用于转导淋巴细胞及调节其活性的方法及组合物与流程
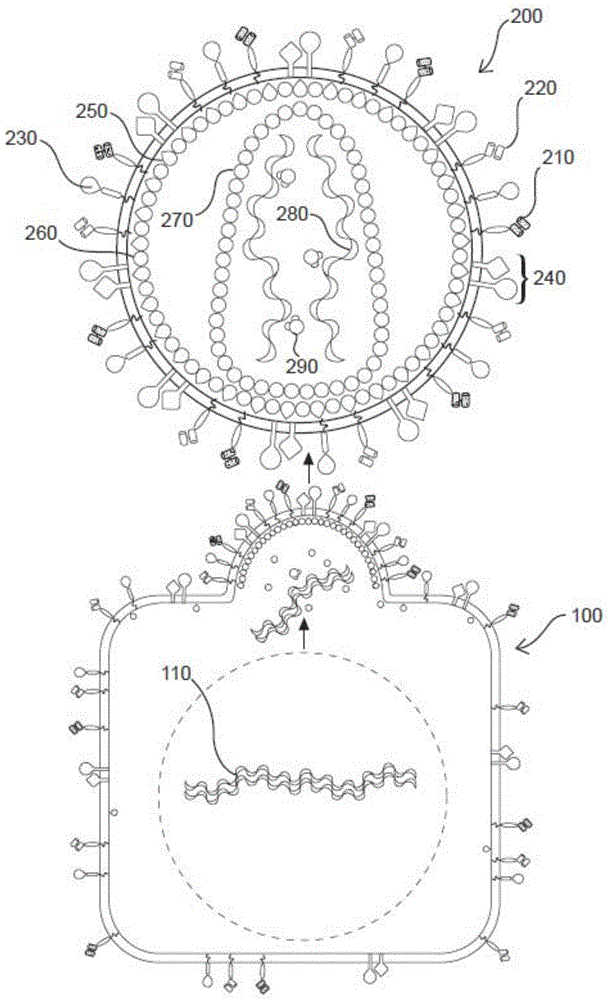
本申请案主张申请案2017年3月19日申请的国际申请案第pct/us2017/023112号,2017年3月19日申请的美国专利申请案第15/462855号,2016年7月8日申请的美国临时申请案第62/360041号及2017年3月3日申请的美国临时申请案第62/467039号的权益;国际申请案第pct/us2017/023112号主张2016年3月19日申请的美国临时申请案第62/390093号,2016年7月8日申请的美国临时申请案第62/360041号及2017年3月3日申请的美国临时申请案第62/467039号的权益;美国申请案第15/462855号主张2016年3月19日申请的美国临时申请案第62/390093号,2016年7月8日申请的美国临时申请案第62/360041号及2017年3月3日申请的美国临时申请案第62/467039号的权益。本段所引用的这些申请案以全文引用的方式并入本文中。序列表本申请案在此通过引用的方式并入与本申请案一起提交的电子序列表材料。电子序列表中的材料提交为在2017年7月8日创建的标题为“f1_001_wo_02_sequence_2017_07_08.txt”的文本(.txt)文档(其文档大小为268kb),且以全文引用的方式并入本文中。本公开内容涉及免疫学领域,更具体的说,涉及t淋巴细胞或其他免疫细胞的遗传修饰,及制备复制缺陷型重组逆转录病毒颗粒及控制其中基因表达的方法。
背景技术:
:从受试者(例如患者)分离的淋巴细胞可在体外活化且经遗传修饰以表达合成蛋白,这样的合成蛋白能够基于所整合入的遗传程序而与其他细胞及环境重定向接合。嵌合抗原受体(car)就是此类合成蛋白的一个例子。目前使用的一种car是由复制缺陷型重组逆转录病毒编码的一个或多个胞内信号传导域、跨膜结构域和胞外识别结构域(例如,抗原结合结构域)的融合。尽管重组逆转录病毒已在感染非分裂细胞上显示功效,但静息cd4及cd8淋巴细胞对这类载体的基因转导不敏感。为了解决这个问题,通常在可出现对car基因载体的遗传修饰之前使用刺激试剂在体外活化这些细胞。在刺激及转导之后,经遗传修饰的细胞在体外扩增并随后重新引入淋巴细胞清除(lymphodepleted)的患者。在体内抗原接合之后,car胞内信号传导部分可在免疫细胞中开始活化相关的反应并释放细胞溶解分子以诱导肿瘤细胞死亡。目前此类的方法需要大量的操作和生产,即在t细胞回输注至患者之前在体外增殖t细胞,以及淋巴细胞清除化学疗法以释放细胞因子并消耗竞争性受体从而促进t细胞植入。一旦引入体内,这种car疗法不能继续控制体内传播速率,也不能安全地定向又在肿瘤外表达的靶标。因此,现今car疗法一般由使用1×105个细胞/公斤至1×108个细胞/公斤的剂量离体扩增12天至28天的细胞输注,且所定向的靶标(例如肿瘤靶标)为在靶脱瘤毒性(offtumorontargettoxicity)一般是可接受的靶标。这些相对长的离体扩增时间产生细胞存活性及无菌性问题,以及样品一致性和可扩展性的挑战。因此,极为需要一种更安全,更有效的可扩展的t细胞或nk细胞疗法。发明概述本文中提供帮助克服用于转导和/或遗传修饰淋巴细胞(例如t细胞和/或nk细胞)以及用这些细胞执行过继性细胞疗法的方法的有效性及安全性有关问题的方法、组合物和试剂盒。因此,在一些方面中,本文中提供用于遗传修饰和/或转导淋巴细胞(尤其t细胞和/或nk细胞)和/或用于调节经转导和/或遗传修饰的t细胞和/或nk细胞的活性的方法,组合物及试剂盒。这些方法,组合物及试剂盒提供较目前技术更加改善的功效及安全性,尤其关于表达嵌合抗原受体(car)及在说明性实施方式中微环境限制的生物car的t细胞和/或nk细胞。利用本文中所提供的方法生产的和/或在本文中所提供的方法中使用的经转导和/或遗传修饰的t细胞和/或nk细胞包括在经由逆转录病毒(例如慢病毒)颗粒自逆转录病毒(例如慢病毒)基因组递送的说明性实施方式中为此类细胞及为利用这类细胞方法(例如过继性细胞疗法)提供经改良特征的功能性及功能性组合。举例而言,这样的细胞可以在更短的时间内离体生产,并且具有可以被更好地调节的改善的生长特性。本文在一些方面中提供了用于调节car的表达的调节元件,mrna,抑制性rna和/或在淋巴细胞(如t细胞及nk细胞)中非抑制性rna的淋巴增殖性元件(lymphoproliferativeelement)。此外,本文在一些方面中提供表达多种功能元件并在其表面上携载多种功能元件的重组逆转录病毒,以及用于生产重组逆转录病毒的方法及包装细胞系。这些重组逆转录病毒及用于生产它们的方法和细胞克服了现有技术对于基因组中在递送至t细胞和/或nk细胞时提供益处的不同功能元件的数目和大小的限制。在一些方面中,提供用于转导和/或遗传修饰淋巴细胞(例如t细胞和/或nk细胞)的方法,并且在说明性实施方式中,提供用于转导和/或遗传修饰静息t细胞和/或nk细胞的离体方法。这些方面中的一些可比先前方法快速得多地执行,这样可以促进改进患者护理方法。此外,本文中提供在一些实施方式中利用本文提供的重组逆转录病毒在一些方面中连同药剂以提供改进的安全机制以帮助调节经转导和/或遗传修饰的淋巴细胞(例如t细胞和/或nk细胞)的活性的方法。这些方法,组合物及试剂盒可用于用表达car的经转导和/或遗传修饰的t细胞和/或nk细胞的过继性细胞疗法。在本专利申请中提供了关于本公开内容的方面和实施方式的更多细节。章节及章节标题并不旨在限制其中方法,组合物及试剂盒或功能元件的组合。附图简述图1显示包括包装细胞(100)及本公开的一个例示性,非限制性实施方式的包装细胞(100)产生的复制缺陷型重组逆转录病毒颗粒(200)的说明性组合物的示意图。在图1中,能够编码本发明方面的多种载体(被称为重组多核苷酸(110))被包装至重组逆转录病毒颗粒(200)中,该重组逆转录病毒颗粒在其基因组中包括第一经改造信号传导多肽(其包括一个或多个淋巴增殖性元件)及在一些实施方式中第二经改造信号传导多肽(其为嵌合抗原受体或car)。复制缺陷型重组逆转录病毒颗粒在其膜上表达:允许复制缺陷型重组逆转录病毒颗粒与靶细胞结合并融合的假型化元件(pseudotypingelement)(在一非限制性实施方式中,麻疹病毒凝血素(h)多肽及麻疹病毒融合(f)多肽,或其细胞质结构域缺失变体)(240);能够结合并活化静息t细胞的活化元件(在非限制性实施方式中,具有能够结合cd28的多肽及能够结合cd3的多肽的活化元件(分别为210及220);及膜结合细胞因子(在一非限制性实施方式中,il-7daf融合多肽)(230)。标记为(250),(260),(270),(280)及(290)的部分分别为src-flag-vpx,hivgag基质,hivgag衣壳,rna及hivpol。图2显示包括由包装细胞(100)产生的复制缺陷型重组逆转录病毒颗粒(200)及由复制缺陷型重组逆转录病毒颗粒(200)转染的静息t细胞(300)的说明性组合物的示意性图。复制缺陷型重组逆转录病毒颗粒(200)表面上的元件与静息t细胞表面上的受体和/或配体结合。在非限制性实施方式中,假型化元件可包括有助于复制缺陷型重组逆转录病毒颗粒(200)与t细胞结合及融合的结合多肽及促融合多肽(fusogenicpolypeptide)(在非限制性实施方式中,麻疹病毒凝血素(h)多肽及麻疹病毒融合(f)多肽,或其细胞质结构域缺失变体)。在非限制性实施方式中,复制缺陷型重组逆转录病毒颗粒(200)在其表面上包括能够通过接合t细胞受体复合物及任选地辅助受体(320)来活化静息t细胞的活化元件(在非限制性实施方式中,具有能够结合cd28的多肽及能够结合cd3的多肽的活化元件)。此外,存在于复制缺陷型重组逆转录病毒颗粒(200)表面上的膜结合细胞因子(在非限制性实施方式中,il-7daf融合多肽)与静息t细胞表面上的il-7rα(310)结合。与t细胞融合的复制缺陷型重组逆转录病毒颗粒(200)及编码包括淋巴增殖性元件(在说明性实施方式中,组成型活化的il-7rα)(370)的第一经改造信号传导多肽的多核苷酸在迁移至细胞核并待整合入经活化t细胞的dna之前在胞溶质中被逆转录。不受理论限制,在一些非限制性实施方式中,用病毒包装的src-flag-vpx(250)进入静息t细胞的胞溶质且促进samhd1(350)降解,从而导致可用于逆转录的细胞质dntp库增加。在一些实施方式中,多核苷酸也可编码包括car(360)的第二经改造信号传导多肽。在一些实施方式中,在化合物与调节其表达的控制元件(在非限制性实例中,该控制元件为结合核苷类似物的核糖开关)结合时,表达淋巴增殖性元件。在一些实施方式中,car的表达也受控制元件调节。部分(330)为slam及cd46。部分(340)为cd3。图3a至图3e显示用于转染包装细胞以产生本文中所描述的复制缺陷型重组逆转录病毒颗粒的非限制性、例示性载体构建体的示意图。图3a显示含有编码融合至nfκbp65活化子结构域(p65ad)的frb结构域及融合至组成型表达的三个fkbp重复的zfhd1dna结合结构域的多核苷酸序列的构建体。图3a中的构建体还包括hiv1rev及作为在雷帕霉素诱导型zfhd1/p65ad启动子下的srcflagvpx融合的vpx。图3b显示含有在zfhd1/p65ad启动子的控制下编码rtta序列的多核苷酸的构建体。图3c显示含有编码侧翼为loxp位点的嘌呤霉素抗性基因及侧翼为lox2272位点的胞外myc标签的多核苷酸的构建体。两个可选标记均在bitre启动子的控制下,其侧翼有frt位点。图3d显示含有编码侧翼为在tre启动子的控制下的loxp位点的rfp及在tre启动子与rfp的5'loxp位点之间的单frt位点的多核苷酸的构建体。图3e显示含编码侧翼有在tre启动子控制下的loxp位点的gfp和在tre启动子与gep的5'loxp位点之间的单个frt位点的多核苷酸的构建体。图3c至3e中的构建体充当其他多核苷酸序列插入至包装细胞系的基因组中的着陆点。图4a至图4c显示用于转染包装细胞以产生本文中所描述的复制缺陷型重组逆转录病毒颗粒的非限制性、例示性载体构建体的示意图。图4a显示含有三顺反子多核苷酸的构建体,所述三顺反子多核苷酸编码具有cd14gpi锚连接位点的抗cd3(克隆ucht1)scfvfc、能够使cd28与cd16bgpi锚连接位点结合的cd80胞外结构域(ecd)及融合至衰变加速因子(daf)的il-7(其中,转座子序列侧接多核苷酸区以用于整合至hek293s基因组中)。图4b显示含有具有bitre启动子的多核苷酸及在一个方向上编码gag多肽及pol多肽的多核苷酸区以及在另一方向上编码麻疹病毒fδx蛋白质及hδy蛋白质的多核苷酸区的构建体。图4c显示含有编码car及在cd3z启动子(其在hek293s细胞中不具有活性)的控制下的淋巴增殖性元件il7rα-insppcl的多核苷酸序列的构建体,其中car及il7rα-insppcl由编码t2a核糖体跳跃序列的多核苷酸序列隔开,并且il7rα-insppcl具有阿昔洛韦核糖开关控制的核酶。含car的构建体进一步包括cppt/cts,rre序列及编码hiv-1psi(ψ)的多核苷酸序列。待被整合至基因组中的含car的构建体上的整个多核苷酸序列侧翼有frt位点。图5a至图5c显示作为选择性核糖开关控制的代表性核苷类似物的阿昔洛韦(图5a),喷昔洛韦(图5b)及2'-脱氧鸟苷(图5c)的分子结构。图6表示mesoplasmaflorumi-a型脱氧鸟苷核糖开关调节区及相关基因产物。序列为m.floruml1基因组dna(ae017263.1)nt624396至nt625670的反向互补物,其与m.florumw37基因组dna(cp006778.1)nt636277至nt637550相同。用于初始筛选的脱氧鸟苷结合适配体序列以粗体及下划线指示。下游基因产物(ib类(需氧)核糖核苷酸还原酶,β亚基)以大写字母指示。图7表示用于定向进化策略的m.florumi-a型脱氧鸟苷核糖开关适配体区。空椭圆内的核苷酸用于随机化。条纹椭圆内的核苷酸用于插入/缺失及随机化。图8a及图8b表示m.florumi-a型脱氧鸟苷核苷开关适配体筛选文库。在图8a中,实线框内的核苷酸是用于随机化的序列区域,虚线框内的核苷酸是用于插入/缺失及随机化的序列区。图8b显示经由突变(随机核苷酸(“n”))及缺失/插入产生的可能序列。图9表示合成为具有额外碱基对(添加其以允许pcr扩增)及t7启动子(添加其以用于在体外转录以用于文库筛选)的反向互补物的m.florumi-a型脱氧鸟苷核糖开关适配体寡核苷酸文库。还显示了对应的t7启动子扩增引物和反向扩增引物。图10表示枯草芽孢杆菌(bacillussubtilis)鸟苷xpt核糖开关调节区及相关基因产物。序列为枯草芽孢杆菌枯草亚种(b.subtilissubsp.subtilis)6051-hgw基因组dna(cp003329.1)nt2319439至nt2320353的反向互补物。用于初始筛选的鸟苷结合适配体序列以粗体及下划线指示。下游基因产物(黄嘌呤磷酸核糖转移酶xpt)以大写字母指示。图11表示用于定向进化策略的枯草芽孢杆菌鸟苷xpt核糖开关适配体区。空椭圆内的核苷酸用于随机化。条纹椭圆内的核苷酸用于插入/缺失及随机化。图12a及图12b表示枯草芽孢杆菌鸟苷xpt核糖开关适配体筛选文库。在图12a中,实线框内的核苷酸是用于随机化的序列区,且虚线框内的核苷酸是用于插入/缺失及随机化的序列区。图12b显示经由突变(随机核苷酸(“n”))及缺失/插入产生的可能序列。图13表示合成为具有额外碱基对(添加其以允许pcr扩增)及t7启动子(添加其以用于在体外转录以用于文库筛选)的反向互补物的枯草芽孢杆菌鸟苷xpt核糖开关适配体寡核苷酸文库。还显示了对应的t7启动子扩增引物及反向扩增引物。图14显示选择文库构建。该文库是基于已知鸟苷结合rna及脱氧鸟苷结合rna来构建(pikovskaya,2013)。图15显示氧化石墨烯(gro)适配体选择的图示。在步骤(1)中,rna被转录和纯化。在步骤(2)中,纯化的rna被洗脱。在步骤(3)中,适配体与反向靶标(counter-target)和缓冲剂一起孵育。在步骤(4)中,用氧化石墨烯移除与反向靶标或缓冲剂组分结合的序列。在步骤(5)中,离心分离上清液中的非特异性反应物质,然后将其丢弃。两次额外5分钟洗涤移除大部分的残余反向靶标结合序列及缓冲剂结合序列。在步骤(6)中,将阿昔洛韦在1×选择缓冲剂中的溶液添加至gro结合文库以用于阳性选择,因此潜在适配体序列经由与阳性靶标相互作用而自gro解吸附。在步骤(7)中,最终离心步骤将上清液中的靶标结合序列与仍吸附于gro的非反应序列分离。在步骤(8)中,对选择的序列进行逆转录,然后经由pcr扩增文库,然后转录以产生用于下一轮选择的文库。图16显示氧化石墨烯平行评估的图示。进行平行评估的富集文库分成四等份。然后,将文库样品添加至氧化石墨烯,且进行孵育以将文库装载在氧化石墨烯上。使用两次5分钟洗涤以移除未结合材料。对于阳性(阿昔洛韦)及特殊靶标(喷昔洛韦)样品,各靶标分别在1×选择缓冲剂中制备成1μm;反向靶标用10μm呈溶液形式的各反向靶标替换阳性靶标;阴性样品用等体积的无核酸酶水代替阳性靶标。然后将样品与其各自的氧化石墨烯制剂组合且进行孵育。孵育后,离心样品以回收其上清液,且通过nanodrop-1000分光光度计读数(thermofisherscientific;wilmington,de)来测定文库回收。在变性page上分析剩余文库样品。在用gel-star染色/脱色之后获取凝胶影像。对应于预期文库大小的条带被回收用于后续轮次的平行评估,其中阳性靶标阿昔洛韦替代阴性,反向及特殊靶标样品的预装载孵育物的反向靶标。自第二平行评估回收的材料用于测序及分析。图17显示针对阿昔洛韦的七种适配体候选物。通过quikfold3.0(zuker2003)在37℃及1mna+下计算每个适配体的自由能。序列使用专有算法进行识别。各序列中的下划线区域为pcr引物退火区域。图18显示针对喷昔洛韦的七种适配体候选物。通过quikfold3.0(zuker2003)在37℃及1mna+下计算每个适配体的自由能。序列使用专有算法进行识别。各序列中的下划线区域为pcr引物退火区域。图19a提供在pbmc中表达时测试淋巴增殖性/存活活性的il7rα变体的示意图。图19b提供显示在存在及不存在il-2下pbmc存活百分比的条形图。图20显示编码gfp,抗cd19嵌合抗原受体及etag(本文中称为f1-0-03)的慢病毒表达载体的示意图。图21a及图21b分别显示在用所指示的慢病毒颗粒转导来自供体12m的新鲜分离且未刺激的pbmc14小时后的第3天,第6天,第9天,第13天,第17天的总cd3+群体中的cd3+gfp+细胞百分比(%)的直方图及cd3+gfp+群体的绝对细胞计数/孔的直方图。每个条柱表示一式两份的平均值+/-sd。图22a及图22b分别显示在用所指示的慢病毒颗粒转导来自供体13f的新鲜分离且未刺激的pbmc14小时后的第3天及第6天的总cd3+群体中的cd3+gfp+细胞(%)的直方图及cd3+gfp+群体的绝对细胞计数/孔的直方图。请注意,“a”显示使用vsv-g假型化慢病毒颗粒(一式三份实验)的结果;“b”显示使用vsv-g假型化慢病毒颗粒以及添加至转导基质的okt3ab(1ug/ml)(一式两份实验)的结果;“c”显示使用在其表面上表达gpi锚定ucht1scfvfc的vsv-g假型化慢病毒颗粒(一式三份实验)的结果;及“d”显示使用在其表面上表达gpi锚ucht1scfvfc及gpi锚cd80或其功能性胞外片段的vsv-g假型化慢病毒颗粒(一式两份实验)的结果。每个条柱表示一式两份或一式三份的平均值+/-sd,如图22a所指示。图23a及图23b分别显示在用所指示的慢病毒颗粒转导来自供体12m的新鲜分离且未刺激的pbmc持续所指示的暴露时间(2h至20h)后的第3天,第6天及第9天的总cd3+群体中的cd3+gfp+细胞百分比(%)的直方图及cd3+gfp+群体的绝对细胞计数/孔的直方图。如所示,在平板或摇瓶中进行转导。每个条柱代表用vsv-g假型化慢病毒颗粒(“[vsv-g]”)的一式两份d平均值+/-sd;其他实验没有重复。图24a为慢病毒载体骨架f1-0-02的示意图,其包括驱动gfp及etag的表达的转基因表达盒以及合成的ef-1α启动子与gfp的上游内含子a.图24b显示将mirna插入至f1-0-02骨架的ef1α内含子a中。“1”表示ef1α重叠;“2”表示5'臂;“3”表示mirna15'茎;“4”表示环;“5”“表示mirna13'茎;“6”表示3'臂;“7”表示接头;“88”表示mirna25'茎;“9”表示mirna23'茎;“10”表示mirna35'茎;“11”表示mirna33'茎;“12”表示mirna45'茎;及“13”表示mirna43'茎。图25为显示靶向处于ef-1α启动子内含子中cd3ζ的mirna能够敲低cd3复合物的表达的图。图26为显示用含有mir-tcrα的复制缺陷型慢病毒颗粒转导的样品的δδct的直方图。δδct值表示相对于未经转导对照的各经转导样品中经处理mir-tcrαmirna的量。图27a至图27c为显示在经ph调节药剂处理或不经其处理的情况下cho-靶标1细胞的特异性溶解百分比的图。在图27a中,cho-靶标1细胞最初在ph6.7下,且在由箭头所指示的时间处,分别用nahco3或不用nahco3来处理实验孔(实线)及对照细胞(虚线)。在图27b中,cho-靶标1细胞最初在ph6.7下,且在由箭头所指示的时间处,分别用naoh或不用naoh来处理实验孔(实线)及对照细胞(虚线)。在图27c中,cho-靶标1细胞最初在ph7.4下,且分别用hcl或不用hcl来处理实验孔(实线)及对照细胞(虚线)。图28是显示通过dsc测量的阿昔洛韦不存在(圆形)或存在(方形)时f1a-795的热通量相对时间的图。图29是显示在施用pbs或碳酸氢盐之前及之后,携带cho异种移植肿瘤的小鼠中来自prosensefast探针的rfu百分比的图。定义如本文中所使用,术语“嵌合抗原受体”或“car”或“cars”是指经改造受体,其将抗原特异性移植至细胞上,例如t细胞,nk细胞,巨噬细胞及干细胞。本发明的car包括至少一个抗原特异性靶向区(astr)及胞内活化结构域(iad)且可包括柄(stalk),跨膜结构域(tm)及一个或多个共刺激结构域(csd)。在另一实施方式中,car为对两种不同抗原或表位具有特异性的双特异性car。在astr与靶标抗原特异性结合之后,iad活化胞内信号传导。举例而言,iad可以以非mhc限制的方式将t细胞特异性及反应性重定向至所选靶标,从而利用抗体的抗原结合特性。非mhc限制的抗原识别赋予表达car的t细胞不依赖于抗原处理而识别抗原的能力,因此绕过肿瘤逃逸的主要机制。此外,当在t细胞中表达时,car有利地不与内源性t细胞受体(tcr)α链及β链二聚化。如本文中所使用,术语“微环境”是指组织或身体的任何部分或区域,其与身体的组织或区域的其他区域相比具有恒定或暂时的,物理的或化学的差异。举例而言,如本文中所使用的“肿瘤微环境”是指存在肿瘤的环境,是肿瘤内的非细胞区域,以及直接位于肿瘤性组织外但并不属于癌细胞本身的胞内区室的区域。肿瘤微环境可指代肿瘤周围的任何及所有条件,包括为恶化过程创建结构性和/或功能性环境以存活和/或扩增和/或扩散的条件。举例而言,肿瘤微环境可包括条件的更改,这些条件例如但不限于:压力,温度,ph,离子强度,渗透压,重量摩尔渗透浓度,氧化应激,一种或多种溶质的浓度,电解质浓度,葡萄糖浓度,透明质酸浓度,乳酸或乳酸盐浓度,白蛋白浓度,腺苷水平,r-2-羟基戊二酸水平,丙酮酸浓度,氧浓度和/或氧化剂,还原剂或辅助因子的存在,以及本领域技术人员将理解的其他条件。如本文中交换地使用,术语“多核苷酸”及术语“核酸”指任何长度的核苷酸(核糖核苷酸或脱氧核糖核苷酸)的聚合形式。因此,该术语包括但不限于:单链,双链或多链的dna或rna,基因组dna,cdna,dna-rna杂交体,或包含嘌呤碱基和嘧啶碱基或其他天然,经化学方式或生物化学方式修饰的,非天然或衍生化的核苷酸碱基的聚合物。如本文中所使用,术语“抗体”包括多克隆抗体及单克隆抗体,包括完整的抗体及保留与抗原特异性结合的抗体片段。抗体片段可以是但不限于:片段抗原结合(fab)片段,fab'片段,f(ab')2片段,fv片段,fab'-sh片段,f(ab')2fv片段,fd片段,重组igg(rigg)片段,单链抗体片段,包括单链可变片段(scfv),二价scfv的,三价的scfv及单域抗体片段(例如,sdab,sdfv,纳米抗体)。该术语包括免疫球蛋白的遗传改造和/或其他修饰的形式,例如:胞内抗体(intrabodies),肽抗体(peptibodies),嵌合抗体,单链抗体,全人抗体,人源化抗体,包括抗体及非抗体蛋白质的抗原特异性靶向区的融合蛋白质,异源偶联抗体,多特异性抗体,例如,双特异性抗体,双功能抗体(diabodies),三功能抗体(triabodies)及四功能抗体(tetrabodies),串联双-scfv及串联三-scfv。除非另外陈述,否则术语“抗体”应理解为包括其功能性抗体片段。该术语亦包括完整或全长抗体,包括任何类别或亚类的抗体,包括igg及其亚类,igm,ige,iga和igd。如本文中所使用,术语“抗体片段”包括完整抗体的一部分,例如,完整抗体的抗原结合区或可变区。抗体片段的实例包括fab,fab',f(ab')2和fv片段;双功能抗体;线性抗体(zapata等,proteineng.8(10):1057-1062(1995));单链抗体分子;和由抗体片段形成的多特异性抗体。木瓜蛋白酶消化抗体产生两个相同的抗原结合片段(称为“fab”片段,每个片段具有单个抗原结合位点)和一个残余“fe”片段(一种反映容易结晶的能力的命名)。胃蛋白酶处理产生具有两个抗原结合位点且仍能够交联抗原的f(ab')2片段。如本文中交换地使用,术语“单链fv”,“scfv”或“sfv”抗体片段包括抗体的vh及vl结构域,其中这些结构域存在于单个多肽链中。在一些实施方式中,fv多肽进一步包括vh及vl结构域之间的多肽接头或间隔区,使得sfv能够形成抗原结合所需的结构。关于sfv的综述,参见pluckthuninthepharmacologyofmonoclonalantibodies,第113卷,rosenburg及moore编,springer-verlag,newyork,第269-315页(1994)。如本文中所使用,“天然存在”的vh结构域及vl结构域是指已从宿主分离而未进一步分子进化以改变其在特殊条件下以scfv的形式产生时的亲和力(例如美国专利8709755b2及申请案wo/2016/033331a1中公开的)的vh结构域及vl结构域。如本文中所使用,术语“亲合力”是指两种试剂的可逆结合的平衡常数,并表示为解离常数(kd)。亲合力可比抗体针对无关氨基酸序列的亲合力大至少1倍,至少2倍,至少3倍,至少4倍,至少5倍,至少6倍,至少7倍,至少8倍,至少9倍,至少10倍,至少20倍,至少30倍,至少40倍,至少50倍,至少60倍,至少70倍,至少80倍,至少90倍,至少100倍或至少1000倍,或更高。抗体对于靶蛋白的亲合力可以是例如,约100纳摩尔(nm)至约0.1nm,约100nm至约1皮摩尔(pm),或约100nm至约1飞摩尔(fm)或更高。如本文中所使用,术语“亲合性”是指两种或更多种试剂的复合物在稀释后对解离的抗性。关于抗体和/或抗原结合片段,术语“免疫反应性”及“优先结合”在本文中可互换使用。如本文中所使用,术语“结合”是指,归因于例如共价相互作用,静电相互作用,疏水相互作用及离子相互作用和/或氢键相互作用(包括例如盐桥及水桥的相互作用)而在两个分子之间的直接关联。非特异性结合指低于约10-7m的亲合力结合,例如,以10-6m,10-5m,10-4m等的亲合力结合。如本文中所使用,提及“细胞表面表达系统”或“细胞表面展示系统”是指细胞表面上展示或表达蛋白质或其部分。通常,生成表达与细胞表面蛋白质融合的目的蛋白质的细胞。举例而言,蛋白质表达为具有跨膜结构域的融合蛋白质。如本文中所使用,术语“元件”包括多肽(包括多肽的融合物,多肽的区域,及其功能性突变体或片段)及多核苷酸(包括微小rna及shrna的,以及其功能性突变体或片段)。如本文中所使用,术语“区”或“区域”是多肽或多核苷酸的任何区段。如本文中所使用,“结构域”是具有功能性和/或结构性特性的多肽或多核苷酸的区域。如本文中所使用,术语“柄(stalk)”或“柄结构域”是指柔性多肽连接区,其提供结构柔性以及与侧翼多肽区的间隔,且可由天然或合成的多肽组成。柄可源自免疫球蛋白(例如,igg1)的铰链或铰链区,其通常定义为从人igg1的glu216至pro230的延伸(burton(1985)molec.immunol,22:161-206)。可通过将形成重链间二硫键(s-s)的第一个半胱氨酸与最后一个半胱氨酸放置在相同位置来将其他的igg同种型的铰链区与igg1序列进行比对。柄可以是天然存在或非天然存在的,包括但不限于经改变的铰链区,如美国专利第5,677,425号所示。柄可包括衍生自任何类别或亚类的抗体的完整铰链区。柄也可包括源自cd8,cd28或在提供柔性且与侧翼区的间隔上提供类似功能的其他受体的区。如本文中所使用术语“分离的”是指将材料从其原始环境(例如,如果是天然存在的,则是天然环境)中移除。举例而言,存在于活体动物中天然存在的多核苷酸或多肽是未分离的,但与天然系统中共存材料的一些或全部分离的相同多核苷酸或多肽则是分离的。此类多核苷酸可以是载体的一部分和/或这类多核苷酸或多肽可以是组合物的一部分并且仍是分离的,因为这类载体或组合物并不是天然环境的一部分。如本文中所使用,“多肽”是通过肽键连接的氨基酸残基的单链。多肽既不折叠成固定结构,亦不具有任何翻译后修饰。“蛋白质”是折叠成固定结构的多肽。“多肽”和“蛋白质”在本文可互换使用。如本文中所使用,多肽可经“纯化”以移除多肽的天然环境的杂质组分,例如,会干扰多肽的诊断或治疗用途的材料,例如酶,激素及其他蛋白质或非蛋白质溶质。多肽可(1)纯化至按抗体重量计高于90%,高于95%或高于98%,如利用洛瑞法(lowrymethod)所测定,例如,高于99重量%,(2)借助于旋转杯顺序分析仪纯化至足以获得n-端或内部氨基酸序列的至少15个残基的程度,或(3)利用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sds-page)在还原条件或非还原条件下使用库马斯蓝或银染料纯化至均一。如本文中所使用,术语“免疫细胞”一般包括源自骨髓中产生的造血干细胞(hsc)的白血细胞(白细胞)。“免疫细胞”包括例如,淋巴细胞(t细胞,b细胞,自然杀伤(nk)细胞)及源自骨髓的细胞(嗜中性粒细胞,嗜酸性粒细胞,嗜碱性粒细胞,单核细胞,巨噬细胞,树突状细胞)。如本文中所使用,“t细胞”包括表达cd3的所有类型的免疫细胞,其包括辅助t细胞(cd4+细胞),细胞毒性t细胞(cd8+细胞),调节性t细胞(treg)及γ-δt细胞。如本文中所用,“细胞毒性细胞”包括cd8+t细胞,自然杀伤(nk)细胞,nk-t细胞,γδt细胞(cd4+细胞亚群)和嗜中性粒细胞(是能够介导细胞毒性反应的细胞)。如本文中所用,术语“干细胞”一般包括多能(pluripotent)或多潜能(multipotent)干细胞。“干细胞”包括例如胚胎干细胞(es),间充质干细胞(msc),诱导型多能干细胞(ips)及定向祖细胞(造血干细胞(hsc),骨髓衍生细胞等)。如本文中所使用,术语“治疗”,“处理”等是指获得期望的药理学和/或生理学效果。该效果就完全或部分地预防疾病或其症状而言可以是预防性的,和/或就部分或完全治愈疾病和/或由疾病引起的副作用而言可以是治疗性的。如本文中所使用的“治疗”涵盖哺乳动物(例如,人类)疾病的任何治疗,并且包括:(1)在可易患疾病但尚未被确诊为患有该疾病的受试者种预防所述疾病;(b)抑制疾病,即阻止其发展;及(c)缓解疾病,即导致疾病消退。如本文中互换地使用,术语“受试者”,“对象”,“宿主”和“患者”是指哺乳动物,包括但不限于人类,鼠类(例如,大鼠,小鼠),兔形目动物(例如,兔),非人类灵长类,人类,犬科动物,猫科动物,有蹄类动物(例如,马,牛,绵羊,猪,山羊)等。如本文中所使用,术语“治疗有效量”或“有效量”是指在向哺乳动物或其他受试者施用以用于治疗疾病时足以影响疾病的此类治疗的试剂的量或两种试剂的组合量。“治疗有效量”将根据试剂,疾病及其严重程度以及待治疗受试者的年龄,体重等而变化。如本文中所使用,术语“进化“或”演进“是指使用一种或多种突变方法以产生编码一种不同的多肽的不同多核苷酸,所述多肽自身为经改良的生物分子和/或有助于产生另一经改良的生物分子。“生理学”或“正常”或“正常生理学”条件为例如但不限于以下的条件:压力,温度,ph,离子强度,渗透压,摩尔渗透压浓度,氧化应激,一种或多种溶质的浓度,电解质浓度,葡萄糖浓度,透明质酸浓度,乳酸或乳酸盐浓度,白蛋白浓度,腺苷水平,r-2-羟基戊二酸水平,丙酮酸浓度,氧浓度和/或氧化剂,还原剂或辅助因子的存在,以及将被视为在给药部位或在作用部位的组织或器官上对于受试者在正常范围内的其他条件。如本文中所使用,“遗传修饰的细胞”包括含有外源性核酸的细胞,无论外源性核酸是否整合至细胞基因组中。如本文中所使用的“多肽”可包括部分或完整蛋白质分子以及任何翻译后或其他修饰。如本文中所使用的假型化元件(pseudotypingelement)可包括:“结合多肽”,其包括识别且结合靶标宿主细胞的一种或多种多肽(通常糖蛋白);及一种或多种“促融合多肽”,其介导逆转录病毒与靶标宿主细胞膜融合,从而使逆转录病毒基因组进入靶标宿主细胞。如本文中所使用的“结合多肽”也可被称为“t细胞和/或nk细胞结合多肽”或“靶标接合(engagement)元件”,“促融合多肽”也可被称为“促融合元件”。“静息”淋巴细胞(如,静息t细胞)是并不表达活化标记(如ki-67)的细胞周期的g0期的淋巴细胞。静息淋巴细胞可包括从未接触特异性抗原的原生t细胞及先前接触抗原而改变的记忆t细胞。“静息”淋巴细胞也可以称为“静态”淋巴细胞。如本文中所使用,“淋巴细胞清除(lymphodepletion)”涉及利用例如施用淋巴细胞清除剂减少受试者中淋巴细胞数目的方法。部分身体或全身分级辐射疗法也可导致淋巴细胞清除。淋巴细胞清除剂可以是能够在施用于哺乳动物时降低该哺乳动物中功能性淋巴细胞数目的化学化合物或组合物。这种药剂的一个实例为一种或多种化学治疗剂。这样的药剂及剂量是已知的,并且可以由治疗医师根据待治疗的对象选择。淋巴细胞清除剂的实例包括但不限于氟达拉滨(fludarabine),环磷酰胺,克拉屈滨(cladribine),地尼白介素(denileukindiftitox)或其组合。rna干扰(rnai)是rna分子通过中和靶向的rna分子而抑制基因表达或翻译的生物学过程。rna靶标可以是mrna,或可以是任何对通过rnai的功能性抑制敏感的其他rna。如本文中所使用,“抑制性rna分子”是指其存在在细胞内导致rnai且致使抑制性rna分子靶向的转录物表达降低的rna分子。如本文中所使用的抑制性rna分子具有能够形成rna双链体的5'茎及3'茎。抑制性rna分子可以是例如,mirna(内源性或人工)或shrna,mirna的前体(即pri-mirna或pre-mirna)或shrna,或dsrna,其作为经分离核酸转录或直接引入至细胞或受试者。如本文中所使用,“双链rna”或“dsrna”或“rna双链体”是指由两条链构成的rna分子。双链分子包括由杂交以形成双链rna结构的两条rna链构成,或由自身折叠以形成双链结构的单链rna构成。大多数(但不一定是全部)双链区中的碱基是碱基配对的。双链区包含与靶标rna互补的序列。与靶标rna互补的序列是反义序列,并且通常为18至29、19至29、19至21或25至28个核苷酸长,或在一些实施方式中在下限上为18、19、20、21、22、23、24、25,上限上为21、22、23、24、25、26、27、28、29或30个之间,其中给定范围总是有低于上限的下限。这种结构通常包括5'茎,环及由与各茎相邻且并非双链体的一部分的环所连接的3'茎。在某些实施方式中,所述环包含至少3、4、5、6、7、8、9或10个核苷酸。在另一些实施方式中,该环包含2至40、3至40、3至21或19至21个核苷酸;或在一些实施方式中,在下限为2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20个与上限为10、11、12、13、14、15、16、17、18、19、20、25、30、35或40个之间,其中给定范围总是有低于上限的下限。如本文中所使用术语“微小rna侧翼序列”是指包括微小rna处理元件的核苷酸序列。微小rna处理元件是有助于由前体微小rna产生成熟微小rna的最小核酸序列。通常这些元件位于侧接微小rna茎环结构的40个核苷酸序列之内。在一些情况下,在侧接微小rna茎环结构的5至4000个核苷酸的长度中的核苷酸序列延伸内发现微小rna处理元件。术语“接头(linker)”在用于提及多重抑制性rna分子时是指连接两个抑制性rna分子的连接构件。如本文中所使用,“重组逆转录病毒”是指不可复制的或“复制缺陷型”逆转录病毒,除非将其明确地指出是可复制逆转录病毒。术语“重组逆转录病毒”及“重组逆转录病毒颗粒”在本文中互换使用。这样的逆转录病毒/逆转录病毒颗粒可以是任何类型的逆转录病毒颗粒,其包括例如γ逆转录病毒及在说明性实施方式中的慢病毒。众所周知,此类逆转录病毒颗粒(例如慢病毒颗粒)通常是在包装细胞中通过用以下转染这类包装细胞来形成的:包括如gag,pol和rev的包装元件的质粒,包膜或假型化质粒(编码假型化元件)及转移的,基因组的或逆转录病毒(例如慢病毒)表达载体,该表达载体通常为在其上编码基因或其他目的编码序列的质粒。因此,逆转录病毒(例如慢病毒)表达载体包括在转染至细胞中后促进表达及包装的序列(例如,侧接例如psi包装元件及靶标异源编码序列的5'ltr及3'ltr)。术语“慢病毒”及“慢病毒颗粒”在本文中互换地使用。mirna的“构架(framework)”由围绕的mirna的“5'微小rna侧翼序列”和/或“3'微小rna侧翼序列”,以及在一些情况下分离mirna中茎环结构的茎的环序列组成。在一些实例中,“构架”源自天然存在的mirna,例如mir-155。术语“5'微小rna侧翼序列”及“5'臂”在本文中互换地使用。术语“3'微小rna侧翼序列”及“3'臂”在本文中互换地使用。如本文中所使用,术语“mirna前体”是指可酶促加工成mirna的任何长度的rna分子,例如初级rna转录物,pri-mirna或pre-mirna。应理解,本公开内容及本文所提供的方面及实施方式不限于所公开的特定示例,因此当然可有变化。还应理解,本文中所使用的术语仅用于公开特定示例和实施方式的目的,并非旨在限制,因为本公开的范围将仅由所附权利要求限制。在提供数值范围的情况下,应理解,除非上下文另有明确规定,否则在该范围的上限与下限之间的各个间值(interveningvalue)(至下限单位的十分之一)以及任何其他陈述的或在所述范围内的间值均涵盖在本公开内。这些较小范围的上限和下限可独立包括在较小范围中,并且也涵盖在本发明内,受限于所述范围内的任何特别排除的端值。当所述范围包括一个或两个端值的情况下,排除那些所包括的端值中的任一个或两个的范围也包括在本发明中。当给出多个范围的低值和多个高值时,本领域技术人员应认识到,所选范围将包括小于高值的低值。本说明书中所有标题均为易于阅读起见,且并非为限制性的。除非另外定义,否则本文中所使用的所有技术及科学术语均具有与本发明所属领域普通技术人员所通常理解相同的含义。尽管与本文所述相似或等效的任何方法和材料也可用于本发明的实践或测试中,但现在描述优选的方法及材料。本文中提及的所有出版物均通过引用并入本文,以公开和描述与所引用出版物相关的方法和/或材料。必须注意的是,除非上下文另外明确指示,否则如本文中所使用及在随附权利要求中所使用的,不使用数量词修饰时也包括复数指示物。因此,例如,提及“嵌合抗原受体”包括多种此类嵌合抗原受体及熟习此项技术者已知的等。应进一步注意,权利要求可被撰写成排除任何任选要素。因此,该陈述旨在用作使用与权利要求要素相关的这种排除性术语(例如“仅”等)或使用“排除性(negative)”限制的在先基础。应理解,为清楚起见,在独立实施方式上下文中描述的本发明的某些特征也可以组合方式提供于单个实施方式中。相反,为简洁起见,在单个实施方式的上下文中描述的本发明的多种特征也可单独提供或以任何适合的子组合方式提供。属于本发明的实施方式的所有组合均具体的包涵在本发明中,且正如各组合及每一组合单独及清楚地公开一样公开于本文中。另外,多种实施方式及其要素的所有子组合也具体包涵在本发明中,且正如各子组合及每个该类子组合单独及清楚地公开一样公开于本文中。发明详述本公开通过提供用于遗传修饰淋巴细胞的方法和组合物以及进行包括转导t细胞和/或nk细胞的过继细胞疗法的方法来克服这些现有技术挑战,其所需的离体时间短得多,例如24、12或者8小时或甚至更短,并且在一些实施方式中不需要预先离体刺激。这些方法非常适合于离体处理来自受试者血液的封闭系统,并且可以在受试者的血液或其分离的血细胞在执行方法的全部时间期间与受试者身处的同一房间和/或一些实施方式中在受试者的视线中的情况下进行。更特别地,本文中公开内容的方面及实施方式通过提供用于转导静息t细胞和/或静息nk细胞的方法来克服与目前过继性细胞疗法相关的问题,所述方法通常利用有助于复制缺陷型重组逆转录病毒颗粒与静息t细胞和/或静息nk细胞结合及融合的假型化元件,以促进复制缺陷型重组逆转录病毒颗粒对静息t细胞和/或静息nk细胞的遗传修饰。此外,本文中所提供的方法通过在说明性实施方式中利用嵌合抗原受体及一个或多个淋巴增殖性元件(其表达是在控制元件的控制下)克服了本领域问题,使得受试者暴露于结合控制元件的化合物,或终止此类暴露促进体内遗传修饰的t细胞和/或nk细胞的扩增。由于本文中详细公开的这些和其他优化,在一个方面中,本文中提供一种用于修饰受试者(例如患有疾病或病症的患者)的静息t细胞和/或静息nk细胞的方法,其中收集来自受试者的血液;通过使静息t细胞和/或nk细胞与复制缺陷型重组逆转录病毒颗粒接触来对其进行遗传修饰;且通常在比现有技术方法中更短的时间(例如在24小时内及在一些非限制性实施方式中为在12小时内)内和/或在未进一步离体扩增经遗传修饰的t细胞和/或nk细胞群的情况下(例如使得遗传修饰的静息t细胞和/或nk细胞未经历超过4次离体细胞分裂)而将经遗传修饰的细胞重新引入至受试者体内。因此,本文中所提供的方法可在比目前car疗法短很多的时间内执行,从而所提供的过程使得受试者可以在离体步骤的整个时间内留在诊所中。这有利于在封闭系统中执行离体步骤,降低了污染和混合患者样品的机会且可更容易地由临床实验室执行。因此,图1及图2提供用于本文中提供的方法的说明性组合物的示意图。图1提供包装细胞(100)和由该包装细胞产生的复制缺陷型重组逆转录病毒颗粒(200)的图。包装细胞(100)包括整合到其基因组中的重组多核苷酸(110),其包括表达逆转录病毒蛋白质的重组转录元件及在可诱导启动子(其由结合配体且由配体活化的反式激活因子调节)的控制下的多种不同膜结合多肽。这些反式激活因子,可诱导启动子及配体用于诱导细胞膜结合多肽的顺序表达和积累,所述细胞膜结合多肽将被整合入至复制缺陷型重组逆转录病毒颗粒的膜以及包装及组装复制缺陷型重组逆转录病毒颗粒所必需的逆转录病毒组分中。由于如下文中详细论述的多种多核苷酸的顺序诱导表达,产生图1中所示的说明性包装细胞(100),且可将该包装细胞用于说明性方法以产生用于本文中所提供的转染静息t细胞和/或nk细胞(图2中的(300))的方法中的复制缺陷型重组逆转录病毒颗粒。在非限制性说明实施方式中,包装细胞(100)在其基因组中包括编码可包装的逆转录病毒rna基因组的核酸,该可包装的逆转录病毒rna基因组包括为包装及组装复制缺陷型重组逆转录病毒颗粒所必需的逆转录病毒基因组元件中的至少一些(作为非限制性说明性实例,逆转录病毒psi元件,逆转录病毒的gag多肽及逆转录病毒pol多肽)。与包装细胞的细胞膜整合或相关联的一些膜结合多肽将整合或关联至复制缺陷型重组逆转录病毒颗粒中,但并不由逆转录病毒基因组编码。举例而言,包装细胞及由其形成的复制缺陷型重组逆转录病毒颗粒可包括:逆转录病毒vpx多肽(250),在非限制性说明性实施方式中其可表达为膜关联的融合蛋白质,例如src-flag-vpx多肽;可包括结合多肽及促融合多肽的假型化元件(240),在非限制性实施方式中,其包括麻疹病毒凝血素(h)多肽及麻疹病毒融合(f)多肽,或其细胞质结构域缺失变体;任选地,一个或多个活化元件(210、220),在非限制性实施方式中,其包括能够与cd3结合的膜结合多肽及能够与cd28结合的膜结合多肽;和/或任选地膜结合细胞因子(230),其非限制性实施方式为包括融合于daf的il-7的融合多肽或其片段。本文中提供了多种其他特定类型的这些膜结合多肽。作为由包装细胞顺序表达转录元件的结果,产生了复制缺陷型重组逆转录病毒颗粒。在包装细胞中且通常整合入包装细胞基因组中的rna逆转录病毒基因组成为复制缺陷型逆转录病毒颗粒的基因组,其包括逆转录病毒的生产,感染和整合至宿主细胞(通常为静息t细胞和/或nk细胞)的基因组中所必需的逆转录病毒组分(作为非限制性说明性实施方式,逆转录病毒的gag及pol多核苷酸)。此外,逆转录病毒基因组还包括编码本文中所提供的一种或通常两种经改造信号传导多肽的多核苷酸。经改造信号传导多肽中的一种通常编码至少一个淋巴增殖性元件(在非限制性实例中,组成型白介素7受体突变体),另一种经改造信号传导多肽通常编码嵌合抗原受体。然后,复制缺陷型重组逆转录病毒颗粒(200)在本文所提供的方法中转导静息t细胞和/或静息nk细胞(300)。如图2所展示,在静息t细胞和/或nk细胞(300)与复制缺陷型重组逆转录病毒颗粒(200)接触后,上述在复制缺陷型重组逆转录病毒颗粒表面上的膜多肽与静息t细胞和/或nk细胞(300)表面上的受体和/或配体结合。举例而言,如上文所指示可包括与静息t细胞和/或静息nk细胞表面分子结合的结合多肽及促融合多肽的假型化元件有助于复制缺陷型重组逆转录病毒颗粒(200)与t细胞和/或nk细胞膜的结合及融合。活化元件(210、220)通过接合t细胞受体复合物来活化静息t细胞和/或nk细胞(300),该过程在接触或之后的孵育过程中发生。此外,膜结合细胞因子(230)可存在于复制缺陷型重组逆转录毒颗粒表面上,并且结合静息t细胞和/或nk细胞(300)表面上的细胞因子受体(310),从而进一步促进结合及活化。因此,不受理论限制,在本文中所提供的说明性实施方式中,由于这种复制缺陷型重组逆转录病毒颗粒(200)组分中的一种或多种,不需要利用未在复制缺陷型重组逆转录病毒颗粒(200)中或其上的元件进行离体刺激或活化。这样帮助缩减本文中所提供的这种说明性方法中完成方法所需的离体时间。在与t细胞和/或nk细胞结合后(200),然后复制缺陷型重组逆转录病毒颗粒与t细胞和/或nk细胞(300)融合,复制缺陷型重组逆转录病毒颗粒中的多肽及核酸进入t细胞和/或nk细胞(300)。如上文所述,复制缺陷型重组逆转录病毒颗粒中的这些多肽之一是vpx多肽(250)。vpx多肽(250)与降解细胞质中游离的dntp的samhd1限制因子(350)结合并且诱导其降解。因此,细胞质中游离的dntp的浓度随着vpx降解samhd1而增加,且增加逆转录活性,从而有助于逆转录病毒基因组的逆转录及整合至t细胞和/或nk细胞基因组中。在将逆转录病毒基因组整合至t细胞和/或nk细胞中(200)之后,t细胞和/或nk细胞基因组包括编码信号传导多肽的核酸,其编码淋巴增殖性元件(370)及任选地信号传导多肽编码car(360)。淋巴增殖性元件及任选地car的表达处于控制元件的控制下。暴露于结合控制元件的化合物(可通过将该化合物施用于其t细胞和/或nk细胞(300)被转导的受试者中来在体外或体内发生)通过表达淋巴增殖性元件及任选地由于car表达和car与其靶细胞的结合来促进体外或体内中的t细胞和/或nk细胞(300)的增殖。因此,本文中用复制缺陷型重组逆转录病毒颗粒转导的t细胞和/或nk细胞具有驱动增殖和/或抑制细胞死亡的一个或多个信号,在说明性实施方式中,这样避免了需要在将经转导t细胞和/或nk细胞返回受试者中之前对宿主进行淋巴细胞清除的在前方法。在说明性实施方式中,这样进一步降低了在将经转导t细胞和/或nk细胞重新引入至受试者中前进行数天处理的需要。因此,在说明性实施方式中,自受试者采集血液至将血液重新引入至受试者所需时间不超过36小时,24小时,12小时,或在一些情况下甚至不超过8小时,这样从根本上改变了现有技术方法的car-t过程。此外,控制元件也提供本文中所提供的安全性机制之一。举例而言,停止施用化合物可下调或甚至终止淋巴增殖性元件及任选地car的表达,从而结束对于经转导t细胞和/或nk细胞及其子代的增殖和/或存活信号。用于执行过继性细胞疗法的方法在某些方面中,本文提供用于对受试者执行过继性细胞疗法的方法,作为一说明性实例,该方法可包括以下:a.采集受试者的血液;b.分离包含静息t细胞和/或静息nk细胞的外周血单核细胞(pbmc);c.使受试者的静息t细胞和/或静息nk细胞在离体的情况下与复制缺陷型重组逆转录病毒颗粒接触,其中所述复制缺陷型重组逆转录病毒颗粒在其表面上包含能够结合静息t细胞和/或nk细胞且有助于复制缺陷型重组逆转录病毒颗粒与其膜融合的假型化元件,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导静息t细胞和/或nk细胞,从而产生遗传修饰的t细胞和/或nk细胞;及d.在从受试者采集血液的36小时,24小时,12小时或甚至8小时内,将所述遗传修饰的细胞重新引入至受试者体内,从而在受试者中执行过继性细胞疗法。在本文中所提供的一些方面中,具有相似步骤的方法被称为用于遗传修饰和扩增受试者的淋巴细胞的方法。本领域技术人员应理解,本文中用于进行过继细胞疗法的方法和组合物的讨论也适用于对受试者的淋巴细胞进行遗传修饰和扩增的方法。通常,本公开的过继性细胞疗法是通过自体性转移来进行,其中细胞分离和/或以其他方式制备自接受细胞疗法的受试者,或制备自来自所述受试者的样品。因此,在一些方面中,细胞是源自需要治疗的受试者(例如,患者),并且在分离和处理后的细胞施用于同一受试者。在本文中所公开的方法及组合物的一些实施方式中,患有疾病或病症的受试者进入使用已知方法(例如静脉穿刺)抽取受试者血液的医疗机构中。在某些实施方式中,从受试者抽取的血液体积在范围下限为10ml,15ml,20ml,25ml,30ml,35ml,40ml,50ml,75ml或100ml至范围上限为200ml,250ml,300ml,350ml,400ml,500ml,750ml,1000ml,2000ml或2500ml之间。在一些实施方式中,从受试者抽取10ml至400ml之间的血液。在一些实施方式中,自受试者抽取20ml与250ml之间的血液。一些实施方式中,血液在处理时为新鲜的。在本文所公开的任何实施方式中,新鲜血液可以是在少于15分钟,30分钟,45分钟,60分钟,90分钟,120分钟,150分钟或180分钟之前从受试者抽取的血液。在一些实施方式中,以本文中所提供方法处理血液而不储存。t细胞和/或nk细胞与复制缺陷型重组逆转录病毒颗粒之间的接触通常有助于利用复制缺陷型重组逆转录病毒颗粒转导t细胞和/或nk细胞。在本公开中,经转导t细胞和/或nk细胞包括离体转导细胞的子代,其保留至少一些在离体转导过程中整合入细胞的核酸或多核苷酸。在本文描述“重新引入”转导细胞的方法中,应理解,当从受试者的血液中收集时,此类细胞通常不处于经转导的状态。本文公开的任何方面的受试者可以是例如动物,哺乳动物,并且在说明性实施方式中是人类。不受理论限制,在非限制性说明性方法中,将编码淋巴增殖性元件的多核苷酸(如il7组成型活化突变体)离体递送至静息t细胞和/或nk细胞(其可整合至t细胞或nk细胞的基因组中)提供了具有用于体内扩增的驱动子的细胞,而不需要宿主的淋巴细胞清除。因此,在说明性实施方式中,受试者在执行接触的1天,2天,3天,4天,5天,6天,7天,10天,14天,21天或28天内或1个月,2个月,3个月或6个月内,接触期间和/或在将经修饰的t细胞和/或nk细胞重新引入回受试者后的1天,2天,3天,4天,5天,6天,7天,10天,14天,21天或28天内或1个月,2个月,3个月或6个月内并不暴露于淋巴细胞清除试剂中。此外,在非限制性说明性实施方式中,可执行本文中所提供的方法而不在复制缺陷型重组逆转录病毒颗粒与受试者的静息t细胞和/或nk细胞接触的步骤期间或在整个离体方法期间使受试者暴露于淋巴细胞清除试剂中。因此,受试者体内扩增遗传修饰的t细胞和/或nk细胞的方法是本公开的一些实施方式的特征。在说明性实施方式中,此类方法为无离体增殖的或基本上无增殖的。在本文的非限制性说明性实施方式中,从受试者抽取血液到在离体转导t细胞和/或nk细胞之后将血液重新引入回至受试者体内的整个方法/过程可在以下时间内发生:少于48小时,少于36小时,少于24小时,少于12小时,少于11小时,少于10小时,少于9小时,少于8小时,少于7小时,少于6小时,少于5小时,少于4小时,少于3小时,2小时或少于2小时。在另一些实施方式中,在本文的非限制性说明性实施方式中,从受试者抽取/采集血液至在离体转导t细胞和/或nk细胞后将血液重新引入回受试者的整个方法/过程在以下时间内发生:1小时与12小时之间,或2小时与8小时之间,或1小时与3小时之间,或2小时与4小时之间,或2小时与6小时之间,或4小时与12小时之间,或4小时与24小时之间,或8小时与24小时之间,或8小时与36小时之间,或8小时与48小时之间,或12小时与24小时之间,或12小时与36小时之间,或12小时与48小时之间;或在以下时间内发生:在范围下限为15分钟,30分钟,60分钟,90分钟,120分钟,180分钟及240分钟与范围上限为120分钟,180分钟,及240分钟,300分钟,360分钟,420分钟及480分钟之间。在另一些实施方式中,从受试者抽取/采集血液至在离体转导t细胞和/或nk细胞之后将血液重新引入回受试者的整个方法/过程在以下时间内发生:在范围下限为1小时,2小时,3小时,4小时,6小时,8小时,10小时及12小时与范围上限为8小时,9小时,10小时,11小时,12小时,18小时,24小时,36小时或48小时之间。在一些实施方式中,经遗传修饰的t细胞和/或nk细胞在发生接触的时间段后与复制缺陷型重组逆转录病毒颗粒分离。在本文中的任何方法(包括血液采集步骤及转导淋巴细胞(在说明性实施方式中,t细胞和/或nk细胞,包括静息t细胞及nk细胞)的步骤)的一些实施方式中,从血液采集至转导t细胞和/或nk细胞的方法并不包括通过在黏着物基质上孵育在一个实施方式中超过4小时,或在另一实施方式中超过6小时或在又一实施方式中超过8小时来移除单核细胞的步骤。在一个说明性实施方式中,从血液采集至转导t细胞和/或nk细胞的方法并不包括在黏着物基质上过夜孵育以移除单核细胞。在另一实施方式中,从血液采集至转导t细胞和/或nk细胞的方法包括通过在黏着物基质上孵育不超过30分钟,1小时或2小时来移除单核细胞的步骤。在另一实施方式中,从受试者进行血液采集至转导淋巴细胞(在说明性实施方式中,t细胞和/或nk细胞,包括静息t细胞和/或nk细胞)的方法不包括通过在黏着物基质上孵育移除单核细胞的步骤。在另一实施方式中,从受试者进行血液采集至转导淋巴细胞(在说明性实施方式中,t细胞和/或nk细胞,包括静息t细胞和/或nk细胞)的方法包括,在该方法期间t细胞和/或nk细胞并不与牛血清(如细胞培养牛血清,例如胎牛血清)一起孵育或并不暴露于其中。在本文中的任何方法(包括血液采集的步骤及转导淋巴细胞(在说明性实施方式中,t细胞和/或nk细胞,包括静息t细胞及nk细胞)的步骤)的一些实施方式中,从受试者进行血液采集至重新引入t细胞和/或nk细胞至受试者中的方法并不包括通过在黏着物基质上孵育在一个实施方式中超过4小时,或在另一实施方式中超过6小时或在又一实施方式中超过8小时来移除单核细胞的步骤。在一个说明性实施方式中,从受试者进行血液采集至重新引入t细胞和/或nk细胞至受试者中的方法并不包括在黏着物基质上孵育过夜以移除单核细胞。在另一实施方式中,从受试者进行血液采集至重新引入t细胞和/或nk细胞至受试者中的方法包括通过在黏着物基质上孵育不超过30分钟,1小时或2小时来移除单核细胞的步骤。在另一实施方式中,从受试者进行血液采集至重新引入t细胞和/或nk细胞至受试者中的方法并不包括通过在黏着物基质孵育来移除单核细胞的步骤。在另一实施方式中,自受试者进行血液采集至重新引入t细胞和/或nk细胞至受试者中的方法,在该方法期间t细胞和/或nk细胞并不与牛血清(例如细胞培养牛血清,例如胎牛血清)一起孵育或并不暴露于其中。在包括转导t细胞和/或nk细胞步骤的本文中任何方法的一些实施方式中,在一些实施方式中,在转导之前,t细胞和/或nk细胞未曾暴露于在黏着单核细胞的基质上的孵育在一个实施方式中超过4小时,或在另一实施方式中超过6小时,或在又一实施方式中超过8小时。在一个说明性实施方式中,在转导之前,t细胞和/或nk细胞已在黏着物基质上孵育过夜以移除单核细胞。在另一实施方式中,该方法可包括,在转导之前在结合单核细胞的黏着物基质上孵育t细胞和/或nk细胞不超过30分钟,1小时或2小时。在另一实施方式中,在该转导步骤之前,t细胞和/或nk细胞并不暴露于通过在黏着物基质上孵育来移除单核细胞的步骤。在另一实施方式中,在转导之前或其期间,t细胞和/或nk细胞并不与牛血清(例如细胞培养牛血清,例如胎牛血清)一起孵育或并不暴露于其中。因为本文中所提供的用于过继性细胞疗法的方法和用于在体内扩增其之前离体修饰静息t细胞和/或静息nk细胞的相关方法可以以显著短于现有方法的时间来执行,使得可以对患者护理及安全性以及产品制造性进行重要改良。因此,在负责批准在体内进行用于治疗性目的的这种方法的管理机构看来,这种方法预期是有利的。举例而言,在非限制性实例中,在经修饰的t细胞和/或nk细胞重新引入至患者中之前的样品处理的整个过程,受试者可留在与处理其血液或样品的仪器所处的相同建筑(例如输注诊所)或房间中。在非限制性说明性实施方式中,在从受试者抽取/收集血液至在离体转导t细胞和/或nk细胞后将血液重新引入至受试者的整个方法/过程中,受试者保持在位置线内和/或正被处理的其血液或细胞的100、50、25或12英尺或手臂距离内。在另一些非限制性说明性实施方式中,在从受试者进行血液抽取/收集至在离体转导t细胞和/或nk细胞后将血液重新引入至受试者的整个方法/过程的整个期间和/或在此期间连续地使受试者保持清醒和/或至少一个人可继续监测受试者的正被处理的血液或细胞。因为本文中所提供的改良,自受试者进行血液抽取/收集至离体转导t细胞和/或nk细胞后将血液重新引入至受试者的用于过继性细胞疗法和/或转导静息t细胞和/或nk细胞的整个方法/过程可在人的持续监控下进行。在另一些非限制性说明性实施方式中,在从受试者进行血液抽取/收集至离体转导t细胞和/或nk细胞后将血液重新引入至受试者的整个方法/过程中从未在无人在场的房间中孵育血液细胞。在另一些非限制性说明性实施方式中,从受试者进行血液抽取/收集至离体转导t细胞和/或nk细胞后将血液重新引入至受试者的整个方法/过程是在受试者旁边和/或在与受试者相同的房间中和/或在受试者的床或椅子旁边处执行。因此,可避免样品混淆,以及避免经数天或数周的长时间且昂贵的孵育。这通过本文中所提供的方法容易适用于封闭及自动化的血液处理系统的事实进一步证实,其中将重新引入至受试者中的血液样品及其组分仅与单次、一次性使用的组件进行接触。用于执行本文中所提供的过继性细胞疗法的方法一般包括1)转导淋巴细胞(诸如t细胞或nk细胞,其在说明性实施方式中为静息t细胞和/或nk细胞)的方法,和/或包括2)用于遗传修饰淋巴细胞(诸如t细胞和/或nk细胞,其在说明性实施方式中为静息t细胞和/或nk细胞)的方法,其两者(1及2)自身各自形成本公开的不同方面。此类方法可在具有或不具有在本文中指出的用于执行过继性细胞疗法的其他步骤的情况下执行。本领域技术人员应认识到,本文在所提供的转导和/或遗传修饰t细胞和/或nk细胞的细节可应用于包括此类步骤的任何方面,包括关于转导和/或遗传修饰淋巴细胞(诸如t细胞和/或nk细胞)的方法的方面。因此,在某些方面中,本文中提供一种转导和/或遗传修饰t细胞和/或nk细胞(一般地,静息t细胞和/或静息nk细胞)的方法,该方法包括使静息t细胞和/或静息nk细胞与复制缺陷型重组逆转录病毒颗粒接触,其中复制缺陷型重组逆转录病毒颗粒一般包含在其表面上的假型化元件,该假型化元件能够结合静息t细胞和/或nk细胞且一般有助于以其自身或结合复制缺陷型重组逆转录病毒颗粒的其他蛋白质的与所述静息t细胞和/或nk细胞的膜融合,其中该接触(及在接触条件下的孵育)有助于利用复制缺陷型重组逆转录病毒颗粒来转导静息t细胞和/或nk细胞,从而产生遗传修饰的t细胞和/或nk细胞。此方法的另外实施方式可包括复制缺陷型重组逆转录病毒颗粒、淋巴增殖性元件、car、假型化元件、核糖开关、活化元件、膜结合细胞因子、mirna和/或本文中所揭示的其他元件的任意实施方式。用于转导t细胞和/或nk细胞的此方法可在体外或离体进行。因此,在一个方面中,本文中提供一种用于转导(和/或遗传修饰)淋巴细胞(一般来自经分离血液的静息t细胞和/或静息nk细胞)的方法,该方法包含:a.采集受试者血液;b.分离包含静息t细胞和/或静息nk细胞的外周血单核细胞(pbmc);及c.使受试者的静息t细胞和/或静息nk细胞在离体的情况下与复制缺陷型重组逆转录病毒颗粒接触,其中所述复制缺陷型重组逆转录病毒颗粒包含在其表面上的能够结合静息t细胞和/或静息nk细胞且有助于复制缺陷型重组逆转录病毒颗粒与其膜融合的假型化元件,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导至少5%的静息t细胞和/或静息nk细胞,从而产生遗传修饰的t细胞和/或nk细胞,从而转导静息t细胞和/或nk细胞。因此,在另一方面中,本文中提供一种用于遗传修饰或转导受试者的淋巴细胞(在说明性实施方式中,t细胞和/或nk细胞,或t细胞和/或nk细胞群)的方法,该方法包括使t细胞和/或nk细胞(通常为受试者的t细胞和/或nk细胞)在离体的情况下与复制缺陷型重组逆转录病毒颗粒接触,该复制缺陷型重组逆转录病毒颗粒在其基因组中包含多核苷酸,该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中该一个或多个核酸序列中的第一核酸序列编码针对一个或多个rna靶标的两个或更多个抑制性rna分子,且该一个或多个核酸序列中的第二核酸序列编码包含抗原特异性靶向区(astr),跨膜结构域及胞内活化结构域的嵌合抗原受体(car),其中所述接触有助于利用复制缺陷型重组逆转录病毒颗粒转导静息t细胞和/或nk细胞或至少一些静息t细胞和/或nk细胞,从而产生遗传修饰的t细胞和/或nk细胞。在另一方面中,本文中提供一种用于遗传修饰或转导受试者的淋巴细胞(例如,t细胞或nk细胞)或其群体的方法,所述方法包括使受试者的淋巴细胞(例如,t细胞或nk细胞)或其群体在离体的情况下与复制缺陷型重组逆转录病毒颗粒接触,该复制缺陷型重组逆转录病毒颗粒在其基因组中包含多核苷酸,该多核苷酸包含可操作地连接至在淋巴细胞(例如,t细胞和/或nk细胞)中有活性的启动子的一个或多个核酸序列,其中该一个或多个核酸序列中的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如两个或更多个)抑制性rna分子,且该一个或多个核酸序列中的第二核酸序列编码包含抗原特异性靶向区(astr),跨膜结构域及胞内活化结构域的嵌合抗原受体(car),其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒遗传修饰和/或转导淋巴细胞(例如,t细胞或nk细胞)或至少一些淋巴细胞(例如,t细胞和/或nk细胞),从而产生遗传修饰和/或经转导的淋巴细胞(例如,t细胞和/或nk细胞)。在此部分提供的方法的一些实施方式中,将遗传修饰和/或经转导的淋巴细胞(例如,t细胞和/或nk细胞)或其群体引入至受试者中。在一些实施方式中,遗传修饰和/或经转导的淋巴细胞(例如,t细胞和/或nk细胞)或其群体在引入或重新引入至受试者中之前进行4次或更少的离体细胞分裂。在一些实施方式中,淋巴细胞是与复制缺陷型重组逆转录病毒颗粒接触1小时与12小时之间的静息t细胞和/或静息nk细胞。在一些实施方式中,自受试者采集血液的时间与将遗传修饰的t细胞和/或nk细胞重新引入至受试者中的时间之间经过不超过8小时。在一些实施方式中,在采集血液后及重新引入血液前的所有步骤均在封闭系统中执行,其中在整个处理期间有人监控该封闭系统。在此部分提供的包括多核苷酸(该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中该一个或多个核酸序列中的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如两个或更多个)抑制性rna分子,且该一个或多个核酸序列中的第二核酸序列编码包含抗原特异性靶向区(astr),跨膜结构域及胞内活化结构域的嵌合抗原受体(car))的方法方面的任一者中,该多核苷酸可进一步包括编码不是抑制性rna分子的至少一个淋巴增殖性元件的第三核酸序列。在一些实施方式中,淋巴增殖性元件可以是细胞因子或细胞因子受体多肽,或其包含信号结构域的片段。在一些实施方式中,淋巴增殖性元件为组成型活化的。在某些实施方式中,淋巴增殖性元件可以是il-7受体或其片段。在说明性实施方式中,淋巴增殖性元件可以是组成型活化的il-7受体或其组成型活化的片段。在此部分提供的任何一种包括多核苷酸(该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中活性的启动子的一个或多个核酸序列,其中该一个或多个核酸序列中的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子)的方法方面中,抑制性rna分子在一些实施方式中可包括与彼此部分或完全互补的5'链及3'链,其中该5'链及该3'链能够形成18至25个核苷酸的rna双链体。在一些实施方式中,5'链的长度可以是18、19、20、21、22、23、24或25个核苷酸,且3'链的长度可以是18、19、20、21、22、23、24或25个核苷酸。在一些实施方式中,5'链及3'链的长度可相同或不同。在一些实施方式中,rna双链体可包括一个或多个错配。在替代实施方式中,rna双链体不具有错配。在此部分提供的任何一种包括多核苷酸(该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中该一个或多个核酸序列中的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子)的方法方面中,抑制性rna分子可以是mirna或shrna。在一些实施方式中,抑制性分子可以是mirna的前体,例如pri-mirna或pre-mirna,或shrna的前体。在一些实施方式中,抑制性分子可以是人工衍生的mirna或shrna。在另一些实施方式中,抑制性rna分子可以是被处理成的sirna的dsrna(经转录或人工引入)或sirna本身。在一些实施方式中,抑制性rna分子可以是mirna或shrna,其具有在自然界中未发现的序列,或具有在自然界中未发现的至少一个功能性区段,或具有在自然界中未发现的功能性区段的组合。在说明性实施方式中,至少一种或全部抑制性rna分子是mir-155。在此部分提供的任何一种包括多核苷酸(该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中该一个或多个核酸序列中的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或多个)抑制性rna分子)的方法方面中,在一些实施方式中,抑制性rna分子可自5'至3'方向包含:5'臂,5'茎,环,与该5'茎部分或完全互补的3'茎,及3'臂。在一些实施方式中,该两个或更多个抑制性rna分子中至少一个具有此排列。在另一些实施方式中,该两个或更多个抑制性分子中的所有均具有此排列。在一些实施方式中,5'茎的长度可以是18、19、20、21、22、23、24或25个核苷酸。在一些实施方式中,3'茎的长度可以是18、19、20、21、22、23、24或25个核苷酸。在一些实施方式中,环的长度可以是3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18,19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39或40个核苷酸。在一些实施方式中,5'臂,3'臂或两者均衍生自天然存在的mirna。在一些实施方式中,5'臂,3'臂或两者均衍生自天然存在的mirna,该mirna的选自:mir-155,mir-30,mir-17-92,mir-122及mir-21。在说明性实施方式中,5'臂,3'臂或两者均衍生自mir-155。在一些实施方式中,5'臂,3'臂或两者均衍生自小家鼠(musmusculus)mir-155或人(homosapiens)mir-155。在一些实施方式中,5'臂具有seqidno:256中所阐述的序列或为其功能性变体,例如这样的序列,其与seqidno:256长度相同,或为seqidno:256长度的95%,90%,85%,80%,75%或50%,或100个核苷酸或更少,95个核苷酸或更少,90个核苷酸或更少,85个核苷酸或更少,80个核苷酸或更少,75个核苷酸或更少,70个核苷酸或更少,65个核苷酸或更少,60个核苷酸或更少,55个核苷酸或更少,50个核苷酸或更少,45个核苷酸或更少,40个核苷酸或更少,35个核苷酸或更少,30个核苷酸或更少,或25个核苷酸或更少;并且与seqidno:256至少50%,55%,60%,65%,70%,75%,80%,85%,90%或95%相同。在一些实施方式中,3'臂具有阐述于seqidno:260中的序列或其功能性变体,例如这样的序列,其与seqidno:260长度相同或为seqidno:260长度的95%,90%,85%,80%,75%或50%或为100个核苷酸或更少,95个核苷酸或更少,90个核苷酸或更少,85个核苷酸或更少,80个核苷酸或更少,75个核苷酸或更少,70个核苷酸或更少,65个核苷酸或更少,60个核苷酸或更少,55个核苷酸或更少,50个核苷酸或更少,45个核苷酸或更少,40个核苷酸或更少,35个核苷酸或更少,30个核苷酸或更少,或25个核苷酸或更少的序列;并且与seqidno:260至少50%,55%,60%,65%,70%,75%,80%,85%,90%或95%相同。在一些实施方式中,3'臂包含小家鼠bic的第221至283位核苷酸。在此部分提供的任何一种包括多核苷酸(包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中该一个或多个核酸序列中的第一核酸序列编码针对一个或多个rna靶标的两个或更多个抑制性rna分子)方法方面中,在一些实施方式中,该两个或更多个抑制性rna分子可以以顺次排列的形式位于第一核酸序列中。在一些实施方式中,抑制性rna分子可利用非功能性接头序列直接或间接地与彼此连接。在一些实施方式中,接头序列的长度可在5个与120个核苷酸之间,或在10个与40个核苷酸之间。在此部分提供的任何一种包括多核苷酸(该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中该一个或多个核酸序列中的第一核酸序列编码针对一个或多个rna靶标的两个或更多个抑制性rna分子)的方法方面中,在一些实施方式中,该第一核酸序列编码两个至四个抑制性rna分子。在说明性实施方式中,第一核酸序列中包括2个与10个之间,2个与8个之间,2个与6个之间,2个与5个之间,2个与4个之间,3个与5个之间或3个与6个之间的抑制性rna分子。在一说明性实施方式中,第一核酸序列中包括四个抑制性rna分子。在此部分提供的任何一种包括多核苷酸(该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中该一个或多个核酸序列中的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如两个或更多个)抑制性rna分子)的方法方面中,该一个或多个(例如两个或更多个)抑制性rna分子可处于内含子中。在一些实施方式中,内含子在启动子中。在说明性实施方式中,内含子是ef-1α内含子a。在一些实施方式中,内含子与启动子相邻且在启动子下游,在说明性实施方式中,该内含子在用于产生复制缺陷型重组逆转录病毒颗粒的包装细胞中是无活性的。在此部分提供的任何一种包括多核苷酸(该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中该一个或多个核酸序列中的第一核酸序列编码针对一个或多个rna靶标的两个或更多个抑制性rna分子)的方法方面中,在一些实施方式中,该两个或更多个抑制性rna分子可针对不同靶标。在一替代实施方式中,两个或更多个抑制性rna分子针对同一靶标。在一些实施方式中,rna靶标是t细胞表达的基因转录的mrna,例如但不限于:pd-1(防止失活);ctla4(防止失活);tcrα(安全-防止自体免疫);tcrb(安全-防止自体免疫);cd3z(安全-防止自体免疫);socs1(防止失活);smad2(防止失活);mir-155靶标(促进活化);ifnγ(减少crs);ccbl(延长信号传导);trail2(防止死亡);pp2a(延长信号传导);abcg1(通过限制胆固醇的清除来增加胆固醇微区含量(microdomaincontent))。在一些实施方式中,rna靶标是从编码t细胞受体(tcr)复合物组分的基因转录的mrna。在一些实施方式中,两个或更多个抑制性rna分子中的至少一种可减少t细胞受体(在说明性实施方式中,t细胞的一个或多个内源性t细胞受体)的表达。在某些实施方式中,rna靶标可以是从t细胞内源性tcrα或tcrβ基因转录的mrna,该t细胞基因组包含编码一个或多个mirna的第一核酸序列。在说明性实施方式中,rna靶标是从tcrα基因转录的mrna。在此部分提供的任何一种包括多核苷酸(该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中该一个或多个核酸序列中的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如两个或更多个)抑制性rna分子,且该一个或多个核酸序列中的第二核酸序列编码包含抗原特异性靶向区(astr),跨膜结构域及胞内活化结构域的嵌合抗原受体(car))的方法方面中,在一些实施方式中,car是微环境限制的生物(mrb)-car。在另一些实施方式中,car的astr与肿瘤相关抗原结合。在另一些实施方式中,car的astr是微环境限制的生物(mrb)-astr。在此部分提供的任何一种包括多核苷酸(该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中该一个或多个核酸序列中的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如两个或更多个)抑制性rna分子,且该一个或多个核酸序列中的第二核酸序列编码包含抗原特异性靶向区(astr),跨膜结构域及胞内活化结构域的嵌合抗原受体(car))的方法方面中,在一些情况下,该一个或多个核酸序列中的第三核酸序列编码不是抑制性rna分子的至少一个淋巴增殖性元件,在一些实施方式中,该第一核酸序列,第二核酸序列及第三核酸序列中任意个或全部可操作地连接至核糖开关。在一些实施方式中,核糖开关能够结合核苷类似物。在一些实施方式中,核苷类似物为抗病毒药物。在用于过继性细胞疗法的方法及本文中提供的包括离体转导静息t细胞和/或静息nk细胞的任何方法中,通常在细胞与复制缺陷型重组逆转录病毒颗粒接触之前,将嗜中性粒细胞/粒细胞从血细胞中分离出来。在一些实施方式中,使用例如血球分离术和/或密度梯度离心法将包括外周血淋巴细胞(pbl)(例如t细胞和/或nk细胞)的外周血单核细胞(pbmc)与血液样品的其他组分分离。在一些实施方式中,在处理pbmc和/或t细胞和/或nk细胞,与复制缺陷型重组逆转录病毒颗粒接触,转导或转染之前,移除嗜中性粒细胞。关于待治疗的受试者,细胞可以是异体性和/或自体性的。作为非限制性实例,在一些实施方式中,在实施中使用sepax或sepax2细胞处理系统(biosafe)分离pbmc。在一些实施方式中,使用clinimacsprodigy细胞处理器(miltenyibiotec)分离pbmc。在一些实施方式中,使用自动采血分离器,其取得受试者的血液,使血液穿过分选出特定细胞类型(例如,pbmc)的装置,且使剩余部分返回至受试者中。密度梯度离心可在血球分离后执行。在一些实施方式中,使用去除白细胞的过滤器装置分离pbmc。在一些实施方式中,然后使用磁珠活化细胞分选以根据细胞表型(即阳性选择)来纯化来自pbmc的特定细胞群(例如,pbl或其子集)。也可使用其他用于纯化的方法,例如,基质黏着,其利用模拟t细胞在募集期间遭遇的环境的基质,从而使其黏着及迁移,或阴性选择,其中不需要的细胞利用抗体复合物(其靶向不需要的细胞)来靶向移除。在一些实施方式中,红细胞花结术(redbloodcellrosetting)可用于纯化细胞。在本文任何相关方面的一些说明性实施方案中,pbl包括t细胞和/或nk细胞。在本文某些实施方式中,在例如修饰淋巴细胞的方法和执行过继性细胞疗法的方法中,与本公开的复制缺陷型重组逆转录病毒颗粒接触的t细胞和/或nk细胞主要为静息t细胞。在一些实施方式中,t细胞和/或nk细胞由95-100%的静息细胞组成(ki-67-)。在一些实施方式中,与复制缺陷型重组逆转录病毒颗粒接触的t细胞和/或nk细胞包括在下限为90%,91%,92%,93%,94%及95%的静息细胞与上限为96%,97%,98%,99%或100%的静息细胞的范围之间。在一些实施方式中,t细胞和/或nk细胞包括原生细胞。在本文中所公开的方法及组合物的一些实施方式中,t细胞和/或nk细胞在离体的情况下与复制缺陷型重组逆转录病毒颗粒接触以遗传修饰t细胞和/或nk细胞,从而在重新引入至受试者中时在受试者中引起靶向免疫反应。在接触期间,复制缺陷型重组逆转录病毒颗粒识别t细胞和/或nk细胞且与t细胞和/或nk细胞结合,此时逆转录病毒与宿主细胞膜开始融合。然后,经由转导程序,来自复制缺陷型重组逆转录病毒颗粒的基因材料进入t细胞和/或nk细胞,且整合入至宿主细胞dna中。慢病毒转导的方法为已知的。例示性方法描述于例如wang等人(2012).j.immunother.35(9):689-701,cooper等人(2003)blood.101:1637-1644,verhoeyen等人(2009)methodsmolbiol.506:97-114,及cavalieri等人(2003)blood.102(2):497-505中。本文中所提供的许多方法包括转导t细胞和/或nk细胞。本领域已知用复制缺陷型重组逆转录病毒颗粒(例如复制缺陷型重组慢病毒颗粒)离体转导t细胞和/或nk细胞的方法。在说明性实施方式中,本文中提供的方法并不需要离体刺激或活化。因此,在本方法中可避免现有方法中这种常规步骤,但在转导期间可存在离体刺激性分子(例如抗cd3和/或抗cd28珠粒)。然而,利用本文中所提供的说明性方法,不需要离体刺激。在某些例示性方法中,可使用3至10感染复数(moi),且在一些实施方式中,5moi与10moi之间的单位的复制缺陷型重组逆转录病毒颗粒,例如慢病毒。如本文中所讨论的,转导反应可在封闭系统(例如sepax系统)中进行,其中转导反应可以在装载在系统上的一次性袋中进行。一旦将血细胞(例如pbmc)从粒细胞(包括嗜中性粒细胞)中分开,分离和/或纯化(这种粒细胞在接触步骤(即转导反应)期间通常不存在),就可以将来自受试者的采集的血液样品的这些血细胞与本文公开的复制缺陷型重组逆转录病毒在袋中接触。复制缺陷型重组逆转录病毒颗粒可被引入至含有经分离pbmc的袋中,从而接触pbmc。从采集受试者血液的时间至将血细胞(例如pbmc)添加至转导反应袋的时间可在30分钟至4小时之间,在30分钟至2小时之间,或在一些实例1小时左右。可将添加物例如培养基,人血清白蛋白,人ab+血清和/或源自受试者的血清添加至转导反应混合物中。通常存在培养基,例如本领域中已知用于离体处理的那些培养基(作为非限制性实例,x-vivo15(lonza)或cts培养基(thermofisherscientific)。可将支持性细胞因子添加至转导反应混合物,例如il2,il7或il15,或hsa中发现的那些。转导反应混合物可在23℃与39℃之间,及在一些说明性实施方式中在37℃下孵育。在某些实施方式中,转导反应可在37℃至39℃下进行以加快融合/转导。可将dgtp添加至转导反应。转导反应混合物可孵育1至12小时,且在一些实施方式中6至12小时。在转导后,在将转导的t细胞和/或nk细胞输注回受试者之前,将细胞从转导反应混合物中洗出。举例而言,在将转导的细胞输注回受试者之前,可以使用系统(例如sepax仪器)来洗涤细胞,例如用10ml至50ml的洗涤溶液。在一些实施方式中,在pbmc和/或t细胞和/或nk细胞被处理、与复制缺陷型重组逆转录病毒颗粒接触、被转导或转染之前,移除嗜中性粒细胞。在用于执行过继性细胞疗法的说明性实施方式中,采集受试者的血液至血袋中,且将该血袋连接至细胞处理系统,例如sepax细胞处理系统。将使用细胞处理系统分离的pbmc收集至袋中,使其与复制缺陷型重组逆转录病毒颗粒在足以转导t细胞和/或nk细胞的条件下接触,并对其进行孵育。在孵育之后,将含有pbmc与复制缺陷型重组逆转录病毒颗粒的混合物的袋连接至细胞处理系统,且洗涤pbmc。将经洗涤的pbmc收集至袋中,重新输注至受试者中。在一些实施方式中,从采集血液至重新输注转导t细胞和/或nk细胞的整个方法是在1小时、2小时、3小时、4小时、5小时、6小时、7小时、8小时、9小时、10小时、11小时、12小时、15小时、18小时或24小时内执行。在说明性实施方式中,整个方法是在12小时内执行。在一些实施方式中,复制缺陷型重组逆转录病毒颗粒的靶标细胞是pbl。在一些实施方式中,靶细胞是t细胞和/或nk细胞。在一些实施方式中,t细胞是辅助t细胞和/或杀伤t细胞。在一些实施方式中,本文中所提供的复制缺陷型重组逆转录病毒颗粒在其表面上具有能够结合t细胞和/或nk细胞并且促进复制缺陷型重组逆转录病毒颗粒与其膜融合的假型化元件。在另一些实施方式中,复制缺陷型重组逆转录病毒颗粒在其表面上具有能够结合静息t细胞和/或nk细胞的活化元件。在另一些实施方式中,复制缺陷型重组逆转录病毒颗粒在其表面上具有膜结合细胞因子。在一些实施方式中,复制缺陷型重组逆转录病毒颗粒包括具有一个或多个编码一种或多种经改造信号传导多肽的转录单元的多核苷酸,其中一种或多种经改造信号传导多肽包括一个或多个淋巴增殖性元件。在另一些实施方式中,当利用两种信号传导多肽时,一种包括至少一个淋巴增殖性元件,且另一种通常是包括抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域的嵌合抗原受体(car)。如本文中所指示,通常与本文中所提供的复制缺陷型重组逆转录病毒颗粒的表面缔合的活化元件能够接触静息t细胞和/或nk细胞足够时间,且由于该接触及适当的条件活化静息t细胞和/或nk细胞。应理解,这样的活化在本文方法的接触步骤期间随时间推移而发生。此外,应理解,在假型化元件在结合t细胞和/或nk细胞的复制缺陷型重组逆转录病毒颗粒的表面上的一些实施方式中,在本文的方法中,活化可通过结合假型化元件来诱导。活化元件在这些实施方式中是任选的。关于假型化元件、活化元件、膜结合细胞因子、经改造信号传导多肽、淋巴增殖性元件及car的更多细节在本文的其他部分中提供。在本文中所公开的方法及组合物的一些实施方式中,从血液收集的总淋巴细胞的5%至90%被转导。在某些实施方式中,经转导淋巴细胞的百分比在下限为5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%及60%与上限为50%、55%、60%、65%、70%、75%、80%、85%及90%的范围之间。在一些实施方式中,经转导的淋巴细胞的百分比为至少5%、至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少55%或至少60%。在本文中所公开的方法及组合物的一些实施方式中,将遗传修饰的t细胞和/或nk细胞引回、重新引入或重新输注至受试者中而无需额外离体操作,例如刺激和/或活化t细胞和/或nk细胞。在现有技术方法中,离体操作用于刺激/活化t细胞和/或nk细胞,及用于在将遗传修饰的t细胞和/或nk细胞引入至受试者中之前扩增遗传修饰的t细胞和/或nk细胞。在现有技术方法中,这通常花费数天或数周,且需要受试者在初始血液抽取数天或数周后返回诊所进行血液输注。在本文中所公开的方法及组合物的一些实施方式中,在t细胞和/或nk细胞与复制缺陷型重组逆转录病毒颗粒接触前,t细胞和/或nk细胞并不通过暴露于抗cd3/抗cd28固体载体(例如,涂布有抗cd3/抗cd28的珠粒)而进行离体刺激。因此,本文中提供一种离体的无繁殖(propagation-free)的方法。在另一些实施方式中,遗传修饰的t细胞和/或nk细胞并不离体扩增,或仅扩增较小数目的细胞分裂(例如1、2、3、4、5、6、7、8、9或10轮细胞分裂),而是在体内(即在受试者内)扩增或主要在体内扩增。在一些实施方式中,不添加额外介质以允许细胞进一步扩增。在一些实施方式中,在pbl与复制缺陷型重组逆转录病毒颗粒接触时,不发生pbl的细胞制造。在说明性实施方式中,当pbl离体时不发生pbl的细胞制造。在过继性细胞疗法的已有方法中,受试者在重新输注遗传修饰的t细胞和/或nk细胞前进行淋巴细胞清除。在一些实施方式中,患者或受试者在抽取血液前并不进行淋巴细胞清除。在一些实施方式中,患者或受试者在重新输注遗传修饰t细胞和/或nk细胞前并不进行淋巴细胞清除。在本文中所公开的任何一种实施方式中,待重新输注至受试者中的t细胞和/或nk细胞的数目可在下限为1×103、2.5×103、5×103、1×104、2.5×104、5×104、1×105、2.5×105、5×105、1×106、2.5×106、5×106及1×107个细胞/公斤与上限为5×104、1×105、2.5×105、5×105、1×106、2.5×106、5×106、1×107、2.5×107、5×107及1×108个细胞/公斤之间的范围。在说明性实施方式中,待重新输注至受试者中的t细胞和/或nk细胞的数目可在下限为1×104、2.5×104、5×104及1×105个细胞/公斤与上限为2.5×104、5×104、1×105、2.5×105、5×105及1×106个细胞/公斤之间的范围。在一些实施方式中,待重新输注至受试者中pbl的数目可少于范围的下限为5×105、1×106、2.5×106、5×106、1×107、2.5×107、5×107及1×108个细胞,与范围的上限为2.5×106、5×106、1×107、2.5×107、5×107、1×108、2.5×108、5×108及1×109个细胞。在一些实施方式中,可用于重新输注至70kg受试者或患者中的t细胞和/或nk细胞的数目在7×105个细胞与2.5×108个细胞之间。在另一些实施方式中,可用于转导的t细胞和/或nk细胞的数目大约为7×106个加或减10%。在本文中所公开的方法中,整个过继性细胞疗法程序(从抽取血液至重新输注遗传修饰的t细胞和/或nk细胞)可有利地在比已有方法更短的时间内执行。在一些实施方式中,整个过继性细胞疗法程序可在少于1小时、2小时、3小时、4小时、5小时、6小时、7小时、8小时、9小时、10小时、11小时、12小时、15小时、18小时或24小时中执行。在说明性实施方式中,整个过继性细胞疗法程序可在少于1小时、2小时、3小时、4小时、5小时、6小时、7小时、8小时、9小时、10小时、11小时或12小时中执行。在一些实施方式中,整个过继性细胞疗法程序可在下限为1小时、2小时、3小时、4小时、5小时、6小时、7小时、8小时、9小时、10小时、11小时、12小时或15小时与上限为1小时、2小时、3小时、4小时、5小时、6小时、7小时、8小时、9小时、10小时、11小时、12小时、15小时、18小时或24小时之间的范围执行。在本文中所提供的一些实施方式中,从受试者抽取血液样品、使t细胞和/或nk细胞与复制缺陷型重组逆转录病毒颗粒接触、和/或将遗传修饰的t细胞和/或nk细胞引入至受试者中的步骤发生在封闭系统中。封闭系统是对污染通常封闭或完全封闭的培养方法。本发明的一个优点是本文中提供的是用于在封闭系统中执行car疗法的方法。在细胞处理程序中的安全和监管控制的最大风险之一是通过频繁暴露于环境中而造成污染的风险,正如在传统开放式细胞培养系统中所发现的。为了减轻这样的风险,尤其在没有使用抗生素的情况下,已研发出致力于使用一次性(单次使用)设备的一些商业流程。然而,即使在无菌条件下使用,仍然存在开瓶以加样或添加额外生长培养基的污染风险。为了克服这个问题,本文中提供了封闭系统方法,一种经设计且可经操作使得产物并不暴露于外部环境的方法。这点是很重要的,因为外部环境通常不是无菌的。材料转移通过无菌连接或管的接合进行。用于气体交换的空气通过透气膜或类似其他添加剂经由0.2μm过滤器进行以防止环境暴露。在一些实施方式中,封闭系统包括连接至受试者的体内循环系统的离体循环系统,使得血液被抽取然后在被引回至受试者之前循环至离体循环系统。在一些实施方式中,离体循环系统包括与用于将细胞暴露于复制缺陷型重组逆转录病毒颗粒的系统或装置结合的用于分离pbl的系统或装置和/或用于分离t细胞和/或nk细胞的系统或装置。在一些实施方式中,封闭系统并不允许t细胞和/或nk细胞暴露于空气。此类封闭系统方法可利用市售装置来执行。举例而言,该方法可在适用于封闭系统t细胞生产的装置中进行。此类装置包括g-rextm、wavebioreactortm、origenpermalifetm袋及袋。在本文中所公开的方法及组合物的一些实施方式中,将受试者体内遗传修饰的t细胞和/或nk细胞暴露于结合其中存在的体内控制元件的化合物,其中控制元件是由复制缺陷型重组逆转录病毒颗粒引入的基因材料的一部分。在一些实施方式中,控制元件可以是核糖开关,且化合物可结合核糖开关的适配体结构域。在一些实施方式中,控制元件可以是分子伴侣。在本文所公开的任何一种实施方式中,化合物可以是核苷类似物。在一些实施方式中,核苷类似物可以是核苷类似抗病毒药物,其中抗病毒药物是由食品与药物管理局(foodanddrugadministration)针对抗病毒治疗批准的化合物或美国抗病毒临床试验中的化合物。在说明性实施方式中,化合物可以是阿昔洛韦或喷昔洛韦。在一些实施方式中,化合物可以是泛昔洛韦(famciclovir)(喷昔洛韦的口服前药)或伐昔洛韦(valaciclovir)(阿昔洛韦的口服前药)。化合物与控制元件的结合影响引入的遗传物质的表达,且因此影响遗传修饰的t细胞和/或nk细胞的增殖。在一些实施方式中,在从受试者的血液分离pbl之前、同时和/或之后,且在t细胞和/或nk细胞与复制缺陷型重组逆转录病毒颗粒接触之前,将核苷类似抗病毒药物或前药(例如阿昔洛韦、伐昔洛韦、喷昔洛韦或泛昔洛韦)施用至受试者。在一些实施方式中,在从血液分离pbl之前或在t细胞和/或nk细胞与复制缺陷型重组逆转录病毒颗粒接触之前,在下限为5分钟、10分钟、15分钟、30分钟及60分钟与上限为1.5小时、2小时、3小时、4小时、5小时、6小时、8小时、12小时或24小时之间的范围,将核苷类似抗病毒药物或前药施用至受试者。在另一些实施方式中,在从血液分离pbl及在t细胞和/或nk细胞与本文所提供的方法中的复制缺陷型重组逆转录病毒颗粒接触之后,在下限为1.5小时、2小时、3小时、4小时、5小时、6小时、8小时、12小时或24小时与上限为1/2天、1天、2天、3天、4天、5天、6天、7天、10天、14天、21天或28天之间的范围,将核苷类似抗病毒药物或前药施用至受试者。在一些实施方式中,在从血液分离pbl及在t细胞和/或nk细胞与本文所提供的方法中的复制缺陷型重组逆转录病毒颗粒接触之后的至少1.5小时、2小时、3小时、4小时、5小时、6小时、8小时、12小时或24小时,或至少2天、3天、4天、5天、6天、7天、10天、14天、21天或28天,将核苷类似抗病毒药物或前药施用至受试者。在一些实施方式中,在已将pbl重新输注至受试者之后的至少1天、2天、3天、4天、5天、7天、10天、14天、21天、28天、30天、60天、90天或120天,或5个月、6个月、9个月、12个月、24个月、36个月、48个月、60个月、72个月、84个月、96个月、120个月或不确定的长时期内,将核苷类似抗病毒药物或前药施用至受试者。在本文所公开的任何一种实施方式中,核苷类似抗病毒药物或前药可在pbl重新输注之前和/或期间和/或已将pbl重新输注之后施用。在一些实施方式中,与控制元件结合的化合物每日一次,两次,三次或四次施用至受试者。在一些实施方式中,提供日剂量的化合物1周、2周、4周、3个月、6个月、1年,直至受试者无病(例如无癌症)或不确定的长时期内地提供。在说明性实施方式中,药物是与核苷类似物结合的核苷类似抗病毒药物,例如核糖开关,如在本文中进一步详细公开的。本领域已知用于递送药物的方法,无论小分子还是生物制剂,并且可以用于本文所提供的方法。任何这样的方法均可用于递送用于本发明的方法中的药物或候选化合物或抗体。举例而言,常见的给药途径包括非侵入性经口(经由口)、局部(皮肤)、经黏膜(鼻腔、颊/舌下、阴道、眼睛及直肠)和吸入途径。许多蛋白质和肽药物(例如单克隆抗体)必须通过注射或纳米针阵列来递送。举例而言,许多免疫接种基于蛋白质药物的递送,且通常通过注射来进行。经改造的信号传导多肽在一些实施方式中,用于接触t细胞和/或nk细胞的复制缺陷型重组逆转录病毒颗粒具有多核苷酸,该多核苷酸具有编码一种或多种经改造信号传导多肽的一个或多个转录单元,其中一个或多个包括至少一个淋巴增殖性元件。在一些实施方式中,信号传导多肽包括以下的任何组合:胞外抗原结合结构域(或抗原特异性靶向区或astr)、柄、跨膜结构域、胞内活化结构域、淋巴增殖性元件、调节结构域(例如共刺激结构域)及t细胞存活基序。在说明性实施方式中,经改造信号传导多肽中至少一个、两个或全部为car。在一些实施方式中,当利用两种信号传导多肽时,一种编码一个或多个淋巴增殖性元件,且另一种编码包括抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域的嵌合抗原受体(car)。在另一些实施方式中,car可包括与抗原特异性靶向区融合的淋巴增殖性元件。在另一些实施方式中,当淋巴增殖性元件为组成型活化的白介素受体(例如il-7rα的已知变体)时,不需要抗原特异性靶向区,这是因为结合并不依赖于配体的存在。本领域普通技术人员将能够针对本文中所公开的方法及组合物,将系统进行再配置以将淋巴增殖性元件及car放在具有类似或不类似的控制元件的不同多核苷酸上。本领域技术人员应认识到,这种经改造多肽亦可称作重组多肽。抗原特异性靶向区在一些实施方式中,经改造信号传导多肽包括特异性结合对的成员,通常为astr,有时在本文中称作抗原结合结构域。特异性结合对包括但不限于抗原-抗体结合对;配体-受体结合对;以及类似者。因此,适用于本公开的经改造信号传导多肽中的特异性结合对成员包括astr,其为抗体、抗原、配体、配体的受体结合结构域、受体、受体的配体结合结构域,以及亲和抗体(affibody)。适用于本公开的经改造信号传导多肽中的astr可以是任何抗原结合多肽。在某些实施方式中,astr为抗体,例如全长抗体、单链抗体、fab片段、fab'片段、(fab')2片段、fv片段,以及二价单链抗体或双功能抗体。在一些实施方式中,astr为单链fv(scfv)。在一些实施方式中,重链位于经改造信号传导多肽中的轻链的n端。在另一些实施方式中,轻链位于经改造信号传导多肽中的重链的n端。在任何公开的实施方式中,重链和轻链可以通过接头隔开,如本文中更详细论述的。在任何公开的实施方式中,重链或轻链可在经改造信号传导多肽的n端且通常为另一结构域(例如信号序列或信号肽)的c端。另一些基于抗体的识别结构域(cabvhh(骆驼抗体可变结构域)及人源化版本,ignarvh(鲨鱼抗体可变结构域)及人源化版本,sdabvh(单一域抗体可变结构域)及“骆驼化”抗体可变结构域)适宜与经改造信号传导多肽一起使用且适用于使用本公开的经改造信号传导多肽的方法中。在一些情况下,也适宜使用基于t细胞受体(tcr)的识别结构域,例如单链tcr(sctv,含有vαvβ的单链两域tcr)。在一些实施方式中,astr可以是多特异性的,例如双特异性抗体。多特异性抗体对至少两个不同位点具有结合特异性。在某些实施方式中,结合特异性中之一针对一种靶标抗原且另一种针对另一种靶标抗原。在某些实施方式中,双特异性抗体可结合至ta靶向抗原的两个不同的表位。双特异性抗体也可用于将细胞毒剂定位至表达靶标抗原的细胞。双特异性抗体可制备为全长抗体或抗体片段。适用于本公开的经改造信号传导多肽的astr可具有多种抗原结合特异性。在一些情况下,抗原结合结构域对于由靶标细胞表达(由其合成)的抗原中存在的表位具有特异性。在一个实例中,靶标细胞是癌细胞相关的抗原。癌细胞相关抗原可以是与以下相关的抗原:例如乳腺癌细胞、b细胞淋巴瘤、霍奇金淋巴瘤细胞、卵巢癌细胞、前列腺癌细胞、间皮瘤、肺癌细胞(例如,小细胞肺癌细胞)、非霍奇金b细胞淋巴瘤(b-nhl)细胞、卵巢癌细胞、前列腺癌细胞、间皮瘤细胞、肺癌细胞(例如,小细胞肺癌细胞)、黑色素瘤细胞、慢性淋巴细胞白血病细胞、急性淋巴细胞白血病细胞、神经母细胞瘤细胞、神经胶质瘤、神经胶母细胞瘤、神经管胚细胞瘤、结直肠癌细胞等。癌细胞相关抗原也可由非癌细胞表达。经改造信号传导多肽的astr可结合的抗原的非限制性实例包括例如cd19、cd20、cd38、cd30、erbb2、ca125、muc-1、前列腺特异性膜抗原(psma)、cd44表面黏着分子、间皮素、癌胚抗原(cea)、表皮生长因子受体(egfr)、egfrviii、血管内皮生长因子受体-2(vegfr2)、高分子量黑色素瘤相关抗原(hmw-maa)、mage-al、il-13r-a2、gd2、axl、ror2,以及类似者。在一些情况下,适用于经改造信号传导多肽的特异性结合对的成员是作为受体的配体的astr。配体包括但不限于:细胞因子(例如,il-13等);生长因子(例如,表皮生长因子受体调节蛋白(heregulin);血管内皮生长因子(vegf);以及类似者);整合素结合肽(例如包含序列arg-gly-asp的肽);以及类似者。当经改造信号传导多肽中的特异性结合对的成员是配体时,可在特异性结合对的第二成员存在下使经改造信号传导多肽活化,其中特异性结合对的第二成员是该配体的受体。举例而言,当配体为vegf时,特异性结合对的第二成员可以是vegf受体,包括可溶性vegf受体。如上所述,在一些情况下,包括在经改造信号传导多肽中的特异性结合对的成员是作为受体的astr,例如配体的受体、共受体等。该受体可以是受体的配体结合片段。合适受体包括但不限于:生长因子受体(例如,vegf受体);杀伤细胞凝集素样受体子家族k;成员1(nkg2d)多肽(mica、micb及ulb6的受体);细胞因子受体(例如,il-13受体;il-2受体等);cd27;自然细胞毒性受体(ncr)(例如,nkp30(ncr3/cd337)多肽(hla-b相关转录物3(bat3)及b7-h6的受体等)等。柄在一些实施方式中,经改造信号传导多肽包括位于经改造信号传导多肽部分中的柄,该部分位于细胞外部且插入在astr与跨膜结构域之间。在一些情况下,该柄与野生型cd8柄区(tttpaprpptpaptiasqplslrpeacrpaaggavhtrgldfa(seqidno:79))具有至少85%、90%、95%、96%、97%、98%、99%或100%同一性,与野生型cd28柄区(fckievmypppyldneksngtiihvkgkhlcpsplfpgpskp(seqidno:80))具有至少85%、90%、95%、96%、97%、98%、99%或100%同一性,或与野生型免疫球蛋白重链柄区具有至少85%、90%、95%、96%、97%、98%、99%或100%同一性。在经改造信号传导多肽中,所用的柄允许抗原特异性靶向区(特别是整个经改造信号传导多肽)保持与靶标抗原的增加的结合。柄区的长度可以是约4个氨基酸至约50个氨基酸,例如约4个氨基酸至约10个氨基酸、约10个氨基酸至约15个氨基酸、约15个氨基酸至约20个氨基酸、约20个氨基酸至约25个氨基酸、约25个氨基酸至约30个氨基酸、约30个氨基酸至约40个氨基酸,或约40个氨基酸至约50个氨基酸。在一些情况下,经改造信号传导多肽的柄包括至少一个半胱氨酸。举例而言,在一些情况下,柄可包括序列cys-pro-pro-cys(seqidno:62)。若存在,第一经改造信号传导多肽的柄中的半胱氨酸可以与第二经改造信号传导多肽中的柄形成二硫键。柄可包括本领域已知的免疫球蛋白铰链区氨基酸序列;参见例如tan等人,(1990)proc.natl.acad.sci.usa87:162;及huck等人(1986)nucl.acidsres.14:1779。作为非限制性实例,免疫球蛋白铰链区可包括与以下氨基酸序列中任何一个的至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:dktht(seqidno:63);cppc(seqidno:62);cpepkscdtpppcpr(seqidno:64);(参见例如glaser等人(2005)j.biol.chem.280:41494);elktplgdttht(seqidno:65);kscdkthtcp(seqidno:66);kccvdcp(seqidno:67);kygppcp(seqidno:68);epkscdkthtcppcp(seqidno:69)(人类iggl铰链);erkccvecppcp(seqidno:70)(人类igg2铰链);elktplgdtthtcprcp(seqidno:71)(人类igg3铰链);spnmvphahhaq(seqidno:72)(人类igg4铰链);以及类似者。柄可包括具有人类igg1、igg2、igg3或igg4铰链区的氨基酸序列的铰链区。柄与野生型(天然存在的)铰链区相比可包括一个或多个氨基酸替换和/或插入和/或缺失。举例而言,人类igg1铰链的his229可被tyr替换,使得柄包括序列epkscdktytcppcp(参见例如yan等人(2012)j.biol.chem.287:5891)。柄可包括来源于人类cd8的氨基酸序列;例如,该柄可包括氨基酸序列:tttpaprpptpaptiasqplslrpeacrpaaggavhtrgldfacd(seqidno:73),或其变体。跨膜结构域本公开的经改造信号传导多肽可包括用于插入至真核细胞膜中的跨膜结构域。跨膜结构域可插入在astr与共刺激结构域之间。跨膜结构域可插入在柄与共刺激结构域之间,使得嵌合抗原受体自氨基端(n端)至羧基端(c端)依次包括:astr、柄、跨膜结构域以及活化结构域。提供将多肽插入至真核(例如,哺乳动物)细胞的细胞膜中的任何跨膜(tm)结构域适用于本发明的方面及实施方式中。适用于任何本文中所提供的方面或实施方式中的tm结构域的非限制性实例包括与以下任一tm结构域中的至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:a)cd*α(iyiwaplagtcgvlllslvitlyc(seqidno:46));b)cd8β(lgllvagvlvllvslgvaihlcc(seqidno:47));c)cd4(alivlggvaglllfiglgiffcvrc(seqidno:48));d)cd3z(lcylldgilfiygviltalflrv(seqidno:49);e)cd28(fwvlvvvggvlacysllvtvafiifwv(seqidno:50));f)cd134(ox40):(vaailglglvlgllgplaillalyll(seqidno:51));g)cd7(alpaalavisfllglglgvacvla(seqidno:52));h)cd8tttpaprpptpaptiasqplslrpeacrpaaggavhtrgldfacdiyiwaplagtcgvlllslvitlyc(seqidno:75);以及i)cd28ievmypppyldneksngtiihvkgkhlcpsplfpgpskpfwvlvvvggvlacysllvtvafiifwv(seqidno:76)。作为非限制性实例,本发明一个方面的跨膜结构域可与以下具有至少80%、90%或95%的序列同一性:seqidno:46跨膜结构域、cd8β跨膜结构域、cd4跨膜结构域、cd3ζ跨膜结构域、cd28跨膜结构域、cd134跨膜结构域,或cd7跨膜结构域。胞内活化结构域适用于本公开的经改造信号传导多肽中的胞内活化结构域在活化时通常诱导产生一种或多种细胞因子;增加细胞死亡;和/或增加cd8+t细胞、cd4+t细胞、自然杀伤t细胞、γδt细胞和/或嗜中性粒细胞的增殖。活化结构域在本文中也可称为活化域。在一些实施方式中,胞内活化结构域包括至少一个(例如,一个、两个、三个、四个、五个、六个等)如下文描述的itam基序。在一些实施方式中,胞内活化结构域包括dap10/cd28型信号传导链。在一些实施方式中,胞内活化结构域并未共价连接至膜结合的经改造信号传导多肽,而是扩散在细胞质中。作为非限制性实例,本发明一个方面的胞内活化结构域可与如下文所描述的cd3z、cd3d、cd3e、cd3g、cd79a、dap12、fcerlg、dap10/cd28或zap70结构域具有至少80%、90%或95%的序列同一性。适用于本公开的经改造信号传导多肽的胞内活化结构域包括含基于免疫受体酪氨酸的活化基序(itam)的细胞内信号传导多肽。itam基序为yx1x2l/i,其中x1及x2独立地是任何氨基酸。在一些情况下,经改造信号传导多肽的胞内活化结构域包括1个、2个、3个、4个或5个itam基序。在一些情况下,itam基序在胞内活化结构域中重复两次,其中itam基序的第一次出现与第二次出现彼此分开6个至8个氨基酸(例如,(yx1x2l/i)(x3)n(yx1x2l/i),其中n为整数6至8,且6个至8个x3中的每一个可以是任何氨基酸。在一些情况下,经改造信号传导多肽的胞内活化结构域包括3个itam基序。合适的胞内活化结构域可以是来源于含有itam基序的多肽的含itam基序部分。举例而言,合适的胞内活化结构域可以是来自任何含有itam基序的蛋白质的含有itam基序的结构域。因此,合适的胞内活化结构域不需要含有其所源自的整个蛋白质的完整序列。合适的含有itam基序的多肽的实例包括但不限于:cd3z(cd3ζ);cd3d(cd3δ);cd3e(cd3ε);cd3g(cd3γ);cd79a(抗原受体复合物相关的蛋白质α链);dap12;以及fcerlg(fcε受体iγ链)。在一些情况下,胞内活化结构域来源于t细胞表面糖蛋白cd3ζ链(也称为cd3z、t细胞受体t3ζ链、cd247、cd3-ζ、cd3h、cd3q、t3z、tcrz等)。举例而言,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链或与以下任一氨基酸序列(2个同功异型物)中的约100个氨基酸至约110个氨基酸(aa)、约110aa至约115aa、约115aa至约120aa、约120aa至约130aa、约130aa至约140aa、约140aa至约150aa或约150aa至约160aa的连续段具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:或其中itam基序为加粗且加下划线的。同样,合适的胞内活化结构域多肽可包括全长cd3ζ氨基酸序列的含itam基序的部分。因此,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链或与以下任一氨基酸序列中的约100个氨基酸至约110个氨基酸(aa)、约110aa至约115aa、约115aa至约120aa、约120aa至约130aa、约130aa至约140aa、约140aa至约150aa或约150aa至约160aa的连续段具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:或其中itam基序为加粗且加下划线的。在一些情况下,胞内活化结构域来源于t细胞表面糖蛋白cd3δ链(也称为cd3d、cd3-δ、t3d、cd3抗原、δ亚单位、cd3δ、cd3d抗原、δ多肽(tit3复合物)、okt3、δ链、t细胞受体t3δ链、t细胞表面糖蛋白cd3δ链等)。因此,合适的胞内活化结构域可包括与以下序列中的至少10个、15个、20个或全部氨基酸的一段链或与以下任何氨基酸序列的约100个氨基酸至约110个氨基酸(aa)、约110aa至约115aa、约115aa至约120aa、约120aa至约130aa、约130aa至约140aa、约140aa至约150aa或约150aa至约160aa的连续段具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:或其中itam基序为加粗且加下划线的。同样,合适的胞内活化结构域多肽可包含全长cd3δ氨基酸序列的含itam基序的部分。因此,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:其中itam基序为加粗且加下划线的。在一些情况下,胞内活化结构域来源于t细胞表面糖蛋白cd3ε链(亦称为cd3e、t细胞表面抗原t3/leu-4ε链、t细胞表面糖蛋白cd3ε链、ai504783、cd3、cd3ε、t3e等)。因此,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链或与以下氨基酸序列的约100个氨基酸至约110个氨基酸(aa)、约110aa至约115aa、约115aa至约120aa、约120aa至约130aa、约130aa至约140aa、约140aa至约150aa或约150aa至约160aa的连续段具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:其中itam基序为加粗且加下划线的。同样,合适的胞内活化结构域多肽可包含全长cd3ε氨基酸序列的含itam基序的部分。因此,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:其中itam基序为加粗且加下划线的。在一些情况下,胞内活化结构域来源于t细胞表面糖蛋白cd3γ链(亦称为cd3g、t细胞受体t3γ链、cd3-γ、t3g、γ多肽(tit3复合物)等)。因此,合适的胞内活化结构域可包括与以下序列中的至少10个、15个、20个或全部氨基酸的一段链或与以下氨基酸序列的约100个氨基酸至约110个氨基酸(aa)、约110aa至约115aa、约115aa至约120aa、约120aa至约130aa、约130aa至约140aa、约140aa至约150aa或约150aa至约160aa的连续段具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:meqgkglavlilaiillqgtlaqsikgnhlvkvydyqedgsvlltcdaeaknitwfkdgkmigfltedkkkwnlgsnakdprgmyqckgsqnkskplqvyyrmcqncielnaatisgflfaeivsifvlavgvyfiagqdgvrqsrasdkqtllpndqlyqplkdreddqyshlqgnqlrrn(seqidno:22),其中itam基序为加粗且加下划线的。同样,合适的胞内活化结构域多肽可包含全长cd3γ氨基酸序列的含itam基序的部分。因此,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:其中itam基序为加粗且加下划线的。在一些情况下,胞内活化结构域来源于cd79a(也称为b细胞抗原受体复合物相关蛋白α链;cd79a抗原(免疫球蛋白相关α);mb-1膜糖蛋白;ig-α;膜结合的免疫球蛋白相关蛋白;表面igm相关蛋白等)。因此,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链或与以下任何氨基酸序列中的约100个氨基酸至约110个氨基酸(aa)、约110aa至约115aa、约115aa至约120aa、约120aa至约130aa、约130aa至约140aa、约140aa至约150aa或约150aa至约160aa的连续段具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:或其中itam基序为加粗且加下划线的。同样,合适的胞内活化结构域多肽可包含全长cd79a氨基酸序列的含itam基序的部分。因此,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:其中itam基序为加粗且加下划线的。在一些情况下,胞内活化结构域来源于dap12(也称为tyrobp;tyro蛋白酪氨酸激酶结合蛋白;karap;plosl;dnax活化蛋白12;kar相关蛋白;tyro蛋白酪氨酸激酶结合蛋白质;杀伤活化受体相关蛋白;杀伤活化受体相关蛋白等)。举例而言,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链或与以下任何氨基酸序列(4个同功异型物)中约100个氨基酸至约110个氨基酸(aa)、约110aa至约115aa、约115aa至约120aa、约120aa至约130aa、约130aa至约140aa、约140aa至约150aa或约150aa至约160aa的连续段具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:或其中itam基序为加粗且加下划线的。同样,合适的胞内活化结构域多肽可包含全长dap12氨基酸序列的含itam基序的部分。因此,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:其中itam基序为加粗且加下划线的。在一些情况下,胞内活化结构域来源于fcerlg(也称为fcrg;fcε受体iγ链;fc受体γ链;fc-εri-γ;fcrγ;fceriγ;高亲和性免疫球蛋白ε受体亚单位γ;免疫球蛋白e受体、高亲和性γ链等)。举例而言,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链或与,以下氨基酸序列的约50个氨基酸至约60个氨基酸(aa)、约60aa至约70aa、约70aa至约80aa或约80aa至约88aa的连续段具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:其中itam基序为加粗且加下划线的。同样,合适的胞内活化结构域多肽可包含全长fcer1g氨基酸序列的含itam基序的部分。因此,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:其中itam基序为加粗且加下划线的。适用于本公开的经改造信号传导多肽中的胞内活化结构域包括dap10/cd28型信号传导链。dap10信号传导链的一个实例是氨基酸序列:在一些实施方式中,合适的胞内活化结构域可包括与以下序列中的至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:cd28信号传导链的一个实例是氨基酸序列:在一些实施方式中,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:适用于本公开的经改造信号传导多肽中的胞内活化结构域包括zap70多肽,例如,合适的胞内活化结构域可包括与以下序列中至少10个、15个、20个或全部氨基酸的一段链或与以下氨基酸序列的约300个氨基酸至约400个氨基酸、约400个氨基酸至约500个氨基酸或约500个氨基酸至约619个氨基酸的连续段具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:mpdpaahlpffygsisraeaeehlklagmadglfllrqclrslggyvlslvhdvrfhhfpierqlngtyaiaggkahcgpaelcefysrdpdglpcnlrkpcnrpsglepqpgvfdclrdamvrdyvrqtwklegealeqaiisqapqvekliattahermpwyhssltreeaerklysgaqtdgkfllrprkeqgtyalsliygktvyhylisqdkagkycipegtkfdtlwqlveylklkadgliyclkeacpnssasnasgaaaptlpahpstlthpqrridtlnsdgytpeparitspdkprpmpmdtsvyespysdpeelkdkklflkrdnlliadielgcgnfgsvrqgvyrmrkkqidvaikvlkqgtekadteemmreaqimhqldnpyivrligvcqaealmlvmemagggplhkflvgkreeipvsnvaellhqvsmgmkyleeknfvhrdlaarnvllvnrhyakisdfglskalgaddsyytarsagkwplkwyapecinfrkfssrsdvwsygvtmwealsygqkpykkmkgpevmafieqgkrmecppecppelyalmsdcwiykwedrpdfltveqrmracyyslaskvegppgstqkaeaaca(seqidno:36)。淋巴增殖性元件尽管连续添加细胞,但外周t淋巴细胞数在整个成年期间维持非常稳定的水平,这是由于自胸腺的移出及回应于抗原遭遇的增殖,以及由于在抗原清除后去除抗原特异性效应的细胞的损耗(marrak,p.等人.2000.natimmunol1:107-111;freitas,aa等人.2000.annurevimmunol18:83-111)。外周t细胞区室的大小由影响增殖及存活的多个因素来调节。然而,在淋巴细胞减少的环境下,由于维持外周t细胞区室大小的“急性稳态增殖”机制,t淋巴细胞分裂独立于同源抗原。淋巴细胞减少症的条件已经在过继性细胞治疗的受试者或患者中通过体外增殖t细胞并将其引入淋巴细胞清除的受试者而建立,导致经转移t细胞的植入及抗肿瘤功能增强。然而,受试者的淋巴细胞清除是不期望的,因为它可引起严重的副作用,包括免疫功能紊乱及死亡。研究已展示,淋巴细胞清除移除作为稳态细胞因子的细胞池的内源性淋巴细胞,从而释放细胞因子以诱导过继转移细胞的存活和增殖。已知一些例如il-7和il-15的细胞因子介导t细胞的抗原非依赖性增殖,因此能够在非淋巴细胞减少的环境中诱发稳态增殖。然而,这些细胞因子及其受体具有在稳态下防止淋巴增殖性疾病的内在控制机制。本文中所提供的很多方面中包括淋巴增殖性元件或其编码核酸,其通常作为经改造信号传导多肽的部分。在本文的说明性实施方式中,通常通过用复制缺陷型重组逆转录病毒颗粒转导静息t细胞和/或静息nk细胞而将一种或多种淋巴增殖性元件引入至静息t细胞和/或静息nk细胞中,所述逆转录病毒颗粒的基因组编码作为部分经改造信号传导多肽的淋巴增殖性元件。淋巴增殖性元件可以是细胞因子或在另一些说明性实施方式中是细胞因子受体,或包括活化stat3途径、stat4途径或在又一说明性实施方式中jak/stat5途径的其信号传导结构域的片段。由此,淋巴增殖性元件在非限制性实例中可以是细胞因子受体或包括其信号传导结构域的活化片段(例如白介素受体),或包括活化stat5的其信号传导结构域的活化片段。因此,淋巴增殖性元件为诱导t细胞和/或nk细胞增殖的多肽。说明性淋巴增殖性元件通过活化stat5来诱导增殖。因此,在说明性实施方式中,这种淋巴增殖性元件的片段保留了通过活化stat5来诱导t细胞和/或nk细胞增殖的能力。在一些本文所呈现的方法及组合物中,淋巴增殖性元件用于促进遗传修饰的t细胞体内增殖或扩增而不必使受试者淋巴细胞清除。由此,可执行本文所提供的方法(包括通常通过转导受试者的静息t细胞和/或nk细胞而将淋巴增殖性元件插入至所述t细胞和/或nk细胞中)的非限制性说明性实施方式而不在执行该方法之前、期间和/或之后使受试者淋巴细胞清除,或不在执行此方法之前在从受试者采集血液之前、期间和/或之后使受试者淋巴细胞清除,或不在离体遗传修饰来自受试者的t细胞和/或nk细胞之前、期间和/或之后,和/或在将遗传修饰t细胞和/或nk细胞重新引入至受试者中之前、期间和/或之后使受试者淋巴细胞清除。促进t细胞体内增殖的因子包括细胞因子及其受体,其中受体通常包括配体结合结构域及信号传导结构域。在一些实施方式中,用于本文所公开的方法及组合物中的淋巴增殖性元件为细胞因子和/或细胞因子受体。细胞因子可以是白介素,且细胞因子受体可以是白介素受体。淋巴增殖性元件可以是细胞因子的功能性片段和/或细胞因子受体的功能性片段(例如其信号传导结构域),其中该片段能够通过例如活化stat5来促进t细胞增殖。在一些实例中,本文中的方法及组合物中的细胞因子淋巴增殖性元件包括以下一种或多种:白介素-7(il-7)或其受体(il-7r),或其信号传导结构域;白介素-12(il-12)或其受体(il-12r),或其信号传导结构域;白介素-23(il-23)或其由il-12rβ1及il-23r构成的受体,或其信号传导结构域;白介素-27(il-27)或其受体(il-27r),或其信号传导结构域;白介素-15(il-15)或其受体(il-15r),或其信号传导结构域;白介素-21(il-21)或其受体(il-21r),或其信号传导结构域;或转化生长因子β(tgfβ)或其受体(tgfβr)或其信号传导结构域;或tgfβ诱饵受体(tgf-β-显性负性受体ii(dnrii))。在一些实施方式中,淋巴增殖性元件为il-12r或tgfβ诱饵受体(tgf-β-显性负性受体ii(dnrii))。il-7结合至il-7受体(由il-7rα及共有γ链受体组成的异二聚体)。结合产生对胸腺内的t细胞发育及外周内存活至关重要的信号级联。已知il-7与il-7受体的结合使jak/stat5途径活化。il-12参与原生t细胞分化成th1细胞(hsiehcs等人.1993.science.260(5107):547-9),并且已知为t细胞刺激因子。il-12与il-12受体结合,il-12受体是il-12r-β1及il-12r-β2形成的异二聚体受体。il12可通过活化stat4而起作用,但经证明同样活化t细胞中的stat5(ahn,h.等人.1998.j.immun.161:5893-5900)。il-12家族由细胞因子il-12、il-23及il-27构成。il-23受体由il-12rβ1及il-23r构成。il-27为由两种不同基因(爱泼斯坦-巴尔二氏病毒诱导基因3(ebi3)及il-27p28)构成的异二聚体细胞因子。il-27与il-27受体相互作用。il-15是在结构及功能上类似于il-2的t细胞及nk细胞刺激因子。两种细胞因子均诱导t细胞增殖;并且认为它们的共有功能是因为两种受体均使用il-2/il-15rβ及共有γ链。il-15的信号传导途径以与il-15rα受体结合开始,随后呈递给细胞表面上带有il-15rβγc复合物的周围细胞。结合il-15β亚基活化詹纳斯激酶1(januskinase1;jak1)和γc亚基詹纳斯激酶3(jak3),其导致stat3和stat5的磷酸化及活化。il-21在经活化的人类cd4+t细胞和nkt细胞中表达,并且il-21表达在t辅助细胞的th2和th17亚群中上调。il-21受体(il-21r)在t细胞、b细胞及nk细胞表面上表达,其结构与其他i型细胞因子如il-2r或il-15的受体结构相似。il-21r需要与共有γ链(γc)二聚化以便结合il-21。当与il-21结合时,il-21受体通过jak/stat途径起作用,从而活化stat1、stat3及stat5。tgfβ诱饵受体(tgf-β-显性负性受体ii(dnrii))通过与tgfβ结合的天然受体竞争来阻断tgfβ信号传导。tgfβ-dnrii是rii的激酶失活截短形式,其含有胞外tgfβ结合结构域和rii的跨膜结构域。tgfβ-dnrii结合配体但并不磷酸化和活化ri,从而减少或消除smad磷酸化。在患有b细胞和t细胞急性淋巴细胞性白血病(b-all和t-all)的受试者中已鉴别出il-7rα的功能获得性突变(zenattipp等人.2011.natgenet43:932-939;snochat,c.等人.2011.jexpmed208:901-908;mcelroy,ca等人.2012.pnas109(7):2503-2508)。突变包括在il-7rαtmd的n端区中的插入和缺失,其中几乎所有的序列均含有额外的cys残基,以及s165至c165突变。半胱氨酸导致受体的组成型活化。t-all群组中的一些突变活化jak1。这些功能获得性il-7r突变体可在本文所提供的任何方面中用作淋巴增殖性元件之一。因此,在一些实施方式中,淋巴增殖性元件是突变的il-7受体。在另一些实施方式中,突变的il-7受体是组成型活化的,在不存在细胞因子配体的情况下激活jak-stat5途径。在另一些实施方式中,突变的il-7受体在包括组成型活化stat5途径的能力的包含半胱氨酸残基的237至254之间的位置包含1个至10个氨基酸插入。在一些实施方式中,突变的il-7受体为il-7rα-insppcl(由seqidno:82表示)。在一些实施方式中,淋巴增殖性元件是嵌合细胞因子受体,例如但不限于系接(tethered)至其受体的细胞因子,其通常组成型活化与对应经活化野生型细胞因子受体如stat3、stat4(以及在说明性实施方式中,stat5)相同的stat途径。在一些实施方式中,嵌合细胞因子受体是通过接头系接至或共价连接至其同源受体或其片段的白介素或其片段。在一些实施方式中,嵌合细胞因子受体是与il-7rα系接的il-7。在另一些实施方式中,嵌合细胞因子受体是与il-7rα结构域系接的il-7,例如il-7rα的胞外结构域和/或il-7rα的跨膜结构域。在一些实施方式中,淋巴增殖性元件是未系接至细胞因子的细胞因子受体,且实际上在一些实施方式中,本文所提供的淋巴增殖性元件是未系接至细胞因子的组成型活化的细胞因子受体。这些嵌合il-7受体在表达时通常组成型活化stat5。在一些实施方式中,淋巴增殖性元件不是细胞因子或细胞因子受体而是抑制性rna,例如通常通过利用使socs途径中的负调节因子降解或下调而增强stat5活化来刺激stat5途径的mirna。在一些实施方式中,mirna靶向编码影响增殖的蛋白质的mrna,所述蛋白质例如但不限于abcg1、socs1、tgfbr2、smad2、ccbl及pd1。在说明性实施方式中,如本文中所例示,此类抑制性rna(例如mirna)可位于包装细胞和/或复制缺陷型重组逆转录病毒颗粒基因组和/或逆转录病毒载体中的内含子中,且通常通过在t细胞和/或nk细胞中有活性的启动子驱动来表达。不受理论限制,认为转录单元中包含内含子导致转录物更高表达和/或稳定性。由此,将mirna置放于逆转录病毒基因组的内含子中的能力增加了本发明的教导,该教导克服了现有技术中试图在逆转录病毒(例如慢病毒基因组)的大小限制下达到最大活性的挑战。在一些实施方式中,1个、2个、3个、4个、5个、6个、7个、8个、9个或10个mirna(在说明性实施方式中,2个与5个之间的(例如4个)mirna(其中一种或多种各自结合编码abcg1、socs1、tgfbr2、smad2、ccbl及pd1中一种或多种的核酸))可包括于重组逆转录病毒基因组中且使用本文所提供的方法递送至靶标细胞,例如t细胞和/或nk细胞。实际上,如本文所提供,1个、2个、3个或4个mirna可在单个内含子(例如ef1α内含子)中递送。abcg1是对胸腺细胞及外周淋巴细胞增殖进行负调节的atp结合盒转运蛋白(armstrong等人.2010.jimmunol184(1):173-183)。socs1是抑制jak/stat途径(例如stat5)的细胞因子信号转导负调因子的socs(细胞因子信号传导抑制因子)家族的成员。socs1也称为jab(janus激酶结合蛋白质)、ssi-1(stat诱导的stat抑制因子-1)及tip3(tec相互作用蛋白)。tgfbr2是丝氨酸/苏氨酸蛋白质激酶家族的成员,其结合tgf-β,从而形成使蛋白质磷酸化的复合物,随后进入细胞核且调节与增殖相关的基因的转录。smad2介导转化生长因子(tgf)-β的信号且调节多个细胞过程,例如细胞增殖、凋亡及分化。ccbl是通过使zap-70去磷酸化且失活且经由tcr的内化来抑制tcr信号传导的e3泛素连接酶。pd1(cd279)是在t细胞和prob细胞上表达的细胞表面受体。pd-1结合两种配体,pd-l1及pd-l2。经由pd-1进行信号传导起防止细胞活化的作用。在一些本文所公开的方法及组合物中,淋巴增殖性元件的表达是通过化合物与控制元件(如本文其他地方所论述)的结合来诱导且甚至可依赖于该结合,该控制元件在非限制性实施方式中为核糖开关。在一些实施方式中,淋巴增殖性元件由在t细胞和/或nk细胞中有活性的启动子表达。对于本文所提供的方法及组合物,熟练技术人员将认识到,已知在t细胞和/或nk细胞中有活性且可用于表达第一经改造信号传导多肽或第二经改造信号传导多肽,或其任何组分的启动子。在说明性实施方式中,此启动子在包装细胞系(例如本文所公开的包装系)中是无活性的。在一些实施方式中,启动子为ef1α启动子或鼠干细胞病毒(mscv)启动子(jones等人,humangenetherapy(2009)20:630-40)。在说明性实施方式中,启动子为t细胞特异性cd3ζ启动子。在一些实施方式中,淋巴增殖性元件为微环境限制的。举例而言,淋巴增殖性元件可以是在异常条件相对于生理条件下差异性结合其对应细胞因子的突变受体。举例而言,可使用在肿瘤环境下比在正常生理环境下可更强烈地结合il7的il-7r。在一些实施方式中,淋巴增殖性元件与识别结构域或消除结构域融合。在本文中更详细地公开这类识别结构域或消除结构域。此融合提供了在逆转录病毒基因组中需要更少的多核苷酸的优势,尤其在使用经截短或其他突变的淋巴增殖性元件时。这在本文所提供的说明性实施方式中为重要的,这是由于其有助于允许编码功能性元件的更多核酸包括于逆转录病毒基因组中。在另一些实施方式中,淋巴增殖性元件与共刺激结构域和/或胞内活化结构域融合。如本文所公开的淋巴增殖性元件不是嵌合抗原受体(car)或其胞内活化结构域或共刺激结构域。然而,在一些实施方式中,淋巴增殖性元件可与抗原特异性靶向区(astr)融合且通过astr与其抗原的结合来活化。在又一些实施方式中,经改造信号传导多肽可包括astr、胞内活化结构域(例如cd3ζ信号传导结构域)、共刺激结构域及淋巴增殖性结构域。关于共刺激结构域、胞内活化结构域、astr及其他car结构域的其他细节公开于本文其他地方中。在本文的说明性实施方式中,通常通过用复制缺陷型重组逆转录病毒颗粒转导静息t细胞和/或静息nk细胞而将t细胞和/或nk细胞存活元件引入至静息t细胞和/或静息nk细胞中,该复制缺陷型重组逆转录病毒颗粒的基因组编码作为经改造信号传导多肽的部分的t细胞和/或nk细胞存活元件。在一些实施方式中,淋巴增殖性元件也是t细胞和/或nk细胞存活元件。如上文所论述,淋巴增殖性元件中的一些不仅促进增殖,而且同样促进细胞存活。在一些实施方式中,t细胞和/或nk存活基序不是淋巴增殖性元件。举例而言,t细胞和/或nk细胞存活基序可以是cd28t细胞存活基序或cd137细胞存活基序。可在包括astr(例如scfv)的经改造信号传导多肽上发现此类t细胞存活基序。在一说明性实施方式中,t细胞存活基序为经由cd8a跨膜结构域或cd28跨膜结构域连接至scfv的cd28t细胞存活基序或cd137基序。在某些实施方式中,该胞内信号传导结构域包含多肽序列,该多肽序列包含基于免疫受体酪氨酸的活化基序(itam)。在一特定实施方式中,该多肽序列为cd3ζ信号传导结构域。调节结构域调节结构域可改变经改造信号传导多肽中的胞内活化结构域的作用,包括增强或抑制活化结构域的下游作用或改变反应的性质。适用于本公开的经改造信号传导多肽中的调节结构域包括共刺激结构域。适合于包含入经改造信号传导多肽中的调节结构域的长度可以是约30个氨基酸至约70个氨基酸(aa),例如,调节结构域的长度可以是约30aa至约35aa、约35aa至约40aa、约40aa至约45aa、约45aa至约50aa、约50aa至约55aa、约55aa至约60aa、约60aa至约65aa或约65aa至约70aa。在另一些情况下,调节结构域的长度可以是约70aa至约100aa、约100aa至约200aa,或大于200aa。共刺激结构域通常增强和/或改变对活化结构域的反应的性质。适用于本公开的经改造信号传导多肽中的共刺激结构域通常为衍生自受体的多肽。在一些实施方式中,共刺激结构域同源二聚化。本发明共刺激结构域可以是跨膜蛋白质的胞内部分(即,共刺激结构域可衍生自跨膜蛋白质)。合适的共刺激多肽的非限制性实例包括但不限于4-lbb(cd137)、cd27、cd28、lck结合缺失(icδ)的cd28、icos、ox40、btla、cd27、cd30、gitr及hvem。举例而言,本发明的一个方面的共刺激结构域可与4-lbb(cd137)、cd27、cd28、lck结合缺失(icδ)的cd28、icos、ox40、btla、cd27、cd30、gitr或hvem的共刺激结构域具有至少80%、90%或95%序列同一性。举例而言,本发明的一个方面的共刺激结构域可与合适共刺激多肽的非限制性实例的共刺激结构域具有至少80%、90%或95%序列同一性,实例包括但不限于4-lbb(cd137)、cd27、cd28、lck结合缺失(icδ)的cd28、icos、ox40、btla、cd27、cd30、gitr及hvem。举例而言,本发明的一个方面的共刺激结构域可与4-lbb(cd137)、cd27、cd28、lck结合缺失(icδ)的cd28、icos、ox40、btla、cd27、cd30、gitr或hvem的共刺激结构域具有至少80%、90%或95%序列同一性。适合于包含入经改造信号传导多肽中的共刺激结构域的长度可以是约30个氨基酸至约70个氨基酸(aa),例如,共刺激结构域的长度可以是约30aa至约35aa、约35aa至约40aa、约40aa至约45aa、约45aa至约50aa、约50aa至约55aa、约55aa至约60aa、约60aa至约65aa或约65aa至约70aa。在另一些情况下,共刺激结构域的长度可以是约70aa至约100aa、约100aa至约200aa,或大于200aa。在一些情况下,共刺激结构域衍生自跨膜蛋白质cd137(也称为tnfrsf9;cd137;4-lbb;cdwl37;ila等)的胞内部分。举例而言,合适的共刺激结构域可包括与以下氨基酸序列中至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:krgrkkllyifkqpfmrpvqttqeedgcscrfpeeeeggcel(seqidno:1)。在一些这类实施方式中,共刺激结构域的长度为约30aa至约35aa、约35aa至约40aa、约40aa至约45aa、约45aa至约50aa、约50aa至约55aa、约55aa至约60aa、约60aa至约65aa或约65aa至约70aa。在一些情况下,共刺激结构域衍生自跨膜蛋白质cd28(也称为tp44)的胞内部分。举例而言,合适的共刺激结构域可包括与以下氨基酸序列中至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:rskrsrllhsdymnmtprrpgptrkhyqpyapprdfaayrs(seqidno:2)。在一些这类实施方式中,共刺激结构域的长度为约30aa至约35aa、约35aa至约40aa、约40aa至约45aa、约45aa至约50aa、约50aa至约55aa、约55aa至约60aa、约60aa至约65aa或约65aa至约70aa。在一些情况下,共刺激结构域衍生自lck结合缺失(icδ)的跨膜蛋白质cd28的胞内部分。举例而言,合适的共刺激结构域可包括与以下氨基酸序列中至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:rskrsrllhsdymnmtprrpgptrkhyqayaaardfaayrs(seqidno:3)。在一些这类实施方式中,共刺激结构域的长度为约30aa至约35aa、约35aa至约40aa、约40aa至约45aa、约45aa至约50aa、约50aa至约55aa、约55aa至约60aa、约60aa至约65aa或约65aa至约70aa。在一些情况下,共刺激结构域衍生自跨膜蛋白质icos(也称为ailim、cd278及cvidl)的胞内部分。举例而言,合适的共刺激结构域可包括与以下氨基酸序列中至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:tkkkysssvhdpngeymfmravntakksrltdvtl(seqidno:4)。在一些这类实施方式中,共刺激结构域的长度为约30aa至约35aa、约35aa至约40aa、约40aa至约45aa、约45aa至约50aa、约50aa至约55aa、约55aa至约60aa、约60aa至约65aa或约65aa至约70aa。在一些情况下,共刺激结构域衍生自跨膜蛋白质ox40(也称为tnfrsf4、rp5-902p8.3、act35、cd134、ox-40、txgpll)的胞内部分。举例而言,合适的共刺激结构域可包括与以下氨基酸序列中的至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:rrdqrlppdahkppgggsfrtpiqeeqadahstlaki(seqidno:5)。在一些这类实施方式中,共刺激结构域的长度为约30aa至约35aa、约35aa至约40aa、约40aa至约45aa、约45aa至约50aa、约50aa至约55aa、约55aa至约60aa、约60aa至约65aa或约65aa至约70aa。在一些情况下,共刺激结构域衍生自跨膜蛋白质cd27(也称为s152、t14、tnfrsf7及tp55)的细胞内部分。举例而言,合适的共刺激结构域可包括与以下氨基酸序列中的至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:hqrrkyrsnkgespvepaepcryscpreeegstipiqedyrkpepacsp(seqidno:6)。在一些这类实施方式中,共刺激结构域的长度为约30aa至约35aa、约35aa至约40aa、约40aa至约45aa、约45aa至约50aa,约。在一些情况下,共刺激结构域衍生自跨膜蛋白质btla(也称为btlal及cd272)的胞内部分。举例而言,合适的共刺激结构域可包括与以下氨基酸序列中的至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:cclrrhqgkqnelsdtagreinlvdahlkseqteastrqnsqvllsetgiydndpdlcfrmqegsevysnpcleenkpgivyaslnhsvigpnsrlarnvkeapteyasicvrs(seqidno:7)。在一些情况下,共刺激结构域衍生自跨膜蛋白质cd30(也称为tnfrsf8、dls166e及ki-1)的胞内部分。举例而言,合适的共刺激结构域可包括与以下氨基酸序列中的约100个氨基酸至约110个氨基酸(aa)、约110aa至约115aa、约115aa至约120aa、约120aa至约130aa、约130aa至约140aa、约140aa至约150aa、约150aa至约160aa或约160aa至约185aa的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:rracrkrirqklhlcypvqtsqpklelvdsrprrsstqlrsgasvtepvaeerglmsqplmetchsvgaayleslplqdaspaggpssprdlpeprvstehtnnkiekiyimkadtvivgtvkaelpegrglagpaepeleeeleadhtphypeqetepplgscsdvmlsveeegkedplptaasgk(seqidno:8)。在一些情况下,共刺激结构域衍生自跨膜蛋白质gitr(也称为tnfrsf18、rp5-902p8.2、aitr、cd357及gitr-d)的胞内部分。举例而言,合适的共刺激结构域可包括与以下氨基酸序列中的至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:hiwqlrsqcmwpretqlllevppstedarscqfpeeergersaeekgrlgdlwv(seqidno:9)。在一些这类实施方式中,共刺激结构域的长度为约30aa至约35aa、约35aa至约40aa、约40aa至约45aa、约45aa至约50aa、约50aa至约55aa、约55aa至约60aa、约60aa至约65aa或约65aa至约70aa。在一些情况下,共刺激结构域衍生自跨膜蛋白质hvem(也称为tnfrsf14、rp3-395m20.6、atar、cd270、hvea、hvem、lightr及tr2)的胞内部分。举例而言,合适的共刺激结构域可包括与以下氨基酸序列中的至少10个、15个、20个或全部氨基酸的一段链具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:cvkrrkprgdvvkvivsvqrkrqeaegeatviealqappdvttvaveetipsftgrspnh(seqidno:10)。在一些这类实施方式中,第一及第二多肽的共刺激结构域的长度为约30aa至约35aa、约35aa至约40aa、约40aa至约45aa、约45aa至约50aa、约50aa至约55aa、约55aa至约60aa、约60aa至约65aa或约65aa至约70aa。接头在一些情况下,经改造信号传导多肽包括在任何两个相邻结构域间的接头。举例而言,接头可在跨膜结构域与第一共刺激结构域之间。作为另一实例,astr可以是抗体,且接头可在重链与轻链之间。作为另一实例,接头可在astr与跨膜结构域及共刺激结构域之间。作为另一实例,接头可在第二多肽的胞内活化结构域与共刺激结构域之间。作为另一实例,接头可在astr与胞内信号传导结构域之间。接头肽可具有多种氨基酸序列中的任何一种。蛋白质可通过通常具有柔性的间隔肽连接,但不排除其他化学键合。接头可以是长度在约1个与约100个氨基酸之间,或长度在约1个与约25个氨基酸之间的肽。这种接头可通过使用合成的编码接头的寡核苷酸偶合这些蛋白质来产生。可使用具有一定程度柔性的肽接头。连接肽实际上可以具有任何氨基酸序列,前提是合适的接头将具有产生通常柔性的肽的序列。小氨基酸(例如甘氨酸及丙氨酸)的用途是用于创建柔性肽。对于本领域技术人员来说,创建这样的序列是常规的。合适的接头可以是容易选择的且可以具有任何合适的不同长度,例如1个氨基酸(例如gly)至20个氨基酸、2个氨基酸至15个氨基酸、3个氨基酸至12个氨基酸,包括4个氨基酸至10个氨基酸、5个氨基酸至9个氨基酸、6个氨基酸至8个氨基酸、或7个氨基酸至8个氨基酸,且可以是1个、2个、3个、4个、5个、6个或7个氨基酸。例示性柔性接头包括甘氨酸聚合物(g)n、甘氨酸-丝氨酸聚合物(包括例如(gs)n、gsggsn、gggsn及ggggsn,其中n是至少为1的整数),甘氨酸-丙氨酸聚合物、丙氨酸-丝氨酸聚合物,及本领域已知的其他柔性接头。甘氨酸及甘氨酸-丝氨酸聚合物是令人感兴趣的,这是由于这两种氨基酸均为相对非结构化的,且因此可作为组分之间的中性系接。甘氨酸聚合物尤其令人感兴趣,是由于甘氨酸甚至比丙氨酸具有显著更多的phi-psi空间,且比具有较长侧链的残基受到少得多的限制(参见scheraga,rev.computationalchem.11173-142(1992))。例示性柔性接头包括但不限于ggggsggggsggggs(seqidno:53)、ggggsggggsggggsggggsggggsggggs(seqidno:54)、ggggsgggsggggs(seqidno:55)、ggsg(seqidno:56)、ggsgg(seqidno:57)、gsgsg(seqidno:58)、gsggg(seqidno:59)、gggsg(seqidno:60)、gsssg(seqidno:61),以及类似者。一般技术人员将认识到,将肽缀合至上文所描述的任何元件的设计可包括为完全或部分柔性的接头,使得接头可包括柔性接头以及赋予较低柔性结构的一个或多个部分。嵌合抗原受体在一些本发明的方面中,经改造信号传导多肽是嵌合抗原受体(car)或编码car的多核苷酸,为简单起见,在本文称为“car”。在一些实施方式中,本公开的car包括:a)至少一个抗原特异性靶向区(astr);b)跨膜结构域;及c)胞内活化结构域。在说明性实施方式中,car的抗原特异性靶向区是抗体针对靶标抗原的scfv部分。本公开的car可存在于真核细胞(例如,哺乳动物细胞)的质膜中,其中合适的哺乳动物细胞包括但不限于细胞毒性细胞、t淋巴细胞、干细胞、干细胞的子代、祖细胞、祖细胞的子代及nk细胞、nk-t细胞及巨噬细胞。当存在于真核细胞的质膜中时,本公开的car在存在一个或多个靶标抗原(在某些条件下,结合astr)的情况下被活化。靶标抗原为特异性结合对的第二成员。特异性结合对的靶标抗原可以是可溶性(例如,未结合至细胞的)因子;存在于细胞例如靶标细胞的表面的因子;存在于固体表面上的因子;存在于脂质双层上的因子;以及类似者。当astr为抗体,且特异性结合对的第二成员为抗原时,抗原可以是可溶性(例如,未结合至细胞的)抗原;存在于细胞例如靶标细胞的表面的抗原;存在于固体表面上的抗原;存在于脂质双层上的抗原;以及类似者。在一些情况下,本公开的car在存在于真核细胞质膜中时且由一种或多种靶标抗原活化时,使细胞中的至少一种核酸的表达增加。举例而言,在一些情况下,本公开的car在存在于真核细胞的质膜中时且由一种或多种靶标抗原活化时,与在不存在一种或多种靶标抗原时的核酸的转录水平相比,使细胞中至少一种核酸的表达增加至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约40%、至少约50%、至少约75%、至少约2倍、至少约2.5倍、至少约5倍、至少约10倍,或大于10倍。作为一实例,本公开的car可包括含有基于免疫受体酪氨酸的活化基序(itam)的胞内信号传导多肽。在一些情况下,本公开的car在存在于真核细胞质膜中时且由一种或多种靶标抗原活化时,可使得细胞一种或多种细胞因子的产生增加。举例而言,本公开的car在存在于真核细胞质膜中时且由一种或多种靶标抗原活化时,与在不存在一种或多种靶标抗原时细胞所产生的细胞因子的量相比,可使得细胞产生的细胞因子增加至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约40%、至少约50%、至少约75%、至少约2倍、至少约2.5倍、至少约5倍、至少约10倍,或大于10倍。其产量可增加的细胞因子包括但不限于干扰素γ(ifn-γ)、肿瘤坏死因子-α(tnf-α)、il-2、il-15、il-12、il-4、il-5、il-10;趋化因子;生长因子;以及类似者。在一些情况下,本公开的car在存在于真核细胞质膜中时且由一种或多种靶标抗原活化时,可导致细胞中的核酸转录的增加及细胞的细胞因子产生增加二者。在一些情况下,本公开的car在存在于真核细胞质膜中时且由一种或多种靶标抗原活化时,产生细胞对靶标细胞的细胞毒性活性,该靶标细胞在其细胞表面上表达抗原,该抗原与car的第一多肽的抗原结合结构域结合。举例而言,当真核细胞为细胞毒性细胞(例如,nk细胞或细胞毒性t淋巴细胞)时,本公开的car在存在于细胞质膜中时且由一种或多种靶标抗原活化时,使细胞对靶标细胞的细胞毒性活性增加,该靶标细胞在其细胞表面上表达一种或多种靶标抗原。举例而言,当真核细胞为nk细胞或t淋巴细胞时,本公开的car在存在于细胞质膜中时且由一种或多种靶标抗原活化时,与不存在一种或多种靶标抗原时的细胞的细胞毒性活性相比,使得细胞的细胞毒性活性增加至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约40%、至少约50%、至少约75%、至少约2倍、至少约2.5倍、至少约5倍、至少约10倍,或大于10倍。在一些情况下,本公开的car在存在于真核细胞质膜中时且由一种或多种靶标抗原活化时,可产生其他与car活化相关的事件,例如增殖及扩增(由于细胞分裂增加或抗凋亡反应增强)。在一些情况下,本公开的car在存在于真核细胞质膜中时且由一种或多种靶标抗原活化时,可产生其他与car活化相关的事件,例如胞内信号传导调节、细胞分化或细胞死亡。本公开的car可存在于真核细胞膜中,其中car的第一及第二多肽并未彼此共价连接。本公开的car可存在于真核细胞膜中作为单个异二聚体,该异二聚体未共价连接至膜中的任何其他多肽。或者,本公开的第一car可存在于真核细胞膜中作为异二聚体,该异二聚体共价连接或非共价连接至本公开的第二car。在一些情况下,第一及第二car经由二硫键共价连接,该二硫键形成在存在于柄中的半胱氨酸之间,该柄存在于第一car的第一多肽及第二car的第一多肽两者中。在一些情况下,本公开的car可存在于真核细胞膜中,其中car的第一多肽包括抗体片段,且car的第二多肽包括衍生自细胞因子受体的信号转导结构域,使得在二聚化后,car可表示异二聚化的信号体car,例如由至少两种独立多肽构成的信号体。“信号体(signalobody)”在本领域是已知的,是由衍生自细胞因子受体的信号转导结构域及抗体片段构成的单个嵌合大分子。在某些情况下,本公开的异二聚化信号体car在存在于真核细胞的细胞膜中,由二聚体二聚化,且由抗原(例如寡聚抗原)活化时,可诱导异二聚化信号体car的寡聚化。异二聚化信号体car的这种配体诱导的寡聚化可活化(例如增强)或保持(例如维持)信号转导,例如,异二聚化信号体car的配体诱导的寡聚化可传输诱发细胞反应的信号。在一些情况下,可组合使用多个异二聚化信号体car以诱发所需的细胞反应。在一些情况下,本公开的car为微环境限制的。这种特性通常是car的astr结构域的微环境限制性质的结果。因此,本公开的car可在微环境条件下比在正常生理环境条件下可具有针对一种或多种靶标抗原的较低结合亲和力,或在说明性实施方式中,更高结合亲和力。序列重组在某些情况下,在本文中可称为重组多肽的经改造信号传导多肽的序列可通过使用位点特异性重组技术在细胞中被重排或缺失。在某些实施方式中,可通过位点特异性重组改变针对特定经改造信号传导多肽的细胞活化相关反应,例如,诱发第一活化相关反应的经改造信号传导多肽的第一胞内活化结构域可被改换为诱发第二活化相关反应的第二胞内活化结构域。本领域技术人员将清楚,可在细胞中使用位点特异性重组以将经改造信号传导多肽的任何结构域或序列与本文所公开的任何其他结构域或序列交换。本领域技术人员也将清楚,可在细胞中使用位点特异性重组以缺失经改造信号传导多肽的任何结构域或序列。序列和结构域的这种交换及去除是本领域已知的,参见例如tone等人.(2013)biotechnologyandbioengineering,3219-3226中所描述的信号体中的结构域切换,其公开内容以引用方式并入本文。在体内执行位点特异性重组的机制及要求在本领域也是公知的,参见例如grindley等人.(2006)annualreviewofbiochemistry,567-605及tropp(2012)molecularbiology(jones&bartlettpublishers,sudbury,ma),其公开内容以引用方式并入本文。在一些实施方式中,经改造信号传导多肽通过将上文所论述的所有不同结构域融合在一起以形成融合蛋白质来产生。经改造信号传导多肽通常通过包含多核苷酸序列的转录单元来产生,该多核苷酸序列编码如本文所论述的经改造信号传导多肽的不同结构域。在一些实施方式中,本发明的起识别和结合靶标细胞上的抗原作用的astr是微环境限制的。可通过产生蛋白质文库且在该文库筛选对靶标抗原具有所需结合亲和力的蛋白质来发现在本发明中作为astr、适宜以全部或至少其针对靶标抗原的结合结构域的部分使用的野生型或天然蛋白质。野生型蛋白质可通过筛选cdna文库来发现。cdna文库为插入至宿主细胞集合中的克隆cdna(互补dna)片段的组合,其一起构成生物体的一部分转录组。cdna是由经完全转录的mrna产生的且因此含有生物体所表达蛋白质的编码序列。cdna文库中的信息是通过筛选文库中对靶标抗原具有所需结合亲和力的蛋白质来发现具有所需特性的蛋白质的强大的且有用的工具。组合在一些实施方式中,通过复制缺陷型重组逆转录病毒颗粒提供的多核苷酸具有编码一种或多种经改造信号传导多肽的某些组合的一个或多个转录单元。在本文所提供的一些方法及组合物中,在通过复制缺陷型重组逆转录病毒颗粒转导t细胞后,遗传修饰的t细胞包括一种或多种经改造信号传导多肽的组合。应理解,提及第一多肽、第二多肽、第三多肽等是为了方便,且“第一多肽”上的元件及“第二多肽”上的元件是指元件位于不同多肽上,这些多肽,特别是在该特定多肽的其他元件或步骤中,被称为第一或第二仅供参照及方便。在一些实施方式中,第一经改造信号传导多肽包括能够结合抗原的胞外抗原结合结构域,及胞内信号传导结构域。在另一些实施方式中,第一经改造信号传导多肽也包括t细胞存活基序和/或跨膜结构域。在一些实施方式中,第一经改造信号传导多肽不包括共刺激结构域,而在另一些实施方式中,第一经改造信号传导多肽的确包括共刺激结构域。在一些实施方式中,第二经改造信号传导多肽包括淋巴增殖性基因产物及任选的胞外抗原结合结构域。在一些实施方式中,第二经改造信号传导多肽也包括以下中的一种或多种:t细胞存活基序、胞内信号传导结构域,及一个或多个共刺激结构域。在另一些实施方式中,当使用两种经改造信号传导多肽时,至少一种是car。在一个实施方式中,一种或多种经改造信号传导多肽在相同转录物中在t细胞特异性启动子或一般启动子下表达,其中在该转录物中,编码经改造信号传导多肽的核酸由编码一种或多种内部核糖体进入位点(ire)或一种或多种蛋白酶切割肽的核酸分隔开。在某些实施方式中,多核苷酸编码两种经改造信号传导多肽,其中第一经改造信号传导多肽包括能够结合第一抗原的第一胞外抗原结合结构域,及第一胞内信号传导结构域而非共刺激结构域,且第二多肽包括能够结合vegf的第二胞外抗原结合结构域,及第二胞内信号传导结构域,例如共刺激分子的信号传导结构域。在某一实施方式中,第一抗原为psca、psma或bcma。在某一实施方式中,第一胞外抗原结合结构域包含抗体或其片段(例如scfv),例如对psca、psma或bcma具有特异性的抗体或其片段。在某一实施方式中,结合vegf的第二胞外抗原结合结构域为vegf的受体,即vegfr。在某些实施方式中,vegfr为vegfr1、vegfr2或vegfr3。在某一实施方式中,vegfr为vegfr2。在某些实施方式中,多核苷酸编码两种经改造信号传导多肽,其中第一经改造信号传导多肽包括胞外肿瘤抗原结合结构域及cd3ζ信号传导结构域,且第二经改造信号传导多肽包括抗原结合结构域(其中该抗原为血管生成(angiogenic)或血管原性(vasculogenic)因子),及一个或多个共刺激分子信号传导结构域。血管生成因子可以是例如vegf。一个或多个共刺激分子信号传导基序可包含例如来自cd27、cd28、ox40、icos及4-1bb各自的共刺激信号传导结构域。在某些实施方式中,多核苷酸编码两种经改造信号传导多肽,其中第一经改造信号传导多肽包括胞外肿瘤抗原结合结构域及cd3ζ信号传导结构域,第二多肽包含能够结合vegf的抗原结合结构域,及来自cd27、cd28、ox40、icos及4-1bb各自的共刺激信号传导结构域。在另一实施方式中,第一信号传导多肽或第二信号传导多肽也具有t细胞存活基序。在一些实施方式中,t细胞存活基序是或来源于il-7受体(il-7r)的胞内信号传导结构域、il-12受体的胞内信号传导结构域、il-15受体的胞内信号传导结构域、il-21受体的胞内信号传导结构域,或转化生长因子β(tgfβ)受体或tgfβ诱饵受体(tgf-β--显性-负性受体ii(dnrii))的胞内信号传导结构域。在某些实施方式中,多核苷酸编码两种经改造信号传导多肽,其中第一经改造信号传导多肽包括胞外肿瘤抗原结合结构域及cd3ζ信号传导结构域,且第二经改造信号传导多肽包含能够结合vegf的抗原结合结构域、il-7受体胞内t细胞存活基序、及来自cd27、cd28、ox40、icos及4-1bb各自的共刺激信号传导结构域。在一些实施方式中,由多核苷酸编码超过两种信号传导多肽。在某些实施方式中,仅经改造信号传导多肽中的一个包括结合至肿瘤相关抗原或肿瘤特异性抗原的抗原结合结构域;这种经改造信号传导多肽中的剩余部分的每一种包含结合至非肿瘤相关抗原或非肿瘤特异性抗原的抗原的抗原结合结构域。在另一些实施方式中,经改造信号传导多肽中的两种或更多包括结合至一种或多种肿瘤相关抗原或肿瘤特异性抗原的抗原结合结构域,其中这种经改造信号传导多肽中的至少一种包含不结合至肿瘤相关抗原或肿瘤特异性抗原的抗原结合结构域。在一些实施方式中,肿瘤相关抗原或肿瘤特异性抗原为her2、前列腺干细胞抗原(psca)、psma(前列腺特异性膜抗原)、b细胞成熟抗原(bcma)、α-胎蛋白(afp)、癌胚抗原(cea)、癌症抗原-125(ca-125)、ca19-9、钙视网膜蛋白、muc-1、上皮膜蛋白(ema)、上皮肿瘤抗原(eta)、酪氨酸酶、黑色素瘤相关抗原(mage)、cd34、cd45、cd99、cd117、嗜铬粒蛋白、细胞角蛋白、结蛋白、胶质纤维酸性蛋白(gfap)、大囊性疾病流体蛋白(gcdfp-15)、hmb-45抗原、蛋白黑色素a(t淋巴细胞识别的黑色素瘤抗原;mart-1)、肌凝蛋白-d1(myo-d1)、肌肉特异性肌动蛋白(msa)、神经纤毛、神经元特异性烯醇酶(nse)、胎盘碱性磷酸酶、突触泡蛋白、甲状腺球蛋白、甲状腺转录因子-1、丙酮酸激酶同工酶m2型的二聚体形式(肿瘤m2-pk)、cd19、cd22、cd27、cd30、cd70、gd2(神经节苷脂g2)、epha2、cspg4、cd138、fap(纤维母细胞活化蛋白)、cd171、κ整合素、λ整合素、5t4整合素、αvβ6整合素、整合素αvβ3(cd61)、泌乳激素、k-ras(v-ki-ras2kirsten大鼠肉瘤病毒致癌基因)、ral-b、b7-h3、b7-h6、caix、cd20、cd33、cd44、cd44v6、cd44v7/8、cd123、egfr、egp2、egp40、epcam、胚胎achr、frα、gd3、hla-a1+mage1、hla-a1+ny-eso-1、il-11rα、il-13rα2、路易斯-y(lewis-y)、muc16、ncam、nkg2d配体、ny-eso-1、prame、ror1、存活素、tag72、tems、vegfr2、egfrviii(表皮生长因子变体iii)、精子蛋白17(sp17)、间皮素、pap(前列腺酸性磷酸酶)、前列腺卵白(prostein)、tarp(t细胞受体γ替代阅读框蛋白)、trp-p8、steap1(前列腺1的六跨膜上皮抗原)、异常ras蛋白或异常p53蛋白。在一些实施方式中,第一经改造信号传导多肽包括结合第一抗原的第一胞外抗原结合结构域,及第一胞内信号传导结构域;且第二经改造信号传导多肽包括结合第二抗原或结合第二抗原的受体的第二胞外抗原结合结构域,及第二胞内信号传导结构域,其中该第二经改造信号传导多肽不包含共刺激结构域。在某一实施方式中,第一抗原结合结构域及第二抗原结合结构域独立地为受体的抗原结合部分或抗体的抗原结合部分。在某一实施方式中,第一抗原结合结构域或第二抗原结合结构域中的一种或两种是scfv抗体片段。在某些实施方式中,第一经改造信号传导多肽和/或第二经改造信号传导多肽额外包含跨膜结构域。在某一实施方式中,第一经改造信号传导多肽或第二经改造信号传导多肽包含t细胞存活基序,例如本文所描述的任何t细胞存活基序。在另一实施方式中,第一经改造信号传导多肽包括结合her2的第一胞外抗原结合结构域,且第二经改造信号传导多肽包括结合muc-1的第二胞外抗原结合结构域。在另一实施方式中,第二经改造信号传导多肽的第二胞外抗原结合结构域结合白介素。在另一实施方式中,第二经改造信号传导多肽的第二胞外抗原结合结构域结合损伤相关的分子模式分子(damp;也称为警报素(alarmin))。在另一些实施方式中,damp为热休克蛋白、染色质相关蛋白高迁移率组盒1(hmgb1)、s100a8(也称为mrp8或钙粒蛋白a)、s100a9(也称为mrp14或钙粒蛋白b)、血清淀粉样蛋白a(saa)、脱氧核糖核酸、三磷酸腺苷、尿酸或硫酸肝素。在某些实施方式中,该第二抗原为抗体上的抗原,该抗体结合由肿瘤细胞呈递的抗原。在一些实施方式中,经由第二经改造信号传导多肽的信号转导活化为非抗原性的,但与低氧有关。在某些实施方式中,低氧是通过低氧诱导因子-1α(hif-1α)、hif-1β、hif-2α、hif-2β、hif-3α或hif-3β活化诱导的。在一些实施方式中,一种或多种经改造信号传导多肽的表达是由在本文中更详细地公开的控制元件调节的。额外序列经改造信号传导多肽(例如car)可进一步包括一种或多种额外多肽结构域,其中这类结构域包括但不限于信号序列、表位标签、亲和结构域,及可例如通过抗体分析法或由于其为产生可检测信号的多肽而检测(可检测标记)其存在或活性的多肽。对于任何本文所提供方面或实施方式中额外结构域的非限制性实例包括与如下文所描述的任何以下序列具有至少50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%或100%序列同一性的结构域:信号序列、表位标签、亲和结构域或产生可检测信号的多肽。适用于本发明car(例如本发明car的第一多肽)中的信号序列包括任何真核信号序列,其包括天然存在的信号序列、合成的(例如,人造的)信号序列等。在一些实施方式中,举例而言,信号序列可以是cd8信号序列malpvtalllplalllhaarp(seqidno:74)。合适的表位标签包括但不限于凝血素(ha;例如ypydvpdya;seqidno:37)、flag(例如dykddddk;seqidno:38)、c-myc(例如eqkliseedl;seqidno:39)以及类似者。亲和结构域包括可与结合搭配物(例如在固体载体上固定的结合搭配物)相互作用的适用于识别或纯化的肽序列。编码多个连续的单一氨基酸(例如组氨酸)的dna序列当与所表达蛋白质融合时,可用于通过高亲和力结合至树脂柱(例如镍琼脂糖)一步纯化重组蛋白质。例示性亲和结构域包括his5(hhhhh;seqidno:40)、hisx6(hhhhhh;seqidno:41)、c-myc(eqkliseedl;seqidno:39)、flag(dykddddk;seqidno:38)、strep标签(wshpqfek;seqidno:42)、凝血素,例如ha标签(ypydvpdya;seqidno:37),gst、硫氧还蛋白、纤维素结合结构域、ryirs(seqidno:43)、phe-his-his-thr(seqidno:44)、甲壳素结合结构域、s-肽、t7肽、sh2结构域、c-末端rna标签、weaaareaccreccara(seqidno:45)、金属结合结构域,例如锌结合结构域或钙结合结构域,例如来自钙结合蛋白的,如钙调蛋白、肌钙蛋白c、钙调神经磷酸酶b、肌球蛋白轻链、恢复蛋白、s-调节素、视锥蛋白、vilip、神经钙蛋白、海马钙结合蛋白、频蛋白(frequenin)、钙牵蛋白(caltractin)、钙蛋白酶大亚基、s100蛋白,小白蛋白、钙结合蛋白d9k、钙结合蛋白d28k及钙视蛋白,内含肽、生物素,链霉亲和素,myod,id,亮氨酸拉链序列以及麦芽糖结蛋白。合适的可检测信号产生蛋白质包括例如荧光蛋白、催化产生可检测信号作为产物的反应的酶,以及类似者。合适的荧光蛋白质包括但不限于绿色荧光蛋白(gfp)或其变体、gfp的蓝色荧光变体(bfp)、gfp的青色荧光变体(cfp)、gfp的黄色荧光变体(yfp)、增强型gfp(egfp)、增强型cfp(ecfp)、增强型yfp(eyfp)、gfps65t、翡翠绿荧光蛋白(emerald)、黄玉荧光蛋白(topaz(tyfp))、维纳斯荧光蛋白(venus)、黄晶荧光蛋白(citrine)、变体黄晶荧光蛋白(mcitrine)、gfpuv、去稳定化的egfp(degfp)、去稳定化的ecfp(decfp)、去稳定化的eyfp(deyfp)、mcfpm、天蓝荧光蛋白(cerulean)、t-蓝宝石荧光蛋白(t-sapphire)、cypet,ypet、mko、hcred、t-hcred、dsred、dsred2、dsred-单体、j-red、二聚体2、t-二聚体2(12)、mrfp1、珊瑚荧光蛋白(pocilloporin)、renillagfp、monstergfp、pagfp、kaede蛋白及点燃蛋白(kindlingprotein)、藻胆蛋白及藻胆蛋白缀合物,包括b-藻红蛋白、r-藻红蛋白及别藻蓝蛋白。荧光蛋白质的另一些实例包括mhoneydew、mbanana、morange、dtomato、tdtomato、mtangerine、mstrawberry、mcherry、mgrapel、mraspberry、mgrape2、mplum(shaner等人.(2005)nat.methods2:905-909),以及类似者。适宜使用来自珊瑚虫(anthozoan)物种的多种荧光及有色蛋白质中的任意种,如例如matz等人.(1999)naturebiotechnol.17:969-973中所描述。合适的酶包括但不限于辣根过氧化物酶(hrp)、碱性磷酸酶(ap)、β-半乳糖苷酶(gal)、葡萄糖-6-磷酸脱氢酶、β-n-乙酰氨基葡糖苷酶、β-葡糖苷酸酶、转化酶、黄嘌呤氧化酶、萤火虫萤光素酶、葡萄糖氧化酶(go),以及类似者。识别和/或消除结构域本文所提供的任何复制缺陷型重组逆转录病毒颗粒可包括编码识别或消除结构域的核酸,该识别或消除结构域作为编码本文所提供的任何经改造信号传导多肽的核酸的一部分或独立于其存在。因此,本文所提供的任何经改造信号传导多肽可包括识别或消除结构域。举例而言,本文所公开的任何car可包括识别或消除结构域。此外,识别或消除结构域可与本文所公开的任何淋巴增殖性元件一起表达,或甚至与其融合。识别或消除结构域在t细胞和/或nk细胞上表达,但不在复制缺陷型重组逆转录病毒颗粒上表达。在一些实施方式中,识别或消除结构域可来源于单纯疱疹病毒衍生的酶胸苷激酶(hsv-tk)或诱导型半胱天冬酶-9。在一些实施方式中,识别或消除结构域可包括经修饰的内源细胞表面分子,例如如美国专利8,802,374中所公开的。经修饰的内源细胞表面分子可以是任何细胞表面相关的受体、配体、糖蛋白、细胞黏着分子、抗原、整合素或经修饰的分化簇(cd)。在一些实施方式中,经修饰的内源细胞表面分子是截短的酪氨酸激酶受体。在一个方面中,截短的酪氨酸激酶受体是表皮生长因子受体(egfr)家族中的成员,例如erbb1、erbb2、erbb3、erbb4。在一些实施方式中,识别结构域可以是由抗体识别的多肽,该抗体识别egfr成员的胞外结构域。在一些实施方式中,识别结构域可以是egfr家族成员的至少20个连续氨基酸,或例如egfr家族成员的20个至50个连续氨基酸。举例而言,seqidno:78为由识别egfr成员的胞外结构域的抗体识别且在适当条件下结合的例示性多肽。这种胞外egfr表位有时在本文中称为etag。在说明性实施方式中,这种表位由可市购的抗egfr单克隆抗体识别。表皮生长因子受体(也称为egfr、erbb1和her1)是表皮生长因子家族成员胞外配体的细胞表面受体。已将egfr活性的改变与某些癌症相联系。在一些实施方式中,编码包括人类表皮生长因子受体(egfr)的egfr多肽的基因通过以下构建:移除编码包括膜远端egf结合结构域及细胞质信号传导尾的多肽的核酸序列,但保留由抗egfr抗体识别的胞外膜近端表位。优选地,抗体为已知可市购的抗egfr单克隆抗体,例如西妥昔单抗(cetuximab)、马妥珠单抗(matuzumab)、尼西珠单抗(necitumumab)或帕尼单抗(panitumumab)。其他人已证明,生物素标记的西妥昔单抗结合抗生物素微珠应用于免疫磁选择成功地从低至群体的2%到大于90%的纯度富集用含有egfrt的构建体慢病毒转导的t细胞,而对细胞制备物无可观测的毒性。此外,其他人已证明,此惰性egfr分子的组成型表达不影响t细胞表型或如由协调表达的嵌合抗原受体(car)cd19r引导的效应功能。此外,其他人已证明,经由流式细胞术分析,egfr成功地用作小鼠中的t细胞移植的体内追踪标记物。此外,已证明egfr具有通过介导的抗体依赖型细胞毒性(adcc)途径的自杀基因潜力。本公开的发明者已使用慢病毒载体在pbmc中成功地表达etag,且已发现暴露于西妥昔单抗的pbmc在体外表达etag,提供一种pbmc的有效消除机制。因此,egfr可用作具有免疫治疗潜力的转导t细胞的非免疫原性选择工具、追踪标记物及自杀基因。egfr核酸也可通过本领域中熟知的方法来检测。在一些本文所提供的实施方式中,egfr作为也包括car的单个多肽的一部分或作为包含淋巴增殖性元件的单个多肽的一部分表达。在一些实施方式中,可通过切割信号和/或核糖体跳跃序列将编码egfr识别结构域的氨基酸序列与编码嵌合抗原受体的氨基酸序列分隔开。核糖体跳跃和/或切割信号可以是本领域已知的任何核糖体跳跃和/或切割信号。不受理论限制,核糖体跳跃序列可以是例如具有氨基酸序列gsgegrgslltcgdveenpgp(seqidno:77)的2a-1。不受理论限制,切割信号及核糖体跳跃序列的其他实例包括fmdv2a(f2a)、马a型鼻病毒2a(简称e2a)、猪捷申病毒-12a(p2a)及扁刺蛾属(thoseaasigna)病毒2a(t2a)。在一些实施方式中,编码识别结构域的多核苷酸序列可与car或淋巴增殖性元件在相同的转录物上,但通过内部核糖体进入位点与编码car或淋巴增殖性元件的多核苷酸序列隔开。在如本文中例示性列举的另一些实施方式中,识别结构域可表达为与淋巴增殖性元件融合的融合多肽的一部分。这种构建体与单独的多肽相比提供在rna基因组上占用更少的基因组空间的优势,尤其结合本文中提供的其他“空间节省”元件的情况下。在一个说明性实施方式中,etag表达为与il7rα突变体融合的融合多肽,如在本文中实验地论证的。假型化元件(pseudotypingelement)本文所提供的许多方法及组合物中包括假型化元件。具有异源包膜糖蛋白的复制缺陷型重组逆转录病毒颗粒的假型化通常改变病毒的向性(tropism)且有助于宿主细胞的转导。如本文中所使用的假型化元件可包括:“结合多肽”,其包括识别和结合靶标宿主细胞的一种或多种多肽(通常糖蛋白);及一种或多种“促融合多肽”,其介导逆转录病毒与靶标宿主细胞膜融合,从而使逆转录病毒基因组进入靶标宿主细胞。在本文所提供的一些实施方式中,假型化元件提供为多肽/蛋白质,或提供为编码多肽/蛋白质的核酸序列。在一些实施方式中,假型化元件为猫内源性病毒(rd114)包膜蛋白、致癌逆转录病毒双嗜性包膜蛋白、致癌逆转录病毒嗜亲性包膜蛋白、水泡性口炎病毒包膜蛋白(vsv-g),和/或副黏液病毒麻疹包膜蛋白h及f。在一些实施方式中,假型化元件包括来源于不同蛋白质的结合多肽及促融合多肽。举例而言,本文所公开的方法及组合物中的复制缺陷型重组逆转录病毒颗粒可通过麻疹病毒(mv)的融合(f)多肽及凝血素(h)多肽来假型化,作为非限制性实例,mv的临床野生型菌株,及包括埃德蒙斯顿菌株(edmonstonstrain,mv-edm)的疫苗菌株或其片段。不受理论限制,认为凝血素(h)及融合(f)多肽两者均在进入宿主细胞中发挥作用,其中该h蛋白将mv与靶标细胞上的受体cd46、slam及nectin-4结合,且f介导逆转录病毒及宿主细胞膜的融合。在一说明性实施方式中,尤其在靶标细胞为t细胞和/或nk细胞时,结合多肽为麻疹病毒h多肽,且促融合多肽为麻疹病毒f多肽。在一些研究中,用截短的f及h多肽假型化的慢病毒颗粒在效价及转导效率上显著增加(funke等人.2008.moleculartherapy.16(8):1427-1436)、(frecha等人.2008.blood.112(13):4843-4852)。在f胞质尾区被截短30个残基(也称为mv(ed)-fδ30(seqidno:105))时获得最高效价。针对h变体,在缺失18个或19个残基(mv(ed)-hδ18(seqidno:106)或(mv(ed)-hδ19))时出现最佳截短,尽管具有24个残基的截短的变体在缺失残基经丙氨酸置换及未置换的情况下(mv(ed)-hδ24(seqidno:235)及mv(ed)-hδ24+a)也产生最佳效价。在一些实施方式中,包括针对转导t细胞和/或nk细胞的那些实施方案中,本文中所公开的方法及组合物中的复制缺陷型重组逆转录病毒颗粒经麻疹病毒融合(f)多肽及凝血素(h)多肽的突变或变体版本(在说明性实例中,麻疹病毒f和h多肽的胞质结构域缺失变体)假型化。在一些实施方式中,经突变f和h多肽为“截短的h”或“截短的f”多肽,其胞质部分已经截短,即氨基酸残基(或编码蛋白质的对应核酸分子的编码核酸)已经缺失。“hδy”及“fδx”分别表示这种截短的h及f多肽,其中“y”是指已从胞质结构域的氨基端缺失的1个至34个残基,且“x”指已从胞质结构域的羧基端缺失的1个至35个残基。在另一实施方式中,“截短的f多肽”是fδ24或fδ30和/或“截短的h蛋白”选自:hδ14、hδ15、hδ16、hδ17、hδ18、hδ19、hδ20、hδ21+a、hδ24及hδ24+4a,更优选地hδ18或hδ24。在一说明性实施方式中,截短的f多肽是mv(ed)-fδ30,且截短的h多肽是mv(ed)-hδ18。在一些实施方式中,促融合多肽包括表达为一种多肽的多种元件。在一些实施方式中,结合多肽和促融合多肽从相同的转录物但从独立的核糖体结合位点翻译;在另一些实施方式中,结合多肽及促融合多肽由切割肽位点(其不受理论限制,在翻译后切割,如文献中常见的)或核糖体跳跃序列隔开。在一些实施方式中,结合多肽及促融合多肽从分离的核糖体结合位点的翻译导致比结合多肽更高量的促融合多肽。在一些实施方式中,促融合多肽与结合多肽的比率为至少2:1、至少3:1、至少4:1、至少5:1、至少6:1、至少7:1或至少8:1。在一些实施方式中,促融合多肽与结合多肽的比率在1.5:1、2:1或3:1的下限与3:1、4:1、5:1、6:1、7:1、8:1、9:1或10:1的上限之间的范围。活化元件本公开的许多方法及组合物方面中包括活化元件,或编码活化元件的核酸。与慢病毒(lv)转导进入静息t细胞中相关的限制归因于一系列的进入前及进入后障碍以及细胞限制因子(strebel等人2009.bmcmedicine7:48)。一个限制为包膜假型化lv颗粒不能识别潜在受体并介导与细胞膜的融合。然而,在某些条件下,用基于hiv-1的慢病毒载体转导静息t细胞在t细胞受体(tcr)cd3复合物及cd28共刺激(korin及zack.1998.journalofvirology.72:3161-8,maurice等人.2002.blood99:2342-50)以及通过暴露于细胞因子(cavalieri等人2003)的情况下最为可行。免疫系统的细胞(例如t淋巴细胞)通过受体或受体复合物识别特异性抗原并与其相互作用,在识别这种抗原或与其相互作用后,使得细胞活化并在体内扩增。此受体的实例是抗原特异性t淋巴细胞受体复合物(tcr/cd3)。t细胞受体(tcr)在t淋巴细胞表面上表达。组分cd3负责在tcr由配体占据之后的胞内信号传导。针对抗原cd3复合物(tcr/cd3)的t淋巴细胞受体识别通过主要组织相容性复合物(mhc)的蛋白质呈递给它的抗原性肽。mhc和肽的复合物在抗原呈递细胞及其他t淋巴细胞标靶的表面上表达。tcr/cd3复合物的刺激导致t淋巴细胞活化及随后的抗原特异性免疫反应。tcr/cd3复合物在免疫系统的效应功能及调节上发挥重要作用。t淋巴细胞也需要第二、共刺激信号以变得完全活化。在无此信号的情况下,t淋巴细胞对结合至tcr的抗原无反应或变得无变应性。此共刺激信号例如由cd28提供,cd28为在产生抗原细胞上与cd80及cd86相互作用的t淋巴细胞蛋白质。如本文所使用,cd80的功能性胞外片段保留其与cd28相互作用的能力。icos(诱导性共刺激剂)(另一种t淋巴细胞蛋白质)在结合至icos配体时提供共刺激信号。t细胞受体(tcr)cd3复合物的活化和cd28的共刺激可通过离体暴露于包被有抗cd3和抗cd28的固体表面(例如,珠粒)而发生。在一些本文所公开方法及组合物的实施方式中,静息t细胞通过离体暴露于包被有抗cd3和抗cd28的固体表面而被活化。在本文所提供的某些方法及组合物的说明性实施方式中,能够结合cd3和/或cd28的多肽在本文所公开的方法及组合物中的复制缺陷型重组逆转录病毒颗粒的表面上呈现为“活化元件”,这也是本发明的方面。结合cd3和/或cd28的多肽称为“活化元件”,这是由于其活化静息t细胞的能力。在一些实施方式中,活化元件是能够结合cd3的多肽。在一些实施方式中,能够结合cd3的多肽是抗cd3抗体,或其保留与cd3结合能力的片段。在说明性实施方式中,抗cd3抗体或其片段是单链抗cd3抗体,例如但不限于抗cd3scfv。在另一说明性实施方式中,能够结合cd3的多肽为抗cd3scfvfc。大量抗人类cd3单克隆抗体及其抗体片段为可得的,且可用于本发明,包括但不限于ucht1、okt-3、hit3a、trx4、x35-3、vit3、bma030(bw264/56)、clb-t3/3、cris7、yth12.5、f111409、clb-t3.4.2、tr-66、wt31、wt32、spv-t3b、11d8、xiii-141、xiii46、xiii-87、12f6、t3/rw2-8c8、t3/rw24b6、okt3d、m-t301、smc2及f101.01。在一些实施方式中,活化元件是能够结合cd28的多肽。在一些实施方式中,能够结合cd28的多肽为抗cd28抗体,或其保留与cd28结合能力的片段。在另一些实施方式中,能够结合至cd28的多肽为cd80、cd86,或其能够结合cd28且诱导cd28介导的akt活化的功能性片段,例如cd80的外部片段。在本文的一些方面中,cd80的外部片段是指保留与cd28结合能力的通常存在于cd80的正常细胞位置处的细胞外部的片段。在说明性实施方式中,抗cd28抗体或其片段是单链抗cd28抗体,例如但不限于抗cd28scfv。在另一说明性实施方式中,能够结合cd28的多肽是cd80,或cd80的片段,例如cd80的外部片段。抗cd28抗体是本领域已知的且可包括作为非限制性实例的单克隆抗体9.3(为igg2a抗体)(dr.jefferyledbetter,bristolmyerssquibbcorporation,seattle,wash.)、单克隆抗体kolt-2(igg1抗体)、15e8(igg1抗体)、248.23.2(igm抗体)及ex5.3d10(igg2a抗体)。在一说明性实施方式中,活化元件包括两种多肽,能够结合cd3的多肽及能够结合cd28的多肽。在某些实施方式中,能够结合cd3或cd28的多肽为抗体、单链单克隆抗体或抗体片段,例如单链抗体片段。因此,抗体片段可以是例如单链片段可变区(scfv)、抗体的抗体结合(fab)片段、单链抗原结合片段(scfab)、无半胱氨酸的单链抗原结合片段(scfabδc)、片段可变区(fv)、对抗原的相邻表位具有特异性的构建体(crab)或单域抗体(vh或vl)。在一些实施方式中,活化元件与异源信号序列和/或异源膜黏附序列融合,两者都有助于将活化元件引导至膜上。异源信号序列将活化元件靶向内质网,其中异源膜黏附序列共价连接至一种或多种脂肪酸(也称为翻译后脂质修饰)使得与异源膜黏附序列融合的活化元件锚定在质膜的脂质筏中。在一些实施方式中,翻译后脂质修饰可经由豆蔻酰化、棕榈酰化或gpi锚来发生。豆蔻酰化是对应于14-碳饱和脂肪酸(肉豆蔻酸)与真核或病毒蛋白质的n端甘氨酸的共价连接的翻译后蛋白质修饰。棕榈酰化是对应于c16酰基链与半胱氨酸,且在较少情况下与蛋白质的丝氨酸及苏氨酸残基的共价连接的翻译后蛋白质修饰。gpi锚是指在翻译后修饰期间糖基磷脂酰肌醇或gpi与蛋白质c端的连接。在一些实施方式中,异源膜连接序列为gpi锚连接序列。异源gpi锚连接序列可来源于任何已知的经gpi锚定的蛋白质(评论于fergusonmaj,kinoshitat,hartgw.glycosylphosphatidylinositolanchors.在varkia,cummingsrd,eskojd等人编.essentialsofglycobiology.第2版.coldspringharbor(ny):coldspringharborlaboratorypress;2009.第11章)。在一些实施方式中,异源gpi锚连接序列是来自cd14、cd16、cd48、cd55(daf)、cd59、cd80及cd87的gpi锚连接序列。在一些实施方式中,异源gpi锚连接序列来源于cd16。在说明性实施方式中,异源gpi锚连接序列来源于fc受体fcγriiib(cd16b)或衰变加速因子(daf),另外称为补体衰变加速因子或cd55。在一些实施方式中,一种或两种活化元件包括帮助将活化元件的表达引导至细胞膜的异源信号序列。可使用在包装细胞系中有活性的任何信号序列。在一些实施方式中,信号序列为daf信号序列。在说明性实施方式中,活化元件在其n端融合至daf信号序列,在其c端融合至gpi锚连接序列。在一说明性实施方式中,活化元件包括与来源于cd14的gpi锚连接序列融合的抗-cd3scfvfc,及与来源于cd16b的gpi锚连接序列融合的cd80;且两者表达于本文所提供的复制缺陷型重组逆转录病毒颗粒的表面上。在一些实施方式中,抗cd3scfvfc在其n端与daf信号序列及在其c端与来源于cd14的gpi锚连接序列融合,且cd80在其n端与daf信号序列及在其c端与来源于cd16b的gpi锚连接序列融合;且两者均表达于本文所提供的复制缺陷型重组逆转录病毒颗粒的表面上。在一些实施方式中,daf信号序列包括daf蛋白的1-30位氨基酸残基。膜结合细胞因子本文所提供的方法及组合物方面中的一些实施方式包括膜结合细胞因子,或编码膜结合细胞因子的多核苷酸。细胞因子通常但不总是分泌性蛋白质。天然分泌的细胞因子可经经改造为膜结合的融合蛋白质。膜结合细胞因子融合多肽包括在本文所公开的方法及组合物中,且也是本发明的方面。在一些实施方式中,复制缺陷型重组逆转录病毒颗粒具有在其表面上的能够结合t细胞和/或nk细胞且促进其增殖及/存活的膜结合细胞因子融合多肽。通常地,将膜结合多肽整合入复制缺陷型重组逆转录病毒颗粒的膜中,且在细胞通过复制缺陷型重组逆转录病毒颗粒转导时,逆转录病毒与宿主细胞膜的融合使得多肽结合至经转导细胞的膜。在一些实施方式中,细胞因子融合多肽包括il-7、il-15或其活化片段。膜结合细胞因子融合多肽通常为与异源信号序列和/或异源膜连接序列融合的细胞因子。在一些实施方式中,异源膜连接序列为gpi锚连接序列。异源gpi锚连接序列可来源于任何已知的经gpi锚定的蛋白质(评论于fergusonmaj,kinoshitat,hartgw.glycosylphosphatidylinositolanchors.varkia,cummingsrd,eskojd等人编.essentialsofglycobiology.第2版.coldspringharbor(ny):coldspringharborlaboratorypress;2009.第11章)。在一些实施方式中,异源gpi锚连接序列为来自cd14、cd16、cd48、cd55(daf)、cd59、cd80及cd87的gpi锚连接序列。在一些实施方式中,异源gpi锚连接序列来源于cd16。在一说明性实施方式中,异源gpi锚连接序列来源于fc受体fcγriiib(cd16b)。在一些实施方式中,gpi锚为daf的gpi锚。在说明性实施方式中,膜结合细胞因子为与daf融合的细胞因子的融合多肽。已知daf累积在整合入自包装细胞出芽的复制缺陷型重组逆转录病毒颗粒的膜中的脂质筏中。因此,不受理论限制,认为daf融合蛋白质优先靶向将变成重组逆转录病毒膜的一部分的包装细胞的膜部分。在非限制说明性实施方式中,细胞因子融合多肽是il-7,或其与daf融合的活性片段。在一特定非限制说明性实施方式中,融合细胞因子多肽依次包括:daf信号序列(daf的第1-31位残基)、无其信号序列的il-7,及daf的第36-525位残基。核糖开关控制元件核糖开关本文所提供的一些组合物及方法中包括一种或多种核糖开关或包括一种或多种核糖开关的多核苷酸,其本身形成本公开的独立方面。核糖开关是细菌中调节基因表达的常见特征,也是实现对生物功能的rna控制的手段。核糖开关是可存在于mrna的5′非翻译区中的多核苷酸且经由结合小分子配体而允许对基因表达的调节控制,该小分子配体诱导或抑制核糖开关活性。通常地,核糖开关控制参与产生小分子配体的基因产物,由此形成反馈环。核糖开关通常以顺式方式作用,尽管核糖开关已经鉴别以反式方式作用。天然核糖开关由两个结构域组成:经由三维折叠rna结构结合配体的适配体结构域及基于不存在或存在配体诱导或抑制核糖开关活性的功能切换结构域。因此,核糖开关存在两种配体敏感的构型,表示开状态及关状态(garst等人,2011)。功能切换结构域可通过调节以下来影响多核苷酸的表达:内部核糖体进入位点、逆转录病毒基因构建体中pre-mrna剪接供体的可及性、翻译、转录终止、转录物降解、mirna表达,或shrna表达(dambach及winkler2009)。适配体结构域及功能切换结构域可用作模组化组件,允许合成性rna器件控制基因表达,作为天然适配体、突变/经进化的天然适配体,或通过筛选随机rna文库所鉴定的全合成适配体(mckeague等人2016)。嘌呤核糖开关家族是发现了超过500个序列的最大家族之一(mandal等人2003;us20080269258及wo2006055351)。嘌呤核糖开关具有类似结构,由三个保守螺旋元件/茎结构(p1、p2、p3)与中间环/接合元件(j1-2、l2、j2-3、l3、j3-1)组成。由于序列改变,嘌呤家族核糖开关的适配体结构域通过不同的嘌呤化合物(例如腺嘌呤、鸟嘌呤、腺苷、鸟苷、脱氧腺苷、脱氧鸟苷(图5)等)而在其亲和力/调节上天然不同(kim等人2007)。在一个方面中,本文提供用于调节靶多核苷酸表达的经分离多核苷酸,包括:编码可操作连接至启动子和核糖开关的靶多核苷酸的多核苷酸,其中该核糖开关包括:a)能够结合核苷类似抗病毒药物且相对于核苷类似抗病毒药物与鸟嘌呤或2′-脱氧鸟苷的结合减少的适配体结构域;及b)能够调节靶多核苷酸表达的功能切换结构域,其中核苷类似物与适配体结构域结合诱导或抑制功能切换结构域的表达调节活性,从而调节靶基因的表达。在一些实施方式中,靶多核苷酸可以是多肽编码区、mirna或shrna。在一非限制性实例中,核糖开关可操作连接至具有体内活性(例如在治疗疾病时有效)的编码多肽、mirna或shrna的核酸。举例而言,在此非限制性实例中,核糖开关可操作连接至编码嵌合抗原受体的核酸。在本文所提供的非限制说明性实例中,靶多核苷酸编码包括在本公开开多个其他方面中的一种或多种经改造信号传导多肽。在这种非限制说明性实例中,核糖开关及编码一种或多种经改造信号传导多肽的靶多核苷酸可存在于包装细胞的基因组中、复制缺陷型重组逆转录病毒颗粒中、t细胞和/或nk细胞中。在一些实施方式中,适配体结构域可在30个、35个、40个、45个、50个、55个、60个、65个及70个核苷酸长度的范围下限与35个、40个、45个、50个、55个、60个、65个、70个、75个、80个、85个、90个、95个及100个核苷酸长度范围上限之间,例如长度在45个与80个核苷酸之间,长度在45个与60个核苷酸之间或长度在45个与58个核苷酸之间。在说明性实施方式中,核苷类似抗病毒药物可以是药物配体阿昔洛韦(acyclovir)(也称为阿昔洛维及无环鸟苷)或喷昔洛韦(penciclovir)(图5)。在一些实施方式中,适配体结构域对核苷类似抗病毒药物的结合亲和力可比对核苷或核苷酸的结合亲和力大例如至少2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍、10倍、20倍、30倍、40倍、50倍、60倍、70倍、80倍、90倍或100倍。在一些实施方式中,控制元件(例如核糖开关)可操作连接至靶基因且可控制靶基因在体内和/或体外的表达,促进经转导t细胞的体内扩增。在一些实施方式中,扩增取决于控制元件的存在。然而,在另一些实施方式中,经转导t细胞的扩增可通过其他因素例如受试者中白介素的存在及重组t细胞上的car的astr与其配体的结合来至少部分地驱动。在一些实施方式中,在从血液分离pbl之前、期间和/或之后且在t细胞和/或nk细胞与包括控制元件的复制缺陷型重组逆转录病毒颗粒接触之前将核苷类似抗病毒药物(例如阿昔洛韦或喷昔洛韦)施用至受试者,该控制元件在说明性非限制性实例中为结合至核苷类似抗病毒药物且调节一种或多种靶多核苷酸的表达的核糖开关。一种或多种靶多核苷酸可编码一种或多种多肽,这种多肽在非限制说明性实例中为一种或多种经改造信号传导多肽,其中至少一种编码至少一种淋巴增殖性元件。在一些实施方式中,在从血液分离pbl之前或在t细胞和/或nk细胞与复制缺陷型重组逆转录病毒颗粒接触之前5分钟、10分钟、15分钟、30分钟与60分钟的范围下限与1.5小时、2小时、3小时、4小时、5小时、6小时、8小时、12小时、24小时、48小时或72小时的范围上限之间将核苷类似抗病毒药物(例如阿昔洛韦或喷昔洛韦)施用至受试者。在一些实施方式中,在从血液分离pbl之后或在t细胞和/或nk细胞与本文所提供的方法中的复制缺陷型重组逆转录病毒颗粒接触之后1.5小时、2小时、3小时、4小时、5小时、6小时、8小时、12小时或24小时的范围下限与1/2天、1天、2天、3天、4天、5天、6天、7天、10天、14天、21天或28天的范围上限之间将核苷类似抗病毒药物(例如阿昔洛韦或喷昔洛韦)施用至受试者。在一些实施方式中,在从血液分离pbl之后或在t细胞和/或nk细胞与本文所提供方法中的复制缺陷型重组逆转录病毒颗粒接触之后的至少1.5小时、2小时、3小时、4小时、5小时、6小时、8小时、12小时或24小时,或至少2天、3天、4天、5天、6天、7天、10天、14天、21天或28天将核苷类似抗病毒药物(例如阿昔洛韦或喷昔洛韦)施用至受试者。在一些实施方式中,在pbl已再输注至受试者之后的至少1天、2天、3天、4天、5天、7天、10天、14天、21天、28天、30天、60天、90天或120天,或5个月、6个月、9个月、12个月、24个月、36个月、48个月、60个月、72个月、84个月、96个月、120个月或无限期将核苷类似抗病毒药物(例如阿昔洛韦或喷昔洛韦)施用至受试者。在任何本文所公开的实施方式中,可在pbl的再输注之前和/或期间和/或pbl已再输注之后施用核苷类似抗病毒药物。在一些实施方式中,施用核苷类似抗病毒药物直至受试者不再经受靶多核苷酸相关的疾病症状,或不再罹患该疾病。在一些实施方式中,相对于阿昔洛韦或替代地另一抗病毒剂,适配体结构域可优先结合喷昔洛韦,使得可利用伴随的抗病毒疗法而不影响核糖开关。在一些实施方式中,适配体结构域可结合喷昔洛韦,且结合亲和力比该适配体结构域结合阿昔洛韦或另一抗病毒剂大例如至少2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍、10倍、20倍、30倍、40倍、50倍、60倍、70倍、80倍、90倍或100倍。在一些实施方式中,相对于喷昔洛韦或替代地另一抗病毒剂,适配体结构域可优先结合阿昔洛韦,使得可利用伴随的抗病毒疗法而不影响核糖开关。在一些实施方式中,适配体结构域可结合阿昔洛韦,且结合亲和力比该适配体结构域结合喷昔洛韦或另一抗病毒剂大例如至少2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍、10倍、20倍、30倍、40倍、50倍、60倍、70倍、80倍、90倍或100倍。在一些实施方式中,可向受试者给予喷昔洛韦(泛昔洛韦)及阿昔洛韦(伐昔洛韦)的口服前药。在一些实施方式中,经分离多核苷酸的适配体结构域可与序列seqidno:87至seqidno:93中任何一种有至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%序列同一性或与其一致,且保留结合阿昔洛韦的能力及相对于核苷类似抗病毒药物结合鸟嘌呤或2′-脱氧鸟苷的减小的能力,且其中该适配体结构域保留在被阿昔洛韦结合时诱导或抑制功能切换结构域的表达调节活性的能力。在一些实施方式中,经分离多核苷酸的适配体结构域可与seqidno:94至seqidno:100的适配体结构域有至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%序列同一性或与其一致,且保留结合喷昔洛韦的能力及相对于核苷类似抗病毒药物结合鸟嘌呤或2’-脱氧鸟苷的减小的能力,且其中该适配体结构域保留在被喷昔洛韦结合时诱导或抑制功能切换结构域的表达调节活性的能力。在一些实施方式中,经分离多核苷酸的区或核糖开关的区可与序列seqidno:87至seqidno:100中任何一种有至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%序列同一性或与其一致。在一些实施方式中,含有经分离多核苷酸的适配体结构域的区的dna序列可与序列seqidno:108至seqidno:221中任何一种有至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%序列同一性或与其一致。在一些实施方式中,经分离多核苷酸的区可与序列seqidno:108至seqidno:221中任何一种有至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%序列同一性或与其一致。在一些实施方式中,含有经分离多核苷酸的适配体结构域的区的dna序列可与seqidno:108有至少80%、85%、90%、91%、91.84%、92%、93%、94%、95%、96%、97%、98%或99%序列同一性或与其一致。在一些实施方式中,含有经分离多核苷酸的适配体结构域的区的dna序列可与seqidno:147有至少80%、85%、90%、91%、92%、93%、94%、95%、95.83%、96%、97%、98%或99%序列同一性或与其一致。在一些实施方式中,含有经分离多核苷酸的适配体结构域的区的dna序列可与seqidno:164有至少80%、85%、90%、91%、92%、93%、93.88%、94%、95%、96%、97%、98%或99%序列同一性或与其一致。在一些实施方式中,含有经分离多核苷酸的适配体结构域的区的dna序列可与seqidno:183有至少80%、85%、90%、91%、92%、93%、94%、95%、95.83%、96%、97%、98%或99%序列同一性或与其一致。在一些实施方式中,含有经分离多核苷酸的适配体结构域的区的dna序列可与seqidno:198有至少80%、85%、90%、91%、91.84%、92%、93%、94%、95%、95.83%、96%、97%、98%或99%序列同一性或与其一致。在一些实施方式中,经分离多核苷酸的区可包括共有序列seqidno:222至seqidno:226中任何一种。在一些实施方式中,经分离多核苷酸的区可与序列seqidno:222至seqidno:226中任何一种有至少80%、85%、90%、91%、92%、93%、94%、95%、95.83%、96%、97%、98%或99%序列同一性或与其一致。在任何本文所公开的实施方式中,经分离多核苷酸可保留结合阿昔洛韦和/或喷昔洛韦的能力。在任何本文所公开的实施方式中,经分离多核苷酸可以是序列seqidno:87至seqidno:100或seqidno:108至seqidno:221中任何一种的反向互补体。在任何本文所公开的实施方式中,经分离多核苷酸可以是dna序列seqidno:108至seqidno:221或与seqidno:108至seqidno:221互补的dna序列的转录物或rna版本。在任何本文所公开的实施方式中,经分离多核苷酸可以是rna序列seqidno:87至seqidno:100中任何一种的反向转录物或dna版本或与rna序列seqidno:87至seqidno:100中任何一种反向转录物互补的dna链。在本文提供的一些实施方式中,核糖开关支架可用于突变分析或分子进化。针对突变分析或分子进化所选择的核糖开关可来自任何已知的生物体,例如细菌。在一些实施方式中,来自mesoplasmaflorum的i-a类脱氧鸟苷核糖开关可用于分子进化。在一些实施方式中,衍生的适配体结构域可与来自mesoplasmaflorum的i-a类脱氧鸟苷核糖开关的适配体结构域至少50%、60%、70%、75%、80%、85%、90%或95%相同(seqidno:237)。在另一些实施方式中,可使用来自枯草芽孢杆菌(bacillussubtilis)的xpt核糖开关。在一些实施方式中,衍生的适配体结构域可与来自枯草芽孢杆菌之xpt核糖开关的适配体结构域(seqidno:243)至少50%、60%、70%、75%、80%、85%、90%或95%相同。适配体结构域可用作分子组件且与功能切换结构域中任何一种组合以影响rna转录物。在本文公开的任何实施方式中,核糖开关可通过调节以下活性中任何一种来影响rna转录物:内部核糖体进入位点(ires)、pre-mrna剪接供体可及性、翻译、转录终止、转录降解、mirna表达,或shrna表达。在一些实施方式中,功能切换结构域可控制抗ires与ires结合(参见例如ogawa,rna(2011),17:478-488,其公开内容以全文引用方式并入本文中)。在本文公开的任何实施方式中,小分子配体的存在或不存在可使得核糖开关影响rna转录物。在一些实施方式中,核糖开关可包括核酶。具有核酶的核糖开关可在存在小分子配体的情况下抑制或增强对靶多核苷酸的转录降解。在一些实施方式中,核酶可以是手枪类核酶,锤头类核酶、扭曲类核酶、斧头类核酶,或hdv(丁型肝炎病毒)核酶。在本文公开的任何实施方式中,核糖开关可定位于相对于靶多核苷酸的多种位置中,如通常对于核糖开关已知的。在一些实施方式中,核糖开关可调节pre-mrna剪接供体可及性且定位于靶多核苷酸之前。在一些实施方式中,核糖开关可调节poly(a)尾部的包含性且定位于靶多核苷酸之后。在一些实施方式中,核糖开关可调节抗ires且可定位于ires上游。在非限制性说明性实施方式中,本文提供的核糖开关可定位于相对于编码本文提供的一个或多个经改造信号传导多肽的核酸的任何这种位置中。在一些实施方式中,核糖开关可在高于37.5℃、38℃、38.5℃、39℃、39.5℃或40℃的温度下去稳定化,从而使得核糖开关不再对配体具有反应性。在一些实施方式中,分子进化可用于选择在高于37.5℃、38℃、38.5℃、39℃、39.5℃或40℃的温度下去稳定化的核糖开关。在一些实施方式中,靶多核苷酸可编码mirna、shrna和/或多肽,其中靶多核苷酸可操作地连接至启动子。在一些实施方式中,靶多核苷酸可编码淋巴增殖性元件。在一些实施方式中,靶多核苷酸可以是mirna或shrna。在一些实施方式中,mirna或shrna可增强stat5途径或抑制socs途径。在一些实施方式中,mirna或shrna可靶向socs1、smad2、tgfb或pd-1的转录物。在一些实施方式中,mirna为mir-115。在一些实施方式中,靶多核苷酸编码多肽且该多肽可包括包含抗原特异性靶向区、跨膜结构域及胞内活化结构域的car。在另一方面中,本文提供用于调节靶多核苷酸表达的经分离多核苷酸,包括:编码可操作连接至启动子及核糖开关的靶多核苷酸的多核苷酸,其中该核糖开关包括:a)适配体结构域,其能够结合核苷类似抗病毒药物,且结合亲和力为适配体结构域结合鸟嘌呤或2′-脱氧鸟苷亲和力的至少两倍;及b)能够调节靶多核苷酸表达的功能切换结构域,其中核苷类似物与适配体结构域的结合诱导或抑制功能切换结构域的表达调节活性。在一些实施方式中,适配体结构域可结合核苷类似抗病毒药物且结合亲和力比该适配体结构域结合鸟嘌呤或2'-脱氧鸟苷亲和力大至少3倍、4倍、5倍、6倍、7倍、8倍、9倍、10倍、20倍、30倍、40倍、50倍、60倍、70倍、80倍、90倍或100倍。在一些实施方式中,适配体结构域的长度可在下限为30个、35个、40个、45个、50个、55个、60个、65个及70个核苷酸与上限为35个、40个、45个、50个、55个、60个、65个、70个、75个、80个、85个、90个、95个及100个核苷酸之间的范围,例如长度在45个与80个核苷酸之间或长度在45个与58个核苷酸之间。在说明性实施方式中,核苷类似抗病毒药物可以是药物配体阿昔洛韦(也称为阿昔洛维及无环鸟苷)或喷昔洛韦。在一些实施方式中,适配体结构域可具有比对核苷或核苷酸的结合亲和力大例如至少2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍、10倍、20倍、30倍、40倍、50倍、60倍、70倍、80倍、90倍或100倍的对核苷类似抗病毒药物的结合亲和力。在一些实施方式中,核苷类似物与适配体结构域的结合可诱导核糖开关中的活性。在一些实施方式中,适配体结构域可对于喷昔洛韦具有特异性且对阿昔洛韦或另一种抗病毒剂缺乏反应性,使得可利用伴随抗病毒治疗而不影响核糖开关。在一些实施方式中,适配体结构域可结合喷昔洛韦且结合亲和力比该适配体结构域结合阿昔洛韦或另一抗病毒剂大至少2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍、10倍、20倍、30倍、40倍、50倍、60倍、70倍、80倍、90倍或100倍。在一些实施方式中,适配体结构域可对于阿昔洛韦具有特异性且对喷昔洛韦或另一抗病毒剂缺乏反应性,使得可利用伴随抗病毒治疗而不影响核糖开关。在一些实施方式中,适配体结构域可结合阿昔洛韦且结合亲和力比该适配体结构域结合喷昔洛韦或另一抗病毒剂大至少2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍、10倍、20倍、30倍、40倍、50倍、60倍、70倍、80倍、90倍或100倍。在一些实施方式中,可向受试者给予喷昔洛韦(泛昔洛韦)及阿昔洛韦(伐昔洛韦)的口服前药。在一些实施方式中,衍生的适配体结构域可与来自mesoplasmaflorum的i-a类脱氧鸟苷核糖开关的适配体结构域至少50%、60%、70%、75%、80%、85%、90%或95%相同。在一些实施方式中,衍生的适配体结构域可与来自枯草芽孢杆菌的xpt核糖开关的适配体结构域至少50%、60%、70%、75%、80%、85%、90%或95%相同。在本文公开的任何实施方式中,核糖开关可通过调节任何以下活性来影响rna转录物:内部核糖体进入位点、逆转录病毒基因构建体中的pre-mrna剪接供体可及性、翻译、转录终止、转录降解、mirna表达,或shrna表达。在一些实施方式中,功能切换结构域可控制抗ires与ires的结合。在本文公开的任何实施方式中,小分子配体的存在或不存在可使得核糖开关影响rna转录物。在一些实施方式中,核糖开关可包括核酶。具有核酶的核糖开关可在存在小分子配体的情况下抑制或增强对目的基因的转录降解。在一些实施方式中,核酶可以是手枪类核酶、锤头类核酶、扭曲类核酶、斧头类核酶,或hdv(丁型肝炎病毒)核酶。在一些实施方式中,核糖开关可在高于37.5℃、38℃、38.5℃、39℃、39.5℃或40℃的温度下去稳定化,从而使得核糖开关不再对配体具有反应性。在一些实施方式中,分子进化可用于选择在高于37.5℃、38℃、38.5℃、39℃、39.5℃或40℃的温度下去稳定化的核糖开关。在一些实施方式中,靶多核苷酸可编码mirna、shrna和/或多肽,其中靶多核苷酸可操作地连接至启动子。在一些实施方式中,靶多核苷酸可编码淋巴增殖性元件。在一些实施方式中,靶多核苷酸可以是mirna,且任选地,mirna可刺激stat5途径或抑制socs途径。在一些实施方式中,mirna可靶向socs1、shp、smad2、tgfb或pd-1的转录物。在这种实施方式中,mirna可以是mir-115。在一些实施方式中,靶多核苷酸编码多肽且该多肽可包括包含抗原特异性靶向区、跨膜结构域,及胞内活化结构域的car。car的其他实施方式公开于本文其他处。在一些实施方式中,适配体的进化可以通过使用selex(通过指数富集的配体系统进化)方法从随机化天然嘌呤或鸟嘌呤适配体文库进行适配体选择来执行,该方法包括但不限于在选择过程及筛选中采用氧化石墨烯的方法。在另一些实施方式中,例如易错pcr的随机突变方法可用以进化适配体构建体或核糖开关构建体,其中该适配体通过在体外或在哺乳动物细胞内筛选来整合入本文所描述的任何核糖开关活性的内容中。在另一些实施方式中,核苷酸的随机文库可用于核糖开关的进化中。在本文所公开的任何实施方式中,可通过在体外或在哺乳动物细胞内筛选此类文库来鉴定核糖开关。在一些实施方式中,进化或衍生的适配体结构域可增强对天然配体的类似物的结合且减弱对天然配体的结合。在一些实施方式中,适配体结构域可被配置以增强对天然配体的类似物的结合且减弱对天然配体的结合。在一些实施方式中,适配体结构域可衍生自嘌呤核糖开关家族。在一些实施方式中,天然配体可以是核苷或核苷酸且类似物可以是核苷类似物或核苷酸类似物。在一些实施方式中,核苷类似物为抗病毒药物。在说明性实施方式中,适配体结构域可衍生自2'-脱氧鸟苷及鸟嘌呤核糖开关支架且衍生的适配体结构域可显示相对于野生型核糖开关对2'-脱氧鸟苷及鸟嘌呤的降低的结合。在一些实施方式中,核糖开关可调节逆转录病毒基因构建体中的pre-mrna剪接供体可及性,其中该逆转录病毒构建体在一般启动子或t细胞特异性启动子下自反向链驱动car基因或其他目的基因。在另一些实施方式中,核糖开关可调节逆转录病毒基因构建体中的ires,其中该逆转录病毒构建体驱动car基因或其他目的基因的翻译。在另一些实施方式中,核糖开关可控制rna、mirna的转录终止或基因转录物,或可控制转录物的翻译。在另一些实施方式中,可将核苷类似物核糖开关与核酶整合以在核苷类似物的存在下抑制或增强car基因或其他目的基因的转录降解。在一些实施方式中,用于调节靶多核苷酸表达的经分离多核苷酸为分子克隆载体,该靶多核苷酸包括编码可操作地连接至启动子的靶多核苷酸及结合核苷类似抗病毒药物的核糖开关的多核苷酸。分子克隆载体可以是本领域已知的任何类型的分子克隆载体。作为非限制性实例,载体可以是质粒、病毒,或复制缺陷型重组逆转录病毒颗粒,其中任何可以是表达载体。该表达载体可编码在上文所提供的任何靶多核苷酸。一个或多个限制位点和/或多克隆位点可包括于分子克隆载体上,在本文所提供的核糖开关的5'或3',从而使得该核糖开关可操作地连接至插入至该限制位点和/或多克隆位点中的靶多核苷酸。分子伴侣在一个方面中,本文中提供一种用于遗传修饰及扩增受试者的淋巴细胞的方法,其包含:a.使受试者的静息t细胞和/或nk细胞在离体的情况下(通常不需要先前离体刺激)与复制缺陷型重组逆转录病毒颗粒接触,该复制缺陷型重组逆转录病毒颗粒包含:i.在其表面上的假型化元件,该假型化元件能够与t细胞和/或nk细胞结合且有助于复制缺陷型重组逆转录病毒颗粒至其上的膜融合;及ii.多核苷酸,其包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码由控制元件调节的第一经改造信号传导多肽,其中该第一经改造信号传导多肽包含至少一个淋巴增殖性元件和/或嵌合抗原受体,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导至少一些静息t细胞和/或nk细胞,从而产生遗传修饰的t细胞和/或nk细胞;b.将遗传修饰的t细胞和/或nk细胞引入至受试者中;及c.使遗传修饰的t细胞和/或nk细胞体内暴露于充当控制元件的化合物,以影响第一经改造信号传导多肽的表达且促进淋巴细胞在体内扩增,从而遗传修饰且扩增受试者的淋巴细胞。在说明性实施方式中,在无需离体刺激的情况下进行转导。在说明性实施方式中,该化合物为分子伴侣,例如小分子分子伴侣。在说明性实施方式中,分子伴侣与淋巴增殖性元件和/或car组分的结合增强淋巴增殖性元件和/或car的增殖活性。可在采集血液之前、在接触期间和/或在将t细胞和/或nk细胞引入至受试者中之后将分子伴侣施用至受试者。此方面的一些实施方式包括从受试者采集血液。在这些实施方式中,引入是将经收集且在重新引入之前遗传修饰的细胞重新引入。在说明性实施方式中,整个过程比现有技术方法的过程更短,本文的其他方面也是如此。举例而言,整个过程可在少于48小时、少于24小时,或少于12小时内完成。在另一些实施方式中,整个过程可在范围下限是2小时、4小时、6小时,或8小时且在范围上限是12小时、24小时、36小时或48小时内完成。因此,在本文所提供的方法及组合物的一些实施方式中,控制元件为分子伴侣。与本文中具有其他体内控制元件(例如通常结合化合物以影响本文中的第一或第二经改造信号传导多肽的淋巴增殖性元件或其他组分的表达的核糖开关)的其他实施方式相比,分子伴侣为这样的化合物,其为控制元件且因此直接影响(通常通过结合)本文中的第一或第二经改造信号传导多肽的淋巴增殖性元件或其他组分的活性。在本文方法的此类实施方式的包括施用分子伴侣的说明性实例中,淋巴增殖性元件、膜结合细胞因子和/或car组分可以是较低活性或非活性的淋巴增殖性元件、膜结合细胞因子,和/或car组分,其通过结合分子伴侣以增加其活性。因此,由分子伴侣结合的靶标通常为靶多肽。在一些实施方式中,如所指示,多肽可以是第一经改造信号传导多肽和/或第二经改造信号传导多肽,或其多肽组分,其活性通过结合至分子伴侣受影响,在说明性实施方式中为小分子分子伴侣。在一些实施方式中,多肽可包括活性被调节的淋巴增殖性元件,在说明性实施方式中其活性被分子伴侣(优选地小分子分子伴侣)上调。本文所提供方法中的分子伴侣可以是结合至突变淋巴增殖性元件和/或非活性car组分,由此使其活化的化合物。在另一些实施方式中,淋巴增殖性元件或其他信号传导结构域已经突变以仅在存在小分子合成伴侣的情况下允许输送至质膜。在另一些实施方式中,伴侣作为增强剂促进淋巴增殖性元件或其他信号传导结构域或蛋白质的稳定性及半衰期。应理解,本发明的方面及实施方式包括本文详细提供的许多相同的步骤和化合物。因此,应理解,贯穿本说明书的涉及这些共同要素的教导适用于利用分子伴侣作为控制元件的方面及实施方式,该分子伴侣通常直接结合淋巴增殖性元件或其他靶分子,附加本文中所提供的其他体内控制元件(例如核糖开关)或作为其的替代,其通常利用结合核糖开关的分子,例如药物。在一些实施方式中,分子伴侣是可调节靶标的亚细胞定位的化合物,例如将例如淋巴增殖性元件和/或car的组分的靶标蛋白质恰当地折叠且自内质网输送至质膜或其表面上的半衰期。在另一些实施方式中,分子伴侣可促进功能失调的靶标的功能性构型,因此充当增强剂。充当天然突变蛋白质的伴侣或增强剂的分子的实例包括鲁玛卡托(lumacaftor)及依伐卡托(ivacaftor)。这种蛋白质对例如g551d或f508del的突变cftr氯通道变体起作用。依伐卡托增强g551d或f508del突变的离子通道的活性,而鲁玛卡托促进突变体氯通道的稳定化及依伐卡托的后续增强。这种伴侣依赖性蛋白质可由天然功能性蛋白质产生且仅在存在分子伴侣的情况下筛选功能活性。因此,此类蛋白质仅在伴侣存在时具有活性。可针对特异性伴侣活性进行筛选这种分子的实例包括对正常人类蛋白质显示无活性的小分子抗病毒剂或抗感染剂。因此,在一个实施方式中,用于本文方法中的分子伴侣为对正常人类蛋白质显示无活性的小分子抗病毒化合物或抗感染化合物。在一些实施方式中,可暴露遗传修饰的淋巴细胞和/或可向受试者施用分子伴侣。在一些实施方式中,在从血液分离pbl之前、期间和/或之后及在使t细胞和/或nk细胞与复制缺陷型重组逆转录病毒颗粒接触之前将化合物施用受试者。在这种实施方式中,复制缺陷型重组逆转录病毒颗粒包括较低活性或非活性的结合至分子伴侣化合物且由该分子伴侣化合物调节的car组分和/或淋巴增殖性元件。对于本文所提供的用于修饰及扩增淋巴细胞任何实施方式中(其可以是过继性细胞疗法方法的一部分),可在自血液分离pbl之前或在使t细胞和/或nk细胞与复制缺陷型重组逆转录病毒颗粒接触之前,可在范围下限为5分钟、10分钟、15分钟、30分钟及60分钟与范围上限为1.5小时、2小时、3小时、4小时、5小时、6小时、8小时、12小时,或24小时之间施用化合物给受试者。在一些实施方式中,在自血液分离pbl之后或在t细胞和/或nk细胞与本文所提供的方法中的复制缺陷型重组逆转录病毒颗粒接触之后,在范围下限为1.5小时、2小时、3小时、4小时、5小时、6小时、8小时、12小时或24小时与范围上限为1/2天、1天、2天、3天、4天、5天、6天、7天、10天、14天、21天或28天之间将化合物施用受试者。在一些实施方式中,在从血液分离pbl之后或在t细胞和/或nk细胞与本文所提供方法中的复制缺陷型重组逆转录病毒颗粒接触之后的至少1.5小时、2小时、3小时、4小时、5小时、6小时、8小时、12小时或24小时,或至少2天、3天、4天、5天、6天、7天、10天、14天、21天或28天将化合物施用受试者。在一些实施方式中,在pbl已再输注至受试者之后的至少1天、2天、3天、4天、5天、7天、10天、14天、21天、28天、30天、60天、90天或120天,或5个月、6个月、9个月、12个月、24个月、36个月、48个月、60个月、72个月、84个月、96个月、120个月或无限期将化合物施用受试者。在本文所公开的任何实施方式中,可在再输注pbl之前和/或期间和/或pbl已再输注之后施用化合物。对于任何本文的实施方式,分子伴侣不在化合物所结合的控制元件中,该化合物对其进行调节和/或活化。分子伴侣为作为控制元件且调节淋巴增殖性元件和/或car的功能性组分的活性的化合物,优选地小分子化合物。制造重组逆转录病毒颗粒的包装细胞系/方法在一个方面中,本文提供逆转录病毒包装系统,其包括:哺乳动物细胞,其包括:a)第一反式活化子,其由组成型启动子表达且能够结合第一配体及第一可诱导启动子以在存在相对于不存在第一配体的情况下影响可操作地连接至其的核酸序列的表达;b)第二反式活化子,其能够结合第二配体及第二可诱导启动子,且在存在相对于不存在第二配体的情况下影响可操作地连接至其的核酸序列的表达;及c)逆转录病毒颗粒的可包装rna基因组,其中该第一反式活化子调节该第二反式活化子的表达,且其中该第二反式活化子调节参与病毒包装的逆转录病毒多肽的表达,该逆转录病毒多肽是例如gag多肽、pol多肽和/或假型化元件及任选地将整合入复制缺陷型重组逆转录病毒颗粒中或整合入复制缺陷型重组逆转录病毒颗粒上且被认为对包装细胞系具有毒性的其他多肽,例如hek-293。在某些方面中,第二反式活化子自身对于包装细胞系具有细胞毒性。假型化元件通常能够结合至靶细胞的细胞膜且有助于其融合,如本文所详细论述。因此,不受理论限制,系统提供累积不抑制或不实质上抑制或不被认为抑制哺乳动物细胞增殖或存活的某些多肽/蛋白质(例如非毒性蛋白质)的能力,同时培养哺乳动物细胞群体持续数天或无限期,且控制期望用于逆转录病毒产物但对哺乳动物细胞的存活和/或增殖具有抑制性或可具有抑制性或已经报道具有抑制性的多肽(例如毒性多肽)的诱导,直至以后临近产生并采集复制缺陷型重组逆转录病毒颗粒时。可包装rna基因组通常由可操作地连接至启动子(出于方便起见有时在本文中称为第三启动子)的多核苷酸编码,其中该第三启动子通常可由第一反式活化子或第二反式活化子诱导。在说明性实施方式中,可包装rna基因组由可操作地连接至第三启动子的多核苷酸编码,其中该第三启动子能够由第二反式活化子诱导。因此,可包装rna基因组可以在以后临近采集复制缺陷型重组逆转录病毒颗粒时产生。本领域技术人员将理解,许多不同的反式活化子、配体,及可诱导启动子可用于逆转录病毒包装系统中。这种可诱导启动子可分离和来源于许多生物,例如真核生物及原核生物。对用于第二生物中的来源于第一生物的可诱导启动子(例如第一原核生物及第二真核生物、第一真核生物及第二原核生物,等)的修饰为本领域中已知的。这种可诱导启动子及基于此类可诱导启动子但也包括其他控制蛋白质的系统包括但不限于醇调节的启动子(例如醇脱氢酶i(alca)基因启动子、对醇反式活化子蛋白质具有反应性的启动子(alcr)等)、四环素调节的启动子(例如包括tetactivators、teton、tetoff等的启动子系统)、类固醇调节的启动子(例如大鼠糖皮质激素受体启动子系统、人类雌性激素受体启动子系统、类视黄素启动子系统、甲状腺启动子系统、蜕皮激素启动子系统、米非司酮启动子系统等)、金属调节的启动子(例如金属硫蛋白启动子系统等)、病原相关调节的启动子(例如水杨酸调节的启动子、乙烯调节的启动子、苯并噻二唑调节的启动子等)、温度调节的启动子(例如热休克可诱导启动子(例如hsp-70、hsp-90、大豆热休克启动子等))、光调节的启动子、合成可诱导启动子及类似者。在一些实施方式中,可使用米非司酮调节的系统。在一些实施方式中,可使用具有自动调节反馈环路的米非司酮可诱导系统。在一些实施方式中,gal4调节融合蛋白质由也含有转座子端重复位点及lox位点及frt位点的一个构建体表达。在一些实施方式中,gal4调节融合蛋白质控制反向tet反式活化子(rtta)及bitre的表达。在一些实施方式中,具有lox位点及frt位点的另一构建体含有gal4上游活化序列(uas)及驱动如mcherry的报告子的e1btata盒启动子。在一些实施方式中,gal4调节融合蛋白在控制gal4调节融合蛋白表达的启动子及控制靶多核苷酸表达的启动子两者上结合gal4上游活化序列(uas)。在一些实施方案中,米非司酮、多西环素及嘌呤霉素将用于包装细胞系的诱导及选择。在一些实施方式中,任何一个或两个反式活化子可以分成两个或更多个多肽。在一些实施方式中,该两个或更多个多肽可包括dna结合结构域及能够刺激单独多肽上的转录的活化结构域。不要将此“活化结构域”与“活化元件”混淆,例如结合cd3的多肽,其能够活化t细胞和/或nk细胞,且在与此t细胞和/或nk细胞接触时通常将其活化,如本文所详细论述。分离的多肽可进一步包括具有能够通过添加配体而二聚化的多肽的融合物。在一些实施方式中,活化结构域可以是p65活化结构域或其功能片段。在本文包装系统的说明性实施方式中,dna结合结构域可以是来自zfhd1的dna结合结构域或其功能片段。在一些实施方式中,一个多肽可以是具有fkbp或其功能突变体和/或其片段或多个fkbp的融合物,且另一多肽可以是具有mtor的frb结构域或其功能突变体和/或其片段的融合物,且配体可以是雷帕霉素或功能性雷帕类似物。在一些实施方式中,frb含有突变k2095p、t2098l和/或w2101f。在一些实施方式中,分离的多肽可以是fkbp或其功能片段,及钙依赖磷酸酶a或其功能片段,且二聚化剂可以是fk506。在一些实施方式中,分离的多肽可以是fkbp或其功能片段,及cyp-fas或其功能片段,且二聚化剂可以是fkcsa。在一些实施方式中,分离的多肽可以是gai或其功能片段,及gid1或其功能片段,且二聚化剂可以是赤霉素。在一些实施方式中,分离的多肽可以是snap-tag及halotag或其功能片段且二聚化剂可以是haxs。在一些实施方式中,分离的多肽可包括相同的多肽。举例而言,dna结合结构域及活化结构域可表达为具有fkbp或gyrb的融合蛋白,且二聚化剂可分别为fk1012或香豆霉素。在一些实施方式中,可诱导启动子可以是dna结合结构域通常结合的dna序列。在一些实施方式中,可诱导启动子可不同于dna结合结构域通常结合的dna序列。在一些实施方式中,反式活化子可以是rtta,配体可以是四环素或多西环素,且可诱导启动子可以是tre。在说明性实施方式中,第一反式活化子为融合至frb的p65活化结构域及融合至三个fkbp多肽的zfhd1dna结合结构域,且第一配体为雷帕霉素。在另一些说明性实施方式中,第二反式活化子可以是rtta,第二配体可以是四环素或多西环素,且可诱导启动子可以是tre。在一些实施方式中,第一反式活化子可调节元件的表达以控制含有共有序列的转录物的核输出,例如hivrev,且该共有序列可以是rev反应元件。在说明性实施方式中,靶细胞为t细胞。在一些实施方式中,假型化元件为逆转录病毒包膜多肽。假型化元件通常包括用于将靶细胞与病毒膜结合并促进靶细胞与病毒膜的膜融合的结合多肽及促融合多肽,如本文更详细地论述。在一些实施方式中,假型化元件为猫内源性病毒(rd114)包膜蛋白质、肿瘤逆转录病毒双嗜性包膜蛋白质、肿瘤逆转录病毒嗜亲性包膜蛋白质和/或水泡性口炎病毒包膜蛋白质(vsv-g)。在说明性实施方式中,假型化元件包括结合多肽及衍生自不同蛋白质的促融合多肽,如本文进一步详细论述。举例而言,在说明性实施方式中,尤其在靶细胞为t细胞和/或nk细胞时,结合多肽为麻疹病毒的凝血素(h)多肽(例如麻疹病毒的埃德蒙斯顿菌株(edmonstonstrain))或其细胞质结构域缺失变体,且其他促融合多肽为麻疹病毒的融合(f)多肽(例如麻疹病毒的埃德蒙斯顿菌株)或其细胞质结构域缺失变体。在一些实施方式中,促融合多肽可包括表达为一个多肽的多个元件。在一些实施方式中,结合多肽及促融合多肽可翻译自同一转录物,但翻译自不同核糖体结合位点,或在翻译后使用肽切割信号或核糖体跳过序列来切割多肽,如本文其他处所公开,以产生结合多肽及促融合多肽。在一些实施方式中,当结合多肽为麻疹病毒h多肽或其细胞质结构域缺失,且融合多肽为麻疹病毒f多肽或其细胞质结构域缺失时,f多肽及h多肽自分离的核糖体结合位点的翻译产生比h多肽更高量的f多肽。在一些实施方式中,f多肽(或其细胞质结构域缺失)与h多肽(或其细胞质结构域缺失)的比率为至少2:1、至少3:1、至少4:1、至少5:1、至少6:1、至少7:1,或至少8:1。在一些实施方式中,第一反式活化子可调节能够结合并活化例如t细胞的靶细胞的活化元件的表达。可表达本文所公开的任何活化元件。举例而言,在这种实施方式中,活化元件可包括:a.)能够结合并活化cd3的膜结合多肽;和/或b.)能够结合并活化cd28的膜结合多肽。在一些实施方式中,能够结合并活化cd28的膜结合多肽为cd80、cd86或其功能片段,例如cd80的胞外结构域。在一些实施方式中,第二反式活化子可调节可包装rna基因组的表达,该可包装rna基因组可包括编码一种或多种靶标多肽的rna,作为非限制性实例包括本文公开的经改造信号传导多肽中的任一种和/或一种或多种(例如两种或更多种)抑制性rna分子。应注意,可预见逆转录病毒包装系统方面及制备复制缺陷型重组逆转录病毒颗粒的方法方面不限于制备用于转导t细胞和/或nk细胞的复制缺陷型重组逆转录病毒颗粒,而也适用于可通过复制缺陷型重组逆转录病毒颗粒转导的任何细胞类型。在一些说明性实施方式中,可包装rna基因组经设计以表达一种或多种靶标多肽,作为非限制性实例包括本文所公开的经改造信号传导多肽中的任一种和/或一种或多种(例如两种或更多种)抑制性rna分子,其与例如gag及pol的逆转录病毒组分相反定向(例如,在相反链上且在相反方向上编码)。举例而言,可包装rna基因组可自5′至3′包括:5'长末端重复或其活性截短片段;编码逆转录病毒顺式作用rna包装元件的核酸序列;编码第一靶标多肽及任选地第二靶标多肽的核酸序列,例如但不限于呈相反定向的经改造信号传导多肽,其可由相对于5'长末端重复及顺式作用rna包装元件呈相反方向的启动子驱动,该启动子在一些实施方式中仅出于方便起见称为“第四”启动子(且有时在本文中称为在t细胞和/或nk细胞中有活性的启动子),其在例如t细胞和/或nk细胞的靶细胞中具有活性,但在说明性实例中,在包装细胞中不具有活性,或在包装细胞中仅具有诱导性或最小活性;及3'长末端重复或其活性截短片段。在一些实施方式中,可包装rna基因组可包括中央聚嘌呤区(cppt)/中央终止序列(cts)元件。在一些实施方式中,逆转录病毒顺式作用rna包装元件可以是hivpsi。在一些实施方式中,逆转录病毒顺式作用rna包装元件可以是rev反应元件。在例示性实施方式中,由与5'长末端重复相反定向的启动子驱动的经改造信号传导多肽是本文所公开的一种或多种经改造信号传导多肽,且可任选地表达如本文更详细公开的一种或多种抑制性rna分子。应理解,例如第一启动子、第二启动子、第三启动子、第四启动子等的启动子编号仅出于方便起见。除非明确地叙述其他启动子,否则称为“第四”启动子的启动子不应被视为暗示存在任何其他启动子,例如第一启动子、第二启动子或第三启动子。应注意,这些启动子中的每一种能够驱动转录物在适当细胞类型中表达,且此类转录物形成转录单元。在一些实施方式中,经改造信号传导多肽可包括第一淋巴增殖性元件。本文其他部分中公开了合适的淋巴增殖性元件。作为非限制性实例,淋巴增殖性元件可表达为具有识别结构域的融合物,例如etag,如本文所公开。在一些实施方式中,可包装rna基因组可进一步包括编码第二经改造多肽的核酸序列,包括编码本文所提供的任何car实施方式的嵌合抗原受体。举例而言,第二经改造多肽可包括第一抗原特异性靶向区、第一跨膜结构域,及第一胞内活化结构域。抗原特异性靶向区、跨膜结构域及胞内活化结构域的实例公开于本文其他处。在靶细胞为t细胞的一些实施方式中,在靶细胞中有活性的启动子在t细胞中具有活性,如本文其他处所公开。在一些实施方式中,可包装rna基因组可进一步包括核糖开关,如本文其它部分所论述。在一些实施方式中,编码经改造信号传导多肽的核酸序列可呈反向定向。在另一些实施方式中,可包装rna基因组可进一步包括核糖开关以及任选地该核糖开关可呈反向定向。在本文所公开的任何实施方式中,包括任何这种元件的多核苷酸可包括引物结合位点。在说明性实施方式中,转录阻断剂或polya序列可置放于基因附近以防止或减少未经调节的转录。在本文所公开的任何实施方式中,编码vpx的核酸序列可位于第二转录单元上或任选地第三转录单元上,或位于可操作地连接至第一可诱导启动子的额外转录单元上。在本文中的另一方面中提供一种包含复制缺陷型逆转录病毒颗粒的可包装rna基因组的哺乳动物包装细胞系,其中该可包装rna基因组包含:a.5′长末端重复或其活化片段;b.编码逆转录病毒顺式作用rna包装元件的核酸序列;c.多核苷酸,其包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一种或多种核酸序列,其中一种或多种核酸中的第一核酸序列编码针对一种或多种rna靶标的一种或多种(例如两种或更多种)抑制性rna分子,且一种或多种核酸序列中的第二核酸序列编码嵌合抗原受体(car),该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域;及d.3′长末端重复或其活化片段。以上方面中的抑制性rna分子可包括在本公开其它部分中所提供的任何一种抑制性rna分子,作为非限制性实例,shrna或mirna。在哺乳动物包装细胞系方面的一些实施方式中,(c)的多核苷酸可与编码逆转录病毒顺式作用rna包装元件(b)、5'长末端重复(a)和/或3'长末端重复(d)的核酸序列呈相反定向。在哺乳动物包装细胞系方面的一些实施方式中,可包装rna基因组的表达通过在哺乳动物包装细胞系中具有活性的诱导型启动子驱动。在说明性实施方式中,在驱动本部分提供的这些方面中的可诱导rna及car表达的t细胞和/或nk细胞中具活性启动子,在包装细胞系中不具有活性或仅具有最小活性或可诱导活性。在说明性实施方式中,在t细胞和/或nk细胞中具有活性的此启动子位于编码一(例如两种)或多种可诱导rna及car及3'ltr的核酸之间的可包装rna基因组上。在涉及编码针对一种或多种rna靶标的一种或多种抑制性rna分子的可包装细胞或细胞系的任何方面中,至少一种抑制性rna分子及在一些实施方式中,所有抑制性rna分子可包括彼此部分地或完全互补的5'链及3'链,其中该5'链及该3'链能够形成18个至25个核苷酸rna双链体体。在一些实施方式中,5'链的长度可以是18、19、20、21、22、23、24或25个核苷酸,且3'链的长度可以是18、19、20、21、22、23、24或25个核苷酸。在一些实施方式中,5'链及3'链的长度可相同或不同。在一些实施方式中,rna双链体可包括一个或多个错配。在替代实施方式中,rna双链体不具有错配。在上文针对本文的编码针对一种或多种rna靶标的抑制性rna分子的可包装细胞或细胞系所提供的方面中的任何一种中,抑制性rna分子可以是mirna或shrna。在一些实施方式中,抑制性分子可以是mirna的前体,例如pri-mirna或pre-mirna,或shrna的前体。在一些实施方式中,一种或多种抑制性rna分子可以是人工衍生的mirna或shrna。在其他实施方式中,抑制性rna分子可以是经处理成sirna的dsrna(经转录或人工引入)或sirna本身。在一些实施方式中,抑制性rna分子可以是mirna或shrna,其具有在自然界中未发现的序列,或具有在自然界中未发现的至少一个功能区段,或具有在自然界中未发现的功能区段的组合。在说明性实施方式中,抑制性rna分子中至少一者或全部为mir-155。在上文针对本文的编码针对一种或多种rna靶标的抑制性rna分子的可包装细胞或细胞系所提供的任何方面中,该一种或多种抑制性rna分子在一些实施方式中可包含自5'至3'方向:5'臂、5'茎、环、与该5'茎部分或完全互补的3'茎,及3'臂。在一些实施方式中,两种或更多种抑制性rna分子中至少一种具有此排列。在其他实施方式中,两个或更多个抑制性rna分子的全部均具有此排列。在一些实施方式中,5'茎的长度可以是18、19、20、21、22、23、24或25个核苷酸。在一些实施方式中,3'茎的长度可以是18、19、20、21、22、23、24或25个核苷酸。在一些实施方式中,环的长度可以是3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39或40个核苷酸。在一些实施方式中,5'臂、3'臂或两者均衍生自天然存在的mirna。在一些实施方式中,5'臂、3'臂或两者均衍生自天然存在的mirna,该mirna选自:mir-155、mir-30、mir-17-92、mir-122及mir-21。在说明性实施方式中,5'臂、3'臂或两者均衍生自mir-155。在一些实施方式中,5'臂、3'臂或两者均衍生自小家鼠mir-155或人mir-155。在一些实施方式中,5'臂具有seqidno:256中所阐述的序列或为其功能变体,例如与seqidno:256长度相同,或为seqidno:256的长度的95%、90%、85%、80%、75%,或50%,或为100个核苷酸或更少、95个核苷酸或更少、90个核苷酸或更少、85个核苷酸或更少、80个核苷酸或更少、75个核苷酸或更少、70个核苷酸或更少、65个核苷酸或更少、60个核苷酸或更少、55个核苷酸或更少、50个核苷酸或更少、45个核苷酸或更少、40个核苷酸或更少、35个核苷酸或更少、30个核苷酸或更少,或25个核苷酸或更少;且与seqidno:256至少50%、55%、60%、65%、70%、75%、80%、85%、90%或95%一致的序列。在一些实施方式中,3'臂具有seqidno:260中所阐述的序列或为其功能变体,例如与seqidno:260相同的长度,或为seqidno:206的长度的95%、90%、85%、80%、75%,或50%,或为100个核苷酸或更少、95个核苷酸或更少、90个核苷酸或更少、85个核苷酸或更少、80个核苷酸或更少、75个核苷酸或更少、70个核苷酸或更少、65个核苷酸或更少、60个核苷酸或更少、55个核苷酸或更少、50个核苷酸或更少、45个核苷酸或更少、40个核苷酸或更少、35个核苷酸或更少、30个核苷酸或更少,或25个核苷酸或更少;且与seqidno:260至少50%、55%、60%、65%、70%、75%、80%、85%、90%或95%一致的序列。在一些实施方式中,3'臂包含小家鼠bic第221至283位核苷酸。在另一方面中,本文提供用于制备复制缺陷型重组逆转录病毒颗粒的方法,其包括:培养包装细胞群以累积第一反式活化子,其中这些包装细胞包括由组成型启动子表达的第一反式活化子,其中该第一反式活化子能够结合第一配体及第一可诱导启动子,以在存在与不存在第一配体的情况下影响可操作地连接至其上的核酸序列的表达,且其中第二反式活化子表达由第一反式活化子调节;在第一配体的存在下孵育包括累积的第一反式活化子的包装细胞群以累积第二反式活化子,其中第二反式活化子能够结合第二配体及第二可诱导启动子,以在存在与不存在第二配体的情况下影响可操作地连接到其上的核酸序列的表达;且在第二配体的存在下孵育包括累积的第二反式活化子的包装细胞群,从而诱导涉及病毒包装的逆转录病毒多肽的现,例如gag多肽、pol多肽和/或假型化元件,及任选地被认为抑制哺乳动物细胞增殖或存活的其他多肽,其将并入复制缺陷型重组逆转录病毒中或并入其上,从而制得复制缺陷型重组逆转录病毒颗粒。在说明性实施方式中,可包装rna基因组由可操作地连接至启动子(出于方便起见有时称作“第三”启动子)的多核苷酸编码,其中该第三启动子具有组成型活化或可由第一反式活化子或在说明性实施方式中由第二反式活化子诱导,从而制得复制缺陷型重组逆转录病毒颗粒。假型化元件通常能够结合靶细胞的细胞膜且促进靶细胞膜与复制缺陷型重组逆转录病毒颗粒膜融合。假型化元件可以是此项技术中已知的任何包膜蛋白质。在一些实施方式中,包膜蛋白质可以是水泡性口炎病毒包膜蛋白质(vsv-g)、猫内源性病毒(rd114)包膜蛋白质、肿瘤逆转录病毒双嗜性包膜蛋白质和/或肿瘤逆转录病毒嗜亲性包膜蛋白质。本领域熟练技术人员将理解,多种不同反式活化子、配体,及可诱导启动子可用于制备复制缺陷型重组逆转录病毒颗粒的方法中。适合的反式活化子、配体,及可诱导启动子公开于本文中其他处,包括上文中。本领域技术人员将进一步理解,与本文提供的逆转录病毒包装系统方面相关的上述教导也适用于制备复制缺陷型重组逆转录病毒颗粒方面的方法,且反之亦然。在一些实施方式中,第一反式活化子可调节元件的表达以控制含有共有序列的转录物的核输出,例如hivrev,且共有序列可以是rev反应元件(rre)。在说明性实施方式中,靶细胞通常为t细胞。在一些实施方式中,hivrre及编码hivrev的多核苷酸区域可分别由hiv-2rre及编码hiv-2rev的多核苷酸区域替代。在一些实施方式中,hivrre及编码hivrev的多核苷酸区域可分别由sivrre及编码sivrev的多核苷酸区域替代。在一些实施方式中,hivrre及编码hivrev的多核苷酸区域可分别由remre及编码β逆转录病毒rem的多核苷酸区域替代。在一些实施方式中,hivrre及编码hivrev的多核苷酸区域可分别由δ逆转录病毒rexrre及编码δ逆转录病毒rex的多核苷酸区域替代。在一些实施方式中,不需要rev类蛋白质且pre可由例如组成型转运元件(cte)的顺式作用rna元件替代。在一些实施方式中,假型化元件为病毒包膜蛋白质。假型化元件通常包括用于将病毒与靶细胞膜结合并促进病毒与靶细胞膜的膜融合的结合多肽及融合多肽。在一些实施方式中,假型化元件可以是猫内源性病毒(rd114)包膜蛋白质、肿瘤逆转录病毒双嗜性包膜蛋白质、肿瘤逆转录嗜亲性包膜蛋白质和/或水泡性口炎病毒包膜蛋白质(vsv-g)。在说明性实施方式中,假型化元件包括结合多肽及衍生自不同蛋白质的促融合多肽,如本文进一步详细论述。举例而言,在一说明性实施方式中,尤其在靶细胞为t细胞和/或nk细胞时,促结合多肽可以是麻疹病毒h多肽的细胞质结构域缺失变体且融合多肽可以是麻疹病毒f多肽的细胞质结构域缺失变体。在一些实施方式中,促融合多肽可包括表达为一个多肽的多个元件。在一些实施方式中,结合多肽及促融合多肽可翻译自同一转录物且翻译自不同核糖体结合位点,或可在翻译后使用肽切割信号或核糖体跳过序列来切割多肽,如本文其他处所公开,以产生结合多肽及促融合多肽。在一些实施方式中,结合多肽及促融合多肽自分离的核糖体结合位点的翻译产生比结合多肽更高量的融合多肽。在一些实施方式中,促融合多肽与结合多肽的比率为至少2:1、至少3:1、至少4:1、至少5:1、至少6:1、至少7:1,或至少8:1。在一些实施方式中,第一反式活化子可调节能够结合并活化例如t细胞的靶细胞的活化元件的表达。在这些实施方式中,活化元件可包括:a.)能够结合并活化cd3的膜结合多肽;和/或b.)能够结合并活化cd28的膜结合多肽。在一些实施方式中,能够结合并活化cd28的膜结合多肽为cd80、cd86或其功能片段。在一些实施方式中,复制缺陷型重组逆转录病毒颗粒可包括逆转录病毒膜上的活化元件及核衣壳内的逆转录病毒rna,从而制得复制缺陷型重组逆转录病毒颗粒。在一些实施方式中,第二反式活化子可调节rna的表达,所述rna自5'至3'包括:5'长末端重复或其活性截短片段;编码逆转录病毒顺式作用rna包装元件的核酸序列;编码第一靶标多肽及任选地第二靶标多肽(作为非限制性实例,一或两种经改造信号传导多肽)的核酸序列;在靶细胞中有活性的启动子;及3'长末端重复或其活性截短片段。在一些实施方式中,rna可包括cppt/cts元件。在一些实施方式中,rna可包括引物结合位点。在一些实施方式中,逆转录病毒顺式作用rna包装元件可以是hivpsi。在一些实施方式中,逆转录病毒顺式作用rna包装元件可以是rev反应元件。在本文公开的任何实施方式中,rna上的逆转录病毒组分(包括rre及psi)可位于任何位置,如熟练技术人员将理解。在说明性实施方式中,经改造信号传导多肽为本文所公开的一种或多种经改造信号传导多肽。在一些实施方式中,经改造信号传导多肽可包括第一淋巴增殖性元件。适合的淋巴增殖性元件公开于本文的其他部分中。在一些说明性实施方式中,淋巴增殖性元件为融合至识别结构域的il-7受体突变体,例如etag。在一些实施方式中,可包装rna基因组可进一步包括编码第二经改造多肽的核酸序列,包括编码本文所提供的任何car实施方式的嵌合抗原受体。举例而言,第二经改造多肽可包括第一抗原特异性靶向区、第一跨膜结构域,及第一胞内活化结构域。抗原特异性靶向区、跨膜结构域及胞内活化结构域的实例公开于本文其他处。在靶细胞为t细胞的一些实施方式中,在靶细胞中有活性的启动子在t细胞中具有活性,如本文其他处所公开。在一些实施方式中,可包装rna基因组可进一步包括核糖开关,如本文其它部分所论述。在一些实施方式中,编码经改造信号传导多肽的核酸序列可呈反向定向。在另一些实施方式中,可包装rna基因组可进一步包括核糖开关,且任选地该核糖开关可呈反向定向。在本文所公开的任何实施方式中,包括任何元件的多核苷酸可包括引物结合位点。在说明性实施方式中,转录阻断剂或polya序列可置放于基因附近以防止或减少未经调节的转录。在本文所公开的任何实施方式中,编码vpx的核酸序列可位于第二转录单元上或任选地第三转录单元上,或位于可操作地连接至第一可诱导启动子的额外转录单元上。在用于制备复制缺陷型重组逆转录病毒颗粒方面的包装系统或方法的一些实施方式中,编码的rna可以包括内含子,其可例如转录自用于表达靶标多肽的相同启动子。在某些说明性实施方式中,此类内含子可编码1种、2种、3种,或4种mirna。在用于制备复制缺陷型重组逆转录病毒颗粒方面的包装系统或方法的这些及其他实施方式中,可包装rna基因组大小为11,000kb或更小,且在一些实例中为10,000kb或更小。在一些实施方式中,第一反式活化子可影响一种或多种非毒性多肽的表达。在一些实施方式中,第二反式活化子可影响一种或多种毒性多肽的表达。举例而言,第一反式活化子可诱导除了将转运至包装细胞的细胞膜的多肽之外的逆转录病毒蛋白质rev及vpx的表达,且第二反式活化子可诱导逆转录病毒蛋白质gag、pol、mv(ed)-fδ30以及mv(ed)-hδ18或mv(ed)-hδ24的表达以及慢病毒基因组的表达。在一些实施方式中,第一反式活化子可影响一种或多种毒性多肽的表达和/或第二反式活化子可影响一种或多种非毒性多肽的表达。在另一方面中,本文提供哺乳动物包装细胞,其包括:a)哺乳动物包装细胞的基因组中的第一转录单元,其包括编码第一反式活化子的核酸序列,其中该第一转录单元可操作地连接至组成型启动子,且其中该反式活化子能够结合第一可诱导启动子且在存在与不存在第一配体的情况下影响可操作地连接至其上的核酸序列的表达,且其中该第一反式活化子能够结合该第一配体;b)哺乳动物包装细胞基因组中的第二转录单元及任选地第三转录单元,其包括编码逆转录病毒rev蛋白质的核酸序列及编码第二反式活化子的核酸序列,该第二反式活化子能够结合第二可诱导启动子且在存在与不存在第二配体的情况下影响可操作地连接至其上的核酸序列的表达,其中该第二反式活化子能够结合该第二配体,且其中该第二转录单元及任选地存的第三转录单元可操作地连接至该第一可诱导启动子;c)哺乳动物包装细胞基因组中的第四转录单元及任选地第五转录单元,其包括编码逆转录病毒gag多肽及逆转录病毒pol多肽的核酸序列,以及能够结合并促进靶细胞膜与逆转录病毒膜融合的结合多肽和融合多肽,其中该第四转录单元及任选地第五转录单元可操作地连接至该第二可诱导启动子;以及d)哺乳动物包装细胞基因组中的第六转录单元,其自5'至3'包括5'ltr或其活性截短片段、编码逆转录病毒顺式作用rna包装元件的核酸序列、cppt/cts元件、编码经改造信号传导多肽的核酸序列的反向互补、内含子、在靶细胞中有活性的启动子,以及3'ltr或其活性截短片段,其中该第六转录单元可操作地连接至该第二可诱导启动子。在另一方面中,本文提供用于制备复制缺陷型重组逆转录病毒颗粒的方法,其包括:1.)培养包装细胞群以累积第一反式活化子,其中这些包装细胞包括:a)哺乳动物包装细胞基因组中的第一转录单元,其包括编码第一反式活化子的核酸序列,其中该第一转录单元可操作地连接至组成型启动子,且其中该反式活化子能够结合第一可诱导启动子且在存在与不存在第一配体的情况下影响可操作地连接至其上的核酸序列的表达,且其中该第一反式活化子能够结合该第一配体;b)哺乳动物包装细胞基因组中的第二转录单元及任选地第三转录单元,其包括编码逆转录病毒rev蛋白质的核酸序列及编码第二反式活化子的核酸序列,该第二反式活化子能够结合第二可诱导启动子且在存在与不存在第二配体的情况下影响可操作地连接至其上的核酸序列的表达,其中该第二反式活化子能够结合该第二配体,且其中该第二转录单元及任选地第三转录单元可操作地连接至该第一可诱导启动子;c)哺乳动物包装细胞基因组中的第四转录单元及任选地第五转录单元,其包括编码逆转录病毒gag多肽及逆转录病毒pol多肽的核酸序列,以及能够结合并促进逆转录病毒膜与靶细胞膜融合的结合多肽和融合多肽,其中该第四转录单元及任选地第五转录单元可操作地连接至该第二可诱导型启动子;以及d)哺乳动物包装细胞基因组中的第六转录单元,其自5'至3'包括5'ltr或其活性截短片段、引物结合位点(pbs)、编码逆转录病毒顺式作用rna包装元件的核酸序列、cppt/cts元件、编码经改造信号传导多肽的核酸序列的反向互补、内含子、在靶细胞中具有活性的靶细胞启动子,以及3'ltr或其活性截短片段,其中该第五转录单元可操作地连接至该第二可诱导启动子;及2)在该第一配体的存在下孵育包括该第一反式活化子的包装细胞群以累积该第二反式活化子及逆转录病毒rev蛋白质;及3)在该第二配体的存在下孵育包括该第二反式活化子及逆转录病毒rev蛋白质的包装细胞群,从而诱导逆转录病毒gag多肽、逆转录病毒pol多肽、结合多肽、融合多肽及逆转录病毒rna(自5'至3'包括5'ltr或其活化片段的逆转录病毒rna、pbs、逆转录病毒顺式作用rna包装元件、编码经改造信号传导多肽的核酸序列的反向互补、靶细胞启动子,以及3'ltr或其活性截短片段)的表达,其中复制缺陷型重组逆转录病毒颗粒由包装细胞形成并释放,且其中该复制缺陷型重组逆转录病毒颗粒包括逆转录病毒膜上的结合多肽和/或融合多肽以及核衣壳内的逆转录病毒rna,而制得复制缺陷型重组逆转录病毒颗粒。在本文所提供的一个方面中,逆转录病毒包装系统可包括哺乳动物细胞,其包括:1.)第一反式活化子,其由组成型启动子表达且能够结合第一配体及第一可诱导启动子来在存在与不存在第一配体的情况下影响可操作地连接至其上的核酸序列的表达;2)第二反式活化子,其能够结合第二配体及第二可诱导启动子,且在存在与不存在第二配体的情况下影响可操作地连接至其上的核酸序列的表达;及3)逆转录病毒颗粒的可包装rna基因组,其中该第一反式活化子调节该第二反式活化子、hivrev、il7gpidaf及活化元件的表达,且其中该第二反式活化子调节gag多肽、pol多肽、逆转录病毒顺式作用rna包装元件,及一种或多种包膜多肽的表达。在说明性实施方式中,第一反式活化子可以是融合至p65活化结构域的frb结构域及融合至zfhd1dna结合结构域的一个或多个fkbp结构域,第一配体可以是雷帕霉素,且第一可诱导启动子可以是一个或多个zfhd1结合位点。在说明性实施方式中,第二反式活化子可以是rtta蛋白质,第二配体可以是四环素或多西环素,且第二可诱导启动子可以是tre启动子或双向tre启动子。在说明性实施方式中,逆转录病毒顺式作用rna包装元件可以是hivpsi。在说明性实施方式中,一种或多种包膜蛋白质包括麻疹病毒的f多肽及h多肽的细胞质结构域缺失变体。在说明性实施方式中,转录阻断剂或polya序列可置放于基因附近以防止或减少未经调节的转录。在一些实施方式中,可使用具有核糖开关的雷帕霉素-多西环素可诱导慢病毒基因组(seqidno:83)。在一些实施方式中,可使用雷帕霉素-多西环素可诱导gagpolenv(seqidno:84)。在一些实施方式中,可使用雷帕霉素可诱导tet活化子(seqidno:85)。在一些实施方式中,可使用雷帕霉素诱导子可诱导revsrcvpx(seqidno:86)。本公开的一些方面包括细胞或是细胞,在说明性实例中为用作包装细胞以制备复制缺陷型重组逆转录病毒颗粒的哺乳动物细胞,例如用于转导t细胞和/或nk细胞的慢病毒颗粒。可选择多种细胞的任何一种用于在体外产生病毒或病毒颗粒,例如根据本发明重定向的重组逆转录病毒颗粒。通常使用真核细胞,尤其哺乳动物细胞,包括人类细胞,猿类细胞,犬类细胞,猫类细胞,马类细胞及啮齿动物细胞。在说明性实例中,细胞为人类细胞。在其他说明性实施方式中,细胞无限地增殖,且因此永生。可有利地用于本发明的细胞的实例包括nih3t3细胞、cos细胞、madin-darby犬肾细胞、人类胚胎293t细胞和任何衍生自这些细胞的细胞,例如gpnlslacz细胞,其衍生自293t细胞。可使用高度可转染细胞,例如人胚胎肾293t细胞。“高度可转染”是指细胞的至少约50%、更优选至少约70%、最优选至少约80%可表达引入的dna的基因。适合的哺乳动物细胞包括原代细胞及固定化细胞系。适合的哺乳动物细胞系包括人类细胞系、非人类灵长类细胞系、啮齿动物(例如小鼠、大鼠)细胞系,及类似者。适合的哺乳动物细胞系包括但不限于hela细胞(例如美国模式培养物保藏所(atcc)第ccl-2号)、cho细胞(例如atcc第crl9618号、第ccl61号、第crl9096号)、293细胞(例如atcc第crl-1573号)、vero细胞、nih3t3细胞(例如atcc第crl-1658号)、huh-7细胞、bhk细胞(例如atcc第ccllo号)、pc12细胞(atcc第crl1721号)、cos细胞、cos-7细胞(atcc第crl1651号)、ratl细胞、小鼠l细胞(atcc第ccli.3号)、人类胚胎肾(hek)细胞(atcc第crl1573号)、hlhepg2细胞、hut-78、jurkat、hl-60、nk细胞系(例如nkl、nk92及yts),及类似者。在本文所公开的任何实施方式中,制备复制缺陷型重组逆转录病毒颗粒的方法可包括使哺乳动物包装细胞生长至50%、60%、70%、80%、90%或95%汇合,或生长至25%、30%、40%、50%、60%、70%、80%、90%或95%的峰值细胞密度,且接着分开或稀释细胞。在一些实施方式中,可使用搅拌孔反应器来使细胞生长。在一些实施方式中,可使用本领域熟练技术人员将理解的方法将细胞以至少约1:2、1:3、1:4、1:5、1:6、1:7、1:8、1:9、1:10、1:12、1:15,或1:20分开。在一些实施方式中,可将细胞稀释至25%、30%、40%、50%、60%、70%、80%、90%或95%的峰值细胞密度。在一些实施方式中,在分开或稀释细胞之后,可在添加第一配体之前使细胞生长持续1小时、2小时、3小时、4小时、5小时、6小时、7小时、8小时、10小时,或16小时,或1天、2天、3天、4天、5天、6天,或7天。在一些实施方式中,在存在第一配体的情况下使细胞生长持续1天、2天、3天、4天、5天、6天、7天、8天、9天、10天、11天、12天、13天、14天、21天,或28天,在说明性实施方式中,该第一配体可以是雷帕霉素或雷帕类似物。在一些实施方式中,可添加第二配体且使细胞生长持续1天、2天、3天、4天、5天、6天、7天、8天、9天、10天、11天、12天、13天、14天、21天,或28天,在说明性实施方式中,该第二配体可以是四环素或多西环素。培养条件将取决于所用的细胞及配体,且方法为本领域已知的。用于培养且诱导hek293s细胞的条件的特定实例展示于实施例8中。如本文所公开,复制缺陷型重组逆转录病毒颗粒为用于基因递送的常用工具(miller,nature(1992)357:455-460)。复制缺陷型重组逆转录病毒颗粒将未经排列的核酸序列递送至广泛范围的啮齿动物、灵长类动物及人类体细胞中的能力使得复制缺陷型重组逆转录病毒颗粒较适用于将基因转移至细胞。在一些实施方式中,复制缺陷型重组逆转录病毒颗粒可衍生自α逆转录病毒属,β逆转录病毒属,γ逆转录病毒属,δ逆转录病毒属,ε逆转录病毒属,慢病毒属或泡沫病毒属。存在许多种适用于本文中所公开方法的逆转录病毒。举例而言,可使用鼠白血病病毒(mlv)、人类免疫缺陷病毒(hiv)、马感染性贫血病毒(eiav)、小鼠乳腺肿瘤病毒(mmtv)、劳斯肉瘤病毒(rsv)、富士纳肉瘤病毒(fusv)、莫洛尼鼠白血病病毒(mo-mlv)、fbr鼠骨肉瘤病毒(fbrmsv)、莫洛尼鼠肉瘤病毒(mo-msv)、艾贝森鼠白血病病毒(abelsonmurineleukemiavirus)(a-mlv)、禽骨髓细胞病变病毒-29(mc29)及禽红细胞增生病毒(aev)。逆转录病毒的详细清单可见于coffin等人的(“retroviruses”,1977,coldspringharborlaboratorypress:编者jmcoffin,smhughes,hevarmus,第758至763页)。可在本领域中找到关于一些逆转录病毒的基因组结构的细节。举例而言,可自ncbigenbank(即genomeaccession第af033819号)中找到关于hiv的细节。在说明性实施方式中,复制缺陷型重组逆转录病毒颗粒可衍生自慢病毒属。在一些实施方式中,复制缺陷型重组逆转录病毒颗粒可衍生hiv、siv或fiv。在另一些说明性实施方式中,复制缺陷型重组逆转录病毒颗粒可衍生自慢病毒属中的人类免疫缺陷病毒(hiv)。慢病毒为复杂的逆转录病毒,除常见的逆转录病毒基因gag、pol及env外,也含有具有调节或结构功能的其他基因。较高复杂性使得慢病毒能够在潜伏感染过程中调节其生命周期。典型的慢病毒是人类免疫缺陷病毒(hiv),aids病原体。在体内,hiv可感染极少分裂的终末分化的细胞,例如淋巴细胞及巨噬细胞。在说明性实施方式中,本文提供的复制缺陷型重组逆转录病毒颗粒含有vpx多肽。vpx多肽可在其基因组中整合vpx编码核酸后在包装细胞系中表达,例如作为并入至逆转录病毒膜中的细胞膜结合蛋白质(durand等人,j.virol(2013)87:234-242)。可通过病毒蛋白酶的处理顺序构建逆转录病毒膜结合vpx,使得游离vpx一旦并入病毒颗粒中即被释放。具有此功能的vpx融合物的这种实例是src-flag-vpx,其包括c-src的头11个氨基酸的膜靶向结构域(mgsskskpkdp)(seqidno:227),随后为病毒蛋白酶切割结构域karvlaea(seqidno:228),随后为flag标签化的vpx。不受理论限制,vpx多肽通过降解限制因子samhd1来刺激逆转录过程的效率,而有助于静息细胞转导。因此,认为在本文提供的方法中,其中vpx存在于用以转导t细胞和/或nk细胞的复制缺陷型重组逆转录病毒颗粒中,vpx在通过含有vpx的复制缺陷型重组逆转录病毒颗粒转导细胞之后经释放至静息t细胞或静息nk细胞的细胞质中。vpx接着降解samhd1,其使得游离dntp增加,这样进而刺激逆转录病毒基因组的逆转录。逆转录病毒基因组大小在本文提供的方法及组合物中,重组逆转录病毒基因组(在非限制性说明性实例中为慢病毒基因组)对可包装至病毒颗粒中的多核苷酸数量具有限制。在本文提供的一些实施方式中,由多核苷酸编码区编码的多肽可以是保留功能活性的截短或其它缺失,使得多核苷酸编码区由比野生型多肽的多核苷酸编码区更少的核苷酸编码。在一些实施方式中,由多核苷酸编码区编码的多肽可以是可由一个启动子表达的融合多肽。在一些实施方式中,融合多肽可具有切割信号以由一个融合多肽及一个启动子产生两个或更多个功能性多肽。此外,在初始体外转导之后不需要的一些功能不包括于逆转录病毒基因组中,而经由包装细胞膜存在于复制缺陷型重组逆转录病毒颗粒表面上。这种不同策略在本文用于使包装于复制缺陷型重组逆转录病毒颗粒内的功能性元件最大化。在一些实施方式中,待包装的重组逆转录病毒基因组可在范围下限1,000个、2,000个、3,000个、4,000个、5,000个、6,000个、7,000个及8,000个核苷酸与范围上限2,000个、3,000个、4,000个、5,000个、6,000个、7,000个、8,000个、9,000个、10,000个及11,000个核苷酸之间。待包装的逆转录病毒基因组包括编码如本文所详细公开的第一及第二经改造信号传导多肽的一个或多个多核苷酸区。在一些实施方式中,待包装的逆转录病毒基因组可少于5,000个、6,000个、7,000个、8,000个、9,000个、10,000个,或11,000个核苷酸。本文其他处所讨论的可包装功能包括用于逆转录病毒装配及包装所需的逆转录病毒序列,例如逆转录病毒rev、gag及pol编码区,以及5'ltr及3'ltr或其活性截短片段,编码逆转录病毒顺式作用rna包装元件的核酸序列及cppt/cts元件。此外,在说明性实施方式中,本文中的复制缺陷型重组逆转录病毒颗粒可包括以下中任何一这种或多种或全部(在一些实施方式中这些逆转录病毒功能区呈相反方向):编码第一及第二工程信号传导多肽的一个或多个多核苷酸区,其中的至少一种包括至少一种淋巴组织增殖元件且可进一步包括astr;可包括嵌合抗原受体的第二经改造信号传导多肽;通常调节第一和/或第二经改造信号传导多肽表达的控制元件,例如核糖开关;识别结构域、内含子、在靶细胞中具活性启动子,例如t细胞,2a切割信号和/或ires。重组逆转录病毒颗粒重组逆转录病毒颗粒公开于本文提供的方法及组合物中,例如用以转导t细胞和/或nk细胞以制备遗传修饰的t细胞和/或nk细胞。重组逆转录病毒颗粒自身为本发明的方面。通常,包括于本文提供的方面中的重组逆转录病毒颗粒为复制缺陷型的,意为重组逆转录病毒颗粒在其离开包装细胞时即无法复制。在说明性实施方式中,重组逆转录病毒颗粒是慢病毒颗粒。在一些方面中,本文提供一种重组逆转录病毒颗粒,其包括(i)假型化元件,其能够结合t细胞和/或nk细胞并促进重组逆转录病毒颗粒的膜融合;(ii)多核苷酸,其具有可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码具有嵌合抗原受体(包括抗原特异性靶向区、跨膜结构域及胞内活化结构域)的第一经改造信号传导多肽,以及包含至少一种淋巴组织增殖性元件的第二经改造信号传导多肽;其中该第一经改造信号传导多肽和/或该第二经改造信号传导多肽的表达由体内控制元件调节;及(iii)在其表面上的活化元件,其中该活化元件能够结合至t细胞和/或nk细胞,且不由重组逆转录病毒颗粒中的多核苷酸编码。在一些实施方式中,在t细胞和/或nk细胞中具活性启动子在包装细胞系中不具有活性,或仅以可诱导的方式在包装细胞系中具有活性。在本文公开的任何实施方式中,第一及第二经改造信号传导多肽中的一种可具有嵌合抗原受体且另一经改造信号传导多肽可具有至少一种淋巴增殖性元件。贯穿本公开提供包括于复制缺陷型重组逆转录病毒颗粒中的多种元件及元件组合,例如假型化元件、活化元件及膜结合细胞因子,以及包括于复制缺陷型重组逆转录病毒颗粒基因组中的核酸序列,例如但不限于编码car的核酸;编码淋巴增殖性元件的核酸;编码控制元件(例如核糖开关)的核酸;启动子,尤其在t细胞中具有组成型活性或可诱导的启动子;及编码抑制性rna分子的核酸。此外,本文提供的多种方面(例如制备重组逆转录病毒颗粒的方法、执行过继细胞治疗的方法,以及转导t细胞的方法)产生和/或包括复制缺陷型重组逆转录病毒颗粒。此类方法中产生和/或包括的复制缺陷型重组逆转录病毒自身形成作为本发明的独立方面的复制缺陷型重组逆转录病毒颗粒组合物,这种组合物可呈分离形式。此类组合物可呈干燥(例如冻干)形式,或者可呈本领域已知的适合溶液或介质形式以用于储存且使用逆转录病毒颗粒。因此,作为非限制性实例,在另一方面中,本文提供一种在其基因组中具有多核苷酸的复制缺陷型重组逆转录病毒颗粒,该多核苷酸具有可操作地连接至t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,在一些情况下,该核酸序列包括编码针对一种或多种rna靶标的一种或多种(例如两种或更多种)抑制性rna分子的第一核酸序列及编码嵌合抗原受体或car的第二核酸序列,如本文所描述。在另一些实施方式中,存在编码本文先前所描述的不为抑制性rna分子的至少一种淋巴增殖性元件的第三核酸序列。在某些实施方式中,多核苷酸包括如本文所提及的一个或多个核糖开关,这种核糖开关可操作地连接至第一核酸序列、第二核酸序列和/或第三核酸序列(若存在)。在此构建体中,一种或多种抑制性rna、car和/或不为抑制性rna的一种或多种淋巴增殖性元件的表达由核糖开关控制。在一些实施方式中,两种至十种抑制性rna分子由第一核酸序列编码。在其他实施方式中,两种至六种抑制性rna分子由第一核酸序列编码。在说明性实施方式中,4种抑制性rna分子由第一核酸序列编码。在一些实施方式中,第一核酸序列编码一种或多种抑制性rna分子且位于内含子内。在某些实施方式中,内含子包括启动子的全部或部分。启动子可以是poli、polii,或poliii启动子。在一些说明性实施方式中,启动子为polii启动子。在一些实施方式中,内含子邻近在t细胞和/或nk细胞中具活性启动子且位于该启动子的下游。在一些实施方式中,内含子为ef1-α内含子a。本文的重组逆转录病毒颗粒实施方式包括其中逆转录病毒颗粒包含基因组的实施方式,该基因组包括编码一种或多种抑制性rna分子的一种或多种核酸。编码可包括于逆转录病毒颗粒的基因组中的抑制性rna分子的此类核酸的多种替代实施方式(包括此类核酸与编码除抑制性rna分子之外的car或淋巴增殖性元件的其他核酸的组合)包括于例如本文提供的抑制性rna部分以及包括于组合此等实施方式的多种其他段落中。此外,此类复制缺陷型重组逆转录病毒的多种替代物可通过在本文中所公开的包装细胞系方面内公开的例示性核酸识别。本领域技术人员将认识到,包括编码一种或多种(例如两种或更多种)抑制性rna分子基因组的重组逆转录病毒颗粒这部分的公开内容可与本文其他部分所提供的编码抑制性rna分子这类核酸的多种替代物组合。此外,本领域技术人员将认识到,编码一种或多种抑制性rna分子的这类核酸可与本文所提供的多种其他功能性核酸元件组合,如例如本文中侧重于抑制性rna分子及编码这种分子的核酸的部分所公开。此外,在本文其他部分中提供的特异性抑制性rna分子的多种实施方式可用于本发明的重组逆转录病毒颗粒方面中。例如慢病毒载体的重组逆转录病毒载体的必需要素为是本领域中已知的。此要素包括于包装细胞系部分中,及在实例部分中所提供的用于制备重组逆转录病毒颗粒的细节中。举例而言,慢病毒颗粒通常包括包装元件rev、gag及pol,其可经由一种或多种包装质粒递送至包装细胞系;假型化元件,本文提供的多种实例,其可经由假型化质粒递送至包装细胞系;及基因组,其由经由转移质粒递送至宿主细胞的多核苷酸产生。此多核苷酸通常包括病毒ltr及psi包装信号。5'ltr可以是与异源启动子融合的嵌合5'ltr,其包括不依赖于tat反式活化的5'ltr。转移质粒可自失活,例如通过移除3'ltr的u3区。在一些非限制性实施方式中,例如src-flag-vpx的vpx包装于逆转录病毒颗粒内。不受理论的限制,在t细胞转导时,vpx进入细胞的细胞溶质且促进samhd1降解,使得可用于逆转录的胞质dntp增加。包括于本发明多种方面中的逆转录病毒颗粒(例如慢病毒颗粒)在说明性实施方式中为复制缺陷型,尤其出于对于包括将用此类逆转录病毒颗粒转导的细胞引入受试者的实施方式的安全原因。当复制缺陷型逆转录病毒颗粒用于转导细胞时,逆转录病毒颗粒并非由转导的细胞产生。对逆转录病毒基因组修饰是本领域中已知的,以确保包括该基因组的逆转录病毒颗粒是复制缺陷型的。然而,应理解,在一些实施方式中,针对本文所提供的任何方面中,可使用复制胜任型重组逆转录病毒颗粒。本领域技术人员将认识到,可使用不同类型的载体(例如表达载体)将本文所论述的功能元件递送至包装细胞和/或至t细胞。本发明的说明性方面利用逆转录病毒载体,及(在一些特定说明性实施方式中)慢病毒载体。另一些适合的表达载体可用于实现本文的某些实施方式。此类表达载体包括但不限于病毒载体(例如基于以下的病毒载体:痘苗病毒;脊髓灰质炎病毒;腺病毒(参见例如li等人,investopthalmolvissci35:25432549,1994;borras等人,genether6:515524,1999;li及davidson,pnas92:77007704,1995;sakamoto等人,hgenether5:10881097,1999;wo94/12649,wo93/03769;wo93/19191;wo94/28938;wo95/11984andwo95/00655);腺相关病毒(参见例如ali等人,humgenether9:8186,1998,flannery等人,pnas94:69166921,1997;bennett等人,investopthalmolvissci38:28572863,1997;jomary等人,genether4:683690,1997,rolling等人,humgenether10:641648,1999;ali等人,hummolgenet5:591594,1996;wo93/09239中的srivastava,samulski等人,j.vi(1989)63:3822-3828;mendelson等人,virol.(1988)166:154-165;及flotte等人,pnas(1993)90:10613-10617);sv40;单纯疱疹病毒;或逆转录病毒载体(例如,小鼠白血病病毒脾坏死病毒及衍生自例如罗斯肉瘤病毒(roussarcomavirus)、哈维肉瘤病毒(harveysarcomavirus)、禽白血病病毒、人类免疫缺陷病毒、骨髓增生性肉瘤病毒及乳腺肿瘤病毒的逆转录病毒的载体),例如γ逆转录病毒;或人类免疫缺陷病毒(参见例如miyoshi等人,pnas94:1031923,1997;takahashi等人,jvirol73:78127816,1999);及类似者。在说明性实施方式中,逆转录病毒颗粒为慢病毒颗粒。此类逆转录病毒颗粒通常包括位于病毒包膜内的衣壳内的逆转录病毒基因组。在一些实施方式中,利用含dna的病毒颗粒代替重组逆转录病毒颗粒。此类病毒颗粒可以是腺病毒、腺相关病毒、疱疹病毒、巨细胞病毒、痘病毒、牛痘病毒、流感病毒、水泡性口炎病毒(vsv),或辛德毕斯病毒(sindbisvirus)。本领域技术人员将理解如何修改本文公开的用于不同病毒及逆转录病毒或逆转录病毒颗粒的方法。在使用包括dna基因组的病毒颗粒时,本领域技术人员将理解,此等基因组中可包括功能单元以诱导将病毒颗粒的全部或部分dna基因组整合至用此类病毒转导t细胞的基因组中。在一些实施方式中,hivrre及编码hivrev的多核苷酸区可由n端rgg盒rna结合基序及编码icp27的多核苷酸区替代。在一些实施方式中,编码hivrev的多核苷酸区可由编码腺病毒e1b55-kda及e4orf6的一个或多个多核苷酸区替代。在一个方面中,本文提供含有复制缺陷型重组逆转录病毒颗粒及使用其治疗受试者肿瘤生长说明书的商用容器,其中该复制缺陷型重组逆转录病毒颗粒在其基因组中包含多核苷酸,该多核苷酸包含可操作地连接在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中该一个或多个核酸序列中的第一核酸序列编码针对一个或多个rna靶标的两种或更多种抑制性rna分子且该一个或多个核酸序列中的第二核酸序列编码包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域的嵌合抗原受体(car)。含有重组逆转录病毒颗粒的容器可以是用于储存重组逆转录病毒颗粒的管、小瓶,孔板,或其他容器。试剂盒可包括两个或更多个容器,其中第二或其他容器可包括例如用于转导t细胞和/或nk细胞的溶液或介质,和/或第二或其他容器可包括ph调节药剂。任何这种容器可具有工业强度及等级。包括试剂盒及编码抑制性rna分子的核酸的此类方面中的复制缺陷型重组逆转录病毒颗粒可以是本文所提供的此类复制缺陷型重组逆转录病毒颗粒的任何实施方式,其包括本文所提供的任何抑制性rna实施方式。遗传修饰的t细胞及nk细胞在本文的方法及组合物的实施方式中,产生本身为本发明的单独方面的遗传修饰淋巴细胞。此类遗传修饰的淋巴细胞可以是经转导的淋巴细胞。在一些实施方式中,遗传修饰的淋巴细胞是已遗传修饰以表达包含至少一种淋巴增殖性元件的第一经改造信号传导多肽和/或包含嵌合抗原受体的第二经改造信号传导多肽的淋巴细胞,例如t细胞或nk细胞,该嵌合抗原受体包括抗原特异性靶向区(astr),跨膜结构域及胞内活化结构域。本公开的遗传修饰的淋巴细胞具有已通过重组dna方法引入至淋巴细胞中的异源核酸序列。举例而言,在用于转导本文所提供的淋巴细胞的方法期间将说明性实施方式中的异源序列插入至淋巴细胞中。异源核酸存在于淋巴细胞内,且在一些实施方式中经整合或未整合至遗传修饰的淋巴细胞的基因组中。在说明性实施方式中,将异源核酸整合至遗传修饰的淋巴细胞的基因组中。在说明性实施方式中,使用利用重组逆转录病毒颗粒的本文所提供的用于转导淋巴细胞的方法来产生此类淋巴细胞。此类重组逆转录病毒颗粒可包括编码嵌合抗原受体的多核苷酸,该嵌合抗原受体通常包括至少一个抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域。在本发明其他部分中,本文提供复制缺陷型重组逆转录病毒颗粒及在复制缺陷型逆转录病毒颗粒的基因组中编码的多核苷酸的多种实施方式,该复制缺陷型逆转录病毒颗粒可用以产生本身形成本公开的另一方面的遗传修饰的淋巴细胞。本发明遗传修饰的淋巴细胞可在体外分离。举例而言,此类淋巴细胞可发现于如本文所提供的用于离体转导的介质及其他溶液中。淋巴细胞可以未遗传修饰的形式存在于以本文所提供的方法从受试者收集的血液中,接着在转导方法期间遗传修饰。遗传修饰的淋巴细胞可在其遗传修饰之后在其经引入或重新引入至受试者之后发现于受试者内部。遗传修饰的淋巴细胞可以是静息t细胞或静息nk细胞,或遗传修饰的t细胞或nk细胞可以主动分裂,尤其在其表达在如本文所公开的转导之后经插入至t细胞或nk细胞中的核酸中所提供的功能元件中的一些后。在一个方面中,本文提供经转导和/或遗传修饰的t细胞或nk细胞,其包含重组多核苷酸,该重组多核苷酸在其基因组中包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,该转录单元表达本发明的任何方面及实施方式中提供的功能元件中的一种或多种。举例而言,一个或多个转录元可表达car,其可包括本文所提供的car元件中任一种,例如astr(作为非限制性实例,为mbr-astr)、跨膜结构域及胞内信号传导结构域,且可以进一步包括作为非限制性实例的调节结构域。此外,本文提供在经转导和/或遗传修饰的t细胞或nk细胞内表达的一个或多个功能元件、淋巴增殖性元件中的一种或多种,例如组成型活化il-7受体突变体或不为抑制性rna分子(例如mirna或shrna)的其他淋巴增殖性元件、识别结构域和/或消除结构域。在一个方面中,本文提供遗传修饰的t细胞或nk细胞,其包含:a.针对一种或多种rna靶标的一种或多种(例如两种或更多种)抑制性rna分子;及b.嵌合抗原受体(car),其包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域,和/或编码针对一种或多种rna靶标的抑制性rna分子及car的核酸,其中该一种(例如两种)或更多种抑制性rna分子及car或编码其的核酸编码由t细胞和/或nk细胞遗传修饰的核酸序列编码或为t细胞和/或nk细胞遗传修饰的核酸序列。遗传修饰的t细胞或nk细胞可以是遗传修饰的t细胞群和/或nk细胞群,其包括针对一种或多种rna靶标的一种(例如两种)或更多种抑制性rna分子;及car。在本方面的t细胞或nk细胞包含一种或多种(例如两种或更多种)抑制性rna分子及car或编码其核酸的方面的一些实施方式中,针对可包括作为car一部分或可与淋巴增殖性元件一起表达或用以控制淋巴增殖性元件的元件,可包括本文所提供的任何具体实施方式。在本方面的t细胞或nk细胞包含一种或多种(例如两种或更多种)抑制性rna分子及car或编码其核酸的方面的一些实施方式中,car为微环境限制的生物(mrb)-car和/或遗传修饰的t细胞或nk细胞可进一步包括不为抑制性rna分子的至少一种淋巴增殖性元件和/或编码淋巴增殖性元件的核酸。在此类实施方式中,淋巴增殖性元件由对t细胞和/或nk细胞的遗传修饰的核酸序列编码。在此类实施方式中,可使用和/或编码本文所公开的任何淋巴增殖性元件。举例而言,至少一种淋巴增殖性元件可以是组成型活化的il-7受体。在本方面t细胞或nk细胞包含一种或多种(例如两种或更多种)抑制性rna分子及car或编码其核酸的方面的一些实施方式中,该抑制性rna分子为mirna或shrna的前体。在此方面的一些实施方式中,该一种(例如两种)或更多种抑制性rna分子为多顺反子的。在此方面的一些实施方式中,该一种(例如两种)或更多种抑制性rna分子针对相同的或(在说明性实施方式中)不同的rna靶标。在此方面的一些实施方式中,该一种(例如两种)或更多种抑制性rna分子中的大部分或全部降低内源性tcr的表达。在本方面t细胞或nk细胞包含一种或多种(例如两种或更多种)抑制性rna分子及car或编码其核酸的方面的一些实施方式中,该rna靶标为自选自以下的基因转录的mirna:pd-1、ctla4、tcrα、tcrβ、cd3ζ、socs、smad2、mir-155靶标、ifnγ、ccbl、trail2、pp2a,及abcg1。在此方面的一些实施方式中,该一种(例如两种)或更多种抑制性rna分子中至少一种为mir-155。在本方面t细胞或nk细胞包含一种或多种(例如两种或更多种)抑制性rna分子及car或编码其核酸的方面的一些实施方式中,car的astr为mrbastr和/或car的astr结合至与肿瘤相关的抗原。此外,在以上方面的一些实施方式中,第一核酸序列可操作地连接至核糖开关,该核糖开关例如能够结合核苷类似物,且在说明性实施方式中为抗病毒药物,例如阿昔洛韦。在本文公开的方法及组合物中,经改造信号传导多肽的表达由控制元件调节,且在一些实施方式中,该控制元件为包含核糖开关的多核苷酸。在某些实施方式中,核糖开关能够结合核苷类似物,且当核苷类似物存在时,表达一种或两种经改造信号传导多肽。本文所公开的遗传修饰淋巴细胞也可在其表面上具有多肽,这些多肽在本文所提供的转导方法期间为复制缺陷型重组逆转录病毒颗粒融合的残留物。此类多肽可包括活化元件、假型化元件和/或包括细胞因子的一种或多种融合多肽。在一个方面中,本文提供表达针对一种或多种rna靶标一种或多种(例如两种或更多种)抑制性rna分子及嵌合抗原受体或car的遗传修饰的t细胞和/或nk细胞,如本文所公开。在一些实施方式中,遗传修饰的t细胞和/或nk细胞进一步表达本文所公开的不为抑制性rna分子的至少一种淋巴增殖性元件。在某些实施方式中,遗传修饰的t细胞和/或nk细胞也表达控制一种或多种抑制性rna分子、car和/或不为抑制性rna分子的至少一种淋巴增殖性元件的表达的一种或多种核糖开关。在一些实施方式中,遗传修饰的t细胞和/或nk细胞表达两种至10种抑制性rna分子。在其他实施方式中,遗传修饰的t细胞和/或nk细胞表达两种至六种抑制性rna分子。在说明性实施方式中,遗传修饰的t细胞和/或nk细胞表达四种抑制性rna分子。核酸本发明提供编码本发明的多肽的核酸。在一些实施方式中,核酸将为dna,包括例如重组性表达载体。在一些实施方式中,核酸为rna,例如体外合成的rna。在一些情况下,核酸提供例如在哺乳动物细胞中产生本发明的多肽。在另一些情况下,本发明的核酸提供编码本公开的多肽的核酸的扩增。编码本公开的多肽的核苷酸序列可操作地连接至转录控制元件,例如启动子及增强子等。适合的启动子及增强子元件为本领域中已知的。为了在细菌细胞中表达,适合的启动子包括但不限于laci、lacz、t3、t7、gpt、λp及trc。为了在真核细胞中表达,适合的启动子包括但不限于轻链和/或重链免疫球蛋白基因启动子及增强子元件;巨细胞病毒立即早期启动子;单纯疱疹病毒胸苷激酶启动子;早期及晚期sv40启动子;逆转录病毒的长末端重复中存在的启动子;小鼠金属硫蛋白-i启动子;及本领域中已知的多种组织特异性启动子。适合的可逆启动子(包括可逆诱导型启动子)为本领域中已知的。此类可逆启动子可分离且衍生自许多生物体,例如真核生物及原核生物。对用于第二生物中的衍生自第一生物(例如第一原核生物及第二真核生物、第一真核生物及第二原核生物,等)的可逆启动子的修饰是本领域中已知的。此类可逆启动子,及基于此类可逆启动子但也包含其他控制蛋白质的系统包括但不限于醇调节启动子(例如醇脱氢酶i(alca)基因启动子、对醇反式活化子蛋白质具有反应性的启动子(alcr)等)、四环素调节的启动子(例如包括tetactivators、teton、tetoff等启动子系统)、类固醇调节的启动子(例如大鼠糖皮质激素受体启动子系统、人类雌性激素受体启动子系统、类视黄素启动子系统、甲状腺启动子系统、蜕皮激素启动子系统、米非司酮启动子系统等)、金属调节的启动子(例如金属硫蛋白启动子系统等)、相关病原调节的启动子(例如水杨酸调节的启动子、乙烯调节的启动子、苯并噻二唑调节的启动子等)、温度调节在启动子(例如热休克可诱导启动子(例如hsp-70、hsp-90、大豆热休克启动子等))、光调节的启动子、合成可诱导启动子,及类似。在一些情况下,含有适合的启动子的基因座或构建体或转基因经由对可诱导系统的诱导来进行不可逆转换。用于诱导不可逆转换的适合的系统是本领域中已知的,例如,不可逆转换的诱导可利用cre-lox介导的重组(参见例如fuhrmann-benzakein等人,pnas(2000)28:e99,其公开内容以引用方式并入本文)。本领域中已知的重组酶、内切核酸酶、连接酶、重组位点等任何适合的组合可用于产生不可逆转换的启动子。本文其他处描述的用于执行位点特异性重组的方法、机制及要求用于产生不可逆转换的启动子且为本领域中所熟知的,参见例如grindley等人(2006)annualreviewofbiochemistry,567-605及tropp(2012)molecularbiology(jones&bartlettpublishers,sudbury,ma),其公开内容以引用方式并入本文中。在一些情况下,启动子为cd8细胞特异性启动子、cd4细胞特异性启动子、嗜中性粒细胞特异性启动子或nk特异性启动子。举例而言,可使用cd4基因启动子,参见例如salmon等人(1993)proc.natl.acad.sci.usa90:7739及marodon等人(2003)blood101:3416。作为另一实例,可使用cd8基因启动子。nk细胞特异性表达可通过使用neri(p46)启动子来实现;参见例如eckelhart等人(2011)blood117:1565。在一些实施方式中,例如,为了在酵母细胞中表达,适合的启动子为组成型启动子,例如adh1启动子、pgk1启动子、eno启动子、pyk1启动子及类似者;或为了可调节启动子,例如gali启动子、gal10启动子、adh2启动子、ph05启动子、cup1启动子、gal7启动子、met25启动子、met3启动子、cyc1启动子、his3启动子、adh1启动子、pgk启动子、gapdh启动子、adc1启动子、trp1启动子、ura3启动子、leu2启动子、eno启动子、tp1启动子及aox1(例如用于毕赤酵母中)。对合适载体及启动子的选择是在本领域普通技术人员的水平之内。用于原核宿主细胞中适合的启动子包括但不限于噬菌体t7rna聚合酶启动子、trp启动子、lac操纵子启动子、杂合启动子(例如lac/tac杂合启动子、tac/trc杂合启动子、trp/lac启动子、t7/lac启动子);trc启动子;tac启动子及类似者;arabad启动子;经体内调节的启动子,例如ssag启动子或相关启动子(参见例如美国专利公开案第20040131637号)、pagc启动子(pulkkinen及miller,j.bacterial.,1991:173(1):86-93;alpuche-aranda等人,pnas,1992;89(21):10079-83)、nirb启动子(harborne等人(1992)mal.micro.6:2805-2813),及类似者(参见例如dunstan等人(1999)infect.immun.67:5133-5141;mckelvie等人(2004)vaccine22:3243-3255;及chatfield等人(1992)biotechnol.10:888-892);σ70启动子,例如共有σ70启动子(参见例如genomeaccession第ax798980号、第ax798961号及第ax798183号);固定相启动子,例如dps启动子、spv动子及类似者;衍生自致病岛spi-2的启动子(参见例如wo96/17951);acta启动子(参见例如shetron-rama等人(2002)infect.immun.70:1087-1096);rpsm启动子(参见例如valdivia及falkow(1996).mal.microbial.22:367);tet启动子(参见例如hillen,w.及wissmann,a.(1989)insaenger,w.及heinemann,u.(编),topicsinmolecularandstructuralbiology,protein-nucleicacidinteraction.macmillan,伦敦,英国,第10卷,第143至162页);sp6启动子(参见例如melton等人(1984)nucl.acidsres.12:7035);及类似者。用于例如大肠杆菌的原核生物的适合的强启动子包括但不限于trc、tac、t5、t7及pλ。用于细菌宿主细胞的操纵子的非限制性实例包括乳糖启动子操纵子(当与乳糖接触时,laci抑制蛋白改变构象,从而防止laci抑制蛋白与操纵子结合)、色氨酸启动子操纵子(当与色氨酸复合时,trpr抑制蛋白具有结合操纵子的构象;在不存在色氨酸的情况下,trpr抑制蛋白具有不结合操纵子的构象)及tac启动子操纵子(参见例如deboer等人(1983)proc.natl.acad.sci.美国80:21-25)。编码本公开多肽的核苷酸序列可存在于表达载体和/或克隆载体中。编码两个单独多肽的核苷酸序列可在相同或不同载体中克隆。表达载体可包括选择性标记、复制起点及提供载体的复制和/或维持的其他部件。适合的表达载体包括例如质粒、病毒载体,及类似者。大量适合的载体及启动子为本领域技术人员已知;许多在商业上可用于产生本发明重组构建体。借助于实例提供以下细菌载体:pbs、phagescript、psixl74、pbluescriptsk、pbsks、pnh8a、pnh16a、pnh18a、pnh46a(stratagene,lajolla,ca,usa);ptrc99a、pkk223-3、pkk233-3、pdr540,及prit5(pharmacia,uppsala,sweden)。借助于实例提供以下真核生物载体:pwlneo、psv2cat、pog44、pxrl、psg(stratagene)psvk3、pbpv、pmsg,及psvl(pharmacia)。表达载体通常具有位于启动子序列附近的方便限制位点,以提供编码异源蛋白质核酸序列的插入。可存在在表达宿主中操作的可选择标记。如上所述,在一些实施方式中,编码本发明多肽的核酸在一些实施方式中将为rna,例如体外合成的rna。体外合成rna的方法为此项技术中已知的;可使用任何已知的方法来合成包括编码本发明多肽的核苷酸序列的rna。将rna引入至宿主细胞中的方法为此项技术中已知的。参见例如zhao等人(2010)cancerres.15:9053。将包括编码本发明多肽的核苷酸序列的rna引入至宿主细胞中可在体外或离体或体内进行。举例而言,可通过包含编码本公开多肽的核苷酸序列的rna对宿主细胞(例如,nk细胞、细胞毒性t淋巴细胞等)进行体外或离体电穿孔。细胞本公开提供哺乳动物细胞系,其产生遗传修饰的靶哺乳动物细胞及靶哺乳动物细胞自身的复制缺陷型重组逆转录病毒颗粒。适合的哺乳动物细胞包括原代细胞及固定化细胞系。适合的哺乳动物细胞系包括人类细胞系、非人类灵长类细胞系、啮齿动物(例如小鼠、大鼠)细胞系,及类似者。适合的哺乳动物细胞系包括但不限于hela细胞(例如美国模式培养物保藏所(atcc)第ccl-2号)、cho细胞(例如atcc第crl9618号、第ccl61号、第crl9096号)、293细胞(例如atcc第crl-1573号)、vero细胞、nih3t3细胞(例如atcc第crl-1658号)、huh-7细胞、bhk细胞(例如atcc第ccllo号)、pc12细胞(atcc第crl1721号)、cos细胞、cos-7细胞(atcc第crl1651号)、ratl细胞、小鼠l细胞(atcc第ccli.3号)、人类胚胎肾(hek)细胞(atcc第crl1573号)、hlhepg2细胞、hut-78、jurkat、hl-60、nk细胞系(例如nkl、nk92及yts),及类似者。在一些情况下,细胞并非固定化细胞系,而是自受试者获得的细胞或离体细胞(例如,原代细胞)。举例而言,在一些情况下,细胞为自受试者获得的免疫细胞。作为另一实例,细胞为自受试者获得的干细胞或祖细胞。活化免疫细胞的方法本公开提供体外、体内或离体活化免疫细胞的方法。该方法通常包括使免疫细胞(体外、体内或离体)与一种或多种靶标抗原接触,其中该免疫细胞已遗传修饰以产生本发明微环境限制的car。在一种或多种靶标抗原存在下,微环境限制的car活化免疫细胞,从而产生经活化的免疫细胞。免疫细胞包括例如细胞毒性t淋巴细胞、nk细胞、cd4+t细胞、t调节(treg)细胞,γδt细胞、nk-t细胞、嗜中性粒细胞等。将遗传修饰的免疫细胞(例如t淋巴细胞、nk细胞)与一种或多种靶标抗原接触可使由免疫细胞产生的细胞因子与在不存在一种或多种靶标抗原时由免疫细胞所产生的细胞因子量相比增加至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约40%、至少约50%、至少约75%、至少约2倍、至少约2.5倍、至少约5倍、至少约10倍,或大于10倍。产量可增加的细胞因子包括但不限于il-2及ifn-γ。将遗传修饰的细胞毒性细胞(例如细胞毒性t淋巴细胞)与aar接触可使细胞毒性细胞的细胞毒性活性与在不存在一种或多种靶标抗原时细胞毒性细胞的细胞毒性活性相比增加至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约40%、至少约50%、至少约75%、至少约2倍、至少约2.5倍、至少约5倍、至少约10倍,或大于10倍。将遗传修饰的细胞毒性细胞(例如细胞毒性t淋巴细胞)与一种或多种靶标抗原接触可使细胞毒性细胞的细胞毒性活性与在不存在一种或多种靶标抗原时细胞毒性细胞的细胞毒性活性相比增加至少约10%、至少约15%、至少约20%、至少约25%、至少约30%、至少约40%、至少约50%、至少约75%、至少约2倍、至少约2.5倍、至少约5倍、至少约10倍,或大于10倍。在另一些实施方式中,例如,取决于宿主免疫细胞,使遗传修饰的宿主细胞与抗原接触可增加或减少细胞增殖、细胞存活、细胞死亡,及类似者。用于制备微环境限制的抗原特异性靶向区的方法在一些实施方式中,car抗原结合结构域(本文中也称作“抗原特异性靶区”或“astr”)组成型地结合其同源抗原(cognateantigen)。在另一些实施方式中,astr可受微环境限制,优选地或仅在某些异常条件下结合其同源抗原,例如存在于肿瘤微环境中的那些条件下,如本文更详细地公开。优选地或仅在肿瘤微环境的异常条件下结合的微环境限制的astr可减少靶向肿瘤外效应,这是因为在正常生理条件下与抗原结合减少,在一些情况下减少至低于通过免疫测定检测的水平。在某些方面中,本文所提供的car包括特异性结合靶蛋白的微环境限制的astr,其中该astr为包括重链可变区域及轻链可变区域的scfv片段。本文所公开方面的某些说明性实施方式,例如本文所公开的方法、细胞、细胞系、复制缺陷型重组逆转录病毒颗粒、多核苷酸或载体,包括car,该car包括微环境限制的抗原特异性靶向区。因此,在一个方面中,本文提供用于结合靶标抗原的嵌合抗原受体,其包括:a)微环境限制的抗原特异性靶向区,与正常生理环境相比在异常条件下其呈现出与靶标抗原结合增加,其中该抗原特异性靶向区结合至该靶标;b)跨膜结构域;及c)胞内活化结构域。在另一方面中,本文提供用于结合靶标抗原的嵌合抗原受体,其包括:a)至少一个微环境限制的抗原特异性靶向区,其通过淘选多肽文库且在与正常生理条件相比的异常条件下在靶标抗原结合测定中活性增加而经选择;b)跨膜结构域;及c)胞内活化结构域。在本文公开的任何方面的一些实施方式中,任何嵌合抗原受体可以是微环境限制的,使得与正常生理条件相比在异常状态下其表达出增加的结合活性。在本文公开的任何方面的一些说明性实施方式中,自初始多肽库识别微环境限制的astr,而无需在筛选/进化之前突变/进化文库的成员和/或无需在任选地重复轮次筛选期间或之间突变。例示性跨膜结构域及胞内活化结构域可以是本文公开的用于car的任何结构域。在一个方面中,本文提供一种用于选择微环境限制的astr的方法,其包含通过以下步骤淘选多肽展示文库:a.使多肽展示文库的多肽在正常生理条件下进行靶标抗原结合测定,且在异常条件下进行靶标抗原结合测定;及b.选择在与生理条件相比的异常条件下显示出靶标抗原结合活性增加的多肽,从而选择微环境限制的抗原特异性靶区。在另一方面中,本文提供一种用于分离微环境限制的astr的方法,其包括通过以下步骤淘选多肽展示文库:在异常条件下使多肽文库与结合至固体载体的靶标抗原接触,其中表达与结合靶标抗原多肽的克隆仍经由靶标抗原结合至固体载体;在生理条件下使固体载体与结合多肽一起孵育;及收集在生理条件下自固体载体洗脱的克隆,从而分离微环境限制的抗原特异性靶向区。在本文公开的任何方面的一些说明性实施方式中,自初始多肽文库筛选识别微环境限制的抗原特异性靶向区,而无需在筛选之前突变/进化库的成员和/或无需在任选地重复轮次筛选或淘选期间或之间突变/进化。正常生理条件可包括针对受试者而言被认为在给药位点或在作用位点的组织或器官处的正常范围内的温度、ph、渗透压、重量摩尔渗透浓度、氧化应力及电解质浓度等。异常条件为偏离正常可接受范围的条件。在一个方面中,微环境限制的抗原特异性靶向区(即多肽)在正常条件下实际上无活性,但在除正常条件以外在与正常条件相同或更好的位准下具有活性。举例而言,在一个方面中,微环境限制的抗原特异性靶向区在体温下实际上无活性,但在更低温度下具有活性。在另一方面中,在正常条件下使微环境限制的抗原特异性靶向区可逆地或不可逆地失活。在另一方面中,微环境限制的抗原特异性靶向区为治疗性蛋白质。在另一方面中,将微环境限制的抗原特异性靶向区用作药物或治疗剂。在又一方面中,微环境限制的抗原特异性靶向区在高度充氧的血液中(例如在通过肺之后或在肾脏中发现的较低ph环境中)或多或少具有活性。在一些实施方式中,执行单轮选择以获得微环境限制的抗原特异性靶向区。在某些实施方式中,在识别在异常条件下结合抗原并且在生理条件下不结合的游离多肽或表达具有这种性质的测试多肽的细胞或通过在初始或上一轮中具有这种性质的测试多肽包被的噬菌体之后重复筛选或淘选方法。在一些方法中,收集的噬菌体用于感染也可通过辅助噬菌体感染的细胞,以便扩增收集噬菌体。在测试细胞表面多肽的其他方法中,可使收集的细胞生长以通过扩增编码多肽的细胞中的多核苷酸来“扩增”由细胞表达的多肽。在一些实施方式中,通过生长表达所识别的多肽的细胞来完成扩增,而无需进行处理以在各轮之间突变编码所识别的多肽的多核苷酸。因此,通过扩增含有编码这些收集的多肽的多核苷酸的细胞来富集前一轮中收集的多肽。淘选或筛选方法可进行单次,或重复1次至1000次。在说明性实施方式中,淘选重复1次至20次或2次至10次或2次至5次。在另一些方法中,在各轮淘选之间使用一轮或多轮突变/进化来执行针对相关抗原(即靶标抗原)微环境限制的astr。在一个方法中,例如通过产生多肽或蛋白质文库并筛选该多肽或蛋白质文库得到具有对靶标抗原有所需结合亲和力的多肽或蛋白质来识别野生型蛋白质。在野生型蛋白质为抗体的一些实施方式中,可通过产生及筛选多克隆或单克隆抗体文库(包括噬菌体展示抗体文库,例如噬菌体展示人源化抗体文库)发现野生型抗体。可通过使野生型蛋白质或编码野生型蛋白质的核酸序列进行突变诱发处理以产生突变多肽群来产生进化的astr,这种突变多肽可经筛选以识别在肿瘤环境和/或体外肿瘤替代测定条件中具有与正常生理环境相比增加的活性(例如与靶标抗原的结合亲和力增强)的astr。此类方法的实例提供于wo2016033331(“conditionallyactivechimericantigenreceptorsformodifiedtcells”)或美国专利第8709755号中,这些专利的全部内容以引用方式并入本文中。产生微环境限制抗体的方法在本文中以全文引用方式并入本文中。在另一些实施方式中,可通过在异常条件及生理条件下筛选初始多肽库且自初始多肽文库识别在异常条件下及生理条件下优先地或专有地结合的测试多肽,来识别微环境限制的抗原特异性多肽(即靶向区,例如抗体)。在一些实例中,在初始多肽库筛选中,自初始多肽文库识别的经识别且分离的微环境限制的抗原特异性多肽(即靶向区,例如抗体)在异常条件下及生理条件下优先地或专有地结合其同源抗原。在此类情况中,不进行一轮突变/进化。因此,进行说明性实施方式中的方法而无需在筛选轮次(例如,淘选轮次)之间使编码经分离微环境限制性抗原特异性靶向区的多肽突变,或在异常条件及生理条件下仅针对单个结合测定进行以分离且识别微环境限制的抗原特异性多肽(即靶向区,例如抗体)。该方法可通过在筛选和/或淘选轮次之间培养、高保真扩增和/或稀释编码抗原特异性靶向区的多核苷酸或包括其的宿主生物体来执行而无需任何突变/进化。此外,该方法可在无需重复筛选和/或淘选的情况下进行,且可在微环境限制的抗原特异性多肽(即靶向区,例如抗体)经分离之后进行而无需使编码经分离的微环境限制的抗原特异性靶向区的多核苷酸突变/进化。用于本文所提供的用以检测多肽与同源结合配偶体结合的方法的测定包括基于细胞的测定,且尤其使用细胞表面展示系统(例如哺乳动物细胞表面显示系统)进行的测定。在例示性方法中,可将编码多肽或变异多肽库(包括经修饰的多肽库)的核酸引入至适于在细胞(例如哺乳动物细胞)中表达的载体中。接着通过载体转染细胞,且由这些细胞表达一种或多种多肽。可使含有表面表达多肽的细胞库与含有可溶性或表面结合的同源结合配偶体的溶液接触。可使用可检测对细胞表面结合的任何测定法来检测结合活性。也可通过评估多肽的功能活性或多肽来评估活性。本领域技术人员已知的任何基于细胞的测定均被考虑用于本文所提供的方法,包括细胞增殖测定、细胞死亡测定、流式细胞术、细胞分离技术、荧光活化细胞分选(facs)、相位显微镜、荧光显微镜、受体结合测定、细胞信号传导测定、免疫细胞化学及报道子基因测定。在一些实例中,测定为荧光活化细胞分选(facs)测定。多肽或蛋白质可由哺乳动物细胞表达为分泌的可溶性分子、细胞表面分子或胞内抗体。在例示性方法中,可在大部分或全部细胞显示锚定于细胞表面上的蛋白质文库成员的条件下通过蛋白质文库来转染细胞。任选地,可使用表达系统,其中大多数哺乳动物细胞转染体在其基因组中仅具有一个质粒。因此,大多数(即,至少约70%或约80%或约90%)转染物表达一种多肽中的一种或多种分子。此可通过例如分离且培养受试者转染子且对表达的序列进行扩增及测序以确定其是否具有单个序列来验证。在本文所提供的方法的一些实例中,多肽为展示于哺乳动物细胞表面的抗体。本文所描述的任何抗体可在哺乳动物细胞表面上表达,包括全长二价功能性抗体,例如igg抗体。抗体可以是片段,例如fab片段或scfv片段。抗体可以包括fc区,例如scfv-fc或包含两个重链及两个轻链的全长抗体。本领域技术人员可选择适合的抗体片段。举例而言,在哺乳动物细胞中制备scfv-fcs及全长抗体可具有优于scfv's或fab片段的若干优点。可用于本文所提供的结合测定中的固体载体包括能够与多肽的结合配偶体(例如配体,受体或抗原)附着的任何载体。为促进高通量筛选,通常将同源结合配偶体附着至固体载体。在本文所提供的方法中用作固体载体的载体实例包括但不限于玻璃、聚苯乙烯、聚丙烯、聚乙烯、葡聚糖、尼龙、直链淀粉、天然纤维素及改质纤维素、聚丙烯酰胺、琼脂及磁性固体载体,例如包括磁铁矿的固体载体。固体载体可以是此项技术中已知的一种或多种珠粒或颗粒、微球、管或板表面、过滤膜,及其他固体载体。例示性固体载体系统包括但不限于由例如玻璃、硅、金属、尼龙、纤维素、塑料或复合材料(包括多孔板或膜)构成的平坦表面;或可呈珠粒形式,例如矽胶、受控孔玻璃、磁性或纤维素珠粒。此外,此类方法可适用于悬浮液或以柱形式使用。在一些实施方式中,通过生物淘选具有固定化靶标抗原的噬菌体展示或酵母表面展示(colby等人,“engineeringantibodyaffinitybyyeastsurfacedisplay,”meth.enzym.388,26(2004))的抗体(例如人源化抗体)文库来识别及分离微环境限制的抗原特异性多肽(即靶向区域,例如抗原)。举例而言,可使用本文中的噬菌体显示或酵母表面显示方法来淘选天然人源化抗体库或合成的人源化抗体文库。在一些实施方式中,初始噬菌体展示过程可将噬菌体克隆转移至哺乳动物载体并用于哺乳动物细胞表面筛选方法(参见例如yoon等人,bmcbiotechnology12:62;1472-6750(2012))。在实施例2中提供用于进行噬菌体显示以分离微环境限制抗原特异性靶区的例示性方法。使用本文提供的方法识别微环境限制的astr可以是抗体、抗原、配体、配体的受体结合结构域、受体、受体的配体结合结构域或亲和体。在微环境限制的astr为抗体的实施方式中,其可以是全长抗体、单链抗体、fab片段、fab'片段、(fab')2片段、fv片段,及二价单链抗体或双功能抗体。其中该抗原特异性靶向区包含来自抗体重链及轻链。在一些实施方式中,微环境限制的astr为单链可变片段。此类单链可变片段可具有由接头隔开的重链及轻链,其中该接头长度在6个至100个氨基酸之间。在一些实施方式中,重链位于嵌合抗原受体上的轻链的n端。在另一些实施方式中,轻链位于重链的n端。微环境限制的astr可以是双特异性astr。使用本文提供的方法识别微环境限制的astr通常为多肽,且更特别地为多肽抗体,且在说明性实施方式中为单链抗体。这些多肽可在异常条件及正常条件下以更高或更低亲和力结合至同源抗原,但在说明性实施方式中在异常条件下以比正常条件下更高的亲和力结合。在一些实施方式中,这些多肽在异常条件下可以与生理(即正常)条件相比高10%、20%、25%、50%、75%、90%、95%或99%的亲和力来结合至其同源抗原。在一些实施方式中,使用本文提供的方法识别的astr在正常生理条件下不结合至同源抗原达到其使用阴性对照(例如阴性对照抗体)获得的背景水平以上的任何可检测水平。通过本文提供的方法分离的编码微环境限制的astr的核苷酸序列可通过对表达微环境限制的抗原特异性靶向的收集细胞的核苷酸进行测序来测定。接着可通过产生编码包含微环境限制的抗原特异性靶向区、跨膜结构域及胞内活化结构域的多肽的多核苷酸来将此核苷酸序列信息用于制备微环境限制的生物嵌合抗原受体(mrb-car)。微环境限制的抗原特异性靶向区可经克隆至car构建体表达系统中,其可用以在其基因组中产生包括car的重组慢病毒,且接着重组慢病毒可用于转导t细胞来在与生理条件相比的肿瘤选择性环境中测试表达靶细胞杀伤的car介导的肿瘤抗原。条件性活性的条件在本文提供的方法中,在两组不同条件下筛选或测试一种或多种多肽(例如单链抗体)的活性,该两组条件模拟两种不同生理环境中的一个或多个条件,例如病态微环境及非病态微环境的正常生理条件。通常,这些条件为可在体外模拟或复制的条件。一组条件可包括用以模拟与疾病相关的微环境的一个或多个条件。疾病可改变细胞内平衡及细胞外平衡。举例而言,病变微环境可模拟肿瘤微环境或癌症微环境中的一个或多个条件。通常,两组条件下的活性差异可产生分子的条件性活性。因此,与第二组条件(例如正常环境或非疾病环境中的模拟条件)相比,在第一组条件(例如模拟肿瘤微环境中的条件)下显示更高活性的分子经识别为微环境限制的候选分子。可选择两组条件以改变在两种生理环境中不同的一个或多个参数,例如本文所描述或熟习此项技术者已知的,包括但不限于化学条件、生物条件,或身体状况。可在两组条件之间变化的参数可包括选自以下的一个或多个条件:压力、温度、ph、离子强度、渗透压、重量摩尔渗透浓度、氧化应力、浊度,暴露于光(包括uv、红外线或可见光)、一种或多种溶质(例如电解质)的浓度、葡萄糖浓度、透明质酸浓度、乳酸或乳酸盐浓度、白蛋白浓度、腺苷含量、r-2-羟基戊二酸含量、丙酮酸浓度、氧浓度和/或氧化剂、还原剂或辅助因子的存在。通过改变校准溶液中的电解质及缓冲体系,可将例如ph、缓冲能力、离子环境、温度、葡萄糖浓度及离子强度的生理条件调节至待模拟生物环境的生理条件。通过本文所描述的一个或多个条件,可将模拟正常生理环境的这组条件选择为与模拟病变微环境(例如肿瘤微环境)的一组条件不同。举例而言,如以下所论述,与非肿瘤微环境相比,肿瘤微环境的多种参数不同,包括但不限于氧浓度、压力、辅助因子的存在、ph、透明质酸浓度、乳酸浓度、白蛋白浓度、腺苷含量、r-2-羟基戊二酸含量及丙酮酸浓度。与在非肿瘤环境或正常环境中存在的条件相比,这些参数中的任一个可在体外复制以模拟存在于肿瘤或癌症环境中的一种或多种条件。可模拟的正常生理条件包括在身体任何位置处健康或非疾病组织(例如胃肠道、皮肤、脉管系统、血液及细胞外基质)中发现的环境。通常,在本文的分析中,生理条件可通过选择用于评估蛋白质活性的缓冲液来在体外进行模拟。举例而言,可通过用以评估测定中活性的测定缓冲液中的差异来模拟病变微环境(例如肿瘤微环境)及非病变环境的任何一个或多个条件。因此,在本文中用以识别微环境限制的多肽的方法中,使测定缓冲液的一种或多种组分或一种或多种特征改变或在用以在第一条件下测试活性的第一测定中与用以在第二个条件下测试活性的第二测定中不同。举例而言,如本文所论述,肿瘤微环境的多种参数与非肿瘤环境相比不同,包括但不限于氧、压力、辅助因子的存在、ph、透明质酸浓度(例如提高或降低透明质酸浓度)、乳酸盐浓度(例如提高或降低乳酸盐浓度)、白蛋白浓度(例如提高或降低白蛋白浓度)、腺苷含量(例如提高或降低腺苷含量)、r-2-羟基戊二酸含量(例如提高或降低r-2-羟基戊二酸含量)及丙酮酸盐浓度(包括提高或降低丙酮酸浓度)。特别地,肿瘤微环境中的条件可包括与非肿瘤微环境相比较低的ph、较高浓度的透明质酸、较高浓度的乳酸盐及丙酮酸盐、较高浓度的白蛋白、增加的腺苷含量,增加的r-2-羟基戊二酸含量、低氧、较低浓度的葡萄糖及略微较高的温度。举例而言,微环境限受限的astr在正常体温下实际上无活性,但在肿瘤微环境中在较高温度下具有活性。在又一方面中,微环境限制的抗体在正常氧合血液中具有较低活性,但在存在于肿瘤中的较少氧合环境下具有较高活性。在又一方面中,微环境限制的抗体在正常生理ph7.2至7.8中具有较低活性,但在肿瘤微环境中存在的酸性ph5.8至7.0或6.0至6.8下具有较高活性。举例而言,微环境限制的抗体在ph6.7下比在ph7.4下具有更高活性。在本领域技术人员已知的肿瘤微环境中存在也可用作本发明的条件的其他条件,在这些条件下条件活性astr具有不同的结合亲和力。模拟这些体内肿瘤条件的体外测定条件在本文中称作体外肿瘤替代测定条件。可通过选择特定测定缓冲液来在体外模拟这些条件中的任一种或多种。模拟病变微环境的测定缓冲液的组合物可选择为与模拟正常环境的测定缓冲液的组合物相同,除本文已知或描述的在病变微环境中改变的一个或多个条件之外。此外,在筛选或识别一种或多种多肽在两组不同条件下的活性时,通常仅在测定中变化的条件与模拟体内微环境的缓冲条件相关。测定的其他条件(例如时间、温度及孵育条件)对于两组条件可相同。通常,在模拟病变微环境的该组条件及模拟正常微环境条件中使用相同的碱性缓冲液,但缓冲组合物的设计可在例如以下的一个或多个参数上不同,例如ph、氧、压力、辅助因子的存在、ph、透明质酸浓度(例如提高或降低透明质酸浓度)、乳酸盐浓度(例如提高或降低乳酸浓度)、白蛋白浓度(例如提高或降低透明质酸浓度)和/或丙酮酸浓度(包括提高或降低丙酮酸浓度)。在模拟病变微环境条件及模拟正常微环境条件下,可使用本领域技术人员已知的任何碱性缓冲液。产生微环境限制的细胞的方法本公开提供一种产生微环境限制细胞的方法。该方法大体上包括通过表达载体(例如质粒或逆转录病毒载体)或包括编码本发明的微环境限制的car的核苷酸序列的rna(例如体外转录的rna)来遗传修饰哺乳动物细胞。遗传修饰的细胞在一种或多种靶标抗原存在下受微环境限制。遗传修饰可在体内、体外或离体进行。细胞可以是免疫细胞(例如t淋巴细胞、t辅助细胞或nk细胞)、干细胞、祖细胞等。在一些情况下,离体进行遗传修饰。举例而言,自受试者获得t淋巴细胞、干细胞、t辅助细胞或nk细胞;且将自受试者获得的细胞进行遗传修饰以表达本发明的car。遗传修饰的细胞在一种或多种靶标抗原存在下可受微环境限制。在一些情况下,遗传修饰的细胞经离体活化。在其他情况下,将遗传修饰的细胞引入受试者(例如自其获得细胞的受试者);且遗传修饰的细胞在体内活化。举例而言,当一个或多个靶标抗原存在于受试者的细胞表面上时,不需要施用抗原。遗传修饰的细胞与存在于受试者的细胞表面抗原接触,且将遗传修饰的细胞活化。举例而言,当遗传修饰的细胞为t淋巴细胞时,遗传修饰的细胞可对于在car结合的表面上表达一种或多种靶标抗原细胞展现细胞毒性。用于通过改变ph来调节mrbcar表达的t细胞和/或nk细胞活性的方法在某些方面中,本文提供通过在微环境(包括靶向组织内或一个或多个非靶向(例如健康/正常)组织内的靶细胞)内产生ph变化或偏移,通过调节mrb-car与这些靶细胞上的同源抗原的结合来调节表达mrbcar的t细胞和/或nk细胞与靶细胞结合及产生靶细胞裂解/杀伤的方法。此类方法通常包括在微环境中使例如哺乳动物细胞(例如人细胞)的靶细胞与表达mrbcar的t细胞和/或nk细胞接触,且接着通过降低或更通常地提高ph来改变微环境ph。微环境可以是靶标微环境,例如肿瘤或脱靶微环境,其中脱靶结合可产生副作用。在一些实施方式中,此类方法可提供短暂降低的对肿瘤微环境敏感的car-t靶向结合。因此,在一个方面中,本文提供用于调节微环境限制的表达生物嵌合抗原受体(mrb-car)的t细胞或nk细胞与表达受试者中mrb-car同源抗原细胞结合的方法,该方法包括以下:a.将包含编码mrb-car的核酸的t细胞和/或nk细胞引入至受试者中,其中在引入之后(以及任选地和/或在其期间),包含编码mrb-car的核酸的t细胞和/或nk细胞表达mrb-car且结合至在受试者中表达同源抗原细胞;及b.将药物试剂以充足量施用至受试者以增加血液ph和/或组织的ph和/或微环境的ph,其中该给药在引入之前、期间或之后执行,且其中血液、组织和/或微环境增加的ph介导表达mrb-car的t细胞和/或nk细胞与表达具有增加ph的血液、组织或微环境中的同源抗原细胞的结合。在包括施用本发明的ph调节药剂步骤的方面中,ph的变化/偏移可通过将靶向或非靶向细胞/组织暴露于ph调节药剂来实现,例如通过向受试者施用ph调节药剂。本文提供ph调节药剂的非限制性实例。在某些方面中,本文提供在用于调节mrb-car与其同源抗原结合或用于调节mrbcar表达的t细胞和/或nk细胞与表达其同源抗原的细胞结合或用于减少或减轻受试者中靶向肿瘤外毒性的方法中使用的药剂。在某些实施方式中,此类方面是关于治疗肿瘤生长、癌症、增生或细胞增殖性疾病。在另一些方面中,本文提供ph调节药剂的用途,其用于制备用于控制经遗传改造的t细胞和/或nk细胞与受试者中的靶标哺乳动物细胞在体内结合的药物或试剂盒中。在另一些方面中,本文提供一种试剂盒,其包括含有复制缺陷型重组逆转录病毒颗粒的容器及其用于执行治疗肿瘤生长方法的说明书,其中该说明书指示通过调节ph来控制t细胞和/或nk细胞与靶标哺乳动物细胞结合的方法。此类方法可以是本文此部分提供的用于通过改变ph来调节表达mrbcar的t细胞和/或nk细胞结合和/或活性方法中的任何一种。含有重组逆转录病毒颗粒的容器可以是用于储存重组逆转录病毒颗粒和/或ph调节药剂的管、小瓶,孔板,或其他容器。这些容器中的任何一种可具有工业强度及等级。在某些实施方式中,该试剂盒可包括两个或更多个容器。一个容器(container/vessel)可包括重组逆转录病毒颗粒且另一容器可包括ph调节药剂。在此类方法中,药剂以足够量经递送/施用以提高血液ph和/或组织ph和/或微环境ph以调节表达mrb-car的经修饰/重组t细胞和/或nk细胞的mrb-car与具有增加ph的血液和/或组织中的同源抗原结合。本文提供用于以足够量施用ph调节药剂且持续足够时间的非限制性例示性细节。在将mrb-car引入至受试者中之后,无论相对于组织在靶或脱靶的靶细胞,均可与例如ph调节药剂的ph调节剂接触。因此,本文提供的用于调节表达mrbcar的t细胞结合和/或细胞毒性活性(例如用于减轻靶向肿瘤外活性和/或用于抑制例如肿瘤细胞增殖的靶细胞增殖)的例示性方面可包括以下步骤:a.将包含编码mrb-car的核酸的t细胞和/或nk细胞引入至受试者中,其中在引入之后,包含编码mrb-car的核酸的t细胞和/或nk细胞表达mrb-car,且任选地结合至在受试者中表达同源抗原细胞;及b.以足够量将药剂施用至受试者以提高血液ph和/或组织ph和/或微环境ph以调节表达mrbcar的t细胞和/或nk细胞与表达具有增加ph的血液、组织或微环境中的mrbcar的同源抗原细胞的结合。将理解,取决于用以将编码mrb-car的核酸引入至t细胞和/或nk细胞中的具体方法,t细胞和/或nk细胞可或可不在其经引入至受试者中之前表达mrb-car。然而,在引入至受试者中之后的一些时间点下,例如,2小时、4小时、8小时、12小时、1天、2天、4天和/或7天或更长时间,包括编码mrb-car的核酸的t细胞和/或nk细胞表达mrb-car。接着此类细胞通常结合至表达mrb-car的同源抗原的靶细胞。本文提供的用于遗传修饰及任选地扩增受试者的淋巴细胞的方法可用于将编码mrb-car的核酸序列引入至受试者的t细胞和/或nk细胞基因组中以产生能够表达mrbcar的t细胞和/或nk细胞,且接着将能够表达mrbcar的t细胞和/或nk细胞引入至受试者中,其中在引入之后,t细胞和/或nk细胞表达mrbcar,以便使mrb-car与靶细胞/组织接触。本发明提供如何执行这些方法以及用于修饰及扩增淋巴细胞的多种替代方案的细节,其中任何一种可用于本发明的方面,包括改变ph以调节表达mrbcar的t细胞和/或nk细胞与表达mrb-car的同源抗原的靶细胞的结合。用于遗传修饰及扩增淋巴细胞的此类方法通常涉及使t细胞和/或nk细胞(在说明性实施方式中可以是静息细胞)与复制缺陷型重组逆转录病毒颗粒接触以转导这些t细胞和/或nk细胞。这种接触通常在自受试者移除淋巴细胞之后离体进行。接着将t细胞和/或nk细胞引入/重新引入至通常已将其移除的受试者中。复制缺陷型重组逆转录病毒颗粒包括具有编码mrb-car的多核苷酸的基因组。关于此类复制缺陷型重组逆转录病毒颗粒的许多替代实施方式及其他细节提供在本文其他部分中,且可用于本文提供的方法中,该方法以取决于ph的方式通过调节表达由mrb-car识别的同源靶标多肽的细胞微环境中的ph来调节mrb-car的结合及产生其裂解/杀灭。包括此类组合物的特别说明性方面可包括用于调节本文中其他部分提供的表达mrbcar的t细胞结合、细胞杀伤活性和/或存活的其他元件。在包括ph变化的方法以及本文提供的另一些实施方式中可与mrbcar表达组合的此类控制元件包括核糖开关及消除结构域,因此,本文提供的此类方法及组合物的组合形成一种强大的多方面途径来确保在将表达car的t细胞(包括表达mrbcar的t细胞)施用受试者之后的受试者的安全性。用于通过表达mrbcar的t细胞和/或nk细胞调节靶细胞结合的此类方法可用以例如通过增加受试者内血液和/或非肿瘤组织的ph来降低在靶脱瘤毒性。举例而言,在受试者内“正常”组织ph变得瞬间降低的情况下,可以正常组织的ph增加同时肿瘤的ph保持较低且仍处于表达mrb-car的t细胞和/或nk细胞结合靶标肿瘤细胞ph下的方式来递送ph调节剂。在这些实施方式中,可以较低浓度或以靶向方式将ph调节剂递送至正常组织。在一些实施方式中,此可完成同时使肿瘤微环境内ph保持足够低以使mrb-cart细胞和/或nk细胞与肿瘤内的其同源靶标表达细胞结合。在本文所提供方法的说明性方面中,组织的ph保持在表达mrbcar的t细胞和/或nk细胞与其靶标结合一段时间的ph下,该时间足以使表达mrbcar的t细胞和/或nk细胞接触且结合表达其同源抗原细胞(例如2小时、4小时、8小时、12小时或24小时,或2天、4天、7天、14天、28天或30天,或1个月、2个月、3个月、4个月、5个月、6个月、12个月、24个月,或更长时间),且接着使ph偏移/改变,例如通过使组织的ph提高至一程度以影响表达mrbcar的t细胞和/或nk细胞与靶细胞结合。因此,在一个方面中,本文提供一种经由药物改变血管及组织ph来瞬时降低肿瘤微环境敏感性car-t细胞靶结合的方法。在不同条件下具有不同结合性的肿瘤微环境敏感性car-t细胞的靶结合部分也称为微环境限制生物学、微环境限制、微环境受控或条件活性,且可指示整个car或其任何靶结合结构域,例如astr、scfv或scfvfc。car-t细胞中的这些微环境受控的astr提供额外位准保护以避免靶向肿瘤外毒性,需要肿瘤局部环境条件实现t细胞接合。尽管对于一些单克隆抗体疗法具有吸引力,但过继性细胞疗法可产生对其car-t靶标瞬时容许的局部环境。举例而言,在具有低ph组织中活化的car-t细胞可进一步降低微环境ph,取决于car构建体中存在的细胞质结构域。在其他情况下,与过继性细胞治疗相关的细胞因子释放综合征及其他病状可导致血液的碳酸氢盐缓冲能力损失,从而引起乳酸性酸中毒。已确立通过静脉内输注施用的过继性细胞疗法产生暂时性肺压迫。对于一些细胞疗法,输注速率需要持续监测溶解氧(fischer等人stemcellsdev.2009年6月;18(5):683-691)。肺压迫程度取决于细胞大小、活化状态、细胞剂量及输注速率。cruz等人(cytotherapy.2010年10月;12(6):743-749)报道来自超过300个t细胞输注的不良发现,低剂量及缓慢输注可减少肺压迫。然而,通过某些高效car-t细胞,甚至以低含量存在于肺内皮上的靶标,例如her2(morgan等人molther.2010年4月;18(4):843-851),可产生不可控制的立即毒性,且由于输注后肺中的初始高car-t细胞浓度以及这些组织中t细胞靶标的存在而使得患者急剧恶化。在另一些情况下,t细胞靶标在例如胆管的其他脱靶组织中存在可产生不可控制的在靶脱瘤毒性(lamersmolther.2013年4月;21(4):904-12)且在可使用例如类固醇的其他试剂或细胞消除表位之前产生严重器官毒性。尽管静脉及动脉血浆对酸中毒具有强缓冲能力,但呼吸性酸中毒、休克、代谢性酸中毒及缺血性酸中毒的条件可发生于通过过继性细胞疗法治疗的癌症患者中。在本文提供的一些方面中,mrb-car在受试者中的结合可通过向受试者施用药剂来调节,以提高或降低血液、组织和/或微环境的ph。在一些方面中,可通过向受试者施用药剂以提高或降低血液ph和/或组织ph和/或微环境ph来在受试者中减轻在靶脱瘤毒性。在一些方面中,t细胞和/或nk细胞与靶标哺乳动物细胞的结合可通过引入药剂以提高或降低血液ph和/或组织ph和/或微环境ph来控制。在一些方面中,可通过向受试者施用ph调节药剂来控制受试者体内经遗传改造的t细胞和/或nk细胞与靶标哺乳动物细胞的结合。在说明性实施方式中,药剂可提高血液ph和/或组织ph和/或微环境ph。在一些实施方式中,微环境可以是体内微环境。在说明性实施方式中,微环境可以是肿瘤微环境。在一些实施方式中,微环境可包括靶标哺乳动物细胞,其中靶标哺乳动物细胞在其表面上表达靶标抗原。在一些实施方式中,向受试者施用药剂可将血液、组织和/或微环境的ph自小于5.8、5.9、6.0、6.1、6.2、6.3、6.4、6.5、6.6、6.7、6.8或6.9ph增加至至少6.6、6.7、6.8、6.9、7.0、7.1、7.2、7.3、7.4、7.5或7.6ph,其中血液、组织和/或微环境ph在施用药剂之前低于施用药剂之后。在一些实施方式中,向受试者施用药剂可将血液、组织和/或微环境ph自高于6.6、6.7、6.8、6.9、7.0、7.1、7.2、7.3、7.4、7.5或7.6ph降低至低于5.8、5.9、6.0、6.1、6.2、6.3、6.4、6.5、6.6、6.7、6.8、6.9或7.0ph,其中血液、组织和/或微环境ph在施用药剂之前高于施用药剂之后。在一些实施方式中,向受试者施用药剂可在受试者血液、组织和/或微环境中产生ph偏移。在一些实施方式中,ph偏移可以是在任一方向上至少0.1ph单位、0.2ph单位、0.3ph单位、0.4ph单位、0.5ph单位、0.6ph单位、0.7ph单位、0.8ph单位、0.9ph单位、1.0ph单位、1.1ph单位、1.2ph单位、1.3ph单位、1.4ph单位、1.5ph单位、1.6ph单位、1.7ph单位,或1.8ph单位,即在施用药剂之后的ph相对于施用药剂之前的ph提高或降低。在说明性实施方式中,ph偏移为ph增加。本发明的mrb-car可在一个ph下而非不同ph下减少与其同源抗原结合。在针对mrb-car差异性结合的说明性ph值未曾提供于广泛方面中的说明性实施方式中及替代地对于代替用于这些方面的那些值的其他实施方式中,mrb-car可在较高ph下而非较低ph下减少结合。举例而言,mrb-car可在高于7.0、7.1、7.2、7.3、7.4或7.5ph下而非低于6.4、6.5、6.6、6.7、6.8、6.9或7.0ph下减少与其同源抗原结合。在其他实施方式中,mrb-car可在较低ph下而非较高ph下减少结合。举例而言,mrb-car可在低于6.4、6.5、6.6、6.7、6.8、6.9或7.0ph下而非高于7.0、7.1、7.2、7.3、7.4或7.5ph下减少与其同源抗原结合。在一些说明性实施方式中,相比于在ph7.4至7.6下,mrb-car在6.5至6.7ph下展现增加的结合。在其他说明性实施方式中,相比于在7.4ph下,mrb-car在6.7ph下展现增加的结合。在另一些实施方式中,相比于在血液ph下,mrb-car在肿瘤ph下展现增加的结合。在一些实施方式中,mrb-car可包括抗原特异性靶向区、柄及胞内活化结构域。在一些实施方式中,mrb-car也可包括共刺激结构域。在一些实施方式中,mrb-car可与肿瘤相关抗原结合。在包括调节血液、组织或微环境ph的方法中,可将微环境ph自低于7.0ph增加至高于7.0ph。举例而言,ph可自低于6.4、6.5、6.6、6.7、6.8、6.9或7.0ph增加至高于7.0、7.1、7.2、7.3或7.4ph。在一些实施方式中,mrb-car可在引入药剂之前在增加的ph下而非微环境ph下与同源抗原结合。在某些实施方式中,ph可自低于7.0增加7.1至8.0ph、或至7.1至7.8ph、或至7.2至7.8ph、或至7.2至7.6ph、或至7.3至7.6ph、或至7.4至7.8ph、或至7.4至7.6ph。视施用的药剂类型及剂量而定,此ph的增加可发生少于1小时、2小时、4小时、6小时、8小时、12小时或24小时,或多于1小时、2小时、4小时、6小时、8小时、12小时或24小时。在某些实施方式中,施用药剂,使得ph在靶标组织(例如肿瘤)中,及例如在靶标组织(例如,肿瘤)微环境的至少一个表面中,在靶标组织(例如,肿瘤)微环境的至少一部分中以及在说明性实施方式中在整个靶标组织(例如,肿瘤)微环境中保持高于7.0、7.1、7.2、7.3、7.4或7.5;或在7.0、7.1、7.2、7.3的范围下限与7.4、7.5、7.6、7.7或7.8的范围上限之间。微环境可以是体内微环境,例如肿瘤、组织、非肿瘤组织、正常组织或经历瞬时ph偏移的组织。举例而言,通常经历短暂ph变化的组织包括厌氧条件中的肌肉组织或经历锻炼的肌肉组织或已发炎组织或正经历发炎的组织。在包括靶标哺乳动物细胞的一些实施方式中,靶标哺乳动物细胞可以是肿瘤细胞或非肿瘤细胞或正常细胞。在一些方面中,提供用于瞬时提高血管ph以降低微环境受控的mrb-car对其抗原亲和力的方法。血液ph的0.4u偏移可使形成mrb-car的一部分的某些scfv对其同源抗原亲和力降低超过10倍。在一些实施方式中,可经由多种药剂的iv或口服施用途径来实现治疗性ph控制。举例而言,在一些实施方式中,可通过碳酸氢盐或碳酸氢钠来实现结合亲和力的失活。在另一些实施方式中,可使用三羟甲基氨基甲烷(也称为缓血酸胺(tromethamine/trometamol)及tham)和/或carbicarbtm(碳酸氢钠和碳酸钠等摩尔高渗溶液)可以足以减轻在靶脱瘤毒性的量来提高血液ph。在另一些实施方式中,可使用小分子质子泵抑制剂以足以减轻靶向肿瘤外毒性量来提高血液ph和/或组织ph。可用于包括调节ph方法中的质子泵抑制剂包括但不限于埃索美拉唑(esomeprazole)(nexium)、埃索美拉唑及萘普生(naproxen)(vimovo)、兰索拉唑(lansoprazole)(prevacid)、奥美拉唑(omeprazole)(prilosec及zegerid)及雷贝拉唑(rabeprazole)(aciphex)。质子泵抑制剂的施用可在更长时间内有效地用于调节抗原结合结构域与其同源抗原结合亲和力持续在数天,数周,数月或数年。在其他实施方式中,抗原结合结构域对其同源抗原亲和力可通过控制转运体及泵的转录、翻译、膜表达及稳定性来改变血液ph和/或组织ph来调节。经改变表达可调节ph的此类转运体及泵的实例包括但不限于质子泵、钠质子交换家族(nhe)、碳酸盐转运体家族(bct)及单羧酸转运体家族成员。在某些实施方式中,在输注患者表达微环境限制生物astr(例如scfv或scfvfcs)的car-t细胞之前或同时施用ph调节药剂,例如碳酸氢盐、tham或caricarbtm。此类治疗将减轻另外与car-t细胞输注的暂时性肺压迫相关的即时细胞毒性。因此,在某些方面中,本文提供通过向受试者施用足以提高血液ph和/或组织ph和/或微环境ph量的药剂;且同时或随后(例如之后1小时、2小时、4小时、6小时、8小时、12小时或24小时,或1天、2天、3天、4天或7天)将表达mrbcar的t细胞或nk细胞引入至受试者中来减少对受试者正常、健康组织造成细胞毒性的方法。在某些实施方式中,在此类引入之后的目标时间(例如之后1小时、2小时、4小时、6小时、8小时、12小时或24小时,或1天、2天、3天、4天或7天),持续在一段时间内或无限期终止施用药剂,以便改变受试者血液、组织或微环境ph并调节表达mrbcar的t细胞结合/活性。可以使用多种有效的给药方案来施用能够调节ph的药剂(例如提高受试者血液ph值和/或组织ph和/或微环境ph),如本领域技术人员将理解。在本文中,给药可指示给予受试者药剂,包括经由iv将药剂注射至受试者中,或向受试者或服用药剂的受试者提供口服剂量的药剂。药剂可施用至受试者或患者持续多种时间长度,例如至少1天、2天、3天、4天、5天或6天;1周、2周、3周、4周、5周、6周、7周、8周、9周、10周或11周;3个月、4个月、5个月、6个月、7个月、8个月、9个月、10个月、12个月、15个月或18个月;或2年、2.5年、3年、3.5年、4年、4.5年或5年或无限期。在一些实施方式中,药剂可以是碳酸氢盐、碳酸氢钠(nahco3)或碳酸氢钠及碳酸钠的溶液,且肠胃外或iv剂量可以是:0.2×受试者体重(kg)×受试者的碱缺失(basedeficit);所需hco3(meq)=0.5×重量(kg)×[24-血清hco3(meq/l)];或2meq/kg至5meq/kgiv输注持续4至8小时。在一些实施方式中,取决于酸中毒严重程度,可使用碳酸氢盐、碳酸氢钠及碳酸氢钠溶液的标准给药方案。举例而言,可经由iv以1公升/小时至1.5公升/小时的速率来施用稀释于1l含5%葡萄糖水溶液中的50meq至150meq碳酸氢盐。在其他非限制性实例中,可经由iv以1公升/小时至1.5公升/小时的速率来施用稀释于1l5%葡萄糖水溶液中的90meq至180meq碳酸氢盐。在药剂是碳酸氢盐或碳酸氢钠(nahco3)的一些实施方式中,肠内或口服剂量可以是例如给予受试者325mg至2000mg碳酸氢钠1次/天至4次/天。在一些实施方式中,药剂可以是三羟甲基氨基甲烷(也称为缓血酸胺(tromethamine/trometamol)和tham),且胃肠外或iv剂量可估计为:所需缓血酸胺溶液(ml,0.3m)=体重(kg)×碱缺失(毫克当量/公升)×1.1。在一些实施方式中,可自借助于siggaard-andersen列线图测定的meq/l细胞外液的缓冲液碱缺失(以meq/l为单位)来估计三羟甲基氨基甲烷的iv剂量。在一些实施方式中,初始剂量可以是通过缓慢iv输注至多1000ml来注射的500ml(150meq)三羟甲基氨基甲烷,其中最大剂量为500mg/kg(227mg/lb)持续时间不少于一小时。在一些实施方式中,药剂可以是小分子质子泵抑制剂,且可施用延长的治疗长度。举例而言,可投与小分子质子泵抑制剂持续至少1天、2天、3天、4天、5天或6天;1周、2周、3周、4周、5周、6周、7周、8周、9周、10周或11周;3个月、4个月、5个月、6个月、7个月、8个月、9个月、10个月、12个月、15个月或18个月;或2年、2.5年、3年、3.5年、4年、4.5年或5年或无限期。在一些实施方式中,质子泵抑制剂可以是埃索美拉唑(nexium),且可每天一次或两次经口施用20mg或40mg埃索美拉唑。在一些实施方式中,质子泵抑制剂可以是埃索美拉唑(nexium)与萘普生(vimovo)的组合,且可每天一次或两次经口施用20mg埃索美拉唑与375mg至500mg萘普生。在一些实施方式中,质子泵抑制剂可以是兰索拉唑(prevacid),且可每天一次或两次经口施用15mg、30mg或60mg兰索拉唑。在一些实施方式中,可通过iv每天一次历经30分钟注射30mg兰索拉唑来施用兰索拉唑并持续至多7天。接着受试者可切换至口服兰索拉唑并继续治疗。在一些实施方式中,质子泵抑制剂可以是奥美拉唑(prilosec及zegerid),且可每天一次或两次经口施用10mg、20mg或40mg奥美拉唑。在一些实施方式中,质子泵抑制剂可以是雷贝拉唑(aciphex),且可每天一次或两次经口施用20mg或60mg雷贝拉唑或可每天一次经口施用100mg雷贝拉唑。在本文所公开的任何实施方式中,药剂可彼此组合使用。在本文所公开的任何实施方式中,可在施用药剂之前、期间或之后量测受试者血液、组织和/或微环境ph。在一些实施方式中,向受试者施用或继续施用药剂以提供或降低ph的决定可基于受试者血液、组织和/或微环境ph量测值。用于量测受试者血液ph和/或血液的碳酸氢盐含量方法为本领域中熟知的。在一些实施方式中,可使用正电子发射断层摄影法(pet)、磁共振光谱(mrs)、磁共振成像(mri)及光学成像来量测微环境中体内ph,例如肿瘤中(关于测量肿瘤ph的细节,参见:zhangx,liny,gilliesrj.tumorphanditsmeasurement.jnuclmed.2010年8月;51(8):1167-70)。在另一方面中,本文提供一种减轻受试者在靶脱瘤毒性的方法,其包括以下:a.将编码微环境限制的生物嵌合抗原受体(mrb-car)的多核苷酸引入至受试者的t细胞或nk细胞中以产生能够表达mrb-car的t细胞和/或nk细胞;b.将能够表达mrb-car的t细胞和/或nk细胞引入至受试者中,其中t细胞和/或nk细胞在受试者中表达mrb-car;及c.将药剂以充足量施用至受试者以增加血液ph和/或组织ph和/或微环境ph以介导mrb-car与其具有增加的ph血液、组织和/或微环境中的同源抗原结合,由此减轻受试者中的在靶脱瘤(ontargetofftumor)毒性。在引入步骤中,t细胞或nk细胞能够表达mrb-car,是因为其遗传修饰以含有编码mrb-car的核酸。此遗传修饰可以是在通过转染或转导而引入t细胞或nk细胞内载体上存在mrb-car编码序列。在说明性实施方式中,编码mrb-car的核酸经整合至t细胞或nk细胞基因组中。设想本领域中已知的用于将多核苷酸引入t细胞和/或nk细胞的多种方法可与本文提供的方法一起用于包括使用例如ph调节药剂来改变ph以影响mrb-cart细胞或nk细胞与细胞上的其同源抗原结合的方面(在本文中有时称为“ph切换方面”)。通常,载体(在说明性实例中为表达载体)用于递送多核苷酸。此类载体可包括此项技术中已知的用于向t细胞和/或nk细胞递送核酸的多种载体。本发明的说明性方面利用逆转录病毒载体及逆转录病毒颗粒,及(在一些特别说明性实施方式中)慢病毒载体及(在说明性实施方式中)重组慢病毒颗粒。另一些适合的表达载体可用于本文提供的ph切换方面。此类表达载体包括但不限于病毒载体(例如基于以下的病毒载体:痘苗病毒;脊髓灰质炎病毒;腺病毒(参见例如li等人,investopthalmolvissci35:25432549,1994;borras等人,genether6:515524,1999;li及davidson,pnas92:77007704,1995;sakamoto等人,hgenether5:10881097,1999;wo94/12649,wo93/03769;wo93/19191;wo94/28938;wo95/11984andwo95/00655);腺相关病毒(参见例如ali等人,humgenether9:8186,1998,flannery等人,pnas94:69166921,1997;bennett等人,investopthalmolvissci38:28572863,1997;jomary等人,genether4:683690,1997,rolling等人,humgenether10:641648,1999;ali等人,hummolgenet5:591594,1996;wo93/09239中的srivastava,samulski等人,j.vi(1989)63:3822-3828;mendelson等人,virol.(1988)166:154-165;及flotte等人,pnas(1993)90:10613-10617);sv40;单纯疱疹病毒;或逆转录病毒载体(例如,小鼠白血病病毒脾坏死病毒及衍生自例如罗斯肉瘤病毒、哈维肉瘤病毒、禽白血病病毒、人类免疫缺陷病毒、骨髓增生性肉瘤病毒及乳腺肿瘤病毒的逆转录病毒的载体),例如γ逆转录病毒;或人类免疫缺陷病毒(参见例如miyoshi等人,pnas94:1031923,1997;takahashi等人,jvirol73:78127816,1999);及类似者。在一些实施方式中,利用含dna的病毒颗粒代替重组逆转录病毒颗粒。此类病毒颗粒可以是腺病毒、腺相关病毒、疱疹病毒、巨细胞病毒、痘病毒、牛痘病毒、流感病毒、水泡性口炎病毒(vsv),或辛德毕斯病毒。本领域技术人员将理解如何修改本文公开的用于不同病毒及逆转录病毒或逆转录病毒颗粒的方法。在使用包括dna基因组的病毒颗粒时,本领域技术人员将理解,这些基因组中可包括功能单元以诱导将病毒颗粒的全部或部分dna基因组整合至用此类病毒转导的t细胞和/或nk细胞基因组中。替代地,功能性dna可经递送至在细胞中表达但不整合至t细胞和/或nk细胞基因组中的t细胞和/或nk细胞。在说明性实施方式中,在本发明的ph切换方面中使用的载体为重组逆转录病毒颗粒,且在某些实施方式中为重组慢病毒颗粒。此类逆转录病毒颗粒通常包括位于病毒包膜内的衣壳内的逆转录病毒基因组。在本文各个部分中本发明提供重组逆转录病毒颗粒的多种实施方式,其公开可包含于重组逆转录病毒颗粒表面上或在重组逆转录病毒颗粒基因组内和/或中的元件。这些重组逆转录病毒颗粒实施方式中任何一种可用于本文所提供的ph切换方面中。抑制性rna分子在某些实施方式中,本文提供的本发明的方法包括抑制在t细胞和/或nk细胞中表达的一种或多种内源基因的表达。本文提供的方法说明制备重组逆转录病毒颗粒的能力,这些重组逆转录病毒颗粒表达一种或多种(且在说明性实施方式中,两种或更多种)抑制性rna分子,例如可用于此类方法的mirna或shrna。事实上,本文提供的方法说明此类抑制性rna分子可在包括例如ef1a内含子的内含子内经编码。此利用本方法教示以使可包括于可包装逆转录病毒基因组中的功能元件最大化,以克服先前教导的缺点,且使此类重组逆转录病毒颗粒在过继性t细胞疗法中的有效性最大化。在一些实施方式中,抑制性rna分子包括与彼此部分或完全互补的5'链及3'链(在一些实例中,有义链及反义链),使得该两个链能够在细胞环境内形成18至25个核苷酸的rna双链体。5'链的长度可以是18、19、20、21、22、23、24或25个核苷酸,且3'链的长度可以是18、19、20、21、22、23、24或25个核苷酸。5'链及3'链可以是相同或不同的长度,且rna双链体可包括一个或多个错配。替代地,rna双链体不具有错配。包括于本文所提供的组合物及方法中抑制性rna分子在某些说明性实施方式中不存在于和/或不天然表达在其插入其基因组的t细胞中。在一些实施方式中,抑制性rna分子为mirna或shrna。在一些实施方式中,当在本文或优先权申请中提及编码sirna的核酸,尤其在核酸为基因组一部分的上下文中,将理解,此类核酸能够在由dicer处理的细胞中形成例如mirna或shrna的sirna前体,以形成通常与risk复合物相互作用或成为risk复合物一部分的双链rna。在一些实施方式中,本发明的一实施方式中的抑制性分子可以是mirna的前体(例如pri-mirna或pre-mirna),或shrna的前体。在一些实施方式中,mirna或shrna为人工衍生的(即人工mirna或sirna)。在其他实施方式中,抑制性rna分子为经处理成sirna的dsrna(经转录或人工引入)或sirna本身。在一些实施方式中,mirna或shrna具有在自然界中未发现的序列,或具有在自然界中未发现的至少一个功能区段,或具有在自然界中未发现的功能区段的组合。在一些实施方式中,抑制性rna分子以一连串或多重排列方式置于第一核酸分子中,使得多个mirna序列同时自单个多顺反子mirna转录物表达。在一些实施方式中,抑制性rna分子可利用非功能性接头序列直接或间接地与彼此连接。在一些实施方式中,接头序列的长度可在5个核苷酸与120个核苷酸之间,且在一些实施方式中可在10个核苷酸与40个核苷酸之间,作为非限制性实例。在说明性实施方式中,编码一种或多种(例如两种或更多种)抑制性rna的第一核酸序列及编码car(例如mrb-car)的第二核酸序列可操作地连接至具有组成型活性或可在t细胞或nk细胞中诱导的启动子。因此,这些抑制性rna分子(例如mirna)以及car以多顺反子的方式表达。另外,功能性序列可自同一转录物表达。举例而言,本文所提供的并非抑制性rna分子的任何淋巴增殖性元件可自与car及一种或多种(例如两种或更多种)抑制性rna分子相同的转录物表达。在一些实施方式中,抑制性rna分子为天然存在的mirna,例如但不限于mir-155。替代地,可产生人造mirna,其中能够形成杂交/互补的茎结构且针对靶标rna的序列置于包括用于微小rna处理微小rna侧翼序列及环的mirna框架中,且可任选地衍生自与茎序列之间的侧翼序列相同的天然存在的mirna。因此,在一些实施方式中,抑制性rna分子自5'至3'定向包括:5'微小rna侧翼序列、5'茎、环、与该5'茎部分或完全互补的3'茎,及3'微小rna侧翼序列。在一些实施方式中,5'茎(本文中也称为5'臂)长度可以是18、19、20、21、22、23、24或25个核苷酸。在一些实施方式中,3'茎(本文中也称为3'臂)长度可以是18、19、20、21、22、23、24或25个核苷酸。在一些实施方式中,环的长度为3至40、10至40、20至40或20至30个核苷酸,且在说明性实施方式中,环的长度可以是18、19、20、21或22个核苷酸。在一些实施方式中,一个茎比另一茎长两个核苷酸。较长茎可以是5′茎或3′茎。在一些实施方式中,5'微小rna侧翼序列、3'微小rna侧翼序列或两者均衍生自天然存在的mirna,例如但不限于mir-155、mir-30、mir-17-92、mir-122及mir-21。在某些实施方式中,5'微小rna侧翼序列、3'微小rna侧翼序列或两者均衍生自mir-155,例如来自小家鼠或人的mir-155。将合成的mirna茎环插入至mir-155框架(即5'微小rna侧翼序列、3'微小rna侧翼序列及mirna5'茎与3'茎之间的环)为本领域普通技术人员已知的(chung,k.等人2006.nucleicacidsresearch.34(7):e53;us7,387,896)。sibr(合成抑制性bic衍生的rna)序列(chung等人.2006,同前文献)例如具有由小家鼠bic非编码mrna(genbankiday096003.1)的核苷酸134至161(seqidno:256)组成的5'微小rna侧翼序列及由小家鼠bic非编码mrna(genbankiday096003.1)第223至283位核苷酸组成的3'微小rna侧翼序列。在一项研究中,sibr序列经修饰(esibr)以增强mirna表达(fowler,d.k.等人.2015.nucleicacidsresearch44(5):e48)。在本发明的一些实施方式中,mirna可置于sibr或esibrmir-155框架中。在本文说明性实施方式中,mirna置于mir-155框架中,其包括由seqidno:256展示的mir-155的5'微小rna侧翼序列,由seqidno:260展示的3'微小rna侧翼序列(小家鼠bic非编码mrna核苷酸221至265);及经修饰mir-155环(seqidno:258)。因此,在一些实施方式中,mir-155的5'微小rna侧翼序列为seqidno:256或其功能性变体,例如与seqidno:256相同的长度,或为seqidno:256长度的95%、90%、85%、80%、75%或50%或长度为100个核苷酸或更少、95个核苷酸或更少、90个核苷酸或更少、85个核苷酸或更少、80个核苷酸或更少、75个核苷酸或更少、70个核苷酸或更少、65个核苷酸或更少、60个核苷酸或更少、55个核苷酸或更少、50个核苷酸或更少、45个核苷酸或更少、40个核苷酸或更少、35个核苷酸或更少、30个核苷酸或更少,或25个核苷酸或更少;且与seqidno:256至少50%、55%、60%、65%、70%、75%、80%、85%、90%或95%一致的序列。在一些实施方式中,mir-155的3'微小rna侧翼序列为seqidno:260或其功能性变体,例如与seqidno:260相同的长度,或为seqidno:260长度的95%、90%、85%、80%、75%,或50%或长度为100个核苷酸或更少、95个核苷酸或更少、90个核苷酸或更少、85个核苷酸或更少、80个核苷酸或更少、75个核苷酸或更少、70个核苷酸或更少、65个核苷酸或更少、60个核苷酸或更少、55个核苷酸或更少、50个核苷酸或更少、45个核苷酸或更少、40个核苷酸或更少、35个核苷酸或更少、30个核苷酸或更少,或25个核苷酸或更少;且与seqidno:260至少50%、55%、60%、65%、70%、75%、80%、85%、90%或95%一致的序列。然而,作用以在插入其中的mirna的细胞内提供适当处理以形成能够抑制其结合的靶标mrna表达的成熟mirna的任何已知的微小rna框架涵盖于本发明中。在一些实施方式中,由复制缺陷型重组逆转录病毒颗粒的多核苷酸中的核酸序列编码的抑制性rna分子中的至少一者、至少两者、至少三者,或至少四者具有呈5'至3'取向的以下排列:5'微小rna侧翼序列、5'茎、环、与该5'茎部分或完全互补的3'茎,及3'微小rna侧翼序列。在一些实施方式中,所有抑制性rna分子具有呈5'至3'取向的以下排列:5'微小rna侧翼序列、5'茎、环、与该5'茎部分或完全互补的3'茎,及3'微小rna侧翼序列。如本文所公开,抑制性rna分子可由一个或多个接头序列分隔开,其在一些实施方式中没有除在这些抑制性rna分子之间作为间隔子之外的功能。在一些实施方式中,当包括两个或更多个抑制性rna分子(在一些实例中,包括1个、2个、3个、4个、5个、6个、7个、8个、9个或10个抑制性rna分子)时,这些抑制性rna分子针对相同或不同的rna靶标(例如自相关基因转录的mrna)。在说明性实施方式中,第一核酸序列中包括2个与10个之间、2个与8个之间、2个与6个之间、2个与5个之间、3个与5个之间或3个与6个之间的抑制性rna分子。在一说明性实施方式中,第一核酸序列中包括四个抑制性rna分子。在一些实施方式中,rna靶标为自由t细胞表达的基因转录mrna,这些mrna例如但不限于:pd-1(防止失活);ctla4(防止失活);tcra(安全地防止自体免疫);tcrb(安全-防止自体免疫);cd3z(安全-防止自体免疫);socs1(防止失活);smad2(防止失活);mir-155靶标(促进活化);ifnγ(减少crs);ccbl(延长信号传导);trail2(防止死亡);pp2a(延长信号传导);abcg1(通过限制胆固醇的清除来增加胆固醇微区(microdomain)含量)。在说明性实例中,在本文提供的方法中插入至t细胞至基因组中的mirna针对靶标,使得t细胞增殖经诱导且/或增强且/或凋亡被抑制。在一些实施方式中,rna靶标包括编码t细胞受体(tcr)复合物组份的mrna。此类组分可包括用于产生和/或形成t细胞受体复合物组分和/或用于t细胞受体复合物的恰当功能组分。因此,在一个实施方式中,两种或更多种抑制性rna分子中的至少一者使得tcr复合物(在说明性实施方式中,t细胞的一种或多种内源性tcr复合物)的形成和/或功能降低。t细胞受体复合物包括tcra、tcrb、cd3d、cd3e、cd3g及cd3z。已知这些组分具有复杂的相互依赖性,使得任何一个亚单位表达的降低将导致复合物的表达及功能降低。因此,在一个实施方式中,rna靶标为表达与经转导t细胞内源性的tcra、tcrb、cd3d、cd3e、cd3g及cd3z中一种或多种mrna。在某些实施方式中,rna靶标为自t细胞的内源性tcrα或tcrβ基因转录mrna,该t细胞基因组包含编码一个或多个mirna的第一核酸序列。在说明性实施方式中,rna靶标为自tcrα基因转录mrna。在某些实施方式中,可将针对自具有相似预期效用的靶基因转录的mrna的抑制性rna分子组合。在其他实施方式中,可将针对自具有互补效用的靶基因转录的靶标mrna的抑制性rna分子组合。在一些实施方式中,两种或更多种抑制性rna分子针对自靶基因cd3z、pd1、socs1和/或ifnγ转录mrna。在本文提供的一些实施方式中,两种或更多种抑制性rna分子可在单个内含子中递送,例如但不限于ef1-αa内含子a。可用于携带本发明的mirna的内含子序列包括在t细胞内处理的任何内含子。如本文所示,此类排列的一个优势在于,此有助于以本文提供的方法使mirna序列包括于用于将此类序列递送至t细胞的逆转录病毒基因组大小内的能力最大化。当第一核酸序列的内含子包括用以以多顺反子方式表达内含子、car序列及本文所提供的其他功能性序列的启动子序列的全部或部分(例如并非抑制性rna分子的淋巴增殖性元件)时,尤其如此。内含子的序列要求为此项技术中已知的。在一些实施方式中,此类内含子处理可操作地连接至核糖开关,例如本文所公开的任何核糖开关。因此,核糖开关可提供用于控制第一核酸序列上的一个或多个mirna序列表达的调节元件。因此,本文所提供的说明性实施方式为针对内源性t细胞受体亚单位的mirna的组合,其中mirna的表达由核糖开关调节,该核糖开关可以是本文所论述的任何核糖开关。在一些实施方式中,抑制性rna分子可提供于可包括于相同或不同转录单元上的多个核酸序列上。举例而言,第一核酸序列可编码一种或多种抑制性rna分子,且自第一启动子表达,且第二核酸序列可编码一种或多种抑制性rna分子且自第二启动子表达。在说明性实施方式中,两种或更多种抑制性rna分子位于自单个启动子表达的第一核酸序列上。用于表达此类mirna的启动子通常为在用以表达逆转录病毒颗粒的包装细胞中无活性的启动子,该逆转录病毒颗粒将其基因组中的mirna递送至靶标t细胞,但此类启动子在t细胞内为组成型活化或呈可诱导方式的活性。启动子可以是poli、polii,或poliii启动子。在一些说明性实施方式中,启动子为polii启动子。治疗方法本公开提供使用car的多种治疗方法。本发明的car在存在与t淋巴细胞或nk细胞中时,可介导对靶细胞的细胞毒性。本发明的car结合靶细胞上存在抗原,从而通过遗传修饰以产生car的t淋巴细胞或nk细胞介导靶细胞杀伤。car的astr结合存在于靶细胞表面上的抗原。本公开提供杀伤或抑制靶细胞生长的方法,该方法包括使遗传修饰以产生受试者car的细胞毒性免疫效应细胞(例如,细胞毒性t细胞或nk细胞)接触,使得t淋巴细胞或nk细胞识别存在于靶细胞表面上的抗原,且介导对靶细胞杀伤。本公开提供治疗患有疾病或病症的受试者的疾病或病症的方法,该方法包括:a.将包括编码car的多核苷酸序列的表达载体引入至自受试者获得的外周血细胞中以产生经基因经改造细胞毒性细胞;及b.向受试者施用该经基因经改造细胞毒性细胞。适于治疗的受试者多种受试者适用于通过本发明的方法及组合物治疗。适合的受试者包括患有疾病或病症、经诊断患有疾病或病症、处于患有疾病或病症的风险、曾患有疾病或病症且处于疾病或病症复发的风险、已通过用于该疾病或病症的药物治疗且未能对此治疗作出反应、或已通过用于该疾病或病症的药物治疗但在对此治疗初步反应之后复发的任何受试者,例如人或非人类动物。适用于通过免疫调节方法治疗的受试者包括具有自身免疫疾病的受试者;为器官或组织移植接受者及类似者的受试者;免疫缺陷受试者;及感染病原体的受试者。示示例例性性实实施施方方式式在一个方面中,本文中提供一种用于遗传修饰及扩增受试者淋巴细胞的方法,其包含:a.使受试者静息t细胞和/或nk细胞在体外在不需要进行预先体外刺激的情况下与复制缺陷型重组逆转录病毒颗粒接触,这些复制缺陷型重组逆转录病毒颗粒包含:iii.在其表面上的假型化元件,该假型化元件能够与t细胞和/或nk细胞结合且有助于复制缺陷型重组逆转录病毒颗粒在其上的膜融合;及iv.多核苷酸,其包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码由控制元件调节的第一经改造信号传导多肽,其中该第一经改造信号传导多肽包含至少一个淋巴增殖性元件,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导静息t细胞和/或nk细胞中的至少一些,从而产生遗传修饰的t细胞和/或nk细胞;b.将遗传修饰的t细胞和/或nk细胞引入至受试者中;及c.将经遗传修饰的t细胞和/或nk细胞在体内暴露于结合控制元件的化合物,以影响第一经改造信号传导多肽的表达且体内促进和/或强化淋巴细胞的扩增、移植和/或持久性,由此遗传修饰及扩增受试者的淋巴细胞。在说明性实施方式中,在无需体外刺激的情况下进行转导。在以上方面及用于遗传修饰及扩增淋巴细胞或用于执行本文的细胞治疗的方法方面中的任何一种,若在最广泛的方面中未叙述,则在某些实施方式中,多核苷酸进一步包含编码包含第一嵌合抗原受体的第二经改造信号传导多肽的转录单元,该第一嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域。在另一方面中,本文提供用于对受试者执行过继性细胞疗法的方法,该方法包含:a.自受试者采集血液;b.使来自受试者血液的静息t细胞及/nk细胞在体外与复制缺陷型重组逆转录病毒颗粒接触,其中该复制缺陷型重组逆转录病毒颗粒包含:i.在其表面上的假型化元件,该假型化元件能够与t细胞和/或nk细胞结合且有助于复制缺陷型重组逆转录病毒颗粒至其上的膜融合;及ii.多核苷酸,其包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码包含至少一个淋巴增殖性元件(其表达由控制元件调节)的第一经改造信号传导多肽,及包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域的嵌合抗原受体的第二经改造信号传导多肽,其中该接触产生变为遗传修饰的静息t细胞和/或nk细胞中的至少一些;及c.将遗传修饰的t细胞和/或nk细胞重新引入至受试者中,其中遗传修饰的t细胞和/或nk细胞扩增、移植和/或持久性出现于受试者体内,且其中在采集血液与重新引入遗传修饰的t细胞和/或nk细胞之间的方法在不超过24小时内进行,从而对受试者执行过继性细胞疗法。本文的另一方面提供用于对受试者执行过继性细胞疗法的方法,该方法包含:a.自受试者采集血液;b.分离包含静息t细胞和/或静息nk细胞的外周血单核细胞(pbmc);c.使受试者静息t细胞和/或静息nk细胞在体外与复制缺陷型重组逆转录病毒颗粒接触,其中复制缺陷型重组逆转录病毒颗粒在其表面上包含能够结合静息t细胞和/或nk细胞且有助于复制缺陷型重组逆转录病毒颗粒至其上的膜融合的假型化元件,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导静息t细胞和/或nk细胞,从而产生遗传修饰的t细胞和/或nk细胞;及d.在自受试者采集血液24小时内,将遗传修饰的细胞重新引入至受试者中,从而在受试者中执行过继性细胞疗法。在本文另一方面中提供转导受试者静息淋巴细胞的方法,该方法包含使受试者静息t细胞和/或静息nk细胞在体外与复制缺陷型重组逆转录病毒颗粒接触,其中复制缺陷型重组逆转录病毒颗粒在其表面上包含能够结合静息t细胞和/或nk细胞且有助于复制缺陷型重组逆转录病毒颗粒至其上的膜融合的假型化元件,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导静息t细胞和/或nk细胞,从而产生遗传修饰的t细胞和/或nk细胞。在此方面的说明性实施方式中,至少10%、20%或25%的静息t细胞和/或nk细胞,或10%与70%之间,或20%与50%之间的t细胞和/或nk细胞经转导,作为处理经转导结果,作为处理结果。在本文的另一方面中提供一种用于转导来自分离血液的静息t细胞和/或静息nk细胞的方法,其包含:a.自受试者采集血液;b.分离包含静息t细胞和/或静息nk细胞的外周血单核细胞(pbmc);c.使受试者的静息t细胞和/或静息nk细胞在体外与复制缺陷型重组逆转录病毒颗粒接触,其中复制缺陷型重组逆转录病毒颗粒在其表面上包含能够结合静息t细胞和/或静息nk细胞且有助于复制缺陷型重组逆转录病毒颗粒至其上的膜融合的假型化元件,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导至少5%静息t细胞和/或静息nk细胞,从而产生遗传修饰的t细胞和/或nk细胞,从而转导静息t细胞和/或nk细胞。在一个方面中,本发明提供复制缺陷型重组逆转录病毒颗粒,其包含:a.一个或多个假型化元件,其能够与t细胞和/或nk细胞结合且有助于复制缺陷型重组逆转录病毒颗粒至其上的膜融合;及b.多核苷酸,其包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码包含嵌合抗原受体(其包含抗原特异性靶向区、跨膜结构域及胞内活化结构域)的第一经改造信号传导多肽;及包含至少一个淋巴增殖性元件的第二经改造信号传导多肽;其中该第一经改造信号传导多肽和/或该第二经改造信号传导多肽的表达由控制元件调节;及c.其表面上的活化元件,其中该活化元件能够结合t细胞和/或nk细胞,且不由复制缺陷型重组逆转录病毒颗粒中的多核苷酸编码。在另一方面中,本文提供复制缺陷型重组逆转录病毒颗粒,每一者包含:a.在其表面上的假型化元件,该假型化元件能够与t细胞和/或nk细胞结合且有助于复制缺陷型重组逆转录病毒颗粒至其上的膜融合,其中该假型化元件包含麻疹病毒f多肽和/或麻疹病毒h多肽的细胞质结构域缺失变体;b.多核苷酸,其包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码包含嵌合抗原受体(其包含抗原特异性靶向区、跨膜结构域及胞内活化结构域)的第一经改造信号传导多肽,及包含组成型活化il-7受体突变体的第二经改造信号传导多肽;其中il-7受体突变体表达由结合核苷类似抗病毒药物的核糖开关调节;及c.能够与cd3结合的多肽及能够与cd28结合的多肽,其中这些多肽在复制缺陷型重组逆转录病毒颗粒表面上表达;能够与t细胞和/或nk细胞结合;且不由复制缺陷型重组逆转录病毒颗粒中的多核苷酸编码。在此方面的说明性实施方式中,核苷类似抗病毒药物与核糖开关结合增加il-7受体突变体表达。在任何本文所提供的方法或组合物方面中,若在最广泛的方面中未叙述,则一个或多个复制缺陷型重组逆转录病毒颗粒包含或进一步包含其表面上的能够活化静息t细胞和/或静息nk细胞的活化元件。在本文叙述t细胞和/或nk细胞,或静息t细胞或静息nk细胞的任何方法或组合物中,在某些说明性实施方式中,该细胞为t细胞。通常,本文所提供的任何方法及组合物中的重组逆转录病毒颗粒为复制缺陷型的,即无法复制。在说明性实施方式中,逆转录病毒为慢病毒,例如复制缺陷的hiv慢病毒。在说明性实施方式中,逆转录病毒颗粒为慢病毒颗粒,例如复制缺陷的hiv慢病毒颗粒。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法或类似方法的任何方法方面中的说明性实施方式中,10%与75%之间、或10%与70%之间、或10%与60%之间、或10%与50%之间、或10%与25%之间、或20%与75%之间、或20%与50%之间,或至少10%、20%或25%的静息t细胞经转导且0%与75%之间的nk细胞经转导。在其他实施方式中,5%与80%之间、或10%与80%之间、或10%与70%之间、或10%与60%之间、或10%与50%之间,或10%与25%之间,或10%与20%之间、或20%与50%的静息nk细胞经转导。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法,或本文所提供的类似方法或任何组合物的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则该第二经改造信号传导多肽的表达由控制元件调节。在用于遗传修饰及扩增淋巴细胞或用于执行过继性细胞疗法或类似方法的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则接触可在范围下限为15分钟、30分钟或45分钟或1小时、2小时、3小时、4小时、5小时、6小时、7小时或8小时,与范围上限为6小时、8小时、10小时、12小时、18小时、24小时、36小时、48小时及72小时之间进行。举例而言,在说明性实施方式中,接触进行2小时与24小时之间,或4小时与12小时之间,或4小时与8小时之间。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法或类似方法的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则该方法可进一步包含将遗传修饰的t细胞和/或nk细胞在体内暴露于结合控制元件的化合物,以影响第一经改造信号传导多肽及任选地第二经改造信号传导多肽表达,且促进体内淋巴细胞的扩增、移植和/或持久性。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法或类似方法的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则遗传修饰的t细胞和/或nk细胞在经引入或重新引入至受试者之前在体外经历8次、7次、6次、5次、4次、3次或更少的细胞分裂。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法或类似方法的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则经基因发生修饰的t细胞和/或nk细胞在体内扩增、移植和/或持久性取决于结合控制元件化合物的存在或不存在,且在说明性实施方式中取决于结合控制元件化合物的存在。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法或类似方法的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则受试者在执行接触7天、14天或21天内或在接触期间或在将经修饰的t细胞和/或nk细胞引入至受试者中之后7天、14天或21天内未暴露于淋巴细胞清除剂。在其他实施方式中,受试者在接触期间未暴露于淋巴细胞清除剂。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法或类似方法的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则静息t细胞和/或静息nk细胞与复制缺陷型重组逆转录病毒颗粒接触15分钟与12小时之间。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法或类似方法的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则该方法进一步包括在接触之后但在引入之前自t细胞和/或nk细胞分离复制缺陷型重组逆转录病毒颗粒的步骤。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法或类似方法的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则该暴露步骤包括在接触之前或期间,和/或在已将遗传修饰的t细胞和/或nk细胞引入至受试者中之后向受试者施用一定剂量化合物。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法或类似方法的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则该方法包含在使t细胞和/或nk细胞在体外与复制缺陷型重组逆转录病毒颗粒接触之前自受试者收集包含t细胞和/或nk细胞的血液,且其中该引入为重新引入。举例而言,自受试者抽取20ml与250ml之间的血液。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法或类似方法的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则在自受试者采集血液的时间与将经修饰的t细胞和/或nk细胞重新引入至受试者中的时间之间不超过8小时、12小时、24小时或48小时。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法或类似方法的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则在自受试者采集血液的时间与将经修饰的t细胞和/或nk细胞重新引入至受试者中的时间之间在下限为4小时或8小时与范围上限为12小时、24小时、36小时或48小时之间。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法或类似方法的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则在采集血液之后且在重新引入血液之前的所有步骤都在一个封闭系统中进行,在该系统中一人在整个处理过程中监控该封闭系统。在另一实施方式中,在采集血液之后及在重新引入血液之前,在与受试者保持在同一房间中的封闭系统中进行。在用于遗传修饰及扩增淋巴细胞或用于执行本文的过继性细胞疗法或类似方法的任何方法方面的说明性实施方式中,若在最广泛方面中未明确地叙述,则这些经改造信号传导多肽中的一种包含或进一步包含抗原特异性靶向区(astr)及将astr连接至淋巴增殖性元件的跨膜结构域。此经改造信号传导多肽的astr能够结合第一肿瘤抗原,且当存在时,第二经改造信号传导多肽的astr能够结合第二肿瘤抗原。在某些实施方式中,第一经改造信号传导多肽和/或第二经改造信号传导多肽进一步包含共刺激结构域。此外,第一经改造信号传导多肽和/或第二经改造信号传导多肽进一步包含柄。此外,第一经改造信号传导多肽进一步包含胞内活化结构域。第一经改造信号传导多肽和/或第二经改造信号传导多肽上的胞内活化结构域可衍生自cd3ζ。在本文提供的包括淋巴增殖性元件的任何方法及组合物的说明性实施方式中,淋巴增殖性元件可包含t细胞存活基序。t细胞存活基序可包含il-7受体、il-15受体或cd28的所有片段或功能性片段。在其他实施方式中,淋巴增殖性元件可包括细胞因子或细胞因子受体多肽,或其包含信号传导结构域片段。举例而言,淋巴增殖性元件可包含经由接头共价附接至其同源白介素受体多肽的白介素多肽。替代地,淋巴增殖性元件可以是il-7受体的胞内信号传导结构域、il-12受体的胞内信号传导结构域,il-23的胞内信号传导结构域、il-27的胞内信号传导结构域、il-15受体的胞内信号传导结构域、il-21受体的胞内信号传导结构域或转化生长因子β(tgfβ)诱饵受体的胞内信号传导结构域。在其他说明性实施方式中,淋巴增殖性元件具有组成型活化。此外,淋巴增殖性元件可包括突变的il-7受体或其片段,其可以进一步包括组成型活化突变的il-7受体或其组成型活化片段。在本文提供的包括复制缺陷型重组逆转录病毒颗粒的任何方法及组合物的说明性实施方式中,若在最广泛方面中未明确地叙述,则复制缺陷型重组逆转录病毒颗粒可在其表面上包含活化元件,该活化元件包含:a.能够与cd3结合的膜结合多肽;和/或b.能够结合cd28的膜结合多肽。此外,能够与cd3结合的膜结合多肽为能够与融合至异源gpi锚连接序列cd3结合的多肽,且能够与cd28结合的膜结合多肽可以是能够与融合至异源gpi锚连接序列cd28结合的多肽。在一些实施方式中,能够与cd28结合的膜结合多肽为cd80、cd86或能够诱导对akt(例如cd80的胞外结构域)经cd28介导活化的其功能性片段。在本文提供的包括复制缺陷型重组逆转录病毒颗粒的任何方法及组合物的说明性实施方式中,能够结合cd3的膜结合多肽可以是与cd14gpi锚连接序列结合的抗cd3scfv,且能够结合cd28的膜结合多肽可以是与cd16bgpi锚连接序列结合的cd80或其胞外结构域。在本文提供的包括复制缺陷型重组逆转录病毒颗粒的任何方法及组合物的说明性实施方式中,复制缺陷型重组逆转录病毒颗粒可在其表面上包含结合cd14gpi锚连接序列的抗cd3scfv,或结合cd16bgpi锚连接序列的cd80或其胞外结构域,及il-7的融合多肽或其活化片段,及包含gpi锚连接序列的daf。在本文提供的包括复制缺陷型重组逆转录病毒颗粒的任何方法及组合物的说明性实施方式中,il-7或其活化片段,及daf融合体、抗cd3scfv及cd80及其胞外结构域各包含daf信号序列。在本文提供的包括复制缺陷型重组逆转录病毒颗粒的任何方法及组合物的说明性实施方式中,若在最广泛方面中未明确地叙述,则复制缺陷型重组逆转录病毒颗粒可在其表面上包含膜结合细胞因子。膜结合细胞因子可以是il-7、il-15或其活化片段。在其他实施方式中,膜结合细胞因子为il-7的融合多肽或其活化片段及daf。举例而言,融合多肽可包含daf信号序列(seqidno:286的核苷酸1至34),无其信号序列(seqidno:286的核苷酸35至186)的il-7及包括其gpi锚连接序列(seqidno:286的核苷酸187至532)的daf片段。在本文提供的方法及组合物方面中的任何说明性实施方式中,假型化元件可包含一种或多种异源包膜蛋白质。在其他实例中,假型化元件可包括由t细胞识别的一种或多种病毒多肽。一个或多个假型化元件可包含麻疹病毒f多肽、麻疹病毒h多肽和/或其片段。该一个或多个假型化元件可以是麻疹病毒f多肽和/或麻疹病毒h多肽的细胞质结构域缺失变体。在本文提供的包括控制元件的任何方法及组合物的说明性实施方式中,该控制元件为可调节淋巴增殖性元件的控制元件,其中淋巴增殖性元件在促进t细胞和/或nk细胞增殖中在不存在化合物的情况下无活性或具有较低活性,且其中该化合物为结合淋巴增殖性元件且诱导淋巴增殖性元件活性的分子伴侣。在本文提供的包括控制元件的任何方法及组合物的说明性实施方式中,控制元件可以是包含核糖开关的多核苷酸。核糖开关可能够结合核苷类似物且结合控制元件的化合物为核苷类似物。核苷类似物可以是抗病毒剂。抗病毒剂可以是阿昔洛韦或喷昔洛韦。在本文提供的包括经改造信号传导多肽(其包括astr)方法及组合物中的任何说明性实施方式中,这些经改造信号重叠多肽中的一种或两种的astr可结合肿瘤相关抗原。在一些说明性实施方式中,第二经改造多肽的抗原特异性靶向区为微环境限制的抗原特异性靶向区。在本文提供的包括一个或多个复制缺陷型重组逆转录病毒颗粒的方法及组合物的任何说明性实施方式中,若在最广泛方面中未明确地叙述,则复制缺陷型重组逆转录病毒颗粒可编码经生物验证的单克隆抗体识别结构域。在一些实施方式中,识别结构域在与嵌合抗原受体相同的转录物上表达,且其中通过核糖体跳跃和/或切割信号将识别结构域与嵌合抗原受体分离。核糖体跳跃和/或切割信号可以是2a-1。识别结构域可包括由识别egfr的抗体识别的多肽或其表位。识别结构域可以是由egfr抗体识别且在经转导的t细胞和/或nk细胞表面上表达为本文提供的另一控制机制的egfr突变体。在相关实施方式中,识别结构域可包括由识别egfr抗体识别的多肽或其表位。在本文提供的包括淋巴增殖性元件的任何方法或组合物中,淋巴增殖性元件可包括刺激stat5途径或抑制socs途径的抑制性rna分子,例如mirna或shrna。举例而言,抑制性rna分子可结合编码选自由以下群蛋白质的核酸:abcg1、socs、tgfbr2、smad2、ccbl及pd1。在本文提供的复制缺陷型重组逆转录病毒颗粒或转导细胞中的任一种或包括其方法的说明性实施方式中,此类复制缺陷型重组逆转录病毒颗粒或转导细胞可编码两种或更多种抑制性rna分子,例如内含子中的mirna或shrna,在一些实施方式中为1种、2种、3种或4种抑制性rna分子,其结合编码以下目标内源性t细胞表达的基因中的一种或多种的核酸:pd-1、ctla4、tcrα、tcrβ、cd3ζ、socs、smad2、mir-155、ifnγ、ccbl、trail2、pp2a或abcg1。举例而言,在一个实施方式中,靶向以下任一种mirna的组合可包括在复制缺陷型重组逆转录病毒颗粒或转导细胞基因组中:tcrα、cd3ζ、ifnγ,及pd-1;且在另一实施方式中为socs1、ifnγ、tcrα及cd3ζ。在本文提供的任何方法及组合物的说明性实施方式中,复制缺陷型重组逆转录病毒颗粒、哺乳动物细胞和/或包装细胞可包含vpx多肽。vpx多肽可以是例如融合多肽,且在一些实例中,尤其在包装细胞中,为膜结合vpx多肽。在本文所提供的任何方法及组合物中,一个或多个假型化元件包括水疱性口炎病毒包膜蛋白(vsv-g)、猫类内源性病毒(rd114)包膜蛋白、致癌逆转录病毒双嗜性包膜蛋白或致癌逆转录病毒亲嗜性包膜蛋白或其功能性片段。在另一方面中,本文提供遗传修饰的t细胞和/或nk细胞,其包含:a.第一经改造信号传导多肽,其包含至少一个淋巴增殖性元件;及b.第二经改造信号传导多肽,其包含嵌合抗原受体,该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域。在本文提供的任何包括哺乳动物包装细胞的方法(包括复制缺陷型重组逆转录病毒颗粒包装系统方面,或任何用于制备复制缺陷型重组逆转录病毒颗粒的方法)中,例如,可包装rna基因组由可操作地连接至启动子的多核苷酸编码,其中该启动子具有组成型活化或可由第一反式活化子或第二反式活化子诱导。可包装rna基因组可由可操作地连接至启动子的多核苷酸编码,其中该启动子可由第二反式活化子诱导。本文中用于驱动第一和/或第二经改造信号传导多肽表达的启动子通常在靶细胞(例如淋巴细胞、pbl、t细胞和/或nk细胞)中具有活性,但在说明性实施方式中,在包装细胞系中不具有活性。第二反式活化子可调节能够结合并活化靶细胞的活化元件的表达。在本文提供的任何包括哺乳动物包装细胞的方法(包括复制缺陷型重组逆转录病毒颗粒包装系统方面,或任何用于制备复制缺陷型重组逆转录病毒颗粒(例如,一些实施方式中可包装rna基因组)的方法)中,可包装rna基因组表达可由第二反式活化子调节。此外,可包装rna基因组自5′至3′包含:1.)5′长末端重复或其活化片段;2.)核酸序列,其编码逆转录病毒顺式作用rna包装元件;3.)编码第一靶标多肽的核酸序列和/或编码一种或多种(例如两种或更多种)抑制性rna分子的核酸序列;4.)在靶细胞中具活性启动子;及5.)3′长末端重复或其活化片段。在一些实施方式中,编码第一靶标多肽的核酸序列与编码用于包装及组装的逆转录病毒组分的rna及5'ltr呈相反定向。在本文提供的任何包括哺乳动物包装细胞的方法(包括复制缺陷型重组逆转录病毒颗粒包装系统方面,或任何用于制备复制缺陷型重组逆转录病毒颗粒的方法)中,第一靶标多肽包含第一工程信号传导多肽且其中该第一经改造信号传导多肽包含至少一个淋巴增殖性元件。可包装rna基因组可进一步包含编码第二靶标多肽的核酸序列。第二靶标多肽可包含包括嵌合抗原受体的第二经改造信号传导多肽,该嵌合抗原受体包含:1.)第一抗原特异性靶向区;2.)第一跨膜结构域;及3.)第一胞内活化结构域。在本文提供的任何包括哺乳动物包装细胞的方法(包括复制缺陷型重组逆转录病毒颗粒包装系统方面,或任何用于制备复制缺陷型重组逆转录病毒颗粒(例如哺乳动物细胞)的方法)中,例如,包装细胞可包括例如在第二转录单元或任选地第三转录单元上或在可操作地连接至第一可诱导启动子的其他转录单元上编码vpx的核酸序列。可以是包装细胞的哺乳动物细胞可以是293细胞。在本文提供的任何包括哺乳动物包装细胞的方法(包括复制缺陷型重组逆转录病毒颗粒包装系统方面,或用于制备复制缺陷型重组逆转录病毒颗粒的方法)中,第一配体可以是雷帕霉素,且第二配体可以是四环素或阿霉素,或第一配体可以是四环素或阿霉素且第二配体可以是雷帕霉素。在一些方面中,本文提供已通过本文提供的任何复制缺陷型重组逆转录病毒颗粒转导的细胞。该细胞可以是例如淋巴细胞,例如t细胞或nk细胞。在说明性实施方式中,该细胞为人类细胞。在一个方面中,本文提供用于扩增受试者中经修饰的t细胞和/或nk细胞的方法,该方法包含:a.使自该受试者获得的经分离静息t细胞和/或静息nk细胞与本文所公开实施方式中的任何复制缺陷型重组逆转录病毒颗粒接触;b.将遗传修饰的静息t细胞和/或nk细胞引入至受试者中;及c.向该受试者提供有效量的阿昔洛韦、阿昔洛韦前药、喷昔洛韦或喷昔洛韦前药,其中该经修饰的t细胞和/或nk细胞在施用阿昔洛韦、阿昔洛韦前药、喷昔洛韦或喷昔洛韦前药后在该受试者中增殖,从而扩增受试者中经修饰的t细胞和/或nk细胞。在另一方面中,本文提供用于停止受试者中经修饰的t细胞和/或nk细胞扩增、移植和/或持久性的方法,该方法包含:a.使自该受试者获得的经分离静息t细胞和/或nk细胞与本文所公开实施方式中任何复制缺陷型重组逆转录病毒颗粒接触;b.将经修饰的t细胞和/或nk细胞引入至受试者中;c.向该受试者施用有效量的阿昔洛韦、阿昔洛韦前药、喷昔洛韦或喷昔洛韦前药以扩增受试者中经修饰t细胞和/或nk细胞,其中该经修饰t细胞和/或nk细胞在施用阿昔洛韦、阿昔洛韦前药、喷昔洛韦、喷昔洛韦前药后在该受试者中增殖,从而扩增受试者中经修饰pbl;及d.停止向该受试者施用阿昔洛韦、阿昔洛韦前药、喷昔洛韦或喷昔洛韦前药,其中该经修饰t细胞和/或nk细胞在停止施用阿昔洛韦、阿昔洛韦前药、喷昔洛韦或喷昔洛韦前药后在该受试者中停止增殖,从而控制受试者中经修饰t细胞和/或nk细胞的扩增、移植和/或持久性。在另一方面中,本文中提供一种治疗受试者癌症的方法,该方法包含:a.使自该受试者获得的经分离静息t细胞和/或nk细胞与根据本文所公开实施方式中任何复制缺陷型重组逆转录病毒颗粒接触;b.将遗传修饰t细胞和/或nk细胞引入至受试者中;及c.向该受试者施用有效量的阿昔洛韦、阿昔洛韦前药、喷昔洛韦或喷昔洛韦前药以扩增受试者中经修饰t细胞和/或nk细胞,其中该经修饰t细胞和/或nk细胞在施用阿昔洛韦、阿昔洛韦前药、喷昔洛韦、喷昔洛韦前药后在该受试者中增殖,且其中该经修饰t细胞和/或nk细胞中的嵌合抗原受体结合受试者中的癌细胞,从而治疗受试者癌症。在另一方面中,本文中提供一种经转导t细胞和/或nk细胞,其包含重组多核苷酸,该重组多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中一个或多个转录单元编码由控制元件调节的第一经改造信号传导多肽,其中该第一经改造信号传导多肽包含组成型活化il-7受体突变体,且其中控制元件能够体内与化合物结合,和/或经设计和/或经组态与化合物结合。在另一方面中,本文中提供一种逆转录病毒包装系统,其包含:哺乳动物细胞,其包含:a.第一反式活化子,其由组成型启动子表达且能够结合第一配体及第一可诱导启动子以供在第一配体存在对比于缺失下影响可操作地连接至其核酸序列的表达;b.第二反式活化子,其能够结合第二配体及第二可诱导启动子,且在第二配体存在下对比于缺失下影响可操作地连接至其核酸序列的表达;及c.针对逆转录病毒颗粒的可包装rna基因组,其中第一反式活化子调节第二反式活化子及逆转录病毒rev蛋白质的表达,其中第二反式活化子调节gag多肽、pol多肽及能够与靶标细胞结合且有助于与其膜融合的一个或多个假型化元件的表达,且其中逆转录病毒蛋白质衍生自逆转录病毒。此方面的实施方式可包括针对其他方面中所述实施方式在本文中提供的任何实施方式中。在另一方面中,本文提供一种用于制造复制缺陷型重组逆转录病毒颗粒的方法,其包含:a.培养包装细胞群体以累积第一反式活化子,其中包装细胞包含由第一组成型启动子表达的第一反式活化子,其中第一反式活化子能够结合第一配体及第一可诱导启动子以供在第一配体存在下对比于缺失下影响可操作地连接至其核酸序列的表达,且其中由第一反式活化子调节第二反式活化子及逆转录病毒rev蛋白质的表达;b.在第一配体存在下孵育包含经累积第一反式活化子的包装细胞群体以累积第二反式活化子及逆转录病毒rev蛋白质,其中第二反式活化子能够结合第二配体及第二可诱导启动子以供在第二配体存在下对比于缺失下影响可操作地连接至其核酸序列的表达;及c.在第二配体存在下孵育包含经累积第二反式活化子及逆转录病毒rev蛋白质包装细胞群体,从而诱导gag多肽、pol多肽及一个或多个假型化元件的表达,从而制造复制缺陷型重组逆转录病毒颗粒,其中可包装rna基因组是由可操作地连接至第三启动子的多核苷酸编码,其中该第三启动子是组成型活化的或可由第一反式活化子或第二反式活化子诱导,且其中一个或多个假型化元件能够与靶标细胞结合和/或有助于膜与复制缺陷型重组逆转录病毒颗粒融合。在本文所提供的逆转录病毒包装系统及用于制造复制缺陷型重组逆转录病毒颗粒的方法的一些实施方式中,哺乳动物细胞进一步包含能够与靶标细胞结合且活化靶标细胞的活化元件,且第一反式活化子调节活化元件的表达。活化元件在复制缺陷型重组逆转录病毒颗粒表面上且其中该活化元件可包括:能够与cd3结合的膜结合多肽;和/或能够与cd28结合的膜结合多肽。能够与cd3结合的膜结合多肽为能够与融合至异源gpi锚连接序列cd3结合的多肽,且能够与cd28结合的膜结合多肽为能够与融合至异源gpi锚连接序列cd28结合的多肽。能够与cd28结合的膜结合多肽在一些实施方式中包含cd80、cd86或能够诱导经cd28介导akt的活化的其功能性片段(例如cd80胞外结构域)。在其他实施方式中,能够结合cd3的膜结合多肽为与cd14gpi锚连接序列结合的抗cd3scfvfc或抗cd3scfvfc,且其中能够与cd28结合的膜结合多肽为cd80,或与cd16bgpi锚连接序列结合的其胞外片段。在本文所提供的逆转录病毒包装系统及用于制造复制缺陷型重组逆转录病毒颗粒方面方法的一些实施方式中,哺乳动物细胞进一步包含膜结合细胞因子,且第一反式活化子调节膜结合细胞因子表达。膜结合细胞因子可以是例如il-7、il-15或其活化片段。实施方式中的膜结合细胞因子可以是il-7或其活化片段及daf的融合多肽。举例而言,融合多肽可包含daf信号序列及不具有其信号序列的il-7,接着为daf的残基36至525。在本文所提供的逆转录病毒包装系统及用于制造复制缺陷型重组逆转录病毒颗粒方面的方法的一些实施方式中,哺乳动物细胞包含与其膜相关联的:活化元件,其包含与cd14gpi锚连接序列结合的抗cd3scfv或抗cd3scfvfc,或与cd16bgpi锚连接序列结合的cd80或其胞外片段;及膜结合细胞因子,其包含il-7或其活化片段及包含gpi锚连接序列的daf的融合多肽,且其中第一反式活化子调节每个活化元件及膜结合细胞因子的表达。在一些实施方式中,il-7或其活化片段及daf融合、抗cd3scfv或抗cd3scfvfc及cd80或其胞外片段各自包含daf信号序列。在本文所提供的逆转录病毒包装系统及用于制造复制缺陷型重组逆转录病毒颗粒方面的方法的一些实施方式中,哺乳动物细胞进一步包含vpx多肽。在这些或其他实施方式中,一个或多个假型化元件包含由t细胞识别的一个或多个病毒多肽。一个或多个假型化元件可包含麻疹病毒f多肽、麻疹病毒h多肽和/或其片段。在某些说明性实施方式中,一个或多个假型化元件为麻疹病毒f多肽和/或麻疹病毒h多肽的细胞质结构域缺失变体。在本文所提供的逆转录病毒包装系统及用于制造复制缺陷型重组逆转录病毒颗粒方面的方法的一些实施方式中,可包装rna基因组由可操作地连接至第三启动子的多核苷酸编码,其中该第三启动子是组成型活化的或可由第一反式活化子或第二反式活化子诱导。在说明性实施方式中,可包装rna基因组由可操作地连接至第三启动子的多核苷酸编码,其中该第三启动子可由第二反式活化子诱导。在本文所提供的逆转录病毒包装系统及用于制造复制缺陷型重组逆转录病毒颗粒方面的方法的一些实施方式中,可包装rna基因组进一步自5′至3′包含:a.5′长末端重复或其活化片段;b.核酸序列,其编码逆转录病毒顺式作用rna包装元件;c.核酸序列,其编码第一靶标多肽及任选地第二靶标多肽;d.第四启动子,其可操作地连接至第一靶标多肽及任选地第二靶标多肽,其中该第四启动子在靶标细胞中有活性但在包装细胞系中为无活性的;及e.3′长末端重复或其活化片段。在包括此部分构建体的本文所提供的逆转录病毒包装系统及用于制造复制缺陷型重组逆转录病毒颗粒方面的方法的一些实施方式中,第三启动子促进与由第四启动子促进的转录及表达相反的方向上的转录或表达。在本文所提供的逆转录病毒包装系统及用于制造复制缺陷型重组逆转录病毒颗粒方面的方法的一些实施方式中,可包装rna基因组编码在本发明中公开的任何实施方式的复制缺陷型重组逆转录病毒颗粒,其中第一靶标多肽及第二靶标多肽分别为第一经改造信号传导多肽及第二经改造信号传导多肽。在一些实施中,例如,可包装rna基因组进一步包含可操作地连接至核酸的控制元件,该核酸编码第一经改造信号传导多肽或第二经改造信号传导多肽。控制元件在说明性实施方式中为核糖开关。核糖开关在说明性实施方式中能够结合化合物且结合控制元件的化合物为核苷类似物,且该核苷类似物可以是抗病毒药物,例如阿昔洛韦或喷昔洛韦。在本文所提供的逆转录病毒包装系统及用于制造复制缺陷型重组逆转录病毒颗粒方面的方法的一些实施方式中,可包装rna基因组进一步包含内含子,该内含子包含编码抑制性rna分子(例如mirna或shrna)的多核苷酸。内含子可邻近于第四启动子或在第四启动子下游。在本文所提供的逆转录病毒包装系统及用于制造复制缺陷型重组逆转录病毒颗粒方面的方法的一些实施方式中,靶标细胞可以是t细胞和/或nk细胞。在本文所提供的逆转录病毒包装系统及用于制造复制缺陷型重组逆转录病毒颗粒方面的方法的一些实施方式中,一个或多个假型化元件包含水疱性口炎病毒包膜蛋白(vsv-g)、猫类内源性病毒(rd114)包膜蛋白、致癌逆转录病毒双嗜性包膜蛋白或致癌逆转录病毒亲嗜性包膜蛋白或其功能性片段。在本文所提供的逆转录病毒包装系统及用于制造复制缺陷型重组逆转录病毒颗粒方面的方法的一些实施方式中,可包装rna基因组大小为11,000kb或更小,或10,000kb或更小。在本文所提供的逆转录病毒包装系统及用于制造复制缺陷型重组逆转录病毒颗粒方面的方法的一些实施方式中,第一靶标多肽包含第一经改造信号传导多肽且其中该第一经改造信号传导多肽包含至少一个淋巴增殖性元件,且第二靶标多肽包含包括car的第二经改造信号传导多肽。在一个方面中,本文中提供一种用于调节多核苷酸靶标多核苷酸表达的经分离多核苷酸,其包含:多核苷酸,其编码可操作地连接至启动子及核糖开关的多核苷酸靶标多核苷酸,其中该核糖开关包含:a.适配体结构域,其能够结合核苷类似物抗病毒药物且相对于核苷类似物抗病毒药物减少与鸟嘌呤或2′-脱氧鸟苷的结合;b.功能切换结构域,其能够调节多核苷酸靶标多核苷酸的表达,其中由适配体结构域结合的核苷类似物诱导或抑制功能切换结构域的表达调节活性,从而调节多核苷酸靶标多核苷酸的表达。在包括控制元件的本文所提供的任何这些方法及组合物的说明性实施方式中可以是包含核糖开关的多核苷酸。核糖开关可能够结合核苷类似物且结合控制元件的化合物为核苷类似物。核苷类似物可以是抗病毒剂。抗病毒剂可以是阿昔洛韦或喷昔洛韦。核糖开关可较佳地经由喷昔洛韦结合阿昔洛韦或较佳地经由阿昔洛韦结合喷昔洛韦。核糖开关可在高于37℃、37.5℃、38℃、38.5℃或39℃(例如,高于39℃)的温度下减少与核苷类似物抗病毒药物的结合。核糖开关可在长度为35、40、45及50个核苷酸的范围下限与长度为60、65、70、75、80、85、90、95及100个核苷酸的范围上限之间,例如在长度为45与80个核苷酸之间。在包括核糖开关的本文所提供的任何这些方法及组合物的说明性实施方式中,由核糖开关调节的多核苷酸靶标多核苷酸可包括编码mirna、shrna和/或多肽的区域。多核苷酸靶标多核苷酸可编码淋巴增殖性元件。多核苷酸靶标多核苷酸可可操作地连接至启动子。多核苷酸靶标多核苷酸可包括编码多肽的区域,且该多肽可包括嵌合抗原受体,该嵌合抗原受体包含抗原特异性靶向区、跨膜结构域及胞内活化结构域。在包括核糖开关的本文所提供的这任何些方法及组合物的说明性实施方式中,功能切换结构域可调节内部核糖体进入位点、病毒基因构建体中的pre-mrna剪接供体可及性、翻译、转录终止、转录降解、mirna表达或shrna表达,从而调节多核苷酸靶标多核苷酸的表达。核糖开关可包括核糖核酸酶。在包括核糖开关的本文所提供的任何这些方法及组合物的说明性实施方式中,经分离多核苷酸可以是分子克隆载体或表达载体。在包括核糖开关的本文所提供的任何这些方法及组合物的说明性实施方式中,可将经分离多核苷酸逆转录至病毒基因组中或至哺乳动物染色体或其片段中。本文中提供的另一方面是一种用于遗传修饰及扩增受试者淋巴细胞的方法,其包含:a.自受试者采集血液b.使来自受试者血液的t细胞和/或nk细胞与复制缺陷型重组逆转录病毒颗粒体外接触,该复制缺陷型重组逆转录病毒颗粒包含:i)在其表面上的假型化元件,该假型化元件能够与t细胞和/或nk细胞结合且有助于与复制缺陷型重组逆转录病毒颗粒膜融合,其中该假型化元件包含麻疹病毒f多肽和/或麻疹病毒h多肽的细胞质结构域缺失变体;ii)能够与cd3结合的多肽及能够与cd28结合的多肽,其中这些多肽在复制缺陷型重组逆转录病毒颗粒表面上表达且能够与t细胞和/或nk细胞结合,且另外其中这些多肽并不由复制缺陷型重组逆转录病毒颗粒中的多核苷酸编码;及iii)多核苷酸,其包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中一个或多个转录单元编码包含组成型活化il-7受体突变体的第一经改造信号传导多肽及包含嵌合抗原受体的第二经改造信号传导多肽,该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域。其中il-7受体突变体的表达是由结合核苷类似物抗病毒药物的核糖开关调节,其中核苷类似物抗病毒药物与核糖开关的结合增加il-7受体突变体表达,且其中该接触使得静息t细胞和/或nk细胞中至少一些变为遗传修饰;a.将遗传修饰t细胞和/或nk细胞重新引入至受试者中;及b.将遗传修饰t细胞和/或nk细胞体内暴露于核苷类似物抗病毒药物以促进t细胞和/或nk细胞扩增,其中采集血液与重新引入遗传修饰t细胞和/或nk细胞之间的方法在不多于24小时内执行和/或不需要进行预先体外刺激,从而遗传修饰且扩增受试者淋巴细胞。在此方法方面的说明性实施方式中,逆转录病毒颗粒为慢病毒颗粒。在另一说明性实施方式中,复制缺陷型重组逆转录病毒颗粒遗传修饰t细胞。在另一说明性实施方式中,能够与cd3结合的多肽及能够与cd28结合的多肽各自与异源性gpi锚连接序列融合。在一些情况下,能够与cd3结合的多肽可以是抗cd3scfvfc或抗cd3scfv,且能够与cd28结合的多肽可以是cd80。抗cd3scfvfc或抗cd3scfv及cd80可各自进一步与daf信号序列融合。在另一说明性实施方式中,复制缺陷型重组逆转录病毒颗粒在其表面上进一步包含融合多肽,该融合多肽包含共价连接至daf的细胞因子。在一些情况下,细胞因子可以是il-7或il-15,且融合多肽可包含daf信号序列、不具有其信号序列的il-7及包含gpi锚连接序列的daf片段。在此部分的此方法方面的另一说明性实施方式中,核糖开关进一步以通过核糖开关与核苷类似物抗病毒药物结合调节的方式来控制嵌合抗原受体表达,该核苷类似物抗病毒药物在一些情况下为阿昔洛韦和/或喷昔洛韦。在另一实施方式中,组成型活化il-7可经mirna或shrna或编码mirna或shrna的核酸替换,且可存在il-7。在一些情况下,mirna或shrna可由内含子内的核酸编码。本文中提供的另一方面为复制缺陷型重组逆转录病毒颗粒,其包含:a.在其表面上的假型化元件,该假型化元件能够与t细胞和/或nk细胞结合且有助于与复制缺陷型重组逆转录病毒颗粒膜融合,其中该假型化元件包含麻疹病毒f多肽和/或麻疹病毒h多肽的细胞质结构域缺失变体;b.多核苷酸,其包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码包含嵌合抗原受体(包含抗原特异性靶向区、跨膜结构域及胞内活化结构域)的第一经改造信号传导多肽及包含组成型il-7受体突变体的第二经改造信号传导多肽;其中il-7受体突变体表达由结合核苷类似物抗病毒药物的核糖开关调节,其中核苷类似物抗病毒药物与核糖开关的结合增加il-7受体突变体表达;及c.能够与cd3结合的多肽及能够与cd28结合的多肽,其中这些多肽在复制缺陷型重组逆转录病毒颗粒表面上表达;能够与t细胞和/或nk细胞结合;且不由复制缺陷型重组逆转录病毒颗粒中的多核苷酸编码。在此部分的复制缺陷型重组逆转录病毒颗粒方面的说明性实施方式中,该逆转录病毒颗粒为慢病毒颗粒。在该方法的其他说明性实施方式中,能够与cd3结合的多肽及能够与cd28结合的多肽各自与异源性gpi锚连接序列融合。在一些情况下,能够与cd3结合的多肽可以是抗cd3scfvfc或抗cd3scfv,且能够与cd28结合的多肽可以是cd80。抗cd3scfvfc或抗cd3scfv及cd80可各自进一步与daf信号序列融合。在另一说明性实施方式中,复制缺陷型重组逆转录病毒颗粒在其表面上进一步包含融合多肽,该融合多肽包含共价连接至daf的细胞因子。在一些情况下,细胞因子可以是il-7或il-15,且融合多肽可包含daf信号序列、不具有其信号序列的il-7及包含gpi锚连接序列的daf片段。在此部分的复制缺陷型重组逆转录病毒颗粒方面的另一说明性实施方式中,核糖开关进一步以通过核糖开关与核苷类似物抗病毒药物结合调节的方式来控制嵌合抗原受体表达,该核苷类似物抗病毒药物在一些情况下为阿昔洛韦和/或喷昔洛韦。在另一实施方式中,组成型活化il-7可经mirna或shrna或编码mirna或shrna的核酸替换,且可存在il-7。mirna或shrna可由内含子内的核酸编码。本文中提供的另一方面为一种用于制造复制缺陷型重组逆转录病毒颗粒的方法,其包含:a.培养包装细胞的群体以累积第一反式活化子,其中包装细胞包含由组成型启动子表达的第一反式活化子,其中第一反式活化子能够结合第一配体及第一可诱导启动子以供在第一配体存在下对比于缺失下影响可操作地连接至其核酸序列表达,且其中由第一反式活化子调节第二反式活化子及逆转录病毒rev蛋白质表达;b.在第一配体存在下孵育包含经累积第一反式活化子的包装细胞群体以累积第二反式活化子及逆转录病毒rev蛋白质及通常在其表面上的活化元件,该活化元件包含能够与cd3结合的多肽及包含能够与cd28结合的多肽,其中第二反式活化子能够结合第二配体及第二可诱导启动子以供在第二配体存在下对比于缺失下影响可操作地连接至其核酸序列表达;及c.在第二配体存在下孵育包含经累积第二反式活化子及逆转录病毒rev蛋白质的包装细胞群体,从而诱导gag多肽、pol多肽及能够与t细胞和/或nk细胞结合且有助于与复制缺陷型重组逆转录病毒颗粒膜融合的假型化元件表达,其中该假型化元件包含麻疹病毒f多肽和/或麻疹病毒h多肽的细胞质结构域缺失变体,其中可包装rna基因组是由可操作地连接至第三启动子的多核苷酸编码,且其中该启动子可由第二反式活化子诱导,其中可包装rna基因组自5′至3′包含:i)5′长末端重复或其活化片段;ii)核酸序列,其编码逆转录病毒顺式作用rna包装元件;iii)核酸序列,其编码由切割信号隔开的包含嵌合抗原受体的第一经改造信号传导多肽及包含组成型活化il-7受体突变体的第二经改造信号传导多肽;iv)第四启动子,其在t细胞和/或nk细胞中有活性;及v)3′长末端重复或其活化片段,且其中可包装rna基因组进一步包含结合核苷类似物抗病毒药物的核糖开关,其中核苷类似物抗病毒药物与核糖开关结合增加il-7受体突变体表达,从而制造复制缺陷型重组逆转录病毒颗粒。在该方法的一说明性实施方式中,核糖开关进一步以通过结合核糖开关与核苷类似物抗病毒药物结合调节的方式来控制嵌合抗原受体表达。在另一说明性实施方式中,核苷类似物抗病毒药物为阿昔洛韦和/或喷昔洛韦。在另一说明性实施方式中,可包装rna基因组进一步包含识别结构域,其中该识别结构域包含由识别egfr或其表位的抗体识别的多肽。在另一说明性实施方式中,第一配体为雷帕霉素,且第二配体为四环素或阿霉素;或第一配体为四环素或阿霉素,且第二配体为雷帕霉素。在另一说明性实施方式中,包装细胞进一步包含在第二或任选地第三转录单元上或可操作地连接至第一可诱导启动子的额外转录单元上编码vpx的核酸序列。在另一说明性实施方式中,能够与cd3结合的多肽及能够与cd28结合的多肽各自与异源性gpi锚连接序列融合。在一些情况下,能够与cd3结合的多肽可以是抗cd3scfvfc或抗cd3scfv,且能够与cd28结合的多肽可以是cd80。抗cd3scfvfc或抗cd3scfv及cd80可各自进一步与daf信号序列融合。在另一说明性实施方式中,也包括包含共价连接至daf的细胞因子的融合多肽的表达。在一些情况下,细胞因子可以是il-7或il-15,且融合多肽可包含daf信号序列、不具有其信号序列的il-7及包含gpi锚连接序列的daf片段。在另一说明性实施方式中,核糖开关进一步以通过结合核糖开关与核苷类似物抗病毒药物结合调节的方式来控制嵌合抗原受体表达,该核苷类似物抗病毒药物在一些情况下为阿昔洛韦和/或喷昔洛韦。在另一实施方式中,组成型活化il-7可经mirna或shrna或编码mirna或shrna的核酸替换,且可存在il-7。mirna或shrna可由内含子内的核酸编码。在一说明性实施方式中,逆转录病毒颗粒为慢病毒颗粒。本文中另一方面中提供一种遗传修饰淋巴细胞,其包含:a.第一经改造信号传导多肽,其包含组成型活化il-7受体突变体;及b.第二经改造信号传导多肽,其包含嵌合抗原受体,该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域。在以上遗传修饰淋巴细胞的说明性实施方式中,遗传修饰的淋巴细胞为t细胞和/或nk细胞。在某些实施方式中,淋巴细胞为t细胞。在另一说明性实施方式中,该第一经改造信号传导多肽和/或该第二经改造信号传导多肽表达是由结合核苷类似物抗病毒药物的核糖开关调节,其中核苷类似物抗病毒药物与核糖开关结合增加il-7受体突变体表达。在另一实施方式中,遗传修饰淋巴细胞表达至少一种(例如,两种)抑制性rna分子,例如mirna或shrna。抑制性rna分子可进一步由内含子内的核酸编码。本文中的另一方面提供一种遗传修饰的t细胞和/或nk细胞,其包含:a.第一经改造信号传导多肽,其包含至少一个淋巴增殖性元件;及b.第二经改造信号传导多肽,其包含嵌合抗原受体,该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域。在遗传修饰t细胞和/或nk细胞方面的说明性实施方式中,淋巴增殖性元件是组成型活化的,且在一些情况下为组成型活化突变的il-7受体或其片段。在另一说明性实施方式中,第一经改造信号传导多肽和/或第二经改造信号传导多肽表达是由控制元件调节。在一些情况下,控制元件为包含核糖开关的多核苷酸。在一些情况下,核糖开关能够结合核苷类似物且当核苷类似物存在时,表达第一经改造信号传导多肽和/或第二经改造信号传导多肽。在其他说明性实施方式中,遗传修饰t细胞和/或nk细胞在其表面上具有活化元件、假型化元件和/或膜结合细胞因子。在一些情况下,活化元件包含能够与cd3结合的膜结合多肽;和/或能够与cd28结合的膜结合多肽。在某些实施方式中,活化元件包含与异源性gpi锚连接序列融合的抗cd3scfv或抗cd3scfvfc和/或与异源性gpi锚连接序列融合的cd80。在一说明性实施方式中,假型化元件包含麻疹病毒f多肽、麻疹病毒h多肽,和/或麻疹病毒f多肽和/或麻疹病毒h多肽的细胞质结构域缺失变体。在其他实施方式中,膜结合细胞因子是融合多肽,其包含与daf融合的il-7或其片段或其包含gpi锚连接序列的片段。在一个方面中,本文中提供一种用于遗传修饰及扩增受试者淋巴细胞的方法,其包含:a.使受试者静息t细胞和/或nk细胞体外在通常不需要进行预先体外刺激的情况下与复制缺陷型重组逆转录病毒颗粒接触,这些复制缺陷型重组逆转录病毒颗粒包含:i)在其表面上的假型化元件,该假型化元件能够与t细胞和/或nk细胞结合且有助于与复制缺陷型重组逆转录病毒颗粒膜融合;及ii)多核苷酸,其包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元:编码由控制元件调节的第一经改造信号传导多肽,其中该第一经改造信号传导多肽包含至少一个淋巴增殖性元件;且任选地编码由控制元件任选地调节的第二经改造信号传导多肽,其中第二经改造信号传导多肽包含胞内活化结构域及任选地car的其他组分,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导至少一些静息t细胞和/或nk细胞,从而产生遗传修饰t细胞和/或nk细胞;b.将遗传修饰t细胞和/或nk细胞引入至受试者中;及将遗传修饰t细胞和/或nk细胞体内暴露于充当控制元件的化合物以影响第一经改造信号传导多肽表达且体内促进淋巴细胞扩增、移植和/或持久性,从而遗传修饰及扩增受试者淋巴细胞。在说明性实施方式中,在无需体外刺激的情况下进行转导。在说明性实施方式中,化合物为分子伴侣,例如小分子伴侣。在说明性实施方式中,分子伴侣与淋巴增殖性元件结合增加淋巴增殖性元件增殖活性。可在采集血液之前、在接触期间和/或在将t细胞和/或nk细胞引入至受试者中之后将分子伴侣施用至受试者。将通过化合物是控制元件的此方面了解,此化合物通常能够与淋巴增殖性元件和/或car的组分结合,且在执行该方法期间与此淋巴增殖性元件或car组分结合。与包括用复制缺陷型重组逆转录病毒颗粒转染t细胞和/或nk细胞的本文所提供方法相关的其他实施方式及教示应用于此方面,也包括分子伴侣实施方式。在另一方面中,本文中提供一种用于选择微环境限制的抗原特异性靶向区的方法,该方法包含通过以下淘选多肽展示文库:a.使多肽展示文库的多肽在正常生理条件下经受结合分析及在异常条件下的结合分析;及b.选择在与生理条件相比的异常条件下展现结合活性增加的多肽,从而选择微环境限制的抗原特异性靶向区。在另一方面中,本文中提供一种用于分离微环境限制的抗原特异性靶向区的方法,其包含:通过以下淘选多肽库:a.在异常条件下使多肽库与结合至固体载体的靶标抗原接触,其中表达与结合靶标抗原多肽的克隆仍经由靶标抗原结合至固体载体;b.在生理条件下将固体载体与结合多肽一起孵育;及c.收集在生理条件下自固体载体洗脱的克隆,从而分离微环境限制的抗原特异性靶向区。在另一方面中,本文中提供一种用于结合靶标抗原的嵌合抗原受体,该嵌合抗原受体包含:a.至少一个微环境限制的抗原特异性靶向区,其通过淘选多肽库选择,且与正常生理条件下相比在异常条件下结合分析的活性增加;b.跨膜结构域;及c.胞内活化结构域。在另一方面中,本文中提供一种用于结合靶标抗原的嵌合抗原受体,该嵌合抗原受体包含:a.微环境限制的抗原特异性靶向区,其与正常生理环境下相比,在异常条件下展现与靶标抗原结合的增加,其中抗原特异性靶向区与靶标结合;b.跨膜结构域;及c.胞内活化结构域。在本文提供的包括微环境限制性抗原特异性靶向区(astr)的任何方法和组合物的说明性实施方案中,与正常条件下相比,astr在异常条件下的分析中对靶标抗原可增加至少5%、10%、15%、20%、25%、30%、40%、50%、60%、70%、80%、90%或100%的结合亲和力。异常条件可以是低氧、酸性ph、较高浓度乳酸、较高浓度透明质酸、较高浓度白蛋白、较高浓度腺苷、较高浓度r-2-羟基戊二酸、较高浓度pad酶、较高压力、较高氧化反应及较低养分可用性。如相比于在7.4ph下,微环境限制性astr在6.7ph下可展现抗原结合的增加。微环境限制性astr可展现在与对应生理条件相关的肿瘤环境和/或体内肿瘤环境分析条件中的抗原结合增加。靶标可以是4-1bb、st4、腺癌抗原、α-胎蛋白、axl、baff、b-淋巴瘤细胞、c242抗原、ca-125、碳酸酐酶-9(ca-ix)、c-met、ccr4、cd152、cd19、cd20、cd200、cd22、cd221、cd23(ige受体)、cd28、cd30(tnfrsf8)、cd33、cd4、cd40、cd44v6、cd51、cd52、cd56、cd74、cd80、cea、cnt0888、ctla-4、drs、egfr、epcam、cd3、fap、纤连蛋白外域-b、叶酸受体1、gd2、gd3神经节苷脂、糖蛋白75、gpnmb、her2/neu、hgf、人类分散因子受体激酶、igf-1受体、igf-i、igg1、ll-cam、il-13、il-6、类胰岛素生长因子i受体、整合素nsp1、整合素nvp3、morab-009、ms4a1、muc1、黏蛋白canag、n葡萄糖代神经胺酸、npc-1c、pdgf-ra、pdl192、磷脂酰丝氨酸、前列腺癌细胞、rankl、ron、ror1、ror2sch900105、sdc1、slamf7、tag-72、腱生蛋白c、tgfβ2、tgf-p、trail-r1、trail-r2、肿瘤抗原ctaa16.88、vegf-a、vegfr-1、vegfr2及波形蛋白。astr可以是抗体、抗原、配体、配体的受体结合结构域、受体、受体的配体结合结构域或亲和体。astr可以是全长抗体、单链抗体、fab片段、fab'片段、(fab')2片段、fv片段以及二价单链抗体或双功能抗体。astr可包括来自抗体的重链及轻链。抗体可以是单链可变片段。在一些实施方式中,重链及轻链可由接头隔开,其中该接头长度在6与100个氨基酸之间。在一些实施方式中,重链可定位于嵌合抗原受体上的轻链n端,且在一些实施方式中轻链可定位于嵌合抗原受体上的重链n端。在包括多肽展展示文库的任何方法的说明性实施方式中,多肽展示文库可以是噬菌体展示文库或酵母展展示文库。多肽展展示文库可以是抗体展示文库。抗体展示文库可以是人类或人源化抗体展示文库。抗体展示文库可以是原生库。这些方法可包括用所收集噬菌体感染细菌细胞以产生精制噬菌体展示文库,且重复接触、孵育及收集1至1000个循环,使用由前述循环产生的精制噬菌体展示文库。在包括分离或选择微环境限制性astr的任何本文所提供的这些方法的说明性实施方式中,该方法可包括测定编码微环境限制的抗原特异性靶向区多核苷酸的核酸序列,从而测定微环境限制性astr的多肽序列。这些方法可包括通过产生编码多肽的多核苷酸来制造微环境限制的生物嵌合抗原受体,该多肽包含微环境限制性astr、跨膜结构域及胞内活化结构域。该文库可以是单链抗体文库。用于分离微环境限制性astr的方法可包括重复淘选1至1000次之间。可执行用于分离微环境限制性astr的方法,而无需在淘选轮次之间突变编码微环境限制的抗原特异性靶向区的多核苷酸。用于分离微环境限制性astr的方法可在淘选轮次之间通过培养、高保真扩增和/或稀释编码抗原特异性靶向区的多核苷酸或包括其的宿主生物体来执行。这些方法可包括在重复之前,诱变处理经选择和/或经分离微环境限制的抗原特异性靶向区。这些方法可包括在经由长久读取dna定序的淘选一个或多个轮次之后测定经选择和/或经分离微环境限制的抗原特异性靶向区和/或编码其的多核苷酸序列。这些方法可包括在经分离微环境限制性astr的扩增之前及之后测定序列。可执行用于分离经分离微环境限制性astr的方法,而无需在不需要重复淘选。可执行用于分离经分离微环境限制性astr的方法,而无需在分离微环境限制性astr之后,突变编码经分离微环境限制性astr的多核苷酸。在本文提供的包括具有微环境限制性astr的嵌合抗原受体的任何组合物的说明性实施方案中,可以通过淘选抗体文库来鉴别微环境限制性astr。在一些实施方式中,微环境限制性astr是通过淘选噬菌体展示文库或酵母展示文库来鉴别。在一些实施方式中,嵌合抗原受体包含双特异性astr。在另一方面中本文中提供一种经转导t细胞和/或nk细胞,其包含重组多核苷酸,该重组多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码由控制元件调节的第一经改造信号传导多肽,其中该第一经改造信号传导多肽包含组成型活化il-7受体突变体,且其中控制元件能够体外或体内与化合物结合,或经组态以体内结合化合物。在另一方面中本文中提供一种复制缺陷型重组逆转录病毒颗粒,其包含重组多核苷酸,该重组多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码由控制元件(其可以是体内控制元件)调节的第一经改造信号传导多肽,其中该第一经改造信号传导多肽包含组成型il-7受体突变体,且其中控制元件能够体内与化合物结合,或经组态以体内结合化合物。在另一方面中本文中提供一种转导t细胞和/或nk细胞的方法,该方法包含:使t细胞和/或nk细胞与包含重组多核苷酸的复制缺陷型重组逆转录病毒颗粒接触,该重组多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码由控制元件调节的第一经改造信号传导多肽,其中该第一经改造信号传导多肽包含组成型il-7受体突变体,且其中体内控制元件能够在转导条件下体内或体外与化合物结合,从而转导t细胞和/或nk细胞。在前述段落提供的经转导t细胞和/或nk细胞方面、复制缺陷型重组逆转录病毒颗粒方面、及方法方面的说明性实施方式中,重组多核苷酸进一步包含编码包含第一嵌合抗原受体的第二经改造信号传导多肽的转录单元,该第一嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域。在其他说明性实施方式中,淋巴增殖性元件包含经突变il-7受体或其片段。在其他说明性实施方式中,控制元件为包含核糖开关的多核苷酸。在一些情况下,核糖开关能够结合核苷类似物且结合控制元件的化合物为核苷类似物。在一些情况下,核苷类似物为抗病毒剂,例如阿昔洛韦或喷昔洛韦。在某些实施方式中,抗病毒剂为阿昔洛韦。在其他说明性实施方式中,组成型活化il-7受体突变体与egfr或其表位融合。在其他说明性实施方式中,组成型活化il-7受体突变体包含etag。在其他说明性实施方式中,组成型活化il-7受体突变体包含ppcl插入。在其他说明性实施方式中,组成型活化il-7受体突变体在与野生型人类il-8受体中的位置243等同的位置处包含ppcl插入。在其他说明性实施方式中,经转导t细胞和/或nk细胞为经转导t细胞。在另一方面中,本文中提供一种用于调节表达微环境限制的生物嵌合抗原受体(mrb-car)t细胞和/或nk细胞与表达受试者中mrb-car的同源抗原细胞结合的方法,该方法包括:a.将包括编码mrb-car的核酸的t细胞和/或nk细胞引入至受试者中,其中在引入后,包括编码mrb-car的核酸的t细胞和/或nk细胞表达mrb-car且与受试者中表达同源抗原的细胞结合;及b.向受试者施用呈充足量的药剂以增加血液ph和/或组织ph和/或微环境ph,其中该施用在引入之前、期间或之后执行,且其中血液、组织和/或微环境的增加的ph调节表达mrb-car的t细胞和/或nk细胞与表达具有增加的ph的血液、组织或微环境中的同源抗原细胞的结合。在另一方面中,本文中提供一种用于靶向减轻受试者中的肿瘤毒性的方法,其包括:a.将编码微环境限制的生物嵌合抗原受体(mrb-car)的核酸引入至受试者t细胞和/或nk细胞中以产生包括编码mrb-car的核酸的t细胞和/或nk细胞;b.将包括编码mrb-car的核酸的t细胞和/或nk细胞引入至受试者中,其中在引入之后,包括编码mrb-car的核酸的t细胞和/或nk细胞表达mrb-car且与表达受试者中同源抗原细胞结合;及c.向受试者施用充足量的药剂以增加血液ph和/或组织ph和/或微环境ph以调节mrb-car与具有增加的ph的血液、组织和/或微环境中的其同源抗原结合,从而减轻受试者中的在靶脱瘤毒性。在一些实施方式中,核酸可以是载体。在说明性实施方式中,载体为逆转录病毒颗粒。在另一方面中,本文中提供一种用于控制t细胞和/或nk细胞与靶标哺乳动物细胞结合的方法,其包括:a.使靶标哺乳动物细胞与微环境中的t细胞和/或nk细胞接触,其中靶标哺乳动物细胞表达同源抗原,且如相比于在ph7.4下,t细胞和/或nk细胞表达在ph6.7下有差异地与同源抗原结合的微环境限制的生物嵌合抗原受体(mrb-car);及b.通过将药剂以充足量引入微环境来增加微环境ph,从而控制t细胞和/或nk细胞与靶标哺乳动物细胞的结合。在另一方面中,本文中提供一种用于控制表达微环境限制的生物嵌合抗原受体(mrb-car)的t细胞和/或nk细胞与受试者中的靶标哺乳动物细胞在体内结合的方法,该方法包括经由增加受试者内微环境ph的有效给药方案向受试者施用ph调节药剂,其中受试者包括表达mrb-car的t细胞和/或nk细胞,其中如相比于在ph7.4下,mrb-car在ph6.7下有差异地与其同源抗原结合,其中微环境包括靶标哺乳动物细胞,其中靶标哺乳动物细胞在其表面上表达同源抗原,且其中t细胞和/或nk细胞在增加微环境ph之前相比于之后有差异地与靶标哺乳动物细胞结合,从而控制t细胞和/或nk细胞与受试者中的靶标哺乳动物细胞在体内的结合。在包括药剂及mrb-car的此部分提供任何方面中,mrb-car可在一个ph下而非不同ph下减少与其同源抗原结合。在针对mrb-car的差异性结合的说明性ph值未曾提供于广泛方面中的说明性实施方式中及替代地对于代替用于这些方面的那些值的其他实施方式,mrb-car可在较高ph下而非较低ph下减少结合。举例而言,mrb-car可在高于ph7.0、7.1、7.2、7.3、7.4或7.5下而非低于ph6.4、6.5、6.6、6.7、6.8、6.9或7.0下减少与其同源抗原结合。在其他实施方式中,mrb-car可在较低ph下而非较高ph下减少结合。举例而言,mrb-car可在低于ph6.4、6.5、6.6、6.7、6.8、6.9或7.0下而非高于ph7.0、7.1、7.2、7.3、7.4或7.5下减少与其同源抗原结合。在一些说明性实施方式中,相比于在ph7.4至7.6下,mrb-car在6.5至ph6.7下展现增加的结合。在其他说明性实施方式中,相比于在ph7.4下,mrb-car在ph6.7下展现增加的结合。在其他实施方式中,相比于在血液ph下,mrb-car在肿瘤ph下展现增加的结合。在一些实施方式中,mrb-car可包括抗原特异性靶向区、柄及胞内活化结构域。在一些实施方式中,mrb-car也可包括共刺激结构域。在一些实施方式中,mrb-car可与肿瘤相关抗原结合。在包括药剂及mrb-car的此部分提供的任何方面中,微环境ph可自低于ph7.0增加至高于ph7.0。举例而言,ph可自低于ph6.4、6.5、6.6、6.7、6.8、6.9或7.0增加至高于ph7.0、7.1、7.2、7.3或7.4。在一些实施方式中,mrb-car可在引入药剂之前在增加的ph下而非微环境ph下与同源抗原结合。在某些实施方式中,ph可自低于7.0增加7.1至8.0、或至7.1至7.8、或至7.2至7.8、或至7.2至7.6、或至7.3至7.6、或至7.4至7.8、或至7.4至7.6。视施用药剂类型及剂量而定,此ph的增加可发生少于1小时、2小时、4小时、6小时、8小时、12小时或24小时,或多于1小时、2小时、4小时、6小时、8小时、12小时或24小时。在某些实施方式中,施用药剂,使得ph在靶标组织(例如肿瘤)中,及例如在靶标组织(例如,肿瘤)微环境的至少一个表面中,在靶标组织(例如,肿瘤)微环境的至少一部分中以及在说明性实施方式中在整个靶标组织(例如,肿瘤)微环境中保持高于7.0、7.1、7.2、7.3、7.4或7.5;或在7.0,7.1、7.2、7.3范围下限与7.4、7.5、7.6、7.7或7.8范围上限之间。在包括药剂及mrb-car的此部分提供的任何方面中,微环境可以是体内微环境,例如肿瘤、组织、非肿瘤组织、正常组织或已经历ph短暂变化的组织。举例而言,通常经历ph短暂变化的组织包括厌氧条件下的肌肉组织或经历锻炼的肌肉组织或已发炎组织或正经历发炎的组织。在包括靶标哺乳动物细胞的一些实施方式中,靶标哺乳动物细胞可以是肿瘤细胞或非肿瘤细胞或正常细胞。在包括药剂及mrb-car的此部分提供的任何方面中,药剂可以是碳酸氢钠、参(羟甲基)氨基甲烷、碳酸氢钠及碳酸钠等摩尔高渗溶液或质子泵抑制剂,这些质子泵抑制剂,例如艾美拉唑(esomeprazole)、艾美拉唑及萘普生(naproxen)、兰索拉唑(lansoprazole)、奥美拉唑(omeprazole)及雷贝拉唑(rabeprazole)。编码本发明的mrb-car的核酸可经由多种手段引入至t细胞和/或nk细胞中。在包括药剂及mrb-car的此部分提供的任何方面中,一个或多个引入步骤可通过以下来执行:a.使受试者静息t细胞和/或nk细胞体外在不需要进行预先体外刺激的情况下与复制缺陷型重组逆转录病毒颗粒接触,该复制缺陷型重组逆转录病毒颗粒包括:i)在其表面上的一个或多个假型化元件,该假型化元件能够与t细胞和/或nk细胞结合且有助于与复制缺陷型重组逆转录病毒颗粒膜融合;及ii)多核苷酸,其包括可操作地连接编码mrb-car的t细胞和/或nk细胞中活性启动子的转录单元,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导至少一些静息t细胞和/或nk细胞,从而产生能够表达mrb-car的t细胞和/或nk细胞,通常是因为其现今包括多核苷酸,该多核苷酸包括在编码mrb-car的t细胞和/或nk细胞中活性启动子的转录单元;及b.将能够表达mrb-car的t细胞和/或nk细胞引入至受试者中。在一些实施方式中,t细胞和/或nk细胞可在引入之前体外经历2、3、4、5、6、7、8、9或10次或更少细胞分裂。在一些实施方式中,静息t细胞和/或静息nk细胞可与复制缺陷型重组逆转录病毒颗粒接触1小时、2小时、3小时、4小时、5小时、6小时、7小时、8小时、9小时、10小时、11或12小时的范围下限与6小时、7小时、8小时、9小时、10小时、11小时、12小时、13小时、14小时、15小时、16小时、17小时、18小时、19小时、20小时、21小时、22小时、23小时或24小时的范围上限之间,其中对于给定范围,较低值低于较高值。在一些实施方式中,静息t细胞和/或静息nk细胞可来自自受试者收集的血液。在说明性实施方式中,在自受试者采集血液的时间与将能够表达mrb-car的t细胞和/或静息nk细胞引入至受试者中的时间之间可不多于12小时、15小时、16小时、18小时、21小时、24小时、30小时、36小时、42小时或48小时。在一些实施方式中,采集血液之后及引入能够表达mrb-car的t细胞和/或静息nk细胞之前的所有步骤在封闭系统中执行。在包括方法(包括mrb-car及药剂)中复制缺陷型重组逆转录病毒颗粒的此部分提供的任何实施方式中,包括可操作地连接编码mrb-car的t细胞和/或nk细胞中活性启动子的转录单元的多核苷酸由t细胞和/或nk细胞吸收,使得此类细胞能够表达mrb-car。在说明性实施方式中t细胞和/或nk细胞将多核苷酸整合至其基因组中。在包括方法(包括mrb-car及药剂)中复制缺陷型重组逆转录病毒颗粒的此部分提供的任何实施方式中,复制缺陷型重组逆转录病毒颗粒可进一步包括在其表面上的活化元件,例如能够活化静息t细胞和/或静息nk细胞的活化元件。在一些实施方式中,活化元件可包括在本发明中提供的任何活化元件。在说明性实施方式中,活化元件可包括能够与cd3结合的膜结合多肽和/或能够与cd28结合的膜结合多肽。在包括方法(包括mrb-car及药剂)中的复制缺陷型重组逆转录病毒颗粒表面上活化元件的此部分提供的任何实施方式中,膜结合多肽中的一个或多个可与异源性gpi锚连接序列融合。在一些实施方式中,能够与cd3结合的膜结合多肽和/或能够与cd28结合的膜结合多肽可以是分别结合cd3或cd28的scfv或scfvfc。在说明性实施方式中,能够与cd3结合的膜结合多肽可以是结合cd3的scfv或scfvfc。在一些实施方式中,能够与cd28结合的膜结合多肽可以是cd80、cd86的胞外结构域或能够诱导cd28介导的akt的活化的其功能性片段。在包括方法(包括mrb-car及药剂)中复制缺陷型重组逆转录病毒颗粒的此部分提供的任何实施方式中,编码mrb-car的多核苷酸可可操作地连接至核糖开关。在一些实施方式中,核糖开关可以能够结合核苷类似物。在一些实施方式中,核苷类似物可以是抗病毒药物,例如阿昔洛韦或喷昔洛韦。在包括方法(包括mrb-car及药剂)中复制缺陷型重组逆转录病毒颗粒的此部分提供的任何实施方式中,复制缺陷型重组逆转录病毒颗粒可在其表面上包括生物批准的单克隆抗体识别结构域。举例而言,识别结构域可包括由识别egfr或其表位的抗体所识别的多肽。在包括方法(包括mrb-car及药剂)中复制缺陷型重组逆转录病毒颗粒的此部分提供的任何实施方式中,一个或多个假型化元件可包括麻疹病毒f多肽、麻疹病毒h多肽和/或保留与静息t细胞和/或静息nk细胞结合能力的其片段。在一些实施方式中,一个或多个假型化元件可包括vsv-g多肽。在一些实施方式中,复制缺陷型重组逆转录病毒颗粒可在其表面上包括il-7或其活化片段及包括gpi锚连接序列的daf的融合多肽。在包括方法(包括mrb-car及药剂)中复制缺陷型重组逆转录病毒颗粒的此部分提供的任何实施方式中,复制缺陷型重组逆转录病毒颗粒基因组可编码一种或多种抑制性rna分子,例如两种或更多种、三种或更多种、四种或更多种、五种或更多种、或六种或更多种抑制性rna分子。在一些实施方式中,抑制性rna分子可针对不同rna靶标。在一些实施方式中,抑制性rna分子可位于内含子内。在一些实施方式中,抑制性rna分子能够形成形成18至25个核苷酸rna双链体的5'茎及3'茎。在一些实施方式中,抑制性rna分子中的至少一者自5'至3'取向可包括:5'微小rna侧翼序列、5'茎、环、3'茎及3'微小rna侧翼序列,其中5'茎或3'茎能够与rna靶标结合。在另一实施方式中,5'茎长度可以是18至25个核苷酸,其中该3'茎长度为18至25个核苷酸,其中该环长度为3至40个核苷酸。在一些实施方式中,5'微小rna侧翼序列及3'微小rna侧翼序列中的一种或多种衍生自天然存在的mirna,例如mir-155。在另一方面中,本文中提供一种ph调节药剂以供在用于控制t细胞和/或nk细胞与受试者中的靶标哺乳动物细胞在体内结合的方法中使用,该方法包括经由增加受试者内微环境ph的有效给药方案向受试者施用ph调节药剂,其中受试者包括t细胞和/或nk细胞,其中如相比于在ph7.4下,t细胞和/或nk细胞表达在ph6.7下有差异地与其同源抗原结合微环境限制的生物嵌合抗原受体(mrb-car),其中t细胞和/或nk细胞表达mrb-car,其中微环境包括靶标哺乳动物细胞,其中靶标哺乳动物细胞其表面上的表达同源抗原,且其中t细胞和/或nk细胞在通过施用ph调节药剂增加微环境ph之前相比于之后有差异地与靶标哺乳动物细胞结合,从而控制t细胞和/或nk细胞与受试者中的靶标哺乳动物细胞在体内结合。在另一方面中,本文中提供一种药剂以供在用于调节表达微环境限制的生物嵌合抗原受体(mrb-car)的t细胞和/或nk细胞与表达受试者中的mrb-car的同源抗原细胞结合的方法中使用,以供治疗肿瘤生长,其中该方法包括:a.将能够表达mrb-car的t细胞和/或nk细胞引入至受试者中,其中mrb-car与表达受试者中的同源抗原细胞结合,其中在引入之后,包括编码mrb-car的核酸的t细胞和/或nk细胞表达mrb-car且与表达受试者中的同源抗原细胞结合;及b.向受试者施用呈有效量的药剂以增加血液ph和/或组织ph和/或微环境ph以调节表达mrb-car的t细胞和/或nk细胞与表达具有增加的ph的血液、组织或微环境中的同源抗原细胞结合。在另一方面中,本文中提供一种药剂以供在用于靶向减轻受试者中的肿瘤毒性的方法中使用,其中该方法包括:a.将编码微环境限制的生物嵌合抗原受体(mrb-car)的核酸引入至受试者的t细胞和/或nk细胞中,以产生能够表达mrb-car的t细胞和/或nk细胞;b.将能够表达mrb-car的t细胞和/或nk细胞引入至受试者中,其中在引入之后,包括编码mrb-car的核酸的t细胞和/或nk细胞表达mrb-car且与表达受试者中的同源抗原细胞结合;及c.向受试者施用呈充足量的药剂以增加血液ph和/或组织ph和/或微环境ph以调节mrb-car与具有增加的ph的血液、组织和/或微环境中的其同源抗原结合,从而靶向减轻受试者中的肿瘤毒性。在另一方面中,本文中提供一种药剂以供在用于控制表达微环境限制的生物嵌合抗原受体(mrb-car)的t细胞和/或nk细胞与靶标哺乳动物细胞结合的方法中使用,以供治疗肿瘤生长,其中该方法包括:a.使靶标哺乳动物细胞与表达微环境中的mrb-car的t细胞和/或nk细胞接触,其中靶标哺乳动物细胞表达同源抗原,且如相比于在ph7.4下,t细胞和/或nk细胞表达在ph6.7下有差异地与同源抗原结合的mrb-car;及b.通过将充足量的药剂引入至微环境来增加微环境ph,从而控制表达mrb-car的t细胞和/或nk细胞与靶标哺乳动物细胞结合。在另一方面中,本文中提供一种药剂以供在用于控制表达微环境限制的生物嵌合抗原受体(mrb-car)的t细胞和/或nk细胞与受试者中的靶标哺乳动物细胞在体内结合的方法中使用,以供治疗肿瘤生长,其中该药剂为ph调节药剂,且其中该方法包括经由增加受试者内微环境ph的有效给药方案向受试者施用ph调节药剂,其中受试者包括表达mrb-car的t细胞和/或nk细胞,其中如相比于在ph7.4下,mrb-car在ph6.7下有差异地与其同源抗原结合,其中微环境包括靶标哺乳动物细胞,其中靶标哺乳动物细胞在其表面上表达同源抗原,且其中t细胞和/或nk细胞在增加微环境ph之前相比于之后有差异地与靶标哺乳动物细胞结合。在另一方面中,本文中提供一种ph调节药剂以供在用于控制表达微环境限制的生物嵌合抗原受体(mrb-car)的t细胞和/或nk细胞与受试者中的靶标哺乳动物细胞在体内结合的方法中使用,以供治疗肿瘤生长,其中该方法包括经由增加受试者内微环境ph的有效给药方案向受试者施用ph调节药剂,其中受试者包括表达mrb-car的t细胞和/或nk细胞,其中如相比于在ph7.4下,mrb-car在ph6.7下有差异地与其同源抗原结合,其中微环境包括靶标哺乳动物细胞,其中靶标哺乳动物细胞在其表面上表达同源抗原,且其中t细胞和/或nk细胞在通过施用ph调节药剂增加微环境ph之前相比于之后有差异地与靶标哺乳动物细胞结合。在另一方面中,本文中提供一种ph调节药剂以供用于制造用于控制表达微环境限制的生物嵌合抗原受体(mrb-car)的t细胞和/或nk细胞与受试者中的靶标哺乳动物细胞在体内结合的药剂,其中经由增加受试者内微环境ph的有效给药方案向受试者施用ph调节药剂,其中受试者包括表达mrb-car的t细胞和/或nk细胞,其中如相比于在ph7.4下,mrb-car在ph6.7下有差异地与其同源抗原结合,其中微环境包括靶标哺乳动物细胞,其中靶标哺乳动物细胞在其表面上表达同源抗原,且其中t细胞和/或nk细胞在通过施用ph调节药剂增加微环境ph之前相比于之后有差异地与靶标哺乳动物细胞结合。在包括用于方法中的ph调节药剂或药剂及mrb-car或包括ph调节药剂及mrb-car用途的此部分提供的任何方面中,mrb-car可在一个ph下而非不同ph下减少与其同源抗原结合。在针对mrb-car的差异性结合的说明性ph值未曾提供于广泛方面中的说明性实施方式中及替代地对于代替用于这些方面的那些值的其他实施方式,mrb-car可在较高ph下而非较低ph下减少结合。举例而言,mrb-car可在高于ph7.0、7.1、7.2、7.3、7.4或7.5下而非低于ph6.4、6.5、6.6、6.7、6.8、6.9或7.0下减少与其同源抗原结合。在其他实施方式中,mrb-car可在较低ph下而非较高ph下减少结合。举例而言,mrb-car可在低于ph6.4、6.5、6.6、6.7、6.8、6.9或7.0下而非高于ph7.0、7.1、7.2、7.3、7.4或7.5下减少与其同源抗原结合。在一些说明性实施方式中,相比于在ph7.4至7.6下,mrb-car在ph6.5至6.7下展现增加的结合。在其他说明性实施方式中,相比于在ph7.4下,mrb-car在ph6.7下展现增加的结合。在其他实施方式中,相比于在血液ph下,mrb-car在肿瘤ph下展现增加的结合。在一些实施方式中,mrb-car可包括抗原特异性靶向区、柄及胞内活化结构域。在一些实施方式中,mrb-car也可包括共刺激结构域。在一些实施方式中,mrb-car可与肿瘤相关抗原结合。在包括用于方法中的ph调节药剂或药剂及mrb-car或包括ph调节药剂及mrb-car用途的此部分提供的任何方面中,微环境ph可自低于ph7.0增加至高于ph7.0。举例而言,ph可自低于6.4、6.5、6.6、6.7、6.8、6.9或7.0增加至高于7.0、7.1、7.2、7.3或7.4。在一些实施方式中,mrb-car可在引入药剂之前在增加的ph下而非微环境ph下与同源抗原结合。在某些实施方式中,ph可自低于7.0增加7.1至8.0、或至7.1至7.8、或至7.2至7.8、或至7.2至7.6、或至7.3至7.6、或至7.4至7.8、或至7.4至7.6。视施用的药剂类型及剂量而定,此ph的增加可发生少于1小时、2小时、4小时、6小时、8小时、12小时或24小时,或多于1小时、2小时、4小时、6小时、8小时、12小时或24小时。在某些实施方式中,施用药剂,使得ph在靶标组织(例如肿瘤)中,及例如在靶标组织(例如,肿瘤)微环境的至少一个表面中,在靶标组织(例如,肿瘤)微环境的至少一部分中以及在说明性实施方式中在整个靶标组织(例如,肿瘤)微环境中保持高于7.0、7.1、7.2、7.3、7.4或7.5;或在7.0,7.1、7.2、7.3的范围下限与7.4、7.5、7.6、7.7或7.8的范围上限之间。在包括用于方法中的ph调节药剂或药剂及mrb-car或包括ph调节药剂及mrb-car用途的此部分提供的任何方面中,微环境可以是体内微环境,例如肿瘤、组织、非肿瘤组织、正常组织或已经历ph短暂变化的组织。举例而言,通常经历ph短暂变化的组织包括厌氧条件下的肌肉组织或经历锻炼的肌肉组织或已发炎组织或正经历发炎的组织。在包括靶标哺乳动物细胞的一些实施方式中,靶标哺乳动物细胞可以是肿瘤细胞或非肿瘤细胞或正常细胞。在包括用于方法中ph调节药剂或药剂及mrb-car或包括ph调节药剂及mrb-car用途的此部分提供的任何一方面中,药剂可以是碳酸氢钠、三(羟甲基)氨基甲烷、碳酸氢钠及碳酸钠的等摩尔高渗溶液或质子泵抑制剂,这些质子泵抑制剂例如艾美拉唑、艾美拉唑及萘普生、兰索拉唑、奥美拉唑及雷贝拉唑。在包括用于方法中的ph调节药剂或药剂及mrb-car或包括ph调节药剂及mrb-car用途的此部分提供的任何方面中,药剂可在用于治疗癌症、肿瘤、肿瘤生长或细胞增殖性病症的方法中使用。在另一方面中,本文中提供一种含有容器(含有复制缺陷型重组逆转录病毒颗粒)及用于治疗肿瘤生长的其使用说明书的试剂盒,其中这些说明书指导用于控制t细胞和/或nk细胞与靶标哺乳动物细胞结合的方法,在该方法中包括:a.用在其基因组中包括在与ph7.4相比的ph6.7下有差异地与同源抗原结合的微环境限制的嵌合抗原受体(mrb-car)的复制缺陷型重组逆转录病毒颗粒来转导t细胞和/或nk细胞,以产生能够表达mrb-car的t细胞和/或nk细胞;b.将能够表达mrb-car的t细胞和/或nk细胞引入至受试者中,其中在引入之后,包括编码mrb-car的核酸的t细胞和/或nk细胞表达mrb-car且与表达受试者中的同源抗原细胞结合;c.使靶标哺乳动物细胞与微环境中表达mrb-car的t细胞和/或nk细胞接触,其中靶标哺乳动物细胞表达mrb-car的同源抗原,且t细胞和/或nk细胞表达mrb-car;及d.通过引入充足量的ph调节药剂至微环境来增加微环境ph,从而影响靶标哺乳动物细胞与t细胞和/或nk细胞结合。在一些实施方式中,试剂盒可进一步包括ph调节药剂。在试剂盒的一些实施方式中,mrb-car可在一个ph下而非不同ph下减少与其同源抗原结合。在针对mrb-car的差异性结合的说明性ph值未曾提供于广泛方面中的说明性实施方式中及替代地对于代替用于这些方面的那些值的其他实施方式,mrb-car可在较高ph下而非较低ph下减少结合。举例而言,mrb-car可在高于ph7.0、7.1、7.2、7.3、7.4或7.5下而非低于ph6.4、6.5、6.6、6.7、6.8、6.9或7.0下减少与其同源抗原结合。在另一些实施方式中,mrb-car可在较低ph下而非较高ph下减少结合。举例而言,mrb-car可在低于ph6.4、6.5、6.6、6.7、6.8、6.9或7.0下而非高于ph7.0、7.1、7.2、7.3、7.4或7.5下减少与其同源抗原结合。在一些说明性实施方式中,相比于在ph7.4至7.6下,mrb-car在ph6.5至6.7下展现增加的结合。在其他说明性实施方式中,相比于在ph7.4下,mrb-car在ph6.7下展现增加的结合。在另一些实施方式中,相比于在血液ph下,mrb-car在肿瘤ph下展现增加的结合。在一些实施方式中,mrb-car可包括抗原特异性靶向区、柄及胞内活化结构域。在一些实施方式中,mrb-car也可包括共刺激结构域。在一些实施方式中,mrb-car可与肿瘤相关的抗原结合。在试剂盒的一些实施方式中,微环境ph可自低于ph7.0增加至高于ph7.0。举例而言,ph可自低于ph6.4、6.5、6.6、6.7、6.8、6.9或7.0增加至高于ph7.0、7.1、7.2、7.3或7.4。在一些实施方式中,mrb-car可在引入药剂之前在增加的ph下而非微环境ph下与同源抗原结合。在某些实施方式中,ph可自低于7.0增加7.1至8.0、或至7.1至7.8、或至7.2至7.8、或至7.2至7.6、或至7.3至7.6、或至7.4至7.8、或至7.4至7.6。视施用的药剂类型及剂量而定,此ph的增加可发生少于1小时、2小时、4小时、6小时、8小时、12小时或24小时,或多于1小时、2小时、4小时、6小时、8小时、12小时或24小时。在某些实施方式中,施用药剂,使得ph在靶标组织(例如肿瘤)中,及例如在靶标组织(例如,肿瘤)微环境的至少一个表面中,在靶标组织(例如,肿瘤)微环境的至少一部分中以及在说明性实施方式中在整个靶标组织(例如,肿瘤)微环境中保持高于7.0、7.1、7.2、7.3、7.4或7.5;或在7.0,7.1、7.2、7.3的范围下限与7.4、7.5、7.6、7.7或7.8的范围上限之间。在一些实施方式中,微环境可以是体内微环境,例如肿瘤、组织、非肿瘤组织、正常组织或已经历ph短暂变化的组织。举例而言,通常经历ph短暂变化的组织包括厌氧条件中的肌肉组织或经历锻炼的肌肉组织或已发炎组织或正经历发炎的组织。在包括靶标哺乳动物细胞的一些实施方式中,靶标哺乳动物细胞可以是肿瘤细胞或非肿瘤细胞或正常细胞。在试剂盒的一些实施方式中,药剂可以是碳酸氢钠、三(羟甲基)氨基甲烷、碳酸氢钠及碳酸钠等摩尔高渗溶液或质子泵抑制剂,这些质子泵抑制剂例如艾美拉唑、艾美拉唑及萘普生、兰索拉唑、奥美拉唑及雷贝拉唑。在一个方面中,本文中提供一种在其基因组中包含多核苷酸的复制缺陷型重组逆转录病毒颗粒,该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中:a一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,及b.该一个或多个核酸序列的第二核酸序列编码嵌合抗原受体(car),该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域。在本文中的另一方面中提供一种包含复制缺陷型重组逆转录病毒颗粒的可包装rna基因组的哺乳动物包装细胞系,其中该可包装rna基因组包含:a.5′长末端重复或其活化片段;b.核酸序列,其编码逆转录病毒顺式作用rna包装元件;c.多核苷酸,其包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中一个或多个核酸的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,且一个或多个核酸序列的第二核酸序列编码嵌合抗原受体(car),该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域;及d.3′长末端重复或其活化片段。在哺乳动物包装细胞系方面的一些实施方式中,(c)的多核苷酸可在与编码逆转录病毒顺式作用rna包装元件(b)、5′长末端重复(a)和/或3′长末端重复(d)的核酸序列呈相反定向。在哺乳动物包装细胞系方面的一些实施方式中,可包装rna基因组表达通过在哺乳动物包装细胞系中有活性的可诱导启动子驱动。在哺乳动物包装细胞系方面的一些实施方式中,逆转录病毒顺式作用rna包装元件可包含中央聚嘌呤区(cppt)/中央终止序列、hivpsi或其组合。本文中的另一方面提供一种逆转录病毒载体,其包含针对复制缺陷型重组逆转录病毒颗粒的可包装rna基因组,其中该可包装rna基因组包含:a.5′长末端重复或其活化片段;b.核酸序列,其编码逆转录病毒顺式作用rna包装元件;c.多核苷酸,其包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中一个或多个核酸的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,且一个或多个核酸序列的第二核酸序列编码嵌合抗原受体(car),该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域;及d.3′长末端重复或其活化片段。在逆转录病毒载体方面的一些实施方式中,(c)的多核苷酸可在与编码逆转录病毒顺式作用rna包装元件的核酸序列(b)、5′长末端重复(a)和/或3′长末端重复(d)相反的取向中。在逆转录病毒载体方面的一些实施方式中,可包装rna基因组的表达是由在哺乳动物包装细胞系中有活性的可诱导启动子驱动。在逆转录病毒载体方面的一些实施方式中,逆转录病毒顺式作用rna包装元件可包含中央聚嘌呤段(cppt)/中央终止序列、hivpsi或其组合。逆转录病毒载体可任选地包括抗生素抗性基因和/或可检测标记。在另一方面中,本文提供一种用于遗传修饰或转导受试者淋巴细胞(例如,t细胞和/或nk细胞)或其群的方法,该方法包含使受试者淋巴细胞(例如,t细胞和/或nk细胞)或其群体外与复制缺陷型重组逆转录病毒颗粒接触,该复制缺陷型重组逆转录病毒颗粒在其基因组中包含多核苷酸,该多核苷酸包含可操作地连接淋巴细胞(例如,t细胞和/或nk细胞)中活性启动子的一个或多个核酸序列,其中该一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,且该一个或多个核酸序列的第二核酸序列编码嵌合抗原受体(car),该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒遗传修饰和/或转导淋巴细胞(例如,t细和/或nk细胞)或淋巴细胞(例如,t细胞和/或nk细胞)中的至少一些,从而产生遗传修饰和/或转导的淋巴细胞(例如,t细胞和/或nk细胞)。在此部分提供的方法的一些实施方式中,将遗传修饰和/或转导的淋巴细胞(例如,t细胞和/或nk细胞)或其群引入至受试者中。在一些实施方式中,遗传修饰和/或转导的淋巴细胞(例如,t细胞和/或nk细胞)或其群体在被引入或再次引入至受试者中之前体外经历4次或更少细胞分裂。在一些实施方式中,淋巴细胞为与复制缺陷型重组逆转录病毒颗粒接触1小时与12小时之间的静息t细胞和/或静息nk细胞。在一些实施方式中,自受试者采集血液的时间与将遗传修饰t细胞和/或nk细胞重新引入至受试者中的时间之间不超过8小时。在一些实施方式中,在采集血液之后及重新引入血液之前的所有步骤均在封闭系统中执行,在整个处理期间有人监测该封闭系统。在另一方面中,本文中提供一种遗传修饰t细胞和/或nk细胞,其包含:a.针对一个或多个rna靶标一个或多个(例如,两个或更多个)抑制性rna分子;及b.嵌合抗原受体(car),其包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域,其中该一个或多个(例如,两个或更多个)抑制性rna分子及car是由t细胞和/或nk细胞遗传修饰的核酸序列编码。在遗传修饰的t细胞和/或nk细胞方面的一些实施方式中,遗传修饰t细胞和/或nk细胞也包含不为抑制性rna分子的至少一个淋巴增殖性元件,其中该淋巴增殖性元件是由t细胞和/或nk细胞的遗传修饰的核酸编码。在一些实施方式中,抑制性rna分子、car和/或至少一个淋巴增殖性元件以多顺反子物质表达。在说明性实施方式中,抑制性rna分子由单个多顺反子转录物表达。在另一方面中本文提供一种复制缺陷型重组逆转录病毒颗粒以供在用于遗传修饰受试者淋巴细胞的方法中使用,以供治疗肿瘤生长,其中复制缺陷型重组逆转录病毒颗粒在其基因组中包含多核苷酸,该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,且一个或多个核酸序列的第二核酸序列编码嵌合抗原受体(car),该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域,其中该方法包含体外接触受试者的t细胞和/或nk细胞,且该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导静息t细胞和/或nk细胞中的至少一些,从而产生遗传修饰的t细胞和/或nk细胞。在此部分提供的用于遗传修饰受试者方面淋巴细胞的方法中,在一些实施方式中,药剂是用于该方法中,其进一步包括将经遗传改造的t细胞和/或nk细胞引入至受试者中。在另一方面中本文提供一种复制缺陷型重组逆转录病毒颗粒以供在用于遗传修饰受试者t细胞和/或nk细胞的方法中使用,以供治疗肿瘤生长,其中该方法包含:a.使受试者t细胞和/或nk细胞在离体的情况下与在其基因组中包含多核苷酸的复制缺陷型重组逆转录病毒颗粒接触,该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,且一个或多个核酸序列的第二核酸序列编码嵌合抗原受体(car),该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导静息t细胞和/或nk细胞中的至少一些,从而产生遗传修饰t细胞和/或nk细胞;及b.将遗传修饰t细胞和/或nk细胞引入至受试者中,从而遗传修饰受试者t细胞和/或nk细胞。在此部分提供的方面中,在一些实施方式中,t细胞和/或nk细胞群体在接触步骤中接触,且在引入步骤中引入至受试者中。在另一方面中本文提供复制缺陷型重组逆转录病毒颗粒在制造用于遗传修饰受试者t细胞和/或nk细胞试剂盒中的用途,其中该试剂盒的用途包含:1.使受试者t细胞和/或nk细胞在离体的情况下与在其基因组中包含多核苷酸的复制缺陷型重组逆转录病毒颗粒接触,该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个靶标的一个或多个(例如,两个或更多个)抑制性rna分子,且一个或多个核酸序列的第二核酸序列编码嵌合抗原受体(car),该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导至少一些静息t细胞和/或nk细胞,从而产生遗传修饰的t细胞和/或nk细胞;及2.将遗传修饰的t细胞和/或nk细胞引入至受试者中,从而遗传修饰受试者的t细胞和/或nk细胞。在另一方面中本文提供复制缺陷型重组逆转录病毒颗粒在制造用于遗传修饰受试者t细胞和/或nk细胞的药剂中的用途,其中该药剂的用途包含:a)使受试者t细胞和/或nk细胞在离体的情况下与在其基因组中包含多核苷酸的复制缺陷型重组逆转录病毒颗粒接触,该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个靶标的一个或多个(例如,两个或更多个)抑制性rna分子,且一个或多个核酸序列的第二核酸序列编码嵌合抗原受体(car),该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导至少一些静息t细胞和/或nk细胞,从而产生遗传修饰的t细胞和/或nk细胞;及b)将遗传修饰的t细胞和/或nk细胞引入至受试者中,从而遗传修饰受试者t细胞和/或nk细胞。在另一方面中本文提供一种含有复制缺陷型重组逆转录病毒颗粒及用以以治疗受试者肿瘤生长的其使用说明书的商用容器,其中复制缺陷型重组逆转录病毒颗粒在其基因组中包含多核苷酸,该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,且一个或多个核酸序列的第二核酸序列编码嵌合抗原受体(car),该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域。在一些实施方式中,在商用容器的方面中,该说明书指导使用者体外接触受试者t细胞和/或nk细胞,以有助于利用复制缺陷型重组逆转录病毒颗粒转导受试者至少一个静息t细胞和/或nk细胞,从而产生遗传修饰的t细胞和/或nk细胞。在包括多核苷酸(包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列)的此部分提供的任何方面中,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,且一个或多个核酸序列的第二核酸序列编码嵌合抗原受体(car),该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域,该多核苷酸可进一步包括第三核酸序列,该第三核酸序列编码不为抑制性rna分子的至少一个淋巴增殖性元件。在一些实施方式中,淋巴增殖性元件可以是细胞因子或细胞因子受体多肽,或其包含信号传导结构域的片段。在一些实施方式中,淋巴增殖性元件为组成型活化。在某些实施方式中,淋巴增殖性元件可以是il-7受体或其片段。在说明性实施方式中,淋巴增殖性元件可以是组成型活化il-7受体或其组成型活化片段。在包括多核苷酸(包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列)的此部分提供的任何方面中,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,抑制性rna分子在一些实施方式中可包括与彼此部分或完全互补的5'链及3'链,其中该5'链及该3'链能够形成18至25个核苷酸rna双链体。在一些实施方式中,5'链长度可以是18、19、20、21、22、23、24或25个核苷酸,且3'链长度可以是18、19、20、21、22、23、24或25个核苷酸。在一些实施方式中,5'链及3'链长度可相同或不同。在一些实施方式中,rna双链体可包括一个或多个错配。在替代实施方式中,rna双链体不具有错配。在包括多核苷酸(包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列)的此部分提供的任何方面中,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,抑制性rna分子可以是mirna或shrna。在一些实施方式中,抑制性分子可以是mirna的前体,例如pri-mirna或前mirna,或shrna的前体。在一些实施方式中,抑制性分子可以是人工衍生的mirna或shrna。在另一些实施方式中,抑制性rna分子可以是经处理成sirna的dsrna(经转录或人工引入)或sirna本身。在一些实施方式中,抑制性rna分子可以是mirna或shrna,其具有在自然界中未发现的序列,或具有在自然界中未发现的至少一个功能性区段,或具有在自然界中未发现的功能性区段组合。在说明性实施方式中,抑制性rna分子中至少一者或全部为mir-155。在包括多核苷酸(包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列)的此部分提供的任何方面中,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,在一些实施方式中,抑制性rna分子可自5'至3'取向包含:5'臂、5'茎、环、与该5'茎部分或完全互补的3'茎,及3'臂。在一些实施方式中,两个或更多个抑制性rna分子中至少一者具有此排列。在另一些实施方式中,两个或更多个抑制性rna分子的全部均具有此排列。在一些实施方式中,5'茎长度可以是18、19、20、21、22、23、24或25个核苷酸。在一些实施方式中,3'茎长度可以是18、19、20、21、22、23、24或25个核苷酸。在一些实施方式中,环长度可以是3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39或40个核苷酸。在一些实施方式中,5'臂、3'臂或两者均衍生自天然存在的mirna。在一些实施方式中,5'臂、3'臂或两者均衍生自天然存在的mirna,该天然存在的mirna选自:mir-155、mir-30、mir-17-92、mir-122及mir-21。在说明性实施方式中,5'臂、3'臂或两者是衍生自mir-155。在一些实施方式中,5'臂、3'臂或两者是衍生自小家鼠mir-155或人mir-155。在一些实施方式中,5'臂具有阐述于seqidno:256中的序列或为其功能性变体,例如长度与seqidno:256相同,或seqidno:256长度的95%、90%、85%、80%、75%或50%或为100个核苷酸或更少、95个核苷酸或更少、90个核苷酸或更少、85个核苷酸或更少、80个核苷酸或更少、75个核苷酸或更少、70个核苷酸或更少、65个核苷酸或更少、60个核苷酸或更少、55个核苷酸或更少、50个核苷酸或更少、45个核苷酸或更少、40个核苷酸或更少、35个核苷酸或更少、30个核苷酸或更少或25个核苷酸或更少的序列;且至少50%、55%、60%、65%、70%、75%、80%、85%、90%或95%与seqidno:256一致。在一些实施方式中,3'臂具有阐述于seqidno:260中的序列或为其功能性变体,例如长度与seqidno:260相同,或seqidno:260长度的95%、90%、85%、80%、75%或50%或为100个核苷酸或更少、95个核苷酸或更少、90个核苷酸或更少、85个核苷酸或更少、80个核苷酸或更少、75个核苷酸或更少、70个核苷酸或更少、65个核苷酸或更少、60个核苷酸或更少、55个核苷酸或更少、50个核苷酸或更少、45个核苷酸或更少、40个核苷酸或更少、35个核苷酸或更少、30个核苷酸或更少或25个核苷酸或更少的序列;且至少50%、55%、60%、65%、70%、75%、80%、85%、90%或95%与seqidno:260一致。在一些实施方式中,3'臂包含小家鼠bic的第221至283位核苷酸。在包括多核苷酸(包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列)的此部分提供的任何方面中,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的两个或更多个抑制性rna分子,在一些实施方式中,两个或更多个抑制性rna分子可连续地定位于第一核酸序列中。在一些实施方式中,抑制性rna分子可利用非功能性接头序列直接地或间接地与彼此邻接。在一些实施方式中,接头序列的长度可在5个与120个核苷酸之间,或长度在10个与40个核苷酸之间。在包括多核苷酸(包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列)的此部分提供的任何方面中,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的两个或更多个抑制性rna分子,在一些实施方式中,第一核酸序列编码两个至四个抑制性rna分子。在说明性实施方式中,第一核酸序列中包括2个与10个之间、2个与8个之间、2个与6个之间、2个与5个之间、2个与4个之间、3个与5个之间或3个与6个之间的抑制性rna分子。在一说明性实施方式中,第一核酸序列中包括四个抑制性rna分子。在包括多核苷酸(包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列)的此部分提供的任何方面中,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,该一个或多个(例如,两个或更多个)抑制性rna分子可在内含子中。在一些实施方式中,内含子在启动子中。在说明性实施方式中,内含子为ef-1α内含子a。在一些实施方式中,内含子邻近启动子且在启动子的下游,在说明性实施方式中,该内含子在用于产生复制缺陷型重组逆转录病毒颗粒的包装细胞中是无活性的。在包括多核苷酸(包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列)的此部分提供的任何方面中,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的两个或更多个抑制性rna分子,在一些实施方式中,该两个或更多个抑制性rna分子可针对不同靶标。在一替代实施方式中,两个或更多个抑制性rna分子针对同一靶标。在一些实施方式中,rna靶标为自由t细胞表达基因转录的mrna,这些mrna例如但不限于:pd-1(防止失活);ctla4(防止失活);tcra(安全地防止自体免疫);tcrb(安全-防止自体免疫);cd3z(安全-防止自体免疫);socs1(防止失活);smad2(防止失活);mir-155靶标(促进活化);ifnγ(减少crs);ccbl(延长信号传导);trail2(防止死亡);pp2a(延长信号传导);abcg1(通过限制胆固醇的清除来增加胆固醇微区含量)。在一些实施方式中,rna靶标为自编码t细胞受体(tcr)复合物组分的基因转录的mrna。在一些实施方式中,两个或更多个抑制性rna分子中至少一种可减少t细胞受体(在说明性实施方式中,t细胞的一个或多个内源性t细胞受体)的表达。在某些实施方式中,rna靶标可以是自t细胞的内源性tcrα或tcrβ基因转录的mrna,该t细胞基因组包含编码一个或多个mirna的第一核酸序列。在说明性实施方式中,rna靶标为自tcrα基因转录的mrna。在包括多核苷酸(包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列)的此部分提供的任何方面中,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,且一个或多个核酸序列的第二核酸序列编码嵌合抗原受体(car),该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域,在一些实施方式中,car为微环境限制的生物(mrb)-car。在另一些实施方式中,car的astr与肿瘤相关抗原结合。在另一些实施方式中,car的astr为微环境限制生物(mrb)-astr。在包括多核苷酸(包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个核酸序列)的此部分提供的任何方面中,其中一个或多个核酸序列的第一核酸序列编码针对一个或多个rna靶标的一个或多个(例如,两个或更多个)抑制性rna分子,且一个或多个核酸序列的第二核酸序列编码嵌合抗原受体(car),该嵌合抗原受体包含抗原特异性靶向区(astr)、跨膜结构域及胞内活化结构域,且在一些情况下,一个或多个核酸序列的第三核酸序列编码不为抑制性rna分子的至少一个淋巴增殖性元件,在一些实施方式中,第一核酸序列、第二核酸序列及第三核酸序列中的任一者或全部可操作地连接至核糖开关。在一些实施方式中,核糖开关能够结合核苷类似物。在一些实施方式中,核苷类似物为抗病毒药物。在包括复制缺陷型重组逆转录病毒颗粒的此部分提供的任何方面中,在一些实施方式中,复制缺陷型重组逆转录病毒颗粒包含在其表面上的假型化元件,该假型化元件能够与t细胞和/或nk细胞结合且有助于与复制缺陷型重组逆转录病毒颗粒膜融合。在一些实施方式中,假型化元件可以是麻疹病毒f多肽、麻疹病毒h多肽、vsv-g多肽,或保留与静息t细胞和/或静息nk细胞结合能力的任何其片段。在说明性实施方式中,假型化元件为vsv-g。在包括复制缺陷型重组逆转录病毒颗粒的此部分提供的任何方面中,在一些实施方式中,复制缺陷型重组逆转录病毒颗粒包含在其表面上的活化元件,该活化元件包含能够与cd3结合的膜结合多肽,和/或能够与cd28结合的膜结合多肽。在一些实施方式中,能够与cd3结合的膜结合多肽与异源性gpi锚连接序列融合,和/或能够与cd28结合的膜结合多肽与异源性gpi锚连接序列融合。在一些实施方式中,能够与cd3结合的膜结合多肽为抗cd3scfv或抗cd3scfvfc。在说明性实施方式中,能够与cd3结合的膜结合多肽为抗cd3scfvfc。在说明性实施方式中,能够与cd28结合的膜结合多肽为cd80,或与cd16bgpi锚连接序列结合的其胞外细胞域。在包括复制缺陷型重组逆转录病毒颗粒的此部分提供的任何方面中,在一些实施方式中,复制缺陷型重组逆转录病毒颗粒在其表面上包含核酸,该核酸编码通过生物批准的单克隆抗体识别的结构域。在一个方面中本文提供一种转导和/或遗传修饰受试者淋巴细胞(例如,t细胞和/或nk细胞)(在说明性实施方式中,静息淋巴细胞(例如,静息t细胞和/或nk细胞))的方法,该方法包含使受试者静息t细胞和/或静息nk细胞体外与复制缺陷型重组逆转录病毒颗粒接触,其中复制缺陷型重组逆转录病毒颗粒包含在其表面上的假型化元件及在其表面上膜结合抗cd3scfvfc抗体,该膜结合抗cd3scfvfc抗体能够结合静息t细胞和/或静息nk细胞且有助于与复制缺陷型重组逆转录病毒颗粒膜融合,其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导静息t细胞和/或nk细胞,从而产生遗传修饰的t细胞和/或nk细胞。在一个方面中本文提供一种用于转导和/或遗传修饰来自经分离血液的静息t细胞和/或静息nk细胞的方法,其包含:自受试者采集血液;分离包含静息t细胞和/或静息nk细胞的外周血单核细胞(pbmc);且使受试者的静息t细胞和/或静息nk细胞在离体的情况下与复制缺陷型重组逆转录病毒颗粒接触有效时间,其中该复制缺陷型重组逆转录病毒颗粒包含在其表面上的假型化元件及在其表面上的膜结合抗cd3scfvfc抗体,从而产生遗传修饰的t细胞和/或nk细胞,从而转导静息t细胞和/或nk细胞。在用于转导和/或遗传修饰包括膜结合抗cd3scfvfc抗体的t淋巴细胞(例如,t细胞和/或nk细胞)的此上段落中的这些方面中,某些实施方式中的假型化元件为水疱性口炎病毒包膜蛋白(vsv-g)。在一些实施方式中,复制缺陷型重组逆转录病毒颗粒进一步包含能够与cd28结合的膜结合多肽,该膜结合多肽可包括例如cd80、cd86的胞外结构域,或保留结合cd28的能力的功能性片段。在一些实施方式中,抗cd3scfvfc抗体与异源性gpi锚连接序列融合。在一些实施方式中,抗cd3scfvfc抗体不由复制缺陷型重组逆转录病毒颗粒中的多核苷酸编码。在用于转导和/或遗传修饰包括膜结合抗cd3scfvfc抗体的t淋巴细胞(例如,t细胞和/或nk细胞)的此上段落中的这些方面中,复制缺陷型重组逆转录病毒颗粒可进一步包括多核苷酸,该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码嵌合抗原受体。在一些实施方式中,能够与cd3结合的膜结合多肽并不由复制缺陷型重组逆转录病毒颗粒中的多核苷酸编码。在一些实施方式中,抗cd3scfvfc抗体不由复制缺陷型重组逆转录病毒颗粒中的多核苷酸编码。在另一方面中本文提供一种转导和/或遗传修饰受试者静息淋巴细胞的方法,该方法包含使受试者静息t细胞和/或静息nk细胞体外与复制缺陷型重组逆转录病毒颗粒接触,其中复制缺陷型重组逆转录病毒颗粒包含在其表面上的假型化元件及能够在其表面上与cd3结合的膜结合多肽(但不为能够在其表面上与cd28结合且活化cd28的膜结合多肽),其中该接触有助于利用复制缺陷型重组逆转录病毒颗粒转导静息t细胞和/或nk细胞,从而产生遗传修饰的t细胞和/或nk细胞。在另一方面中,本文提供一种用于转导和/或遗传修饰来自经分离血液的静息t细胞和/或静息nk细胞的方法,该方法包含:自受试者采集血液;分离包含静息t细胞和/或静息nk细胞的外周血单核细胞(pbmc);且使受试者静息t细胞和/或静息nk细胞体外与复制缺陷型重组逆转录病毒颗粒接触有效时间,其中这些复制缺陷型重组逆转录病毒颗粒包含在其表面上的假型化元件及能够在其表面上与cd3结合的膜结合多肽(但不为能够在其表面上与cd28结合且活化cd28的膜结合多肽),从而产生经遗传修饰的t细胞和/或nk细胞,从而转导t细胞和/或nk细胞。在用于转导和/或遗传修饰包括能够在其表面上与cd3结合的膜结合多肽(但不为能够在其表面上与cd28结合且活化cd28的膜结合多肽)的静息t淋巴细胞的此上段落中的这些方面中,假型化元件可以是例如水疱性口炎病毒包膜蛋白(vsv-g)。在说明性实施方式中,能够与cd3结合的膜结合多肽为抗cd3scfvfc抗体,其在一些实施方式中与异源性gpi锚连接序列融合。在此方面的一些实施方式中,执行该接触至少2小时,或在2小时与24小时之间,或在2小时与6小时之间。在一些实施方式中,可检测标记是由复制缺陷型重组逆转录病毒颗粒的基因组编码,且在转导之后在t细胞和/或nk细胞中检测到。在一些实施方式中,能够与cd3结合的膜结合多肽并不由复制缺陷型重组逆转录病毒颗粒中的多核苷酸编码。在一些实施方式中,可检测标记是由复制缺陷型重组逆转录病毒颗粒的基因组编码,且在转导之后在t细胞和/或nk细胞中检测到。在另一方面中,本文中提供一种复制缺陷型重组逆转录病毒颗粒,其包含:一个或多个假型化元件;多核苷酸,其包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码嵌合抗原受体;及在其表面上的假型化元件以及在其表面上的活化元件,其中该活化元件能够与t细胞和/或nk细胞结合且不由复制缺陷型重组逆转录病毒颗粒中的多核苷酸编码,且其中该活化元件为抗cd3scfvfc抗体。在另一方面中,本文中提供一种复制缺陷型重组逆转录病毒颗粒,其包含:一个或多个假型化元件,其能够与t细胞和/或nk细胞结合且有助于与复制缺陷型重组逆转录病毒颗粒至其上的膜融合;多核苷酸,其包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码嵌合抗原受体;及在其表面上的假型化元件以及在其表面上的活化元件,其中该活化元件能够与t细胞和/或nk细胞结合且不由复制缺陷型重组逆转录病毒颗粒中的多核苷酸编码,且其中该活化元件为能够在其表面上与cd3结合的膜结合多肽,但不为能够在其表面上与cd28结合且活化cd28的膜结合多肽。在此上段落中的复制缺陷型重组逆转录病毒颗粒中,该复制缺陷型重组逆转录病毒颗粒进一步包含多核苷酸,该多核苷酸包含可操作地连接至在t细胞和/或nk细胞中有活性的启动子的一个或多个转录单元,其中该一个或多个转录单元编码嵌合抗原受体。在这些方面的一些实施方式中,能够与cd3结合的膜结合多肽并不由复制缺陷型重组逆转录病毒颗粒中的多核苷酸编码。在这些方面的一些实施方式中,抗cd3scfvfc抗体并不由复制缺陷型重组逆转录病毒颗粒中的多核苷酸编码。以下非限制性实施例仅借助于说明例示性实施例而提供,且绝不限制本发明的范畴及精神。此外,应理解,本文所公开或主张的任何发明涵盖本文所描述的任何一个或多个特征的所有变体、组合及排列。任何一个或多个特征可明确地自权利要求排除,即使未在本文中明确地阐述特定排除。也应理解,除非一般熟习此项技术者将理解,否则根据本文公开的特定方法或本领域中已知的其他方法,用于方法中的试剂公开内容意欲与(及为其提供支援)涉及试剂用途的方法同义。另外,除非一般熟习此项技术者将理解,否则说明书和/或权利要求公开一种方法,本文所公开的任何试剂可用于该方法。实施例实施例1.特异性反应于核苷类似物抗病毒药物的核糖开关的产生。此实施例提供一种基于结合鸟苷及脱氧鸟苷的天然结构性核糖开关来筛选文库的方法。这些核糖开关是用作支架以开发偏差文库(biasedlibraries)以供选择与配体核苷类似物特异性结合的适配体。以前,等温滴定热量测定法已用于展示这些天然核糖开关与其天然配体结合。额外测试展示,脱氧鸟苷开关也微弱地与核苷类似物阿昔洛韦及喷昔洛韦相互作用,从而导致将此序列重新设计为新的文库。靶向核糖开关的单链区以选择特异性回应于阿昔洛韦或喷昔洛韦的突变及变体序列。材料自sigma-aldrich(st.louis,mo)订购选择组分:鸟嘌呤、鸟苷、脱氧鸟苷、阿昔洛韦及喷昔洛韦。阿昔洛韦为初始靶标,而喷昔洛韦为在稍后轮次中使用的特别关注的分析物,且鸟嘌呤、鸟苷及脱氧鸟苷是用作反向靶标。待用作分配介质的氧化石墨烯(gro)是购自angstronmaterials(dayton,oh)。hepes(ph7.3)及mgcl2是购自amerscollc.(solon,oh)。kcl是购自teknova(hollister,ca)。在5×(1×为50mmhepes,100mmkcl,20mmmgcl2,ph7.3)下制备选择缓冲液。在无核酸酶的水中重构靶标、反向靶标及寡核苷酸以供初步分析及适配体筛选。制备所有靶标的等份且储存在-20℃下以最大化保质期。适配体文库的产生最初的适配体文库模板由ibagmbh(gottingen,germany)合成,作为图14中序列的反向互补序列。在图14中,框中的核苷酸在已知序列中是单链的,在合成期间引入“突变”以使与原始靶标的类似物更好结合。对于用实线框内的核苷酸,允许替换突变;对于虚线框内的核苷酸,允许替换突变以及插入或缺失。引物由idt(coralville,ia)以单链dna的形式合成。将t7引物(seqidno:240)与文库模板序列组合以供利用titaniumtaqdna聚合酶(clontech;mountainview,ca)进行引物延伸。使用ampliscribet7高产率转录试剂盒(epicentre;madison,wi)转录经引物延伸的材料,然后在用于选择之前在具有8m脲素的10%变性聚丙烯酰胺凝胶电泳(page)上进行纯化。在选择期间,使用superscriptiv逆转录酶(invitrogen;carlsbad,ca)使用反向引物(seqidno:241)来逆转录文库且使用titaniumtaqdna聚合酶(clontech;mountainview,ca)进行扩增。具有seqidno:248的适配体具有-3至-1的j2-3环变化及~2.25×1010的多样性。具有seqidno:250的适配体具有0(天然)至+5的j2-3环变化及~9.38×1014的多样性。两种寡核苷酸(seqidno:249及seqidno:250)以1:4160的比率混合以在经组合文库池中产生等摩尔多样性,具有~9.38×1013的总多样性。文库筛选使用氧化石墨烯-通过指数富集的配体系统进化(go-selex)方法(图15)(park等人,2012),利用使氧化石墨烯针对单链核酸有高亲和力的π-π相互作用(zeng等人,2015)来进行文文库筛选。目标是选择不与1×选择缓冲液或不与反向靶标(鸟嘌呤、鸟苷及脱氧鸟苷)相互作用但与阳性靶标阿昔洛韦结合的序列。对于每个轮次,首先在1×选择缓冲液中对给定量文库进行重新折叠(在90℃下变性5分钟,在4℃下5分钟,然后在室温下)。然后将反向靶标添加至经重新折叠的文库中且在37℃下孵育30分钟。例外情况为轮次1及轮次2,其中仅短暂地(<1分钟)包括反向靶标以帮助将文库装载至gro上。在使文库与反向靶标及缓冲液组分相互作用之后,历经在37℃下孵育10分钟将未结合文库装载至gro上(质量等于轮次开始时文库质量的100倍)。然后以7,000×g离心溶液以沉降gro。移除含有与反向靶标和/或缓冲液结合的序列的上清液。沉降物然后用200μl1×选择缓冲液洗涤两次,以7,000×g离心且在每次洗涤后移除上清液。然后添加含阳性靶标的溶液且允许在37℃下多至60分钟的表1中指示条件下自gro洗脱文库,基本上允许靶标针对文库结合与氧化石墨烯竞争。与靶标更强有力地结合的序列将自氧化石墨烯解吸附且在孵育末期保持与靶标结合。最后的离心步骤将位于上清液中的释放材料与保持与氧化石墨烯结合的非响应文库分开。在阳性选择后,用8m尿素用10%变性page纯化回收的rna,然后使用分光光度计读数(表1)进行定量,用superscriptiv逆转录,并用titaniumtaqdna聚合酶进行pcr扩增。扩增产物转录成rna用于下一轮选择。在选择的过程中实施三级严格性(表1)。前两轮选择不包括针对反向靶标进行筛选,以最大限度地使文库装载上gro。另外,阳性孵育中使用大量过量阿昔洛韦以最大化文库回收率,因此是低严格性设计。在实现文库回收之后引入反向靶标孵育,作为中等严格性条件。也在这些三个轮次期间降低阿昔洛韦与文库的比率以增加针对靶标结合的文库竞争。一旦实现大于10%的回收率,则实施高严格性选择的最终轮次。保持高的反向靶标/文库比率并使阳性靶标/文库比率为1:1,同时减少阳性孵育时间,以选择更快结合的序列。一旦文库回收率在高严格性条件的多于两个轮次之后展示为保持超过10%,则进行平行评估。表1.选择和评估条件。用于每轮选择或孵育的条件,回收率为针对各轮次的经回收样品与输入文库之间的比率。在选择期间监测文库富集。*反向靶标用于装载,而非延伸孵育。与合并的反向靶标进行预加载孵育。用阳性靶标阿昔洛韦进行预装载孵育。这是为了尽量减少交叉反应物质的回收。本表中使用以下缩写:“x-靶标”为反向靶标;“(+)靶标”为阿昔洛韦或喷昔洛韦;“(+)孵育时间(分钟)”为“文库:(+)靶标”溶液于gro上孵育的时间。g0为第0代,以此类推;r1是第1轮,以此类推。对于平行评估(平行1及平行2),用以下执行孵育:(-)为仅1×选择缓冲液,(×)为含反向靶标的1×选择缓冲液,(+)为含阿昔洛韦的1×选择缓冲液,及(p)为含喷昔洛韦的1×选择缓冲液。对于两个平行评估,将待评估的文库分为四个相等量以用于如上制备及重新折叠(图16)。对于每种条件,将50皮摩尔的文库与1×选择缓冲液组合,重新折叠(90℃下5分钟,4℃下5分钟),且然后在37℃下与200μl10μm含经合并的反向靶标的1×选择缓冲液一起孵育30分钟。然后将这些样品装载等于样品中文库质量100倍的量的氧化石墨烯中,并在37℃下孵育10分钟,且然后如前述用200μl的1×选择缓冲液洗涤两次。然后在37℃下将经装载氧化石墨烯样品与含200μl的合适评估条件(仅1×选择缓冲液,10μm经合并反向靶标,1μm喷昔洛韦,1μm阿昔洛韦(第一平行评估),或0.5μm阿昔洛韦(第二平行评估);在表1中,这些条件分别展示为:(-)、(x)、(p)、(+)及(+))的1×选择缓冲液一起平行孵育30分钟。最终离心步骤将无反应的经氧化石墨烯结合的文库分离经解吸附反应文库。反应文库使用分光光度计读数来定量(表1),使用具有8m脲素的10%变性page来确认,且准备用于第二平行评估。该后续评估继续使用反向靶标来进行阳性样品的预加载孵育,但对于每个其他样品的预孵育使用了阳性靶标阿昔洛韦。这样做是为了最小化给定样品中的交叉反应序列的表示(即,反应于阳性样品中的反向靶标,反应于阴性反向靶标或喷昔洛韦样品中的阿昔洛韦)。自第二平行评估回收的材料是使用分光光度计读数来定量(表1),使用具有8m尿素的10%变性page来确认,且利用逆转录及pcr以产生双链dna进行测序。测序初始文库经10个轮次以上的基于gro的选择及和平行评估(表1)。go-selex方法被设计为在多轮选择中富集序列,其结合给定的目标靶标并去除结合非靶标化合物或缓冲剂组分的序列。因此,要测序的群体预计会包含潜在候选适配体的多个拷贝。实施illuminamiseq系统(sandiego,ca)以在平行评估之后使用单端读取技术测序适配体文库。深度测序与后续数据分析减少传统selex所需的大量筛选轮次,这可由于筛选方法引入误差及偏差(schütze等人,2011)。测序以下五个样品:反应于阿昔洛韦的最后一代文库,反应于反向靶标的最后一代文库,反应于1×选择缓冲液(阴性条件)的最后一代文库,反应于阿昔洛韦的倒数第二代文库,及反应于感兴趣的额外靶标(喷昔洛韦)的最后一代文库。根据这些数据集,以95%同源性(考虑到突变、缺失及插入的序列相似性)构建序列家族以用于适配体候选物鉴别。存在来自反应于阿昔洛韦的文库的1,711,535个原始序列(124,600个唯一序列)及来自反应于喷昔洛韦的文库的2,074,832个原始序列(110,149个唯一序列)。适配体候选物选择序列家族构建主要聚焦于序列相似性。这意味着计入阳性靶标群体中序列的频率,但强调的是相似序列之间的变化程度,其中95%同源性为最小要求(在整个序列中100%匹配对于加入家族并不是必需的,可错配、插入或缺失至多2个碱基)。因此,预期拥有具有最多成员的家族能够作为适配体候选物有高的评级。在构建家族之后,考虑到家族在某一给定群体中的相对存在-频繁出现在阴性及反向靶标群体中的家族被视为较弱的候选物,这是因为其在与缓冲液或反向靶标组分结合时显示一定程度的非特异性相互作用。另外,显示高富集率(即,最终阳性群体与倒数第二阳性群体之间的较大比率)的家族提高其候选资格,这是因为富集率已与候选物相对于剩余群体的结合亲和力相关(levay等人,2015;wang等人,2014)。在这些条件下,若干候选物家族显示出为针对结合阿昔洛韦(表2)及喷昔洛韦的强有力候选物。表2.用于结合阿昔洛韦及喷昔洛韦的候选物。dna序列对应于结合阿昔洛韦的rna核糖开关的非茎区。在筛选时鉴别到七个家族:582、769、795、935、946、961及996,其中在各家族中具有在1个与39个之间的序列。将家族中各序列的同一性百分比与各家族内最常见的序列(582-1、769-1、795-1、935-1、946-1、961-1及996-1)进行比较。还将家族中各序列的同一性百分比与野生型序列进行比较。阳性靶标阿昔洛韦产生对应于582-1(seqidno:108)、769-1(seqidno:147)、795-1(seqidno:164)、935-1(seqidno:183)、946-1(seqidno:198)、961-1(seqidno:220)及996-1(seqidno:221)的七个强有力候选物(seqidno:87至seqidno:93;包括茎区的rna序列),各自命名为f1a(图17)。这些序列为各家族中最常见的序列(各家族所有成员dna序列为:582(seqidno:108至seqidno:146);769(seqidno:147至seqidno:163);795(seqidno:164至seqidno:182);935(seqidno:183至seqidno:197);946(seqidno:198至seqidno:219);961(seqidno:220);及996(seqidno:221))。共有序列展示各家族各核苷酸位置处所有可能的替换或空位(seqidno:222至seqidno:226)。由于目标为从基于已知与脱氧鸟苷结合的rna的文库鉴定适配体,所以强有力候选物需要在反向靶标群体中具有最少存在。候选物f1a-795、f1a-935及f1a-946非常符合此标准,这是因为未在反向靶标群体中检测到这些候选物。就此而言,将f1a-996及f1a-961视为第二最佳候选物,尽管其在反向靶标群体中有较小程度的出现。另外,候选物应最低程度地出现于阴性群体中,这是因为这些序列自gro解吸附而没有阿昔洛韦的影响且可表示假阳性。f1a-935及f1a-946在此标准下非常好的理想地表现,因为其在阴性群体中没有发现。候选物f1a-769在阴性群体中被最小程度地检测到,而候选物f1a-961、f1a-795及f1a-996表现地优异程度较低。富集率是要考虑的最终条件,其中f1a-935、f1a-946及f1a-769的表现足够。包括候选物f1a-582,这是因为它展现最大富集率,尽管其在其他标准下表现并不优异。剩余候选物没有优于这四个的表现,但展现可接受的特性。额外靶标喷昔洛韦产生七个强有力候选物(seqidno:94至seqidno:100),各自命名为f1p(图18)。如前,目标是从基于已知与脱氧鸟苷结合的rna的文库来鉴别适配体,在第10轮次之后与在阿昔洛韦结合(阿昔洛韦)方面富集的文库偏离。强有力候选物需要在阿昔洛韦及反向靶标群体两者中具有最小存在以最小化交叉反应性。候选物f1p-923符合第一个标准,候选物f1p-710符合第二个标准,且候选物f1p-584在一定程度上符合两个标准。候选物f1p-584也显示相对于阴性条件对喷昔洛韦的适中偏好,以及相对于针对阿昔洛韦的前代反应的适中富集。剩余候选物显示相对于阿昔洛韦对喷昔洛韦的最小偏好或相对于反向靶标对喷昔洛韦的最小偏好(分别为f1p-837及f1p-932;f1p-991及f1p-718)。这四个候选物显示相对于阴性条件对喷昔洛韦的一些偏好,这最小化假阳性的机会,但在候选物不显示喷昔洛韦优于其类似物的选择性时,该标准并不重要。富集率是要考虑的最终条件,其中f1p-923、f1p-932及f1p-584表现足够。对所选择的适配体进行定性page评估。经个别合成及转录的适配体在生理mg++(0.5mm)下经受氧化石墨烯(gro)选择,且用阿昔洛韦(+)或反向靶标(x)洗脱。将各样品的特异性洗脱的适配体级分进行page分析。将100皮摩尔的各适配体候选物(每实验/泳道)重悬于1×修饰的选择缓冲液(50mmhepes,100mmkcl,0.5mmmgcl2,ph7.3)中且重新折叠(90℃下5分钟,然后4℃下5分钟),然后在37℃下与200皮摩尔(每一)合并的反向靶标或靶标一起孵育30分钟。最终文库浓度为0.5μm,靶标/反向靶标浓度为1μm(孵育体积为200μl)。在靶标/反向靶标孵育之后,添加250μggro(angstronmaterials(dayton,oh))以吸附未结合的候选物(在37℃下孵育10分钟)。以7,000×g离心样品5分钟。回收上清液,使用具有40mmedta的2×甲酰胺变性,且在具有8m尿素的10%变性page上电泳(供应商:americanbioanalytical;目录号#ab13021-01000.ab13022-01000)。电泳缓冲液为1×tbe(供应商:amresco/vwr;目录号#0658-20l,使用di水稀释)。dna梯为20/100dna梯(idt)。凝胶用凝胶星型(lonza,50535)染色且在蓝光透射仪上成像。候选物f1a-769、f1a-795、f1a-946及f1a-996在该定性page评估中表现出选择性阳性应答(使用阿昔洛韦靶标从gro良好的洗脱且用反向靶标的相对较低或最小洗脱的适配体)。结论经过12轮反复筛选和平行评估后,确定了阿昔洛韦的强有力候选物;经过2轮筛选和平行评估后确定了喷昔洛韦的合理候选物。实施例2.条件性scfv's的分离移除潜在剪接位点文库,并通过重叠寡核苷酸合成来合成肿瘤抗原特异性scfv's,且将其克隆至含有阿昔洛韦反应性元件及灵长类cd3ζ启动子的car穿梭式构建体中。作为初始原型,利用具有cd8-α信号肽、柄及跨膜结构域的erbb2scfv或epcam的抗ecd。使用如美国专利第8,709,755号及pct公开案第wo/2016/033331a1号中所描述的方法或通过在许可及非许可条件下自人类噬菌体文库直接选择来产生受实体肿瘤微环境限制的car产物。简而言之,在以下肿瘤许可条件下淘选来自creativebiolabs(shirley,ny)的人类vh×vl文库:100μg/ml透明质酸,100kda级分(lifecorebiomedical,chaska,mn),20mg/ml重组hsa(cyagen,santaclara,ca),含200ng/ml重组人类vegf的25mm碳酸氢钠缓冲液,2μm腺苷,10mm乳酸钠ph6.7,然后用与生物素化人类igg结合的链霉亲和素珠(thermofisher,carlsbad,ca)清除。在许可条件下进行37℃下与epcam及erbb2缀合珠的生物素化靶标受体ecd的结合,然后在许可条件下系列洗涤。用生理条件(1μg/ml透明质酸,20mg/mlhsa,25mm碳酸氢盐,1mm乳酸钠ph7.2),然后用酸性洗脱液洗脱封闭性变体(tightvariant)且用1mtris快速中和来释放噬菌体。扩增噬菌体且使用长读取测序(pacbio,menlopark,ca)分裂基因组dna以供vh×vl链的深度序列分析。可重复淘选以供富集。在经由对靶标富集的噬菌体培养过程期间展示较佳扩增读数的vh×vl序列被排除用于进一步分析。与在肿瘤许可条件下富集但在生理条件下释放的靶标选择性结合的噬菌体被选择以供进一步通过以下表征:克隆至car构建体表达系统中,慢病毒世代且转导至t细胞中以供测试表达car介导的肿瘤抗原的靶标细胞在肿瘤选择性环境中相比于生理条件下的杀伤。实施例3.使用微环境限制的scfv's产生mrb-car。通过利用如申请案wo/2016/033331a1中所描述的进化使vh及vl序列对低ph微环境具有低选择性而获得微环境限制性astr。通过将微环境限制的单链抗体的重链及轻链与其他car结构域一起整合入慢病毒表达载体中以产生一系列候选mrb-car来制得用于结合两种同源肿瘤抗原(靶标1或靶标2)中任何一种的嵌合抗原受体(car),其相比于正常组织mrb-car的微环境,在肿瘤微环境降低的ph下具有增加的活性。这些mrb-car包括模块的多种组合。mrb-car从氨基至羧基末端在位置1至位置9中包括:cd8信号肽(sp)(p1)(seqidno:74);微环境限制的抗靶标1astr或抗靶标2astr(p2至p4);在t2a及t2b情况下来自cd8的柄及跨膜(tm)结构域(seqidno:75)(p5)及来自cd137的共刺激结构域(p6)(seqidno:1)或在t1a情况下来自cd28的柄及跨膜(tm)结构域(seqidno:76)(p5)及来自icδ的共刺激结构域(seqidno:3)(p6);来自cd3z的活化结构域(seqidno:13)(p7);2a-1核糖体跳跃序列(seqidno:77)(p8);及例示性etag(seqidno:78)(p9)。用重组慢病毒颗粒转导泛性t细胞(allcells,alameda,ca)以表达该系列的候选mrb-car,且通过使用facs测定表达etag的细胞的百分比来计算经转导细胞的百分比。用编码候选mrb-car的重组慢病毒颗粒成功地转导了泛性t细胞。利用xcelligence系统(acea)在ph7.4(正常组织)或ph6.7(肿瘤微环境下降低的ph)下分析候选mrb-car针对表达靶标1或靶标2的靶标细胞(分别为cho-靶标1及cho-靶标2)的细胞毒性活性。简而言之,在实验前一天将表达靶标1或靶标2的靶标细胞以20,000个细胞/孔接种至具有肿瘤条件介质或正常介质的96-孔e-盘中。使效应细胞在含有100iu/ml的il-2的人类t细胞介质中静息两天且以3:1、1:1及0.3:1的效应细胞/靶标细胞比率(e/t)添加至含有靶标细胞的实验孔中。在添加效应细胞之后每5分钟进行阻抗读取持续大约40小时且阻抗报告为细胞指数(ci)。如下计算特异性细胞溶解百分比:((ci靶标+经对照病毒转导的效应t细胞)-(ci靶标+用针对靶标1或靶标2的car转导的效应t细胞))/(ci靶标+经对照病毒转导的效应t细胞)×100。结果许多候选mrb-car在ph6.7下比在ph7.4下对靶标细胞具有更高细胞毒性活性。在ph6.7下比在ph7.4下在裂解靶标细胞时更有效的例示性mrb-car包括mrb-cart1a、mrb-cart2a及mrb-cart2b。mrb-cart1a的astr自5'至3'包含由接头1(seqidno:283)隔开的靶标1mrbvh(seqidno:281)及靶标1mrbvl(seqidno:282)。mrb-cart2a的astr自5'至3'包含由接头2(seqidno:291)隔开的靶标2mrbvh(seqidno:289)及靶标2mrbvl(seqidno:290)。除交换vh及vl的位置以外,mrb-cart2b的astr与mrb-cart2a的astr相同。实施例4.配体可诱导的核糖开关的构建。将脱氧鸟苷核糖开关适配体及鸟苷核糖开关适配体(pikovskaya,2014;kim,2007)或其他嘌呤核糖开关适配体合成为寡核苷酸。在一个实施例中,选择来自mesoplasmaflorum的脱氧鸟苷ia核糖开关(在图6及图7中加下划线且呈粗体)以用于进化产生阿昔洛韦反应性核糖开关。在另一实施例中,选择来自枯草芽孢杆菌的鸟苷xpt核糖开关(在图10及图11中加下划线且呈粗体)以进化产生阿昔洛韦反应性核糖开关。对于这些两个实施例中的每一个,在p2、p3、j1-2及j2-3区段(图7及图11)中用替代核苷酸在靶向序列位置处产生随机rna文库。各区段在各靶向序列位置处允许3个替代核酸,或替代地在各靶向序列位置的+1位点中允许4个核苷酸的碱基缺失及插入以供如图8a至图8b及图9(m.florumia)与图12a至图12b及图13(枯草芽孢杆菌xpt)中所指示的饱和诱变。引物延伸及试剂制备是在rna转录后。所得rna文库在鸟嘌呤、鸟苷及脱氧鸟苷的存在下在氧化石墨烯上进行阴性选择,然后为用阿昔洛韦或喷昔洛韦进行阳性选择。在阴性及阳性选择过程中,使用人类细胞生理镁水平(0.5mm至1.2mm)且温度保持在37℃下。对经回收适配体进行逆转录且pcr扩增,然后进行转录且随后筛选至少8个连续轮次的选择。在平行方法中,在40℃下用额外阴性筛选来筛选适配体。通过nextgen测序及分析来检查所得阳性池。合成各个适配体且利用在35℃至40℃下在人类细胞生理镁水平的等温热量测定法来检查亲和力。然后进行阳性阿昔洛韦及喷昔洛韦特异性适配体的选择,将适配体与核酶锤头型及枪式核酶整合在一起。将阳性阿昔洛韦选择性适配体与枪式核酶组合以鉴别被阿昔洛韦调节的核酶。(harriskarna.2015nov;21(11):1852-8.doi:10.1261/rna)。变体在存在阿昔洛韦下及不存在喷昔洛韦下经受基于凝胶移位的page纯化。另外,将无环鸟苷选择性核糖开关置放于环中紧邻car/il-7构建体的剪接受体上游的3′。在不存在阿昔洛韦下,剪接位点位置在核糖开关复合物中结合,但在存在阿昔洛韦下变得可接近,从而产生功能性car转录物。实施例5.体内增殖结构域的构建。利用重叠寡核苷酸合成(dna2.0,newark,california)来合成来自t细胞淋巴母细胞白血病的一系列组成型活化il7受体(il7r)跨膜突变体(243insppcl(seqidno:82);246inskch(seqidno:101);241insfscgp(seqidno:102);244inschl(seqidno:103);及244insppvcsvt(seqidno:104);全部均来自shochat等人2011,j.exp.med.vol.208no.5901-908)。将经合成的组成型活化il7r跨膜突变体插入至紧邻在2a核糖体跳跃序列后面的组成型表达慢病毒载体骨架中,然后为抗cd19cd3ζ表达盒,该表达盒包括cd8a柄(seqidno:79)及前导肽(seqidno:74)。hek293包装细胞用il7r跨膜突变体慢病毒载体及慢病毒包装构建体来转染,进行生长,并使用本领域中已知方法采集病毒上清液。经cd3/cd28刺激的t细胞用病毒上清液来转导且在il2缺陷型aimv、ctsoptmizert细胞扩增sfm或x-vivo15介质中生长4周,每周补充来自相同供体的冷冻pbmc。通过facs分选克隆表达il7r变体的所得扩增的转导t细胞,并通过测序rt-pcr产物鉴定il7r构建体的序列。选择243insppcl变体(ppcl)(seqidno:82)以供进一步进化以产生条件性活化的car。实施例6.用于car-t活化及增殖的辅助组分的筛选。一系列蛋白编码结构域(abcg1、socs1、smad2、tgfbr2、ccbl及pd1)及mirna序列被构建用于整合入cd3-启动子驱动的car盒的反向链上的合成内含子中。含有cd3-启动子驱动的car盒及蛋白编码结构域或mirna序列的各构建体包括用于深度测序的唯一条码且使用gibson组装来组装,然后转化,及在大肠杆菌(e.coli.)中进行文库扩增。制备病毒原液且用于在不具有il2的aimv、ctsoptmizert细胞扩增sfm或x-vivo15介质中转导经cd3/cd28刺激的t细胞,且使其在连续取样dna的情况下在培养物中生长4周,所述dna用于扩增及密码鉴别的深度测序。文库也经受pacbio全长测序以测定文库多样性且解码条码组分。针对carcd3ζ结构域的协同活化测试mirna序列及蛋白编码结构域。实施例7.将逆转录病毒包装及转导系统改造以靶向静息t细胞以供选择t细胞整合及由pbmc表达。尽管利用瞬时转染产生高效价慢病毒载体系是可行的,但此方法带有产生复制缺陷型逆转录病毒(rcr)的风险且对于临床应用而言不可扩大规模。在此,通过将编码可诱导启动子及其调节子的多个构建体同步引入至hek293悬浮适应性细胞(hek293s)中以稳定地产生病毒组分、car基因及其调节组分来产生稳定的逆转录病毒包装细胞系。两个不同的可诱导系统可用于对基因表达进行时间性控制。一个系统是基于两种转录因子的雷帕霉素-或雷帕霉素类似物诱导的二聚化。一个转录因子由与zfhd1dna结合结构域融合的三个拷贝的fkpb蛋白质组成,且另一转录因子由与p65活化结构域融合的frb蛋白质组成。雷帕霉素或雷帕霉素类似物将转录因子二聚化以形成zfhd1/p65ad,且可在12xzfhd1结合位点处活化基因转录。如图3a至图3e中所展示的一系列载体是通过侧翼转座子序列产生的以用于整合至hek293s基因组中。一旦整合至细胞的基因组中,这些序列充当调节组分及lox位点和/或frt位点(在此被称为着陆点)以供随后使用cre和/或flp重组酶进行整合。初始5个构建体含有编码嘌呤霉素抗性、gfp、rfp及经由n端plss(牛促乳素信号肽)靶向细胞膜且经由衍生自血小板的生长因子受体(pdgfr)c端跨膜锚定结构域锚定至细胞膜的胞外myc标签的多核苷酸序列。初始5个构建体也可包括组成型最小cmv启动子及最小il-2启动子、雷帕霉素调节的基于zfhd1的启动子、四环素反应性元件(tre)启动子或双向tre(bitre)启动子。图3a中的构建体含有编码与nfκbp65活化子结构域(p65ad)融合的frb结构域及与组成型表达的三个fkbp重复融合的zfhd1dna结合结构域的多核苷酸序列。图3a中的构建体也包括在雷帕霉素可诱导的zfhd1/p65ad启动子下的hiv1rev及hsvvp65结构域srcflagvpx。图3b中的构建体包括编码在zfhd1/p65ad启动子控制下的rtta序列的多核苷酸。图3c中的构建体包括编码侧接有loxp位点的嘌呤霉素抗性基因及侧接有lox2272位点的胞外myc标签的多核苷酸。这些可选标记二者均在bitre启动子的控制下,该bitre启动子侧接有frt位点。图3d中的构建体包括编码在tre启动子控制下侧接有loxp位点的gfp的多核苷酸。图3d中的构建体也包括在tre启动子与gfp的5′loxp位点间的单个frt位点。图3e中的构建体包括编码在zfhd1/p65ad启动子控制下侧接有loxp位点的rfp的多核苷酸。图3e中的构建体也包括在zfhd1/p65ad启动子与rfp的5′loxp位点间的单个frt位点。图3c至图3e中的构建体充当其他多核苷酸序列的着陆点以插入至包装细胞系的基因组中。待插入的多核苷酸序列可侧接有lox位点且使用cre重组酶及loxp位点插入至基因组中。这导致编码嘌呤霉素抗性、胞外myc标签、gfp及rfp的基因组区的插入及同步移除。替代地,多核苷酸序列可侧接有frt位点,且使用flp重组酶及frt位点来插入至基因组中,然后使用cre重组酶移除编码嘌呤霉素抗性、胞外myc标签、gfp及rfp的多核苷酸序列。为用整合至基因组中的着陆点产生包装细胞系,用等摩尔浓度的5种质粒(图3a至图3e)加5μg经体外转录的piggybac转座酶mrna或5μg具有用于在pei存在下以2:1或3:1的pei比dna(w/w)比率表达piggybac转座酶的启动子的质粒或2μg至5μgpiggybac转座酶蛋白使用阳离子肽混合物来共转染hek293s细胞。在100nm雷帕霉素及1μg/ml多西环素的存在下用嘌呤霉素选择经转染细胞2至5天,然后荧光活化的细胞分选以收集表达gfp及rfp的细胞。经分选细胞在不存在嘌呤霉素、雷帕霉素及多西环素的情况下生长5天,且移除表达gfp及rfp的细胞,也用myc珠粒移除myc阳性细胞。然后筛选来自经阴性分选的细胞的各个克隆以筛选雷帕霉素及多西环素诱导gfp及rfp,以及克隆单个细胞。采集来自克隆的dna且经测序以用于整合分析。在存在雷帕霉素及多西环素下具有gfp及rfp的强有力诱导表达且在不存在雷帕霉素及多西环素下具有有限背景表达的阳性克隆进行扩增并保存。然后用含有三顺反子多核苷酸的构建体转染基因组中整合了来自图3a至图3e的构建体的hek293s细胞,该三顺反子多核苷酸编码daf信号序列/抗cd3scfvfc(ucht1)/cd14gpi锚连接位点(seqidno:287)、能够结合cd28/cd16bgpi锚连接位点的daf信号序列/cd80胞外结构域(seqidno:286)及daf信号序列/il-7/daf(seqidno:286)及侧接用于整合至hek293s基因组中的多核苷酸区的转座子序列(图4a)。在转染后,细胞在不存在雷帕霉素及多西环素下扩增2天且选择是组成型红色的克隆。然后用用于表达cre重组酶的构建体瞬时转染阳性克隆以移除剩余基因组dna及rfp编码区。同时转染另一构建体(图4b),其含有在一个方向上的具有bitre启动子的多核苷酸及编码gag多肽及pol多肽的多核苷酸区以及在另一方向上的编码麻疹病毒f蛋白质及h蛋白质的多核苷酸区。cre重组酶将构建体整合至基因组中以产生展示于图4b中的经整合序列。针对在多西环素及雷帕霉素存在下的蛋白表达评估所得克隆,且通过深度测序进行分析用于基因组整合。保留剩余的tre反应性gfp位点以用于慢病毒基因组插入。实施例8.慢病毒载体及逆转录病毒包装的产生。用用于表达flp重组酶的构建体(图4c)及含有多核苷酸序列的构建体转染实施例7中产生的逆转录病毒包装稳定细胞系,该多核苷酸序列编码在cd3z启动子(其在hek293s细胞中无活性)控制下的淋巴增殖性元件il7rα-insppcl及car,其中car及il7rα-insppcl由编码t2a核糖体跳跃序列的多核苷酸序列隔开,且il7rα-insppcl具有阿昔洛韦核糖开关控制的核酶。含car的构建体进一步包括cppt/cts及rre序列以及编码hiv-1psi的多核苷酸序列。待被整合至基因组中的含car的构建体的整个多核苷酸序列侧接有frt位点。含car的构建体的成功整合造成gfp组成型表达,其因此通过用用于表达cre重组酶的构建体瞬时转染而移除。hek293s系在无血清介质中生长。在经搅拌罐反应器中生长至峰值细胞密度后,将细胞稀释至70%峰值细胞密度且用100nm雷帕霉素处理2天以诱导早期基因rev、vpx及acd3scfvcd16bgpi、acd28scfvcd16bgpi及il-7sdgpidaf的表达,然后在介质中添加1μg/ml多西环素以诱导如gagpol、mv(ed)-fδ30、mv(ed)-hδ18的结构化元件及包括治疗性靶标的慢病毒基因组的表达。病毒产量的水平是由包装序列的qpcr及p24elisa来检查。通过细胞深度过滤及使用tff盒的浓缩/渗滤,然后快速冷冻以供装入小瓶来采集病毒。实施例9.外周血单核细胞(pbmc)分离、转导及扩增以下实施例说明在体内扩增前用于离体处理pbmc的封闭系统的用途。作为一实施例,用酸性柠檬酸盐葡萄糖溶液(acd)作为抗凝剂自受试者抽取30ml至200ml人类血液至血液收集袋中。替代地,将血液抽取至vacutainer管、注射器或等效物中且转移至空血液收集袋或iv袋中。根据制造商说明书,在sepax2细胞处理系统(biosafe)上使用纯细胞试剂盒(目录号#cs-900.2,omniamed)处理全血。将外周血单核细胞(pbmc)收集至培养袋中,或替代地注射器中。无菌地等份采集以用于细胞计数以测定存活细胞数目。在具有至多200ml最终体积的具有10iu/ml至300iu/mlil-2(目录号#202-il-010,r&dsystems)的x-vivo15(目录号#08-879h,lonza)或ctsoptmizer细胞扩增sfm(目录号#a1048501,thermofisherscientific)介质中将最终浓度为0.1×106至1.0×106个存活细胞/毫升的pbmc转移至g-rex100mcs透气细胞培养系统器件(wilsonwolf)。除il-2以外,可将cts免疫细胞sr(目录号#a2596101,thermofisherscientific)添加至介质。根据制造商说明书,封闭的g-rex透气细胞培养系统器件可预涂布有retronectin(目录号#ch-296,takara)或类似衍生自纤连蛋白的等效物。将自外周血液分离的pbmc装载至pallpbmc过滤器上,经由过滤器用10mlaimv(thermofisherscientific)或x-vivo15介质洗涤一次,然后在37℃下以5毫升/小时用10ml至60ml慢病毒原液(如实施例8中所制备)灌注。然后再次用含有重组人类dna酶(pulmozyme,genentech)的aimv、ctsoptmizert细胞扩增sfm或x-vivo15介质洗涤pbmc,然后用无dna酶林格氏乳酸盐(lactatedringers)(目录号#l7500,braun)洗涤。然后经由过滤器将pbmc反灌注至注射器中。然后经由静脉内输注将细胞(细胞的目标水平为5×105至1×106个细胞/kg)重新输注至受试者中。视逆转录病毒基因组内含有的核糖开关而定,将相应核苷类似物抗病毒药物或核苷类似物抗病毒前药(阿昔洛韦、伐昔洛韦、喷昔洛韦或泛昔洛韦)给予受试者。可经口每天三次将任何治疗有效剂量(例如500mg核苷类似物抗病毒药物或前药)给予受试者。用核苷类似物抗病毒药物或前药治疗优选地在重新输注前(例如2小时前)开始,也可在重新输注时或重新输注一些时间后开始。治疗可继续至少1、2、3、4、5、7、10、14、21、28、30、60、90、120天或5、6、9、12、24、36或48个月或更长。治疗可包括每天一次、两次、三次或四次施用核苷类似物抗病毒药物或前药。在重新输注及治疗开始后,使用qpcr以定量病毒基因组的量在重新输注后第2天、第5天、第7天、第11天、第13天、第18天、第28天及第56天经由血液计数来测定经感染细胞数目。经历发热或细胞因子释放综合症的受试者可使核苷类似物抗病毒药物或前药的剂量或频率减少或停止。若经感染t细胞未能在第18天扩增10,000至100,000倍,则可增加核苷类似物抗病毒药物或前药的剂量或频率。可经由fdgpet成像及系列ct扫描来测量受试者的临床反应。可在延长的缓解期后或在过量t细胞增殖超出总外周t细胞计数的30%的情形下减少或停止核苷类似物抗病毒药物或前药的口服剂量实施例10.增加血管或组织ph的治疗性干预。为减少抗原结合结构域与其同源抗原的结合,将nahco3作为iv团注或通过iv输注来施用。标准剂量为1mg/kg体重作为初始剂量,然后每10分钟0.5mg/kg。50毫升nahco3团注将增加血清ph大约0.1ph单位。若ph为7.0,则其需要四个50meq安瓿nahco3以将ph校正为7.40。实施例11.测试pbmc中il-7受体淋巴增殖性/存活元件的活性。为测试il-7rα变体介导t细胞的抗原非依赖性存活的能力,用枸橼酸葡萄糖(acd)作为抗凝剂将三十毫升人类血液抽取至vacutainer管中。遵循制造商说明书使用用ficoll-pacquetm(generalelectric)的密度梯度离心来处理全血,以获得外周血单核细胞(pbmc)。将pbmc的等份与x-vivotm15介质(lonza)一起无菌转移至12孔组织培养盘的孔中,至含50万存活细胞/毫升的1ml最终体积中的最终浓度。在一些样品中也将重组人类白介素-2(il-2)(novoprotein)添加至浓度为100iu/ml。在50ng/ml浓度下添加活化抗cd3ab(okt3,novoprotein),以活化pbmc以供病毒转导。在37℃及5%二氧化碳下在标准湿润组织培养孵育箱中孵育盘过夜。在孵育过夜之后,在5重感染(moi)下将含有所要测试构建体(图19a)的慢病毒颗粒制剂添加至各个孔。在37℃及5%二氧化碳下在标准湿润组织培养孵育箱中孵育盘过夜。在孵育过夜之后,收集12孔盘各孔中的内含物且离心以获得沉淀。用d-pbs+2%人类血清白蛋白(hsa)洗涤样品一次,将其再悬浮于x-vivo15tm介质中,且转移至g-6-孔透气细胞培养器件(wilsonwolf)的孔中。添加额外x-vivotm15介质以使各孔的最终体积达至30ml。将每一构建体的匹配对照样品转移至g-6-孔透气细胞培养器件(wilsonwolf)的孔中且添加额外介质以使最终体积达至30ml,针对一些对照样品用100iu/mlil-2。在37℃及5%二氧化碳下在标准湿润组织培养孵育箱中孵育g-器件7天。在培养期间每2至3天将新鲜il-2添加至含有il-2的对照样品。不具有il-2的经匹配测试样品不进行补充。移除样品以供在第7天追踪在扩增期间的细胞数目及存活率(countess,thermofisher)。图19a提供所测试的il7rα构建体的示意图。将这些构建体插入至重组慢病毒基因组中。复制缺陷型重组逆转录病毒颗粒用于转导pbmc。图19a展示野生型il7rα(seqidno:229)的示意图,其由信号序列(ss)、胞外结构域(ecd)、跨膜(tm)结构域及胞内结构域(icd)组成。“1”指示纤连蛋白iii型结构域位点;“2”指示wsxws基序的位点;“3”指示box1位点;“4”指示蛋白激酶c(pkc)磷酸化位点的位点;及“5”指示box2位点。变体“a”为在位置243处具有insppcl(shochat等人2011,j.exp.med.vol.208no.5901-908)但不具有s185c突变的il-7rα,其在具有gfp多肽、gsg接头及与其n端融合的p2a核糖体跳跃序列的转录物上表达。变体“b”为具有gfp多肽、gsg接头及与其n端融合的p2a核糖体跳跃序列以及信号序列与胞外结构域之间的myc标签的il-7rαinsppcl。变体“c”类似于变体“b”,除其胞内结构域在位置292处截短以外。变体“d”类似于变体“a”,除其胞内结构域在位置292处截短以外。变体“e”为在其n端处截短使得信号序列及大部分胞外结构域(残基1至残基228)不存在的il-7rαinsppcl变体;变体“e”自氨基端依次也具有gfp多肽、gsg接头、p2a核糖体跳跃序列及与n端融合的etag。氨基酸残基编号基于il7rα(ncbigi第002176.2号)。使用trypanblue排除法针对在存在或不存在il-2下的存活率测试含有每一变体的t细胞。如图19b中所展示,pbmc需要il-2以供体外存活。如图19b中所说明,未转染的pbmc在存在il-2下具有约80%存活率且在不存在il-2下具有0%存活率。具有il-7rαinsppcl全长版本(图19a中的il-7rα变体a及b)的pbmc在不存在il-2下具有大于20%的存活率,从而指示组成型活化的il-7rαinsppcl受体的表达在这些细胞中具有存活活性。此外,相比于野生型il-7受体,表达具有经截短的胞内结构域(icd)的il-7rαinsppcl变体(图19a中的il-7rα变体c及d)的t细胞具有增加的存活率。最后,如图19b中展示的n端il-7受体突变体(图19a中的il-7rα变体e)在这些细胞中具有存活活性。因此,此实施例说明,当在pbmc中表达时il-7受体具有存活活性。实施例12.通过用vsv-g假型化且在其表面上表达抗cd3scfvfc的逆转录病毒颗粒转导新鲜分离及未经刺激的人类t细胞的效率。重组慢病毒颗粒是通过用编码gag/pol及rev的单独慢病毒包装质粒及编码vsv-g的假型化质粒瞬时转染293t细胞(lenti-xtm293t,clontech)来产生。与包装质粒共转染编码gfp、抗cd19嵌合抗原受体及etag的第三代慢病毒表达载体,在此称为f1-0-03(图20)。细胞通过在freestyletm293表达介质(thermofisherscientific)中连续生长来适应悬浮培养。以1×106个细胞/毫升(30ml)在125mlerlenmeyer烧瓶中接种悬浮液中的细胞,并立即使用溶解于弱酸中的聚乙烯亚胺(pei)(polysciences)转染。对于30ml细胞将质粒dna稀释于1.5mlgibcotmopti-memtm介质中。为获得用vsv-g假型化的慢病毒颗粒,所使用的总dna(1μg/ml培养体积)为具有以下摩尔比的4种质粒的混合物:2×基因组质粒(f1-0-03)、1×含rev的质粒、1×含vsv-g的质粒及1×含gag/pol的质粒。为获得用vsv-g假型化且在其表面上表达抗cd3-scfvfc的慢病毒颗粒,所使用的总dna(1μg/ml培养体积)为具有以下摩尔比的5种质粒的混合物:2×基因组质粒(f1-0-03)、1×含rev质粒、1×含vsv-g质粒、1×含抗cd3-scfvfc-gpi质粒及1×含gag/pol质粒。为获得用vsv-g假型化且在其表面上表达抗cd3-scfvfc及cd80的慢病毒颗粒,所使用的总dna(1μg/ml培养体积)为具有以下摩尔比的6种质粒的混合物:2×基因组质粒(f1-0-03)、1×含rev质粒、1×含vsv-g质粒、1×含抗cd3-scfvfc质粒、1×含cd80质粒及1×含gag/pol质粒。单独地,在1.5mlgibcotmopti-memtm中稀释pei至2μg/ml(培养体积,与dna为2:1比率)。在5分钟室温孵育之后,将两种溶液彻底地混合在一起,且再在室温下孵育20分钟。将最终体积(3ml)添加至细胞。然后伴随125rpm旋转及8%co2在37℃下孵育细胞72小时。含有抗cd3-scfvfc的质粒包括衍生自okt3或ucht1的scfv及gpi锚连接序列。ucht1scfvfc-gpi载体编码包括人类igκ信号肽(ncbigi第caa45494.1号氨基酸1至氨基酸22)的肽(seqidno:278),该人类igκ信号肽与ucht1scfv(ncbigi第cah69219.1号氨基酸21至氨基酸264)融合,与具有位置115处的a替换为t的人类igg1fc(ncbigi第aev43323.1号氨基酸1至氨基酸231)融合,与人类dafgpi锚连接序列(ncbigi第np_000565号氨基酸345至氨基酸381)融合。okt3scfvfc-gpi载体编码包括人类igκ信号肽(ncbigi第caa45494.1号氨基酸1至氨基酸22)的肽(seqidno:279),该人类igκ信号肽与okt3scfv(seqidno:285)融合,与人类igg1fc(ncbigi第aev43323.1号氨基酸1至氨基酸231)融合,与人类dafgpi锚连接序列(ncbigi第np_000565号氨基酸345至氨基酸381)融合。含cd80的质粒编码包括人类cd80信号肽及与人类cd16bgpi锚连接序列(ncbigi第np_000561号氨基酸196至氨基酸233)融合的胞外结构域(ncbigi第np_005182号氨基酸1至氨基酸242)的肽(seqidno:280)。在72小时后,采集上清液且通过以1,200g离心10分钟来澄清。将经澄清上清液倒入新试管中。在4℃下,通过以3,300g过夜离心来沉淀慢病毒颗粒。舍弃上清液,且将慢病毒颗粒沉淀再悬浮于1:100初始体积的x-vivotm15介质(lonza)中。利用系列稀释及在转导72小时后的293t及jurkat细胞中利用流式细胞术分析gfp表达来滴定慢病毒颗粒。将外周血单核细胞(pbmc)首先自供体12f及供体12m的acd(枸橼酸葡萄糖)试管中新鲜血液或自供体13f的血块黄层分离,利用sandiegobloodbank,ca收集并分配。根据制造商说明书,执行基于sepmatetm50(stemcelltm)的ficoll-paque(gehealthcarelifesciences)上的pbmc的梯度密度分离。每个sepmatetm试管中层积稀释于pbs-2%hifcs(热灭活胎牛血清)中的30ml血液或血块黄层。在室温下以1,200g离心20分钟之后,收集pbmc层,合并且用45mlpbs-2%hifcs洗涤三次,并在室温下以400g离心10分钟。然后在室温下在10mlrbc裂解缓冲液(alfaaesar)中孵育沉淀10分钟,且用45mlpbs-2%hifcs洗涤额外两次,并在室温下以400g离心10分钟。在以下转导及培养介质中执行最终洗涤:供体13f的x-vivotm15或供体12f及供体12m的rpmi-1640+10%hifcs。不采取额外步骤以移除单核细胞。在分离之后,新鲜及未经刺激pbmc被再悬浮在相应介质中达到1e6/ml的最终浓度,且用先前公开的慢病毒颗粒一式两份或一式三份转导。使传导在37℃、5%co2下在供体13f的x-vivotm15介质中或在供体12f及供体12m的rpmi-1640+10%hifcs中进行14小时。传导通常在1毫升/孔的12孔盘形式中在moi1下进行。对于动力学实验,0.5e6pbmc/ml在7ml最终溶液、在moi1下,伴随呈125rpm及具有8%co2的旋转,在37℃下在125ml振荡烧瓶中孵育2小时至20小时来转导。在与慢病毒一起孵育所选择时间后,细胞用供体13f的x-vivotm15介质或供体12f及供体12m的pbs+2%hifcs洗涤三次,且最终在37℃,5%co2下在13f的x-vivotm15介质或供体12f及供体12m的rpmi-1640+10%hifcs中在1e6/ml的细胞密度下孵育。在转导后不同天(第3天至第17天)收集样品,以利用gfp表达水平评估所产生的各类型慢病毒的转导效率。在转导后的不同天,对于具有或不具有1μg/m的okt3抗体(biolegend)下用vsv-g假型化的慢病毒颗粒,vsv-g假型化且在其表面上表达抗cd3scfvfc的慢病毒颗粒,或用vsv-g假型化且在其表面上表达抗cd3scfvfc及cd80的慢病毒颗粒,收集100μl细胞且利用流式细胞术对于cd3+细胞群体中gfp的表达进行分析。图21a及图21b分别展示在用所指示慢病毒颗粒转导自供体12m的新鲜经分离及未经刺激pbmc14小时后第3天、第6天、第9天、第13天及第17天的总cd3+群体中cd3+gfp+细胞百分比(%)直方图及cd3+gfp+群体的每孔绝对细胞计数直方图。各棒表示重复的平均值+/-sd。图21a及图21b展示,用vsv-g假型化慢病毒颗粒且在慢病毒颗粒表面上表达抗cd3-scfvfc有效地转导新鲜经分离及未经刺激的pbmc。衍生自okt3或ucht1的抗cd3scfv's在呈scfvfc形式时是有效的。图22a及图22b分别展示在用所指示慢病毒颗粒转导自供体13f的新鲜经分离及未经刺激的pbmc14小时后第3天及第6天的总cd3+群体中cd3+gfp+细胞百分比(%)直方图及cd3+gfp+群体的每孔绝对细胞计数直方图。请注意,“a”为使用经vsv-g假型化的慢病毒颗粒的结果(一式三份实验);“b”为在向转导介质添加okt3抗体(1μg/ml)的情况下使用经vsv-g假型化的慢病毒颗粒的结果(一式两份实验);“c”为使用在其表面上表达经gpi锚定的ucht1scfvfc的经vsv-g假型化的慢病毒颗粒的结果(一式三份实验);及“d”为使用在其表面上表达经gpi锚定的ucht1scfvfc及经gpi锚定的cd80的经vsv-g假型化的慢病毒颗粒的结果(一式两份实验)。各棒表示一式两份或一式三份的平均值+/-sd,如图22a所指示。图22a及图22b展示,当执行转导14小时时,用vsv-g假型化慢病毒颗粒且在其表面上表达抗cd3-scfvfc及cd80也有效地转导新鲜经分离及未经刺激的pbmc。图23a及图23b分别展示在用所指示慢病毒颗粒转导自供体12m的新鲜经分离及未经刺激pbmc持续所指示暴露时间(2小时至20小时)后第3天、第6天及第9天的总cd3+群体中cd3+gfp+细胞百分比(%)直方图及cd3+gfp+群体的每孔绝对细胞计数直方图。转导是在如所指示的盘或振荡器烧瓶中执行。各棒表示用vsv-g(“[vsv-g]”)假型化的慢病毒颗粒的两个重复的平均值+/-sd;其他实验不具有重复。图23a及图23b展示,可用用vsv-g假型化及在其表面上表达抗cd3scfvfc及cd80慢病毒颗粒在少至2小时内有效地转导新鲜经分离及未经刺激的pmbc。实施例13.插入至ef-1α启动子内含子中的mirna的功能性。设计四个单独基因片段,各自含有包括mir-1555′侧接序列或“5′臂”(seqidno:256)及mir-1553′侧接序列或“3′臂”(seqidno:260)的mir-155构架。对于各靶向cd3zetamrna转录物的唯一mirna片段用于替换mir-155茎环前体。各含有经设计以有助于将全部四个作为单链组装至ef-1α启动子内含子中的40bp重叠序列。使用用于执行超级组装的商用试剂盒(nebuilder,newenglandbiolabs,inc.)来组装含有mirna(在seqidno:255中)的内含子a及合成的ef-1α启动子为驱动慢病毒载体骨架中所含有的gfp及etag表达的转基因表达盒的部分(具有由西妥昔单抗识别的例示性etag及gfp的慢病毒载体骨架在此称为f1-0-02;图24a及24b)。在表3中指示seqidno:255中各及其相应组分的核苷酸位置及图24b中各“特征”位置。将四个mirna适当组装至慢病毒载体骨架中是通过对经修饰的ef-1α启动子及内含子区进行全面测序来确认。表3.seqidno:255中特征的核苷酸位置在其基因中含有编码针对cd3zeta的四个mirna的核酸的复制缺陷型慢病毒颗粒是通过瞬时共转染以下四个质粒至悬浮hek293细胞中来产生:含有编码经修饰以包括靶向cd3zetamrna转录物的四个mirna的f1-0-02的核酸的质粒,编码vsv-g的质粒,编码rev的质粒及编码gag-pol的质粒。在48小时后采集慢病毒颗粒上清液且经peg沉淀24小时。离心上清液,且将沉淀的慢病毒颗粒再悬浮于不具有il-2的完全pbmc生长介质中。利用jurkat细胞48小时转导来计算慢病毒颗粒效价。对于转导,在第0天融解pbmc且与100u/ml的hril-2一起孵育24小时。在第1天,通过经cd3/cd28缀合的珠粒活化pbmc。在第2天,在10moi下用含有具有编码mirna的核酸序列的基因组的慢病毒颗粒转导经活化pbmc。扩增细胞直至第11天,其中每两天添加新鲜hril-2。在第7天、第9天及第11天,采集1百万个细胞以供facs分析。针对cd3ε表面表达,使用经pe缀合的okt-3抗体(biolegend)染色细胞。表达水平是利用gfp阳性群体(经转导细胞)中pe的平均荧光强度来测定。在衍生自f1-0-02的逆转录病毒颗粒与其中将编码连续定位的cd3zmirna核酸序列插入至ef-1α启动子及内含子a中的衍生自f1-0-02的逆转录病毒颗粒之间比较经转导细胞表达水平。结果展示于图25中。此数据展示,由ef-1α启动子内含子a内的核酸序列编码的靶向cd3zeta的连续mirna在敲低cd3复合物的表达中有效。实施例14.插入至ef-1α启动子内含子中的连续抑制性rna的位置独立性克隆四个表达mirna的慢病毒载体构建体经设计以测试在以顺次排列的形式(inseries)包含4个mirna前体的结构中各个mirna前体的加工。表4展示各个构建体名称及各构建体中mir-tcrα的位置。构建体位置1位置2位置3位置4tcra-p1mir-tcramir-155mir-pd-1mir-ctla-4tcra-p2mir-155mir-tcramir-pd-1mir-ctla-4tcra-p3mir-155mir-pd-1mir-tcramir-ctla-4tcra-p4mir-155mir-ctla-4mir-pd-1mir-tcra表4.含有多顺反子mirna的构建体各mirna含有用于实施例13中的mir-155构架,即,mir-1555′臂(seqidno:256)、mir-1553′臂(seqidno:260)、环(seqidno:258)及如表5中所展示的茎序列特异次序。类型ii组装方法是用于达成将四个mirna片段组装至慢病毒载体构建体(f1-0-02;提供于实施例13中且展示于图24a中)的ef-1α内含子内的其合适位置中。其中:seqidno:267=tcrαmirna茎1;atatgtacttggctggacagcseqidno:268=tcrαmirna茎2;gctgtccacaagtacatatseqidno:269=接头1;cacattggtgccggatgaagctcttatgttgccggtcatseqidno:270=mir-155茎1;ctgttaatgctaatcgtgataseqidno:271=mir-155茎2;tatcacgattattaacagseqidno:272=接头2;gttgccggagtcttggcagcgagagatcactatcaactaaseqidno:273=pd-1mirna茎1;taccagtttagcacgaagctcseqidno:274=pd-1mirna茎2;gagcttcgctaaactggtaseqidno:275=接头3;gtgttaattgtccatgtagcgaggcatccttatggcgtggseqidno:276=ctla-4mirna茎1;tgccgctgaaatccaaggcaaseqidno:277=ctla-4mirna茎2;ttgccttgtttcagcggca表5:mirna构建体中的序列慢病毒颗粒产生四个构建体及对照f1-0-02(其不包括编码mirna的核酸序列)用于在293t细胞30ml悬浮培养物中产生慢病毒颗粒。采集慢病毒颗粒且利用peg沉淀浓缩。功能性慢病毒颗粒的效价是通过在多次稀释(1:1000、1:10000、1:100000)下转导jurkat细胞获得的,在37℃下孵育慢病毒颗粒及细胞2天,用facs缓冲液洗涤细胞2×及利用流式细胞术分析gfp。关于慢病毒颗粒产生的其他细节提供于本文实施例13中。转导对于转导,融解pbmc且在含有100u/mlhril-2的完全介质中回收过夜。经由暴露于cd3/cd28缀合的珠粒24小时来活化1e5pbmc。在moi10下用四个mirna构建体中每一种或用对照逆转录病毒颗粒f1-0-02在一式两份孔中转导细胞。每3天用100u/mlhril-2供应细胞且扩增这些细胞直至第10天。facs采集细胞以供facs分析,该facs分析确认用复制缺陷型慢病毒载体转导了细胞。结果展示,在实验中将大约等量含有mirna的病毒递送至各孔。细胞-对-ctmirnart-qpcr及分析rt-qpcr分析经设计以检测表达及将mirna前体加工为成熟加工的mir。通过首先将所有mir-tcract值归一化为rnu48内部对照以产生δct值来进行分析。然后,自未经转导的对照δct减去各经转导样品δct值以产生δδct。此值表示各经转导样品相对于未经转导对照中的经加工mir-tcramirna的量。如图26中所展示,rt-qpcr分析成功地检测到用含有mir-tcrα的复制缺陷型慢病毒颗粒转导的样品中的经加工的mir-tcrα。此外,结果清晰地指示,所测试的四个位置中任一处的mirnatcrα加工无显著差异。实施例15.表达微环境限制的生物car的t细胞的细胞毒性活性可通过改变ph来控制。以下实施例说明,表达mrb-car的经转导t细胞(也称为效应细胞)的细胞毒性活性可如何通过改变微环境ph来调节。在此实施例中,编码能够结合同源抗原靶标1(抗靶标1)的mrb-car的核酸用于产生复制缺陷型重组慢病毒颗粒。用慢病毒颗粒转导泛性t细胞,且在改变介质ph之前及之后使用实时细胞分析(rtca)来比较效应细胞的细胞毒性活性。复制缺陷型重组慢病毒颗粒的产生测试编码t1b的核酸(来自实施例3中候选mrb-car系列的抗靶标1mrb-car)。复制缺陷型重组慢病毒颗粒通过用慢病毒表达载体及核酸瞬时转染lenti-x293t细胞(clontech,mountainview,ca)来产生,这些核酸包括编码mrb-car或对照物c1(含有gmcfsp及etag(seqidno:284)但不包括抗靶标1mrb-car)的区段。细胞通过在freestyle293表达介质(thermofisherscientific,waltham,ma)中连续生长来适应悬浮培养。使用溶解于弱酸中的pei(polysciences,warminster,pa)转染悬浮液中的细胞。细胞(30ml)在125mlerlenmeyer烧瓶中生长至1×106个细胞/毫升。对于30ml细胞将总dna稀释于1.5mloptimem介质中。总dna(1μg/ml培养体积)为具有以下摩尔比的4种质粒的混合物:包括慢病毒包装元件、ltr及编码t1b的核酸的2×基因组质粒,1×编码rev的质粒,1×编码vsvg的质粒及1×编码gagpol的质粒。单独地,在1.5mloptimem中稀释pei至2μg/ml(培养体积,与dna2:1比率)。在5分钟室温孵育之后,将两种溶液充分混合在一起,且在室温下孵育20分钟。将最终体积(3ml)添加至细胞。然后伴随呈120rpm及具有5%至8%co2的旋转在37℃下孵育细胞72小时。在72小时后,通过以1,000g离心10分钟来采集上清液。将上清液倒入新试管且添加peg溶液(peg-it,systembiosciences)中1/4上清液体积。通过在4℃下孵育过夜来沉淀复制缺陷型重组慢病毒颗粒,然后在4℃下以1,500g离心20分钟。移除上清液,且将病毒再悬浮于1:100体积的x-vivo15介质中。利用jurkat细胞中etag表达滴定病毒。t细胞转导/扩增自allcells获得泛性t细胞。如上论述制造抗靶标1mrb-car复制缺陷型重组慢病毒颗粒。在慢病毒转导前两天,融解细胞且在x-vivo15介质(lonza,basel,switzerland)中与5%人类ab血清(valleybiomedicalinc.,winchester,va)及10mmn-乙酰基l-半胱氨酸(sigma-aldrich,st.louis,mo)一起培养。添加重组人类il-2(r&dsystems,minneapolis,mn)达到100iu/ml的最终浓度。在病毒转导前二十四小时,将原初人类t细胞以0.5×106个细胞/孔接种至12-孔盘中且在1:3细胞:珠粒比率使用dynabeads人类t-活化子cd3/cd28(thermofisherscientific)来活化。在转导当天,在5moi下将慢病毒颗粒溶液添加至孔。使经转导的泛性t细胞在x-vivo15介质中维持~106个/ml持续3天,然后与100iu/mlil-2转移至具有30毫升/孔x-vivo15介质的6-孔g-rex盘中。在进行实验之前培养细胞至少10天,且每隔一天添加il-2。ph变化细胞毒性分析通过添加nahco3或naoh改变ph之前及之后的经转导t细胞的细胞毒性活性使用xcelligence系统来测量。简言之,在实验前一天,用含有40mmhepes及40mmpipes(ph6.7)的x-vivo15介质将靶标细胞(用在细胞表面上表达靶标1的构建体稳定转染的cho细胞(cho-靶标1细胞))以10,000个细胞/孔接种至96-孔e-盘(acea;sandiego,ca)中。融解先前用含有编码如上论述产生的t1b或c1(分别为t1bvp及c1vp)的核酸的慢病毒颗粒转导的经冷冻保存的效应细胞且在含有100iu/mlil-2(r&dsystems,minneapolis,mn)的x-vivo15介质中培养两天。在实验当天,洗涤用t1bvp或c1vp转导的细胞且将其再悬浮于含有40mmhepes及40mmpipes(ph6.7)的x-vivo15介质中,然后以1:1的效应细胞/靶标细胞比率(e/t)添加至实验孔中。每5分钟获取在xcelligence系统(acea)上测量的阻抗读数且报告为细胞指数(ci)以定量细胞汇合度作为细胞增殖/细胞裂解的测量。添加效应细胞后大约3小时,将8μl7.5%nahco3或14μl0.5mnaoh添加至具有含有40mmhepes及40mmpipes(ph6.7)的x-vivo15介质的孔中以将ph自6.7增加至7.4。在添加效应细胞后继续阻抗读数大约20小时。如下计算特异性细胞溶解百分比:((ci靶标+经c1vp转导的效应t细胞)-(ci靶标+经t1bvp转导的效应t细胞))/(ci靶标+经c1vp转导的效应t细胞)×100。hcl的切换rtca杀伤分析通过添加hcl改变ph之前及之后的经转导t细胞的细胞毒性活性使用xcelligence系统来测量。简言之,在实验前一天,用含有40mmhepes及40mmpipes(ph7.4)的x-vivo15介质将cho-靶标1细胞以10,000个细胞/孔接种至96-孔e-盘。融解先前用c1vp或t1bvp转导的经冷冻保存的效应细胞且在含有100iu/mlil-2的x-vivo15介质中培养两天。在实验当天,洗涤用t1bvp或c1vp转导的细胞且将其再悬浮于含有40mmhepes及40mmpipes(ph7.4)的x-vivo15介质中,且然后以1:1效应细胞/靶标细胞比率(e/t)添加至实验孔中。每5分钟获取阻抗读数且报告为细胞指数(ci)。添加效应细胞后大约3小时,将8μl1mhcl添加至具有含有40mmhepes及40mmpipes(ph7.4)的x-vivo15介质的孔中以将ph自7.4切换至6.7。在添加效应细胞后继续阻抗读数大约20小时。如下计算特异性细胞溶解百分比:((ci靶标+经c1vp转导的效应t细胞)-(ci靶标+经t1bvp转导的效应t细胞))/(ci靶标c1vp)×100。结果在ph6.7及ph7.4下比较能够在降低的ph下以增加的活性结合同源抗原靶标1的mrb-car的细胞毒性活性。用编码抗靶标1mrb-car的慢病毒颗粒转导的t细胞被用于杀灭表达靶标1的cho细胞,且然后增加ph以测定细胞毒性活性是否可由ph变化抑制。如图27a及图27b中所展示,添加nahco3或naoh至活性car-t细胞的微环境以增加介质ph抑制了表达mrb-car的t细胞的细胞毒性活性。这些结果展示,表达活性mrb-car的t细胞可杀灭表达靶标的细胞,且然后此杀灭活性可通过增加微环境的ph来抑制。也测定表达待利用ph变化活化的mrb-car的t细胞的细胞毒性活性的能力。如图27c中所展示,表达抗靶标1mrb-car的t细胞的对于cho-靶标1细胞的细胞毒性活性在ph7.4时低,且通过添加hcl以降低微环境ph来增加。此外,这些结果显示,表达mrb-car的t细胞的细胞毒性活性可由微环境内ph的变化(通过在增加ph后降低细胞毒性活性及在减少ph后增加细胞毒性活性两者)来调节。在此非限制实施例中,ph自ph6.7增加且自ph7.4减少。实施例16.碳酸氢盐施用可增加小鼠中的肿瘤微环境ph。以下实施例显示,体内肿瘤微环境的ph可通过施用药剂来调节。在此实施例中,药剂为碳酸氢钠,且肿瘤微环境为小鼠中cho异种移植肿瘤。该实施例包括测量体内及离体两者的肿瘤微环境ph的两种方法。大多数实体肿瘤的胞外微环境是酸性的,ph值通常在6.5与6.9之间。相反,正常组织ph是碱性的,ph通常在7.2与7.5之间。然而,直接地测量肿瘤微环境的体内ph可能是困难的。幸运的是,组织蛋白酶的相关蛋白酶活性在较低ph下较高且在较高ph下较低。因此,肿瘤内组织蛋白酶活性的测量可充当肿瘤微环境ph的替代测量。为测量组织蛋白酶b、组织蛋白酶l、组织蛋白酶s、组织蛋白酶k、组织蛋白酶v及组织蛋白酶d的体内活性,使用近红外prosense750fast探针(perkinelmer)。为进一步确认利用碳酸氢钠施用的肿瘤微环境中ph的调节,用酚红处理所切除肿瘤并注意颜色。酚红为经历ph依赖性颜色转换的ph指示剂。酚红的钠盐广泛用于细胞培养介质中以鉴别ph值。酚红溶液在ph6.4或更低时呈黄色颜色,或在约ph7.0下呈橙色颜色,在约ph7.4下呈红色颜色及在高于ph7.8下紫色颜色。根据研究所动物看护与使用委员会(institutionalanimalcareandusecommittee)批准方案来处理小鼠。皮下(sc)中国仓鼠卵巢(cho)肿瘤异种移植是建立于12周龄至14周龄雌性b-nsg小鼠(nod-prkdcscidil2rgtm1/bcgen(beijingbiocytogenco.ltd.))的后腿中。简言之,在dpbs(thermofisher)中洗涤经培养cho细胞(atcc,manassas,va),经计数,并再悬浮于冷dpbs中且在冰上在1.5×106个细胞/200微升浓度下与合适体积的matrigelecm(corning;最终浓度5mg/ml)混合。动物在注射之前移除毛发(nair)的情况下使用标准经批准的麻醉注射。将200μl含细胞悬浮液的ecm皮下注射至小鼠后腿中。一旦肿瘤可触,使用侧径计2次/周测量肿瘤。使用以下方程式计算肿瘤体积:(最长直径*最短直径2)/2。当平均肿瘤体积达到200mm3时,将小鼠随机地分配至各别处理组。在施用碳酸氢盐前两天,将b-nsg小鼠的饮用水自酸性改变为常规ph的经高压灭菌的纯化水。在随后一天,经由100μl尾部静脉注射(4nmolprosense750fast探针/100μlpbs)将750prosensefast探针施用至6个携带有cho-异种移植肿瘤的小鼠。使单独一组携带有cho-异种移植肿瘤的小鼠未处理。在随后一天,施用碳酸氢钠且使用caliperivisluminaxr对用prosense750fast探针处理的小鼠执行成像。简言之,使用含3%o22l/min异氟烷的o2载气以2l/min麻醉小鼠,且然后置放为在成像期间经鼻锥供应1.5%异氟烷以麻醉小鼠。图像采集由5秒近红外探针(745/810nm激发/发射波长)的暴露组成。将荧光图像覆盖在小鼠正常光图像上。在施用pbs(对照)或碳酸氢钠之前获取时间0(处理前)图像。然后经由腹膜内注射(ip)向小鼠施用1毫升/小鼠pbs(对照,thermofisher)或1毫升/小鼠1m碳酸氢钠(shanghaiexperimentreagentco.,ltd)。然后在施用pbs或碳酸氢盐后30min对小鼠进行成像。调整所收集荧光图像以具有相同的最小值、最大值及临限值。在此研究中将光子计数定义为相对荧光单位(rfu)。通过将各小鼠中来自30min时间点的光子计数归一化至处理前时间点(时间0;100%)来计算rfu。由于时间0处理前值处的各小鼠中荧光值之间的不同,不同时间点处所观测的荧光强度值仅针对各个小鼠归一化而不针对处理前值的平均值归一化。在实验的单独臂中,未接受nir组织蛋白酶探针的6只小鼠在腹膜内施用pbs或碳酸氢钠后1.5小时利用颈椎脱位来安乐死。自各小鼠切除cho异种移植肿瘤。用解剖刀将异种移植肿瘤分成两半且置放于皮式培养皿(petridish)上。然后使用解剖刀重复地切割/分割肿瘤组织半部。将水或0.05%酚红溶液(50mg酚红/100ml水)分别逐滴添加至各肿瘤半部。注意颜色,且在移除肿瘤异种移植样品后,即获取经处理肿瘤异种移植及留在皮式培养皿上的酚红溶液的图像。结果图29展示在施用pbs(对照;n=3)或碳酸氢盐(n=3)之前及之后携带cho-异种移植肿瘤小鼠中的肿瘤内组织蛋白酶活性成像的rfu结果(具有sem平均值)。这些结果表明,碳酸氢钠施用可增加体内肿瘤微环境ph,这由在腹膜内碳酸氢钠施用后观测到的降低的组织蛋白酶活性所证明。相对于经pbs处理的小鼠(n=3),使用自经碳酸氢钠处理的小鼠(n=3)切除的肿瘤组织观测到酚红指示剂自黄色/橙色变为红色的颜色改变。这些结果表明,在腹膜内施用后,碳酸氢钠施用增加体内肿瘤微环境ph,这由酚红指示剂自黄色/橙色变为红色的颜色改变所证明。实施例17.利用差示扫描量热法(dsc)在阿昔洛韦不存在及存在下的f1a-795的热变性在此实施例中,通过在核苷类似物抗病毒药物阿昔洛韦不存在相比于存在下比较适配体结构域的热变性来证实核苷类似物抗病毒药物与核糖开关(seqidno:87)适配体结构域的结合。适配体制备利用idt(coralville,ia)将t7引物(seqidno:246)以及所鉴别候选序列的模板(反向互补物)合成为单链dna。通过组合组分至1μm模板、2μmt7引物、200μmdntp、1×titaniumtaqdna聚合酶及1×titaniumtaq缓冲液(clontechlaboratories;mountainview;ca),然后在95℃下加热3分钟,在55℃下加热1分钟及在68℃下加热2分钟(无循环)来使候选物引物延伸。42皮摩尔的双链dna用于根据标准试剂盒指示(1×反应缓冲液,7.5mm各ntp,10mmdtt,1×riboguardrnase抑制剂,1×ampliscribet7酶溶液)用ampliscribet7高产率转录试剂盒(lucigen;middleton,wi)进行十二个20-μl反应。在42℃下孵育反应物过夜,然后通过添加1μldna酶i终止且在37℃下孵育15分钟。在具有8m脲素的10%变性page上纯化转录产物。冻干至少10纳摩尔的各候选物,且在结合评估当天将其再悬浮于1×结合缓冲液(50mmhepes,100mmkcl,0.5mmmgcl2,ph7.3)中。简言之,dsc用于分析在阿昔洛韦不存在下的f1a-795(7.2μm)或在阿昔洛韦(29.4μm)存在下的f1a-795(2.77μm)。所有分析均是在1×结合缓冲液中进行且所使用的所有水均经depc处理。将所有分析物首先再悬浮于经depc处理的水中,然后在1×结合缓冲液中稀释至其最终所列浓度。在装载dsc(gehealthcaremicrocalvp-dsc)之前,在25℃下对样品进行脱气10分钟。以每分钟1℃的速率自10℃至115℃扫描不具有阿昔洛韦的样品。以每分钟1℃的速率自10℃至105℃扫描具有阿昔洛韦的样品。结果由dsc测量的在阿昔洛韦不存在下的f1a-795热变性具有中心在约60℃下的转换(图28)。此转换表明,适配体结构域在阿昔洛韦不存在下结构化。在阿昔洛韦不存在下,f1a-795具有中心在约75℃下的转换(图28)。此稳定化效果指示,核苷类似物抗病毒药物阿昔洛韦与适配体结构域f1a-795结合。因此,此实验确认,f1a-795与阿昔洛韦结合。所公开的实施例、实例及实验不意欲限制本发明范畴或表示以下实验为执行的全部或唯一的实验。已努力确保关于所使用数字(例如,量、温度等)的准确度,但应计入一些实验性误差及偏差。应理解,可在不改变实验意欲说明基本方面的情况下对如所描述方法进行变化。本领域技术人员可在本发明范畴及精神下设计许多修改及其他实施方式。实际上,可在不改变本发明基本方面情况下由本领域技术人员对所描述的材料、方法、图式、实验、实例及实施例进行变化。任何所公开实施方式可结合其他所公开实施方式来使用。在一些情况下,参考特定实施方式描述一些概念。然而,普通技术人员了解,可在不背离如以下权利要求所阐述的技术实现要素:范畴的情况下进行多种修改及改变。因此,说明书及图式应视为呈说明性意义而非限制性意义,且所有这些修改意欲包括在本发明范畴内。序列表<110>f1肿瘤医学公司<120>用于转导淋巴细胞及调节其活性的方法及组合物<130>f1.001.wo.02<150>62/467,039<151>2017-03-03<150>62/390,093<151>2016-03-19<150>62/360,041<151>2016-07-08<150>pct/us2017/023112<151>2017-03-19<150>15/462,855<151>2017-03-19<160>291<170>patentinversion3.5<210>1<211>42<212>prt<213>homosapiens<400>1lysargglyarglyslysleuleutyrilephelysglnprophemet151015argprovalglnthrthrglnglugluaspglycyssercysargphe202530proglugluglugluglyglycysgluleu3540<210>2<211>41<212>prt<213>homosapiens<400>2argserlysargserargleuleuhisserasptyrmetasnmetthr151015proargargproglyprothrarglyshistyrglnprotyralapro202530proargaspphealaalatyrargser3540<210>3<211>41<212>prt<213>homosapiens<400>3argserlysargserargleuleuhisserasptyrmetasnmetthr151015proargargproglyprothrarglyshistyrglnalatyralaala202530alaargaspphealaalatyrargser3540<210>4<211>35<212>prt<213>homosapiens<400>4thrlyslyslystyrserserservalhisaspproasnglyglutyr151015metphemetargalavalasnthralalyslysserargleuthrasp202530valthrleu35<210>5<211>37<212>prt<213>homosapiens<400>5argargaspglnargleuproproaspalahislysproproglygly151015glyserpheargthrproileglnglugluglnalaaspalahisser202530thrleualalysile35<210>6<211>49<212>prt<213>homosapiens<400>6hisglnargarglystyrargserasnlysglygluserprovalglu151015proalagluprocysargtyrsercysproarggluglugluglyser202530thrileproileglngluasptyrarglysprogluproalacysser354045pro<210>7<211>114<212>prt<213>homosapiens<400>7cyscysleuargarghisglnglylysglnasngluleuseraspthr151015alaglyarggluileasnleuvalaspalahisleulysserglugln202530thrglualaserthrargglnasnserglnvalleuleusergluthr354045glyiletyraspasnaspproaspleucyspheargmetglnglugly505560sergluvaltyrserasnprocysleuglugluasnlysproglyile65707580valtyralaserleuasnhisservalileglyproasnserargleu859095alaargasnvallysglualaprothrglutyralaserilecysval100105110argser<210>8<211>187<212>prt<213>homosapiens<400>8argargalacysarglysargileargglnlysleuhisleucystyr151015provalglnthrserglnprolysleugluleuvalaspserargpro202530argargserserthrglnleuargserglyalaservalthrglupro354045valalaglugluargglyleumetserglnproleumetgluthrcys505560hisservalglyalaalatyrleugluserleuproleuglnaspala65707580serproalaglyglyproserserproargaspleuprogluproarg859095valserthrgluhisthrasnasnlysileglulysiletyrilemet100105110lysalaaspthrvalilevalglythrvallysalagluleuproglu115120125glyargglyleualaglyproalagluprogluleugluglugluleu130135140glualaasphisthrprohistyrprogluglngluthrglupropro145150155160leuglysercysseraspvalmetleuservalgluglugluglylys165170175gluaspproleuprothralaalaserglylys180185<210>9<211>54<212>prt<213>homosapiens<400>9hisiletrpglnleuargserglncysmettrpproarggluthrgln151015leuleuleugluvalproproserthrgluaspalaargsercysgln202530pheprogluglugluargglygluargseralagluglulysglyarg354045leuglyaspleutrpval50<210>10<211>60<212>prt<213>homosapiens<400>10cysvallysargarglysproargglyaspvalvallysvalileval151015servalglnarglysargglnglualagluglyglualathrvalile202530glualaleuglnalaproproaspvalthrthrvalalavalgluglu354045thrileproserphethrglyargserproasnhis505560<210>11<211>163<212>prt<213>homosapiens<400>11metlystrplysalaleuphethralaalaileleuglnalaglnleu151015proilethrglualaglnserpheglyleuleuaspprolysleucys202530tyrleuleuaspglyileleupheiletyrglyvalileleuthrala354045leupheleuargvallyspheserargseralaaspalaproalatyr505560glnglnglyglnasnglnleutyrasngluleuasnleuglyargarg65707580gluglutyraspvalleuasplysargargglyargaspproglumet859095glyglylysproargarglysasnproglngluglyleutyrasnglu100105110leuglnlysasplysmetalaglualatyrsergluileglymetlys115120125glygluargargargglylysglyhisaspglyleutyrglnglyleu130135140serthralathrlysaspthrtyraspalaleuhismetglnalaleu145150155160proproarg<210>12<211>164<212>prt<213>homosapiens<400>12metlystrplysalaleuphethralaalaileleuglnalaglnleu151015proilethrglualaglnserpheglyleuleuaspprolysleucys202530tyrleuleuaspglyileleupheiletyrglyvalileleuthrala354045leupheleuargvallyspheserargseralaaspalaproalatyr505560glnglnglyglnasnglnleutyrasngluleuasnleuglyargarg65707580gluglutyraspvalleuasplysargargglyargaspproglumet859095glyglylysproglnargarglysasnproglngluglyleutyrasn100105110gluleuglnlysasplysmetalaglualatyrsergluileglymet115120125lysglygluargargargglylysglyhisaspglyleutyrglngly130135140leuserthralathrlysaspthrtyraspalaleuhismetglnala145150155160leuproproarg<210>13<211>112<212>prt<213>homosapiens<400>13argvallyspheserargseralaaspalaproalatyrglnglngly151015glnasnglnleutyrasngluleuasnleuglyargarggluglutyr202530aspvalleuasplysargargglyargaspproglumetglyglylys354045proargarglysasnproglngluglyleutyrasngluleuglnlys505560asplysmetalaglualatyrsergluileglymetlysglygluarg65707580argargglylysglyhisaspglyleutyrglnglyleuserthrala859095thrlysaspthrtyraspalaleuhismetglnalaleuproproarg100105110<210>14<211>21<212>prt<213>homosapiens<400>14asnglnleutyrasngluleuasnleuglyargarggluglutyrasp151015valleuasplysarg20<210>15<211>22<212>prt<213>homosapiens<400>15gluglyleutyrasngluleuglnlysasplysmetalaglualatyr151015sergluileglymetlys20<210>16<211>21<212>prt<213>homosapiens<400>16aspglyleutyrglnglyleuserthralathrlysaspthrtyrasp151015alaleuhismetgln20<210>17<211>171<212>prt<213>homosapiens<400>17metgluhisserthrpheleuserglyleuvalleualathrleuleu151015serglnvalserprophelysileproileglugluleugluasparg202530valphevalasncysasnthrserilethrtrpvalgluglythrval354045glythrleuleuseraspilethrargleuaspleuglylysargile505560leuaspproargglyiletyrargcysasnglythraspiletyrlys65707580asplysgluserthrvalglnvalhistyrargmetcysglnsercys859095valgluleuaspproalathrvalalaglyileilevalthraspval100105110ilealathrleuleuleualaleuglyvalphecysphealaglyhis115120125gluthrglyargleuserglyalaalaaspthrglnalaleuleuarg130135140asnaspglnvaltyrglnproleuargaspargaspaspalaglntyr145150155160serhisleuglyglyasntrpalaargasnlys165170<210>18<211>127<212>prt<213>homosapiens<400>18metgluhisserthrpheleuserglyleuvalleualathrleuleu151015serglnvalserprophelysileproileglugluleugluasparg202530valphevalasncysasnthrserilethrtrpvalgluglythrval354045glythrleuleuseraspilethrargleuaspleuglylysargile505560leuaspproargglyiletyrargcysasnglythraspiletyrlys65707580asplysgluserthrvalglnvalhistyrargthralaaspthrgln859095alaleuleuargasnaspglnvaltyrglnproleuargaspargasp100105110aspalaglntyrserhisleuglyglyasntrpalaargasnlys115120125<210>19<211>21<212>prt<213>homosapiens<400>19aspglnvaltyrglnproleuargaspargaspaspalaglntyrser151015hisleuglyglyasn20<210>20<211>206<212>prt<213>homosapiens<400>20metglnserglythrhistrpargvalleuglyleucysleuleuser151015valglyvaltrpglyglnaspglyasngluglumetglyglyilethr202530glnthrprotyrlysvalserileserglythrthrvalileleuthr354045cysproglntyrproglysergluileleutrpglnhisasnasplys505560asnileglyglyaspgluaspasplysasnileglyseraspgluasp65707580hisleuserleulysgluphesergluleugluglnserglytyrtyr859095valcystyrproargglyserlysprogluaspalaasnphetyrleu100105110tyrleuargalaargvalcysgluasncysmetglumetaspmetser115120125valalathrilevalilevalaspilecysilethrglyglyleuleu130135140leuleuvaltyrtyrtrpserlysasnarglysalalysalalyspro145150155160valthrargglyalaglyalaglyglyargglnargglyglnasnlys165170175gluargproproprovalproasnproasptyrgluproilearglys180185190glyglnargaspleutyrserglyleuasnglnargargile195200205<210>21<211>21<212>prt<213>homosapiens<400>21asnproasptyrgluproilearglysglyglnargaspleutyrser151015glyleuasnglnarg20<210>22<211>182<212>prt<213>homosapiens<400>22metgluglnglylysglyleualavalleuileleualaileileleu151015leuglnglythrleualaglnserilelysglyasnhisleuvallys202530valtyrasptyrglngluaspglyservalleuleuthrcysaspala354045glualalysasnilethrtrpphelysaspglylysmetileglyphe505560leuthrgluasplyslyslystrpasnleuglyserasnalalysasp65707580proargglymettyrglncyslysglyserglnasnlysserlyspro859095leuglnvaltyrtyrargmetcysglnasncysilegluleuasnala100105110alathrileserglypheleuphealagluilevalserilepheval115120125leualavalglyvaltyrpheilealaglyglnaspglyvalarggln130135140serargalaserasplysglnthrleuleuproasnaspglnleutyr145150155160glnproleulysasparggluaspaspglntyrserhisleuglngly165170175asnglnleuargargasn180<210>23<211>21<212>prt<213>homosapiens<400>23aspglnleutyrglnproleulysasparggluaspaspglntyrser151015hisleuglnglyasn20<210>24<211>226<212>prt<213>homosapiens<400>24metproglyglyproglyvalleuglnalaleuproalathrilephe151015leuleupheleuleuseralavaltyrleuglyproglycysglnala202530leutrpmethislysvalproalaserleumetvalserleuglyglu354045aspalahispheglncysprohisasnserserasnasnalaasnval505560thrtrptrpargvalleuhisglyasntyrthrtrpproprogluphe65707580leuglyproglygluaspproasnglythrleuileileglnasnval859095asnlysserhisglyglyiletyrvalcysargvalglngluglyasn100105110glusertyrglnglnsercysglythrtyrleuargvalargglnpro115120125proproargpropheleuaspmetglygluglythrlysasnargile130135140ilethralagluglyileileleuleuphecysalavalvalprogly145150155160thrleuleuleuphearglysargtrpglnasnglulysleuglyleu165170175aspalaglyaspglutyrgluaspgluasnleutyrgluglyleuasn180185190leuaspaspcyssermettyrgluaspileserargglyleuglngly195200205thrtyrglnaspvalglyserleuasnileglyaspvalglnleuglu210215220lyspro225<210>25<211>188<212>prt<213>homosapiens<400>25metproglyglyproglyvalleuglnalaleuproalathrilephe151015leuleupheleuleuseralavaltyrleuglyproglycysglnala202530leutrpmethislysvalproalaserleumetvalserleuglyglu354045aspalahispheglncysprohisasnserserasnasnalaasnval505560thrtrptrpargvalleuhisglyasntyrthrtrpproprogluphe65707580leuglyproglygluaspproasngluproproproargpropheleu859095aspmetglygluglythrlysasnargileilethralagluglyile100105110ileleuleuphecysalavalvalproglythrleuleuleuphearg115120125lysargtrpglnasnglulysleuglyleuaspalaglyaspglutyr130135140gluaspgluasnleutyrgluglyleuasnleuaspaspcyssermet145150155160tyrgluaspileserargglyleuglnglythrtyrglnaspvalgly165170175serleuasnileglyaspvalglnleuglulyspro180185<210>26<211>21<212>prt<213>homosapiens<400>26gluasnleutyrgluglyleuasnleuaspaspcyssermettyrglu151015aspileserarggly20<210>27<211>113<212>prt<213>homosapiens<400>27metglyglyleugluprocysserargleuleuleuleuproleuleu151015leualavalserglyleuargprovalglnalaglnalaglnserasp202530cyssercysserthrvalserproglyvalleualaglyilevalmet354045glyaspleuvalleuthrvalleuilealaleualavaltyrpheleu505560glyargleuvalproargglyargglyalaalaglualaalathrarg65707580lysglnargilethrgluthrgluserprotyrglngluleuglngly859095glnargseraspvaltyrseraspleuasnthrglnargprotyrtyr100105110lys<210>28<211>107<212>prt<213>homosapiens<400>28metglyglyleugluprocysserargleuleuleuleuproleuleu151015leualavalserglyleuargprovalglnalaglnalaglnserasp202530cyssercysserthrvalserproglyvalleualaglyilevalmet354045glyaspleuvalleuthrvalleuilealaleualavaltyrpheleu505560glyargleuvalproargglyargglyalaalaglualathrarglys65707580glnargilethrgluthrgluserprotyrglngluleuglnglygln859095argseraspvaltyrseraspleuasnthrgln100105<210>29<211>102<212>prt<213>homosapiens<400>29metglyglyleugluprocysserargleuleuleuleuproleuleu151015leualavalseraspcyssercysserthrvalserproglyvalleu202530alaglyilevalmetglyaspleuvalleuthrvalleuilealaleu354045alavaltyrpheleuglyargleuvalproargglyargglyalaala505560glualaalathrarglysglnargilethrgluthrgluserprotyr65707580glngluleuglnglyglnargseraspvaltyrseraspleuasnthr859095glnargprotyrtyrlys100<210>30<211>101<212>prt<213>homosapiens<400>30metglyglyleugluprocysserargleuleuleuleuproleuleu151015leualavalseraspcyssercysserthrvalserproglyvalleu202530alaglyilevalmetglyaspleuvalleuthrvalleuilealaleu354045alavaltyrpheleuglyargleuvalproargglyargglyalaala505560glualathrarglysglnargilethrgluthrgluserprotyrgln65707580gluleuglnglyglnargseraspvaltyrseraspleuasnthrgln859095argprotyrtyrlys100<210>31<211>21<212>prt<213>homosapiens<400>31gluserprotyrglngluleuglnglyglnargseraspvaltyrser151015aspleuasnthrgln20<210>32<211>86<212>prt<213>homosapiens<400>32metileproalavalvalleuleuleuleuleuleuvalgluglnala151015alaalaleuglygluproglnleucystyrileleuaspalaileleu202530pheleutyrglyilevalleuthrleuleutyrcysargleulysile354045glnvalarglysalaalailethrsertyrglulysseraspglyval505560tyrthrglyleuserthrargasnglngluthrtyrgluthrleulys65707580hisglulysproprogln85<210>33<211>21<212>prt<213>homosapiens<400>33aspglyvaltyrthrglyleuserthrargasnglngluthrtyrglu151015thrleulyshisglu20<210>34<211>20<212>prt<213>homosapiens<400>34argproargargserproalaglnaspglylysvaltyrileasnmet151015proglyarggly20<210>35<211>68<212>prt<213>homosapiens<400>35phetrpvalleuvalvalvalglyglyvalleualacystyrserleu151015leuvalthrvalalapheileilephetrpvalargserlysargser202530argleuleuhisserasptyrmetasnmetthrproargargprogly354045prothrarglyshistyrglnprotyralaproproargasppheala505560alatyrargser65<210>36<211>619<212>prt<213>homosapiens<400>36metproaspproalaalahisleuprophephetyrglyserileser151015argalaglualaglugluhisleulysleualaglymetalaaspgly202530leupheleuleuargglncysleuargserleuglyglytyrvalleu354045serleuvalhisaspvalargphehishispheproilegluarggln505560leuasnglythrtyralailealaglyglylysalahiscysglypro65707580alagluleucysgluphetyrserargaspproaspglyleuprocys859095asnleuarglysprocysasnargproserglyleugluproglnpro100105110glyvalpheaspcysleuargaspalametvalargasptyrvalarg115120125glnthrtrplysleugluglyglualaleugluglnalaileileser130135140glnalaproglnvalglulysleuilealathrthralahisgluarg145150155160metprotrptyrhisserserleuthrarggluglualagluarglys165170175leutyrserglyalaglnthraspglylyspheleuleuargproarg180185190lysgluglnglythrtyralaleuserleuiletyrglylysthrval195200205tyrhistyrleuileserglnasplysalaglylystyrcysilepro210215220gluglythrlyspheaspthrleutrpglnleuvalglutyrleulys225230235240leulysalaaspglyleuiletyrcysleulysglualacysproasn245250255serseralaserasnalaserglyalaalaalaprothrleuproala260265270hisproserthrleuthrhisproglnargargileaspthrleuasn275280285seraspglytyrthrprogluproalaargilethrserproasplys290295300proargprometprometaspthrservaltyrgluserprotyrser305310315320aspproglugluleulysasplyslysleupheleulysargaspasn325330335leuleuilealaaspilegluleuglycysglyasnpheglyserval340345350argglnglyvaltyrargmetarglyslysglnileaspvalalaile355360365lysvalleulysglnglythrglulysalaaspthrgluglumetmet370375380argglualaglnilemethisglnleuaspasnprotyrilevalarg385390395400leuileglyvalcysglnalaglualaleumetleuvalmetglumet405410415alaglyglyglyproleuhislyspheleuvalglylysarggluglu420425430ileprovalserasnvalalagluleuleuhisglnvalsermetgly435440445metlystyrleugluglulysasnphevalhisargaspleualaala450455460argasnvalleuleuvalasnarghistyralalysileseraspphe465470475480glyleuserlysalaleuglyalaaspaspsertyrtyrthralaarg485490495seralaglylystrpproleulystrptyralaproglucysileasn500505510phearglyspheserserargseraspvaltrpsertyrglyvalthr515520525mettrpglualaleusertyrglyglnlysprotyrlyslysmetlys530535540glyprogluvalmetalapheilegluglnglylysargmetglucys545550555560proproglucysproprogluleutyralaleumetseraspcystrp565570575iletyrlystrpgluaspargproasppheleuthrvalgluglnarg580585590metargalacystyrtyrserleualaserlysvalgluglypropro595600605glyserthrglnlysalaglualaalacysala610615<210>37<211>9<212>prt<213>artificialsequence<220><223>synthetichaepitope<400>37tyrprotyraspvalproasptyrala15<210>38<211>8<212>prt<213>artificialsequence<220><223>syntheticflagepitope<400>38asptyrlysaspaspaspasplys15<210>39<211>10<212>prt<213>artificialsequence<220><223>syntheticc-mycepitope<400>39gluglnlysleuileserglugluaspleu1510<210>40<211>5<212>prt<213>artificialsequence<220><223>synthetichis5affinity<400>40hishishishishis15<210>41<211>6<212>prt<213>artificialsequence<220><223>synthetichisx6affinity<400>41hishishishishishis15<210>42<211>8<212>prt<213>artificialsequence<220><223>syntheticstreptagaffinity<400>42trpserhisproglnpheglulys15<210>43<211>5<212>prt<213>artificialsequence<220><223>synthetichisx6affinity<400>43argtyrileargser15<210>44<211>4<212>prt<213>artificialsequence<220><223>syntheticaffinity<400>44phehishisthr1<210>45<211>17<212>prt<213>artificialsequence<220><223>syntheticaffinity<400>45trpglualaalaalaargglualacyscysargglucyscysalaarg151015ala<210>46<211>24<212>prt<213>homosapiens<400>46iletyriletrpalaproleualaglythrcysglyvalleuleuleu151015serleuvalilethrleutyrcys20<210>47<211>23<212>prt<213>homosapiens<400>47leuglyleuleuvalalaglyvalleuvalleuleuvalserleugly151015valalailehisleucyscys20<210>48<211>25<212>prt<213>homosapiens<400>48alaleuilevalleuglyglyvalalaglyleuleuleupheilegly151015leuglyilephephecysvalargcys2025<210>49<211>23<212>prt<213>homosapiens<400>49leucystyrleuleuaspglyileleupheiletyrglyvalileleu151015thralaleupheleuargval20<210>50<211>27<212>prt<213>homosapiens<400>50phetrpvalleuvalvalvalglyglyvalleualacystyrserleu151015leuvalthrvalalapheileilephetrpval2025<210>51<211>26<212>prt<213>homosapiens<400>51valalaalaileleuglyleuglyleuvalleuglyleuleuglypro151015leualaileleuleualaleutyrleuleu2025<210>52<211>24<212>prt<213>homosapiens<400>52alaleuproalaalaleualavalileserpheleuleuglyleugly151015leuglyvalalacysvalleuala20<210>53<211>15<212>prt<213>artificialsequence<220><223>syntheticlinker1<400>53glyglyglyglyserglyglyglyglyserglyglyglyglyser151015<210>54<211>30<212>prt<213>artificialsequence<220><223>syntheticlinker2<400>54glyglyglyglyserglyglyglyglyserglyglyglyglysergly151015glyglyglyserglyglyglyglyserglyglyglyglyser202530<210>55<211>14<212>prt<213>artificialsequence<220><223>syntheticlinker3<400>55glyglyglyglyserglyglyglyserglyglyglyglyser1510<210>56<211>4<212>prt<213>artificialsequence<220><223>syntheticlinker4<400>56glyglysergly1<210>57<211>5<212>prt<213>artificialsequence<220><223>syntheticlinker5<400>57glyglyserglygly15<210>58<211>5<212>prt<213>artificialsequence<220><223>syntheticlinker6<400>58glyserglysergly15<210>59<211>5<212>prt<213>artificialsequence<220><223>syntheticlinker7<400>59glyserglyglygly15<210>60<211>5<212>prt<213>artificialsequence<220><223>syntheticlinker8<400>60glyglyglysergly15<210>61<211>5<212>prt<213>artificialsequence<220><223>syntheticlinker9<400>61glysersersergly15<210>62<211>4<212>prt<213>artificialsequence<220><223>synthetichinge1<400>62cysproprocys1<210>63<211>5<212>prt<213>artificialsequence<220><223>synthetichinge2<400>63asplysthrhisthr15<210>64<211>15<212>prt<213>artificialsequence<220><223>synthetichinge3<400>64cysprogluprolyssercysaspthrproproprocysproarg151015<210>65<211>12<212>prt<213>artificialsequence<220><223>synthetichinge4<400>65gluleulysthrproleuglyaspthrthrhisthr1510<210>66<211>10<212>prt<213>artificialsequence<220><223>synthetichinge5<400>66lyssercysasplysthrhisthrcyspro1510<210>67<211>7<212>prt<213>artificialsequence<220><223>synthetichinge6<400>67lyscyscysvalaspcyspro15<210>68<211>7<212>prt<213>artificialsequence<220><223>synthetichinge7<400>68lystyrglyproprocyspro15<210>69<211>15<212>prt<213>artificialsequence<220><223>synthetichinge8<400>69gluprolyssercysasplysthrhisthrcysproprocyspro151015<210>70<211>12<212>prt<213>artificialsequence<220><223>synthetichinge9<400>70gluarglyscyscysvalglucysproprocyspro1510<210>71<211>17<212>prt<213>artificialsequence<220><223>synthetichinge10<400>71gluleulysthrproleuglyaspthrthrhisthrcysproargcys151015pro<210>72<211>12<212>prt<213>artificialsequence<220><223>synthetichinge11<400>72serproasnmetvalprohisalahishisalagln1510<210>73<211>45<212>prt<213>artificialsequence<220><223>synthetichinge12<400>73thrthrthrproalaproargproprothrproalaprothrileala151015serglnproleuserleuargproglualacysargproalaalagly202530glyalavalhisthrargglyleuaspphealacysasp354045<210>74<211>21<212>prt<213>homosapiens<400>74metalaleuprovalthralaleuleuleuproleualaleuleuleu151015hisalaalaargpro20<210>75<211>24<212>prt<213>homosapiens<400>75iletyriletrpalaproleualaglythrcysglyvalleuleuleu151015serleuvalilethrleutyrcys20<210>76<211>27<212>prt<213>homosapiens<400>76phetrpvalleuvalvalvalglyglyvalleualacystyrserleu151015leuvalthrvalalapheileilephetrpval2025<210>77<211>21<212>prt<213>artificialsequence<220><223>synthetic2a-1cleavagesignal<400>77glyserglygluglyargglyserleuleuthrcysglyaspvalglu151015gluasnproglypro20<210>78<211>357<212>prt<213>homosapiens<400>78metleuleuleuvalthrserleuleuleucysgluleuprohispro151015alapheleuleuileproarglysvalcysasnglyileglyilegly202530gluphelysaspserleuserileasnalathrasnilelyshisphe354045lysasncysthrserileserglyaspleuhisileleuprovalala505560pheargglyaspserphethrhisthrproproleuaspproglnglu65707580leuaspileleulysthrvallysgluilethrglypheleuleuile859095glnalatrpprogluasnargthraspleuhisalaphegluasnleu100105110gluileileargglyargthrlysglnhisglyglnpheserleuala115120125valvalserleuasnilethrserleuglyleuargserleulysglu130135140ileseraspglyaspvalileileserglyasnlysasnleucystyr145150155160alaasnthrileasntrplyslysleupheglythrserglyglnlys165170175thrlysileileserasnargglygluasnsercyslysalathrgly180185190glnvalcyshisalaleucysserprogluglycystrpglyproglu195200205proargaspcysvalsercysargasnvalserargglyargglucys210215220valasplyscysasnleuleugluglygluproarggluphevalglu225230235240asnserglucysileglncyshisproglucysleuproglnalamet245250255asnilethrcysthrglyargglyproaspasncysileglncysala260265270histyrileaspglyprohiscysvallysthrcysproalaglyval275280285metglygluasnasnthrleuvaltrplystyralaaspalaglyhis290295300valcyshisleucyshisproasncysthrtyrglycysthrglypro305310315320glyleugluglycysprothrasnglyprolysileproserileala325330335thrglymetvalglyalaleuleuleuleuleuvalvalalaleugly340345350ileglyleuphemet355<210>79<211>45<212>prt<213>homosapiens<400>79thrthrthrproalaproargproprothrproalaprothrileala151015serglnproleuserleuargproglualacysargproalaalagly202530glyalavalhisthrargglyleuaspphealacysasp354045<210>80<211>39<212>prt<213>homosapiens<400>80ilegluvalmettyrproproprotyrleuaspasnglulysserasn151015glythrileilehisvallysglylyshisleucysproserproleu202530pheproglyproserlyspro35<210>81<211>113<212>prt<213>homosapiens<400>81argvallyspheserargseralaaspalaproalatyrglnglngly151015glnasnglnleutyrasngluleuasnleuglyargarggluglutyr202530aspvalleuasplysargargglyargaspproglumetglyglylys354045proglnargarglysasnproglngluglyleutyrasngluleugln505560lysasplysmetalaglualatyrsergluileglymetlysglyglu65707580argargargglylysglyhisaspglyleutyrglnglyleuserthr859095alathrlysaspthrtyraspalaleuhismetglnalaleupropro100105110arg<210>82<211>463<212>prt<213>homosapiens<400>82metthrileleuglythrthrpheglymetvalpheserleuleugln151015valvalserglygluserglytyralaglnasnglyaspleugluasp202530alagluleuaspasptyrserphesercystyrserglnleugluval354045asnglyserglnhisserleuthrcysalaphegluaspproaspval505560asnthrthrasnleugluphegluilecysglyalaleuvalgluval65707580lyscysleuasnphearglysleuglngluiletyrpheilegluthr859095lyslyspheleuleuileglylysserasnilecysvallysvalgly100105110glulysserleuthrcyslyslysileaspleuthrthrilevallys115120125proglualapropheaspleuservaliletyrarggluglyalaasn130135140aspphevalvalthrpheasnthrserhisleuglnlyslystyrval145150155160lysvalleumethisaspvalalatyrargglnglulysaspgluasn165170175lystrpthrhisvalasnleuserserthrlysleuthrleuleugln180185190arglysleuglnproalaalamettyrgluilelysvalargserile195200205proasphistyrphelysglyphetrpserglutrpserprosertyr210215220tyrpheargthrprogluileasnasnserserglyglumetasppro225230235240ileleuleuproprocysleuthrileserileleuserphepheser245250255valalaleuleuvalileleualacysvalleutrplyslysargile260265270lysproilevaltrpproserleuproasphislyslysthrleuglu275280285hisleucyslyslysproarglysasnleuasnvalserpheasnpro290295300gluserpheleuaspcysglnilehisargvalaspaspileglnala305310315320argaspgluvalgluglypheleuglnaspthrpheproglnglnleu325330335glugluserglulysglnargleuglyglyaspvalglnserproasn340345350cysprosergluaspvalvalvalthrprogluserpheglyargasp355360365serserleuthrcysleualaglyasnvalseralacysaspalapro370375380ileleuserserserargserleuaspcysarggluserglylysasn385390395400glyprohisvaltyrglnaspleuleuleuserleuglythrthrasn405410415serthrleupropropropheserleuglnserglyileleuthrleu420425430asnprovalalaglnglyglnproileleuthrserleuglyserasn435440445glngluglualatyrvalthrmetserserphetyrglnasngln450455460<210>83<211>13673<212>dna<213>artificialsequence<220><223>syntheticdox-rapamycininduciblelentiviralgenomewithriboswitch<400>83gatatctataacaagaaaatatatatataataagttatcacgtaagtagaacatgaaata60acaatataattatcgtatgagttaaatcttaaaagtcacgtaaaagataatcatgcgtca120ttttgactcacgcggtcgttatagttcaaaatcagtgacacttaccgcattgacaagcac180gcctcacgggagctccaagcggcgactgagatgtcctaaatgcacagcgacggattcgcg240ctatttagaaagagagagcaatatttcaagaatgcatgcgtcaattttacgcagactatc300tttctagggttaagacggatcgggagatctcccgatcccctatggtgcactctcagtaca360atctgctctgatgccgcatagttaagccagtatctgctccctgcttgtgtgttggaggtc420gctgagtagtgcgcgagcaaaatttaagctacaacaaggcaaggcttgaccgacaattgc480atgaagaatctgcttagggttaggcgttttgcgctgcttcgcgatgtacgggccagatat540acgcgttgacattgattattgactagttattaatagtaatcaattacggggtcattagtt600catagcccatatatggagttccgcgttacataacttacggtaaatggcccgcctggctga660ccgcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacgcca720atagggactttccattgacgtcaatgggtggagtatttacggtaaactgcccacttggca780gtacatcaagtgtatcatatgccaagtacgccccctattgacgtcaatgacggtaaatgg840cccgcctggcattatgcccagtacatgaccttatgggactttcctacttggcagtacatc900tacgtattagtcatcgctattaccatggtgatgcggttttggcagtacatcaatgggcgt960ggatagcggtttgactcacggggatttccaagtctccaccccattgacgtcaatgggagt1020ttgttttggaaccaaaatcaacgggactttccaaaatgtcgtaacaactccgccccattg1080acgcaaatgggcggtaggcgtgtacggtgggaggtctatataagcagagctctccctatc1140agtgatagagatctccctatcagtgatagagatcgtcgacgagctcgtttagtgaaccgt1200cagatcgcctggagacgccctcgaagccgcggtgcgggtgccagggcgtgccctgggtct1260ctctggttagaccagatctgagcctgggagctctctggctaactagggaacccactgctt1320aagcctcaataaagcttgccttgagtgcttcaagtagtgtgtgcccgtctgttgtgtgac1380tctggtaactagagatccctcagacccttttagtcagtgtggaaaatctctagcagtggc1440gcccgaacagggacttgaaagcgaaagggaaaccagaggagctctctcgacgcaggactc1500ggcttgctgaagcgcgcacggcaagaggcgaggggcggcgactggtgagtacgccaaaaa1560ttttgactagcggaggctagaaggagagagatgggtgcgagagcgtcagtattaagcggg1620ggagaattagatcgcgatgggaaaaaattcggttaaggccagggggaaagaaaaaatata1680aattaaaacatatagtatgggcaagcagggagctagaacgattcgcagttaatcctggcc1740tgttagaaacatcagaaggctgtagacaaatactgggacagctacaaccatcccttcaga1800caggatcagaagaacttagatcattatataatacagtagcaaccctctattgtgtgcatc1860aaaggatagagataaaagacaccaaggaagctttagacaagatagaggaagagcaaaaca1920aaagtaagaccaccgcacagcaagcggccgctgatcttcagacctggaggaggagatatg1980agggacaattggagaagtgaattatataaatataaagtagtaaaaattgaaccattagga2040gtagcacccaccaaggcaaagagaagagtggtgcagagagaaaaaagagcagtgggaata2100ggagctttgttccttgggttcttgggagcagcaggaagcactatgggcgcagcgtcaatg2160acgctgacggtacaggccagacaattattgtctggtatagtgcagcagcagaacaatttg2220ctgagggctattgaggcgcaacagcatctgttgcaactcacagtctggggcatcaagcag2280ctccaggcaagaatcctggctgtggaaagatacctaaaggatcaacagctcctggggatt2340tggggttgctctggaaaactcatttgcaccactgctgtgccttggaatgctagttggagt2400aataaatctctggaacagatttggaatcacacgacctggatggagtgggacagagaaatt2460aacaattacacaagcttaatacactccttaattgaagaatcgcaaaaccagcaagaaaag2520aatgaacaagaattattggaattagataaatgggcaagtttgtggaattggtttaacata2580acaaattggctgtggtatataaaattattcataatgatagtaggaggcttggtaggttta2640agaatagtttttgctgtactttctatagtgaatagagttaggcagggatattcaccatta2700tcgtttcagacccacctcccaaccccgaggggacccgacaggcccgaaggaatagaagaa2760gaaggtggagagagagacagagacagatccattcgattagtgaacggatctcgacggtat2820cgattagactgtagcccaggaatatggcagctagattgtacacatttagaaggaaaagtt2880atcttggtagcagttcatgtagccagtggatatatagaagcagaagtaattccagcagag2940acagggcaagaaacagcatacttcctcttaaaattagcaggaagatggccagtaaaaaca3000gtacatacagacaatggcagcaatttcaccagtactacagttaaggccgcctgttggtgg3060gcggggatcaagcaggaatttggcattccctacaatccccaaagtcaaggagtaatagaa3120tctatgaataaagaattaaagaaaattataggacaggtaagagatcaggctgaacatctt3180aagacagcagtacaaatggcagtattcatccacaattttaaaagaaaaggggggattggg3240gggtacagtgcaggggaaagaatagtagacataatagcaacagacatacaaactaaagaa3300ttacaaaaacaaattacaaaaattcaaaattttcgggtttattacagggacagcagagat3360ccagtttggctgcattgatcacgtgaggggctctagactctagacacacaaaaaaccaac3420acacagatctaatgaaaataaagatcttttattgagaaacttatacagggtagcataatg3480ggctactgaccccgccttcaaacctatttggagactataagtgaaaattatcactggttt3540tggtagaagctggacatggtgacatatgcttcttcttgatttgatcccagggaagtaaga3600atgggctgaccctgagcaactgggttcaatgtcaggattccagattggagagaaaatgga3660gggggcagcgtgctgtttgtagtcccaaggctaagcaggaggtcctggtacacatgaggc3720ccattcttgccactctccctgcagtctagggacctggaagaggagagaataggggcgtca3780catgcactgacattcccagccaggcatgtgagggatgaatctcttccaaagctttctgga3840gtgacgactacatcctcagatgggcagttggggctctgcacatcccctccaagcctctgc3900ttctcagattcttctagttgctgaggaaacgtatcttgcagaaaaccttccacttcatct3960ctagcttgaatgtcatccaccctatgaatctggcagtccaggaaactttcaggattgaaa4020ctcacatttaaattttttcttggtttcttacaaagatgttccagagtcttcttatgatcg4080gggagactgggccatacgataggcttaatcctttttttccataacacacaggccaagatg4140accaacagagcgacagagaaaaaactcaaaatgctgatggtcaggcatggtggtagtaag4200ataggatccatctcccctgagctattattgatctctggagttctgaagtaataacttgga4260ctccattcactccagaagcctttaaaatagtgatcagggatggatcgaactttaatctca4320tacattgctgccggttggagctttctctgcaggagtgtcagctttgtgctggataaattc4380acatgcgtccatttgttttcatccttttcctggcggtaagctacatcatgcattaaaact4440tttacatacttcttttgcaagtgtgatgtattaaatgtcaccacaaagtcattggctcct4500tcccgatagatgacactcaggtcaaaaggagcctcaggtttaactatagtggttaggtct4560atttttttgcaggttagactcttttctccaaccttcacacatatattgctctttccaatc4620agtaagaatttctttgtctcgatgaaatatatctcttgtagtttcctgaaattcaggcac4680tttacctccacgagggccccacatatttcaaattccagattggtggtgttgacatctggg4740tcctcaaaagcacatgtcagtgaatgctgcgatccattcacttccaactggctatagcat4800gagaatgagtagtcatccagttctgcatcttccaagtctccattttgagcatagccactt4860tctccagaaacgacttgaagtaaagaaaaaaccatgccaaaagttgtacctagaattgtc4920attgggccgggattttcctccacgtccccgcatgttagtagacttcccctgccctcgccg4980gagcgagggggcagggcctgcatgtgaagggcgtcgtaggtgtccttggtggctgtactg5040agaccctggtaaaggccatcgtgccccttgcccctccggcgctcgcctttcatcccaatc5100tcactgtaggcctccgccatcttatctttctgcagttcattgtacaggccttcctgaggg5160ttcttccttctcggctttccccccatctcagggtcccggccacgtctcttgtccaaaaca5220tcgtactcctctcttcgtcctagattgagctcgttatagagctggttctggccctgcttg5280tacgcgggggcgtctgcgctcctgctgaacttcactctcagttcacatcctccttcttct5340tcttctggaaatcggcagctacagccatcttcctcttgagtagtttgtactggtctcata5400aatggttgtttgaatatatacaggagtttctttctgccccgtttgcagtaaagggtgata5460accagtgacaggagaaggaccccacaagtcccggccaagggcgcccagatgtagatatca5520caggcgaagtccagccccctcgtgtgcactgcgccccccgccgctggccggcacgcctct5580gggcgcagggacaggggctgcgacgcgatggtgggcgccggtgttggtggtcgcggcgct5640ggcgtcgtggttgaggagacggtgactgaggttccttggccccagtagtccatagcatag5700ctaccaccgtagtaataatgtttggcacagtagtaaatggctgtgtcatcagtttgcaga5760ctgttcatttttaagaaaacttggctcttggagttgtccttgatgatggtcagtctggat5820ttgagagctgaattatagtatgtggtttcactaccccatattactcccagccactccaga5880ccctttcgtggaggctggcgaatccagcttacaccatagtcgggtaatgagacccctgag5940acagtgcatgtgacggacaggctctgtgagggcgccaccaggccaggtcctgactcctgc6000agtttcacctcagatccgccgccacccgacccaccaccgcccgagccaccgccacctgtg6060atctccagcttggtcccccctccgaacgtgtacggaagcgtattaccctgttggcgaagt6120aggtggcaatatcttcttgctccaggttgctaattgtcagagaataatctgttccagacc6180cactgccactgaaccttgatgggactcctgagtgtaatcttgatgtatggtagatcagga6240gtttaacagttccatctggtttctgctgataccaatttaaatatttactaatgtcctgac6300ttgccctgcaactgatggtgactctgtctcccagagaggcagacagggaggatgtagtct6360gtgtcatctggatgtccggcctggcggcgtggagcagcaaggccagcggcaggagcaagg6420cggtcactggtaaggccatggatcctctagatcacgacacctgaaatggaagaaaaaaac6480tttgaaccactgtctgaggcttgagaatgaaccaagatccaaactcaaaaagggcaaatt6540ccaaggagaattacatcaagtgccaagctggcctaacttcagtctccacccactcagtgt6600ggggaaactccatcgcataaaacccctccccccaacctaaagacgacgtactccaaaagc6660tcgagaactaatcgaggtgcctggacggcgcccggtactcagtggagtcacatgaagcga6720cggctgaggacggaaaggcccttttcctttgtgtgggagaaacttatacagggtagcata6780atgggctactgaccccgccttcaaacctatttggagactataagtgaaaatgactcaccc6840gcccgctctcccggcaccttcatcttgtcctttccctcagaaagaggctgggaggcagag6900gctgaggcagcggtggccgggacggttaggagaaaaggagtctctgctggttttattctg6960cagctacctccccaggaagtggaggactgtggggcctttgagaagcacctgccgacaggg7020ccaagaaattcgcactccccctttcggttcacaggcaggaagccctggaggtttgagggt7080ttggggtgtgtgtatgtatctgtctgtctgaattttgctttttctctcatttgaccattg7140ttttaatgctcctttttttaaaaaaaataattcttatctaattcctatcttgattggtaa7200agtccatctctaggcaaatacaagttctcgatggaaaacaataagtaatgtaaaatacag7260catagcaaaactttaacctccaaatcaagcctctacttgaatccttttctgagggatgaa7320taaggcataggcatcaggggctgttgccaatgtgcattagctgtttgcagcctcaccttc7380tttcatggagtttaagatatagtgtattttcccaaggtttgaactagctcttcatttctt7440tatgttttaaatgcactgacctcccacattccctttttagtaaaatattcagaaataatt7500taaatacatcattgcaatgaaaataaatgttttttattaggcagaatccagatgctcaag7560gcccttcataatatcccccagtttagtagttggacttagggaacaaaggaacctttaata7620gaaattggacagcaagaaagcgagcttagtgatacttgtgatcctctagatcacgacacc7680tgaaatggaagaaaaaaactgcaccttcatcttgtcctttccctcagaaagaggctggga7740ggcagaggctgaggcagcggtggccgggacggttaggagaaaaggagtctctgctggttt7800tattctgcagctacctccccaggaagtggaggactgtggggcctttgagaagcacctgcc7860gacagggccaagaaattcgcactccccctttcggttcacaggcaggaagccctggaggtt7920tgagggtttggggtgtgtgtatgtatctgtctgtctgaattttgctttttctctcatttg7980accattgttttaatgctcctttttttaaaaaaaataattcttatctaattcctatcttga8040ttggtaaagtccatctctaggcaaatacaagttctcgatggaaaacaataagtaatgtaa8100aatacagcatagcaaaactttaacctccaaatcaagcctctacttgaatccttttctgag8160ggatgaataaggcataggcatcaggggctgttgccaatgtgcattagctgtttgcagcct8220caccttctttcatggagtttaagatatagtgtattttcccaaggtttgaactagctcttc8280atttctttatgttttaaatgcactgacctcccacattccctttttagtaaaatattcaga8340aataatttaaatacatcattgcaatgaaaataaatgttttttattaggcagaatccagat8400gctcaaggcccttcataatatcccccagtttagtagttggacttagggaacaaaggaacc8460tttaatagaaattggacagcaagaaagcgagcttagtgatacttgtaaaaagagacgcgt8520ctctaaaagtcctttccatggctgctcgcctgtgttgccacctggattctgcgcgggacg8580tccttctgctacgtcccttcggccctcaatccagcggaccttccttcccgcggcctgctg8640ccggctctgcggcctcttccgcgtcttcgccttcgccctcagacgagtcggatctccctt8700tgggccgcctccccgcctggaattcgagctcggtacctttaagaccaatgacttacaagg8760cagctgtagatcttagccactttttaaaagaaaaggggggactggaagggctaattcact8820cccaacgaagacaagatctgctttttgcttgtactgggtctctctggttagaccagatct8880gagcctgggagctctctggctaactagggaacccactgcttaagcctcaataaagcttgc8940cttgagtgcttcaagtagtgtgtgcccgtctgttgtgtgactctggtaactagagatccc9000tcagacccttttagtcagtgtggaaaatctctagcagtagtagttcatgtcatcttatta9060ttcagtatttataacttgcaaagaaatgaatatcagagagtgagaggaacttgtttattg9120cagcttataatggttacaaataaagcaatagcatcacaaatttcacaaataaagcatttt9180tttcactgcattctagttgtggtttgtccaaactcatcaatgtatcttatcatgtctggc9240tctagctatcccgcccctaactccgcccatcccgcccctaactccgcccagttccgccca9300ttctccgccccatggctgactaattttttttatttatgcagaggccgaggccgcctcggc9360ctctgagctattccagaagtagtgaggaggcttttttggaggcctagggacgtatggcca9420caacctgggctccccgggcgcgtactccacctcacccatcatccacgctgttttatgagt9480aaaggagaagaacttttcactggagttgtcccaattcttgttgaattagatggtgatgtt9540aatgggcacaaattttctgtcagtggagagggtgaaggtgatgcaacatacggaaaactt9600acccttaaatttatttgcactactggaaaactacctgttccatggccaacacttgtcact9660actttctcttatggtgttcaatgcttttcaagatacccagatcatatgaaacggcatgac9720tttttcaagagtgccatgcccgaaggttatgtacaggaaagaactatatttttcaaagat9780gacgggaactacaagacacgtgctgaagtcaagtttgaaggtgatacccttgttaataga9840atcgagttaaaaggtattgattttaaagaagatggaaacattcttggacacaaattggaa9900tacaactataactcacacaatgtatacatcatggcagacaaacaaaagaatggaatcaaa9960gttaacttcaaaattagacacaacattgaagatggaagcgttcaactagcagaccattat10020caacaaaatactccaattggcgatggccctgtccttttaccagacaaccattacctgtcc10080acacaatctgccctttcgaaagatcccaacgaaaagagagaccacatggtccttcttgag10140tttgtaacagctgctgggattacacatggcatggatgaactatacaaataggacctccat10200agaagacaccgggaccgatccaataacttcgtatagcatacattatacgaagttatgcct10260ccggactctagcgtttaaacttaagcttgggaagttcctattccgaagttcctattctct10320agaaagtataggaacttctaccgagctcggatccactagtccagtgtggtggaattctgc10380agatatccagcacagtggcggccgctcgagtctagagggcccgtttaaacccgctgatca10440gcctcgactgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttcc10500ttgaccctggaaggtgccactcccactgtcctttcctaataaaatgaggaaattgcatcg10560cattgtctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaagggg10620gaggattgggaagacaatagcaggcatgctggggatgcggtgggctctatggcttctgag10680gcggaaagttaaccctagaaagataatcatattgtgacgtacgttaaagataatcatgcg10740taaaattgacgcatgtgttttatcggtctgtatatcgaggtttatttattaatttgaata10800gatattaagttttattatatttacacttacatactaataataaattcaacaaacaattta10860tttatgtttatttatttattaaaaaaaaacaaaaactcaaaatttcttctataaagtaac10920aaaaaccagctggggctcgaagttcctatactttctagagaataggaacttctatagtga10980gtcgaataagggcgacacaaaatttattctaaatgcataataaatactgataacatctta11040tagtttgtattatattttgtattatcgttgacatgtataattttgatatcaaaaactgat11100tttccctttattattttcgagatttattttcttaattctctttaacaaactagaaatatt11160gtatatacaaaaaatcataaataatagatgaatagtttaattataggtgttcatcaatcg11220aaaaagcaacgtatcttatttaaagtgcgttgcttttttctcatttataaggttaaataa11280ttctcatatatcaagcaaagtgacaggcgcccttaaatattctgacaaatgctctttccc11340taaactccccccataaaaaaacccgccgaagcgggtttttacgttatttgcggattaacg11400attactcgttatcagaaccgcccagggggcccgagcttaagactggccgtcgttttacaa11460cacagaaagagtttgtagaaacgcaaaaaggccatccgtcaggggccttctgcttagttt11520gatgcctggcagttccctactctcgccttccgcttcctcgctcactgactcgctgcgctc11580ggtcgttcggctgcggcgagcggtatcagctcactcaaaggcggtaatacggttatccac11640agaatcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggccaggaa11700ccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgacgagcatca11760caaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggc11820gtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggata11880cctgtccgcctttctcccttcgggaagcgtggcgctttctcatagctcacgctgtaggta11940tctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccgttca12000gcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggtaagacacga12060cttatcgccactggcagcagccactggtaacaggattagcagagcgaggtatgtaggcgg12120tgctacagagttcttgaagtggtgggctaactacggctacactagaagaacagtatttgg12180tatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgatccgg12240caaacaaaccaccgctggtagcggtggtttttttgtttgcaagcagcagattacgcgcag12300aaaaaaaggatctcaagaagatcctttgatcttttctacggggtctgacgctcagtggaa12360cgacgcgcgcgtaactcacgttaagggattttggtcatgagcttgcgccgtcccgtcaag12420tcagcgtaatgctctgcttttagaaaaactcatcgagcatcaaatgaaactgcaatttat12480tcatatcaggattatcaataccatatttttgaaaaagccgtttctgtaatgaaggagaaa12540actcaccgaggcagttccataggatggcaagatcctggtatcggtctgcgattccgactc12600gtccaacatcaatacaacctattaatttcccctcgtcaaaaataaggttatcaagtgaga12660aatcaccatgagtgacgactgaatccggtgagaatggcaaaagtttatgcatttctttcc12720agacttgttcaacaggccagccattacgctcgtcatcaaaatcactcgcatcaaccaaac12780cgttattcattcgtgattgcgcctgagcgaggcgaaatacgcgatcgctgttaaaaggac12840aattacaaacaggaatcgagtgcaaccggcgcaggaacactgccagcgcatcaacaatat12900tttcacctgaatcaggatattcttctaatacctggaacgctgtttttccggggatcgcag12960tggtgagtaaccatgcatcatcaggagtacggataaaatgcttgatggtcggaagtggca13020taaattccgtcagccagtttagtctgaccatctcatctgtaacatcattggcaacgctac13080ctttgccatgtttcagaaacaactctggcgcatcgggcttcccatacaagcgatagattg13140tcgcacctgattgcccgacattatcgcgagcccatttatacccatataaatcagcatcca13200tgttggaatttaatcgcggcctcgacgtttcccgttgaatatggctcatattcttccttt13260ttcaatattattgaagcatttatcagggttattgtctcatgagcggatacatatttgaat13320gtatttagaaaaataaacaaataggggtcagtgttacaaccaattaaccaattctgaaca13380ttatcgcgagcccatttatacctgaatatggctcataacaccccttgtttgcctggcggc13440agtagcgcggtggtcccacctgaccccatgccgaactcagaagtgaaacgccgtagcgcc13500gatggtagtgtggggactccccatgcgagagtagggaactgccaggcatcaaataaaacg13560aaaggctcagtcgaaagactgggcctttcgcccgggctaattagggggtgtcgcccttat13620tcgactctatagtgaagttcctattctctagaaagtataggaacttctgaagt13673<210>84<211>15025<212>dna<213>artificialsequence<220><223>syntheticdox-rapamycininduciblegagpolenv<400>84gatatctataacaagaaaatatatatataataagttatcacgtaagtagaacatgaaata60acaatataattatcgtatgagttaaatcttaaaagtcacgtaaaagataatcatgcgtca120ttttgactcacgcggtcgttatagttcaaaatcagtgacacttaccgcattgacaagcac180gcctcacgggagctccaagcggcgactgagatgtcctaaatgcacagcgacggattcgcg240ctatttagaaagagagagcaatatttcaagaatgcatgcgtcaattttacgcagactatc300tttctagggttaagacggatcgggagatctcccgatcccctatggtgcactctcagtaca360atctgctctgatgccgcatagttaagccagtatctgctccctgcttgtgtgttggaggtc420gctgagtagtgcgcgagcaaaatttaagctacaacaaggcaaggcttgaccgacaattgc480atgaagaatctgcttagggttaggcgttttgaagttcctatactttctagagaataggaa540cttcggaataggaacttcggatgcaatttcctcattttattaggaaaggacagtgggagt600ggcaccttccagggtcaaggaaggcacgggggaggggcaaacaacagatggctggcaact660agaaggcacagcgttagtgatacttgtgggccagggcattagccacaccagccaccactt720tctgataggcagcctgcactggtggggtgaattccgcggaagcttgtgtaattgttaatt780tctctgtcccactccatccaggtcgtgtgattccaaatctgttccagagatttattactc840caactagcattccaaggcacagcagtggtgcaaatgagttttccagagcaaccccaaatc900cccaggagctgttgatcctttaggtatctttccacagccaggattcttgcctggagctgc960ttgatgccccagactgtgagttgcaacagatgctgttgcgcctcaatagccctcagcaaa1020ttgttctgctgctgcactataccagacaataattgtctggcctgtaccgtcagcgtcatt1080gacgctgcgcccatagtgcttcctgctgctcccaagaacccaaggaacaaagctcctgcg1140gccgctccggaattccatgtgttaatcctcatcctgtctacttgccacacaatcatcacc1200tgccatctgttttccataatccctgatgatctttgcttttcttcttggcactacttttat1260gtcactattatcttgtattactactgccccttcacctttccagaggagctttgctggtcc1320tttccaaactggatctctgctgtccctgtaataaacccgaaaattttgaatttttgtaat1380ttgtttttgtaattctttagtttgtatgtctgttgctattatgtctactattctttcccc1440tgcactgtaccccccaatccccccttttcttttaaaattgtggatgaatactgccatttg1500tactgctgtcttaagatgttcagcctgatctcttacctgtcctataattttctttaattc1560tttattcatagattctattactccttgactttggggattgtagggaatgccaaattcctg1620cttgatccccgcccaccaacaggcggccttaactgtagtactggtgaaattgctgccatt1680gtctgtatgtactgtttttactggccatcttcctgctaattttaagaggaagtatgctgt1740ttcttgccctgtctctgctggaattacttctgcttctatatatccactggctacatgaac1800tgctaccaagataacttttccttctaaatgtgtacaatctagctgccatattcctgggct1860acagtctacttgtccatgcatggcttccccttttagctgacatttatcacagctggctac1920tatttcttttgctactacaggtggtaggttaaaatcactagccattgctctccaattact1980gtgatatttctcatgttcttcttgggccttatctattccatctaaaaatagtactttcct2040gattccagcactgaccaatttatctacttgttcatttcctccaattcctttgtgtgctgg2100tacccatgccaggtagactttttccttttttattaactgctctattatttgactgactaa2160ctctgattcactcttatctggttgtgcttgaatgattcccaatgcatattgtgagtctgt2220cactatgtttacttctaatcccgaatcctgcaaagctagatgaattgcttgtaactcagt2280cttctgatttgttgtgtccgttagggggacaactttttgtcttcctctgtcagttacata2340tcctgcttttcctaatttagtttccctattggctgccccatctacatagaaagtttctgc2400tcctattatgggttctttctctaactggtaccataacttcactaagggaggggtattgac2460aaactcccactcaggaatccaggtggcttgccaatactctgtccaccatgcttcccatgt2520ttccttttgtatgggtaatttaaatttaggagtctttccccatattactatgctttctgt2580ggctattttttgtactgcctctgttaattgtttcacatcattagtgtgggcacccttcat2640tcttgcatactttcctgttttcagatttttaaatggctcttgataaatttgatatgtcca2700ttggccttgcccctgcttctgtatttctgctattaagtcttttgatgggtcataatacac2760tccatgtaccggttcttttagaatctccctgttttctgccagttctagctctgcttcttc2820tgttagtggtactacttctgttagtgctttggttcccctaagaagtttacataattgcct2880tactttaatccctgcataaatctgacttgcccaattcaattttcccactaatttctgtat2940gtcattgacagtccagctgtccttttctggcagcactataggctgtactgtccatttatc3000aggatggagttcataacccatccaaaggaatggaggttctttctgatgttttttgtctgg3060tgtggtaaatccccacctcaacagatgttgtctcagttcctctatttttgttctatgctg3120ccctatttctaagtcagatcctacatacaaatcatccatgtattgatagatgactatgtc3180tggattttgttttctaaaaggctctaagatttttgtcatgctacactggaatattgctgg3240tgatcctttccatccctgtggaagcacattgtactgatatctaatccctggtgtctcatt3300gtttatactaggtatggtaaatgcagtatacttcctgaagtctttatctaagggaactga3360aaaatatgcatcgcccacatccagtactgttactgattttttctgttttaaccctgcagg3420atgtggtattcctaattgaacttcccagaaatcttgagttctcttattaagttctctgaa3480atctactaattttctccatttagtactgtcttttttctttatggcaaatactggagtatt3540gtatggattttcaggcccaatttttgaaatttttccttccttttccatttctgtacaaat3600ttctactaatgcttttattttttcttctgtcaatggccattgtttaacttttgggccatc3660cattcctggctttaattttactggtacagtctcaataggactaatgggaaaatttaaagt3720gcagccaatctgagtcaacagatttcttccaattatgttgacaggtgtaggtcctactaa3780tactgtacctatagctttatgtccgcagatttctatgagtatctgatcatactgtcttac3840tttgataaaacctccaattccccctatcatttttggtttccatcttcctggcaaattcat3900ttcttctaatactgtatcatctgctcctgtatctaatagagcttcctttaattgcccccc3960tatctttattgtgacgaggggtcgctgccaaagagtgatctgagggaagctaaaggatac4020agttccttgtctatcggctcctgcttctgagagggagttgttgtctcttccccaaacctg4080aagctctcttctggtggggctgttggctctggtctgctctgaagaaaattccctggcctt4140cccttgtgggaaggccagatcttccctaaaaaattagcctgtctctcagtacaatctttc4200atttggtgtccttcctttccacatttccaacagccctttttcctaggggccctgcaattt4260ttggctatgtgcccttctttgccacaattgaaacacttaacagtctttctttggttccta4320aaattgcctttctgtatcattatggtagctggatttgttacttggctcattgcttcagcc4380aaaactcttgctttatggccgggtccccccactccctgacatgctgtcatcatttcttct4440agtgtcgctcctggtcccaatgcttttaaaatagtcttacaatctgggttcgcattttgg4500accaacaaggtttctgtcatccaattttttacctcttgtgaagcttgctcggctcttaga4560gttttatagaatcggtctacatagtctctaaagggttcctttggtccttgtcttatgtcc4620agaatgctggtagggctatacattcttactattttatttaatcccaggattatccatctt4680ttatagatttctcctactgggataggtggattatgtgtcatccatcctatttgttcctga4740agggtactagtagttcctgctatgtcacttccccttggttctctcatctggcctggtgca4800ataggccctgcatgcactggatgcactctatcccattctgcagcttcctcattgatggtc4860tcttttaacatttgcatggctgcttgatgtccccccactgtgtttagcatggtgtttaaa4920tcttgtggggtggctccttctgataatgctgaaaacatgggtatcacttctgggctgaaa4980gccttctcttctactacttttacccatgcatttaaagttctaggtgatatggcctgatgt5040accatttgcccctggatgttctgcactatagggtaattttggctgacctgattgctgtgt5100cctgtgtcagctgctgcttgctgtgcttttttcttacttttgttttgctcttcctctatc5160ttgtctaaagcttccttggtgtcttttatctctatcctttgatgcacacaatagagggtt5220gctactgtattatataatgatctaagttcttctgatcctgtctgaagggatggttgtagc5280tgtcccagtatttgtctacagccttctgatgtttctaacaggccaggattaactgcgaat5340cgttctagctccctgcttgcccatactatatgttttaatttatattttttctttccccct5400ggccttaaccgaattttttcccatcgatctaattctcccccgcttaatactgacgctctc5460gcacccatggcggcggcagatctcgaattcagatctcacgtgctttgccaaagtgatggg5520ccagcacacagaccagcacgttgcccaggagctgtgggaggaagataagaggtatgaaca5580tgattagcaaaagggcctagcttggactcagaataatccagccttatcccaaccataaaa5640taaaagcagaatggtagctggattgtagctgctattagcaatatgaaacctcttacatca5700gttacaatttatatgcagaaatatttatatgcagaaatattgctattgccttaacccaga5760aattatcactgttattctttagaatggtgcaaagaggcatgatacattgtatcattattg5820ccctgaaagaaagagattagggaaagtattagaaataagataaacaaaaaagtatattaa5880aagaagaaagcattttttaaaattacaaatgcaaaattaccctgatttggtcaatatgtg5940taccctgttacttctccccttcctatgacatgaacttaaccatagaaaagaaggggaaag6000aaaacatcaagggtcccatagactcaccctgaagttctcaggatccgagctcggtaccac6060atgtaagcttcgaggggaggctggatcggtcccggtgtcttctatggaggtcaaaacagc6120gtggatggcgtctccaggcgatctgacggttcactaaacgctgcttcgcgatgtacgggc6180cagatatacgcgttgcgatctgacggttcactaaacgagctctgcttatataggcctccc6240accgtacacgccacctcgacatactcgagtttactccctatcagtgatagagaacgtatg6300aagagtttactccctatcagtgatagagaacgtatgcagactttactccctatcagtgat6360agagaacgtataaggagtttactccctatcagtgatagagaacgtatgaccagtttactc6420cctatcagtgatagagaacgtatctacagtttactccctatcagtgatagagaacgtata6480tccagtttactccctatcagtgatagagaacgtataagctttaggcgtgtacggtgggcg6540cctataaaagcagagctcgtttagtgaaccgtcagatcgcctggagcaattccacaacac6600ttttgtcttataccaactttccgtaccacttcctaccctcgtaaatcgtcgacgagctcg6660tttagtgaaccgtcagatcgcctggagacgccctcgaagccgcggtgcgggtgccagggc6720gtgcccttgggctccatgtccatcatgggtctcaaggtgaacgtctctgccatattcatg6780gcagtactgttaactctccaaacacccaccggtcaaatccattggggcaatctctctaag6840ataggggtggtaggaataggaagtgcaagctacaaagttatgactcgttccagccatcaa6900tcattagtcataaaattaatgcccaatataactctcctcaataactgcacgagggtagag6960attgcagaatacaggagactactgagaacagttttggaaccaattagagatgcacttaat7020gcaatgacccagaatataagaccggttcagagtgtagcttcaagtaggagacacaagaga7080tttgcgggagtagtcctggcaggtgcggccctaggcgttgccacagctgctcagataaca7140gccggcattgcacttcaccagtccatgctgaactctcaagccatcgacaatctgagagcg7200agcctggaaactactaatcaggcaattgaggcaatcagacaagcagggcaggagatgata7260ttggctgttcagggtgtccaagactacatcaataatgagctgataccgtctatgaaccaa7320ctatcttgtgatttaatcggccagaagctcgggctcaaattgctcagatactatacagaa7380atcctgtcattatttggccccagcttacgggaccccatatctgcggagatatctatccag7440gctttgagctatgcgcttggaggagacatcaataaggtgttagaaaagctcggatacagt7500ggaggtgatttactgggcatcttagagagcagaggaataaaggcccggataactcacgtc7560gacacagagtcctacttcattgtcctcagtatagcctatccgacgctgtccgagattaag7620ggggtgattgtccaccggctagagggggtctcgtacaacataggctctcaagagtggtat7680accactgtgcccaagtatgttgcaacccaagggtaccttatctcgaattttgatgagtca7740tcgtgtactttcatgccagaggggactgtgtgcagccaaaatgccttgtacccgatgagt7800cctctgctccaagaatgcctccgggggtccaccaagtcctgtgctcgtacactcgtatcc7860gggtcttttgggaaccggttcattttatcacaagggaacctaatagccaattgtgcatca7920atcctttgcaagtgttacacaacaggaacgatcattaatcaagaccctgacaagatccta7980acatacattgctgccgatcactgcccggtagtcgaggtgaacggcgtgaccatccaagtc8040gggagcaggaggtatccagacgctgtgtacttgcacagaattgacctcggtcctcccata8100tcattggagaggttggacgtagggacaaatctggggaatgcaattgctaagttggaggat8160gccaaggaattgttggagtcatcggaccagatattgaggagtatgaaaggtttatcgagc8220actagcatagtctacatcctgattgcagtgtgtcttggagggttgatagggatccccgct8280ttaatatgttgctgcagggggcgttgacccctctccctcccccccccctaacgttactgg8340ccgaagccgcttggaataaggccggtgtgcgtttgtctatatgttattttccaccatatt8400gccgtcttttggcaatgtgagggcccggaaacctggccctgtcttcttgacgagcattcc8460taggggtctttcccctctcgccaaaggaatgcaaggtctgttgaatgtcgtgaaggaagc8520agttcctctggaagcttcttgaagacaaacaacgtctgtagcgaccctttgcaggcagcg8580gaaccccccacctggcgacaggtgcctctgcggccaaaagccacgtgtataagatacacc8640tgcaaaggcggcacaaccccagtgccacgttgtgagttggatagttgtggaaagagtcaa8700atggctctcctcaagcgtattcaacaaggggctgaaggatgcccagaaggtaccccattg8760tatgggatctgatctggggcctcggtacacatgctttacatgtgtttagtcgaggttaaa8820aaaacgtctaggccccccgaaccacggggacgtggttttcctttgaaaaacacgatgata8880atatggccacaaccatgggaagtaggatagtcattaacagagaacatcttatgattgata8940gaccttatgttttgctggctgttctgtttgtcatgtttctgagcttgatcgggttgctag9000ccattgcaggcattagacttcatcgggcagccatctacaccgcagagatccataaaagcc9060tcagcaccaatctagatgtaactaactcaatcgagcatcaggtcaaggacgtgctgacac9120cactcttcaaaatcatcggtgatgaagtgggcctgaggacacctcagagattcactgacc9180tagtgaaattcatctctgacaagattaaattccttaatccggatagggagtacgacttca9240gagatctcacttggtgtatcaacccgccagagagaatcaaattggattatgatcaatact9300gtgcagatgtggctgctgaagagctcatgaatgcattggtgaactcaactctactggaga9360ccagaacaaccaatcagttcctagctgtctcaaagggaaactgctcagggcccactacaa9420tcagaggtcaattctcaaacatgtcgctgtccctgttagacttgtatttaggtcgaggtt9480acaatgtgtcatctatagtcactatgacatcccagggaatgtatgggggaacttacctag9540tggaaaagcctaatctgagcagcaaaaggtcagagttgtcacaactgagcatgtaccgag9600tgtttgaagtaggtgttatcagaaatccgggtttgggggctccggtgttccatatgacaa9660actatcttgagcaaccagtcagtaatgatctcagcaactgtatggtggctttgggggagc9720tcaaactcgcagccctttgtcacggggaagattctatcacaattccctatcagggatcag9780ggaaaggtgtcagcttccagctcgtcaagctaggtgtctggaaatccccaaccgacatgc9840aatcctgggtccccttatcaacggatgatccagtgatagacaggctttacctctcatctc9900acagaggtgttatcgctgacaaycaagcaaaatgggctgtcccgacaacacgaacagatg9960acaagttgcgaatggagacatgcttccaacaggcgtgtaagggtaaaatccaagcactct10020gcgagaatcccgagtgggcaccattgaaggataacaggattccttcatacggggtcttgt10080ctgttgatctgagtctgacagttgagcttaaaatcaaaattgcttcgggattcgggccat10140tgatcacacacggttcagggatggacctatacaaatccaaccacaacaatgtgtattggc10200tgactatcccrccaatgaagaacctagccttaggtgtaatcaacacattggagtggatac10260cgagattcaaggttagtccctacctcttcamtgtcccaattaaggaagcaggcgaagact10320gccatgccccaacatacctacctgcggaggtggatggtgatgtcaaactcagttccaatc10380tggtgattctacctggtcaagatctccaatatgttttggcaacctacgatacttccaggg10440ttgaacatgctgtggtttattacgtttacagcccaagccgctcattttcttacttttatc10500cttttaggttgcctataaagggggtccccatcgaattacaagtggaatgcttcacatggg10560accaaaaactctggtgccgtcacttctgtgtgcttgcggactcagaatctggtggacata10620tcactcactctgggatggtgggcatgggagtcagctgcacagtcacccgggaagatggaa10680ccaatcgcagatagggctgctagtgaacyaatcwcatgatgtcacccagacatcaggcat10740acccactagtgtgaaatagacatcagaattaagaaaaatgggctccccgggcgcgtactc10800cacctcacccatcatccacgctcggcaataaaaagacagaataaaacgcacgggtgttgg10860gtcgtttgttcgccgggcgcgtactccacctcacccatcatccacgctgttttatggata10920gcactgagaacgtcatcaagcccttcatgcgcttcaaggtgcacatggagggctccgtga10980acggccacgagttcgagatcgagggcgagggcgagggcaagccctacgagggcacccaga11040ccgccaagctgcaggtgaccaagggcggccccctgcccttcgcctgggacatcctgtccc11100cccagttccagtacggctccaaggtgtacgtgaagcaccccgccgacatccccgactaca11160agaagctgtccttccccgagggcttcaagtgggagcgcgtgatgaacttcgaggacggcg11220gcgtggtgaccgtgacccaggactcctccctgcaggacggcaccttcatctaccacgtga11280agttcatcggcgtgaacttcccctccgacggccccgtaatgcagaagaagactctgggct11340gggagccctccaccgagcgcctgtacccccgcgacggcgtgctgaagggcgagatccaca11400aggcgctgaagctgaagggcggcggccactacctggtggagttcaagtcaatctacatgg11460ccaagaagcccgtgaagctgcccggctactactacgtggactccaagctggacatcacct11520cccacaacgaggactacaccgtggtggagcagtacgagcgcgccgaggcccgccaccacc11580tgttccagtaggacctccatagaagacaccgggaccgatccaataacttcgtatagcata11640cattatacgaagttatgcctccggactctagcgtttaaacttaagcttggtaccgagctc11700ggatccactagtccagtgtggtggaattctgcagatatccagcacagtggcggccgctcg11760agtctagagggcccgtttaaacccgctgatcagcctcgactgtgccttctagttgccagc11820catctgttgtttgcccctcccccgtgccttccttgaccctggaaggtgccactcccactg11880tcctttcctaataaaatgaggaaattgcatcgcattgtctgagtaggtgtcattctattc11940tggggggtggggtggggcaggacagcaagggggaggattgggaagacaatagcaggcatg12000ctggggatgcggtgggctctatggcttctgaggcggaaagttaaccctagaaagataatc12060atattgtgacgtacgttaaagataatcatgcgtaaaattgacgcatgtgttttatcggtc12120tgtatatcgaggtttatttattaatttgaatagatattaagttttattatatttacactt12180acatactaataataaattcaacaaacaatttatttatgtttatttatttattaaaaaaaa12240acaaaaactcaaaatttcttctataaagtaacaaaaaccagctggggctcgaagttccta12300tactttctagagaataggaacttctatagtgagtcgaataagggcgacacaaaatttatt12360ctaaatgcataataaatactgataacatcttatagtttgtattatattttgtattatcgt12420tgacatgtataattttgatatcaaaaactgattttccctttattattttcgagatttatt12480ttcttaattctctttaacaaactagaaatattgtatatacaaaaaatcataaataataga12540tgaatagtttaattataggtgttcatcaatcgaaaaagcaacgtatcttatttaaagtgc12600gttgcttttttctcatttataaggttaaataattctcatatatcaagcaaagtgacaggc12660gcccttaaatattctgacaaatgctctttccctaaactccccccataaaaaaacccgccg12720aagcgggtttttacgttatttgcggattaacgattactcgttatcagaaccgcccagggg12780gcccgagcttaagactggccgtcgttttacaacacagaaagagtttgtagaaacgcaaaa12840aggccatccgtcaggggccttctgcttagtttgatgcctggcagttccctactctcgcct12900tccgcttcctcgctcactgactcgctgcgctcggtcgttcggctgcggcgagcggtatca12960gctcactcaaaggcggtaatacggttatccacagaatcaggggataacgcaggaaagaac13020atgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgttt13080ttccataggctccgcccccctgacgagcatcacaaaaatcgacgctcaagtcagaggtgg13140cgaaacccgacaggactataaagataccaggcgtttccccctggaagctccctcgtgcgc13200tctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcgggaagc13260gtggcgctttctcatagctcacgctgtaggtatctcagttcggtgtaggtcgttcgctcc13320aagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaac13380tatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagccactggt13440aacaggattagcagagcgaggtatgtaggcggtgctacagagttcttgaagtggtgggct13500aactacggctacactagaagaacagtatttggtatctgcgctctgctgaagccagttacc13560ttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtagcggtggt13620ttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcctttg13680atcttttctacggggtctgacgctcagtggaacgacgcgcgcgtaactcacgttaaggga13740ttttggtcatgagcttgcgccgtcccgtcaagtcagcgtaatgctctgcttttagaaaaa13800ctcatcgagcatcaaatgaaactgcaatttattcatatcaggattatcaataccatattt13860ttgaaaaagccgtttctgtaatgaaggagaaaactcaccgaggcagttccataggatggc13920aagatcctggtatcggtctgcgattccgactcgtccaacatcaatacaacctattaattt13980cccctcgtcaaaaataaggttatcaagtgagaaatcaccatgagtgacgactgaatccgg14040tgagaatggcaaaagtttatgcatttctttccagacttgttcaacaggccagccattacg14100ctcgtcatcaaaatcactcgcatcaaccaaaccgttattcattcgtgattgcgcctgagc14160gaggcgaaatacgcgatcgctgttaaaaggacaattacaaacaggaatcgagtgcaaccg14220gcgcaggaacactgccagcgcatcaacaatattttcacctgaatcaggatattcttctaa14280tacctggaacgctgtttttccggggatcgcagtggtgagtaaccatgcatcatcaggagt14340acggataaaatgcttgatggtcggaagtggcataaattccgtcagccagtttagtctgac14400catctcatctgtaacatcattggcaacgctacctttgccatgtttcagaaacaactctgg14460cgcatcgggcttcccatacaagcgatagattgtcgcacctgattgcccgacattatcgcg14520agcccatttatacccatataaatcagcatccatgttggaatttaatcgcggcctcgacgt14580ttcccgttgaatatggctcatattcttcctttttcaatattattgaagcatttatcaggg14640ttattgtctcatgagcggatacatatttgaatgtatttagaaaaataaacaaataggggt14700cagtgttacaaccaattaaccaattctgaacattatcgcgagcccatttatacctgaata14760tggctcataacaccccttgtttgcctggcggcagtagcgcggtggtcccacctgacccca14820tgccgaactcagaagtgaaacgccgtagcgccgatggtagtgtggggactccccatgcga14880gagtagggaactgccaggcatcaaataaaacgaaaggctcagtcgaaagactgggccttt14940cgcccgggctaattagggggtgtcgcccttattcgactctatagtgaagttcctattctc15000tagaaagtataggaacttctgaagt15025<210>85<211>6584<212>dna<213>artificialsequence<220><223>syntheticrapamycin-inducibletetactivator<400>85gatatctataacaagaaaatatatatataataagttatcacgtaagtagaacatgaaata60acaatataattatcgtatgagttaaatcttaaaagtcacgtaaaagataatcatgcgtca120ttttgactcacgcggtcgttatagttcaaaatcagtgacacttaccgcattgacaagcac180gcctcacgggagctccaagcggcgactgagatgtcctaaatgcacagcgacggattcgcg240ctatttagaaagagagagcaatatttcaagaatgcatgcgtcaattttacgcagactatc300tttctagggttaagacggatcgggagatctcccgatcccctatggtgcactctcagtaca360atctgctctgatgccgcatagttaagccagtatctgctccctgcttgtgtgttggaggtc420gctgagtagtgcgcgagcaaaatttaagctacaacaaggcaaggcttgaccgacaattgc480atgaagaatctgcttagggttaggcgttttgcgctgcttcgcgatgtacgggccagatat540acgcgttctccatagaagacaccgaataaaatatctttattttcattacatctgtgtgtt600ggttttttgtgtgaatcgatagtactaacatacgctctccatcaaaacaaaacgaaacaa660aacaaactagcaaaataggctgtccccagtgcaagtgcaggtgccagaacatttctctgg720accgatccaataacttcgtatagcatacattatacgaagttatgcctccggactctagcg780ttttagttattactagcgctaccggactcagatctcgagctcaagcttcgaattctgcag840tcgacggtaccgcggcttacgcgtgctagctaatgatgggcgctcgagtaatgatgggcg900gtcgactaatgatgggcgctcgagtaatgatgggcgtctagctaatgatgggcgctcgag960taatgatgggcggtcgactaatgatgggcgctcgagtaatgatgggcgtctagctaatga1020tgggcgctcgagtaatgatgggcggtcgactaatgatgggcgctcgagtaatgatgggcg1080tctagaacgcgaattaattcaacattttgacacccccataatatttttccagaattaaca1140gtataaattgcatctcttgttcaagagttccctatcactctctttaatcactactcacag1200taacctcaactcctgccacaagcttgccctgcagcgggaattccaaacttaagcttggta1260ccgagctcggatccactagtccagtgtggtggaattctgcagatatccagcacagtggcg1320gccgctcgagtctagagggcccgtttaaacccgctgatcaatgtctagactggacaagag1380caaagtcataaactctgctctggaattactcaatggagtcggtatcgaaggcctgacgac1440aaggaaactcgctcaaaagctgggagttgagcagcctaccctgtactggcacgtgaagaa1500caagcgggccctgctcgatgccctgccaatcgagatgctggacaggcatcatacccactc1560ctgccccctggaaggcgagtcatggcaagactttctgcggaacaacgccaagtcataccg1620ctgtgctctcctctcacatcgcgacggggctaaagtgcatctcggcacccgcccaacaga1680gaaacagtacgaaaccctggaaaatcagctcgcgttcctgtgtcagcaaggcttctccct1740ggagaacgcactgtacgctctgtccgccgtgggccactttacactgggctgcgtattgga1800ggaacaggagcatcaagtagcaaaagaggaaagagagacacctaccaccgattctatgcc1860cccacttctgaaacaagcaattgagctgttcgaccggcagggagccgaacctgccttcct1920tttcggcctggaactaatcatatgtggcctggagaaacagctaaagtgcgaaagcggcgg1980gccgaccgacgcccttgacgattttgacttagacatgctcccagccgatgcccttgacga2040ctttgaccttgatatgctgcctgctgacgctcttgacgattttgaccttgacatgctccc2100cgggtaagtccctcccccccccctaacgttactggccgaagccgcttggaataaggccgg2160tgtgcgtttgtctatatgttattttccaccatattgccgtcttttggcaatgtgagggcc2220cggaaacctggccctgtcttcttgacgagcattcctaggggtctttcccctctcgccaaa2280ggaatgcaaggtctgttgaatgtcgtgaaggaagcagttcctctggaagcttcttgaaga2340caaacaacgtctgtagcgaccctttgcaggcagcggaaccccccacctggcgacaggtgc2400ctctgcggccaaaagccacgtgtataagatacacctgcaaaggcggcacaaccccagtgc2460cacgttgtgagttggatagttgtggaaagagtcaaatggctctcctcaagcgtattcaac2520aaggggctgaaggatgcccagaaggtaccccattgtatgggatctgatctggggcctcgg2580tgcacatgctttacatgtgtttagtcgaggttaaaaaacgtctaggccccccgaaccacg2640gggacgtggttttcctttgaaaaacacgatgataaatggatagcactgagaacgtcatca2700agcccttcatgcgcttcaaggtgcacatggagggctccgtgaacggccacgagttcgaga2760tcgagggcgagggcgagggcaagccctacgagggcacccagaccgccaagctgcaggtga2820ccaagggcggccccctgcccttcgcctgggacatcctgtccccccagttccagtacggct2880ccaaggtgtacgtgaagcaccccgccgacatccccgactacaagaagctgtccttccccg2940agggcttcaagtgggagcgcgtgatgaacttcgaggacggcggcgtggtgaccgtgaccc3000aggactcctccctgcaggacggcaccttcatctaccacgtgaagttcatcggcgtgaact3060tcccctccgacggccccgtaatgcagaagaagactctgggctgggagccctccaccgagc3120gcctgtacccccgcgacggcgtgctgaagggcgagatccacaaggcgctgaagctgaagg3180gcggcggccactacctggtggagttcaagtcaatctacatggccaagaagcccgtgaagc3240tgcccggctactactacgtggactccaagctggacatcacctcccacaacgaggactaca3300ccgtggtggagcagtacgagcgcgccgaggcccgccaccacctgttccagtagctcgact3360gtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgaccctg3420gaaggtgccactcccactgtcctttcctaataaaatgaggaaattgcatcgcattgtctg3480agtaggtgtcattctattctggggggtggggtggggcaggacagcaagggggaggattgg3540gaagacaatagcaggcatgctggggatgcggtgggctctatggcttctgaggcggaaagt3600taaccctagaaagataatcatattgtgacgtacgttaaagataatcatgcgtaaaattga3660cgcatgtgttttatcggtctgtatatcgaggtttatttattaatttgaatagatattaag3720ttttattatatttacacttacatactaataataaattcaacaaacaatttatttatgttt3780atttatttattaaaaaaaaacaaaaactcaaaatttcttctataaagtaacaaaaaccag3840ctggggctcgaagttcctatactttctagagaataggaacttctatagtgagtcgaataa3900gggcgacacaaaatttattctaaatgcataataaatactgataacatcttatagtttgta3960ttatattttgtattatcgttgacatgtataattttgatatcaaaaactgattttcccttt4020attattttcgagatttattttcttaattctctttaacaaactagaaatattgtatataca4080aaaaatcataaataatagatgaatagtttaattataggtgttcatcaatcgaaaaagcaa4140cgtatcttatttaaagtgcgttgcttttttctcatttataaggttaaataattctcatat4200atcaagcaaagtgacaggcgcccttaaatattctgacaaatgctctttccctaaactccc4260cccataaaaaaacccgccgaagcgggtttttacgttatttgcggattaacgattactcgt4320tatcagaaccgcccagggggcccgagcttaagactggccgtcgttttacaacacagaaag4380agtttgtagaaacgcaaaaaggccatccgtcaggggccttctgcttagtttgatgcctgg4440cagttccctactctcgccttccgcttcctcgctcactgactcgctgcgctcggtcgttcg4500gctgcggcgagcggtatcagctcactcaaaggcggtaatacggttatccacagaatcagg4560ggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaa4620ggccgcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcg4680acgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggcgtttccccc4740tggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgc4800ctttctcccttcgggaagcgtggcgctttctcatagctcacgctgtaggtatctcagttc4860ggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccg4920ctgcgccttatccggtaactatcgtcttgagtccaacccggtaagacacgacttatcgcc4980actggcagcagccactggtaacaggattagcagagcgaggtatgtaggcggtgctacaga5040gttcttgaagtggtgggctaactacggctacactagaagaacagtatttggtatctgcgc5100tctgctgaagccagttaccttcggaaaaagagttggtagctcttgatccggcaaacaaac5160caccgctggtagcggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaagg5220atctcaagaagatcctttgatcttttctacggggtctgacgctcagtggaacgacgcgcg5280cgtaactcacgttaagggattttggtcatgagcttgcgccgtcccgtcaagtcagcgtaa5340tgctctgcttttagaaaaactcatcgagcatcaaatgaaactgcaatttattcatatcag5400gattatcaataccatatttttgaaaaagccgtttctgtaatgaaggagaaaactcaccga5460ggcagttccataggatggcaagatcctggtatcggtctgcgattccgactcgtccaacat5520caatacaacctattaatttcccctcgtcaaaaataaggttatcaagtgagaaatcaccat5580gagtgacgactgaatccggtgagaatggcaaaagtttatgcatttctttccagacttgtt5640caacaggccagccattacgctcgtcatcaaaatcactcgcatcaaccaaaccgttattca5700ttcgtgattgcgcctgagcgaggcgaaatacgcgatcgctgttaaaaggacaattacaaa5760caggaatcgagtgcaaccggcgcaggaacactgccagcgcatcaacaatattttcacctg5820aatcaggatattcttctaatacctggaacgctgtttttccggggatcgcagtggtgagta5880accatgcatcatcaggagtacggataaaatgcttgatggtcggaagtggcataaattccg5940tcagccagtttagtctgaccatctcatctgtaacatcattggcaacgctacctttgccat6000gtttcagaaacaactctggcgcatcgggcttcccatacaagcgatagattgtcgcacctg6060attgcccgacattatcgcgagcccatttatacccatataaatcagcatccatgttggaat6120ttaatcgcggcctcgacgtttcccgttgaatatggctcatattcttcctttttcaatatt6180attgaagcatttatcagggttattgtctcatgagcggatacatatttgaatgtatttaga6240aaaataaacaaataggggtcagtgttacaaccaattaaccaattctgaacattatcgcga6300gcccatttatacctgaatatggctcataacaccccttgtttgcctggcggcagtagcgcg6360gtggtcccacctgaccccatgccgaactcagaagtgaaacgccgtagcgccgatggtagt6420gtggggactccccatgcgagagtagggaactgccaggcatcaaataaaacgaaaggctca6480gtcgaaagactgggcctttcgcccgggctaattagggggtgtcgcccttattcgactcta6540tagtgaagttcctattctctagaaagtataggaacttctgaagt6584<210>86<211>11528<212>dna<213>artificialsequence<220><223>syntheticrapamycininducerinduciblerevsrcvpx<400>86gatatctataacaagaaaatatatatataataagttatcacgtaagtagaacatgaaata60acaatataattatcgtatgagttaaatcttaaaagtcacgtaaaagataatcatgcgtca120ttttgactcacgcggtcgttatagttcaaaatcagtgacacttaccgcattgacaagcac180gcctcacgggagctccaagcggcgactgagatgtcctaaatgcacagcgacggattcgcg240ctatttagaaagagagagcaatatttcaagaatgcatgcgtcaattttacgcagactatc300tttctagggttaagacggatcgggagatctcccgatcccctatggtgcactctcagtaca360atctgctctgatgccgcatagttaagccagtatctgctccctgcttgtgtgttggaggtc420gctgagtagtgcgcgagcaaaatttaagctacaacaaggcaaggcttgaccgacaattgc480atgaagaatctgcttagggttaggcgttttgcgctgcttcgcgatgtacgggccagatat540acgcgttgacattgattattgactagttattaatagtaatcaattacggggtcattagtt600catagcccatatatggagttccgcgttacataacttacggtaaatggcccgcctggctga660ccgcccaacgacccccgcccattgacgtcaataatgacgtatgttcccatagtaacgcca720atagggactttccattgacgtcaatgggtggagtatttacggtaaactgcccacttggca780gtacatcaagtgtatcatatgccaagtacgccccctattgacgtcaatgacggtaaatgg840cccgcctggcattatgcccagtacatgaccttatgggactttcctacttggcagtacatc900tacgtattagtcatcgctattaccatggtgatgcggttttggcagtacatcaatgggcgt960ggatagcggtttgactcacggggatttccaagtctccaccccattgacgtcaatgggagt1020ttgttttggaaccaaaatcaacgggactttccaaaatgtcgtaacaactccgccccattg1080acgcaaatgggcggtaggcgtgtacggtgggaggtctatataagcagagctcggcattga1140ttattgactagttattaatagtaatcaattacggggtcattagttcatagcccatatatg1200gagttccgcgttacataacttacggtaaatggcccgcctggctgaccgcccaacgacccc1260cgcccattgacgtcaataatgacgtatgttcccatagtaacgccaatagggactttccat1320tgacgtcaatgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtat1380catatgccaagtccgccccctattgacgtcaatgacggtaaatggcccgcctggcattat1440gcccagtacatgaccttacgggactttcctacttggcagtacatctacgtattagtcatc1500gctattaccatggtgatgcggttttggcagtacaccaatgggcgtggatagcggtttgac1560tcacggggatttccaagtctccaccccattgacgtcaatgggagtttgttttggcaccaa1620aatcaacgggactttccaaaatgtcgtaacaactgcgatcgcccgccccgttgacgcaaa1680tgggcggtaggcgtgtacggtgggaggtctatataagcagagctcgtttagtgaaccgtc1740agatcactagaagctttattgcggtagtttatcacagttaaattgctaacgcagtcagtg1800cttctgacacaacagtctcgaacttaagctgcagtgactctcttaaggtagccttgcaga1860agttggtcgtgaggcactgggcaggtaagtatcaaggttacaagacaggtttaaggagac1920caatagaaactgggcttgtcgagacagagaagactcttgcgtttctgataggcacctatt1980ggtcttactgacatccactttgcctttctctccacaggtgtccactcccagttcaattac2040agctcttaaggctagagtacttaatacgactcactataggctagcctcgagaattcatgg2100cttctagaatcctctggcatgagatgtggcatgaaggcctggaagaggcatctcgtttgt2160actttggggaaaggaacgtgaaaggcatgtttgaggtgctggagcccttgcatgctatga2220tggaacggggcccccagactctgaaggaaacatcctttaatcaggcctatggtcgagatt2280taatggaggcccaagagtggtgcaggaagtacatgaaatcagggaatgtcaaggacctcc2340tccaagcctgggacctctattatcatgtgttccgacgaatctcaaagactagagatgagt2400ttcccaccatggtgtttccttctgggcagatcagccaggcctcggccttggccccggccc2460ctccccaagtcctgccccaggctccagcccctgcccctgctccagccatggtatcagctc2520tggcccaggccccagcccctgtcccagtcctagccccaggccctcctcaggctgtggccc2580cacctgcccccaagcccacccaggctggggaaggaacgctgtcagaggccctgctgcagc2640tgcagtttgatgatgaagacctgggggccttgcttggcaacagcacagacccagctgtgt2700tcacagacctggcatccgtcgacaactccgagtttcagcagctgctgaaccagggcatac2760ctgtggccccccacacaactgagcccatgctgatggagtaccctgaggctataactcgcc2820tagtgacaggggcccagaggccccccgacccagctcctgctccactgggggccccggggc2880tccccaatggcctcctttcaggagatgaagacttctcctccattgcggacatggacttct2940cagccctgctgagtcagatcagctccactagttattaagaattcacgcgtcgagcatgca3000tctagggcggccaattccgcccctctcccccccccccctctccctcccccccccctaacg3060ttactggccgaagccgcttggaataaggccggtgtgcgtttgtctatatgttattttcca3120ccatattgccgtcttttggcaatgtgagggcccggaaacctggccctgtcttcttgacga3180gcattcctaggggtctttcccctctcgccaaaggaatgcaaggtctgttgaatgtcgtga3240aggaagcagttcctctggaagcttcttgaagacaaacaacgtctgtagcgaccctttgca3300ggcagcggaaccccccacctggcgacaggtgcctctgcggccaaaagccacgtgtataag3360atacacctgcaaaggcggcacaaccccagtgccacgttgtgagttggatagttgtggaaa3420gagtcaaatggctctcctcaagcgtattcaacaaggggctgaaggatgcccagaaggtac3480cccattgtatgggatctgatctggggcctcggtgcacatgctttacatgtgtttagtcga3540ggttaaaaaaacgtctaggccccccgaaccacggggacgtggttttcctttgaaaaacac3600gatgataagcttgccacaacccgggatcctctagagtcgacatggactatcctgctgcca3660agagggtcaagttggactctagagaacgcccatatgcttgccctgtcgagtcctgcgatc3720gccgcttttctcgctcggatgagcttacccgccatatccgcatccacacaggccagaagc3780ccttccagtgtcgaatctgcatgcgtaacttcagtcgtagtgaccaccttaccacccaca3840tccgcacccacacaggcggcggccgcaggaggaagaaacgcaccagcatagagaccaaca3900tccgtgtggccttagagaagagtttcttggagaatcaaaagcctacctcggaagagatca3960ctatgattgctgatcagctcaatatggaaaaagaggtgattcgtgtttggttctgtaacc4020gccgccagaaagaaaaaagaatcaacactagaggagtgcaggtggaaaccatctccccgg4080gagacgggcgcaccttccccaagcgcggccagacctgcgtggtgcactacaccgggatgc4140ttgaagatggaaagaaatttgattcctcccgggacagaaacaagccctttaagtttatgc4200taggcaagcaggaggtgatccgaggctgggaagaaggggttgcccagatgagtgtgggtc4260agagagccaaactgactatatctccagattatgcctatggtgccactgggcacccaggca4320tcatcccaccacatgccactctcgtcttcgatgtggagcttctaaaactggaagtcgagg4380gcgtgcaggtggaaaccatctccccaggagacgggcgcaccttccccaagcgcggccaga4440cctgcgtggtgcactacaccgggatgcttgaagatggaaagaaatttgattcctcccggg4500acagaaacaagccctttaagtttatgctaggcaagcaggaggtgatccgaggctgggaag4560aaggggttgcccagatgagtgtgggtcagagagccaaactgactatatctccagattatg4620cctatggtgccactgggcacccaggcatcatcccaccacatgccactctcgtcttcgatg4680tggagcttctaaaactggaaactagaggagtgcaggtggaaaccatctccccaggagacg4740ggcgcaccttccccaagcgcggccagacctgcgtggtgcactacaccgggatgcttgaag4800atggaaagaaatttgattcctcccgggacagaaacaagccctttaagtttatgctaggca4860agcaggaggtgatccgaggctgggaagaaggggttgcccagatgagtgtgggtcagagag4920ccaaactgactatatctccagattatgcctatggtgccactgggcacccaggcatcatcc4980caccacatgccactctcgtcttcgatgtggagcttctaaaactggaaactagttattaag5040tcgacccgggcggccgcttccctttagtgagggttaatgcttcgagcagacatgataaga5100tacattgatgagtttggacaaaccacaactagaatgcagtgaaaaaaatgctttatttgt5160gaaatttgtgatgctattgctttatttgtaaccattataagctgcaataaacaagttaac5220ctccatagaagacaccgggaccgatccaataacttcgtatagcatacattatacgaagtt5280atggctgctagtgaacyaatcwcatgatgtcacccagacatcaggcatacccactagtgt5340gaaatagacatcagaattaagaaaaatgggctccccgggcgcgtactccacctcacccat5400catccacgctcggcaataaaaagacagaataaaacgcacgggtgttgggtcgtttgttcg5460ttttatggatagcactgagaacgtcatcaagcccttcatgcgcttcaaggtgcacatgga5520gggctccgtgaacggccacgagttcgagatcgagggcgagggcgagggcaagccctacga5580gggcacccagaccgccaagctgcaggtgaccaagggcggccccctgcccttcgcctggga5640catcctgtccccccagttccagtacggctccaaggtgtacgtgaagcaccccgccgacat5700ccccgactacaagaagctgtccttccccgagggcttcaagtgggagcgcgtgatgaactt5760cgaggacggcggcgtggtgaccgtgacccaggactcctccctgcaggacggcaccttcat5820ctaccacgtgaagttcatcggcgtgaacttcccctccgacggccccgtaatgcagaagaa5880gactctgggctgggagccctccaccgagcgcctgtacccccgcgacggcgtgctgaaggg5940cgagatccacaaggcgctgaagctgaagggcggcggccactacctggtggagttcaagtc6000aatctacatggccaagaagcccgtgaagctgcccggctactactacgtggactccaagct6060ggacatcacctcccacaacgaggactacaccgtggtggagcagtacgagcgcgccgaggc6120ccgccaccacctgttccagtaggacctccatagaagacaccgaataaaatatctttattt6180tcattacatctgtgtgttggttttttgtgtgaatcgatagtactaacatacgctctccat6240caaaacaaaacgaaacaaaacaaactagcaaaataggctgtccccagtgcaagtgcaggt6300gccagaacatttctctggaccgatccaataacttcgtatagcatacattatacgaagtta6360tgcctccggactctagcgttttagttattactagcgctaccggactcagatctcgagctc6420aagcttcgaattctgcagtcgacggtaccgcggcttacgcgtgctagctaatgatgggcg6480ctcgagtaatgatgggcggtcgactaatgatgggcgctcgagtaatgatgggcgtctagc6540taatgatgggcgctcgagtaatgatgggcggtcgactaatgatgggcgctcgagtaatga6600tgggcgtctagctaatgatgggcgctcgagtaatgatgggcggtcgactaatgatgggcg6660ctcgagtaatgatgggcgtctagaacgcgaattaattcaacattttgacacccccataat6720atttttccagaattaacagtataaattgcatctcttgttcaagagttccctatcactctc6780tttaatcactactcacagtaacctcaactcctgccacaagcttgccctgcagcgggaatt6840ccaaacttaagcttggtaccgagctcggatccactagtccagtgtggtggaattctgcag6900atatccagcacagtggcggccgctcgagtctagagggcccgtttaaacccgctgatcaga6960tggcaggaagaagcggagacagcgacgaagacctcctcaaggcagtcagactcatcaagt7020ttctctatcaaagcaacccacctcccaaccccgaggggacccgacaggcccgaaggaata7080gaagaagaaggtggagagagagacagagacagatccattcgattagtgaacggatcctta7140gcacttatttgggacgatctgcggacgctgtgcctcttcagctaccaccgcttgagagac7200ttactcttgattgtgacgaggattgtggaacttctgggacgcagggggtgggaagccctc7260aaatattggtggaatctcctacaatattggagtcaggagctaaagaatagtccctccccc7320ccccctaacgttactggccgaagccgcttggaataaggccggtgtgcgtttgtctatatg7380ttattttccaccatattgccgtcttttggcaatgtgagggcccggaaacctggccctgtc7440ttcttgacgagcattcctaggggtctttcccctctcgccaaaggaatgcaaggtctgttg7500aatgtcgtgaaggaagcagttcctctggaagcttcttgaagacaaacaacgtctgtagcg7560accctttgcaggcagcggaaccccccacctggcgacaggtgcctctgcggccaaaagcca7620cgtgtataagatacacctgcaaaggcggcacaaccccagtgccacgttgtgagttggata7680gttgtggaaagagtcaaatggctctcctcaagcgtattcaacaaggggctgaaggatgcc7740cagaaggtaccccattgtatgggatctgatctggggcctcggtgcacatgctttacatgt7800gtttagtcgaggttaaaaaacgtctaggccccccgaaccacggggacgtggttttccttt7860gaaaaacacgatgataatggggaggaggaagaggaagccgaaggatccgaaggcgagggt7920gttggcggaggcggattacaaggacgacgatgacaagatgtcagatcccagggagagaat7980cccacctggaaacagtggagaagagacaataggagaggccttcgaatggctaaacagaac8040agtagaggagataaacagagaggcagtaaaccacctaccaagggagctgattttccaggt8100ttggcaaaggtcttgggaatactggcatgatgaacaagggatgtcacaaagctatgtaaa8160atacagatacttgtgtttaatgcaaaaggctttatttatgcattgcaagaaaggctgtag8220atgtctaggggaaggacacggggcaggaggatggagaccaggacctcctcctcctccccc8280tccaggactagcataacctcgactgtgccttctagttgccagccatctgttgtttgcccc8340tcccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctaataaaat8400gaggaaattgcatcgcattgtctgagtaggtgtcattctattctggggggtggggtgggg8460caggacagcaagggggaggattgggaagacaatagcaggcatgctggggatgcggtgggc8520tctatggcttctgaggcggaaagttaaccctagaaagataatcatattgtgacgtacgtt8580aaagataatcatgcgtaaaattgacgcatgtgttttatcggtctgtatatcgaggtttat8640ttattaatttgaatagatattaagttttattatatttacacttacatactaataataaat8700tcaacaaacaatttatttatgtttatttatttattaaaaaaaaacaaaaactcaaaattt8760cttctataaagtaacaaaaaccagctggggctcgaagttcctatactttctagagaatag8820gaacttctatagtgagtcgaataagggcgacacaaaatttattctaaatgcataataaat8880actgataacatcttatagtttgtattatattttgtattatcgttgacatgtataattttg8940atatcaaaaactgattttccctttattattttcgagatttattttcttaattctctttaa9000caaactagaaatattgtatatacaaaaaatcataaataatagatgaatagtttaattata9060ggtgttcatcaatcgaaaaagcaacgtatcttatttaaagtgcgttgcttttttctcatt9120tataaggttaaataattctcatatatcaagcaaagtgacaggcgcccttaaatattctga9180caaatgctctttccctaaactccccccataaaaaaacccgccgaagcgggtttttacgtt9240atttgcggattaacgattactcgttatcagaaccgcccagggggcccgagcttaagactg9300gccgtcgttttacaacacagaaagagtttgtagaaacgcaaaaaggccatccgtcagggg9360ccttctgcttagtttgatgcctggcagttccctactctcgccttccgcttcctcgctcac9420tgactcgctgcgctcggtcgttcggctgcggcgagcggtatcagctcactcaaaggcggt9480aatacggttatccacagaatcaggggataacgcaggaaagaacatgtgagcaaaaggcca9540gcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgccc9600ccctgacgagcatcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggact9660ataaagataccaggcgtttccccctggaagctccctcgtgcgctctcctgttccgaccct9720gccgcttaccggatacctgtccgcctttctcccttcgggaagcgtggcgctttctcatag9780ctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgca9840cgaaccccccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaa9900cccggtaagacacgacttatcgccactggcagcagccactggtaacaggattagcagagc9960gaggtatgtaggcggtgctacagagttcttgaagtggtgggctaactacggctacactag10020aagaacagtatttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttgg10080tagctcttgatccggcaaacaaaccaccgctggtagcggtggtttttttgtttgcaagca10140gcagattacgcgcagaaaaaaaggatctcaagaagatcctttgatcttttctacggggtc10200tgacgctcagtggaacgacgcgcgcgtaactcacgttaagggattttggtcatgagcttg10260cgccgtcccgtcaagtcagcgtaatgctctgcttttagaaaaactcatcgagcatcaaat10320gaaactgcaatttattcatatcaggattatcaataccatatttttgaaaaagccgtttct10380gtaatgaaggagaaaactcaccgaggcagttccataggatggcaagatcctggtatcggt10440ctgcgattccgactcgtccaacatcaatacaacctattaatttcccctcgtcaaaaataa10500ggttatcaagtgagaaatcaccatgagtgacgactgaatccggtgagaatggcaaaagtt10560tatgcatttctttccagacttgttcaacaggccagccattacgctcgtcatcaaaatcac10620tcgcatcaaccaaaccgttattcattcgtgattgcgcctgagcgaggcgaaatacgcgat10680cgctgttaaaaggacaattacaaacaggaatcgagtgcaaccggcgcaggaacactgcca10740gcgcatcaacaatattttcacctgaatcaggatattcttctaatacctggaacgctgttt10800ttccggggatcgcagtggtgagtaaccatgcatcatcaggagtacggataaaatgcttga10860tggtcggaagtggcataaattccgtcagccagtttagtctgaccatctcatctgtaacat10920cattggcaacgctacctttgccatgtttcagaaacaactctggcgcatcgggcttcccat10980acaagcgatagattgtcgcacctgattgcccgacattatcgcgagcccatttatacccat11040ataaatcagcatccatgttggaatttaatcgcggcctcgacgtttcccgttgaatatggc11100tcatattcttcctttttcaatattattgaagcatttatcagggttattgtctcatgagcg11160gatacatatttgaatgtatttagaaaaataaacaaataggggtcagtgttacaaccaatt11220aaccaattctgaacattatcgcgagcccatttatacctgaatatggctcataacacccct11280tgtttgcctggcggcagtagcgcggtggtcccacctgaccccatgccgaactcagaagtg11340aaacgccgtagcgccgatggtagtgtggggactccccatgcgagagtagggaactgccag11400gcatcaaataaaacgaaaggctcagtcgaaagactgggcctttcgcccgggctaattagg11460gggtgtcgcccttattcgactctatagtgaagttcctattctctagaaagtataggaact11520tctgaagt11528<210>87<211>76<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>87gggcggcacuuauacagcgaagcauaauggcuacugacgcccucaaacccuauuugcaga60cuauaagugucgcgcg76<210>88<211>73<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>88gggcggcacuuauacaggguagcauaauggcuuaggacgccuucaaaccuaucaagacua60uaagugucgcgcg73<210>89<211>75<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>89gggcggcacuuauacaggguagcauaaugggcuacuugacgccuucaccuauuuguagac60uauaagugucgcgcg75<210>90<211>76<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>90gggcggcacuuauacagcguagcauaaugggcugcagacgccgucaaaccuauuugcaga60cuauaagugucgcgcg76<210>91<211>74<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>91gggcggcacuuauacaccguagcauaaugggcuacugccgccgucgaccuuuuggagacu60auaagugucgcgcg74<210>92<211>75<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>92gggcggcacuuauacaggucagcauaaugugcuagugcgccuucaaaccuauuuagagac60uauaagugucgcgcg75<210>93<211>76<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>93gggcggcacuuauacagcuuagcguaauggcuacugacgccguccaaaccuauuuacaga60cuauaagugucgcgcg76<210>94<211>76<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>94gggcggcacuuauacaggggagcauaaugggcuacugacgccuuuaaaccuauuugagga60cuauaagugucgcgcg76<210>95<211>74<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>95gggcggcacuuauacauggaagcauaaugggcugccgacggcccuuaaccuuuggagacu60auaagugucgcgcg74<210>96<211>79<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>96gggcggcacuuauacagauuagcauaaugggcuacugaccccgccggcaaaccuauuuga60agacuauaagugucgcgcg79<210>97<211>75<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>97gggcggcacuuauacaguguagcauaaugggcuacugucgcaucaaaccuauuuggagac60uauaagugucgcgcg75<210>98<211>76<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>98gggcggcacuuauacagugaagcauaaugggcuacugacacccuuaaaccuauuugcaga60cuauaagugucgcgcg76<210>99<211>76<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>99gggcggcacuuauacagaguagcauaaugggcuacagacgccgucaaaccuauuuaccga60cuauaagugucgcgcg76<210>100<211>75<212>rna<213>artificialsequence<220><223>syntheticevolvedaptamer<400>100gggcggcacuuauacagggugcauaaugggcuagugacgccuucaaaccuauuuguagac60uauaagugucgcgcg75<210>101<211>461<212>prt<213>homosapiens<400>101metthrileleuglythrthrpheglymetvalpheserleuleugln151015valvalserglygluserglytyralaglnasnglyaspleugluasp202530alagluleuaspasptyrserphesercystyrserglnleugluval354045asnglyserglnhisserleuthrcysalaphegluaspproaspval505560asnthrthrasnleugluphegluilecysglyalaleuvalgluval65707580lyscysleuasnphearglysleuglngluiletyrpheilegluthr859095lyslyspheleuleuileglylysserasnilecysvallysvalgly100105110glulysserleuthrcyslyslysileaspleuthrthrilevallys115120125proglualapropheaspleuservaliletyrarggluglyalaasn130135140aspphevalvalthrpheasnthrserhisleuglnlyslystyrval145150155160lysvalleumethisaspvalalatyrargglnglulysaspgluasn165170175lystrpthrhisvalasnleuserserthrlysleuthrleuleugln180185190arglysleuglnproalaalamettyrgluilelysvalargserile195200205proasphistyrphelysglyphetrpserglutrpserprosertyr210215220tyrpheargthrprogluileasnasnserserglyglumetasppro225230235240ileleuleuthrileserlyscyshisleuserphepheservalala245250255leuleuvalileleualacysvalleutrplyslysargilelyspro260265270ilevaltrpproserleuproasphislyslysthrleugluhisleu275280285cyslyslysproarglysasnleuasnvalserpheasnprogluser290295300pheleuaspcysglnilehisargvalaspaspileglnalaargasp305310315320gluvalgluglypheleuglnaspthrpheproglnglnleugluglu325330335serglulysglnargleuglyglyaspvalglnserproasncyspro340345350sergluaspvalvalvalthrprogluserpheglyargaspserser355360365leuthrcysleualaglyasnvalseralacysaspalaproileleu370375380serserserargserleuaspcysarggluserglylysasnglypro385390395400hisvaltyrglnaspleuleuleuserleuglythrthrasnserthr405410415leupropropropheserleuglnserglyileleuthrleuasnpro420425430valalaglnglyglnproileleuthrserleuglyserasnglnglu435440445glualatyrvalthrmetserserphetyrglnasngln450455460<210>102<211>463<212>prt<213>homosapiens<400>102metthrileleuglythrthrpheglymetvalpheserleuleugln151015valvalserglygluserglytyralaglnasnglyaspleugluasp202530alagluleuaspasptyrserphesercystyrserglnleugluval354045asnglyserglnhisserleuthrcysalaphegluaspproaspval505560asnthrthrasnleugluphegluilecysglyalaleuvalgluval65707580lyscysleuasnphearglysleuglngluiletyrpheilegluthr859095lyslyspheleuleuileglylysserasnilecysvallysvalgly100105110glulysserleuthrcyslyslysileaspleuthrthrilevallys115120125proglualapropheaspleuservaliletyrarggluglyalaasn130135140aspphevalvalthrpheasnthrserhisleuglnlyslystyrval145150155160lysvalleumethisaspvalalatyrargglnglulysaspgluasn165170175lystrpthrhisvalasnleuserserthrlysleuthrleuleugln180185190arglysleuglnproalaalamettyrgluilelysvalargserile195200205proasphistyrphelysglyphetrpserglutrpserprosertyr210215220tyrpheargthrprogluileasnasnserserglyglumetasppro225230235240ilephesercysglyproleuthrileserileleuserphepheser245250255valalaleuleuvalileleualacysvalleutrplyslysargile260265270lysproilevaltrpproserleuproasphislyslysthrleuglu275280285hisleucyslyslysproarglysasnleuasnvalserpheasnpro290295300gluserpheleuaspcysglnilehisargvalaspaspileglnala305310315320argaspgluvalgluglypheleuglnaspthrpheproglnglnleu325330335glugluserglulysglnargleuglyglyaspvalglnserproasn340345350cysprosergluaspvalvalvalthrprogluserpheglyargasp355360365serserleuthrcysleualaglyasnvalseralacysaspalapro370375380ileleuserserserargserleuaspcysarggluserglylysasn385390395400glyprohisvaltyrglnaspleuleuleuserleuglythrthrasn405410415serthrleupropropropheserleuglnserglyileleuthrleu420425430asnprovalalaglnglyglnproileleuthrserleuglyserasn435440445glngluglualatyrvalthrmetserserphetyrglnasngln450455460<210>103<211>462<212>prt<213>homosapiens<400>103metthrileleuglythrthrpheglymetvalpheserleuleugln151015valvalserglygluserglytyralaglnasnglyaspleugluasp202530alagluleuaspasptyrserphesercystyrserglnleugluval354045asnglyserglnhisserleuthrcysalaphegluaspproaspval505560asnthrthrasnleugluphegluilecysglyalaleuvalgluval65707580lyscysleuasnphearglysleuglngluiletyrpheilegluthr859095lyslyspheleuleuileglylysserasnilecysvallysvalgly100105110glulysserleuthrcyslyslysileaspleuthrthrilevallys115120125proglualapropheaspleuservaliletyrarggluglyalaasn130135140aspphevalvalthrpheasnthrserhisleuglnlyslystyrval145150155160lysvalleumethisaspvalalatyrargglnglulysaspgluasn165170175lystrpthrhisvalasnleuserserthrlysleuthrleuleugln180185190arglysleuglnproalaalamettyrgluilelysvalargserile195200205proasphistyrphelysglyphetrpserglutrpserprosertyr210215220tyrpheargthrprogluileasnasnserserglyglumetasppro225230235240ileleuleuthrcyshisleuileserileleuserphepheserval245250255alaleuleuvalileleualacysvalleutrplyslysargilelys260265270proilevaltrpproserleuproasphislyslysthrleugluhis275280285leucyslyslysproarglysasnleuasnvalserpheasnproglu290295300serpheleuaspcysglnilehisargvalaspaspileglnalaarg305310315320aspgluvalgluglypheleuglnaspthrpheproglnglnleuglu325330335gluserglulysglnargleuglyglyaspvalglnserproasncys340345350prosergluaspvalvalvalthrprogluserpheglyargaspser355360365serleuthrcysleualaglyasnvalseralacysaspalaproile370375380leuserserserargserleuaspcysarggluserglylysasngly385390395400prohisvaltyrglnaspleuleuleuserleuglythrthrasnser405410415thrleupropropropheserleuglnserglyileleuthrleuasn420425430provalalaglnglyglnproileleuthrserleuglyserasngln435440445gluglualatyrvalthrmetserserphetyrglnasngln450455460<210>104<211>466<212>prt<213>homosapiens<400>104metthrileleuglythrthrpheglymetvalpheserleuleugln151015valvalserglygluserglytyralaglnasnglyaspleugluasp202530alagluleuaspasptyrserphesercystyrserglnleugluval354045asnglyserglnhisserleuthrcysalaphegluaspproaspval505560asnthrthrasnleugluphegluilecysglyalaleuvalgluval65707580lyscysleuasnphearglysleuglngluiletyrpheilegluthr859095lyslyspheleuleuileglylysserasnilecysvallysvalgly100105110glulysserleuthrcyslyslysileaspleuthrthrilevallys115120125proglualapropheaspleuservaliletyrarggluglyalaasn130135140aspphevalvalthrpheasnthrserhisleuglnlyslystyrval145150155160lysvalleumethisaspvalalatyrargglnglulysaspgluasn165170175lystrpthrhisvalasnleuserserthrlysleuthrleuleugln180185190arglysleuglnproalaalamettyrgluilelysvalargserile195200205proasphistyrphelysglyphetrpserglutrpserprosertyr210215220tyrpheargthrprogluileasnasnserserglyglumetasppro225230235240ileleuleuthrproprovalcysservalthrileserileleuser245250255phepheservalalaleuleuvalileleualacysvalleutrplys260265270lysargilelysproilevaltrpproserleuproasphislyslys275280285thrleugluhisleucyslyslysproarglysasnleuasnvalser290295300pheasnprogluserpheleuaspcysglnilehisargvalaspasp305310315320ileglnalaargaspgluvalgluglypheleuglnaspthrphepro325330335glnglnleuglugluserglulysglnargleuglyglyaspvalgln340345350serproasncysprosergluaspvalvalvalthrprogluserphe355360365glyargaspserserleuthrcysleualaglyasnvalseralacys370375380aspalaproileleuserserserargserleuaspcysarggluser385390395400glylysasnglyprohisvaltyrglnaspleuleuleuserleugly405410415thrthrasnserthrleupropropropheserleuglnserglyile420425430leuthrleuasnprovalalaglnglyglnproileleuthrserleu435440445glyserasnglngluglualatyrvalthrmetserserphetyrgln450455460asngln465<210>105<211>523<212>prt<213>artificialsequence<220><223>syntheticfc-delta-30<400>105metserilemetglyleulysvalasnvalseralailephemetala151015valleuleuthrleuglnthrprothrglyglnilehistrpglyasn202530leuserlysileglyvalvalglyileglyseralasertyrlysval354045metthrargserserhisglnserleuvalilelysleumetproasn505560ilethrleuleuasnasncysthrargvalgluilealaglutyrarg65707580argleuleuargthrvalleugluproileargaspalaleuasnala859095metthrglnasnileargprovalglnservalalaserserargarg100105110hislysargphealaglyvalvalleualaglyalaalaleuglyval115120125alathralaalaglnilethralaglyilealaleuhisglnsermet130135140leuasnserglnalaileaspasnleuargalaserleugluthrthr145150155160asnglnalaileglualaileargglnalaglyglnglumetileleu165170175alavalglnglyvalglnasptyrileasnasngluleuileproser180185190metasnglnleusercysaspleuileglyglnlysleuglyleulys195200205leuleuargtyrtyrthrgluileleuserleupheglyproserleu210215220argaspproileseralagluileserileglnalaleusertyrala225230235240leuglyglyaspileasnlysvalleuglulysleuglytyrsergly245250255glyaspleuleuglyileleugluserargglyilelysalaargile260265270thrhisvalaspthrglusertyrpheilevalleuserilealatyr275280285prothrleusergluilelysglyvalilevalhisargleuglugly290295300valsertyrasnileglyserglnglutrptyrthrthrvalprolys305310315320tyrvalalathrglnglytyrleuileserasnpheaspgluserser325330335cysthrphemetprogluglythrvalcysserglnasnalaleutyr340345350prometserproleuleuglnglucysleuargglyserthrlysser355360365cysalaargthrleuvalserglyserpheglyasnargpheileleu370375380serglnglyasnleuilealaasncysalaserileleucyslyscys385390395400tyrthrthrglythrileileasnglnaspproasplysileleuthr405410415tyrilealaalaasphiscysprovalvalgluvalasnglyvalthr420425430ileglnvalglyserargargtyrproaspalavaltyrleuhisarg435440445ileaspleuglyproproileserleugluargleuaspvalglythr450455460asnleuglyasnalailealalysleugluaspalalysgluleuleu465470475480gluserseraspglnileleuargsermetlysglyleuserserthr485490495serilevaltyrileleuilealavalcysleuglyglyleuilegly500505510ileproalaleuilecyscyscysargglyarg515520<210>106<211>599<212>prt<213>artificialsequence<220><223>synthetichc-delta-18<400>106metglyserargilevalileasnarggluhisleumetileasparg151015protyrvalleuleualavalleuphevalmetserleuserleuile202530glyleuleualailealaglyileargleuhisargalaalailetyr354045thralagluilehislysserleuserthrasnleuaspvalthrasn505560serilegluhisglnvallysaspvalleuthrproleuphelysile65707580ileglyaspgluvalglyleuargthrproglnargphethraspleu859095vallyspheileserasplysilelyspheleuasnproaspargglu100105110tyrasppheargaspleuthrtrpcysileasnproprogluargile115120125lysleuasptyraspglntyrcysalaaspvalalaalaglugluleu130135140metasnalaleuvalasnserthrleuleugluthrargthrthrasn145150155160glnpheleualavalserlysglyasncysserglyprothrthrile165170175argglyglnpheserasnmetserleuserleuleuaspleutyrleu180185190serargglytyrasnvalserserilevalthrmetthrserglngly195200205mettyrglyglythrtyrleuvalglulysproasnleuserserlys210215220argsergluleuserglnleusermettyrargvalphegluvalgly225230235240valileargasnproglyleuglyalaprovalphehismetthrasn245250255tyrleugluglnprovalserasnaspleuserasncysmetvalala260265270leuglygluleulysleualaalaleucyshisglygluaspserile275280285thrileprotyrglnglyserglylysglyvalserpheglnleuval290295300lysleuglyvaltrplysserprothraspmetglnsertrpvalpro305310315320leuserthraspaspprovalileaspargleutyrleuserserhis325330335argglyvalilealaaspasnglnalalystrpalavalprothrthr340345350argthraspasplysleuargmetgluthrcyspheglnglnalacys355360365lysglylysileglnalaleucysgluasnproglutrpalaproleu370375380lysaspasnargileprosertyrglyvalleuservalaspleuser385390395400leuthrvalgluleulysilelysilealaserglypheglyproleu405410415ilethrhisglyserglymetaspleutyrlysserasnhisasnasn420425430valtyrtrpleuthrileproprometlysasnleualaleuglyval435440445ileasnthrleuglutrpileproargphelysvalserproasnleu450455460phethrvalproilelysglualaglygluaspcyshisalaprothr465470475480tyrleuproalagluvalaspglyaspvallysleuserserasnleu485490495valileleuproglyglnaspleuglntyrvalleualathrtyrasp500505510thrserargvalgluhisalavalvaltyrtyrvaltyrserprogly515520525argserphesertyrphetyrpropheargleuproilelysglyval530535540proilegluleuglnvalglucysphethrtrpaspglnlysleutrp545550555560cysarghisphecysvalleualaaspsergluserglyglyhisile565570575thrhisserglymetvalglymetglyvalsercysthrvalthrarg580585590gluaspglythrasnargarg595<210>107<211>529<212>prt<213>artificialsequence<220><223>syntheticdafss-il7-daffusion<400>107metthrvalalaargproservalproalaalaleuproleuleugly151015gluleuproargleuleuleuleuvalleuleucysleuproalaasp202530cysaspilegluglylysaspglylysglntyrgluservalleumet354045valserileaspglnleuleuaspsermetlysgluileglyserasn505560cysleuasnasnglupheasnphephelysarghisilecysaspala65707580asnlysgluglymetpheleupheargalaalaarglysleuarggln859095pheleulysmetasnserthrglyasppheaspleuhisleuleulys100105110valsergluglythrthrileleuleuasncysthrglyglnvallys115120125glyarglysproalaalaleuglyglualaglnprothrlysserleu130135140glugluasnlysserleulysgluglnlyslysleuasnaspleucys145150155160pheleulysargleuleuglngluilelysthrcystrpasnlysile165170175leumetglythrlysgluhiscysglyleuproproaspvalproasn180185190alaglnproalaleugluglyargthrserpheprogluaspthrval195200205ilethrtyrlyscysglugluserphevallysileproglyglulys210215220aspservalilecysleulysglyserglntrpseraspilegluglu225230235240phecysasnargsercysgluvalprothrargleuasnseralaser245250255leulysglnprotyrilethrglnasntyrpheprovalglythrval260265270valglutyrglucysargproglytyrargarggluproserleuser275280285prolysleuthrcysleuglnasnleulystrpserthralavalglu290295300phecyslyslyslyssercysproasnproglygluileargasngly305310315320glnileaspvalproglyglyileleupheglyalathrileserphe325330335sercysasnthrglytyrlysleupheglyserthrserserphecys340345350leuileserglyserservalglntrpseraspproleuproglucys355360365arggluiletyrcysproalaproproglnileaspasnglyileile370375380glnglygluargasphistyrglytyrargglnservalthrtyrala385390395400cysasnlysglyphethrmetileglygluhisseriletyrcysthr405410415valasnasnaspgluglyglutrpserglyproproproglucysarg420425430glylysserleuthrserlysvalproprothrvalglnlysprothr435440445thrvalasnvalprothrthrgluvalserprothrserglnlysthr450455460thrthrlysthrthrthrproasnalaglnalathrargserthrpro465470475480valserargthrthrlyshisphehisgluthrthrproasnlysgly485490495serglythrthrserglythrthrargleuleuserglyhisthrcys500505510phethrleuthrglyleuleuglythrleuvalthrmetglyleuleu515520525thr<210>108<211>49<212>dna<213>artificialsequence<220><223>synthetic582-1<400>108acagcttagcgtaatggctactgacgccgtccaaacctatttacagact49<210>109<211>49<212>dna<213>artificialsequence<220><223>synthetic582-2<400>109acagcttaggataatggctactgacgccgtccaaacctatttacagact49<210>110<211>49<212>dna<213>artificialsequence<220><223>synthetic582-3<400>110acagcttagcataatggctactgacgccgtccaaacctattcacagact49<210>111<211>49<212>dna<213>artificialsequence<220><223>synthetic582-4<400>111acagcttagcataatggctactgacgccgtccaaacctattgacagact49<210>112<211>49<212>dna<213>artificialsequence<220><223>synthetic582-5<400>112acagcatagcataatggctactgacgccgtccaaacctatttacagact49<210>113<211>49<212>dna<213>artificialsequence<220><223>synthetic582-6<400>113acagcttagcataatggctactgacgccgtccaaacctatgtacagact49<210>114<211>48<212>dna<213>artificialsequence<220><223>synthetic582-7<400>114acagctagcgtaatggctactgacgccgtccaaacctatttacagact48<210>115<211>49<212>dna<213>artificialsequence<220><223>synthetic582-8<400>115acagcttagcattatggctactgacgccgtccaaacctatttacagact49<210>116<211>48<212>dna<213>artificialsequence<220><223>synthetic582-9<400>116acagttagcataatggctactgacgccgtccaaacctatttacagact48<210>117<211>49<212>dna<213>artificialsequence<220><223>synthetic582-10<400>117acagcttagcataatggctactgacgcggtccaaacctatttacagact49<210>118<211>49<212>dna<213>artificialsequence<220><223>synthetic582-11<400>118acagcttagcttaatggctactgacgccgtccaaacctatttacagact49<210>119<211>49<212>dna<213>artificialsequence<220><223>synthetic582-12<400>119acagcttagcataatggctactgacgccgtccaaacccatttacagact49<210>120<211>49<212>dna<213>artificialsequence<220><223>synthetic582-13<400>120acagcttagcataatggctactgacgccgtccaaaccaatttacagact49<210>121<211>49<212>dna<213>artificialsequence<220><223>synthetic582-14<400>121acagcttagcataatggatactgacgccgtccaaacctatttacagact49<210>122<211>49<212>dna<213>artificialsequence<220><223>synthetic582-15<400>122acagcttagcattgtggctactgacgccgtccaaacctatttacagact49<210>123<211>49<212>dna<213>artificialsequence<220><223>synthetic582-16<400>123acaggttagcataatggctaccgacgccgtccaaacctatttacagact49<210>124<211>49<212>dna<213>artificialsequence<220><223>synthetic582-17<400>124acagcttagcgtaatggctactgacgccgcccaaacctatttacagact49<210>125<211>49<212>dna<213>artificialsequence<220><223>synthetic582-18<400>125acagcttagcataatggctactgacgccgtccaaaactatttccagact49<210>126<211>49<212>dna<213>artificialsequence<220><223>synthetic582-19<400>126acagcctagcataagggctactgacgccgtccaaacctatttacagact49<210>127<211>49<212>dna<213>artificialsequence<220><223>synthetic582-20<400>127acagcttagcataatggctactgaggccgtccaaacctatttacagact49<210>128<211>49<212>dna<213>artificialsequence<220><223>synthetic582-21<400>128acagcttaccttaatggctactgacgccgtccaaacctatttacagact49<210>129<211>49<212>dna<213>artificialsequence<220><223>synthetic582-22<400>129acagcttagcataatggctaccgacgctgtccaaacctatttacagact49<210>130<211>49<212>dna<213>artificialsequence<220><223>synthetic582-23<400>130acagcttagcgtaatggctactggcgccgtccaaacctatttacagact49<210>131<211>49<212>dna<213>artificialsequence<220><223>synthetic582-24<400>131acagcttagcatactggctactgacgccgcccaaacctatttacagact49<210>132<211>49<212>dna<213>artificialsequence<220><223>synthetic582-25<400>132acagcttagcataatggctactgacgccgtcctaacctatttacagact49<210>133<211>49<212>dna<213>artificialsequence<220><223>synthetic582-26<400>133acaggttagcataatgcctactgacgccgtccaaacctatttacagact49<210>134<211>49<212>dna<213>artificialsequence<220><223>synthetic582-27<400>134acagcttagcataattgctactgacgccgttcaaacctatttacagact49<210>135<211>49<212>dna<213>artificialsequence<220><223>synthetic582-28<400>135acagcttagcataaaggctactgacgccgtccaaacctatttacagact49<210>136<211>49<212>dna<213>artificialsequence<220><223>synthetic582-29<400>136acagcttagcgtaatggctactgacgccgtctaaacctatttccagact49<210>137<211>49<212>dna<213>artificialsequence<220><223>synthetic582-30<400>137acaggttagcataatggctactgacgccgtccaaacctatttagagact49<210>138<211>49<212>dna<213>artificialsequence<220><223>synthetic582-31<400>138acagggtagcgtaatggctactgacgccgtccaaacctatttacagact49<210>139<211>49<212>dna<213>artificialsequence<220><223>synthetic582-32<400>139acagcgtagcataatggctactgacgccgttcaaacctatttacagact49<210>140<211>49<212>dna<213>artificialsequence<220><223>synthetic582-33<400>140acagcttagcataatggctactgacgccgtccaaactcatttacagact49<210>141<211>49<212>dna<213>artificialsequence<220><223>synthetic582-34<400>141acagcgtagcatagtggctactgacgccgtccaaacctatttacagact49<210>142<211>49<212>dna<213>artificialsequence<220><223>synthetic582-35<400>142acagcttagtgtaatggctactgacgctgtccaaacctatttacagact49<210>143<211>49<212>dna<213>artificialsequence<220><223>synthetic582-36<400>143acagcttagcataatggctactgacggcgttcaaacctatttacagact49<210>144<211>49<212>dna<213>artificialsequence<220><223>synthetic582-37<400>144acaggttagcataatggctactgacgccgtccaaacctatttatagact49<210>145<211>48<212>dna<213>artificialsequence<220><223>synthetic582-38<400>145acagcttagcataatggctactgacgccgtccaaacctattgtcgact48<210>146<211>48<212>dna<213>artificialsequence<220><223>synthetic582-39<400>146acagcttagcataatggctactgacgccgtccaaacctatttacgact48<210>147<211>48<212>dna<213>artificialsequence<220><223>synthetic769-1<400>147acaggtcagcataatgtgctagtgcgccttcaaacctatttagagact48<210>148<211>48<212>dna<213>artificialsequence<220><223>synthetic769-2<400>148acaggtcagcataatgtgctagtgcgccctcaaacctatttagagact48<210>149<211>48<212>dna<213>artificialsequence<220><223>synthetic769-3<400>149acaggttagcataatgtgctattgcgccttcaaacctatttagagact48<210>150<211>48<212>dna<213>artificialsequence<220><223>synthetic769-4<400>150acaggtcagcataatgtgctagtgcgcattcaaacctatttagagact48<210>151<211>48<212>dna<213>artificialsequence<220><223>synthetic769-5<400>151acaggttagcataatgtgctagtgcgccttcaaacctattttgagact48<210>152<211>48<212>dna<213>artificialsequence<220><223>synthetic769-6<400>152acaggttatcataatgtgctagtgcgccttcaaacctatttagagact48<210>153<211>48<212>dna<213>artificialsequence<220><223>synthetic769-7<400>153acaggttagcatgatgtgctagtgcgccttcaaacctatttagagact48<210>154<211>48<212>dna<213>artificialsequence<220><223>synthetic769-8<400>154acaggttagcataatgggctagtgcgccttcaaacctatttagagact48<210>155<211>48<212>dna<213>artificialsequence<220><223>synthetic769-9<400>155acaggtcagcaaaatgtgcaagtgcgccttcaaacctatttagagact48<210>156<211>48<212>dna<213>artificialsequence<220><223>synthetic769-10<400>156acaggtcagcataatgtgctagtgcgccttcaaacctatctggagact48<210>157<211>48<212>dna<213>artificialsequence<220><223>synthetic769-11<400>157acagcttagcataatgtgctagtgcgccttcaaacctatttagagact48<210>158<211>48<212>dna<213>artificialsequence<220><223>synthetic769-12<400>158acaggtcagcataatgtgctagtgcgccttcaaacctatttacagact48<210>159<211>48<212>dna<213>artificialsequence<220><223>synthetic769-13<400>159acaggtcagcataatgtgctagtgcgccttcaaacatatttagagact48<210>160<211>48<212>dna<213>artificialsequence<220><223>synthetic769-14<400>160acagggtagcataatgtgctagtgcgccttcaaacctatttagagact48<210>161<211>48<212>dna<213>artificialsequence<220><223>synthetic769-15<400>161acaggttagcataatgtgctagtgcgccctcaaacctatttagagact48<210>162<211>48<212>dna<213>artificialsequence<220><223>synthetic769-16<400>162acaggttagcataatgtgccagtgcgccttcaaacctatttagagact48<210>163<211>48<212>dna<213>artificialsequence<220><223>synthetic769-17<400>163acaggtcagcataatgggctagtgcgccttcaaacctatttagagact48<210>164<211>49<212>dna<213>artificialsequence<220><223>synthetic795-1<400>164acagcgaagcataatggctactgacgccctcaaaccctatttgcagact49<210>165<211>49<212>dna<213>artificialsequence<220><223>synthetic795-2<400>165acagcgaagcataatggctactgacgccctcaaaccctatttacagact49<210>166<211>49<212>dna<213>artificialsequence<220><223>synthetic795-3<400>166acagcgaagcataatggcttctgacgccctcaaaccctatttgcagact49<210>167<211>49<212>dna<213>artificialsequence<220><223>synthetic795-4<400>167acagccaagcatactggctactgacgccctcaaaccctatttgcagact49<210>168<211>49<212>dna<213>artificialsequence<220><223>synthetic795-5<400>168acagcgaagcataatggctactgacgcccgcaaaccctatttgcagact49<210>169<211>49<212>dna<213>artificialsequence<220><223>synthetic795-6<400>169acagcgaagcataatggctactgacggcctcaaaccctatttgcagact49<210>170<211>49<212>dna<213>artificialsequence<220><223>synthetic795-7<400>170acagcgaggcataatggctactgacgccctcaaaccctatttgcagact49<210>171<211>49<212>dna<213>artificialsequence<220><223>synthetic795-8<400>171acagcgaagcataatggctactgacgccttcaaaccctatttgcagact49<210>172<211>49<212>dna<213>artificialsequence<220><223>synthetic795-9<400>172acagcgaagcataatggctacagacgccctcaaaacctatttgcagact49<210>173<211>48<212>dna<213>artificialsequence<220><223>synthetic795-10<400>173acagcgaagcataatggctactgacgccctcaaaccctatttgagact48<210>174<211>48<212>dna<213>artificialsequence<220><223>synthetic795-11<400>174acagcgaagcataatggctactgacgccctcaaaccctattgtcgact48<210>175<211>49<212>dna<213>artificialsequence<220><223>synthetic795-12<400>175acagccaagcataatggctactgacgccctcaaaccctatttgcagact49<210>176<211>49<212>dna<213>artificialsequence<220><223>synthetic795-13<400>176acagcgaagcataatggctactgacgccctcaaaccctatttggcgact49<210>177<211>49<212>dna<213>artificialsequence<220><223>synthetic795-14<400>177acagcgaagcataatgtctactgacgccctcaaaccctatttgcagact49<210>178<211>49<212>dna<213>artificialsequence<220><223>synthetic795-15<400>178acagcgaagcataatggctactgacgccgtcaaaccctatttgtagact49<210>179<211>49<212>dna<213>artificialsequence<220><223>synthetic795-16<400>179acagcgaagcataatggctactgacgccctcaaaccttatttgcagact49<210>180<211>48<212>dna<213>artificialsequence<220><223>synthetic795-17<400>180acaggtagcataatggctactgacgccctcaaaccctatttgcagact48<210>181<211>49<212>dna<213>artificialsequence<220><223>synthetic795-18<400>181acagcgaagcataatggctactgacgccctcaaaccctatttctagact49<210>182<211>49<212>dna<213>artificialsequence<220><223>synthetic795-19<400>182acagcgaagcataatggctactgacgccctcaaaccctatttgtagact49<210>183<211>48<212>dna<213>artificialsequence<220><223>synthetic935-1<400>183acagggtagcataatgggctacttgacgccttcacctatttgtagact48<210>184<211>47<212>dna<213>artificialsequence<220><223>synthetic935-2<400>184acagggtagcataatgggctacttgacgccttcacctatttgagact47<210>185<211>48<212>dna<213>artificialsequence<220><223>synthetic935-3<400>185acagggtagcataatgggctactttacgccttcacctatttgtagact48<210>186<211>48<212>dna<213>artificialsequence<220><223>synthetic935-4<400>186acagggtagcataatgggctacttgacgccttcacctatttctagact48<210>187<211>48<212>dna<213>artificialsequence<220><223>synthetic935-5<400>187acagggtagcataatgggctacttgacgccttcacctatttggagact48<210>188<211>48<212>dna<213>artificialsequence<220><223>synthetic935-6<400>188acagggtagcatagtgggctacttgacgccttcacctatttgtagact48<210>189<211>48<212>dna<213>artificialsequence<220><223>synthetic935-7<400>189acagggtagcatgatgggctacttgacgccttcacctatttgtagact48<210>190<211>48<212>dna<213>artificialsequence<220><223>synthetic935-8<400>190acagggtagcataatgggctacttgacgccttcacctattagtagact48<210>191<211>48<212>dna<213>artificialsequence<220><223>synthetic935-9<400>191acagggtagcataatgggctatttgacgccttcacctatttgtagact48<210>192<211>48<212>dna<213>artificialsequence<220><223>synthetic935-10<400>192acagggtagcataatgggctacttgccgccttcacctatttgtagact48<210>193<211>48<212>dna<213>artificialsequence<220><223>synthetic935-11<400>193acagtgtagcataattggctacttgacgccttcacctatttgtagact48<210>194<211>48<212>dna<213>artificialsequence<220><223>synthetic935-12<400>194acagggtagcataatgggctacttgacgctttcacctttttgtagact48<210>195<211>48<212>dna<213>artificialsequence<220><223>synthetic935-13<400>195acagggtagcataaggggctacttgacgccttcacctatttgtagact48<210>196<211>48<212>dna<213>artificialsequence<220><223>synthetic935-14<400>196acagggtagcataatggactacttgacgcctccacctatttgtagact48<210>197<211>48<212>dna<213>artificialsequence<220><223>synthetic935-15<400>197acagggtagcataatgggctacttgtcgccttcacctatttgtagact48<210>198<211>49<212>dna<213>artificialsequence<220><223>synthetic946-1<400>198acagcgtagcataatgggctgcagacgccgtcaaacctatttgcagact49<210>199<211>49<212>dna<213>artificialsequence<220><223>synthetic946-2<400>199acagcgtagcataatgggctgcagacgcagtcaaacctatttgcagact49<210>200<211>48<212>dna<213>artificialsequence<220><223>synthetic946-3<400>200acatgtagcataatgggctactgacgccgtcaaacctatttgcagact48<210>201<211>49<212>dna<213>artificialsequence<220><223>synthetic946-4<400>201acagcgtagcatagtgggctgcagacgccgtcaaacctatttgcagact49<210>202<211>49<212>dna<213>artificialsequence<220><223>synthetic946-5<400>202acagtgtagcataatgggctgcagacgccttcaaacctatttggagact49<210>203<211>49<212>dna<213>artificialsequence<220><223>synthetic946-6<400>203acagtgtagcataatgggctgctgacgccgtcaaacctatttgaagact49<210>204<211>49<212>dna<213>artificialsequence<220><223>synthetic946-7<400>204acagcgtagcataatgggctacaggcgccgtcaaacctatttgcagact49<210>205<211>49<212>dna<213>artificialsequence<220><223>synthetic946-8<400>205acagcgtagcataatgggctactggcgccgtcaaacctatttgcagact49<210>206<211>48<212>dna<213>artificialsequence<220><223>synthetic946-9<400>206acagcgtagcataatgggctgcagacgccgtcaaacctatttgagact48<210>207<211>48<212>dna<213>artificialsequence<220><223>synthetic946-10<400>207acaggtagcataatgggctgcagacgccgtcaaacctatttgcagact48<210>208<211>48<212>dna<213>artificialsequence<220><223>synthetic946-11<400>208acaggtagcataatgggctgctgacgccgtcaaacctatttacagact48<210>209<211>49<212>dna<213>artificialsequence<220><223>synthetic946-12<400>209acagcgtagcatattgggctgcagacgccgtcaaacctatttgcagact49<210>210<211>49<212>dna<213>artificialsequence<220><223>synthetic946-13<400>210acagcgtagcataatgggctgcagacgccttcaaacctatttggagact49<210>211<211>48<212>dna<213>artificialsequence<220><223>synthetic946-14<400>211acagtgtagcataatgggctgcagacgccgtcaaacctatttgagact48<210>212<211>49<212>dna<213>artificialsequence<220><223>synthetic946-15<400>212acagcgtagcataatgggctgctgacgccgtcaaacctatttggagact49<210>213<211>49<212>dna<213>artificialsequence<220><223>synthetic946-16<400>213acagcgtagcataatgggctgcagacgccgtcaaacctatttacagact49<210>214<211>49<212>dna<213>artificialsequence<220><223>synthetic946-17<400>214acagcgtagcataatgggctgctgacgccgtcaaacctatttgcagact49<210>215<211>49<212>dna<213>artificialsequence<220><223>synthetic946-18<400>215acagggtagcataatgggctgcagacgccgtcaaacctatttggagact49<210>216<211>49<212>dna<213>artificialsequence<220><223>synthetic946-19<400>216acagcgtagcataatgggctacagacgccgtcaaacctatttgcagact49<210>217<211>49<212>dna<213>artificialsequence<220><223>synthetic946-20<400>217acagcgtcgcataatgggctgcagacgccgtcaaatctatttgcagact49<210>218<211>49<212>dna<213>artificialsequence<220><223>synthetic946-21<400>218acagcgtagcataatgggcttcagacgccgtcaaacctatttgcagact49<210>219<211>48<212>dna<213>artificialsequence<220><223>synthetic946-22<400>219acatgtagcataatgggctgcagacgccgtcaaacctatttggagact48<210>220<211>47<212>dna<213>artificialsequence<220><223>synthetic961-1<400>220acaccgtagcataatgggctactgccgccgtcgaccttttggagact47<210>221<211>46<212>dna<213>artificialsequence<220><223>synthetic996-1<400>221acagggtagcataatggcttaggacgccttcaaacctatcaagact46<210>222<211>49<212>dna<213>artificialsequence<220><223>syntheticfamily582consensus<220><221>misc_feature<222>(5)..(5)<223>natposition5canbec,g,ornonucleotide<220><221>misc_feature<222>(6)..(6)<223>natposition6canbea,c,g,t,ornonucleotide<220><221>misc_feature<222>(45)..(45)<223>natposition45canbeaornonucleotide<400>222acagnntasbdtwvdksmtacygrsgsbgyyywaamyhatkbhbngact49<210>223<211>48<212>dna<213>artificialsequence<220><223>syntheticfamily769consensus<400>223acagskyakcawratgkgchaktgcgcmytcaaacmtatytdsagact48<210>224<211>49<212>dna<213>artificialsequence<220><223>syntheticfamily795consensus<220><221>misc_feature<222>(5)..(5)<223>natposition5canbecornonucleotide<220><221>misc_feature<222>(40)..(40)<223>natposition40canbetornonucleotide<220><221>misc_feature<222>(44)..(44)<223>natposition44canbec,g,t,ornonucleotide<400>224acagnswrgcatamtgkctwcwgacgscbkcaaamcytanttvnmgact49<210>225<211>48<212>dna<213>artificialsequence<220><223>syntheticfamily935consensus<220><221>misc_feature<222>(43)..(43)<223>natposition43canbeg,t,ornonucleotide<400>225acagkgtcgcatrrkkgrctayttkhcgcytycacctwttwsnagact48<210>226<211>49<212>dna<213>artificialsequence<220><223>syntheticfamily946consensus<220><221>misc_feature<222>(4)..(4)<223>natposition4canbegornonucleotide<220><221>misc_feature<222>(5)..(5)<223>natposition5canbec,g,t,ornonucleotide<220><221>misc_feature<222>(44)..(44)<223>natposition44canbea,c,g,ornonucleotide<400>226acanngtmgcatadtgggctdcwgrcgcmktcaaayctatttrnagact49<210>227<211>11<212>prt<213>artificialsequence<220><223>syntheticmembrane-targetingdomainofsrc-flag-vpx<400>227metglyserserlysserlysprolysasppro1510<210>228<211>8<212>prt<213>artificialsequence<220><223>syntheticviralproteasecleavagedomain<400>228lysalaargvalleualagluala15<210>229<211>458<212>prt<213>homosapiens<400>229metthrileleuglythrthrpheglymetvalpheserleuleugln151015valvalserglygluserglytyralaglnasnglyaspleugluasp202530alagluleuaspasptyrserphesercystyrserglnleugluval354045asnglyserglnhisserleuthrcysalaphegluaspproaspval505560asnilethrasnleugluphegluilecysglyalaleuvalgluval65707580lyscysleuasnphearglysleuglngluiletyrpheilegluthr859095lyslyspheleuleuileglylysserasnilecysvallysvalgly100105110glulysserleuthrcyslyslysileaspleuthrthrilevallys115120125proglualapropheaspleuservalvaltyrarggluglyalaasn130135140aspphevalvalthrpheasnthrserhisleuglnlyslystyrval145150155160lysvalleumethisaspvalalatyrargglnglulysaspgluasn165170175lystrpthrhisvalasnleuserserthrlysleuthrleuleugln180185190arglysleuglnproalaalamettyrgluilelysvalargserile195200205proasphistyrphelysglyphetrpserglutrpserprosertyr210215220tyrpheargthrprogluileasnasnserserglyglumetasppro225230235240ileleuleuthrileserileleuserphepheservalalaleuleu245250255valileleualacysvalleutrplyslysargilelysproileval260265270trpproserleuproasphislyslysthrleugluhisleucyslys275280285lysproarglysasnleuasnvalserpheasnprogluserpheleu290295300aspcysglnilehisargvalaspaspileglnalaargaspgluval305310315320gluglypheleuglnaspthrpheproglnglnleuglugluserglu325330335lysglnargleuglyglyaspvalglnserproasncysproserglu340345350aspvalvalilethrprogluserpheglyargaspserserleuthr355360365cysleualaglyasnvalseralacysaspalaproileleuserser370375380serargserleuaspcysarggluserglylysasnglyprohisval385390395400tyrglnaspleuleuleuserleuglythrthrasnserthrleupro405410415propropheserleuglnserglyileleuthrleuasnprovalala420425430glnglyglnproileleuthrserleuglyserasnglnglugluala435440445tyrvalthrmetserserphetyrglnasn450455<210>230<211>723<212>prt<213>artificialsequence<220><223>syntheticgfp-linker-p2a-il7raincppclcodon(exceptp2a)andspliceoptimized<400>230metserglyglyglugluleuphealaglyilevalprovalleuile151015gluleuaspglyaspvalhisglyhislyspheservalargglyglu202530glygluglyaspalaasptyrglylysleugluilelyspheilecys354045thrthrglylysleuprovalprotrpprothrleuvalthrthrleu505560cystyrglyileglncysphealaargtyrprogluhismetlysmet65707580asnaspphephelysseralametprogluglytyrileglngluarg859095thrileglnpheglnaspaspglylystyrlysthrargglygluval100105110lysphegluglyaspthrleuvalasnargilegluleulysglylys115120125aspphelysgluaspglyasnileleuglyhislysleuglutyrser130135140pheasnserhisasnvaltyrileargproasplysalaasnasngly145150155160leuglualaasnphelysthrarghisasnilegluglyglyglyval165170175glnleualaasphistyrglnthrasnvalproleuglyaspglypro180185190valleuileproileasnhistyrleuserthrglnthrlysileser195200205lysaspargasnglualaargasphismetvalleuleugluserphe210215220seralacyscyshisthrhisglymetaspgluleutyrargglyser225230235240glyalathrasnpheserleuleulysglnalaglyaspvalgluglu245250255asnproglyprometthrileleuglythrthrpheglymetvalphe260265270serleuleuglnvalvalserglygluserglytyralaglnasngly275280285aspleugluaspalagluleuaspasptyrserphesercystyrser290295300glnleugluvalasnglyserglnhisserleuthrcysalapheglu305310315320aspproaspvalasnilethrasnleugluphegluilecysglyala325330335leuvalgluvallyscysleuasnphearglysleuglngluiletyr340345350pheilegluthrlyslyspheleuleuileglylysserasnilecys355360365vallysvalglyglulysserleuthrcyslyslysileaspleuthr370375380thrilevallysproglualapropheaspleuservalvaltyrarg385390395400gluglyalaasnaspphevalvalthrpheasnthrserhisleugln405410415lyslystyrvallysvalleumethisaspvalalatyrargglnglu420425430lysaspgluasnlystrpthrhisvalasnleuserserthrlysleu435440445thrleuleuglnarglysleuglnproalaalamettyrgluilelys450455460valargserileproasphistyrphelysglyphetrpserglutrp465470475480serprosertyrtyrpheargthrprogluileasnasnsersergly485490495glumetaspproileleuleuproprocysleuthrileserileleu500505510serphepheservalalaleuleuvalileleualacysvalleutrp515520525lyslysargilelysproilevaltrpproserleuproasphislys530535540lysthrleugluhisleucyslyslysproarglysasnleuasnval545550555560serpheasnprogluserpheleuaspcysglnilehisargvalasp565570575aspileglnalaargaspgluvalgluglypheleuglnaspthrphe580585590proglnglnleuglugluserglulysglnargleuglyglyaspval595600605glnserproasncysprosergluaspvalvalilethrprogluser610615620pheglyargaspserserleuthrcysleualaglyasnvalserala625630635640cysaspalaproileleuserserserargserleuaspcysargglu645650655serglylysasnglyprohisvaltyrglnaspleuleuleuserleu660665670glythrthrasnserthrleupropropropheserleuglnsergly675680685ileleuthrleuasnprovalalaglnglyglnproileleuthrser690695700leuglyserasnglngluglualatyrvalthrmetserserphetyr705710715720glnasngln<210>231<211>733<212>prt<213>artificialsequence<220><223>syntheticgfp-linker-p2a-myctag-il7raincppclcodon(exceptp2a)andspliceoptimized<400>231metserglyglyglugluleuphealaglyilevalprovalleuile151015gluleuaspglyaspvalhisglyhislyspheservalargglyglu202530glygluglyaspalaasptyrglylysleugluilelyspheilecys354045thrthrglylysleuprovalprotrpprothrleuvalthrthrleu505560cystyrglyileglncysphealaargtyrprogluhismetlysmet65707580asnaspphephelysseralametprogluglytyrileglngluarg859095thrileglnpheglnaspaspglylystyrlysthrargglygluval100105110lysphegluglyaspthrleuvalasnargilegluleulysglylys115120125aspphelysgluaspglyasnileleuglyhislysleuglutyrser130135140pheasnserhisasnvaltyrileargproasplysalaasnasngly145150155160leuglualaasnphelysthrarghisasnilegluglyglyglyval165170175glnleualaasphistyrglnthrasnvalproleuglyaspglypro180185190valleuileproileasnhistyrleuserthrglnthrlysileser195200205lysaspargasnglualaargasphismetvalleuleugluserphe210215220seralacyscyshisthrhisglymetaspgluleutyrargglyser225230235240glyalathrasnpheserleuleulysglnalaglyaspvalgluglu245250255asnproglyprometthrileleuglythrthrpheglymetvalphe260265270serleuleuglnvalvalserglygluglnlysleuilesergluglu275280285aspleugluserglytyralaglnasnglyaspleugluaspalaglu290295300leuaspasptyrserphesercystyrserglnleugluvalasngly305310315320serglnhisserleuthrcysalaphegluaspproaspvalasnile325330335thrasnleugluphegluilecysglyalaleuvalgluvallyscys340345350leuasnphearglysleuglngluiletyrpheilegluthrlyslys355360365pheleuleuileglylysserasnilecysvallysvalglyglulys370375380serleuthrcyslyslysileaspleuthrthrilevallysproglu385390395400alapropheaspleuservalvaltyrarggluglyalaasnaspphe405410415valvalthrpheasnthrserhisleuglnlyslystyrvallysval420425430leumethisaspvalalatyrargglnglulysaspgluasnlystrp435440445thrhisvalasnleuserserthrlysleuthrleuleuglnarglys450455460leuglnproalaalamettyrgluilelysvalargserileproasp465470475480histyrphelysglyphetrpserglutrpserprosertyrtyrphe485490495argthrprogluileasnasnserserglyglumetaspproileleu500505510leuproprocysleuthrileserileleuserphepheservalala515520525leuleuvalileleualacysvalleutrplyslysargilelyspro530535540ilevaltrpproserleuproasphislyslysthrleugluhisleu545550555560cyslyslysproarglysasnleuasnvalserpheasnprogluser565570575pheleuaspcysglnilehisargvalaspaspileglnalaargasp580585590gluvalgluglypheleuglnaspthrpheproglnglnleugluglu595600605serglulysglnargleuglyglyaspvalglnserproasncyspro610615620sergluaspvalvalilethrprogluserpheglyargaspserser625630635640leuthrcysleualaglyasnvalseralacysaspalaproileleu645650655serserserargserleuaspcysarggluserglylysasnglypro660665670hisvaltyrglnaspleuleuleuserleuglythrthrasnserthr675680685leupropropropheserleuglnserglyileleuthrleuasnpro690695700valalaglnglyglnproileleuthrserleuglyserasnglnglu705710715720glualatyrvalthrmetserserphetyrglnasngln725730<210>232<211>572<212>prt<213>artificialsequence<220><223>syntheticgfp-linker-p2a-il7rasp-myctag-il7raincppclc-terminaltruncationcodon(exceptp2a)andsplice<400>232metserglyglyglugluleuphealaglyilevalprovalleuile151015gluleuaspglyaspvalhisglyhislyspheservalargglyglu202530glygluglyaspalaasptyrglylysleugluilelyspheilecys354045thrthrglylysleuprovalprotrpprothrleuvalthrthrleu505560cystyrglyileglncysphealaargtyrprogluhismetlysmet65707580asnaspphephelysseralametprogluglytyrileglngluarg859095thrileglnpheglnaspaspglylystyrlysthrargglygluval100105110lysphegluglyaspthrleuvalasnargilegluleulysglylys115120125aspphelysgluaspglyasnileleuglyhislysleuglutyrser130135140pheasnserhisasnvaltyrileargproasplysalaasnasngly145150155160leuglualaasnphelysthrarghisasnilegluglyglyglyval165170175glnleualaasphistyrglnthrasnvalproleuglyaspglypro180185190valleuileproileasnhistyrleuserthrglnthrlysileser195200205lysaspargasnglualaargasphismetvalleuleugluserphe210215220seralacyscyshisthrhisglymetaspgluleutyrargglyser225230235240glyalathrasnpheserleuleulysglnalaglyaspvalgluglu245250255asnproglyprometthrileleuglythrthrpheglymetvalphe260265270serleuleuglnvalvalserglygluglnlysleuilesergluglu275280285aspleugluserglytyralaglnasnglyaspleugluaspalaglu290295300leuaspasptyrserphesercystyrserglnleugluvalasngly305310315320serglnhisserleuthrcysalaphegluaspproaspvalasnile325330335thrasnleugluphegluilecysglyalaleuvalgluvallyscys340345350leuasnphearglysleuglngluiletyrpheilegluthrlyslys355360365pheleuleuileglylysserasnilecysvallysvalglyglulys370375380serleuthrcyslyslysileaspleuthrthrilevallysproglu385390395400alapropheaspleuservalvaltyrarggluglyalaasnaspphe405410415valvalthrpheasnthrserhisleuglnlyslystyrvallysval420425430leumethisaspvalalatyrargglnglulysaspgluasnlystrp435440445thrhisvalasnleuserserthrlysleuthrleuleuglnarglys450455460leuglnproalaalamettyrgluilelysvalargserileproasp465470475480histyrphelysglyphetrpserglutrpserprosertyrtyrphe485490495argthrprogluileasnasnserserglyglumetaspproileleu500505510leuproprocysleuthrileserileleuserphepheservalala515520525leuleuvalileleualacysvalleutrplyslysargilelyspro530535540ilevaltrpproserleuproasphislyslysthrleugluhisleu545550555560cyslyslysproarglysvalservalpheglyala565570<210>233<211>562<212>prt<213>artificialsequence<220><223>syntheticgfp-linker-p2a-il7rasp-il7raincppclc-terminaltruncationcodon(exceptp2a)andspliceoptimized<400>233metserglyglyglugluleuphealaglyilevalprovalleuile151015gluleuaspglyaspvalhisglyhislyspheservalargglyglu202530glygluglyaspalaasptyrglylysleugluilelyspheilecys354045thrthrglylysleuprovalprotrpprothrleuvalthrthrleu505560cystyrglyileglncysphealaargtyrprogluhismetlysmet65707580asnaspphephelysseralametprogluglytyrileglngluarg859095thrileglnpheglnaspaspglylystyrlysthrargglygluval100105110lysphegluglyaspthrleuvalasnargilegluleulysglylys115120125aspphelysgluaspglyasnileleuglyhislysleuglutyrser130135140pheasnserhisasnvaltyrileargproasplysalaasnasngly145150155160leuglualaasnphelysthrarghisasnilegluglyglyglyval165170175glnleualaasphistyrglnthrasnvalproleuglyaspglypro180185190valleuileproileasnhistyrleuserthrglnthrlysileser195200205lysaspargasnglualaargasphismetvalleuleugluserphe210215220seralacyscyshisthrhisglymetaspgluleutyrargglyser225230235240glyalathrasnpheserleuleulysglnalaglyaspvalgluglu245250255asnproglyprometthrileleuglythrthrpheglymetvalphe260265270serleuleuglnvalvalserglygluserglytyralaglnasngly275280285aspleugluaspalagluleuaspasptyrserphesercystyrser290295300glnleugluvalasnglyserglnhisserleuthrcysalapheglu305310315320aspproaspvalasnilethrasnleugluphegluilecysglyala325330335leuvalgluvallyscysleuasnphearglysleuglngluiletyr340345350pheilegluthrlyslyspheleuleuileglylysserasnilecys355360365vallysvalglyglulysserleuthrcyslyslysileaspleuthr370375380thrilevallysproglualapropheaspleuservalvaltyrarg385390395400gluglyalaasnaspphevalvalthrpheasnthrserhisleugln405410415lyslystyrvallysvalleumethisaspvalalatyrargglnglu420425430lysaspgluasnlystrpthrhisvalasnleuserserthrlysleu435440445thrleuleuglnarglysleuglnproalaalamettyrgluilelys450455460valargserileproasphistyrphelysglyphetrpserglutrp465470475480serprosertyrtyrpheargthrprogluileasnasnsersergly485490495glumetaspproileleuleuproprocysleuthrileserileleu500505510serphepheservalalaleuleuvalileleualacysvalleutrp515520525lyslysargilelysproilevaltrpproserleuproasphislys530535540lysthrleugluhisleucyslyslysproarglysvalservalphe545550555560glyala<210>234<211>824<212>prt<213>artificialsequence<220><223>syntheticgfp-linker-p2a-etag-il7raincppcln-terminaldeletioncodon(exceptp2a)andspliceoptimized<400>234metserglyglyglugluleuphealaglyilevalprovalleuile151015gluleuaspglyaspvalhisglyhislyspheservalargglyglu202530glygluglyaspalaasptyrglylysleugluilelyspheilecys354045thrthrglylysleuprovalprotrpprothrleuvalthrthrleu505560cystyrglyileglncysphealaargtyrprogluhismetlysmet65707580asnaspphephelysseralametprogluglytyrileglngluarg859095thrileglnpheglnaspaspglylystyrlysthrargglygluval100105110lysphegluglyaspthrleuvalasnargilegluleulysglylys115120125aspphelysgluaspglyasnileleuglyhislysleuglutyrser130135140pheasnserhisasnvaltyrileargproasplysalaasnasngly145150155160leuglualaasnphelysthrarghisasnilegluglyglyglyval165170175glnleualaasphistyrglnthrasnvalproleuglyaspglypro180185190valleuileproileasnhistyrleuserthrglnthrlysileser195200205lysaspargasnglualaargasphismetvalleuleugluserphe210215220seralacyscyshisthrhisglymetaspgluleutyrargglyser225230235240glyalathrasnpheserleuleulysglnalaglyaspvalgluglu245250255asnproglyprometleuleuleuvalthrserleuleuleucysglu260265270leuprohisproalapheleuleuileproarglysvalcysasngly275280285ileglyileglygluphelysaspserleuserileasnalathrasn290295300ilelyshisphelysasncysthrserileserglyaspleuhisile305310315320leuprovalalapheargglyaspserphethrhisthrproproleu325330335aspproglngluleuaspileleulysthrvallysgluilethrgly340345350pheleuleuileglnalatrpprogluasnargthraspleuhisala355360365phegluasnleugluileileargglyargthrlysglnhisglygln370375380pheserleualavalvalserleuasnilethrserleuglyleuarg385390395400serleulysgluileseraspglyaspvalileileserglyasnlys405410415asnleucystyralaasnthrileasntrplyslysleupheglythr420425430serglyglnlysthrlysileileserasnargglygluasnsercys435440445lysalathrglyglnvalcyshisalaleucysserprogluglycys450455460trpglyprogluproargaspcysvalsercysargasnvalserarg465470475480glyargglucysvalasplyscysasnleuleugluglygluproarg485490495gluphevalgluasnserglucysileglncyshisproglucysleu500505510proglnalametasnilethrcysthrglyargglyproaspasncys515520525ileglncysalahistyrileaspglyprohiscysvallysthrcys530535540proalaglyvalmetglygluasnasnthrleuvaltrplystyrala545550555560aspalaglyhisvalcyshisleucyshisproasncysthrtyrgly565570575cysthrglyproglyleugluglycysprothrasnglyprogluile580585590asnasnserserglyglumetaspproileleuleuproprocysleu595600605thrileserileleuserphepheservalalaleuleuvalileleu610615620alacysvalleutrplyslysargilelysproilevaltrpproser625630635640leuproasphislyslysthrleugluhisleucyslyslysproarg645650655lysasnleuasnvalserpheasnprogluserpheleuaspcysgln660665670ilehisargvalaspaspileglnalaargaspgluvalgluglyphe675680685leuglnaspthrpheproglnglnleuglugluserglulysglnarg690695700leuglyglyaspvalglnserproasncysprosergluaspvalval705710715720ilethrprogluserpheglyargaspserserleuthrcysleuala725730735glyasnvalseralacysaspalaproileleuserserserargser740745750leuaspcysarggluserglylysasnglyprohisvaltyrglnasp755760765leuleuleuserleuglythrthrasnserthrleuproproprophe770775780serleuglnserglyileleuthrleuasnprovalalaglnglygln785790795800proileleuthrserleuglyserasnglngluglualatyrvalthr805810815metserserphetyrglnasngln820<210>235<211>593<212>prt<213>artificialsequence<220><223>syntheticmv(ed)-hd24<400>235metasnarggluhisleumetileaspargprotyrvalleuleuala151015valleuphevalmetserleuserleuileglyleuleualaileala202530glyileargleuhisargalaalailetyrthralagluilehislys354045serleuserthrasnleuaspvalthrasnserilegluhisglnval505560lysaspvalleuthrproleuphelysileileglyaspgluvalgly65707580leuargthrproglnargphethraspleuvallyspheileserasp859095lysilelyspheleuasnproaspargglutyrasppheargaspleu100105110thrtrpcysileasnproprogluargilelysleuasptyraspgln115120125tyrcysalaaspvalalaalaglugluleumetasnalaleuvalasn130135140serthrleuleugluthrargthrthrasnglnpheleualavalser145150155160lysglyasncysserglyprothrthrileargglyglnpheserasn165170175metserleuserleuleuaspleutyrleuserargglytyrasnval180185190serserilevalthrmetthrserglnglymettyrglyglythrtyr195200205leuvalglulysproasnleuserserlysargsergluleusergln210215220leusermettyrargvalphegluvalglyvalileargasnprogly225230235240leuglyalaprovalphehismetthrasntyrleugluglnproval245250255serasnaspleuserasncysmetvalalaleuglygluleulysleu260265270alaalaleucyshisglygluaspserilethrileprotyrglngly275280285serglylysglyvalserpheglnleuvallysleuglyvaltrplys290295300serprothraspmetglnsertrpvalproleuserthraspasppro305310315320valileaspargleutyrleuserserhisargglyvalilealaasp325330335asnglnalalystrpalavalprothrthrargthraspasplysleu340345350argmetgluthrcyspheglnglnalacyslysglylysileglnala355360365leucysgluasnproglutrpalaproleulysaspasnargilepro370375380sertyrglyvalleuservalaspleuserleuthrvalgluleulys385390395400ilelysilealaserglypheglyproleuilethrhisglysergly405410415metaspleutyrlysserasnhisasnasnvaltyrtrpleuthrile420425430proprometlysasnleualaleuglyvalileasnthrleuglutrp435440445ileproargphelysvalserproasnleuphethrvalproilelys450455460glualaglygluaspcyshisalaprothrtyrleuproalagluval465470475480aspglyaspvallysleuserserasnleuvalileleuproglygln485490495aspleuglntyrvalleualathrtyraspthrserargvalgluhis500505510alavalvaltyrtyrvaltyrserproglyargserphesertyrphe515520525tyrpropheargleuproilelysglyvalproilegluleuglnval530535540glucysphethrtrpaspglnlysleutrpcysarghisphecysval545550555560leualaaspsergluserglyglyhisilethrhisserglymetval565570575glymetglyvalsercysthrvalthrarggluaspglythrasnarg580585590arg<210>236<211>1275<212>dna<213>mesoplasmaflorum<400>236aaaaaaaataaaatcaatagcaaatattaagatttttaagaaataaaaaattaatattaa60tttacaactgaatataaaagaaacttatacagggtagcataatgggctactgaccccgcc120ttcaaacctatttggagactataagtgaaaaaccactctttaattattaaagtttctttt180tatgtccaaaagacaagaagaaacttttttatttagttgaatttataataagagaaaaag240aaaggatattatatggcaaaaataaaaaaccaatattacaacgagtctgtttcgccaatt300gaatatgcgcaacaaggatttaaaggaaaaatgcgttcagtaaactgaaacgtagtaaat360gatgaaaaagatttagaggtatgaaatagaattacacaaaacttctgattgcctgaaaaa420attccagtttcaaatgatttaacttcatgaagaactttgacaccagaatgacaagaatta480attacaagaacttttacaggattaacattgttagatacaattcaagctactgttggtgat540gtggctcaagttcctaactcattaactgaccatgaacaagtaatttacacaaactttgca600tttatggttgcagttcacgctagatcatatggttcaatcttttcaactttatgttcaagt660gaacaaattgaagaggctcatgaatgagttatcaatacagaaacattacaagaaagagct720aaagcattaattccttattatgtgaatgatgaccctttaaagtcaaaagttgcagctgct780ttaatgccaggcttcttattatatggaggcttctatttaccattttacctatcagctaga840ggtaaattaccaaacacttcagatattattagattaatattaagagataaagttatacat900aactactatagtggttataaatatcaaaagaaagttgctaaactttctccagaaaaacaa960gctgaaatgaaagaatttgtttttaaattattatatgaattaatagatttagaaaaagct1020tatttgaaagaattgtatgaggattttggattagctgatgatgctattagatttagtgtt1080tacaacgcaggtaaatttttacaaaatttaggttatgattcaccgtttacagaagaagaa1140acaagaattgagccagaaatattcacacaattatcagctagagctgatgaaaaccatgat1200ttcttttcagggaatggctcatcatatattatgggagtttcagaagaaactgaagatgac1260gattgggagttttaa1275<210>237<211>64<212>rna<213>mesoplasmaflorum<400>237acuuauacaggguagcauaaugggcuacugaccccgccuucaaaccuauuuggagacuau60aagu64<210>238<211>69<212>rna<213>artificialsequence<220><223>syntheticdeoxyguanosineriboswitchaptamermutations<220><221>misc_feature<222>(10)..(13)<223>natpositions10-13canbea,c,g,ort<220><221>misc_feature<222>(27)..(29)<223>natpositions27-29canbea,c,g,ort<220><221>misc_feature<222>(31)..(32)<223>natpositions31-32canbea,c,g,ort<220><221>misc_feature<222>(33)..(40)<223>natpositions33-40canbea,c,g,t,ornonucleotide<220><221>misc_feature<222>(44)..(46)<223>natpositions44-46canbea,c,g,ort<220><221>misc_feature<222>(57)..(59)<223>natpositions57-59canbea,c,g,ort<400>238acuuauacannnnagcauaaugggcunnngnnnnnnnnnngccnnnaaaccuauuunnng60acuauaagu69<210>239<211>84<212>dna<213>artificialsequence<220><223>syntheticoligolibraryforscreen<220><221>misc_feature<222>(19)..(21)<223>natpositions19-21canbea,c,g,ort<220><221>misc_feature<222>(32)..(34)<223>natpositions32-34canbea,c,g,ort<220><221>misc_feature<222>(38)..(39)<223>natpositions38-39canbea,c,g,ort<220><221>misc_feature<222>(40)..(47)<223>natpositions40-47canbea,c,g,t,ornonucleotide<220><221>misc_feature<222>(49)..(51)<223>natpositions49-51canbea,c,g,ort<220><221>misc_feature<222>(65)..(68)<223>natpositions65-68canbea,c,g,ort<400>239cgcgcgacacttatagtcnnnaaataggtttnnnggcnnnnnnnnnncnnnagcccatta60tgctnnnntgtataagtgccgccc84<210>240<211>33<212>dna<213>artificialsequence<220><223>synthetict7promoteramplificationprimer<400>240taatacgactcactatagggcggcacttataca33<210>241<211>18<212>dna<213>artificialsequence<220><223>syntheticreverseamplificationprimer<400>241cgcgcgacacttatagtc18<210>242<211>915<212>dna<213>bacillussubtilis<400>242tcaaaagcctggcggcgcggtcgtcagactcttttatatcgaatccccttgaaatacgaa60tgatatctaaaaaaacaaaattaaagttcgggaatttttattttcagcctatgcaagaga120ttagaatcttgatataatttattacaatataataggaacactcatataatcgcgtggata180tggcacgcaagtttctaccgggcaccgtaaatgtccgactatgggtgagcaatggaaccg240cacgtgtacggttttttgtgatatcagcattgcttgctctttatttgagcgggcaatgct300ttttttattctcataacggaggtagacaggatggaagcactgaaacggaaaatagaggaa360gaaggcgtcgtattatctgatcaggtattgaaagtggattcttttttgaatcaccaaatt420gatccgctgcttatgcagagaattggtgatgaatttgcgtctaggtttgcaaaagacggt480attaccaaaattgtgacaatcgaatcatcaggtatcgctcccgctgtaatgacgggcttg540aagctgggtgtgccagttgtcttcgcgagaaagcataaatcgttaacactcaccgacaac600ttgctgacagcgtctgtttattcctttacgaagcaaacagaaagccaaatcgcagtgtct660gggacccacctgtcggatcaggatcatgtgctgattatcgatgattttttggcaaatgga720caggcagcgcacgggcttgtgtcgattgtgaagcaagcgggagcttctattgcgggaatc780ggcattgttattgaaaagtcatttcagccgggaagagatgaacttgtaaaactgggctac840cgagtggaatctttggcaagaattcagtctttagaagaaggaaaagtgtccttcgtacag900gaggttcattcatga915<210>243<211>69<212>rna<213>bacillussubtilis<400>243cacucauauaaucgcguggauauggcacgcaaguuucuaccgggcaccguaaauguccga60cuaugggug69<210>244<211>74<212>rna<213>artificialsequence<220><223>syntheticguanosinexptriboswitchaptamermutations<220><221>misc_feature<222>(11)..(14)<223>natpositions11-14canbea,c,g,ort<220><221>misc_feature<222>(30)..(35)<223>natpositions30-35canbea,c,g,ort<220><221>misc_feature<222>(36)..(43)<223>natpositions36-43canbea,c,g,t,ornonucleotide<220><221>misc_feature<222>(47)..(49)<223>natpositions47-49canbea,c,g,ort<220><221>misc_feature<222>(61)..(63)<223>natpositions61-63canbea,c,g,ort<400>244cacucauauannnncguggauauggcacgnnngnnnnnnnnnnaccnnnuaccguaaaug60nnngacuaugggug74<210>245<211>89<212>dna<213>artificialsequence<220><223>syntheticoligolibraryforscreen<220><221>misc_feature<222>(20)..(22)<223>natpositions20-22canbea,c,g,ort<220><221>misc_feature<222>(34)..(36)<223>natpositions34-36canbea,c,g,ort<220><221>misc_feature<222>(40)..(41)<223>natpositions40-41canbea,c,g,ort<220><221>misc_feature<222>(42)..(49)<223>natpositions42-49canbea,c,g,t,ornonucleotide<220><221>misc_feature<222>(51)..(53)<223>natpositions51-53canbea,c,g,ort<220><221>misc_feature<222>(69)..(72)<223>natpositions69-72canbea,c,g,ort<400>245cgcgcgaccacccatagtcnnncatttacggtgnnnggtnnnnnnnnnncnnncgtgcca60tatccacgnnnntatatgagtggccgccc89<210>246<211>34<212>dna<213>artificialsequence<220><223>synthetict7promoteramplificationprimer<400>246taatacgactcactatagggcggccactcatata34<210>247<211>19<212>dna<213>artificialsequence<220><223>syntheticreverseamplificationprimer<400>247cgcgcgaccacccatagtc19<210>248<211>152<212>rna<213>artificialsequence<220><223>syntheticoligo1transcribedrna<400>248gggcggcacuuauacaggguagcauaaugggcuacugacgccuucaaaccuauuuggaga60cuauaagugucgcgcggggcggcacuuauacaggguagcauaaugggcuacugacgccuu120caaaccuauuuggagacuauaagugucgcgcg152<210>249<211>76<212>dna<213>artificialsequence<220><223>syntheticoligo1synthesizedtemplate<400>249cgcgcgacacttatagtctccaaataggtttgaaggcgtcagtagcccattatgctaccc60tgtataagtgccgccc76<210>250<211>79<212>rna<213>artificialsequence<220><223>syntheticoligo2transcribedrna<400>250gggcggcacuuauacaggguagcauaaugggcuacugaccccgccuucaaaccuauuugg60agacuauaagugucgcgcg79<210>251<211>79<212>dna<213>artificialsequence<220><223>syntheticoligo2synthesizedtemplate<400>251cgcgcgacacttatagtctccaaataggtttgaaggcggggtcagtagcccattatgcta60ccctgtataagtgccgccc79<210>252<211>560<212>prt<213>artificialsequence<220><223>syntheticdafss-acd3scfv(ucht1)igg1fc-cd14gpi<400>252metthrvalalaargproservalproalaalaleuproleuleugly151015gluleuproargleuleuleuleuvalleuleucysleuproaspile202530glnmetthrglnserproserserleuseralaservalglyasparg354045valthrilethrcysargalaserglnaspileargasntyrleuasn505560trptyrglnglnlysproglylysalaprolysleuleuiletyrtyr65707580thrserargleugluserglyvalproserargpheserglysergly859095serglythrasptyrthrleuthrileserserleuglnprogluasp100105110phealathrtyrtyrcysglnglnglyasnthrleuprotrpthrphe115120125glyglnglythrlysvalgluilelysglyglyglyglyserglygly130135140glyglyserglyglyglyglysergluvalglnleuvalglusergly145150155160glyglyleuvalglnproglyglyserleuargleusercysalaala165170175serglytyrserphethrglytyrthrmetasntrpvalargglnala180185190proglylysglyleuglutrpvalalaleuileasnprotyrlysgly195200205valserthrtyrasnglnlysphelysaspargphethrileserval210215220asplysserlysasnthralatyrleuglnmetasnserleuargala225230235240gluaspthralavaltyrtyrcysalaargserglytyrtyrglyasp245250255serasptrptyrpheaspvaltrpglyglnglythrleuvalthrval260265270sersergluprolyssercysasplysthrhisthrcysproprocys275280285proalaprogluleuleuglyglyproservalpheleuphepropro290295300lysprolysaspthrleumetileserargthrprogluvalthrcys305310315320valvalvalaspvalserhisgluaspprogluvallyspheasntrp325330335tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu340345350gluglntyrasnserthrtyrargvalvalservalleuthrvalleu355360365hisglnasptrpleuasnglylysglutyrlyscyslysvalserasn370375380lysalaleuproalaproileglulysthrileserlysalalysgly385390395400glnproarggluproglnvaltyrthrleuproproserarggluglu405410415metthrlysasnglnvalserleuthrcysleuvallysglyphetyr420425430proseraspilealavalglutrpgluserasnglyglnprogluasn435440445asntyrlysthrthrproprovalleuaspseraspglyserphephe450455460leutyrserlysleuthrvalasplysserargtrpglnglnglyasn465470475480valphesercysservalmethisglualaleuhisasnhistyrthr485490495glnlysserleuserleuserproglyvalaspasnleuthrleuasp500505510glyasnpropheleuvalproglythralaleuprohisgluglyser515520525metasnserglyvalvalproalacysalaargserthrleuserval530535540glyvalserglythrleuvalleuleuglnglyalaargglypheala545550555560<210>253<211>276<212>prt<213>artificialsequence<220><223>syntheticdafss-cd80ecd-cd16gpi<400>253metthrvalalaargproservalproalaalaleuproleuleugly151015gluleuproargleuleuleuleuvalleuleucysleuprovalile202530hisvalthrlysgluvallysgluvalalathrleusercysglyhis354045asnvalservalglugluleualaglnthrargiletyrtrpglnlys505560glulyslysmetvalleuthrmetmetserglyaspmetasniletrp65707580proglutyrlysasnargthrilepheaspilethrasnasnleuser859095ilevalileleualaleuargproseraspgluglythrtyrglucys100105110valvalleulystyrglulysaspalaphelysarggluhisleuala115120125gluvalthrleuservallysalaasppheprothrproserileser130135140aspphegluileprothrserasnileargargileilecysserthr145150155160serglyglypheprogluprohisleusertrpleugluasnglyglu165170175gluleuasnalaileasnthrthrvalserglnaspprogluthrglu180185190leutyralavalserserlysleuasppheasnmetthrthrasnhis195200205serphemetcysleuilelystyrglyhisleuargvalasnglnthr210215220pheasntrpasnthrthrlysglngluhispheproaspasnvalser225230235240thrileserserpheserproproglytyrglnvalserphecysleu245250255valmetvalleuleuphealavalaspthrglyleutyrpheserval260265270lysthrasnile275<210>254<211>533<212>prt<213>artificialsequence<220><223>syntheticdafss-il7-daf<400>254metalathrthrmetthrvalalaargproservalproalaalaleu151015proleuleuglygluleuproargleuleuleuleuvalleuleucys202530leuproalaaspcysaspilegluglylysaspglylysglntyrglu354045servalleumetvalserileaspglnleuleuaspsermetlysglu505560ileglyserasncysleuasnasnglupheasnphephelysarghis65707580ilecysaspalaasnlysgluglymetpheleupheargalaalaarg859095lysleuargglnpheleulysmetasnserthrglyasppheaspleu100105110hisleuleulysvalsergluglythrthrileleuleuasncysthr115120125glyglnvallysglyarglysproalaalaleuglyglualaglnpro130135140thrlysserleuglugluasnlysserleulysgluglnlyslysleu145150155160asnaspleucyspheleulysargleuleuglngluilelysthrcys165170175trpasnlysileleumetglythrlysgluhiscysglyleupropro180185190aspvalproasnalaglnproalaleugluglyargthrserphepro195200205gluaspthrvalilethrtyrlyscysglugluserphevallysile210215220proglyglulysaspservalilecysleulysglyserglntrpser225230235240aspileglugluphecysasnargsercysgluvalprothrargleu245250255asnseralaserleulysglnprotyrilethrglnasntyrphepro260265270valglythrvalvalglutyrglucysargproglytyrargargglu275280285proserleuserprolysleuthrcysleuglnasnleulystrpser290295300thralavalgluphecyslyslyslyssercysproasnproglyglu305310315320ileargasnglyglnileaspvalproglyglyileleupheglyala325330335thrileserphesercysasnthrglytyrlysleupheglyserthr340345350serserphecysleuileserglyserservalglntrpserasppro355360365leuproglucysarggluiletyrcysproalaproproglnileasp370375380asnglyileileglnglygluargasphistyrglytyrargglnser385390395400valthrtyralacysasnlysglyphethrmetileglygluhisser405410415iletyrcysthrvalasnasnaspgluglyglutrpserglypropro420425430proglucysargglylysserleuthrserlysvalproprothrval435440445glnlysprothrthrvalasnvalprothrthrgluvalserprothr450455460serglnlysthrthrthrlysthrthrthrproasnalaglnalathr465470475480argserthrprovalserargthrthrlyshisphehisgluthrthr485490495proasnlysglyserglythrthrserglythrthrargleuleuser500505510glyhisthrcysphethrleuthrglyleuleuglythrleuvalthr515520525metglyleuleuthr530<210>255<211>1823<212>dna<213>artificialsequence<220><223>syntheticef-1apromoterwithmirs<400>255ggctccggtgcccgtcagtgggcagagcgcacatcgcccacagtccccgagaagttgggg60ggaggggtcggcaattgaaccggtgcctagagaaggtggcgcggggtaaactgggaaagt120gatgtcgtgtactggctccgcctttttcccgagggtgggggagaaccgtatataagtgca180gtagtcgccgtgaacgttctttttcgcaacgggtttgccgccagaacacaggtaagtgcc240gtgtgtggttcccgcgggcctggcctctttacgggttatggcccttgcgtgccttgaatt300acttccacctggctgcagtacgtgattcttgatcccgagcttcgggttggaagtgggtgg360gagagttcgaggccttgcgcttaaggagccccttcgcctcgtgcttgagttgaggcctgg420cctgggcgctggggccgccgcgtgcgaatctggtggcaccttcgcgcctgtctcgctgct480ttcgataagtctctagccatttaaaatttttgatgacctgctgcgacgctttttttctgg540caagatagtcttgtaaatgcgggccaagatctgcacactggtatttcggtttttggggcc600gcgggcggcgacggggcccgtgcgtcccagcgcacatgttcggcgaggcggggcctgcga660gcgcggccaccgagaatcggacgggggtagtctcaagctggccggcctgctctggtgcct720ggcctcgcgccgccgtgtatcgccccgccctgggcggcaaggctggcccggtcggcacca780gttgcgtgagcggaaagatggccgcttcccggccctgctgcagggagctcaaaatggagg840acgcggcgctcgggagagcgggcgggtgagtcacccacacaaaggaaaagggcctttccg900tcctcagccgtcgcttcatgtgactccactgagtaccgggcgccgtccaggcacctcgat960tagttcctggaggcttgctgaaggctgtatgctgacatggtacagttcaatggtggtttt1020ggccactgactgaccaccattgctgtaccatgtcaggacacaaggcctgttactagcact1080cacatggaacaaatggcccacattggtgccggatgaagctcttatgttgcacggtcatct1140ggaggcttgctgaaggctgtatgctgtcagtctgttcatcttctggcgttttggccactg1200actgacgccagaaggaacagactgacaggacacaaggcctgttactagcactcacatgga1260acaaatggccgttgccggagtcttggcagcgagagatcactatcaactaactggaggctt1320gctgaaggctgtatgctgaagcgtgaagtgaatcaacgggttttggccactgactgaccc1380gttgatacttcacgcttcaggacacaaggcctgttactagcactcacatggaacaaatgg1440ccgtgttaattgtccatgtagcgaggcatccttatggcgtggctggaggcttgctgaagg1500ctgtatgctggcagtatcctagtacattgacgttttggccactgactgacgtcaatgtta1560ggatactgccaggacacaaggcctgttactagcactcacatggaacaaatggccgctttt1620ggagtacgtcgtctttaggttggggggaggggttttatgcgatggagtttccccacactg1680agtgggtggagactgaagttaggccagcttggcacttgatgtaattctccttggaatttg1740ccctttttgagtttggatcttggttcattctcaagcctcagacagtggttcaaagttttt1800ttcttccatttcaggtgtcgtga1823<210>256<211>28<212>dna<213>musmusculus<400>256ctggaggcttgctgaaggctgtatgctg28<210>257<211>21<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>257acatggtacagttcaatggtg21<210>258<211>19<212>dna<213>artificialsequence<220><223>syntheticdnaencodingloop<400>258gttttggccactgactgac19<210>259<211>19<212>dna<213>artificialsequence<220><223>syntheticdnaencodingstem<400>259caccattgctgtaccatgt19<210>260<211>45<212>dna<213>musmusculus<400>260caggacacaaggcctgttactagcactcacatggaacaaatggcc45<210>261<211>21<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>261tcagtctgttcatcttctggc21<210>262<211>19<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>262gccagaaggaacagactga19<210>263<211>21<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>263aagcgtgaagtgaatcaacgg21<210>264<211>19<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>264ccgttgatacttcacgctt19<210>265<211>21<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>265gcagtatcctagtacattgac21<210>266<211>19<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>266gtcaatgttaggatactgc19<210>267<211>21<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>267atatgtacttggctggacagc21<210>268<211>19<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>268gctgtccacaagtacatat19<210>269<211>39<212>dna<213>artificialsequence<220><223>syntheticdnaencodinglinker<400>269cacattggtgccggatgaagctcttatgttgccggtcat39<210>270<211>21<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>270ctgttaatgctaatcgtgata21<210>271<211>18<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>271tatcacgattattaacag18<210>272<211>40<212>dna<213>artificialsequence<220><223>syntheticdnaencodinglinker<400>272gttgccggagtcttggcagcgagagatcactatcaactaa40<210>273<211>21<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>273taccagtttagcacgaagctc21<210>274<211>19<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>274gagcttcgctaaactggta19<210>275<211>40<212>dna<213>artificialsequence<220><223>syntheticdnaencodinglinker<400>275gtgttaattgtccatgtagcgaggcatccttatggcgtgg40<210>276<211>21<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>276tgccgctgaaatccaaggcaa21<210>277<211>19<212>dna<213>artificialsequence<220><223>syntheticdnaencodingmirnastem<400>277ttgccttgtttcagcggca19<210>278<211>534<212>prt<213>artificialsequence<220><223>syntheticucht1scfvfc-gpi<400>278metaspmetargvalproalaglnleuleuglyleuleuleuleutrp151015leuserglyalaargcysaspileglnmetthrglnserproserser202530leuseralaservalglyaspargvalthrilethrcysargalaser354045glnaspileargasntyrleuasntrptyrglnglnlysproglylys505560alaprolysleuleuiletyrtyrthrserargleugluserglyval65707580proserargpheserglyserglyserglythrasptyrthrleuthr859095ileserserleuglnprogluaspphealathrtyrtyrcysglngln100105110glyasnthrleuprotrpthrpheglyglnglythrlysvalgluile115120125lysglyglyglyglyserglyglyglyglyserglyglyglyglyser130135140gluvalglnleuvalgluserglyglyglyleuvalglnproglygly145150155160serleuargleusercysalaalaserglytyrserphethrglytyr165170175thrmetasntrpvalargglnalaproglylysglyleuglutrpval180185190alaleuileasnprotyrlysglyvalserthrtyrasnglnlysphe195200205lysaspargphethrileservalasplysserlysasnthralatyr210215220leuglnmetasnserleuargalagluaspthralavaltyrtyrcys225230235240alaargserglytyrtyrglyaspserasptrptyrpheaspvaltrp245250255glyglnglythrleuvalthrvalsersergluprolyssercysasp260265270lysthrhisthrcysproprocysproalaprogluleuleuglygly275280285proservalpheleupheproprolysprolysaspthrleumetile290295300serargthrprogluvalthrcysvalvalvalaspvalserhisglu305310315320aspprogluvallyspheasntrptyrvalaspglyvalgluvalhis325330335asnalalysthrlysproargglugluglntyrasnserthrtyrarg340345350valvalservalleuthrvalleuhisglnasptrpleuasnglylys355360365glutyrlyscyslysvalserasnlysalaleuprothrproileglu370375380lysthrileserlysalalysglyglnproarggluproglnvaltyr385390395400thrleuproproserarggluglumetthrlysasnglnvalserleu405410415thrcysleuvallysglyphetyrproseraspilealavalglutrp420425430gluserasnglyglnprogluasnasntyrlysthrthrproproval435440445leuaspseraspglyserphepheleutyrserlysleuthrvalasp450455460lysserargtrpglnglnglyasnvalphesercysservalmethis465470475480glualaleuhisasnhistyrthrglnlysserleuserleuserpro485490495glyproasnlysglyserglythrthrserglythrthrargleuleu500505510serglyhisthrcysphethrleuthrglyleuleuglythrleuval515520525thrmetglyleuleuthr530<210>279<211>530<212>prt<213>artificialsequence<220><223>syntheticokt3scfvfc-gpi<400>279metaspmetargvalproalaglnleuleuglyleuleuleuleutrp151015leuserglyalaargcysglnilevalleuthrglnserproalaile202530metseralaserproglyglulysvalthrmetthrcysseralaser354045serservalsertyrmetasntrptyrglnglnlysserglythrser505560prolysargtrpiletyraspthrserlysleualaserglyvalpro65707580alahispheargglyserglyserglythrsertyrserleuthrile859095serglymetglualagluaspalaalathrtyrtyrcysglnglntrp100105110serserasnprophethrpheglyserglythrlysleugluileasn115120125glyglyglyglyserglyglyglyglyserglyglyglyglysergln130135140valglnleuglnglnserglyalagluleualaargproglyalaser145150155160vallysmetsercyslysalaserglytyrthrphethrargtyrthr165170175methistrpvallysglnargproglyglnglyleuglutrpilegly180185190tyrileasnproserargglytyrthrasntyrasnglnlysphelys195200205asplysalathrleuthrthrasplysserserserthralatyrmet210215220glnleuserserleuthrsergluaspseralavaltyrtyrcysala225230235240argtyrtyraspasphistyrcysleuasptyrtrpglyglnglythr245250255thrleuthrvalsersergluprolyssercysasplysthrhisthr260265270cysproprocysproalaprogluleuleuglyglyproservalphe275280285leupheproprolysprolysaspthrleumetileserargthrpro290295300gluvalthrcysvalvalvalaspvalserhisgluaspprogluval305310315320lyspheasntrptyrvalaspglyvalgluvalhisasnalalysthr325330335lysproargglugluglntyrasnserthrtyrargvalvalserval340345350leuthrvalleuhisglnasptrpleuasnglylysglutyrlyscys355360365lysvalserasnlysalaleuproalaproileglulysthrileser370375380lysalalysglyglnproarggluproglnvaltyrthrleupropro385390395400serarggluglumetthrlysasnglnvalserleuthrcysleuval405410415lysglyphetyrproseraspilealavalglutrpgluserasngly420425430glnprogluasnasntyrlysthrthrproprovalleuaspserasp435440445glyserphepheleutyrserlysleuthrvalasplysserargtrp450455460glnglnglyasnvalphesercysservalmethisglualaleuhis465470475480asnhistyrthrglnlysserleuserleuserproglyproasnlys485490495glyserglythrthrserglythrthrargleuleuserglyhisthr500505510cysphethrleuthrglyleuleuglythrleuvalthrmetglyleu515520525leuthr530<210>280<211>280<212>prt<213>artificialsequence<220><223>synthetichcd80-cd16gpi<400>280metglyhisthrargargglnglythrserproserlyscysprotyr151015leuasnphepheglnleuleuvalleualaglyleuserhisphecys202530serglyvalilehisvalthrlysgluvallysgluvalalathrleu354045sercysglyhisasnvalservalglugluleualaglnthrargile505560tyrtrpglnlysglulyslysmetvalleuthrmetmetserglyasp65707580metasniletrpproglutyrlysasnargthrilepheaspilethr859095asnasnleuserilevalileleualaleuargproseraspglugly100105110thrtyrglucysvalvalleulystyrglulysaspalaphelysarg115120125gluhisleualagluvalthrleuservallysalaasppheprothr130135140proserileseraspphegluileprothrserasnileargargile145150155160ilecysserthrserglyglypheprogluprohisleusertrpleu165170175gluasnglyglugluleuasnalaileasnthrthrvalserglnasp180185190progluthrgluleutyralavalserserlysleuasppheasnmet195200205thrthrasnhisserphemetcysleuilelystyrglyhisleuarg210215220valasnglnthrpheasntrpasnthrthrlysglngluhisphepro225230235240aspasnvalserthrileserserpheserproproglytyrglnval245250255serphecysleuvalmetvalleuleuphealavalaspthrglyleu260265270tyrpheservallysthrasnile275280<210>281<211>120<212>prt<213>artificialsequence<220><223>synthetictarget1mrbvh<400>281gluvalglnleuvalglnserglyalagluvallyslysproglyala151015thrvallysilesercyslysvalserglytyrserphetrpglyala202530thrmetasntrpileargglnproproglylysglyleuglutrpile354045glyleuilelysproserasnglyglythrsertyrasnglnlysphe505560lysglyargvalthrileseralaasplysserileserthralatyr65707580leuglntrpserserleulysalaseraspthralamettyrtyrcys859095alahisglyhistyrglusertyrglualametasptyrtrpglygln100105110glythrleuvalthrvalserser115120<210>282<211>107<212>prt<213>artificialsequence<220><223>synthetictarget1mrbvl<400>282aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcyslysalaserglnaspvalvalserala202530valalatrptyrglnglnlysproglyglnalaproargleuleuile354045tyrtrpglnaspthrarghisthrglyvalproserargphesergly505560serglyserglythrgluphethrleuthrileserserleuglnpro65707580aspaspphealathrtyrtyrcysglngluhispheserproproleu859095thrpheglyglnglythrlysvalgluilelys100105<210>283<211>15<212>prt<213>artificialsequence<220><223>syntheticlinker1<400>283glyglyglyglyserglyglyglyglyserglyglyglyglyser151015<210>284<211>357<212>prt<213>artificialsequence<220><223>syntheticc1controlpolypeptidecontaininggmcsfalphachainsignalsequence(aminoacids1-22)andetag(aminoacids23-357)<400>284metleuleuleuvalthrserleuleuleucysgluleuprohispro151015alapheleuleuileproarglysvalcysasnglyileglyilegly202530gluphelysaspserleuserileasnalathrasnilelyshisphe354045lysasncysthrserileserglyaspleuhisileleuprovalala505560pheargglyaspserphethrhisthrproproleuaspproglnglu65707580leuaspileleulysthrvallysgluilethrglypheleuleuile859095glnalatrpprogluasnargthraspleuhisalaphegluasnleu100105110gluileileargglyargthrlysglnhisglyglnpheserleuala115120125valvalserleuasnilethrserleuglyleuargserleulysglu130135140ileseraspglyaspvalileileserglyasnlysasnleucystyr145150155160alaasnthrileasntrplyslysleupheglythrserglyglnlys165170175thrlysileileserasnargglygluasnsercyslysalathrgly180185190glnvalcyshisalaleucysserprogluglycystrpglyproglu195200205proargaspcysvalsercysargasnvalserargglyargglucys210215220valasplyscysasnleuleugluglygluproarggluphevalglu225230235240asnserglucysileglncyshisproglucysleuproglnalamet245250255asnilethrcysthrglyargglyproaspasncysileglncysala260265270histyrileaspglyprohiscysvallysthrcysproalaglyval275280285metglygluasnasnthrleuvaltrplystyralaaspalaglyhis290295300valcyshisleucyshisproasncysthrtyrglycysthrglypro305310315320glyleugluglycysprothrasnglyprolysileproserileala325330335thrglymetvalglyalaleuleuleuleuleuvalvalalaleugly340345350ileglyleuphemet355<210>285<211>240<212>prt<213>artificialsequence<220><223>syntheticokt3scfv<400>285glnilevalleuthrglnserproalailemetseralaserprogly151015glulysvalthrmetthrcysseralaserserservalsertyrmet202530asntrptyrglnglnlysserglythrserprolysargtrpiletyr354045aspthrserlysleualaserglyvalproalahispheargglyser505560glyserglythrsertyrserleuthrileserglymetglualaglu65707580aspalaalathrtyrtyrcysglnglntrpserserasnprophethr859095pheglyserglythrlysleugluileasnglyglyglyglysergly100105110glyglyglyserglyglyglyglyserglnvalglnleuglnglnser115120125glyalagluleualaargproglyalaservallysmetsercyslys130135140alaserglytyrthrphethrargtyrthrmethistrpvallysgln145150155160argproglyglnglyleuglutrpileglytyrileasnproserarg165170175glytyrthrasntyrasnglnlysphelysasplysalathrleuthr180185190thrasplysserserserthralatyrmetglnleuserserleuthr195200205sergluaspseralavaltyrtyrcysalaargtyrtyraspasphis210215220tyrcysleuasptyrtrpglyglnglythrthrleuthrvalserser225230235240<210>286<211>532<212>prt<213>artificialsequence<220><223>syntheticdafss-il7-daffusion<400>286metthrvalalaargproservalproalaalaleuproleuleugly151015gluleuproargleuleuleuleuvalleuleucysleuproalaval202530trpglyaspcysaspilegluglylysaspglylysglntyrgluser354045valleumetvalserileaspglnleuleuaspsermetlysgluile505560glyserasncysleuasnasnglupheasnphephelysarghisile65707580cysaspalaasnlysgluglymetpheleupheargalaalaarglys859095leuargglnpheleulysmetasnserthrglyasppheaspleuhis100105110leuleulysvalsergluglythrthrileleuleuasncysthrgly115120125glnvallysglyarglysproalaalaleuglyglualaglnprothr130135140lysserleuglugluasnlysserleulysgluglnlyslysleuasn145150155160aspleucyspheleulysargleuleuglngluilelysthrcystrp165170175asnlysileleumetglythrlysgluhiscysglyleuproproasp180185190valproasnalaglnproalaleugluglyargthrserpheproglu195200205aspthrvalilethrtyrlyscysglugluserphevallysilepro210215220glyglulysaspservalilecysleulysglyserglntrpserasp225230235240ileglugluphecysasnargsercysgluvalprothrargleuasn245250255seralaserleulysglnprotyrilethrglnasntyrpheproval260265270glythrvalvalglutyrglucysargproglytyrargargglupro275280285serleuserprolysleuthrcysleuglnasnleulystrpserthr290295300alavalgluphecyslyslyslyssercysproasnproglygluile305310315320argasnglyglnileaspvalproglyglyileleupheglyalathr325330335ileserphesercysasnthrglytyrlysleupheglyserthrser340345350serphecysleuileserglyserservalglntrpseraspproleu355360365proglucysarggluiletyrcysproalaproproglnileaspasn370375380glyileileglnglygluargasphistyrglytyrargglnserval385390395400thrtyralacysasnlysglyphethrmetileglygluhisserile405410415tyrcysthrvalasnasnaspgluglyglutrpserglypropropro420425430glucysargglylysserleuthrserlysvalproprothrvalgln435440445lysprothrthrvalasnvalprothrthrgluvalserprothrser450455460glnlysthrthrthrlysthrthrthrproasnalaglnalathrarg465470475480serthrprovalserargthrthrlyshisphehisgluthrthrpro485490495asnlysglyserglythrthrserglythrthrargleuleusergly500505510histhrcysphethrleuthrglyleuleuglythrleuvalthrmet515520525glyleuleuthr530<210>287<211>564<212>prt<213>artificialsequence<220><223>syntheticdafss-acd3scfv(ucht1)igg1fc-cd14gpi<400>287metthrvalalaargproservalproalaalaleuproleuleugly151015gluleuproargleuleuleuleuvalleuleucysleuproalaval202530trpglyaspileglnmetthrglnserproserserleuseralaser354045valglyaspargvalthrilethrcysargalaserglnaspilearg505560asntyrleuasntrptyrglnglnlysproglylysalaprolysleu65707580leuiletyrtyrthrserargleugluserglyvalproserargphe859095serglyserglyserglythrasptyrthrleuthrileserserleu100105110glnprogluaspphealathrtyrtyrcysglnglnglyasnthrleu115120125protrpthrpheglyglnglythrlysvalgluilelysglyglygly130135140glyserglyglyglyglyserglyglyglyglysergluvalglnleu145150155160valgluserglyglyglyleuvalglnproglyglyserleuargleu165170175sercysalaalaserglytyrserphethrglytyrthrmetasntrp180185190valargglnalaproglylysglyleuglutrpvalalaleuileasn195200205protyrlysglyvalserthrtyrasnglnlysphelysaspargphe210215220thrileservalasplysserlysasnthralatyrleuglnmetasn225230235240serleuargalagluaspthralavaltyrtyrcysalaargsergly245250255tyrtyrglyaspserasptrptyrpheaspvaltrpglyglnglythr260265270leuvalthrvalsersergluprolyssercysasplysthrhisthr275280285cysproprocysproalaprogluleuleuglyglyproservalphe290295300leupheproprolysprolysaspthrleumetileserargthrpro305310315320gluvalthrcysvalvalvalaspvalserhisgluaspprogluval325330335lyspheasntrptyrvalaspglyvalgluvalhisasnalalysthr340345350lysproargglugluglntyrasnserthrtyrargvalvalserval355360365leuthrvalleuhisglnasptrpleuasnglylysglutyrlyscys370375380lysvalserasnlysalaleuproalaproileglulysthrileser385390395400lysalalysglyglnproarggluproglnvaltyrthrleupropro405410415serarggluglumetthrlysasnglnvalserleuthrcysleuval420425430lysglyphetyrproseraspilealavalglutrpgluserasngly435440445glnprogluasnasntyrlysthrthrproprovalleuaspserasp450455460glyserphepheleutyrserlysleuthrvalasplysserargtrp465470475480glnglnglyasnvalphesercysservalmethisglualaleuhis485490495asnhistyrthrglnlysserleuserleuserproglyvalaspasn500505510leuthrleuaspglyasnpropheleuvalproglythralaleupro515520525hisgluglysermetasnserglyvalvalproalacysalaargser530535540thrleuservalglyvalserglythrleuvalleuleuglnglyala545550555560argglypheala<210>288<211>280<212>prt<213>artificialsequence<220><223>syntheticdafss-cd80ecd-cd16gpi<400>288metthrvalalaargproservalproalaalaleuproleuleugly151015gluleuproargleuleuleuleuvalleuleucysleuproalaval202530trpglyvalilehisvalthrlysgluvallysgluvalalathrleu354045sercysglyhisasnvalservalglugluleualaglnthrargile505560tyrtrpglnlysglulyslysmetvalleuthrmetmetserglyasp65707580metasniletrpproglutyrlysasnargthrilepheaspilethr859095asnasnleuserilevalileleualaleuargproseraspglugly100105110thrtyrglucysvalvalleulystyrglulysaspalaphelysarg115120125gluhisleualagluvalthrleuservallysalaasppheprothr130135140proserileseraspphegluileprothrserasnileargargile145150155160ilecysserthrserglyglypheprogluprohisleusertrpleu165170175gluasnglyglugluleuasnalaileasnthrthrvalserglnasp180185190progluthrgluleutyralavalserserlysleuasppheasnmet195200205thrthrasnhisserphemetcysleuilelystyrglyhisleuarg210215220valasnglnthrpheasntrpasnthrthrlysglngluhisphepro225230235240aspasnvalserthrileserserpheserproproglytyrglnval245250255serphecysleuvalmetvalleuleuphealavalaspthrglyleu260265270tyrpheservallysthrasnile275280<210>289<211>119<212>prt<213>artificialsequence<220><223>synthetictarget2mrbvh<400>289glnvalglnleuglngluserglyproglyleuvallysprosergln151015thrleuserleuthrcysthrvalserglytyrserilethrthrgly202530glutyrtrpasntrpvalargglnalaargglyglnargleuglutrp354045ileglytyrilethrtyraspglyserlysasntyrasnproserleu505560lysasnargvalthrileservalaspthrserlysasnglnpheser65707580leulysleuserservalthralaalaaspthralavaltyrtyrcys859095serargphegluglyvaltrptyrglyleuasptyrtrpglyglngly100105110thrleuvalthrvalserser115<210>290<211>111<212>prt<213>artificialsequence<220><223>synthetictarget2mrbvl<400>290alaileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalasergluservalaspargtyr202530glyasnserpheilehistrptyrglnglnlysproglylysalapro354045lysleuleuiletyrargthrtyrasnleugluserglyileproala505560argpheserglyserglyserglythrgluphethrleuthrileser65707580serleuglnsergluaspphealavaltyrtyrcysglnglnthrasn859095gluaspprotrpthrpheglyglnglythrlysvalgluilelys100105110<210>291<211>30<212>prt<213>artificialsequence<220><223>syntheticlinker2<400>291glyglyglyglyserglyglyglyglyserglyglyglyglysergly151015glyglyglyserglyglyglyglyserglyglyglyglyser202530当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1