一种CMKLR1拮抗多肽及其衍生物与应用的制作方法
本发明涉及生物技术和生物医药领域,具体而言,本发明是雌性生殖疾病靶点cmklr1受体拮抗多肽lrh12-c5及其衍生物与应用。
背景技术:
雌性生殖癌症中,有的与内分泌相关。如,乳腺癌(breastcancer),子宫内膜癌(endometrialcancer)和卵巢癌(ovariancancer,oac)等。其中,卵巢癌是发生于卵巢的一种恶性肿瘤,90%~95%为卵巢原发性的癌,另外5%~10%为其它部位原发的癌转移到卵巢。虽然我国的卵巢癌发病率不如欧美国家高,但根除性的手术治疗,以及细胞毒性的化疗,均缺乏有效降低卵巢癌的死亡率的效用。由于卵巢癌早期缺少症状,即使有症状也不特异,筛查的作用又有限,因此早期诊断比较困难,就诊时60%~70%已为晚期,而晚期病例又疗效不佳。因此,虽然卵巢癌的发病率仅次于宫颈癌和子宫内膜癌,居妇科恶性肿瘤的第三位,但死亡率却超过宫颈癌及子宫内膜癌之和,高居妇科癌症首位,是严重威胁妇女健康的最大疾患。
在以往的临床研究中认为,卵巢癌在早期是没有明显的症状表现。但也一些研究表明,卵巢癌在早期是会表现出一些临床的症状,例如异常的腹胀、饱胀,腹部疼痛或背部疼痛,精神不振等,但这些症状往往被患者所忽视,当诊断时往往已发生转移,因而卵巢癌往往被形容为“无声的杀手”,进而也导致了卵巢癌的预后不良。
大量研究已表明,卵巢癌常通过直接蔓延及盆、腹腔播散种植到远处的器官。卵巢癌转移的主要途径是腹腔内种植性转移,据临床观察,转移的主要部位为大网膜,在患有卵巢癌的妇女中80%伴有网膜转移。此时,大网膜上的癌细胞以远快于原发病灶处的速度增长。大网膜是连接胃大弯至横结肠的腹膜,内含有吞噬细胞,有重要的防御功能,同时也是含有大量脂滴的脂肪组织,是体内一个重要的内分泌器官,并参与体内环境稳态。尽管卵巢癌向大网膜转移的事实很明显,但是其机理一直尚不明晰。因大网膜部位以脂肪组织为主,脂肪组织是体内最大的内分泌器官,可以分泌脂肪因子、细胞因子等,脂肪组织内的脂肪细胞与卵巢癌细胞之间的关系及其机制,仍需要更多的证据阐明。
随着对肿瘤研究的不断深入,越来越多的证据表明,肥胖会增加癌症患者致死的风险。比如,白色脂肪组织分泌的瘦素(leptin)能够促进乳腺癌的发展:一方面,瘦素能够通过激活jak/stat3、mapk-erk1/2或者pi3k通路,从而促进乳腺癌的生长;另外,瘦素还能通过诱导血管生成素的表达促进肿瘤相关血管的生成,并且瘦素还能诱导人表皮生长因子受体2(erbb-2)的转录,并且参与三阴性乳腺癌细胞中胰岛素样生长受体1(igf-1)的反应,激活表皮生长因子受体(egfr)从而促进细胞的侵袭与转移。目前的研究还表明,瘦素还能在前列腺癌、甲状腺癌等多种癌症过程中有着促进作用,其表达水平和肿瘤发生呈正相关,但胰腺癌中的瘦素水平则比较低,并且两者间的关系并不是很清楚。也有研究发现脂联素对肿瘤的发生发展具有抑制作用。
有报道称,肥胖也能增加女性患上卵巢癌的风险。脂肪组织会影响肥胖妇女的卵巢癌的病情,研究人员对216名妇女进行研究,其中35名是肥胖妇女,108名是正常体重的妇女,发现在卵巢癌患者中,与正常体重的妇女相比,肥胖妇女的存活率较低,存活时间较短。科学家发现除了他们之间的癌症致死率和癌症复发率有区别外,他们的肿瘤细胞也表现不同,提示脂肪组织分泌的激素或者蛋白可能引起卵巢癌细胞增殖迅速。还有研究表明,腹腔中大网膜的脂肪细胞分泌的il-6和il-8能够促进卵巢癌细胞向其转移。
关于脂肪组织,尤其是大网膜脂肪,与卵巢癌的关系,2011年10月30日的《自然—医学》(naturemedicine)杂志上ernstlengyel教授发表的一篇文章提供了详实的证据。chemerin是新近发现的一种脂肪细胞因子,又称chemerin,在免疫应答、炎症反应、脂肪细胞分化成熟、脂质代谢等方面发挥作用,与肥胖和代谢综合征相关。chemerin基因也称为他扎罗汀诱导基因2(tazaroteneinducedgene2,tig2),1997年被首次克隆发现,随后,2003年,wittamer等在卵巢癌继发的腹水中通过反相高压液相层析分离得到其活性蛋白。
2012年,reverchon等的研究证实chemerin及其受体cmklr1(类趋化因子受体-1)在主要的人卵巢颗粒细胞(hgcs)和人卵巢颗粒样肿瘤细胞(kgn)中有表达,再次提到chemerin与卵巢肿瘤的关系。
2011年10月,ernstlengyel博士发表在naturemedicine的研究发现卵巢癌在向大网膜转移过程中,fabp4起到了至关重要的作用,且大网膜的趋化因子参与了卵巢癌迁移至大网膜脂肪的过程,并检测到了大网膜脂肪细胞中的趋化因子脂联素和细胞因子il-6、il-8等。但文章中并没有记载检测到同样是趋化因子的chemerin。
脂肪因子chemerin
脂肪因子chemerin也称为他扎罗汀诱导基因2(tig2)或视黄醇受体反应蛋白2。在1997年被nagpal等克隆发现后,在2003年,wittamer等在卵巢癌继发的腹水中通过反相高压液相层析分离得到其活性蛋白。chemerin蛋白前体全长163个氨基酸,具有6个半胱氨酸残基,形成3个二硫键,去掉n端信号肽和c端的几个氨基酸,才具有生物活性,其血液中的chemerin活化形式含有134个氨基酸(约16kda),在结构上属于杀菌肽/半胱氨酸蛋白酶抑制剂家族,当发生炎症反应时,在几种蛋白酶作用下chemerin能迅速转变成活化形式。chemerin在人体的多种组织广泛表达,主要表达于白色脂肪组织、胎盘和肝脏,与3个受体相结合:①g蛋白偶联受体cmklr1;②g蛋白偶联受体gpr1;③ccrl-2。cmklr1在巨噬细胞、未成熟树突细胞和白色脂肪组织高表达,是chemerin发挥生物功能的主要受体,chemerin可以趋化高表达有cmklr1的树突状细胞和巨噬细胞,在免疫和适应性免疫之间起桥梁作用,在免疫应答、炎症反应、脂肪细胞分化成熟、脂质代谢等方面发挥作用,与肥胖和代谢综合征相关。chemerin结合cmklr1后,使cmklr1阳性细胞内的钙离子释放,抑制camp的聚集,使map激酶磷酸化。百日咳毒素的预先处理可以阻断cmklr1的细胞内信号转导,cmklr1的细胞内信号转导可能与gi族有关。chemerin一方面作为趋化因子,趋化树突状细胞和巨噬细胞,在免疫和适应性免疫之间起桥梁作用,趋化天然杀伤细胞到达炎症部位参与炎症反应;另一方面作为新的脂肪因子,由脂肪组织分泌产生,调节脂肪细胞的分化、脂解,可促进脂肪细胞内胰岛素信号传导途径等生物学效应。chemerin在肥胖和代谢综合征的病理生理机制中占重要的地位。
缓减素e1(resolvine1)
2002年,美国学者serhancn等研究表明epa和阿司匹林治疗可以减少小鼠急性炎症模型炎性渗出物中25%-60%中性粒细胞数量,他们使用液相色谱串联质谱法(lc-ms/ms)分析炎性渗出物,发现了新型含三羟基化合物(5,12,18-trihepe)以及从epa衍生出单羟基脂肪酸,18-羟基-epa(18-hepe)及5-hepe。当静脉给药时(100ng/鼠),这种新的三羟基-epa是中性粒细胞浸润的强效抑制剂。它具有缓减炎症的特性,被命名为rve1。基于其合成化合物的物理和生物属性通过完整的立体结构分析,rve1被确定为5s,12r,18r-羟基-6z,8e,10e,14z,16e-eicosapentaenoic。缓减素类化合物(rvs)是一组由epa或dha在血管内皮细胞内通过环氧化作用产生。根据所需底物不同,可分d、e等型,每型分为数类。rve1属于e族rvs,resolvine1为cmklr1另一个脂类配体,能够通过与cmklr1和白三烯b4受体blt1结合减轻炎症反应、减少树突状细胞的迁移和减少白介素12的产量。rve1可以通过阿司匹林依赖途径和非依赖阿司匹林途径合成。
rve1应刺激而生,在局部产生作用,并且通过酶迅速进一步代谢灭活。在哺乳动物组织中,rve1至少存在四个单独的途径代谢。rve1具有强烈的抗炎作用,它可以缓减白细胞介导的组织损伤和下调炎症基因过度表达。rve1是中性粒细胞重要的调节因子。在炎症模型中,包括酵母诱导性腹膜炎和2,4,6-三硝基苯诱导性结肠炎,毫微克剂量rve1就可以显著降低炎症部位中性粒细胞浸润。在cmklr1转染细胞rve1与cmklr1相互作用,抑制肿瘤坏死因子-α诱导的nf-κb活化。在体内rve1可以阻断dc迁移和il-12的产生。同时,rve1可以有效的抑制脂多糖(lps)刺激骨髓源树突状细胞(bmdcs)释放tnf-α和il-6和il-23。rve1可以阻断病理状态下过度血小板聚集,但不干扰胶原蛋白激发的生理凝血。rve1可诱导l-选择素脱落,下调人粒细胞和单核细胞表面分子cd18的表达。此外在小鼠体内,rve1可快速减少白细胞睾肌静脉上皮细胞滚动。rve1选择性诱导的上皮细胞表达抗粘附分子cd55的,从而促进依赖上皮细胞表面cd55的中性粒细胞清除。它可以阻止炎症细胞跨上皮和跨内皮迁移;促进巨噬细胞吞噬凋亡的中性粒细胞;下调树突状细胞白介素-12的分泌;上调t细胞ccr5的表达;调节白细胞粘附分子的表达减少白细胞滚动;选择性的阻断腺苷和血栓素受体激动剂u46619促进血小板聚集。已在兔牙周炎、小鼠腹膜炎、小鼠视网膜病变、结肠炎等多种疾病模型中展现出良好的预防和治疗效果。
cmklr1受体
cmklr1,也称chemr23。2003年,meder等通过反向药理学的方法证明chemerin是cmklr1的天然配体。人cmklr1基因定位于12号染色体上臂的23.3,有3个外显子和2个内含子组成。在获得性免疫系统高表达,其中在巨噬细胞、未成熟树突细胞(immaturedcs)、单核细胞、中性粒细胞、自然杀伤细胞均检测到高表达,但在成熟dcs的表达减少;加小板细胞、脂肪细胞、内皮细胞、口腔上皮细胞、破骨细胞、血管平滑肌细胞、肌细胞和神錢胶质瘤纳胞(dbtrg-05mg)同样表达cmklr1。chemerin/rve1与cmklr1结合激活后内吞,可促使细胞内钙(ca2+)离子的释放,使细胞外信号调节激酶1/2(erk1/2)磷酸化,通过结合g蛋白偶联的异源三聚体来抑制环磷酸腺苷(camp)的积聚;上调pi3k/akt信号通路,下调核转录因子κb(nfκb)信号通路:募集未成熟树突细胞和巨噬细胞到炎症位点,从而实现对各种代谢过程、炎性反应和癌症的调控作用。
越来越多的研究表明chemerin及其受体的信号异常与癌症发生和发展之间有着密切关联,chemerin与内皮细胞膜上的cmklr1受体结合后,还可诱导内皮细胞增殖和新生血管丛形成,对肿瘤细胞生长增殖起到促进作用。chemerin在大多数肿瘤内部的表达水平很低,有的甚至检测不到,但是在癌旁组织,及远端的正常组织中,chemerin的表达水平较高,比如:结肠直癌,肺癌(包括非小细胞肺癌),肝癌,黑色素瘤,胃癌以及卵巢癌。yu等发现chemerin在胃癌中通过激活p38和erk1/2信号通路,上调vegf,mmp-7和il-6蛋白水平,促进肿瘤的侵袭和迁移。2012年,reverchon等的研究证实chemerin和cmklr1在主要的人卵巢颗粒细胞(hgcs)和人卵巢颗粒样肿瘤细胞(kgn)中有表达。2011年10月,ernstlengyel博士发表在naturemedicine的研究发现卵巢癌在向大网膜转移过程中,fabp4起到了至关重要的作用,且大网膜的趋化因子参与了卵巢癌迁移至大网膜脂肪的过程,并检测到了大网膜脂肪细胞中的趋化因子脂联素和细胞因子il-6,il-8等,再一次提示:脂肪组织中的chemerin可能与卵巢癌细胞迁移有关。
g蛋白偶联受体超家族(gpcrs)在胞外信号向胞内转导过程中起到重要的作用,调控着细胞运动、生长和基因转录—这三个癌症生物学中至关重要的因素。过去的十几年中,介导的信号通路已经被证明是原癌基因信号的关键调控者,并且是很好的药物靶点,目前市面上出售的药物60%都是针对该受体设计的。而cmklr1作为gpcrs成员之一,目前仍然没有任何药物是直接针对cmklr1来治疗临床癌症的。因此,以cmklr1为靶标,筛选抗癌的新型多肽药物和人源化抗体具有重要的社会意义和广阔的经济市场。
噬菌体展示技术(phagedisplaytechnology)是一项特异性多肽或蛋白的筛选技术,此技术可将目的基因编码的多肽以融合蛋白的形式展示于噬菌体表面,被展示的多肽或蛋白可以保持相对独立的空间结构和生物活性,使大量随机多肽与其dna编码序列之间建立了直接联系,使得各种靶分子(如抗体、酶和细胞表面受体等)的多肽配体通过体外亲和淘选程序得以快速鉴定。噬菌体展示技术已被广泛用于肿瘤诊断标志物和抗肿瘤先导化合物的筛选、肿瘤特异性抗体和肿瘤药物靶向运输等方面的研究。
技术实现要素:
为了克服现有技术的不足和缺点,本发明的首要目的在于提供一种由噬菌体展示库筛选得到的cmklr1拮抗多肽,该拮抗多肽与chemerin/rve1受体cmklr1具有特异性高亲和力,能够通过抑制chemerin/rve1与cmklr1的结合来阻断chemerin/rve1-cmklr1的信号通路,证明该多肽在靶向抑制卵巢癌细胞增殖、促进卵巢癌细胞凋亡等方面起着重要的作用,其在卵巢癌靶向治疗方面具有巨大的应用价值。
本发明的另一目的在于提供上述cmklr1受体拮抗多肽的衍生物,该衍生物也能够与cmklr1受体具有特异性高亲和力,且特异性竞争chemerin/rve1与cmklr1的结合位点,能够抑制chemerin/rve1结合cmklr1。
本发明的再一目的在于提供上述cmklr1拮抗多肽及其衍生物的应用。
为了实现上述任务,本发明采取如下的技术解决方案:
本发明一个方面提供了一种cmklr1拮抗多肽,其特征在于,其氨基酸序列lrh12-c5为phe-tyr-ser-his-ser-met-pro-arg-leu-pro-pro-ala(fyshsmprlppa);其中所述的tyr为d构型,末端进行酰胺化修饰,即为phe-(d)tyr-ser-his-ser-met-pro-arg-leu-pro-pro-ala-nh2。
上述cmklr1拮抗多肽的筛选方法,该方法利用噬菌体随机肽库,首先采用cmklr1质粒转染293t细胞,获得永久高表达cmklr1的稳定细胞系,以野生型293t细胞为对照吸附细胞,进行5轮全细胞消减筛选,随机挑取50个阳性噬菌体扩增,提取克隆单链dna测序。分析多肽的氨基酸序列的基本特征,多肽同源性比较,检索出现频率高的多肽基序。blast检索蛋白质数据库,检测多肽基序同源性较高的蛋白质,发现含有大量该多肽的生物物种,及可能结合的细胞表面受体和配体,有利于后续大量提取和纯化获得多肽。
亲水性分析表明,lrh12-c5为亲水性多肽;
高效液相色谱法(hplc)和质谱(ms)检测lrh12-c5合成的纯度达到了99.85%;
本发明另一个方面提供了本发明所述的cmklr1拮抗多肽的衍生物为cmklr1拮抗多肽氨基酸侧链基团上、cmklr1拮抗多肽片段的氨基端或羧基端进行常规修饰得到的产物,或者为cmklr1拮抗多肽上连接用于多肽或蛋白检测或纯化的标签所得到的产物;
优选地,所述的常规修饰为氨基化、酰胺化、羟基化、羧基化、羰基化、烷基化、乙酰化、磷酸化、硫酸化、酯化、糖基化、环化、生物素化、荧光基团修饰、聚乙二醇peg修饰或固定化修饰等;
优选地,所述的标签为his6、gst、egfp、mbp、nus、ha、igg、flag、c-myc或profinityexact。
本发明再一个方面提供了本发明所述的cmklr1拮抗多肽的生物活性片段或类似物,其如seqidno.2-24所示:
fyx1x2x3x4x5rx6x7paseqidno.2,其中:
⑴x1为谷氨酸、苏氨酸、脯氨酸、缬氨酸(e、t、p、v);
⑵x3为天冬酰胺、天冬氨酸、脯氨酸(n、d、p);
⑶x6为亮氨酸或谷氨酰胺(l、q);
⑷x7为丝氨酸、苏氨酸、谷氨酰胺(s、t、q);
⑸x2、x4、x5依次为谷氨酸、异亮氨酸、赖氨酸(e、i、k)。
fx8x9x10x11x12x13x14x15mx16x17lppseqidno.3,其中:
⑴x8为色氨酸、苯丙氨酸(w、f);
⑵x9为脯氨酸、苏氨酸、天冬氨酸、组氨酸(p、t、d、h);
⑶x10为谷氨酰胺或异亮氨酸(q、i);
⑷x11为缬氨酸、精氨酸、天冬酰胺、苏氨酸、天冬氨酸(v、r、n、t、d);
⑸x12为亮氨酸、丙氨酸、缬氨酸、天冬氨酸、天冬酰胺(l、a、v、d、n);
⑹x13为丝氨酸、谷氨酰胺、谷氨酸(s、q、e);
⑺x14为异亮氨酸、天冬酰胺、脯氨酸、蛋氨酸(i、n、p、m);
⑻x15为异亮氨酸、丙氨酸、蛋氨酸(i、a、m);
⑼x16为亮氨酸、丙氨酸、赖氨酸(l、a、k);
⑽x17为蛋氨酸、赖氨酸(m、k);
yx18x19x20x21x22x23x24x25paseqidno.4,其中:
⑴x18、x19、x20、x21为丙氨酸、天冬酰胺、亮氨酸、亮氨酸(a、n、l、l);
⑵x22为精氨酸或天冬氨酸(r、d);
⑶x23为脯氨酸或谷氨酰胺(p、q);
⑷x24、x25独立地选自丙氨酸、精氨酸(a、r)。
x26x27x28x29x30px31x32x33x34lx35x36x37ppseqidno.5,其中:
⑴x26为苯丙氨酸(f);
⑵x27为苯丙氨酸、丙氨酸、组氨酸(f、a、h);
⑶x28为赖氨酸或谷氨酰胺(k、q);
⑷x29、x30、x31、x32、x33、x34、x35、x36、x37依次为天冬氨酸、异亮氨酸、天冬酰胺、丝氨酸、脯氨酸、脯氨酸、甘氨酸、天冬氨酸、天冬酰胺(d、i、n、s、p、p、g、d、n)。
sx38x39x40x41x42x43x44lpx45x46x47paseqidno.6,其中:
⑴x38、x40、x41、x42、x44、x45、x46、x47依次为谷氨酰胺、亮氨酸、丝氨酸、谷氨酸、赖氨酸、天冬氨酸、甘氨酸、苏氨酸(q、l、s、e、k、d、g、t);
⑵x39为半胱氨酸、赖氨酸、甘氨酸(c、k、g);
⑶x43为色氨酸或丝氨酸(w、s)。
sx48sx49x50x51x52x53mx54x55px56lppseqidno.7,其中:
⑴x48为谷氨酰胺、天冬酰胺、精氨酸(q、n、r);
⑵x49、x50、x51、x52、x53、x54、x55依次为缬氨酸、苏氨酸、甘氨酸、丙氨酸、天冬氨酸、甘氨酸、亮氨酸(v、t、g、a、d、g、l);
⑶x56为脯氨酸、异亮氨酸、甘氨酸(p、i、g)。
7.yx57hx58mx59x60x61x62rlpseqidno.8,其中:
⑴x57、x58、x59、x60、x61依次为天冬酰胺、丙氨酸、半胱氨酸、丙氨酸、天冬酰胺(n、a、c、a、d);
⑵x62为精氨酸、天冬氨酸(r、d)。
hsmpx63lpx64aseqidno.9,其中:
⑴x63、x64依次为脯氨酸、谷氨酰胺(p、q);
x65x66mprlppseqidno.10,其中:
⑴x65、x66依次为谷氨酰胺、天冬酰胺(q、n);
fyshx67x68prseqidno.11,其中:
⑴x67、x68依次为苏氨酸、亮氨酸(t、l)。
yshx69x70mx71x72x73x74x75x76prlseqidno.12,其中:
⑴x69为丙氨酸、半胱氨酸、天冬酰胺(a、c、n);
⑵x70、x71、x72、x73、x74、x75、x76依次为天冬酰胺、色氨酸、缬氨酸、苏氨酸、丝氨酸、天冬氨酸、天冬酰胺(n、w、v、t、s、d、n)。
所述的cmklr1拮抗多肽及其衍生物可以应用于制备预防和/或治疗雌性生殖疾病方面的药物,其存在形式有:①含有5个氨基酸;②由7个氨基酸组成;③由8个氨基酸组成;④由9个氨基酸组成;⑤由11个氨基酸组成;⑥由12个氨基酸组成;⑦由15个氨基酸组成;⑧由16个氨基酸组成。所述的cmklr1拮抗多肽的衍生物为cmklr1拮抗多肽氨基酸侧链基团上、cmklr1拮抗多肽片段的氨基端或羧基端进行常规修饰得到的产物,或者为cmklr1拮抗多肽上连接用于多肽或蛋白检测或纯化的标签所得到的产物;所述的常规修饰优选为氨基化、酰胺化、羟基化、羧基化、羰基化、烷基化、乙酰化、磷酸化、酯化、糖基化、环化、生物素化、荧光基团修饰、聚乙二醇peg修饰或固定化修饰等;所述的标签优选为his6、gst、egfp、mbp、nus、ha、igg、flag、c-myc或profinityexact等;
所述的cmklr1拮抗多肽及其衍生物可以来源于细菌、线虫、哺乳类动物或者鸟类,例如灵长类动物(人类);啮齿类动物,包括小鼠,大鼠,仓鼠,兔,马,牛,犬类,猫等。
所述的cmklr1拮抗多肽的衍生物优选为:上述cmklr1拮抗多肽seqidno.1第2个氨基酸残基为d构型酪氨酸,末端进行酰胺化修饰,即为phe-(d)tyr-ser-his-ser-met-pro-arg-leu-pro-pro-ala-nh2seqidno.1。
所述的cmklr1拮抗多肽及其衍生物的获得,采用现有技术中的公知方法进行,既可以用多肽自动合成仪进行化学合成;通过将短肽序列推导出核苷酸序列,然后克隆到载体中进行生物合成;也可以从现有存在的生物体内进行大量提取和纯化。
具体来说,cmklr1拮抗多肽lrh12-c5及其衍生物具有以下序列:
1.fyx1x2x3x4x5rx6x7pa(seqidno.2),其中:
⑴x1为谷氨酸、苏氨酸、脯氨酸、缬氨酸(e、t、p、v);
⑵x3为天冬酰胺、天冬氨酸、脯氨酸(n、d、p);
⑶x6为亮氨酸或谷氨酰胺(l、q);
⑷x7为丝氨酸、苏氨酸、谷氨酰胺(s、t、q);
⑸x2、x4、x5为谷氨酸、异亮氨酸、赖氨酸(e、i、k)。
2.fx8x9x10x11x12x13x14x15mx16x17lpp(seqidno.3),其中:
⑴x8为色氨酸、苯丙氨酸(w、f);
⑵x9为脯氨酸、苏氨酸、天冬氨酸、组氨酸(p、t、d、h);
⑶x10为谷氨酰胺或异亮氨酸(q、i);
⑷x11为缬氨酸、精氨酸、天冬酰胺、苏氨酸、天冬氨酸(v、r、n、t、d);
⑸x12为亮氨酸、丙氨酸、缬氨酸、天冬氨酸、天冬酰胺(l、a、v、d、n);
⑹x13为丝氨酸、谷氨酰胺、谷氨酸(s、q、e);
⑺x14为异亮氨酸、天冬酰胺、脯氨酸、蛋氨酸(i、n、p、m);
⑻x15为异亮氨酸、丙氨酸、蛋氨酸(i、a、m);
⑼x16为亮氨酸、丙氨酸、赖氨酸(l、a、k);
⑽x17为蛋氨酸、赖氨酸(m、k);
3.yx18x19x20x21x22x23x24x25pa(seqidno.4),其中:
⑴x18、x19、x20、x21依次为丙氨酸、天冬酰胺、亮氨酸、亮氨酸(a、n、l、l);
⑵x22为精氨酸或天冬氨酸(r、d);
⑶x23为脯氨酸或谷氨酰胺(p、q);
⑷x24、x25为丙氨酸、精氨酸(a、r)。
4.x26x27x28x29x30px31x32x33x34lx35x36x37pp(seqidno.5),其中:
⑴x26为苯丙氨酸(f);
⑵x27为苯丙氨酸、丙氨酸、组氨酸(f、a、h);
⑶x28为赖氨酸或谷氨酰胺(k、q);
⑷x29、x30、x31、x32、x33、x34、x35、x36、x37依次为天冬氨酸、异亮氨酸、天冬酰胺、丝氨酸、脯氨酸、脯氨酸、甘氨酸、天冬氨酸、天冬酰胺(d、i、n、s、p、p、g、d、n)。
5.sx38x39x40x41x42x43x44lpx45x46x47pa(seqidno.6),其中:
⑴x38、x40、x41、x42、x44、x45、x46、x47依次为谷氨酰胺、亮氨酸、丝氨酸、谷氨酸、赖氨酸、天冬氨酸、甘氨酸、苏氨酸(q、l、s、e、k、d、g、t);
⑵x39为半胱氨酸、赖氨酸、甘氨酸(c、k、g);
⑶x43为色氨酸或丝氨酸(w、s)。
6.sx48sx49x50x51x52x53mx54x55px56lpp(seqidno.7),其中:
⑴x48为谷氨酰胺、天冬酰胺、精氨酸(q、n、r);
⑵x49、x50、x51、x52、x53、x54、x55依次为缬氨酸、苏氨酸、甘氨酸、丙氨酸、天冬氨酸、甘氨酸、亮氨酸(v、t、g、a、d、g、l);
⑶x56为脯氨酸、异亮氨酸、甘氨酸(p、i、g)。
7.yx57hx58mx59x60x61x62rlp(seqidno.8),其中:
⑴x57、x58、x59、x60、x61依次为天冬酰胺、丙氨酸、半胱氨酸、丙氨酸、天冬酰胺(n、a、c、a、d);
⑵x62为精氨酸、天冬氨酸(r、d)。
8.hsmpx63lpx64a(seqidno.9),其中:
⑴x63、x64分别为脯氨酸、谷氨酰胺(p、q);
9.x65x66mprlpp(seqidno.10),其中:
⑴x65、x66分别为谷氨酰胺、天冬酰胺(q、n);
10.fyshx67x68pr(seqidno.11),其中:
⑴x67、x68分别为苏氨酸、亮氨酸(t、l)。
11.yshx69x70mx71x72x73x74x75x76prl(seqidno.12),其中:
⑴x69为丙氨酸、半胱氨酸、天冬酰胺(a、c、n);
⑵x70、x71、x72、x73、x74、x75、x76依次为天冬酰胺、色氨酸、缬氨酸、苏氨酸、丝氨酸、天冬氨酸、天冬酰胺(n、w、v、t、s、d、n)。
另外,cmklr1拮抗多肽lrh12-c5及其衍生物也包括以下天然存在于生物中的多肽:
本发明再一个方面提供了一种多聚核苷酸,其编码seqidno.1-24任一项所述多肽。
本发明再一个方面提供了一种载体,其包含了本发明所述的核苷酸,可以通过基因技术手段与启动子序列链接。
本发明再一个方面提供了一种宿主细胞,其转染了本发明所述的载体。
本发明再一个方面提供了本发明所述的cmklr1拮抗多肽、cmklr1拮抗多肽的衍生物、cmklr1拮抗多肽的生物活性片段或类似物及其衍生物在制备治疗chemerin/rve1-cmklr1介导疾病的药物中的用途。
本发明再一个方面提供了本发明所述的cmklr1拮抗多肽、cmklr1拮抗多肽的衍生物、cmklr1拮抗多肽的生物活性片段或类似物及其衍生物在治疗chemerin/rve1-cmklr1介导疾病中的用途。
在本发明的技术方案中,所述chemerin/rve1-cmklr1介导疾病选自卵巢癌、多囊卵巢综合征、脂肪肝、糖尿病、炎症反应。
本发明再一个方面提供了本发明所述的cmklr1拮抗多肽、cmklr1拮抗多肽的衍生物、cmklr1拮抗多肽的生物活性片段或类似物及其衍生物在制备抑制chemerin造成的camp浓度的降低的药物中的用途。
本发明再一个方面提供了本发明所述的cmklr1拮抗多肽、cmklr1拮抗多肽的衍生物、cmklr1拮抗多肽的生物活性片段或类似物及其衍生物在抑制chemerin造成的camp浓度的降低中的用途。
本发明再一个方面提供了本发明所述的cmklr1拮抗多肽、cmklr1拮抗多肽的衍生物、cmklr1拮抗多肽的生物活性片段或类似物及其衍生物在制备抑制chemerin引起的钙(ca2+)内流作用的药物中的用途。
本发明再一个方面提供了本发明所述的cmklr1拮抗多肽、cmklr1拮抗多肽的衍生物、cmklr1拮抗多肽的生物活性片段或类似物及其衍生物在抑制chemerin引起的钙(ca2+)内流作用中的用途。
本发明再一个方面提供了本发明所述的cmklr1拮抗多肽、cmklr1拮抗多肽的衍生物、cmklr1拮抗多肽的生物活性片段或类似物及其衍生物在制备抑制chemerin引起的细胞趋化作用的药物中的用途。
本发明再一个方面提供了本发明所述的cmklr1拮抗多肽、cmklr1拮抗多肽的衍生物、cmklr1拮抗多肽的生物活性片段或类似物及其衍生物在抑制chemerin引起的细胞趋化作用中的用途。
本发明再一个方面提供了本发明所述的cmklr1拮抗多肽、cmklr1拮抗多肽的衍生物、cmklr1拮抗多肽的生物活性片段或类似物及其衍生物在制备治疗卵巢癌、多囊卵巢综合征、脂肪肝、糖尿病、炎症反应的药物中的用途。
本发明再一个方面提供了本发明所述的cmklr1拮抗多肽、cmklr1拮抗多肽的衍生物、cmklr1拮抗多肽的生物活性片段或类似物及其衍生物在治疗卵巢癌、多囊卵巢综合征、脂肪肝、糖尿病、炎症反应中的用途。
本发明再一个方面提供了一种药物组合物,其中包含本发明所述的cmklr1拮抗多肽、cmklr1拮抗多肽的衍生物、cmklr1拮抗多肽的生物活性片段或类似物及其衍生物中的一种或多种作为活性成分。
所述的药物组合物可以含有一种或者是多种药学上可以接受的载体;
所述药学上可以接受的载体优选为稀释剂、赋形剂、填充剂、粘合剂、湿润剂、崩解剂、吸收促进剂、吸附载体、表面活性剂或润滑剂等。
所述的药物组合物可以进一步制成片剂、粒剂、胶囊、口服液或注射剂等多种形式,各种剂型的药物可以按照药学领域的常规方法制备。
一种预防和/或治疗雌性生殖疾病方面的药物,包含本发明所述的cmklr1拮抗多肽、cmklr1拮抗多肽的衍生物、cmklr1拮抗多肽的生物活性片段或类似物及其衍生物中的至少一种;
在本发明的一具体实验中,本发明利用所获得的cmklr1拮抗多肽lrh12-c5可以有效的缓解chemerin/rve1对camp信号通路的抑制作用。其中,cmklr1拮抗多肽lrh12-c5衍生物(seqidno.1~24)具有相同作用。
在本发明的另一具体实验中,利用cmklr1拮抗多肽lrh12-c5可以有效的抑制chemerin引起的钙(ca2+)内流作用。其中,cmklr1拮抗多肽lrh12-c5衍生物(seqidno.1~24)具有相同作用。
此外,chemerin可以通过与cmklr1受体结合发挥作用,趋化细胞进行迁移。在本发明的另一具体实验中,发明人利用transwell试验,发现cmklr1拮抗多肽lrh12-c5具有显著抑制chemerin引起的细胞迁移作用。其中,cmklr1拮抗多肽lrh12-c5衍生物(seqidno.1~24)具有相同作用。
在进一步试验中,发明人利用高表达cmklr1的卵巢癌细胞skov-3作为细胞模型验证了cmklr1拮抗多肽lrh12-c5的功能,与对照组相比,发现lrh12-c5可以显著抑制卵巢癌细胞skov-3的增殖。其中,cmklr1拮抗多肽lrh12-c5衍生物(seqidno.1~24)具有相同作用。
在本发明的另一具体实验中,发明人采用流式细胞技术,检测发现cmklr1拮抗多肽lrh12-c5可以阻断卵巢癌细胞skov-3周期,促进skov-3发生凋亡。其中,cmklr1拮抗多肽lrh12-c5衍生物(seqidno.1~24)具有相同作用。
本发明相对于现有技术具有如下的优点及效果:
(1)本发明提供了一种cmklr1拮抗多肽lrh12-c5及其衍生物(seqidno.1~24),所述的拮抗多肽及其衍生物能够专一性与cmklr1结合,并特异性竞争chemerin/rve1与cmklr1结合位点,能够抑制chemerin/rve1-cmklr1信号通路。
(2)本发明筛选得到的cmklr1拮抗多肽及其衍生物(seqidno.1~24)可以通过阻断chemerin/rve1-cmklr1的结合而抑制乳腺癌细胞增殖,促进乳腺癌细胞发生凋亡,可以作为chemerin/rve1-cmklr1结合位点的生物类多肽药物,可用于制备预防和/或治疗雌性生殖疾病方面的药物,例如:卵巢癌、多囊卵巢综合征、脂肪肝、糖尿病以及炎症反应。能够在医学与生物学领域得到广泛的应用,并产生巨大的社会与经济效益。
附图说明
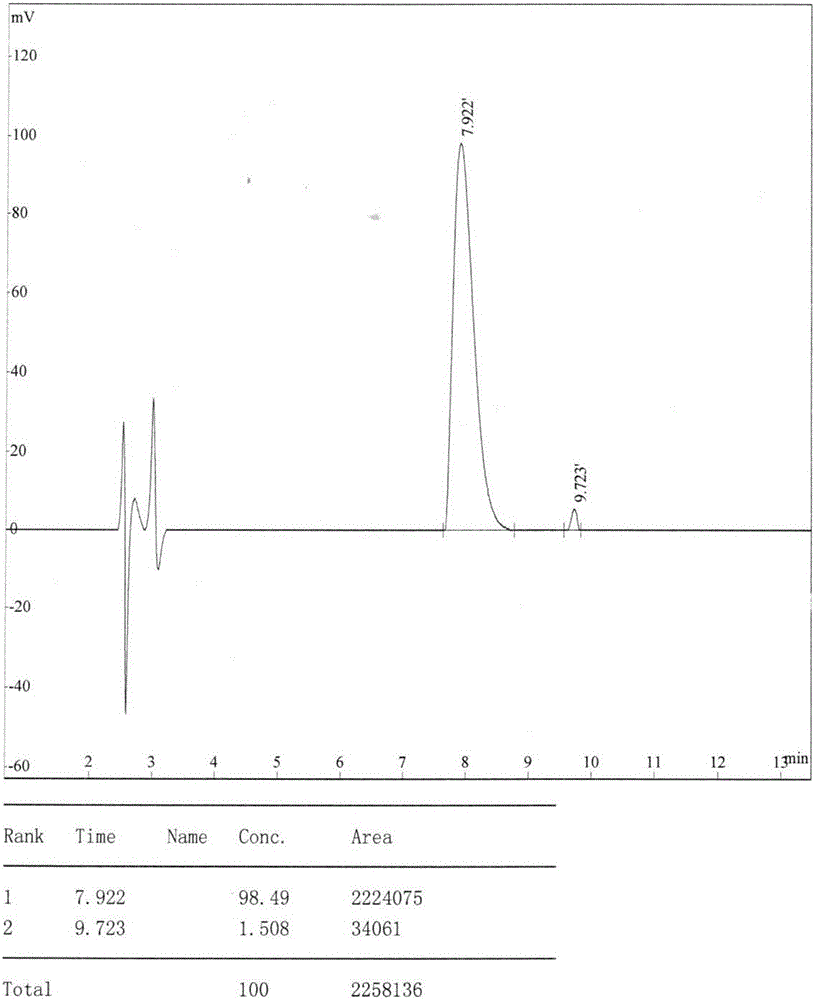
图1:lrh12-c5高效液相(hplc)检测图。
图2:lrh12-c5质谱(ms)检测图。
图3:cmklr1,chemerin,rve1以及lrh12-c5多肽疏水轮廓比较分析。其中:a:cmklr1疏水轮廓图;b:chemerin疏水轮廓图;c:rve1疏水轮廓图;d:lrh12-c5多肽疏水轮廓图;e:cmklr1,chemerin,rve1以及lrh12-c5多肽疏水轮廓比较图。cmklr1轮廓为蓝色,chemerin轮廓为绿色,rve1轮廓为红色,lrh12-c5轮廓为紫色;
图4:lrh12-c5缓解chemerin对camp信号通路的抑制作用。其中,a,lrh12-c5与forsklin和chemerin在293t野生型细胞中的作用;b,lrh12-c5单独在293tcmklr1+/+中的作用;c,lrh12-c5与forsklin和chemerin在293tcmklr1+/+中的作用;
图5:lrh12-c5抑制chemerin引起的钙(ca2+)内流作用。
图6:lrh12-c5抑制chemerin引起的细胞趋化作用。
图7:lrh12-c5抑制卵巢癌细胞skov-3增殖作用。
图8:lrh12-c5阻断卵巢癌细胞skov-3细胞周期进程。a:流式细胞仪检测细胞周期;b:细胞周期数据统计。
图9:lrh12-c5促进卵巢癌细胞skov-3凋亡作用。a:流式细胞仪检测细胞凋亡;b:细胞凋亡数据统计。
具体实施方式
为了更清楚地理解本发明,现参照下列实施例及附图进一步描述本发明。实施例仅用于解释而不以任何方式限制本发明。实施例中,各原始试剂材料均可商购获得,未注明具体条件的实验方法为所属领域熟知的常规方法和常规条件,或按照仪器制造商所建议的条件。
实施例1:进行cmklr1拮抗多肽lrh12-c5的淘选、扩增、纯化、测序及合成。
本实施例主要是为了筛选获得与cmklr1特异性结合的阳性噬菌体,再通过将阳性噬菌体扩增、纯化,提取噬菌体单链dna(ssdna)进行测序,将所获得的序列分析对比,最后合成高纯度的拮抗多肽lrh12-c5。
具体如下:
1.建立永久高表达cmklr1的293t细胞系:293t-cmklr1+/+/lrh
①选取生长旺盛的可发光的人293t细胞,在转染前一天以5×105个/孔,接种于6孔板中,培养至第二日后,细胞融合度为60%;
②第二日进行转染,以6孔板的一个培养孔为单位,用200μl的opti-mem培养基稀释3μg质粒,另以200μl的opti-mem培养基稀释6μl脂质体lipofectamine2000,分别轻轻混匀后,室温放置5分钟;
③将两管稀释液轻轻混合,室温静置20分钟后,向混合后的稀释液中轻轻加入600μl的opti-mem培养基;
④将待转染的细胞用pbs轻轻漂洗一次,然后将混合好的稀释液轻轻加入培养孔中,置于二氧化碳培养箱中培养;
⑤培养4~6小时后弃尽转染所用培养基,向孔中加入3ml完全培养基;
⑥48小时后选用含有1μg/ml嘌呤霉素(puromycin)的培养基进行筛选;待细胞不再出现死亡后即得到稳定表达cmklr1的293t细胞系。
⑦用trizol提取总rna,定量2μgrna进行逆转录(逆转录试剂盒,购于promega公司),并用特异引物序列进行qpcr。所用特异引物序列为hu-cmklr1引物序列:
fw5’-gaggcgtgacatagaatgga-3’seqidno.25;
rv5’-tgatatggattgggaggaagac-3’seqidno.26;
⑧与转染了psm2c-hu-scramblerna的做比较,检测cmklr1的高表达水平,并命名为:293t-cmklr1+/+/lrh,即可以用于阳性噬菌体筛选。
2.进行cmklr1拮抗多肽的淘选、扩增、纯化、测序及合成。
①er2738宿主菌液的制备:无菌技术操作,先取200μllb-tet液体培养基于1.5ml灭菌离心管中,再从e.colier2738的甘油冻存物中取0.2μl菌液与之充分混匀,全部吸取涂布于lb-tet平板上,标记平板,室温放置3min,然后置于37℃恒温培养箱倒置过夜培养。次日观察,长出克隆后用封口膜封口,4℃避光保存备用。用灭菌枪头以无菌技术挑取单菌落,放入已预先加有3mllb-tet液体培养基的10ml灭菌离心管中,标记后于恒温摇床37℃,300rpm/min振荡培养过夜。次日,细菌扩增液于4℃储存备用。取10ml灭菌离心管,无菌操作加入3mllb-tet液体培养基,取30μl过夜培养的细菌接种其中,恒温摇床37℃,300rpm/min振荡培养2~3h,细菌处于指数生长期,肉眼观察呈雾状(od600~0.5)。
②cmklr1拮抗肽的淘选:将高表达cmklr1细胞按105个/培养皿接种于预先包被多聚赖氨酸的60×15mm2培养皿中,常规培养至细胞密度80%~90%时,用于淘洗(同时用不表达cmklr1的细胞系作为空白对照)每轮洗脱液先取1μl测滴度,剩下的将其加入到20mllb培养液中扩增,再纯化、最后再测扩增后的滴度,扩增物于4℃短期保存,并取相等数量级用于下一轮淘选,剩下的扩增物用50%的甘油于-20℃保存。
③测定噬菌体的滴度:取4支灭菌的10ml离心管,每个噬菌体稀释度准备1个灭菌离心管,微波炉熔化顶层琼脂(agarosetop),每管加入3ml顶层琼脂,45℃水浴备用。每个噬菌体稀释度准备1块lb/iptg/xgal平板,37℃恒温培养箱预热备用。将od600~0.5的e.colier2738大肠杆菌按照噬菌体稀释度200μl/管分装,4℃保存备用。取4支灭菌的1.5ml离心管,分别盛有100μl、90μl、90μl、90μllb-tet培养基,将待测噬菌体吸取1μl入100μllb-tet培养基中,按10倍梯度稀释,分别标记为10-1、10-2、10-3、10-4,每个稀释度轻轻振荡混匀,瞬间离心。取10μl待滴定的各稀释度的噬菌体与200μle.colier2738混合,轻轻振荡混匀,瞬间离心,室温孵育5min。将混合菌液迅速加入顶层琼脂中,快速振荡混匀,立即倒入预热的lb/iptg/xgal平板,将其均匀展平,室温冷却5min,37℃恒温培养箱中,倒置平板培养过夜。
④洗脱噬菌体的扩增及纯化:取250ml的锥形瓶,按1:100比例,将过夜培养的er2738宿主菌液加入至20mllb液体培养基中,37℃,250rpm剧烈振荡培养2h;然后将待扩增的噬菌体液加入到锥形瓶中,37℃,250rpm剧烈振荡培养4.5h;将培养物转入50ml离心管中,4℃10,000rpm离心10min。上清液转入另一干净离心管中,4℃10,000rpm再次离心10min;取上清的80%转入另一干净离心管中,加入1/4体积的peg/nacl,颠倒混匀后于4℃沉淀过夜;第二天,将沉淀于4℃,12,000rpm离心20min。用干净枪头小心吸取上清液,再4℃12,000rpm离心1min,去掉残留上清液;然后用1mltbs重悬沉淀物,轻轻吹打100次。然后将悬液转入2ml离心管中,4℃10,000rpm离心5min除去残余细胞;将上清加入1/4体积的peg/nacl后,于冰上孵育60min再次沉淀;取出离心管,4℃12,000rpm离心20min,去掉上清;用200μltbs重悬沉淀,4℃10,000rpm离心1min。上清转入另一离心管中。4℃短期保存,也可以用50%的甘油于-20℃长期保存。单克隆噬菌体的扩增,包括⑴按1:100比例,将过夜培养的er2738宿主菌液加入至2mllb液体培养基中,37℃,250rpm剧烈振荡培养2h;用灭菌牙签,从第四轮滴度平板中选取少于100个噬菌斑的平板,挑取分隔良好的蓝色噬菌斑,加入到培养管中,37℃250r/min剧烈震荡培养4.5h;然后将培养物转入到新鲜离心管中,4℃10,000rpm离心30sec。上清转入以新鲜管中,再同样离心一次;将上清的80%转入新鲜离心管中,4℃贮存,也可以用50%的甘油于-20℃长期保存。
⑤琼脂糖凝胶电泳鉴定m13噬菌体ssdna:将凝胶成形模具水平放置,将选好的梳子放好,梳子底部与模具之间留1mm空间;称取dna电泳用琼脂糖1g放入250ml的三角烧瓶中,加入100ml1×tae缓冲液,混匀后,将烧瓶置于微波炉中,加热煮沸,直至琼脂糖完全溶解;关闭电磁炉,取出三角烧瓶,将其置室温下冷却至室温(手握烧瓶可以耐受),再加入溴化乙锭5μl,混匀后,即将凝胶溶液倒入胶板铺板。本实验所用制胶板约需胶液100ml;室温下待凝胶完全凝固,需时约30分钟,拔出梳齿,将胶板放入电泳槽中;在电泳槽加入1×tae缓冲液,以高出凝胶表面2mm为宜;将样品用loadingbuffer稀释以后加入胶板中,注意加样器吸头应恰好置于凝胶点样孔中,不可刺穿凝胶,也要防止将样品溢出孔外;接通电源,调节电压至50伏,电泳90min后,将凝胶板取出,在紫外灯下观察结果。
⑥ssdna测序及序列分析:将提取的m13噬菌体ssdna送到上海英潍捷基生物技术有限公司进行dna测序。测序以后用bioedit软件进行序列分析。通过分析结果可知,样品序列为phe-(d)tyr-ser-his-ser-met-pro-arg-leu-pro-pro-ala-nh2,其中第二位酪氨酸为d构型,以lrh12-c5表示,最后短肽由上海强耀生物公司合成。
图1高效液相色谱法(hplc)及图2质谱(ms)检测lrh12-c5多肽的纯度为99.85%,并且分子量大小与预测值一致。图3的疏水性轮廓分析,证明lrh12-c5多肽与cmklr1有一定的相似性,相似度达到0.0053619(pam250)。
实施例2cmklr1拮抗多肽lrh12-c5可以有效的缓解chemerin对camp信号通路的抑制作用。
(1)环磷酸腺苷(camp)酶联免疫吸附实验:
①细胞铺板:将野生型293t细胞和高表达cmklr1的293t细胞(293tcmklr1+/+),分别以5x105个/孔接种于6孔细胞培养板中,每孔培养基体积为1ml,放置于培养箱中培养24h后,饥饿过夜,加入不同浓度梯度(3μm,0.3μm,0.03μm)的lrh12-c5多肽,fosklin(25μm)以及chemerin(30nm)作用6h;
②样品制备:每孔加入300μl的细胞裂解液,4℃放置20分钟,用细胞刮刀刮取收集细胞,上下颠倒混匀后,12,000rpm离心10分钟,收集上清;
③样品浓度测定:样品浓度通过bca法测定;
④环磷酸腺苷(camp)酶联免疫吸附测试:
a,准备所需试剂,每个样品准备3个复孔;
b,将50μl的样品或标准品加入包被有抗体的96孔板中;再在每孔中加入25μlcamp过氧化物酶示踪剂缀合物;
c,每孔加入50μlanti-camp抗体,室温,摇床上缓慢孵育30分钟;
d,使用洗脱液清洗5次,每孔加入100μl化学反光剂,室温孵育5分钟;
e,酶标仪读板,记录发光值。
图4显示,chemerin在浓度为30nm的时候可以降低细胞camp浓度,但是在高表达cmklr1的293t细胞(293tcmklr1+/+)中加入不同浓度的lrh12-c5(3μm,0.3μm,0.03μm)作用后,可以使camp浓度显著增高。而在野生型293t细胞中,不表达cmklr1受体,因此lrh12-c5对chemerin降低camp浓度的作用无明显抑制作用。同样的,在lrh12-c5短肽单独作用组中,lrh12-c5也不能使camp浓度增高。综上所述,可以推断出,lrh12-c5可以特异性的通过与cmklr1作用来进一步抑制chemerin对camp浓度的降低作用。*p<0.05,**p<0.01,***p<0.001与forskolin+chemerin组相比较。
实施例3cmklr1拮抗多肽lrh12-c5可以有效的抑制chemerin引起的钙(ca2+)内流作用。
①细胞铺板:将野生型293t细胞和高表达cmklr1的293t细胞(293tcmklr1+/+),分别以5x103个/孔接种于96孔细胞培养板中,每孔培养基体积为200μl,放置于培养箱中培养24h后,饥饿过夜;
②试剂配置:将丙磺舒融入1ml缓冲溶液中,配置成浓度为250nm的丙磺舒,摇匀后,加入荧光试剂中准备待用;
③去掉细胞培养基,加入不同浓度梯度(30μm,3μm,0.3μm,0.03μm,0.003μm)的lrh12-c5多肽以及chemerin(0.3nm)作用30分钟,然后每孔加入100μl上述荧光试剂;
④37℃放置30分钟,然后室温放置30分钟;
⑤在激发光494nm和发射光516nm处测量荧光吸光值。
图5显示,在高表达cmklr1的293t细胞(293tcmklr1+/+)中,chemerin在作用浓度为0.3nm时,可以促进钙(ca2+)流信号通路,增高钙离子(ca2+)浓度。而在加入不同浓度的lrh12-c5短肽后,可以显著降低钙离子(ca2+)浓度,抑制chemerin对钙(ca2+)流信号通路的激活。但是在野生型不表达cmklr1的293t细胞中,chemerin对钙(ca2+)流信号通路无激活作用,lrh12-c5对钙(ca2+)流信号通路也无作用效果,通过该实验可以推断出,chemerin可以通过与受体cmklr1结合来激活钙(ca2+)流信号通路,而lrh12-c5短肽可以特异性的抑制chemerin/cmklr1信号通路来降低钙离子(ca2+)浓度。*p<0.05,**p<0.01,***p<0.001与chemerin组相比较。
实施例4transwell试验检测cmklr1拮抗多肽lrh12-c5抑制chemerin引起的细胞趋化作用。
①将野生型l1.2细胞和高表达cmklr1的l1.2细胞(l1.2cmklr1+/+),分别以1x106个/孔接种于5μm大小的96孔transwell细胞培养板中,培养6h后,饥饿1h;
②在培养板的下室中加入不同浓度梯度(30μm,3μm,0.3μm,0.03μm,0.003μm)的lrh12-c5多肽以及chemerin(0.3nm)作用2h;
③用流式细胞仪计数从上室趋化到下室的细胞数。
图6显示,在高表达cmklr1的l1.2细胞(l1.2cmklr1+/+)中,chemerin可以显著促进细胞从上室趋化到下室,而在加入同浓度的lrh12-c5短肽后,可以显著降低细胞趋化数目,抑制chemerin对细胞的趋化作用。但是在野生型不表达cmklr1的l1.2细胞中,chemerin对细胞无趋化作用,同时lrh12-c5短肽对细胞也无作用。通过该实验可以推断出,chemerin可以通过与受体cmklr1结合来促进细胞发生趋化作用,而lrh12-c5短肽可以特异性的阻断chemerin/cmklr1信号通路来抑制细胞发生趋化。*p<0.05,**p<0.01,***p<0.001与chemerin组相比较。
实施例5lrh12-c5可以显著抑制人卵巢癌skov-3细胞的增殖。
①将人卵巢癌细胞skov-3以5x103个/孔接种于96孔细胞培养板中,每孔培养基体积为200μl,培养24h,然后饥饿过夜;
②加入不同浓度梯度(100μm,10μm,1μm,0.1μm,0.01μm,0.001μm)的lrh12-c5多肽分别培养24小时、48小时、72小时;
③每孔加入20μl的mtt工作溶液,继续放入二氧化碳培养箱中培养4小时;
④弃培养板中的上清,加入150μldmso(二甲基亚砜),震荡10分钟,在酶标仪上选择490nm波长进行检测,绘制细胞的生长曲线。
发明人实验室在前期工作中,通过q-pcr试验检测发现任卵巢癌细胞skov-3高表达cmklr1。图7显示,不同浓度的lrh12-c5短肽可以显著抑制skov-3细胞增殖,并且随着作用时间的增加,短肽抑制skov-3细胞增殖的作用也更为显著。
实施例6lrh12-c5可以阻断人卵巢癌skov-3细胞周期进程。
①将人卵巢癌细胞skov-3以5×105个/孔接种于6孔细胞培养板中,每孔培养基体积为1ml,培养24h,然后饥饿过夜;
②加入不同浓度梯度(1μm,0.1μm,0.01μm)的lrh12-c5多肽分别培养24小时、48小时、72小时;
③小心收集细胞培养液到一离心管内备用。用胰酶消化细胞,至细胞可以被轻轻用移液管或枪头吹打下来时,加入前面收集的细胞培养液,吹打下所有的贴壁细胞,并轻轻吹散细胞。再次收集到离心管内。1000g左右离心3-5分钟,沉淀细胞;
④小心吸除上清,可以残留约50微升左右的培养液,以避免吸走细胞。加入约1毫升冰浴预冷的pbs,重悬细胞,并转移到1.5毫升离心管内。再次离心沉淀细胞,小心吸除上清,可以残留约50微升左右的pbs,以避免吸走细胞。轻轻弹击离心管底以适当分散细胞,避免细胞成团;
⑤加入1毫升冰浴预冷70%乙醇中,轻轻吹打混匀,4℃固定12h。1000g左右离心3-5分钟,沉淀细胞。小心吸除上清,可以残留约50微升左右的70%乙醇,以避免吸走细胞。加入约1毫升冰浴预冷的pbs,重悬细胞。再次离心沉淀细胞,小心吸除上清,可以残留约50微升左右的pbs,以避免吸走细胞。轻轻弹击离心管底以适当分散细胞,避免细胞成团;
⑥每管细胞样品中加入0.5毫升碘化丙啶染色液,缓慢并充分重悬细胞沉淀,37℃避光温浴30分钟。随后可以4℃或冰浴避光存放;
⑦用流式细胞仪在激发波长488nm波长处检测红色荧光,同时检测光散射情况。分析细胞结果。
图8显示,通过流式细胞仪分析发现,一方面随着作用浓度的升高,lrh12-c5短肽可以明显降低人卵巢癌细胞skov-3g2/m期细胞数目,增加g0/g1和s期数目,阻断细胞周期正常进展;另一方面随着作用时间的延长,lrh12-c5短肽也显著降低g2/m期细胞数目,增加g0/g1和s期数目,阻断细胞周期正常进展。综上所述,可以推断出lrh12-c5短肽通过阻断细胞周期来抑制人卵巢癌细胞skov-3的增殖。*p<0.05,**p<0.01与对照组g0/g1期相比较;#p<0.005与对照组s期相比较;&p<0.005与对照组g2/m期相比较。
实施例7lrh12-c5可以促进人卵巢癌skov-3细胞凋亡。
①将人卵巢癌细胞skov-3以5ⅹ105个/孔接种于6孔细胞培养板中,每孔培养基体积为1ml,培养24h,然后饥饿过夜;
②加入不同浓度梯度(1μm,0.1μm,0.01μm)的lrh12-c5多肽分别培养24小时、48小时、72小时;
③先将上清收集到离心管中,再用不含edta的胰酶小心消化收集细胞培养液到离心管。500g左右离心5分钟,沉淀细胞;
④用预冷的pbs洗涤细胞两次,500g左右离心5分钟收集细胞;
⑤加入100μl预冷的1ⅹannexinvbindingbuffer,重悬细胞;
⑥加入5μlannexinv-fitc和5μlpi,轻轻混匀,室温避光反应15分钟;
⑦加入400μl预冷的1ⅹannexinvbindingbuffer,轻轻混匀,将样品至于冰上避光放置,1h内用流式细胞仪检测。分析检测结果。
图9显示,通过流式细胞仪分析发现,一方面随着作用浓度的升高,lrh12-c5短肽可以明显促进人卵巢癌细胞skov-3发生凋亡,另一方面随着作用时间的延长,lrh12-c5短肽也显著增高了人卵巢癌细胞skov-3凋亡数目。综上所述,lrh12-c5短肽可以显著促进人卵巢癌细胞skov-3发生凋亡。*p<0.05,**p<0.01与对照组相比较。
sequencelisting
<110>深圳先进技术研究院
<120>一种cmklr1拮抗多肽及其衍生物与应用
<130>cp11701233c
<160>26
<170>patentinversion3.3
<210>1
<211>12
<212>prt
<213>人工序列
<400>1
phetyrserhissermetproargleuproproala
1510
<210>2
<211>12
<212>prt
<213>人工序列
<220>
<221>misc_feature
<222>(3)..(3)
<223>谷氨酸、苏氨酸、脯氨酸或缬氨酸
<220>
<221>misc_feature
<222>(4)..(4)
<223>谷氨酸
<220>
<221>misc_feature
<222>(5)..(5)
<223>天冬酰胺、天冬氨酸或脯氨酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>异亮氨酸
<220>
<221>misc_feature
<222>(7)..(7)
<223>赖氨酸
<220>
<221>misc_feature
<222>(9)..(9)
<223>亮氨酸或谷氨酰胺
<220>
<221>misc_feature
<222>(10)..(10)
<223>丝氨酸、苏氨酸或谷氨酰胺
<400>2
phetyrxaaxaaxaaxaaxaaargxaaxaaproala
1510
<210>3
<211>15
<212>prt
<213>人工序列
<220>
<221>misc_feature
<222>(2)..(2)
<223>色氨酸或苯丙氨酸
<220>
<221>misc_feature
<222>(3)..(3)
<223>脯氨酸、苏氨酸、天冬氨酸或组氨酸
<220>
<221>misc_feature
<222>(4)..(4)
<223>谷氨酰胺或异亮氨酸
<220>
<221>misc_feature
<222>(5)..(5)
<223>缬氨酸、精氨酸、天冬酰胺、苏氨酸或天冬氨酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>亮氨酸、丙氨酸、缬氨酸、天冬氨酸或天冬酰胺
<220>
<221>misc_feature
<222>(7)..(7)
<223>丝氨酸、谷氨酰胺或谷氨酸
<220>
<221>misc_feature
<222>(8)..(8)
<223>异亮氨酸、天冬酰胺、脯氨酸或蛋氨酸
<220>
<221>misc_feature
<222>(9)..(9)
<223>异亮氨酸、丙氨酸或蛋氨酸
<220>
<221>misc_feature
<222>(11)..(11)
<223>亮氨酸、丙氨酸或赖氨酸
<220>
<221>misc_feature
<222>(12)..(12)
<223>蛋氨酸或赖氨酸
<400>3
phexaaxaaxaaxaaxaaxaaxaaxaametxaaxaaleupropro
151015
<210>4
<211>11
<212>prt
<213>人工序列
<220>
<221>misc_feature
<222>(2)..(2)
<223>丙氨酸
<220>
<221>misc_feature
<222>(3)..(3)
<223>天冬酰胺
<220>
<221>misc_feature
<222>(4)..(5)
<223>亮氨酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>精氨酸或天冬氨酸
<220>
<221>misc_feature
<222>(7)..(7)
<223>脯氨酸或谷氨酰胺
<220>
<221>misc_feature
<222>(8)..(8)
<223>丙氨酸
<220>
<221>misc_feature
<222>(9)..(9)
<223>精氨酸
<400>4
tyrxaaxaaxaaxaaxaaxaaxaaxaaproala
1510
<210>5
<211>16
<212>prt
<213>人工序列
<220>
<221>misc_feature
<222>(1)..(1)
<223>苯丙氨酸
<220>
<221>misc_feature
<222>(2)..(2)
<223>苯丙氨酸、丙氨酸或组氨酸
<220>
<221>misc_feature
<222>(3)..(3)
<223>赖氨酸或谷氨酰胺
<220>
<221>misc_feature
<222>(4)..(4)
<223>天冬氨酸
<220>
<221>misc_feature
<222>(5)..(5)
<223>异亮氨酸
<220>
<221>misc_feature
<222>(7)..(7)
<223>天冬酰胺
<220>
<221>misc_feature
<222>(8)..(8)
<223>丝氨酸
<220>
<221>misc_feature
<222>(9)..(10)
<223>脯氨酸
<220>
<221>misc_feature
<222>(12)..(12)
<223>甘氨酸
<220>
<221>misc_feature
<222>(13)..(13)
<223>天冬氨酸
<220>
<221>misc_feature
<222>(14)..(14)
<223>天冬酰胺
<400>5
xaaxaaxaaxaaxaaproxaaxaaxaaxaaleuxaaxaaxaapropro
151015
<210>6
<211>15
<212>prt
<213>人工序列
<220>
<221>misc_feature
<222>(2)..(2)
<223>谷氨酰胺
<220>
<221>misc_feature
<222>(3)..(3)
<223>半胱氨酸、赖氨酸或甘氨酸
<220>
<221>misc_feature
<222>(4)..(4)
<223>亮氨酸
<220>
<221>misc_feature
<222>(5)..(5)
<223>丝氨酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>谷氨酸
<220>
<221>misc_feature
<222>(7)..(7)
<223>色氨酸或丝氨酸
<220>
<221>misc_feature
<222>(8)..(8)
<223>赖氨酸
<220>
<221>misc_feature
<222>(11)..(11)
<223>天冬氨酸
<220>
<221>misc_feature
<222>(12)..(12)
<223>甘氨酸
<220>
<221>misc_feature
<222>(13)..(13)
<223>苏氨酸
<400>6
serxaaxaaxaaxaaxaaxaaxaaleuproxaaxaaxaaproala
151015
<210>7
<211>16
<212>prt
<213>人工序列
<220>
<221>misc_feature
<222>(2)..(2)
<223>谷氨酰胺、天冬酰胺或精氨酸
<220>
<221>misc_feature
<222>(4)..(4)
<223>缬氨酸
<220>
<221>misc_feature
<222>(5)..(5)
<223>苏氨酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>甘氨酸
<220>
<221>misc_feature
<222>(7)..(7)
<223>丙氨酸
<220>
<221>misc_feature
<222>(8)..(8)
<223>天冬氨酸
<220>
<221>misc_feature
<222>(10)..(10)
<223>甘氨酸
<220>
<221>misc_feature
<222>(11)..(11)
<223>亮氨酸
<220>
<221>misc_feature
<222>(13)..(13)
<223>脯氨酸、异亮氨酸或甘氨酸
<400>7
serxaaserxaaxaaxaaxaaxaametxaaxaaproxaaleupropro
151015
<210>8
<211>12
<212>prt
<213>人工序列
<220>
<221>misc_feature
<222>(2)..(2)
<223>天冬酰胺
<220>
<221>misc_feature
<222>(4)..(4)
<223>丙氨酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>半胱氨酸
<220>
<221>misc_feature
<222>(7)..(7)
<223>丙氨酸
<220>
<221>misc_feature
<222>(8)..(8)
<223>天冬酰胺
<220>
<221>misc_feature
<222>(9)..(9)
<223>精氨酸或天冬氨酸
<400>8
tyrxaahisxaametxaaxaaxaaxaaargleupro
1510
<210>9
<211>9
<212>prt
<213>人工序列
<220>
<221>misc_feature
<222>(5)..(5)
<223>脯氨酸
<220>
<221>misc_feature
<222>(8)..(8)
<223>谷氨酰胺
<400>9
hissermetproxaaleuproxaaala
15
<210>10
<211>8
<212>prt
<213>人工序列
<220>
<221>misc_feature
<222>(1)..(1)
<223>谷氨酰胺
<220>
<221>misc_feature
<222>(2)..(2)
<223>天冬酰胺
<400>10
xaaxaametproargleupropro
15
<210>11
<211>8
<212>prt
<213>人工序列
<220>
<221>misc_feature
<222>(5)..(5)
<223>苏氨酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>亮氨酸
<400>11
phetyrserhisxaaxaaproarg
15
<210>12
<211>15
<212>prt
<213>人工序列
<220>
<221>misc_feature
<222>(4)..(4)
<223>丙氨酸、半胱氨酸或天冬酰胺
<220>
<221>misc_feature
<222>(5)..(5)
<223>天冬酰胺
<220>
<221>misc_feature
<222>(7)..(7)
<223>色氨酸
<220>
<221>misc_feature
<222>(8)..(8)
<223>缬氨酸
<220>
<221>misc_feature
<222>(9)..(9)
<223>苏氨酸
<220>
<221>misc_feature
<222>(10)..(10)
<223>丝氨酸
<220>
<221>misc_feature
<222>(11)..(11)
<223>天冬氨酸
<220>
<221>misc_feature
<222>(12)..(12)
<223>天冬酰胺
<400>12
tyrserhisxaaxaametxaaxaaxaaxaaxaaxaaproargleu
151015
<210>13
<211>8
<212>prt
<213>细菌
<400>13
tyrserhissermetproargleu
15
<210>14
<211>8
<212>prt
<213>细菌
<400>14
tyrserhissermetproargleu
15
<210>15
<211>8
<212>prt
<213>细菌
<400>15
sermetproargleuproproala
15
<210>16
<211>8
<212>prt
<213>细菌
<400>16
sermetproargleuproproala
15
<210>17
<211>7
<212>prt
<213>虫类
<400>17
phetyrserhissermetpro
15
<210>18
<211>7
<212>prt
<213>虫类
<400>18
phetyrserhissermetpro
15
<210>19
<211>7
<212>prt
<213>树
<400>19
tyrserhissermetproarg
15
<210>20
<211>7
<212>prt
<213>树
<400>20
tyrserhissermetproarg
15
<210>21
<211>7
<212>prt
<213>细菌
<400>21
hissermetproargleupro
15
<210>22
<211>7
<212>prt
<213>细菌
<400>22
hissermetproargleupro
15
<210>23
<211>5
<212>prt
<213>细菌
<400>23
metproargleupro
15
<210>24
<211>5
<212>prt
<213>细菌
<400>24
metproargleupro
15
<210>25
<211>20
<212>dna
<213>人工序列
<400>25
gaggcgtgacatagaatgga20
<210>26
<211>22
<212>dna
<213>人工序列
<400>26
tgatatggattgggaggaagac22
- 还没有人留言评论。精彩留言会获得点赞!