赋予鞘翅目和半翅目害虫抗性的COPI外被体GAMMA亚单位核酸分子的制作方法
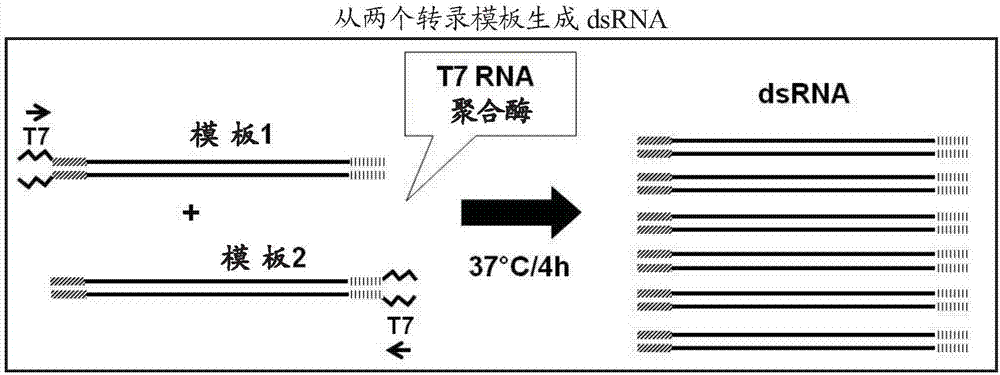
优先权声明本申请要求2014年10月13日提交的题为“copicoatomergammasubunitnucleicacidmoleculesthatconferresistancetocoleopteranandhemipteranpests”的美国临时专利申请系列号62/063192的申请日的利益,在此将其公开全体并入本文。公开领域本发明一般性地涉及由昆虫害虫(例如鞘翅目害虫和半翅目害虫)引起的植物损害的遗传控制。在特定的实施方案中,本发明涉及目标编码和非编码多核苷酸的鉴定、及其重组dna技术在昆虫害虫细胞中转录后阻遏或抑制目标编码多核苷酸和非编码多核苷酸表达从而提供植保作用的用途。背景西方玉米根虫(wcr)(diabroticavirgiferavirgiferaleconte,玉米根萤叶甲)是北美的一种破坏性最大的玉米根虫物种,在美国中西部玉米种植区受到特别关注。北方玉米根虫(ncr)(巴氏根萤叶甲)是与wcr共栖大致相同范围的近缘物种。有几种叶甲属的其他相关亚种是美洲的严重害虫:墨西哥玉米根虫(mcr)[墨西哥玉米根萤叶甲(d.virgiferazeaekrysanandsmith)];南方玉米根虫(scr)(十一星根萤叶甲);黄瓜条根萤叶甲(d.balteataleconte);d.undecimpunctatatenella;南美叶甲(d.speciosagermar)、和d.u.undecimpunctatamannerheim。美国农业部估计玉米根虫每年造成10亿美元收入损失,包括8亿美元产量损失和2亿美元处理成本。wcr和ncr两者都以卵的形式在夏季期间潜伏在土壤中。这些昆虫在整个冬季都停留在卵阶段。卵是椭圆的、白色、且长度小于0.004英寸。幼虫在5月下旬或6月上旬孵出,卵孵化的精确时间由于温度差异和位置而在各年间有所变化。新孵出的幼虫是白色的蠕虫,长度小于0.125英寸。一旦孵出,幼虫便开始以玉米根为食。玉米根虫经历三个幼虫龄。在进食几周后,幼虫蜕皮,进入若虫阶段。它们在土壤中化蛹,然后它们在7月和8月中以成虫从土壤出现。成体根虫长度约0.25英寸。玉米根虫幼虫在玉米和几种其他禾本科物种上完成发育。在黄色狗尾草上饲养的幼虫较晚出现,并且作为成虫比玉米上饲养的幼虫具有更小的头壳尺寸。ellsbury等人,(2005)environ.entomol.34:627-634。wcr成虫以玉米穗丝、花粉、和暴露的穗尖上的籽粒为食。如果wcr成虫在存在玉米生殖组织之前出现,则它们可以以叶组织为食,由此减缓植物生长,且偶而杀死宿主植物。然而,当优选的穗丝和花粉变得可得到时,成虫会快速向其移动。ncr成虫也以玉米植物的生殖组织为食,但相比之下很少以玉米叶为食。玉米中的大部分根虫损伤由幼虫进食引起。新孵出的根虫最初以细的玉米根毛为食,并钻入根尖中。随着幼虫长得更大,它们以初生根为食并钻入其中。在有大量玉米根虫时,幼虫啮食往往导致根部被一直修剪到玉米杆基部。严重的根损伤干扰根将水和养分转运到植物中的能力,降低植物生长,并导致籽粒产生减少,由此经常急剧降低总产量。严重的根损伤还经常导致玉米植物的倒伏,其使收获变得更难,并进一步降低产量。此外,成虫以玉米生殖组织为食可以导致穗尖的穗丝修剪。如果这种“穗丝剪切”在花粉脱落期间足够严重,则传粉可能受到破坏。可以通过作物轮作、化学杀虫剂、生物杀虫剂(例如,孢子形成革兰氏阳性细菌苏云金芽孢杆菌(bt))、表达bt毒素的转基因植物、或这些手段的组合来尝试控制玉米根虫。作物轮作的重大缺陷是过度限制了农田的用途。此外,一些根虫物种可以在玉米之外的作物田间产卵,或者长期的滞育(diapause)可导致卵孵化经历多年,因此会降低用玉米和大豆实施的作物轮作的效率。化学杀虫剂是最为人们所倚重的实现玉米根虫控制的手段。然而,使用化学杀虫剂并不是完美的玉米根虫控制策略;如果把化学杀虫剂的成本与使用杀虫剂后仍可能发生的根虫害所致的损失相加,则美国每年由于玉米根虫可能损失超过10亿美元。大的幼虫群体、大雨、和不适当的杀虫剂应用均可导致玉米根虫控制不充分。此外,杀虫剂的连续使用可能选择杀虫剂抗性根虫品系,并且由于杀虫剂中许多对非靶物种有毒性,有严重的影响环境之虞。椿象和其他半翅目昆虫(heteroptera)构成了另一大类重要的农业害虫。已知全世界有超过50个椿象的近缘物种引起作物损害。mcpherson&mcpherson(2000)stinkbugsofeconomicimportanceinamericanorthofmexicocrcpress。这些昆虫孳生于许多重要作物,包括玉米、大豆、水果、蔬菜、和谷物。椿象在达到成体期之前要经历多个蛹期。从卵发育到成虫的时间约为30-40天。若虫和成体均以来自软组织的汁液为食,它们还将消化酶注入到软组织中,引起口外组织的消化和坏死。然后摄入消化的植物材料和养分。植物维管系统水分和养分的耗竭导致植物组织破坏。对于籽粒和种子发育的损害最为显著,因为产量和萌发显著减少。在温暖的气候下会出现多个世代,导致显著的虫害压力。当前的椿象管理依赖于单块田的杀虫剂处理。因此,迫切需要替代的管理策略,以将正在发生的作物损失降低到最低限度。rna干扰(rnai)是一种利用内源细胞途径的方法,由此对足够大小的整个或任何部分靶基因序列特异性的干扰rna(irna)分子(例如,双链rna(dsrna)分子)导致由其编码的mrna的降解。在最近几年,在许多物种和实验系统,例如线虫秀丽隐杆线虫、植物、昆虫胚胎、和组织培养物中的细胞中,已经使用rnai进行基因“敲低”。参见,例如fire等人,(1998)nature391:806-811;martinez等人,(2002)cell110:563-574;mcmanus和sharp(2002)naturerev.genetics3:737-747。rnai通过包括切丁酶(dicer)蛋白质复合物的内源途径完成mrna的降解。切丁酶将长的dsrna分子切割成约20个核苷酸的短片段,称作小干扰rna(sirna)。sirna解旋成两个单链rna:乘客链(passengerstrand)和引导链(guidestrand)。乘客链被降解,引导链则被整合到rna诱导的沉默复合物(risc)中。美国专利7,612,194和美国专利公开文本2007/0050860、2010/0192265、和2011/0154545披露了自玉米根萤叶甲若虫分离的9112种表达序列标签(est)序列的文库。在美国专利7,612,194和美国专利公开号2007/0050860中提出将与其中披露的玉米根萤叶甲液泡型h+-atp酶(v-atp酶)的几个特定部分序列之一互补的核酸分子与启动子可操作连接,以在植物细胞中表达反义rna。美国专利公开文本no.2010/0192265提出将启动子与核酸分子可操作连接以在植物细胞中表达反义rna,所述核酸分子与具有未知且未公开的功能的玉米根萤叶甲基因的特定部分序列互补(该部分序列据称与秀丽隐杆线虫中的c56c10.3基因产物58%相同)。美国专利公开文本no.2011/0154545提出将启动子与核酸分子可操作连接以在植物细胞中表达反义rna,所述核酸分子与玉米根萤叶甲外被体(coatomer)gamma亚单位基因的两个特定部分序列互补。此外,美国专利7,943,819公开了自玉米根萤叶甲幼虫、若虫、和切开的中肠分离的906种表达序列标签(est)序列的文库,并且提出了将启动子与核酸分子可操作连接以在植物细胞中表达双链rna,所述核酸分子与玉米根萤叶带电的多泡体蛋白4b基因的特定部分序列互补。除了v-atp酶的几个特定部分序列和未知功能的基因的特定部分序列之外,在美国专利7,612,194和美国专利公开文本2007/0050860、2010/0192265和2011/0154545中没有进一步提出使用其中列出的超过9000个序列中的任何特定序列进行rna干扰。此外,美国专利7,612,194、以及美国专利公开文本2007/0050860、2010/0192265、和2011/0154545都没有教示提供的超过9000个序列中哪些其他序列在用作dsrna或sirna时在玉米根虫物种中会是致命的、甚至没有教示其有任何其他方面的用处。除了带电多泡体蛋白4b基因的特定部分序列之外,美国专利7,943,819没有提出使用其文中的超过900个序列的任何特定序列进行rna干扰。此外,美国专利7,943,819没有教示其提供的超过900个序列中哪些其他序列在用作dsrna或sirna时在玉米根虫物种中会是致命的、甚至没有教示其有任何其他方面的用处。美国专利申请公开文本u.s.2013/040173和pct申请公开文本wo2013/169923描述了源自玉米根萤叶甲snf7基因的序列在玉米中进行rna干扰的用途。(还公开于bolognesi等人,(2012)plosone7(10):e47534.doi:10.1371/journal.pone.0047534)。与玉米根虫dna互补的绝大多数序列(如上述)用作dsrna或sirna时不提供对玉米根虫物种的植保作用。例如,baum等人(2007,naturebiotechnology25:1322-1326)描述了rnai对几个wcr基因靶的抑制作用。这些作者报告,在超过520ng/cm2的极高irna(例如,dsrna)浓度下,他们测试的26个靶基因中有8个不能提供实验上显著的鞘翅目害虫死亡率。美国专利7,612,194和美国专利申请公开2007/0050860的作者们首次报道了在玉米植株中靶向西方玉米根虫的植物体内(inplanta)rnai。baumetal.(2007)nat.biotechnol.25(11):1322-6。这些作者们描述了一种高通量体内饵食rnai系统,用来筛选用于开发转基因rnai玉米的潜在靶基因。在290种靶物的初始基因池中,只有14种基因展现了幼虫控制潜力。最有效的双链rna(dsrna)中之一靶向了编码囊泡性atp酶亚单位a(v-atpase)的基因,导致相应的内源mrna的迅速阻遏,并以低浓度的dsrna引发了特异性的rnai应答。由此,这些作者首次记录了植物体内rnai作为害虫管理工具的潜力,同时证明,有效的靶物是无法先验地准确鉴定的,即使从较小的一组候选基因鉴定亦然。公开概要本文中公开了用于控制昆虫害虫的核酸分子(例如,靶基因、dna、dsrna、sirna、mirna、shrna、和hprna)及其使用方法,所述昆虫害虫包括,例如,鞘翅目害虫,诸如玉米根萤叶甲(d.virgiferaleconte)(西方玉米根虫,“wcr”);巴氏根萤叶甲(d.barberismithandlawrence,北方玉米根虫,“ncr”);十一星根萤叶甲(d.u.howardibarber;南方玉米根虫,“scr”);墨西哥玉米根萤叶甲(d.v.zeaekrysanandsmith,墨西哥玉米根虫,“mcr”);黄瓜条根萤叶甲(d.balteataleconte);d.u.tenella;南美叶甲(d.speciosagermar);d.u.undecimpunctatamannerheim;和半翅目害虫,诸如英雄美洲蝽(euschistusheros(fabr.)(新热带棕椿象,“bsb”)、褐椿象(e.servus)(say)(棕椿象);稻绿蝽(nezaraviridula(l.)(南部绿椿象)、盖德拟壁蝽(piezodorusguildinii(westwood)(红带椿象)、和茶翅蝽(halyomorphahylys(棕翅椿象)、chinaviahilare(say)(绿椿象);c.marginatum(palisotdebeauvois);dichelopsmelacanthus(dallas);d.furcatus(f.);edessameditabunda(f.);thyantaperditor(f.)(新热带红肩椿象);horciasnobilellus(berg)(棉红蝽);taediastigmosa(berg);dysdercusperuvianus(guérin-méneville);neomegalotomusparvus(westwood);leptoglossuszonatus(dallas);niesthreasidae(f.);lygushesperus(knight)(西方牧草盲蝽);和牧草盲蝽[l.lineolaris(palisotdebeauvois)]。在具体的实例中,公开了示例性核酸分子,其与昆虫害虫中的一个或多个天然核酸序列的至少一部分可以同源。在这些和进一步的实例中,天然核酸可以是靶基因,其产物可以,例如但不限于:涉及代谢过程;或者涉及幼虫/若虫发育。在一些实例中,借助于包含与靶基因同源的多核苷酸的核酸分子,靶基因表达的翻译后抑制在鞘翅目和/或半翅目害虫中可以是致命的,或导致其生长和/或发育的降低。在具体的实例中,可选择由外壳蛋白复合物gamma亚单位(本文中称为copigamma亚单位和copigamma)构成的基因作为转录后沉默的靶基因。在具体实例中,有用于转录后抑制的靶基因是本文称为copigamma的新基因。因此本文中公开了包含:copigamma(seqidno:1和seqidno:87)、copigamma(seqidno:1和seqidno:87)的互补物、以及前述任一者的片段的分离的核酸分子。还公开了包含编码与靶基因产物内的氨基酸序列(例如本文中称为copigamma的基因的产物)至少约85%相同的多肽的多核苷酸的核酸分子。例如,核酸分子可以包含编码与seqidno:2或seqidno:88(copigamma蛋白)至少85%相同的多肽的多核苷酸。在具体的实例中,核酸分子包含的核苷酸序列编码与copigamma的产物内的氨基酸序列至少85%相同的多肽。进一步公开了包含编码这样的多核苷酸的核酸分子,所述多核苷酸是编码与靶基因产物内的氨基酸序列至少85%相同的多肽的多核苷酸的反向互补物。还公开了用于产生irna(例如,dsrna、sirna、mirna、shrna、和hprna)分子的cdna多核苷酸,所述irna与鞘翅目和/或半翅目害虫靶基因(例如copigamma)的全部或部分互补。在特定的实施方案中,dsrna、sirna、mirna、shrna、和/或hprna可以在体外产生,或由遗传修饰的生物(如植物或细菌)在体内产生。在特定的实例中,公开了可用于产生与copigamma(seqidno:1和seqidno:87)的全部或部分互补的irna分子。进一步公开了用于抑制鞘翅目和/或半翅目害虫中必需基因的表达的手段、以及用于给植物提供鞘翅目和/或半翅目害虫抗性的手段。用于抑制鞘翅目和/或半翅目害虫中必需基因表达的手段是由下述至少一者构成的单链或双链rna分子:seqidno:3(叶甲属copigamma区域1,本文中有时称作copigammareg1)或seqidno:4(叶甲属copigamma区域2,本文中有时称作copigammareg2),或seqidno:5(叶甲属copigamma区域3,本文中有时称作copigammareg3),或seqidno:75(叶甲属copigamma版本1,本文中有时称作copigammaver1),或seqidno:76(叶甲属copigamma版本2,本文中有时称作copigammaver2),或seqidno:77(叶甲属copigamma版本3,本文中有时称作copigammaver3),或seqidno:78(叶甲属copigamma版本4,本文中有时称作copigammaver4),或seqidno:89(英雄美洲蝽copigamma区域2,本文中有时称作bsb_copigamma-2),或它们的互补物。用于抑制鞘翅目和/或半翅目害虫中的必需基因表达的手段的功能等同物包括与包含seqidno:1或seqidno:87的wcr或bsb基因的全部或部分基本上同源的单链或双链rna分子。给植物提供鞘翅目和/或半翅目害虫抗性的手段是这样的dna分子,其包含与启动子可操作连接的、编码用于抑制鞘翅目和/或半翅目害虫中的必需基因的表达的手段的核苷酸序列,其中所述dna分子能够整合到玉米或大豆植物的基因组中。公开了控制昆虫害虫(例如鞘翅目和/或半翅目害虫)群体的方法,所述方法包括向昆虫害虫(例如鞘翅目和/或半翅目害虫)提供irna(例如,dsrna、sirna、shrna、mirna、和hprna)分子,所述irna分子在被所述害虫摄取之后发挥作用而抑制该害虫体内的生物功能,其中所述irna分子包含选自以下的核苷酸序列的全部或部分:seqidno:1、seqidno:3、seqidno:4、seqidno:5、seqidno:75、seqidno:76、seqidno:77、seqidno:78、seqidno:87和seqidno:89;seqidno:1、seqidno:3、seqidno:4、seqidno:5、seqidno:75、seqidno:76、seqidno:77、seqidno:78、seqidno:87和seqidno:89的互补物;叶甲属生物(例如wsr)或半翅目生物(例如bsb)的包含seqidno:1、seqidno:3、seqidno:4、seqidno:5、seqidno:75、seqidno:76、seqidno:77、seqidno:78、seqidno:87和seqidno:89中任一者之全部或部分的天然编码序列;叶甲属生物或半翅目生物的包含seqidno:1、seqidno:3、seqidno:4、seqidno:5、seqidno:75、seqidno:76、seqidno:77、seqidno:78、seqidno:87和seqidno:89中任一者之全部或部分的天然编码序列的互补物;叶甲属生物或半翅目生物的转录为包含seqidno:1、seqidno:3、seqidno:4、seqidno:5、seqidno:75、seqidno:76、seqidno:77、seqidno:78、seqidno:87和seqidno:89中任一者之全部或部分的天然rna分子的天然非编码序列;以及叶甲属生物或半翅目生物的转录为包含seqidno:1、seqidno:3、seqidno:4、seqidno:5、seqidno:75、seqidno:76、seqidno:77、seqidno:78、seqidno:87和seqidno:89中任一者之全部或部分的天然rna分子的天然非编码序列的互补物。本文中还公开了方法,其中可将dsrna、mirna、sirna、shrna、和/或hprna在基于饵食的测定中(diet-basedassay)提供给鞘翅目和/或半翅目害虫,或提供在表达dsrna、mirna、sirna、shrna、和/或hprna的遗传修饰的植物细胞中。在这些和另外的实例中,dsrna、shrna、mirna、sirna、和/或hprna可被鞘翅目幼虫和/或半翅目害虫蛹摄入。然后,本发明的dsrna、sirna、shrna、mirna、和/或hprna的摄入可在该幼虫/蛹中导致rnai,进而可导致对于该鞘翅目和/或半翅目害虫的活力必需的基因的沉默,并最终导致幼虫/蛹死亡。因此,公开了方法,其中给鞘翅目和/或半翅目害虫提供核酸分子,所述核酸分子包含可用于控制鞘翅目和/或半翅目害虫的示例性核酸序列。在特定的实例中,通过使用本发明的核酸分子控制的鞘翅目和/或半翅目害虫可以是wcr、ncr、scr,mcr、d.balteata、d.u.tenella、南美叶甲、d.u.undecimpunctata、英雄美洲蝽(euchistusheros)、褐椿象(e.servus)、盖德拟壁蝽(piezodorusguildinii)、茶翅蝽(halyomorphahalys)、稻绿蝽(nezaraviridula)、绿蝽(chinaviahilare)、c.marginatum,dichelopsmelacanthus,d.furcatus,edessameditabunda,thyantaperditor,horciasnobilellus,taediastigmosa,dysdercusperuvianus,neomegalotomusparvus,leptoglossuszonatus,niesthreasidae,和/或牧草盲蝽(lyguslineolari)。参考下文结合附图进行的对若干实施方案的详细说明,前述的特征以及其他特征将会更加自明。附图简述图1包括用于使用用单一一对引物从单个转录模板生成dsrna的策略的描述。图2是用于自两个转录模板的生成dsrna的策略的描述。序列表中的序列的简述使用如37c.f.r.§1.822规定的核苷酸碱基的标准字母缩写,示出了所附序列表中列出的核酸序列。所列出的核酸和氨基酸序列定义了具有以所记载的方式排列的核苷酸和氨基酸单体的分子(即分别为多核苷酸和多肽)。每个列出的核酸和氨基酸序列也定义了包含以所记载的方式排列的核苷酸和氨基酸单体的多核苷酸或多肽的类属(genus)。鉴于遗传密码的冗余,可以理解的是,某个包含编码序列的核苷酸序列同样可还描述一个多核苷酸类属,其中的多核苷酸和由参考序列组成的多核苷酸编码相同的多肽。还应当理解,某个氨基酸序列可描述编码该多肽的多核苷酸orf的类属。对于每个核酸序列仅显示了一条链,仅显示每个核酸序列的一条链,任何对所展示的链的提述应理解为包括对互补链。由于一级核酸序列的互补序列和反向互补序列必然被该一级序列所公开,所以对核酸序列的任何提述也包括核酸序列的互补序列和反向互补序列,但另有明确说明或者从该序列出现的上下文可以明确看出并非如此的除外。此外,如本领域所知,rna链的核苷酸序列决定于转录该rna的dna的序列(除了胸腺嘧啶(t)被尿嘧啶(u)核碱基替换之外),对于某个rna序列而言,任何对编码它的dna序列的提述也包括了该序列。在所附序列表中:seqidno:1显示了来自玉米根萤叶甲的包含copigamma亚单位的dna。seqidno:2显示来自玉米根萤叶甲的copigamma蛋白质的氨基酸序列。seqidno:3显示被用于体外dsrna合成的来自玉米根萤叶甲的copigammareg1(区域1)的dna序列(未显示5’和3’末端的t7启动子序列)。seqidno:4显示被用于体外dsrna合成的来自玉米根萤叶甲的copigammareg2(区域2)的dna序列(未显示5’和3’末端的t7启动子序列)。seqidno:5显示被用于体外dsrna合成的来自玉米根萤叶甲的copigammareg3(区域3)的dna序列(未显示5’和3’末端的t7启动子序列)。seqidno:6显示t7噬菌体启动子的dna序列。seqidno:7显示被用于体外dsrna合成的yfp编码区区段的dna序列(未显示5’和3’末端的t7启动子序列)。seqidno:8至13显示引物,这些引物被用于扩增来自玉米根萤叶甲的copigamma亚单位序列中包含copigammareg1、copigammareg2和copigammareg3的部分。seqidno:14呈现如pdab117221中所见的来自玉米根萤叶甲的copigamma发夹v3-rna-形成序列。大写字母的碱基为copigamma有义链,下划线的小写字母碱基包含st-ls1内含子,无下划线的小写字母碱基为copigamma反义链。cgacctcctccggtgtctagagaagaaaacttcgccgaaaaacttagtaacgttccgggtatacaacagttaggacctttgttcaaaacttccgacgtcgttgaactcacgactagtaccggttgggaaaggtatgtttctgcttctacctttgatatatatataataattatcactaattagtagtaatatagtatttcaagtatttttttcaaaataaaagaatgtagtatatagctattgcttttctgtagtttataagtgtgtatattttaatttataacttttctaatatatgaccaaaacatggtgatgtgcaggttgatccgcggttagtgagttcaacgacgtcggaagttttgaacaaaggtcctaactgttgtatacccggaacgttactaagtttttcggcgaagttttcttctctagacaccggaggaggtcgseqidno:15呈现如pdab117222中所见的来自玉米根萤叶甲的copigamma发夹v4-rna-形成序列。大写字母碱基为copigamma有义链,下划线的小写字母碱基包含st-ls1内含子,无下划线的小写字母碱基为copigamma反义链.agttgcactataacgaaaccggtaccacatatgtagtagttaagttgcctgatgatgatctccccaactctgttggtacgtgtggagccgtgttgaagttcttagtgaaagattgtgatccatcaacgggaataccagattctgatgagggttacgatgatgaatatacactggaagacatcgaaataacattaggggacgactagtaccggttgggaaaggtatgtttctgcttctacctttgatatatatataataattatcactaattagtagtaatatagtatttcaagtatttttttcaaaataaaagaatgtagtatatagctattgcttttctgtagtttataagtgtgtatattttaatttataacttttctaatatatgaccaaaacatggtgatgtgcaggttgatccgcggttagtcccctaatgttatttcgatgtcttccagtgtatattcatcatcgtaaccctcatcagaatctggtattcccgttgatggatcacaatctttcactaagaacttcaacacggctccacacgtaccaacagagttggggagatcatcatcaggcaacttaactactacatatgtggtaccggtttcgttatagtgcaactseqidno:16显示如pdab110853中所见的yfp发夹-rna-形成序列v2。大写字母碱基为yfp有义链,下划线的碱基包含st-ls1内含子,小写字母、无下划线的碱基为yfp反义链。atgtcatctggagcacttctctttcatgggaagattccttacgttgtggagatggaagggaatgttgatggccacacctttagcatacgtgggaaaggctacggagatgcctcagtgggaaaggactagtaccggttgggaaaggtatgtttctgcttctacctttgatatatatataataattatcactaattagtagtaatatagtatttcaagtatttttttcaaaataaaagaatgtagtatatagctattgcttttctgtagtttataagtgtgtatattttaatttataacttttctaatatatgaccaaaacatggtgatgtgcaggttgatccgcggttactttcccactgaggcatctccgtagcctttcccacgtatgctaaaggtgtggccatcaacattcccttccatctccacaacgtaaggaatcttcccatgaaagagaagtgctccagatgacatseqidno:17显示包含st-ls1内含子的序列。seqidno:18显示如pdab101556中所见的yfp蛋白编码序列。seqidno:19显示膜联蛋白区域1的dna序列。seqidno:20显示膜联蛋白区域2的dna序列。seqidno:21显示β血影蛋白2区域1的dna序列。seqidno:22显示β血影蛋白2区域2的dna序列。seqidno:23显示mtrp-l4区域1的dna序列。seqidno:24显示mtrp-l4区域2的dna序列。seqidno:25至52显示被用于扩增yfp、膜联蛋白、β血影蛋白2及mtrp-l4的基因区域以供dsrna合成的引物。seqidno:53显示编码tip41样蛋白的玉米dna序列。seqidno:54显示寡核苷酸t20nv的dna序列。seqidno:55至59显示用于测量玉米转录物水平的引物和探针的序列。seqidno:60显示被用于二元载体骨架检测的specr编码区域的一部分的dna序列。seqidno:61显示被用于基因组拷贝数分析的aad1编码区域的一部分的dna序列。seqidno:62显示玉米转化酶基因的dna序列。seqidno:63至71显示被用于基因拷贝数分析的引物和探针的序列。seqidno:72至74显示被用于玉米表达分析的引物和探针的序列。seqidno:75显示被用于体外dsrna合成的来自玉米根萤叶甲的copigammaver1(版本1)的dna序列(未显示5’和3’末端的t7启动子序列)。seqidno:76显示被用于体外dsrna合成的来自玉米根萤叶甲的copigammaver2(版本2)的dna序列(未显示5’和3’末端的t7启动子序列)。seqidno:77显示被用于体外dsrna合成的来自玉米根萤叶甲的copigammaver3(版本3)dna序列(未显示5’和3’末端的t7启动子序列)。seqidno:78显示seqidno:77显示被用于体外dsrna合成的来自玉米根萤叶甲的copigammaver4(版本4)dna序列(未显示5’和3’末端的t7启动子序列)。seqidno:79-86显示引物,所述引物被用于扩增来自玉米根萤叶甲copigamma序列的包含copigammaver1、copigammaver2、gammaver3和copigammaver4的部分。seqidno:87显示示例性的来自新热带褐椿象(英雄美洲蝽)的bsbcopigamma转录物的dna序列。seqidno:88显示来自英雄美洲蝽copigamma蛋白的氨基酸序列。seqidno:89显示被用于体外dsrna合成的来自英雄美洲蝽的bsb_copigamma-2的dna序列(未显示5’和3’末端的t7启动子序列)。seqidno:90-91显示引物,所述引物被用于扩增来自英雄美洲蝽copigamma的包含bsb_copigamma-2的序列的部分。seqidno:92是靶定yfp的dsrna:yfpv2的有义链。seqidno:93-94显示被用于扩增靶定yfp的dsrna:yfpv2的部分的引物。seqidno:95呈现yfp发夹序列(yfpv2-1)。大写字母碱基为yfp有义链,下划线的小写字母碱基包含rtm1内含子,无下划线的小写字母碱基为yfp反义链。atgtcatctggagcacttctctttcatgggaagattccttacgttgtggagatggaagggaatgttgatggccacacctttagcatacgtgggaaaggctacggagatgcctcagtgggaaagtccggcaacatgtttgacgtttgtttgacgttgtaagtctgatttttgactcttcttttttctccgtcacaatttctacttccaactaaaatgctaagaacatggttataactttttttttataacttaatatgtgatttggacccagcagatagagctcattactttcccactgaggcatctccgtagcctttcccacgtatgctaaaggtgtggccatcaacattcccttccatctccacaacgtaaggaatcttcccatgaaagagaagtgctccagatgacat详细说明i.若干实施方案的概览本文中公开了用于遗传控制昆虫(例如鞘翅目或半翅目)害虫侵染的方法和组合物。还提供了用于鉴定对于昆虫害虫的生命周期必需的一个或多个基因,作为靶基因用于rnai介导的昆虫害虫群体控制的方法。可设计编码rna分子的dna质粒载体来抑制生长、存活、和/或发育所必需的靶基因。在一些实施方案中,所述rna分子可能能够形成dsrna分子。在一些实施方案中,提供了通过与昆虫害虫中靶基因的编码或非编码序列互补的核酸分子来转录后阻抑靶基因表达或抑制靶基因的方法。在这些和另外的实施方案中,昆虫害虫可以摄取一种或多种从与靶基因的编码或非编码序列互补的核酸分子的全部或部分转录的dsrna、sirna、shrna、mirna、和/或hprna分子,由此提供植物保护效果。因此,一些实施方案涉及使用与靶基因的编码和/或非编码序列互补的irna(例如dsrna、sirna、shrna、mirna、和/或hprna)对靶基因产物的表达进行序列特异性抑制,以实现对昆虫(例如鞘翅目和/或半翅目)害虫的至少部分控制。公开了一组分离纯化的核酸分子,其包含,例如,如seqidno:1,seqidno:3,seqidno:4,seqidno:5,seqidno:75,seqidno:76,seqidno:77,seqidno:78,seqidno:87,seqidno:89任一者中列出的多核苷酸及其片段。在一些实施方案中,可以从该序列、其片段、或包含这些多核苷酸中一种或多种的基因表达稳定化的dsrna分子,用于靶基因的转录后沉默或抑制。在某些实施方案中,分离和纯化的核酸分子包含seqidno:1的全部或部分。在其他的实施方案中,分离和纯化的核酸分子包含seqidno:3的全部或部分。在进一步的实施方案中,分离和纯化的核酸分子包含seqidno:4的全部或部分。在其他的实施方案中,分离和纯化的核酸分子包含seqidno:5的全部或部分。在另外的实施方案中,分离和纯化的核酸分子包含seqidno:75,seqidno:76,seqidno:77,seqidno:78,seqidno:87或seqidno:89的全部或部分。一些实施方案涉及在其基因组中具有编码至少一种irna(例如,dsrna)分子的至少一种重组dna的重组宿主细胞(例如,植物细胞)。在特定的实施方案中,dsrna分子可以在被鞘翅目和/或半翅目害虫摄取时产生,使该害虫中的靶基因发生转录后沉默或抑制其表达。重组dna可以包含,例如:seqidno:1、seqidno:3、seqidno:4、seqidno:5、seqidno:75、seqidno:76、seqidno:77、seqidno:78、seqidno:87或seqidno:89中任一者;seqidno:1、seqidno:3、seqidno:4、seqidno:5、seqidno:75、seqidno:76、seqidno:77、seqidno:78、seqidno:87或seqidno:89中任一者的片段;包含seqidno:1、seqidno:3、seqidno:4、seqidno:5、seqidno:75、seqidno:76、seqidno:77、seqidno:78、seqidno:87或seqidno:89中一者或更多者的基因的部分序列;或其互补物。一些实施方案涉及重组宿主细胞,所述细胞在其基因组中具有重组dna,所述重组dna具有编码至少一种irna(例如,dsrna)分子,所述irna分子包含由seqidno:1和/或seqidno:87编码的rna的全部或部分;和/或其互补物。当被昆虫(例如鞘翅目和/或半翅目)害虫摄入时,irna分子可以在该鞘翅目和/或半翅目害虫中沉默或抑制包含seqidno:1和/或seqidno:87的靶基因的表达,从而导致害虫的进食、生长和/或发育停止。在一些实施方案中,在其基因组中具有编码至少一种能够形成dsrna分子的rna分子的至少一种重组dna的重组宿主细胞可以是转化的植物细胞。一些实施方案涉及包含这样的转化的植物细胞的转基因植物。除了这样的转基因植物之外,还提供了任何转基因植物世代的后代植物、转基因种子、和转基因植物产品,它们均包含重组dna。在特定的实施方案中,可以在转基因植物细胞中表达能够形成dsrna分子的rna分子。因此,在这些和其他实施方案中,可以从转基因植物细胞分离dsrna分子。在特定的实施方案中,转基因植物是选自下组的植物,包括:玉米(zeamays)、大豆(glycinemax)、和禾本科(poaceae)的植物。一些实施方案涉及用于调控昆虫(例如鞘翅目和/或半翅目)害虫细胞中靶基因表达的方法。在这些和其他实施方案中,可以提供核酸分子,其中所述核酸分子包含编码能够形成dsrna分子的rna分子的多核苷酸。在特定的实施方案中,编码能够形成dsrna分子的rna分子的多核苷酸可与启动子可操作连接,并且还可与转录终止序列可操作连接。在特定的实施方案中,用于调控昆虫害虫细胞中靶基因表达的方法可包括:(a)用包含编码能够形成dsrna分子的rna分子的多核苷酸的载体转化植物细胞;(b)在足以容许形成包含多个转化的植物细胞的植物细胞培养物的条件下培养所述转化的植物细胞;(c)选择已将载体整合到其基因组中的转化的植物细胞;和(d)确定选择的转化的植物细胞包含由载体的所述多核苷酸序列编码的、能够形成dsrna分子的rna分子。可以从基因组中整合有载体、且包含由载体的所述多核苷酸编码的dsrna分子的植物细胞再生植物。因此,还公开了在其基因组中整合有载体的转基因植物,所述载体具有编码能够形成dsrna分子的rna分子的多核苷酸,其中转基因植物包含由所述载体的所述多核苷酸编码的dsrna分子。在特定的实施方案中,植物中能够形成dsrna分子的rna分子的表达足以调控接触所述转化的植物或植物细胞(例如通过以转化的植物、植物的一部分(例如,根)或植物细胞为食)的昆虫(例如鞘翅目和/或半翅目)害虫细胞中靶基因的表达,使得害虫的生长和/或存活被抑制。本文中公开的转基因植物可以展现出对昆虫害虫侵染的抗性和/或增强的耐受性。特定的转基因植物可以展现出对一种或多种选自下组的鞘翅目和/或半翅目害虫的抗性和/或增强的保护:wcr、ncr、scr、mcr、玉米根萤叶甲、d.u.tenella、南美叶甲、d.u.undecimpunctatamannerheim、英雄美洲蝽、盖德拟壁蝽、茶翅蝽、稻绿蝽、绿蝽、褐椿象、dichelopsmelacanthus;dichelopsfurcatus;edessameditabunda;thyantaperditor;chinaviamarginatum;horciasnobilellus;taediastigmosa;dysdercusperuvianus;neomegalotomusparvus;leptoglossuszonatus;niesthreasidae;lygushesperus;和lyguslineolaris。本文中还公开了用于将控制剂(如irna分子等)投送给昆虫(例如鞘翅目和/或半翅目)害虫的方法。这样的控制剂可以直接或间接地阻碍昆虫害虫群体进食、生长或以其他方式致害宿主的能力。在一些实施方案中,提供了这样的方法,包括将稳定化的dsrna分子递送至昆虫害虫以抑制该害虫中的至少一种靶基因,从而导致rnai并减少或消除害虫宿主中的植物损害。在一些实施方案中,抑制昆虫害虫中靶基因表达的方法可以导致害虫中生长、存活、和/或发育的停止。在一些实施方案中,提供了包含irna(例如,dsrna)分子的组合物(例如,局部用组合物),供用于植物、动物、和/或植物或动物的环境以消除或降低昆虫(例如鞘翅目和/或半翅目)害虫的侵染。在特定的实施方案中,组合物可以是要喂给昆虫害虫的营养组合物或食物来源。一些实施方案包括使害虫可得到所述营养组合物或食物来源。包含irna分子的组合物的摄入可导致一个或多个害虫细胞摄取所述分子,进而可导致害虫细胞中至少一种靶基因表达的抑制。通过在害虫的宿主中提供一种或多种包含irna分子的组合物,可以在任何存在昆虫害虫的宿主组织或环境之中或之上限制或消除该害虫侵染对植物或植物细胞的摄入或损伤。本文中公开的组合物和方法可与针对其他靶(例如ras相对物或rop(美国专利申请公开号20150176025)和rnapii(美国专利申请公开号20150176009))的irna分子组合一起使用。这种影响害虫中(例如幼虫中)多种靶序列的潜力可以增加功效,并且还可以改善涉及irna技术的可持续的昆虫害虫管理策略。本文中公开的组合物和方法可与用于控制昆虫(例如鞘翅目和/或半翅目)害虫所致损害的其他方法和组合物组合一起使用。例如,本文中所描述的用于针对昆虫害虫保护植物的irna分子可以在这样的方法中使用,所述方法包括另外使用一种或多种针对昆虫害虫有效的化学剂、针对这样的害虫有效的生物杀虫剂、作物轮作、展现出与rnai介导的方法和rnai组合物的特征不同的特征(例如,在植物中重组产生对于昆虫害虫有害的蛋白质(例如,bt毒素))的重组遗传技术。ii.缩写bsb新热带棕椿象(英雄美洲蝽,euschistusheros)dsrna双链核糖核酸est表达序列标签gi生长抑制ncbi美国国家生物技术信息中心gdna基因组dnairna抑制性核糖核酸orf开放阅读框rnai核糖核酸干扰mirna微小核糖核酸sirna小抑制性核糖核酸shrna短发夹核糖核酸hprna发夹核糖核酸utr非翻译区wcr西方玉米根虫(玉米根萤叶甲)ncr北方玉米根虫(巴氏根萤叶甲)mcr墨西哥玉米根虫(墨西哥玉米根萤叶甲)pcr聚合酶链式反应qpcr定量聚合酶链式反应riscrna诱导的沉默复合物scr南方玉米根虫(十一星根萤叶甲)sem均值标准误yfp绿色荧光蛋白iii.术语在以下描述和表中,使用了许多术语。为了提供对说明书和权利要求书的清楚且一致的理解,包括要赋予此类术语的范围,提供了以下定义:鞘翅目害虫:如本文中使用的,术语“鞘翅目害虫”指以农业作物和作物产品,包括玉米和其它真正的禾本科为食的、属于鞘翅目(coleoptera)的昆虫,包括属于叶甲属的害虫。在具体的例子中,鞘翅目害虫选自下列清单,包括:玉米根萤叶甲(wcr);巴氏根萤叶甲(ncr);十一星根萤叶甲(scr);墨西哥玉米根萤叶甲(mcr);黄瓜条根萤叶甲;d.u.tenella;和d.u.undecimpunctatamannerheim。(与生物)接触:如本文中使用的,与生物(例如,鞘翅目害虫和/或半翅目害虫)“接触”或被生物“摄取”等术语,就核酸分子而言,包括核酸分子内化到生物中,例如但不限于:生物摄取分子(例如,通过进食);使生物与包含核酸分子的组合物接触;以及用包含核酸分子的溶液浸泡生物。重叠群:如本文中使用的,术语“重叠群”指从一组来源于单一遗传来源的重叠dna区段重建的dna序列。玉米植物:如本文中使用的,术语“玉米植物”指物种玉蜀黍(玉米)的植物。表达:如本文中使用的,编码多核苷酸(例如,基因或转基因)的“表达”是指这样的过程,通过该过程,核酸转录单元(包括,例如gdna或cdna)的编码信息被转化为细胞的操作部分、非操作部分、或结构部分(常常包括蛋白质的合成)。基因表达可能受到外部信号的影响,例如细胞、组织、或生物暴露于增加或降低基因表达的物质。基因表达也可以在从dna到rna再到蛋白质的途径中的任何位置受到调节。例如,通过控制对转录、翻译、rna运输和加工、中间分子比如mrna的降解的作用,或通过在特异性蛋白分子已经产生之后的活化、失活、区室化、或降解,或它们的组合,由此发生基因表达的调节。通过本领域已知的任何方法,包括但不限于,northern印迹、rt-pcr、western印迹,或体外、原位或体内蛋白活性测定,可以在rna水平或蛋白质水平测量基因表达。遗传物质:如本文中使用的,术语“遗传物质”包括所有基因和核酸分子,诸如dna和rna。半翅目害虫:如本文中使用的,术语“半翅目害虫”指属于半翅目(hemiptera)的昆虫,包括但不限于蝽科(pentatomidae),盲蝽科(miridae),红蝽科(pyrrhocoridae),缘蝽科(coreidae),蛛蝽科(alydidae),和姬缘蝽科(rhopalidae)等科中的昆虫,其以范围很广的宿主植物为食,具有刺吸式口器。在特定的实例中,半翅目宿主选自下组:英雄美洲蝽[euschistusheros(fabr.)](新热带棕椿象),稻绿蝽nezaraviridula(l.)(南方绿椿象),盖德拟壁蝽[piezodorusguildinii(westwood)](红带椿象),茶翅蝽[halyomorphahalys](棕色大理石椿象),绿蝽[chinaviahilare(say)](绿色椿象),褐椿象[euschistusservus(say)](棕色椿象),dichelopsmelacanthus(dallas),dichelopsfurcatus(f.),edessameditabunda(f.),thyantaperditor(f.)(新热带红肩椿象),chinaviamarginatum(palisotdebeauvois),horciasnobilellus(berg)(棉红蝽),taediastigmosa(berg),dysdercusperuvianus(guérin-méneville),neomegalotomusparvus(westwood),leptoglossuszonatus(dallas),niesthreasidae(f.),lygushesperus(knight)(西方牧草盲蝽),和牧草盲蝽[lyguslineolaris(palisotdebeauvois)]。抑制:如本文中使用的,术语“抑制”在用于描述对编码多核苷酸(例如基因)的影响时是指从编码多核苷酸转录的mrna和/或编码多核苷酸的肽、多肽、或蛋白质产物的细胞水平的可测量的降低。在一些实例中,抑制编码多核苷酸的表达可以使得表达大致消失。“特异性抑制”是指在实现特异性抑制的细胞中对靶编码多核苷酸的抑制不对其他编码多核苷酸(例如,基因)的表达产生影响。昆虫:如本文用于害虫的,术语“害虫”特别包括鞘翅目昆虫害虫和半翅目害虫。在一些实施方案中,该术语还包括某些其它害虫。分离的:“分离的”生物学组分(诸如核酸、肽或蛋白质)是已经从该组分所天然存在的生物细胞中的其他生物学组分(即,其他染色体和染色体外的dna和rna、和蛋白质)实质性分开、分开产生、或纯化出来,同时实现组分中的化学或功能变化(例如,核酸可以通过断开将核酸连接到染色体中的其余dna的化学键而从染色体分离)。已经“分离的”核酸分子和蛋白质包括通过标准纯化方法纯化的核酸分子和蛋白质。该术语还包括通过在宿主细胞内重组表达制备的核酸分子和蛋白质,以及化学合成的核酸分子、蛋白质和肽。核酸分子:如本文中使用的,术语“核酸分子”可以是指核苷酸的聚合物形式,可包括rna、cdna、gdna的有义和反义链,以及上述情形的合成形式和混合聚合物。核苷酸或核碱基可以是指核糖核苷酸、脱氧核糖核苷酸、或这两种类型核苷酸的任一者的修饰形式。如本文中使用的“核酸分子”与“核酸”和“多核苷酸”是同义词。除非另外指明,核酸分子通常在长度上具有至少10个碱基。根据惯例,核酸分子的核苷酸序列从分子的5’至3’端阅读。核酸分子的“互补物”(即互补序列)是指具有可以与该核酸分子的核碱基形成碱基对(即,a-t/u,和g-c)的核碱基的多核苷酸。一些实施方案包括包含转录为rna分子的模板dna的核酸,所述rna分子是mrna分子的互补序列。在这些实施方案中,转录为mrna分子的核酸的互补物以5'至3'方向存在,使得rna聚合酶(其以5'至3'方向转录dna)将从互补物转录出可以与mrna分子杂交的核酸。因此,除非另有明确说明或者从上下文中可以清楚地看出另有所指,术语“互补物”是指具有可以与特定核苷酸序列的核碱基形成碱基对的从5'至3'的核碱基的多核苷酸。类似地,除非另有明确说明或者从上下文中可以清楚地看出另有所指,核酸的“反向互补物”是指相反取向的互补物。前述内容在以下图示中示出:5’atgatgatg3’多核苷酸5’tactactac3’该多核苷酸的“互补物”5’catcatcat3’该多核苷酸的“反向互补物”本发明的一些实施方案包括形成发夹rna的irna分子。在这些irna分子中,rna干扰的靶核酸的互补物和反向互补物可以出现在同一分子中,使得单链rna分子在包含互补和反向互补的多核苷酸的区域上可以“回折”并与自身杂交,如下面的图示所示:5’augaugaug–接头多核苷酸–caucaucau3’,其杂交而形成:“核酸分子”包括所有多核苷酸,例如:dna的单链和双链形式;rna的单链形式;和rna的双链形式(dsrna)。术语“核苷酸序列”或“核酸序列”是指作为个别单链或在双链体中的核酸有义和反义链两者。术语“核糖核酸”(rna)包括irna(抑制性rna)、dsrna(双链rna)、sirna(小干扰rna)、mrna(信使rna)、mirna(微小rna)、shrna(小发夹rna)、hprna(发夹rna)、trna(转运rna)(无论是装载还是卸下相应的酰化氨基酸)、和crna(互补rna)。术语“脱氧核糖核酸”(dna)包括cdna、gdna、和dna-rna杂交体。术语“多核苷酸”、“核酸”、其“区段”,或其“片段”,如本领域技术人员将理解的,包括,例如,gdna、核糖体rna、转运rna、rna、信使rna、操纵子、和较小的工程化多核苷酸,其编码或可适用于编码肽、多肽、或蛋白质;以及核酸分子中的功能和/或结构元件,由其相应的核苷酸序列界定。寡核苷酸:寡核苷酸是短的核酸聚合物。寡核苷酸可通过切割较长核酸区段、或通过使单独的核苷酸前体聚合而形成。自动合成仪能够合成长度多达几百个碱基的寡核苷酸。由于寡核苷酸可与互补的核酸结合,因此它们可用作检测dna或rna的探针。由dna(寡脱氧核糖核苷酸)组成的寡核苷酸可以用于pcr,pcr是一种用于扩增dna和rna(逆转录成cdna)序列的技术。在pcr中,寡核苷酸一般被称为“引物”,其允许dna聚合酶延伸寡核苷酸并复制互补链。核酸分子可以包括天然存在的核苷酸和/或修饰核苷酸,它们通过天然存在和/或非天然存在的核苷酸连接而连接在一起。本领域技术人员易于理解,核酸分子可被化学修饰或生物化学修饰,或者可以含有非天然的或衍生化的核苷酸碱基。这些修饰包括,例如,标记物、甲基化、用类似物取代一个或多个天然存在的核苷酸、核苷酸间修饰(例如,如不带电连接(unchargedlinkage)的修饰:例如甲基膦酸盐、磷酸三酯、氨基磷酸酯、氨基甲酸酯等;带电连接(chargedlinkage)的修饰:例如,硫代磷酸酯、二硫代磷酸酯等;悬垂部分的修饰:例如肽类;嵌入剂修饰:例如吖啶、补骨脂素等;螯合剂修饰;烷化剂(alkylator)修饰;和修饰的连接:例如α异头核酸等)。术语“核酸分子”还包括任何拓扑结构,包括单链的、双链的、部分双链体的、三链体的、发夹形的(hairpinned)、圆形的、挂锁形的结构。如本文中使用的,就dna而言,术语“编码序列”、“结构核苷酸序列”、或“结构核酸分子”是指这样的核苷酸序列,当被置于适当的调节序列控制下时,其通过转录和mrna最终翻译成多肽。就rna而言,术语“编码序列”指翻译成肽、多肽或蛋白质的核苷酸序列。编码序列的边界由5’端的翻译起始密码子和3’端的翻译终止密码子所界定。编码多核苷酸包括但不限于:基因组dna;cdna;est;和重组核苷酸序列。基因组:如本文中所使用的,术语“基因组”是指存在于细胞核内的染色体dna,并且也指存在于细胞的亚细胞组成部分内的细胞器dna。在本发明的一些实施方案中,可以将dna分子导入植物细胞中,使得dna分子整合到植物细胞的基因组中。在这些和另外的实施方案中,dna分子可以整合到植物细胞的核dna中,或整合到植物细胞的叶绿体或线粒体的dna中。术语“基因组”在其应用于细菌时,指细菌细胞内的染色体和质粒两者。在本发明的一些实施方案中,可以将dna分子导入细菌中,使得dna分子整合到细菌的基因组中。在这些和另外的实施方案中,dna分子可以整合于染色体,或者作为稳定的质粒定位或位于稳定的质粒中。序列同一性:如本文中使用的术语“序列同一性”或“同一性”,在两个核酸或多肽序列的情况下,是指在指定比较窗口上以最大对应性比对时,这两个序列中相同的残基。如本文中使用的,术语“序列同一性百分比”可以是指通过在比较窗口上比较分子的两个最优比对序列(例如,核酸序列或多肽序列)确定的值,其中为了实现这两个序列的最优比对,该比较窗口中的序列部分相比于参考序列(其不包含添加或缺失)可以包含添加或缺失(即,空位)。通过确定在两个序列中出现相同核苷酸或氨基酸残基的位置的数目而产生匹配位置数,用该匹配位置数除以比较窗口中位置的总数,将结果乘以100而产生序列同一性的百分比,从而计算出该百分比。每个位置与参考序列相比均相同的序列被认为是与参考序列是100%相同的,反之亦然。用于比较的序列比对方法是本领域技术人员熟知的。各种程序和比对算法例如描述于smith和waterman(1981)adv.appl.math.2:482;needleman和wunsch(1970)j.mol.biol.48:443;pearson和lipman(1988)proc.natl.acad.sci.u.s.a.85:2444;higgins和sharp(1988)gene73:237-244;higgins和sharp(1989)cabios5:151-153;corpet等人,(1988)nucleicacidsres.16:10881-10890;huang等人,(1992)comp.appl.biosci.8:155-165;pearson等人,(1994)methodsmol.biol.24:307-331;tatiana等人(1999)femsmicrobiol.lett.174:247-250。序列比对方法和同源性计算的详细考虑事项可见于例如,altschul等人,(1990)j.mol.biol.215:403-410。美国国家生物技术信息中心(ncbi)基本局部比对搜索工具(blasttm;altschul等人(1990))可在几个来源访问,包括美国国家生物技术信息中心(bethesda,md)、以及在互联网上,其与几种序列分析程序结合使用。怎样使用该程序确定序列同一性的描述可在互联网上blasttm的“帮助”部分获得。为了比较核酸序列,可以采用利用默认参数的默认blosum62矩阵集的blasttm(blastn)程序的“blast2序列”函数。在用这种方法评估时,与参考序列具有较大相似性的核酸序列将显示出同一性百分比的增加。可特异性杂交/特异性互补:如本文中使用的,术语“可特异性杂交”和“特异性互补”是表明互补性的足够程度的术语,该互补性足以使得在核酸分子与靶核酸分子之间发生稳定且特异的结合。在两个核酸分子之间的杂交两种涉及核酸分子的核酸序列之间的反平行比对的形成。然后,两个分子能够与相反链上的相应碱基形成氢键以形成双链体分子,如果该双链体分子足够稳定,则可使用本领域中熟知的方法检出。核酸分子与其可特异性杂交的靶序列不一定是100%互补的。然而,对于特异性杂交而言必须存在的序列互补性的量依赖所使用的杂交条件而变化。导致特定严格程度的杂交条件会根据选择的杂交方法的性质和杂交的核酸序列的组成和长度而变化。通常,杂交的温度和杂交的离子强度(尤其是na+和/或mg++浓度)将决定杂交严格性。洗涤缓冲液的离子强度和洗涤温度也会影响严格性。关于获得特定严格性程度所需要的杂交条件的计算是本领域普通技术人员已知的,并且论述于例如sambrook等人(编辑)molecularcloning:alaboratorymanual,2nded.,vol.1-3,coldspringharborlaboratorypress,coldspringharbor,ny,1989,chapters9and11和更新;以及hames和higgins(编辑)nucleicacidhybridization,irlpress,oxford,1985。关于核酸杂交的进一步详细说明和指导例如可见于tijssen,"overviewofprinciplesofhybridizationandthestrategyofnucleicacidprobeassays,"inlaboratorytechniquesinbiochemistryandmolecularbiology-hybridizationwithnucleicacidprobes,parti,chapter2,elsevier,ny,1993;以及ausubel等人编辑,currentprotocolsinmolecularbiology,chapter2,greenepublishingandwiley-interscience,ny,1995和更新。如本文中使用的,“严格条件”涵盖仅在杂交分子与靶核酸分子内的同源序列之间存在大于80%的序列匹配时才发生杂交的条件。“严格条件”包括另外的特定严格性水平。因此,如本文中使用的,“中等严格性”条件为具有超过80%的以上序列匹配(即具有少于20%错配)的分子将会杂交的条件;“高严格性”条件为具有超过90%的匹配(即具有少于10%错配)的序列将会杂交的条件(即,具有小于10%的错配);并且“极高严格性”条件为具有超过95%的匹配(即具有少于15%错配)的序列将会杂交的条件。以下是代表性的非限制性杂交条件:高严格性条件(检测出共享至少90%序列同一性的序列):在5xssc缓冲液中在65℃杂交16小时;在2xssc缓冲液中在室温下洗涤两次,每次15分钟;并且在0.5xssc缓冲液中在65℃洗涤两次,每次20分钟。中等严格性条件(检测出共享至少80%序列同一性的序列):在5x到6xssc缓冲液中在65--70℃杂交16-20小时;在2xssc缓冲液中在室温下洗涤两次,每次5-20分钟;并且在1xssc缓冲液中在55-70℃洗涤两次,每次30分钟。非严格对照条件(共享至少50%序列同一性的序列会杂交):在6xssc缓冲液中在室温到55℃杂交16-20小时;在2x到3xssc缓冲液中在室温到55℃洗涤至少两次,每次20-30分钟。如本文中使用的,术语“基本上同源”或“实质同源性”就核酸而言是指具有这样的连续核碱基的多核苷酸,所述连续核碱基在严格条件下与参考核酸杂交。例如,与seqidno:1中任一个的参考核酸序列基本上同源的序列的核酸是在严格条件(例如,上文列出的中等严格性条件)下与seqidno:1中任一个的参考核酸杂交的那些核酸。基本上同源的多核苷酸可以具有至少80%序列同一性。例如,基本上同源的多核苷酸可以具有约80%到100%序列同一性,诸如约79%;80%;约81%;约82%;约83%;约84%;约85%;约86%;约87%;约88%;约89%;约90%;约91%;约92%;约93%;约94%;约95%;约96%;约97%;约98%;约98.5%;约99%;约99.5%;和约100%。实质性同源性的特性与特异性杂交紧密相关。例如,当存在足够的互补程度时,核酸分子可特异性地杂交,以便避免核酸在特异性结合是期望的情况下(例如,在严格杂交条件下)与非靶多核苷酸的非特异性结合。如本文中使用的,术语“直系同源物”是指两种或更多种物种中已经从共同的祖先核酸进化并且可以在所述两种或更多种物种中保留相同功能的基因。如本文中使用的,当一个多核苷酸以5’至3’方向阅读的每个核苷酸与另一多核苷酸以3’至5’方向阅读时的每个核苷酸序列互补时,两个核酸分子被认为展现出“完全互补性”。与参考多核苷酸互补的多核苷酸将展现出与参考多核苷酸的反向互补物相同的序列。这些术语和描述是本领域中定义明确的,并且是本领域普通技术人员容易理解的。可操作(地)连接:当第一多核苷酸与第二多核苷酸处于功能性关系中时,则第一多核苷酸与第二多核苷酸是可操作连接的。当以重组方式产生时,可操作连接的多核苷酸通常是连续的,并且在需要将两个蛋白质编码区连接在一起的场合,二者在同一个阅读框内(例如,在翻译融合的orf中)。然而,可操作连接的核酸不一定是连续的。在有关调节性遗传元件和编码多核苷酸的语境下使用时,术语“可操作连接”意味着该调节元件影响该连接的编码序列的表达。“调节元件”或“控制元件”是指影响转录的周期和水平/量、rna加工或稳定性、或相关编码多核苷酸的翻译的多核苷酸。调节元件可以包括启动子;翻译前导;内含子;增强子;茎环结构;阻抑物结合多核苷酸;具有终止序列的多核苷酸;具有多腺苷酸化识别序列的多核苷酸等。特定的调节元件可位于与其可操作连接的编码多核苷酸的上游和/或下游。并且,与编码多核苷酸可操作连接的特定调节序列可位于双链核酸分子的相关互补链上。启动子:如本文中使用的,术语“启动子”是指可以在转录起始上游的、且可能涉及rna聚合酶与其他蛋白质的识别和结合而启动转录的dna区域。启动子可与用于在细胞中表达的编码多核苷酸可操作连接,或者启动子可与编码信号序列的多核苷酸可操作连接,所述多核苷酸可与用于在细胞中表达的编码多核苷酸可操作连接。“植物启动子”可以是能够启动植物细胞转录的启动子。在发育控制下的启动子的实例包括优先启动某些组织中的转录的启动子,所述组织例如叶、根、种子、纤维、木质部导管、管胞、或厚壁组织。这样的启动子称为“组织优先”启动子。仅启动某些组织中的转录的启动子被称为“组织特异性”启动子。“细胞类型特异性”启动子主要驱动一个或多个器官中某些细胞类型(例如,根或叶中的维管细胞)中的表达。“诱导型”启动子可以是可以在环境控制之下的启动子。可通过诱导型启动子启动转录的环境条件的实例包括厌氧条件和光的存在。组织特异性启动子、组织优先启动子、细胞类型特异性启动子、和诱导型启动子构成“非组成型”启动子类别。“组成型”启动子是可在大多数环境条件下或在大多数组织或细胞类型中有活性的启动子。在本发明的一些实施方案中可以使用任何诱导型启动子。参见ward等人,(1993)plantmol.biol.22:361-366。利用诱导型启动子,转录速率响应于诱导剂而增加。诱导型启动子的实例包括但不限于:来自响应于铜的acei系统的启动子;响应于苯磺酰胺除草剂安全剂的来自玉米的in2基因启动子;来自tn10的tet阻抑物;和来自类固醇激素基因的诱导型启动子,其转录活性可通过糖皮质激素诱导(schena等人,(1991)proc.natl.acad.sci.usa88:0421)。示例性的组成型启动子包括,但不限于:来自植物病毒的启动子,例如来自花椰菜花叶病毒(camv)的35s启动子;来自水稻肌动蛋白基因的启动子;泛素启动子;pemu;mas;玉米h3组蛋白启动子;和als启动子,欧洲油菜als3结构基因5'的xba1/ncoi片段(或与所述xba1/ncoi片段相似的多核苷酸)(国际pct公布号美国专利wo96/30530)。另外,在本发明的一些实施方案中可以利用任何组织特异性或组织优先启动子。用包含与组织特异性启动子可操作连接的编码多核苷酸的核酸分子转化的植物可唯一地或优先地在特异性组织中产生所述编码多核苷酸的产物。示例性的组织特异性或组织优先启动子包括但不限于:种子特异性启动子,如来自菜豆蛋白基因的启动子;叶特异性和光诱导型启动子,如来自cab或rubisco的启动子;花药特异性启动子,如来自lat52的启动子;花粉特异性启动子,如来自zm13的启动子;以及小孢子优先启动子,如来自apg的启动子。大豆植物:如本文中使用的,术语“大豆植物”是指属于大豆属物种(glycinesp.)的植物,例如大豆(glycinemax)。转化:如本文中使用的,术语“转化”或“转导”是指将一个或多个核酸分子转移到细胞中。当核酸分子通过核酸分子并入细胞基因组中、或通过附加型复制而由该细胞稳定复制时,细胞被转导到该细胞中的核酸分子“转化”。如本文中使用的,术语“转化”涵盖可将核酸分子导入这种细胞中的所有技术。实例包括但不限于:用病毒载体转染;用质粒载体转化;电穿孔(fromm等人,(1986)nature319:791-3);脂质体转染(felgner等人,(1987)proc.natl.acad.sci.usa84:7413-7);显微注射(mueller等人,(1978)cell15:579-85);土壤杆菌介导的转移(fraley等人,(1983)proc.natl.acad.sci.usa80:4803-7);直接dna摄取;和微粒轰击(klein等人,(1987)nature327:70)。转基因:外源核酸。在一些实例中,转基因可以是编码rna的dna,所述rna能够形成dsrna分子的一条或两条链,包含与存在于鞘翅目和/或半翅目害虫中的核酸分子互补的多核苷酸。在另外的实例中,转基因可以为基因(例如除草剂耐性基因、编码工业或药学上有用的化合物的基因、或编码期望的农业性状的基因)。在这些和其他实例中,转基因可以含有与转基因的编码多核苷酸可操作连接的调节元件(例如,启动子)。载体:引入到细胞中,例如产生转化的细胞的核酸分子。载体可以包含允许其在宿主细胞中复制的遗传元件,诸如复制起点。载体的实例包括但不限于:质粒;粘粒;噬菌体;和携带外源dna或rna进入细胞中的病毒。载体还可包括一种或多种基因、包括产生反义分子的基因、和/或选择标志物基因和本领域已知的其他遗传元件。载体可以转导、转化或感染细胞,由此引起细胞表达核酸分子和/或由该载体编码的蛋白质。任选地,载体包括辅助核酸分子实现进入细胞中的物质(例如脂质体、蛋白质包衣等)。产量:相对于在相同生长位置在相同时间且在相同条件下生长的检验品种的产量,约100%或更大的稳定化产量。在特定的实施方案中,“提高的产量”或“提高产量”意指相对于在含有相当大密度的对该作物有害的鞘翅目和/或半翅目害虫(本申请的组合物和方法以所述害虫为目标)的相同生长位置在相同时间且在相同条件下生长的检验品种的产量,具有105%或更大的稳定化产量的栽培种。除非特别指明或暗示,如本文中使用的术语“一个”、“一种”和“该”表示“至少一个/种”。除非另外特别地解释,本文中使用的全部技术术语和科学术语具有与属于本公开的领域之内的普通技术人员通常所理解的相同的含义。可以在以下出版物中找到分子生物学中常见术语的定义:例如,lewin’sgenesx,jones&bartlettpublishers,2009(isbn100763766321);krebs等人(编辑),theencyclopediaofmolecularbiology,blackwellscienceltd.,1994(isbn0-632-02182-9);和meyersr.a.(编辑),molecularbiologyandbiotechnology:acomprehensivedeskreference,vchpublishers,inc.,1995(isbn1-56081-569-8)。除非另有说明,所有百分比均以重量计,所有溶剂混合物比例以体积计。所有温度以摄氏度计。iv.包含昆虫害虫多核苷酸的核酸分子a.概述本文中描述了可用于控制昆虫害虫的核酸分子。在一些实例中,昆虫害虫是鞘翅目和/或半翅目昆虫害虫。描述的核酸分子包括靶多核苷酸(例如,天然基因、和非编码多核苷酸)、dsrna、sirna、shrna、hprna、和mirna。例如,在一些实施方案中描述了dsrna、mirna、sirna、shrna和/或hprna分子,其可与鞘翅目和/或半翅目害虫中的一种或多种天然核酸的全部或部分特异性地互补。在这些和另外的实施方案中,天然核酸可以是一种或多种靶基因,其产物可以例如但不限于:涉及幼虫/若虫发育。本文中描述的核酸分子在导入包含至少一种与该核酸分子特异性互补的天然核酸的细胞中时,可以在细胞中启动rnai,随后降低或消除该天然核酸的表达。在一些实例中,借助于与靶基因特异性互补的核酸分子降低或消除靶基因表达,可导致害虫的生长、发育和/或进食停止。在一些实施方案中,可以选择昆虫害虫中的至少一种靶基因,其中靶基因包含copigamma(seqidno:1或seqidno:87)。在具体的实例中,选自鞘翅目和/或半翅目害虫中的靶基因,其中该靶基因包含新的核苷酸序列,该序列包含copigamma(seqidno:1或seqidno:87)。在一些实施方案中,靶基因可以是包含这样的多核苷酸的核酸分子,该多核苷酸能够在计算机上翻译成包含连续氨基酸序列的多肽:所述连续氨基酸序列与copigamma(seqidno:1或seqidno:87)多核苷酸的蛋白质产物的氨基酸序列至少约85%相同(例如,至少84%、85%、约90%、约95%、约96%、约97%、约98%、约99%、约100%、或100%相同)。靶基因可以是昆虫害虫中的任何核酸,对该序列的转录后抑制会对该害虫的生长和/或存活造成有害影响,从而,例如,给植物提供针对该害虫的保护效益。在具体的实例中,靶基因是这样的核酸分子,其包含能够在计算机上反向翻译为多肽的多核苷酸,所述多肽包含与新核苷酸序列seqidno:1或seqidno:87的蛋白产物的氨基酸序列至少约85%相同、约90%相同、约95%相同、约96%相同、约97%相同、约98%相同、约99%相同、约100%相同、或100%相同的连续氨基酸序列。依照一些实施方案提供了dna,其表达生成包含这样的多核苷酸的rna分子,该多核苷酸与由昆虫(例如鞘翅目和/或半翅目)害虫中的编码多核苷酸编码的天然rna分子的全部或部分特异性地互补。在一些实施方案中,在昆虫害虫摄入表达的rna分子后,可以获得害虫细胞中编码多核苷酸的下调。在特定的实施方案中,害虫细胞中编码序列的下调可以对害虫的生长、发育、和/或存活产生有害影响。在一些实施方案中,靶多核苷酸包括转录的非编码性rna,诸如5’utr;3’utr;剪接前导;内含子;末端内含子(outron)(例如,随后在反式剪接中修饰的5'utrrna);donatron(例如,提供反式剪接的供体序列需要的非编码rna);和昆虫害虫靶基因的其他非编码转录rna。这样的多核苷酸可以衍生自单顺反子和多顺反子基因两者。因此,本文中结合一些实施方案还描述了包含至少一种多核苷酸的irna分子(例如,dsrna、mirna、sirna、shrna和hprna),所述多核苷酸与昆虫(例如鞘翅目和/或半翅目)害虫中的靶核酸的全部或部分特异性地互补。在一些实施方案中,irna分子可以包含与多个靶核酸,例如2、3、4、5、6、7、8、9、10、或更多个靶核酸的全部或部分互补的多核苷酸。在特定的实施方案中,irna分子可以在体外产生,或通过遗传修饰的生物(如植物或细菌)在体内产生。还公开了cdna,其可以用于产生与昆虫害虫中的靶核酸的全部或部分特异性地互补的dsrna分子、sirna分子、mirna分子、shrna分子、和/或hprna分子。进一步描述了在实现特定宿主靶标的稳定转化中使用的重组dna构建体。转化的宿主靶标可以从重组dna构建体表达有效水平的dsrna、sirna、mirna分子、shrna分子、和/或hprna分子。因此,还描述了植物转化载体,其包含与在植物细胞中功能性的异源启动子可操作连接的至少一个多核苷酸,其中多核苷酸的表达生成包含与昆虫害虫中的靶核酸的全部或部分特异性地互补的连续核碱基串的rna分子。在特定实例中,可用于控制昆虫(例如鞘翅目和/或半翅目)害虫的核酸分子可以包括:从叶甲属或半翅目生物分离的包含copigamma(seqidno:1或seqidno:87)的天然核酸的全部或部分;核苷酸序列,其在表达时生成这样的rna分子:所述rna分子包含与由copigamma(seqidno:1或seqidno:87)编码的天然rna分子的全部或部分特异性地互补的多核苷酸;irna分子(例如,dsrna、mirna、sirna、shrna和hprna),所述irna分子包含与copigamma(seqidno:1或seqidno:87)的全部或部分特异性地互补的至少一个多核苷酸;可用于产生dsrna分子、sirna分子、mirna、shrna和/或hprna分子的cdna序列,所述分子与copigamma(seqidno:1或seqidno:87)的全部或部分特异性地互补;以及用于在实现特定宿主靶标的稳定转化中使用的重组dna构建体,其中转化的宿主靶标包含一种或多种前述的核酸分子。b.核酸分子本发明提供了,除其他事项外,抑制昆虫(例如鞘翅目和/或半翅目)害虫的细胞、组织、或器官中的靶基因表达的irna(例如,dsrna、mirna、sirna、shrna和hprna)分子;和能够在细胞或微生物中表达为irna分子以抑制昆虫害虫的细胞、组织、或器官中的靶基因表达的dna分子。本发明的一些实施方案提供了分离的核酸分子,其包含至少一个(例如,一个、两个、三个、或更多个)选自下组的多核苷酸:seqidno:1或seqidno:87中任一者;seqidno:1或seqidno:87中任一者的互补物;seqidno:1或seqidno:87中任一者的至少15个连续核苷酸的片段;seqidno:1或seqidno:87中任一者的至少15个连续核苷酸的片段的互补物;叶甲属生物(例如wcr)的包含seqidno:1的天然编码多核苷酸;半翅目生物的包含seqidno:87的天然编码多核苷酸;叶甲属生物的包含seqidno:1的天然编码多核苷酸的互补物;半翅目生物的包含seqidno:87的天然编码多核苷酸的互补物;叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列;半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列;叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列的互补物;半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列的互补物;叶甲属生物的包含seqidno:1的天然编码多核苷酸的至少15个连续核苷酸的片段;半翅目生物的包含seqidno:87的天然编码多核苷酸的至少15个连续核苷酸的片段;叶甲属生物的包含seqidno:1的天然编码多核苷酸的至少15个连续核苷酸的片段的互补物;半翅目生物的包含seqidno:87的天然编码多核苷酸的至少15个连续核苷酸的片段的互补物;叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段;半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列至少15个连续核苷酸的片段;叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段的互补物;以及半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段的互补物。在特定实施方案中,所述分离的多核苷酸与鞘翅目和/或半翅目害虫接触或被所述昆虫害虫摄取抑制该害虫的生长、发育和/或进食。在一些实施方案中,本发明的核酸分子可以包含至少一个(例如,一个、两个、三个、或更多个)dna,其能够在细胞或微生物中表达为irna分子以抑制鞘翅目和/或半翅目害虫的细胞、组织、或器官中的靶基因表达。这样的dna可以与在包含该dna分子的细胞中具备功能的启动子可操作连接,以启动或增强编码的能够形成dsrna分子的rna的转录。在一个实施方案中,所述至少一个(例如,一个、两个、三个、或更多个)dna可衍生自选自包含seqidno:1或seqidno:87的群组的多核苷酸。seqidno:1或seqidno:87的衍生物包括seqidno:1或seqidno:87的片段。在一些实施方案中,这样的片段可包含例如seqidno:1或seqidno:87的至少约15个连续核苷酸,或其互补物。因此,这样的片段可包含,例如,seqidno:1或seqidno:87的15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、40、50、60、70、80、90、100、110、120、130、140、150、160、170、180、190、200个或更多个连续核苷酸,或其互补物。在某些实例中,这样的片段可包含,例如,seqidno:1或seqidno:87的至少19个连续核苷酸,或其互补物。因此,seqidno:1或seqidno:87的片段可以包含,例如,seqidno:1或seqidno:87的15,16,17,18,19,20,21,约25个(例如,22,23,24,25,26,27,28,29),约30,约40,(例如,35,36,37,38,39,40,41,42,43,44和45),约50,约60,约70,约80,约90,约100,约110,约120,约130,约140,约150,约160,约170,约180,约190,约200或更多个连续核苷酸,或其互补物。一些实施方案包括将部分或完全稳定化的dsrna分子导入鞘翅目和/或半翅目害虫中以抑制鞘翅目和/或半翅目害虫的细胞、组织或器官中靶基因的表达。当表达为irna分子(例如,dsrna、mirna、sirna、shrna、和hprna)并被鞘翅目和/或半翅目害虫摄取时,包含seqidno:1或seqidno:87的一个或多个片段的多核苷酸可以引起下列一项或多项:鞘翅目和/或半翅目害虫的死亡、发育停止、生长抑制、性别比变化、产卵量(broodsize)减少、感染停止、和/或进食停止。例如,在一些实施方案中,提供这样的dsrna分子:该dsrna分子包含与鞘翅目和/或半翅目害虫靶基因序列基本同源的约15个至约300个、或约19个至约300个核苷酸的核苷酸序列,并且包含含有seqidno:1或seqidno:87的核苷酸序列的一个或多个片段。这种dsrna分子的表达可能例如导致摄取dsrna分子的鞘翅目和/或半翅目害虫中的死亡和/或生长抑制。在某些实施方案中,本发明提供的dsrna分子包含与来自包含seqidno:1或seqidno:87的靶基因的转录物互补的多核苷酸,和/或与seqidno:1或seqidno:87的片段互补的核苷酸序列,所述靶基因在昆虫害虫中的抑制导致对于害虫的生长、发育、或其他生物学功能必需的蛋白质或多核苷酸物质的减少或消除。选择的核苷酸序列可以与下列各项展现出约80%到约100%序列同一性:seqidno:1或seqidno:87中任一者;seqidno:1或seqidno:87中所示的核苷酸序列的连续片段;或前述任一者的互补物。例如,选定的多核苷酸可以与下列任一项展现出79%、80%、约81%、约82%、约83%、约84%、约85%、约86%、约87%、约88%、约89%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约98.5%、约99%、约99.5%、或约100%的序列同一性:seqidno:1或seqidno:87;seqidno:1或seqidno:87中所示的核苷酸序列的连续片段;和前述任一者的互补物。在一些实施方案中,能够在细胞或微生物中表达为irna分子以抑制靶基因表达的dna分子可以包含与存在于一个或多个靶昆虫害虫物种(例如鞘翅目或半翅目害虫物种)中的天然多核苷酸的全部或部分特异性地互补的单个多核苷酸,或者dna分子可以是从多个这样的特异性互补多核苷酸构建而成的嵌合体。在一些实施方案中,核酸分子可以包含以“间隔物”分开的第一和第二多核苷酸。间隔物可以是任何包含促进第一和第二多核苷酸之间的二级结构形成(在期望的情况下)的核苷酸序列的区域。在一个实施方案中,间隔物是mrna的有义或反义编码多核苷酸的一部分。或者,间隔物可以包含能够与核酸分子可操作连接的核苷酸或其同源物的任何组合。在一些实例中,间隔物可以是内含子(例如st-ls1内含子或rtm1内含子)。例如,在一些实施方案中,dna分子可以包含编码一种或多种不同irna分子的多核苷酸,其中每种不同irna分子分别包含第一多核苷酸和第二多核苷酸,其中第一和第二多核苷酸彼此互补。第一和第二多核苷酸在rna分子内可以通过间隔物连接。间隔物可以构成第一多核苷酸或第二多核苷酸的一部分。包含第一和第二多核苷酸的rna分子的表达可以通过第一和第二多核苷酸的特异性分子内碱基配对而导致dsrna分子的形成。第一多核苷酸或第二多核苷酸可以与对于昆虫害虫(例如鞘翅目和/或半翅目害虫)而言天然的多核苷酸(例如,靶基因、或转录的非编码多核苷酸)、其衍生物、或其互补多核苷酸基本上相同。dsrna核酸分子包含聚合核糖核苷酸的双链,并且可以包含对磷酸-糖主链或核苷的修饰。可以适宜调整rna结构的修饰以使得特异性抑制能够发生。在一个实施方案中,可以通过普遍存在的酶促过程来修饰dsrna分子,以便能够生成sirna分子。此酶促过程可以利用体外或体内的rna酶iii酶,如真核生物中的切丁酶。参见elbashir等人,(2001)nature411:494-8;以及hamilton和baulcombe(1999)science286(5441):950-2。切丁酶或功能等效的rna酶iii酶将较大的dsrna链和/或hprna分子切割成较小的寡核苷酸(例如,sirna),每个所述寡核苷酸长度为约19-25个核苷酸。由这些酶生成的sirna分子具有2到3个核苷酸3’突出物、和5’磷酸和3’羟基末端。由rna酶iii酶生成的sirna分子在细胞中解旋,并分成单链rna。然后,sirna分子与从靶基因转录的rna特异性地杂交,并且这两种rna分子随后通过固有的细胞rna降解机制而被降解。这个过程可以导致由靶生物中的靶基因编码的rna的有效降解或消除。结果导致靶向基因的转录后沉默。在一些实施方案中,通过内源rna酶iii酶从异源核酸分子产生的sirna分子可以有效介导鞘翅目和/或半翅目害虫中靶基因的下调。在一些实施方案中,核酸分子可以包括至少一种可被转录成单链rna分子的非天然存在的多核苷酸,所述单链rna分子能够在体内通过分子间杂交形成dsrna分子。这样的dsrna常常进行自组装,并且可以在昆虫(例如鞘翅目和/或半翅目)害虫的营养来源中提供,以实现靶基因的转录后抑制。在这些和另外的实施方案中,核酸分子可以包含两种不同的非天然存在的核苷酸序列,其中每种序列与昆虫害虫中的不同靶基因特异性地互补。当向例如鞘翅目和/或半翅目害虫以dsrna分子的形式提供这样的核酸分子时,dsrna分子抑制害虫中至少两种不同靶基因的表达。c.获得核酸分子可以使用昆虫(例如鞘翅目或半翅目)害虫中的多种多核苷酸作为靶标来设计核酸分子,如irna和编码irna的dna分子。然而,天然多核苷酸的选择并非是直截了当的过程。例如,鞘翅目或半翅目害虫的天然多核苷酸中仅有少数会是有效的靶标。特定的天然多核苷酸是否可以被本发明的核酸分子有效下调,抑或特定的天然多核苷酸的下调是否会对昆虫害虫的生长、发育和/或生存具有有害影响,都是无法肯定地预测的。绝大多数的鞘翅目和半翅目害虫天然多核苷酸,如自这些害虫分离的est(例如,如美国专利7,612,194中所列出的鞘翅目害虫多核苷酸),对害虫的生长、发育和/或存活没有有害影响。同样无法预测的是,在可以对害虫具有有害影响的天然多核苷酸中哪些能够在重组技术中使用,从而在宿主植物中表达与这样的天然多核苷酸互补的核酸分子,并且在进食后对害虫造成有害影响,同时不对宿主植物引起损害。在一些实施方案中,核酸分子(例如,要在昆虫(例如鞘翅目和/或半翅目)害虫的宿主植物中提供的dsrna分子)被选择为靶向这样的cdna,其编码对于害虫发育必需的蛋白质或蛋白质部分(如涉及代谢或分解代谢生物化学途径、细胞分裂、能量代谢、消化、宿主植物识别等的多肽)。如本文中描述的,靶害虫生物摄入含有一种或多种dsrna——所述dsrna的至少一个区段与靶害虫生物细胞中产生的rna的至少一个基本上相同的区段特异地互补——的组合物,可以导致靶生物的死亡或其他抑制。可以使用从昆虫害虫衍生的多核苷酸(dna或rna)来构建对该害虫侵染有抗性的植物细胞。例如,可以转化鞘翅目和/或半翅目害虫的宿主植物(例如,玉米或大豆),使其含有从鞘翅目和/或半翅目害虫衍生的如本文中提供的一种或多种多核苷酸。转化到宿主中的多核苷酸可以编码一种或多种rna,其在转化的宿主内的细胞或生物学流体中形成dsrna结构,这样,如果/当害虫与转基因宿主形成营养关系时,就有dsrna可用。这可以导致害虫细胞中一种或多种基因表达的阻抑,最终导致死亡或其生长或发育的抑制。因此,在一些实施方案中,靶向实质参与昆虫(例如鞘翅目和/或半翅目)害虫的生长、发育和生殖的基因。本发明中使用的其他靶基因可以包括例如那些在害虫的运动、迁移、生长、发育、感染性、和进食位置的建立中发挥重要作用的靶基因。因此,靶基因可以是管家基因或转录因子。另外,本发明中使用的天然昆虫害虫多核苷酸也可以从植物、病毒、细菌或昆虫基因的同源物(例如,直系同源物)衍生,所述同源物的功能对于本领域技术人员是已知的,且其多核苷酸与害虫的基因组中的靶基因可特异性地杂交。通过杂交鉴定已知核苷酸序列的基因同源物的方法是本领域技术人员已知的。在一些实施方案中,本发明提供了用于获得包含用于产生irna(例如,dsrna、mirna、sirna、shrna、和hprna)分子的多核苷酸的核酸分子的方法。一种这样的实施方案包括:(a)对一种或多种靶基因分析其在昆虫(例如鞘翅目和/或半翅目)害虫中在dsrna介导的基因阻抑后的表达、功能和表型;(b)用探针探查cdna或gdna文库,所述探针包含来自靶向的害虫的多核苷酸的全部或部分或其同源物,所述靶向的害虫在dsrna介导的阻抑分析中展现出改变的(例如,降低的)生长或发育表型;(c)鉴定与探针特异性杂交的dna克隆;(d)分离步骤(b)中鉴定的dna克隆;(e)对包含步骤(d)中分离的克隆的cdna或gdna片段测序,其中测序的核酸分子包含rna的全部或大部分或其同源物;和(f)化学合成基因的全部或大部分、mirna、sirna、hprna、mrna、shrna、或dsrna。在另外的实施方案中,用于获得包含用于产生大部分的irna(例如,dsrna、mirna、sirna、shrna、和hprna)分子的多核苷酸的核酸片段的方法包括:(a)合成第一和第二寡核苷酸引物,其与来自靶向的昆虫(例如鞘翅目和/或半翅目)害虫的天然多核苷酸的一部分特异性地互补;和(b)使用步骤(a)的第一和第二寡核苷酸引物扩增克隆载体中存在的cdna或gdna插入物,其中扩增的核酸分子包含mirna、sirna、mrna、hprna、shrna或dsrna分子的大部分。核酸可以通过多种途径分离、扩增、或产生。例如,可以通过pcr扩增从gdna或cdna文库衍生的靶多核苷酸(例如,靶基因或靶转录的非编码多核苷酸)、或其部分而获得irna(例如,dsrna、mirna、sirna、shrna、和hprna)分子。可以从靶生物提取dna或rna,并且可以使用本领域普通技术人员已知的方法从其制备核酸文库。可使用自靶生物生成的gdna或cdna文库进行靶基因的pcr扩增和测序。可以使用确认的pcr产物作为体外转录的模板以在最低限度的启动子的情况下生成有义和反义rna。或者,可以使用标准化学,如亚磷酰胺化学,通过许多技术(参见,例如ozaki等人,(1992)nucleicacidsresearch,20:5205-5214;和agrawal等人,(1990)nucleicacidsresearch,18:5419-5423),包括使用自动化dna合成仪(例如,p.e.biosystems,inc.(fostercity,calif.)392或394型dna/rna合成仪)的任一者来合成核酸分子。参见,例如,beaucage等人,(1992)tetrahedron,48:2223-2311;美国专利4,980,460、4,725,677、4,415,732、4,458,066、和4,973,679。也可以采用备选化学,其生成非天然主链基团,如硫代磷酸酯、氨基磷酸酯,等等。本发明的rna、dsrna、mirna、sirna、shrna、或hprna分子可以由本领域技术人员可以通过手动或自动反应以化学或酶促方式产生,或者在包含含有编码rna、dsrna、mirna、sirna、shrna、或hprna分子的多核苷酸的核酸分子的细胞中体内产生。还可以通过部分或完全有机合成产生rna,可以通过体外酶促或有机合成引入任何经修饰的核糖核苷酸。可以通过细胞rna聚合酶或噬菌体rna聚合酶(例如,t3rna聚合酶、t7rna聚合酶、和sp6rna聚合酶)合成rna分子。可用于克隆和表达多核苷酸的表达构建体是本领域中已知的。参见,例如,国际pct公布号wo97/32016;及美国专利5,593,874、5,698,425、5,712,135、5,789,214、和5,804,693。可以在导入细胞中之前纯化化学合成或通过体外酶促合成合成的rna分子。例如,可以通过用溶剂或树脂提取、沉淀、电泳、色谱法、或其组合从混合物纯化rna分子。或者,可以在不纯化或最小程度纯化的情况下使用化学合成或通过体外酶促合成而合成的rna分子,例如,以避免由于样品加工所致的损失。可以将rna分子干燥贮存或溶解在水溶液中。溶液可以含有缓冲剂或盐以促进dsrna分子双链体链的退火和/或稳定化。在实施方案中,可以通过单一自身互补的rna链或者从两条互补rna链形成dsrna分子。dsrna分子可以在体内或在体外合成。细胞的内源rna聚合酶可以在体内介导一条或两条rna链的转录,或者可以使用克隆的rna聚合酶在体内或在体外介导转录。通过宿主器官、组织、或细胞类型中的特异性转录(例如,通过使用组织特异性启动子进行);刺激宿主中的环境条件(例如,通过使用响应于感染、胁迫、温度、和/或化学诱导物的诱导型启动子);和/或在宿主的某个发育阶段或年龄人工造成转录(例如,通过使用发育阶段特异性启动子),在昆虫害虫中靶基因的转录后抑制可以是宿主靶向性的。形成dsrna分子的rna链(不论是体外还是体内转录的)可以也可以不是多聚腺苷酸化的,并且可以能够也可以不能被细胞翻译机制翻译成多肽。d.重组载体和宿主细胞转化在一些实施方案中,本发明还提供了用于导入细胞(例如,细菌细胞、酵母细胞、或植物细胞)中的dna分子,其中dna分子包含这样的多核苷酸,该多核苷酸在表达为rna并被昆虫(例如鞘翅目和/或半翅目)害虫摄取后,可实现对害虫的细胞、组织或器官中靶基因的阻抑。因此,一些实施方案提供了重组核酸分子,其包含能够在植物细胞中表达为irna(例如,dsrna、mirna、sirna、shrna、和hprna)分子以抑制昆虫害虫中靶基因表达的多核苷酸。为了启动或增强表达,这样的重组核酸分子可以包含一种或多种调节元件,所述调节元件可以与能够表达为irna的多核苷酸可操作连接。在植物中表达基因阻抑分子的方法是已知的,并且可以用于表达本发明的多核苷酸。参见,例如,国际pct公开号wo06/073727;和美国专利公开号2006/0200878a1)。在具体的实施方案中,本发明的重组dna分子可以包含编码可形成dsrna分子的rna的多核苷酸。这样的重组dna分子可以编码可形成dsrna分子的rna,其被摄取后能够抑制昆虫(例如鞘翅目和/或半翅目)害虫细胞中内源靶基因的表达。在许多实施方案中,转录的rna可以形成dsrna分子,所述dsrna分子可以以稳定化形式提供;例如,以发夹和茎和环结构的形式提供。在某些实施方案中,dsrna分子的一条链可以通过从与下述任意多核苷酸基本上同源的多核苷酸转录而形成:seqidno:1;seqidno:1的互补物;seqidno:1的至少15个连续核苷酸的片段;seqidno:1的至少15个连续核苷酸的片段的互补物;叶甲属生物(例如wcr)的包含seqidno:1的天然编码序列;叶甲属生物的包含seqidno:1的天然编码序列的互补物;叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列;叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列的互补物;叶甲属生物的包含seqidno:1的天然编码序列的至少15个连续核苷酸的片段;叶甲属生物的包含seqidno:1的天然编码序列的至少15个连续核苷酸的片段的互补物;叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段;和叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段的互补物。在其他实施方案中,dsrna分子的一条链可以通过从与下述选自下组的多核苷酸基本上同源的多核苷酸转录而形成:seqidno:87;seqidno:87的互补物;seqidno:87的至少15个连续核苷酸的片段;seqidno:87的至少15个连续核苷酸的片段的互补物;半翅目生物的包含seqidno:87的天然编码序列;半翅目生物的包含seqidno:87的天然编码序列的互补物;半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列;半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列的互补物;半翅目生物的包含seqidno:87的天然编码序列的至少15个连续核苷酸的片段;半翅目生物的包含seqidno:87的天然编码序列的至少15个连续核苷酸的片段的互补物;半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段;和半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段的互补物。在特定的实施方案中,编码可形成dsrna分子的重组dna分子可包含编码区,其中至少两个多核苷酸如此排列,使得相对于至少一个启动子,一个多核苷酸处于有义取向,另一多核苷酸处于反义取向,其中该有义多核苷酸和该反义多核苷酸通过间隔物连接或联接,所述间隔物具有,例如,约5个(~5)至约一千个(~1000)核苷酸。间隔物可以在有义和反义多核苷酸之间形成环。有义多核苷酸或反义多核苷酸可以与靶基因(例如,包含seqidno:1或seqidno:87的基因)或其片段基本上同源。然而,在一些实施方案中,重组dna分子可以编码可以形成没有间隔物的dsrna分子的rna。在实施方案中,有义编码多核苷酸和反义编码多核苷酸可以是不同的长度。通过在本发明的重组核酸分子中创建适当的表达盒,可以容易地将鉴定为对昆虫害虫具有有害影响或者针对该害虫的植物保护效果的多核苷酸掺入表达的dsrna分子中。例如,可以通过如下步骤将这样的多核苷酸表达为具有茎和环结构的发夹:采用对应于靶基因多核苷酸(例如,seqidno:1或seqidno:87,或其片段)的第一区段;将此多核苷酸与第二区段间隔物区连接,所述第二区段间隔物区与第一区段不是同源或互补的;并将这与第三区段连接,其中第三区段的至少一部分与第一区段基本上互补。这样的构建体通过第一区段与第三区段的分子内碱基配对而形成茎和环结构,其中环结构形式包含第二区段。参见,例如,美国专利公开文本2002/0048814和2003/0018993;以及国际pct公开文本wo94/01550和wo98/05770。可以例如以双链结构诸如茎环结构(例如,发夹)形式生成dsrna分子,由此通过共表达靶基因的片段(例如在另外的植物表达盒上的靶基因的片段)增强靶向天然鞘翅目和/或半翅目害虫序列的sirna的产生,这可导致sirna产生增强,或者降低甲基化以防止dsrna发夹启动子的转录基因沉默。本发明的实施方案包括将本发明的重组核酸分子导入植物中(即,转化)以实现一种或多种irna分子的昆虫(例如鞘翅目和/或半翅目)害虫抑制水平的表达。重组dna分子可以例如是载体,如线性或闭合环状质粒。载体系统可以是单一载体或质粒,或者共同含有要导入宿主基因组中的总dna的两个或更多个载体或质粒。另外,载体可以是表达载体。可以例如将本发明的核酸适当地插入载体中,在一种或多种宿主中发挥功能以驱动连接的编码多核苷酸或其他dna元件表达的合适启动子控制下。许多载体可用于此目的,适当载体的选择主要将取决于要插入载体中的核酸的大小和要用载体转化的特定宿主细胞。每种载体含有不同的组分,根据其功能(例如,dna扩增或dna表达)及与其相容的特定宿主细胞而定。为了对转基因植物赋予对昆虫(例如鞘翅目和/或半翅目)害虫的保护,例如可以在重组植物的组织或流体内将重组dna转录成irna分子(例如,形成dsrna分子的rna分子)。irna分子可包含与可引起宿主植物物种损害的害虫内的相应转录多核苷酸基本上同源且可特异性杂交的多核苷酸。例如,害虫可以通过摄取包含irna分子的转基因宿主植物的细胞或流体而接触在转基因宿主植物细胞中转录的irna分子。因此,在特定实例中,靶基因的表达在侵染转基因宿主植物的鞘翅目和/或半翅目害虫内受到irna分子阻抑。在一些实施方案中,对靶鞘翅目和/或半翅目害虫中靶基因表达的阻抑可以导致植物被保护免遭害虫攻击。为了使irna分子能够被投送给与已经用本发明的重组核酸分子转化的植物细胞存在营养关系的害虫,需要在植物细胞中表达(即,转录)irna分子。因此,重组核酸分子可以包含与一种或多种调节元件(诸如在宿主细胞中发挥功能的异源启动子元件)可操作连接的本发明的多核苷酸,所述宿主细胞诸如其中要扩增核酸分子的细菌细胞,或其中要表达核酸分子的植物细胞。适合于用于本发明的核酸分子的启动子包括诱导型、病毒的、合成的、或组成型的那些启动子,它们都是本领域中熟知的。描述这样的启动子的非限制性实例包括美国专利6,437,217(玉米rs81启动子);5,641,876(水稻肌动蛋白启动子);6,426,446(玉米rs324启动子);6,429,362(玉米pr-1启动子);6,232,526(玉米a3启动子);6,177,611(组成型玉米启动子);5,322,938、5,352,605、5,359,142、和5,530,196(camv35s启动子);6,433,252(玉米l3油质蛋白启动子);6,429,357(水稻肌动蛋白2启动子和稻肌动蛋白2内含子);6,294,714(光诱导型启动子);6,140,078(盐诱导型启动子);6,252,138(病原体诱导型启动子);6,175,060(磷缺乏诱导型启动子);6,388,170(双向启动子);6,635,806(γ薏苡辛(coixin)启动子);及美国专利公开号2009/757,089(玉米叶绿体醛缩酶启动子)。其他启动子包括胆脂碱合酶(nos)启动子(ebert等人,(1987)proc.natl.acad.sci.usa84(16):5745-9)和章鱼碱合酶(ocs)启动子(两者都在根癌土壤杆菌的肿瘤诱导质粒上携带);花椰菜花叶病毒组(caulimovirus)启动子,如花椰菜花叶病毒(camv)19s启动子(lawton等人,(1987)plantmol.biol.9:315-24);camv35s启动子(odell等人,(1985)nature313:810-2;玄参花叶病毒35s启动子(walker等人,(1987)proc.natl.acad.sci.usa84(19):6624-8);蔗糖合酶启动子(yang和russell(1990)proc.natl.acad.sci.usa87:4144-8);r基因复合物启动子(chandler等人,(1989)plantcell1:1175-83);叶绿素a/b结合蛋白基因启动子;camv35s(美国专利5,322,938、5,352,605、5,359,142、和5,530,196);fmv35s(美国专利6,051,753和5,378,619);pclsv启动子(美国专利5,850,019);scp1启动子(美国专利6,677,503);和agrtu.nos启动子(genbanktm登录号v00087;depicker等人,(1982)j.mol.appl.genet.1:561-573;bevan等人,(1983)nature304:184-7)。在特定的实施方案中,本发明的核酸分子包含组织特异性启动子,如根特异性启动子。根特异性启动子驱动专门或优先在根组织中表达可操作连接的编码多核苷酸。根特异性启动子的实例是本领域中已知的。参见,例如,美国专利5,110,732;5,459,252和5,837,848;opperman等人,(1994)science263:221-3;和hirel等人,(1992)plantmol.biol.20:207-18。在一些实施方案中,可以在两个根特异性启动子之间克隆依照本发明的用于鞘翅目和/或半翅目害虫控制的多核苷酸或片段,所述根特异性启动子相对于所述多核苷酸或片段以相反的转录方向排布,在转基因植物细胞中可操作,并且在转基因植物细胞中表达以在转基因植物细胞中产生rna分子,rna分子随后可形成dsrna分子,如上文所述。昆虫害虫可以摄取植物组织中表达的irna分子,从而实现对靶基因表达的阻抑。其他可任选地与核酸可操作连接的调节元件包括5'utr,5'utr作为位于启动子元件和编码多核苷酸之间的翻译前导元件发挥功能。翻译前导元件存在于完全加工的mrna中,并且其可以影响初级转录物的加工和/或rna的稳定性。翻译前导元件的例子包括玉米和矮牵牛热休克蛋白前导物(美国专利no.5,362,865)、植物病毒外壳蛋白前导序列、植物rubisco前导序列等。参见,例如,turner和foster(1995)molecularbiotech.3(3):225-36。5'utr的非限制性实例包括gmhsp(美国专利no.5,659,122);phdnak(美国专利5,362,865);atant1;tev(carrington和freed(1990)j.virol.64:1590-7);和agrtunos(genbanktm登录号v00087;和bevan等人,(1983)nature304:184-7)。其他任选地与核酸可操作连接的调节元件还包括3'非翻译元件、3'转录终止区、或多腺苷酸化区。这些是位于多核苷酸下游的遗传元件,并且包括提供多腺苷酸化信号的多核苷酸、和/或能够影响转录或mrna加工的其他调节信号。多腺苷酸化信号在植物中发挥功能,导致多腺苷酸化的核苷酸被添加至mrna前体的3'端。多腺苷酸化元件可以源自多种植物基因或t-dna基因。3'转录终止区的一个非限制性实例是胆脂碱合酶3'区(nos3';fraley等人,(1983)proc.natl.acad.sci.usa80:4803-7)。在ingelbrecht等人,(1989)plantcell1:671-80中提供了使用不同3'非翻译区的实例。多腺苷酸化信号的非限制性实例包括来自豌豆rbcs2基因的信号(ps.rbcs2-e9;coruzzi等人,(1984)emboj.3:1671-9)和agrtu.nos(登录号e01312)。一些实施方案可以包括植物转化载体,该植物转化载体包含分离纯化的dna分子,该dna分子包含与本发明的一种或多种多核苷酸可操作连接的至少一种上文描述的调节元件。在表达时,一种或多种多核苷酸生成一种或多种包含多核苷酸的irna分子,所述多核苷酸与昆虫(例如鞘翅目和/或半翅目)害虫中的天然rna分子的全部或部分特异性地互补。因此,多核苷酸可以包含编码靶向的鞘翅目和/或半翅目害虫rna转录物内存在的多核糖核苷酸的全部或部分的区段,并且可以包含被靶向的害虫转录物的全部或部分的反向重复。植物转化载体可以含有与超过一种靶多核苷酸特异性互补的多核苷酸,由此容许产生超过一种dsrna以抑制靶害虫的一个或多个群体或物种的细胞中两种或更多种基因的表达。可以将与不同基因中存在的多核苷酸特异性互补的多核苷酸区段组合成单一复合核酸分子,以便在转基因植物中表达。这样的区段可以是连续的或者由间隔物分开。在一些实施方案中,可以通过在同一质粒中依次插入另外的多核苷酸来修饰已经含有本发明的至少一个多核苷酸的本发明质粒,其中所述另外的多核苷酸与原有的至少一个多核苷酸可操作连接于相同的调节元件。在一些实施方案中,核酸分子可以设计为抑制多种靶基因。在一些实施方案中,要抑制的多种基因可以获自相同的昆虫(例如鞘翅目和/或半翅目)害虫物种,这样做可以增强核酸分子的有效性。在其他实施方案中,基因可以来自不同的昆虫害虫,这样可以拓宽药剂有效的害虫的范围。当靶向多种基因以实现阻抑、或表达和阻抑的组合时,可以工程化多顺反子dna元件。本发明的重组核酸分子或载体可以包含赋予转化的细胞,诸如植物细胞可选择表型的选择标志物。也可以使用选择标志物来选择包含本发明的重组核酸分子的植物或植物细胞。标志物可以编码杀生物剂抗性、抗生素抗性(例如,卡那霉素、遗传霉素(g418)、博来霉素、潮霉素,等等)、或除草剂耐性(例如,草甘膦等)。选择标志物的实例包括但不限于编码卡那霉素抗性并且可以使用卡那霉素、g418等选择的neo基因;编码双丙氨磷抗性的bar基因;编码草甘膦耐性的突变体epsp合酶基因;赋予对溴苯腈的抗性的腈水解酶基因;赋予咪唑啉酮或磺酰脲耐性的突变体乙酰乳酸合酶基因(als);和甲氨蝶呤抗性dhfr基因。可用多种选择标志物,其赋予对氨苄青霉素、博来霉素、氯霉素、庆大霉素、潮霉素、卡那霉素、林可霉素、甲氨蝶呤、膦丝菌素、嘌呤霉素、大观霉素、利福平、链霉素和四环素等的抗性。这样的选择标志物的实例展示于,例如美国专利5,550,318;5,633,435;5,780,708和6,118,047。本发明的重组核酸分子或载体还可以包含可筛选标志物。可以使用可筛选标志物来监测表达。示例性筛选标志物包括β-葡糖醛酸糖苷酶或uida基因(gus),其编码已知的多种生色底物的酶(jefferson等人,(1987)plantmol.biol.rep.5:387-405);r基因座基因,其编码调节植物组织中花色素苷色素(红色)产生的产物(dellaporta等人,(1988)"molecularcloningofthemaizer-njallelebytransposontaggingwithac."收录于18thstadlergeneticssymposium,p.gustafson和r.appels编辑,(newyork:plenum),pp.263-82);β-内酰胺酶基因(sutcliffe等人,(1978)proc.natl.acad.sci.usa75:3737-41);编码已知多种生色底物的酶(例如,padac,一种生色头孢菌素)的基因;萤光素酶基因(ow等人,(1986)science234:856-9);xyle基因,其编码能转化生色儿茶酚的儿茶酚双加氧酶(zukowski等人,(1983)gene46(2-3):247-55);淀粉酶基因(ikatu等人,(1990)bio/technol.8:241-2);酪氨酸酶基因,其编码能够将酪氨酸氧化为dopa和多巴醌(dopaquinone)(其继而缩合成黑色素)的酶(katz等人,(1983)j.gen.microbiol.129:2703-14));和α-半乳糖苷酶。在一些实施方案中,在用于创建转基因植物和在植物中表达异源核酸的方法中,可以使用如上文所描述的重组核酸分子来制备表现出对昆虫(例如鞘翅目和/或半翅目)害虫的易感性降低的转基因植物。例如,可以通过将编码irna分子的核酸分子插入植物转化载体中,并将这些导入植物中来制备植物转化载体。用于宿主细胞转化的适合方法包括任何能将dna导入细胞中的方法,如通过原生质体转化(参见,例如,美国专利5,508,184),通过干燥/抑制介导的dna摄取(参见,例如,potrykus等人.(1985)mol.gen.genet.199:183-8),通过电穿孔(参见,例如,美国专利5,384,253),通过用碳化硅纤维搅拌(参见,例如,美国专利5,302,523和5,464,765),通过土壤杆菌介导的转化(参见,例如,美国专利5,563,055;5,591,616;5,693,512;5,824,877;5,981,840;和6,384,301),以及通过加速dna包被的颗粒(参见,例如,美国专利5,015,580、5,550,318、5,538,880、6,160,208、6,399,861、和6,403,865),等。特别可用于转化玉米的技术描述于例如美国专利7,060,876和5,591,616;以及国际pct公布wo95/06722中。通过应用诸如此类的这些技术,几乎任何物种的细胞都可被稳定转化。在一些实施方案中,转化dna被整合到宿主细胞的基因组中。在多细胞物种的情况下,转基因细胞可再生为转基因生物。可以使用任何这些技术来产生转基因植物,例如在转基因植物的基因组中包含编码一种或多种irna分子的一种或多种核酸。用于将表达载体导入植物中的广泛使用的方法是基于土壤杆菌的天然转化系统。根癌土壤杆菌和发根土壤杆菌是将植物细胞遗传转化的植物致病性土壤细菌。根癌土壤杆菌和发根土壤杆菌的ti和ri质粒分别携带负责植物遗传转化的基因。ti(诱导肿瘤)-质粒含有被称为t-dna的大片段,其被转移到转化的植物中。ti质粒的另一个片段,vir区,负责t-dna的转移。t-dna区的边界为末端重复序列。在修饰的二元载体中,肿瘤诱导基因已缺失,vir区的功能用于转移以t-dna边界元件为界的外来dna。t区还可含有用于转基因细胞和植物有效恢复的选择标志物、以及插入用于转移诸如编码核酸的dsrna之类多核苷酸的多克隆位点。因此,在一些实施方案中,植物转化载体来源于根癌土壤杆菌的ti质粒(参见,例如,美国专利4,536,475、4,693,977、4,886,937、和5,501,967;以及欧洲专利no.ep0122791)或发根土壤杆菌的ri质粒。其他的植物转化载体包括,例如但不限于,由herrera-estrella等人,(1983)nature303:209-13;bevan等人,(1983)nature304:184-7;klee等人,(1985)bio/technol.3:637-42;以及在欧洲专利no.ep0120516中所描述的那些载体,以及从任何前述文献来源的那些载体。可以修饰天然地与植物相互作用的其他细菌,如中华根瘤菌属、根瘤菌属、和中慢生根瘤菌属,以介导到许多各种各样的植物中的基因转移。通过获取卸甲ti质粒和适合的二元载体,可以使这些植物相关的共生细菌能够胜任基因转移。在提供外源dna到受体细胞的递送之后,通常鉴定出转化的细胞用于进一步培养和植物再生。为了提高鉴定转化细胞的能力,人们可能期望采用如前提出的选择或筛选标志物基因,与用来再生转化体的转化载体一道使用。在采用选择标志物的情况下,通过使细胞暴露于选择剂或药剂,鉴定出在潜在转化的细胞群中的转化细胞。在采用筛选标志物的情况下,可针对期望的标志物基因性状来筛选细胞。暴露于选择剂后存活的细胞、或者在筛选测定中已被评分为阳性的细胞,可以在支持植物再生的介质中进行培养。在一些实施方案中,可通过包含其他物质,如生长调节剂来改良任何适合的植物组织培养基(例如,ms和n6培养基)。可将组织维持在具有生长调节剂的基本培养基上,直到可得到足够的组织用于启动植物再生工作时为止,或者在重复多轮的手动选择之后,直到组织形态适合于再生时为止(例如,至少2周),然后转移到有助于芽形成的介质中。定期转移培养物,直到已经出现充分的芽形成时为止。一旦形成芽,将它们转移到有助于根形成的介质中。一旦形成足够的根,可将植物转移到土壤中,以便进一步生长和成熟。为了证实再生植物中感兴趣核酸分子(例如,编码一种或多种irna分子的dna,所述irna分子抑制鞘翅目和/或半翅目害虫中的靶基因表达)的存在,可以进行多种测定法。这样的测定法例如包括:分子生物学测定,如southern和northern印迹、pcr、和核酸测序;生物化学测定,如检测蛋白质产物的存在,例如通过免疫学手段(elisa和/或western印迹)或借助于酶功能;植物部分测定,如叶或根测定;和再生的全植物的表型分析。可例如通过使用对感兴趣核酸分子特异的寡核苷酸引物进行pcr扩增来分析整合事件。pcr基因分型应当理解为包括但不限于,来源于分离的宿主植物愈伤组织的gdna的聚合酶链反应(pcr)扩增,所述愈伤组织预期含有整合到基因组中的感兴趣核酸分子,然后进行标准克隆和pcr扩增产物的序列分析。pcr基因分型方法已被很好地描述(例如rios,g等人,(2002)plantj.32:243-53),并可应用于来源于任何植物物种(例如,玉米或大豆)或组织类型(包括细胞培养物)的gdna。采用依赖于土壤杆菌的转化方法形成的转基因植物一般含有插入一个染色体中的单个重组dna。该单个重组dna的多核苷酸被称为“转基因事件”或“整合事件”。这样的转基因植物对于插入的外源多核苷酸而言是杂合的。在一些实施方案中,通过含有单个外源基因的独立分离的转基因植物与自身(例如t0植物)有性交配(自交)以产生tl种子,可获得相对于转基因为纯合的转基因植物。所产生的tl种子的四分之一相对于所述转基因是纯合的。萌发tl种子产生的植物可用于测试杂合性,所述测试一般使用snp测定或热扩增测定,使得允许在杂合子和纯合子之间进行区分(即,接合型测定)。在特定的实施方案中,在植物细胞中产生具有昆虫(例如鞘翅目和/或半翅目)害虫-抑制效果的至少2、3、4、5、6、7、8、9种或10种或更多种不同irna分子。可以从引入不同转化事件中的多种核酸、或从引入单一转化事件中的单一核酸表达irna分子(例如,dsrna分子)。在一些实施方案中,在单一启动子的控制下表达多个irna分子。在其他实施方案中,在多个启动子的控制下表达多个irna分子。可以表达包含多个多核苷酸的单一irna分子,所述多核苷酸各自与在相同昆虫害虫物种的不同群体中或在不同的昆虫害虫物种中的一个或多个昆虫害虫内的不同基因座(例如由seqidno:1或seqidno:87定义的基因座)同源。除了用重组核酸分子直接转化植物之外,可通过使具有至少一个转基因事件的第一植物与缺乏这种事件的第二植物杂交来制造转基因植物。例如,可将包含编码irna分子的多核苷酸的重组核酸分子导入易于转化的第一植物品系中而产生转基因植物,其中转基因植物可与第二植物品系杂交而使编码irna分子的多核苷酸渗入到第二植物品系中。本发明还包括含有本发明的一种或多种序列的商业产品。具体的实施方案包括自含有本发明一种或多种核苷酸序列的重组植物或种子产生的商业产品。包含本发明的一种或多种序列的商业产品意在包括,但不限于:植物的粕、油类、碾碎的或完整的籽粒或种子,或包含含有本发明的一种或多种序列的重组植物或种子的任何粕、油、或碾碎的或完整的籽粒的任何食物产品或动物饲料产品。在一种或多种商品或商业产品中的检出本发明的一种或多种序列,事实上证明该商品或商业产品是由为了利用dsrna介导的基因阻遏方法来控制鞘翅目和/或半翅目植物害虫的目的而设计为表达本发明的一种或多种序列的转基因植物生产的。在一些方面,包括由自转化的植物细胞衍生的转基因植物产生的种子和商业产品,其中种子或商业产品包含可检出量的本发明的核酸。在一些实施方案中,例如,可以通过获得转基因植物并从它们制备食物或饲料来生产这样的商业产品。包含本发明的一种或多种多核苷酸的商业产品包括例如但不限于:植物的粕、油类、碾碎的或完整的籽粒或种子,和包含含有本发明的一种或多种核酸的重组植物或种子的任何粕、油、或碾碎的或完整的籽粒的任何食物产品。在一种或多种商品或商业产品中的检出本发明的一种或多种核酸,事实上证明该商品或商业产品是由为了控制昆虫(例如鞘翅目和/或半翅目)害虫的目的,设计为表达本发明的一种或多种irna分子的转基因植物生产的。在一些实施方案中,包含本发明的核酸分子的转基因植物或种子也可在其基因组中包含至少一个其他的转基因事件,包括但不限于:自其转录靶向昆虫害虫中与由seqidno:1或seqidno:87定义者不同的基因座的irna分子的转基因事件,所述不同的基因座举例而言可为选自下组的一个或多个基因座:caf1-180(美国专利申请公开号2012/0174258)、vatpasec(美国专利申请公开号2012/0174259)、rho1(美国专利申请公开号2012/0174260)、vatpaseh(美国专利申请公开号2012/0198586)、ppi-87b(美国专利申请公开号2013/0091600)、rpa70(美国专利申请公开号2013/0091601)、和rps6(美国专利申请公开号2013/0097730);自其转录靶向与鞘翅目和/或半翅目害虫不同的生物(例如,植物寄生性线虫)中的基因的irna分子的转基因事件;编码杀虫蛋白(例如,苏云金芽孢杆菌、产碱杆菌属(例如美国专利申请公开号2014/0033361)、或假单胞菌属(例如pct申请公开号wo2015038734)杀虫蛋白)的基因;除草剂耐性基因(例如提供对草甘膦耐性的基因);以及促成转基因植物中的期望表型的基因,所述期望表型例如产量增加、脂肪酸代谢改变、或细胞质雄性不育的恢复。在特定的实施方案中,可以在植物中将编码本发明irna分子的多核苷酸与其他昆虫控制和疾病性状组合以实现增强的植物疾病和昆虫损害的期望性状。例如,由于对所述性状的抗性的概率在田间将发生降低,组合采用独特作用模式的昆虫控制性状可以对受保护的转基因植物提供优越的持久性,该持久性优于含有单一控制性状的植物。v.鞘翅目和/或半翅目害虫中的靶基因阻抑a.概述在本发明的一些实施方案中,可以对鞘翅目和/或半翅目害虫提供至少一种可用于控制鞘翅目和/或半翅目害虫的核酸分子,其中所述核酸分子在害虫中导致rnai介导的基因沉默。在特定的实施方案中,可以对鞘翅目和/或半翅目宿主提供irna分子(例如,dsrna、mirna、sirna、shrna、和hprna)。在一些实施方案中,可以通过使核酸分子与害虫接触来对害虫提供可用于控制鞘翅目和/或半翅目害虫的核酸分子。在这些和另外的实施方案中,可以在害虫的进食基质,例如营养组合物中提供可用于控制鞘翅目和/或半翅目害虫的核酸分子。在这些和另外的实施方案中,可以通过摄取被害虫摄取的包含核酸分子的植物材料来提供可用于控制鞘翅目和/或半翅目害虫的核酸分子。在某些实施方案中,核酸分子通过表达导入植物材料中的重组核酸而存在于植物材料中,所述导入例如通过用包含重组核酸的载体转化植物细胞,并从转化的植物细胞再生植物材料或全植物来进行。b.rna介导的靶基因阻抑在实施方案中,本发明提供了irna分子(例如,dsrna、mirna、sirna、shrna、和hprna),可以设计此类分子使之靶向昆虫害虫(例如,鞘翅目(例如wcr或ncr)或半翅目(例如bsb)害虫)的转录组中的必需天然多核苷酸(例如,必需基因),例如设计有至少一条链包含与靶多核苷酸特异性互补的多核苷酸的irna分子。如此设计的irna分子的序列可以与靶多核苷酸相同,或者可以含有不会阻止irna分子与其靶多核苷酸之间的特异性杂交的错配。本发明的irna分子可以在用于昆虫(例如鞘翅目和/或半翅目)害虫中基因阻抑的方法中使用,由此降低由害虫对植物(例如,包含irna分子的受保护的转化植物)引起的损害水平或发生率。如本文中使用的,术语“基因阻抑”是指用于降低由于基因转录成mrna及随后mrna翻译而产生的蛋白质水平,包括降低基因或编码多核苷酸的蛋白质表达,包括任何公知的转录后抑制表达和转录阻抑表达的方法。通过从靶向阻抑的基因转录的mrna的全部或部分与用于阻抑的相应irna分子之间的特异性同源性而介导转录后抑制。另外,转录后抑制是指通过核糖体结合的细胞中可用的mrna量的实质性且可测量的降低。在其中rnai分子是dsrna分子的实施方案中,切丁酶这种酶可以将dsrna分子切割成短sirna分子(大约20个核苷酸长)。借助于切丁酶对dsrna分子的活性而生成的双链sirna分子可以分成两个单链sirna:“乘客链”和“引导链”。乘客链可以被降解,而引导链可以掺入risc中。通过引导链与mrna分子的特异性互补多核苷酸的特异性杂交,随后通过酶argonaute(risc复合物的催化组分)切割而发生转录后抑制。在本发明的实施方案中,可以使用任何形式的irna分子。本领域技术人员会理解的是,在制备过程中及在对细胞提供irna分子的步骤过程中,dsrna分子一般比单链rna分子更稳定,并且一般在细胞中也是更稳定的。因此,例如,虽然在一些实施方案中sirna和mirna分子可能是同等有效的,但是dsrna分子可以由于其稳定性而被选用。在特定的实施方案中,提供了包含多核苷酸的核酸分子,所述多核苷酸可以在体外表达以产生irna分子,所述irna分子与由昆虫(例如鞘翅目和/或半翅目)害虫基因组内的多核苷酸编码的核酸分子基本上同源。在某些实施方案中,体外转录的irna分子可以是包含茎环结构的稳定化的dsrna分子。在昆虫害虫接触体外转录的irna分子后,可以发生对该昆虫害虫中靶基因(例如,必需基因)的转录后抑制。在本发明的一些实施方案中,在用于对昆虫(例如鞘翅目和/或半翅目)害虫中靶基因的转录后抑制的方法中利用包含多核苷酸的至少15个连续核苷酸(例如至少19个连续核苷酸)的核酸分子的表达,其中所述多核苷酸选自下组:seqidno:1;seqidno:1的互补物;seqidno:1的至少15个连续核苷酸的片段;seqidno:1的至少15个连续核苷酸的片段的互补物;叶甲属生物的包含seqidno:1的天然编码多核苷酸;自叶甲属生物的包含seqidno:1的天然编码多核苷酸表达的rna的互补物;叶甲属生物的包含seqidno:1的天然编码多核苷酸;自叶甲属生物的包含seqidno:1的天然编码多核苷酸表达的rna的互补物;自叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列表达的rna的互补物;叶甲属生物(例如wcr)的包含seqidno:1的天然编码序列的至少15个连续核苷酸的片段;叶甲属生物的包含seqidno:1的天然编码序列的至少15个连续核苷酸的片段的互补物;叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段;和叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段的互补物。在某些实施方案中,可以使用与前述任一项至少约80%相同(例如,79%,约80%、约81%、约82%、约83%、约84%、约85%、约86%、约87%、约88%、约89%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%、约100%、和100%)的核酸分子的表达。在本发明的特定实施方案中,在用于转录后抑制半翅目害虫中的靶基因的方法中利用包含核苷酸序列的至少15个连续核苷酸的核酸分子的表达,其中所述核苷酸序列选自下组:seqidno:87;seqidno:87的互补物;seqidno:87的至少15个连续核苷酸的片段;seqidno:87的至少15个连续核苷酸的片段的互补物;半翅目生物的包含seqidno:87的天然编码序列;半翅目生物的包含seqidno:87的天然编码序列的互补物;半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列;半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列的互补物;半翅目生物的包含seqidno:87的天然编码序列的至少15个连续核苷酸的片段;半翅目生物的包含seqidno:87的天然编码序列的至少15个连续核苷酸的片段的互补物;半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段;和半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段的互补物。在某些实施方案中,可以使用与前述任一项至少约80%相同(例如,80%、约81%、约82%、约83%、约84%、约85%、约86%、约87%、约88%、约89%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%、约100%、和100%)的核酸分子的表达。在这些实施方案和进一步的实施方案中,可以表达这样的核酸分子,其与昆虫(例如鞘翅目和/或半翅目)害虫的至少一个细胞中存在的rna分子特异性杂交。在本发明的特定实施方案中,在用于转录后抑制鞘翅目害虫中的靶基因的方法中可以利用包含核苷酸序列的至少15个连续核苷酸的核酸分子的表达,其中所述核苷酸序列选自下组:seqidno:1;seqidno:1的互补物;seqidno:1的至少15个连续核苷酸的片段;seqidno:1的至少15个连续核苷酸的片段的互补物;叶甲属生物(例如wcr)的包含seqidno:1的天然编码序列;叶甲属生物(例如wcr)的包含seqidno:1的天然编码序列的互补物;叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列;叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列的互补物;叶甲属生物(例如wcr)的包含seqidno:1的天然编码序列的至少15个连续核苷酸的片段;叶甲属生物的包含seqidno:1的天然编码序列的至少15个连续核苷酸的片段的互补物;叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段;和叶甲属生物的转录为包含seqidno:1的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段的互补物。在某些实施方案中,可以使用与前述任一项至少约80%相同(例如,80%、约81%、约82%、约83%、约84%、约85%、约86%、约87%、约88%、约89%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%、约100%、和100%)的核酸分子的表达。在这些实施方案和进一步的实施方案中,可以表达这样的核酸分子,其与鞘翅目害虫的至少一个细胞中存在的rna分子特异性杂交。在具体的实例中,这样的核酸分子可包含包含seqidno:1的核苷酸序列。在本发明的特定实施方案中,在用于转录后抑制半翅目害虫中的靶基因的方法中利用包含核苷酸序列的至少15个连续核苷酸的核酸分子的表达,其中所述核苷酸序列选自下组:seqidno:87;seqidno:87的互补物;seqidno:87的至少15个连续核苷酸的片段;seqidno:87的至少15个连续核苷酸的片段的互补物;半翅目生物的包含seqidno:87的天然编码序列;半翅目生物的包含seqidno:87的天然编码序列的互补物;半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列;半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列的互补物;半翅目生物的包含seqidno:87的天然编码序列的至少15个连续核苷酸的片段;半翅目生物的包含seqidno:87的天然编码序列的至少15个连续核苷酸的片段的互补物;半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段;和半翅目生物的转录为包含seqidno:87的天然rna分子的天然非编码序列的至少15个连续核苷酸的片段的互补物。在某些实施方案中,可以使用与前述任一项至少约80%相同(例如,80%、约81%、约82%、约83%、约84%、约85%、约86%、约87%、约88%、约89%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%、约100%、和100%)的核酸分子的表达。在这些实施方案和进一步的实施方案中,可以表达这样的核酸分子,其与半翅目害虫的至少一个细胞中存在的rna分子特异性杂交。在具体的实例中,这样的核酸分子可包含包含seqidno:87的核苷酸序列。本发明的一些实施方案的重要特征在于,rnai转录后抑制系统能够容忍靶基因中预期由于遗传突变、株系多态性、或进化趋异而可能发生的序列变异。导入的核酸分子可以不必与靶基因的初级转录产物或完全加工的mrna绝对同源,只要导入的核酸分子与靶基因的初级转录产物或完全加工的mrna可特异性杂交即可。而且,相对于靶基因的初级转录产物或完全加工的mrna,导入的核酸分子可以不必是全长的。使用本文的irna技术抑制靶基因是序列特异性的;即,靶向与irna分子基本上同源的多核苷酸进行遗传抑制。在一些实施方案中,可以使用包含具有与靶基因的一部分相同的核苷酸序列的多核苷酸的rna分子进行抑制。在这些和另外的实施方案中,可以使用包含相对于靶多核苷酸具有一个或多个插入、缺失和/或点突变的多核苷酸的rna分子。在特定的实施方案中,irna分子和靶基因的一部分可以共享例如至少约80%、至少约81%、至少约82%、至少约83%、至少约84%、至少约85%、至少约86%、至少约87%、至少约88%、至少约89%、至少约90%、至少约91%、至少约92%、至少约93%、至少约94%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%、至少约100%、和100%的序列同一性。或者,dsrna分子的双链体区可以与靶基因转录物的一部分可特异性地杂交。在可特异性杂交的分子中,展现出较大同源性的小于全长的多核苷酸可补偿较长的、同源性较低的多核苷酸。dsrna分子的双链体区中与靶基因转录物的一部分相同的多核苷酸的长度可以是至少约15、25、50、100、200、300、400、500个、或至少约1000个碱基。在一些实施方案中,可以使用大于20个到100个核苷酸的多核苷酸。在特定的实施方案中,可以使用大于约200个到300个核苷酸的多核苷酸。在特定的实施方案中,根据靶基因的大小,可以使用大于约500个到1000个核苷酸的多核苷酸。在某些实施方案中,可以将害虫(例如鞘翅目或半翅目害虫)中靶基因的表达在害虫细胞内抑制至少10%;至少33%;至少50%;或至少80%,使得发生显著抑制。显著抑制是指高于阈值的抑制,所述抑制导致可检出的表型(例如,生长停止、进食停止、发育停止、和诱导性死亡,等等),或与所抑制的靶基因对应的rna和/或基因产物的可检出降低。虽然在本发明的某些实施方案中,在害虫的基本上所有细胞中均发生抑制,但在其他实施方案中,仅在表达靶基因的细胞的一个子集中发生抑制。在一些实施方案中,细胞中的转录阻抑由细胞中展现出与启动子dna或其互补物的实质性序列同一性的dsrna分子的存在所介导,从而产生所谓的“启动子反式阻抑”。基因阻抑可以在可以摄取或接触此类dsrna分子的昆虫害虫中针对靶基因发挥效果,例如通过害虫摄取或接触含有dsrna分子的植物材料。在启动子反式阻抑中使用的dsrna分子可以特异性地设计为抑制或阻抑昆虫害虫细胞中一种或多种同源或互补多核苷酸的表达。美国专利5,107,065;5,759,829;5,283,184;和5,231,020中公开了通过反义或有义取向的rna进行转录后基因阻抑以调节植物细胞中的基因表达。c.对昆虫害虫提供的irna分子的表达可以许多体外或体内形式的任一种进行表达用于在昆虫(例如鞘翅目和/或半翅目)害虫中rnai介导的基因抑制的irna分子。然后,可以对害虫提供irna分子,例如通过使irna分子与害虫接触,或通过引起害虫摄取或以别的方式内在化irna分子。一些实施方案包括鞘翅目和/或半翅目害虫的经转化的宿主植物、经转化的植物细胞、和经转化的植物的后代。经转化的植物细胞和经转化的植物可以工程化改造为,例如,在异源启动子控制下表达一种或多种irna分子,以提供害虫保护效果。因此,当在昆虫害虫在进食期间食用转基因植物或植物细胞时,害虫可以摄取转基因植物或细胞中表达的irna分子。也可以将本发明的多核苷酸导入广泛而多样的原核和真核微生物宿主中以产生irna分子。术语“微生物”包括原核和真核物种,如细菌和真菌。基因表达的调变可以包括对此类表达的部分或完全阻抑。在另一个实施方案中,用于阻抑昆虫(例如鞘翅目和/或半翅目)害虫中基因表达的方法包括在害虫宿主的组织中提供基因阻抑量的至少一种如本文中所描述的多核苷酸在转录后形成的dsrna分子,其至少一个区段与害虫细胞内的mrna互补。昆虫害虫所摄取的dsrna分子,包括其修饰的形式,如mirna、sirna、shrna、或hprna分子,可以与包含含有seqidno:1或seqidno:87的核苷酸序列的分子转录的rna分子至少约80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、或约100%相同。因此,提供了用于制备dsrna分子的分离的且基本上纯化的核酸分子,包括但不限于非天然存在的多核苷酸和重组dna构建体,其在导入昆虫害虫时阻抑或抑制其中的内源编码多核苷酸或靶编码多核苷酸的表达。特定的实施方案提供了用于递送irna分子以转录后抑制昆虫(例如鞘翅目和/或半翅目)植物害虫中的一种或多种靶基因并控制该植物害虫群体的递送系统。在一些实施方案中,所述递送系统涉及包括对宿主转基因植物细胞或包含在宿主细胞中转录的rna分子的宿主细胞内容物的摄取。在这些和另外的实施方案中,创建转基因植物细胞或转基因植物,其含有可提供本发明的稳定化dsrna分子的重组dna构建体。包含编码特定irna分子的核酸的转基因植物细胞和转基因植物可以如下产生:采用重组dna技术(这些基础技术是本领域中熟知的)构建包含编码本发明的irna分子(例如,稳定化的dsrna分子)的多核苷酸的植物转化载体;以此转化植物细胞或植物;并以此生成含有转录的irna分子的转基因植物细胞或转基因植物。为了对转基因植物赋予针对昆虫(例如鞘翅目和/或半翅目)害虫的保护,可以例如将重组dna分子转录成irna分子,如dsrna分子、sirna分子、shrna分子、mirna分子、或hprna分子。在一些实施方案中,从重组dna分子转录的rna分子可以在重组植物的组织或流体内形成dsrna分子。这样的dsrna分子可以包含在多核苷酸的一部分中,所述多核苷酸与可侵染宿主植物的类型的昆虫害虫内的dna转录的相应多核苷酸相同。害虫内靶基因的表达被所述dsrna分子所阻抑,并且该害虫中靶基因表达的阻抑导致该转基因植物对该害虫有抗性。已经显示dsrna分子的调控作用可适用于害虫中表达的多种基因,包括例如负责细胞代谢或细胞转化的内源基因,包括管家基因;转录因子;蜕皮相关基因;和编码涉及细胞代谢或正常生长和发育的多肽的其他基因。为了从体内转基因或表达构建体转录,可以在一些实施方案中使用调节区(例如,启动子、增强子、沉默子、和多聚腺苷酸化信号)以转录一条或多条rna链。因此,在一些实施方案中,如上文提出的,在产生irna分子中使用的多核苷酸可以与在植物宿主细胞中有功能的一种或多种启动子元件可操作连接。启动子可以是通常驻留于宿主基因组中的内源启动子。在可操作连接的启动子元件控制下的本发明的多核苷酸可以进一步被其他有利地影响其转录和/或所得转录物的稳定性的元件所侧翼。这样的元件可以位于可操作连接启动子的上游、表达构建体3'端的下游、并且可以同时存在于启动子的上游和表达构建体3'端的下游。一些实施方案提供了用于降低由以植物为食的昆虫(例如鞘翅目和/或半翅目)害虫引起的对宿主植物(例如,玉米植物)的损害的方法,其中所述方法包括在宿主植物中提供表达本发明的至少一种核酸分子的转化植物细胞,其中所述核酸分子在被害虫摄取后发挥功能以抑制该害虫内靶多核苷酸的表达,该表达抑制导致该害虫的死亡和/或生长降低,由此降低由该害虫引起的对宿主植物的损害。在一些实施方案中,核酸分子包含dsrna分子。在这些和另外的实施方案中,核酸分子包含dsrna分子,所述dsrna分子各自包含超过一种与鞘翅目和/或半翅目害虫细胞中表达的核酸分子可特异性地杂交的多核苷酸。在一些实施方案中,核酸分子由一种多核苷酸组成,所述多核苷酸与昆虫害虫细胞中表达的核酸分子可特异性地杂交。在一些实施方案中,提供了用于提高玉米作物产量的方法,其中所述方法包括将本发明的至少一种核酸分子导入玉米植物中;栽培玉米植物以容许表达包含核酸的irna分子,其中包含所述核酸的irna分子的表达可抑制昆虫(例如鞘翅目和/或半翅目)害虫损害和/或生长,由此降低或消除由于害虫侵染所致的产量损失。在一些实施方案中,irna分子是dsrna分子。在这些和另外的实施方案中,核酸分子包含dsrna分子,所述dsrna分子各自包含超过一种与昆虫害虫细胞中表达的核酸分子特异性可杂交的多核苷酸。在一些实例中,所述核酸分子包含与鞘翅目和/或半翅目害虫细胞中表达的核酸分子可特异性地杂交的多核苷酸。在一些实施方案中,提供了用于调变昆虫(例如鞘翅目和/或半翅目)害虫中靶基因表达的方法,所述方法包括:用包含编码本发明的至少一种irna分子的多核苷酸的载体转化植物细胞,其中所述多核苷酸与启动子和转录终止元件可操作连接;在足以容许形成包含多个转化植物细胞的植物细胞培养物的条件下培养经转化的植物细胞;选择已经将核酸分子整合到其基因组中的转化植物细胞;对转化植物细胞筛选由整合的核酸分子编码的irna分子的表达;选择表达irna分子的转基因植物细胞;并且用选择的转基因植物细胞喂养昆虫害虫。也可以从表达由整合的核酸分子编码的irna分子的转化植物细胞再生植物。在一些实施方案中,irna分子是dsrna分子。在这些和另外的实施方案中,核酸分子包含dsrna分子,所述dsrna分子各自包含超过一种与昆虫害虫细胞中表达的核酸分子可特异性地杂交的多核苷酸。在一些实例中,核酸分子包含与鞘翅目和/或半翅目害虫细胞中表达的核酸分子可特异性地杂交的多核苷酸。可以将本发明的irna分子作为来自掺入植物细胞基因组中的重组基因的表达产物、或者掺入应用于种植前种子的包衣或种子处理中而掺入植物物种(例如,玉米)的种子之内。包含重组基因的植物细胞被视为转基因事件。本发明的实施方案中还包括用于将irna分子投送给昆虫(例如鞘翅目和/或半翅目)害虫的递送系统。例如,可以将本发明的irna分子直接导入害虫的细胞中。用于导入的方法可以包括将irna与来自害虫的宿主的植物组织直接混合,以及对宿主植物组织应用包含本发明的irna分子的组合物。例如,可将irna分子喷雾到植物表面上。或者,可以通过微生物表达irna分子,并且可以将微生物施加到植物表面上,或通过物理手段诸如注射导入根或茎中。如上文论述的,也可以对转基因植物进行遗传工程改造,使其以足以杀死已知侵染植物的昆虫害虫的量表达至少一种irna分子。通过化学或酶促合成产生的irna分子也可以以符合常见农业实践的方式配制,并且作为喷雾产品使用以控制昆虫害虫所致的植物损害。配制剂可以包含有效的叶覆盖需要的适当辅料(例如粘着剂(sticker)和湿润剂),以及保护irna分子(例如,dsrna分子)免于uv损伤的uv保护剂。这样的添加剂通常在生物杀虫剂产业中使用,并且是本领域技术人员熟知的。这样的应用可以与其他喷雾杀虫剂应用(基于生物学的或其他方面)组合以增强针对害虫的植物保护。本文中所讨论的所有参考文献,包括本文引用的出版物,专利和专利申请,均通过引用并入本文,只要与本公开的明确细节不冲突,如同单独地和具体地明示将每篇参考文献通过引用并入本文一样。本文所讨论的参考文献仅为其在本申请的申请日之前的公开内容而提供。本文中任何内容不应被解释为承认发明人无权凭借在先发明而先于这些公开。提供以下实施例以展示某些具体特征和/或方面。这些实施例不应被理解为将本公开限于本文中描述的具体特征或方面。实施例实施例1昆虫饵食生物测定样品制备和生物测定使用rnai试剂盒合成并纯化了若干dsrna分子(包括对应于copigammareg1(seqidno:3),copigammareg2(seqidno:4),copigammareg3(seqidno:5),copigammaver1(seqidno:75),copigammaver2(seqidno:76),copigammaver3(seqidno:77),和copigammaver4(seqidno:78)的那些)。将纯化的dsrna分子准备在te缓冲液中,并且所有生物测定均包含由该缓冲液组成的对照处理,用作wcr(玉米根萤叶甲)的死亡率或生长抑制的背景检查。使用nanodroptm8000分光光度计(thermoscientific,wilmington,de)测量生物测定缓冲液中dsrna分子的浓度。在使用饲喂人工昆虫饵食的新生昆虫幼虫的生物测定中测试样品的昆虫活性。wcr卵获自cropcharacteristics,inc(farmington,mn)。生物测定在特别为昆虫生物测定设计的128孔塑料托盘(c-dinternational,pitman,nj)中进行。每个孔含有约1.0ml设计用于鞘翅目昆虫生长的人工饵食。通过移液管将60μl等份的dsrna样品递送到每个孔的饵食的表面(40μl/cm2)。dsrna样品浓度作为孔中每平方厘米(ng/cm2)表面积(1.5cm2)的dsrna量来计算。将处理的托盘保持在通风橱中,直到饵食表面上的液体蒸发或吸收到饵食中。在破壳后的几个小时内,用湿润的骆驼毛刷拾取幼虫个体,并且将其放置在处理的饵食(每孔一个或两个幼虫)上。然后用透明塑料粘合片密封128孔塑料托盘上带虫的孔,并通气以允许气体交换。将生物测定盘在受控环境条件(28℃,~40%相对湿度,16:8(光照:黑暗))下保持9天,而后记录暴露于每个样品的昆虫总数、死亡的昆虫数、和存活昆虫的重量。计算每个处理的平均百分比死亡率和平均生长抑制。生长抑制(gi)计算如下:gi=[1-(twit/tnit)/(twibc/tnibc)],其中twit是处理中活昆虫的总重量;tnit是处理中的昆虫总数;twibc是背景检查中的活昆虫的总重量(缓冲液对照);和tnibc是背景检查中的昆虫总数(缓冲液对照)。统计学分析使用jmptm软件sas,cary,nc)进行。lc50(致死浓度)定义为50%的测试昆虫被杀死时的剂量。gi50(生长抑制)定义为试验昆虫的平均生长(例如活体重)为在背景检查样品中观察到的平均值的50%时的剂量。重复进行的生物测定证明,特定样品被摄取后导致了玉米根虫幼虫的令人惊讶且意想不到的死亡率和生长抑制。实施例2候选靶基因的鉴定选择多个wcr(玉米根萤叶甲)发育期进行汇集的转录组分析,以提供通过rnai转基因植物抗性技术控制的候选靶基因序列。在一个范例中,从约0.9gm的完整第一龄wcr幼虫(孵化后4到5天;保持在16℃)分离总rna,并使用下述基于苯酚/tri-的方法(molecularresearchcenter,cincinnati,oh)进行纯化:将幼虫于室温在具有10mltri的15ml匀浆器中均质化,直到获得均匀的悬浮液时为止。在室温温育5分钟后,将匀浆分配到1.5ml微量离心管中(每管1ml),添加200μl氯仿,并将混合物强力振摇15秒。在室温提取10分钟之后,通过在4℃以12,000xg离心分离各个相。将上层相(包含约0.6ml)小心转移到另一个无菌的1.5ml管中,并添加等体积的室温异丙醇。在室温温育5到10分钟之后,将混合物在12,000xg(4℃或25℃)离心8分钟。将上清液小心取出并弃去,并将rna离心沉淀通过用75%乙醇涡旋振荡洗涤两次,在每次洗涤后通过在7,500xg(4℃或25℃)离心5分钟回收。小心去除乙醇,让离心沉淀风干3至5分钟,然后溶解在无核酸酶的无菌水中。通过在260nm和280nm测量吸光度(a)测定rna浓度。从约0.9gm幼虫的典型提取产生超过1mg的总rna,其中a260/a280比为1.9。如此提取的rna在80℃贮存,直到进一步处理。通过使等份跑1%琼脂糖凝胶确定rna质量。在经高温灭菌的容器中,使用经depc(焦碳酸二乙酯)处理的水稀释的高温灭菌的10xtae缓冲液(tris乙酸盐edta;1x浓度为0.04mtris乙酸盐,1mmedta(乙二胺四乙酸钠盐,ph8.0)制成琼脂糖凝胶溶液。使用1xtae作为运行缓冲液。在使用前,用rnaseawaytm(invitrogeninc.,carlsbad,ca)清洁电泳槽和造孔梳。将2μlrna样品与8μlte缓冲液(10mmtrishclph7.0;1mmedta)和10μlrna样品缓冲液(目录号70606;emd4bioscience,gibbstown,nj)混合。将样品于70℃加热3分钟,冷却至室温,每孔上样5μl(含有1μg到2μgrna)。将市售的rna分子量标记物在分离的孔中同时运行以进行分子大小比较。在60伏特下跑胶2小时。由服务提供商(eurofinsmwgoperon,huntsville,al)利用随机引发从幼虫总rna制备标准化的cdna文库。在eurofinsmwgoperon通过gsflx454titaniumtm系列化学以1/2板的规模对标准化的幼虫cdna文库进行测序,其产生具有平均读段长度348bp的超过600,000个读段。将350,000个读段装配成超过50,000个重叠群。使用公众可访问的程序f0rmatdb(可在ncbi访问)将未装配的读段和重叠群两者都转换成可blast的数据库。类似地从其他wcr发育期收获的材料制备了总rna和标准化的cdna文库。合并代表各个发育期的cdna文库成员,构建了用于靶基因筛选的汇集的转录组文库。利用关于其他昆虫如果蝇属(drosophila)和拟谷盗属(tribolium)中的致死性rnai效应的信息选择了用于rnai靶向的候选基因。这些基因被假设为对于鞘翅目昆虫的存活和生长是必需的。对于选定的靶基因,在转录物组序列数据库中鉴定其同源物,如下文所述。通过pcr扩增靶基因的全长或部分序列来制备用于产生双链rna(dsrna)的模板。使用候选蛋白质编码多核苷酸针对含有未装配的叶甲属序列读段或已装配的重叠群的可blast数据库进行tblastn搜索。对于与叶甲属序列的显著命中(定义为:对于重叠群同源性,好于e-20;对于未装配的序列读段同源性,好于e-10),使用blastx针对ncbi非冗余数据库加以确认。此blastx搜索的结果确认,在tblastn搜索中鉴定的叶甲属同源物候选基因序列确实包含叶甲属基因,或者是针对叶甲属序列中存在的非叶甲属候选基因序列的最佳命中。在大多数情况下,被注释为编码蛋白质的拟谷盗属候选基因呈现出明确的与叶甲属转录物组序列中的一种或多种序列的序列同源性。在少数情况下可明显看出,某些基于与非叶甲属候选基因的同源性选择的叶甲属重叠群或未装配的序列读段有重叠,而且重叠群的装配未能连接起这些重叠。在这些情况下,使用sequenchertmv4.9(genecodescorporation,annarbor,mi)将序列装配成更长的重叠群。一种编码叶甲属copigamma(seqidno:1)的候选靶基因被鉴定为可能导致鞘翅目害虫死亡、生长抑制、发育抑制、或wcr中的生殖抑制的候选靶基因。与wcrcopigamma有同源性的基因copi指的是抑制顺式高尔基体膜上出芽过程的特异性外壳蛋白复合物(nickel,etal.2002.journalofcellscience115,3235-3240)。copi外被体复合物由七个亚单位组成。copi外被体γ是这些亚单位之一。该复合物的功能是将囊泡从高尔基复合体的顺式末端转移回到粗糙内质网——这些囊泡原来被合成之处。其它同样含有该结构域的玉米根萤叶甲蛋白可能有共同的结构和/或功能性质,因此编码这些蛋白质之一的基因可能包括了可能导致鞘翅目害虫死亡、生长抑制、发育抑制、或抑制wcr中的繁殖的候选靶基因。seqidno:1的序列是新的。该序列未提供在公共数据库中,并且没有在wo/2011/025860;美国专利申请号20070124836;美国专利申请号20090306189;美国专利申请号us20070050860;美国专利申请no.20100192265;或美国专利no.7,612,194中公开。叶甲属copigamma序列(seqidno:1)与来自一种线虫,旋毛虫(trichinellaspiralis)的一个序列(genbank登录号xm_003381124.1)的某个片段有一定相关。叶甲属copigamma氨基酸序列(seqidno:2)的最接近的同系物是具有genbank登录号xp_973414.1的金龟子(triboliumcasetanum)蛋白(90%相似;在同源区域上81%相同)。copigammadsrna转基因可与其他dsrna分子组合以提供冗余的rnai靶向和协同的rnai效应。表达靶向copigamma的dsrna的转基因玉米事件用于防止玉米根虫引起的噬根性损害(rootfeedingdamage)。copigammadsrna转基因提供了新的作用模式,可以用来与苏云金芽孢杆菌、产碱杆菌属、或假单胞菌属杀虫蛋白技术在昆虫抗性管理基因叠加(insectresistancemanagementgenepyramid)中组合,以减缓根虫群体对这两种根虫控制技术中的任一者发展抗性。利用该叶甲属候选基因,本文中称为copigamma,的序列的全长或部分克隆来生成用于dsrna合成的pcr扩增子。seqidno:1显示叶甲属copigamma的2840bpdna序列。seqidno:3显示copigammareg1的303bpdna序列。seqidno:4显示copigammareg2的332bpdna序列。seqidno:5显示copigammareg3的350bpdna序列。seqidno:75显示copigammaver1的108bpdna序列。seqidno:76显示copigammaver2的140bpdna序列。seqidno:77显示copigammaver3的110bpdna序列。seqidno:78显示copigammaver4的200bpdna序列。实施例3扩增靶基因以产生dsrna设计引物来通过pcr扩增每个靶基因的编码区部分。参见表1。在适当的情况下,将t7噬菌体启动子元件(ttaatacgactcactatagggaga;seqidno:6)引入扩增的有义或反义链的5'端。参见表1。从wcr提取总dna,并使用相反向的引物,使用第一链cdna作为模板进行pcr反应,引物的位置可扩增天然靶基因序列的全部或部分。还从包含黄色荧光蛋白(yfp)(seqidno:7;shagin等人,(2004)mol.biol.evol.21(5):841-50)编码区的dna克隆扩增了dsrna。表1.用于扩增示例性copigamma靶基因和yfp阴性对照基因的编码区部分的引物和引物对实施例4rnai构建体通过pcr制备模板和dsrna合成。图1中显示了一种提供特异性模板用于产生copigamma和yfpdsrna的策略。使用表1中的引物对、以及从分离自wcr第一龄幼虫的总rna制备的第一链cdna(作为pcr模板),制备了准备在copigammadsrna合成中使用的模板dna。对于每个选定的copigamma和yfp靶基因区,pcr扩增在扩增的有义和反义链的5'端导入了一个t7启动子元件(yfp区段扩增自yfp编码区的dna克隆)。将在有义链和反义链5’末端均具有t7启动子序列的pcr产物用作模板来产生dsrna。见图1。被特定引物对扩增的dsrna模板序列是:seqidno:3(copigammareg1)、seqidno:4(copigammareg2)、seqidno:5(copigammareg3)、seqidno:75(copigammaver1)、seqidno:76(copigammaver2),seqidno:77(copigammaver3),seqidno:78(copigammaver4)、和yfp(seqidno:7)。合成用于昆虫生物测定的双链rna,并使用rnai试剂盒遵循制造商的说明书(invitrogen)进行纯化。使用nanodroptm8000分光光度计(thermoscientific,wilmington,de)测量dsrna的浓度。植物转化载体的构建.使用化学合成的片段(dna2.0,menlopark,ca)的组合以及标准分子克隆方法,组装包含具有copigamma(seqidno:1)的区段的用于发夹形成的靶基因构建体的入门载体(pdab117215和pdab117216)。通过(在单一转录单位内)安排两个拷贝的copigamma靶基因序列的区段彼此相反取向来易化rna初级转录物形成分子内发夹,其中两个区段相隔一个st-ls1内含子序列(seqidno:17,vancanneyt等人,(1990)mol.gen.genet.220(2):245-50)。因此,初级mrna转录物含有被内含子序列分开的两个copigamma基因区段,其中两个区段互为大的反向重复序列。用一个拷贝玉米泛素1启动子(美国专利5,510,474)来驱动初级mrna发夹转录物的产生,并用包含玉米过氧化物酶5基因的(zmper53'utrv2;美国专利号6,699,984)的3'非翻译区的片段来终止表达发夹rna的基因的转录。入门载体pdab117215包含copigamma发夹v3rna构建体(seqidno:14),其含有copigamma(seqidno:1)的一个区段。入门载体pdab117216包含copigamma发夹v4-rna构建体(seqidno:15),其含有不同于pdab117215中所见的copigamma(seqidno:1)区段。使用上述入门载体pdab117215和pdab117216与典型的二元目标载体(pdab109805)进行标准重组反应,分别产生了用于土壤杆菌介导的玉米胚转化的copigamma发夹rna表达转化载体(分别为pdab117221和pdab117222)。通过典型的二元目标载体(pdab109805)与入门载体pdab101670的标准重组反应,构建包含表达yfp发夹dsrna的基因的阴性对照二元载体pdab110853。入门载体pdab101670包含处于玉米泛素1启动子(如上文)的表达控制之下的yfp发夹序列(seqidno:16)和包含来自玉米过氧化物酶5基因(如上文)的3'非翻译区的片段。二元目标载体pdab109805包含处于甘蔗杆状dna病毒(scbv)启动子(schenk等人,(1999)plantmolec.biol.39:1221-30)的调节之下的除草剂抗性基因(芳氧基链烷酸酯双加氧酶;aad-1v3)(美国专利7838733(b2)和wrightetal.(2010)proc.natl.acad.sci.u.s.a.107:20240-5)。一段人造5’utr序列,其由来自玉米条纹病毒(msv)外壳蛋白基因5’utr以及来自玉米醇脱氢酶1(adh1)基因的内含子6组成,定位在上述scbv启动子区段的3’末端和aad-1编码区的起始密码子之间。利用一个包含玉米脂肪酶基因的3'非翻译区(zmlip3'utr;美国专利号7,179,902)的片段来终止aad-1mrna的转录。通过典型的二元目标载体(pdab9989)与入门载体pdab100287的标准重组反应,构建了另一个包含表达yfp蛋白的基因的阴性对照二元载体pdab101556。二元目标载体pdab9989包含处于玉米泛素1启动子(如上文)的表达调节之下的除草剂抗性基因(芳氧基链烷酸酯双加氧酶;aad-1v3)(如上文)和一个包含来自玉米脂肪酶基因的3'非翻译区(zmlip3'utr;如上文)的片段。入门载体pdab100287包含处于玉米泛素1启动子(如上文)的表达控制之下的yfp编码区(seqidno:18)和一个包含来自玉米过氧化物酶5基因(如上文)的3'非翻译区的片段。seqidno:14呈现如pdab117221中所见的copigamma发夹v3形成序列。seqidno:14呈现如pdab117222中所见的copigamma发夹v4形成序列。实施例5候选靶基因的筛选在基于饵食的测定中,当将设计为抑制实施例2中鉴定的靶基因序列的合成dsrna给予至wcr时,引起了死亡和生长抑制。观察到copigammareg1,copigammareg2,copigammareg3,copigammaver1,copigammaver2,copigammaver3和copigammaver4在此测定中相比于其他被筛选的dsrna显示出大大增加的功效。重复的生物测定证明,对分别来源于copigammareg1,copigammareg2,copigammareg3,opigammaver1,copigammaver2,copigammaver3和copigammaver4的dsrna制备物的摄取导致西方玉米根虫幼虫的死亡和/或生长抑制。表2和表3显示了在wcr幼虫暴露于这些dsrna9天之后的基于饵食的饲喂生物测定的结果,以及从黄色荧光蛋白(yfp)编码区(seqidno:7)制备的dsrna的阴性对照样品获得的结果。表2.利用西方玉米根虫幼虫进食9天之后获得的copigammadsrna饵食进食测定的结果。anova分析发现在平均百分比死亡率和平均生长抑制(gi)上的显著差异。使用tukey--kramer检验分离平均值。*sem=均值标准误。刮弧中的字母指明统计水平。不是由相同字母连接的水平是显著不同的(p<0.05)。**te=trishcl(1mm)加edta(1mm)缓冲液,ph7.2。***yfp=黄色荧光蛋白表3.copigammadsrna对wcr幼虫的口服效力(ng/cm2)的总结。以前有人提出,某些叶甲属物种基因可以用于rnai介导的昆虫控制。参见美国专利公开号2007/0124836,其公开了906个序列,以及美国专利7,612,194,其公开了9,112个序列。然而确定了,许多被提示对rnai介导的昆虫控制基因对控制叶甲属无效。还确定了,与表明其他对于rnai介导的昆虫控制有用的基因相比,序列copigammareg1,copigammareg2,copigammareg3,opigammaver1,copigammaver2,copigammaver3和copigammaver4各自提供了令人惊奇和出乎意料的优越的叶甲属控制。例如,美国专利7,612,194提出,膜联蛋白、β血影蛋白2和mtrp-l4都在rnai介导的昆虫控制中有效。seqidno:19为膜联蛋白区1(reg1)的dna序列,seqidno:20为膜联蛋白区2(reg2)的dna序列。seqidno:21为β血影蛋白2区1(reg1)的dna序列,seqidno:22为β血影蛋白2区2(reg2)的dna序列。seqidno:23为mtrp-l4区1(reg1)的dna序列,并且seqidno:24为mtrp-l4区2(reg2)的dna序列。还使用了yfp序列(seqidno:7)产生用作阴性对照的dsrna。分别利用上述序列通过实施例3中的方法来产生dsrna。提供用于产生dsrna的特异性模板的策略显示在图2中。使用表4中的引物对和从分离自wcr第一龄幼虫的总rna制备的第一链cdna(作为pcr模板),制备了准备了在dsrna合成中使用的模板dna。(yfp从dna克隆扩增。)对于每个选择的靶基因区,进行两个单独的pcr扩增。第一个pcr扩增在扩增的有义链的5'端引入t7启动子元件。第二个反应在反义链的5'端引入t7启动子元件。然后将每个靶基因区的两个pcr扩增片段以大致相等的量混合,并将混合物用作dsrna产生的转录模板。参见图2。合成双链rna,并使用rnai试剂盒遵循制造商的说明书(invitrogen)进行纯化。使用nanodroptm8000分光光度计(thermoscientific,wilmington,de)测量dsrna的浓度。并通过上述相同的基于饵食的生物测定方法测试每一种dsrna。表4列出了用来产生yfp、膜联蛋白reg1、膜联蛋白reg2、β血影蛋白2reg1、β血影蛋白2reg2、mtrp-l4reg1、和mtrp-l4reg2dsrna分子的引物的序列。用于图2显示的方法的yfp引物序列也在表4中列出。表5呈现了wcr幼虫在暴露于这些dsrna分子9天之后的基于饵食的饲喂生物测定的结果。重复的生物测定证明,在以上西方玉米根虫幼虫中,摄取这些dsrna没有导致超过te缓冲液、水、或yfp蛋白等对照样品中所见的死亡或生长抑制。表4.用来扩增基因的编码区的多个部分的引物和引物对。表5.利用西方玉米根虫幼虫9天之后获得的饵食进食测定的结果。*te=trishcl(10mm)加edta(1mm)缓冲液,ph8。**yfp=黄色荧光蛋白实施例6包含杀虫发夹dsrna的转基因玉米组织的产生土壤杆菌介导的转化.在土壤杆菌介导的转化之后,制成了通过表达稳定地整合到植物基因组中的嵌合基因而产生一种或多种杀虫dsrna分子(例如,至少一种dsrna分子,其包含靶向包含copigamma,seqidno:1)的基因的dsrna分子)的转基因玉米细胞、组织、和植物。采用超二元或二元转化载体的玉米转化方法是本领域中已知的,正如例如美国专利号8,304,604中(在此通过提述并入其全部内容)所描述。通过转化的组织在含吡氟氯禾灵的培养基上生长的能力选择它们,并视情况适宜筛选其dsrna产生。可以将一部分这样的转化的组织培养物提供给新生玉米根虫幼虫以进行生物测定,基本上如实施例1中所描述。土壤杆菌培养启动.将包含上述(实施例4)二元转化载体pdab114515、pdab115770、pdab110853或pdab101556的土壤杆菌菌株dat13192细胞(wo2012/016222a2)的甘油储划线接种在含有适当抗生素的ab基本培养基平板(watson等人,(1975)j.bacteriol.123:255-264)上,并在20℃培养3天。然后将培养物划线接种在含有相同抗生素的yep平板(g/l:酵母提取物10;蛋白胨10;nacl,5)上,并在20℃温育1天。土壤杆菌培养.在实验当天,以适合于实验中的构建体数目的体积制备接种培养基和乙酰丁香酮的储液,并将其移液到无菌的一次性250ml烧瓶中。接种培养基(frame等人,(2011)genetictransformationusingmaizeimmaturezygoticembryos.收录于plantembryoculturemethodsandprotocols:methodsinmolecularbiology.t.a.thorpeande.c.yeung,(eds),springerscienceandbusinessmedia,llc.pp327-341)含有:2.2gm/lms盐;1xisu改良的ms维生素(frame等人,同上);68.4gm/l蔗糖;36gm/l葡萄糖;115mg/l脯氨酸;和100mg/l肌醇;ph为5.4)。将乙酰丁香酮以200μm的终浓度(从100%二甲亚砜中的1m储液)添加到含有接种培养基的烧瓶中,并充分混合溶液。对于每种构建体,来自yep平板的1或2个满接种环的土壤杆菌悬浮在一次性50ml无菌离心管内15ml的接种培养基/乙酰丁香酮储液中,在分光光度计中在550nm(od550)处测量溶液的光密度。然后使用另外的接种培养基/乙酰丁香酮混合物将悬液稀释到0.3到0.4的od550。然后将土壤杆菌悬液的管水平放置在设置为在室温下约75rpm的平台摇床上,并在进行胚切开的同时振摇1到4小时。玉米棒消毒和胚分离.未成熟玉米胚从玉米近交系b104(hallauer等人,(1997)cropscience37:1405-1406)植物获得,所述植物在温室中培养,并进行自花授粉或近亲授粉以产生玉米棒。在授粉后大约10到12天收获玉米棒。在实验当天,将玉米棒脱壳,并通过浸没在20%市售漂白剂(ultragermicidalbleach,6.15%次氯酸钠;加两滴吐温tm20)的溶液中并振摇20到30分钟进行表面消毒,随后在层流罩内在无菌去离子水中冲洗三次。从每个玉米棒无菌切下未成熟合子胚(1.8到2.2mm长)并随机分配到微量离心管中,每根微量离心管含有2.0ml的液体接种培养基中的适当土壤杆菌细胞的悬液,其中含200μm乙酰丁香酮,并添加了2μl的10%s233表面活性剂(evonikindustries;essen,germany)。对于一套给定的实验,每次转化均使用来自汇集的玉米棒的胚。土壤杆菌共培养.在分离之后,将胚在摇动平台上放置5分钟。然后将管的内容物倾倒到共培养基的平板上,所述培养基含有4.33gm/lms盐;1xisu改良的ms维生素;30gm/l蔗糖;700mg/ll-脯氨酸;在koh中的3.3mg/l的麦草畏(3,6-二氯-o-茴香酸或3,6-二氯-2-甲氧基苯甲酸);100mg/l肌醇(myo-inositol);100mg/l酪蛋白酶水解物;15mg/lagno3;200μm溶于dmso中的乙酰丁香酮;和3gm/lgelzantm,ph为5.8。用无菌一次性移液管移除液态土壤杆菌悬液。然后借助于显微镜,使用无菌镊子使胚定向为盾片面向朝上。盖上平板,用3mtmmicroporetm医用胶带密封,并放置在具有大约60μmolm-2s-1的光合有效辐射(par)的连续光照的25℃培养箱中。愈伤组织选择和转基因事件的再生.在共培养期之后,将胚转移到静息培养基上,静息培养基的组成为:4.33gm/lms盐;1xisu改良的ms维生素;30gm/l蔗糖;700mg/ll-脯氨酸;溶于koh中的3.3mg/l的麦草畏;100mg/l肌醇;100mg/l酪蛋白酶水解物;15mg/lagno3;0.5g/lmes(2-(n-吗啉代)乙磺酸一水合物;phytotechnologieslabr.;lenexa,ks);250mg/l羧苄青霉素;和2.3gm/lgelzantm;ph为5.8。将不超过36个胚移到每个平板上。将平板放置在透明的塑料盒中,在27℃下在大约50μmolm-2s-1par的连续光照下温育7到10天。然后将愈伤的胚转移(<18个/板)到选择培养基i上,所述培养基由具有100nm吡氟氯禾灵酸(r-haloxyfopacid)(0.0362mg/l;用于选择包含aad-1基因的愈伤组织)的静息培养基(上文)构成。将平板返回到透明盒中,在27℃下在大约50μmolm-2s-1par的连续光照下温育7天。然后将愈伤的胚转移(<12个/板)到选择培养基ii上,所述培养基由具有500nm吡氟氯禾灵酸(0.181mg/l)的静息培养基组成。将平板返回到透明盒中,在27℃下在大约50μmolm-2s-1par的连续光照下温育14天。这一选择步骤允许转基因愈伤组织进一步增殖和分化。将增殖中的胚性愈伤组织转移到(<9个/板)预再生培养基上。预再生培养基含有4.33g/lms盐;1xisu改良的ms维生素;45gm/l蔗糖;350mg/ll-脯氨酸;100mg/l肌醇;50mg/l酪蛋白酶水解物;1.0mg/lagno3;0.25gm/lmes;溶于naoh中的0.5mg/l萘乙酸;溶于乙醇中的2.5mg/l脱落酸;1mg/l6-苄氨基嘌呤;250mg/l羧苄青霉素;2.5gm/lgelzantm;和0.181mg/l吡氟氯禾灵酸;ph为5.8。将平板保存在透明盒中,在27℃下在大约50μmolm-2s-1par的连续光照下温育7天。然后将再生中的愈伤组织转移到(<6个/板)phytatraystm(sigma-aldrich)中的再生培养基上,在28℃以每天16小时光照/8小时黑暗(以大约160μmolm-2s-1par)温育14天,直到发出芽和根为止。再生培养基含有4.33gm/lms盐;1xisu改良的ms维生素;60gm/l蔗糖;100mg/l肌醇;125mg/l羧苄青霉素;3gm/lgellantm胶;和0.181mg/l吡氟氯禾灵酸;ph为5.8。然后分离具有主根的小芽,并不经选择直接转移到伸长培养基上。伸长培养基含有4.33gm/lms盐;1xisu改良的ms维生素;30gm/l蔗糖;和3.5gm/lgelritetm:ph为5.8。通过它们在含有吡氟氯禾灵的培养基上生长的能力而选择出转化植物芽,将所述芽从phytatraystm移栽到填充有生长培养基(promixbx;premiertechhorticulture)的小盆中,小盆用杯子或humi-domes(arcoplastics)覆盖,然后在conviron生长室中炼苗(27℃昼/24℃夜,16小时光周期,50-70%rh,200μmolm-2s-1par)。在一些情况下,分析推定的转基因小植物的转基因相对拷贝数,这使用设计用于检测整合到玉米基因组中的aad1除草剂耐受基因的引物通过定量实时pcr测定来完成。此外,利用rnaqpcr测定来检测在推定的转化体表达的dsrna中st-ls1内含子序列的存在。然后将选定的转化小植物移动到温室中,以便进一步生长和测试。转移t0植物和在温室中定植以进行生物测定和产生种子.当植物达到v3-v4期时,将其移栽到iecustomblend(profile/metromix160)土壤混合物中,在温室中(光曝露类型:光或同化作用;高光限值:1200par;16-小时昼长;27℃昼/24℃夜)培植至开花。将要用于昆虫生物测定的植物从小盆移栽到tinustm350-4(spencer-lemaireindustries,acheson,alberta,canada)(每每事件一个植物)。在移栽到约四天后,侵染植物以进行生物测定。通过对t0转基因植物的穗丝授粉,其中花粉从非转基因良种近交系b104或其他适当的花粉供体采集而来,并种植所得的种子而获得t1代植物。在可能时进行互交。实施例7转基因玉米组织的分子分析对在评估噬根损害的同日从温室培植的植物采集的叶和根的样品进行了玉米组织的分子分析(例如,rnaqpcr)。用对per53'utr的rnaqpcr测定结果来验证发夹转基因的表达。(在非转化的玉米植物中预期会检出低水平的per53'utr,因为在玉米组织中通常存在内源per5基因表达)。用针对表达的rna中的st-ls1内含子序列(为形成dsrna发夹分子所必需)的rnaqpcr测定结果来验证发夹转录物的存在。测量了相对于内源玉米基因的rna水平的转基因rna表达水平。通过dnaqpcr分析检测基因组dna中aad1编码区的一部分,用来估计转基因插入拷贝数。从培植在环境室中的植物采集样品用于这些分析。将结果与旨在检测单拷贝天然基因的一部分的测定法的dnaqpcr结果进行比较,并将简单事件(具有一个或两个copigamma转基因拷贝)推进到温室中的进一步研究。另外,用旨在检测大观霉素抗性基因(specr;包含在t-dna外部的二元载体质粒上)的一部分的qpcr测定法来确定转基因植物是否含有无关的整合的质粒骨架序列。发夹rna转录物表达水平:per53'utrqpcr.通过per53'utr序列的实时定量pcr(qpcr)来分析愈伤组织细胞事件或转基因植物,以确定全长发夹转录物的相对表达水平,与内部玉米基因(seqidno:53;genbank登录号bt069734)的转录物水平比较,所述内部玉米基因编码tip41-样蛋白(即,genbank登录号at4g34270的玉米同源物;具有74%同一性的tblastx得分)。使用rnaeasytm96试剂盒(qiagen,valencia,ca)分离rna。洗脱后,根据试剂盒建议的方案进行总rna的dnase1处理。然后在nanodroptm8000分光光度计(thermoscientific)上将rna定量,并将浓度归一化为25ng/μl。大体上按照制造商推荐的方案,使用高容量cdna合成试剂盒(invitrogen)制备第一链cdna,反应体积10μl,含有5μl变性rna。略微改变该方案以包括将10μl的100μmt20vn寡核苷酸(idt)(seqidno:54;ttttttttttttttttttttvn,其中v是a、c、或g,n是a、c、g、或t/u)添加到随机引物储备预混物的1ml管中,以制备随机引物和寡dt混合的工作储备液。在cdna合成后,以无核酸酶的水将样品以1:3稀释,并储存在-20℃直到用于测定时为止。在lightcyclertm480(rochediagnostics,indianapolis,in)上以10μl的反应体积分别进行per53'utr和tip41样转录物的实时pcr测定。对于per53'utr测定,利用引物p5u76s(f)(seqidno:55)和p5u76a(r)(seqidno:56),以及rocheuniversalprobetm(upl76;目录号4889960001;用fam标记)运行反应。对于tip41-样参考基因测定,使用引物tipmxf(seqidno:57)和tipmxr(seqidno:58)、以及标记有hex(六氯荧光素)的探针hxtip(seqidno:59)。所有测定都包括没有模板的阴性对照(仅有预混物)。为制备标准曲线,在源板中还包括空白(源孔中加水),以检查样品交叉污染。在表6中列出了引物和探针序列。用于检测各种转录物的反应成分配方在表7中公开,pcr反应条件概述于表8中。在465nm处激发fam(6-羧基荧光素亚磷酰胺)荧光部分,并测量在510nm处的荧光;hex(六氯荧光素)荧光部分的对应值为533nm和580nm。表6.被用于转基因玉米中转录物水平的分子分析的多多核苷酸。*tip41样蛋白**nav序列无法从供应商获得表7.用于转录物检测的pcr反应配方。表8.用于rnaqpcr的热循环仪条件。使用lightcyclertm软件v1.5,根据供应商的推荐利用二阶导数最大算法计算cq值,通过相对定量分析数据。对于表达分析,使用δδct方法(即,2-(cqtarget–cqref))计算表达值,所述方法依赖于比较两个靶之间的cq值的差异,其中假定对于优化的pcr反应条件,每个循环产物都加倍,基值选择为2。发夹转录物大小和完整性:northern印迹测定.在一些情况下,利用northern印迹(rna印迹)分析来确定在表达copigamma发夹dsrna的转基因植物中copigamma发夹rna的分子大小,从而获得转基因植物的另外的分子表征。所有的材料和设备在使用之前用rnazap(ambion/invitrogen)处理。将组织样品(100mg到500mg)采集到2mlsafelockeppendorf管中,用配备三个钨珠的kleckotm组织粉碎器(garciamanufacturing,visalia,ca)在1mltrizol(invitrogen)中破碎5分钟,然后在室温(rt)下温育10分钟。任选地,将样品在4℃在11,000rpm下离心10分钟,并将上清液转移到新鲜的2mlsafelockeppendorf管中。在将200μl的氯仿添加到匀浆中之后,通过翻转该管2到5分钟进行混合,然后在rt下温育10分钟,并在4℃在12,000xg下离心15分钟。将上层相转移到无菌的1.5mleppendorf管中,添加600μl的100%异丙醇,在rt温育10分钟到2小时之后,然后在4°到25℃在12,000xg下离心10分钟。弃去上清液,用1ml的70%乙醇将rna离心沉淀物洗涤两次,在两次洗涤之间,在4°到25℃在7,500xg下离心10分钟。弃去乙醇,将离心沉淀物快速风干3到5分钟,然后重悬在50μl无核酸酶的水中。使用(thermo-fisher)对总rna定量,并将样品归一化为5μg/10μl。然后向每个样品中加入10μl的乙二醛(ambion/invitrogen)。将5到14ng的digrna标准标志物预混物(rocheappliedscience,indianapolis,in)分配并添加到等体积的乙二醛中。在50℃使样品和标志物rna变性45分钟,并保存在冰上,直到上样到在northernmaxtm10x乙二醛运行缓冲液(ambion/invitrogen)中的1.25%seakemgoldtm琼脂糖(lonza,allendale,nj)凝胶上为止,通过在65伏特/30ma下电泳2小时15分钟分离rna。电泳之后,在2xssc冲洗凝胶5分钟,在凝胶doc工作站(biorad,hercules,ca)上成像,然后在rt过夜使rna被动转移到尼龙膜(millipore)上,其中使用10xssc作为转移缓冲液(由3m氯化钠和300mm柠檬酸三钠组成的20xssc,ph7.0)。转膜后,在2xssc中冲洗膜5分钟,通过uv使rna与该膜交联(agilent/stratagene),并使该膜在室温下干燥2天。使膜在缓冲液(ambion/invitrogen)中预杂交1到2小时。探针由含有感兴趣序列(例如,seqidno:14或seqidno:15的反义序列部分,视情况适宜)的pcr扩增产物组成,其通过rocheappliedsciencedig程序用地高辛配基标记。在杂交管中推荐的缓冲液中60℃的温度下杂交过夜。杂交后,对印迹进行dig洗涤,包装,暴露于胶片1到30分钟,然后将胶片显影,所有这些都通过dig试剂盒的供应商推荐的方法来进行。转基因拷贝数确定.将大约等同于2个叶冲孔块的玉米叶片收集在96孔的收集平板(qiagen)中。用配备一个不锈钢珠的kleckotm组织粉碎器(garciamanufacturing,visalia,ca)在biosprint96ap1溶解缓冲液(与biosprint96plantkit一起提供;qiagen)中进行组织破碎。组织浸软之后,使用biosprint96plantkit和biosprint96提取机器人以高通量形式分离基因组dna(gdna)。在设立qpcr反应之前,以2:3的dna:水稀释基因组dna。qpcr分析.通过使用480系统的实时pcr,借助水解探针测定来进行转基因检测。使用probedesignsoftware2.0设计了要在水解探针测定中用于检测st-ls1内含子序列(seqidno:17)、或检测specr基因(即,负载在二元载体质粒上的大观霉素抗性基因;seqidno:71;在表9中的spc1寡核苷酸)的一部分的寡核苷酸。此外,使用primerexpress软件(appliedbiosystems)设计了要在水解探针测定中用于检测aad-1耐除草剂基因(seqidno:65;在表9中的gaad1寡核苷酸)的区段的寡核苷酸。表9显示了引物和探针序列。测定用内源玉米染色体基因(转化酶(seqidno:62;genbank登录号u16123;在本文中称为ivr1)的试剂多重化,所述基因用作内部参考序列以确保在每个测定中都存在gdna。为了扩增,制备了在10μl体积的多重反应物中的1x终浓度的480probes母液混合物(rocheappliedscience),其含有每种引物各0.4μm,以及每种探针各0.2μm(表10)。如表11中概述进行两步扩增反应。fam-和hex-标记探针的荧光团活化和发射如上文所述;cy5偶联物在650nm处最大激发,并且在670nm处发最大荧光。使用拟合点算法(软件发行版本1.5)和相对定量模块(基于δδct方法),根据实时pcr数据确定cp得分(荧光信号与背景阈值杂交的点)。如前所述处理数据(上文;rnaqpcr)。表9.用于基因拷贝数确定和二元载体质粒骨架检测的引物和探针(具有荧光偶联物)的序列。cy5=花青-5表10.用于基因拷贝数分析和质粒骨架检测的反应成分成分量(μl)储液终浓度2x缓冲液5.02x1x适当的正向引物0.410μm0.4适当的反向引物0.410μm0.4适当的探针0.45μm0.2ivr1-正向引物0.410μm0.4ivr1-反向引物0.410μm0.4ivr1-探针0.45μm0.2h2o0.6na*nagdna2.0nd**nd总计10.0*na=不适用**nd=未确定表11.用于dnaqpcr的热循环仪条件实施例8转基因玉米的生物测定体外昆虫生物测定.通过生物测定方法来证实在植物细胞中产生的本主题发明的dsrna的生物活性。参见,例如,baum等人,(2007)nat.biotechnol.25(11):1322-1326。人们能够例如通过在受控的饲喂环境下给靶昆虫喂食来源于产生杀虫dsrna的植物的各种植物组织或组织片来证实效力。或者,从来源于产生杀虫dsrna的植物的各种植物组织制备提取物,并将提取的核酸分配在用于如以前在本文中描述的生物测定的人工饵食上面。将这样的饲喂测定的结果与类似方式进行的采用来自不产生杀虫dsrna的宿主植物的适当对照组织、或其他对照样品的生物测定进行比较。关于转基因玉米事件的昆虫生物测定.选择从经洗涤的卵孵化的两条西方玉米根虫幼虫(1到3日龄)并将其置于生物测定托盘的每个孔中。然后将这些孔用“拉-揭”(pulln’peel)护盖(bio-cv-16,bio-serv)覆盖,并置于具有18小时/6小时光照/黑暗周期的28℃培养箱中。在初始侵染9日之后,评估幼虫死亡率,其按照死亡昆虫占的每次处理中昆虫总数的百分比来计算。将昆虫样品在-20℃冷冻两天,然后汇集来自每个处理的昆虫幼虫并称重。生长抑制百分比按照实验处理的平均重量除以两个对照孔处理的平均重量的均值来计算。数据表示为(阴性对照的)生长抑制百分比。将超过对照平均重量的平均重量归一化为零。温室中的昆虫生物测定.从cropcharacteristics(farmington,mn)收到带有西方玉米根虫(wcr,玉米根萤叶甲)卵的土壤。在28℃温育wcr卵10到11天。洗掉卵上的土壤,将卵置于0.15%琼脂溶液中,浓度调节至每0.25ml等份大约75个到100个卵。将一份卵悬浮液加入培养皿中以设置孵化平板,用于监测孵化速率。用150到200个wcr卵侵染中生长的玉米植物周围的土壤。允许昆虫摄食2周,在这段时间之后,对每个植物给出“根评级”。利用一个节损伤量表进行分级,大体上如oleson等人,(2005,j.econ.entomol.98:1-8)所述。将通过了这个生物测定的植物移栽到5加仑的盆中供产生种子。用杀虫剂处理移植物以防止进一步根虫损害和昆虫释放到温室中。将植物人工授粉以产生种子。保存由这些植物产生的种子以评估植物的t1和后续世代。温室生物测定包括两种阴性对照植物。转基因阴性对照植物通过用包含设计为产生黄色荧光蛋白(yfp)的基因或yfp发夹dsrna的载体转化而生成(参见实施例4)。非转化的阴性对照植物从品系7sh382或b104的种子培植而来。在两个不同的日期进行生物测定,其中每一组植物材料中均包括阴性对照。表12显示了copigamma发夹植物的分子分析和生物测定的结果汇总。考察总结在表12中的生物测定结果揭示了一项令人惊奇和出乎意料的观察结果,即,大多数包含表达含有seqidno:1区段(例如,如seqidno:14和seqidno:15所例示)的copigamma发夹dsrna的构建体的转基因玉米植物受到保护而免于通过喂养西方玉米根虫幼虫招致的根损害。37个被分级的事件中的二十二个具有0.5或更低的根评级。表13显示了阴性对照植物的分子分析和生物测定的合并结果。大多数植物不具有针对wcr幼虫喂养的保护,虽然34株被评级的植物中有5株具有0.75或更低的根评级。有时观察到阴性对照植物组中存在一些具有根评级分数低的植物,这反映了在温室环境中进行这种类型生物测定的易变性和困难性。表12.表达copigamma发夹v3和copigamma发夹v4的玉米植物的温室生物测定和分子分析结果。*rtl=针对tip4-样基因转录物水平测量的相对转录物水平。**ng=由于植物尺寸小而未评级。表13.包含转基因的阴性对照植物和非转化玉米植物的温室生物测定和分子分析结果。*rtl=针对tip4样基因转录物水平测量的相对转录物水平。**ng=由于植物尺寸小未予分级***nd=未进行。实施例9包含鞘翅目害虫序列的转基因玉米如实施例6中所述生成10个到20个转基因t0玉米植物。获得另外的10--20个表达针对rnai构建体的发夹dsrna的t1玉米独立品系用于玉米根虫攻击。发夹dsrna可按照seqidno:14、seqidno:15中所列出而衍生,或者进一步包含seqidno:1。更多的发夹dsrna衍生自例如鞘翅目害虫序列,例如像caf1-180(美国专利申请公开号2012/0174258)、vatpasec(美国专利申请公开号2012/0174259)、rho1(美国专利申请公开号2012/0174260)、vatpaseh(美国专利申请公开号2012/0198586)、ppi-87b(美国专利申请公开号2013/0091600)、rpa70(美国专利申请公开号2013/0091601)、或rps6(美国专利申请公开号2013/0097730)。这些通过rt-pcr或其他分子分析方法得以证实。来自选定的独立t1系的总rna制备物在某些情况下用于rt-pcr,其中引物被设计为在每个rnai构建体中的发夹表达盒的st-ls1内含子中结合。另外,用于rnai构建体中每个靶基因的特异性引物在某些情况下用于扩增在植物内sirna产生所需要的预加工mrna,并用于确认所述预加工mrna的产生。对于每个靶基因的期望条带的扩增可确认每个转基因玉米植物中发夹rna的表达。随后在某些情况下利用rna印迹杂交在独立转基因系中确认靶基因的dsrna发夹加工成sirna。而且,与靶基因具有80%以上序列同一性、具有错配序列的rnai分子对玉米根虫的影响类似于与靶基因具有100%序列同一性的rnai分子。错配序列与天然序列的配对在同一个rnai构建体中形成发夹dsrna,由此产生出能够影响摄食中的鞘翅目害虫的生长、发育和生存力的植物加工的sirna。对应于靶基因的dsrna、sirna、或mirna的植物体内递送以及被鞘翅目害虫进食后的摄取,导致鞘翅目害虫中的靶基因由于rna介导的基因沉默而下调。当靶基因在发育的一个或多个阶段发挥重要作用的时候,鞘翅目害虫的生长、发育和生殖受到影响,而且在wcr、ncr、scr、mcr、玉米根萤叶甲、d.u.tenella、和d.u.undecimpunctatamannerheim的至少一者的情况下,会导致鞘翅目害虫无法成功侵染、摄食、发育、和/或生殖,或导致其死亡。然后通过靶基因的选择和rnai的成功应用来控制鞘翅目害虫。转基因rnai品系和非转化玉米的表型比较.选用于创建发夹dsrna的靶鞘翅目害虫基因或序列与任何已知的植物基因bn序列都没有相似性。因此不期望靶向这些鞘翅目害虫基因或序列的构建体的(系统性)rnai的产生或活化对转基因植物会有任何有害影响。然而,将转基因系的发育和形态特征与非转化植物、以及用没有发夹表达基因的“无效”(null)载体转化的那些转基因系进行比较。比较了植物根、芽、叶和生殖特征。转基因植物和非转化植物在根长和生长模式上没有可观察到的差异。植物地上部特征,诸如高度、叶数和大小、开花时间、花大小和外观之类,是相似的。总的来说,在体外以及在温室土壤中培养时,在转基因系和没有表达靶irna分子的那些品系之间没有可观察的形态差异。实施例10包含鞘翅目害虫序列和另外的rnai构建体的转基因玉米转基因玉米植物在其基因组中包含异源编码多核苷酸,所述异源编码多核苷酸被转录成靶向鞘翅目害虫之外的生物的irna分子,对这样的转基因植物通过土壤杆菌或whiskerstm方法(参见petolino和arnold(2009)methodsmol.biol.526:59-67)进行二次转化,以产生一种或多种杀虫dsrna分子(例如,至少一种dsrna分子,包括靶向包含seqidno:1的基因的dsrna分子)。基本上如实施例4中所述制备植物转化质粒载体,经由土壤杆菌或whiskerstm-介导的转化方法递送到获自转基因hiii或b104玉米植物的玉米悬浮细胞或未成熟玉米胚中,所述玉米植物在其基因组中包含异源编码多核苷酸,所述异源编码多核苷酸被转录成靶向鞘翅目害虫之外的生物的irna分子。实施例11包含rnai构建体和另外的鞘翅目害虫控制序列的转基因玉米转基因玉米植物在其基因组中包含被转录成靶向鞘翅目害虫生物的irna分子的异源编码多核苷酸(例如,至少一种dsrna分子,包括靶向包含seqidno:1的基因的dsrna分子),对于这样的转基因玉米植物,通过土壤杆菌或whiskerstm方法(参见petolino和arnold(2009)methodsmol.biol.526:59-67)进行二次转化以产生一种或多种杀虫蛋白分子,例如,cry1b、cry1i、cry2a、cry3、cry7a、cry8、cry9d、cry14、cry18、cry22、cry23、cry34、cry35、cry36、cry37、cry43、cry55、cyt1a和cyt2c杀虫蛋白。基本上如实施例4中所述制备植物转化质粒载体,经由土壤杆菌或whiskerstm-介导的转化方法递送到获自转基因b104玉米植物的玉米悬浮细胞或未成熟玉米胚中,所述玉米植物在其基因组中包含被转录成靶向鞘翅目害虫生物的irna分子的异源编码多核苷酸。获得产生用于控制鞘翅目害虫的irna分子和杀虫蛋白的双重转化植物。实施例12新热带棕椿象(英雄美洲蝽)在copigammarnai注射后的死亡新热带棕椿象(bsb;英雄美洲蝽)群落.将bsb饲养在27℃、相对湿度65%、16:8小时的光照:黑暗循环的培养箱中。将经2-3天收集的1克卵接种在5l容器中,容器底部有滤纸盘,并用#18目筛盖住容器以便通风。每个饲养容器产生约300-400只成虫bsb。在所有阶段,昆虫均每周三次喂养新鲜青豆,每周更换一小袋含有向日葵种子,大豆和花生(3:1:1重量比)的种子混合物。水在带有棉塞作为捻子的小瓶中补充。经最初两周后,每周一次将昆虫转移到新的容器上。bsb人工饵食.用如下制备的bsb人工饵食(在制备两周内使用)。在magic混合器中将冻干绿豆混合成细粉,同时在另一台magic混合器中将生(有机)花生混合。在大的magic混合器中合并混合的干成分(重量百分比:绿豆35%;花生35%;蔗糖5%;维生素复合物(例如用于昆虫的vanderzant维生素混合物,sigma-aldrich,目录号v1007)0.9%),加盖并充分振摇以将这些成分混合。然后将混合的干成分添加到混合碗。在另一容器中,将水和苯菌灵抗真菌剂(50ppm;25μl20,000ppm溶液/50ml饵食液)充分混合,然后添加到干成分混合物中。人工混合所有成分,直到溶液完全混合时为止。将饵食成形为期望的大小,松散地包装在铝箔中,在60℃加热4小时,然后冷却,在4℃储存。bsb转录组组装.选择六个bsb发育期用于mrna文库制备。从冷冻在-70℃的昆虫提取总rna,并将其在-24设备(mpbiomedicals)上的lysingmatrixa2ml管(mpbiomedicals,santaana,ca)中的10倍体积的溶解/结合缓冲液中均质化。使用mirvanatmmirna分离试剂盒(ambion;invitrogen)根据制造商的方案提取总mrna。使用hiseqtm系统(sandiego,ca)的rna测序提供了用于在rnai昆虫控制技术中使用的候选靶基因序列。hiseqtm为六个样品生成了总共约3.78亿个读段。使用trinity汇编程序软件(grabherr等人,(2011)naturebiotech.29:644-652)针对每个样品将读段逐一汇编。合并汇编的转录物以生成汇集的转录组。这个bsb汇集的转录组含有378,457个序列。bsbcopigamma直系同源物鉴定.使用果蝇属copigamma蛋白序列γcop-pa、γcop-pb及γcop-pc:genbank登录号np_524608、np_733432和np_001163784,分别作为查询进行bsb汇集的转录组的tblastn搜索。bsbcopigamma(seqidno:87)被鉴定为英雄美洲蝽候选靶基因产物,其预测的肽序列为seqidno:88。模板制备和dsrna合成.使用试剂(lifetechnologies),从提取自单一年轻成虫(约90mg)的总bsbrna制备cdna。使用微量管研磨棒(pelletpestle)(fisherbrand目录号12-141-363)和研磨棒混合器(pestlemotormixer)(cole-parmer,vernonhills,il),在装有200μl的的1.5ml微量离心管中将昆虫均质化。均质化之后,再添加800μl的使匀浆涡旋,然后在室温下温育五分钟。通过离心去除细胞碎片,上清液转移到新的管中。按照制造商推荐的提取方案,将rna离心沉淀在室温下干燥,并重悬于来自gfxpcrdnaandgelextractionkit(illustratm;gehealthcarelifesciences)的200μltris缓冲液中,使用洗脱缓冲液类型4(即10mmtris-hcl,ph8.0)。使用nanodroptm8000分光光度计(thermoscientific,wilmington,de)测定rna浓度。cdna扩增.使用针对rt-pcr的superscriptiiifirst-strandsynthesissystemtm(invitrogen),根据供应商的推荐方案,从5μg的bsb总rna模板和寡dt引物反向转录而成cdna。用无核酸酶的水将转录反应的终体积调节为100μl。使用引物bsb_γcop-2-for(seqidno:90)和bsb_γcop-2-rev(seqidno:91)来扩增bsb_copigamma区域2,又称bsb_copigamma-2模板。用1μl的cdna(上文)进行触地(touch-down)pcr(退火温度从60℃降至50℃,以10℃/循环降低)来扩增该dna模板。在35个循环的pcr中产生了包含bsb_copigamma-2(seqidno:89)的495bp区段的片段。上述程序也被用于使用yfpv2-f(seqidno:93)和yfpv2-r(seqidno:94)引物扩增301bp阴性对照模板yfpv2(seqidno:92)。bsb_copigamma和yfpv2引物在其5'端含有t7噬菌体启动子元件(seqidno:6),因此使得yfpv2和bsb_copigammadna片段能够用于dsrna转录。dsrna合成.利用megascripttmrnai试剂盒(ambion),根据制造商的说明,使用2μl的pcr产物(上文)作为模板合成dsrna。(参见图1)。在nanodroptm8000分光光度计上将dsrna定量并在无核酸酶的0.1xte缓冲液(1mmtrishcl,0.1mmedta,ph7.4)中稀释到500ng/μl。dsrna注射到bsb血腔中.在27℃培养箱中,在65%相对湿度和16:8小时的光:暗光周期下,用绿豆和种子饵食喂养作为群落的bsb。用小刷子轻轻操作第二龄若虫(每只重1到1.5mg)以防损伤,并将它们置于冰上培养皿中,以使昆虫预冷和固定。给每只昆虫注射55.2nl的500ng/μldsrna溶液(即,27.6ngdsrna;18.4到27.6μg/g体重的剂量)。注射使用配备有从drummond3.5英寸#3-000=203-g/x玻璃毛细管拉制而成的注射针的nanojecttmii注入器(drummondscientific,broomhall,pa)。将针尖打破,用轻矿油装填毛细管,然后充填2到3μl的dsrna。将dsrna注射到若虫的腹部(每次试验每dsrna注射10个昆虫),在不同的三天重复试验。经过注射的昆虫(每孔5个)被转移到含有一团人工bsb饵食并覆盖有pull-n-peeltm盖片(bio-cv-4;bio-serv)的32孔托盘(bio-rt-32rearingtray;bio-serv,frenchtown,nj)中。借助于带有棉花芯的装有1.25ml水的1.5ml微量离心管提供水分。将这些托盘在26.5℃、60%湿度和16:8小时光:暗光周期下温育。在注射之后7天,获取生存力计数和重量。注射鉴定了bsbcopidelta是致死性dsrna靶.在bsb注射实验中使用靶向yfp编码区的区段yfpv2的dsrna作为阴性对照。如表14中总结的,将27.6ng的bsb_copigamma-2dsrna注射到第二龄bsb若虫的血腔中,在七天内产生了高死亡率。bsb_copigamma-2dsrna导致的死亡率均与相同量注射的yfpv2dsrna(阴性对照)所见到的显著不同,其p=0.001935(student'st-检验)。表14.bsb_copigamma-2dsrna注射到第二龄棕椿象若虫的血腔中注射七天之后的结果。*每种dsrna每次试验注射10只昆虫实施例13包含半翅目害虫序列的转基因玉米如实施例7中所述,生成10到20个包含表达载体的转基因t0玉米植物,所述表达载体含有seqidno:87和/或seqidno89的核酸。获得另外的10-20个t1玉米独立系,其表达针对rnai构建体的发夹dsrna,用于bsb攻击。可以获得包含如seqidno:89中所示,或进一步包含seqidno:87的发夹dsrna。这些通过rt-pcr或其他分子分析方法得以证实。任选地用选定的独立t1系的总rna制备物进行rt-pcr,其中引物设计为结合每个rnai构建体中的发夹表达盒的st-ls1内含子中。另外,任选地用针对rnai构建体中每个靶基因的特异性引物进行扩增,并且在植物体内确认产生sirna所需要的预加工mrna的产生。每个靶基因的各期望条带的扩增可确认每个转基因玉米植物中发夹rna的表达。任选地随后在独立转基因系中利用rna印迹杂交来确认靶基因的dsrna发夹加工成sirna。而且,与靶基因具有80%以上序列同一性的具有错配序列的rnai分子以类似于与靶基因具有100%序列同一性的rnai分子所见的方式影响玉米根虫。错配序列与天然序列的配对在同一个rnai构建体中形成发夹dsrna,从而投放出能够影响摄食的半翅目害虫的生长、发育和生存力的植物加工的sirna。在植物体内投予对应于靶基因的dsrna、sirna、shrna、或mirna,随后被半翅目害虫通过摄食而摄取,结果导致半翅目害虫中的靶基因由于rna介导的基因沉默作用而下调。当靶基因在所影响的半翅目害虫的生长、发育和生殖的一个或多个阶段中发挥重要功能时,并且在英雄美洲蝽、盖德拟壁蝽、茶翅蝽、稻绿蝽、拟绿蝽、和褐椿象中至少一者的情况下,可导致半翅目害虫无法成功侵染、进食、发育、和/或生殖,或导致其死亡。然后选定的靶基因的和rnai的成功应用用来控制半翅目害虫。转基因rnai系和非转化玉米的表型比较.选用于创建发夹dsrna的靶半翅目害虫基因或序列与任何已知的植物基因序列都没有相似性。因此可以预期,靶向这些半翅目害虫基因或序列的构建体的(系统性)rnai的产生或者激活不会对转基因植物产生任何有害影响。尽管如此,将转基因系的发育和形态特征与非转化植物、以及用没有发夹表达基因的“空”载体转化的转基因系进行比较。比较植物根、芽、叶和生殖特征。转基因植物和非转化植物在根长和生长模式上没有可观察到的差异。诸如高度、叶数和大小、开花时间、花大小和外观之类的植物地上部特征是相似的。总的来说,在体外以及在温室土壤中培养时,在转基因品系和没有表达靶irna分子的品系之间没有可观察到的形态差异。实施例14包含半翅目害虫序列的转基因大豆生成10到20个包含下述核酸的表达载体的转基因t0大豆植物,所述核酸含有seqidno:87和/或89,所述植物按照本领域已知的方式生成,包括例如通过如下所述的土壤杆菌介导的转化。用氯气将成熟大豆(glycinemax)种子消毒十六小时过夜。用氯气消毒之后,将种子置于laminartm层流罩内的开放容器中以驱散氯气。接着,使用黑盒,在黑暗中24℃下使消毒过的种子吸收无菌h2o十六小时。分割种子大豆的制备.包含部分胚轴的大豆种子分割方案需要制备纵向切开的大豆种子材料,纵向切开使用固定在解剖刀上的10号刀片,沿着种子的种脐分离并去除种皮,并将种子分开为两个子叶部分。小心地部分移除胚轴,其中约1/2-1/3的胚轴仍然附着在子叶的节端。接种.然后将包含部分胚轴的分割大豆种子在根癌土壤杆菌(例如,菌株eha101或eha105)溶液中浸没约30分钟,根癌土壤杆菌携带包含seqidno:87和/或seqidno:89的二元质粒。在浸入带胚轴的子叶之前,将根癌土壤杆菌溶液稀释到λ=0.6od650的终浓度。共培养.接种后,将分割的大豆种子与根癌土壤杆菌菌株在共培养基(wang,kan.agrobacteriumprotocols.2.1.newjersey:humanapress,2006.print.)上共培养5天,共培养基置于培养皿中,培养皿用一片滤纸覆盖。芽诱导.在共培养5天之后,将分割的大豆种子在含有b5盐、b5维生素、28mg/l二价铁、38mg/lna2edta、30g/l蔗糖、0.6g/lmes、1.11mg/lbap、100mg/ltimentintm、200mg/l头孢噻肟、和50mg/l万古霉素的液体芽诱导(si)(ph5.7)培养基中洗涤。然后将分割的大豆种子在含有b5盐、b5维生素、7g/lnoble琼脂、28mg/l二价铁、38mg/lna2edta、30g/l蔗糖、0.6g/lmes、1.11mg/lbap、50mg/ltimentintm、200mg/l头孢噻肟、50mg/l万古霉素的芽诱导i(sii)培养基(ph5.7)上培养,其中子叶的平坦一侧向上,而子叶的节端埋入培养基中。在培养2周之后,将来自转化的分割大豆种子的外植体转移到芽诱导ii(siii)培养基中,该培养基含有补充了6mg/l草丁膦的sii培养基。芽伸长.在siii培养基上培养2周之后,将子叶从外植体移除,通过在子叶的基部做切口切下含有胚轴的齐平的(flush)芽垫。将分离自子叶的芽垫转移到芽伸长(se)培养基上。该se培养基含有ms盐、28mg/l二价铁、38mg/lna2edta、30g/l蔗糖和0.6g/lmes、50mg/l天冬酰胺、100mg/ll--焦谷氨酸、0.1mg/liaa、0.5mg/lga3、1mg/l玉米素核苷、50mg/ltimentintm、200mg/l头孢噻肟、50mg/l万古霉素、6mg/l草丁膦、7g/lnoble琼脂,(ph5.7)。每2周将培养物转移到新鲜的se培养基上。培养物在生长室中在24℃下培养,使用18h光周期,光强为80-90μmol/m2sec。生根.对于从子叶芽垫发出的伸长芽,通过在子叶芽垫基部切割伸长芽加以分离,并将伸长芽浸入1mg/liba(吲哚-3-丁酸)中1-3分钟促进生根。接着,将伸长芽转移到phyta托盘中的生根培养基(ms盐、b5维生素、28mg/l二价铁、38mg/lna2edta、20g/l蔗糖和0.59g/lmes、50mg/l天冬酰胺、100mg/ll-焦谷氨酸、7g/lnoble琼脂,ph5.6)中。栽培.在convirontm生长室24℃、18h光周期培养1-2周之后,将已经生根的芽转移到带盖的圣代杯内的土壤混合物中,放到convirontm生长室(型号cmp4030和cmp3244,controlledenvironmentslimited,winnipeg,manitoba,canada)内,置于长的白昼条件下(16小时光照/8小时黑暗),光强度为120-150μmol/m2sec,温度(22℃)和湿度(40-50%)恒定,以驯化小植物。生根的小植物在圣代杯中驯化数周之后,转移到温室中进行进一步的驯化并定植强健的转基因大豆植物。获得另外的10-20个表达针对rnai构建体的发夹dsrna的t1大豆独立系用于bsb攻击。可以获得如seqidno:89中所示,或进一步包含seqidno:87的发夹dsrna。这些通过rt-pcr或其他分子分析方法加以确认。任选用来自选择的独立t1系的总rna制备物进行rt-pcr,其中引物设计为结合每个rnai构建体中的发夹表达盒中的st-ls1内含子。另外,任选用针对rnai构建体中每个靶基因的特异性引物进行扩增,并且在植物体内确认产生sirna所需要的预加工mrna的产生。每个靶基因的期望条带的扩增可确认每个转基因玉米植物中发夹rna的表达。任选地随后在独立转基因系中利用rna印迹杂交来确认靶基因的dsrna发夹加工成sirna。此外,与靶基因具有80%以上序列同一性且具有错配序列的rnai分子影响玉米根虫的方式类似于与靶基因具有100%序列同一性的rnai分子。错配序列与天然序列的配对在同一个rnai构建体中形成发夹dsrna,从而投放出能够影响摄食的半翅目害虫的生长、发育和生存力的植物加工的sirna。在植物内投放对应于靶基因的dsrna、sirna、shrna、或mirna,随后被半翅目害虫通过摄食而摄取,结果导致半翅目害虫中的靶基因由于rna介导基因沉默作用而下调。当靶基因在发育的一个或多个阶段中重要时,半翅目害虫的生长、发育和/或存活受影响,并且在英雄美洲蝽、盖德拟壁蝽、茶翅蝽、稻绿蝽、绿蝽(acrosternumhilare)、褐椿象中至少一者的情况下,可导致半翅目害虫无法成功侵染、进食、发育和/或生殖,或导致其死亡。然后选定的靶基因的和rnai的成功应用可用来控制半翅目害虫。转基因rnai系和非转化玉米的表型比较.选用于创建发夹dsrna的靶半翅目害虫基因或序列与任何已知的植物基因序列都没有相似性。因此可以预期,靶向这些半翅目害虫基因或序列的构建体的(系统性)rnai的产生或者激活不会对转基因植物产生任何有害影响。尽管如此,将转基因系的发育和形态特征与非转化植物、以及用没有发夹表达基因的“空”载体转化的转基因系进行比较。比较植物根、芽、叶和生殖特征。转基因植物和非转化植物在根长和生长模式上没有可观察到的差异。诸如高度、叶数和大小、开花时间、花大小和外观之类的植物地上部特征是相似的。总的来说,在体外以及在温室土壤中培养时,在转基因品系和没有表达靶irna分子的品系之间没有可观察到的形态差异。实施例15人工饮食的英雄美洲蝽生物测定在使用人工饵食的dsrna饲喂测定中,与注射实验(实施例12)相同,设置具有一颗约18mg的人工饲料小丸和水的32孔托盘。将浓度为200ng/μl的dsrna加入到食物小丸和水样品中,向两个孔中的每一个加入100μl。向每个孔中加入5只第2龄的英雄美洲蝽若虫。以水样品和靶向yfp转录物的dsrna作为阴性对照。在三个不同的日期重复实验。称重存活的昆虫,并在处理8天后测定死亡率。实施例16包含半翅目害虫序列的转基因拟南芥使用与实施例4相似的标准分子方法产生包含用于发夹形成的靶基因构建体的拟南芥转化载体,所述构建体包含copigamma(seqidno:87)的区段。使用标准的基于土壤杆菌的方法进行拟南芥转化。用草铵膦耐受性选择标志物选择t1种子。产生转基因t1拟南芥植物,并产生纯合的单拷贝t2转基因植物用于昆虫研究。对具有花序的生长中的拟南芥植物进行生物测定。将五至十只昆虫放置在每株植物上,并在14天内监测存活。拟南芥转化载体的构建.使用化学合成的片段(dna2.0,menlopark,ca)的组合,以及标准的分子克隆方法,组装基于入门载体pdab3916的入门克隆,这些入门克隆包含用于发夹形成的靶基因构建体,靶基因构建体包含copigamma(seqidno:87)。通过将靶基因区段的两个拷贝以相反方向排列(在单个转录单元内),两个区段被st-ls1内含子序列(seqidno:17)(vancanneytetal.(1990)mol.gen.genet.220(2):245-50)所分隔,可以易化rna初级转录物的分子内发夹形成。因此,初级mrna转录物包含两个copigamma基因区段序列,它们彼此作为对方的大反向重复序列,由内含子序列分隔开。使用一个拟南芥泛素10启动子(callisetal.(1990)j.biologicalchem.265:12486-12493))的拷贝来驱动初级mrna发夹转录物的产生,并且使用包含来自根癌土壤杆菌的开放阅读框23(atuorf233'utrv1;美国专利号5,428,147)的3'非翻译区的片段来终止表达发夹rna的基因的转录。将上述pdab3916载体内的发夹克隆用于标准重组反应,使用典型的二元目的载体pdab101836,以产生用于土壤杆菌介导的拟南芥转化的发夹rna表达转化载体。二元目的载体pdab101836包含在木薯叶脉花叶病毒启动子(csvmv启动子v2,美国专利号us7601885;verdaguer等人(1996)plantmol.biol.31:1129-1139))控制下的除草剂耐受基因dsm-2v2(美国专利申请号2011/0107455)。使用包含来自根癌土壤杆菌开放阅读框1的3'非翻译区(atuorf13'utrv6;huang等人(1990)j.bacteriol,172:1814-1822)的片段来终止dsm2v2mrna的转录。通过标准重组反应用典型的二元目的载体(pdab101836)和入门载体pdab3916构建包含表达yfp发夹rna的基因的阴性对照二元构建体pdab114507。入门构建体pdab112644包含在拟南芥泛素10启动子(如上)的表达控制下的yfp发夹序列(hpyfpv2-1,seqidno:95)和包含来自根癌土壤杆菌的orf233'非翻译区的片段(如上所述)。包含杀虫发夹rna的转基因拟南芥的产生:土壤杆菌介导的转化.将含有发夹序列的二元质粒电穿孔到土壤杆菌菌株gv3101(pmp90rk)中。通过重组土壤杆菌菌落的质粒制备物的限制性分析确认重组土壤杆菌克隆。使用qiagenplasmidmaxkit(qiagen,cat#12162)根据制造商推荐的方案从土壤杆菌培养物中提取质粒。拟南芥转化和t1选择.将12至15株拟南芥植物(columbia栽培种)在温室中的4”盆中生长,光强度为250μmol/m2,25℃和18:6小时光照:黑暗条件。转化前一周修剪主花茎。通过将10μl的重组土壤杆菌甘油储在28℃、225rpm振荡下在100mllb肉汤(sigmal3022)+100mg/l壮观霉素+50mg/l卡那霉素中温育72小时来制备土壤杆菌接种物。收获土壤杆菌细胞,并将其悬浮于5%蔗糖+0.04%silwet-l77(lehleseedscat#vis-02)+10μg/l苯甲氨基嘌呤(ba)溶液中至od6000.8~1.0,然后浸花。将植株的地上部分浸入土壤杆菌溶液5-10分钟,轻柔搅动,然后将植株转移到温室中进行正常生长,并定期浇水和施肥,直到种子定植。实施例17转基因拟南芥的生长和生物测定选择用发夹rnai构建体转化的t1拟南芥.将来自每次转化的至多达200mg的t1种子在0.1%琼脂糖溶液中分层。将种子种植在装有5号阳光培养基(sunshinemedia)的发芽盘(10.5”×21”×1”;t.o.plasticsinc.,clearwater,mn)中。在种植后6天和9天,选择对280g/ha的(草铵膦)具有耐受性的转化体。将选出的事件移植到4”直径的盆中。在移植的一周内进行插入拷贝分析,插入拷贝分析使用rochelightcycler480的水解定量实时pcr(qpcr)。使用lightcyclerprobedesignsoftware2.0(roche),针对dsm2v2选择标记设计pcr引物和水解探针。将植物维持在24℃,在强度为100-150me/m2×s的荧光和白炽灯下以16:8小时光:暗光周期。英雄美洲蝽植物饲养生物测定.为每个构建体选择至少四个低拷贝事件(1-2个插入)、四个中拷贝事件(2-3个插入)和四个高拷贝(≥4个插入)事件。使植物生长至开花期(植物有花和长角果)。土壤表面覆盖有~50ml体积的白色沙子,便于鉴别昆虫。将五到十二龄的英雄美洲蝽若虫引入每株植物。植物用直径3”,高16”,壁厚0.03”的塑料管(产品号484485,visipackfentonmo)覆盖;用尼龙网盖住管子以隔离昆虫。在conviron培养箱中将植物保持在正常温度、光照和浇水条件下。在14天中,收集昆虫并称重,计算死亡率百分比以及生长抑制(1-处理重量/对照重量)。用表达yfp发夹的植物作为对照。t2拟南芥种子生成和t2生物测定.对于每个构建体,从选定的低拷贝(1-2个插入)事件产生t2种子。如上所述,对植物(纯合的和/或杂合的)进行英雄美洲蝽饲养生物测定。从纯合子收获t3种子并储存用于将来分析。上文已经通过实施例详细描述了具体实施方案,但本公开可能容许各种修改和替代形式。然而,应当理解,本公开不旨在限于所公开的特定形式。相反,本公开应覆盖落入由附随的权利要求及其法律上等同物限定的本公开的范围内的所有修改、等同物和替代物。序列表<110>美国陶氏益农公司k·纳瓦h·李c·耿n·埃兰戈m·j·亨利m·兰加萨米a·t·伍斯利k·阿罗拉p·甘德拉s·e·沃登e·菲什里维奇<120>赋予鞘翅目和半翅目害虫抗性的copi外被体gamma亚单位核酸分子<130>74227<150>62/063192<151>2014-10-13<160>95<170>patentinversion3.5<210>1<211>2840<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<400>1tggtttttttttatcaaagttttatatggaaacttataaattggttttttgcttaaaatt60tgttttcagttttgacgtttattttggtcgtcggctgatgtgacgtgtaaagaaattaaa120tcaaattattttaaagtttttaaattaaaatgggtacttttaaaagagatactcatgatg180aggacgggggatcaagtgcttttcaaaatctggagaaaactactgttttgcaggaagcta240gagtttttaatgaaactagtgtaaatccaagaaaatgtacaccgatactaacaaaactgt300tgtacttattgaaccagggtgaaactttaagtgccaaagaggccacagatgttttctttg360ccatgaccaaactgttccaatcaaaagatgtaatattgagaaggatggtttatttgggaa420ttaaagaactcagttctgttgctgatgatgtcattattgtaacatccagtcttacaaaag480atatgactggtaaagaagacatgtacagagcagctgctataagagcattatgcagtatta540ctgatgctactatgcttcaagctatagaacgttatatgaagcaagctattgtagatagaa600acgcagctgtcagttcagcagcactaattagttcattacatatgagcaaattagctccag660atgtagtaaaaagatgggtaaatgaagctcaggaagcagtaaatagtgataatgcaatgg720tacagtatcacgcattaggtcttctataccatattaggaagactgataagctagcagtga780caaaattgatttccaagctgaattcaatgggtttaaagagcccttatgctttgtgtatgt840tgataagaatcactgcaaaacttttagaagaagaggaccaagagtcactcctcaactccc900catatacaataatatttacaatgggcttaaggaacaaatctgaaatggtggtgtatgaag960ctgcacatgccatggttaacctgaagttcacgagtagtaatgtgctagcacccgctataa1020gtgttctacaactattttgtggatctcctaaagccacactcagatttgctgctgttagaa1080ctttaaatcaagtggccaccacccaccctgcgtcagtgacagcttgtaatttggatctag1140aaaatttgattactgatcctaataggtcaattgctacactggccattactactcttttga1200aaacaggtgccgaatcttctgttgacagactaatgaaacaaatcgctacttttgtatctg1260aaatcagtgatgaatttaaagtggttgtcattcaggcaattaaggtattagctttgaaat1320ttccaaggaaacatagcacgcttatgaatttcctatccgccatgttaagagatgagggag1380gtttagaatataaagcatccatagcagataccattataaccctaatcgaagataatcccg1440aagctaaagaatctggtttggcgcatctttgcgagttcattgaagactgtgaacatgttt1500ctttggctgtgagaatcttgcatttgttaggaaaggaaggacccaagaccaaacaaccat1560cgagatacatccgttttatctacaatcgcgtcatattggaatgtccttctgtaagagctg1620ctgcagtctccgccatggcacaattcggagcctcttgtcccgatttgttagaaaatatcc1680aaatattactttcgaggtgtcagatggattcagacgatgaagttagggacagagctacat1740attatagtaatatacttaacaaaaatgataaaagtttatacaacaattacattttggatt1800ctttgcaggtttcaattccttcactagaaagatcgcttagagaatacattcaaaatccaa1860ctgacgaaccatttgacattaagtccgtacctgtagcagcagtgccaacagcagaagaac1920gagaagttaaaaacaaatctgaaggactgctagtctctcaaggtccagtccgacctcctc1980cggtgtctagagaagaaaacttcgccgaaaaacttagtaacgttccgggtatacaacagt2040taggacctttgttcaaaacttccgacgtcgttgaactcactgaatctgaaacagagtatt2100ttgtccgctgtatcaagcactgtttcaaacatcacatcgtcctccaattcgattgtctga2160ataccttgccagaccagcttttagaaaacgttagagtggagatagacgccggtgaaacct2220tcgaaattttggcagaaataccttgtgaaaagttgcactataacgaaaccggtaccacat2280atgtagtagttaagttgcctgatgatgatctccccaactctgttggtacgtgtggagccg2340tgttgaagttcttagtgaaagattgtgatccatcaacgggaataccagattctgatgagg2400gttacgatgatgaatatacactggaagacatcgaaataacattaggggaccaaattcaaa2460aagtaagcaaagtaaattgggctgcagcctgggaagaagctgcagctacttatgtagaaa2520aagaggatacatactccttgaccatcaatacgctaagtggcgctgttaagaatattattc2580agttcttgggattacagcctgcggaaaggactgacagagtaccggagggtaaatctacgc2640acacattacttcttgctggtgtattcaggggaggtattgacatactagtaagagcgaaac2700tagctttgggcgaatgtgttacgatgcaactaacagtcaggtcgccagatcctgacgttg2760ctgagcttataacttcaactgtaggttaagtttaaaggctacgttaatgattatattgta2820ttacaatttttccatatgcg2840<210>2<211>879<212>prt<213>玉米根萤叶甲(diabroticavirgifera)<400>2metglythrphelysargaspthrhisaspgluaspglyglyserser151015alapheglnasnleuglulysthrthrvalleuglnglualaargval202530pheasngluthrservalasnproarglyscysthrproileleuthr354045lysleuleutyrleuleuasnglnglygluthrleuseralalysglu505560alathraspvalphephealametthrlysleupheglnserlysasp65707580valileleuargargmetvaltyrleuglyilelysgluleuserser859095valalaaspaspvalileilevalthrserserleuthrlysaspmet100105110thrglylysgluaspmettyrargalaalaalaileargalaleucys115120125serilethraspalathrmetleuglnalailegluargtyrmetlys130135140glnalailevalaspargasnalaalavalserseralaalaleuile145150155160serserleuhismetserlysleualaproaspvalvallysargtrp165170175valasnglualaglnglualavalasnseraspasnalametvalgln180185190tyrhisalaleuglyleuleutyrhisilearglysthrasplysleu195200205alavalthrlysleuileserlysleuasnsermetglyleulysser210215220protyralaleucysmetleuileargilethralalysleuleuglu225230235240glugluaspglngluserleuleuasnserprotyrthrileilephe245250255thrmetglyleuargasnlysserglumetvalvaltyrglualaala260265270hisalametvalasnleulysphethrserserasnvalleualapro275280285alaileservalleuglnleuphecysglyserprolysalathrleu290295300argphealaalavalargthrleuasnglnvalalathrthrhispro305310315320alaservalthralacysasnleuaspleugluasnleuilethrasp325330335proasnargserilealathrleualailethrthrleuleulysthr340345350glyalagluserservalaspargleumetlysglnilealathrphe355360365valsergluileseraspgluphelysvalvalvalileglnalaile370375380lysvalleualaleulyspheproarglyshisserthrleumetasn385390395400pheleuseralametleuargaspgluglyglyleuglutyrlysala405410415serilealaaspthrileilethrleuilegluaspasnprogluala420425430lysgluserglyleualahisleucysglupheilegluaspcysglu435440445hisvalserleualavalargileleuhisleuleuglylysglugly450455460prolysthrlysglnproserargtyrileargpheiletyrasnarg465470475480valileleuglucysproservalargalaalaalavalseralamet485490495alaglnpheglyalasercysproaspleuleugluasnileglnile500505510leuleuserargcysglnmetaspseraspaspgluvalargasparg515520525alathrtyrtyrserasnileleuasnlysasnasplysserleutyr530535540asnasntyrileleuaspserleuglnvalserileproserleuglu545550555560argserleuargglutyrileglnasnprothraspglupropheasp565570575ilelysservalprovalalaalavalprothralaglugluargglu580585590vallysasnlyssergluglyleuleuvalserglnglyprovalarg595600605proproprovalserargglugluasnphealaglulysleuserasn610615620valproglyileglnglnleuglyproleuphelysthrseraspval625630635640valgluleuthrglusergluthrglutyrphevalargcysilelys645650655hiscysphelyshishisilevalleuglnpheaspcysleuasnthr660665670leuproaspglnleuleugluasnvalargvalgluileaspalagly675680685gluthrphegluileleualagluileprocysglulysleuhistyr690695700asngluthrglythrthrtyrvalvalvallysleuproaspaspasp705710715720leuproasnservalglythrcysglyalavalleulyspheleuval725730735lysaspcysaspproserthrglyileproaspseraspgluglytyr740745750aspaspglutyrthrleugluaspilegluilethrleuglyaspgln755760765ileglnlysvalserlysvalasntrpalaalaalatrpglugluala770775780alaalathrtyrvalglulysgluaspthrtyrserleuthrileasn785790795800thrleuserglyalavallysasnileileglnpheleuglyleugln805810815proalagluargthraspargvalprogluglylysserthrhisthr820825830leuleuleualaglyvalpheargglyglyileaspileleuvalarg835840845alalysleualaleuglyglucysvalthrmetglnleuthrvalarg850855860serproaspproaspvalalagluleuilethrserthrvalgly865870875<210>3<211>303<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<400>3gacatgtacagagcagctgctataagagcattatgcagtattactgatgctactatgctt60caagctatagaacgttatatgaagcaagctattgtagatagaaacgcagctgtcagttca120gcagcactaattagttcattacatatgagcaaattagctccagatgtagtaaaaagatgg180gtaaatgaagctcaggaagcagtaaatagtgataatgcaatggtacagtatcacgcatta240ggtcttctataccatattaggaagactgataagctagcagtgacaaaattgatttccaag300ctg303<210>4<211>332<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<400>4gggtttaaagagcccttatgctttgtgtatgttgataagaatcactgcaaaacttttaga60agaagaggaccaagagtcactcctcaactccccatatacaataatatttacaatgggctt120aaggaacaaatctgaaatggtggtgtatgaagctgcacatgccatggttaacctgaagtt180cacgagtagtaatgtgctagcacccgctataagtgttctacaactattttgtggatctcc240taaagccacactcagatttgctgctgttagaactttaaatcaagtggccaccacccaccc300tgcgtcagtgacagcttgtaatttggatctag332<210>5<211>530<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<400>5gtggttgtcattcaggcaattaaggtattagctttgaaatttccaaggaaacatagcacg60cttatgaatttcctatccgccatgttaagagatgagggaggtttagaatataaagcatcc120atagcagataccattataaccctaatcgaagataatcccgaagctaaagaatctggtttg180gcgcatctttgcgagttcattgaagactgtgaacatgtttctttggctgtgagaatcttg240catttgttaggaaaggaaggacccaagaccaaacaaccatcgagatacatccgttttatc300tacaatcgcgtcatattggaatgtccttctgtaagagctgctgcagtctccgccatggca360caattcggagcctcttgtcccgatttgttagaaaatatccaaatattactttcgaggtgt420cagatggattcagacgatgaagttagggacagagctacatattatagtaatatacttaac480aaaaatgataaaagtttatacaacaattacattttggattctttgcaggt530<210>6<211>24<212>dna<213>人工序列<220><223>合成的启动子寡核苷酸<400>6ttaatacgactcactatagggaga24<210>7<211>503<212>dna<213>人工序列<220><223>合成部分编码区<400>7caccatgggctccagcggcgccctgctgttccacggcaagatcccctacgtggtggagat60ggagggcaatgtggatggccacaccttcagcatccgcggcaagggctacggcgatgccag120cgtgggcaaggtggatgcccagttcatctgcaccaccggcgatgtgcccgtgccctggag180caccctggtgaccaccctgacctacggcgcccagtgcttcgccaagtacggccccgagct240gaaggatttctacaagagctgcatgcccgatggctacgtgcaggagcgcaccatcacctt300cgagggcgatggcaatttcaagacccgcgccgaggtgaccttcgagaatggcagcgtgta360caatcgcgtgaagctgaatggccagggcttcaagaaggatggccacgtgctgggcaagaa420tctggagttcaatttcaccccccactgcctgtacatctggggcgatcaggccaatcacgg480cctgaagagcgccttcaagatct503<210>8<211>46<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>8ttaatacgactcactatagggagaaccatggcgttaaagaaccaag46<210>9<211>44<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>9ttaatacgactcactatagggagagggtggtggcacaaggtact44<210>10<211>42<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>10ttaatacgactcactatagggagactcgaccgaggtttcgac42<210>11<211>46<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>11ttaatacgactcactatagggagataactgaaggttggcgatggtc46<210>12<211>47<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>12ttaatacgactcactatagggagacaccatgggctccagcggcgccc47<210>13<211>47<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>13ttaatacgactcactatagggagaagatcttgaaggcgctcttcagg47<210>14<211>445<212>dna<213>人工序列<220><223>synthesized人工序列<400>14cgacctcctccggtgtctagagaagaaaacttcgccgaaaaacttagtaacgttccgggt60atacaacagttaggacctttgttcaaaacttccgacgtcgttgaactcacgactagtacc120ggttgggaaaggtatgtttctgcttctacctttgatatatatataataattatcactaat180tagtagtaatatagtatttcaagtatttttttcaaaataaaagaatgtagtatatagcta240ttgcttttctgtagtttataagtgtgtatattttaatttataacttttctaatatatgac300caaaacatggtgatgtgcaggttgatccgcggttagtgagttcaacgacgtcggaagttt360tgaacaaaggtcctaactgttgtatacccggaacgttactaagtttttcggcgaagtttt420cttctctagacaccggaggaggtcg445<210>15<211>626<212>dna<213>人工序列<220><223>合成的人工序列<400>15agttgcactataacgaaaccggtaccacatatgtagtagttaagttgcctgatgatgatc60tccccaactctgttggtacgtgtggagccgtgttgaagttcttagtgaaagattgtgatc120catcaacgggaataccagattctgatgagggttacgatgatgaatatacactggaagaca180tcgaaataacattaggggacgactagtaccggttgggaaaggtatgtttctgcttctacc240tttgatatatatataataattatcactaattagtagtaatatagtatttcaagtattttt300ttcaaaataaaagaatgtagtatatagctattgcttttctgtagtttataagtgtgtata360ttttaatttataacttttctaatatatgaccaaaacatggtgatgtgcaggttgatccgc420ggttaggtcccctaatgttatttcgatgtcttccagtgtatattcatcatcgtaaccctc480atcagaatctggtattcccgttgatggatcacaatctttcactaagaacttcaacacggc540tccacacgtaccaacagagttggggagatcatcatcaggcaacttaactactacatatgt600ggtaccggtttcgttatagtgcaact626<210>16<211>471<212>dna<213>人工序列<220><223>合成的人工序列<400>16atgtcatctggagcacttctctttcatgggaagattccttacgttgtggagatggaaggg60aatgttgatggccacacctttagcatacgtgggaaaggctacggagatgcctcagtggga120aaggactagtaccggttgggaaaggtatgtttctgcttctacctttgatatatatataat180aattatcactaattagtagtaatatagtatttcaagtatttttttcaaaataaaagaatg240tagtatatagctattgcttttctgtagtttataagtgtgtatattttaatttataacttt300tctaatatatgaccaaaacatggtgatgtgcaggttgatccgcggttactttcccactga360ggcatctccgtagcctttcccacgtatgctaaaggtgtggccatcaacattcccttccat420ctccacaacgtaaggaatcttcccatgaaagagaagtgctccagatgacat471<210>17<211>225<212>dna<213>solanumtuberosum<400>17gactagtaccggttgggaaaggtatgtttctgcttctacctttgatatatatataataat60tatcactaattagtagtaatatagtatttcaagtatttttttcaaaataaaagaatgtag120tatatagctattgcttttctgtagtttataagtgtgtatattttaatttataacttttct180aatatatgaccaaaacatggtgatgtgcaggttgatccgcggtta225<210>18<211>705<212>dna<213>人工序列<220><223>合成的人工序列<400>18atgtcatctggagcacttctctttcatgggaagattccttacgttgtggagatggaaggg60aatgttgatggccacacctttagcatacgtgggaaaggctacggagatgcctcagtggga120aaggttgatgcacagttcatctgcacaactggtgatgttcctgtgccttggagcacactt180gtcaccactctcacctatggagcacagtgctttgccaagtatggtccagagttgaaggac240ttctacaagtcctgtatgccagatggctatgtgcaagagcgcacaatcacctttgaagga300gatggcaacttcaagactagggctgaagtcacctttgagaatgggtctgtctacaatagg360gtcaaactcaatggtcaaggcttcaagaaagatggtcatgtgttgggaaagaacttggag420ttcaacttcactccccactgcctctacatctggggtgaccaagccaaccacggtctcaag480tcagccttcaagatctgtcatgagattactggcagcaaaggcgacttcatagtggctgac540cacacccagatgaacactcccattggtggaggtccagttcatgttccagagtatcatcac600atgtcttaccatgtgaaactttccaaagatgtgacagaccacagagacaacatgtccttg660aaagaaactgtcagagctgttgactgtcgcaagacctacctttga705<210>19<211>218<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<400>19tagctctgatgacagagcccatcgagtttcaagccaaacagttgcataaagctatcagcg60gattgggaactgatgaaagtacaatmgtmgaaattttaagtgtmcacaacaacgatgaga120ttataagaatttcccaggcctatgaaggattgtaccaacgmtcattggaatctgatatca180aaggagatacctcaggaacattaaaaaagaattattag218<210>20<211>424<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<220><221>misc_feature<222>(393)..(393)<223>nisa,c,g,ort<220><221>misc_feature<222>(394)..(394)<223>nisa,c,g,ort<220><221>misc_feature<222>(395)..(395)<223>n为a、c、g或t<400>20ttgttacaagctggagaacttctctttgctggaaccgaagagtcagtatttaatgctgta60ttctgtcaaagaaataaaccacaattgaatttgatattcgacaaatatgaagaaattgtt120gggcatcccattgaaaaagccattgaaaacgagttttcaggaaatgctaaacaagccatg180ttacaccttatccagagcgtaagagatcaagttgcatatttggtaaccaggctgcatgat240tcaatggcaggcgtcggtactgacgatagaactttaatcagaattgttgtttcgagatct300gaaatcgatctagaggaaatcaaacaatgctatgaagaaatctacagtaaaaccttggct360gataggatagcggatgacacatctggcgactannnaaaagccttattagccgttgttggt420taag424<210>21<211>397<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<400>21agatgttggctgcatctagagaattacacaagttcttccatgattgcaaggatgtactga60gcagaatagtggaaaaacaggtatccatgtctgatgaattgggaagggacgcaggagctg120tcaatgcccttcaacgcaaacaccagaacttcctccaagacctacaaacactccaatcga180acgtccaacaaatccaagaagaatcagctaaacttcaagctagctatgccggtgatagag240ctaaagaaatcaccaacagggagcaggaagtggtagcagcctgggcagccttgcagatcg300cttgcgatcagagacacggaaaattgagcgatactggtgatctattcaaattctttaact360tggtacgaacgttgatgcagtggatggacgaatggac397<210>22<211>490<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<400>22gcagatgaacaccagcgagaaaccaagagatgttagtggtgttgaattgttgatgaacaa60ccatcagacactcaaggctgagatcgaagccagagaagacaactttacggcttgtatttc120tttaggaaaggaattgttgagccgtaatcactatgctagtgctgatattaaggataaatt180ggtcgcgttgacgaatcaaaggaatgctgtactacagaggtgggaagaaagatgggagaa240cttgcaactcatcctcgaggtataccaattcgccagagatgcggccgtcgccgaagcatg300gttgatcgcacaagaaccttacttgatgagccaagaactaggacacaccattgacgacgt360tgaaaacttgataaagaaacacgaagcgttcgaaaaatcggcagcggcgcaagaagagag420attcagtgctttggagagactgacgacgttcgaattgagagaaataaagaggaaacaaga480agctgcccag490<210>23<211>330<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<400>23agtgaaatgttagcaaatataacatccaagtttcgtaattgtacttgctcagttagaaaa60tattctgtagtttcactatcttcaaccgaaaatagaataaatgtagaacctcgcgaactt120gcctttcctccaaaatatcaagaacctcgacaagtttggttggagagtttagatacgata180gacgacaaaaaattgggtattcttgagctgcatcctgatgtttttgctactaatccaaga240atagatattatacatcaaaatgttagatggcaaagtttatatagatatgtaagctatgct300catacaaagtcaagatttgaagtgagaggt330<210>24<211>320<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<400>24caaagtcaagatttgaagtgagaggtggaggtcgaaaaccgtggccgcaaaagggattgg60gacgtgctcgacatggttcaattagaagtccactttggagaggtggaggagttgttcatg120gaccaaaatctccaacccctcatttttacatgattccattctacacccgtttgctgggtt180tgactagcgcactttcagtaaaatttgcccaagatgacttgcacgttgtggatagtctag240atctgccaactgacgaacaaagttatatagaagagctggtcaaaagccgcttttgggggt300ccttcttgttttatttgtag320<210>25<211>47<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>25ttaatacgactcactatagggagacaccatgggctccagcggcgccc47<210>26<211>23<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>26agatcttgaaggcgctcttcagg23<210>27<211>23<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>27caccatgggctccagcggcgccc23<210>28<211>47<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>28ttaatacgactcactatagggagaagatcttgaaggcgctcttcagg47<210>29<211>46<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>29ttaatacgactcactatagggagagctccaacagtggttccttatc46<210>30<211>29<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>30ctaataattcttttttaatgttcctgagg29<210>31<211>22<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>31gctccaacagtggttccttatc22<210>32<211>53<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>32ttaatacgactcactatagggagactaataattcttttttaatgttcctgagg53<210>33<211>48<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>33ttaatacgactcactatagggagattgttacaagctggagaacttctc48<210>34<211>24<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>34cttaaccaacaacggctaataagg24<210>35<211>24<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>35ttgttacaagctggagaacttctc24<210>36<211>48<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>36ttaatacgactcactatagggagacttaaccaacaacggctaataagg48<210>37<211>47<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>37ttaatacgactcactatagggagaagatgttggctgcatctagagaa47<210>38<211>22<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>38gtccattcgtccatccactgca22<210>39<211>23<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>39agatgttggctgcatctagagaa23<210>40<211>46<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>40ttaatacgactcactatagggagagtccattcgtccatccactgca46<210>41<211>46<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>41ttaatacgactcactatagggagagcagatgaacaccagcgagaaa46<210>42<211>22<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>42ctgggcagcttcttgtttcctc22<210>43<211>22<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>43gcagatgaacaccagcgagaaa22<210>44<211>46<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>44ttaatacgactcactatagggagactgggcagcttcttgtttcctc46<210>45<211>51<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>45ttaatacgactcactatagggagaagtgaaatgttagcaaatataacatcc51<210>46<211>26<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>46acctctcacttcaaatcttgactttg26<210>47<211>27<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>47agtgaaatgttagcaaatataacatcc27<210>48<211>50<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>48ttaatacgactcactatagggagaacctctcacttcaaatcttgactttg50<210>49<211>50<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>49ttaatacgactcactatagggagacaaagtcaagatttgaagtgagaggt50<210>50<211>25<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>50ctacaaataaaacaagaaggacccc25<210>51<211>26<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>51caaagtcaagatttgaagtgagaggt26<210>52<211>49<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>52ttaatacgactcactatagggagactacaaataaaacaagaaggacccc49<210>53<211>1150<212>dna<213>zeamays<400>53caacggggcagcactgcactgcactgcaactgcgaatttccgtcagcttggagcggtcca60agcgccctgcgaagcaaactacgccgatggcttcggcggcggcgtgggagggtccgacgg120ccgcggagctgaagacagcgggggcggaggtgattcccggcggcgtgcgagtgaaggggt180gggtcatccagtcccacaaaggccctatcctcaacgccgcctctctgcaacgctttgaag240atgaacttcaaacaacacatttacctgagatggtttttggagagagtttcttgtcacttc300aacatacacaaactggcatcaaatttcattttaatgcgcttgatgcactcaaggcatgga360agaaagaggcactgccacctgttgaggttcctgctgcagcaaaatggaagttcagaagta420agccttctgaccaggttatacttgactacgactatacatttacgacaccatattgtggga480gtgatgctgtggttgtgaactctggcactccacaaacaagtttagatggatgcggcactt540tgtgttgggaggatactaatgatcggattgacattgttgccctttcagcaaaagaaccca600ttcttttctacgacgaggttatcttgtatgaagatgagttagctgacaatggtatctcat660ttcttactgtgcgagtgagggtaatgccaactggttggtttctgcttttgcgtttttggc720ttagagttgatggtgtactgatgaggttgagagacactcggttacattgcctgtttggaa780acggcgacggagccaagccagtggtacttcgtgagtgctgctggagggaagcaacatttg840ctactttgtctgcgaaaggatatccttcggactctgcagcgtacgcggacccgaacctta900ttgcccataagcttcctattgtgacgcagaagacccaaaagctgaaaaatcctacctgac960tgacacaaaggcgccctaccgcgtgtacatcatgactgtcctgtcctatcgttgcctttt1020gtgtttgccacatgttgtggatgtacgtttctatgacgaaacaccatagtccatttcgcc1080tgggccgaacagagatagctgattgtcatgtcacgtttgaattagaccattccttagccc1140tttttccccc1150<210>54<211>22<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<220><221>misc_feature<222>(22)..(22)<223>n为a、c、g或t<400>54ttttttttttttttttttttvn22<210>55<211>20<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>55ttgtgatgttggtggcgtat20<210>56<211>24<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>56tgttaaataaaaccccaaagatcg24<210>57<211>21<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>57tgagggtaatgccaactggtt21<210>58<211>24<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>58gcaatgtaaccgagtgtctctcaa24<210>59<211>32<212>dna<213>人工序列<220><223>合成的探针寡核苷酸<400>59tttttggcttagagttgatggtgtactgatga32<210>60<211>151<212>dna<213>escherichiacoli<400>60gaccgtaaggcttgatgaaacaacgcggcgagctttgatcaacgaccttttggaaacttc60ggcttcccctggagagagcgagattctccgcgctgtagaagtcaccattgttgtgcacga120cgacatcattccgtggcgttatccagctaag151<210>61<211>69<212>dna<213>人工序列<220><223>合成部分编码区<400>61tgttcggttccctctaccaagcacagaaccgtcgcttcagcaacacctcagtcaaggtga60tggatgttg69<210>62<211>4233<212>dna<213>zeamays<400>62agcctggtgtttccggaggagacagacatgatccctgccgttgctgatccgacgacgctg60gacggcgggggcgcgcgcaggccgttgctcccggagacggaccctcgggggcgtgctgcc120gccggcgccgagcagaagcggccgccggctacgccgaccgttctcaccgccgtcgtctcc180gccgtgctcctgctcgtcctcgtggcggtcacagtcctcgcgtcgcagcacgtcgacggg240caggctgggggcgttcccgcgggcgaagatgccgtcgtcgtcgaggtggccgcctcccgt300ggcgtggctgagggcgtgtcggagaagtccacggccccgctcctcggctccggcgcgctc360caggacttctcctggaccaacgcgatgctggcgtggcagcgcacggcgttccacttccag420ccccccaagaactggatgaacggttagttggacccgtcgccatcggtgacgacgcgcgga480tcgtttttttcttttttcctctcgttctggctctaacttggttccgcgtttctgtcacgg540acgcctcgtgcacatggcgatacccgatccgccggccgcgtatatctatctacctcgacc600ggcttctccagatccgaacggtaagttgttggctccgatacgatcgatcacatgtgagct660cggcatgctgcttttctgcgcgtgcatgcggctcctagcattccacgtccacgggtcgtg720acatcaatgcacgatataatcgtatcggtacagagatattgtcccatcagctgctagctt780tcgcgtattgatgtcgtgacattttgcacgcaggtccgctgtatcacaagggctggtacc840acctcttctaccagtggaacccggactccgcggtatggggcaacatcacctggggccacg900ccgtctcgcgcgacctcctccactggctgcacctaccgctggccatggtgcccgatcacc960cgtacgacgccaacggcgtctggtccgggtcggcgacgcgcctgcccgacggccggatcg1020tcatgctctacacgggctccacggcggagtcgtcggcgcaggtgcagaacctcgcggagc1080cggccgacgcgtccgacccgctgctgcgggagtgggtcaagtcggacgccaacccggtgc1140tggtgccgccgccgggcatcgggccgacggacttccgcgacccgacgacggcgtgtcgga1200cgccggccggcaacgacacggcgtggcgggtcgccatcgggtccaaggaccgggaccacg1260cggggctggcgctggtgtaccggacggaggacttcgtgcggtacgacccggcgccggcgc1320tgatgcacgccgtgccgggcaccggcatgtgggagtgcgtggacttctacccggtggccg1380cgggatcaggcgccgcggcgggcagcggggacgggctggagacgtccgcggcgccgggac1440ccggggtgaagcacgtgctcaaggctagcctcgacgacgacaagcacgactactacgcga1500tcggcacctacgacccggcgacggacacctggacccccgacagcgcggaggacgacgtcg1560ggatcggcctccggtacgactatggcaagtactacgcgtcgaagaccttctacgaccccg1620tccttcgccggcgggtgctctgggggtgggtcggcgagaccgacagcgagcgcgcggaca1680tcctcaagggctgggcatccgtgcaggtacgtctcagggtttgaggctagcatggcttca1740atcttgctggcatcgaatcattaatgggcagatattataacttgataatctgggttggtt1800gtgtgtggtggggatggtgacacacgcgcggtaataatgtagctaagctggttaaggatg1860agtaatggggttgcgtataaacgacagctctgctaccattacttctgacacccgattgaa1920ggagacaacagtaggggtagccggtagggttcgtcgacttgccttttcttttttcctttg1980ttttgttgtggatcgtccaacacaaggaaaataggatcatccaacaaacatggaagtaat2040cccgtaaaacatttctcaaggaaccatctagctagacgagcgtggcatgatccatgcatg2100cacaaacactagataggtctctgcagctgtgatgttcctttacatataccaccgtccaaa2160ctgaatccggtctgaaaattgttcaagcagagaggccccgatcctcacacctgtacacgt2220ccctgtacgcgccgtcgtggtctcccgtgatcctgccccgtcccctccacgcggccacgc2280ctgctgcagcgctctgtacaagcgtgcaccacgtgagaatttccgtctactcgagcctag2340tagttagacgggaaaacgagaggaagcgcacggtccaagcacaacactttgcgcgggccc2400gtgacttgtctccggttggctgagggcgcgcgacagagatgtatggcgccgcggcgtgtc2460ttgtgtcttgtcttgcctatacaccgtagtcagagactgtgtcaaagccgtccaacgaca2520atgagctaggaaacgggttggagagctgggttcttgccttgcctcctgtgatgtctttgc2580cttgcatagggggcgcagtatgtagctttgcgttttacttcacgccaaaggatactgctg2640atcgtgaattattattattatatatatatcgaatatcgatttcgtcgctctcgtggggtt2700ttattttccagactcaaacttttcaaaaggcctgtgttttagttcttttcttccaattga2760gtaggcaaggcgtgtgagtgtgaccaacgcatgcatggatatcgtggtagactggtagag2820ctgtcgttaccagcgcgatgcttgtatatgtttgcagtattttcaaatgaatgtctcagc2880tagcgtacagttgaccaagtcgacgtggagggcgcacaacagacctctgacattattcac2940tttttttttaccatgccgtgcacgtgcagtcaatccccaggacggtcctcctggacacga3000agacgggcagcaacctgctccagtggccggtggtggaggtggagaacctccggatgagcg3060gcaagagcttcgacggcgtcgcgctggaccgcggatccgtcgtgcccctcgacgtcggca3120aggcgacgcaggtgacgccgcacgcagcctgctgcagcgaacgaactcgcgcgttgccgg3180cccgcggccagctgacttagtttctctggctgatcgaccgtgtgcctgcgtgcgtgcagt3240tggacatcgaggctgtgttcgaggtggacgcgtcggacgcggcgggcgtcacggaggccg3300acgtgacgttcaactgcagcaccagcgcaggcgcggcgggccggggcctgctcggcccgt3360tcggccttctcgtgctggcggacgacgacttgtccgagcagaccgccgtgtacttctacc3420tgctcaagggcacggacggcagcctccaaactttcttctgccaagacgagctcaggtatg3480tatgttatgacttatgaccatgcatgcatgcgcatttcttagctaggctgtgaagcttct3540tgttgagttgtttcacagatgcttaccgtctgctttgtttcgtatttcgactaggcatcc3600aaggcgaacgatctggttaagagagtatacgggagcttggtccctgtgctagatggggag3660aatctctcggtcagaatactggtaagtttttacagcgccagccatgcatgtgttggccag3720ccagctgctggtactttggacactcgttcttctcgcactgctcattattgcttctgatct3780ggatgcactacaaattgaaggttgaccactccatcgtggagagctttgctcaaggcggga3840ggacgtgcatcacgtcgcgagtgtaccccacacgagccatctacgactccgcccgcgtct3900tcctcttcaacaacgccacacatgctcacgtcaaagcaaaatccgtcaagatctggcagc3960tcaactccgcctacatccggccatatccggcaacgacgacttctctatgactaaattaag4020tgacggacagataggcgatattgcatacttgcatcatgaactcatttgtacaacagtgat4080tgtttaatttatttgctgccttccttatccttcttgtgaaactatatggtacacacatgt4140atcattaggtctagtagtgttgttgcaaagacacttagacaccagaggttccaggagtat4200cagagataaggtataagagggagcagggagcag4233<210>63<211>20<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>63tgttcggttccctctaccaa20<210>64<211>22<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>64caacatccatcaccttgactga22<210>65<211>24<212>dna<213>人工序列<220><223>合成的探针寡核苷酸<400>65cacagaaccgtcgcttcagcaaca24<210>66<211>18<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>66tggcggacgacgacttgt18<210>67<211>19<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>67aaagtttggaggctgccgt19<210>68<211>26<212>dna<213>人工序列<220><223>合成的探针寡核苷酸<400>68cgagcagaccgccgtgtacttctacc26<210>69<211>19<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>69cttagctggataacgccac19<210>70<211>19<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>70gaccgtaaggcttgatgaa19<210>71<211>21<212>dna<213>人工序列<220><223>合成的探针寡核苷酸<400>71cgagattctccgcgctgtaga21<210>72<211>25<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>72gtatgtttctgcttctacctttgat25<210>73<211>29<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>73ccatgttttggtcatatattagaaaagtt29<210>74<211>34<212>dna<213>人工序列<220><223>合成的探针寡核苷酸<400>74agtaatatagtatttcaagtatttttttcaaaat34<210>75<211>108<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<400>75aatgcaatggtacagtatcacgcattaggtcttctataccatattaggaagactgataag60ctagcagtgacaaaattgatttccaagctgaattcaatgggtttaaag108<210>76<211>140<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<400>76atgggcttaaggaacaaatctgaaatggtggtgtatgaagctgcacatgccatggttaac60ctgaagttcacgagtagtaatgtgctagcacccgctataagtgttctacaactattttgt120ggatctcctaaagccacact140<210>77<211>110<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<400>77cgacctcctccggtgtctagagaagaaaacttcgccgaaaaacttagtaacgttccgggt60atacaacagttaggacctttgttcaaaacttccgacgtcgttgaactcac110<210>78<211>200<212>dna<213>玉米根萤叶甲(diabroticavirgifera)<400>78agttgcactataacgaaaccggtaccacatatgtagtagttaagttgcctgatgatgatc60tccccaactctgttggtacgtgtggagccgtgttgaagttcttagtgaaagattgtgatc120catcaacgggaataccagattctgatgagggttacgatgatgaatatacactggaagaca180tcgaaataacattaggggac200<210>79<211>46<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>79ttaatacgactcactatagggagaaatgcaatggtacagtatcacg46<210>80<211>47<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>80ttaatacgactcactatagggagactttaaacccattgaattcagct47<210>81<211>46<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>81ttaatacgactcactatagggagaatgggcttaaggaacaaatctg46<210>82<211>46<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>82ttaatacgactcactatagggagaagtgtggctttaggagatccac46<210>83<211>42<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>83ttaatacgactcactatagggagacgacctcctccggtgtct42<210>84<211>43<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>84ttaatacgactcactatagggagagtgagttcaacgacgtcgg43<210>85<211>46<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>85ttaatacgactcactatagggagaagttgcactataacgaaaccgg46<210>86<211>46<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>86ttaatacgactcactatagggagagtcccctaatgttatttcgatg46<210>87<211>3106<212>dna<213>英雄美洲蝽(euschistusheros)<400>87taacctgcgagaaggagtgttctgttgacgttgacgtgggatgtgtagttgatgtttaag60gattagtagagtaatttttaatttataatacgttaaatacaaatttatatctactaaaaa120tggcaataaaacgagataagaaggaggaagaagatggtggaaacccctttcagagtcttg180ataagaccagtgttcttcaggatgccagaacttttaatgaaacaccagtagaacctcgca240agtgcaccccaatattgaccaaaattctgtatcttttgaaccaaggagagcagcttggtc300ctgcggaagcaacagaaacatttttcgctgttacgaagctatttcaatcaaataacactt360tgcttcgacgcatggtatatcttggcataaaagagctatctctgattgctcaagatgtca420tcattgttacctccagtcttacaaaagatatgactggaaaagaagatttatacagagcag480ctgcgattcgggcattatgtagtataacagatgctactatgttacaaacaattgaaagat540atatgaaacaagcaattgtcgatagaaatccagctgttgccagtgcagctcttgtaagtt600cattacacatgagtagaattgcaagtgacgtcgtcaaaagatgggttaatgaagcccaag660aagctgttaattctgacagcataatggtccaatatcatgcgctgggccttctttaccaca720ttagaaaaaatgacaggctggctgtcacaaaattagttgccaagctaactcgaatgtcat780tgaaatctccatttgctgtttgtatgctgattcgaattgcatgtaaattgttggaagaag840aaagctcaggagaatatgcagattcacctctttttgactttattgaatcgtgtttacgcc900acaaaagtgaaacagttgtttatgaagcagctgctgctcttgtaaacttacgtcacacta960cagcccggcaaatcacgcctgctgtcagtgtcctccaattgttttgttcttctcccaaac1020cggcgcttcgttttgctgctgtgagaacacttaataaggtagccatgacacacccagctg1080cagtaacatcatgcaatattgacttggagaacctcataactgattccaataggtccatcg1140ctactctggccataacaactcttctgaaaacaggagcggaatcagctgtcgataggctta1200tgaaacagatagcatcattcgtttctgaaattagtgatgaattcaaaattgtagttgtac1260aggcgattagagcattatgtttgaaatttcctcgaaaacatggaacattgatgacatttt1320tatctgctatgctaagggacgagggaggcttggagtataaggcttcgattgctgacactc1380tcatatccctgattgaaggtaaccctgaagcgaaagaatctggacttgcacatttgtgtg1440aattcatcgaggattgtgagcacacttcactagcagtgaggatattacatttgctcggta1500aagaaggacccaaaacaaaacaaccttcaaggtacattaggttcatctataatagggtaa1560tcttggaaaatgcagtggtacgagcagcagctgtttccgcattggcacaatttggagcac1620aatgtcctgatcttcttgaaaatatacttgtcctcttagcacgttgccagatggatacag1680atgatgaagtgagggacagggctacatattactacagtattttacaatttcaagatcgac1740atttgattaataattatatagttgaaccacctcaggtgtgtgtcgccagtttggaaaaag1800ccttaattgtgcatttgatggaaagtccggaagaagtatttgatatgagttccgtaccat1860tagctcctccacctctaactgatgaagttcaagctgctccagtggtaccagaaccattag1920cagctttgggccgcactgtctcaaaagaggagagtgcttcggatagacttcgagcaattc1980ctgaactctcttggatccagggcccactgttcagaagttccgatcccgtcagtcttactg2040aatctgagactgaatatcaagttagagttactaagcatgtcttcaagaaccatattgttc2100ttcagtttgactgtacaaataccatgagtgaccagctgcttgagaaagtgcgagttcagc2160tagaagtgagtgaaggatatcagatcgtagctgaggtcccatgccaaagattagcctgtt2220cggaaacatcccctacttacattgccctgcagttcccagaagcccctaatcttactgtca2280caaactttggtgctaccctgaggtttgttgtgaaagattgtgatccaatgactggcatcc2340ctaactctgatgatggttatgaagacgattatatgctggaggatgttgaagtgatgcttg2400ctgatcagatgcagcggttgactaaaagcaactttggtgctgcatgggaggaagccgaat2460cttatagtgagctagaagacacttataacttgtctggaataaacagcctagaagaggcag2520tgaggagcgttgtcagtttcatgggaatgcaacctgcggacaggagtgacagggtacaac2580ctgataaatcttcacatactgtctatcttggaggtatgttccgcggtggagttgaagtat2640tagcaagagccaaactggcaatgggcaattcccctggtgttgccatgcaacttacagtcc2700gctctccaaatccagatatttgcgagctgattatttctgtagtcgggtaaaagagatact2760tatatacatatttatgagctacagttttctcagaagttgtacagaatcaaacattgaaca2820taaagtaaatatcgtatgaaatgtattagttgactagctgcttgggaaaattttggtcac2880tcaatgttttataagtatctgtttttattaaagtgtaaacatttctgtatatgttgattt2940atgaaacatttttaagatagctaagtcttacactttgtattgagtaaatttatcagctga3000tttttatgttacccctgtggaattttattttagtttaagtaaattgaattttttttttgt3060tttttgtttaggtataacttaaaaactgaaatttttccaatggcat3106<210>88<211>876<212>prt<213>英雄美洲蝽(euschistusheros)<400>88metalailelysargasplyslysgluglugluaspglyglyasnpro151015pheglnserleuasplysthrservalleuglnaspalaargthrphe202530asngluthrprovalgluproarglyscysthrproileleuthrlys354045ileleutyrleuleuasnglnglygluglnleuglyproalagluala505560thrgluthrphephealavalthrlysleupheglnserasnasnthr65707580leuleuargargmetvaltyrleuglyilelysgluleuserleuile859095alaglnaspvalileilevalthrserserleuthrlysaspmetthr100105110glylysgluaspleutyrargalaalaalaileargalaleucysser115120125ilethraspalathrmetleuglnthrilegluargtyrmetlysgln130135140alailevalaspargasnproalavalalaseralaalaleuvalser145150155160serleuhismetserargilealaseraspvalvallysargtrpval165170175asnglualaglnglualavalasnseraspserilemetvalglntyr180185190hisalaleuglyleuleutyrhisilearglysasnaspargleuala195200205valthrlysleuvalalalysleuthrargmetserleulysserpro210215220phealavalcysmetleuileargilealacyslysleuleugluglu225230235240gluserserglyglutyralaaspserproleupheasppheileglu245250255sercysleuarghislyssergluthrvalvaltyrglualaalaala260265270alaleuvalasnleuarghisthrthralaargglnilethrproala275280285valservalleuglnleuphecysserserprolysproalaleuarg290295300phealaalavalargthrleuasnlysvalalametthrhisproala305310315320alavalthrsercysasnileaspleugluasnleuilethraspser325330335asnargserilealathrleualailethrthrleuleulysthrgly340345350alagluseralavalaspargleumetlysglnilealaserpheval355360365sergluileseraspgluphelysilevalvalvalglnalailearg370375380alaleucysleulyspheproarglyshisglythrleumetthrphe385390395400leuseralametleuargaspgluglyglyleuglutyrlysalaser405410415ilealaaspthrleuileserleuilegluglyasnproglualalys420425430gluserglyleualahisleucysglupheilegluaspcysgluhis435440445thrserleualavalargileleuhisleuleuglylysgluglypro450455460lysthrlysglnproserargtyrileargpheiletyrasnargval465470475480ileleugluasnalavalvalargalaalaalavalseralaleuala485490495glnpheglyalaglncysproaspleuleugluasnileleuvalleu500505510leualaargcysglnmetaspthraspaspgluvalargaspargala515520525thrtyrtyrtyrserileleuglnpheglnasparghisleuileasn530535540asntyrilevalgluproproglnvalcysvalalaserleuglulys545550555560alaleuilevalhisleumetgluserproglugluvalpheaspmet565570575serservalproleualaproproproleuthraspgluvalglnala580585590alaprovalvalprogluproleualaalaleuglyargthrvalser595600605lysglugluseralaseraspargleuargalaileprogluleuser610615620trpileglnglyproleupheargserseraspprovalserleuthr625630635640glusergluthrglutyrglnvalargvalthrlyshisvalphelys645650655asnhisilevalleuglnpheaspcysthrasnthrmetseraspgln660665670leuleuglulysvalargvalglnleugluvalsergluglytyrgln675680685ilevalalagluvalprocysglnargleualacyssergluthrser690695700prothrtyrilealaleuglnpheproglualaproasnleuthrval705710715720thrasnpheglyalathrleuargphevalvallysaspcysasppro725730735metthrglyileproasnseraspaspglytyrgluaspasptyrmet740745750leugluaspvalgluvalmetleualaaspglnmetglnargleuthr755760765lysserasnpheglyalaalatrpgluglualaglusertyrserglu770775780leugluaspthrtyrasnleuserglyileasnserleuglugluala785790795800valargservalvalserphemetglymetglnproalaaspargser805810815aspargvalglnproasplysserserhisthrvaltyrleuglygly820825830metpheargglyglyvalgluvalleualaargalalysleualamet835840845glyasnserproglyvalalametglnleuthrvalargserproasn850855860proaspilecysgluleuileileservalvalgly865870875<210>89<211>495<212>dna<213>英雄美洲蝽(euschistusheros)<400>89gtttgaaatttcctcgaaaacatggaacattgatgacatttttatctgctatgctaaggg60acgagggaggcttggagtataaggcttcgattgctgacactctcatatccctgattgaag120gtaaccctgaagcgaaagaatctggacttgcacatttgtgtgaattcatcgaggattgtg180agcacacttcactagcagtgaggatattacatttgctcggtaaagaaggacccaaaacaa240aacaaccttcaaggtacattaggttcatctataatagggtaatcttggaaaatgcagtgg300tacgagcagcagctgtttccgcattggcacaatttggagcacaatgtcctgatcttcttg360aaaatatacttgtcctcttagcacgttgccagatggatacagatgatgaagtgagggaca420gggctacatattactacagtattttacaatttcaagatcgacatttgattaataattata480tagttgaaccacctc495<210>90<211>49<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>90ttaatacgactcactatagggagagtttgaaatttcctcgaaaacatgg49<210>91<211>58<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>91ttaatacgactcactatagggagagaggtggttcaactatataattattaatcaaatg58<210>92<211>301<212>dna<213>人工序列<220><223>合成的人工序列<400>92catctggagcacttctctttcatgggaagattccttacgttgtggagatggaagggaatg60ttgatggccacacctttagcatacgtgggaaaggctacggagatgcctcagtgggaaagg120ttgatgcacagttcatctgcacaactggtgatgttcctgtgccttggagcacacttgtca180ccactctcacctatggagcacagtgctttgccaagtatggtccagagttgaaggacttct240acaagtcctgtatgccagatggctatgtgcaagagcgcacaatcacctttgaaggagatg300g301<210>93<211>47<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>93ttaatacgactcactatagggagagcatctggagcacttctctttca47<210>94<211>46<212>dna<213>人工序列<220><223>合成的引物寡核苷酸<400>94ttaatacgactcactatagggagaccatctccttcaaaggtgattg46<210>95<211>410<212>dna<213>人工序列<220><223>合成的人工序列<400>95atgtcatctggagcacttctctttcatgggaagattccttacgttgtggagatggaaggg60aatgttgatggccacacctttagcatacgtgggaaaggctacggagatgcctcagtggga120aagtccggcaacatgtttgacgtttgtttgacgttgtaagtctgatttttgactcttctt180ttttctccgtcacaatttctacttccaactaaaatgctaagaacatggttataacttttt240ttttataacttaatatgtgatttggacccagcagatagagctcattactttcccactgag300gcatctccgtagcctttcccacgtatgctaaaggtgtggccatcaacattcccttccatc360tccacaacgtaaggaatcttcccatgaaagagaagtgctccagatgacat410当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 诱导果实抗性控制病害的方法和所用制剂与流程
- 水稻白叶枯抗性基因Xa27的分子标记引物及其应用的制作方法与工艺
- 赋予对鞘翅目有害生物的抗性的核酸分子的制造方法与工艺
- 避免抗微生物剂抗性的抗腹泻制剂的制造方法与工艺
- 一种选育小麦条锈病持久抗性材料的方法与流程
- 一种产生PVY抗性的NteIF4E‑1基因突变体及其应用的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体GAMMA亚单位核酸分子的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体BETA亚单位核酸分子的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体ALPHA亚单位核酸分子的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体DELTA亚单位核酸分子的制造方法与工艺
- 一种选育小麦条锈病持久抗性材料的方法与流程
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体GAMMA亚单位核酸分子的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体BETA亚单位核酸分子的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体ALPHA亚单位核酸分子的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体DELTA亚单位核酸分子的制造方法与工艺
- 一个影响猪生长和PRRSV抗性的SLA‑DRB1启动子区分子标记及其引物和应用的制造方法与工艺
- 一个影响猪生长和PRRSV抗性的分子标记、检测方法及其应用与制造工艺
- 赋予鞘翅目害虫抗性的DRE4核酸分子的制造方法与工艺
- 一种提高茶树抗性且肥效高的茶树肥料及其制备方法与制造工艺
- 抗穿透的复合件的制作方法