubacteriumlimosum)、desulfitbacteriumhafhierise、产生消化链球菌(peptostreptococcusproductus)、深红红螺菌(rhodospirillumrubrum)、thermoanaerobacterkiuvi、oxobacterpfennigii、阿拉伯糖醋盐杆菌(acetohalobiumarabaticum)、carbophiluscarboxidus、cloacibacillusevryensis、类黄色产氢噬胞菌(hydrogenophagapseudoflava)、沼泽红假单胞菌(rhodopseudomonaspalustris)、pseudomonasgazotropha、真氧产碱杆菌(ralstoniaeutropha)、calderihabitansmaritimus、caloribacteriumcisternae、carboxydobrachiumpacificum、desulfurisporathermophila、嗜热网球菌(dictyoglomusthermophilum)、hydrogenophilusislandicus、thermincolacarboxydiphila、thermincolaferriacetica、thermincolapotens、thermoacetogeniumphaeum、凯伍热厌氧菌(thermoanaerobacterkivui)、thermoanaerobacterthermohydrosulfuricussubsp.carboxydovorans、thermosinuscarboxydivorans、食羧寡养菌(oligotrophacarboxidovorans)、desulfosporosinusmeridiei、dehalococcoidesmccartyi、desulfatibacillumaliphaticivorans、自养脱硫杆菌(desulfobacteriumautotrophicum)、desulfobaculatoluolica、desulfospirajoergensenii、desulfosporosinusorientis、desulfosporosinusyoungiae、desulfovermiculushalophilus、desulfurisporathermophila、臭味全噬菌(holophagafoetida)、嗜树木甲烷短杆菌(methanobrevibacterarboriphilus)、oreniasalinaria、多粘类芽孢杆菌(paenibacilluspolymyxa)、tindalliacaliforniensis、anoxybacillusflavithermus、desulfovirgulathermocuniculi、thermosediminibacteroceani、candidatusscalinduabrodae、潮湿厌氧醋菌(acetoanaerobiumnoterae)、ammonifexdegensii、瘤胃聚乙酸菌(acetitomaculumruminis)、acetoanaerobiumromashkovil、长醋丝菌(acetonemalongum)、bryanellaformatexigens、灼热喜热菌(caloramatorfervidus)、natroniellaacetigena、natronincolahistinovorans、糖变共养球菌(syntrophococcussucromutans)、treponemaprimitia、pseudomonascarboxydohyrogena、热食碳酸假单胞菌(pseudomonasthermocarboxydovorans)、大豆慢生根瘤菌(bradyrhizobiumjaponicum)、热自养链霉菌(streptomycesthermoautotrophicus)、施氏芽孢杆菌(bacillusschlegelii)、caldanaerobactersubterraneus、thermolithobactercarboxydivorans、thermococcusonnurineus、thermofilumcarboxyditrophus、闪烁古生球菌(archaeoglobusfulgidus)、蒂杰氏脱硫项圈菌(desulfomoniletiedjei)、附着热变形菌(thermoproteustenax)或rubrivivaxgelatinosa。根据第二方面,提供重组核酸分子。该重组核酸分子包含编码使乙酰coa转化为主链具有2至6个碳原子的一种或多种烷烃和/或烯烃的一种或多种酶的核酸序列。在该重组核酸分子包含编码催化乙酰coa转化为主链具有2至6个碳原子的一种或多种烷烃和/或烯烃的一种或多种酶的一种以上编码序列或一种以上编码序列的组合的实施方案中,该重组核酸分子可以不包含该编码序列中的至少一种。在一些实施方案中,该重组核酸分子不包含编码催化乙酰coa转化为主链具有2至6个碳原子的一种或多种烷烃和/或烯烃的一种或多种酶的至少一种编码序列或编码序列的组合。在一些实施方案中,该重组核酸分子包含编码使乙酰coa转化为主链具有2至5个碳原子的一种或多种烯烃的一种或多种酶的核酸序列。该重组核酸分子可以包含编码催化乙酰coa转化为巴豆酰coa和进一步催化巴豆酰coa转化为烯烃的一种或多种酶的一种或多种编码序列。该重组核酸分子还可以包含编码催化乙酰coa转化为3-甲基巴豆酰coa和进一步催化3-甲基巴豆酰coa转化为烯烃的一种或多种酶的一种或多种编码序列。该重组核酸分子还可以包含编码催化乙酰coa转化为丙酰coa和进一步催化丙酰coa转化为烯烃的一种或多种酶的一种或多种编码序列。包含在该重组核酸分子中的各编码序列与转录启动子有效连接。在一些实施方案中,该重组核酸分子包含编码固氮酶ec1.18.6.1或ec1.19.6.1的编码序列。在第二方面的重组核酸分子的一些实施方案中,该催化乙酰coa转化为巴豆酰coa的一种或多种酶包括:乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);乙酰乙酰coa还原酶(ec1.1.1.36);和烯酰coa水合酶2(ec4.2.1.119)。在第二方面的重组核酸分子的一些实施方案中,该催化乙酰coa转化为巴豆酰coa的一种或多种酶包括:乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);和3-羟基丁酰coa脱水酶。在一些实施方案中,该催化乙酰coa转化为巴豆酰coa的一种或多种酶包括:4-氨基丁酸氨基转移酶(ec2.6.1.19);谷氨酸脱氢酶(ec1.4.1.2);2-羟基戊二酸脱氢酶(ec1.1.99.2);戊烯二酸coa转移酶(ec2.8.3.12);2-羟基戊二酰coa脱水酶(ec4.2.1.-);和戊烯二酸单酰coa脱羧酶(ec4.1.1.70)。在一些实施方案中,该催化乙酰coa转化为巴豆酰coa的一种或多种酶包括:4-氨基丁酸氨基转移酶(ec2.6.1.19);4-羟基丁酸脱氢酶(ec1.1.1.61);4-羟基丁酸coa转移酶(ec2.8.3.-);乙烯乙酰coaδ异构酶(ec4.2.1.120);和4-羟基丁酰coa脱水酶(ec5.3.3.3)。在第二方面的重组核酸分子的一些实施方案中,该催化乙酰coa转化为3-甲基巴豆酰coa的一种或多种酶包括:乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10);3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18);和3-甲基巴豆酰coa羧化酶(ec6.4.1.4)。在一些实施方案中,该催化乙酰coa转化为3-甲基巴豆酰coa的一种或多种酶包括:乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10);3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18);和3-甲基巴豆酰coa羧化酶(ec6.4.1.4)。在第二方面的重组核酸分子的一些实施方案中,该催化乙酰coa转化为丙酰coa的一种或多种酶包括:乙酰coa羧化酶(ec6.4.1.2);丙二酸单酰coa还原酶(ec1.2.1.75);丙二酰coa还原酶(ec1.1.1.298);丙酰coa合酶(ec6.2.1.36);丙酰coa合酶/丙烯酰coa合酶(ec4.2.1.116);和丙酰coa合酶(ec1.3.1.84)。在第二方面的重组核酸分子的一些实施方案中,该催化乙酰coa转化为丙酰coa的一种或多种酶包括:乙酰coa羧化酶(ec6.4.1.2);甲基丙二酸单酰coa变位酶(ec5.4.99.2);和甲基丙二酸单酰coa脱羧酶(ec4.1.1.41)。在一些实施方案中,该催化乙酰coa转化为丙酰coa的一种或多种酶包括:2-氧代戊二酸/丁酸铁氧还蛋白氧化还原酶(ec1.2.7.3)和/或丙酮酸/2-氧代丁酸铁氧还蛋白氧化还原酶(ec1.2.7.1)。在一些实施方案中,该催化乙酰coa转化为丙酰coa的一种或多种酶包括:乳酸coa转移酶(ec2.8.3.-);乳酰coa脱水酶(ec4.2.1.54);和丙酰coa合酶(ec1.3.1.84)。在第二方面的重组核酸分子的一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括:乙酰coa羧化酶(ec6.4.1.2);丙二酸单酰coa还原酶(ec1.2.1.75);丙二酰coa还原酶(ec1.1.1.298);丙酰coa合酶(ec6.2.1.36);和丙酰coa合酶/丙烯酰coa合酶(ec4.2.1.116)。在一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括乳酸coa转移酶(ec2.8.3.-);和乳酰coa脱水酶(ec4.2.1.54)。在一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括酰基coa硫酯酶(ec3.1.2.-)和(b)苯基丙烯酸脱羧酶(ec4.1.1.-)。在第二方面的重组核酸分子的一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括:磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括:反式-2-烯酰coa还原酶(ec1.1.1.36);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-)和/或磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)的组合;及苯基丙烯酸脱羧酶(ec4.1.1.-)。在第二方面的重组核酸分子的一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括:醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括:醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在第二方面的重组核酸分子的一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括以下的组合:酰基coa硫酯酶(ec3.1.2.-)和醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括以下的组合:酰基coa硫酯酶(ec3.1.2.-)和醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在第二方面的重组核酸分子的一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括以下的组合:醛铁氧还蛋白氧化还原酶(ec1.2.7.5)和酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括以下的组合:醛铁氧还蛋白氧化还原酶(ec1.2.7.5)和酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在第二方面的重组核酸分子的一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括以下的组合:醛铁氧还蛋白氧化还原酶(ec1.2.7.5)和酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括以下的组合:醛铁氧还蛋白氧化还原酶(ec1.2.7.5)和酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在第二方面的重组核酸分子的一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括以下的组合:磷酸丁酰转移酶(ec2.3.1.19)、丁酸激酶(ec2.7.2.7)和醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括以下的组合:磷酸丁酰转移酶(ec2.3.1.19)、丁酸激酶(ec2.7.2.7)和醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在第二方面的重组核酸分子的一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括以下的组合:磷酸丁酰转移酶(ec2.3.1.19)、丁酸激酶(ec2.7.2.7)和醛铁氧还蛋白氧化还原酶(ec1.2.7.5);(m)醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化乙酰coa转化为丙烯酰coa的一种或多种酶包括以下的组合:磷酸丁酰转移酶(ec2.3.1.19)、丁酸激酶(ec2.7.2.7)和醛铁氧还蛋白氧化还原酶(ec1.2.7.5);(m)醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在第二方面的重组核酸分子的一些实施方案中,该催化3-甲基巴豆酰coa转化为烯烃的一种或多种酶包括:苯基丙烯酸脱羧酶(ec4.1.1.-);和酰基coa硫酯酶(ec3.1.2.-);和/或磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)的组合。在第二方面的重组核酸分子的一些实施方案中,该催化3-甲基巴豆酰coa转化为烯烃的一种或多种酶包括以下的组合:酰基coa硫酯酶(ec3.1.2.-)和醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化3-甲基巴豆酰coa转化为烯烃的一种或多种酶包括以下的组合:酰基coa硫酯酶(ec3.1.2.-)和醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在第二方面的重组核酸分子的一些实施方案中,该催化3-甲基巴豆酰coa转化为烯烃的一种或多种酶包括:醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化3-甲基巴豆酰coa转化为烯烃的一种或多种酶包括:醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在第二方面的重组核酸分子的一些实施方案中,该催化3-甲基巴豆酰coa转化为烯烃的一种或多种酶包括以下的组合:醛铁氧还蛋白氧化还原酶(ec1.2.7.5)和酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在第二方面的重组核酸分子的一些实施方案中,该催化3-甲基巴豆酰coa转化为烯烃的一种或多种酶包括以下的组合:醛铁氧还蛋白氧化还原酶(ec1.2.7.5)和酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在第二方面的重组核酸分子的一些实施方案中,该催化3-甲基巴豆酰coa转化为烯烃的一种或多种酶包括以下的组合:磷酸丁酰转移酶(ec2.3.1.19)、丁酸激酶(ec2.7.2.7)和醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化3-甲基巴豆酰coa转化为烯烃的一种或多种酶包括以下的组合:磷酸丁酰转移酶(ec2.3.1.19)、丁酸激酶(ec2.7.2.7)和醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在第二方面的重组核酸分子的一些实施方案中,该催化3-甲基巴豆酰coa转化为烯烃的一种或多种酶包括以下的组合:磷酸丁酰转移酶(ec2.3.1.19)、丁酸激酶(ec2.7.2.7)和醛铁氧还蛋白氧化还原酶(ec1.2.7.5);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该催化3-甲基巴豆酰coa转化为烯烃的一种或多种酶包括以下的组合:磷酸丁酰转移酶(ec2.3.1.19)、丁酸激酶(ec2.7.2.7)和醛铁氧还蛋白氧化还原酶(ec1.2.7.5);醛脱氢酶(ec1.2.1.-)和/或醇脱氢酶(ec1.1.1.-);及(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在第二方面的重组核酸分子的一些实施方案中,该催化丙酰coa转化为烯烃的一种或多种酶包括:乙酰coac-乙酰转移酶(ec2.3.1.9);乙酰乙酰coa还原酶(ec1.1.1.36)和烯酰coa水合酶2(ec4.2.1.119)的组合;酰基coa硫酯酶(ec3.1.2.-)和/或磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)的组合;及苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该催化丙酰coa转化为烯烃的一种或多种酶包括:乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157)和3-羟基丁酰coa脱水酶(ec4.2.1.55)的组合;酰基coa硫酯酶(ec3.1.2.-)和/或磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)的组合;及苯基丙烯酸脱羧酶(ec4.1.1.-)。在第二方面的重组核酸分子的一些实施方案中,该催化丙烯酰coa转化为烯烃的一种或多种酶包括:酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该催化丙烯酰coa转化为烯烃的一种或多种酶包括:磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该重组核酸分子包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:4-氨基丁酸氨基转移酶(ec2.6.1.19);谷氨酸脱氢酶(ec1.4.1.2);2-羟基戊二酸脱氢酶(ec1.1.99.2);戊烯二酸coa转移酶(ec2.8.3.12);2-羟基戊二酰coa脱水酶(ec4.2.1.-);戊烯二酸单酰coa脱羧酶(ec4.1.1.70);酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:4-氨基丁酸氨基转移酶(ec2.6.1.19);4-羟基丁酸脱氢酶(ec1.1.1.61);4-羟基丁酸coa转移酶(ec2.8.3.-);乙烯乙酰coaδ异构酶(ec4.2.1.120);4-羟基丁酰coa脱水酶(ec5.3.3.3);酰基coa硫酯酶(ec3.1.2.-);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:乙酰coac-乙酰转移酶(ec2.3.1.9)和/或乙酰coa羧化酶(ec6.4.1.2)和乙酰乙酰coa合酶(ec2.3.1.194)、3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10)、3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18)和3-甲基巴豆酰coa羧化酶(ec6.4.1.4);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127),和/或(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的组合的编码序列:乙酰coac-乙酰转移酶(ec2.3.1.9)、3-羟基丁酰coa脱氢酶(ec1.1.1.157)和3-羟基丁酰coa脱水酶(ec4.2.1.55);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127),和/或(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的组合的编码序列:乙酰coa羧化酶(ec6.4.1.2)、乙酰乙酰coa合酶(ec2.3.1.194)、乙酰乙酰coa还原酶(ec1.1.1.36)和烯酰coa水合酶2(ec4.2.1.119);醛脱氢酶(ec1.2.1.-)或醇脱氢酶(ec1.1.1.-);及牻牛儿醇异构酶(ec5.4.4.4),其也是芳樟醇脱水酶(ec4.2.1.127),和/或(i)法尼醇激酶(ec2.7.1.-)、牻牛儿基牻牛儿醇激酶(ec2.7.1.b19)、羟乙基噻唑激酶(ec2.7.1.50)或十一异戊烯醇激酶(ec2.7.1.66)中的一种或多种和(ii)2-甲基-1,3-丁二烯合酶(ec4.2.3.27)或甲基丁烯醇合酶中的一种或多种的组合。在一些实施方案中,该第二方面的重组核酸分子包含编码以下的编码序列:(i)乙酰coac-乙酰转移酶(ec2.3.1.9)、3-羟基丁酰coa脱氢酶(ec1.1.1.157)和3-羟基丁酰coa脱水酶(ec4.2.1.55)的组合,和/或乙酰coa羧化酶(ec6.4.1.2)、乙酰乙酰coa合酶(ec2.3.1.194)、乙酰乙酰coa还原酶(ec1.1.1.36)和烯酰coa水合酶2(ec4.2.1.119)的组合;(ii)磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)和/或酰基coa硫酯酶(ec3.1.2.-)的组合;和(iii)苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下的编码序列:(i)乙酰coac-乙酰转移酶(ec2.3.1.9)和/或乙酰coa羧化酶(ec6.4.1.2)和乙酰乙酰coa合酶(ec2.3.1.194)的组合;3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10);3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18);和3-甲基戊烯二酸单酰coa脱羧酶;(ii)磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)和/或酰基coa硫酯酶(ec3.1.2.-)的组合;和(iii)苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:乳酸coa转移酶(ec2.8.3.-);乳酰coa脱水酶(ec4.2.1.54);和酰基coa硫酯酶(ec3.1.2.-)。在一些实施方案中,该重组核酸分子包含编码以下酶的编码序列:乳酸coa转移酶(ec2.8.3.-)、乳酰coa脱水酶(ec4.2.1.54);磷酸丁酰转移酶(ec2.3.1.19);丁酸激酶(ec2.7.2.7);和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:4-氨基丁酸氨基转移酶(ec2.6.1.19);4-羟基丁酸脱氢酶(ec1.1.1.61);4-羟基丁酸coa转移酶(ec2.8.3.-);乙烯乙酰coaδ异构酶(ec4.2.1.120);4-羟基丁酰coa脱水酶(ec5.3.3.3);磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)和/或酰基coa硫酯酶(ec3.1.2.-)的组合;和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该重组核酸分子包含编码以下酶的编码序列:4-氨基丁酸氨基转移酶(ec2.6.1.19);谷氨酸脱氢酶(ec1.4.1.2);2-羟基戊二酸脱氢酶(ec1.1.99.2);戊烯二酸coa转移酶(ec2.8.3.12);2-羟基戊二酰coa脱水酶(ec4.2.1.-);戊烯二酸单酰coa脱羧酶(ec4.1.1.70);磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)和/或酰基coa硫酯酶(ec3.1.2.-)的组合;和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);反式-2-烯酰coa还原酶(ec1.1.1.36);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)和/或酰基coa硫酯酶(ec3.1.2.-)的组合;和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);乙酰乙酰coa合酶(ec2.3.1.194);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);反式-2-烯酰coa还原酶(ec1.1.1.36);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)和/或酰基coa硫酯酶(ec3.1.2.-)的组合;和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);甲基丙二酸单酰coa变位酶(ec5.4.99.2);甲基丙二酸单酰coa脱羧酶(ec4.1.1.41);乙酰coac-乙酰转移酶(ec2.3.1.9);(i)乙酰乙酰coa还原酶(ec1.1.1.36)和烯酰coa水合酶2(ec4.2.1.119)和/或(ii)3-羟基丁酰coa脱氢酶(ec1.1.1.157)和3-羟基丁酰coa脱水酶(ec4.2.1.55)的组合;(vi)酰基coa硫酯酶(ec3.1.2.-);乙酰coac-乙酰转移酶(ec2.3.1.9);乙酰乙酰coa还原酶(ec1.1.1.36);烯酰coa水合酶2(ec4.2.1.119);磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)和/或酰基coa硫酯酶(ec3.1.2.-)的组合;和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);甲基丙二酸单酰coa变位酶(ec5.4.99.2);甲基丙二酸单酰coa脱羧酶(ec4.1.1.41);乙酰coac-乙酰转移酶(ec2.3.1.9);(i)乙酰乙酰coa还原酶(ec1.1.1.36)和烯酰coa水合酶2(ec4.2.1.119)和/或(ii)3-羟基丁酰coa脱氢酶(ec1.1.1.157)和3-羟基丁酰coa脱水酶(ec4.2.1.55)的组合;酰基coa硫酯酶(ec3.1.2.-);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)和/或酰基coa硫酯酶(ec3.1.2.-)的组合;和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:2-氧代戊二酸/丁酸铁氧还蛋白氧化还原酶(ec1.2.7.3);丙酮酸/2-氧代丁酸铁氧还蛋白氧化还原酶(ec1.2.7.1);乳酰coa脱水酶(ec4.2.1.54);乙酰coac-乙酰转移酶(ec2.3.1.9);(i)乙酰乙酰coa还原酶(ec1.1.1.36)和烯酰coa水合酶2(ec4.2.1.119)和/或(ii)3-羟基丁酰coa脱氢酶(ec1.1.1.157)和3-羟基丁酰coa脱水酶(ec4.2.1.55)的组合;磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)和/或酰基coa硫酯酶(ec3.1.2.-)的组合;和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:乳酸coa转移酶(ec2.8.3.-);乳酰coa脱水酶(ec4.2.1.54);丙酰coa合酶(ec1.3.1.84);乳酰coa脱水酶(ec4.2.1.54);乙酰coac-乙酰转移酶(ec2.3.1.9);(i)乙酰乙酰coa还原酶(ec1.1.1.36)和烯酰coa水合酶2(ec4.2.1.119)和/或(ii)3-羟基丁酰coa脱氢酶(ec1.1.1.157)和3-羟基丁酰coa脱水酶(ec4.2.1.55)的组合;磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)和/或酰基coa硫酯酶(ec3.1.2.-)的组合;和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);丙二酸单酰coa还原酶(ec1.2.1.75);丙二酰coa还原酶(ec1.1.1.298);丙酰coa合酶(ec6.2.1.36);丙酰coa合酶/丙烯酰coa合酶(ec4.2.1.116);丙酰coa合酶(ec1.3.1.84);乳酰coa脱水酶(ec4.2.1.54);乙酰coac-乙酰转移酶(ec2.3.1.9);(i)乙酰乙酰coa还原酶(ec1.1.1.36)和烯酰coa水合酶2(ec4.2.1.119)和/或(ii)3-羟基丁酰coa脱氢酶(ec1.1.1.157)和3-羟基丁酰coa脱水酶(ec4.2.1.55)的组合;磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)和/或酰基coa硫酯酶(ec3.1.2.-)的组合;和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:4-氨基丁酸氨基转移酶(ec2.6.1.19);(i)谷氨酸脱氢酶(ec1.4.1.2)、2-羟基戊二酸脱氢酶(ec1.1.99.2)、戊烯二酸coa转移酶(ec2.8.3.12)、2-羟基戊二酰coa脱水酶(ec4.2.1.-)和/或(ii)4-羟基丁酸脱氢酶(ec1.1.1.61)、4-羟基丁酸coa转移酶(ec2.8.3.-)、乙烯乙酰coaδ异构酶(ec4.2.1.120)和戊烯二酸单酰coa脱羧酶(ec4.1.1.70)的组合;4-羟基丁酰coa脱水酶(ec5.3.3.3);反式-2-烯酰coa还原酶(ec1.1.1.36);乙酰coac-乙酰转移酶(ec2.3.1.9);3-羟基丁酰coa脱氢酶(ec1.1.1.157);3-羟基丁酰coa脱水酶(ec4.2.1.55);酰基coa硫酯酶(ec3.1.2.-);和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该第二方面的重组核酸分子包含编码以下酶的编码序列:乙酰coa羧化酶(ec6.4.1.2);丙二酸单酰coa还原酶(形成丙二酸半醛)(ec1.2.1.75);3-羟基丙酸脱氢酶(ec1.1.1.298);3-羟基丙酰coa合酶(ec6.2.1.36);羟基丙酰coa脱水酶(ec4.2.1.116);磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)和/或酰基coa硫酯酶(ec3.1.2.-)的组合;和苯基丙烯酸脱羧酶(ec4.1.1.-)。在一些实施方案中,该重组核酸分子包含编码以下的编码序列:(i)乙酰coa羧化酶(ec6.4.1.2)、乳酸coa转移酶(ec2.8.3.-)和乳酰coa脱水酶(ec4.2.1.54)和/或(ii)乳酸coa转移酶(ec2.8.3.-)和乳酰coa脱水酶(ec4.2.1.54)的组合;磷酸丁酰转移酶(ec2.3.1.19)和丁酸激酶(ec2.7.2.7)和/或酰基coa硫酯酶(ec3.1.2.-)的组合;和苯基丙烯酸脱羧酶(ec4.1.1.-)。根据第三方面,提供产生主链具有2至5个碳原子的一种或多种烯烃的方法。该方法包括使第二方面的重组微生物与一氧化碳和/或二氧化碳接触。该第三方面的方法可以是产生乙烯的方法。该方法还可以是产生1-丙烯的方法。在一些实施方案中,该方法可以是产生1-丁烯的方法。在一些实施方案中,该方法可以是产生2-甲基丙烯的方法。在一些实施方案中,该方法可以是产生1,3-丁二烯的方法。在一些实施方案中,该方法可以是产生1-戊烯的方法。在一些实施方案中,该方法可以是产生2-甲基-1,3-丁二烯的方法。其他优势将部分显示在以下描述中,部分将从该描述和附图变得显而易见,或者可以通过实施本文中公开的方法或微生物而了解到。该方法和微生物的优势将借助所附权利要求书中具体指出的要素和组合来实现和获得。应理解,以上发明概述和以下发明详述仅是示例性和说明性的,不是所主张的发明的限制。附图简述图1a显示用于从co、co2和/或cn及电子产生烃的四条固氮酶途径。图1b显示用于从一氧化碳和/或从二氧化碳以及从二氧化碳和电子和/或从一氧化碳和电子形成乙烯的途径,包括关键酶。图1c显示用于从一氧化碳和/或从二氧化碳以及从二氧化碳和电子和/或从一氧化碳和电子形成丙烯的途径,包括关键酶。图1d显示用于从一氧化碳和/或从二氧化碳以及从二氧化碳和电子和/或从一氧化碳和电子形成1-丁烯的途径,包括关键酶。图1e显示用于从一氧化碳和/或从二氧化碳以及从二氧化碳和电子和/或从一氧化碳和电子形成异丁烯的途径,包括关键酶。图1f显示用于从一氧化碳和/或从二氧化碳以及从二氧化碳和电子和/或从一氧化碳和电子形成1,3-丁二烯的途径,包括关键酶。图1g显示用于从一氧化碳和/或从二氧化碳以及从二氧化碳和电子和/或从一氧化碳和电子形成2-甲基-1,3-丁二烯的途径,包括关键酶。(dmap=二甲基烯丙基单磷酸)图1h显示用于从一氧化碳和/或从二氧化碳以及从二氧化碳和电子和/或从一氧化碳和电子形成1-戊烯的途径,包括关键酶。图2a显示用于由随机切割和粘贴转座酶介导的dna整合入细菌染色体的基于质粒的系统的基本设计原理。图2b显示正选择在分离目的突变体中的用途。5’itr(转座酶识别和切割的反向末端重复)和lox66位点作为红霉素/克拉霉素(clarythromycin)抗生素抗性标记misr的启动子。在定位在质粒上时,结合lac操纵基因序列并阻断转录的阻抑蛋白laci失活misr的表达。转座后,操纵基因序列阻抑蛋白lac操纵基因序列丧失,并赋予抗性,因为misr得以表达,并可以针对其进行选择(rbs=核糖体结合位点)。图3a示意性显示所实施的正选择系统的作用方式。阻抑蛋白结合操纵基因位点阻断了抗生素抗性基因的表达。32:3-羟基丁酰coa脱氢酶(ec1.1.1.157);34:3-羟基丁酰coa脱水酶(ec4.2.1.55);36:酰基coa硫酯酶[ec3.1.2.-];38:醛/醇脱氢酶(ec1.1.1.-和ec1.2.1.-);40:牻牛儿醇异构酶/芳樟醇脱水酶(ec5.4.4.4和ec4.2.1.127);42:乙酰coa羧化酶(ec6.4.1.2)。图3b示意性显示用于由随机切割和粘贴转座酶介导的dna整合入细菌染色体的基于质粒的系统的基本设计原理。36:酰基coa硫酯酶(ec3.1.2.-);38:醛/醇脱氢酶(ec1.1.1.-和ec1.2.1.-);40:牻牛儿醇异构酶/芳樟醇脱水酶(ec5.4.4.4和ec4.2.1.127);44:乙酰乙酰coa合酶(ec2.3.1.194)。图4a显示质粒sg193(seqidno:1),其包含编码钼固氮酶途径的酶的序列。图4b显示质粒sg211(seqidno:2),其包含编码钒固氮酶途径的酶的序列。图4c显示质粒sg278(seqidno:3),其包含编码铁固氮酶途径的酶的序列。图4d显示质粒sg323(seqidno:4),其包含编码改造固氮酶的序列。图5a显示质粒sg123(seqidno:5),其包含编码mep途径的酶连同kudzu2-甲基-1,3-丁二烯合酶的序列。图5b显示质粒sg124(seqidno:6),其包含编码mep途径的酶连同populus2-甲基-1,3-丁二烯合酶的序列。图6显示质粒sg156(seqidno:7),其包含编码mep途径的酶连同kudzu2-甲基-1,3-丁二烯合酶的序列。图7显示质粒sg157(seqidno:8),其包含编码mep途径的酶连同populus2-甲基-1,3-丁二烯合酶的序列。图8a显示以二氧化碳和电子作为反应物,用包含质粒sg156的杨氏梭菌发酵产生而获得的产物2-甲基-1,3-丁二烯的产物分析中的gc-ms层析图。图8b显示图8a中测量和显示的鉴定为2-甲基-1,3-丁二烯的峰的质谱。图9示意性显示载体pant(seqidno:9),其是遗传转化丙酮丁醇梭菌(clostridiumacetobutylicum)所需的甲基化质粒。图10示意性显示载体bg132(缩写为t1,seqidno:10),其是包含染色体整合的正选择系统的用于经转座酶进行染色体整合的质粒。图11显示载体bg133(缩写为t2,seqidno:11)的载体图谱。图12显示载体bg134(缩写为t3,seqidno:12)的载体图谱。图13显示载体bg135(缩写为t4,seqidno:13)的载体图谱。图14显示载体bg136(缩写为h,seqidno:14)的载体图谱。图15a显示丙酮丁醇梭菌dnapcr(产物)的0.8%分析型dna琼脂糖凝胶电泳分离。泳道1:标准品;泳道2-6:整合体(integrand)1pcr1-5;泳道7-11:整合体2pcr1-5;泳道12:标准品;泳道13-17:整合体3pcr1-5;泳道18-22:整合体4pcr1-5;泳道23-27:整合体5pcr1-5;泳道28:标准品。图15b显示pcr(产物)的0.8%分析型dna琼脂糖凝胶电泳分离。泳道1:标准品;泳道2-6:整合体6pcr1-5;泳道7-11:整合体7pcr1-5;泳道12-16:整合体8pcr1-5;泳道17:标准品;泳道18-22:野生型pcr1-5;泳道23-27:整合体5pcr1-5;泳道28:标准品。图16显示整合体的荧光显微镜照片。图16a、16b、16c:丙酮丁醇梭菌整合体1;图16d:丙酮丁醇梭菌整合体2;图16e、16f、16g:丙酮丁醇梭菌整合体2;图16h:丙酮丁醇梭菌野生型对照。图17显示载体bg168(缩写为“i”,seqidno:15)的载体图谱。图18显示载体bg182(seqidno:16)的载体图谱。图19显示载体bg282(seqidno:17)的载体图谱。图20显示载体bg281(seqidno:18)的载体图谱。图21显示载体bg287(seqidno:19)的载体图谱。图22显示载体bg288(seqidno:20)的载体图谱。图23显示载体bg289(seqidno:21)的载体图谱。图24显示载体bg290(seqidno:22)的载体图谱。图25显示载体bg291(seqidno:23)的载体图谱。图26显示载体bg292(seqidno:24)的载体图谱。图27显示载体bg178(seqidno:25)的载体图谱。图28显示杨氏梭菌dnapcr产物的0.8%分析型dna琼脂糖凝胶电泳分离。泳道1:标准品;泳道2-9:整合体lbi105-111pcr;泳道10:标准品;泳道11-18:整合体lbi112-120pcr;泳道19:标准品;泳道24-25:阳性对照pcr;泳道26-27:阴性对照gdna杨氏梭菌gdnapcr;泳道28-29:阴性对照gdnac.3。图29显示clostridiumautoethanogenumdnapcr产物的0.8%分析型dna琼脂糖凝胶电泳分离。泳道1:标准品;泳道2-5:整合体a1-a4pcr;泳道6-7:阳性对照pcr;泳道8-9:阴性对照gdna杨氏梭菌gdnapcr;泳道10-11:阴性对照gdnac.autoethanogenumgdnapcr;泳道12:标准品。图30示意性显示质粒sg193(seqidno:26),其包含编码钼固氮酶途径的酶的序列。图31示意性显示质粒sg211(seqidno:27),其包含编码钒固氮酶途径的酶的序列。图32示意性显示质粒sg278(seqidno:28),其包含编码铁固氮酶途径的酶的序列。图33示意性显示质粒sg323(seqidno:29),其包含改造固氮酶复合体。图34示意性显示质粒sg387(seqidno:30),其包含编码1,3-丁二烯途径1型(巴豆酰coa生物合成源自丙酮丁醇梭菌)的酶的序列。图35示意性显示质粒sg411(seqidno:31),其包含编码1,3-丁二烯途径2型(经丙二酸单酰coa的巴豆酰coa生物合成源自大肠杆菌(escherichiacoli)和其他细菌)的酶的序列。图36示意性显示质粒sg455(seqidno:32),其包含编码用于丙烯生物合成的酶的序列。图37示意性显示质粒sg479(seqidno:33),其包含编码用于丙烯生物合成的酶的序列。图38示意性显示质粒sg539(seqidno:34),其包含编码用于1-丁烯生物合成的酶的序列。图39示意性显示质粒sg523(seqidno:35),其包含编码用于1-丁烯生物合成的酶的序列。图40示意性显示质粒sg582(seqidno:36),其包含编码用于异丁烯生物合成的酶的序列。图41示意性显示质粒sg601(seqidno:37),其包含编码用于异丁烯生物合成的酶的序列。图42示意性显示质粒sg498(seqidno:38),其包含编码用于1-戊烯生物合成的酶的序列。图43示意性显示质粒sg513(seqidno:39),其包含编码用于1-戊烯生物合成的酶的序列。图44示意性显示质粒sg557(seqidno:40),其包含编码用于乙烯生物合成的酶的序列。图45示意性显示质粒sg598(seqidno:41),其包含编码用于乙烯烯生物合成的酶的序列。图46示意性显示质粒sg661(seqidno:42),其包含编码1,3-丁二烯途径3型(经丙二酸单酰coa的巴豆酰coa合成源自大肠杆菌和其他细菌,经典途径源自丙酮丁醇梭菌)的酶的序列。图47示意性显示质粒sg696(seqidno:43),其包含编码1,3-丁二烯途径4型(经谷氨酸和4-氨基丁酸的巴豆酰coa合成)的酶的序列。图48示意性显示质粒sg726(seqidno:44),其包含编码2-甲基-1,3-丁二烯生物合成途径的酶的序列和染色体整合的正选择系统。图49示意性显示质粒sg705(seqidno:45),其包含编码2-甲基-1,3-丁二烯生物合成途径的酶的序列和染色体整合的正选择系统。图50示意性显示质粒sg714(seqidno:46),其包含编码1,3-丁二烯生物合成途径的酶的序列和染色体整合的正选择系统。图51示意性显示质粒sg739(seqidno:47),其包含编码1,3-丁二烯生物合成途径的酶的序列和染色体整合的正选择系统。图52示意性显示质粒sg755(seqidno:48),其包含编码丙烯生物合成途径的酶的序列和染色体整合的正选择系统。图53示意性显示质粒sg779(seqidno:49),其包含编码丙烯生物合成途径的酶的序列和染色体整合的正选择系统。图54示意性显示质粒sg839(seqidno:50),其包含编码1-丁烯生物合成途径的酶的序列和染色体整合的正选择系统。图55示意性显示质粒sg823(seqidno:51),其包含编码1-丁烯生物合成途径的酶的序列和染色体整合的正选择系统。图56示意性显示质粒sg882(seqidno:52),其包含编码异丁烯生物合成途径的酶的序列和染色体整合的正选择系统。图57示意性显示质粒sg901(seqidno:53),其包含编码异丁烯生物合成途径的酶的序列和染色体整合的正选择系统。图58示意性显示质粒sg798(seqidno:54),其包含编码1-戊烯生物合成途径的酶的序列和染色体整合的正选择系统。图59示意性显示质粒sg813(seqidno:55),其包含编码1-戊烯生物合成途径的酶的序列和染色体整合的正选择系统。图60示意性显示质粒sg857(seqidno:56),其包含编码乙烯生物合成途径的酶的序列和染色体整合的正选择系统。图61示意性显示质粒sg898(seqidno:57),其包含编码乙烯生物合成途径的酶的序列和染色体整合的正选择系统。图62列出用质粒n和质粒i通过系列转移(正选择)进行的初始整合测试。图63显示在图62中所示的测试中获得的整合体的gdna的制备。r/c:副本/克隆;cla:克拉霉素(μg/ml)。图64显示通过pcr基因组步移进行的图63中所分析的整合体的基因座的测定。图65总结用于分析整合体的基因组dna的流程。gdna浓度用nanodrop分光光度计(thermofisherscientificinc.,darmstadt,德国)测定。图66显示由本文中公开的示例性质粒编码的酶。图66a显示由包含导致异丁烯生物合成的途径的酶的核酸序列的四种质粒编码的酶。图66b显示由包含导致异丙烯生物合成的途径的酶的核酸序列的两种质粒编码的酶。图66c显示由包含导致异丙烯生物合成的途径的酶的核酸序列的六种质粒编码的酶。图67a显示以二氧化碳和电子作为反应物,用包含质粒sg601的杨氏梭菌dsm13528发酵产生而获得的产物异丁烯的产物分析中的gc-ms层析图。(inten.=强度)图67b显示图67a中测量和显示的鉴定为异丁烯的峰的质谱。(int.=强度)在附图通篇中,以下参考数字适用:20-转座酶(例如himar1)22-转座酶识别位点(5’itr反向末端重复)24-抗生素抗性标记(mlsr)26-目的基因(负载)28-转座酶识别位点(3’itr反向末端重复)30-复制起点31-乙酰乙酰辅酶a合酶硫解酶(ec2.3.1.9)=atob32-3-羟基丁酰coa脱氢酶(ec1.1.1.157)=hbd34-3-羟基丁酰coa脱水酶(ec4.2.1.55)=crt36-酰基coa硫酯酶(ec3.1.2.-)=ycia,ydii38-苯基丙烯酸脱羧酶(ec4.1.1.-)=pad1,padc40-牻牛儿醇异构酶/芳樟醇脱水酶(ec5.4.4.4和ec4.2.1.127)=gim,git42-乙酰coa羧化酶(ec6.4.1.2)=accabcd44-乙酰乙酰coa合酶(ec2.3.1.194)=npht746-乙酰乙酰coa还原酶(ec1.1.1.36)=phab148-(r)-特异性烯酰coa水合酶(ec4.2.1.119)=phaj50-4-氨基丁酸氨基转移酶(ec2.6.1.19)=gabt52-谷氨酸脱氢酶(ec1.4.1.2)=gdh54-2-羟基戊二酸脱氢酶(ec1.1.99.2)=ygaf56-戊烯二酸coa转移酶(ec2.8.3.12)=gcta58-2-羟基戊二酰coa脱水酶(ec4.2.1.-)=hdgab60-戊烯二酸单酰coa脱羧酶(ec4.1.1.70)=gcdabcd62-4-羟基丁酸脱氢酶,也称为nad依赖型4-羟基丁酸脱氢酶(ec1.1.1.61)=4hbd64-4-羟基丁酸coa转移酶(ec2.8.3.-)=abft66-乙烯乙酰coaδ异构酶(ec4.2.1.120)=abfd68-4-羟基丁酰coa脱水酶(ec5.3.3.3)=abfd70-乙酰乙酰辅酶a合酶硫解酶(ec2.3.1.9)=bktb72-2-氧代戊二酸/丁酸铁氧还蛋白氧化还原酶,也称为2-酮戊二酸氧化还原酶(ec1.2.7.3)=korab74-丙酮酸/2-氧代丁酸铁氧还蛋白氧化还原酶,也称为丙酮酸/酮异戊酸氧化还原酶(ec1.2.7.1)=pfor76-甲基丙二酸单酰coa变位酶(ec5.4.99.2)=scpa78-甲基丙二酸单酰coa脱羧酶(ec4.1.1.41)=scpb80-3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10)=mvas82-3-甲基戊烯二酸单酰coa水合酶(ec4.2.1.18)=liuc84-3-甲基巴豆酰coa羧化酶(ec6.4.1.4)=liubd86-反式-2-烯酰coa还原酶(ec1.1.1.36)=ter88-乙酰coa乙酰转移酶(ec2.3.1.9)=bktb90-3-羟基丁酰coa脱氢酶(ec1.1.1.157)=paah192-酰基coa硫酯酶(ec3.1.2.-)=mbach94-磷酸丁酰转移酶(ec2.3.1.19)=ptb96-丁酸激酶(ec2.7.2.7)=buk100-乳酸coa转移酶(ec2.8.3.-)=pct102-乳酰coa脱水酶(ec4.2.1.54)=lcdab104-丙二酸单酰coa还原酶(ec1.2.1.75)=mcr106-丙二酸单酰coa还原酶(ec1.1.1.298)=mcr108-丙酰coa合酶(ec6.2.1.36)=pcr110-丙酰coa合酶/丙烯酰coa合酶(ec4.2.1.116)=pcr,msed12112-丙酰coa合酶(ec1.3.1.84)=pcr132-阻抑蛋白(例如laci)134-操纵基因位点(例如lac操纵基因)136-转座酶识别位点(5’itr反向末端重复)138-抗生素抗性基因(mlsr)140-抗生素抗性蛋白(mlsr)142-含正选择特征的质粒144-含随机整合的dna盒的染色体pmb1=复制起点(大肠杆菌)reph=复制起点(梭菌属)catp=氯霉素/甲砜氯霉素抗性标记laci=lac阻抑蛋白tetr=四环素阻抑蛋白xylr=木糖阻抑蛋白himar=转座酶发明详述在公开和描述本微生物、组合物、核酸分子、载体、宿主细胞和/或方法之前,应理解它们除另有说明外不限于具体合成方法,或除另有说明外不限于具体试剂,因为这些天然可变。还应理解,本文所用的术语仅是为了描述具体方面的目的,并非旨在限制。虽然可以使用与本文中所述的那些相似或等同的任何方法和材料,但现在描述示例性方法和材料。本文中提供的微生物的使用容许获得链长为6个或更少碳原子的一种或多种烷烃和/或烯烃的新方法。该方法利用气态c1碳源,包括气态c1碳源和电子。本文公开的是从一氧化碳和/或二氧化碳产生一种或多种烷烃和/或烯烃的方法等。该方法基于使用一种或多种非天然存在的微生物。本文还公开该微生物。本文公开的微生物可以具有烯烃生物合成的能力。该微生物还可以具有烷烃生物合成的能力。在一些实施方案中,该微生物可以具有烷烃生物合成和烯烃生物合成的能力。本文公开的微生物是包含异源核酸序列的重组微生物。该异源核酸序列包含编码一种或多种酶的一种或多种序列。该一种或多种酶为该重组微生物提供了转化一氧化碳和/或二氧化碳以产生一种或多种烷烃和/或烯烃的能力。在一些实施方案中,按照本文公开的方法产生的烯烃可以用以下化学式表示:在此化学式中,r1可以是可包含一个双键的脂肪族碳链。在一些实施方案中,r1不包含双键。通常,r1不分支。在一些实施方案中,r1可以包含4个碳原子。在一些实施方案中,r1可以包含2或3个碳原子。在一些实施方案中,r1可以包含单个碳原子。r2可以是不包含双键的不分支脂肪族碳链。通常,r2可以包含1或2个碳原子。在一些实施方案中,r2是甲基。r2也可以是氢原子。通常,改造该微生物来产生一种或多种烷烃和/或烯烃。在一些实施方案中,改造该微生物来从一氧化碳和/或二氧化碳产生一种或多种烷烃和/或烯烃。本文公开的微生物可以在缺乏烯烃生物合成能力的天然存在的微生物基础上改造。在一些实施方案中,本文公开的微生物可以在缺乏从一氧化碳和/或二氧化碳生物合成烯烃的能力的天然存在的微生物基础上改造。本文公开的微生物还可以在缺乏烷烃生物合成能力的天然存在的微生物基础上改造。在一些实施方案中,本文公开的微生物可以在缺乏从一氧化碳和/或二氧化碳生物合成烷烃的能力的天然存在的微生物基础上改造。本文公开的微生物可以在缺乏烷烃生物合成和烯烃生物合成两种能力的天然存在的微生物基础上改造。在一些实施方案中,本文公开的微生物可以在缺乏从一氧化碳和/或二氧化碳生物合成烷烃和生物合成烯烃两种能力的天然存在的微生物基础上改造。还公开制备并用这类微生物来从一氧化碳和/或二氧化碳产生一种或多种烷烃和/或烯烃的方法,包括设计和产生具有烷烃和/或烯烃生物合成能力的微生物。定义除非另有说明,本文件(包括说明书和权利要求书)中所用的以下术语具有下文给出的定义。除非文中清楚地另有说明,说明书和所附权利要求书中所用的单数形式“一”、“一个”和“该”包括复数指代物。本文中所用的词语“或”意指具体列表的任一成员,且还包括该列表的成员的任何组合。本文中所用的词语“约”指值处于本领域普通技术人员确定的具体值的可接受误差范围内,该可接受误差范围将部分取决于如何测量或测定该值,即测量系统的限制。例如,按照本领域的实践,“约”可以指1或1个以上标准差之内。术语“约”还用来表示所讨论的量或值可以是指定值或大致相同的某个其他值。该短语旨在传达,相似的值促成与所述等同的结果或效应。在此背景中,“约”可以指高和/或低至多10%的范围。在一些实施方案中,词语“约”指高于和低于某个值至多5%的范围,如高于或低于该值至多2%、至多1%或至多0.5%。在一个实施方案中,“约”指高于和低于给定值至多0.1%的范围。术语“等位基因变体”在本文中指占据同一染色体基因座的基因的两种或多种备选形式中的任一种。等位基因变异通过突变天然出现,且可以在群体内产生多态性。基因突变可以是沉默突变(所编码的多肽中无改变)或可以编码氨基酸序列改变的多肽。多肽的等位基因变体是由基因的等位基因变体编码的多肽。除非另有说明,术语“烷烃”指饱和脂肪族分子。各脂肪族分子具有直的或分支的烃链,其可以包含杂原子。在本文公开的方法中,烷烃通常不包含杂原子。烃链的分支可以包含线性链。通常,在本文所述的方法中形成的烷烃不包含环元件。除非另有说明,烃链可以为任意长度,包含任意数目的分支。分支可以定义为烷基,如甲基、乙基或丙基。通常,在本文所述的方法中形成的烷烃的烃(主)链包含1至8个,如2至6个碳原子。除非另有说明,术语“烯烃”指不饱和脂肪族分子,其包含一个或多个双键。各脂肪族分子具有直的或分支的烃链,其可以是单不饱和或多不饱和,且可以包含杂原子。在本文公开的方法中,烯烃通常不包含杂原子。上文已经提到在烃链中存在分支。通常,在本文所述的方法中形成的烯烃不包含环元件。通常,在本文所述的方法中形成的烯烃的烃(主)链包含1至8个,如2至6个碳原子。“厌氧菌”是生长不需要氧气的生物。厌氧菌可以是专性厌氧菌、兼性厌氧菌或耐氧生物。“专性厌氧菌”是大气水平的氧气可以致死的厌氧菌。专性厌氧菌的实例包括但不限于梭菌属、eurobacterium、类杆菌属(bacteroides)、消化链球菌属(peptostreptococcus)、丁酸杆菌属(butyribacterium)、韦荣菌属(veillonella)和放线菌属(actinomyces)。“兼性厌氧菌”是能够在氧气存在下进行有氧呼吸且能够在氧气限制或无氧气条件下进行厌氧发酵的厌氧菌。兼性厌氧菌的实例包括但不限于埃希氏菌属(escherichia)、泛菌属(pantoea)和链霉菌属(streptomyces)。术语“一氧化碳营养生物(carboxidotrophicorganism)”和“一氧化碳营养(carboxidotroph)”指可以耐受高浓度一氧化碳且通常能够利用一氧化碳进行代谢的生物。一氧化碳营养微生物还可从co的氧化获得能量和碳。这种一氧化碳营养微生物可以是能够将co氧化为二氧化碳的需氧细菌。一氧化碳营养微生物还可以是专性厌氧菌,其可以能够将二氧化碳还原为co和/或将co转化为二氧化碳和氢。一些作为专性厌氧菌的一氧化碳营养微生物是产乙酸菌(例如热醋穆尔氏菌),可以从co和/或co2形成乙酸。在本文中使用时,术语“编码序列”指核苷酸序列,其直接指定其蛋白质产物的氨基酸序列。编码序列的边界通常由可读框决定,可读框通常始于atg起始密码子或备选起始密码子如gtg和ttg,结束于终止密码子如taa、tag和tga。编码序列可以是dna、cdna、合成或重组核苷酸序列。本文所用的术语“保守修饰”和“保守取代”分别指修饰和取代,就对应的参考而言,其在物理、生物学、化学或功能上维持特性。包含具有保守取代的序列的分子例如具有相似的大小、形状、电荷、化学特性,包括形成共价键或氢键的相当的能力,和/或相当的极性。这类保守修饰包括但不限于一个或多个核碱基和氨基酸的取代、添加和缺失。例如,保守氨基酸取代包括其中用具有与某氨基酸残基侧链相似侧链的氨基酸残基替换该氨基酸残基的那些。例如,可以用来自同一侧链家族的另一氨基酸残基替换就与抗原的结合而言非必需的氨基酸残基,例如可以用苏氨酸取代丝氨酸。氨基酸残基通常根据共同、相似的侧链特性分为家族,如:1.非极性侧链(例如甘氨酸、丙氨酸、缬氨酸、亮氨酸、异亮氨酸、甲硫氨酸);2.不带电荷的极性侧链(例如天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸、酪氨酸、脯氨酸、半胱氨酸、色氨酸);3.碱性侧链(例如赖氨酸、精氨酸、组氨酸);4.酸性侧链(例如天冬氨酸、谷氨酸);5.β-分支侧链(例如苏氨酸、缬氨酸、异亮氨酸);和6.芳香族侧链(例如酪氨酸、苯丙氨酸、色氨酸、组氨酸)。此分类可以进一步划分。作为另一个取向,以下八个组包含通常可用来定义彼此的保守取代的氨基酸:1)丙氨酸(ala)、甘氨酸(gly);2)天冬氨酸(asp)、谷氨酸(glu);3)天冬酰胺(asn)、谷氨酰胺(gln);4)精氨酸(arg)、赖氨酸(lys);5)异亮氨酸(ile)、亮氨酸(leu)、甲硫氨酸(met)、缬氨酸(val);6)苯丙氨酸(phe)、酪氨酸(tyr)、色氨酸(trp);7)丝氨酸(ser)、苏氨酸(thr);和8)半胱氨酸(cys)、甲硫氨酸(met)。保守取代可以认为是用以上六个组之一内的另一氨基酸取代该六个组的同一组内的第一氨基酸。保守取代通常是以下取代,按照待突变的氨基酸后各跟随可以认为是保守取代的一个或多个替换列出:ala→gly、ser、val;arg→lys;asn→gln、his;asp→glu;cys→ser;gln→asn;glu→asp;gly→ala;his→arg、asn、gln;ile→leu、val;leu→ile、val;lys→arg、gln、glu;met→leu、tyr、ile;phe→met、leu、tyr;ser→thr;thr→ser;trp→tyr;tyr→trp、phe;val→ile、leu。其他取代也可容许,且可以根据经验或按照其他已知的保守或非保守取代来确定。保守取代还可以涉及非天然氨基酸的使用。例如就电荷、偶极矩、大小、亲水性、疏水性或结合分子的构象而言,非保守取代(即一个家族的成员与另一家族的成员交换)可以导致实质性改变,其可以导致结合活性显著下降,尤其是如果受影响的氨基酸是与靶分子的结合所必需的。非保守取代也可以涉及非天然氨基酸的使用。保守和非保守修饰可以通过本领域已知的标准技术引入亲本结合分子,如组合化学、定位dna诱变、pcr介导和/或盒诱变、肽/蛋白质化学合成、特异性修饰亲本结合分子中的反应性基团的化学反应。变体可以通过常规方法针对其化学、生物学、生物物理和/或生物化学特性进行测试。优选地,保守氨基酸取代不实质性改变亲本序列的功能特征,通常也不实质性改变亲本序列的结构特征。因此,包含保守取代的结合分子的结合特征至少基本不变。此外,保守氨基酸取代通常不实质性改变或破坏亲本序列的二级结构。本文所用的术语“内源的”指所提到的分子天然存在于宿主中。因此,本文在微生物的背景中所用的“内源”酶是从天然是相同微生物的部分的核酸序列(如该物种的基因)产生或合成的酶。本文所用的术语“外源的”旨在意指将或已将所提到的分子、序列或活性引入宿主微生物。因此,外源核酸分子或序列指并非源自见于自然界中的该具体生物的任何核酸分子或序列。因此,在引入该生物时可认为非天然存在的核酸分子或序列对细胞而言是外源的。该分子或序列可以例如通过将编码核酸分子引入宿主遗传物质来引入,如通过整合入宿主染色体或作为非染色体遗传物质(如质粒)。因此,在用于提及编码核酸的表达时,该术语指将编码核酸以可表达的形式引入微生物。在用于提及生物合成活性时,该术语指引入宿主参考生物的活性。来源可以是例如引入宿主微生物后表达所提到的活性的同源或异源编码核酸。重要的是要指出,非天然存在的核酸分子或序列可以包含见于自然界中的核酸序列或核酸序列的片段,只要该核酸作为整体不存在于自然界中。例如,在表达载体内包含基因组dna序列的核酸分子是非天然存在的核酸分子,因此在引入细胞时对该细胞而言是外源的,因为该核酸分子作为整体(基因组dna加上载体dna)不存在于自然界中。因此,可认为作为整体不存在于自然界中的任何载体、自主复制质粒或病毒(例如反转录病毒、腺病毒或疱疹病毒)都是非天然存在的核酸。由此得出结论,认为通过pcr或限制性核酸内切酶处理产生的基因组dna片段以及cdna也是非天然存在的核酸,因为它们以不见于自然界中的分开的分子存在。由此还得出结论,以不见于自然界中的排列方式包含启动子序列和多肽编码序列(例如cdna或基因组dna)的任何核酸也是非天然存在的核酸。天然存在的核酸对具体生物而言可以是外源的。例如,在将染色体引入生物的细胞时,从x生物的细胞分离的整个染色体对y生物的细胞而言是外源核酸。提到蛋白质时的术语“表达”定义了将包含在核酸分子中且编码肽/蛋白质的序列转化为其肽/蛋白质产物。在核酸是dna时,细胞经以下表达蛋白质(如酶):将核酸转录为mrna,然后翻译为多肽,该多肽进行折叠并可能进一步加工。在核酸是rna时,表达可以包括将此rna复制为其他rna拷贝和/或将rna反转录为dna并可选地将此dna转录为其他rna分子。在任何一种情况下,rna的表达包括将所提供/产生的任意rna种类翻译为蛋白质。因此,细胞正在表达蛋白质的陈述意指已通过该细胞的表达机器合成该蛋白质。就该生物学过程本身而言,术语“表达”、“基因表达”指将基因的核酸序列中所编码的信息先转化为信使rna(mrna),然后转化为蛋白质的调节途径的整体。因此,基因的表达包括其转录为初级hnrna,此hnrna加工为成熟rna,及mrna序列翻译为相应的蛋白质的氨基酸序列。在此背景中,还应指出,术语“基因产物”不仅指蛋白质,包括例如该基因所编码的最终蛋白质(包括其剪接变体)和各前体蛋白质(适用时),还指各mrna,其可认为是基因表达过程期间的“第一基因产物”。蛋白质或肽的表达可以用体外表达系统进行。这种表达系统可以包括细胞提取物,通常来自细菌、兔网织红细胞或麦胚。许多适宜的系统可购得。所使用的氨基酸混合物可以根据需要包含合成氨基酸,以增加文库中产生的蛋白质可能的数目或多样性。这可以通过用人工氨基酸负载trna并用这些trna进行待选则的蛋白质的体外翻译来达到。用于表达目的的适宜实施方案是使用载体,尤其是表达载体。本文所用的术语“表达盒”指能够在适当的宿主细胞中直接表达具体核酸序列的核酸分子。所有表达盒都包含与目的核苷酸序列有效连接的启动子,目的核苷酸序列与一个或多个终止信号有效连接。它还可以包含正确翻译核苷酸序列所需的序列。编码区可以编码目的多肽,且还可以编码目的功能性rna,包括但不限于有义或反义方向的反义rna或非翻译rna。包含目的核苷酸序列的表达盒可以是嵌合的(意指其成分中的至少一种对其其他成分中的至少一种而言是异源的)。表达盒还可以是这样的表达盒,其天然存在,但以用于异源表达的重组形式获得。但是,在一些实施方案中,表达盒对宿主而言是异源的,即表达盒的具体核酸序列并非天然存在于宿主细胞中,而是通过转化事件引入宿主细胞或宿主细胞的祖先。表达盒中核苷酸序列的表达可以处在组成型启动子或仅在宿主细胞暴露于某种具体外部刺激时才起始转录的诱导型启动子的控制之下。在多细胞生物(如植物或动物)的情况下,该启动子也可以为具体组织、器官或发育阶段特异。除非另有说明,在本文中使用时,短语“发酵”、“发酵方法”和“发酵反应”指化合物的酶促转化。该化合物通常是有机化合物,包括气态碳化合物,如一氧化碳和二氧化碳。反应物酶促转化为产物化合物通常涉及包含和/或产生酶促转化所需的酶的细胞。发酵方法包括细胞培养,不考虑其阶段、密度和/或生长条件。例如,旨在涵盖该方法的生长阶段和产物生物合成阶段二者。在这方面,在一些实施方案中,生物反应器可以包括第一生长反应器和第二发酵反应器。因此,向发酵反应加入金属或组合物应理解为包括向这些反应器中的任一个或二者中加入。本文所用的“发酵液”是包含营养培养基和细菌细胞的培养基。“基因”意指遗传单位,其占据染色体上的具体基因座,是与生物学功能相关的核酸区段。基因涵盖转录和/或翻译调节序列,以及编码区。除编码序列外,基因可以包括启动子区、顺式调节序列、作为调节蛋白特异性识别序列的非表达dna区段、促成基因表达的非表达dna区段、设计为具有所希望得到的参数的dna区段或其组合。基因可以通过多种方法获得,包括从生物样品克隆、根据已知或预测的序列信息合成及已有序列的重组衍生。本文在微生物的背景中所用的“同源”酶是从天然是同一物种的微生物部分的核酸序列产生或合成的酶。因此,同源酶可以是这样的酶,其与微生物的基因编码的酶相同,但它可以是通过基因工程的方式引入微生物的。在这种情况下,该酶可以与该微生物自身的该酶没有区别。一些酶在同一物种内以不同同种型的形式存在。因此,在这种情况下,同一物种的一些成员具有第一同种型,而该物种的其他成员具有第二同种型。同源酶可以是任一同种型。换言之,同源酶可以是酶的同种型,其不同于具体微生物的基因编码的酶,但它可以是见于同一物种的一些其他成员中的酶的同种型。本文所用的术语“同源序列”指两个聚合物分子间(例如两个核酸分子间,例如两个dna分子或两个rna分子间,或两个多肽分子间)的序列相似性。如果给定序列的tfasty搜索针对另一序列得到低于0.001的期望分值(e值),则可以称两个序列同源。本文所用的术语“异源的”指源自所提到的物种之外的来源的分子或活性,而“同源的”指源自宿主微生物的分子或活性。因此,本文所述编码核酸的外源表达可以利用异源或同源编码核酸中的任一种或二者。本文在微生物背景中所用的“异源”酶是从天然存在于不同于该微生物所属物种的物种中的核酸序列产生或合成的酶。氨基酸序列或核酸序列背景中的术语“同一性”或“序列同一性”指两个蛋白质或核酸间的序列匹配。因此,该术语定义了衡量其相似性或关系的序列特性。比对所要比较的蛋白质或核酸序列以给出最大同一性。用于比对序列进行比较的方法为本领域公知。示例性方法是ncbibasiclocalalignmentsearchtool(blasttm;altschul等,j.mol.biol.(1990)215,403-410),其可从若干来源获得,包括nationalcenterforbiotechnologyinformation(ncbi,bethesda,md,u.s.a.),其可以与序列分析程序blastp、blastn、blastx、tblastn和tblastx结合使用。blasttm可以在互联网上在ncbi网站登陆。另一实例是工具embossneedle(配对比对;可在www.ebi.ac.uk处获得)。在要比较的序列中的相同位置由相同核碱基或氨基酸残基占据时,该分子在该位置处相同。“序列同一性”或“同一性”可用于定义,在具体多肽的序列与参考序列的(同源性)比对之后,就这两个序列中较长的序列中的残基数目而言,配对的相同残基的百分比。通过用相同残基的数目除以残基的总数并将结果乘以100来测量同一性。例如,如果10个序列位置中由6个相同,则同一性为60%。例如,可以使用blosum62矩阵、“缺口开放罚分”为10、“缺口延伸罚分”为0.5、“末端缺口罚分”关闭、“末端缺口开放罚分”为10和“末端缺口延伸”为0.5,用已并入embossneedle的needleman和wunsch算法(needleman,s.b.andwunsch,c.d.,jmb(1970,)48,443-453)测定两个蛋白质序列间的百分比同一性。不考虑任何化学和/或生物学修饰,具有相同的一级氨基酸或核酸序列的两个分子是相同的。例如,按照此定义,具有相同的一级氨基酸序列但不同的糖基化模式的两种酶是相同的。在核酸的情况下,按照此定义,例如具有相同的序列但不同的连接成分(如硫代磷酸而不是磷酸)的两个分子是相同的。序列同源性或序列同一性的百分比可以例如用blastp程序(例如版本blastp2.2.5)(2002年11月16日;参见altschul,s.f.等(1997)nucl.acidsres.25,3389-3402))测定。在此实施方案中,在配对比较中用已知的天然蛋白质作为参考,百分比同源性可以基于包括前肽序列的整个多肽序列的比对(矩阵:blosum62;缺口代价:11.1;截止值设为10-3)。将它计算为在blastp程序输出中作为结果显示的“正值”(同源氨基酸)的数目除以该程序选择进行比对的氨基酸总数的百分比。关于这一点应指出,所选择的氨基酸的此总数可以不同于已知的天然蛋白质的长度。提到核酸或多肽时的术语“突变的”或“突变体”分别指与天然存在的核酸分子或多肽相比交换、缺失或插入了一个或多个核苷酸或氨基酸。与相应的天然氨基酸序列或核酸序列相比,该突变体可以例如包含一个或多个取代。本文所用的术语“诱变”意指这样选择实验条件,使得天然存在于所选择的蛋白质的给定序列位置处的氨基酸可被不存在于该天然多肽序列中的此具体位置处的至少一个氨基酸取代。如上文已指出,术语“诱变”还包括通过缺失或插入一个或多个氨基酸来进行序列区段长度的(附加)修饰。因此,例如,可以用三个随机突变的一段序列替换所选择的序列位置处的一个氨基酸,导致与野生型蛋白质的该区段的长度相比插入了两个氨基酸残基。这种插入或缺失可以在可进行诱变的任何肽区段中相互独立地引入。术语“随机诱变”意指没有预先确定的单个氨基酸(突变)存在于某个序列位置处,但可以在诱变期间在预定的序列位置处以某种概率掺入至少两个氨基酸。可以用所选择的编码序列作为诱变的起点。对于所选择的氨基酸位置的诱变,本领域技术人员有多种用于定位诱变的标准方法可供使用。常用的技术是使用在所希望的序列位置处具有简并碱基组成的合成寡核苷酸的混合物,借助pcr(聚合酶链反应)引入突变。例如,密码子nnk或nns(其中n=腺嘌呤、鸟嘌呤或胞嘧啶或胸腺嘧啶;k=鸟嘌呤或胸腺嘧啶;s=腺嘌呤或胞嘧啶)的使用允许在诱变期间掺入全部20种氨基酸加琥珀终止密码子,而密码子vvs使可能掺入的氨基酸的数目限于12种,因为它排除了在所选择的多肽序列位置处掺入氨基酸cys、ile、leu、met、phe、trp、tyr、val;密码子nms(其中m=腺嘌呤或胞嘧啶)的使用例如使所选择的序列位置处可能的氨基酸数目限于11种,因为它排除了在所选择的序列位置处掺入氨基酸arg、cys、gly、ile、leu、met、phe、trp、val。在这方面,应指出,用于不同于常规的20种天然存在的氨基酸的氨基酸(硒代半胱氨酸或吡咯赖氨酸)的密码子也可以掺入核酸。还可能按wang,l.等science(2001)292,498-500或wang,l.和schultz,p.g.chem.comm.(2002)1,1-11所述使用通常识别为终止密码子的“人工”密码子,如uag,以插入其他不常见氨基酸,例如邻-甲基-l酪氨酸或对-氨基苯丙氨酸。使用碱基对特异性降低的核苷酸构件,例如肌苷、8-氧代-2’脱氧鸟苷或6(2-脱氧-β-d-呋喃核糖基)-3,4-二氢-8h-嘧啶-1,2-嗪-7-酮(zaccolo等j.mol.biol.(1996)255,589-603)是在所选择的序列区段中引入突变的另一种选择。另一种可能性是所谓的三联体诱变。此方法用各编码一种氨基酸的不同核苷酸三联体的混合物来掺入编码序列(等,nucleicacidsres(1994)22,5600-5607)。本文中在微生物的背景中使用时,术语“非天然存在的”旨在意指微生物具有通常不见于所提到的物种的天然存在的菌株(包括所提到的物种的野生型菌株)中的一种或多种遗传改变。遗传改变包括例如引入可表达的编码代谢多肽尤其是一种或多种酶的核酸序列。该可表达的核酸序列可以包含在微生物的基因组中,或者它可以包含在不同于微生物基因组的核酸分子中。遗传改变还可以包括核酸添加、核酸缺失和/或微生物遗传物质的其他功能性破坏。对于所提到的物种的异源多肽、同源多肽或异源和同源多肽二者,这类修饰包括例如编码区及其功能片段。其他修饰包括例如非编码调节区,其中修饰改变基因或操纵子的表达。示例性代谢多肽包括烯烃生物合成途径中的酶或蛋白质。代谢修饰指从其天然存在的状态改变的生物化学反应。因此,非天然存在的微生物可以具有对编码代谢多肽或其功能片段的核酸进行的修饰。本文所用的术语“核酸分子”指处于任意可能的构型(如单链、双链或其组合)的任意核酸。核酸的实例包括例如dna分子、rna分子、用核苷酸类似物或用核酸化学产生的dna或rna类似物、锁定核酸分子(lna)、蛋白质核酸分子(pna)、烷基磷酸酯和烷基磷酸三酯核酸分子及tecto-rna分子(例如liu,b.等,j.am.chem.soc.(2004)126,4076-4077)。lna具有在c4'和o2'间具有亚甲基桥的修饰rna主链,为该分子提供更高的双链体稳定性和核酸酶抗性。烷基链酸酯和烷基磷酸三酯核酸分子可以视为dna或rna分子,其中通过将核酸主链中磷酸基团的p-oh基团分别换为烷基和烷氧基来中和核酸主链中的磷酸基团。dna或rna可以具有基因组来源或合成来源,可以是单链或双链。这种核酸可以是例如mrna、crna、合成rna、基因组dna、cdna、合成dna、dna和rna的共聚物、寡核苷酸等。该核酸可以进一步包含非天然核苷酸类似物和/或与亲和力标记或标签连接。许多核苷酸类似物是已知的,且可以用于本文所述的方法中所用的核酸。核苷酸类似物是在例如碱基、糖或磷酸部分上含有修饰的核苷酸。作为说明性实例,已知用2'f、2'o-me或2'h残基取代sirna的2'-oh残基改善该rna的体内稳定性。碱基部分上的修饰可以是a、c、g和t/u的天然或合成修饰,不同的嘌呤或嘧啶碱基,如尿嘧啶-5-基、次黄嘌呤-9-基和2-氨基腺嘌呤-9-基,以及非嘌呤或非嘧啶核苷酸碱基。其他核苷酸类似物作为通用碱基。通用碱基的实例包括3-硝基吡咯和5-硝基吲哚。通用碱基能够与任何其他碱基形成碱基对。碱基修饰常可与例如糖修饰组合,例如2'-o-甲氧乙基,例如以达到独特特性,如提高的双链体稳定性。本文所用的术语“核酸构建体”指单链或双链的核酸分子,其分离自天然存在的基因,或者其修饰为以否则不存在于自然界中的方式包含核酸的区段,或者其是合成的。在核酸构建体包含表达具体编码序列所需的控制序列时,术语核酸构建体与术语“表达盒”是同义词。术语“与…有效连接”指核酸序列与另一核酸序列的功能关系。启动子、增强子、转录和翻译终止位点及其他信号序列是与其他序列有效连接的核酸序列的实例。例如,dna与转录控制元件的有效连接指dna和启动子间的物理和功能关系是这样,使得通过特异性识别、结合和转录dna的rna聚合酶从该启动子起始这种dna的转录。本文所用的术语“多肽”和“蛋白质”指氨基酸残基的聚合物,且不限于某个最小产物长度。在同时使用两个术语时,这种双重命名导致在本领域中一起使用这两个术语。本文件通篇使用的术语“启动子”指基因序列表达所需的核酸序列。启动子区在不同生物间不同,但对不同生物而言为本领域技术人员公知。例如,在原核生物中,启动子区包含启动子(其指导起始rna转录)以及在转录为rna时将发出合成起始信号的dna序列二者。这类区域通常将包含涉及转录和翻译的起始的那些5'非编码序列,如tata框、帽化序列、caat序列等。在本文公开的方法的背景中,组成型和诱导型启动子都可以使用。多种潜在的宿主细胞识别的大量启动子是公知的。通过经限制酶从来源dna去除启动子并将分离的启动子序列插入所选择的载体,可以将所选择的启动子有效连接至编码本文所述多肽的顺反子dna。天然启动子序列和许多异源启动子都可以用于指导所选择的核酸序列的扩增和/或表达。本文中用词语“重组”来描述由于其来源、操作或来源和操作二者而与自然界中与之相关的全部或部分核酸分子不相关的核酸分子。通常,重组核酸分子包含并非天然存在于该野生型生物或细胞中的序列。通常,通过基因工程获得重组核酸分子,通常在细胞外构建。通常,重组核酸分子与自然界中存在的相应核酸分子的至少一部分基本相同和/或基本互补。重组核酸分子可以具有任何来源,如基因组、cdna、哺乳动物、细菌、病毒、半合成或合成来源。就蛋白质/多肽使用的术语“重组”意指通过表达重组多核苷酸而产生的多肽。“相似的”蛋白质序列是这样的蛋白质序列,在比对时,其在待比较的序列的相同位置上具有相似的氨基酸残基,且最常是但并非强制是相同的氨基酸残基。相似的氨基酸残基按侧链化学特征分为家族。这些家族在下文中针对“保守氨基酸取代”进行描述。序列间的“百分比相似性”是在待比较的序列的相同序列位置上包含相同或相似的残基的位置数除以所比较的位置总数,并乘以100%。例如,如果10个序列位置中的6个具有相同的氨基酸残基,且10个位置中的2个包含相似的残基,则序列具有80%相似性。两个序列间的相似性可以例如用embossneedle确定。术语“严格条件”指杂交核酸分子的技术。核酸分子杂交反应可以在具有不同严格性的条件下进行。“严格条件”在本领域中广为人知和公开。通常,杂交反应期间,可以使用基于ssc的缓冲液,其中ssc是ph为7.0的0.15mnacl和15mm柠檬酸缓冲液。提高缓冲液浓度和存在变性剂可提高杂交步骤的严格性。例如,高严格性杂交条件可以涉及:(i)在42℃使用50%(v/v)甲酰胺、5xssc(0.75mnacl、0.075m柠檬酸钠)、50mm磷酸钠(ph6.8)、0.1%焦磷酸钠、5xdenhardt's溶液、超声处理的鲑精dna(50μg/ml)、0.1%sds和10%硫酸葡聚糖,在42℃在0.2xssc和0.1%sds中洗涤;(ii)在42℃使用含0.1%牛血清白蛋白的50%(v/v)甲酰胺/0.1%ficoll/0.1%聚乙烯吡咯烷酮/含750mm氯化钠、75mm柠檬酸钠的50mm磷酸钠缓冲液ph6.5;或(iii)在55℃使用10%硫酸葡聚糖、2xssc和50%甲酰胺,然后在55℃进行包含含有edta的0.1xssc高严格性洗涤。此外或备选地,可以在杂交流程中包括一个、两个或多个使用低离子强度洗涤溶液和高温的洗涤步骤,例如在50℃使用0.015m氯化钠/0.0015m柠檬酸钠/0.1%十二烷基硫酸钠。术语“合成气”指一氧化碳、二氧化碳和氢气的混合物。本文所用的术语“转座子”或“可转座元件”意在指能够从供体多核苷酸(例如载体)切下或拷贝并整合入靶位点(例如细胞的基因组或染色体dna)的核苷酸序列。从一个核酸分子切下或拷贝确定的dna区段并将该区段移动至同一核酸分子或不同核酸分子中的另一位点的过程称为转座。转座子可以是“2类”元件,也称为“dna转座子”。2类元件从供体多核苷酸切下并插入靶核酸分子。此过程由功能性转座酶蛋白质催化。在一些情况下,转座酶蛋白质由转座子编码。这种转座子也称为自主转座子。转座子包含侧翼为转座子末端的顺式作用核苷酸序列的核酸序列。如果至少一个顺式作用核苷酸序列位于核酸序列的5'且至少一个顺式作用核苷酸序列位于核酸序列的3',则该核酸序列“侧翼”为该顺式作用核苷酸序列。对于2类元件,顺式作用核苷酸序列在转座子的每个末端包含至少一个反向重复(本文中也称为反向末端重复,或itr),转座酶与该反向重复结合。转座子也可以是“1类”元件,也称为“反转录转座子”。1类元件不编码转座酶;相反,它产生rna转录物,然后依赖反转录酶将该rna序列反转录为dna,然后该dna插入靶位点。1类元件拷贝自供体多核苷酸,即它离开原拷贝,并产生插入靶位点的第二拷贝。取决于它们是否包含编码反转录转座所需的蛋白质的序列,1类元件分为自主类和非自主类。转座子从供体多核苷酸切下或拷贝并整合入靶位点的能力可以例如通过甲基化和/或小干扰rna分子来沉默。“变体”指这样的氨基酸或核酸序列,其由于添加(包括插入)、缺失和/或取代了一个或多个氨基酸残基或核碱基而不同于亲本序列,但保留本文公开的亲本序列的至少一种希望得到的活性。在酶的情况下,这种希望得到的活性可以包括特异性底物结合。类似地,与亲本序列相比,变体核酸序列可以由于添加、缺失和/或取代了一个或多个核碱基而改变,但所编码的蛋白质(例如酶)保留上文所述的希望得到的活性。变体可以天然存在,如等位基因变体或剪接变体,或者可以人工构建。术语“载体”有时也称为基因递送系统或基因转移载体,指包含待在体外、离体或在体内递送至宿主细胞的多核苷酸的大分子或分子复合体。通常,载体是允许或便于将核酸序列转移入细胞的单链或双链环状核酸分子。载体通常可以转入细胞,并在细胞基因组内或独立于细胞基因组复制。环状双链核酸分子可以通过限制酶处理来切割,从而线性化。各种核酸载体、限制酶及限制酶切割的核苷酸序列的知识对本领域技术人员易于获得。通过用限制酶切割载体并将两个部分连接在一起,可以将编码肽的核酸分子(如编码genbank检索号wp_003251320.1或其同源物的氨基酸序列的序列,和/或编码genbank检索号wp_003243190.1或其同源物的氨基酸序列的序列)插入载体。载体可以是例如病毒载体,如反转录病毒载体、慢病毒载体、基于疱疹病毒的载体或腺病毒载体。载体也可以是质粒载体,其也是原核载体的典型实例。在一些实施方案中,该载体可以是能够在大肠杆菌中复制的质粒,例如pbr322、colel、psc101、pacyc184或πvx。芽孢杆菌属(bacillus)质粒包括pc194、pc221或pt127。适宜的链霉菌属质粒包括1θ101和链霉菌属噬菌体如φc31。载体也可以是基于脂质体的染色体外载体,也称为附加型载体。附加型载体的两个说明性实例是基于orip的载体和编码ebna-1衍生物的载体。淋巴细胞营养(lymphoblast)疱疹病毒是疱疹病毒,其在淋巴母细胞中复制,并在其自然生命周期的一部分中称为质粒。载体也可以基于有机修饰的硅酸盐。在一些实施方案中,载体可以是上文所述的基于转座子的系统。术语“包含”、“包括”、“含有”、“具有”等应扩展性或开放性和无限制性地理解。除非文中清楚地另有说明,诸如“一”、“一个”或“该”的单数形式包括复数指代物。因此,例如,提到一个“载体”包括单个载体,以及多个相同(例如同一操纵子)或不同的载体。同样,提到“细胞”包括单个细胞以及多个细胞。除非另有说明,一系列元件之前的术语“至少”理解为指该系列中的每个元件。术语“至少一个”和“…中的至少一个”包括例如一个、两个、三个、四个、或五个或更多个元件。还应理解,高于或低于所陈述的范围的轻微变异可以用来达到与该范围内的值基本相同的结果。另外,除非另有说明,范围的公开旨在作为包括最小值和最大值之间的每个值的连续范围。任何用法的术语的范围和含义将从使用该术语的具体文字显而易见。根据具体情况,本文通篇使用的所选择的术语的某些其他定义在发明详述的适当文字中给出。除非另有说明,说明书、附图和权利要求书中使用的所有其他科学和技术术语具有如本领域普通技术人员通常理解的其普通含义。本公开的多种方面在以下章节中进一步详述。应理解,多种实施方案、偏好和范围可以随意组合。另外,取决于具体实施方案,所选择的定义、实施方案或范围可以不适用。酶和途径本文所述的重组微生物通常包含提供、完善或增强生物合成途径的一种或多种异源核酸序列。本文所述的重组微生物通常能够表达许多酶,这些酶一起确定导致形成烷烃和/或烯烃的生物合成途径。该生物合成途径的一种或多种酶由异源核酸序列编码。通常,生物合成途径的酶以足以允许产生烷烃和/或烯烃的量表达。由此,该重组微生物可以能够将一氧化碳和/或二氧化碳转化为烷烃和/或烯烃。本文所述的重组微生物可以包含编码导致形成甲烷的途径的酶的核酸序列。在一些实施方案中,本文所述的重组微生物可以包含编码导致形成乙烷的途径的酶的核酸序列本文所述的重组微生物可以包含编码导致形成乙烯的途径的酶的核酸序列。这些酶中的一种或多种可以由异源核酸序列编码。该异源核酸序列可以源自异源核酸分子。在一些实施方案中,该重组微生物可以包含编码导致形成乙烷的途径的所有酶的核酸序列。在一些实施方案中,该重组微生物不包含编码导致形成乙烷的途径的所有酶的核酸序列。在一些实施方案中,本文所述的重组微生物可以能够产生乙烯。在一些实施方案中,本文所述的重组微生物不能产生乙烷。本文公开的重组微生物可以包含编码导致形成丙烷的途径的酶的核酸序列。这些酶中的一种或多种可以由异源核酸序列编码。在一些实施方案中,该重组微生物可以包含编码导致形成丙烷的途径的所有酶的核酸序列。在一些实施方案中,该重组微生物不包含编码导致形成丙烷的途径的所有酶的核酸序列。在一些实施方案中,本文所述的重组微生物能够产生丙烷。本文所述的重组微生物可以包含编码导致形成丙烯的途径的酶的核酸序列。这些酶中的一种或多种可以由异源核酸序列编码。在一些实施方案中,这些酶中的两种或三种可以由异源核酸序列编码。如上文所指出,该异源核酸序列可以源自异源核酸分子。在一些实施方案中,该重组微生物可以包含编码导致形成丙烯的途径的所有酶的核酸序列。在一些实施方案中,该重组微生物不包含编码导致形成丙烯的途径的所有酶的核酸序列。在一些实施方案中,本文所述的重组微生物可以能够产生丙烯。在一些实施方案中,本文所述的重组微生物不能产生丙烯。本文公开的重组微生物可以包含编码导致形成正丁烷的途径的酶的核酸序列。这些酶中的一种或多种可以由异源核酸序列编码。在一些实施方案中,该重组微生物可以包含编码导致形成正丁烷的途径的所有酶的核酸序列。在一些实施方案中,该重组微生物不包含编码导致形成正丁烷的途径的所有酶的核酸序列。在一些实施方案中,本文所述的重组微生物能够产生丙烷。本文所述的重组微生物可以包含编码导致形成1-丁烯的途径的酶的核酸序列。这些酶中的一种或多种可以由异源核酸序列编码。在一些实施方案中,此途径的酶中的两种或三种可以由异源核酸序列编码。如上文所指出,该异源核酸序列可以源自异源核酸分子。在一些实施方案中,该重组微生物可以包含编码导致形成1-丁烯的途径的所有酶的核酸序列。在一些实施方案中,该重组微生物不包含编码导致形成1-丁烯的途径的所有酶的核酸序列。在一些实施方案中,本文所述的重组微生物可以能够产生1-丁烯。在一些实施方案中,本文所述的重组微生物不能产生1-丁烯。在一些实施方案中,本文所述的重组微生物可以包含编码导致形成异丁烯的途径的酶的核酸序列。此途径的酶中的一种或多种可以由异源核酸序列编码。在一些实施方案中,导致形成异丁烯的途径的酶中的至少两种或三种可以由异源核酸序列编码。该异源核酸序列可以源自异源核酸分子。在一些实施方案中,该重组微生物可以包含编码导致形成异丁烯的途径的所有酶的核酸序列。在一些实施方案中,该重组微生物不包含编码导致形成异丁烯的途径的所有酶的核酸序列。在一些实施方案中,本文所述的重组微生物可以能够产生异丁烯。在一些实施方案中,本文所述的重组微生物不能产生异丁烯。本文公开的重组微生物也可以包含编码导致形成1,3-丁二烯的途径的酶的核酸序列。此途径的酶中的一种或多种可以由异源核酸序列编码。在一些实施方案中,此途径的酶中的至少两种或三种可以由异源核酸序列编码。如上文所指出,该异源核酸序列可以源自异源核酸分子。在一些实施方案中,该重组微生物可以包含编码导致形成1,3-丁二烯的途径的所有酶的核酸序列。在一些实施方案中,该重组微生物不包含编码导致形成1,3-丁二烯的途径的所有酶的核酸序列。在一些实施方案中,本文所述的重组微生物可以能够产生1,3-丁二烯。在一些实施方案中,本文所述的重组微生物不能产生1,3-丁二烯。本文所述的重组微生物可以包含编码导致形成正戊烷的途径的酶的核酸序列。在一些实施方案中,该重组微生物可以包含编码导致形成正戊烷的途径的所有酶的核酸序列。在一些实施方案中,该重组微生物不包含编码导致形成正戊烷的途径的所有酶的核酸序列。在一些实施方案中,本文所述的重组微生物能够产生正戊烷。本文所述的重组微生物可以包含编码导致形成1-戊烯的途径的酶的核酸序列。此途径的酶中的一种或多种可以由异源核酸序列编码。在一些实施方案中,此途径的酶中的至少两种或三种可以由异源核酸序列编码。该异源核酸序列可以源自异源核酸分子。在一些实施方案中,该重组微生物可以包含编码导致形成1-戊烯的途径的所有酶的核酸序列。在一些实施方案中,该重组微生物不包含编码导致形成1-戊烯的途径的所有酶的核酸序列。在一些实施方案中,本文所述的重组微生物可以能够产生1-戊烯。在一些实施方案中,本文所述的重组微生物不能产生1-戊烯。在一些实施方案中,本文所述的重组微生物可以包含编码导致形成正己烷的途径的酶的核酸序列。此途径的酶中的一种或多种可以由异源核酸序列编码。在一些实施方案中,该重组微生物可以包含编码导致形成正己烷的途径的所有酶的核酸序列。在一些实施方案中,该重组微生物不包含编码导致形成正己烷的途径的所有酶的核酸序列。在一些实施方案中,本文所述的重组微生物能够产生正己烷。本文所述的重组微生物可以包含编码导致形成1-己烯的途径的酶的核酸序列。此途径的酶中的一种或多种可以由异源核酸序列编码。在一些实施方案中,此途径的酶中的至少两种或三种可以由异源核酸序列编码。在一些实施方案中,该重组微生物可以包含编码导致形成1-己烯的途径的所有酶的核酸序列。在一些实施方案中,该重组微生物不包含编码导致形成1-己烯的途径的所有酶的核酸序列。在一些实施方案中,本文所述的重组微生物能够产生1-己烯。在一些实施方案中,本文所述的重组微生物可以包含编码导致形成异戊二烯(也称为2-甲基-1,3-丁二烯)的途径的酶的核酸序列。导致形成异戊二烯的途径的酶中的一种或多种可以由异源核酸序列编码。在一些实施方案中,导致形成2-甲基-1,3-丁二烯的途径的酶中的至少两种或三种可以由异源核酸序列编码。该异源核酸序列可以源自异源核酸分子。在一些实施方案中,该重组微生物可以包含编码导致形成2-甲基-1,3-丁二烯的途径的所有酶的核酸序列。在一些实施方案中,该重组微生物不包含编码导致形成1-己烯的途径的所有酶的核酸序列。在一些实施方案中,本文所述的重组微生物可以能够产生2-甲基-1,3-丁二烯。在一些实施方案中,本文所述的重组微生物不能产生2-甲基-1,3-丁二烯。该异源核酸序列编码使乙酰coa酶促转化为主链具有2至6个碳原子的一种或多种烷烃和/或烯烃的一种或多种酶。在一些实施方案中,该异源核酸序列编码使乙酰coa酶促转化为主链具有2至5个碳原子的一种或多种烯烃的两种或多种酶。乙酰coa或乙酰辅酶a是乙酸和辅酶a的-巯基乙胺部分的硫酯酶。它在代谢中作为通用成分,可以视为乙酰基团的活化载体。乙酰coa通过尤其是脂肪酸以及糖类的降解而产生,在多种生物合成途径中作为起始物质。乙酰coa不易跨过膜,使得它保留在细胞内并可作为反应物发挥作用。附图中显示可以用本文公开的方法、核酸序列和载体建立、完善或增强的生物合成途径的说明性实例。在一些实施方案中,该重组微生物进一步表达wood-ljungdahl途径的酶。因此,该重组微生物能够从一氧化碳和/或二氧化碳合成乙酰coa。在一些实施方案中,该微生物表达的wood-ljungdahl途径的酶可以由内源核酸序列编码并从内源核酸序列表达。本文所述的重组微生物可以包含编码下文中所述酶的异源核酸序列。所编码的蛋白质可以是该酶的任何等位基因变体。在一些实施方案中,该异源核酸序列可以是见于序列数据库中的核苷酸序列。包含在该重组微生物中的异源核酸序列可以是编码下文所述酶的核酸的同源物。在一些实施方案中,包含在该重组微生物中的异源核酸序列可以包含基本保持该肽/蛋白质的酶活性的保守修饰。在一些实施方案中,该保守修饰使该肽/蛋白质的酶活性基本不变。包含在该重组微生物中的异源核酸序列可以是密码子优化的,即为了在给定生物的细胞中表达而通过用在该生物的基因中更常使用的一种或多种密码子替换了一个或多个、如两个或三个、或更多数目的密码子来对它进行了修改。密码子优化为本领域公知。通常,生物中的高表达基因偏向由该生物中最高丰度的trna种类识别的密码子。这种偏向的一种测量是“密码子适应指数”或“cat”,其测量具体基因中用于编码各氨基酸的密码子是最常出现在来自生物的高表达基因的参考集中的那些密码子的程度。许多生物显示偏向用特定密码子来编码正在生长的肽链中的特定氨基酸的插入。密码子偏好或密码子偏倚(生物间密码子选择的差异)由遗传密码的简并性提供,在许多生物中记录翔实。密码子偏倚常与信使rna(mrna)的翻译效率相关,转而认为信使rna的翻译效率尤其依赖于所翻译的密码子的特性和特定转运rna分子(trna)的可获得性。所选trna在细胞中占优势通常反映肽合成中最常使用的密码子。在一些实施方案中,与下文所述肽/蛋白质的数据库条目的序列相比,包含在该重组微生物中的异源核酸序列可以具有90%或更高的序列同一性。在一些实施方案中,与下文所述肽/蛋白质的数据库条目的序列相比,该重组微生物中的异源核酸序列可以具有95%或更高的序列同一性。在一些实施方案中,与下文所述肽/蛋白质的数据库条目的序列相比,该重组微生物中的异源核酸序列可以具有99%或更高的序列同一性。在一些实施方案中,该重组微生物中的异源核酸序列包含在双链核酸分子中。在一些实施方案中,与异源核酸序列的编码链(即与有义链)互补的链可以能够在严格条件下结合编码下文所述肽/蛋白质的序列的链。在一些实施方案中,本文所述的重组微生物产生自已经包含合成该烷烃和/或烯烃的酶促能力的宿主。在这种实施方案中,它可以用来例如增加该途径的一种或多种中间产物的合成或累积,以例如驱动烷烃/烯烃途径反应朝向烯烃产生。增加的合成或累积可以通过例如过量表达编码上述酶中的一种或多种的一种或多种核酸序列来达到。烷烃或烯烃途径的一种或多种酶和/或一种或多种蛋白质的过量表达可以例如通过外源表达一个或多个内源基因或通过表达一个或多个异源基因而发生。因此,通过过量表达编码该烷烃/烯烃途径的酶的一种或多种(包括全部)核酸序列,可以容易地将天然存在的微生物产生为例如产生烷烃和/或烯烃的非天然存在的微生物。在一些实施方案中,可以通过诱变内源基因来产生重组微生物,该诱变导致烷烃/烯烃生物合成途径中的酶的活性提高。在一些实施方案中,该重组微生物包含编码使乙酰coa转化为乙烯的酶的异源核酸序列。在一些实施方案中,该微生物包含编码乳酸coa转移酶(ec2.8.3.-)和乳酰coa脱水酶(ec4.2.1.54)的一种或多种异源核酸序列。乳酰coa脱水酶的说明性实例是来自丙酸梭菌(clostridiumpropionicum)的酶,其具有2014年11月26日版本13的swissprot检索号g3kim4(lcda_clopr)(该序列的版本1)的α亚基,及2014年10月29日版本20的swissprot检索号q9l3f8(q9l3f8_clopr)(该序列的版本1)的β亚基。乳酸coa转移酶(ec2.8.3.-)可以是酰基coa转移酶,例如酶ec2.8.3.1。适宜的实例是来自丙酸梭菌的乙酸coa转移酶ydif,其对具有至多约4个碳原子的脂肪酸具有宽底物特异性,具有2015年4月1日版本38的swissprot检索号q9l3f7(q9l3f7_clopr)(该序列的版本1)。在一些实施方案中,该微生物包含编码醇脱氢酶的异源核酸序列。该酶的实例是2015年4月1日版本10的swissprot检索号w6cx26(该序列的版本1)的,来自约克氏菌属物种(yokenellasp.)wzy002的醇脱氢酶,其具有genbank检索号ahi87872.1,参见图66。在一些实施方案中,该微生物包含编码nadp依赖型丙二酸单酰coa还原酶(ec1.2.1.75)的异源核酸序列。该酶的实例是2015年5月27日版本56的swissprot检索号a4yen2(该序列的版本1)的,来自勤奋生金球菌(metallosphaerasedula)的丙二酸单酰coa还原酶。另一实例是2015年5月27日版本79的swissprot检索号q96yk1(该序列的版本1)的,来自sulfolobustokodaii的丙二酸单酰coa还原酶。在一些实施方案中,该微生物包含编码丙二酸单酰coa还原酶(ec1.1.1.298)的异源核酸序列。这种酶的实例是2015年7月22日版本114的swissprot检索号p39831(该序列的版本1)的,来自大肠杆菌的nadp依赖型3-羟基酸脱氢酶还原酶。这种酶的另一实例是2015年7月22日版本47的swissprot检索号a7mfx7(该序列的版本1)的,来自cronobactersakazakii的丙二酸半醛还原酶rute。另一适宜的酶是2015年7月22日版本22的swissprot检索号j7r133(该序列的版本1)的,来自大肠杆菌的3-羟基酸脱氢酶。在一些实施方案中,该微生物包含编码3-羟基丙酰coa合酶(ec6.2.1.36)的异源核酸序列。3-羟基丙酰coa合酶的说明性实例是2015年6月24日版本53的swissprot检索号a4a7v6(该序列的版本1)的,来自congregibacterlitoralis的酶。3-羟基丙酰coa合酶的另一实例是2015年7月22日版本49的swissprot检索号a4ygr1(该序列的版本1)的,来自勤奋生金球菌的酶。3-羟基丙酰coa合酶的另一实例是2015年7月22日版本75的swissprot检索号q973w5(该序列的版本1)的,来自sulfolobustokodaii的酶。在一些实施方案中,该微生物包含编码3-羟基丙酰辅酶a脱水酶(ec4.2.1.116)的异源核酸序列。3-羟基丙酰辅酶a脱水酶的说明性实例是2015年7月22日版本11的swissprot检索号f9vng3(该序列的版本1)的,来自sulfolobustokodaii的酶。3-羟基丙酰辅酶a脱水酶的另一实例是2015年7月22日版本52的swissprot检索号a4yi89(该序列的版本1)的,来自勤奋生金球菌的酶。3-羟基丙酰辅酶a脱水酶的另一实例是2015年4月29日版本4的swissprot检索号a0a090iwi4(该序列的版本1)的,来自热噬淀粉芽孢杆菌(bacillusthermoamylovorans)的酶。在一些实施方案中,该微生物包含编码酰基coa硫酯酶(ec3.1.2.-)的异源核酸序列。该酶可以是例如2015年7月22日版本121的swissprot检索号p77781(该序列的版本1)的,来自大肠杆菌的e.c.3.1.2.28的1,4-二羟基-2-萘酰coa水解酶,其与genbank检索号wp_000637982.1相同(参见图66)。此酶对一系列酰基coa硫酯酶具有活性。e.c.3.1.2.28的1,4-二羟基-2-萘酰coa水解酶的另一实例是2015年7月22日版本53的swissprot检索号a2bnw8(该序列的版本1)的,来自海洋原绿球藻(prochlorococcusmarinus)的酶。作为另一实例,该酰基coa硫酯酶可以是2015年7月22日版本82的swissprot检索号p0agg2(该序列的版本2)的,来自大肠杆菌的e.c.3.1.2.2的酰基coa硫酯酶。ec3.1.2.-的酰基coa硫酯酶的另一实例是2015年7月22日版本87的swissprot检索号p0a8z3(该序列的版本1)的,来自大肠杆菌的酰基coa硫酯水解酶ybgc。ec3.1.2.-的酰基coa硫酯酶的另一实例是2015年7月22日版本101的swissprot检索号p94842(该序列的版本2)的,来自幽门螺旋杆菌(helicobacterpylori)的酰基coa硫酯酶ybgc(ec3.1.2.-)。ec3.1.2.-的酰基coa硫酯酶的另一实例是2015年6月24日版本81的swissprot检索号q45061(该序列的版本2)的,来自枯草芽孢杆菌(bacillussubtilis)的酰基coa硫酯酶ynep。ec3.1.2.-的酰基coa硫酯酶的另一实例是2015年7月22日版本15的swissprot检索号w9bnh2(该序列的版本1)的,来自肺炎克雷伯氏菌(klebsiellapneumonia)的酰基coa硫酯酶tesb(ec3.1.2.-)。在一些实施方案中,该微生物包含编码3-羟基丙酰coa脱水酶(ec4.2.1.116)的异源核酸序列。3-羟基丙酰coa脱水酶的说明性实例是2015年7月22日版本52的swissprot检索号a4yi89(该序列的版本1)的,来自勤奋生金球菌的酶。ec4.2.1.116的3-羟基丙酰coa脱水酶的另一实例是2015年4月29日版本4的swissprot检索号a0a090iwi4(该序列的版本1)的,来自勤奋生金球菌的酶。3-羟基丙酰辅酶a脱水酶的另一实例是2015年4月29日版本4的swissprot检索号a0a090iwi4(该序列的版本1)的,来自热噬淀粉芽孢杆菌的酰基coa硫酯酶ybgc。在一些实施方案中,该微生物包含编码ec3.1.2(ec3.1.2.-)的酰基coa硫酯酶(也称为酰基coa硫酯水解酶)的异源核酸序列。酰基coa硫酯酶的说明性实例是2015年6月24日版本81的swissprot检索号q45061(该序列的版本2)的,来自枯草芽孢杆菌的酰基coa硫酯酶ynep(ec:3.1.2.-)。酰基coa硫酯酶的另一实例是2015年7月22日版本2的swissprot检索号a0a0e3jh21(该序列的版本1)的,来自流感嗜血杆菌(haemophilusinfluenza)的酰基coa硫酯酶tesb(ec:3.1.2.-),其具有与genbank检索号wp_005652441.1相同的序列,参见图66。酰基coa硫酯酶的另一实例是2015年7月22日版本17的swissprot检索号v5v9l9(该序列的版本1)的,来自鲍氏不动杆菌(acinetobacterbaumannii)的酰基coa硫酯酶tesb(ec:3.1.2.-)。酰基coa硫酯酶的另一实例是2015年7月22日版本35的swissprot检索号c8u9q5(该序列的版本1)的,来自大肠杆菌的1,4-二羟基-2-萘酰coa水解酶(ec:3.1.2.28),其具有与genbank检索号wp_000637982.1相同的序列,参见图66。酰基coa硫酯酶的另一实例是2015年7月22日版本10的swissprot检索号g7en07(该序列的版本1)的,来自假交替单胞菌属物种(pseudoalteromonassp.)bsi20311的酰基coa硫酯酶tesb(ec:3.1.2.-)。酰基coa硫酯酶的另一实例是2015年7月22日版本82的swissprot检索号p0agg2(该序列的版本2)的,来自大肠杆菌的酰基coa硫酯酶tesb(ec:3.1.2.-)。酰基coa硫酯酶的另一实例是2015年7月22日版本83的swissprot检索号p0ada1(该序列的版本1)的,来自大肠杆菌的酰基coa硫酯酶tesa(ec:3.1.2.-)。酰基coa硫酯酶的另一实例是2015年7月22日版本3的swissprot检索号a0a0e1lws5(该序列的版本1)的酶,其具有与genbank检索号wp_000108160.1相同的序列,参见图66。在一些实施方案中,该微生物可以包含编码磷酸丁酰转移酶(ec2.3.1.19)的异源核酸序列。磷酸丁酰转移酶的实例是2015年7月22日版本78的swissprot检索号p58255(该序列的版本1)的,来自丙酮丁醇梭菌的磷酸丁酰转移酶,其具有genbank检索号wp_010966357.1(参见图66)。磷酸丁酰转移酶的另一实例是2013年10月16日版本85的swissprot检索号p54530(该序列的版本2)的,来自枯草芽孢杆菌的该酶(ec:2.3.1.19)。另一实例是2015年7月22日版本22的swissprot检索号f2npe1(该序列的版本1)的,来自marinithermushydrothermalis的磷酸丁酰转移酶(ec2.3.1.19)。在一些实施方案中,该微生物可以包含编码丁酸激酶(ec2.7.2.7)的异源核酸序列。丁酸激酶的说明性实例是2015年1月7日版本51的swissprot检索号p81337(该序列的版本1)的,来自巴氏梭菌(clostridiumpasteurianum)的酶。丁酸激酶的另一实例是2015年7月22日版本88的swissprot检索号q05619(该序列的版本1)的,来自拜氏梭菌(clostridiumbeijerinckii)的酶。另一实例是2015年7月22日版本31的swissprot检索号q05619(该序列的版本1)的,来自thermosediminibacteroceani的丁酸激酶。在一些实施方案中,该微生物可以包含编码苯基丙烯酸脱羧酶(ec4.1.1.-)(其也可充当酚酸脱羧酶)的异源核酸序列。苯基丙烯酸脱羧酶的说明性实例是2014年2月19日版本113的swissprot检索号p33751(该序列的版本2)的,来自酿酒酵母(saccharomycescerevisiae)的线粒体酶,其与genbank检索号np_010827.3相同(参见图66)。苯基丙烯酸脱羧酶的另一实例是2015年7月22日版本24的swissprot检索号g7xva3(该序列的版本1)的,来自白曲霉(aspergilluskawachii)的酶。苯基丙烯酸脱羧酶的另一实例是2015年5月27日版本5的swissprot检索号u1lp54(该序列的版本1)的,来自pseudoalteromonasrubra的酶。目前命名为酚酸脱羧酶的酶的说明性实例是2015年6月24日版本93的swissprot检索号o07006(该序列的版本1)的,来自枯草芽孢杆菌的酶,其与genbank检索号wp_003243190.1相同(参见图66)。图1b显示导致乙烯合成的示例性途径,其中可以存在上文所示酶中的一些或全部。在一些实施方案中,该微生物可以包含编码4-氨基丁酸氨基转移酶(ec2.6.1.19)的异源核酸序列。4-氨基丁酸氨基转移酶的说明性实例是2014年5月14日版本132的swissprot检索号p22256(该序列的版本1)的,来自大肠杆菌的该酶。4-氨基丁酸氨基转移酶的另一实例是2015年7月22日版本28的swissprot检索号e6slg7(该序列的版本1)的,来自thermaerobactermarianensis的酶。在一些实施方案中,该微生物包含编码谷氨酸脱氢酶(ec1.4.1.2)的异源核酸序列。谷氨酸脱氢酶的说明性实例是2015年7月22日版本111的swissprot检索号p39633(该序列的版本3)的,来自枯草芽孢杆菌的酶。谷氨酸脱氢酶的另一实例是2015年7月22日版本72的swissprot检索号p27346(该序列的版本1)的,来自peptoclostridiumdifficile(艰难梭菌)的酶。在一些实施方案中,该微生物包含编码2-羟基戊二酸脱氢酶(ec1.1.99.2)的异源核酸序列。2-羟基戊二酸脱氢酶的说明性实例是2015年7月22日版本84的swissprot检索号q9n4z0(该序列的版本2)的,来自秀丽新小杆线虫(caenorhabditiselegans)的线粒体酶。2-羟基戊二酸脱氢酶的另一实例是2015年7月22日版本22的swissprot检索号g8q888(该序列的版本1)的,来自荧光假单胞菌(pseudomonasfluorescens)的酶。在一些实施方案中,该微生物包含编码戊烯二酸coa转移酶(ec2.8.3.12)的异源核酸序列。该酶的实例是2015年7月22日版本21的swissprot检索号e3pwj5(该序列的版本1)的,来自斯氏梭菌(clostridiumsticklandii)的戊烯二酸coa转移酶。戊烯二酸coa转移酶的另一实例是这样的酶,该酶具有2015年4月1日版本95的swissprot检索号q59111(该序列的版本3)的来自发酵氨基酸球菌(acidaminococcusfermentans)的a亚基,及2015年7月22日版本95的swissprot检索号q59112(该序列的版本3)的来自发酵氨基酸球菌的b亚基。另一实例是2015年7月22日版本50的swissprot检索号q1d4i4(该序列的版本1)的来自黄色粘球菌(myxococcusxanthus)的酶的a亚基,其具有genbank检索号wp_011554267.1,参见图66。此酶具有2015年7月22日版本46的swissprot检索号q1d4i3(该序列的版本1)的b亚基,其具有genbank检索号wp_011554268.1,参见图66。在一些实施方案中,该微生物可以包含编码2-羟基戊二酰coa脱水酶(ec4.2.1.-)的异源核酸序列。2-羟基戊二酰coa脱水酶的说明性实例是具有2015年7月22日版本83的swissprot检索号p11569(该序列的版本3)的来自发酵氨基酸球菌的α亚基的酶。2-羟基戊二酰coa脱水酶的另一实例是2015年7月22日版本19的swissprot检索号d8gt18(该序列的版本1)的来自杨氏梭菌的酶。在一些实施方案中,该微生物包含编码戊烯二酸单酰coa脱羧酶(ec4.1.1.70)的异源核酸序列。该酶的实例是来自发酵氨基酸球菌的酶,其具有2015年4月1日版本82的swissprot检索号q06700(该序列的版本1)的α亚基,2015年7月22日版本81的swissprot检索号q9zaa6(该序列的版本1)的β亚基,2014年11月26日版本82的swissprot检索号q9zaa7(该序列的版本1)的γ亚基,及2015年7月22日版本78的swissprot检索号q9zaa8(该序列的版本1)的δ亚基。swissprot检索号q9zaa8的δ亚基具有genbank检索号wp_012939173.1,参见图66。戊烯二酸单酰coa脱羧酶的另一实例是2015年5月27日版本50的swissprot检索号q2rn28(该序列的版本1)的,来自深红红螺菌的酶。戊烯二酸单酰coa脱羧酶的另一实例是2015年7月22日版本40的swissprot检索号a9klq1(该序列的版本1)的,来自clostridiumphytofermentans的酶。在一些实施方案中,该微生物可以包含编码nad依赖型4-羟基丁酸脱氢酶(也称为4-羟基丁酸脱氢酶)(ec1.1.1.61)的异源核酸序列。nad依赖型4-羟基丁酸脱氢酶的说明性实例是2015年7月22日版本72的swissprot检索号p38945(该序列的版本3)的,来自克氏梭菌(clostridiumkluyveri)的酶。nad依赖型4-羟基丁酸脱氢酶的另一实例是2015年7月22日版本16的swissprot检索号h6lhy2(该序列的版本1)的,来自伍氏醋杆菌的酶。在一些实施方案中,该微生物可以包含编码4-羟基丁酸coa转移酶(ec2.8.3.-)的异源核酸序列。这种酶的实例是2015年7月22日版本81的swissprot检索号q8x5x6(该序列的版本1)的,来自大肠杆菌的乙酸coa转移酶ydif。此酶对短链脂肪酸具有宽底物特异性,催化丙酸、乙酰乙酸、丁酸、异丁酸和4-羟基丁酸的反应。4-羟基丁酸coa转移酶的另一实例是2015年7月22日版本2的swissprot检索号a0a0e8xk97(该序列的版本1)的,来自假结核耶尔森氏菌(yersiniapseudotuberculosis)的酶。4-羟基丁酸coa转移酶的另一实例是2015年6月24日版本48的swissprot检索号q9rm86(该序列的版本2)的,来自氨基丁酸梭菌(clostridiumaminobutyricum)的酶。在一些实施方案中,该微生物可以包含编码乙烯乙酰coaδ异构酶(ec5.3.3.3)(在许多实施方案中,其也是4-羟基丁酰coa脱水酶(ec4.2.1.120))的异源核酸序列。这种具有两种活性的酶的实例是2015年4月1日版本79的swissprot检索号p55792(该序列的版本3)的,来自氨基丁酸梭菌的酶。乙烯乙酰coaδ异构酶的另一实例是2015年1月7日版本3的swissprot检索号a0a011pck9(该序列的版本1)的,来自candidatusaccumulibactersp.sk-11的酶。乙烯乙酰coaδ异构酶和4-羟基丁酰coa脱水酶的另一实例是2015年6月24日版本8的swissprot检索号w6nes0(该序列的版本1)的,来自酪丁酸梭菌(clostridiumtyrobutyricum)的酶。具有乙烯乙酰coaδ异构酶活性(ec5.3.3.3)和4-羟基丁酰coa脱水酶活性(ec4.2.1.-)的适宜酶的另一实例是2015年7月22日版本67的swissprot检索号q3aci6(该序列的版本1)的,来自生氢氧化碳嗜热菌的酶。具有乙烯乙酰coaδ异构酶活性(ec5.3.3.3)和4-羟基丁酰coa脱水酶活性(ec4.2.1.-)的适宜酶的另一实例是2015年7月22日版本39的swissprot检索号b9e5e4(该序列的版本1)的,来自克氏梭菌的酶,其具有genbank检索号wp_012103363.1,参见图66。仅已知其作为4-羟基丁酰coa脱水酶落在ec4.2.1.120之下的酶的实例是2015年7月22日版本14的swissprot检索号k0ip19(该序列的版本1)的,来自nitrososphaeragargensis的酶。仅已知其具有4-羟基丁酰coa脱水酶活性的酶的另一实例是2015年7月22日版本20的swissprot检索号g4rjk0(该序列的版本1)的,来自附着热变形菌的酶。在一些实施方案中,该微生物可以包含编码乙酰乙酰coa硫解酶或乙酰coa乙酰转移酶(ec2.3.1.9)的异源核酸序列。该酶的实例是2015年1月7日版本74的swissprot检索号p45362(该序列的版本2)的,来自peptoclostridiumdifficile(艰难梭菌)的酶。乙酰coa乙酰转移酶的两个其他实例是2015年7月22日版本12的swissprot检索号a0a024lb55(该序列的版本1)的,及2015年7月22日版本17的swissprot检索号h9uug8(该序列的版本1)的,来自大肠杆菌的酶,其包含genbank检索号akr29896.1和wp_000786547.1的序列,参见图66。适宜的乙酰coa乙酰转移酶的另一实例是2015年7月22日版本62的swissprot检索号q0kbp1(该序列的版本1)的,来自钩虫贪铜菌(cupriavidusnecator)的β-酮硫解酶bktb,其与genbank检索号wp_011615089.1的序列相同,参加图66。乙酰coa乙酰转移酶的另一实例是2015年7月22日版本4的swissprot检索号a0a0d0f6v4(该序列的版本1)的,来自热噬淀粉芽孢杆菌的酶。在一些实施方案中,该微生物可以包含编码3-羟基丁酰coa脱氢酶(ec1.1.1.157)的异源核酸序列。3-羟基丁酰coa脱氢酶的说明性实例是2015年5月27日版本101的swissprot检索号p52041(该序列的版本2)的,来自丙酮丁醇梭菌的酶。3-羟基丁酰coa脱氢酶的另一实例是2015年7月22日版本52的swissprot检索号b5xrj5(该序列的版本1)的,来自肺炎克雷伯氏菌的酶。3-羟基丁酰coa脱氢酶的另一实例是2015年7月22日版本45的swissprot检索号b7l6b5(该序列的版本1)的,来自大肠杆菌的酶。在一些实施方案中,该微生物可以包含编码3-羟基丁酰coa脱水酶(ec4.2.1.55)的异源核酸序列。3-羟基丁酰coa脱水酶的实例是2014年10月29日版本34的swissprot检索号c5az74(该序列的版本1)的,来自扭脱甲基杆菌(methylobacteriumextorquens)的酶。另一实例是2015年7月22日版本60的swissprot检索号q189z6(该序列的版本1)的,来自peptoclostridiumdifficile的酶。在一些实施方案中,该微生物可以包含编码乙酰coa羧化酶(ec6.4.1.2)的异源核酸序列。此酶通常是酶复合物。示例性亚基是2015年7月22日版本13的swissprot检索号w8zll8(该序列的版本1)的来自大肠杆菌的β亚基,其对应于genbank检索号abe08068.1(参见图66)。此酶的来自大肠杆菌的α亚基具有2015年7月22日版本12的swissprot检索号w8zde6(该序列的版本1)。在一些实施方案中,该微生物可以包含编码生物素羧化酶(ec6.3.4.14)的异源核酸序列。适宜的生物素羧化酶的实例是2015年7月22日版本5的swissprot检索号a0a0a7a003(该序列的版本1)的来自痢疾志贺氏菌(shigelladysenteriae)的酶,其对应于genbank检索号aha67379.1(参见图66)。在一些实施方案中,该微生物可以包含编码乙酰乙酰coa合酶(ec2.3.1.194)的异源核酸序列。乙酰乙酰coa合酶的说明性实例是2015年7月22日版本23的swissprot检索号d7urv0(该序列的版本1)的来自链霉菌属物种(streptomycessp.)(菌株cl190)的酶,其与genbank检索号d7urv0.1相同(参见图66)。乙酰乙酰coa合酶的另一实例是2015年7月22日版本5的swissprot检索号a0a0b0i6s7(该序列的版本1)的来自类芽孢杆菌属物种(paenibacillussp.)p1xp2的酶。在一些实施方案中,该微生物可以包含编码乙酰乙酰coa还原酶(ec1.1.1.36)的异源核酸序列。乙酰乙酰coa还原酶的说明性实例是2015年1月7日版本70的swissprot检索号p50203(该序列的版本1)的来自不动杆菌属物种(acinetobactersp.)(菌株ra3849)的酶。乙酰乙酰coa还原酶的另一实例是2015年7月22日版本41的swissprot检索号c5azn8(该序列的版本1)的来自扭脱甲基杆菌的酶。乙酰乙酰coa还原酶的另一实例是2015年7月22日版本7的swissprot检索号w0a092(该序列的版本1)的来自嗜水气单胞菌(aeromonashydrophila)的酶。在一些实施方案中,该微生物可以包含编码烯酰coa水合酶如(r)-特异性烯酰coa水合酶(ec4.2.1.119)的异源核酸序列。烯酰coa水合酶的说明性实例是2015年7月22日版本70的swissprot检索号q88fm3(该序列的版本1)的,来自恶臭假单胞菌(pseudomonasputida)的酶,其与genbank检索号wp_003251320.1的酶相同,参见图66。烯酰coa水合酶的另一实例是2015年7月22日版本9的swissprot检索号w0a1x1(该序列的版本1)的,来自嗜水气单胞菌的酶,其与genbank检索号wp_025327110.1的酶相同,参见图66。(r)-特异性烯酰coa水合酶(ec4.2.1.119)的实例是2015年6月24日版本59的swissprot检索号q2rq36(该序列的版本1)的,来自深红红螺菌的酶。(r)-特异性烯酰coa水合酶(ec4.2.1.119)的另一实例是2015年5月27日版本123的swissprot检索号q02207(该序列的版本1)的,来自酿酒酵母的酶。(r)-特异性烯酰coa水合酶(ec4.2.1.119)的另一实例是2015年6月24日版本7的swissprot检索号x7yi38(该序列的版本1)的,来自蟾蜍分枝杆菌(mycobacteriumxenopi)的过氧化物酶体酶。图1c显示导致丙烯合成的示例性途径,其中可以存在上文所示酶中的一种或多种。在一些实施方案中,该微生物可以包含编码丙烯酰辅酶a还原酶(ec1.3.1.84)(其也可以称为丙酰coa合酶)的异源核酸序列。丙烯酰辅酶a还原酶的实例是2015年7月22日版本96的swissprot检索号q975c8(该序列的版本1)的,来自sulfolobustokodaii的酶。丙烯酰辅酶a还原酶的另一实例是2015年4月29日版本65的swissprot检索号q3j6k9(该序列的版本1)的,来自球形红细菌(rhodobactersphaeroides)的酶。在一些实施方案中,该微生物可以包含编码甲基丙二酸单酰coa变位酶(ec5.4.99.2)的异源核酸序列。甲基丙二酸单酰coa变位酶的实例是2015年7月22日版本124的swissprot检索号p27253(该序列的版本2)的,来自大肠杆菌的酶。甲基丙二酸单酰coa变位酶的另一实例是2015年6月24日版本77的swissprot检索号q05065(该序列的版本1)的,来自肉桂地链霉菌(streptomycescinnamonensis)的酶。甲基丙二酸单酰coa变位酶的另一实例是2015年6月24日版本29的swissprot检索号c3n159(该序列的版本1)的,来自冰岛硫化叶菌(sulfolobusislandicus)的酶。在一些实施方案中,该微生物可以包含编码甲基丙二酸单酰coa脱羧酶(ec4.1.1.41)的异源核酸序列。甲基丙二酸单酰coa脱羧酶的说明性实例是2015年7月22日版本108的swissprot检索号p52045(该序列的版本1)的,来自大肠杆菌的酶。甲基丙二酸单酰coa脱羧酶的另一实例是2015年7月22日版本4的swissprot检索号a0a0c6fwg2(该序列的版本1)的,来自酿脓链球菌(streptococcuspyogenes)的酶。在一些实施方案中,该微生物可以包含编码醛-醇脱氢酶的异源核酸序列。这种酶通常包含醇脱氢酶活性(ec1.1.1.1),且还可以包含乙醛脱氢酶活性(ec1.2.1.10)。醛-醇脱氢酶的说明性实例是2015年5月27日版本24的swissprot检索号q7dfn2(该序列的版本1)的,来自丙酮丁醇梭菌的酶,其具有genban检索号wp_010890720.1(参见图66)。图1d显示导致正丁烯合成的示例性途径,其中可以存在上文所示酶中的一种或多种。在一些实施方案中,该微生物可以包含编码3-羟基-3-甲基戊二酰coa合酶(ec2.3.3.10)的异源核酸序列。3-羟基-3-甲基戊二酰coa合酶的实例是2015年7月22日版本37的swissprot检索号d4gwr6(该序列的版本1)的,来自沃氏盐杆菌(haloferaxvolcanii)的酶。3-羟基-3-甲基戊二酰coa合酶的另一实例是2015年7月22日版本3的swissprot检索号a0a0e1xb83(该序列的版本1)的,来自金黄色葡萄球菌(staphylococcusaureus)的酶,其与genban检索号wp_000172190.1相同。3-羟基-3-甲基戊二酰coa合酶的另一实例是2015年7月22日版本18的swissprot检索号i2c5e4(该序列的版本1)的,来自解淀粉芽孢杆菌(bacillusamyloliquefaciens)y2的酶。在一些实施方案中,该微生物可以包含编码甲基戊烯二酸单酰coa水合酶(也称为3-甲基戊烯二酸单酰coa水合酶)(ec4.2.1.18)的异源核酸序列。甲基戊烯二酸单酰coa水合酶的说明性实例是2015年7月22日版本67的swissprot检索号q54hg7(该序列的版本1)的,来自盘基网柄菌(dictyosteliumdiscoideum)的线粒体酶。甲基戊烯二酸单酰coa水合酶的另一实例是2015年7月22日版本5的swissprot检索号a0a068n7t5(该序列的版本1)的,来自蜡样芽孢杆菌(bacilluscereus)的酶。甲基戊烯二酸单酰coa水合酶的另一实例是2015年7月22日版本36的swissprot检索号b8fc62(该序列的版本1)的,来自desulfatibacillumalkenivorans的酶。在一些实施方案中,该微生物可以包含编码甲基巴豆酰coa羧化酶(也称为3-甲基巴豆酰coa羧化酶)(ec6.4.1.4)的异源核酸序列。甲基巴豆酰coa羧化酶的说明性实例是来自盘基网柄菌的线粒体酶,其具有2013年5月29日版本80的swissprot检索号q54ke6(该序列的版本1)的α亚基,及2015年6月24日版本76的swissprot检索号q8t2j9(该序列的版本2)的β亚基。甲基巴豆酰coa羧化酶的另一实例是来自octadecabacterarcticus的酶,其具有2015年7月22日版本14的swissprot检索号m9rsw8(该序列的版本1)的α亚基,及2015年4月29日版本15的swissprot检索号m9rl62(该序列的版本1)的β亚基。甲基巴豆酰coa羧化酶的另一实例是2015年6月24日版本91的swissprot检索号q9i299(该序列的版本1)的,来自铜绿假单胞菌(pseudomonasaeruginosa)的酶,其与genbank检索号wp_003113506.1的酶相同,参见图66。甲基巴豆酰coa羧化酶的另一实例是2015年7月22日版本8的swissprot检索号w1mjk9(该序列的版本1)的,来自铜绿假单胞菌的酶,其与genbank检索号wp_003100387.1的酶相同,参见图66。图1e显示导致异丁烯合成的示例性途径,其中可以存在上文所示酶中的一种或多种。在一些实施方案中,该微生物可以包含编码丙酮酸/2-氧代丁酸铁氧还蛋白氧化还原酶(也称为丙酮酸/酮异戊酸氧化还原酶或丙酮酸合酶)(ec1.2.7.1)的异源核酸序列。2-氧代丁酸铁氧还蛋白氧化还原酶的说明性实例是来自巴氏甲烷八叠球菌(methanosarcinabarkeri)的丙酮酸合酶,其具有2015年7月22日版本83的swissprot检索号p80521(该序列的版本2)的pora亚基,2015年7月22日版本81的swissprot检索号p80522(该序列的版本3)的porb亚基,及2015年7月22日版本84的swissprot检索号p80523(该序列的版本2)的porc亚基。丙酮酸/2-氧代丁酸铁氧还蛋白氧化还原酶的另一实例是2014年10月1日版本21的swissprot检索号p84819(该序列的版本1)的,来自pyrococcusendeavori的称为丙酮酸/酮异戊酸氧化还原酶共同亚基γ(porg)的酶。另一实例是在一些实施方案中,该微生物可以包含编码2-氧代戊二酸/丁酸铁氧还蛋白氧化还原酶(也称为2-酮戊二酸氧化还原酶或2-氧代戊二酸合酶)(ec1.2.7.3)的异源核酸序列。2-氧代戊二酸铁氧还蛋白氧化还原酶的说明性实例是来自methanocaldococcusjannaschii的丙酮酸合酶,其具有2015年6月24日版本88的swissprot检索号q57724(该序列的版本1)的kora亚基,2015年7月22日版本83的swissprot检索号q57957(该序列的版本1)的korb亚基,及2015年6月24日版本85的swissprot检索号q57956(该序列的版本1)的korc亚基。2-氧代戊二酸/丁酸铁氧还蛋白氧化还原酶的另一实例是来自thermococcussibiricus的2-酮戊二酸:铁氧还蛋白氧化还原酶(kgor),其具有2015年5月27日版本32的swissprot检索号c6a4h7(该序列的版本1)的α亚基,2015年5月27日版本32的swissprot检索号c6a4h6(该序列的版本1)的β亚基,及2015年5月27日版本31的swissprot检索号c6a4h5(该序列的版本1)的γ亚基。在一些实施方案中,该微生物可以包含编码醛铁氧还蛋白氧化还原酶(ec1.2.7.5)的异源核酸序列。醛铁氧还蛋白氧化还原酶的说明性实例是2015年7月22日版本10的swissprot检索号u5rp02(该序列的版本1)的,来自clostridiumautoethanogenum的酶,其具有genbank检索号wp_013238665.1,参见图66。该酶也可以是genbank检索号wp_010966356.1(参见图66)的来自丙酮丁醇梭菌的丁酸激酶,其完全包含在swissprot检索号u5rp02的醛铁氧还蛋白氧化还原酶中。在一些实施方案中,该微生物可以包含编码牻牛儿醇异构酶(ec5.4.4.4)(也称为牻牛儿醇羟基变位酶)的异源核酸序列。此酶是双功能酶,且还作为芳樟醇脱水酶(ec4.2.1.127),也称为芳樟醇脱水酶-异构酶。牻牛儿醇异构酶/芳樟醇脱水酶的实例是2015年4月1日版本12的swissprot检索号e1xuj2(该序列的版本1)的,来自castellanielladefragrans的酶。牻牛儿醇异构酶/芳樟醇脱水酶的另一实例是2015年3月4日版本11的swissprot检索号l2frh3(该序列的版本1)的,来自胶孢炭疽菌(colletotrichumgloeosporioides)的酶。牻牛儿醇异构酶/芳樟醇脱水酶的另一实例是2015年6月24日版本5的swissprot检索号a0a024jqr5(该序列的版本1)的,来自mycobacteriumtriplex的酶。在一些实施方案中,该微生物可以包含编码反式-2-烯酰coa还原酶(也称为乙酰乙酰coa还原酶)(ec1.1.1.36)的异源核酸序列。乙酰乙酰coa还原酶的说明性实例是2015年6月24日版本76的swissprot检索号p50204(该序列的版本1)的,来自脱氮副球菌(paracoccusdenitrificans)的酶。乙酰乙酰coa还原酶的另一实例是2015年7月22日版本31的swissprot检索号e3hwi9(该序列的版本1)的,来自氧化木糖无色杆菌(achromobacterxylosoxidans)的酶。在一些实施方案中,该微生物可以包含编码丁酸激酶(ec2.7.2.7)的异源核酸序列。丁酸激酶的实例是2015年3月27日版本90的swissprot检索号q45829(该序列的版本2)的,来自丙酮丁醇梭菌的丁酸激酶1。丁酸激酶的另一实例是2015年7月22日版本80的swissprot检索号q97ii1(该序列的版本1)的,来自丙酮丁醇梭菌的丁酸激酶2。丁酸激酶的另一实例是2015年7月22日版本34的swissprot检索号g4fge0(该序列的版本1)的,来自海栖热袍菌(thermotogamaritima)的酶。在一些实施方案中,该微生物可以包含编码异戊二烯合酶(ec4.2.3.27)的异源核酸序列。异戊二烯合酶的实例是2015年4月1日版本42的swissprot检索号q6ej97(该序列的版本1)的,来自puerariamontanavar.lobate的酶,其还具有genbank检索号q6ej97.1,参见图66。异戊二烯合酶的另一实例是2015年3月27日版本38的swissprot检索号q50l36(该序列的版本1)的,来自银白杨(populusalba)的酶。在一些实施方案中,该微生物可以包含编码二酰甘油激酶的异源核酸序列。二酰甘油激酶的说明性实例是来自变形链球菌(streptococcusmutans)gs-5的酶,其具有2015年6月24日版本106的swissprot检索号q05888(该序列的版本2),2010年6月23日条目版本5的genbank检索号aaa26867,参见图66。此酶可以例如是具有二酰甘油激酶活性的十一异戊烯醇激酶(ec2.7.1.66)。该十一异戊烯醇激酶的实例是2015年6月24日版本106的swissprot检索号q05888(该序列的版本2)的,来自变形链球菌血清型c的酶,其与genbank检索号aaa26867.1的二酰甘油激酶98%同一。在一些实施方案中,该微生物可以包含编码2-甲基-3-丁烯-2-醇合酶的异源核酸序列。2-甲基-3-丁烯-2-醇合酶的说明性实例是2015年4月1日版本14的swissprot检索号i3qdt7(该序列的版本1)的,来自加州沼松(pinusmuricata)的酶。2-甲基-3-丁烯-2-醇合酶的另一实例是2015年4月1日版本14的swissprot检索号i3qdv8(该序列的版本1)的,来自辐射松(pinusradiata)的酶,其具有genbank检索号afj73575.1,参见图66。图1f显示导致丁二烯合成的途径,其中可以存在上文所示酶中的一种或多种。图1g显示导致2-甲基-1,3-丁二烯合成的途径,其中可以存在上文所示酶中的一种或多种。图1h显示导致1-戊烯合成的途径,其中可以存在上文所示酶中的一种或多种。图66显示由下文所述质粒编码的示例性酶。此附图中所示的任意酶可以由包含在重组微生物中的异源核酸序列编码,并由该异源核酸序列表达。在一些实施方案中,该重组微生物可以包含表达图66中所示的两种或多种酶的一种或多种异源核酸序列。在一些实施方案中,利用一种或多种编码核酸序列的外源表达。外源表达赋予了为宿主量身定制表达和/或调节元件并用来达到受使用者控制的希望得到的表达水平的能力。在一些实施方案中,例如通过去除负调节效应子或在与诱导型启动子或其他调节元件连接时通过诱导基因的启动子来利用内源表达。具有天然存在的诱导型启动子的内源基因可以通过提供适当的诱导剂来上调,或者可以改造内源基因的调节区,以掺入诱导型调节元件,从而允许在所希望的时间调节内源基因的提高的表达。类似地,可以包含诱导型启动子作为引入非天然存在的微生物的外源基因的调节元件。对于一些已知具有某种特定活性的酶,已报道该酶也催化非常不同的功能。作为实例,swissprot检索号m9rsw8的来自octadecabacterarcticus的甲基巴豆酰coa羧化酶具有甲基巴豆酰coa羧化酶活性和生物素羧化酶活性二者。作为另外两个实例,swissprot检索号b8fc62的来自desulfatibacillumalkenivorans的甲基戊烯二酸单酰coa水合酶具有甲基戊烯二酸单酰coa水合酶活性和烯酰coa异构酶活性二者,swissprot检索号a0a0c6fwg2的来自酿脓链球菌的甲基丙二酸单酰coa脱羧酶具有甲基丙二酸单酰coa脱羧酶活性、烯酰coa异构酶活性和烯酰coa水合酶活性。另一方面,还可以用定向进化修饰酶,以扩宽其对非天然底物的特异性。备选地,还可以用定向进化来改变酶的底物偏好。由此可针对对天然底物(例如改善的效率)或非天然底物(例如提高的效率)的有效功能改造给定的酶。在这方面,已用算法设计了逆醛缩酶(retro-aldolase),其可以用于催化非天然和非生物底物4-羟基-4-(6-甲氧基-2-萘基)-2-丁酮中的碳-碳键切割(jiang,l.等,science(2008)319,1387-1391)。这些算法用四种不同催化基序的不同组合来涉及新的酶。与未催化的反应相比,所选择的用于实验表征的设计中有20种的速率提高四倍(jiang等,2008,上文)。这样,不仅可以改变酶可以使用的潜在底物的系列,还可以进行酶的设计和构建。作为实例,已报道了dna混编法(临时模板随机嵌合或rachitt)产生经改造的单加氧酶,其在复杂底物上具有改善的脱硫速率,以及快20倍的天然底物转化(coco,wm等,nat.biotechnol.(2001)19,4,354-359)。类似地,使缓慢型突变体磷酸丙糖异构酶的比活从1.3倍提高了至多19倍(hermes,jd等,proc.nat.acad.sciu.s.a.(1990)87,2,696-700)。这种比活的增强是通过在蛋白质全长内使用随机诱变达到的,该改善可以追溯至六个氨基酸残基中的突变。如上文所指出,该重组微生物可以包含wood-ljungdah途径的内源酶。可以在天然存在的微生物作为已包含wood-ljungdahl途径的功能性内源酶的宿主的基础上改造得到该重组微生物。wood-ljungdahl途径也称为还原性乙酰coa途径,是使co和/或co2转化为乙酰coa的酶促反应途径。此途径在图1b至图1f的任一个中显示。在用co2作为反应物时,通过一氧化碳(co)脱氢酶/乙酰coa合酶将一个co2分子还原为co。还原另一个co2分子产生羰基。该重组微生物可以是产乙酸菌,即专性厌氧菌,其中还原性乙酰coa或wood-ljungdahl途径作为细菌进行能量转化的主要机制。以co2作为底物,在产乙酸菌中,乙酰coa的形成通常需要氢分子。因此该微生物能够利用co和合成气,以及通常还能够利用co2和co2/h2混合物。获得重组微生物许多原核生物适合用于获得本文所述的重组微生物,如革兰氏阴性菌。在一些实施方案中,所使用的微生物是可以发酵一碳反应物如一氧化碳或二氧化碳的细菌。适宜的物种可以属于以下属:醋杆菌属、醋盐杆菌属(acetohalobium)、厌氧醋菌属(acetoanaerobium)、acetitomaculum、醋丝菌属(acetonema)、alkalibaculum、anoxybacillus、ammonifex、古生球菌属(archaeoglobus)、芽孢杆菌属(bacillus)、blautia、慢生根瘤菌属(bradyrhizobium)、bryanella、丁酸杆菌属、喜热菌属(caloramator)、calderihabitans、caloribacterium、caldanaerobacter、candidatus、嗜碳菌属(carbophilus)、carboxydobrachium、carboxydocella、氧化碳嗜热菌属、cloacibacillus、梭菌属、脱硫项圈菌属(desulfomonile)、desulfovirgula、全噬菌属(holophaga)、真杆菌属(eubacterium)、desulfitbacterium、脱硫肠状菌属、desulfurispora、desulfosporosinus、dehalococcoides、desulfatibacillum、脱硫杆菌属(desulfobacterium)、desulfobacula、脱硫螺菌属(desulfospira)、desulfovermiculus、脱硫弧菌属、网球菌属(dictyoglomus)、hydrogenophilus、产氢噬胞菌属(hydrogenophaga)、穆尔氏菌属、甲烷短杆菌属(methanobrevibacter)、分枝杆菌属、natroniella、natronincola、寡养菌属(oligotropha)、醋菌属(oxobacter)、orenia、类芽孢杆菌属、消化链球菌属、假单胞菌属(pseudomonas)、ralstonia、红假单胞菌属(rhodopseudomonas)、红螺菌属(rhodospirillum)、红长命菌属(rubrivivax)、瘤胃球菌属、scalindua、共养球菌属(syntrophococcus)、链霉菌属、鼠孢菌属、热厌氧杆菌属(thermoanaerobacter)、thermincola、thermoacetogenium、thermosinus、tindallia、thermosediminibacter、密螺旋体属(treponema)、thermolithobacter、热球菌属(thermococcus)、热丝菌属(thermofilum)或热变形菌属(thermoproteus)。通常,首先分析选择用于开发为产烷烃/烯烃菌株的菌株,以确定哪些产烯烃基因是该菌株内源的,哪些基因不存在。将该菌株中不存在其内源副本的基因组装在一个或多个重组构建体中,然后将该重组构建体转化入该菌株以提供一种或多种缺失的功能。根据需要,在该菌株中存在其内源副本的基因可以修饰或用一个或多个重组基因补充,例如以增强特定途径的通量或特定步骤。在一些实施方案中,该重组微生物是源自天然存在的一氧化碳营养细胞的细胞。该宿主细胞因此可以是一氧化碳营养的。在一些实施方案中,该重组细胞源自天然存在的耐热宿主细胞。该宿主细胞因此可以是耐热的。由于其在相对高的温度下生长的能力,耐热宿主细胞用于生物技术中的方法应用。在一些实施方案中,该宿主细胞可以是嗜热的,即在约45℃或更高温度下繁盛的细胞。在一些实施方案中,该宿主细胞可以是嗜中温的,即在从约20至约45℃的范围内繁盛的细胞。可以用来获得本文公开的微生物的许多完善的基因工程技术在本领域可得。这类技术包括分子方法,如异源基因表达、基因组插入或缺失、改变基因表达、失活基因或酶工程方法。在本文公开的方法中,核酸分子可以通过本领域可得的用于转化细胞的任何适宜的核酸递送技术引入宿主细胞。适宜的技术的实例包括但不限于直接递送dna,例如,通过转染、注射(包括显微注射)、电穿孔、磷酸钙沉淀、通过使用deae葡聚糖后使用聚乙二醇、直接超声负荷、脂质体介导转染、受体介导转染、微粒轰击、碳化硅纤维搅拌、农杆菌介导转化、脱水/抑制介导dna摄取或其任意组合。外源核酸分子可以作为裸核酸分子递送至微生物,或者它可以用一种或多种物质配制以便于转化过程。作为说明性实例,可以提供脂质体缀合的核酸。本文所述的微生物可以通过将一种或多种外源核酸引入微生物而获得。多种将核酸分子引入细胞的技术在本领域可得。例如,可以通过电穿孔、超声处理、聚乙二醇介导转化、化学或天然感受态、原生质体转化、原噬菌体诱发或接合来达到转化,包括转导或转染。已针对几种一氧化碳营养产乙酸菌描述了电穿孔,如杨氏梭菌(等,poc.nat.acad.sci.u.s.a.(2010)107,13087-13092;wo2012/053905;leang等,applenvironmicrobiol.(2013)79,4,1102-1109)、c.autoethanogenum(wo2012/053905)、伍氏醋杆菌(straetz等,appl.environ.microbiol.(1994)60,1033-37)或热醋穆尔氏菌(kita等,jbioscibioeng.(2013)115,4,347-352),电穿孔是用于许多梭菌的标准方法,如丙酮丁醇梭菌(c.acetobutylicum)(mermelstein等,biotechnology(1992)10,190-195)、解纤维梭菌(c.cellulolyticum)(jennert等,2000,microbiology,146,3071-3080)或热纤维梭菌(c.thermocellum)(tyurin等,appl.environ.microbiol.(2004)70,883-890)。接合已描述为包括艰难梭菌(herbert等,femsmicrobiol.lett.(2003)229,103-110)或丙酮丁醇梭菌(williams等,j.gen.microbiol.(1990)136,819-826)的许多梭菌的方法选择,且可以以类似的方式用于一氧化碳营养产乙酸菌。在某些实施方案中,由于在待转化的微生物中有活性的限制系统,有必要甲基化待引入该微生物的核酸。这可以用本领域已知的多种技术进行。载体和基因整合在一些实施方案中,本文所述的微生物可以是细菌。在一些实施方案中,本文所述的细菌可以包含细菌染色体和附加核酸分子。该附加核酸分子可以按上文所述引入。在一些实施方案中,本文所述的细菌包含细菌染色体,不包含附加核酸分子。在一些实施方案中,该细菌染色体可以是经过修饰的,且包含本文所述的外源核酸序列。该外源核酸序列可以是异源核酸序列。在一个实施方案中,该细菌染色体包含本文所述的外源(包括异源)核酸序列,不包含附加核酸分子。在一些实施方案中,本文所述的微生物包含编码本文所述的酶的两种或多种异源核酸序列。在一些实施方案中,本文所述的微生物包含编码本文所述的酶的三种或多种异源核酸序列。本文所述的微生物还可以包含编码本文所述的两种或多种酶的异源核酸序列。本文所述的微生物还可以包含编码本文所述的三种或多种酶的异源核酸序列。在一些实施方案中,本文公开的核酸分子包含能够诱导和/或调节酶的表达的表达盒。在一些实施方案中,本文公开的核酸分子包含在载体内,该载体包含对在该宿主细胞中起始转录有效的启动子(无论内源或外源来源)。在微生物中表达酶可以包括产生载体,该载体具有含有编码该酶的序列的构建体。可以利用的大量适宜的载体是已知的。一旦制备出用于表达的包含一个或多个构建体的载体或核酸分子,即可通过多种适宜手段(即转化、转染、接合、原生质体融合、电穿孔、微粒枪技术、磷酸钙沉淀、直接显微注射等)中的任一种将该一个或多个构建体引入所选择的适宜宿主细胞。在一些实施方案中,该载体包含选择标记。选择标记的实例包括但不限于抗生素抗性核酸(例如卡那霉素、氨苄青霉素、羧苄青霉素、庆大霉素、潮霉素、腐草霉素、博来霉素、新霉素或氯霉素)和/或赋予宿主细胞代谢优势(如营养优势)的核酸。在这种实施方案中,在引入载体后,受体细胞可以在选择性培养基中生长,该选择性培养基针对含有载体的微生物的生长进行选择。所克隆的一个或多个基因的表达导致该酶的产生。这可以这样或在诱导这些细胞分化后发生在转化细胞中。可以用多种孵育条件来形成本文公开的肽。在一些实施方案中,可以希望使用模拟生理条件的条件。产生本文所述的微生物可以涉及使用多种重组核酸构建体,如克隆载体、表达载体或合成操纵子。克隆载体和表达载体都包含允许该载体在一种或多种适宜的重组微生物中复制的核苷酸序列。在克隆载体中,此序列通常是这样的序列,其使得该载体能够独立于重组微生物染色体复制,且还包含复制起点或自主复制序列。本领域中使用多种细菌和病毒复制起点。通常,在这方面,任何适于维持、繁殖或表达多核苷酸来在重组微生物中表达多肽的载体都可以用于表达。举几个例子,本领域中使用染色体、附加体和病毒衍生的载体,包括衍生自细菌质粒、噬菌体、酵母附加体、酵母染色体元件、病毒(如杆状病毒、papovirus(如sv40)、牛痘病毒、腺病毒、禽痘病毒、假狂犬病毒和反转录病毒)的载体,及衍生自其组合的载体,如衍生自质粒和噬菌体遗传元件的那些,如黏粒和噬粒。通过多种公知和常规的技术将适当的dna序列插入该载体。通常,通过用一种或多种限制性核酸内切酶切割dna序列和表达载体,然后用例如t4-dna连接酶将限制片段连接在一起,来将用于表达的dna序列与表达载体连接。连接通过常规技术达到,如soepcr、dna合成、平端连接或限制酶位点处连接。如果没有适宜的限制位点可用,则可以使用合成的寡核苷酸衔接物或接头。使用gibson等(gibson,d.g.等naturemethods(2009)6,5,343-345;在此以其整体引入作为参考)所述的外切核酸酶、聚合酶固定化dna聚合酶的组合,通过体外重组,可以达到将包含大量基因的多个序列组装为单个核酸分子。许多启动子在细菌细胞中具有功能,并已在文献中描述,包括组成型、诱导型、发育调控型和环境调控型启动子。尤其有利的是使用在适当的重组微生物中具有功能的启动子(也称为转录起始区)。例如,如果用大肠杆菌作为重组微生物,则可以使用的示例性启动子包括但不限于:λ噬菌体pl启动子,大肠杆菌lac、trp和tac启动子,sv40早期和晚期启动子,反转录病毒lrt的启动子,或camv35s启动子。编码本文所述的一种或多种酶的多核苷酸序列也可以是表达盒的一部分,该表达盒最少包含启动子、编码本文公开的酶的一种或多种核酸和在重组微生物中具有功能的转录终止信号序列。根据需要,启动子可以是任何类型的启动子。例如,诱导型启动子或组成型启动子和表达盒可以进一步包含有效连接的能够指导所产生的蛋白质转运的靶向序列或易位或分泌肽编码区。表达盒还可以进一步包含编码选择标记的核苷酸序列。在一些实施方案中,使用能够在两种或多种不同宿主物种中繁殖的穿梭载体。示例性穿梭载体能够在大肠杆菌和/或枯草芽孢杆菌中及在专性厌氧菌(如梭菌属)中复制。酶插入穿梭载体后,可以将该穿梭载体引入大肠杆菌宿主细胞进行该载体的扩增和选择。还可以为了获得甲基化载体的目的而将穿梭载体(例如包含编码酶的一种或多种序列的穿梭载体)引入含有甲基转移酶的宿主细胞(例如表达甲基转移酶的大肠杆菌宿主细胞)。然后可以分离载体,并引入所选择的微生物来表达该一种或多种酶。可以使用任何适宜的穿梭载体或质粒,如本公开下文中所述的穿梭质粒中的任一种。在一些实施方案中,本文所述的任何细胞都引入了单个载体,例如包含编码酶的一种或多种核酸序列的穿梭质粒dna分子。也可以使用双质粒系统。可以使用携带编码酶的一种或多种序列的两种不同质粒。两种不同质粒中的每一种可以携带不同的选择标记。在一些实施方案中,该一种或多种质粒稳定转化在厌氧细胞中。为了产生烷烃和/或烯烃,可以希望使用异源核酸序列在其中稳定维持的重组微生物。在这方面,可以使用复制型质粒。这种质粒是具有复制起点的质粒,该复制起点由宿主细胞识别,并允许质粒作为稳定、自主的染色体外元件复制,在细胞分裂为子细胞期间分割。在一些实施方案中,使编码一种或多种酶的一种或多种序列整合入所选择的微生物的基因组。整合可以与表达盒一起进行。在一些实施方案中,整合入宿主基因组是稳定整合。核酸整合入基因组可以用转座酶进行。该宿主基因组可以是细菌染色体。在编码酶的一种或多种核酸序列整合入宿主基因组的实施方案中,可为此目的而使用一种或多种转座元件。作为实例,可将2类转座元件(其可编码转座酶)包含在载体中,并包含编码本文公开的一种或多种酶的一种或多种编码序列。来自从细菌至人的多种生物的转座酶是已知的。人转座酶的实例是转座酶thap9,其包含thap型锌指。大多数转座酶分类在ec编号ec2.7.7之下。常用且经过检验的转座酶是见于细菌如大肠杆菌中的转座子tn10的转座酶。该转座子包含五个基因,包括转座酶和赋予对抗生素四环素的抗性的两种蛋白质。tn10的转座酶例如包含在质粒pwh1(a31v)-mrfp或质粒pxt107中。适宜的转座酶的另一实例是见于大肠杆菌和希瓦氏菌属(shewanella)中的细菌转座子tn5的转座酶(ec:3.1.-.-)。该转座子编码转座酶tn5和转座抑制物两种蛋白质。转座酶tn5是rna酶蛋白质超家族的成员,该家族包括反转录病毒整合酶。该转座子编码对卡那霉素和其他氨基糖苷类抗生素的抗生素抗性。转座子tn5例如包含在质粒po86a1中。另一个实例是见于原核生物中的tn3转座子的tn3转座酶。该转座子编码β-内酰胺酶(赋予对β-内酰胺抗生素的抗性的酶)、tn3转座酶和tn3解离酶。可以使用的转座酶的另一个实例是tc1/mariner型转座酶,如睡美人(sleepingbeauty)转座子系统。睡美人转座子系统是设计用于将确定的dna序列精确引入脊椎动物染色体的合成dna转座子。它包含睡美人(sb)转座酶。许多sb转座子已发展为非病毒载体,尤其用于将基因引入脊椎动物基因组。可以利用的另外两个基于转座子的系统是例如国际专利申请wo2003/100070中所述的青蛙王子(frogprince)转座子/转座酶系统或ttaa特异性转座子piggybac系统。另一实例是来自大肠杆菌的转座子tnia的转座酶。tnia的转座酶包含在质粒pec279、pec54和peh4h中。见于大肠杆菌中的另一实例——转座子tn1000转座酶,也称为“转座子γ-δ的转座酶”——包含在例如大肠杆菌f质粒中。其他实例可以是来自大肠杆菌的is186a转座酶、rep相关酪氨酸转座酶或insh转座酶。整合入基因组可以与正选择结合进行,例如通过提供编码重组宿主(例如细菌)存活所需的具体特性或功能的核酸分子,重组宿主依赖于由引入该生物的dna编码的具体功能的存在或缺乏。该特性或功能可以在于对特定抗生素的抗性,也参见上文。作为说明性实例,在整合入宿主基因组(例如染色体dna)时,抗性标记可以表达并因此具有活性,使得可以针对携带整合核酸的宿主细胞(也称为“整合体”)进行正选择。在一些实施方案中,在包含在载体中并因此未整合入基因组时,该抗性标记可以不表达并因此无活性。该载体(例如质粒)可以包含能够紧密结合抗生素抗性基因的启动子的阻抑蛋白。因此,在包含在载体中时,该抗性标记无活性。一旦例如通过转座插入细菌基因组,即不存在阻抑蛋白能够结合的操纵基因序列。抗性标记因此在宿主基因组中具有功能。测定酶的表达根据需要,可以通过检测核酸或蛋白质的量或在酶的情况下通过检测酶活性来验证蛋白质的表达。在本文公开的方法背景中,任何可用的方法都可用来检测核酸或蛋白质的存在。这种方法可以包括本领域公知的已建立的标准方法。这种技术的实例包括但不限于rt-pcr、rna酶保护测定、northern分析、western分析、elisa、放射免疫测定或荧光滴定测定。对于酶,可以用酶活性来评估蛋白质的存在或估计所存在的蛋白质的量。在可获得可以引入细胞的染料时,可以用该染料在细胞中体内进行该活性测试。该染料的检测可以用荧光显微术进行,包括比例荧光显微术。能够处理从生物学制备至最终数据的整个测定的集成光学成像系统可购得。具有多种生理学上重要的特性的灵敏的荧光指示剂是可得的,荧光试剂可以用来灵敏地和特异地表征蛋白质、核苷酸、离子和脂质的胞内分布。在比例荧光显微术中,采集两张荧光图像,将目的参数定量为一张图像中的荧光与另一张图像中的荧光的比值。通过顺次用两种不同波长的光激发样品并顺次采集两张不同的图像,通过用单个波长的光激发样品并采集形成自两种不同发射波长的光的图像,或通过用两个波长激发样品并采集两个波长的发射,可以采集荧光比值图像。细胞中蛋白质的量的评估可以包括评估细胞中编码该蛋白质的核酸(例如rna)的量。本文公开的方法可进一步包括测量编码genbank检索号wp_003113506.1或其同源物的序列和/或genbank检索号np_010827.3或其同源物的序列的序列的表达。这可以例如通过测定从处于所选择的启动子控制下的编码核酸分子转录的rna分子的数目来达到。本领域常用的方法是用反转录酶将rna拷贝为cdna,然后将该cdna分子与荧光染料偶联。该分析可以例如以dna微阵列的形式进行。许多这种服务和试剂盒可购得,例如来自affymetrix的genechip表达阵列。测定转录因子的基因表达的其他手段包括但不限于寡核苷酸阵列和定量实时聚合酶链反应(rt-pcr)。可以通过任意常用的杂交方法用核酸探针来探测样品,以检测蛋白质的核酸分子的量。为了获得核酸探针,可以进行化学合成。合成的核酸探针可以首先用作利用适当的模板、基本按照标准pcr流程、按照公认的pcr技术进行的聚合酶链反应(pcr)中的引物,以获得该探针。本领域技术人员能够容易地根据可获得的生物标志序列设计这种探针。杂交探针可以通过标准标记技术标记,如放射性标记、酶标记、荧光标记、生物素-抗生物素蛋白标记、化学发光或纳米颗粒。杂交后,可以用标准技术显示探针。最常用于测定核酸浓度的方法包括通过放射自显影、荧光、化学发光或生物发光以及电化学和电学技术检测。另一种适宜的技术是国际专利申请wo2009/041917和wo2008/097190(二者都在此以其整体引入作为参考)中公开的靶核酸分子的电学检测。在有冲突的情况下,将以本说明书(包括定义)为准。本领域中完善的用于特异性检测所选择的核酸的技术基于核酸结合配偶体和靶核酸之前的杂交。通常,将该核酸结合配偶体固定在固体支持物上,然后利用上文提到的检测方法之一。在一些实施方案中,测定目的基因的表达水平包括测定转录为mrna的水平。编码目的蛋白质的rna可以用任何可用的扩增技术扩增,如聚合酶链反应(pcr),包括多重pcr、嵌套式pcr和扩增阻滞突变特异性(arms)pcr(也称为等位基因特异性pcr(as-pcr))、滚环扩增(rca)、基于核酸序列的扩增(nasba)、连接酶链反应(lcr)、qb复制酶链反应、环介导等温扩增(lamp)、转录介导扩增(tma)和链置换扩增(sda)(包括基因组链置换扩增(wgsda)、多链置换扩增(msda)和基因特异性链置换扩增(gs-msda))。所获得的扩增产物的检测可以以本领域已知的许多方式进行。实例包括但不限于电泳方法,如琼脂糖凝胶电泳结合染色,如溴化乙锭染色。在其他实施方案中,本文公开的方法伴随着实时检测,如实时pcr。在这些实施方案中,监测扩增过程的时程。本领域常用的实时检测手段涉及在扩增过程之前加入染料。这种染料的实例是荧光染料sybrgreen,其仅在结合于双链核酸时发射荧光信号。作为说明性实例,可以用实时pcr来测定编码目的蛋白质(如酶)的rna的水平。这种pcr方法在实时检测下进行,使得扩增过程的时程得到监测。pcr的特征在于靶序列的对数扩增。对于rna的扩增,使用反转录酶pcr。检测蛋白质的表达所需的引物和探针的设计在本领域普通从业人员的技术范围之内。在一些实施方案中,在无rna酶条件下从样品分离rna,然后通过使用反转录酶转化为dna。反转录可以在rt-pcr分析之前进行或在单个反应容器内同时进行。rt-pcr探针是具有附着于5’端的荧光部分(也称为报道染料)和偶联于3’端的猝灭剂部分(或反之)的寡核苷酸。这些探针通常设计用于杂交至pcr产物的内部区域。在非杂交状态下,荧光分子和猝灭分子的靠近阻止了来自探针的荧光信号的检测。在pcr扩增期间,在聚合酶复制rt-pcr探针结合于其上的模板时,聚合酶的5'-3'核酸酶活性切割探针。荧光部分和猝灭部分由此分隔开。然后荧光在每个循环中以与探针切割的量成比例的方式增加。从反应发射的荧光信号可以用可购得的设备用常规和惯常的技术随时间推移进行测量或跟踪。所评价的样品中生物标志rna的定量可以通过将扩增信号与一条或多条标准曲线(其中以类似的方式评价已知量的rna)的扩增信号相比较来进行。在一些实施方案中,将生物标志表达的差异测量为达到阈值荧光的pcr循环时间的差异或“dct”。用重组微生物产生烯烃使用本文所述的重组微生物,可以产生一种或多种烯烃和/或烷烃产物。一种或多种烯烃和/或烷烃的产生可以涉及提供可通过该微生物转化为乙酰coa的底物。如上文所解释,乙酰coa是中心代谢基石。取决于所使用的微生物,多种底物可以是潜在适宜的。在一些实施方案中,该产生可以涉及提供包含co的底物。在一些实施方案中,可以提供包含co2的底物。底物可以是气态的,且包含某个量的co和/或co2。可以例如提供包含25%(v/v)或更多一氧化碳的底物。还可以提供包含25%(v/v)或更多二氧化碳的底物。在一些实施方案中,可以例如提供包含35%(v/v)或更多一氧化碳的底物。在一些实施方案中,可以例如提供包含35%(v/v)或更多二氧化碳的底物。co可以是包含在合成气(也称为合成气体,其是基本由氢气和一氧化碳组成的气体混合物)中的成分。在一些实施方案中,co和co2都包含在合成气中。本文公开的产生烷烃和/或烯烃的方法通常涉及在co和/或co2存在下(例如在包含co和co2的大气下)、在适宜的培养基中、在适宜的条件下培养非天然微生物。在一些实施方案中,本文所述的培养非天然微生物可以在基本厌氧的培养条件下进行。培养基可以包含足以允许所使用的微生物生长的维生素可矿物质。适合用于用co发酵的厌氧培养基为本领域已知。产生一种或多种烯烃和/或烷烃可以包括发酵过程。该发酵过程可以在例如在生物反应器中在水性培养基中进行。用培养基在生物反应器中培养是本领域中完善的技术。一氧化碳和/或二氧化碳(包括含有一氧化碳和/或二氧化碳的底物)以及培养基可以以连续、分批或补料分批方式饲提供至该生物反应器。所产生的烷烃或烯烃可以由重组微生物分泌入周围培养基。该烷烃或烯烃因此可以从发酵液回收。在该烷烃或烯烃为气态的实施方案中,一部分烷烃或烯烃可以溶解在培养基中。作为实例,丁二烯在水中具有约7.4g/l的溶解度。丁烯在水中具有约2.2g/l的溶解度,而丙烯在水中具有约0.6g/l的溶解度。在该烷烃或烯烃为气态时,通常从培养基上方的气相收集该产物。所产生的烷烃或烯烃可以用本领域已知的任何适宜的分离和/或纯化技术分离和富集(包括纯化)。在一些实施方案中,所产生的烷烃或烯烃为气态,如正丁烯或1,3-丁二烯。在一些实施方案中,所产生的烷烃或烯烃为液体,如异戊二烯。在气态产物的情况下,该烷烃或烯烃可以通过滤器、气体分离膜和/或气体纯化器如吸附柱,然后保存在一个或多个气瓶中。在一些实施方案中,可以利用蒸馏来纯化产物气体。在一些实施方案中,可以利用气-液萃取(脱气(stripping))。在液体产物的情况下,该烷烃或烯烃的分离可以包括离心。还可以通过用有机相萃取来富集产物。可以通过向发酵液加入去乳化剂和/或成核剂来从发酵液分离含有烷烃和/或烯烃的该有机相。去乳化剂的说明性实例包括絮凝剂和促凝剂。成核剂的说明性实例包括乙酰coa衍生化合物本身和有机溶剂(如十二烷、肉豆蔻酸异丙酯和油酸甲酯)的小液滴。在一些实施方案中,含有乙酰coa衍生化合物的有机相自发从发酵液分开。纯化所获得的烷烃或烯烃的方法还可以涉及其他标准技术,如超滤,及一种或多种层析技术。本说明书中对此前公开的文件的罗列或讨论不应必然视为承认该文件是现有技术的部分或是普通常识。本文说明性描述的发明可在缺乏本文未明确公开的任意一个或多个元件、一个或多个限制的情况下实施。此外,本文所用的术语和表述作为描述性术语而非限制性术语使用,在这类术语和表述的使用中不存在排除所显示和描述的特征或其部分的任何等同物的意图,而应理解,在所要求的发明的范围内,多种修改是可能的。因此,应理解,虽然已通过示例性实施方案和可选特征明确公开了本发明,但本领域技术人员可以求助于本文公开的包含于其中的发明的修改和变动,认为这类修改和变动也在本发明的范围之内。本文宽泛和概括地描述了本发明。落在此类属公开之内的每一较窄的类别和亚属分组也形成本发明的部分。这包括具有从该类属去除任意主题的条件或阴性限制的本发明的类属描述,无论所删除的材料是否在本文中明确引用。其他实施方案在所附权利要求书内。此外,在按照markush组描述本发明的特征和方面时,本领域技术人员应理解,因此还按照markush组的单个成员或成员亚组描述本发明。为了本发明可易于理解和实施,现将通过以下非限制性实施例描述具体实施方案。实施例实施例阐述可以用于本文公开的方法和用于获得上述重组微生物的技术。实施例1:产生甲烷、乙烷、乙烯、丙烷、丙烯、正丁烷、1-丁烯、2-甲基丙烯、正戊烷、1-戊烯、正己烷和1-己烯用来自棕色固氮菌(azotobactervinelandii)ca的基因通过标准重组dna和分子克隆技术进行表达质粒sg323(及本文所述的所有其他质粒)的构建。图4d显示质粒sg323(seqidno:4),其包含改造的钼固氮酶途径。通过以下流程将此质粒电穿孔入杨氏梭菌:杨氏梭菌细胞的遗传转化电感受态(electrocompetent)杨氏梭菌细胞的制备:制备杨氏梭菌电感受态细胞的方法修改自之前报道的流程(,m.等procnatlacadsciusa.(2010)107,29,13087-13092)。除离心外的所有操作均在厌氧培养室中在冰上进行。所有缓冲液和离心管均在使用前进行冰预冷和无氧处理。所有塑料器皿均在使用前至少24小时放入厌氧培养室,以消除任何残余氧气。感受态细胞-80℃维持在含10%dmso的smp缓冲液(270mm蔗糖、1mmmgcl2、7mm磷酸钠、ph6)中直至使用。从新鲜接种自冻存物,然后在ytf液体培养基中转移两次的培养物制备电感受态杨氏梭菌细胞。制备感受态细胞前约15至16个小时,将中至晚对数期培养物转入两个血清瓶,血清瓶包含100ml补充了40mmdl-苏氨酸的新鲜ytf培养基(最终600nm光密度[od500]=0.004)。37℃过夜生长后,通过4℃、10,000转/分钟离心10分钟来收集早对数期细胞(od600=0.2至0.3;200ml)。用200mlsmp洗涤缓冲液洗涤细胞两次,并按1010至1011细胞/ml的终浓度悬浮在相同缓冲液中。向感受态细胞加入最终悬液体积的五分之一的防冻缓冲液(60%dmso-40%smp,ph6),以达到10%dmso的终浓度。得到的感受态细胞(25μl/管)保存在-80℃供以后使用。这些冷冻感受态细胞的感受性保持稳定约1个月。用于杨氏梭菌的电转化方法:所有流程都在厌氧培养室内进行。将电感受态杨氏梭菌细胞(25μl)于冰上快速从-80℃冰箱转移至厌氧培养室。在冰上融化后(约1分钟),将细胞与1至5μgdna混合,并转移至预冷的0.1cm间隙的小杯。通过使用微生物电穿孔系统,以600ohm电阻和25μf电容在0.625kv脉冲处理细胞。脉冲处理后立即用0.5ml新鲜ytf培养基回收细胞,转移至含有10ml补充了5mml-半胱氨酸(ph7)的ytf培养基的压力管,并在37℃孵育。使电穿孔的细胞在37℃恢复,直至其细胞密度高于电穿孔后即刻(约9至12小时)的细胞密度。将5ml体积的增生(outgrowth)培养物或适当稀释的培养物与20ml含有适当抗生素的ytf融化的琼脂(1.5%)混合,并倒入培养皿。琼脂混合物固化后,将平板倒置孵育。2-3天后,菌落变得可见,并可以接种在液体培养基中。ytf培养基:10g/l酵母提取物;16g/l细菌培养用胰蛋白胨;4g/l氯化钠;5g/l果糖;补充2mml-半胱氨酸;最终ph为6.0。用该遗传改造的含有质粒sg323的杨氏梭菌细胞进行以电极提供电子,通过气体发酵来从合成气(co、co2和h2的混合物)和电子产生甲烷、乙烷、乙烯、丙烷、丙烯、正丁烷、1-丁烯、2-甲基丙烯、正戊烷、1-戊烯、正己烷和1-己烯。下文描述所使用的材料和方法:所使用的含气体的生长培养基:痕量元素:将氨三乙酸加至水中,并用koh调节至ph6.0。加入其余成分。wolfe’s维生素溶液还原剂:l-半胱氨酸(自由碱)4.0g蒸馏水100.0ml所用气体:60%co、10%co2、30%h2。在提供1bar压力的合成气(60%co、10%co2、30%h2)和具有供电电源的电极的cstr生物反应器中,在补充10μg/ml甲砜霉素(抗生素)的以上培养基中37℃培养转化的细胞。在细胞密度达到0.3od600时,通过10mmiptg诱导基因表达。通过gc-ms(shimadzugc-msqp5050a)进行产物分析。用500illhamilton气密注射器进行顶空气体取样。甲烷、乙烷、乙烯、丙烷、丙烯、正丁烷、1-丁烯、2-甲基丙烯、正戊烷、1-戊烯、正己烷和1-己烯的产生可通过gc-ms检测。备选实施方案:质粒sg193(图4a和seqidno:1)、质粒sg211(图4b和seqidno:2)和质粒sg278(图4c和seqidno:3)也用于改造杨氏梭菌以及clostridiumautoethanogenum细胞来产生甲烷、乙烷、乙烯、丙烷、丙烯、正丁烷、1-丁烯、2-甲基丙烯、正戊烷、1-戊烯、正己烷。进行与优选实施方案中所述相同的基本流程和方法。使用下下文列出的不同底物:对于所使用的所有底物,及后续实施例的那些:60%co、10%co2、30%h2100%co30%co2和60%h260%co、10%co2、30%h2和电子100%co和电子100%co2和电子30%co2和60%h2和电子实施例2:梭菌中经转座酶发生的染色体整合和多拷贝染色体整合的正选择以下实施例阐述使异源dna(代谢基因簇或任何其他功能性或非功能性核酸序列)整合入给定宿主或基质(chassis)微生物的方法。在重组细菌的存活依赖于引入该生物的dna编码的特定功能的存在或缺乏时,可能进行突变细菌的正选择(或直接遗传选择)。选择方法优于筛选方法的优势是,含有希望得到的具体突变的细菌的生长极大地超过缺乏该具体突变的细菌,从而便于鉴定优选的突变体。由于转座所需的所有功能(如转座酶和侧翼为转座识别碱基序列的整合盒)是在质粒上引入,通过作为整合盒的一部分的抗生素选择标记的直接正选择是不可能的。抗性标记在质粒上具有功能,因此(大多数质粒还具有高拷贝数)未从整合事件产生生长优势。但是,在标记仅在整合在染色体dna中时才表达并因此具有活性时,整合体的正选择是可能的。图2a提供用于通过切割和粘贴转座酶20来将核酸序列整合入细菌染色体的基本系统的概览。所有必要功能都定位在单个质粒上。主链抗生素抗性标记32和用于繁殖该载体的复制起点30在转化的细菌细胞中。整合盒侧翼为转座酶识别位点22和28;整合盒还包含第二抗生素抗性基因。转座酶20在两个识别位点处切割dna,并将此核酸序列整合入细菌染色体。在定位在质粒上时,紧密结合抗生素基因启动子的阻抑蛋白失活抗性标记的表达。转座后,阻抑蛋白结合的操纵基因序列丧失,并赋予抗性。此原理更详细地显示在图2b中。图3a显示在图2a的最小系统之上实施的正选择系统。在此处,附加的阻抑蛋白32结合操纵基因位点34,阻断抗生素抗性基因38的表达。相应的抗生素抗性蛋白质不表达,不赋予抗性。图3b显示通过转座酶的作用将该盒引入细菌染色体44的情况。操纵位点34不再存在,抗生素抗性基因38的表达产生抗生素抗性。因此可能选择插入染色体突变体。5’itr(转座酶识别和切割的反向末端重复)和lox66位点也是红霉素/克拉霉素抗生素抗性标记mlsr的启动子,参见图2b。阻抑蛋白(laci)结合lac操纵基因序列,并阻断转录(这还可以通过加入iptg来逆转)。转座后,lac操纵基因序列丧失,mlsr得以表达,并可针对其进行选择(rbs=核糖体结合位点)。令人惊奇地,该方法还便于多拷贝整合的产生和高载量转座。可以达到整合体的正选择和发生的转座事件的延时二者。用于在梭菌中通过转座酶进行随机染色体整合的初始系统通过测试包含转座所必需的一切(侧翼为转座识别碱基序列(也称为反向末端重复)的含有抗生素抗性标记和bsfbfp的整合盒,及处于木糖诱导型启动子控制下的转座酶)的四种不同的大肠杆菌-梭菌穿梭质粒,和仅由大肠杆菌-梭菌穿梭质粒组成而无附加功能的阴性对照,确定了厌氧荧光蛋白质(bsfbfp)的染色体整合。测试了野生型转座酶himar1以及高活性变体himar1c9。对于丙酮丁醇梭菌的遗传转化,通过质粒pant体内甲基化该质粒。所使用的载体是seqidno:9至14。它们的载体图谱可从图9至图14取得。丙酮丁醇梭菌的转化按照以下流程进行:1.质粒dna的甲基化在大肠杆菌xl1bluemr中与质粒pant(φ3t甲基转移酶)一起转化质粒含抗生素的平板:氨苄青霉素、氯霉素和四环素midipreperation(dna的浓度不应太低)2.含2xytg培养基的5ml起子液体培养物丙酮丁醇梭菌在晚上,用50-200μl预培养物接种50ml2xytg培养基,并在36℃过夜培养。od600=0.8-1时收集细胞2200g离心细胞10分钟(4℃)用40ml冰冷的heb+缓冲液洗涤细胞2200xg离心细胞10分钟(4℃)用heb+缓冲液洗涤细胞三次悬浮在2mlheb缓冲液中;600μl(一次转化)分装使用冷的4mm电穿孔杯和至多4μl质粒(600μl细胞)冰上孵育2分钟2500v电穿孔加入1.4ml2xytg培养基,并在36℃孵育4小时接种在含适当抗生素(例如克拉霉素(cla)或甲砜霉素(tm))的选择平板上36℃孵育3天观察菌落并接种液体培养基3.培养基2xytg:5g/l葡萄糖16g/l胰蛋白胨10g/l酵母提取物5g/lnaclph7.0heb+缓冲液:7mmhepes272mm蔗糖5mmmgcl2ph7.4不含mgcl2的heb缓冲液所使用的甲基化质粒的浓度:pant甲基化的质粒bg00132:350ng/μlpant甲基化的质粒bg00133:300ng/μlpant甲基化的质粒bg00134:280ng/μlpant甲基化的质粒bg00135:385ng/μlpant甲基化的质粒bg00136:500ng/μl所有构建体的电穿孔都成功,每块平板上观察到了10至50个菌落。整合按以下进行:将每个转化(1、2、3、4、5)的4个菌落接种在如下新鲜培养基(5ml)中:培养基1=2xytg5g/l葡萄糖培养基2=2xytgx5g/l葡萄糖+2.5g/l木糖培养基3=2xytg5g/l葡萄糖+10g/l木糖观察:含有质粒bg132、bg134、bg135的预培养物比其他培养物生长更快。将2μl各预培养物接种在含有以下的2xyt平板上:-5g/l葡萄糖-5g/l葡萄糖,含2.5g/l木糖-10g/l木糖将22个不同菌落涂布培养(struckout)在以下平板上(第1轮):1.2xytg平板2.含5g/l葡萄糖和2.5g/l木糖的2xytgx3.含10g/l木糖的2xytx将22个菌落涂布培养如下(第2轮):1.2xytg平板2.含5g/l葡萄糖和2.5g/l木糖的2xytgx3.含10g/l木糖的2xytx将从平板取得的22个菌落涂布培养如下(第3轮):1.2xytg平板2.含5g/l葡萄糖和2.5g/l木糖的2xytgx3.含10g/l木糖的2xytx将从平板取得的22个菌落涂布培养如下(第4轮):1.不含抗生素的2xytg平板2.含10μg/ml克拉霉素(cla)的2xytg平板3.含20μg/ml甲砜霉素(tm)的2xytg平板检查第4轮的平板:总结:整合体可以分离,对照全为阴性,但与大量所测试的克隆相比,整合体非常少。许多克隆仍未丢失质粒。整合基因座的测定按以下进行:用来自macherey-nagel(用于难以裂解的细菌的特殊流程)的试剂盒制备丙酮丁醇梭菌整合菌株(参见表格)的基因组dna,使用了约20ml液体培养物。通过nanodrop测定dna浓度。整合体1=470ng/μl整合体2=460ng/μl整合体3=520ng/μl整合体4=710ng/μl整合体5=380ng/μl整合体6=460ng/μl整合体7=440ng/μl整合体8=450ng/μl整合体9=420ng/μl整合体10=400ng/μl整合体11=570ng/μl整合体12=370ng/μl样品寄送至lgcgenomicsgmbh(柏林,德国)进行pcr步移测定基因座。进行附加的pcr来验证整合体:聚合酶链反应(pcr)整合检验pcr1:测试荧光蛋白质bsfbfp的存在模板:整合体(丙酮丁醇梭菌)的基因组dna和野生型gdna,阳性对照载体bg132引物bg272正向:gcacttcctcttgttggaaat(seqidno:58)引物bg273反向:acttgtgcaagtccacttaaa(seqidno:59)pcr2:检验整合标记模板:整合体(丙酮丁醇梭菌)的基因组dna和野生型gdna,阳性对照载体bg132引物bg274正向:ctatgtggcgcggtattatc(seqidno:60)bg275反向:gcatttaagcgtcagagcatgg(seqidno:61)pcr3:检验整合标记2模板:整合体(丙酮丁醇梭菌)的基因组dna和野生型gdna,阳性对照载体bg132引物bg276正向:tttaatcgtggaatacgggtttg(seqidno:62)引物bg277反向:gtgagctattcactttaggtttagg(seqidno:63)pcr4:检验主链模板:整合体(丙酮丁醇梭菌)的基因组dna和野生型gdna,阳性对照载体bg132引物bg278正向:caaaaggccaggaaccgtaa(seqidno:64)引物bg279反向:gcgtcagaccccgtagaaaa(seqidno:65)pcr5:检验主链2模板:整合体(丙酮丁醇梭菌)的基因组dna和野生型gdna,阳性对照载体bg132引物bg280正向:attgtaaaccgccattcagag(seqidno:66)引物bg281反向:ataccgttgcgtatcactttc(seqidno:67)以30个循环、1分钟延伸、55℃退火温度、50μl总体积、phusion聚合酶进行pcr1-5。然后通过0.8%分析型dna琼脂糖凝胶电泳检查pcr产物。结果显示在图15a和15b中。凝胶1,图15a:各上样10μl;位置、名称、结果如下:泳道1:3μl购自roth(carlrothgmbh&co.kg、karlsruhe、德国)的1kb梯泳道2-6:整合体1pcr1-5;结果:1和3阳性,2、4、5阴性泳道7-11:整合体2pcr1-5;结果:1和3阳性,2、4、5阴性泳道12:3μl购自roth的100bp梯泳道13-17:整合体3pcr1-5;1和3阳性,2、4、5阴性泳道18-22:整合体4pcr1-5;1和3阳性,2、4、5阴性泳道23-27:整合体5pcr1-5;1和3阳性,2、4、5阴性泳道28:3μl购自roth的1kb梯凝胶2,图15b:各上样10μl;位置、名称、结果如下:泳道1:3μl购自roth的1kb梯泳道2-6:整合体6pcr1-5;结果:1和3阳性,2、4、5阴性泳道7-11:整合体7pcr1-5;结果:1和3阳性,2、4、5阴性泳道12-16:整合体8pcr1-5;1和3阳性,2、4、5阴性泳道17:3μl购自roth的100bp梯泳道18-22:野生型pcr1-5;全阴性泳道23-27:整合体5pcr1-5;1,3,4,5阳性,2阴性泳道28:3μl购自roth的1kb梯基因组步移-来自lgcgenomics的第一批结果。读出结果(第一个数字对应于上文的整合体)可从seqidno:68至78取得。初步分析表:结果:lgcgenomics进行的pcr基因组步移取得成功,但并非所有基因座都可测定出。lgc用优化的设置进行了第二次运行。整合体的荧光显微术在leica荧光显微镜上分析了整合体。丙酮丁醇梭菌整合体1的图像显示在图16a、16b和16c中,放大倍数为1000x。丙酮丁醇梭菌整合体2的图像显示在图16d中。丙酮丁醇梭菌整合体10的图像显示在图16e、16f和16g中。丙酮丁醇梭菌野生型对照的图像显示在图16h中。检查的所有整合体都显示荧光。野生型对照为阴性。基因组步移-由lgcgenomics进行的第二轮pcr基因组步移给出的读出结果可从seqidno:79至85取得。实施例3:为梭菌中通过转座酶进行的随机染色体整合和随机多拷贝染色体整合开发正选择系统为了进一步改进初始的、艰难的染色体整合系统,构建了具有阻断整合标记的转录的阻抑蛋白的新质粒。随后,为了测试新系统,将以下大肠杆菌-丙酮丁醇梭菌穿梭载体转化入丙酮丁醇梭菌。为了将质粒转化入丙酮丁醇梭菌,首先借助上文所述的pant质粒在体内对它进行甲基化。为了达到这一点,将质粒转化入大肠杆菌xl1-blue细胞(参见上文)。载体bg133(缩写为“n”)(来自实施例2的表现最好的载体,见上文)具有序列seqidno:11。载体bg168(缩写为“t”,参见图17)具有序列seqidno:15。温度敏感型质粒bg182(参见图18)具有序列seqidno:16。制备质粒后,可以通过以上流程用例如质粒bg168遗传转化丙酮丁醇梭菌:在2xytg培养基(16g/l胰蛋白胨、10g/l酵母提取物、5g/lnacl、5g/l葡萄糖)中厌氧培养丙酮丁醇梭菌,直至光密度达到0.6至0.8。所有后续步骤也都在厌氧条件下进行。2000g离心细胞10分钟(4℃),然后用电穿孔缓冲液(7mmhepes、270mm蔗糖、5mmmgcl2、ph7.4)洗涤。在重复此处理两次。然后,准备含有5kg质粒dna、500μl感受态细胞的冷电转杯,并在2500v电穿孔3ms以上。用新鲜2xytg培养基再生2小时后,将细胞接种在补充适宜抗生素的2xytg琼脂平板上。2天后,遗传转化的菌落变得可见。通过正选择系统分离染色体整合体:培养细胞,加入木糖诱导转座酶,并加入第二抗生素。两次系列转移后,再次平板接种细胞,然后挑选进行单个测试。通过丧失对第一(主链)标记(其是甲砜霉素)的抗性同时保留对第二(整合)标记克拉霉素的抗性来鉴定整合体。可以分离出超过100个整合体。仅进一步分析了一部分。用来自macherey-nagel的试剂盒(用于难以裂解的细菌的特殊流程)进行丙酮丁醇梭菌整合菌株的基因组dna的制备;使用了约20ml液体培养物。通过系列转移比较染色体整合(正选择)系列转移方案:图例:n=质粒n(见上文)i=质粒i(见上文)v=液体培养物(副本)7ml体积#=数目cla=克拉霉素整合体的分离:在液体培养物中系列转移后,将5μl各副本平板接种在2xytg上。然后将28个菌落涂布培养在以下平板上:1.不含抗生素的2xytg2.含5μg/ml克拉霉素的2xytg3.含20μg/ml甲砜霉素的2xytg在比较平板时,所有来自“n”系列的克隆都显示甲砜霉素抗性(主链标记),意味着质粒仍在。但是,所有来自“i”系列的克隆都只具有克拉霉素抗性(整合标记)而没有甲砜霉素抗性(主链标记),意味着它们全都是染色体整合体。结果总结在图62中所示的表格中。随后,按图63中所示制备gdna(流程见上文)。基因座的测定:lgcgenomics进行的第一轮pcr基因组步移给出了以下结果:不可测定:6、8、10、11、12、13、14、15、17、18、20、21、22、23、24、25、26、27、28、29、32、35guent01:4-羟基-3-甲基丁-2-烯基二磷酸还原酶/s1rna结合结构域蛋白质1、2、3、4、5guent07:pploop-atpase7、9guent16:保守膜蛋白,可能为转运蛋白,ypaa枯草芽孢杆菌直向同源物16guent19:未表征的蛋白质,degv家族19guent30:含纤连蛋白iii型结构域的蛋白质30、31guent34:金黄色葡萄球菌金黄亚种(staphylococcusaureussubsp.aureus)st228质粒pi5s534所获得的读出结果如下:>guent_07_gw2bk_261112.0.10具有序列seqidno:86;>guent01.gw2bk_261112.0.2具有序列seqidno:87;>guent_19_gw2bk_261112.0.15具有序列seqidno:88;>guent_30_gw2bk_261112.0.18具有序列seqidno:89;>guent_34_gw2bk_261112.0.21具有序列seqidno:90;>guent_16_gw2bk_261112.0.14具有序列seqidno:91。第二轮基因座测定给出了图64中所示的结果。几乎所有基因座都测定出。结果也非常一致。用最高选择压力产生的4个克隆中有一个显示多拷贝整合。实施例4:用于通过转座酶进行的随机染色体整合和随机多拷贝染色体整合的正选择系统在杨氏梭菌和clostridiumautoethanogenum中的应用将以下具有与实施例2中所示相同的正选择系统(见上文)的质粒用于杨氏梭菌和clostridiumautoethanogenum:具有序列seqidno:17的载体bg282(图19);具有序列seqidno:18的载体bg281(图20);具有序列seqidno:19的载体bg287(图21);具有序列seqidno:20的载体bg288(图22);具有序列seqidno:21的载体bg289(图23);具有序列seqidno:22的载体bg290(图24);具有序列seqidno:23的载体bg291(图25);具有序列seqidno:24的载体bg292(图26);具有序列seqidno:25的载体bg178(图27)。质粒bg281例如用于将大的甲羟戊酸生物合成基因簇整合入杨氏梭菌的染色体。作为另一个实例,质粒bg282用于将略微不同的mva途径整合入clostridiumautoethanogenum的染色体。杨氏梭菌的遗传转化按照leang,l.等,applenvironmicrobiol.(2013)79,4,1102-1109进行转化。用大肠杆菌bl21替代neb表达细胞,用ytf培养基替代petc培养基。流程如下:电感受态杨氏梭菌细胞的制备除离心外的所有操作均在厌氧培养室中在冰上进行。所有缓冲液和离心管均在使用前进行冰预冷和无氧处理。所有塑料器皿均在使用前至少24小时放入厌氧培养室,以消除任何残余氧气。感受态细胞-80℃维持在含10%dmso的smp缓冲液(270mm蔗糖、1mmmgcl2、7mm磷酸钠、ph6)中直至使用。从新鲜接种自冻存物,然后在petc液体培养基中转移两次的培养物制备电感受态杨氏梭菌细胞。可以用ytf培养基替换petc培养基而不显著损失转化效率。制备感受态细胞前约15至16个小时,将中至晚对数期培养物转入两个血清瓶,血清瓶包含100ml补充了40mmdl-苏氨酸的新鲜petc培养基(最终600nm光密度[od500]=0.004)。37℃过夜生长后,通过4℃、10,000转/分钟离心10分钟来收集早对数期细胞(od600=0.2至0.3;200ml)。用200mlsmp洗涤缓冲液洗涤细胞两次,并按1010至1011细胞/ml的终浓度悬浮在相同缓冲液中。向感受态细胞加入最终悬液体积的五分之一的防冻缓冲液(60%dmso-40%smp,ph6),以达到10%dmso的终浓度。得到的感受态细胞(25μl/管)保存在-80℃供以后使用。这些冷冻感受态细胞的感受性保持稳定约1个月。用于杨氏梭菌的电转化方法所有流程都在厌氧培养室内进行。将电感受态杨氏梭菌细胞(25μl)于冰上快速从-80℃冰箱转移至厌氧培养室。在冰上融化后(约1分钟),将细胞与1至5μgdna混合,并转移至预冷的0.1cm间隙的genepulser小杯(bio-rad)。通过使用genepulserxcell微生物电穿孔系统,以600ohm电阻和25μf电容在0.625kv脉冲处理细胞。脉冲处理后立即用0.5ml新鲜petc培养基回收细胞,转移至含有10ml补充了5mml-半胱氨酸(ph7)的petc培养基的压力管,并在37℃孵育。使电穿孔的细胞在37℃恢复,直至其细胞密度高于电穿孔后即刻(约9至12小时)的细胞密度。将5ml体积的增生培养物或适当稀释的培养物与20ml含有适当抗生素的rcm融化的琼脂(1.5%)混合,并倒入培养皿。琼脂混合物固化后,将平板倒置孵育。所转化的质粒:编号名称(序列)1bg282(seqidno:17)2bg281(seqidno:18)3bg287(seqidno:19)4bg288(seqidno:20)5bg289(seqidno:21)6bg290(seqidno:22)7bg291(seqidno:23)8bg292(seqidno:24)9bg178(seqidno:25)除bg178(9)外的所有质粒都可以转化入杨氏梭菌和clostridiumautoethanogenum。杨氏梭菌中的染色体整合用含5μg/ml克拉霉素的ytx(5g/l木糖)在液体中一式三份培养转化体(transformand)。用新鲜培养基(含5g/l木糖和5μg/ml克拉霉素的ytx)进行3次系列转移。然后,将5μl点在不含抗生素的ytg平板上。每块平板挑选4个菌落,同时接种在含5μg/ml克拉霉素的ytg平板、含5μg/ml甲砜霉素的ytg平板和不含抗生素的平板上。培养整合体(对克拉霉素具有抗性但对甲砜霉素无抗性的那些)进行基因组dna的制备。用来自macherey-nagel的试剂盒(用于难以裂解的细菌的特殊流程)进行杨氏梭菌整合菌株基因组dna的制备。使用了约10ml液体培养物。通过nanodrop测定gdna的浓度。结果显示在图65的表格中。整合体的pcr验证使用以下引物(扩增整合盒),以退火温度55℃、延伸时间5分钟和30个循环进行假定的整合体的gdna上的pcr:正向引物:ctgtctcttatacacatctgctgataagtccccggtc(seqidno:92)。反向引物:ctgtctcttatacacatctgctgataagtccccggtc(seqidno:93)。然后通过0.8%分析型dna琼脂糖凝胶电泳来检查pcr产物。结果可从图28取得。泳道1:3μl购自roth的1kb梯;泳道2-9:整合体lbi105-111pcr;结果:4、5、6、7、8阳性,1、2、3阴性;泳道10:3μl购自roth的1kb梯;泳道11-18:整合体lbi112-120pcr;结果:1、2、5、7、8阳性,3、4、6阴性;泳道19:3μl购自roth的1kb梯;泳道24-25:阳性对照pcr;结果:1、2阳性;泳道26-27:阴性对照gdna杨氏梭菌gdnapcr;阴性;泳道28-29:阴性对照gdnac.autoethanogenumgdnapcr;阴性;泳道30:3μl购自roth的1kb梯。分析:至少一些整合体的整合盒可以通过pcr扩增。这显示,借助正选择系统,在这种情况下可以整合至多10kb的负载。clostridiumautoethanogenum中的染色体整合选择单个构建体bg289进行整合。在含5μg/ml克拉霉素的ytx(5g/l木糖)中培养bg289的转化体。用新鲜培养基(含5g/l木糖和5μg/ml克拉霉素的ytx)进行3次系列转移。然后,将5μl点在不含抗生素的ytg平板上。挑选10个菌落,同时接种在含5μg/ml克拉霉素的ytg平板、含5μg/ml甲砜霉素的ytg平板和不含抗生素的平板上。这产生4个整合体。培养整合体(对克拉霉素具有抗性但对甲砜霉素无抗性的那些)进行基因组dna的制备。用来自macherey-nagel的试剂盒(用于难以裂解的细菌的特殊流程)进行clostridiumautoethanogenum整合体基因组dna的制备。使用了约10ml液体培养物。通过nanodrop测定gdna的浓度。整合体的pcr验证使用以下引物(扩增整合盒中的酶),以退火温度55℃、延伸时间60秒和30个循环进行假定的整合体的gdna上的pcr:正向引物:gggttgccttactggttagseqidno:94。反向引物:gcagtatcggttcggtaatcseqidno:95。然后通过0.8%分析型dna琼脂糖凝胶电泳来检查pcr产物。结果显示在图29中。泳道1:3μl购自roth的1kb梯;泳道2-5:整合体a1-a4pcr;结果:3阳性,1、2、4阴性;泳道6-7:阳性对照pcr;结果:1、2阳性;泳道8-9:异性对照gdna杨氏梭菌gdnapcr;阴性;泳道10-11:阴性对照gdnac.autoethanogenumgdnapcr,阴性;泳道12:3μl购自roth的1kb梯。分析:可以从gdna样品3扩增出所整合的酶。这表明该系统对梭菌属内很宽的宿主范围有用。实施例5:2-甲基-1,3-丁二烯的合成图48和图49分别显示质粒sg726(seqidno:44)和sg705(seqidno:45),其各包含编码2-甲基-1,3-丁二烯途径的酶的序列(对于基因,还参见参考数字)。图49显示质粒sg705(seqidno:45),其包含编码2-甲基-1,3-丁二烯途径的酶的序列(对于基因,还参见参考数字)。按实施例1中所述(还参见实施例3)进行电感受态杨氏梭菌细胞的制备和电转化。按实施例1中所述进行2-甲基-1,3-丁二烯产生。可以通过上文所述gc-ms来检测2-甲基-1,3-丁二烯产生。这两个质粒还用于在染色体上改造杨氏梭菌和clostridiumautoethanogenum细胞来产生2-甲基-1,3-丁二烯。进行实施例1中所述的流程和方法。此外,还成功地将co及co2和co的混合物、以及co和电子和/或co2和电子用作底物。实施例6:2-甲基-1,3-丁二烯的合成图6和图5a分别显示质粒sg156(seqidno:7)和sg123(seqidno:5),其各包含编码2-甲基-1,3-丁二烯途径的酶的序列(对于基因,还参见参考数字)。图5b和图7分别显示质粒sg124(seqidno:6)和sg157(seqidno:8),其各包含编码进一步改造的2-甲基-1,3-丁二烯途径的酶的序列(对于基因,还参见参考数字)。按实施例1中所述(还参见实施例3)进行电感受态杨氏梭菌细胞的制备和电转化。按实施例1中所述用100%co2进行2-甲基-1,3-丁二烯产生。图8a和8b显示用shimadzugc-msqp5050a通过gc-ms进行的产物分析。通过500μlhamilton气密注射器采集样品。收集500μl顶空,并上样在rxi-5ms柱30m、0,25mmid和0,25μmdf(restek)上,使用-1的分割。在30℃等度进行分离(注射温度60℃;以m/z=67记录sim)。图谱显示与2-甲基-1,3-丁二烯的完美匹配。这四个质粒还用于在染色体上改造杨氏梭菌和clostridiumautoethanogenum细胞来产生2-甲基-1,3-丁二烯。进行实施例1中所述的流程和方法。还成功地将100%co2及50%co2和50%co的混合物用作底物。实施例7:1,3-丁二烯的合成图50显示质粒sg714(seqidno:46),其包含编码改造的1,3-丁二烯途径的酶的序列(对于基因,还参见参考数字)。图51显示质粒sg739(seqidno:47),其包含编码第二改造的1,3-丁二烯途径的酶的序列(对于基因,还参见参考数字)。按实施例1中所述(还参见实施例3)进行电感受态杨氏梭菌细胞的制备和电转化。按实施例1中所述进行1,3-丁二烯产生。可以通过上文所述gc-ms来检测1,3-丁二烯产生。这两个质粒还用于在染色体上改造杨氏梭菌和clostridiumautoethanogenum细胞来产生1,3-丁二烯。进行与实施例1中所述相同的流程和方法。此外,还成功地将co及co2和co的混合物、以及co和电子和/或co2和电子用作底物。实施例8:1,3-丁二烯的合成图34显示质粒sg387(seqidno:30),其包含编码改造的1,3-丁二烯途径的酶的序列(对于基因,还参见参考数字)。图35显示质粒sg411(seqidno:31),其包含编码第二改造的1,3-丁二烯途径的酶的序列(对于基因,还参见参考数字)。用来自以下的基因通过标准重组dna和分子克隆技术进行表达质粒的构建:丙酮丁醇梭菌atcc824、clostridiumcarboxidivorans、大肠杆菌、真氧产碱杆菌h16、豚鼠气单胞菌(aeromonascaviae)、链霉菌属物种菌株cl190、酿酒酵母s288c、枯草芽孢杆菌、杨氏梭菌dsm13528dsm13528、大肠杆菌bl21(de3)、梭菌属物种(clostridiumsp.)m62/1、肉毒梭菌(clostridiumbotulinum)bkt015925、克氏梭菌dsm555、发酵氨基酸球菌dsm20731、丙酸梭菌、橙色绿屈挠菌(chloroflexusaurantiacus)j-10-fl、流感嗜血杆菌r2866、恶臭假单胞菌kt2440、齿垢密螺旋体(treponemadenticola)atcc35405、thaueralinaloolentis和分枝杆菌属jdm601。按实施例1中所述(还参见实施例3)进行电感受态杨氏梭菌细胞的制备和电转化。对于质粒sg714,按实施例1中所述用100%co2进行1,3-丁二烯产生。对于质粒sg739,按实施例1中所述用30%co2和60%h2进行1,3-丁二烯产生。两种情况下都可以通过上文所述gc-ms来检测1,3-丁二烯产生。这两个质粒还用于在染色体上改造杨氏梭菌和clostridiumautoethanogenum细胞来产生1,3-丁二烯。进行与实施例1中所述相同的流程和方法。此外,还成功地将co及co2和co的混合物、以及co和电子和/或co2和电子用作底物。实施例9:丙烯的合成图36显示质粒sg455(seqidno:32),其包含编码改造的丙烯途径的酶的序列(对于基因,还参见参考数字)。图52显示质粒sg755(seqidno:48),其包含编码另一改造的丙烯途径的酶的序列。图53显示质粒sg779(seqidno:49),其包含编码另一改造的丙烯途径的酶的序列。图37显示质粒sg479(seqidno:33),其包含编码另一改造的丙烯途径的酶的序列。用来自以下的基因通过标准重组dna和分子克隆技术进行表达质粒的构建:丙酮丁醇梭菌atcc824、clostridiumcarboxidivorans、大肠杆菌、真氧产碱杆菌h16、豚鼠气单胞菌、链霉菌属物种菌株cl190、酿酒酵母s288c、枯草芽孢杆菌、杨氏梭菌dsm13528dsm13528、大肠杆菌bl21(de3)、梭菌属物种m62/1、肉毒梭菌bkt015925、克氏梭菌dsm555、发酵氨基酸球菌dsm20731、丙酸梭菌、橙色绿屈挠菌j-10-fl、流感嗜血杆菌r2866、恶臭假单胞菌kt2440、齿垢密螺旋体atcc35405、小家鼠(musmusculus)、勤奋生金球菌dsm5348和金黄色葡萄球菌。按实施例1中所述进行电感受态杨氏梭菌dsm13528细胞的制备和电转化。按实施例1中所述进行丙烯产生。可以通过上文所述gc-ms来检测丙烯产生。这三个质粒还用于改造clostridiumautoethanogenumdsm10061和醋酸梭菌细胞来产生丙烯。进行与实施例1中所述相同的流程和方法。此外,还成功地将co及co2和co的混合物、以及co和电子和/或co2和电子用作底物。实施例10:1-丁烯的合成图38显示质粒sg539(seqidno:34),其包含编码改造的1-丁烯途径的酶的序列(对于基因,还参见参考数字)。图39显示质粒sg523(seqidno:35),其包含编码另一改造的1-丁烯途径的酶的序列。图54显示质粒sg839(seqidno:50),其包含编码另一改造的1-丁烯途径的酶的序列。图55显示质粒sg823(seqidno:51),其包含编码另一改造的1-丁烯途径的酶的序列。用来自以下的基因通过标准重组dna和分子克隆技术进行表达质粒的构建:丙酮丁醇梭菌atcc824、clostridiumcarboxidivorans、大肠杆菌、真氧产碱杆菌h16、豚鼠气单胞菌、链霉菌属物种菌株cl190、酿酒酵母s288c、枯草芽孢杆菌、杨氏梭菌dsm13528dsm13528、大肠杆菌bl21(de3)、梭菌属物种m62/1、肉毒梭菌bkt015925、克氏梭菌dsm555、发酵氨基酸球菌dsm20731、丙酸梭菌、橙色绿屈挠菌j-10-fl、流感嗜血杆菌r2866、恶臭假单胞菌kt2440、齿垢密螺旋体atcc35405、小家鼠、勤奋生金球菌dsm5348和金黄色葡萄球菌。按实施例1中所述(还参见实施例3)进行电感受态c.autoethanogenumdsm10061细胞的制备和电转化。按实施例1中所述进行1-丁烯产生。可以通过上文所述gc-ms来检测1-丁烯产生。这三个质粒还用于改造杨氏梭菌dsm13528和醋酸梭菌细胞来产生1-丁烯。进行与实施例1中所述相同的流程和方法。此外,还成功地将co及co2和co的混合物、以及co和电子和/或co2和电子用作底物。实施例11:异丁烯(2-甲基丙烯)的合成图41显示质粒sg601(seqidno:37),其包含编码改造的异丁烯途径的酶的序列(对于基因,还参见参考数字)。图40显示质粒sg582(seqidno:36),其包含编码另一改造的异丁烯途径的酶的序列(对于基因,还参见参考数字)。图56显示质粒sg882(seqidno:52),其包含编码另一改造的异丁烯途径的酶的序列(对于基因,还参见参考数字)。图57显示质粒sg901(seqidno:53),其包含编码另一改造的异丁烯途径的酶的序列(对于基因,还参见参考数字)。用来自以下的基因通过标准重组dna和分子克隆技术进行表达质粒的构建:丙酮丁醇梭菌atcc824、clostridiumcarboxidivorans、大肠杆菌、真氧产碱杆菌h16、豚鼠气单胞菌、链霉菌属物种菌株cl190、酿酒酵母s288c、枯草芽孢杆菌、杨氏梭菌dsm13528dsm13528、大肠杆菌bl21(de3)、梭菌属物种m62/1、肉毒梭菌bkt015925、克氏梭菌dsm555、发酵氨基酸球菌dsm20731、丙酸梭菌、橙色绿屈挠菌j-10-fl、流感嗜血杆菌r2866、恶臭假单胞菌kt2440、齿垢密螺旋体atcc35405、小家鼠、勤奋生金球菌dsm5348和金黄色葡萄球菌。按实施例1中所述(还参见实施例3)进行电感受态杨氏梭菌dsm13528细胞的制备和电转化。按实施例1中所述用30%co2和60%h2进行异丁烯产生。可以通过上文所述gc-ms来检测异丁烯产生。图67显示在agilent5977egcmsd上测量的所获得的产物的gs-ms分析。所使用的柱子是restekrt-aluminabond/mapd,30mx0.32mm;df5μm。载气:氦气。sim56m/z。使用gerstel-multipurposesampler2xl-xt的500μl顶空注射。图谱显示与异丁烯(2-甲基丙烯)的完美匹配。这两个质粒还用于改造clostridiumautoethanogenumdsm10061和醋酸梭菌细胞来产生乙烯。进行与实施例1中所述相同的流程和方法。此外,还成功地将co及co2和co的混合物、以及co和电子和/或co2和电子用作底物。实施例12:1-戊烯的合成图42和图43分别显示质粒sg498(seqidno:38)和sg513(seqidno:39),其各包含编码改造的1-戊烯途径的酶的序列(对于基因,还参见参考数字)。图58和图59分别显示质粒sg798(seqidno:54)和sg813(seqidno:55),其各包含编码另一改造的1-戊烯途径的酶的序列。用来自以下的基因通过标准重组dna和分子克隆技术进行表达质粒的构建:丙酮丁醇梭菌atcc824、clostridiumcarboxidivorans、大肠杆菌、真氧产碱杆菌h16、豚鼠气单胞菌、链霉菌属物种菌株cl190、酿酒酵母s288c、枯草芽孢杆菌、杨氏梭菌dsm13528dsm13528、大肠杆菌bl21(de3)、梭菌属物种m62/1、肉毒梭菌bkt015925、克氏梭菌dsm555、发酵氨基酸球菌dsm20731、丙酸梭菌、橙色绿屈挠菌j-10-fl、流感嗜血杆菌r2866、恶臭假单胞菌kt2440、齿垢密螺旋体atcc35405、小家鼠、勤奋生金球菌dsm5348和金黄色葡萄球菌。按实施例1中所述进行电感受态clostridiumautoethanogenumdsm10061细胞的制备和电转化。按实施例1中所述用100%co2进行1-戊烯产生。可以通过上文所述gc-ms来检测1-戊烯产生。这两个质粒还用于改造杨氏梭菌dsm13528和醋酸梭菌细胞来产生1-戊烯。进行与实施例1中所述相同的流程和方法。此外,还成功地将co及co2和co的混合物、以及co和电子和/或co2和电子用作底物。实施例13:乙烷的合成图44、图45、图60和图61分别显示质粒sg557(seqidno:40)、sg598(seqidno:41)、sg857(seqidno:56)和sg898(seqidno:57),其各包含编码改造的乙烯途径的酶的序列(对于基因,还参见参考数字)。用来自以下的基因通过标准重组dna和分子克隆技术进行表达质粒的构建:丙酮丁醇梭菌atcc824、clostridiumcarboxidivorans、大肠杆菌、真氧产碱杆菌h16、豚鼠气单胞菌、链霉菌属物种菌株cl190、酿酒酵母s288c、枯草芽孢杆菌、杨氏梭菌dsm13528dsm13528、大肠杆菌bl21(de3)、梭菌属物种m62/1、肉毒梭菌bkt015925、克氏梭菌dsm555、发酵氨基酸球菌dsm20731、丙酸梭菌、橙色绿屈挠菌j-10-fl、流感嗜血杆菌r2866、恶臭假单胞菌kt2440、齿垢密螺旋体atcc35405、小家鼠、勤奋生金球菌dsm5348和金黄色葡萄球菌。按实施例1中所述进行电感受态杨氏梭菌dsm13528细胞的制备和电转化。按实施例1中所述用30%co2和60%h2进行乙烯产生。可以通过上文所述gc-ms来检测乙烯产生。这两个质粒还用于改造clostridiumautoethanogenumdsm10061和醋酸梭菌细胞来产生乙烯。进行与实施例1中所述相同的流程和方法。此外,还成功地将co及co2和co的混合物、以及co和电子和/或co2和电子用作底物。本文提到的所有出版物在此引用作为参考,以公开和描述与这些出版物所被引用的方法和/或材料有关的方法和/或材料,就如同明确地和单独地说明引入每一单个出版物(包括专利申请)作为参考一样。在有冲突的情况下,将以本说明书(包括定义)为准。本文中讨论的出版物仅针对其在本申请提交日期之前的公开内容提供。不应将本文的任何内容解释为承认本发明没有资格由于优先发明而先于这种出版物。此外,本文提供的出版日期可以不同于实际出版日期,实际公开日期可需要独立确认。除非另有明确说明,本文给出的任何方法或方面绝无解释为需要以特定顺序进行其步骤的意图。因此,在权利要求书或说明书中未针对所要求保护的方法明确陈述步骤限于特定顺序时,绝无在任何方面暗示其顺序的意图。这适用于任何可能的非明确解释基础,包括关于步骤排列或操作流程的逻辑问题、源于语法组织或标点符号的明显意义、或本说明书中所述方面的数目或类型。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1