抑制胞吞作用的单链寡核苷酸的制作方法
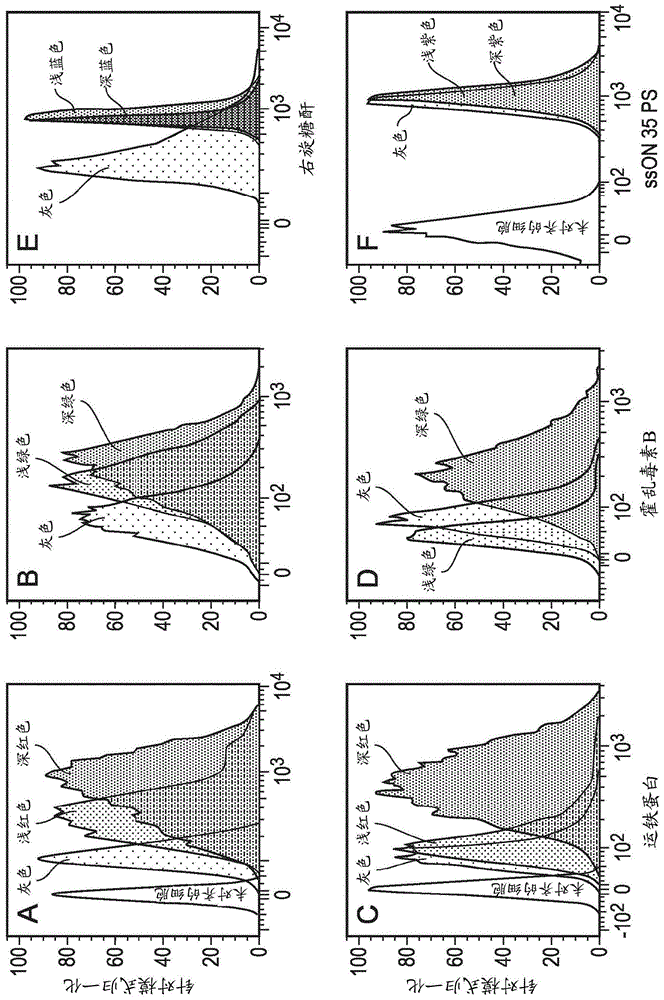
本发明涉及寡核苷酸,并且特别是在预防或治疗流感病毒感染和皮肤和/或皮下组织的紊乱中有用的寡核苷酸。流感病毒(iv)是一种具有单链负义rna基因组的包膜病毒。它是高度传染性的(contagious),频繁全球爆发。该病毒属于正粘病毒(orthomyxovirus)科,并且被分类为三种抗原类型:a、b和c(vandewakker等人,eurjpharmacol,809:178-190)。仅iva和b亚型引起大规模爆发和严重疾病。季节性iv流感影响多达~20%的世界人口,并且每年导致超过250,000-500,000人死亡(vandewakker等人,eurjpharmacol,809:178-190)。由于rna聚合酶的不可靠性,病毒蛋白不断突变。因为遗传漂变和遗传漂移,新的iv病毒不断出现,使得先前获得的免疫力不再适宜且抗病毒药物可能不太有效。由于iv的高突变率,现有疫苗的效力是可变的,并且需要每年开发新的疫苗。目前,有两类抗iv药物被fda批准用于临床使用:m2蛋白抑制剂金刚烷胺和金刚乙胺,以及神经氨酸酶抑制剂奥司他韦和扎那米韦。针对这些药物产生了越来越多的耐药毒株,并且这些药物的使用往往伴随着不利的副作用。因此,对于开发具有新的抗iv作用机制的替代抗iv剂存在迫切需求。寡核苷酸是短的dna或rna分子或寡聚物,其具有广泛的应用。“cpg寡核苷酸”(或cpg-sson)是短的单链的合成dna或rna分子,其包含胞嘧啶三磷酸酯核苷酸(“c”)、胞嘧啶三磷酸酯核苷酸(“c”)之后为鸟嘌呤三磷酸酯核苷酸(“g”)。含非甲基化cpg的核酸刺激免疫系统,并且可以用于治疗感染性疾病、过敏、哮喘和其他紊乱(scheiermann,j.和klinman,d.m.2014.vaccine32:6377-6389;aryan,z.和rezaei,n.2015.curropinallergyclinimmunol.15:568-74)。已经显示ssdna-odn配体中的cpg序列是toll样受体9(tlr9)活化不可或缺的,tlr9在病原体识别和固有免疫活化中发挥重要作用。当cpg重复序列被去除时,配体的刺激作用丧失。缺少cpg基序的单链寡核苷酸(“非cpgsson”)未显示出tlr9介导的免疫刺激作用。单链dna寡核苷酸(sson)在人类单核细胞衍生的树突状细胞和在食蟹猴体内抑制tlr3介导的应答(等人,2012.blood120:768-777)。tlr3是识别双链rna并且启动针对病毒感染的免疫应答的关键受体。等人显示,在用合成的tlr3配体聚肌苷-聚胞苷酸(poly(i:c))处理时,人类单核细胞衍生的树突状细胞上调成熟标志物并分泌促炎性细胞因子。poly(i:c)是针对tlr3的合成激动剂,并且通常被用作疫苗的佐剂(参见例如duthie,m.s.等人,2011.immunol.rev.239(1):178-196)。等人报道,在使用sson的培养物中,tlr3介导的事件被抑制,同样非造血细胞的poly(i:c)活化也被抑制。在sson的存在下,poly(i:c)到细胞中的摄取减少,阻止了tlr3接合的发生。在食蟹猴中,当通过鼻内途径施用sson时,鼻分泌物中促炎性细胞因子的水平减少。因此,根据等人的研究,用sson处理可以导致哺乳动物中细胞因子水平的显著变化。等人使用的sson序列是:5’-gtcgttttgtcgttttgtcgttgttggtggtggtg-3’(cpgsson;seqidno:25);和5’-gaagttttgaggttttgaagttgttggtggtggtg-3’(非cpgsson;seqidno:1)。本发明人已经显示(wo2016/202779)非cpgsson,诸如示出为seqidno:1的非cpgsson,在治疗或预防皮肤和/或皮下组织的紊乱,包括瘙痒症中有用。对皮肤活组织检查物的转录谱分析揭示了猕猴中dsrna(poly(i:c))诱导的促炎性应答的sson依赖性的选择性阻抑。通过对来自皮肤活组织检查物进行离体直接蛋白分析证实了sson调节的细胞因子模式,并且揭示了il-10的诱导和il-6分泌的抑制。这进一步证明,当被施用至哺乳动物时,sson可以产生细胞因子产生的显著差异。baharom,f.等人(j.immunol.194:4422-4430)报道了脂多糖或poly(i:c)对单核细胞衍生的树突状细胞的刺激导致i型ifn诱导并且减少对a型流感病毒感染的易感性。这些结果暗示,能够减少poly(i:c)刺激的作用的sson将增加对病毒感染诸如流感病毒感染的易感性。在临床前模型中已经开发并研究了能够下调免疫系统细胞元件的合成的寡脱氧核苷酸。这些剂在序列、作用机制和细胞靶方面不同,但通常对它们的作用机制缺乏了解(bayik,d.等人,2016.pharmacolres.105:216-225)。例如,已经显示(gursel,i.等人,2003.jimmunol.171(3):1393-1400)哺乳动物端粒中以高频率存在的重复元件,诸如ttaggg,下调了cpg诱导的免疫活化。hanecak等人(us2011/0124715)公开了用于抑制病毒活性的化学修饰的寡核苷酸。该修饰的寡核苷酸具有不多于27个核酸碱基单元,并且包含至少一个gggg序列或至少两个ggg序列。vaillant等人(us2012/0184601)公开了具有抗病毒活性的随机序列寡核苷酸。公开了可以通过sson体外抑制一组病毒,该组病毒限于hsv-1、hsv-2、cmv、柯萨奇b2病毒、dhbv、汉坦病毒、副流感病毒、hiv-1和痘苗病毒。认为对于其中病毒基因组的组装和/或包装和/或包壳是复制中所需步骤的病毒,随机sson具有广谱抗病毒活性。然而,作者还指出,虽然他们可以通过使用sson抑制rsv,但他们未能抑制病毒诸如a型流感病毒(h3n2)、冠状病毒(hcov-oc43)、bvdv、鼻病毒(hgp)和腺病毒(人类ad5)。wo2006/042418中描述了其他sson。总之,在治疗不同病毒感染中有用的单链寡核苷酸的结构和机制仍不清楚。本发明人现已发现sson抑制胞吞作用(endocytosis),并且从而抑制流感病毒感染。如在实施例1中示出的,发现sson抑制人类单核细胞衍生细胞(modc)中的一些胞吞途径,而不影响其他途径。这些研究揭示了细胞外sson在toll样受体(tlr)配体的胞吞摄取中的调节作用,并为它们的免疫调节提供了机制解释。在实施例2中,树突状细胞用sson预处理,并且然后暴露于a型流感病毒(iav)。与未处理的树突状细胞相比,病毒载量明显且高度显著地减少。此外,如在实施例3中示出的,当给予sson作为预防性处理或在流感病毒攻击后三天给予sson作为治疗性处理后,小鼠中的病毒载量显著减少。实施例4显示出,在a型流感病毒感染的小鼠中发生病毒载量减少而细胞因子反应没有显著变化。不希望受任何理论的束缚,本发明人认为抑制胞吞作用的sson将比其他通过直接结合至病毒来阻止细胞融合的寡核苷酸具有更高的抗病毒效力。相应地,本发明的第一方面提供了用于预防或治疗受试者的流感病毒感染的单链寡核苷酸(sson),其中单链寡核苷酸是抑制胞吞作用的寡核苷酸。述及sson,我们包括了任何单链寡核苷酸的含义。寡核苷酸可以是寡脱氧核苷酸(odn)或寡核糖核苷酸或寡脱氧核糖核苷酸。术语寡核苷酸包括核糖核酸(rna)或脱氧核糖核酸(dna)或其模拟物的寡聚物或聚合物。该术语包括包含天然存在的核碱基、糖和共价的核苷(骨架)间键的寡核苷酸,以及具有功能类似的非天然存在的部分的寡核苷酸。这些经修饰或取代的寡核苷酸通常因为期望的特性而优于天然形式,所述期望的特性诸如,例如,增强的细胞摄取、增强的对核酸靶的亲和力和在核酸酶存在下增加的稳定性。以下进一步描述可以使用的修饰的实例。包含骨架和/或其他修饰的寡核苷酸也可以被称为寡核苷酸。胞吞作用是一种主动转运的形式,其中,细胞通过以使用能量的过程吞噬分子(诸如蛋白)来将它们转运到细胞中(内(endo-)+吞排作用(cytosis))。胞吞途径可以被细分为四类:即,“经典的”受体介导的胞吞作用(也被称为网格蛋白介导的胞吞作用)、小窝(caveolae)(也可以是受体介导的,如下文描述的)、巨胞饮作用(macropinocytosis)和吞噬作用。网格蛋白介导的胞吞作用通过产生小的(直径为约100nm)囊泡介导,所述囊泡具有由细胞质蛋白网格蛋白构成的形态特征包被。网格蛋白包被的囊泡(ccv)见于几乎所有细胞中,并且形成被称为网格蛋白有被小窝的质膜的区域。有被小窝可以浓缩具有不同受体的大的细胞外分子,这些受体负责配体的受体介导的胞吞作用,所述配体例如低密度脂蛋白、运铁蛋白(transferrin)、生长因子、抗体和许多其他物质。小窝是最常被报道的非网格蛋白包被的质膜芽,其存在于许多但不是所有细胞类型的表面。它们由胆固醇结合蛋白小窝蛋白(vip21)和富含胆固醇和糖脂的双层组成。小窝是膜中的小的(直径约50nm)瓶状凹陷,类似洞穴形状(因此称为小窝)。它们可以构成一些组织细胞的多达三分之一的质膜面积,在平滑肌细胞、i型肺细胞、成纤维细胞、脂肪细胞和内皮细胞中尤其丰富。还认为细胞外分子的摄取是通过小窝中的受体特异性介导的。胞饮作用(potocytosis)是一种受体介导的胞吞作用的形式,其使用小窝囊泡将多种尺寸的分子带入细胞中。与使用小窝将囊泡内容物递送至溶酶体或其他细胞器的大多数胞吞作用不同,通过胞饮作用胞吞的物质被释放到胞质溶胶中。通常发生于质膜的高度褶皱区域的巨胞饮作用,是细胞膜内陷以形成袋,然后袋掐断到细胞中以形成囊泡(直径为0.5-5μm),囊泡内填充有大量细胞外液和细胞外液中的分子(相当于~100个ccv)。袋的填充以非特异性的方式发生。然后囊泡移动到胞质溶胶中并且与其他囊泡诸如内体和溶酶体融合。吞噬作用是细胞借以结合并且内化直径大于约0.75μm的颗粒物质的过程,颗粒物质诸如小尺寸的灰尘颗粒、细胞碎片、微生物和凋亡细胞。这些过程涉及比网格蛋白介导的胞吞作用和小窝途径更大的膜面积的摄取。哺乳动物细胞的胞吞途径由不同的膜区室组成,这些膜区室将来自质膜的分子内化,并且将其再循环回表面(如在早期内体和再循环内体中),或将其分选为降解(如在晚期内体和溶酶体中)。被抑制的胞吞作用可能是特定胞吞途径或诸如cme和/或cde的途径的胞吞作用,如以下描述的。优选地,被抑制的胞吞作用是cme和cde。应理解,并非所有的胞吞途径都将被抑制。例如,sson通常不影响巨胞饮作用。因此,在优选的实施方案中,sson抑制cde和cme,但不抑制巨胞饮作用。述及抑制胞吞作用的sson,我们包括这样的sson的含义,所述sson使细胞的胞吞作用水平减少到低于没有抑制剂时细胞的胞吞作用水平的水平。例如,sson可以使细胞的胞吞作用水平减少到低于没有sson时细胞的胞吞作用水平的95%或90%,诸如低于没有sson时细胞的胞吞作用水平的80%、70%、60%、50%、40%、30%、20%或10%。优选地,sson使细胞的胞吞作用水平减少到基本上不可检测的水平。应理解,所述细胞是利用胞吞作用以将分子转运到细胞中的细胞。优选地,所述细胞是可能被流感病毒感染的细胞,并且其中病毒利用胞吞作用来感染所述细胞(例如为了避免被细胞质rna传感器识别,所述rna传感器诸如触发高inf-α产生的rig-1和mda/5)。因此,所述细胞可以是流感病毒宿主细胞。流感病毒感染呼吸道上皮细胞、单核细胞和单核细胞衍生的树突状细胞,并且因此优选的是sson抑制选自由以下组成的组的细胞的胞吞作用:单核细胞衍生的树突状细胞、单核细胞和上皮细胞。胞吞作用可能受到抑制的其他细胞包括角质形成细胞和成纤维细胞。测量胞吞作用的方法是本领域熟知的并且可以使用任何合适的方法。通常,这些方法包括:直接标记宿主细胞的胞吞机构(machinery)的组分,诸如早期内体、晚期内体或溶酶体;测量已知依赖于胞吞作用进入细胞的分子的内化;或测量已知其活化依赖于胞吞作用的下游信号传导事件。在直接标记胞吞机构方面,可以通过转基因表达加标记的融合蛋白(例如加荧光标记的融合蛋白诸如gfp)来靶向细胞内的特定结构。例如,已经鉴定出被认为参与胞吞作用的蛋白机构的若干组分,诸如某些rab同种型以及蛋白hrs/stam、tsg101和escrt复合物。标记这样的蛋白可以允许直接监测胞吞机构,并且信号的变化可以被用作为胞吞作用的读出(readout)。应当理解,可以在活细胞中监测胞吞结构,或者可以将细胞固定,例如用于另外的分析,诸如免疫组织化学分析。关于已知依赖于胞吞作用进入细胞的分子的内化,可以使用并且技术人员可以容易地鉴定任何这样的分子,例如通过查询科学文献。合适的胞吞标志物包括低密度脂蛋白(ldl)、运铁蛋白、dsrna、poly(i:c)、白蛋白、破伤风毒素、霍乱毒素亚基b(ctb)和右旋糖酐。应当理解,通过基于感兴趣的途径选择标志物,可以使用不同的标志物来监测不同的胞吞途径。例如,低密度脂蛋白、运铁蛋白、dsrna和poly(i:c)通过网格蛋白介导的胞吞作用(cme)被内化,并且因此可以用于评估cme;白蛋白、破伤风毒素和ctb通过小窝蛋白依赖性胞吞作用(cde)被内化,并且因此可以用于评估cde;以及液相标志物诸如右旋糖酐通过巨胞饮作用被内化,并且因此可以用于评估巨胞饮作用。通常,将分子可检测地标记,使得它们的命运可以被容易地检测。例如,可以用荧光部分、放射性部分、化学发光标记、抗体或生物素、或可检测的酶标记分子。优选地,分子用荧光部分来标记,荧光部分诸如alexa荧光团(647,488等)、texasred、6/5’-fam、dil、pe、apc、percp和fitc。应当理解,当分子被标记后,内化可以通过评估细胞外例如细胞膜处信号的降低(当标记的分子被内化时)和/或细胞内例如胞质溶胶或特定区室诸如内体中信号的增加(当标记的分子被内化时))来监测。相应地,在一种实施方案中,sson是抑制胞吞标志物或分子内化到细胞中的sson,胞吞标志物或分子诸如低密度脂蛋白(ldl)、运铁蛋白、dsrna、poly(i:c)、白蛋白、破伤风毒素和霍乱毒素亚基b(ctb)中的任一种或更多种。如在实施例1中描述的,本发明人证明了本发明的sson抑制运铁蛋白和ctb的内化,并且因此在优选的实施方案中,sson是抑制cme(例如,如通过抑制运铁蛋白的内化证明的)和/或cde(例如,如通过抑制ctb的内化证明的)的sson。本发明人还证明了sson不抑制右旋糖酐的内化,并且因此在优选的实施方案中,sson是不抑制巨胞饮作用的sson(例如,如用右旋糖酐证明的)。流式细胞术是测量分子内化的优选技术,因为数据可以被有效地定量,但是也可以使用其他方法,诸如基于显微术的方法。评估胞吞作用的又另一种方法是通过测量已知其活化依赖于胞吞作用的下游信号传导事件。例如,启动信号传导级联反应的配体可以依赖于胞吞作用以进入细胞,并且因此该信号传导级联反应的报道物可以被用作胞吞作用的读出。如果胞吞作用被抑制,被内化的配体将更少,并且因此通过该信号传导级联反应的信号传导将降低。通常,信号传导级联反应是涉及内体的信号传导级联反应,并且因此可以被认为是“信号传导内体”的下游细胞途径。因此,评估胞吞作用可以包括测量信号传导内体下游的细胞途径。来自内体的细胞信号传导针对受体,诸如notch家族成员、肿瘤坏死因子受体(tnfr)1和一些toll样受体(例如tlr3、tlr7、tlr8和tlr9)发生,并且因此应当理解,可以使用通过notch、tnfr1、tlr3、tlr7、tlr8和tlr9途径中的任一种的信号传导来评估胞吞作用。测量细胞信号传导是本领域的标准实践,并且可以包括测量信号传导级联反应中分子介质(molecularmediator)(例如蛋白)的量和/或信号传导级联反应的产物(例如mrna和/或蛋白)的量。例如,可以定量已知其表达受特定信号传导途径控制的特定蛋白。测定生物样品中的蛋白水平可以使用本领域已知的任何合适的方法来进行。例如,可以通过一系列基于抗体的方法来研究蛋白浓度,包括免疫测定,诸如elisa和放射性免疫测定。在一些信号传导途径中,分子介质的细胞定位可以变化(例如从细胞质到细胞核),并且因此评估信号传导途径的一种或更多种分子介质的细胞定位也可用于评估细胞信号传导,例如使用免疫组织化学。另外,如本领域熟知的,许多信号传导途径控制特定启动子,并且因此对于这些信号传导途径,报道基因可以用于评估信号传导。本领域中有若干种技术可用于检测和测量报道基因的表达,这些技术将适用于本发明。这些技术中的许多种可以以试剂盒获得,用于确定体外和体内的表达。相应地,在另一种实施方案中,sson是抑制下游信号传导事件的sson,已知所述下游信号传导事件的活化依赖于胞吞作用。如实施例1中描述的,本发明人通过使用共刺激分子cd86、cd83和cd80的表达以及促炎性细胞因子il-6的分泌作为读出来评估tlr3信号传导,以测量胞吞作用。因此,在优选的实施方案中,sson是抑制tlr3信号传导的sson。因此,优选地,本发明涉及如本文所定义的sson、方法和/或用途,其中sson是tlr3的抑制剂。因此,在实施方案中,本发明提供的sson是用于预防或治疗受试者的流感病毒感染的tlr3抑制剂,其中单链寡核苷酸是抑制胞吞作用的单链寡核苷酸。已知依赖于胞吞作用的其他tlr是tlr7和tlr8,并且因此在另外的或可选的实施方案中,sson是抑制tlr7和tlr8中的任何一种或两种的sson。tlr4部分地依赖于胞吞作用,因为它具有两种不同的信号传导途径,一种依赖于cme,而另一种来自质膜是myd88依赖性的。因此,sson可以是抑制tlr4信号传导途径的cme介导的信号传导的sson。应当理解,也可以使用其他技术来评估胞吞作用。一种直接的方法是电容技术(capacitancetechnique),该技术测量与囊泡融合和恢复相关的膜表面积的变化(llobet等人,neuron40:1075-1086)。另一种方法涉及用膜染料诸如fm1-43或可检测的融合蛋白(例如gfp融合蛋白)标记的囊泡的荧光成像(llobet等人,neuron40:1075-86)。胞吞作用的实际测量可以利用光的干涉来进行。该方法使用干涉反射显微术(irm),以使当光从与玻璃盖玻片紧密并置的表面膜区域反射时出现的相消干涉区域可视化。(llobet等人,neuron40:1075-86)。其他测定包括生物化学测定,诸如用生物素共价修饰表面受体,以及使用识别特定受体的细胞外结构域的表位的抗体沿着内体途径向下跟踪受体的免疫荧光测定。可以使用任何合适的方法。如在实施例中展示的,本发明人已经证明,sson抑制胞吞作用,并且从而阻断了流感病毒进入细胞的关键途径。不希望受任何理论的束缚,本发明人认为这使得所述sson的抗病毒活性比依赖于直接结合至病毒体来发挥作用的已知抗病毒sson(例如来自wo2006/042418)更有效。应当理解,电子显微术可以用于评估病毒在感染期间的定位,例如通过超延时显微术,并且因此可以用于探测sson是否通过抑制胞吞作用或通过直接结合至病毒来抑制病毒感染。优选的是,用于本文描述的发明上下文中的sson不刺激单核细胞衍生的树突状细胞的固有免疫应答。例如,优选的是sson不刺激一种或更多种固有免疫传感器诸如tlr3、tlr7或tlr9。用于评估sson是否将刺激一种或更多种固有免疫传感器的方法是本领域熟知的,并且包括,例如,测量与固有免疫传感器相关的信号传导途径诸如tlr9信号传导途径。用于评估信号传导途径的活性的方法是本领域熟知的,并且在本文中其他地方描述。也可以通过测量已知与固有免疫应答相关的细胞因子的分泌来评估sson是否刺激一种或更多种免疫传感器。可以使用的技术还包括通过elisa、luminex或细胞内染色测量细胞因子分泌和通过流式细胞术的定量,以及rna定量诸如通过rt-pcr、rnaseq或微阵列。因此,应当理解,用于本发明上下文中的sson优选地是抑制胞吞作用并且不刺激单核细胞衍生的树突状细胞的固有免疫应答的sson。不希望受任何理论的束缚,本发明人认为人类基因组中天然存在的非编码序列会抑制人类细胞的胞吞作用,并且因此本发明基于暂时性关闭胞吞作用的天然机制。通过基于天然序列,寡核苷酸不太可能触发固有免疫应答。然后如下文进一步描述的,这样的天然序列可以被修饰以使其更稳定。在实施方案中,sson的长度在20个和70个核苷酸之间,诸如在25个和70个核苷酸之间。优选的是sson的长度为至少25个核苷酸。因此,本发明提供了用于预防或治疗受试者的流感病毒感染的sson,其中sson是抑制胞吞作用的sson,并且长度为至少25个核苷酸。优选地,sson的长度在25个和70个核苷酸之间,诸如在25个和60个核苷酸之间、25个和50个核苷酸之间、25个和40个核苷酸之间或25个和35个核苷酸之间。在优选的实施方案中,sson的长度在28个和70个核苷酸之间,诸如在28个和60个核苷酸之间、28个和50个核苷酸之间、28个和40个核苷酸之间或28个和35个核苷酸之间。在特别优选的实施方案中,sson的长度在25个和40个核苷酸之间,诸如在25个和39个核苷酸之间,或在28个和40个核苷酸之间。例如,sson的长度可以在29个和38个核苷酸之间或在30个和36个核苷酸之间。sson的长度可以为至少25个、26个、27个、28个、29个、30个、31个、32个、33个、34个、35个、36个、37个、38个、39个或40个核苷酸,和/或sson的长度可以为不多于40个、39个、38个、37个、36个、35个、34个、33个、32个、31个、30个、29个、28个、27个、26个或25个核苷酸。其他优选的长度范围包括30-70个,诸如30-60个、30-50个、30-40个或30-35个核苷酸。进一步优选的范围是25-39个核苷酸。在以下表1中示出的许多sson的长度为35个核苷酸,并且因此在优选的实施方案中,sson的长度为35个核苷酸。优选的是sson中至少50%、60%、70%、80%或90%的核苷酸间键是硫代磷酸酯核苷酸间键。术语“硫代磷酸酯核苷酸间键”和“ps键”是指其中dna磷酸(po)骨架中的非桥氧中的一个被硫取代的核苷酸间键。优选地,根据本发明使用的sson中95%或更优选地100%的核苷酸间键是ps核苷酸间键。因此,本发明包括使用sson,所述sson中一些核苷酸间键(诸如1个、2个、3个或更多个核苷酸间键)是没有硫的po键,而其余键是ps键。在其中sson在5’-末端和/或3’-末端中包含磷酸酯基团的情况下,这样的磷酸酯基团可以是修饰的(ps)或未修饰的(po)基团。应当理解,sson可以包含2’-o-甲基修饰,诸如至少2个、3个、4个、5个或6个2’-o-甲基修饰。述及2’-o-甲基修饰,我们包括了这样的核苷酸修饰的含义,所述核苷酸修饰中甲基基团被添加至核苷的核糖部分的2’-羟基基团。在一种实施方案中,sson包含至少4个硫代磷酸酯核苷酸间键(例如至少5个或6个ps键)和至少4个2’-o-甲基修饰(例如至少5个或6个2’-o-甲基修饰)。在另一种实施方案中,sson包含至少6个硫代磷酸酯核苷酸间键和至少6个2’-o-甲基修饰。根据本发明使用的sson可以包含另外的化学修饰。化学修饰的寡核苷酸是本领域已知的,并且公开于例如jarver,p.等人,2014.nucleicacidtherapeutics24:37-47;和deleavey,g.f.&damha,m.j.2012.chemistry&biology19:937-954中。可能的化学修饰包括例如lna(锁核酸(lockednucleicacid)),其中核糖部分用连接2'氧和4'碳的另外的桥修饰。此外,sson可以包含核糖和脱氧核糖的混合物作为五碳糖。另外,sson中的一个或更多个核碱基可以被修饰。寡核苷酸碱基修饰包括胞嘧啶甲基化以形成5-甲基胞嘧啶,以及腺苷甲基化以形成n6-甲基腺苷。优选的是sson不包含cpg基序。述及cpg基序,我们包括了包含胞嘧啶三磷酸酯脱氧核苷酸(“c”)、胞嘧啶三磷酸酯脱氧核苷酸(“c”)之后为鸟嘌呤三磷酸酯脱氧核苷酸(“g”)的寡核苷酸(例如短单链核酸分子)的含义。“p”是指连续核苷酸之间的磷酸二酯或硫代磷酸酯连接。认为cpg基序是病原体相关分子模式(pamp),因为它们在微生物基因组中丰富,但在脊椎动物基因组中罕见。cpgpamp被模式识别受体(prr)toll样受体9(tlr9)识别,在人类和其他高级灵长类动物中tlr9主要在b细胞和浆细胞样树突状细胞(pdc)中组成型表达。因此,优选地,本发明不包括使用包含能够刺激tlr9反应的cpg基序的sson。优选的是,sson具有“序列非依赖性”作用模式,不具有反义活性,并且不与基因互补。更特别地,优选的是,sson中不多于16个连续核苷酸与任何人类mrna序列(例如人类mrna编码序列)或病毒(例如流感病毒,诸如a型流感病毒)基因互补。优选地,sson基本上与任何人类mrna序列(例如人类mrna编码序列)或病毒(例如流感病毒,诸如a型流感病毒)基因“非互补”。如本领域熟知的,术语“非互补”应被理解为指在中等或严格的杂交条件下(例如低于tm5-10℃)(核酸序列的两条链之间的嘌呤或嘧啶碱基)不能精确配对的核酸序列。例如,优选地sson与编码受体蛋白或识别damp(损伤相关的分子模式)或pamp(病原体相关的分子模式分子)的任何其它蛋白的核苷酸序列不互补,所述受体蛋白例如toll样受体,诸如tlr3或tlr9。因此,应当理解,根据本发明使用的sson不是与细胞内转录的或由病毒(例如流感病毒,诸如a型流感病毒)核酸编码的信使rna(mrna)链互补的“反义”分子。本领域普通技术人员将能够鉴定如根据本发明定义的“非互补”寡核苷酸序列。例如,技术人员可以使用熟知的工具,诸如美国国家生物技术信息中心(usnationalcenterforbiotechnologyinformation)在线执行的blast算法。参见例如,madden,t.2013.theblastsequenceanalysistool.thencbihandbook[互联网]第2版(www.ncbi.nlm.nih.gov/bookinbk153387)。在优选的实施方案中,根据本发明使用的sson不靶向(例如特异性结合至)任何特定病毒(例如流感病毒,诸如a型流感病毒)基因或多肽。如上文提及的,本领域已知可以用特异性靶向病毒编码的分子的寡核苷酸、多核苷酸或适体来治疗病毒感染。例如,arnon等人(us2013/0079390)公开了能够结合至血凝素(流感病毒上的表面抗原)的核酸适配体分子。因此,特别优选的是,sson不靶向(例如特异性结合至)流感(例如a型流感)病毒基因或多肽,诸如神经氨酸酶(na)、血凝素和包膜蛋白诸如m2中的任一种或更多种。寡核苷酸与其他核酸分子和多肽的结合可以使用本领域的常规技术来测试。因此应当理解,根据本发明,sson通常基于其三级结构,例如空间组织,是有用的,而不仅仅依赖于其一级结构,即核苷酸的序列。在又另一种优选实施方案中,根据本发明用于使用的sson不是自身互补的。术语“不是自身互补的”应当被理解为意指sson不具有任何将允许两个sson二聚化或将允许寡核苷酸的部分折叠并与其自身配对以形成茎环的自身互补序列。熟知的是,茎环(也被称为“发夹”环)碱基配对可以出现于单链dna或单链dna中。它发生在同一链的两个区域碱基配对以形成以未配对的环结尾的双螺旋时,所述两个区域在以相反方向读取时通常是互补的。本领域普通技术人员将能够通过比较sson序列的部分并且检测是否能发生沃森-克里克碱基配对(cg和at/au)来鉴定自身互补序列。可选地,可以使用软件工具诸如oligocalc:oligonucleotidepropertiescalculator(www.basic.northwestern.edu/biotools/oligocalc.html)来检测自身互补序列。寡核苷酸中自身二聚化和发夹形成的模型是本领域已知的,并且描述于例如hilbers,c.w.1987.analchem327:70;serra,m.j.1993.nucleicacidsres21:3845-3849;和vallone,p.m.1999.biopolymers.50:425-442中。作为通常规则,自身二聚化将需要至少5个碱基对,而发夹形成将需要至少4个碱基对。因此,优选地,如上文定义的sson不包含可以与同一sson分子内另一3个连续核苷酸的序列形成碱基对的多于3个连续的核苷酸。优选地,所述sson是单链寡脱氧核苷酸(ssodn)。然而,本发明还提供了是稳定化的单链rna(核糖核酸)分子的sson的用途。如本领域技术人员将理解的,当sson是寡脱氧核苷酸时,sson中的单糖是2’-脱氧核糖。因此,sson中的单糖可以选自由2’-脱氧核糖和2’-o-甲基核糖组成的组。在本发明上下文中,术语“ssodn”还包括包含一个或更多个经修饰的单糖诸如2’-o-甲基核糖的寡核苷酸。优选的是根据本发明用于使用的sson不包含ttaggg。优选的是,根据本发明用于使用的sson不包含至少一个gggg序列和/或不包含至少一个ggg序列(例如ttaggg)。还优选的是,根据本发明用于使用的sson包含至少3个不同的选自由a、c、g和t组成的组的核苷酸,或至少4个不同的选自由a、c、g和t组成的组的核苷酸。在这种情况下,应当理解sson不包含单核苷酸重复序列或二核苷酸重复序列,诸如式a(x)、c(x)、g(x)、t(x)、ac(x)、ag(x)、at(x)、cg(x)、ct(x)或gt(x)的重复序列,其中x是整数。在实施方案中,sson中至少30%的核碱基选自a(腺嘌呤)和t(胸腺嘧啶)以及u(尿嘧啶)。优选地,sson中至少35%、40%、45%、50%、55%或60%的核碱基选自a、t和u。当sson是含有脱氧核糖作为其戊糖组分的寡脱氧核苷酸(ssodn)时,核碱基通常选自a和t。当sson是含有核糖的核糖核苷酸时,核碱基通常选自a和u。然而,根据本发明的sson可以包括可以与天然存在的寡核苷酸不同的合成变体。例如,sson可以包含脱氧尿苷部分(即与脱氧核糖结合的尿嘧啶)。sson也可以包含核碱基类似物,核碱基类似物是本领域熟知的并且包括例如黄嘌呤、次黄嘌呤、7-甲基鸟嘌呤、5-甲基胞嘧啶和5-羟基甲基胞嘧啶。在一种特别优选的实施方案中,sson是这样的sson,其中:(a)sson的长度为至少25个核苷酸(例如至少28个核苷酸);(b)(i)所述sson中至少90%的核苷酸间键是硫代磷酸酯核苷酸间键;或(ii)所述sson包含至少4个硫代磷酸酯核苷酸间键和至少4个2’-o-甲基修饰;以及(c)所述sson不包含任何cpg基序。优选地,(i)sson不包含至少一个gggg序列和/或不包含至少一个ggg序列(例如ttaggg);和/或(ii)sson包含至少3个不同的选自由a、c、g和t组成的组的核苷酸,或至少4个不同的选自由a、c、g和t组成的组的核苷酸。在特别优选的实施方案中,sson(例如ssodn)是这样的sson,其中:(a)sson的长度为至少25个核苷酸(例如至少28个核苷酸);(b)(i)sson中至少90%的核苷酸间键是硫代磷酸酯核苷酸间键;或(ii)sson包含至少4个硫代磷酸酯核苷酸间键和至少4个2’-o-甲基修饰;(c)所述sson不包含任何cpg基序;以及(d)(i)sson与任何人类mrna和/或流感病毒基因非互补,和/或(ii)sson不是自身互补的。优选地,(i)sson不包含至少一个gggg序列和/或不包含至少一个ggg序列(例如ttaggg);和/或(ii)sson包含至少3个不同的选自由a、c、g和t组成的组的核苷酸,或至少4个不同的选自由a、c、g和t组成的组的核苷酸。在特别优选的实施方案中,sson(例如ssodn)是这样的sson,其中:(a)sson的长度为至少25个核苷酸(例如至少28个核苷酸);(b)(i)所述sson中至少90%的核苷酸间键是硫代磷酸酯核苷酸间键;或(ii)所述sson包含至少4个硫代磷酸酯核苷酸间键和至少4个2’-o-甲基修饰;并且(c)所述sson不包含任何cpg基序;以及(d)sson不靶向(例如特异性结合至)任何特定病毒(例如流感病毒,诸如a型流感病毒)基因或多肽。优选地,(i)sson不包含至少一个gggg序列和/或不包含至少一个ggg序列(例如ttaggg);和/或(ii)sson包含至少3个不同的选自由a、c、g和t组成的组的核苷酸,或至少4个不同的选自由a、c、g和t组成的组的核苷酸。在特别优选的实施方案中,sson(例如ssodn)是这样的sson,其中:(a)sson的长度为至少25个核苷酸(例如至少28个核苷酸);(b)(i)所述sson中至少90%的核苷酸间键是硫代磷酸酯核苷酸间键;或(ii)所述sson包含至少4个硫代磷酸酯核苷酸间键和至少4个2’-o-甲基修饰;并且(c)所述sson不包含任何cpg基序;(d)sson不靶向(例如特异性结合至)任何特定病毒(例如流感病毒,诸如a型流感病毒)基因或多肽;以及(e)(i)sson与任何人类mrna和/或流感病毒基因非互补,和/或(ii)sson不是自身互补的。优选地,(i)sson不包含至少一个gggg序列和/或不包含至少一个ggg序列(例如ttaggg);和/或(ii)sson包含至少3个不同的选自由a、c、g和t组成的组的核苷酸,或至少4个不同的选自由a、c、g和t组成的组的核苷酸。如实施例中描述的,本发明人已经以24种sson的情况举例说明了本发明。因此,在特别优选的实施方案中,sson包含具有表1中列出的seqidno:1-24中的任一序列或其互补序列的多核苷酸或由该多核苷酸组成。在这些序列中,特别优选的sson是seqidno:1、3、4、7、8、10、11、13-22和24。在另一种实施方案中,sson包含具有与表1中列出的seqidno:1-24中的任一序列或其互补序列共有至少60%序列同一性,诸如与表1中列出的seqidno:1-24中的任一序列或其互补序列共有至少65%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%的序列同一性,并且更优选地至少81%、82%、83%、84%、85%、86%、87%、88%、89%或90%的序列同一性,以及甚至更优选地至少91%、92%、93%、94%、95%、96%、97%、98%或99%的序列同一性的序列的多核苷酸或由该多核苷酸组成。同样,在这些序列中,特别优选的sson是seqidno:1、3、4、7、8、10、11、13-22和24。述及术语“序列同一性”,我们包括了定量测量两种核酸序列之间或相同长度的两种核酸序列之间的同源性程度的含义。如果待被比较的两种序列的长度不相等,则它们必须被对齐以给出最佳的可能拟合,允许插入空位,或可选地,在核苷酸序列的末端处截短。可以计算序列同一性,其中ndif是在比对时两种序列中非相同残基的总数,并且其中nref是其中一种序列中的残基数,优选地,通过本文提供的全长参考来计算序列同一性。空位通常被计为特定残基的非同一性。关于本发明涉及核苷酸序列的所有实施方案,一种或更多种序列之间的序列同一性百分比也可以基于比对,使用ncbiblast软件(https://blast.ncbi.nlm.nih.gov/blast.cgi?page_type=blastsearch&blast_spec=blast2seq&link_loc=align2seq)或clustalw软件(http:/www.ebi.ac.uk/clustalw/index.html),例如用默认设置。在clustalw中核苷酸序列比对的默认设置为:比对=3dfull,空位开启10.00,空位延伸0.20,空位分隔距离4,dna权重矩阵:同一性(iub)。在另一种实施方案中,sson包含具有表1中列出的seqidno:1-24中的任一序列或其互补序列的多核苷酸或由该多核苷酸组成,其中多达10个核苷酸被另一核苷酸取代(例如多达2个、3个、4个、5个、6个、7个、8个、9个或10个核苷酸被另一核苷酸取代)。例如,sson可以包含具有表1中列出的seqidno:1-24中的任一序列或其互补序列的多核苷酸或由该多核苷酸组成,其中2-10个之间,诸如3-9个或4-8个核苷酸被另一核苷酸取代。同样,在这些序列中,特别优选的sson是seqidno:1、3、4、7、8、10、11、13-22和24。为了避免疑问,当通过参考以下表1中的任一序列来定义sson时,应当理解,sson不需要包含表1中描述的关于那些序列中的任一个的每一个(eachandevery)修饰。优选地,当通过参考以下表1中的任一序列来定义sson时,它们确实包含表1中描述的关于任一序列的每一个修饰,然而,这不是必需的。sson可以简单地通过参考表1中描述的特定序列的核苷酸序列来定义,而没有表1中描述的任何化学修饰或具有与表1中描述的化学修饰不同的化学修饰。本发明的第一方面包括sson在制备用于预防或治疗受试者的流感病毒感染的药物中的用途,其中所述sson是抑制胞吞作用的sson。本发明的第一方面还包括用于预防或治疗受试者的流感病毒感染的方法,所述方法包括向所述受试者施用sson,其中所述sson是抑制胞吞作用的sson。在这两种情况下,sson的优选项包括以上描述的那些。特别优选的sson是seqidno:1、3、4、7、8、10、11、13-22和24。为了避免疑问,应当理解,在本文描述的本发明的任何方面,sson不包括通过引用并入本文的wo2006/042418的表21中提及的任何sson(参见第127-129c页,例如rep2006、rep2018、rep2029、rep2028、rep2033、rep2055、rep2056和rep2057),并且不包括通过引用并入本文的us2011/0124715的表1和/或表7中提及的任何sson(参见第6页、第7页、第15页和第16页)。在优选的实施方案中,流感病毒是a型流感病毒(例如h1n1亚型)。然而,应当理解,本发明也适用于b型流感病毒和c型流感病毒,并且因此在另一种实施方案中,流感病毒是b型流感病毒或c型流感病毒或它们的亚型。还应当理解,流感病毒优选地依赖于胞吞作用(例如cde和/或cme)以进入细胞。用于评估流感病毒的胞吞途径的方法包括分子生物学和光学显微术技术,并且在lakadamyali等人,2004(microbesinfect6(1):929)和matlin等人,1981(jcellbiol91(3):601)中有描述。述及预防或治疗流感病毒感染,我们包括了这样的含义,即减少或减轻患者的症状(即缓解性使用)、防止症状恶化或进展、治疗紊乱(例如通过抑制或消除致病病毒)或预防无状况或紊乱的受试者的状况或紊乱。应当理解,在本发明的这种治疗方面,所述方法或用途可以包括鉴定患有或可能患有流感病毒感染或没有流感病毒感染或处于发展流感病毒感染风险(例如在流行或大流行期间)的受试者,向受试者施用根据本发明的第一方面的sson,并且通过进行测试以确定受试者的病毒载量或通过监测受试者的临床症状来监测受试者中的流感病毒水平。取决于监测步骤的结果,可能需要施用更多的sson。述及病毒载量,也称为病毒负荷、病毒滴度(titre)或病毒滴度(titer),我们包括了给定体积中病毒的量的数值表示的含义。它通常取决于测定的类型而被表示为病毒颗粒或感染性颗粒/ml。受试者中的病毒载量可以根据本领域已知的任何合适的方法来确定,并且通常包括核酸测试。例如,可以从受试者获得样品(例如肺液或血液样品),并且通过pcr扩增病毒核酸,或者可以添加对病毒核酸特异性的探针,并且在洗涤步骤之后进行扩增或检测。这样的测试的进一步的细节和实例在以下实施例中提供。还应当理解,本发明有助于选择最合适的sson以向患者施用,例如在病毒流行或大流行期间。在这种情况下,可以测试sson(或一系列sson)针对所讨论的病毒毒株(例如从患者分离的特定流感病毒毒株,例如在流行或大流行期间)的效力。通常,这将在体外或在动物模型中被进行,由此用病毒感染体外细胞或动物模型,并且监测一种或更多种sson对病毒载量的影响。然后,在流行或大流行期间,最有效的sson可以用于治疗患者或预防一个或更多个受试者的流感病毒感染。在以下实施例中详细描述了涉及人类单核细胞衍生的树突状细胞的合适的体外测试。优选地,在本文描述的本发明的任何方面的上下文中,待治疗的受试者是人类。可选地,受试者可以是动物,例如家养动物(例如狗或猫)、实验动物(例如实验啮齿动物,例如鼬、小鼠、大鼠、兔、猴、猪、羊或鸟)、农业上重要的动物(即家畜),例如马、牛、绵羊、猪(pig)、羊、鸡、猪(swine)或山羊、鸟或非人类灵长类动物(例如猴)。应当理解,可以在实验室环境中或在野生环境中向非人类灵长类动物施用sson。在一种实施方案中,sson作为单一疗法被施用。在另一种实施方案中,sson与另外的治疗剂组合施用。因此,本发明提供了用于预防或治疗受试者的流感病毒感染的sson,其中sson是抑制胞吞作用的sson,并且其中受试者被施用另外的治疗剂。应当理解,sson和另外的治疗剂不必是同一组合物的一部分,而是可以如下文另外讨论的那样单独施用。类似地,本发明包括sson在制备用于预防或治疗受试者的流感病毒感染的药物中的用途,其中sson是抑制胞吞作用的sson,并且其中受试者被施用另外的治疗剂。同样,本发明包括预防或治疗受试者的流感病毒感染的方法,所述方法包括向受试者施用抑制胞吞作用的sson和另外的治疗剂。sson和流感病毒的优选项如上文描述的。在一种实施方案中,将sson和另外的治疗剂基本上同时向受试者施用。在可选的实施方案中,将sson和另外的治疗剂依次向受试者施用。因此,个体可以是已经被施用了sson的个体,并且随后将另外的治疗剂向受试者施用。可选地,受试者可以是已经被施用了另外的治疗剂的受试者,并且随后将sson施用至该个体。应当理解,受试者可以是罹患哮喘的受试者,并且因此可能期望也向受试者施用类固醇、β-2-激动剂、白三烯拮抗剂或抗组胺药。因此,在一种实施方案中,另外的治疗剂是类固醇、β-2-激动剂、白三烯拮抗剂或抗组胺药中的一种或更多种。类似地,应当理解,受试者可能是罹患发热的受试者,并且因此可能期望向受试者施用非类固醇抗炎药(nsaid)或扑热息痛。因此,在另一种实施方案中,另外的治疗剂是非类固醇抗炎药或扑热息痛中的一种或更多种。另外的治疗剂可以是另外的抗病毒剂。术语“抗病毒剂”包括减少或阻断病毒的功能或生长或引起病毒破坏的任何物质的含义。例如,可以使用靶向病毒生命周期中的不同步骤的其他抗病毒剂,诸如神经氨酸酶抑制剂、rna聚合酶抑制剂、酸性聚合酶(pa)抑制剂、内切核酸酶抑制剂、聚合酶碱性蛋白2(pb2)抑制剂、α-葡萄糖苷酶抑制剂、靶向血凝素或m2的治疗,例如治疗性抗体(包括抗体结合片段)、基于干扰素α或β的产品。抗病毒剂的特定实例是阿昔洛韦、喷昔洛韦、拉米夫定、利巴韦林、扎那米韦、拉那米韦、帕拉米韦、碘苷、奥司他韦、roflual、金刚烷胺、金刚乙胺(remantidine)、瑞乐沙、gr-121167、gc167、neu5ac2en、二盐酸组胺、帕拉米韦或胸腺法新。本文列出的任何一种或更多种抗病毒剂可以构成本发明上下文中的另外的治疗剂。在一种实施方案中,不向受试者施用dsrna类似物诸如poly(i:c)。通常,施用的途径选自肠胃外、肌内、皮下、表皮、皮内、腹膜内、静脉内、粘膜递送、口服、舌下、皮肤、透皮、局部(topical)、含服、吸入、鼻内、气雾剂、眼内、气管内、直肠内、阴道、基因枪、皮肤贴剂、滴眼剂或漱口水。期望地,所述sson可溶于水,并且在局部递送(localdelivery)至粘膜表面后具有效果。可以将sson以从约1μg/kg至10mg/kg体重;优选地从约1μg/kg至约1mg/kg;并且更优选地从约1μg/kg至约100μg/kg的量全身施用。其他优选的范围包括从约10μg/kg至约10mg/kg;从约10μg/kg至约1mg/kg;和从约10μg/kg至约100μg/kg体重。本发明人已经鉴定出抑制胞吞作用的新的sson,并且因此其在预防或治疗流感病毒感染中有用。相应地,本发明的第二方面提供了sson,所述sson包含具有与表1中列出的seqidno:13-21中的任一序列或其互补序列共有至少60%序列同一性,诸如与表1中列出的seqidno:13-21中的任一序列或其互补序列共有至少65%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%的序列同一性,并且更优选地至少81%、82%、83%、84%、85%、86%、87%、88%、89%或90%的序列同一性,以及甚至更优选地至少91%、92%、93%、94%、95%、96%、97%、98%或99%的序列同一性的序列的多核苷酸或由该多核苷酸组成。在优选的实施方案中,sson包含具有与表1中列出的seqidno:13-21中的任一序列或其互补序列共有至少85%序列同一性,并且更优选地与表1中列出的seqidno:13-21中的任一序列或其互补序列共有90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的序列同一性的序列的多核苷酸或由该多核苷酸组成。应当理解,本发明的这方面包括sson,所述sson包含具有表1中列出的seqidno:13-21中的任一序列或其互补序列的多核苷酸或由该多核苷酸组成,其中多达10个核苷酸被另一核苷酸取代(例如多达2个、3个、4个、5个、6个、7个、8个、9个或10个核苷酸被另一核苷酸取代)。例如,sson可以包含具有表1中列出的seqidno:13-21中的任一序列或其互补序列的多核苷酸或由该多核苷酸组成,其中2-10个之间,诸如3-9个或4-8个核苷酸被另一核苷酸取代。最优选地,本发明的第二方面的sson包含具有表1中列出的seqidno:13-21中的任一序列的多核苷酸或由该多核苷酸组成。本发明的第二方面的sson的长度可以在20个和70个核苷酸之间,诸如在25个和70个核苷酸之间。优选的是sson的长度为至少25个核苷酸。优选地,sson的长度在25个和70个核苷酸之间,诸如在25个和60个核苷酸之间、25个和50个核苷酸之间、25个和40个核苷酸之间或25个和35个核苷酸之间。优选地,sson的长度在28个和70个核苷酸之间,诸如在28个和60个核苷酸之间、28个和50个核苷酸之间、28个和40个核苷酸之间或28个和35个核苷酸之间。在特别优选的实施方案中,sson的长度在25个和40个核苷酸之间,或在28个和40个核苷酸之间。例如,sson的长度可以在29个和38个核苷酸之间或在30个和36个核苷酸之间。sson的长度可以为至少25个、26个、27个、28个、29个、30个、31个、32个、33个、34个、35个、36个、37个、38个、39个或40个核苷酸,和/或sson的长度可以为不多于40个、39个、38个、37个、36个、35个、34个、33个、32个、31个、30个、29个、28个、27个、26个或25个核苷酸。其他优选的长度范围包括30-70个,诸如30-60个、30-50个、30-40个或30-35个核苷酸。在以下表1中示出的seqidno:13-21中的许多sson的长度为35个核苷酸,并且因此在优选的实施方案中,sson的长度为35个核苷酸。如上文描述的,优选的是本发明的第二方面的sson中至少50%、60%、70%、80%或90%的核苷酸间键是硫代磷酸酯核苷酸间键。优选地,sson中95%或更优选地100%的核苷酸间键是ps核苷酸间键。因此,本发明的第二方面的sson可以具有的一些核苷酸间键(诸如1个、2个、3个或更多个核苷酸间键)是没有硫的po键,而其余键是ps键。本发明的第二方面的sson的其他优选项包括上文关于本发明的第一方面所描述的那些。例如,本发明的第二方面的sson可以包含2’-o-甲基修饰和/或其他化学修饰。优选地,sson包含至少4个硫代磷酸酯核苷酸间键(例如至少5个或6个ps键)和至少4个2’-o-甲基修饰(例如至少5个或6个2’-o-甲基修饰)。此外,优选的是本发明的第二方面的sson不包含cpg基序。优选的是,本发明的第二方面的sson具有“序列非依赖性”作用模式,不具有反义活性,并且不与基因互补。更特别地,优选的是,sson中不多于16个连续核苷酸与任何人类mrna序列(例如人类mrna编码序列)或病毒(例如流感病毒,诸如a型流感病毒)基因互补。在实施方案中,本发明的第二方面的sson不靶向(例如特异性结合至)任何特定病毒(例如流感病毒,诸如a型流感病毒)基因或多肽。在又另一种优选的实施方案中,根据本发明的第二方面的sson不是自身互补的。优选地,本发明的第二方面的sson是单链寡脱氧核苷酸(ssodn),但应理解它可以是如上文描述的单链rna(核糖核酸)分子。在实施方案中,本发明的第二方面的sson不包含ttaggg。在实施方案中,本发明的第二方面的sson不包含至少一个gggg序列和/或不包含至少一个ggg序列(例如ttaggg)。优选的是,本发明的第二方面的sson包含至少3个不同的选自由a、c、g和t组成的组的核苷酸,或至少4个不同的选自由a、c、g和t组成的组的核苷酸。在这种情况下,应当理解sson不包含单核苷酸重复序列或二核苷酸重复序列,诸如式a(x)、c(x)、g(x)、t(x)、ac(x)、ag(x)、at(x)、cg(x)、ct(x)或gt(x)的重复序列,其中x是整数。如以下实施例中描述的,本发明人已经证明了sson抑制胞吞作用,并且因此应当理解,本发明的第二方面的sson可以抑制胞吞作用。胞吞作用亚型的优选项,以及如何测量抑制作用,在上文关于本发明的第一方面中进行了描述。在特别优选的实施方案中,本发明的第二方面的sson是这样的sson,其中:(a)sson包含具有与表1中列出的seqidno:13-21中的任一序列或其互补序列共有至少60%序列同一性(优选地100%序列同一性)的序列的多核苷酸或由该多核苷酸组成;(b)(i)sson中至少90%的核苷酸间键是硫代磷酸酯核苷酸间键;或(ii)sson包含至少4个硫代磷酸酯核苷酸间键和至少4个2’-o-甲基修饰;并且(c)sson不包含任何cpg基序。优选地,(i)sson不包含至少一个gggg序列和/或不包含至少一个ggg序列(例如ttaggg);和/或(ii)sson包含至少3个不同的选自由a、c、g和t组成的组的核苷酸,或至少4个不同的选自由a、c、g和t组成的组的核苷酸。在特别优选的实施方案中,本发明的第二方面的sson是这样的sson,其中:(a)sson包含具有与表1中列出的seqidno:13-21中的任一序列或其互补序列共有至少60%序列同一性(优选地100%序列同一性)的序列的多核苷酸或由该多核苷酸组成;(b)(i)sson中至少90%的核苷酸间键是硫代磷酸酯核苷酸间键;或(ii)sson包含至少4个硫代磷酸酯核苷酸间键和至少4个2’-o-甲基修饰;(c)所述sson不包含任何cpg基序;以及(d)(i)sson与任何人类mrna和/或流感病毒基因非互补,和/或(ii)sson不是自身互补的。优选地,(i)sson不包含至少一个gggg序列和/或不包含至少一个ggg序列(例如ttaggg);和/或(ii)sson包含至少3个不同的选自由a、c、g和t组成的组的核苷酸,或至少4个不同的选自由a、c、g和t组成的组的核苷酸。在特别优选的实施方案中,本发明的第二方面的sson是这样的sson,其中:(a)sson包含具有与表1中列出的seqidno:13-21中的任一序列或其互补序列共有至少60%序列同一性(优选地100%序列同一性)的序列的多核苷酸或由该多核苷酸组成;(b)(i)sson中至少90%的核苷酸间键是硫代磷酸酯核苷酸间键;或(ii)sson包含至少4个硫代磷酸酯核苷酸间键和至少4个2’-o-甲基修饰;并且(c)sson不包含任何cpg基序;以及(d)sson不靶向(例如特异性结合至)任何特定病毒(例如流感病毒,诸如a型流感病毒)基因或多肽。优选地,(i)sson不包含至少一个gggg序列和/或不包含至少一个ggg序列(例如ttaggg);和/或(ii)sson包含至少3个不同的选自由a、c、g和t组成的组的核苷酸,或至少4个不同的选自由a、c、g和t组成的组的核苷酸。在特别优选的实施方案中,本发明的第二方面的sson是这样的sson,其中:(a)sson包含具有与表1中列出的seqidno:13-21中的任一序列或其互补序列共有至少60%序列同一性(优选地100%序列同一性)的序列的多核苷酸或由该多核苷酸组成;(b)(i)sson中至少90%的核苷酸间键是硫代磷酸酯核苷酸间键;或(ii)sson包含至少4个硫代磷酸酯核苷酸间键和至少4个2’-o-甲基修饰;并且(c)sson不包含任何cpg基序;(d)sson不靶向(例如特异性结合至)任何特定病毒(例如流感病毒,诸如a型流感病毒)基因或多肽;以及(e)(i)sson与任何人类mrna和/或流感病毒基因非互补,和/或(ii)sson不是自身互补的。优选地,(i)sson不包含至少一个gggg序列和/或不包含至少一个ggg序列(例如ttaggg);和/或(ii)sson包含至少3个不同的选自由a、c、g和t组成的组的核苷酸,或至少4个不同的选自由a、c、g和t组成的组的核苷酸。本发明的第三方面提供了根据本发明的第二方面的sson,所述sson用于在药物中使用。本发明的第四方面提供了包含根据本发明的第二方面的sson和药学上可接受的载体的药物组合物。该药物组合物通常适用于治疗或预防由流感病毒引起的感染。述及药物组合物,我们包括了与常规药物载体和/或赋形剂(例如媒介物)混合并且以水溶液、片剂、胶囊等形式使用的药物活性化合物的含义。任选地,药物组合物可以包含其它药学上可接受的组分,诸如缓冲剂、表面活性剂、抗氧化剂、粘度调节剂、防腐剂等。本发明的第五方面提供了包含根据本发明的第二方面的sson和另外的治疗剂的组合物。合适的另外的治疗剂包括上文关于本发明的第一方面描述的那些。例如,另外的治疗剂可以是另外的抗病毒剂。本发明第五方面的组合物也可以是还包含药学上可接受的载体的药物组合物。本发明人还证明了sson在治疗或预防皮肤和/或皮下组织的疾病或状况方面具有效用。因此,本发明的第六方面提供了根据本发明的第二方面的sson,所述sson用于治疗或预防皮肤和/或皮下组织的疾病或状况。应当理解,本发明的这方面包括根据本发明的第二方面的sson在制备用于治疗或预防皮肤和/或皮下组织的疾病或状况的药物中的用途。类似地,应当理解,本发明的这方面包括预防或治疗皮肤和/或皮下组织的疾病或状况的方法,所述方法包括向受试者施用根据本发明的第二方面的sson。术语皮肤和/或皮下组织的疾病或状况包括icd-10(internationalstatisticalclassificationofdiseasesandrelatedhealthproblems,第10版)中编码的医学状况。这样的状况包括例如皮肤和皮下组织的感染(例如蜂窝组织炎);皮炎和湿疹(例如特应性皮炎和/或瘙痒症);大疱性紊乱(例如天疱疮);丘疹性紊乱(如银屑病);荨麻疹和红斑;皮肤附件紊乱(例如酒渣鼻);或者皮肤和皮下组织的其他紊乱(例如红斑狼疮)。这些实例仅是来自每种类别的说明性的,并不意味着限制本发明的范围。在优选的实施方案中,皮肤和/或皮下组织的疾病或状况包括icd-10,第十二章,l20-l30“dermatitisandeczema(皮炎和湿疹)”,诸如特别是l20“atopicdermatitis(特应性皮炎)”和/或l29“pruritus(瘙痒症)”中编码的医学疾病或状况。因此,特别优选的是,疾病或状况是皮炎和/或湿疹,诸如特应性皮炎或瘙痒症。应当理解,感染可以与皮肤和/或皮下组织的疾病或状况相关。例如,感染可以由皮肤屏障破坏、初始治疗或由免疫系统的变化引起。在这种情况下,可能期望也向受试者施用另外的治疗剂来治疗感染。在一种实施方案中,sson与另外的治疗剂组合施用。因此,本发明提供了根据本发明的第二方面的sson,所述sson用于预防或治疗皮肤和/或皮下组织的疾病或状况,其中受试者被施用另外的治疗剂。应当理解,sson和另外的治疗剂不必是同一组合物的一部分,而是可以如下文另外讨论的那样单独施用。类似地,本发明包括根据本发明的第二方面的sson在制备用于预防或治疗受试者的皮肤和/或皮下组织的疾病或状况的药物中的用途,其中受试者被施用另外的治疗剂。同样,本发明包括预防或治疗受试者的皮肤和/或皮下组织的疾病或状况的方法,所述方法包括向受试者施用根据本发明的第二方面的sson和另外的治疗剂。在一种实施方案中,将sson和另外的治疗剂基本上同时向受试者施用。在可选的实施方案中,将sson和另外的治疗剂依次向受试者施用。因此,个体可以是已经被施用了sson的个体,并且随后将另外的治疗剂向受试者施用。可选地,受试者可以是已经被施用了另外的治疗剂的受试者,并且随后将sson施用至该个体。另外的治疗剂的优选项包括上文关于本发明的第一方面描述的那些。此外,另外的治疗剂可以是已知适于预防或治疗皮肤和/或皮下组织的疾病或状况的治疗剂。另外的治疗剂可以是抗炎剂或抗瘙痒剂。另外的治疗剂的实例包括钙调磷酸酶抑制剂、皮质类固醇、抗il31抑制剂、pde-4抑制剂、il-4r抗体、抗il13抗体、抗il22抗体、抗il12/23抗体、sb011(其裂解gata-3mrna)、去除或遏抑/抑制ige作用的试剂、dp2拮抗剂、神经激肽-1受体拮抗剂、局部非类固醇抗炎试剂诸如le032731和gsk2894512、可乐定、纳曲酮、5-ht2b受体拮抗剂和/或抗组胺药治疗。本发明的第七方面提供了用于鉴定用于治疗或预防流感病毒感染的sson的方法,所述方法包括:(a)提供测试sson;并且(b)评估所述测试sson是否抑制胞吞作用;其中抑制胞吞作用的测试sson是在治疗或预防流感病毒感染中有用的sson。应当理解,测试sson优选地是具有上文描述的关于本发明的第一方面的sson的许多特征的sson。例如,测试sson的长度可以在20个和70个核苷酸之间,诸如在25个和70个核苷酸之间。优选的是,测试sson的长度为至少25个核苷酸。优选地,测试sson的长度在25个和70个核苷酸之间,诸如在25个和60个核苷酸之间、25个和50个核苷酸之间、25个和40个核苷酸之间或25个和35个核苷酸之间。优选地,测试sson的长度在28个和70个核苷酸之间,诸如在28个和60个核苷酸之间、28个和50个核苷酸之间、28个和40个核苷酸之间或28个和35个核苷酸之间。在特别优选的实施方案中,测试sson的长度在25个和40个核苷酸之间,或在28个和40个核苷酸之间。例如,测试sson的长度可以在29个和38个核苷酸之间或在30个和36个核苷酸之间。测试sson的长度可以为至少25个、26个、27个、28个、29个、30个、31个、32个、33个、34个、35个、36个、37个、38个、39个或40个核苷酸,和/或测试sson的长度可以为不多于40个、39个、38个、37个、36个、35个、34个、33个、32个、31个、30个、29个、28个、27个、26个或25个核苷酸。其他优选的长度范围包括30-70个,诸如30-60个、30-50个、30-40个或30-35个核苷酸。应当理解,测试sson可以是以下表1中示出的seqidno:1-24中的任一sson的变体或衍生物。如上文描述的,优选的是测试sson中至少50%、60%、70%、80%或90%的核苷酸间键是硫代磷酸酯核苷酸间键。优选地,测试sson中95%或更优选地100%的核苷酸间键是ps核苷酸间键。因此,测试sson可以具有的一些核苷酸间键(诸如1个、2个、3个或更多个核苷酸间键)是没有硫的po键,而其余键是ps键。本发明的第二方面的测试sson可以包含2’-o-甲基修饰和/或其他化学修饰。优选地,测试sson包含至少4个硫代磷酸酯核苷酸间键(例如至少5个或6个ps键)和至少4个2’-o-甲基修饰(例如至少5个或6个2’-o-甲基修饰)。此外,优选的是,测试sson不包含cpg基序。优选的是,测试sson具有“序列非依赖性”作用模式,不具有反义活性,并且不与基因互补。更特别地,优选的是,sson中不多于16个连续核苷酸与任何人类mrna序列(例如人类mrna编码序列)或病毒(例如流感病毒,诸如a型流感病毒)基因互补。在实施方案中,测试sson不靶向(例如特异性结合至)任何特定病毒(例如流感病毒,诸如a型流感病毒)基因或多肽。在又另一种优选实施例中,测试sson不是自身互补的。期望的是,测试sson不形成任何环结构。优选地,测试sson是单链寡脱氧核苷酸(ssodn),但应理解它可以是如上文描述的单链rna(核糖核酸)分子。在实施方案中,测试sson不包含ttaggg。在实施方案中,测试sson不包含至少一个gggg序列和/或不包含至少一个ggg序列(例如ttaggg)。优选的是,测试sson包含至少3个不同的选自由a、c、g和t组成的组的核苷酸,或至少4个不同的选自由a、c、g和t组成的组的核苷酸。在这种情况下,应当理解sson不包含单核苷酸重复序列或二核苷酸重复序列,诸如式a(x)、c(x)、g(x)、t(x)、ac(x)、ag(x)、at(x)、cg(x)、ct(x)或gt(x)的重复序列,其中x是整数。可以通过本领域已知的任何合适的方法,包括上文关于本发明的第一方面描述的方法,来评估测试sson是否抑制胞吞作用。例如,所述方法可以包括直接标记宿主细胞的胞吞机构的组分,诸如早期内体、晚期内体或溶酶体,并且评估测试sson对胞吞机构的组分的影响。所述方法可以包括测量已知依赖于胞吞作用以进入细胞的分子的内化,并且评估测试sson对这样的内化的影响。所述方法可以包括测量已知依赖于胞吞作用而活化的下游信号传导事件,并且评估测试sson对这样的下游信号传导事件的影响。在实施例中更详细地描述了可以使用的特定方法。应理解,能够高通量操作的筛选测定是特别优选的。测定通常基于细胞。在体外方便地进行测定。例如,可以在体外对人类单核细胞衍生的树突状细胞进行测定。这样的测定在以下实施例中有更详细的描述。可选地,可以在体内进行测定。应当理解,如果测试sson使细胞的胞吞作用水平减少到低于不存在抑制剂时细胞的胞吞作用水平的水平,则测试sson可能是在预防或治疗流感病毒感染(例如a型流感病毒感染)中有用的sson。例如,测试sson可以使细胞的胞吞作用水平减少到低于没有测试sson时细胞的胞吞作用水平的95%或90%,诸如低于没有测试sson时细胞的胞吞作用水平的80%、70%、60%、50%、40%、30%、20%或10%。优选地,测试sson使细胞的胞吞作用水平减少到不可检测的水平。应理解,鉴定抑制胞吞作用的测试sson可以是药物筛选途径中的起始步骤,并且可以进一步选择所鉴定的sson的例如用于预防或治疗流感病毒感染的能力。因此,所述方法还可以包括以下步骤:在流感病毒感染测定(例如基于体外细胞的测定)中测试所述测试sson和/或在流感病毒感染的动物模型(例如小鼠模型)中测试所述测试sson的效力。合适的模型是本领域已知的,并且在以下的实施例中给出了实例。因此,所述方法还可以包括评估已经用流感病毒感染的细胞或动物中的病毒载量的步骤。应当理解,在本发明的第七方面的方法中,上文描述的关于本发明的第一和第二方面的sson可以被用作阳性对照,例如用于帮助优化实验设置。本发明可以包括合成和/或纯化所鉴定的测试sson的另外的步骤。本发明还可以包括将测试sson配制为药学上可接受的组合物的步骤。如本领域技术人员所熟知的,也可以对sson进行其他测试,例如毒理学或代谢测试。本发明包括用于制备在治疗或预防流感病毒感染中有用的sson的方法,所述方法包括使用上文描述的筛选方法鉴定sson,并且合成、纯化和/或配制所鉴定的sson。本发明还包括制备药物组合物的方法,所述方法包括将使用上文描述的方法鉴定的sson与药学上可接受的载体混合的步骤。本说明书中明显在先公布的文献的列表或讨论不必被认为承认该文献是现有技术的一部分或是公知常识。现在将参考以下附图来描述体现本发明某些方面的优选的、非限制性的实例:图1.细胞外sson抑制人类modc中的cme和cde,而不抑制mpc,本图需要更新以便在黑色/白色情况下可理解在+37℃或+4℃,用以下处理人类modc45min:胞吞摄取标志物(运铁蛋白-红色、霍乱毒素b-绿色、右旋糖酐-蓝色和ldl-黄色),添加或不添加sson35;或单独的sson35(紫色)。直方图示出了来自至少三个供体的代表性数据。深着色区域(红色、绿色、黄色、蓝色和紫色)描绘了在+37℃时所指示分子的摄取,而sson的添加用虚线(浅色)描绘。背景,在+4℃进行的实验,被展示为灰色。(a、b)10μmsson35po(medon02)部分抑制了cme和cde依赖性摄取(运铁蛋白、霍乱毒素b的摄取)。(c、d)0.5μmsson35ps(medon01)完全抑制了cme和cde依赖性摄取(运铁蛋白、霍乱毒素b的摄取)。(e)0.5μmsson35ps(medon01)不影响mpc(右旋糖酐)。深蓝色和浅蓝色完全重叠。(f)荧光标记的sson35ps在+37℃和+4℃具有相同的摄取。深紫色和浅紫色完全重叠。(g、h)对在+37℃或+4℃在存在10μmsson35po、0.5μmsson35ps的情况下的运铁蛋白、霍乱毒素b摄取的定量分析。(i)对在+37℃和+4℃的荧光标记的sson35ps摄取的定量分析。(j)0.5μmsson35ps(medon-01)完全抑制了ldl的摄取。以sem给出误差棒。使用非参数mann-whitney检验来比较数据。p值:不显著(n.s)p>0.05;*p≤0.05;**p≤0.01;***p≤0.001。图2.sson抑制了polyi:cpi:c的细胞摄取,并且降低了人类modc的随后成熟(a)0.5μmsson35ps完全抑制了pi:c诱导的cd86表达。(b)sson35ps介导的pi:c暴露的modc中cd86表达的抑制是浓度依赖性的(ec50在100-200nm之间)。(c)sson35ps完全抑制了pi:c诱导的il-6分泌。(d)ps修饰对于抑制作用不是必要的。(e)modc中的荧光pi:c摄取,(f)在0.5μmsson35ps的存在下modc中的荧光pi:c摄取。(g)pi:c诱导的cd86表达的抑制是序列非依赖性的(对于序列,参见表a)。(h、i)pi:c诱导的cd86表达的抑制依赖于sson长度(ssonps15-35)。(j)modc中的荧光sson15ps的摄取,(k)在0.5μmsson35ps的存在下modc中的荧光sson15ps的摄取。(l)荧光sson15ps摄取的抑制依赖于sson35ps浓度。以sem给出误差棒。使用非参数mann-whitney检验来比较数据。p值:不显著(n.s)p>0.05;*p≤0.05;**p≤0.01;***p≤0.00。图3.sson介导的tlr3信号传导的抑制的生物动力学(biokinetics)(a、b)来自暴露于pi:c并且随后用sson35ps攻击的modc在从5min至24h的时间点的cd86表达和il-6分泌。(c、d)以相反的方式处理,即sson35ps并且随后用pi:c攻击的modc。(e)通过添加0.5μmsson35ps逆转了modc中pi:c诱导的细胞死亡(n>25)。(f)互补链(ssoncomplpo)的引入消除了0.5μmsson35ps对pi:c诱导的cd86表达的抑制作用。(g)sson介导的对pbmc中il-6分泌的抑制局限于通过胞吞作用摄取的激动剂。以sem给出误差棒。使用非参数mann-whitney检验来比较数据。p值:不显著(n.s)p>0.05;*p≤0.05;**p≤0.01;***p≤0.001。图4.在具有或没有sson35ps的情况下,暴露于tlr4激动剂lps的modc中的免疫刺激(a)tlr4介导的细胞信号传导的示意图。(b、c)0.5μmsson35ps对nfkb介导的信号传导途径下游的细胞因子/趋化因子(il-6和il-10)的分泌没有影响。(d、e)0.5μmsson35ps减少了trif介导的信号传导途径下游的细胞因子/趋化因子(il-29和cxcl10)的分泌。(f)0.5μmsson35ps抑制了荧光标记的lps的摄取。直方图示出了来自至少三个供体的代表性数据。(g)对在+37℃或+4℃在存在10μmsson35po、0.5μmsson35ps的情况下的lps摄取的定量分析。(h)modc中的荧光lps摄取,(i)在0.5μmsson35ps的存在下modc中的荧光lps摄取。以sem给出误差棒。使用非参数mann-whitney检验来比较数据。p值:不显著(n.s)p>0.05;*p≤0.05;**p≤0.01;***p≤0.001。图5.rna具有与dna相似的抑制人类modc的胞吞摄取的能力在+37℃或+4℃,在添加或不添加sson2’omeps(表a)的情况下,用运铁蛋白(红色)或lps(橙色)处理人类modc45min。直方图示出了来自至少三个供体的代表性数据。(a、b)0.5μmsson2’omeps完全抑制了荧光标记的运铁蛋白和lps的摄取。(c)在阻断pi:c诱导的modc的il-6分泌方面,sson2’omeps展示出与sson35ps相同的效力。(d)总rna以浓度依赖性方式抑制了pi:c诱导的modc的il-6分泌。以sem给出误差棒。使用非参数mann-whitney检验来比较数据。p值:不显著(n.s)p>0.05;*p≤0.05;**p≤0.01;***p≤0.001。图6.在猕猴皮肤中,sson35ps处理局部地阻抑了pi:c介导的炎性特征(a)通过微阵列分析在sson35ps存在或不存在的情况下注射pi:c后皮肤中的rna表达。(b)示出了基因的log2倍数变化和统计显著性的火山图:倍数变化>1或<-1并且未校正的p值<0.05的基因以红色突出显示(b1、b2)。(c)如通过火山图(b1-上图,b2-下图)确定的,将仅pi:c处理或与sson35ps联合处理进行比较时显示出最高的差异表达的基因的聚类。(a、c中)每一行代表一个基因(探针);每一列代表一个样品。将每个基因的表达水平通过减去行平均表达并且除以标准偏差来标准化。将该换算(scaled)表达值(行z-评分)以红色-蓝色颜色标尺绘图,红色表示高表达,而蓝色表示低表达。对基因的无监督层次聚类基于斯皮尔曼相关,并且以平均值±sem示出。(d)刺激后24小时收集的个体猕猴皮肤活组织检查物的相对il-6mrna表达值。(e)通过bio-plex技术测量酶消化的真皮的上清液中存在的il-6蛋白的浓度。从每只动物获得两份皮肤活组织检查物(背部的左侧和右侧),其中一份活组织检查物是在注射pbs后获取的,而另一份活组织检查物是从注射如附图中指示的化合物的部位获取的。通过非参数kruskal-wallis检验和dunn后检验评估显著性差异(*p<0.05,**p<0.01,以及***p<0.001)。使用非参数mann-whitney非配对检验对不同的处理组进行比较,如指示的(虚线箭头)。图7.medon01以剂量依赖性方式抑制人类modc中poly(i:c)的摄取。在+37℃或+4℃,在添加或不添加medon01(浓度范围为500-62.5nm)的情况下,用荧光标记的polyi:c-cy3处理人类modc45min。通过流式细胞术对摄取进行定量(mfi)。对在+37℃存在500-62.5nm的medon01的情况下、在+4℃仅存在对照polyi:c的情况下(基线阴性对照)或在37℃仅存在polyi:c的阳性对照的情况下的polyi:c摄取的定量分析。图8.抑制poly(i:c)摄取的能力是序列非依赖性的在+37℃或+4℃,在添加或不添加medon01-23(浓度为500nm)的情况下,用荧光标记的polyi:c-cy3处理人类modc45min。通过流式细胞术对摄取进行定量(mfi)。对在+37℃存在500nmmedon01-23的情况下、在+4℃仅存在对照polyi:c的情况下(基线阴性对照)或在37℃仅存在polyi:c的阳性对照情况下的polyi:c摄取的定量分析。图9.通过用medon-01预处理抑制a型流感用流感病毒h1n1cal2009(iav)使用moi0.2或moi2体外感染人类单核细胞衍生的dc。在感染前,modc未被处理或用medon-01(0.5μm)预处理2小时。然后在另外的2小时期间用iav进行感染。洗涤细胞并且在再添加或不添加medon-01的情况下孵育24小时(m=培养基)。培养上清液中的病毒载量通过实时pcr来定量。在使用六个不同的供体以一式三份进行分析的四组独立实验中获得数据。使用非参数配对wilcoxon检验进行统计计算。图10.对于抑制a型流感病毒感染的长度要求和化学修饰要求。用流感病毒h1n1cal2009(iav)使用moi1体外感染人类单核细胞衍生的dc。在感染前,modc未被处理或用示出为seqid表1的medon01-21(1μm)预处理2小时。然后在另外的2小时期间用iav进行感染。洗涤细胞并且在再添加或不添加medon的情况下孵育24小时(m=培养基)。培养上清液中的病毒载量通过实时pcr来定量。使用两个不同的供体以一式三份进行分析来获得数据。图11.体内medon-01处理对a型流感病毒载量的抑制。用流感病毒h1n1(cal09)鼻内感染balb/c小鼠。一组未被处理(图11a;圆圈),第二组在病毒攻击的同时鼻内接受25μg的sson(medon01,seqidno1,表1)(图11a;正方形),而第三组在第3天接受25μgsson的鼻内处理(图11a;三角形)。感染后第4天处死小鼠,并且在肺中测量病毒载量。使用单因素anova进行统计计算。未处理的小鼠在第3天具有明显减轻的体重(图11b;圆圈),而接受了用medon-01预防性处理的小鼠能够保持其体重(图11b;正方形)。图12.medon-01处理的a型流感病毒感染的小鼠中tnf-α和il-6的产生。用流感病毒h1n1(cal09)鼻内感染balb/c小鼠。一组未被处理(图12;圆圈)。第二组在病毒攻击的同时鼻内接受25μg的sson(medon01)(图12;正方形)。第三组在第3天接受25μgmedon-01的鼻内处理(图12;三角形)。感染后第4天处死小鼠。通过elisa测量从肺细胞释放的(肺)或支气管肺泡灌洗液中存在的(bal)tnf-α和il-6的水平。使用单因素anova进行统计计算。图13.sson对modc的il-6分泌的作用。在0.5μmmedon不存在或存在的情况下用tlr3激动剂polyi:c(25μg/ml)处理modc,并且在+37℃孵育。处理后24h,收集上清液并且通过使用elisa来评估促炎性细胞因子il-6的分泌。medon-01和medon-13-20能够抑制polyi:c介导的il-6的诱导。图14.sson对cd86的表达的作用。在0.5μmmedon不存在或存在的情况下用tlr3激动剂polyi:c(25μg/ml)处理modc,并且在+37℃孵育。处理后48h,收获细胞并且对免疫成熟标志物cd86染色。通过流式细胞术定量cd86的表面表达。medon-01和medon-13-20能够抑制polyi:c介导的免疫成熟。图15.至少20-25个碱基的sson而不是dsdna抑制tlr3活化。(a)抑制modc中pi:c诱导的cd86表达(48h)不依赖于经典序列(对于序列,参见表2)。(b、c)抑制modc中pi:c诱导的cd86表达(48h)(b)和il-6产生(24h)(c)依赖于sson长度(ssonps15-35)。(d)modc中荧光sson-颜色cy515ps的摄取(e)在0.5μmsson35ps的存在下modc中荧光sson-颜色cy515ps的摄取。(f)荧光sson15ps摄取的抑制依赖于sson35ps浓度。(g)互补链(ssoncomplpo)的引入消除了0.5μmsson35ps对pi:c诱导的cd86表达的抑制作用(48h)。所有数据来自一式两份的至少三个供体。以sem给出误差棒。使用非参数mann-whitney检验来比较数据。p值:不显著(n.s)p>0.05;*p≤0.05;**p≤0.01;***p≤0.001。图16.ssrna展示出与ssdna相似的抑制人类modc的胞吞摄取的能力。在+37℃或+4℃,在添加或不添加sson2’omernaps(表2)的情况下,用tf(alexa647)或lps(alexa488)处理人类modc45min。(a)本研究中使用的寡核苷酸和寡核苷酸类似物的重复单元的结构。(b、c)0.5μmsson2’omernaps抑制了荧光标记的tf和lps的摄取。(d)具有天然po骨架的sson2’omerna能够阻断modc中的tf摄取。(e)在阻断pi:c诱导的modc的il-6分泌方面,sson2’omernaps展示出与sson35dnaps相同的效力。(f)总rna以浓度依赖性方式抑制了pi:c诱导的modc的il-6分泌。所有数据来自一式两份的至少三个供体。以sem给出误差棒。使用非参数mann-whitney检验来比较数据。p值:不显著(n.s)p>0.05;*p≤0.05;**p≤0.01;***p≤0.001。图17.sson35ps抑制pi:c诱导的modc中转录组谱和蛋白质组谱的变化着色的圆圈表示特定类别中包括的基因(绿色=isg,蓝色=炎性体相关的)。(a-c)来自处理后24h的modc的rna-seq数据(3个供体)。ma-图log2倍数变化,低于0.05的经调整的p值为红色。(a)pi:c处理与未处理的细胞相比显示出isg的高的上调。(b)用pi:c和sson35ps(0.5μm)联合处理与未处理的细胞相比显示出微小的差异。(c)sson35ps处理(0.5μm)与未处理的细胞相比显示出微小的差异。(d-f)处理后24h的modc的全细胞蛋白质组学数据(3个供体)。全细胞蛋白质组学数据的ma-图(蛋白log强度相比于平均强度的比率)。(d)pi:c处理与未处理的细胞相比显示出isg的高的上调。(e)用pi:c和sson35ps(0.5μm)联合处理与未处理的细胞相比显示出微小的差异。(f)sson35ps处理(0.5μm)与未处理的细胞相比显示出微小的差异。图18.sson阻抑了猕猴皮肤中dsrna介导的炎性特征(a)在sson35ps存在或不存在的情况下,注射dsrna(pi:c)后皮肤中的rna表达。火山图示出了log2倍数变化并且p<0.05为红色。(b)将pi:c+sson与仅sson35ps处理进行比较时显示出最高的差异表达的基因聚类(b、c)。基因表达被表示为gene-wise标准化的表达(z评分),p<0.05。红色和蓝色分别对应于上调和下调的基因。对基因无监督层次聚类基于斯皮尔曼相关。(c)刺激后24h,来自对个体猕猴皮肤活组织检查物的微阵列分析的相对mrna表达值(平均值±sem)。(d)酶消化的皮肤活组织检查物的上清液中存在的所示细胞因子蛋白的浓度(体内刺激后24h)(来自个体动物的平均值±sem)。右下图示出了范围为每次注射从85-680μg的sson的剂量递增实验中的il-10产生(n=2)。通过非参数kruskal-wallis检验和dunn后检验评估显著性差异(*p<0.05,**p<0.01,以及***p<0.001)。如指示的,使用非参数mann-whitney非配对检验对不同的处理组进行比较。实施例1:细胞外单链寡核苷酸调节胞吞摄取和下游toll样受体反应概述为了维持基本的细胞功能及控制运往内体的分子的摄取,需要严格调节胞吞途径。然而,缺乏对调节胞吞摄取的细胞外分子的了解。在本研究中,我们证明了单链寡核苷酸(sson)抑制人类单核细胞衍生细胞(modc)中的一些胞吞途径,而不影响其他途径。单链dna和rna两者都赋予胞吞抑制。它受浓度和sson长度的影响。sson介导的抑制调节定位于受影响的内体途径中的模式识别受体(prr)的下游细胞信号传导。引入非天然键和/或修饰的核苷对于抑制不是必需的,但是被容许的,并且为了在较长的时间段内保持效果而可能是必要的。我们提供了证据表明,在猕猴中皮内注射后,sson发挥了免疫调节作用。这些研究揭示了细胞外sson在toll样受体(tlr)配体的胞吞摄取中的调节作用,并为它们的免疫调节提供了机制解释。所鉴定的sson介导的胞吞干扰(somie)是暂时性阻抑tlr3/4/7信号传导的调节过程,从而避免过度的免疫应答。引言胞吞机制控制质膜的脂质和蛋白组成,从而调节细胞如何与其周围环境相互作用及交流。为了适当地响应及影响其环境,细胞必须严格地调节质膜组成以及将配体摄取到内体中(doherty和mcmahon,2009)。例如,细胞表面受体的去除使得对特定配体的长期敏感性成为可能(irannejad和zastrow,2014)。然而,胞吞作用不仅负调节与外部环境的相互作用。这些过程还控制基本机制,诸如有丝分裂、细胞迁移和免疫调节过程,诸如启动固有免疫应答和抗原呈递(doherty和mcmahon,2009)。免疫调节源于这样的事实:许多病原体使用胞吞途径来介导其细胞内化(cossart和helenius,2014)。同样,通过胞吞作用摄取从微生物或死亡细胞释放的核酸能够活化定位于内体中的tlr(kawai和akira,2010)。尽管对不同胞吞结构的货物有很多了解,但是这些货物藉以被接合、内化的具体机制以及它们的摄取是如何被调节的,还不太清楚。文献中报道了越来越多的胞吞途径,描述和说明的程度不同。在本研究中,已经评估了细胞外sson是否调节三种明确的途径:网格蛋白介导的胞吞作用(cme)、小窝蛋白依赖性胞吞作用(cde)和巨胞饮作用(mpc)(doherty和mcmahon,2009)。cme由小的(100nm直径)囊泡介导,囊泡具有由网格蛋白构成的包被。网格蛋白包被的囊泡(ccv)见于几乎所有细胞中,并且形成被称为网格蛋白有被小窝的质膜的区域。这些ccv负责从外质膜去除gcpr,减少它们的信号传导,并且使细胞对特定配体去敏感化。此外,这些小窝可以集中负责受体介导的配体胞吞作用的受体,所述配体诸如低密度脂蛋白、运铁蛋白和若干其他配体(kumari等人,2010),包括dsrna及其类似物pi:c(itoh等人,2008)。cde是研究最深入的非网格蛋白包被的胞吞途径之一。它存在于许多细胞类型的表面,并且由小窝蛋白以及富含胆固醇和糖脂的双层组成。小窝是膜中较小的(直径为约50nm)的瓶状凹陷。小窝途径负责配体诸如白蛋白、破伤风毒素和霍乱毒素亚基b(ctb)的胞吞作用(kumari等人,2010)。mpc发生于质膜的高度褶皱区域,然后质膜掐断到细胞中以形成填充有大量细胞外液和分子的较大的囊泡(直径为0.5-5μm)。摄取指示物包括液相标志物,诸如右旋糖酐(doherty和mcmahon,2009)。核酸传感tlr独特地定位于内体并且在体外刺激modc成熟(isomura等人,2005)。tlr3、tlr7、tlr8和tlr9通过识别内体中的微生物核酸来启动对感染的免疫应答。tlr在不同的细胞内区室的这一定位在活化对外来核酸的反应,同时保持对自身核酸的耐受性方面发挥重要作用(roers等人,2016)。tlr信号传导在病毒感染期间构成响应于复制周期期间存在于例如病毒基因组或其中间体中的核酸的防御机制(amarante和watanabe,2010;vercammen等人,2008)。dsrna类似物pi:c经常被用作tlr3激动剂,并且已经证明它依赖于cme以进入胞吞途径(itoh等人,2008)。为了评估细胞外sson是否具有调节胞吞途径的能力,我们在此使用了细胞模型系统,该系统允许使用胞吞标志物(例如运铁蛋白ctb和右旋糖酐)进行基于荧光的摄取研究,以及来自“信号传导内体”的下游细胞途径(scita和difiore,2010;sorkin和zastrow,2009)。来自内体的细胞信号传导针对受体,诸如notch家族成员、肿瘤坏死因子受体1和一些tlr(由sorkin等人综述(sorkin和zastrow,2009))发生。定位于内体中的tlr(tlr3、tlr7、tlr8和tlr9)在复制期间被存在于例如病毒或细菌基因组以及病毒中间体中的on活化(matsumoto等人,2011)。然而,宿主产生的on也具有在从受损细胞或组织释放时启动tlr信号传导的能力(amarante和watanabe,2010;pelka等人,2016)。虽然感应on的tlr存在于特定的内体区室中并且从特定的内体区室发出信号,但感应细菌脂多糖(lps)的tlr4具有两种不同的信号传导途径;一种依赖于myd88的途径从质膜发出信号,以及一种trif依赖性途径依赖于cme并且仅在胞吞作用已经发生后才能发出信号(kagan等人,2008;rajaiah等人,2015;tatematsu等人,2016)。树突状细胞(dc)是抗原呈递细胞,其整合来自环境的刺激,导致免疫活化或诱导并且维持免疫耐受。dc由发育和功能上不同的亚群组成,这些亚群差异性地调节t细胞应答,从而在固有免疫系统和适应性免疫系统之间充当信使(merad等人,2013)。dc亚群存在于与外部环境接触的组织中,诸如皮肤和呼吸系统以及肠的内衬。由于其作为外部环境传感器的作用,dc表现出多种多样的胞吞途径。此外,作为固有免疫系统和适应性免疫系统之间的信使,它们对在胞吞途径中被摄取和加工的若干触发分子作出反应,这些触发分子诸如感应on的tlr。相反,包含源自哺乳动物端粒的重复性ttaggg基序的dna可以抑制免疫应答(gursel等人,2003)。正在临床前模型中研究具有免疫遏抑功能的合成的ss-dna分子,并且它们在尺寸、序列和核苷酸骨架上有所不同,但通常对它们的作用机制缺乏了解(bayik等人,2016)。得到确认的是,on不会被动穿过质膜,并且细胞内递送需要转染试剂或其他膜可通透方法(sharma和watts,2015;silva等人,2015)。然而,认为游离on是通过整联蛋白和/或清道夫受体被摄取的(juliano和carver,2015)。就我们所知,sson与dson之间的细胞摄取是否存在差异尚不清楚。在此,我们比较了具有不同长度和序列的sson的胞吞能力。我们显示长度为35个碱基的sson未被modc胞吞,反而抑制了运铁蛋白、ctb和dsrna类似物聚肌苷-聚胞苷酸(pi:c)的胞吞,但未抑制右旋糖酐的摄取。非天然硫代磷酸酯键(ps)的引入增加了抑制作用,但是天然存在的天然磷酸二酯(po)sson和从细胞纯化的rnapi:c在短期培养物中诱导了il-6产生。我们先前报道了,包含cpg的sson抑制表达tlr3/4/8但缺少tlr7/9的原代人类单核细胞衍生细胞(modc)中的tlr3信号传导。在本研究中,我们合成了一组缺少cpg基序的sson,以鉴定抑制dsrna介导的modc活化的要求(表a)。我们发现sson不仅抑制tlr3活化,而且还抑制tlr7的活化。此外,我们显示sson调节来自依赖于内体摄取的tlr4的信号传导,而不影响来自质膜的信号传导。我们提供的证据表明,某些sson暂时性关闭了cme,而不会对细胞引起重大伤害,如通过存活力测定、细胞因子产生、rnaseq和全细胞蛋白质组学分析证明的。最后,我们证明sson可以在猕猴皮肤中局部地塑造(shape)通过dsrna诱导的免疫应答。这些发现结果显示,细胞外sson可以抑制cme和cde,提供了暂时性关闭货物到内体的摄取、随后改变炎性特征的机制。结果细胞外sson抑制运铁蛋白和ctb的胞吞作用在此,我们研究了细胞外sson是否能够在人类modc中抑制常用的胞吞途径标志物的摄取(不用任何转染方法或递送系统)。在+37℃或+4℃,在添加或不添加0.5μmsson(表a中的序列)的情况下,用胞吞摄取标志物(运铁蛋白、ctb和右旋糖酐以及ldl)处理人类modc细胞45分钟。在+37℃,如通过流式细胞术测量的,运铁蛋白、ctb和右旋糖酐以及ldl被摄取(图1a-d、j)。通过添加0.5μmsson35po显著抑制了运铁蛋白和ctb的荧光信号(图1a、b、g和h)。此外,我们发现通过添加0.5μm修饰的sson35ps完全阻断了运铁蛋白和ctb以及ldl的荧光摄取信号(图1c、d、g、h和j),而mpc保持不受sson35ps处理的影响,如通过添加或不添加0.5μmsson35ps在+37℃对右旋糖酐的相同摄取被证明的(图1e)。在+4℃未发生胞吞作用,显示添加0.5μmsson35ps(在37℃)完全抑制了运铁蛋白和ctb摄取(图1a、b、g和h)。sson35ps本身在+4℃和+37℃显示出相同的摄取,显示荧光标记的sson35ps停留在细胞外,并且未被内化到胞吞途径中(图1f和i)。总之,这些结果显示,天然ssonpo和ssonps都可以显著抑制cme和cde,但mpc保持不受影响,表明sson在货物到内体的摄取中起着调节作用。细胞外sson抑制modc中tlr3配体摄取和随后tlr3活化的能力的长度要求dsrna类似物pi:c是一种通过cme被摄取的tlr3激动剂(itoh等人,2008)。我们接下来提问:sson35ps是否能够阻断pi:c介导的modc的成熟,并且如果能够阻断pi:c介导的modc的成熟,那么其是否通过抑制胞吞摄取而发生(图2)。在处理后48小时,通过测量cd1a阳性细胞中共刺激分子cd86、cd83和cd80的表达(cd83和cd80数据未示出),以及在处理后24小时促炎性细胞因子il-6的分泌,来监测pi:c依赖性modc成熟。我们发现sson以浓度依赖性方式显著阻断pi:c介导的cd86的诱导,ec50在100-200nm之间(图2a、b)。我们还检测到sson35ps显著抑制pi:c介导的modc的il-6分泌(图2c)。与内体标志物运铁蛋白和ctb的摄取类似,对il-6的抑制通过但不依赖于sson的稳定性修饰而增强(图2d)。当使用天然存在的po骨架时,在初始处理后48小时,抑制作用不存在(数据未示出)。然而,当在较早的时间点研究抑制作用时,具有po骨架的sson具有与pssson相同的效力。该作用持续3小时,并且4小时后仍然存在(图2d)。5小时后,该作用完全消失(数据未示出)。显微术研究还证实,通过添加0.5μmsson35ps有效阻断了modc中荧光标记的pi:c的摄取(图2e、f)。然后,我们提问:观察到的胞吞作用的抑制和下游tlr3活化是否可以与特定的sson序列相关。我们比较了三种不同的sson序列(sson35ps、ssongtaps和ssoncomplps;表a),它们在用pi:c处理48小时的modc中全都显示出相同的抑制作用(图2g)。ssongta基于亲本序列sson35,但所有鸟苷(g)碱基都被腺苷(a)取代,而ssoncompl是sson35的互补序列。这三种sson长度全都是35个碱基,并且具有完全由ps取代的骨架。在得出抑制作用不严格依赖于sson序列的结论后,我们接下来评估了sson长度是否会影响效力。我们发现较短的sson不具有与较长的sson相同的抑制作用,sson的长度截止值在20个和25个碱基之间(图2h、i)。显微术研究显示,荧光标记的sson15ps容易被modc摄取,并且通过添加0.5μmsson35ps有效地阻断了这种细胞摄取(图2j、k)。较短的sson(ssonps15)对胞吞摄取的阻断依赖于sson35ps的浓度(图2l)。总之,这些数据证明,sson能够以浓度依赖性的方式抑制tlr3激动剂pi:c的胞吞摄取和下游作用,并且这些作用不与特定的sson序列或基序相关。sson长度确实影响抑制作用,并且具有20-25个碱基之间的截止值。sson介导的对tlr信号传导的抑制的动力学如果细胞外sson能够抑制某些胞吞途径,那么生物动力学应该遵循先前在modc中进行的内体摄取研究(choi等人,2013;smole等人,2015)。因此,我们通过将细胞暴露于tlr3激动剂pi:c或抑制性sson35ps,并且随后在5分钟至24小时的时间段范围内用sson35ps/pi:c脉冲处理细胞,来评估modc成熟的活化和抑制的动力学。共刺激cd86的表达和il-6的分泌被认为是pi:c诱导的modc成熟的标志。将细胞暴露于pi:c,随后用sson35ps脉冲处理,显示出抑制作用存在多达50分钟,并且然后在初始pi:c处理后约2h迅速减少并消失(图3a、b)。以相反的方式处理,即先用sson35ps处理,并且然后用pi:c脉冲处理的modc,在最初始理后多达24小时未显示出成熟或细胞因子产生的迹象(图3c、d)。在暴露于激动剂后,添加sson35ps抑制tlr3活化多达50-60min(图3a)。根据先前的研究,人类modc花费约35-60min来胞吞细胞外化合物(smole等人,2015)。总之,我们的发现表明,在tlr3激动剂被胞吞并且tlr3信号传导的启动已经被启动的情况下,添加sson35ps不能抑制pi:c诱导的modc成熟。然而,在较早的时间点,sson35ps抑制了该过程。sson防止人类modc中pi:c介导的细胞死亡pi:c可以诱导人类细胞凋亡(sun等人,2011)。为了评估sson是否能够防止这种情况,在添加或不添加sson35ps的情况下,用pi:c处理modc,并且通过live/dead细胞染色试剂盒使用流式细胞术测量凋亡。仅用pi:c处理的细胞具有的存活力减少了约20%。通过添加0.5μmsson35ps,在初始处理后48小时,细胞死亡的频率减少回到未处理的细胞的水平(图3e)。pi:c摄取的抑制针对sson而非dson发生我们在此提供的数据证明,细胞外sson抑制胞吞摄取的能力依赖于on长度,而不依赖于其序列(图2)。为了评估dson的引入是否影响胞吞摄取,将sson35ps保持在恒定浓度(0.5μm),同时引入递增浓度的互补链(ssoncompl35po)(图3f)。用pi:c处理modc,并且当引入互补链时,sson的抑制活性(通过初始处理后48小时的cd86表达来监测)减少。当所有sson35ps都具有杂交的互补链时(ssoncomplpo1μm),抑制作用完全消失。sson的抑制作用针对定位于内体中的tlr3和tlr7发生,但不抑制膜可透过的tlr激动剂的作用由于感应sson的tlr依赖于胞吞作用,并且信号传导由信号传导内体发生(sorkin和zastrow,2009),我们接下来评估了sson是否具有阻断由膜可透过的tlr激动剂引起的tlr诱导的modc成熟的能力。r848是一种膜可透过的tlr7/8激动剂,并且因此不依赖于胞吞作用来活化tlr信号传导(govindaraj等人,2011)。cl307是一种与精胺偶联的tlr7激动剂,并且因此其细胞摄取依赖于胞吞作用(soulet等人,2002)。modc缺少感应sson的tlr7的表达,因此这些实验是在外周血单核细胞(pbmc)中进行的。在不存在或存在sson35ps的情况下,用pi:c(tlr3激动剂)、cl307(tlr7激动剂)或r848(膜可透过的tlr7/8激动剂)处理pbmc(图3g)。sson35ps使来自用pi:c和cl307处理的pbmc的il-6分泌显著减少,但在r848刺激后无此现象。这些数据证明,细胞外sson35ps仅在激动剂依赖于胞吞作用来到达其受体时,才具有抑制tlr诱导的促炎性il-6分泌的能力。sson介导的对tlr4信号传导的抑制依赖于内体摄取tlr4对脂多糖刺激有反应,并且具有两种不同的信号传导途径(kagan等人,2008;rajaiah等人,2015;tatematsu等人,2016)。一种通过myd88介导并且发生于质膜,导致nfkb活化。第二种途径通过trif介导,刺激强烈的干扰素应答。trif介导的信号传导发生于网格蛋白包被的内体,并且因此依赖于cme(图4a)。因此,我们推断,添加sson35ps连同tlr4激动剂应主要抑制modc的干扰素应答,因为细胞外sson抑制cme途径(图1)。为了验证这个假设,我们用tlr4激动剂lps和sson35ps同时处理modc。通过il-6和il-10的分泌来评估通过myd88依赖性途径的信号传导,并且通过il-29和cxcl10的分泌来评估trif依赖性途径(rosadini等人,2015)。当modc被暴露于lps和sson35pssson时,myd88依赖性途径下游的il-6和il-10的分泌保持不受影响(图4b和c),但我们发现cxcl10和il-29受到显著抑制(图4d和4e)。这证明sson35ps的添加选择性地抑制了源自相同tlr的两种途径中的一种。一种是依赖于胞吞作用的,而另一种不是。为了证实lps被摄取到细胞中,并且sson抑制这种摄取,我们将modc暴露于alexa-488标记的lps,并且通过流式细胞术对摄取定量。我们发现在37℃孵育后与4℃相比荧光信号显著增加,并且此外,在37℃,在sson35ps的存在下,lps的摄取被显著抑制(图4f和g)。通过显微术证实了lps的摄取,并且还显示出在37℃,在sson的存在下,摄取受到抑制(图4h和i)。用dna和rna两者都可以实现对内体摄取的抑制为了阐明抑制胞吞摄取的能力是否为dna所独有,还是rna也具有这种活性,我们合成了35merssonrna和dna类似物(表a)。一种天然的未修饰的35-merrna对modc的胞吞摄取没有作用(数据未示出)。然而,ondna类似物2’ome和从modc纯化的总rna具有抑制内体摄取的能力(图5a和b)。pi:c诱导的il-6的分泌被sson2’omeps减少到与sson35ps相当的水平(图5c)。使用相同的测定,用低于200b的截止过滤器纯化的总rna(totrna)以浓度依赖性方式减少了il-6的产生(图5d)。这些数据显示,具有抑制人类modc的胞吞摄取的能力的sson可以源自dna或rna。sson35ps的施用调节了猕猴皮肤中dsrna介导的炎症为了评估稳定化的sson35ps是否能在体内介导作用,我们通过对取自注射部位的皮肤活组织检查物进行转录谱分析,测量了sson35ps存在或不存在时对pi:c的整体固有应答。我们发现皮内注射pi:c与注射pbs相比导致了多种固有免疫应答基因的诱导(图6a)。与pbs对照相比,pi:c注射后显著调节的基因(p<0.05)的基因本体(geneontology,go)-注释显示出与go过程“防御应答”(错误发现率(fdr)q值:2.85e-48)和“免疫系统过程”(fdrq值:1.14e-47)相关的基因富集。此外,go功能“细胞因子活性”和“趋化因子活性”被富集(fdrq值:分别为7.95e-14和2.25e-10)。为了更深入地了解皮肤中哪些基因显示出受调节的表达,我们将仅接受皮内注射pi:c的组与接受pi:c/sson的组进行比较,并且计算了这两组之间的倍数变化以及差异的显著性(图6b)。结果显示,将pi:c/sson35ps共施用与仅pi:c处理进行比较,与炎性状况相关的趋化因子和基因在下调最多的基因中(图6c上图)。可以注意到,il-6的表达在下调最多的基因中(负的fc>2p<0.05)(图6d),支持了体外数据(图2c)。我们还通过分析在没有另外体外刺激的情况下收集的过滤的真皮上清液的等分试样,验证了il-6的分泌(图6e)。在pi:c处理后,检测到il-6的显著诱导(p<0.05),在pi:c/sson35ps处理后,il-6的诱导减少。综合起来,我们的数据显示,pi:c在猕猴皮肤中局部诱发了显著的炎性特征,该现象被共施用sson35ps阻抑。实验程序单链寡核苷酸完全ps取代的sson35ps和2’omeps由integrateddnatechnologies制备。其他修饰的寡核苷酸购自eurofins。对于序列,参见表a。使用rneasy总rna纯化试剂盒根据制造商的说明书(qiagen)从modc纯化总rna。tlr配体和标记的内体标志物tlr3配体高分子量pi:c(25μg/ml,除非另有说明)、tlr7配体cl307和tlr7/8配体r848均购自invivogen。内体标志物;运铁蛋白-alexa647、霍乱毒素b-alexa488、右旋糖酐-texasred10kda和lps-alexa488均购自lifetechnologies。modc的摄取研究使用rosettesepmonocyteenrichment试剂盒从血沉棕黄层负选择出人类单核细胞(1ml/10ml血沉棕黄层;stemcelltechnologies),并且在补充有10%fcs、1mm丙酮酸钠、10mmhepes、2mml-谷氨酰胺以及1%链霉素和青霉素的rpmi1640(均来自invitrogenlifetechnologies)中,以5x105个细胞/ml的密度,用gm-csf(250ng/ml;peprotech)和il-4(6.5ng/ml;r&dsystems)处理6天,使人类单核细胞分化为modc。在冰上将modc暴露于完整的10%rpmi培养基(或无血清培养基,用于poon摄取研究)中的内体标志物和sson,并且然后转移至37℃,持续45分钟。孵育后,将细胞放置回冰上,洗涤并且固定,并且通过流式细胞术监测摄取。通过50μg/ml的alexa647标记的运铁蛋白的摄取来监测cme。通过1μg/ml的alexa488标记的霍乱毒素b来监测cde,并且通过0.5mg/ml的texas-red标记的右旋糖酐来监测mpc。通过1μg的alexa488标记的lps来监测tlr4的胞吞摄取。通过定量dil标记的ldl(molecularprobes)的摄取来测量ldl的摄取。modc成熟研究如上文描述的使人类单核细胞分化。将modc暴露于pi:c和sson,并且在48小时后使用靶向cd1a和modc成熟标志物cd86、cd83和cd80的单克隆抗体(ab)(所有ab均来自bdbiosciences)来评估成熟。在fortessa(bdbiosciences)上获得流式细胞术样品数据,并且用flowjo软件(treestar)进行分析。细胞因子/趋化因子分泌研究在处理(pi:c、cl307、r848(invivogen)或lps(sigma))后的特定时间点从细胞收集上清液,并且通过标准elisa根据制造商的说明书(mabtech;il-6、il-10和il-29。lifetechnologies:cxcl10)来测量细胞因子/趋化因子的分泌。通过450nm处的3,3',5,5'-四甲基联苯胺(tmb)吸光度来监测蛋白量,并且以pg/ml或相对吸光度单位给出。宽场显微术使modc在不含酚红的完整rpmi培养基中粘附至聚-l-赖氨酸包被的玻璃载玻片2-4h。在37℃,在sson35ps存在或不存在的情况下,用指示的配体处理细胞45分钟。洗涤掉未结合的配体,并且用完整培养基覆盖细胞,并且在37℃在宽场cellobserver显微镜中使用40x透镜来获得图像。使用slidebook6程序(intelligentimaginginnovations)获得和分析图像。tlr3抑制的动力学分析为了阐明所观察到的tlr3信号传导抑制背后的动力学,将modc暴露于25μg/mlpi:c/0.5μmsson35ps,并且然后分别用0.5μmsson35ps或pi:c脉冲处理从5分钟至24小时的时间段。在初始暴露后48小时,通过流式细胞术分析细胞的modc成熟标志物cd80和cd86的表达。另外,在处理后48小时,从细胞收集上清液,并且通过elisa(mabtech)测量促炎性il-6的分泌。存活力测试在用25μg/mlpi:c/0.5μmsson35ps初始处理后48小时,从总细胞群体中选择存活细胞。使用live/dead可固定近ir死细胞染色试剂盒根据制造商的方案(lifetechnologies)评估存活细胞。样品数据使用fortessa(bdbiosciences)来获得,并且使用flowjo软件(treestar公司)来分析。用dson进行modc成熟研究在pbs中将5μm的sson35ps与1-10μm的互补sson35po在55℃预孵育,并且然后在环境温度留置30分钟。将on混合物以0.5μmsson35ps的终浓度连同25μg/mlpi:c添加至modc。如上文描述的评估modc成熟。动物和注射按照欧洲nhp护理指南(europeanguidelinesfornhpcare)(eudirectiven63/2010)处理成年食蟹猴(猕猴(macacafascicularis))(n=18)。本研究由cea伦理动物委员会(ethicalanimalcommitteeofthecea)“comitéd’ethiqueenexpérimentationanimale”根据12-013号声明批准和认可,该委员会由法国研究部(frenchresearchministry)根据44号注册。在麻醉情况下处理动物,并且在左上和右后侧腹仅用170μgpi:c或用100μlpbs中的170μgsson或仅用pbs进行皮内注射。注射后24小时,从麻醉动物收集皮肤活组织检查物(直径为8mm)。猕猴中的细胞因子分泌测定为了直接离体评估来自猕猴皮肤活组织检查物的细胞因子和趋化因子的产生,在bio-plex设备(bio-rad)上用milliplexmapnhpcytokinemagneticbeadpanel(millipore)根据制造商的说明书收集并测量过滤的真皮上清液的等分试样。微阵列分析使用tissue随后使用rneasyplusuniversal试剂盒(qiagen),根据制造商的说明书,从猕猴皮肤活组织检查物提取总体皮肤rna,在rnalater中在4℃保存至少24小时。在agilent2100生物分析仪上对总rna进行质量检查。使用nanodropnd-1000分光光度计测量rna量。使用quickamplabeling试剂盒(agilent)根据制造商的说明书从200ng总rna制备花菁-3(cy3)标记的crna,随后进行rneasy柱纯化(qiagen,valencia,ca)。用nanodropnd-1000分光光度计检查染料掺入和crna产量。将1.65μgcy3标记的crna在60℃在55μl的包含1xagilent片段化缓冲液和2xagilent封闭剂的反应体积中遵循制造商的说明书片段化30分钟。完成片段化反应后,将55μl的2xagilent杂交缓冲液添加至片段化混合物,并且在旋转的agilent杂交炉中在65℃杂交至agilent猕猴表达微阵列v2持续17h。杂交后,将微阵列在室温用ge洗涤缓冲液1(agilent)洗涤1min,并且在37℃用ge洗涤缓冲液2(agilent)洗涤1min。洗涤后,立即在agilentdna微阵列扫描仪(g2505c)上扫描载玻片,对4x44k阵列载玻片使用单色扫描设置(扫描区域61x21.6mm,扫描分辨率5μm,染料通道设置为绿色,pmt设置为100%)。用featureextraction软件10.7.3.1(agilent)使用默认参数来分析扫描图像,以获得减去背景且空间上去趋势的经处理的信号强度。将信号通过robustmulti-arrayaverage(rma)方法进行背景校正,并且进行分位数归一化。对于热图生成和聚类分析,将数据进行log2转换和平均值中心化。统计分析使用非参数mann-whitney检验来比较呈现的数据。在每个附图图例中说明了供体和重复实验的数目。p值:不显著(n.s)p>0.05;*p≤0.05;**p≤0.01;***p≤0.001。实施例2-8的实验方法寡核苷酸的合成合成的、不含内毒素的寡核苷酸是根据本领域已知方法合成的,如在例如artificialdna:methodsandapplications(khudyakov,y.e.&howarda.fields,h.a.,eds.)crcpress,2002(isbn9780849314261)中描述的。人类体外衍生的dc如先前描述的(a.e.等人,2012blood120:768-777),使用rosetteseptmmonocyteenrichmentcocktail(stemcelltechnologies)从血沉棕黄层负选择单核细胞,并且在补充有10%fcs、1mm丙酮酸钠、10mmhepes、2mml-谷氨酰胺以及1%链霉素和青霉素的rpmi1640(均来自invitrogenlifetechnologies)中,以5x105个细胞/ml的密度,用gm-csf(250ng/ml;peprotech)和il-4(6.5ng/ml;r&dsystems)处理6天或7天,使人类单核细胞分化为modc。将1x105个modc在37℃或+4℃暴露于100μl完整的10%rpmi培养基中的cy3标记的polyi:c(使用mirusbionucleicacidlabelingkit3600l根据制造商的方案来标记polyi:c)和medon寡核苷酸45分钟。孵育后,将细胞用冰冷的pbs洗涤,固定,并且通过流式细胞术监测摄取。在facscaliburtm或fortessatm(bdbiosciences)上获得样品数据;用flowjotm软件(treestar)进行分析。流感病毒感染dc将在gm-csf和ril-4存在的情况下分化6-7天的单核细胞衍生的树突状细胞(modc)用sson(参见表1;0.5μm)预处理2小时,并且然后暴露于不同moi(感染复数)的流感病毒h1n1cal07/09。将modc(500,000/ml)暴露于0.2或2的moi的流感病毒2小时。其后洗涤细胞并且与sson(参见表1;0.5μm)培养24小时。从细胞培养上清液分离rna,并且通过qt-pcr测量病毒载量。在指示的时间点收获上清液,并且保存在-80℃,直到进行rna纯化。将viralrna小型试剂盒(qiagen)遵循制造商的方案用于上清液中的病毒rna纯化。使用iavcal07/09作为内部标准,通过qrt-pcr(iiione-stepquantitativert-pcr系统,lifetechnologies)来确定病毒载量。使用对iavcal07/09ha区段特异性的引物和探针进行taqman实时pcr。使用了对iavha区段特异性的以下引物序列(5’-3’):正向:ggctgctttgaattttaccacaa(seqidno:26);反向:tttgggtagtcataagtcccatttt(seqidno:27);以及探针fam-tgcgataacacgtgcatggaaagtgtc-tamra(seqidno:28)modc成熟研究如上文描述的使人类单核细胞分化。将modc暴露于polyi:c和medon,并且在48小时后使用靶向cd1a和modc成熟标志物cd86的单克隆抗体(ab)(所有ab均来自bdbiosciences)来评估成熟。在fortessa(bdbiosciences)上获得流式细胞术样品数据,并且用flowjo软件(treestar)进行分析。细胞因子/趋化因子分泌研究在polyi:c(invivogen)和medon处理后的特定时间点,从细胞收集上清液。通过标准elisa根据制造商的说明书(mabtech)测量il-6的分泌。通过450nm处的3,3’,5,5’-四甲基联苯胺(tmb)吸光度来监测蛋白量,并且以pg/ml或相对吸光度单位给出。体内a型流感病毒感染小鼠将balb/c小鼠分为n=5的组,并且安置在完全适应的室中。将5x105pfu流感h1n1a/eng/195/2009病毒以100μl经鼻施用至麻醉的小鼠。在指示的组中,将sson(25μg)(medon-01)与病毒共施用。在第三组中,在病毒感染后第3天,将pbs中的sson(25μl中25μg)经鼻施用至麻醉的小鼠。每天监测小鼠体重,通过体重减轻来确定生产性感染。为了确定肺病毒载量,在感染后4天处死小鼠。确定肺病毒滴度和细胞因子收集支气管肺泡灌洗液(bal)后,将肺取出并且通过100μm细胞过滤器匀浆,然后以200xg离心5分钟。取出上清液并储存用于elisa分析,并且用红细胞裂解缓冲液(ack;0.15m氯化铵、1m碳酸氢钾和0.01mmedta,ph7.2)处理细胞沉淀物,然后以200xg离心5分钟。将剩余的细胞重悬于含有10%胎牛血清的rpmi1640培养基中,并且通过台盼蓝排除法确定存活细胞数。使用从tissuelyzer(qiagen,manchester,uk)中破碎的冷冻肺组织经trizol提取的rna来评估体内病毒载量。将rna转化为cdna,并且在stratagenemx3005p(agilenttechnologies,santaclara,ca,usa)上使用批量病毒rna进行定量rt-pcr,对于流感病毒m基因和mrna,使用0.1μm正向引物:5’-aagacaagaccaatyctgtcacctct-3’(seqidno:29),0.1μm反向引物:5’-tctacgytgcagtccycgct-3’(seqidno:30),以及0.2μm探针:5’-fam-tyacgctcaccgtgcccagtg-tamra-3’(seqidno:31)。使用流感病毒m基因标准质粒来确定m-特异性rna拷贝数。统计分析用prism5.0tm(graph-padsoftwareinc.)进行统计分析,使用非参数kruskal-wallis非配对检验,随后使用dunn后检验(*p<0.05,**p<0.01以及***p<0.001)或单因素anova。当被指示时,使用非参数mann-whitney非配对测试来比较不同的处理组。实施例2:medon-01抑制polyi:c的摄取将人类单核细胞衍生细胞在+37℃或+4℃在添加或不添加medon-01的情况下用荧光标记的胞吞摄取标志物(polyi:c)处理45min。在+37℃,如通过流式细胞术测量的,polyi:c被摄取(图7)。图7示出了用medon-01体外处理对原代人类单核细胞衍生细胞摄取polyi:c的剂量依赖性抑制。实施例3:抑制polyi:c摄取的能力不严格依赖于序列,而是需要寡核苷酸的一定长度和化学修饰将人类单核细胞衍生细胞在+37℃或+4℃在添加或不添加medon-01的情况下用荧光标记的胞吞摄取标志物(polyi:c)处理45min。在+37℃,如通过流式细胞术测量的,polyi:c被摄取(图8)。图8示出了使用0.5μm的medon-01、03、04、07、08、10、11、13-21体外处理对原代人类单核细胞衍生细胞摄取polyi:c的抑制。实施例4:通过用medon-01预处理抑制a型流感用流感病毒h1n1cal2009(iav)使用moi0.2或moi2体外感染人类单核细胞衍生的dc。在感染前,modc未被处理或用示出为seqidno:1的medon-01(0.5μm)预处理2小时。然后在另外的2小时期间用iav进行感染。洗涤细胞并且在再添加或不添加medon-01(seqidno:1)的情况下孵育24小时(m=培养基)。培养上清液中的病毒载量通过实时pcr来定量。在使用六个不同的供体以一式三份进行分析的四组独立实验中获得数据。使用非参数配对wilcoxon检验进行统计计算。结果(图9)显示,使用两种剂量的iav(0.2和2moi),病毒载量均明显且高度显著地减少。实施例5:抑制a型流感病毒感染的能力是序列非依赖性的,但需要寡核苷酸的一定长度和化学修饰用流感病毒h1n1cal2009(iav)使用moi1体外感染人类单核细胞衍生的dc。在感染前,modc未被处理或用示出的medon(参见图10和表1(1μm))预处理2小时。然后在另外的2小时期间用iav进行感染。洗涤细胞并且在再添加或不添加medon的情况下孵育24小时(m=培养基)。培养上清液中的病毒载量通过实时pcr来定量。使用两个不同的供体以一式三份进行分析来获得数据。结果(图10)显示,使用medon01、03、04、07、10、11、13-21,iav(1moi)的病毒载量减少而不引起细胞死亡。实施例6:通过体内medon-01处理对a型流感病毒载量的抑制用流感病毒h1n1(cal09)鼻内感染balb/c小鼠。一组未被处理(图11a;圆圈),第二组在病毒攻击的同时鼻内接受25μg的sson(medon01,seqidno:1)(图11a;正方形),而第三组在第3天接受25μgsson的鼻内处理(图11a;三角形)。感染后第4天处死小鼠,并且在肺中测量病毒载量。使用单因素anova进行统计计算。如图11a中示出的,当给予medon-01作为预防性处理(正方形)或在第3天给予medon-01作为治疗性处理(三角形)时,小鼠的病毒载量存在显著减少。每天对小鼠称重,这是它们在病毒感染期间健康状况的灵敏的度量。接受medon-01预防性处理的小鼠能够保持其体重(图11b;正方形)。实施例7:medon-01处理的a型流感病毒感染的小鼠中的tnf-α和il-6产生用流感病毒h1n1(cal09)鼻内感染balb/c小鼠。一组未被处理(图12;圆圈)。第二组在病毒攻击的同时鼻内接受25μg的sson(medon01,seqidno:1)(图12;正方形)。第三组在第3天接受25μgmedon-01的鼻内处理(图12;三角形)。感染后第4天处死小鼠。通过elisa测量从肺细胞释放的(肺)或支气管肺泡灌洗液中存在的(bal)tnf-α和il-6的水平。使用单因素anova进行统计计算。结果(图12)未显示a型流感病毒感染的小鼠的不同组之间tnf-α或il-6产生的任何显著差异。实施例8:sson对modc的il-6分泌和cd86表达的作用在0.5μmmedon不存在或存在的情况下用tlr3激动剂polyi:c(25μg/ml)处理modc,并且在+37℃孵育。处理后48h,收获细胞并且对免疫成熟标志物cd86染色。medon-01和medon-13-20能够抑制polyi:c介导的免疫成熟。在图13中示出了结果。在0.5μmmedon不存在或存在的情况下用tlr3激动剂polyi:c(25μg/ml)处理modc,并且在+37℃孵育。处理后24h,收集上清液并且评估促炎性细胞因子il-6的分泌。medon-01和medon-13-20能够抑制polyi:c介导的il-6的诱导。在图14中示出了结果。另外的实验数据另外的实验数据的实验方法原代人类细胞人类血沉棕黄层从karolinskainstitutet,stockholm,sweden获得。所有关于人体外周血细胞的实验工作都是根据瑞典的指南和规定进行的。所有实验方案均由stockholm当地伦理委员会批准(“regionalaistockholm”,伦理许可dnr2006/229-31/3)。根据瑞典的规定,用来自血沉棕黄层的细胞进行体外实验工作不需要知情同意。如先前描述的(等人(2012),使用rosettesepmonocyteenrichment试剂盒产生人类单核细胞(1ml/10ml血沉棕黄层;stemcelltechnologies),并且在37℃、5%co2,在补充有10%fcs、1mm丙酮酸钠、10mmhepes、2mml-谷氨酰胺以及1%链霉素和青霉素的rpmi1640(均来自invitrogenlifetechnologies)中,以5x105个细胞/ml的密度,用gm-csf(250ng/ml;peprotech)和il-4(6.5ng/ml;r&dsystems)处理6天,使人类单核细胞分化为modc。通过cd1a表达监测细胞的分化。ficoll分离(stemcell)后,从血沉棕黄层分离pbmc。寡核苷酸完全ps取代的sson35ps和2’omeps/po由integrateddnatechnologies制备。其他修饰的寡核苷酸购自eurofins。对于序列,参见表2。使用rneasy总rna纯化试剂盒根据制造商的说明书(qiagen)从modc中纯化总rna。tlr配体和标记的内体标志物tlr3配体pi:c(25μg/ml,除非另有说明)、tlr7配体cl307(1μg/ml)和tlr7/8配体r848(1μg/ml)均购自invivogen。lps(100ng/ml)购自sigma。运铁蛋白-alexa647、右旋糖酐-texasred10kda和lps-alexa488均购自molecularprobes。使用寡核苷酸labelit-试剂盒(mirus)根据制造商的说明书产生cy3标记的pi:c。modc的摄取研究在冰上将modc暴露于完整的10%rpmi培养基(或无血清培养基,用于poon摄取研究)中添加或不添加sson的内体标志物,并且然后转移至37℃,持续45min。用冷的pbs洗涤细胞并且固定(cytofix,bdbiosciences)。通过流式细胞术(fortessa,bdbiosciences)监测荧光信号。用flowjo软件(treestar,9.6.4版)分析数据。通过以下监测cme:25μg/ml运铁蛋白-alexa647,5μg/mlldl-dil或1μg/ml至5μg/mlpi:c-cy3。mpc:0.5mg/ml10kda右旋糖酐-texasred。tlr4的胞吞摄取:1μglps-alexa488。以1:6的比用lyovec(invivogen)根据制造商的说明书进行pi:c-cy3的转染。对于显微术,使modc粘附至多聚-l-赖氨酸包被的玻璃载玻片2-4h。在37℃,在0.5μmsson35ps存在或不存在的情况下,用配体处理细胞45min。洗涤细胞,并且在37℃在宽场cellobserver显微镜(zeiss)中使用40x透镜来获得图像。使用slidebook6程序(intelligentimaginginnovations)获得和分析图像。可选地,在细胞粘附后,在37℃,在0.5μmsson35ps存在或不存在的情况下,用polyi:c-cy3以及polyi:c-cy3与lyovec处理细胞40min。用pbs洗涤细胞,并且用麦胚芽凝集素alexa633(invitrogen)染色10min。洗涤细胞,然后在3.7%甲醛(sigma)中固定。在lsm800airyscan共聚焦显微镜(zeiss)中使用63x油浸透镜获得图像,并且使用zenblue软件(zeiss)分析图像。modc成熟研究将modc暴露于添加或不添加sson的pi:c(25μg/ml)、cl307(1μg/ml)、r848(1μg/ml)lps(100ng/ml)或pi:c/lyovec(1μg/ml)(invivogen),并且在48h后使用靶向cd1a以及modc成熟标志物cd86、cd83和cd80的单克隆抗体(ab)(cd1a-bv510、cd86-apc、cd83-fitc、cd80-pe;全部来自bdbiosciences)通过流式细胞术进行评估。在使用氯喹的情况下,在37℃将细胞用10μm氯喹预孵育1h。使用live/dead可固定近ir死细胞染色试剂盒(lifetechnologies)排除死细胞。细胞因子/趋化因子分泌研究在处理后的特定时间点收集上清液,并且通过标准elisa通过spectramaxi3x,moleculardevices根据制造商的说明书(mabtech;il-6、il-10和il-29。lifetechnologies:cxcl10),来测量细胞因子/趋化因子的分泌。hek细胞的tlr3活化研究将tlr3转染的hek-bluetm细胞(invivogen)以20,000个细胞/孔接种在384孔板(corning)中,并且留置附着1h。用1μg/mlpi:c、指定浓度的sson35ps处理细胞并且孵育过夜。在hek-bluetm检测培养基(invivogen)中用envision酶标仪(perklinelmer)在640nm处测量tlr3活性,并且以没有sson35ps的pi:c处理的细胞的百分比给出。sson35po和ps的核酸酶降解与ciafre等人类似地进行核酸酶降解,并且在无血清rpmi培养基中进行。以1u/μgdna的比使用dna酶i(qiagen)。以10μmsson35po/ps的浓度在37℃进行反应0-24h。在特定的时间点取出等分试样,冷冻于-80℃,并且以后在2%琼脂糖凝胶上分析。tbe缓冲液中以100伏进行电泳1h。凝胶用溴化乙锭(0.5μg/ml)来染色,并且使用凝胶docez成像仪(biorad)拍照。tlr3抑制的动力学分析将modc暴露于25μg/mlpi:c/0.5μmsson35ps,并且然后分别用0.5μmsson35ps或25μg/mlpi:c脉冲处理5min至24h的时间段。在初始暴露后48小时,在cd1a阳性细胞中通过流式细胞术分析细胞的成熟标志物cd80和cd86的表达。另外,在处理后48小时收集上清液,并且通过elisa(mabtech)测量il-6。存活力测试在用25μg/mlpi:c/0.5μmsson35ps初始处理后48h,使用live/dead可固定近ir死细胞染色试剂盒根据制造商的方案(lifetechnologies)评估存活细胞。通过流式细胞术获得数据。根据制造商的方案(sigma)进行wst-1测定,并且在spectramaxi3x上进行测量。用dson进行modc成熟研究在pbs中将5μm的sson35ps与1-10μm的互补sson35po在55℃预孵育,并且然后在环境温度留置30min。将on混合物以0.5μmsson35ps的终浓度连同25μg/mlpi:c添加至modc。如上文描述的评估modc成熟。rna测序和差异表达分析使用rneasy总rna纯化试剂盒根据制造商的说明书(qiagen)从modc纯化总rna,并且提交至瑞典斯德哥尔摩国家基因组基础设施(nationalgenomicsinfrastructureswedenstockholm)(ngi)进行测序。生物分析仪痕量和浓度值是遵循ngi网站上可获得的“基因组学样品要求”文件中的指南获得的。用来自illumina的truseqribozero试剂盒进行rna测序。基于质谱的modc蛋白质组学分析在用pi:c(25μg/ml)或sson35ps(0.5μm)刺激modc后,在特定时间点收集2x8组样品,并且快速冷冻。对于完整的液相色谱串联质谱术(lc-ms/ms),使用在线耦合至qexactiveorbitrap(thermofischerscientific)的agilent1200nano-lc系统进行分析。使用包括用于改进鉴定的sequest-percolator(l.等人(2007)的软件proteomediscoverervs.1.4.0.288将数据针对人类ensembl数据库(版本37)进行检索,以进行蛋白鉴定(错误发现率(fdr)为<1%)。层次聚类和网络生成使用软件morpheus(https://software.broadinstitute.org/morpheus/index.html)对log2转换数据进行分析。使用metacoretm6.22版(thomsonreuters)进行网络分析,以鉴定层次聚类中直接相互关联的基因,并且使用cytoskape3.2.1版(shannonp.等人(2003))来展示结果。动物和注射按照欧洲nhp护理指南(eudirectiven63/2010)处理成年食蟹猴(猕猴)(n=18,雌性和雄性两者)。本研究由cea伦理动物委员会“comitéd’ethiqueenexpérimentationanimale”根据12-013号声明批准和认可,该委员会由法国研究部根据44号注册。在麻醉情况下处理动物,并且在左上和右后侧腹仅用170μgpi:c或用100μlpbs中的170μgsson35ps或仅用pbs进行皮内注射。可选地,如附图图例中指示的,进行sson35ps的剂量递增。注射后24h,从麻醉动物收集皮肤活组织检查物(直径为8mm)。nhp组织收集和流式细胞术从注射后24h收集的新鲜的皮肤活组织检查物提取细胞。将表皮和真皮细胞用单克隆抗体混合物(hla-drv500、cd123-pecy7、cd45-v450、cd11c-apc、cd14-apc-h7来自bdbioscience;cd66-apc、cd66-fitc来自miltenyi;cd1a来自dako;cd163-pcpcy5.5来自biolegend)染色,并且通过流式细胞术来获得。组织化学对活的cd66+cd45+nhp细胞进行facs分选(facsaria,bdbiosciences),并且根据karolinskauniversityhospitalhuddinge临床病理学实验室的临床常规程序,通过细胞离心放置在plus金载玻片(thermoscientific)上,以及使用吉姆萨染色。细胞因子分泌测定在未进行另外刺激的情况下,在bio-plex设备(bio-rad,france)上,用milliplexmapnhpcytokinemagneticbeadpanel(millipore,france)测量离体分离的过滤真皮上清液的等分试样。微阵列分析从猕猴皮肤活组织检查物提取总体皮肤dna。对于详细的样品制备和方案,参见补充材料和方法部分。在旋转的agilent杂交炉中,在65℃将rna杂交至agilent猕猴表达微阵列v2持续17h。将微阵列洗涤并且在agilentdna微阵列扫描仪(g2505c)上扫描,对4x44k阵列载玻片使用单色扫描设置(扫描区域61x21.6mm,扫描分辨率5μm,染料通道设置为绿色,pmt设置为100%)。用featureextraction软件10.7.3.1(agilent)使用默认参数来分析图像,以获得每个基因的背景调整的信号(gprocessedsignal)。随后将信号分位数标准化,以增加样品间的可比性。途径分析使用ingenuitypathwayanalysis软件(build版456367m,content版39480507发布日期20170914)(ingenuitysystems)来鉴定经典的信号途径。为了计算富集的显著性(在该软件中执行fisher精确检验),参考分子集为ingenuityknowledgebase(仅基因)。取决于分析的三个输入分子集:(i)基于层次聚类选择的具有相关表达的蛋白的蛋白质组聚类(质谱数据,体外人类modc)(ii)通过大于2的绝对log2倍数变化过滤的rnaseq数据(体外人类modc)和(iii)通过大于5的绝对log2倍数变化过滤的来自猕猴皮肤活组织检查物的微阵列数据。使用gorilla(基因本体富集分析和可视化工具,geneontologyenrichmentanalysisandvisualizationtool)网络工具(edene.等人(2009))进行基因本体(go)富集分析。定量和统计分析使用非参数kruskal-wallis非配对检验,随后使用dunn后检验或mann-whitney检验来比较图15-16和图18b中呈现的数据。用于统计计算的数据来自独立实验中生物复制中的至少三个供体。示出了来自个体动物的猕猴研究的数据。转录组和蛋白质组数据的统计计算如上文所述。p值:不显著(n.s)p>0.05;*p≤0.05;**p≤0.01;***p≤0.001。使用graphpadprism7软件计算统计数据。另外的实验数据sson抑制tlr3活化的能力的长度要求为至少25个碱基dna中的cpg基序可以与tlr9结合,导致tlr9+细胞诸如浆细胞样树突状细胞和b细胞的活化。我们先前显示,包含cpg的sson(35个碱基)抑制缺少tlr7和tlr9两者的modc中的tlr3活化(等人(2012))。在目前的研究中,我们完全使用不含cpg基序的sson来解决它们抑制tlr3活化的能力。为了进一步了解关于观察到的胞吞作用抑制和下游tlr3活化是否可以被关联至特定的sson序列,比较了三种不同的序列(表2)。ssongta35ps基于亲本序列sson35ps,但所有鸟苷(g)碱基都被腺苷(a)取代,而ssoncompl35ps是sson35ps的互补序列。在用pi:c处理48h的modc中,它们全都显示出相同的抑制作用(图15a)。为了进一步了解结构要求,我们评估了sson的长度是否影响其效力。我们发现较短的sson(表2)不具有与较长的形式相同的抑制能力,有效的sson的长度截止值在20个和25个碱基之间(图15b和15c)。在没有转染剂的情况下递送(裸递送)的用于反义目的的治疗性sson通常少于20个碱基(soiferhs等人(2012)和steinca等人(2010))。因此,我们接下来评估了包含15个碱基的sson的摄取。显微术研究显示,荧光标记的sson15ps容易被modc内化,并且通过添加sson35ps(图15d和15e)以浓度依赖性方式有效地阻断了这种细胞摄取(图15f)。因此,这些数据提供了关于为何依赖于胞吞摄取的治疗性反义sson应当短于20个碱基的线索。我们接下来研究了其他种类的细胞外核酸是否诱导抑制作用,或者抑制作用是否需要sson。为了评估引入双链(ds)dna是否影响胞吞摄取,将sson35ps保持在恒定浓度(0.5μm),同时引入递增浓度的非稳定化互补链(ssoncompl35po)(图15g)。用pi:c处理modc,并且当引入互补链时,sson的抑制活性减少。当所有sson35ps都具有杂交的互补链时(ssoncomplpo1μm),抑制作用消失。天然存在的posson被降解,除非它能够与sson35ps杂交。在初始处理后48h,包含po骨架的sson(sson35po)对pi:c诱导的成熟没有抑制作用,与其降解一致。在添加非互补sson35po的情况下,sson35ps保持抑制作用(图15g)。总之,这些数据显示,sson而非dsdna能够以浓度依赖性的方式抑制tlr3激动剂pi:c的胞吞摄取和下游作用,并且这些作用不与特定的sson序列或基序相关。然而,抑制依赖于sson的长度,在25个碱基或多于25个碱基时达到完全效果。ssdna和ssrna两者都抑制网格蛋白依赖性胞吞途径为了阐明抑制胞吞作用的能力是否为ssdna独有,还是ssrna也具有类似的抑制能力,我们合成了包含常见于核糖体rna和小核rna的无毒修饰2’-o-甲基rna(2’ome)的35meron(图16a)(表2)。这种具有ps骨架的sson2’omerna类似物有效地抑制了tf和lps两者的摄取(图16b和16c)。与ssdna一样,具有未修饰的po骨架的2’omessrna能够抑制tf的摄取,尽管其效力较低(图16d)。通过显示在与pi:c和sson2’omeps一起孵育后,从modc分泌的il-6降低,降低程度与sson35ps相当,发现了暂时性抑制的其他证据(图16e)。另外,当在3-4h后测量时,使用低于200个碱基的截止过滤器纯化的总rna(totrna)以浓度依赖性方式减少了il-6的产生(图16f)。这些数据显示,具有暂时性抑制人类modc中网格蛋白介导的胞吞摄取的能力的sson可以源自ssdna或ssrna。抑制modc中网格蛋白介导的胞吞作用后,转录组或蛋白质组中无重大变化为了了解modc中网格蛋白介导的胞吞作用关闭后发生的细胞反应,在暴露于sson后,我们进行了转录组学和蛋白质组学的联合研究(图17)。在具有和没有sson的情况下,用pi:c刺激后也进行了这些研究,以获得sson抑制tlr3活化的能力的总体情况(图17)。rnaseq分析揭示,在24h后,pi:c刺激的modc中ifn刺激基因(isg)而不是炎性体(inflammasome)相关基因强烈上调(barouch等人(2016))(图17a)。ingenuitypathwayanalysis(ipa)显示,经典途径“干扰素信号传导”、“模式识别受体在识别细菌和病毒中的作用”和“树突状细胞与自然杀伤细胞之间的交互作用”中的基因上调(图17a)。sson35ps的添加消除了pi:c介导的isg诱导,支持sson对tlr3活化的有效抑制(图17b)。仅用sson35ps处理未导致转录组的任何明显变化,对转录组的测量多达24h(图17c)。蛋白质组学分析类似地显示出,在24h后pi:c刺激的modc中isg而不是炎性体相关蛋白上调(图17d)。蛋白质组学分析证实,sson35ps消除了pi:c介导的isg诱导(图17e)。仅用sson35ps刺激未导致蛋白质组的任何重大变化(图17f)。在7个时间点测量了modc中整体蛋白质组变化的时间进程:刺激前,在用sson处理后15min、30min和60min,以及3h、8h和24h(数据未示出)。为了用相关蛋白表达水平来选择蛋白子集以进一步解释数据,进行了层次聚类和网络分析(图17)。ipa分析揭示,对modc的pi:c刺激导致与“干扰素信号传导”、“模式识别受体在识别细菌和病毒中的作用”和“树突状细胞与自然杀伤细胞之间的交互作用”相关的蛋白子集的明显的时间依赖性上调(图17d),与rnaseq数据和先前的研究一致(amit等人(2009)和shalek等人(2014))。pi:c刺激后上调的蛋白包括mhci类和ii类分子,以及与modc成熟相关的标志物,诸如cd83(数据未示出)。如预期的,检测到在prr接合、干扰素信号传导和irf活化后发挥作用的许多分子。对在pi:c刺激后上调的聚类进行的网络分析鉴定出irf1、7、8和stat1为最相互关联的蛋白(>10个边)(数据未示出),而在sson刺激后蛋白水平变化不太明显(图17f)。在sson刺激稳态人类modc后,在上调或下调的蛋白的蛋白子集中未能检测到明显的趋势或聚类。总之,这些数据显示,pi:c刺激导致modc中转录和蛋白质组水平方面都检测到的isg的上调,而sson的添加减弱了这种反应。此外,仅用sson处理的modc中转录组未发生重大变化,在8-24h后蛋白质组开始发生微小变化。然而,蛋白质组和转录组缺乏变化并不排除sson诱导细胞中发生其他变化。细胞骨架、磷酸化模式的调节或蛋白-蛋白相互作用的改变都可能导致somie作用。施用sson35ps调节猕猴皮肤中dsrna介导的炎症为了评估sson35ps是否能够介导体内作用,在sson35ps存在或不存在的情况下皮内注射pi:c后,在取自注射部位的猕猴皮肤活组织检查物中,我们测量了细胞的局部浸润及其固有免疫特征。使用多颜色流式细胞术对从表皮层和真皮层分离的细胞进行表型分型(adam等人(2015)和epaulard等人(2014))(图18),并且揭示了注射pi:c导致多形核中性粒细胞(pmn)和hla-dr+cd1抗原呈递细胞(apc)的内流,而添加sson减少表皮hla-dr+cd1a细胞的浸润,但不减少pmn的浸润(图18)。为了评估存在或不存在sson时对pi:c的整体固有应答,对皮肤活组织检查物进行了转录谱分析。ipa分析揭示,在注射pi:c后启动了固有免疫的若干途径,诸如“固有免疫细胞与适应性免疫细胞之间的通信”、“trem1信号传导”、“模式识别受体在识别细菌和病毒中的作用”和“树突状细胞与自然杀伤细胞之间的交互作用”(图18)。接下来,我们将仅接受皮内注射pi:c的组与接受pi:c连同sson的组进行比较,并且计算了两组间的倍数变化以及显著性差异(图18a)。结果显示,在添加sson35ps后,与炎性状况相关的趋化因子和基因在下调最多的基因中(图18a)。单独的pi:c处理导致趋化因子ccl5、cxcl9和cxcl1以及rax的显著诱导,rax是dsrna依赖性丝氨酸/苏氨酸蛋白激酶的细胞活化物(bennetrl等人(2006)),它们在联合的pi:c和sson处理中均显著减少(图18b)。类似地,在pi:c注射后,组织中参与细胞粘附的icam-1和lrg1上调,但这种诱导通过与sson共注射而降低(图18b)。ifn-γ调节许多趋化因子(antonelli等人(2014))。pi:c注射确实导致了ifn-γ和il-6两者的表达增加,而pi:c和sson联合处理减少了这种表达,这与趋化因子表达的调节一致(图18b)。为了验证皮肤中细胞因子蛋白的分泌,收集过滤的真皮上清液的等分试样,并且在没有另外的体外刺激的情况下,使用bio-plex分析进行分析(图18c)。在pi∶c处理后,检测到il-6和ifn-γ的显著诱导。明显的趋势是,添加sson降低了il-6和ifn-γ的产生,而相反地引起了显著的il-10释放。值得注意的是,单独的sson在体内诱导了剂量依赖性的il-10分泌(图18c)。总之,这些结果显示,与单独的pi∶c相比,接受pi∶c和sson联合处理的动物中存在局部阻抑的炎性特征,这与sson抑制tlr3活化的能力一致。表2寡核苷酸的一级结构。名称序列长度sson35pogaagttttgaggttttgaagttgttggtggtggtg35sson35psg*a*a*g*t*t*t*t*g*a*g*g*t*t*t*t*g*a*a*g*t*t*g*t*t*g*g*t*g*g*t*g*g*t*g35sson30psa*g*t*t*t*t*g*a*g*g*t*t*t*t*g*a*a*g*t*t*g*t*t*g*g*t*g*g*t*g30sson25pst*t*t*g*a*g*g*t*t*t*t*g*a*a*g*t*t*g*t*t*g*g*t*g*g25sson20pst*g*a*g*g*t*t*t*t*g*a*a*g*t*t*g*t*t*g*g20sson15psg*g*t*t*t*t*g*a*a*g*t*t*g*t*t15ssongtapsa*a*a*a*t*t*t*t*a*a*a*a*t*t*t*t*a*a*a*a*t*t*a*t*t*a*a*t*a*a*t*a*a*t*a35ssoncomplpocaccaccaccaacaacttcaaaacctcaaaacttc35ssoncomplpsc*a*c*c*a*c*c*a*c*c*a*a*c*a*a*c*t*t*c*a*a*a*a*c*c*t*c*a*a*a*a*c*t*t*c35sson2’omepogaaguuuugagguuuugaaguuguuggugguggug35sson2’omepsg*a*a*g*u*u*u*u*g*a*g*g*u*u*u*u*g*a*a*g*u*u*g*u*u*g*g*u*g*g*u*g*g*u*g35所有序列都以5′到3′书写*=ps斜体=rna_=2′ome实验结果的讨论在本文中,我们揭示了某些sson能够消除配体的摄取,并且从而抑制tlr3、4和7信号传导内体的活化,这支持并且扩展了我们先前的公布内容,即cpgsson(35个碱基)抑制tlr3信号传导(等人(2012))。我们在此还证明了,如通过显微术示出的以及通过流式细胞术定量的,sson阻断tf、ldl、lps和pi∶c的摄取。通过测量来自经刺激的modc和pbmc的细胞因子分泌和共刺激分子的表达,显示了对tlr3、4和7信号传导的功能抑制。我们还通过进行rnaseq和全细胞蛋白质组分析评估了整体变化,这支持了内体信号传导的阻断。对抑制网格蛋白介导的胞吞作用的结构要求的表征揭示,ssdna或ssrna,而不是dsdna,赋予了该抑制。该能力并不严格依赖于sson序列,但要求它包含至少25个碱基,以具有完全的抑制作用。因此,抑制作用不依赖于cpg基序。通过测量注射后皮肤的局部反应,我们还提供了证据,sson35ps在猕猴体内调节tlr3活化。网格蛋白介导的胞吞作用的暂时性关闭并未严重妨碍细胞。这得到了以下结果的支持:在sson的存在下发生细胞因子分泌,条件是pi:c在sson添加之前被摄取。此外,我们显示了sson并未诱导任何明显的毒性,并且它可以挽救细胞避免pi:c诱导的细胞死亡。rnaseq和全细胞蛋白质组学分析支持这个结论,并且在单独sson处理后,转录组或蛋白质组未发生重大变化,在刺激后对其测量多达24h。在猕猴中注射sson并未诱发炎症,相反,观察到对dsrna(pi:c)介导的炎症的阻抑。我们还显示巨胞饮作用(通过右旋糖酐的摄取测量的)不受sson影响,允许持续摄取液相中的营养。我们发现,至少25个核苷酸的ssdna或ssrna具有暂时(条件是它们没有被降解)关闭网格蛋白介导的胞吞作用的能力,这在宿主-病毒相互作用、rna生物学和自身免疫中提出了有趣的问题。我们的数据涉及1)病毒胞吞作用2)细胞摄取mirna3)内源性rna的稳定化修饰的作用4)非编码rna的免疫调节功能5)开发基于sson的治疗剂(反义on、剪接转换on、免疫遏抑性on、sirna、cpg佐剂)。总之,这些研究揭示了细胞外sson在tlr配体的胞吞摄取中的调节作用,并且我们称这种调节过程为somie(sson-mediatedinterferenceofendocytosis,sson介导的胞吞作用的干扰)。病原体感染和细胞损伤导致细胞外核酸的积累,细胞外核酸可以被rna酶和dna酶降解,但也被内化到内体中用于随后的tlr活化。为了避免在这些系统负荷较重的情况下过多的内体tlr活化,严格控制固有免疫应答是至关重要的,从而减轻潜在的自毁事件。已经表明数目越来越多的分子,诸如a20、cyld、usp4、usp25、otub1/2、duba和mysm1,参与阻抑prr触发的细胞内信号传导级联反应(panda等人(2015))。然而,越来越多的理解到,这些酶发挥的作用不能单独解释prr活化的严格调节。在此,我们证明了sson,天然ssdna和ssrna2’ome两者,都暂时干扰了cme和下游内体tlr活化。somie充当调节过程,暂时限制固有免疫系统的活化。用稳定化的sson35ps处理显示出更长时间的胞吞作用阻断,而mpc保持不受影响,显示出抑制作用对某些胞吞途径是特异性的。已知吞噬作用受活化性(促吞噬;“吃我”)和抑制性(“不要吃我”)受体控制。抑制性信号调节蛋白(sirp)α在与吞噬作用的检查点配体cd47结合后,提供了对吞噬作用的阻断(veilette等人(2018))。胞吞作用的货物通过流式细胞术来定量并且通过显微术来可视化,而tlr的功能活化通过诱导modc中的共刺激分子cd80和cd86以及细胞因子的分泌来测量。为了揭示sson是否特异性影响定位于内体区室中的tlr的活化,我们选择了具有不同特性的tlr激动剂,并且评估了sson35ps是否抑制il-6的分泌。tlr7/8激动剂r848是膜可透过的(govindaraj等人(2011)),并且不受sson的添加的影响。相比之下,胞吞作用依赖性tlr7激动剂cl307的活性受sson35ps显著抑制。此外,我们研究了感应lps的tlr4,tlr4在modc中具有两种不同的信号传导途径。sson35ps选择性地降低了lps介导的il-29/cxcl10的分泌,该分泌主要位于trif下游并且依赖于胞吞作用,而sson35ps不影响myd88依赖性il-6/il-10分泌。仍有待于进一步阐明的是,在哪些细胞类型中tlr4被偶联至myd88和trif信号传导途径两者,并且揭示潜在的物种差异。动力学分析揭示,如果在用pi:c刺激后多于50min时添加,则sson35ps的抑制能力降低,并且在2小时后几乎可忽略。因此,sson看来似乎不能抑制正在进行的tlr3信号传导。当以相反的方式处理modc时,以sson35ps的单一处理开始,则pi:c诱导的成熟被完全阻断多达24h。ps取代的sson在含血清的细胞培养基中具有超过24h的半衰期(ciafrè等人(1995))。另一方面,pi:c在含血清培养基中迅速降解(levy等人(1975)),并且不能在用sson35ps预处理后的modc中触发tlr3。可以预期,对胞吞活性的干扰允许modc对最近摄取的货物作出反应,而不会在完成对第一批胞吞货物的反应之前接受过多的另外刺激或相反的指令。另外,发现modc可以在sson的存在下成熟并且产生il-6产物,条件是货物(pi:c)首先被允许摄取,表明网格蛋白介导的胞吞作用的暂时关闭不会严重损害细胞。转录组学和蛋白质组学modc数据提供了进一步的证据,表明sson有效地减弱了pi:c介导的反应,并且sson处理的细胞在多达24h内没有发生重大变化。我们合成了一组on(表2)来确定somie的结构要求。somie作用并不严格依赖于序列或基序,因为sson35ps的互补链和其中所有g都变成a的突变的sson显示出相似的效力。sson对内体tlr活化的干扰很强,ic50为约150nm。这导致我们推测我们正在监测一种生物学上保守的功能。然而,据我们所知,细胞中ssdna的来源是有限的,并且主要可见于外泌体中(balaj等人(2011)),而另一方面,ssrna则是丰富的。我们证明了35mer完全ps取代的rna类似物2’omesson具有阻断pi:c诱导的modc成熟的能力,并且总rna(totrna)抑制pi:c诱导的il-6的分泌。综合起来,somie活性并不严格依赖于sson序列,ssdna和ssrna两者均可发生,并且稳定性修饰(即ps)对抑制不是必需的,但对于随时间推移保持活性是需要的。仍有待于进一步阐明的是,寡核苷酸的代谢和稳定化碱基修饰诸如2’ome或锁核酸(lna)的差异添加,是否可以通过设置内体tlr活化的阈值来影响自身免疫的易感性。哺乳动物细胞包含大量具有不同长度的编码和非编码on(cech等人(2014))。免疫调节微rna(mirna)由18-23个碱基的sson组成(mehta等人(2016))。然而,细胞也包含尺寸更长的sson(cech等人(2014))。在细胞死亡并释放细胞内容物期间,可以想象不同长度的编码和非编码的dna和rna都可以被释放到细胞外空间,变得易于被dna酶和rna酶降解。可以假定,这些酶的活性将极大地影响细胞外空间中的on的大小组成,并且最终影响感应核酸的prr的触发。我们在此显示,为了发挥somie活性,存在至少20-25个碱基的长度要求。sson(25-35个碱基)抑制较短sson(15个碱基)的摄取,而sson15ps未显示出somie活性。这与最近的数据一致,这些数据表明缺血期间释放到循环和细胞外空间中的mirna(18-23个碱基)可能反而通过内体tlr7/myd88信号传导触发细胞因子产生(feng等人(2017))。所揭示的somie的长度要求暗示了更长的非编码rna的未知功能,并且显示mirna太短而不能提供胞吞检查点控制。我们发现至少20-25个碱基的sson干扰胞吞作用,这为关于为何sson被用作反义核酸或外显子跳读sson被裸递送时应当短于20个碱基以允许通过胞吞途径细胞摄取提供了合理的解释。同样,针对tlr9激动作用的cpgsson应当少于20个碱基(scheiermann等人(2014)),以避免抑制胞吞作用,这可能会在缺少tlr9的细胞中无意中造成脱靶效应(等人(2012)。可以想象的是,细胞外空间中的on的尺寸和相对ds:ss组成将取决于局部微生物环境,在感染期间on溢出。据我们所知,目前还没有可用的技术能够测量天然存在的不同长度的on的细胞内或细胞外浓度。因此,在目前的时间,还不能真正估计观察到的通过天然配体发生的sson介导的tlr抑制的程度和时间范围。但是,我们提供了证据:在猕猴中皮内注射后,sson35ps在体内发挥了免疫调节作用。皮肤活组织检查显示注射sson后猕猴真皮中的pi∶c介导的炎症被阻抑,并且在从离体皮肤活组织检查物获得的上清液中证实了il-6分泌减少(没有另外的体外刺激)。值得注意的是,sson在体内诱导了剂量依赖性的il-10分泌,表明用sson接种可能改变局部免疫应答的方向。仍有待于阐明的是,在sson处理后,皮肤中的哪种(些)细胞类型产生il-10。在用sson刺激后的基因组学和蛋白质组学研究中,我们没有发现modc中il-10产生的任何迹象。我们假设局部环境和细胞交互作用可能是诱导il-10的关键,并且可以诱导广泛的细胞群体产生人类il-10,包括t细胞(treg、th1、th2和th17)、dc亚群、巨噬细胞、中性粒细胞、b细胞、肥大细胞、成纤维细胞和角质形成细胞(berti等;(2017))。已经表明了免疫遏抑性sson阻抑或预防特征为病理性免疫刺激和自身免疫的疾病(dong等人(2004);klinman等人(2005);peter等人(2008)。先前显示了若干免疫遏抑性sson包含直接结合至例如tlr7或tlr9所需的序列或基序(bayik等人(2016)和sarvestani等人(2015))或与stat1和stat4相互作用的重复序列(ttaggg)(shirota等人(2015))。我们和其他研究者先前已经提出,可以通过sson抑制tlr3信号传导,以便阻抑导致tlr3相关发病机制的过度反应性应答(等人(2012)和ranjith-kumar等人(2008))。在此,我们提供了证据:sson通过充当摄取货物到内体中的守门者,可能具有比先前预期的更广泛的免疫调节作用,从而为通过内体tlr3/4/7活化触发的潜在的自身免疫应答提供波动性阈值。参考文献adam,l.,rosenbaum,p.,cosma,a.,legrand,r.&martinon,f.identificationofskinimmunecellsinnon-humanprimates.j.immunol.methods426,42-49(2015).akira,s.&takeda,k.toll-likereceptorsignalling.nat.rev.immunol.4,499-511(2004).amarante,m.k.,oda,j.m.m.,reiche,e.m.v.,morimoto,h.k.,aoki,m.n.,watanabe,m.a.e.,2011.humanendogenousrnas:implicationsfortheimmunomodulationoftoil-likereceptor3.expthermed2,925-929.doi:10.3892/etm.2011.303amarante,m.k.,watanabe,m.a.e.,2010.toll-likereceptor3:involvementwithexogenousandendogenousrna.int.rev.immunol.29,557-573.doi:10.3109/08830185.2010.525723amit,i.etal.unbiasedreconstructionofamammaliantranscriptionalnetworkmediatingpathogenresponses.science326,257-263(2009).antonelli,a.etal.chemokine(c-x-cmotif)ligand(cxcl)10inautoimmunediseases.autoimmunrev13,272-280(2014).balaj,l.,lessard,r.,dai,l.,cho,y.-j.,pomeroy,s.l.,breakefield,x.o.,skog,j.,2011.turnourmicrovesiclescontainretrotransposonelementsandamplifiedoncogenesequences.naturecommunications2,180.doi:10.1038/ncomms1180barouch,d.h.etal.rapidinflammasomeactivationfollowingmucosalsivinfectionofrhesusmonkeys.cell165,656-667(2016).bayik,d.,gursel,i.,klinman,d.m.,2016.structure,mechanismandtherapeuticutilityofimmunosuppressiveoligonucleotides.pharmacol.res.105,216-225.doi:10.1016/j.phrs.2015.11.010bennett,r.l.etal.rax,thepkractivator,sensitizescellstoinflammatorycytokines,serumwithdrawal,chemotherapy,andviralinfection.blood108,821-829(2006).berti,f.c.b.,pereira,a.p.l.,cebinelli,g.c.m.,trugilo,k.p.&deoliveira,k.theroleofinterleukin10inhumanpapillomavirusinfectionandprogressiontocervicalcarcinoma.cytokinegrowthfactorrev.34,1-13(2017).cavassani,k.a.etal.tlr3isanendogenoussensoroftissuenecrosisduringacuteinflammatoryevents.j.exp.med.205,2609-2621(2008).cech,t.r.&steitz,j.a.thenoncodingrnarevolution-trashingoldrulestoforgenewones.cell157,77-94(2014).choi,c.h.j.,hao,l.,narayan,s.p.,auyeung,e.,mirkin,c.a.,2013.mechanismfortheendocytosisofsphericalnucleicacidnanoparticleconjugates.pnas110,7625-7630.doi:10.1073/pnas.1305804110ciafrè,s.a.,rinaldi,m.,gasparini,p.,seripa,d.,bisceglia,l.,zelante,l.,farace,m.g.,fazio,v.m.,1995.stabilityandfunctionaleffectivenessofphosphorothioatemodifiedduplexdnaandsynthetic′mini-genes′.nucleicacidsres.23,4134-4142.cossart,p.,helenius,a.,2014.endocytosisofvirusesandbacteria.coldspringharbperspectbiol6,a016972-a016972.doi:10.1101/cshperspect.a016972debouteiller,o.etal.recognitionofdouble-strandedrnabyhumantoll-likereceptor3anddownstreamreceptorsignalingrequiresmultimerizationandanacidicph.jbiolchem.280,38133-38145(2005).doherty,g.j.,mcmahon,h.t.,2009.mechanismsofendocytosis.annu.rev.biochem.78,857-902.doi:10.1146/annurev.biochem.78.081307.110540dong,l.,ito,s.-i.,ishii,k.j.,klinman,d.m.,2004.suppressiveoligonucleotidesprotectagainstcollagen-inducedarthritisinmice.arthritisrheum.50,1686-1689.doi:10.1002/art.20263duan,y.,zhang,s.,wang,b.,yang,b.&zhi,d.thebiologicalroutesofgenedeliverymediatedbylipid-basednon-viralvectors.expertopindrugdeliv6,1351-1361(2009).eden,e.,navon,r.,steinfeld,i.,lipson,d.&yakhini,z.gorilla:atoolfordiscoveryandvisualizationofenrichedgotermsinrankedgenelists.bmcbioinformatics10,48(2009).epaulard,o.etal.macrophage-andneutrophil-derivedtnf-αinstructsskinlangerhanscellstoprimeantiviralimmuneresponses.j.immunol.193,2416-2426(2014).feng,y.etal.extracellularmicrornasinducepotentinnateimmuneresponsesviatlr7/myd88-dependentmechanisms.j.immunol.199,2106-2117(2017).gitlin,l.etal.essentialroleofmda-5intypeiifnresponsestopolyriboinosinic:polyribocytidylicacidandencephalomyocarditispicornavirus.proc.natl.acad.sci.103,8459-8464(2006).govindaraj,r.g.,manavalan,b.,basith,s.,choi,s.,2011.comparativeanalysisofspecies-specificligandrecognitionintoll-likereceptor8signaling:ahypothesis.plosone6,e25118.doi:10.1371/journal.pone.0025118gursel,i.,gursel,m.,yamada,h.,ishii,k.j.,takeshita,f.,klinman,d.m.,2003.repetitiveelementsinmammaliantelomeressuppressbacterialdna-inducedimmuneactivation.thejournalofimmunology171,1393-1400.husebye,h.etal.endocyticpathwaysregulatetoll-likereceptor4signalingandlinkinnateandadaptiveimmunity.emboj.25,683-692(2006).irannejad,r.,zastrow,von,m.,2014.gpcrsignalingalongtheendocyticpathway.curr.opin.cellbiol.27,109-116.doi:10.1016/j.ceb.2013.10.003isomura,i.,yasuda,y.,tsujimura,k.,takahashi,t.,tochikubo,k.,morita,a.,2005.recombinantcholeratoxinbsubunitactivatesdendriticcellsandenhancesantitumorimmunity.microbiol.immunol.49,79-87.itoh,k.,watanabe,a.,funami,k.,seya,t.,matsumoto,m.,2008.theclathrin-mediatedendocyticpathwayparticipatesindsrna-inducedifn-betaproduction.j.immunol.181,5522-5529.juliano,r.l.,carver,k.,2015.cellularuptakeandintracellulartraffickingofoligonucleotides.advanceddrugdeliveryreviews87,35-45.doi:10.1016/j.addr.2015.04.005kagan,j.c.,su,t.,homg,t.,chow,a.,akira,s.,medzhitov,r.,2008.tramcouplesendocytosisoftoll-likereceptor4totheinductionofinterferon-beta.naturepublishinggroup9,361-368.doi:10.1038/ni1569kato,h.etal.celltype-specificinvolvementofrig-iinantiviralresponse.immunity23,19-28(2005).kawai,t.,akira,s.,2010.theroleofpattern-recognitionreceptorsininnateimmunity:updateontoll-likereceptors.naturepublishinggroup11,373-384.doi:10.1038/ni.1863l.,canterbury,j.d.,weston,j.,noble,w.s.&maccoss,m.j.semisupervisedlearningforpeptideidentificationfromshotgunproteomicsdatasets.nat.methods4,923-925(2007).klein,d.c.g.etal.cd14,tlr4andtramshowdifferenttraffickingdynamicsduringlpsstimulation.traffic16,677-690(2015).klinman,d.m.,gursel,i.,klaschik,s.,dong,l.,currie,d.,shirota,h.,2005.therapeuticpotentialofoligonucleotidesexpressingimmunosuppressivettagggmotifs.ann.n.y.acad.sci.1058,87-95.doi:10.1196/annals.1359.015kumari,s.,mg,s.,mayor,s.,2010.endocytosisunplugged:multiplewaystoenterthecell.cellres.20,256-275.doi:10.1038/cr.2010.19levy,h.b.,baer,g.,baron,s.,buckler,c.e.,gibbs,c.j.,ladarola,m.j.,london,w.t.,rice,j.,1975.amodifiedpolyriboinosinic-polyribocytidylicacidcomplexthatinducesinterferoninprimates.j.infect.dis.132,434-439.matsumoto,m.,oshiumi,h.,seya,t.,2011.antiviralresponsesinducedbythetlr3pathway.rev.med.virol.21,67-77.doi:10.1002/rmv.680mehta,a.,baltimore,d.,2016.micrornasasregulatoryelementsinimmunesystemlogic.natrevimmunol16,279-294.doi:10.1038/nri.2016.40merad,m.,sathe,p.,helft,j.,miller,j.,mortha,a.,2013.thedendriticcelllineage:ontogenyandfunctionofdendriticcellsandtheirsubsetsinthesteadystateandtheinflamedsetting.annu.rev.immunol.31,563-604.doi:10.1146/annurev-immunol-020711-074950panda,s.,nilsson,j.a.&gekara,n.o.deubiquitinasemysm1regulatesinnateimmunitythroughinactivationoftraf3andtraf6complexes.immunity43,647-659(2015).pandey,s.,kawai,t.&akira,s.microbialsensingbytoll-likereceptorsandintracellularnucleicacidsensors.coldspringharbperspectbiol7,a016246(2014).pelka,k.,shibata,t.,miyake,k.,latz,e.,2016.nucleicacid-sensingtlrsandautoimmunity:novelinsightsfromstructuralandcellbiology.immunol.rev.269,60-75.doi:10.1111/imr.12375peter,m.,bode,k.,lipford,g.b.,eberle,f.,heeg,k.,dalpke,a.h.,2008.characterizationofsuppressiveoligodeoxynucleotidesthatinhibittoll-likereceptor-9-mediatedactivationofinnateimmunity.immunology123,118-128.doi:10.1111/j.1365-2567.2007.02718.xpolitz,j.c.,taneja,k.l.,singer,r.h.,1995.characterizationofhybridizationbetweensyntheticoligodeoxynucleotidesandrnainlivingcells.nucleicacidsres.23,4946-4953.rajaiah,r.,perkins,d.j.,ireland,d.d.c.,vogel,s.n.,2015.cd14dependenceoftlr4endocytosisandtrifsignalingdisplaysligandspecificityandisdissociableinendotoxintolerance.pnas112,8391-8396.doi:10.1073/pnas.1424980112ranjith-kumar,c.t.,duffy,k.e.,jordan,j.l.,eaton-bassiri,a.,vaughan,r.,hoose,s.a.,lamb,r.j.,sarisky,r.t.,kao,c.c.,2008.single-strandedoligonucleotidescaninhibitcytokineproductioninducedbyhumantoil-likereceptor3.mol.cell.biol.28,4507-4519.doi:10.1128/mcb.00308-08roers,a.,hiller,b.,hornung,v.,2016.recognitionofendogenousnucleicacidsbytheinnateimmunesystem.immunity44,739-754.doi:10.1016/j.immuni.2016.04.002rosadini,c.v.,zanoni,i.,odendall,c.,green,e.r.,paczosa,m.k.,philip,n.h.,brodsky,i.e.,mecsas,j.,kagan,j.c.,2015.asinglebacterialimmuneevasionstrategydismant1esbothmyd88andtrifsignalingpathwaysdownstreamoftlr4.cellhostandmicrobe18,682-693.doi:10.1016/j.chom.2015.11.006sacre,s.,lo,a.,gregory,b.,stephens,m.,chamberlain,g.,stott,p.,brennan,f.,2015.oligodeoxynucleotideinhibitionoftoll-likereceptors3,7,8and9suppressescytokineproductioninahumanrheumatoidarthritismodel.eur.j.immunol.n/a-n/a.doi:10.1002/eji.201546123sarvestani,s.t.etal.sequence-dependentoff-targetinhibitionoftlr7/8sensingbysyntheticmicrornainhibitors.nucleicacidsres.43,1177-1188(2015).scita,g.,difiore,p.p.,2010.theendocyticmatrix.nature463,464-473.doi:10.1038/nature08910scheiermann,j.&klinman,d.m.clinicalevaluationofcpgoligonucleotidesasadjuvantsforvaccinestargetinginfectiousdiseasesandcancer.vaccine32,6377-6389(2014).shalek,a.k.etal.single-cellrna-seqrevealsdynamicparacrinecontrolofcellularvariation.nature510,363-369(2014).shannon,p.etal.cytoscape:asoftwareenvironmentforintegratedmodelsofbiomolecularinteractionnetworks.genomeres.13,2498-2504(2003).sharma,v.k.,watts,j.k.,2015.oligonucleotidetherapeutics:chemistry,deliveryandclinicalprogress.futuremedchem7,2221-2242.doi:10.4155/fmc.15.144shirota,h.,gursel,i.,gursel,m.,klinman,d.m.,2005.suppressiveoligodeoxynucleotidesprotectmicefromlethalendotoxicshock.thejournalofimmunology174,4579-4583.doi:10.4049/jimmunol.174.8.4579silva,a.c.,lopes,c.m.,sousalobo,j.m.,amaral,m.h.,2015.nucleicacidsdeliverysystems:achallengeforpharmaceuticaltechnologists.curr.drugmetab.16,3-16.a.e.,hasan,m.,vargas,l.,saidi,h.,bosquet,n.,legrand,r.,smith,c.i.e.,spetz,a.-l.,2012.single-strandeddnaoligonucleotidesinhibittlr3-mediatedresponsesinhumanmonocyte-deriveddendriticcellsandinvivoincynomolgusmacaques.blood120,768-777.doi:10.1182/blood-2011-12-397778smole,u.,radauer,c.,lengger,n.,svoboda,m.,rigby,n.,bublin,m.,gaier,s.,hoffmann-sommergruber,k.,jensen-jarolim,e.,mechtcheriakova,d.,breiteneder,h.,2015.themajorbirchpollenallergenbetv1inducesdifferentresponsesindendriticcellsofbirchpollenallergicandhealthyindividuals.plosone10,e0117904.doi:10.1371/journal.pone.0117904soifer,h.s.,koch,t.,lai,j.,hansen,b.,hoeg,a.,oerum,h.,stein,c.a.,2012.silencingofgeneexpressionbygymnoticdeliveryofantisenseoligonucleotides.methodsmol.biol.815,333-346.doi:10.1007/978-1-61779-424-7_25sorkin,a.,zastrow,von,m.,2009.endocytosisandsignalling:intertwiningmolecularnetworks.nat.rev.mol.cellbiol.10,609-622.doi:10.1038/nrm2748soulet,d.,covassin,l.,kaouass,m.,charest-gaudreault,r.,audette,m.,poulin,r.,2002.roleofendocytosisintheinternalizationofspermidine-c(2)-bodipy,ahighlyfluorescentprobeofpolyaminetransport.biochem.j.367,347-357.doi:10.1042/bj20020764stein,c.a.,hansen,j.b.,lai,j.,wu,s.,voskresenskiy,a.,a.,worm,j.,m.,souleimanian,n.,miller,p.,soifer,h.s.,castanotto,d.,benimetskaya,l.,h.,koch,t.,2010.efficientgenesilencingbydeliveryoflockednucleicacidantisenseoligonucleotides,unassistedbytransfectionreagents.nucleicacidsres.38,e3-e3.doi:10.1093/nar/gkp841sun,r.,zhang,y.,lv,q.,liu,b.,jin,m.,zhang,w.,he,q.,deng,m.,liu,x.,li,g.,li,y.,zhou,g.,xie,p.,xie,x.,hu,j.,duan,z.,2011.toil-likereceptor3(tlr3)inducesapoptosisviadeathreceptorsandmitochondriabyup-regulatingthetransactivatingp63isoformalpha(tap63alpha).j.biol.chem.286,15918-15928.doi:10.1074/jbc.m110.178798takeda,k.,kaisho,t.&akira,s.toll-likereceptors.annu.rev.immunol.21,335-376(2003).tatematsu,m.,yoshida,r.,morioka,y.,ishii,n.,funami,k.,watanabe,a.,saeki,k.,seya,t.,matsumoto,m.,2016.raftlincontrolslipopolysaccharide-inducedtlr4internalizationandticam-1signalinginacelltype-specificmanner.j.immunol.196,3865-3876.doi:10.4049/jimmunol.1501734veillette,a.&chen,j.sirpα-cd47immunecheckpointblockadeinanticancertherapy.trendsinimmunology39,173-184(2018).vercammen,e.,staal,j.,beyaert,r.,2008.sensingofviralinfectionandactivationofinnateimmunitybytoll-likereceptor3.clin.microbiol.rev.21,13-25.doi:10.1128/cmr.00022-07weber,c.,müller,c.,podszuweit,a.,montino,c.,vollmer,j.,forsbach,a.,2012.toll-likereceptor(tlr)3immunemodulationbyunformulatedsmallinterferingrnaordnaandtheroleofcd14(intlr-mediatedeffects).immunology136,64-77.doi:10.1111/j.1365-2567.2012.03559.xzhu,f.-g.,reich,c.f.,pisetsky,d.s.,2002.inhibitionofmurinemacrophagenitricoxideproductionbysyntheticoligonucleotides.j.leukoc.biol.71,686-694.当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1