双功能谷胱甘肽合成酶突变体、核苷酸序列及其制备方法和应用与流程
本发明涉及双功能谷胱甘肽合成酶领域,特别地,涉及一种双功能谷胱甘肽合成酶突变体。此外,本发明还涉及一种包括上述双功能谷胱甘肽合成酶突变体的核苷酸序列及其制备方法和应用。
背景技术:
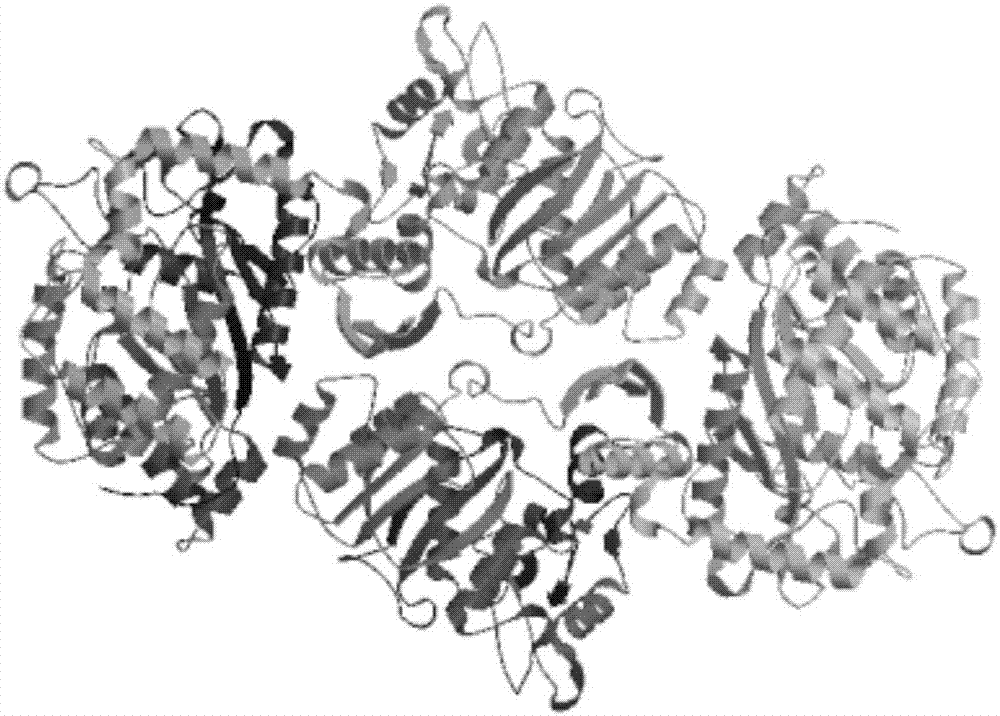
:谷胱甘肽(glutathione,简称gsh)是由谷氨酸、半胱氨酸和甘氨酸经肽键缩合而成的,具有多种重要生理功能的活性三肽。在自然环境中,gsh存在2种形式,分别是还原型和氧化型,其中还原型gsh是细胞内主要还原物质,对于维持生物体内适宜的氧化还原环境起着至关重要的作用,能保护细胞免受氧化型、毒害性化合物和辐射的伤害。同时,gsh分子小,很容易被机体吸收,而且不容易被消化道酶破坏,gsh还是细胞内某些酶的辅因子,参与细胞内的代谢循环。由于其重要的生理功能,gsh在医学、食品、保健品及化妆品方面都有着广泛用途。gsh的生产方法有很多,工业生产主要有溶剂萃取法、化学合成法、酶法、发酵法。目前,在所有生产gsh的方法中,酶法和发酵法应用的最为广泛,他们同属于生物合成法。但是,发酵法相比酶法具有明显的缺点,比如:发酵生产周期长(通常为2-5天),发酵液组分复杂后期提取困难,gsh产量低(4-8g/l),发酵菌体总量大,废菌废渣废水难处理,不能进行多批次连续生产等。酶法生产gsh分为两步酶法和一步酶法。两步酶法是先以l-谷氨酸和l-半胱氨酸为底物,利用γ-谷氨酰半胱氨酸合成酶(γ-ecl或gshi)将其催化合成γ-谷氨酰基-l-半胱氨酸(γ-glu-l-cys),再加入l-甘氨酸,通过谷胱甘肽合成酶(gs或gshii)催化合成谷胱甘肽,具体反应如反应式a所示。一步酶法是利用双功能谷胱甘肽合成酶(gshf),将底物l-谷氨酸、l-半胱氨酸和l-甘氨酸直接催化合成谷胱甘肽,具体反应如反应式b所示。由于双功能谷胱甘肽合成酶(gshf)同时具有gshi和gshii的活性,而且受产物gsh的反馈抑制作用比较小,催化效率更高,因此,gshf日益受到重视。双功能谷胱甘肽合成酶来源于非常广泛,许多微生物来源的gshf被国内外科研工作者发现、挖掘、筛选、克隆、表达和应用。对于双功能谷胱甘肽合成酶,国外研究人员已经开展相关工作并取得很大的进展,如bjornvergauwen等(thejournalofbiologicalchemistryvol.281,no:7,pp.4380-4394)发现来源于多杀巴斯德菌(pasteurellamultocida)的谷胱甘肽合成酶同时具有γ-ecl和gs两种酶的活性,其与多种来源的γ-ecl进行序列比对,发现它们之间的同源性不高并将该酶命名为gshf,构建pgem-gshf载体并在大肠杆菌k12中高表达,发现细胞内有46.2nmol/mg总蛋白的gsh存在,而对照组中没有发现gsh;griffith等也报道了在无乳链球菌(streptococcusagalactiae)中存在类似的gshf;aharonowitz等也发现了在单增李斯特氏菌(listeriamonocytogenes)中也有相关的gshf;近几年,国内科研工作者也对gshf开展相应的研究并应用于生产gsh,如中国专利cn102586369a中,李志敏等对嗜热链球菌(streptococcusthermophilus)来源的gshf进行基因克隆,表达并构建大肠杆菌基因工程菌,该重组菌经优化发酵条件,其gsh产量最高可以达到4.5g/l;如中国专利cn102071171a中,叶勤等克隆并表达了7种来源的gshf,其中重组菌jm109(ptrc99a-gshfap)比野生型大肠杆菌jm109在同样条件下反应合成的gsh高40多倍,经37℃体外反应6小时,gsh浓度可达到7.1g/l;如中国专利cn103122322a中,朱泰承等将来源于单增李斯特氏菌(listeriamonocytogenes)的gshf编码基因插入pgapzαa载体并转入毕赤酵母gs115中形成重组基因工程菌,该重组菌经发酵培养42小时后,其gsh浓度可以达到190mg/l。目前,在酶法生产gsh的过程中发现,野生型gshf的合成活性偏低,比活不高,受产物反馈抑制作用强,稳定性差等缺点,导致酶法生产gsh技术未能得到广泛应用。随着酶工程、基因工程和生物信息技术的发展,许多科研工作者运用理性设计、半理性设计和定向进化等手段对野生型gshf进行突变改造。如中国专利cn104328092a中,傅荣昭对来源于melissococcusplutoniusatcc35311的gshf中的第128位、第256位及第320位进行定点突变,使其催化活性比野生型提高了2倍,其突变体固定化酶生产gsh的浓度可达到50mmol/l;如中国发明专利cn105238797a中,徐志南等对无乳链球菌来源的gshf进行突变,得到3种突变体gshfm1、gshfm2和gshfm3,所得的突变体编码基因被导入毕赤酵母菌中形成gsh重组生产菌,重组菌经发酵培养,最高gsh浓度达到837mg/l。虽然,国内外科研工作者利用基因工程及蛋白工程技术对双功能谷胱甘肽合成酶进行突变改造,使其合成活性有了成倍的提高,但是突变酶的合成活性还偏低、固定化酶使用批次还需进一步提高。因此,开发一种合成活性高和操作稳定性好的酶具有重要意义。技术实现要素:本发明提供了一种双功能谷胱甘肽合成酶突变体、核苷酸序列及其制备方法和应用,以解决突变酶的合成活性还偏低、固定化酶使用批次低的技术问题。本发明采用的技术方案如下:一种双功能谷胱甘肽合成酶突变体,双功能谷胱甘肽合成酶突变体的氨基酸序列第3位、第123位、第161位、第194位、第382位和第390位中的至少一个位点的氨基酸与seqidno:2所示的唾液链球菌野生型双功能谷胱甘肽合成酶氨基酸序列的相应位点氨基酸不同,其余位点的氨基酸相同。进一步地,双功能谷胱甘肽合成酶突变体的氨基酸序列除第3位和/或第161位之外其余位点的氨基酸与唾液链球菌野生型双功能谷胱甘肽合成酶氨基酸序列相应位点的氨基酸相同。当第3位不同时,双功能谷胱甘肽合成酶突变体的氨基酸序列第3位的氨基酸为l。当第161位不同时,双功能谷胱甘肽合成酶突变体的氨基酸序列第161位的氨基酸为g或d。当第3和161位均不同时,双功能谷胱甘肽合成酶突变体的氨基酸序列第3位的氨基酸为l且第161位为d,即为seqidno:4所示的氨基酸序列gshf-2。进一步地,双功能谷胱甘肽合成酶突变体的氨基酸序列除第123位和/或第194位之外其余位点的氨基酸与氨基酸序列gshf-2相应位点的氨基酸相同。当第123位不同时,双功能谷胱甘肽合成酶突变体的氨基酸序列第123位的氨基酸为f或y。当第194位不同时,双功能谷胱甘肽合成酶突变体的氨基酸序列第6位的氨基酸为s。当第123和194位均不同时,双功能谷胱甘肽合成酶突变体的氨基酸序列第123位的氨基酸为y且第194位为s,即为seqidno:6所示的氨基酸序列gshf-3。进一步地,双功能谷胱甘肽合成酶突变体的氨基酸序列除第382位和第390位之外其余位点的氨基酸与氨基酸序列gshf-3相应位点的氨基酸相同。双功能谷胱甘肽合成酶突变体的氨基酸序列第382位的氨基酸为q且第390位的氨基酸为p,即为seqidno:8所示的氨基酸序列gshf-4。本发明另一方面提供了双功能谷胱甘肽合成酶突变体的核苷酸序列,双功能谷胱甘肽合成酶突变体的氨基酸序列包括gshf-2、gshf-3和gshf-4,双功能谷胱甘肽合成酶突变体的核苷酸序列为以下的任一种:1)、seqidno:3所示的编码gshf-2的核苷酸序列、seqidno:5所示的编码gshf-3的核苷酸序列和seqidno:7所示的编码gshf-4的核苷酸序列。2)、在严格条件下与1)中任一核苷酸序列杂交且编码具有双功能谷胱甘肽合成酶活性的蛋白质的核苷酸序列。3)、与1)或2)中任一项限定的核苷酸序列具有95%以上的同源性且编码具有gshf活性的蛋白质的核苷酸序列。本发明还提供了一种上述的双功能谷胱甘肽合成酶突变体的制备方法,包括以下步骤:将seqidno:1的唾液链球菌野生型双功能谷胱甘肽合成酶核苷酸序列进行随机突变、饱和突变和迭代饱中的一种或几种,得到编码上述双功能谷胱甘肽合成酶突变体的核苷酸序列。将突变体的核苷酸序列插入到载体中得到重组表达载体。将重组表达载体转化进入表达菌株中得到重组基因工程菌。将重组基因工程菌进行发酵诱导,破碎,分离得到双功能谷胱甘肽合成酶突变体。本发明还提供了一种双功能谷胱甘肽合成酶突变体的制备方法,还包括纯化步骤,纯化步骤包括:将双功能谷胱甘肽合成酶突变体通过固定化金属螯合亲和层析法进行纯化得到双功能谷胱甘肽合成酶突变体纯酶。进一步地,还包括将双功能谷胱甘肽合成酶突变体固定化的步骤,固定化的步骤包括:将双功能谷胱甘肽合成酶突变体纯酶用磷酸盐缓冲液溶解,然后加入50g经活化处理后的载体,于25℃、120rpm条件下低速搅拌固定化48h,将所得固定化酶用去离子水反复清洗3~5遍,真空滤干后即得双功能谷胱甘肽合成酶突变体的固定化酶,磷酸盐缓冲液的ph为8.0,浓度为0.1mol/l,载体为环氧基载体ecep或氨基载体echa/s,优选为氨基载体echa/s。本发明还提供了一种上述双功能谷胱甘肽合成酶突变体的固定化酶在制备谷胱甘肽中的应用。进一步地,以双功能谷胱甘肽合成酶突变体的固定化酶为催化剂,以l-cys,l-glu,l-gly,atp和mg2+为底物,反应过程ph恒定8.00,反应温度为37℃或50℃,优选为50℃。本发明具有以下有益效果:上述双功能谷胱甘肽合成酶突变体,与唾液链球菌野生型gshf氨基酸序列存在一个或多个位点的氨基酸不同,具有谷胱甘肽合成活性更高,反应速率更快,转化效率更好的优点,可以更有效,快速地将催化合成gsh。除了上面所描述的目的、特征和优点之外,本发明还有其它的目的、特征和优点。下面将参照图,对本发明作进一步详细的说明。附图说明构成本申请的一部分的附图用来提供对本发明的进一步理解,本发明的示意性实施例及其说明用于解释本发明,并不构成对本发明的不当限定。在附图中:图1是本发明优选实施例的重组表达载体pet-gshf-wt构建流程图;图2是本发明优选实施例的gshf-3突变体蛋白三维结构图;图3是本发明实施例3中的gshf-4突变体固定化酶合成gsh的hplc图。具体实施方式以下结合附图对本发明的实施例进行详细说明,但是本发明可以由权利要求限定和覆盖的多种不同方式实施。本申请中下述实施例中所使用的方法如无特殊说明均为常规方法,如《分子克隆实验指南》(j.萨姆布鲁克,d.w.拉塞尔著,黄培堂,汪嘉玺,朱厚础,等译。第3版,北京:科学出版社,2002)中所述的方法进行。基因合成、突变引物合成以及序列测序均为泰和永昌(长沙)生物技术有限公司完成。大肠杆菌宿主菌为e.colibl21(de3)购于merck公司。大肠杆菌宿主菌还可以是e.colibl21(de3)plys,购于天根公司。原核表达载体pet30a(+)购于merck公司。dna内切酶ecori、xhoi和dpni及dnat4连接酶购于fermentas公司,其余试剂及材料均为市售。同时,本发明中的氨基酸无特别说明外均用其缩写或代号标明(氨基酸中英文名称及其缩写和代号见表1)。表1氨基酸中英文名称及其缩写和代号中文名称英文名称缩写代号中文名称英文名称缩写代号丙氨酸alaninealaa脯氨酸prolineprop精氨酸arginineargr亮氨酸leucineleul天冬酰胺asparagineasnn异亮氨酸isoleucineilei天冬氨酸asparticacidaspd甘氨酸glycineglyg半胱氨酸cysteinecysc苯丙氨酸phenylalaninephef谷氨酰胺glutamineglnq蛋氨酸methioninemetm谷氨酸glutamicacidglue赖氨酸lysinelysk苏氨酸threoninethrt组氨酸histidinehish色氨酸tryptophantrpw缬氨酸valinevalv丝氨酸serinesers酪氨酸tyrosinetyry双功能谷胱甘肽合成酶酶活力的检测(即hplc法)如下所示:(1)溶液配置a、底物溶液的配置精确称取半胱氨酸1.40g、谷氨酸1.77g、甘氨酸0.90g、atp6.09g和mgcl2·6h2o3.28g,用0.1mol/l、ph8.0的tris-hcl缓冲液溶解,调ph至8.0,最后定容100ml。b、流动相的配置色谱柱:shim-packxr-ods2.2μm4.6×30mm流动相:称取3.4gkh2po4,溶于一定体积超纯水中,用酸调节ph为3.00±0.02,用0.22μm的水系滤膜过滤后,加入15ml色谱级甲醇,脱气20min。(2)酶活力的测定精确量取0.5ml的液态酶或称取0.2g固定化酶,加入含有已预热至37℃的底物溶液10ml,于37℃的水浴中搅拌反应10min后,加入25%的三氯乙酸溶液2ml终止反应(固定化酶采用过滤离心法终止反应),摇匀后离心,精确量取上清液0.5ml于25ml容量瓶中,用流动相定容后进行hplc分析,以gsh标准品作为对照。酶活力计算注:w标:谷胱甘肽标准品称量,mg;p标:谷胱甘肽标准品含量,%;a样:样品hplc检测中谷胱甘肽面积;m:谷胱甘肽分子量;t:反应时间,min;v:液态酶取样量,ml;w:固定化酶称量,g。酶活力单位(u)定义为:在37℃,ph8.0条件下,在1min内催化底物生成1μmol产物gsh的所需的酶量为1u。本发明的优选实施例提供了一种双功能谷胱甘肽合成酶突变体,双功能谷胱甘肽合成酶突变体的氨基酸序列第3位、第123位、第161位、第194位、第382位和第390位中的至少一个位点的氨基酸与seqidno:2所示的唾液链球菌野生型gshf氨基酸序列的相应位点氨基酸不同,其余位点的氨基酸相同。与唾液链球菌野生型gshf氨基酸序列相比,双功能谷胱甘肽合成酶突变体(gshf突变体)的氨基酸序列中第3位、第123位、第161位、第194位、第382位和第390位有一个或多个位点的氨基酸不同,其余位点的氨基酸相同。因此gshf突变体的氨基酸序列可以唾液链球菌野生型gshf氨基酸序列为基础,通过定向进化技术结合计算机模拟辅助设计手段等非理性及半理性设计的酶工程改造技术对seqidno:2所示的唾液链球菌(streptococcussalivarius)来源的gshf进行突变改造得到。唾液链球菌(streptococcussalivarius)野生型gshf氨基酸序列中的第3位、第123位、第161位、第194位、第382位和第390位氨基酸有1个或多个位点发生取代,得到gshf突变体氨基酸序列。当然也可采用其它的基因合成及构建方式得到上述的gshf突变体氨基酸序列,如采用全基因合成的方式。以下均以酶工程改造技术对野生型gshf氨基酸序列进行突变的方式获得gshf突变体氨基酸序列。在获知相应的氨基酸序列的情况,无疑可相应采用全基因合成的方式获得双功能谷胱甘肽合成酶突变体。由以下的实施例中可知,gshf突变体氨基酸序列在第3位、第123位、第161位、第194位、第382位和第390位中有一个位点的氨基酸与野生型gshf的氨基酸不同,gshf突变体在应用于酶法生产gsh时即可使得谷胱甘肽合成活性更高,反应速率更快,转化效率更好。本发明具有以下有益效果:上述双功能谷胱甘肽合成酶突变体,与唾液链球菌野生型gshf氨基酸序列存在一个或多个位点的氨基酸不同,具有谷胱甘肽合成活性更高,反应速率更快,转化效率更好的优点,可以更有效,快速地将催化合成gsh。可选地,双功能谷胱甘肽合成酶突变体的氨基酸序列除第3位和/或第161位之外其余位点的氨基酸与唾液链球菌野生型gshf氨基酸序列相应位点的氨基酸相同。当第3位不同时,双功能谷胱甘肽合成酶突变体的氨基酸序列第3位的氨基酸为l。当第161位不同时,双功能谷胱甘肽合成酶突变体的氨基酸序列第161位的氨基酸为g或d。当第3和161位均不同时,双功能谷胱甘肽合成酶突变体的氨基酸序列第3位的氨基酸为l且第161位为d,即为seqidno:4所示的氨基酸序列gshf-2。在本实施例中,gshf突变体氨基酸序列在seqidno:2的唾液链球菌(streptococcussalivarius)野生型gshf氨基酸序列基础上对第3位和/或第161位上的氨基酸进行取代。在第3位进行取代时,野生型gshf氨基酸序列的第3位氨基酸由i突变为l,得到gshf突变体。在第161位进行取代时,野生型gshf氨基酸序列的第161位氨基酸由n突变为g或d,得到gshf突变体。并且相比之下,第161位氨基酸由n突变为d为更优选的方案。在第3位和161位同时进行取代时,野生型gshf氨基酸序列的第3位氨基酸由i突变为l且第161位氨基酸由n突变为d,得到gshf突变体gshf-2,其氨基酸序列如seqidno:4所示。本申请碍于篇幅未在序列表中直接提供上述所有gshf突变体的氨基酸序列,但显然也公开上述的其余的gshf突变体的氨基酸序列。如仅第3位进行取代得到的gshf突变体的氨基酸序列,将野生型gshf氨基酸序列的第3位氨基酸由i突变为l即可得到相应的gshf突变体的氨基酸序列。其它位点突变如(如第161位进行取代、123位和/或第194位进行取代)的gshf突变体根据其突变结果,均可得到相应的氨基酸序列。可选地,双功能谷胱甘肽合成酶突变体的氨基酸序列除第123位和/或第194位之外其余位点的氨基酸与氨基酸序列gshf-2相应位点的氨基酸相同。当第123位不同时,双功能谷胱甘肽合成酶突变体的氨基酸序列第123位的氨基酸为f或y。当第194位不同时,双功能谷胱甘肽合成酶突变体的氨基酸序列第6位的氨基酸为s。当第123和194位均不同时,双功能谷胱甘肽合成酶突变体的氨基酸序列第123位的氨基酸为y且第194位为s,即为seqidno:6所示的氨基酸序列gshf-3。在本实施例中,gshf突变体在gshf-2突变体氨基酸序列基础上对第123位和/或第194位上的氨基酸进行取代。在第123位进行取代时,gshf-2突变体氨基酸序列的第123位氨基酸由h突变为f或y,得到gshf突变体。相比之下,第123位氨基酸由h突变为y为更优选的方案。在第194位进行取代时,gshf-2突变体氨基酸序列的第194位氨基酸由a突变为s,得到gshf突变体。在第123位和第194位同时进行取代时,gshf-2突变体氨基酸序列的第123位氨基酸由h突变为y且第194位氨基酸由a突变为s,得到gshf突变体gshf-3,其氨基酸序列如seqidno:6所示。可选地,的双功能谷胱甘肽合成酶突变体除第382位和第390位之外其余位点的氨基酸与氨基酸序列gshf-3相应位点的氨基酸相同。双功能谷胱甘肽合成酶突变体的第382位的氨基酸为q且第390位的氨基酸为p,即为seqidno:8所示的氨基酸序列gshf-4。本实施例中gshf突变体在gshf-3的氨基酸序列基础上对第382位和第390位上的氨基酸同时进行取代,其中第382位氨基酸由h突变为q,第390位氨基酸由s突变为p,即gshf-4,其氨基酸序列如seqidno:8所示。本发明另一方面提供了双功能谷胱甘肽合成酶突变体的核苷酸序列,双功能谷胱甘肽合成酶突变体的氨基酸序列包括gshf-2、gshf-3和gshf-4,双功能谷胱甘肽合成酶突变体的核苷酸序列为以下的任一种:1)、seqidno:3所示的编码gshf-2的核苷酸序列、seqidno:5所示的编码gshf-3的核苷酸序列和seqidno:7所示的编码gshf-4的核苷酸序列。2)、在严格条件下与1)中任一核苷酸序列杂交且编码具有gshf活性的蛋白质的核苷酸序列。3)、与1)或2)中任一项限定的核苷酸序列具有95%以上的同源性且编码具有gshf活性的蛋白质的核苷酸序列。本发明还提供了一种上述的双功能谷胱甘肽合成酶突变体的制备方法,包括以下步骤:将seqidno:1的唾液链球菌野生型gshf核苷酸序列进行随机突变、饱和突变和迭代饱中的一种或几种,得到编码如前的gshf的突变体的核苷酸序列。将突变体的核苷酸序列插入到载体中得到重组表达载体。将重组表达载体转化进入表达菌株中得到重组基因工程菌。将重组基因工程菌进行发酵诱导,破碎,分离得到双功能谷胱甘肽合成酶突变体。将具有序列表中seqidno:1的唾液链球菌野生型gshf核苷酸序列进行随机突变、饱和突变、迭代饱和突变等步骤得到编码如前所述的gshf的突变体的基因序列。将突变体基因序列插入到载体中得到重组表达载体。载体可以是质粒。插入方法可按照本领域的常规技术手段进行。本发明还提供了一种双功能谷胱甘肽合成酶突变体的制备方法,还包括纯化步骤,纯化步骤包括:将双功能谷胱甘肽合成酶突变体通过固定化金属螯合亲和层析法进行纯化得到双功能谷胱甘肽合成酶突变体纯酶。可选地,还包括将双功能谷胱甘肽合成酶突变体固定化的步骤,固定化的步骤包括:将双功能谷胱甘肽合成酶突变体纯酶用磷酸盐缓冲液溶解,然后加入50g经活化处理后的载体,于25℃、120rpm条件下低速搅拌固定化48h,将所得固定化酶用去离子水反复清洗3~5遍,真空滤干后即得双功能谷胱甘肽合成酶突变体的固定化酶,磷酸盐缓冲液的ph为8.0,浓度为0.1mol/l,载体为环氧基载体ecep或氨基载体echa/s,优选为氨基载体echa/s。本发明还提供了一种上述双功能谷胱甘肽合成酶突变体的固定化酶在制备谷胱甘肽中的应用。可选地,以双功能谷胱甘肽合成酶突变体的固定化酶为催化剂,以l-cys,l-glu,l-gly,atp和mg2+为底物,反应过程ph恒定8.00,反应温度为37℃或50℃,优选为50℃。实施例1:野生型gshf重组基因工程菌的构建、表达及重组蛋白的纯化和固定化1-1野生型gshf基因的获得为了得到合成活力高、产物抑制弱、耐热性强及操作稳定性好的突变型gshf,本发明采用的野生型gshf基因及氨基酸序列是来源于唾液链球菌(streptococcussalivarius)(genebank登录号:wp_038676473),通过大肠杆菌密码子偏爱性将该编码基因进行优化并进行全基因合成,并将该野生型双功能谷胱甘肽合成酶命名为gshf-wt,其编码基因命名为gshf-wt,优化后的核苷酸序列及氨基酸序列见seqidno:1和seqidno:2。1-2野生型gshf原核表达载体的构建及重组基因工程菌的构建参照图1,将全基因合成的野生型gshf编码基因gshf-wt与原核表达载体pet30a(+)分别进行ecori和xhoi双酶切,酶切3小时后分别进行切胶回收,将回收产物按照产物:载体为3:1的摩尔比例进行混合,加入t4dnaligase于16℃过夜连接。将连接产物5μl转入50μl的dh5α感受态中,涂布于含50μg/ml卡那霉素的lb固体培养基平板于37℃中进行过夜培养。挑选单菌落进行菌落pcr验证,将阳性克隆接种于含50μg/ml卡那霉素的lb培养基过夜培养,提质粒,进行ecori和xhoi双酶切验证,将大小正确的克隆子送往测序公司进行dna测序验证,序列比对正确后,将该重组表达载体命名为pet-gshf-wt,这样就得到了在n端和c端都带有一个his-tag的质粒,其表达的野生型gshf蛋白带有两个组氨酸标签,可用固定化金属螯合亲和层析(imac)的方式进行蛋白纯化。通过化学转化的方法将上述重组表达载体pet-gshf-wt转化入e.colibl21(de3)或者e.colibl21(de3)plys感受态细胞,转化细胞涂布于含50μg/ml卡那霉素的lb固体培养基平板于37℃中进行过夜培养即获得重组野生型gshf基因工程菌。1-3、重组野生型gshf蛋白的分离纯化及固定化(1)重组野生型gshf蛋白的分离纯化由于在表达载体构建过程中引入了原核表达载体pet30a(+)中的n和c端的2个his-tag,因此,本发明人利用该组氨酸标签进行固定化金属螯合亲和层析(imac)来纯化重组蛋白,具体方法如下。取100ml过夜诱导后的野生型gshf发酵液,离心后弃上清收集菌体(10000rpm、4℃、10min),用磷酸盐缓冲液(ph8.0、0.1mol/l)反复洗涤菌体两次,离心收集菌体,浓缩5倍重悬于20ml磷酸盐缓冲液(ph8.0、0.1mol/l)中。将上述处理后的菌液置于冰水中进行超声破碎直至澄清,其中的超声破碎条件为:工作2s,间隔5s。将上述破碎后的裂解液置于低温高速离心机中离心(12,000rpm、4℃、20min),收集上清,得到重组野生型gshf蛋白,将该粗重组蛋白进样到已活化并结合ni+的ida树脂上,用梯度浓度的咪唑进行梯度洗脱,利用蛋白层析系统(bio-rad)进行实时监控,当计算机中出现稳定的蛋白峰时,开始收集直到该峰消失为止。重组酶蛋白经过分离纯化后密封于无菌袋中放置于4℃冰箱以备后续实验(2)重组野生型gshf蛋白的固定化a、固定化载体的活化准确量取60%的戊二醛30ml、磷酸氢二钾(k2hpo4·3h2o)4.76g溶解于600ml去离子水中,最后用去离子水定容至1000ml,同时用磷酸溶液调节其ph为8.0,灭菌后备用;将环氧基载体ecep或氨基载体echa/s(意大利resindions.r.l公司)250g投入到上述溶液中,并于37℃低速搅拌活化2h,过滤收集载体,并用无菌去离子水反复冲洗2~3次后真空滤干备用。b、酶的固定化取一定量上述纯化后的酶液,用磷酸盐缓冲液(ph8.0、0.1mol/l)溶解,然后加入50g经活化处理后的载体,于37℃、120rpm条件下低速搅拌固定化48h,将所得固定化酶用去离子水反复清洗3~5遍,真空滤干后即得固定化酶终产品。精确称取1g上述固定化酶进行活力测定,以ecep和echa/s为载体的固定化酶酶活力分别为18u/g和23u/g,氨基载体echa/s固定化酶活力明显高于环氧基载体ecep固定化酶,因此选择氨基载体echa/s作为双功能谷胱甘肽合成酶的固定化载体。实施例2:gshf突变体的制备2-1高合成活力gshf突变体的制备2-1-1gshf突变体文库的构建为了提高野生型gshf的合成活力,本发明人以重组表达载体pet-gshf-wt为dna模板,其中引物为t7通用引物(seqidno:15和16),通过易错pcr的方法构建一个随机突变体文库,并通过调整易错pcr反应体系中mg2+和mn2+浓度以及dctp和dttp寡核苷酸浓度,使该突变体文库的碱基错配率为千分之五,即保证一个突变体有1到3个氨基酸发生突变,构建突变体文库的具体过程如下。易错pcr反应体系:10×buffer5μl2mmol/ldntps5μl100mmol/ldctp0.5μl100mmol/ldttp0.5μl10mmol/lmncl25μl50mmol/lmgso45μl引物t7promoter2μl引物t7terminator2μl模板pet-gshf-wt1μltaqdna聚合酶1.5μlddh2o22.5μl易错pcr反应条件是:先95℃预变性5min;然后94℃变性30s,55℃退火1min,72℃1.5min,共30个循环;最后72℃延伸10min。将上述得到的易错pcr产物进行电泳并切胶回收纯化,将纯化后的产物与原核表达载体pet30a(+)分别进行ecori和xhoi双酶切,酶切3小时分别进行切胶回收,将回收产物按照产物:载体为3:1的摩尔比例进行混合,加入t4dnaligase于16℃过夜连接。次日,按照实施例1-2的方法构建重组基因工程菌,即可得到一个库容量大的突变体文库。2-1-2gshf突变体文库的高通量筛选方法本发明中gshf突变体文库的高通量筛选方法是采用磷钼杂多蓝法,其原理为:还原型gsh将磷钼杂多酸还原为磷钼杂多蓝,在710nm处有最大吸收峰。具体方法及操作步骤如下所述:(1)酶液的制备用高温灭菌后的牙签,小心挑取突变体文库中的单菌落接种于装有lb培养基的96孔细胞培养板中,其中lb培养基体积为200μl/孔且含有50μg/ml的卡那霉素,于37℃,250rmp的恒温摇床中培养8个小时后,加入1%的乳糖,于25℃,250rmp的恒温摇床中诱导培养8小时。诱导完毕后,将96孔细胞培养板放入-86℃的超低温冰箱中冷冻2个小时,取出放置于室温中半个小时,后置于96孔细胞培养板离心机中于4,000rmp,4℃离心20分钟。(2)反应取20μl离心酶液加入含有180μl的底物溶液的96孔细胞培养板中,放置于37℃的恒温恒湿培养箱中反应2个小时,加入50μl三氯乙酸(25%,w/v)终止反应,后置于96孔细胞培养板离心机中于4,000rmp,4℃离心20分钟。(3)显色取40μl上清液加入含有160μl显色液(60μl1%的na2moo4,4μl1mol/l的h2so4,20μl0.5%磷酸酸铵,76μlddh2o)的96孔pcr管中置于pcr仪中于99℃放置30分钟,后立即降温至37℃,取100μl处理液置于酶标板中,放入酶标仪,于波长710nm处测定其吸光度,读取数值。2-1-3高合成活力gshf突变体的制备从上述突变子文库中筛选了约10000个克隆,得到6个颜色变化明显且数值比野生型gshf高的突变子。接着对这6个突变子进行摇瓶筛选,具体过程为:将这6个突变子接种于含100mllb培养基的500ml摇瓶中进行发酵及诱导,并通过hplc法测定活力,得到2个比对照活力高1.5倍和2倍的克隆子,分别命名为gshf-1a和gshf-1b,经过测序结果显示gshf-1a在第3位点氨基酸发生了改变,由i突变成了l,而gshf-1b在第161位点氨基酸发生了改变,由n突变成了g。gshf-1a和gshf-1b单点随机突变效果明显,为了得到合成活力更好的突变体,本发明人对第3和第161这2个氨基酸位点进行双点迭代饱和突变,具体方法如下。第3位和第161位氨基酸双点迭代饱和突变引物p1:ttcatgactnnkaatcaactgctgcaaaaactggaacc(下划线处为饱和突变位点)(n为a或t或c或g,k是g或t)p2:gtcggtttgmnngctttctttgaacagagcttcaaccag(下划线处为饱和突变位点)(n为a或t或c或g,m是a或c)以pet-gshf-wt为模板,分别以p1/p2为突变引物,扩增出同时含有第3位和第161位突变点的大引物片段,再以pet-gshf-wt为模板运用大引物pcr技术进行全长扩增,将扩增后的全长基因进行dpni酶切消化去掉模板pet-gshf-wt,后进行纯化,化转bl21(de3),于37℃过夜培养即得到含有第3位和第161位的全饱和突变体文库,其中pcr所用的kod-plus-neo高保真dna聚合酶及相应pcr缓冲液、mg2+、dntps溶液均购买于toyobo公司,并参照其说明书进行pcr反应体系的配置及pcr反应条件的设定。按照上述实施例2-1-2的方法,对突变体文库筛选了3600个克隆子,得到多个活性提高的突变体,其中提高倍数最高达5倍,其突变后酶活性为10u/ml,并将该突变体测序,测序结果显示该突变体的第3位点氨基酸由i突变成了l,161位点氨基酸由n突变成了d,并将其命名为gshf-2,其编码基因序列为seqidno:3,氨基酸序列为seqidno:4。同时按照实施例1-3的方法对上述gshf-1a、gshf-1b和gshf-2突变子进行蛋白纯化及性质分析,具体比较结果见表2。表2中,i3l+n161d,表示第3位由i突变成l且第161位由n突变成d。其余表述和其余表格的相应表与之类似。表2高活力gshf突变体与野生型gshf初步比较编号氨基酸突变点酶活力比活提高倍数gshf-wt野生型2u/ml2.5u/mg0gshf-1ai3l3u/ml3.6u/mg1.5gshf-1bn161g4u/ml4.8u/mg2gshf-2i3l+n161d10u/ml11.2u/mg52-2产物抑制性弱的gshf突变体的制备众多文献表明,在gshf催化生产gsh过程中,gshf会受到产物gsh的强烈反馈抑制,导致gshf活性下降且不能连续进行批次反应,这就是目前酶法制备gsh单位产量不高的原因。因此,需要通过对gshf进行蛋白工程改造以降低其受产物gsh反馈抑制的作用。本发明人,沿着这一目标设定开发路线,对上述gshf突变体进行进一步改造。以gshf-2基因为模板,加入t7通用引物进行易错pcr,按照实施例2-1-1方法得到一个随机突变体文库,同时按照实施例2-1-2方法进行突变体文库筛选,只是在反应过程中加入1%(w/v)的产物gsh以筛选耐受高浓度产物gsh的突变子。在所述随机突变体文库中共筛选了约20000个克隆子,得到2个活力比gshf-2有显著提升的突变子,通过摇瓶发酵复筛选,测定其活力分别为13.6u/ml和14.8u/ml,并分别命名为gshf-2a和gshf-2b,经测序后,显示gshf-2a在第123位点氨基酸由h突变为f,gshf-2b在第194位点氨基酸由a突变为s。为了得到合成活性更好、受产物抑制作用更弱的突变体,本发明人对第123位点和第194位点这2个氨基酸位点进行双点迭代饱和突变,具体方法如下。第123位点和第194位点氨基酸双点迭代饱和突变引物p3:ttcgaacgtnnktaccgtaactacctggcggagaagtatg(下划线处为饱和突变位点)(n为a或t或c或g,k是g或t)p4:ggcaattggmnntgcgccaaacagataggtgataacccag(下划线处为饱和突变位点)(n为a或t或c或g,m是a或c)按照上述实施例2-1-3的方法构建含有123位点和194位点的全饱和突变文库,对突变体文库筛选了3600个克隆子,得到多个活性提高的突变体,其中提高倍数最高达11倍,其突变后酶活性为22.4u/ml,比活为24u/mg,并将该突变体测序,测序结果显示该突变体的第123位点氨基酸由h突变成了y,194位点氨基酸由a突变成了s,并将其命名为gshf-3,其编码基因序列为seqidno:5,氨基酸序列为seqidno:6。gshf-3突变体蛋白三维结构图,如图2所示。同时按照实施例1-3的方法对上述突变子进行蛋白纯化及性质分析,具体比较结果见表3。表3产物抑制性弱的gshf突变体与野生型gshf初步比较将上述野生型及突变型gshf按照实施例1-3的方法制备固定化酶,并将所得到的固定化酶以相同酶量(1000u)分别置于一定浓度(5g/l、10g/l、20g/l和30g/l)的gsh溶液中,于37℃温育1个小时,后取出固定化酶用无菌去离子水反复冲洗4~5遍,以确保固定化酶中不存在gsh残留,将处理后的固定化酶用hplc法测定其残余活力,具体数据如表4所示。结果表明,gshf-3突变体比野生型gshf有着更好的gsh耐受性,其受gsh的反馈抑制作用减弱非常多。表4野生型gshf与gshf突变体固定化酶对gsh浓度耐受性比较2-3热稳定性提高的gshf突变体的制备在工业生产gsh的过程中发现,高温反应具有提高底物溶解度、降低反应液粘度、缩短反应时间和提高gsh产量等优点。但是,目前大多数gshf在高温反应条件下,其活力迅速下降、稳定性较差,因此急需开发一种热稳定性提高的gshf突变体。为了解决工业生产中的这一难题,本发明人设定新的开发路线,对上述gshf突变体进行更进一步改造,具体方法及操作步骤如下所述。(1)确定与温度相关的氨基酸位点利用swiss-model(http://swissmodel.expasy.org)在线同源建模软件对gshf-3突变体进行同源模拟,获得gshf-3突变体的蛋白三维结构;将gshf-3突变体氨基酸序列输入ncbi数据库中进行同源序列搜索,找出与gshf-3突变体高度相似的氨基酸序列并下载保存;采用多序列比对软件clustalw2程序对这些氨基酸序列进行比对;利用分子对接软件autodock4.0对gshf-3突变体蛋白三维结构模型和产物gsh进行分子对接得到gshf-gsh蛋白产物复合体;通过对该复合体进行空间结构分析及多序列比对结果分析,确定要突变的与温度相关的氨基酸位点,这些位点分别为第382位组氨酸和第390位丝氨酸。(2)定点饱和突变及筛选以gshf-3突变体基因为模板,对382和390这2个氨基酸位点进行双点迭代饱和突变,具体方法如下。第382位和第390位氨基酸双点饱和突变引物:p5:gacccgtccnnkaccctggctgcacaactgctgnnktacatcaaagacaaaagc(下划线处为饱和突变位点)(n为a或t或c或g,k是g或t)p6:tttgatgtamnncagcagttgtgcagccagggtmnnggacgggtccgcgaatgc(下划线处为饱和突变位点)(n为a或t或c或g,m是a或c)按照实施例2-1方法进行突变体文库的构建及筛选,只是在酶液制备过程中将粗酶液于60℃温育1小时后再进行后续实验。在所述双点迭代饱和突变体文库筛选了3600个克隆子,得到了一个酶活力为25.6u/ml的突变体,经过测序结果验证,该六点gshf突变体相对于gshf-3突变体来说,第382位点氨基酸由h突变成了q,第390位点氨基酸由s突变成了p,并将其命名为gshf-4,其编码基因序列为seqidno:7,氨基酸序列为seqidno:8。(3)野生型gshf与gshf突变体的最适反应温度为了测定野生型gshf与gshf突变体的最适反应温度,本发明人分别于25℃、30℃、37℃、45℃、50℃、55℃和60℃条件进行反应下,采用hplc的方法测定野生型gshf与gshf突变体的酶力,得到不同温度条件下的酶活力,从而确定其最适反应温度,具体结果见表5。其结果表明,野生型gshf的最适反应温度为30℃;gshf-3突变体的最适反应温度为37℃;gshf-4突变体的最适反应温度为50℃。并且突变体gshf-4相对于野生型gshf的酶活力由的2u/ml提高至25.6u/ml,活力提高了12.8倍,比活由2.5u/g提高至27.5u/g,提高了11倍。由此可见,gshf-4突变体的最适反应温度和高温合成活性得到了显著提高。表5热稳定性提高的gshf突变体与野生型gshf初步比较(4)野生型gshf与gshf突变体的热稳定性为了评价gshf突变体的热稳定性,本发明人将上述野生型及突变型gshf按照实施例1-3的方法制备纯酶,并将所得到的纯酶以相同酶量(100u)分别置于一定温度中(37℃、50℃、55℃和60℃)温育1个小时,后取出纯酶降温至37℃测定其残余活力,具体数据如表6所示。其结果表明,野生型gshf,在37℃时,其活力就开始下降,当温度升高到50℃时,其活力已经丢失将近一半,将温度升高到60℃时,其活力基本丢失完全;gshf-4突变体,当温度升高到50℃时,其活力都没有明显下降,当温度升高到55℃时,其活力仅下降了8.7%,显示出良好的热稳定性。表6野生型gshf与突变型gshf固定化酶热稳定性比较实施例3突变体gshf-4固定化酶制备gsh的应用3-1野生型gshf与gshf-4固定化酶制备gsh实验将上述野生型gshf及gshf-4固定化酶,以相同酶量(5000u)分别置于反应液中(70mmoll-cys,105mmoll-glu,105mmoll-gly,105mmolatp,140mmolmg2+),于37℃或50℃,ph8.00条件下反应,反应过程中不断滴加3mol/l的碳酸钠,使ph恒定在8.00,当反应达到终点后(判定终点的方法是:碳酸钠自动停止补加,ph值维持在3min以上不变),记录下总反应时间。将上述反应液过滤,固定化酶用无菌去离子水反复冲洗4~5遍,备用。取上述反应液,通过hplc方法,检测gsh含量,具体实验对比数据见表7。表7野生型gshf与gshf-4固定化酶转化应用实验对比酶反应温度反应时间gsh的浓度gshf-wt37℃120分钟7.2g/lgshf-437℃90分钟16.1g/lgshf-wt50℃100分钟5.7g/lgshf-450℃60分钟21g/l结果表明,野生型的gshf固定化酶,在37℃,反应120分钟,所含gsh浓度仅为7.2g/l,这是由于随着反应的进行,gsh的浓度慢慢增加,高浓度的gsh对野生型gshf有较强的反馈抑制,使其不能反应完全;在50℃,反应80分钟,所含gsh浓度为5.7g/l,增加反应时间,gsh浓度不会增加,这是因为在50℃条件下,野生型gshf易失活且受gsh的反馈抑制作用,导致反应不能继续进行。gshf-4突变体固定酶,在37℃,反应90分钟,gsh的产量为16.1g/l;在50℃,其反应时间从90分钟缩短到60分钟,且gsh的浓度可以达到21g/l;这是由于gshf-4突变体降低了产物gsh的反馈抑制作用,同时其热稳定性显著提高使得该反应能更快更好的进行,从而得到更多的gsh。3-2突变体gshf-4固定化酶操作稳定性批次实验由表7可知,gshf-4固定化酶在50℃反应的效果比在37℃的效果更好,因此,为了更好地反映该突变体的稳定性,本发明人实验过程中将其反应条件设定如下:gshf-4固定化酶(280u/g)总投量为5000u,反应ph为8.0,反应温度为50℃,反应体积为1000ml,参照图3,具体实验结果如表8所示。表8突变体gshf-4固定化酶操作稳定性批次实验批次反应时间残余活力批次反应时间残余活力160min280u/g20068min231u/g5061min275u/g25072min213u/g10063min263u/g30080min191u/g15065min249u/g40090min151u/g通过gshf-4固定化酶操作稳定性批次实验可知,本发明制备的gshf-4固定化酶经过连续300批次转化实验,其反应时间没有明显延长,固定化酶活力没有明显下降,显示出本发明制备的gshf-4固定化酶具有良好的操作稳定性。以上所述仅为本发明的优选实施例而已,并不用于限制本发明,对于本领域的技术人员来说,本发明可以有各种更改和变化。凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。序列表<110>湖南福来格生物技术有限公司<120>一种双功能谷胱甘肽合成酶突变体及其制备方法和应用<130>2017429<160>16<170>patentinversion3.3<210>1<211>2262<212>dna<213>优化后的唾液链球菌(streptococcussalivarius)野生型gshf编码基因<400>1atgactatcaatcaactgctgcaaaaactggaaccgacttctccgattctgcaggccaac60ttcggcatcgaacgtgagtccctgcgtgttgatcgtcagggtaaactggcacacactccg120cacccgtcttgcctgggtgctcgttcttttcatccgtacattcagaccgacttctgtgag180ttccagatggagctgatcactccggtagcgaaatccaccaccgaagcgcgtcgcttcctg240ggtgccatcaccgacgttgcaggtcgttctatctccaaagacgagctgctgtggccactg300tctatgccaccacgtatcaaagcccaagagatccaggttgcacagctggagaacgagttc360gaacgtcactaccgtaactacctggcggagaagtatggtaccaaactgcaggcaatttcc420ggcatccactacaacatggaactgggcaaagacctggttgaagctctgttcaaagaaagc480aaccaaaccgacatgatcgccttcaaaaacgcactgtacctgaaactggcgcaaaactac540ctgcgttaccgctgggttatcacctatctgtttggcgcagcgccaattgccgagcagggt600ttcttcgaccaagaggttccggaaccggtgcgcagcttccgtaactctgatcacggttat660gtgaacaaggaggagatccaggtgagctttgcatctctggaggactatgttagcgcgatt720gaaaactacatcgagcagggtgacctgattgcagaaaaagagttctattctgcggtacgt780ttccgtggtcagaaagtgaaccgtagcttcctggataaaggcattacctatctggagttc840cgcaactttgatctgaatccattcgaacgtattggtatcagccagaccaccatggacacc900gttcacctgctgctgctggcgttcctgtggatggatgctccggagaacgtggaccaagcg960ctggcacagggtcatgcactgaacgagaaaatcgccctgagccacccgctggaaccgctg1020ccatctgaggcggaaacccagaacatcactactgcgctggaccagctggtgcagcatttc1080ggtctgggcgattatcatcagggcctggttaaacaggttaaggatgcattcgcggacccg1140tcccacaccctggctgcacaactgctgagctacatcaaagacaaaagcctggccgatttc1200gcgctggacaaagctctggcgtatcacgattatgactggaccgctcactacgcgctgaaa1260ggctacgaggagatggaactgagcactcagatgctgctgtttgacgcgatccagaaaggc1320atccacttcgagattctggatgaacaggatcagtttctgaaactgtggcataaagatcat1380gttgagtacgtgaaaaacggtaacatgacctccaaagacaactacgtggtaccgctggct1440atggcgaacaaaaccgtaaccaagaaaattctggcagacgcaggttttccggtaccggca1500ggcgacgagtttaccagcctggaacaaggtctggcgtattacccactgatcaaagacaag1560cagatcgttgtgaaaccgaaatctactaacttcggtctgggtatctccatctttcaggag1620ccagcttctctggacaactataaaaaggctctggaaattgcctttgctgaggatactgcg1680gttctggttgaagagttcattccaggcaccgagtatcgcttctttattctggatggtcgc1740tgtgaggcggtgctgctgcgcgtggctgctaacgtagttggcgacggtaaacacactatc1800cgcgaactggtggcacagaagaacgcgaacccgctgcgtggtcgtgatcaccgttcccca1860ctggaaatcatcgcactgggcgatattgagcaactgatgctgactcagcagggctacact1920ccggacgacatcctgccggaaggcaaaaaggttaatctgcgtcgcaactccaacatctcc1980accggtggtgatagcattgacgtgaccgaaaccatggattccagctaccaggaactggca2040gccgcaatggcaacttctatgggtgcttgggcgtgtggcgttgacctgatcattccggac2100gaaacccagccagcatctaaggagaacccacactgtacctgcattgaactgaacttcaat2160ccatctatgtacatgcacacttattgcgctgaaggtccgggtcaagccatcacttctaaa2220atcctggacaaactgttcccggaagttgctaccaatcagaac2262<210>2<211>754<212>prt<213>优化后的唾液链球菌(streptococcussalivarius)野生型gshf氨基酸序列<400>2metthrileasnglnleuleuglnlysleugluprothrserproile151015leuglnalaasnpheglyilegluarggluserleuargvalasparg202530glnglylysleualahisthrprohisprosercysleuglyalaarg354045serphehisprotyrileglnthraspphecysglupheglnmetglu505560leuilethrprovalalalysserthrthrglualaargargpheleu65707580glyalailethraspvalalaglyargserileserlysaspgluleu859095leutrpproleusermetproproargilelysalaglngluilegln100105110valalaglnleugluasngluphegluarghistyrargasntyrleu115120125alaglulystyrglythrlysleuglnalaileserglyilehistyr130135140asnmetgluleuglylysaspleuvalglualaleuphelysgluser145150155160asnglnthraspmetilealaphelysasnalaleutyrleulysleu165170175alaglnasntyrleuargtyrargtrpvalilethrtyrleuphegly180185190alaalaproilealagluglnglyphepheaspglngluvalproglu195200205provalargserpheargasnserasphisglytyrvalasnlysglu210215220gluileglnvalserphealaserleugluasptyrvalseralaile225230235240gluasntyrilegluglnglyaspleuilealaglulysgluphetyr245250255seralavalargpheargglyglnlysvalasnargserpheleuasp260265270lysglyilethrtyrleuglupheargasnpheaspleuasnprophe275280285gluargileglyileserglnthrthrmetaspthrvalhisleuleu290295300leuleualapheleutrpmetaspalaprogluasnvalaspglnala305310315320leualaglnglyhisalaleuasnglulysilealaleuserhispro325330335leugluproleuproserglualagluthrglnasnilethrthrala340345350leuaspglnleuvalglnhispheglyleuglyasptyrhisglngly355360365leuvallysglnvallysaspalaphealaaspproserhisthrleu370375380alaalaglnleuleusertyrilelysasplysserleualaaspphe385390395400alaleuasplysalaleualatyrhisasptyrasptrpthralahis405410415tyralaleulysglytyrgluglumetgluleuserthrglnmetleu420425430leupheaspalaileglnlysglyilehisphegluileleuaspglu435440445glnaspglnpheleulysleutrphislysasphisvalglutyrval450455460lysasnglyasnmetthrserlysaspasntyrvalvalproleuala465470475480metalaasnlysthrvalthrlyslysileleualaaspalaglyphe485490495provalproalaglyaspgluphethrserleugluglnglyleuala500505510tyrtyrproleuilelysasplysglnilevalvallysprolysser515520525thrasnpheglyleuglyileserilepheglngluproalaserleu530535540aspasntyrlyslysalaleugluilealaphealagluaspthrala545550555560valleuvalgluglupheileproglythrglutyrargphepheile565570575leuaspglyargcysglualavalleuleuargvalalaalaasnval580585590valglyaspglylyshisthrilearggluleuvalalaglnlysasn595600605alaasnproleuargglyargasphisargserproleugluileile610615620alaleuglyaspilegluglnleumetleuthrglnglnglytyrthr625630635640proaspaspileleuprogluglylyslysvalasnleuargargasn645650655serasnileserthrglyglyaspserileaspvalthrgluthrmet660665670aspsersertyrglngluleualaalaalametalathrsermetgly675680685alatrpalacysglyvalaspleuileileproaspgluthrglnpro690695700alaserlysgluasnprohiscysthrcysilegluleuasnpheasn705710715720prosermettyrmethisthrtyrcysalagluglyproglyglnala725730735ilethrserlysileleuasplysleupheprogluvalalathrasn740745750glnasn<210>3<211>2262<212>dna<213>双功能谷胱甘肽合成酶gshf-2突变体编码基因<400>3atgactctgaatcaactgctgcaaaaactggaaccgacttctccgattctgcaggccaac60ttcggcatcgaacgtgagtccctgcgtgttgatcgtcagggtaaactggcacacactccg120cacccgtcttgcctgggtgctcgttcttttcatccgtacattcagaccgacttctgtgag180ttccagatggagctgatcactccggtagcgaaatccaccaccgaagcgcgtcgcttcctg240ggtgccatcaccgacgttgcaggtcgttctatctccaaagacgagctgctgtggccactg300tctatgccaccacgtatcaaagcccaagagatccaggttgcacagctggagaacgagttc360gaacgtcactaccgtaactacctggcggagaagtatggtaccaaactgcaggcaatttcc420ggcatccactacaacatggaactgggcaaagacctggttgaagctctgttcaaagaaagc480gatcaaaccgacatgatcgccttcaaaaacgcactgtacctgaaactggcgcaaaactac540ctgcgttaccgctgggttatcacctatctgtttggcgcagcgccaattgccgagcagggt600ttcttcgaccaagaggttccggaaccggtgcgcagcttccgtaactctgatcacggttat660gtgaacaaggaggagatccaggtgagctttgcatctctggaggactatgttagcgcgatt720gaaaactacatcgagcagggtgacctgattgcagaaaaagagttctattctgcggtacgt780ttccgtggtcagaaagtgaaccgtagcttcctggataaaggcattacctatctggagttc840cgcaactttgatctgaatccattcgaacgtattggtatcagccagaccaccatggacacc900gttcacctgctgctgctggcgttcctgtggatggatgctccggagaacgtggaccaagcg960ctggcacagggtcatgcactgaacgagaaaatcgccctgagccacccgctggaaccgctg1020ccatctgaggcggaaacccagaacatcactactgcgctggaccagctggtgcagcatttc1080ggtctgggcgattatcatcagggcctggttaaacaggttaaggatgcattcgcggacccg1140tcccacaccctggctgcacaactgctgagctacatcaaagacaaaagcctggccgatttc1200gcgctggacaaagctctggcgtatcacgattatgactggaccgctcactacgcgctgaaa1260ggctacgaggagatggaactgagcactcagatgctgctgtttgacgcgatccagaaaggc1320atccacttcgagattctggatgaacaggatcagtttctgaaactgtggcataaagatcat1380gttgagtacgtgaaaaacggtaacatgacctccaaagacaactacgtggtaccgctggct1440atggcgaacaaaaccgtaaccaagaaaattctggcagacgcaggttttccggtaccggca1500ggcgacgagtttaccagcctggaacaaggtctggcgtattacccactgatcaaagacaag1560cagatcgttgtgaaaccgaaatctactaacttcggtctgggtatctccatctttcaggag1620ccagcttctctggacaactataaaaaggctctggaaattgcctttgctgaggatactgcg1680gttctggttgaagagttcattccaggcaccgagtatcgcttctttattctggatggtcgc1740tgtgaggcggtgctgctgcgcgtggctgctaacgtagttggcgacggtaaacacactatc1800cgcgaactggtggcacagaagaacgcgaacccgctgcgtggtcgtgatcaccgttcccca1860ctggaaatcatcgcactgggcgatattgagcaactgatgctgactcagcagggctacact1920ccggacgacatcctgccggaaggcaaaaaggttaatctgcgtcgcaactccaacatctcc1980accggtggtgatagcattgacgtgaccgaaaccatggattccagctaccaggaactggca2040gccgcaatggcaacttctatgggtgcttgggcgtgtggcgttgacctgatcattccggac2100gaaacccagccagcatctaaggagaacccacactgtacctgcattgaactgaacttcaat2160ccatctatgtacatgcacacttattgcgctgaaggtccgggtcaagccatcacttctaaa2220atcctggacaaactgttcccggaagttgctaccaatcagaac2262<210>4<211>754<212>prt<213>双功能谷胱甘肽合成酶gshf-2突变体氨基酸序列<400>4metthrleuasnglnleuleuglnlysleugluprothrserproile151015leuglnalaasnpheglyilegluarggluserleuargvalasparg202530glnglylysleualahisthrprohisprosercysleuglyalaarg354045serphehisprotyrileglnthraspphecysglupheglnmetglu505560leuilethrprovalalalysserthrthrglualaargargpheleu65707580glyalailethraspvalalaglyargserileserlysaspgluleu859095leutrpproleusermetproproargilelysalaglngluilegln100105110valalaglnleugluasngluphegluarghistyrargasntyrleu115120125alaglulystyrglythrlysleuglnalaileserglyilehistyr130135140asnmetgluleuglylysaspleuvalglualaleuphelysgluser145150155160aspglnthraspmetilealaphelysasnalaleutyrleulysleu165170175alaglnasntyrleuargtyrargtrpvalilethrtyrleuphegly180185190alaalaproilealagluglnglyphepheaspglngluvalproglu195200205provalargserpheargasnserasphisglytyrvalasnlysglu210215220gluileglnvalserphealaserleugluasptyrvalseralaile225230235240gluasntyrilegluglnglyaspleuilealaglulysgluphetyr245250255seralavalargpheargglyglnlysvalasnargserpheleuasp260265270lysglyilethrtyrleuglupheargasnpheaspleuasnprophe275280285gluargileglyileserglnthrthrmetaspthrvalhisleuleu290295300leuleualapheleutrpmetaspalaprogluasnvalaspglnala305310315320leualaglnglyhisalaleuasnglulysilealaleuserhispro325330335leugluproleuproserglualagluthrglnasnilethrthrala340345350leuaspglnleuvalglnhispheglyleuglyasptyrhisglngly355360365leuvallysglnvallysaspalaphealaaspproserhisthrleu370375380alaalaglnleuleusertyrilelysasplysserleualaaspphe385390395400alaleuasplysalaleualatyrhisasptyrasptrpthralahis405410415tyralaleulysglytyrgluglumetgluleuserthrglnmetleu420425430leupheaspalaileglnlysglyilehisphegluileleuaspglu435440445glnaspglnpheleulysleutrphislysasphisvalglutyrval450455460lysasnglyasnmetthrserlysaspasntyrvalvalproleuala465470475480metalaasnlysthrvalthrlyslysileleualaaspalaglyphe485490495provalproalaglyaspgluphethrserleugluglnglyleuala500505510tyrtyrproleuilelysasplysglnilevalvallysprolysser515520525thrasnpheglyleuglyileserilepheglngluproalaserleu530535540aspasntyrlyslysalaleugluilealaphealagluaspthrala545550555560valleuvalgluglupheileproglythrglutyrargphepheile565570575leuaspglyargcysglualavalleuleuargvalalaalaasnval580585590valglyaspglylyshisthrilearggluleuvalalaglnlysasn595600605alaasnproleuargglyargasphisargserproleugluileile610615620alaleuglyaspilegluglnleumetleuthrglnglnglytyrthr625630635640proaspaspileleuprogluglylyslysvalasnleuargargasn645650655serasnileserthrglyglyaspserileaspvalthrgluthrmet660665670aspsersertyrglngluleualaalaalametalathrsermetgly675680685alatrpalacysglyvalaspleuileileproaspgluthrglnpro690695700alaserlysgluasnprohiscysthrcysilegluleuasnpheasn705710715720prosermettyrmethisthrtyrcysalagluglyproglyglnala725730735ilethrserlysileleuasplysleupheprogluvalalathrasn740745750glnasn<210>5<211>2262<212>dna<213>双功能谷胱甘肽合成酶gshf-3突变体编码基因<400>5atgactctgaatcaactgctgcaaaaactggaaccgacttctccgattctgcaggccaac60ttcggcatcgaacgtgagtccctgcgtgttgatcgtcagggtaaactggcacacactccg120cacccgtcttgcctgggtgctcgttcttttcatccgtacattcagaccgacttctgtgag180ttccagatggagctgatcactccggtagcgaaatccaccaccgaagcgcgtcgcttcctg240ggtgccatcaccgacgttgcaggtcgttctatctccaaagacgagctgctgtggccactg300tctatgccaccacgtatcaaagcccaagagatccaggttgcacagctggagaacgagttc360gaacgttattaccgtaactacctggcggagaagtatggtaccaaactgcaggcaatttcc420ggcatccactacaacatggaactgggcaaagacctggttgaagctctgttcaaagaaagc480gatcaaaccgacatgatcgccttcaaaaacgcactgtacctgaaactggcgcaaaactac540ctgcgttaccgctgggttatcacctatctgtttggcgcatctccaattgccgagcagggt600ttcttcgaccaagaggttccggaaccggtgcgcagcttccgtaactctgatcacggttat660gtgaacaaggaggagatccaggtgagctttgcatctctggaggactatgttagcgcgatt720gaaaactacatcgagcagggtgacctgattgcagaaaaagagttctattctgcggtacgt780ttccgtggtcagaaagtgaaccgtagcttcctggataaaggcattacctatctggagttc840cgcaactttgatctgaatccattcgaacgtattggtatcagccagaccaccatggacacc900gttcacctgctgctgctggcgttcctgtggatggatgctccggagaacgtggaccaagcg960ctggcacagggtcatgcactgaacgagaaaatcgccctgagccacccgctggaaccgctg1020ccatctgaggcggaaacccagaacatcactactgcgctggaccagctggtgcagcatttc1080ggtctgggcgattatcatcagggcctggttaaacaggttaaggatgcattcgcggacccg1140tcccacaccctggctgcacaactgctgagctacatcaaagacaaaagcctggccgatttc1200gcgctggacaaagctctggcgtatcacgattatgactggaccgctcactacgcgctgaaa1260ggctacgaggagatggaactgagcactcagatgctgctgtttgacgcgatccagaaaggc1320atccacttcgagattctggatgaacaggatcagtttctgaaactgtggcataaagatcat1380gttgagtacgtgaaaaacggtaacatgacctccaaagacaactacgtggtaccgctggct1440atggcgaacaaaaccgtaaccaagaaaattctggcagacgcaggttttccggtaccggca1500ggcgacgagtttaccagcctggaacaaggtctggcgtattacccactgatcaaagacaag1560cagatcgttgtgaaaccgaaatctactaacttcggtctgggtatctccatctttcaggag1620ccagcttctctggacaactataaaaaggctctggaaattgcctttgctgaggatactgcg1680gttctggttgaagagttcattccaggcaccgagtatcgcttctttattctggatggtcgc1740tgtgaggcggtgctgctgcgcgtggctgctaacgtagttggcgacggtaaacacactatc1800cgcgaactggtggcacagaagaacgcgaacccgctgcgtggtcgtgatcaccgttcccca1860ctggaaatcatcgcactgggcgatattgagcaactgatgctgactcagcagggctacact1920ccggacgacatcctgccggaaggcaaaaaggttaatctgcgtcgcaactccaacatctcc1980accggtggtgatagcattgacgtgaccgaaaccatggattccagctaccaggaactggca2040gccgcaatggcaacttctatgggtgcttgggcgtgtggcgttgacctgatcattccggac2100gaaacccagccagcatctaaggagaacccacactgtacctgcattgaactgaacttcaat2160ccatctatgtacatgcacacttattgcgctgaaggtccgggtcaagccatcacttctaaa2220atcctggacaaactgttcccggaagttgctaccaatcagaac2262<210>6<211>754<212>prt<213>双功能谷胱甘肽合成酶gshf-2突变体氨基酸序列<400>6metthrleuasnglnleuleuglnlysleugluprothrserproile151015leuglnalaasnpheglyilegluarggluserleuargvalasparg202530glnglylysleualahisthrprohisprosercysleuglyalaarg354045serphehisprotyrileglnthraspphecysglupheglnmetglu505560leuilethrprovalalalysserthrthrglualaargargpheleu65707580glyalailethraspvalalaglyargserileserlysaspgluleu859095leutrpproleusermetproproargilelysalaglngluilegln100105110valalaglnleugluasngluphegluargtyrtyrargasntyrleu115120125alaglulystyrglythrlysleuglnalaileserglyilehistyr130135140asnmetgluleuglylysaspleuvalglualaleuphelysgluser145150155160aspglnthraspmetilealaphelysasnalaleutyrleulysleu165170175alaglnasntyrleuargtyrargtrpvalilethrtyrleuphegly180185190alaserproilealagluglnglyphepheaspglngluvalproglu195200205provalargserpheargasnserasphisglytyrvalasnlysglu210215220gluileglnvalserphealaserleugluasptyrvalseralaile225230235240gluasntyrilegluglnglyaspleuilealaglulysgluphetyr245250255seralavalargpheargglyglnlysvalasnargserpheleuasp260265270lysglyilethrtyrleuglupheargasnpheaspleuasnprophe275280285gluargileglyileserglnthrthrmetaspthrvalhisleuleu290295300leuleualapheleutrpmetaspalaprogluasnvalaspglnala305310315320leualaglnglyhisalaleuasnglulysilealaleuserhispro325330335leugluproleuproserglualagluthrglnasnilethrthrala340345350leuaspglnleuvalglnhispheglyleuglyasptyrhisglngly355360365leuvallysglnvallysaspalaphealaaspproserhisthrleu370375380alaalaglnleuleusertyrilelysasplysserleualaaspphe385390395400alaleuasplysalaleualatyrhisasptyrasptrpthralahis405410415tyralaleulysglytyrgluglumetgluleuserthrglnmetleu420425430leupheaspalaileglnlysglyilehisphegluileleuaspglu435440445glnaspglnpheleulysleutrphislysasphisvalglutyrval450455460lysasnglyasnmetthrserlysaspasntyrvalvalproleuala465470475480metalaasnlysthrvalthrlyslysileleualaaspalaglyphe485490495provalproalaglyaspgluphethrserleugluglnglyleuala500505510tyrtyrproleuilelysasplysglnilevalvallysprolysser515520525thrasnpheglyleuglyileserilepheglngluproalaserleu530535540aspasntyrlyslysalaleugluilealaphealagluaspthrala545550555560valleuvalgluglupheileproglythrglutyrargphepheile565570575leuaspglyargcysglualavalleuleuargvalalaalaasnval580585590valglyaspglylyshisthrilearggluleuvalalaglnlysasn595600605alaasnproleuargglyargasphisargserproleugluileile610615620alaleuglyaspilegluglnleumetleuthrglnglnglytyrthr625630635640proaspaspileleuprogluglylyslysvalasnleuargargasn645650655serasnileserthrglyglyaspserileaspvalthrgluthrmet660665670aspsersertyrglngluleualaalaalametalathrsermetgly675680685alatrpalacysglyvalaspleuileileproaspgluthrglnpro690695700alaserlysgluasnprohiscysthrcysilegluleuasnpheasn705710715720prosermettyrmethisthrtyrcysalagluglyproglyglnala725730735ilethrserlysileleuasplysleupheprogluvalalathrasn740745750glnasn<210>7<211>2262<212>dna<213>双功能谷胱甘肽合成酶gshf-4突变体编码基因<400>7atgactctgaatcaactgctgcaaaaactggaaccgacttctccgattctgcaggccaac60ttcggcatcgaacgtgagtccctgcgtgttgatcgtcagggtaaactggcacacactccg120cacccgtcttgcctgggtgctcgttcttttcatccgtacattcagaccgacttctgtgag180ttccagatggagctgatcactccggtagcgaaatccaccaccgaagcgcgtcgcttcctg240ggtgccatcaccgacgttgcaggtcgttctatctccaaagacgagctgctgtggccactg300tctatgccaccacgtatcaaagcccaagagatccaggttgcacagctggagaacgagttc360gaacgttattaccgtaactacctggcggagaagtatggtaccaaactgcaggcaatttcc420ggcatccactacaacatggaactgggcaaagacctggttgaagctctgttcaaagaaagc480gatcaaaccgacatgatcgccttcaaaaacgcactgtacctgaaactggcgcaaaactac540ctgcgttaccgctgggttatcacctatctgtttggcgcatctccaattgccgagcagggt600ttcttcgaccaagaggttccggaaccggtgcgcagcttccgtaactctgatcacggttat660gtgaacaaggaggagatccaggtgagctttgcatctctggaggactatgttagcgcgatt720gaaaactacatcgagcagggtgacctgattgcagaaaaagagttctattctgcggtacgt780ttccgtggtcagaaagtgaaccgtagcttcctggataaaggcattacctatctggagttc840cgcaactttgatctgaatccattcgaacgtattggtatcagccagaccaccatggacacc900gttcacctgctgctgctggcgttcctgtggatggatgctccggagaacgtggaccaagcg960ctggcacagggtcatgcactgaacgagaaaatcgccctgagccacccgctggaaccgctg1020ccatctgaggcggaaacccagaacatcactactgcgctggaccagctggtgcagcatttc1080ggtctgggcgattatcatcagggcctggttaaacaggttaaggatgcattcgcggacccg1140tcccagaccctggctgcacaactgctgccgtacatcaaagacaaaagcctggccgatttc1200gcgctggacaaagctctggcgtatcacgattatgactggaccgctcactacgcgctgaaa1260ggctacgaggagatggaactgagcactcagatgctgctgtttgacgcgatccagaaaggc1320atccacttcgagattctggatgaacaggatcagtttctgaaactgtggcataaagatcat1380gttgagtacgtgaaaaacggtaacatgacctccaaagacaactacgtggtaccgctggct1440atggcgaacaaaaccgtaaccaagaaaattctggcagacgcaggttttccggtaccggca1500ggcgacgagtttaccagcctggaacaaggtctggcgtattacccactgatcaaagacaag1560cagatcgttgtgaaaccgaaatctactaacttcggtctgggtatctccatctttcaggag1620ccagcttctctggacaactataaaaaggctctggaaattgcctttgctgaggatactgcg1680gttctggttgaagagttcattccaggcaccgagtatcgcttctttattctggatggtcgc1740tgtgaggcggtgctgctgcgcgtggctgctaacgtagttggcgacggtaaacacactatc1800cgcgaactggtggcacagaagaacgcgaacccgctgcgtggtcgtgatcaccgttcccca1860ctggaaatcatcgcactgggcgatattgagcaactgatgctgactcagcagggctacact1920ccggacgacatcctgccggaaggcaaaaaggttaatctgcgtcgcaactccaacatctcc1980accggtggtgatagcattgacgtgaccgaaaccatggattccagctaccaggaactggca2040gccgcaatggcaacttctatgggtgcttgggcgtgtggcgttgacctgatcattccggac2100gaaacccagccagcatctaaggagaacccacactgtacctgcattgaactgaacttcaat2160ccatctatgtacatgcacacttattgcgctgaaggtccgggtcaagccatcacttctaaa2220atcctggacaaactgttcccggaagttgctaccaatcagaac2262<210>8<211>754<212>prt<213>双功能谷胱甘肽合成酶gshf-4突变体氨基酸序列<400>8metthrleuasnglnleuleuglnlysleugluprothrserproile151015leuglnalaasnpheglyilegluarggluserleuargvalasparg202530glnglylysleualahisthrprohisprosercysleuglyalaarg354045serphehisprotyrileglnthraspphecysglupheglnmetglu505560leuilethrprovalalalysserthrthrglualaargargpheleu65707580glyalailethraspvalalaglyargserileserlysaspgluleu859095leutrpproleusermetproproargilelysalaglngluilegln100105110valalaglnleugluasngluphegluargtyrtyrargasntyrleu115120125alaglulystyrglythrlysleuglnalaileserglyilehistyr130135140asnmetgluleuglylysaspleuvalglualaleuphelysgluser145150155160aspglnthraspmetilealaphelysasnalaleutyrleulysleu165170175alaglnasntyrleuargtyrargtrpvalilethrtyrleuphegly180185190alaserproilealagluglnglyphepheaspglngluvalproglu195200205provalargserpheargasnserasphisglytyrvalasnlysglu210215220gluileglnvalserphealaserleugluasptyrvalseralaile225230235240gluasntyrilegluglnglyaspleuilealaglulysgluphetyr245250255seralavalargpheargglyglnlysvalasnargserpheleuasp260265270lysglyilethrtyrleuglupheargasnpheaspleuasnprophe275280285gluargileglyileserglnthrthrmetaspthrvalhisleuleu290295300leuleualapheleutrpmetaspalaprogluasnvalaspglnala305310315320leualaglnglyhisalaleuasnglulysilealaleuserhispro325330335leugluproleuproserglualagluthrglnasnilethrthrala340345350leuaspglnleuvalglnhispheglyleuglyasptyrhisglngly355360365leuvallysglnvallysaspalaphealaaspproserglnthrleu370375380alaalaglnleuleuprotyrilelysasplysserleualaaspphe385390395400alaleuasplysalaleualatyrhisasptyrasptrpthralahis405410415tyralaleulysglytyrgluglumetgluleuserthrglnmetleu420425430leupheaspalaileglnlysglyilehisphegluileleuaspglu435440445glnaspglnpheleulysleutrphislysasphisvalglutyrval450455460lysasnglyasnmetthrserlysaspasntyrvalvalproleuala465470475480metalaasnlysthrvalthrlyslysileleualaaspalaglyphe485490495provalproalaglyaspgluphethrserleugluglnglyleuala500505510tyrtyrproleuilelysasplysglnilevalvallysprolysser515520525thrasnpheglyleuglyileserilepheglngluproalaserleu530535540aspasntyrlyslysalaleugluilealaphealagluaspthrala545550555560valleuvalgluglupheileproglythrglutyrargphepheile565570575leuaspglyargcysglualavalleuleuargvalalaalaasnval580585590valglyaspglylyshisthrilearggluleuvalalaglnlysasn595600605alaasnproleuargglyargasphisargserproleugluileile610615620alaleuglyaspilegluglnleumetleuthrglnglnglytyrthr625630635640proaspaspileleuprogluglylyslysvalasnleuargargasn645650655serasnileserthrglyglyaspserileaspvalthrgluthrmet660665670aspsersertyrglngluleualaalaalametalathrsermetgly675680685alatrpalacysglyvalaspleuileileproaspgluthrglnpro690695700alaserlysgluasnprohiscysthrcysilegluleuasnpheasn705710715720prosermettyrmethisthrtyrcysalagluglyproglyglnala725730735ilethrserlysileleuasplysleupheprogluvalalathrasn740745750glnasn<210>9<211>38<212>dna<213>第3位和第161位氨基酸双点迭代饱和突变引物p1<220><221>misc_feature<222>(10)..(11)<223>nisa,c,g,ort<400>9ttcatgactnnkaatcaactgctgcaaaaactggaacc38<210>10<211>39<212>dna<213>第3位和第161位氨基酸双点迭代饱和突变引物p2<220><221>misc_feature<222>(11)..(12)<223>nisa,c,g,ort<400>10gtcggtttgmnngctttctttgaacagagcttcaaccag39<210>11<211>40<212>dna<213>第123位点和第194位点氨基酸双点迭代饱和突变引物p3<220><221>misc_feature<222>(10)..(11)<223>nisa,c,g,ort<400>11ttcgaacgtnnktaccgtaactacctggcggagaagtatg40<210>12<211>40<212>dna<213>第123位点和第194位点氨基酸双点迭代饱和突变引物p4<220><221>misc_feature<222>(11)..(12)<223>nisa,c,g,ort<400>12ggcaattggmnntgcgccaaacagataggtgataacccag40<210>13<211>54<212>dna<213>第382位和第390位氨基酸双点饱和突变引物p5<220><221>misc_feature<222>(10)..(11)<223>nisa,c,g,ort<220><221>misc_feature<222>(34)..(35)<223>nisa,c,g,ort<400>13gacccgtccnnkaccctggctgcacaactgctgnnktacatcaaagacaaaagc54<210>14<211>54<212>dna<213>第382位和第390位氨基酸双点饱和突变引物p6<220><221>misc_feature<222>(11)..(12)<223>nisa,c,g,ort<220><221>misc_feature<222>(35)..(36)<223>nisa,c,g,ort<400>14tttgatgtamnncagcagttgtgcagccagggtmnnggacgggtccgcgaatgc54<210>15<211>20<212>dna<213>t7promoterprimer<400>15taatacgactcactataggg20<210>16<211>19<212>dna<213>t7terminatorprimer<400>16gctagttattgctcagcgg19当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1