一种耐酸性玉米赤霉烯酮解毒酶及其编码基因与应用的制作方法
本发明涉及酶工程领域,具体地说,涉及一种耐酸性玉米赤霉烯酮解毒酶及其编码基因与应用。
背景技术:
玉米赤霉烯酮(zearalenone,zen),又称f-2毒素,为2,4-二羟基苯甲酸内酯类化合物,主要是由禾谷镰刀菌次级代谢产生的一种真菌毒素。zen具有雌激素活性,对生殖系统有毒害作用,摄入过量zen导致动物外阴和乳腺肿大甚至阴道及直肠脱垂;影响哺乳动物乳房发育,导致哺乳推迟、假孕,甚至不孕或导致流产,胎儿畸形和死胎。zen可以引发肿瘤,不仅通过增加细胞色素cyp1a1酶活性来刺激人类乳腺癌mcf-7细胞的生长,同时也是食管癌发病率增加的病因之一。zen对人和动物肾脏、肝脏和血液也具有明显的毒害作用。
zen的脱毒方法可以分为物理、化学和生物三大类,其中物理法有高压加热、辐照和吸附剂吸附等,化学法有臭氧、双氧水和碳酸钠处理等,生物法有微生物降解、微生物吸附和酶降解等。但传统的物理和化学脱毒方法会破坏营养物质、引入化学物质造成二次污染等问题,其应用受到一定的局限。生物降解法具有反应条件温和、专一性高、无残留、环境友好等优点,具有广阔的应用前景。
2002年,kakeya分离到一株粉红粘帚霉(clonostachysroseaifo7063)菌株,能够破坏zen的内酯环,将zen完全转化为无任何雌激素活性的代谢产物,并确定由单拷贝基因zhd101所编码的碱性乳糖水解酶是降解zen的功能性蛋白质(kakeya等,一株粉红粘帚霉将玉米赤霉烯酮转化无雌激素活性的化合物.生物科学、生物技术和生物化学杂志,2002)。utermark和karlovsky的插入失活实验证明了zhd101编码酶对zen的降解作用(utermak和karlovsky,玉米赤霉烯酮内酯水解酶在保护粉红粘帚霉免受玉米赤霉烯酮杀菌作用中的角色,应用与环境微生物学,2007),tomok等把zhd101转化到玉米中,发现玉米也表现出对zen的降解能力(tomok等,通过降解基因来减少玉米籽粒中真菌毒素玉米赤霉烯酮的污染,应用与环境微生物,2007)。
中国发明专利zl201110082679.x同样从粉红粘帚霉(clonostachysrosea)中克隆获得玉米赤霉烯酮解毒酶zlhy-6(与2002年kakeya获得的zhd101编码酶只有4个氨基酸差异,蛋白同源性98%)及其编码基因。然而,该酶的最适ph为8.5,ph5.0和6.5时该酶的降解能力分别仅为ph8.5时的15%和25%,由此可见酸性条件严重抑制玉米赤霉烯酮解毒酶的活性;该解毒酶主要是作为添加剂混到猪、鸡饲料中,在猪、鸡体内降解玉米赤霉烯酮,而猪、鸡肠道ph值在5.5~6.5之间,因此亟需对玉米赤霉烯酮解毒酶进行耐酸改造,使其活性最适ph由8.5变为偏酸性。不仅如此,上述技术方案还将克隆获得的玉米赤霉烯酮解毒酶编码基因连接到质粒ppic9k并转化毕赤酵母,获得能够表达玉米赤霉烯酮解毒酶的重组毕赤酵母菌株,但是玉米赤霉烯酮解毒酶的活性(表达量)很低,1ml重组酵母发酵上清液在2h时只可以降解18.8μg的zen。
技术实现要素:
为了解决现有技术中存在的问题,本发明的第一个目的是实现玉米赤霉烯酮解毒酶的耐酸性,扩大其解毒活性ph范围,本发明的第二个目的是实现该耐酸性玉米赤霉烯酮解毒酶的高效表达。
为了实现本发明目的,本发明的技术方案如下:
本发明首先提供一种耐酸性玉米赤霉烯酮解毒酶zde-3,是如下(1)或(2)的蛋白质:
(1)由seqidno.1所示的氨基酸序列组成的蛋白质;
(2)由seqidno.1所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加,且在ph为5.0~6.5时仍然具有降解玉米赤霉烯酮活性的由seqidno.1衍生的蛋白质。
进一步地,编码前述耐酸性玉米赤霉烯酮解毒酶的基因也属于本发明的保护范围。
为了提高玉米赤霉烯酮解毒酶在酵母菌株中的表达量,本发明在所述耐酸性玉米赤霉烯酮解毒酶的基础上,按照酵母密码子的偏好性进行优化,在795bp中改变了181bp,提供了一种可显著增加表达量的编码基因zde-3,其含有如seqidno.2所示的核苷酸序列。当其为seqidno.2所示的核苷酸序列时,其与原序列的差异性为22.8%。
进一步地,含有所述编码基因及优化后的编码基因的载体也属于本发明的保护范围。
当所述载体含有上述优选编码基因时,所述载体的构建方法包括如下步骤:
(1)用限制性内切酶ecori对seqidno.3所示的核苷酸序列α-zde-3和质粒pao815进行酶切反应;
其中,seqidno.3中274bp~1068bp是zde-3基因,8bp~273bp是α信号肽序列,1bp~7bp及1069bp~1074bp是ecori酶切位点;
其中,酶切体系优选为:目的片段或质粒15μl,10×ecorⅰbuffer2μl,ecori酶1μl,ddh2o2μl,酶切条件37℃反应4h;
(2)将酶切后的质粒pao815去磷酸化;
其中,去磷酸化反应体系优选为:酶切后的质粒pao81515μl,antarcticphosphatase1μl,10×reactionbuffer2μl,ddh2o2μl,反应条件37℃水浴1h,在65℃加热5min,使磷酸化失活,处理好的载体备用;
(3)将酶切回收得到的基因片段和去磷酸化的质粒pao815进行酶连接反应,得到重组表达载体。
其中,酶连接反应体系优选为:载体pao8154μl,基因片段3μl,5×ligationbuffer2μl,t4ligase酶1μl,反应条件22℃链接2h,连接产物用于转化;
(4)将重组表达载体转化大肠杆菌top10,经氨苄青霉素抗性筛选,挑取菌落培养,分别进行pcr鉴定和重组表达载体的酶切鉴定,对pao815-α-zde-3进行测序,证明融合连接到质粒pao815的dna序列与序列表seqidno.3相同,获得含有玉米赤霉烯酮解毒酶基因序列的重组表达载体pao815-α-zde-3;
(5)单拷贝表达框的扩增:以重组表达载体pao815-α-zde-3为模板,用seqidno.4~5所示的引物进行pcr扩增,扩增出完整表达框5’aox-α-zde-3-3’aox-tt3’;
seqidno.4~5所示的引物分别为上游引物p1和下游引物p2:
上游引物p1:5’-ggaagatctaacatccaaagacg-3’,引入bglⅱ酶切位点;下游p2:5’-taggatccgcacaaacgaac-3’,引入bamhⅰ酶切位点。
其中,步骤(5)中的pcr反应程序优选为:95℃预变性5min;95℃30s,56℃30s,72℃60s,共35个循环;72℃延伸7min。
(6)单拷贝表达框的ta克隆:将pcr扩增得到的完整表达框连接到pgm-t载体,并转化大肠杆菌top10,经氨苄青霉素抗性筛选获得重组菌株,挑取菌落37℃震荡培养12h,提取质粒,用bamhi单酶切,琼脂糖凝胶电泳检查片段大小,确定含有完整表达框;
(7)多拷贝串联表达框表达载体构建:将步骤(6)中获得的完整表达框经bglⅱ和bamhⅰ双酶切后,试剂盒回收,得到的表达框与经bamhⅰ酶切处理并去磷酸化的重组质粒pao815-α-zde-3连接,转化top10,提取质粒进行酶切鉴定筛选2拷贝表达载体pao815-(α-zde-3)2。得到的2拷贝表达载体再与表达框进行酶切、酶连、转化,如此反复筛选4、6拷贝数的表达载体pao815-(α-zde-3)4和pao815-(α-zde-3)6。
本发明还进一步提供一种高效表达所述玉米赤霉烯酮解毒酶的方法,是将前述玉米赤霉烯酮解毒酶的编码基因连接到pao815载体中,构建多拷贝(2-6拷贝)重组表达载体,接着将该重组表达载体导入宿主细胞,高效表达得到玉米赤霉烯酮解毒酶。
其中所述宿主可为酵母菌、大肠杆菌、哺乳动物细胞、昆虫细胞或枯草杆菌等,优选为酵母菌,所述酵母菌优选为毕赤酵母(pichiapastoris)。
用于构建所述重组大肠杆菌及重组酵母表达载体的出发载体可为在上述宿主中表达外源基因的表达载体,如可在大肠杆菌及巴氏德毕赤酵母(pichiapastoris)中表达的ppic9k、ppic9、ppic3.5k、peb、pao815等。
应当理解的是,含有所述载体的工程菌或宿主细胞也属于本发明的保护范围。
最后,本发明提供了所述耐酸性玉米赤霉烯酮解毒酶及其编码基因在降解谷物或饲料中zen的应用。
本发明涉及到的操作如无特殊说明均为本领域常规操作。
在符合本领域常识的基础上,上述各优选条件,可以相互组合,得到具体实施方式。
本发明的有益效果在于:
本发明解决了现有技术中玉米赤霉烯酮解毒酶不耐酸的问题。本发明通过对玉米赤霉烯酮解毒酶的氨基酸序列进行优化,改变了解毒酶的8个氨基酸残基(与zlhy-6解毒酶有8个氨基酸差异),使重组表达玉米赤霉烯酮解毒酶最适反应ph由8.5降低到6.5,与猪、鸡肠道ph非常吻合,当ph低至5.0时,解毒酶酶活仍保留40%以上,大大提高该玉米赤霉烯酮解毒酶作为饲料添加剂在实际应用中的解毒效率。氨基酸改造后1ml酵母发酵上清液处理24h,可以将2g玉米碴中的zen含量由2000μg/kg降低到489.80μg/kg,降解率为75.51%,表明重组表达的玉米赤霉烯酮降解酶对玉米碴等谷物和饲料中zen具有非常好的降解效果。
不仅如此,本发明还通过对玉米赤霉烯酮解毒酶基因进行密码子优化,并将该基因在重组酵母基因组中的拷贝数增加到4,大大提高了玉米赤霉烯酮解毒酶的表达量,使同体积重组酵母发酵上清液的酶活是专利zl201110082679.x中酶活的72倍。
附图说明
图1为本发明实施例3中不同拷贝重组表达载体pao815-α-zde-3、pao815-(α-zde-3)2、pao815-(α-zde-3)4、pao815-(α-zde-3)6、转化子表达产物的sds-page图。
图2为本发明实施例4中重组玉米赤霉烯酮解毒酶的降解活性测定。
图3为本发明实施例5中不同ph对玉米赤霉烯酮解毒酶降解活性的影响。
图4为本发明实施例6中玉米赤霉烯酮解毒酶对玉米碴中zen的降解。
具体实施方式
下面将结合实施例对本发明的优选实施方式进行详细说明。需要理解的是以下实施例的给出仅是为了起到说明的目的,并不是用于对本发明的范围进行限制。本领域的技术人员在不背离本发明的宗旨和精神的情况下,可以对本发明进行各种修改和替换。
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
实施例1玉米赤霉烯酮解毒酶的耐酸改造和密码子偏好性优化
1、对玉米赤霉烯酮解毒酶进行耐酸改造:在zlhy-6解毒酶的基础上(zl201110082679.x,seqidno.6),将52位精氨酸r、66位赖氨酸k、118位精氨酸r、148位赖氨酸k、175位精氨酸r、185位精氨酸r、189位氨基酸r和204位精氨酸替换为谷氨酸。得到玉米赤霉烯酮解毒酶(zde-3),具体氨基酸序列见seqidno.1。
2、玉米赤霉烯酮解毒酶的编码基因,在氨基酸残基改造的基础上,按照酵母密码子的偏好性进行优化,得到玉米赤霉烯酮解毒酶基因(zde-3),具体核苷酸序列见seqidno.2。
实施例2重组表达载体的构建
1、在实施例1中设计的玉米赤霉烯酮解毒酶基因(zde-3)前端添加α信号肽编码序列,即α-zde-3,并在序列两端引入ecori(gaattc)酶切位点。全序列核苷酸序列见seqidno.3。该核苷酸序列通过化学方法合成获得。
2、含有玉米赤霉烯酮解毒酶编码基因序列的重组表达载体的构建
(1)用限制性内切酶ecori对合成的α-zde-3核苷酸序列和质粒pao815(美国invitrogen公司)进行酶切反应,酶切体系20μl:目的片段或质粒15μl,10×ecorⅰbuffer2μl,ecori酶1μl,ddh2o2μl,酶切条件37℃反应4h。
(2)载体pao815去磷酸化反应体系:酶切后载体15μl,antarcticphosphatase1μl,10×reactionbuffer2μl,ddh2o2μl,反应条件37℃水浴1h,在65℃加热5min,使磷酸化失活,处理好的载体备用。
(3)将酶切回收得到的基因片段和载体进行酶连接反应,反应体系10μl:载体pao8154μl,基因片段3μl,5×ligationbuffer2μl,t4ligase酶1μl,反应条件22℃链接2h,连接产物用于转化。
(4)连接产物转化大肠杆菌top10,经氨苄青霉素抗性筛选,挑取菌落37℃震荡培养12h,分别进行pcr鉴定和重组表达载体的酶切鉴定。获得的重组表达载体命名为pao815-α-zde-3。对pao815-α-zde-3进行测序,证明融合连接到质粒pao815的dna序列与序列表seqidno.3相同,构建含有玉米赤霉烯酮解毒酶基因序列的重组表达载体pao815-α-zde-3正确。
(5)单拷贝表达框的扩增。设计上游引物p1:5’-ggaagatctaacatccaaagacg-3’,引入bglⅱ酶切位点;下游p2:5’-taggatccgcacaaacgaac-3’,引入bamhⅰ酶切位点;以重组表达载体pao815-α-zde-3为模板,用引物p1/p2pcr扩增完整表达框5’aox-α-zde-3-3’aox-tt3’。pcr反应程序:95℃预变性5min,(95℃30s,56℃30s,72℃60s)共35个循环,72℃延伸7min。
(6)单拷贝表达框的ta克隆。将pcr扩增得到的完整表达框连接到pgm-t载体,并转化大肠杆菌top10,经氨苄青霉素抗性筛选获得重组菌株,挑取菌落37℃震荡培养12h,提取质粒,用bamhi单酶切,琼脂糖凝胶电泳检查片段大小,确定含有完整表达框。
(7)多拷贝串联表达框表达载体构建。步骤(6)中获得的完整表达框经bglⅱ、bamhⅰ双酶切后,试剂盒回收。得到的表达框与经bamhⅰ酶切处理并去磷酸化的重组表达载体pao815-α-zde-3连接,转化top10,提取质粒进行酶切鉴定筛选2拷贝表达载体pao815-(α-zde-3)2。得到的2拷贝表达载体再与表达框进行酶切、酶连、转化,如此反复筛选4、6拷贝数的表达载体pao815-(α-zde-3)4和pao815-(α-zde-3)6。
实施例3玉米赤霉烯酮解毒酶的高效表达
将重组表达载体pao815-α-zde-3、pao815-(α-zde-3)2、pao815-(α-zde-3)4和pao815-(α-zde-3)6用sali使之线性化后,采用电击方式,将线性化载体的导入毕赤酵母gs115中,经md选择性培养基(1.34%ynb无氨基酵母氮源(yeastnitrogenbasewithoutaminoacids)、2%葡萄糖(d-glucose)、4×10-5生物素(biotin)、2%琼脂(agar))筛选高表达菌株。
挑取选择性培养基上长出的单菌落接种于5mlbmgy(1%甘油,1%酵母提取物(yeastextract),2%蛋白胨(peptone),1.34%ynb,4×10-5生物素,100mm磷酸钾ph6.0)培养基中,30℃、250rpm培养36h,5000rpm室温离心5min收集菌体。菌体用2mlbmmy(0.5%甲醇,1%酵母提取物(yeastextract),2%蛋白胨(peptone),1.34%ynb,4×10-5生物素,100mm磷酸钾ph6.0)培养基重悬,30℃、250rpm诱导培养3d,期间每隔24h补加甲醇至0.5%,培养结束后,12000rpm离心5min收集上清液进行sds-page电泳筛选。
为比较不同拷贝转化子的蛋白表达量,用相同菌密度的转化子在同体积的bmmy培养基中诱导培养,sds-page电泳比较pao815-α-zde-3、pao815-(α-zde-3)2、pao815-(α-zde-3)4、pao815-(α-zde-3)6转化子上清液的蛋白量。结果如图1所示(泳道1、2、3、4分别为1、2、4、6拷贝的转化子上清液蛋白条带),表明重组菌株在甲醇诱导下表达的蛋白质的分子量约30kda,回收后经ms-ms鉴定分子量为28.8kda,与理论蛋白分子量29.3kda大小相当;通过比较蛋白条带的亮度,发现4拷贝转化子的玉米赤霉烯酮解毒酶表达量最高。
实施例4玉米赤霉烯酮解毒酶的活性检测(液体中)
重组4拷贝毕赤酵母菌株(解毒酶)和野生型毕赤酵母菌株(对照组、空质粒整合的毕赤酵母菌株)接入bmgy培养基中继续培养至菌液od600=2-3,室温离心收集菌体,重悬于bmmy培养基,甲醇诱导培养3d,期间每隔24h补加甲醇至0.5%,培养结束后,12000rpm离心5min收集上清液进行zen降解活性测定。
反应体系1000μl:上清液40μl、0.5mg/mlzen40μl、0.05mtris-hcl(ph7.5)920μl,zen终浓度为20μg/ml。反应体系混匀后37℃反应30、60、120min加入等体积甲醇终止反应,利用高效液相色谱(hplc)检测zen残留量,检测方法参考gb/t23504-2009方法进行。hplc检测条件:waters2695高效液相色谱仪,配备waters2475荧光检测器,激发光exc=360nm、发射光em=440nm;agilenttc-c18色谱柱(4.6mm×150mm,5μm),柱温30℃;流动相(水:甲醇:乙腈,4:1:1),流速0.5ml/min,进样量20μl。
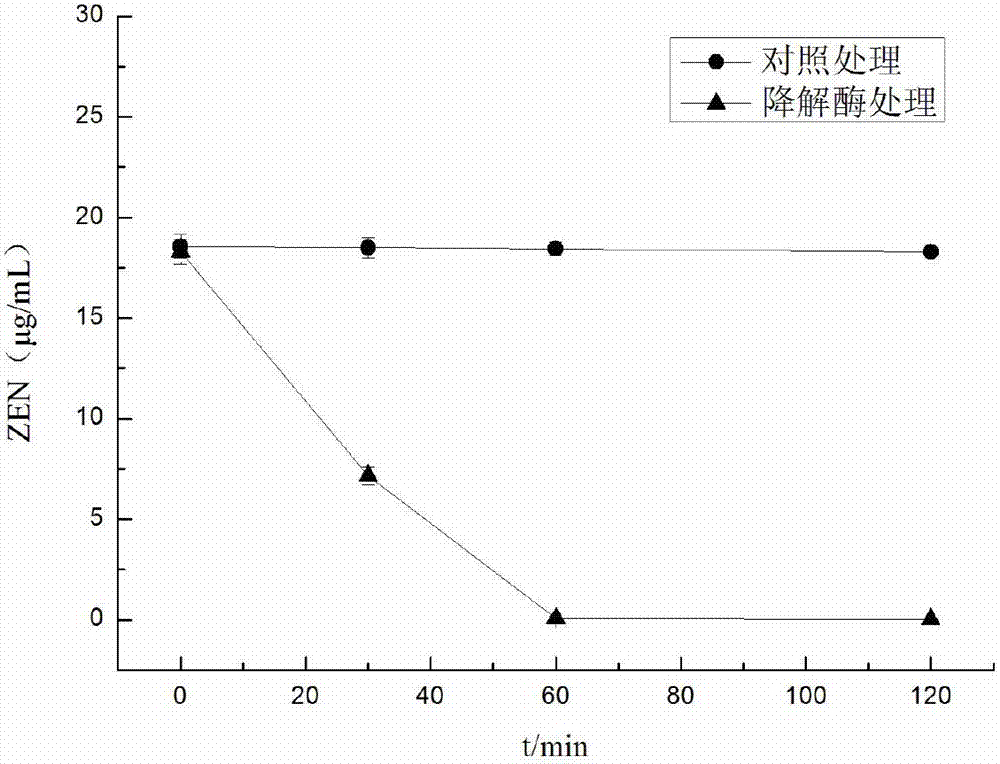
通过hplc检测得知,重组酵母上清液(酶液)处理zen60分钟时,检测不到zen的残留,如图2所示,而对照组控制质粒的发酵上清液中zen基本上没有降解。可以表明,该优化后的zde-3基因重组表达出的解毒酶具有高效、快速降解zen的活性。
对重组解毒酶活力进行定义:在上述条件下,每毫升发酵液每分钟降解1μgzen为1个活力单位(u)。
本发明:40μl上清液在30min降解了13.8μg的zen,计算该上清液的酶活力为:11.5u。
专利zl201110082679.x:980μl上清液在2h降解了18.8μg的zen,该上清液的酶活力为:0.16u。
经过对玉米赤霉烯酮解毒酶基因的密码子进行优化,并将该基因在重组酵母基因组中的拷贝数增加到4,大大提高了玉米赤霉烯酮解毒酶的表达量,最终使同体积重组酵母发酵上清液的zen解毒酶活性达到专利zl201110082679.x中降解酶酶活的72倍。
实施例5玉米赤霉烯酮解毒酶的最适ph
配制0.05m的tris-hcl缓冲液,ph分别设5.0、5.5、6.0、6.5、7.0、7.5、8.0、8.5共8个水平;反应体系1000μl:40μl酶上清液+40μl0.5mg/mlzen+920μl设定的tris-hcl缓冲液;空白对照为40μl沸水浴5分钟的上清液+40μl0.5mg/mlzen+920μl0.05mtris-hcl缓冲液。反应体系混匀后37℃水浴30min,再加入等体积(1000μl)甲醇终止反应。样品过0.22μm微孔滤膜后用hplc检测zen残留量,计算zen的降解率。
如图3所示,氨基酸改造后重组玉米赤霉烯酮解毒酶的最适ph由8.5降低到6.5,与猪、鸡肠道ph非常相近,大大提高该重组玉米赤霉烯酮解毒酶作为饲料添加剂在实际应用中的降解效率;而且在ph5.0-8.0范围内玉米赤霉烯酮解毒酶保持较高的降解活性,zen降解率均在40%以上,扩大了该解毒酶的ph适用范围。
实施例6玉米赤霉烯酮解毒酶对玉米中zen的降解
收集污染zen的玉米碴,检测zen含量为2450μg/kg。用缓冲液将酵母发酵上清液(酶液)分别稀释2、5、10倍。按照发酵液与玉米碴(v:w)1:1的比例混合均匀(即酶液添加量分别为0.1、0.2、0.5ml/g),37℃反应0、3、6、12、24h,不加酶液作为对照。按照gb/t23504~2009提取玉米碴中的zen,利用hplc检测zen的残留量,计算玉米碴中zen的降解率。
如图4所示,随着降解时间的延长,样品中zen含量逐渐降低,反应前6h降解速率较快,6h以后降解速率趋于平缓。酶液添加量越高降解速率越快,降解率也越高,当酶液的添加量为0.1ml/g玉米时,反应6hzen的降解率仅为15.32%,反应24h后,zen降解率为45.18%。而当酶液添加量为0.5ml/g玉米时,处理6h玉米碴中zen含量由2450μg/kg降低到712.5μg/kg,降解率为70.9%;处理24h,玉米碴中zen含量降低到483.5μg/kg,降解率为80.3%。可以表明,重组表达的玉米赤霉烯酮降解酶对玉米碴等谷物和饲料中zen具有较好的降解效果。
虽然,上文中已经用一般性说明及具体实施方案对本发明作了详尽的描述,但在本发明基础上,可以对之作一些修改或改进,这对本领域技术人员而言是显而易见的。因此,在不偏离本发明精神的基础上所做的这些修改或改进,均属于本发明要求保护的范围。
序列表
<110>中国农业科学院农产品加工研究所
<120>一种耐酸性玉米赤霉烯酮解毒酶及其编码基因与应用
<130>khp171111989.8tq
<160>6
<170>patentinversion3.5
<210>1
<211>264
<212>prt
<213>zde-3
<400>1
metargthrargserthrileserthrproasnglyilethrtrptyr
151015
tyrgluglngluglythrglyproaspvalvalleuvalproaspgly
202530
leuglyglucysglnmetpheaspargservalserglnilealaala
354045
glnglyphegluvalthrthrpheaspmetproglymetserargser
505560
valglualaproprogluthrtyrthrgluvalthralaglnlysleu
65707580
alasertyrvalileservalleuaspalaleuaspilelyshisala
859095
thrvaltrpglycysserserglyalaserthrvalvalalaleuleu
100105110
leuglytyrproaspgluileargasnalametcyshisgluleupro
115120125
thrlysleuleuasphisleuserasnthralavalleugluaspglu
130135140
gluilesergluileleualaasnvalmetleuasnaspvalsergly
145150155160
glyserglualatrpglnalametglyaspgluvalhisalagluleu
165170175
hislysasntyrprovaltrpalagluglytyrprogluthrilepro
180185190
proseralaprovallysaspleuglualaleugluglylysproleu
195200205
asptrpthrvalglyalaalathrprothrgluserphepheaspasn
210215220
ilevalthralathrlysalaglyvalasnileglyleuleuprogly
225230235240
methispheprotyrvalserhisproaspvalphealalystyrval
245250255
valgluphethrglnlystyrleu
260
<210>2
<211>795
<212>dna
<213>zde-3
<400>2
atgagaactcgttccactatttccactcctaacggtatcacctggtactacgagcaagag60
ggtactggtccagacgtcgtacttgttcctgacggtttaggtgaatgtcagatgttcgac120
agatccgtctctcaaatcgcagctcagggtttcgaagtcactaccttcgacatgccaggt180
atgtccagatccgtcgaggctccacctgagacttacactgaagtaaccgctcaaaagctg240
gcctcttacgtcatttccgttctggacgctttggacattaagcacgctactgtctggggt300
tgctcttctggtgctagtactgttgtcgctttattgctcggctaccctgacgaaatcaga360
aacgctatgtgtcacgagttgccaaccaagcttttggaccacctttctaataccgccgtt420
ttggaagatgaggaaatctctgaaatcttggctaacgttatgcttaacgacgtctccgga480
ggttctgaagcttggcaagccatgggtgacgaagtccacgccgaattgcacaagaattac540
ccagtctgggccgaaggatacccagaaactatcccaccctccgctccagttaaggacttg600
gaagccttggaaggtaagccattagactggaccgttggtgctgctactccaaccgaatcc660
ttcttcgacaacatcgttactgccaccaaggctggtgttaacatcggtctgttgccaggt720
atgcactttccatacgtctcccacccagacgtcttcgccaaatacgttgtcgagttcacg780
cagaagtacttgtaa795
<210>3
<211>1074
<212>dna
<213>α-zde-3
<400>3
caattgatgagatttccttcaatttttactgcagttttattcgcagcatcctccgcatta60
gctgctccagtcaacactacaacagaagatgaaacggcacaaattccggctgaagctgtc120
atcggttactcagatttagaaggggatttcgatgttgctgttttgccattttccaacagc180
acaaataacgggttattgtttataaatactactattgccagcattgctgctaaagaagaa240
ggggtatctctcgagaaaagagaggctgaagctatgagaactcgttccactatttccact300
cctaacggtatcacctggtactacgagcaagagggtactggtccagacgtcgtacttgtt360
cctgacggtttaggtgaatgtcagatgttcgacagatccgtctctcaaatcgcagctcag420
ggtttcgaagtcactaccttcgacatgccaggtatgtccagatccgtcgaggctccacct480
gagacttacactgaagtaaccgctcaaaagctggcctcttacgtcatttccgttctggac540
gctttggacattaagcacgctactgtctggggttgctcttctggtgctagtactgttgtc600
gctttattgctcggctaccctgacgaaatcagaaacgctatgtgtcacgagttgccaacc660
aagcttttggaccacctttctaataccgccgttttggaagatgaggaaatctctgaaatc720
ttggctaacgttatgcttaacgacgtctccggaggttctgaagcttggcaagccatgggt780
gacgaagtccacgccgaattgcacaagaattacccagtctgggccgaaggatacccagaa840
actatcccaccctccgctccagttaaggacttggaagccttggaaggtaagccattagac900
tggaccgttggtgctgctactccaaccgaatccttcttcgacaacatcgttactgccacc960
aaggctggtgttaacatcggtctgttgccaggtatgcactttccatacgtctcccaccca1020
gacgtcttcgccaaatacgttgtcgagttcacgcagaagtacttgtaagaattc1074
<210>4
<211>23
<212>dna
<213>人工序列
<400>4
ggaagatctaacatccaaagacg23
<210>5
<211>20
<212>dna
<213>人工序列
<400>5
taggatccgcacaaacgaac20
<210>6
<211>264
<212>prt
<213>zhly-6
<400>6
metargthrargserthrileserthrproasnglyilethrtrptyr
151015
tyrgluglngluglythrglyproaspvalvalleuvalproaspgly
202530
leuglyglucysglnmetpheaspargservalserglnilealaala
354045
glnglypheargvalthrthrpheaspmetproglymetserargser
505560
vallysalaproprogluthrtyrthrgluvalthralaglnlysleu
65707580
alasertyrvalileservalleuaspalaleuaspilelyshisala
859095
thrvaltrpglycysserserglyalaserthrvalvalalaleuleu
100105110
leuglytyrproaspargileargasnalametcyshisgluleupro
115120125
thrlysleuleuasphisleuserasnthralavalleugluaspglu
130135140
gluileserlysileleualaasnvalmetleuasnaspvalsergly
145150155160
glyserglualatrpglnalametglyaspgluvalhisalaargleu
165170175
hislysasntyrprovaltrpalaargglytyrproargthrilepro
180185190
proseralaprovallysaspleuglualaleuargglylysproleu
195200205
asptrpthrvalglyalaalathrprothrgluserphepheaspasn
210215220
ilevalthralathrlysalaglyvalasnileglyleuleuprogly
225230235240
methispheprotyrvalserhisproaspvalphealalystyrval
245250255
valgluphethrglnlystyrleu
260
- 还没有人留言评论。精彩留言会获得点赞!
- 一种能降低青贮中玉米赤霉烯酮和呕吐毒素的微生物制剂的制作方法与工艺
- 一种乳制品中玉米赤霉醇类的可视化单分子检测方法与流程
- 一种快速检测植物油中玉米赤霉烯酮毒素的方法与流程
- 一种玉米赤霉烯酮单克隆抗体及其应用的制作方法与工艺
- 一种赤霉烯酮分解酶及其编码基因和用途的制造方法与工艺
- 检测黄曲霉毒素B1、玉米赤霉烯酮、呕吐毒素的检测卡的制造方法与工艺
- 用于以酶促方式使玉米赤霉烯酮解毒的多肽,以及分离的多核苷酸,以及相关的添加剂、用途和方法与流程
- 一种耐酸性玉米赤霉烯酮解毒酶及其编码基因与应用的制造方法与工艺
- 水飞蓟素在缓解动物玉米赤霉烯酮生殖毒性中的应用的制造方法与工艺
- 检测黄曲霉毒素B1、玉米赤霉烯酮、呕吐毒素的三联检测组件的制造方法与工艺