一种柳枝稷腺苷高半胱氨酸在改变木质素单体和提高细胞壁降解效率方面的应用的制作方法
本发明属于植物基因工程技术领域,具体涉及柳枝稷一碳代谢腺苷高半胱氨酸水解酶(s-adenosyl-l-homocysteinehydrolase1,sahh1)的分子调控,以研究其对木质素代谢以及细胞壁降解效率的影响。
背景技术:
细胞壁是植物储存生物质和能量的主要场所。而细胞壁中的纤维素(cellulose)和半纤维素(hemicellulose),不但可以作为牧草饲料,还可用于微生物发酵生产乙醇和丁醇等生物能源。植物细胞壁成分除纤维素、半纤维素以为,还存在另一成分木质素(lignin)。木质素在生物体内起到支撑细胞、维持水分运输、增强植物抗逆和抵抗病虫害入等生理功能,但是在生物能源转化方面和牧草消化方面,木质素是重要的限制因子,它的存在使得纤维素降解效率显著降低。柳枝稷(panicumvirgatuml.)属于禾本科(gramineae)黍属(panicum)的c4多年生草本植物。其产量稳定、适应性强、地理分布广且不占用耕地,既是重要的牧草来源也是理想的第二代能源植物。近年来,柳枝稷的基因组数据、转录组数据以及相关芯片数据已陆续释放,同时,柳枝稷的遗传转化体系也已经成熟稳定,因此柳枝稷已经成为能源草和禾本科牧草研究的理想模式植物。因此,解析柳枝稷木质素代谢的调控机制,为分子育种提供新的靶向位点具有重要的现实意义。
一碳代谢指的是含有一个碳原子的基团(如甲基、甲烯基、甲炔基、甲酰基等)的生成、转变、运输以及参与物质合成的反应过程。主要包括两个循环:四氢叶酸循环(thf)和甲硫氨酸循环(met)。一碳代谢合成的甲硫氨酸可以进一步生成甲基活性供体s-腺苷甲硫氨酸(s-adenosylmethionine,sam)。sam为木质素甲基化反应提供甲基供体,最终生成甲基化程度不同的g型和s型单体,从而将一碳代谢和木质素代谢偶联起来。s-腺苷高半胱氨酸(s-adenosylhomocysteine,sah)是sam的去甲基化形式,研究发现sah能够与sam竞争性结合甲基化酶,从而抑制甲基化反应的进行(keatingetal.,evidenceofbrainmethyltransferaseinhibitionandearlybraininvolvmentinhiv-positivepatients,1991,lancet,337(8747):935-9;nguyenetal.,hypodipsichypernatremiaanddiabetesinsipidusfollowinganteriorcommunicatingarteryaneurysmclipping:diagnosticandtherapeuticchallengesintheamnesticrehabilitationpatient,braininj,15(11):975-80)。sahh对于维持植物体内sah的水平具有重要作用(hansonetal.,one-carbonmetabolisminhigherplants,2001,annualreviewofplantphysiologyandplantmolecularbiology,52:119-137)。拟南芥中sahh1突变体表现出明显的发育表型(rochaetal.,thearabidopsishomology-dependentgenesilencing1genecodesforans-adenosyl-l-homocysteinehydrolaserequiredfordnamethylation-dependentgenesilencing,plantcell,17(2):404-417),但是sahh对木质素合成的影响目前尚未有报道。
木质素合成途径中单体的甲基化反应是由咖啡酰-o-甲基转移酶(ccoaomt)和咖啡酸-o-甲基转移酶(comt)催化进行的,而甲基基团是由一碳代谢的产物sam来提供。研究报道,改变玉米四氢叶酸运输循环中的亚甲基四氢叶酸还原酶(mthfr)和拟南芥中叶酰多谷氨酸合成酶(fpgs1)编码基因的表达量能够显著影响木质素单体累积(tangetal.,themaizebrownmidrib2(bm2)geneencodesamethylenetetrahydrofolatereductasethatcontributestoligninaccumulation.plantjournal.2014,77(3):380-392;zhouetal.,folatepolyglutamylationisinvolvedinchromatinsilencingbymaintainingglobaldnamethylationandhistoneh3k9dimethylationinarabidopsis.plantcell.2013,25(7):2545-2559)。但是甲硫氨酸途径对木质素的影响,目前尚未有相关报道。本发明主要针对柳枝稷的sahh1,通过干扰沉默技术获得pvsahh在体内表达抑制的转基因柳枝稷,研究表明sah显著升高,甲基化木质素单体显著降低,最终使得细胞壁降解效率显著升高,这为深入理解柳枝稷和其他禾本科植物木质素调控机理奠定理论基础,同时也为采用基因工程手段改变木质素提供了新的靶基因源和方向指导。
技术实现要素:
本发明的第一个目的是提供一种柳枝稷一碳代谢s-腺苷高半胱氨酸水解酶pvsahh1的编码基因;其核苷酸序列和氨基酸序列分别如seqidno.1和seqidno.2所示。考虑到密码子的简并性,在不改变氨基酸序列的前提下,对上述核苷酸序列进行同义突变,也属于本发明的保护范围内。
本发明的第二个目的提供一种针对柳枝稷一碳代谢酶类基因pvsahh1调控区片段,其核苷酸序列如seqidno.3所示。利用此片段能够显著降低甲基化木质素单体g型和s型的含量,增加细胞壁降解效率。
本发明的第三个目的提供一种柳枝稷一碳代谢酶类基因pvsahh1的干扰沉默表达载体,该表达载体克隆上述seqidno.3片段。
本发明的第四个目的是提供柳枝稷一碳代谢酶类基因pvsahh1对调控甲基活性供体sam以及其去甲基化形式sah方面的应用。
本发明的第五个目的是提供柳枝稷一碳代谢酶类基因pvsahh1在调控柳枝稷木质素代谢方面的应用。
本发明的第六个目的是提供柳枝稷一碳代谢酶类基因pvsahh1在提高细胞壁降解效率方面的应用。
所述柳枝稷pvsahh1的调控区片段,基于gateway技术重组整合到超表达载体panic8b中;采用农杆菌agl1介导的遗传转化方法,将干扰载体导入柳枝稷低地型品种alamo的胚性愈伤细胞中,通过潮霉素抗性筛选,获得抗性再生植株,并通过pcr分析最终确定阳性转基因植株;代谢物测定数据表明转基因柳枝稷中sah水平显著升高,sam/sah比值显著降低;木质素数据表明甲基化木质素单体g型和s型显著降低;细胞壁降解效率显著升高。
本发明的核心特点和发明理念包括:
1、本发明针对柳枝稷木质素这一制约生物能源转化的主要限制因子,通过先进的基因工程技术对其进行调控,能够获得显著效果。
2、本发明探索的pvsahh1通过改变sah以及sam/sah在体内的比例影响了下游木质素的积累,拓宽了对木质素调控机理的认识和研究。
本发明的有益效果如下:
1.本发明中获得的柳枝稷一碳代谢酶类基因pvsahh1,是调控木质素单体成分的关键基因,这对于通过分子定向设计获得利于能源转化的柳枝稷植株具有重要贡献;
2.本发明中对pvsahh1进行分子调控,能够降低木质素单体含量,提高细胞壁降解效率,对于能源植物和禾本科牧草糖化效率的改变具有重要的参考意义;
3.本发明中所产生的遗传改良植物能够整合到常规育种项目,从而为能源植物和禾本科牧草作物的品种培育提供新的种质资源。
附图说明
图1柳枝稷panic8b-pvsahh-rnai干扰载体简图;
图2转pvsahh-rnai基因柳枝稷植株的pcr鉴定;
图3转pvsahh-rnai基因柳枝稷植株的qrt-pcr分析;
图4pvsahhri转基因植株代谢物sam/sah测定;
图5pvsahhri转基因植株木质素单体测定;
图6pvsahhri转基因植株细胞壁降解效率测定。
具体实施方式
下面以针对pvsahh的干扰沉默降低木质素甲基化单体,提高细胞壁降解效率为实施例对本发明的原理和特征进行描述,所举实例只用于解释本发明,并非用于限定发明的范围。下述实施例中所用的材料、试剂、双元载体和农杆菌等,如无特殊说明,均可从公司通过商业途径购买,ms培养基来自phytotechnologylaboratories(货号m519)。
实施例1:pvsahh基因的克隆与序列测定
取柳枝稷alamo的嫩茎部位,用trizolreagent(invitrogen公司,货号15596026)提取嫩茎总rna,使用琼脂糖凝胶电泳和紫外分光光度计检测总rna的含量和纯度,取2.0μg总rna做逆转录反应,所采用的逆转录酶为m-mlv(promega公司,货号m1701),逆转录反应的步骤参考该逆转录酶的使用说明。以逆转录反应合成的第一链cdna为模板,使用引物:
5’-actcccaatccaatccgcgag-3’
5’-ggtgttctagttgtgctgggc-3’
进行常规pcr扩增;pcr反应体系为:2μlcdna,5μl10×buffer,4μldntp(2.5mm),正/反向引物(10μm)各1μl,0.5μltaq酶(5u/μl)和36.5μlddh2o。在冰上加样后混匀。pcr反应条件为:94oc5min;94oc30sec,56oc30sec;72oc90sec,32个循环;72oc10min。pcr扩增获得约1.4kb的片段。琼脂糖凝胶电泳回收该片段后,于pmd-18t载体(takara公司,货号6011)进行连接反应,反应体系为:4.5μlpcr产物,0.5μlpmd-18t载体和5μlsolutioni,16oc连接过夜。取5μl连接产物,用热激法转化大肠杆菌dh5α感受态细胞,加800μl液体lb培养基,复苏1hr,涂于氨苄抗生素的lb平板上(已涂布x-gal/iptg),37oc培养14hr。挑选白色单克隆菌落,在含氨苄抗生素的液体lb培养基中扩增培养,测序。经测序分析表明,该序列含有一个完整的开放阅读框,全长1458个碱基,序列如seqidno.1所示,其编码蛋白含有485个氨基酸残基;序列如seqidno.2所示。
实施例2:pvsahh基因干扰沉默表达载体构建和遗传转化
本发明所用的入门载体为pgwc,rna干扰表达载体为单子叶表达载体panic系列中的panic8b。通过pcr反应克隆获得pvsahh干扰片段(seqidno.3),使用引物:
5’-ctgtgcccttcaagccctga-3’
5’-ccaagcttgcccaagtggaga-3’
进行pcr扩增,扩增模板为上述pvsahh-pmd18t重组质粒。pcr反应体系为:2μl模板,5μl10×buffer,4μldntp(2.5mm),1μl正/反向引物(10μm)各,0.5μltaq酶(5u/μl)和36.5μlddh2o。pcr反应条件为:94oc5min;94oc30sec,56oc30sec;72oc90sec,32个循环;72oc10min。pcr扩增获得约450bp的片段,片段序列如seqidno.3所示。采用琼脂糖凝胶电泳回收该片段后,连接pgwc载体(invitrogen)。连接反应参考该载体使用说明。取5μl连接产物,用热激法转化大肠杆菌dh5α感受态细胞,加800μl液体lb培养基,复苏1hr,涂于卡那霉素抗性的lb平板上(已涂布x-gal/iptg),37oc培养14hr。挑选白色单克隆菌落,在氯霉素抗性的lb培养基中扩增培养,测序。碱裂解法提取测序正确的重组菌株质粒,ecorv核酸内切酶37oc酶切1hr,使用gateway技术,将重组质粒酶切回收片段转入干扰载体panic8b(图1)。重组反应为:100ng酶切回收片段,50ngpanic8b载体质粒,1μllr酶(invitrogen,货号11791020),然后用ddh2o补足至10μl。16oc连接过夜。取5μl连接产物,用热激法转化大肠杆菌dh5α感受态细胞,加800μl液体lb培养基,复苏1hr,涂于卡那霉素抗性的lb平板上(已涂布x-gal/iptg),37oc培养14hr。挑选白色单克隆菌落,在卡那霉素抗性的lb培养基中扩增培养,测序。碱裂解法提取测序正确的重组菌株质粒,电击法转化农杆菌agl1感受态细胞,涂布于含卡那霉素和利福平的lb平板上,28oc,暗培养2天,使用上述引物检测阳性单克隆,在卡那霉素和利福平抗性的液体lb培养基中扩增培养,用于柳枝稷胚性愈伤的遗传转化。
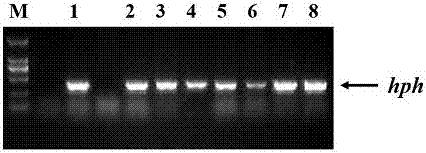
采用农杆菌介导的柳枝稷胚性愈伤的遗传转化方法(xietal,agrobacterium-mediatedtransformationofswitchgrassandinheritanceofthetransgenes.2009,bioenergyresearch,2:275-283)将panic8b-pvsahh-rnai导入低地型野生型柳枝稷alamo,获得抗性苗,经pcr鉴定扩增hph基因最终确定阳性转基因株系(图2)。
实施例3:转基因植株分子鉴定
取上述鉴定转基因阳性植株嫩茎组织,使用trizol(invitrogen公司,货号15596026)法提取总rna,逆转录酶(promega公司,货号m1701)合成第一链cdna,以柳枝稷ubiquitin基因作为内参基因,分别采用引物pvsahh-qf1/qr1检测阳性转基因植株sahh基因的表达情况(图3),引物序列如下:
pvubiquitin-qf:5’-ttcgtggtggccagtaag-3’
pvubiquitin-qr:5’-agagaccagaagacccaggtacag-3’
pvsahh-qf1:5’-gaagttagccacatcgggttatctt-3’
pvsahh-qr1:5’-actcaaccaaactaaaccgctcaag-3’
如图3所示,转基因植株中pvsahh基因的表达量明显低于野生型,说明本发明设计的带有pvsahh区段的干扰表达载体在柳枝稷体内确实发挥了功能。
实施例4:转基因植株代谢物测定
sam为木质素甲基化反应提供甲基生成甲基化程度不同的单体,并转换为去甲基化形式sah,因此为了追踪改变pvsahh基因表达量之后,对这些代谢中间物的影响,我们进一步采用elisa酶联免疫法检测各转基因植株中sam、sah和hcy等的含量变化,elisa酶联免疫试剂盒购自上海百趣生物有限公司。sahhri的三个株系中sah稳定升高(图4a),最终导致sam/sah降低(图4b)。
实施例5:转基因植株的木质素单体测定
木质素单体测定采用脱硫法(thioacidolysis)。当野生型和pvsahhrnai转基因植株生长至r1(reproduction1)时期,取茎节研磨,分别经过烘干、提取木质素以及气相色谱-质谱(gc-ms)分析等步骤,获得不同植株的木质素单体g型、s型和h型的含量,如图5所示,转基因植株中g型、s型木质素单体显著低于野生型。
实施例6:转基因植株的细胞壁降解效率测定
利用苯酚-硫酸比色法测定转基因株系以及对照柳枝稷中细胞壁多糖含量,利用纤维素酶降解细胞壁残基cwr,苯酚-硫酸比色法测定酶解纤维素含量,细胞壁降解效率=酶解纤维素含量/细胞壁多糖含量×100%。结果表明,相比于对照组,pvsahhrnai转基因柳枝稷中细胞壁降解效率显著提高(图6)。
序列表
<110>中国科学院青岛生物能源与过程研究所
<120>一种柳枝稷腺苷高半胱氨酸在改变木质素单体和提高细胞壁降解效率方面的应用
<160>3
<170>siposequencelisting1.0
<210>1
<211>1458
<212>dna
<213>柳枝稷(panicumvirgatuml.)
<400>1
atggcgctctccgtggagaagacctcgtcggggcgggagtacaaggtcaaggacctgtcc60
caggccgacttcggccgcctcgagatcgagctggccgaggtcgagatgcccggactcatg120
gcgtgccgcgccgagttcggcccatccaagcccttcgccggcgccaggatctcgggctcc180
ctccacatgaccatccagaccgccgtcctcatcgagaccctcaccgcgctcggcgccgag240
gtccgctggtgctcctgcaacatcttctccacgcaggaccacgccgccgccgccatcgcg300
cgggactccgccgcggtcttcgcgtggaagggggagacgctcgaggagtactggtggtgc360
accgagcgctgcctcgactggggcgagggcggcggccccgacctcatcgtcgacgacggt420
ggcgacgccacgctcctcatccacgagggcgtcaaggccgaggaggagtacgagaagacc480
ggcaaggtccccgatccggagtccaccgacaacgccgagttcaagatcgtgctcaccatc540
atccgcgacgggatcaaggctgaccccaagaagtaccgcaagatgaaggataggctcgtt600
ggagtctcggaggagaccactaccggagttaagaggctctaccagatgcaggagaccggc660
actctcctcttccccgccatcaacgtcaacgactccgtcaccaagagcaagttcgacaac720
ctgtatggttgccgccactcccttcctgatggtctgatgagggccaccgatgttatgatt780
gctggcaaggttgcggtagtatgcggttatggtgatgttggcaagggctgtgctgctgcc840
ctcaagcaggctggtgcccgtgtcatcgtcactgagattgatcccatctgtgcccttcaa900
gccctgatggagggtcttcaggtccttaccttggaggatgtcgtttctgaagctgacatc960
tttgtgaccaccactggcaacaaggacatcatcatggttgaccacatgaggaagatgaag1020
aacaatgcaattgtctgcaacatcggtcactttgacaatgagattgacatgcatggtctt1080
gagacctaccctggtgtgaaacgcatcaccatcaagcctcagacggaccgctgggtgttc1140
cctgagaccaagactggcatcattgtccttgctgagggtcgtctgatgaaccttggatgc1200
gctactggtcaccccagctttgtcatgtcctgctcattcactaaccaggtcattgctcag1260
cttgagctgtggaaggagaagagctctggcaagtatgagaagaaggtgtacgtgctcccc1320
aagcaccttgatgagaaggttgctgctctccacttgggcaagcttggtgccaagctcacc1380
aagctcaccaagtctcaggccgactacatcagcgtcccgattgagggtccctacaagccc1440
gctcactaccggtactag1458
<210>2
<211>485
<212>prt
<213>柳枝稷(panicumvirgatuml.)
<400>2
metalaleuservalglulysthrserserglyargglutyrlysval
151015
lysaspleuserglnalaasppheglyargleugluilegluleuala
202530
gluvalglumetproglyleumetalacysargalaglupheglypro
354045
serlysprophealaglyalaargileserglyserleuhismetthr
505560
ileglnthralavalleuilegluthrleuthralaleuglyalaglu
65707580
valargtrpcyssercysasnilepheserthrglnasphisalaala
859095
alaalailealaargaspseralaalavalphealatrplysglyglu
100105110
thrleugluglutyrtrptrpcysthrgluargcysleuasptrpgly
115120125
gluglyglyglyproaspleuilevalaspaspglyglyaspalathr
130135140
leuleuilehisgluglyvallysalaglugluglutyrglulysthr
145150155160
glylysvalproaspprogluserthraspasnalagluphelysile
165170175
valleuthrileileargaspglyilelysalaaspprolyslystyr
180185190
arglysmetlysaspargleuvalglyvalserglugluthrthrthr
195200205
glyvallysargleutyrglnmetglngluthrglythrleuleuphe
210215220
proalaileasnvalasnaspservalthrlysserlyspheaspasn
225230235240
leutyrglycysarghisserleuproaspglyleumetargalathr
245250255
aspvalmetilealaglylysvalalavalvalcysglytyrglyasp
260265270
valglylysglycysalaalaalaleulysglnalaglyalaargval
275280285
ilevalthrgluileaspproilecysalaleuglnalaleumetglu
290295300
glyleuglnvalleuthrleugluaspvalvalserglualaaspile
305310315320
phevalthrthrthrglyasnlysaspileilemetvalasphismet
325330335
arglysmetlysasnasnalailevalcysasnileglyhispheasp
340345350
asngluileaspmethisglyleugluthrtyrproglyvallysarg
355360365
ilethrilelysproglnthraspargtrpvalpheprogluthrlys
370375380
thrglyileilevalleualagluglyargleumetasnleuglycys
385390395400
alathrglyhisproserphevalmetsercysserphethrasngln
405410415
valilealaglnleugluleutrplysglulysserserglylystyr
420425430
glulyslysvaltyrvalleuprolyshisleuaspglulysvalala
435440445
alaleuhisleuglylysleuglyalalysleuthrlysleuthrlys
450455460
serglnalaasptyrileservalproilegluglyprotyrlyspro
465470475480
alahistyrargtyr
485
<210>3
<211>480
<212>dna
<213>柳枝稷(panicumvirgatuml.)
<400>3
ctgtgcccttcaagccctgatggagggtcttcaggtccttaccttggaggatgtcgtttc60
tgaagctgacatctttgtgaccaccactggcaacaaggacatcatcatggttgaccacat120
gaggaagatgaagaacaatgcaattgtctgcaacatcggtcactttgacaatgagattga180
catgcatggtcttgagacctaccctggtgtgaaacgcatcaccatcaagcctcagacgga240
ccgctgggtgttccctgagaccaagactggcatcattgtccttgctgagggtcgtctgat300
gaaccttggatgcgctactggtcaccccagctttgtcatgtcctgctcattcactaacca360
ggtcattgctcagcttgagctgtggaaggagaagagctctggcaagtatgagaagaaggt420
gtacgtgctccccaagcaccttgatgagaaggttgctgctctccacttgggcaagcttgg480
- 还没有人留言评论。精彩留言会获得点赞!