一种异养硝化-好氧反硝化菌及其培养方法和应用与流程
本发明涉及微生物
技术领域:
,尤其涉及一种异养硝化-好氧反硝化菌及其培养方法和应用。
背景技术:
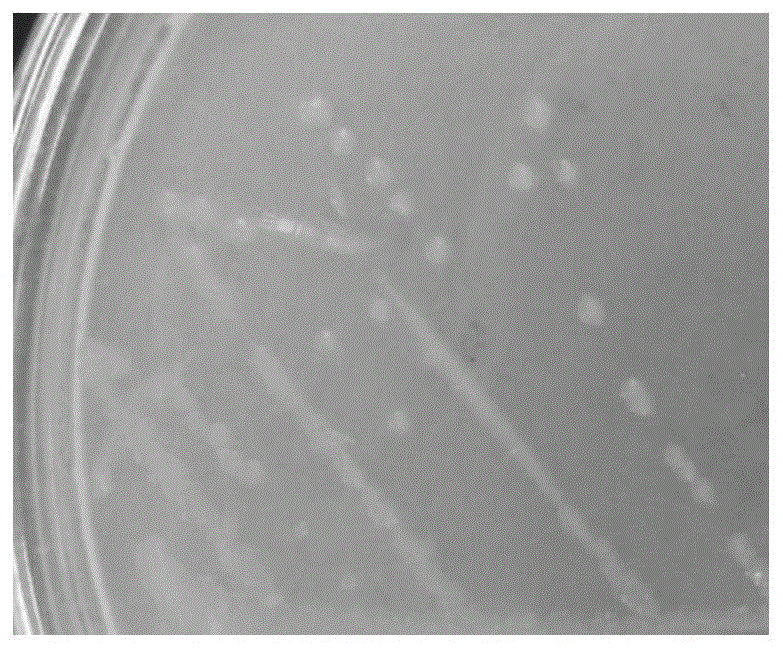
:生物脱氮技术具有高效、低耗、安全稳定和脱氮彻底等众多优点,是目前最具发展前景的水体脱氮技术。传统的生物脱氮技术包括硝化和反硝化作用两个过程。硝化作用包括两步连续的氧化反应:nh4+-n在氨氧化菌的作用下转化为no2--n;no2--n在亚硝酸盐氧化菌的作用下进一步转化为no3--n;反硝化过程是指no3--n被反硝化细菌转化成气体氮产物的过程。虽然传统生物脱氮工艺可有效的去除水体中的氮素,但目前存在以下不足:(1)硝化过程产生酸,需加碱中和,使处理成本增加,易造成二次污染;(2)自养硝化菌系统抗冲击能力差,其生长在高浓度有机物下受到抑制;(3)硝化和反硝化对环境的要求不同,难以在时间和空间上统一,通常需要分开,使运行成本和基建面积增大;(4)自养硝化细菌在电子传递过程中需消耗大量能量、产能少、生长缓慢、世代时间长、在环境中对氧和营养物利用低,生物浓度难以处于较高水平,使废水处理水力停留时间长,造成运行费用增加;(5)其对高浓度含氮废水脱氮效果不理想,常需要对高浓度的含氮废水先进行稀释和吹脱,但该工艺处理时间长、费用高,同时反硝化作用也会因碳源不足而受到限制,处理过程很难保持连续高效稳定进行。随着脱氮理论研究的不断深入,随之涌现出新型脱氮技术和新型脱氮微生物,目前已报道的新型脱氮技术有厌氧氨氧化、短程硝化反硝、同步硝化反硝化、异养硝化、好氧反硝化等。新型脱氮微生物异养硝化-好氧反硝化菌(heterotrophicnitrification-aerobicdenitrification,hn-ad)可以同时进行硝化和反硝化作用,避免了传统工艺中好氧硝化和缺氧反硝化分离、投入大等缺点,在废水处理应用中具有成本低、无二次污染、生长快速、耐有机负荷、脱氮效率高等特点,具有广阔的研究价值和应用前景。异养硝化-好氧反硝化菌生长快,脱氮效率高,易保存于处理系统中;菌株基质和产物的多样性,可能有利于其与其他菌株共存,扩大了其应用范围。随着研究人员对同步硝化反硝化的深层次研究,目前将从河流、土壤、生活污水、活性污泥等环境中分离并鉴定近20种不同菌属的异养硝化-好氧反硝化菌,如raoultellasp.sari01、alcaligenesfaecaliswy-01、zobellellasp.b307等。目前对hn-ad的研究,除了对脱氮优良菌株的筛选外,还主要集中在碳源、c/n、温度、ph、溶解氧浓度等环境影响因素对菌株脱氮性能的影响方面。例如,大部分的hn-ad菌都能以乙酸盐、柠檬酸盐、琥珀酸盐等小分子有机物为碳源,部分菌也能利用葡萄糖,如bacilluspumilusbp-171sf16;大多数hn-ad菌c/n范围较宽泛,最适c/n范围多在2-20之间,如pseudomonassp.hn、acinetobactersp.y1、acinetobacter.spyn3,最适c/n分别为7、10、15;hn-ad菌主要在中温条件下进行生长,最适温度范围为28-37℃,如enterobacterasburiaeyt和acinetobactersp.sq2最适温度分别为30、28℃;hn-ad菌适应的ph范围一般为中性或偏碱性,当ph<5或>10,hn-ad菌不具有生长活性。目前报道的很多hn-ad菌株具有能够在高浓度nh4-n条件下生长代谢的特点,例如,acinetobactorsp.jq1004能够耐受的初始nh4+-n浓度为2500mg·l-1。但目前对于环境适应性强且能耐受高浓度no3--n和no2--n的假单胞菌研究报道较少。分离筛选到耐受高浓度no3--n和no2--n的异养硝化-好氧反硝化菌可为环境治理提供菌源保障,其脱氮性能研究为其在高浓度有机氮污水脱氮处理过程中的应用奠定理论基础。大量研究表明,hn-ad菌在转化nh4+-n为气态氮化物的同时,还利用氮源发生同化作用,转化为菌株细胞内的含氮有机化合物。如不动杆菌acinetobactesp.hy2在异养硝化-好氧反硝化过程中的氮平衡分析结果表明,接种该菌48h后,去除的nh4+-n中32.00%经过同化作用转化为菌体细胞内氮,66%经异养硝化-好氧反硝化作用完全去除。由此可见,hn-ad菌的生物同化作用成为限制总氮去除率的一个主要原因。技术实现要素:本发明要解决的技术问题是克服现有技术的不足,提供一种具有耐高温和耐碱特性且在低c/n条件下具有很强的异养硝化-好氧反硝化性能,同时具有耐受高浓度no3--n和no2--n能力的异养硝化-好氧反硝化菌,还相应提供该菌的培养方法,以及在处理含氮废水中的应用。为解决上述技术问题,本发明采用以下技术方案:一种异养硝化-好氧反硝化菌,所述异养硝化-好氧反硝化菌为假单胞菌xn2(pseudomonasstutzerixn2),保藏于广东省微生物菌种保藏中心,保藏时间为2019年1月29日,保藏编号为gdmccno:60531。作为一个总的发明构思,本发明还提供一种上述的异养硝化-好氧反硝化菌的培养方法,包括如下步骤:将假单胞菌xn2接种于lb培养基中培养至指数生长期,所得菌悬液离心去除上清液,洗涤后加无菌水制成od600为0.6~0.8的菌悬液。作为一个总的发明构思,本发明还提供一种上述的异养硝化-好氧反硝化菌在处理含氮废水中的应用。作为应用的优选方案:假单胞菌xn2以有机酸为碳源、nh4+为氮源进行异养硝化脱氮。含氮有机废水的ph值为7~11,温度为20~45℃,c/n比为10~50,nh4+-n浓度为≤500mg·l-1,异养硝化脱氮的转速为100~200rpm,假单胞菌xn2的接种量为3%~15%。假单胞菌xn2以有机酸为碳源、no3-为氮源进行好氧反硝化脱氮。所述含氮有机废水的ph值为7~11,温度为20~45℃,c/n比为10~50,no3--n浓度≤2000mg·l-1,好氧反硝化脱氮的转速为100~200rpm,假单胞菌xn2的接种量为3%~15%。假单胞菌xn2以有机酸为碳源、no2-为氮源进行好氧亚硝化脱氮。含氮有机废水的ph值为7~11,温度为20~45℃,c/n比为10~50,no2--n浓度为≤100mg·l-1,好氧反硝化脱氮的转速为100~200rpm,假单胞菌xn2的接种量为3%~15%。所述有机酸为柠檬酸钠、乙酸钠和琥珀酸钠中的一种或多种。与现有技术相比,本发明的优点在于:1、本发明的菌株xn2在ph为10-11范围内,nh4+-n、tn仍能达到89.77%、79.08%以上;45℃时,nh4+-n去除率为60.67%,tn去除率为55.38%;在c/n=10时,nh4+-n去除率达到100%,tn去除率达到90.34%。说明菌株xn2具有耐高温和耐碱特性且在低c/n条件下具有很强的异养硝化性能。2、菌株xn2在c/n范围较宽(10-50)的条件下,具有较强的异养硝化、好氧反硝化和好氧亚硝化脱氮能力。菌株xn2能耐受的最高nh4+-n浓度为500mg·l-1,nh4+-n去除率为69.77%;菌株xn2能耐受的最高no3--n和no2--n浓度分别为2000mg·l-1和1000mg·l-1,且具有高效的no3--n和no2--n去除能力。3、菌株xn2脱氮的主要形式是细胞同化作用和好氧反硝化作用。初始no3--n浓度较低时,菌株xn2在培养初期主要转化成胞内氮,少部分转化成含氮气体去除,随着培养时间的延长,细胞内的氮逐渐减少,而转化为含氮气体的总氮量逐渐增加;初始no3--n浓度较高时,随着培养时间的延长,同化作用转化成细胞内的氮增加量不明显,而转化为含氮气体的总氮量增加显著,菌株xn2好氧反硝化过程中的no3--n主要不是同化作用转化为细胞内氮,而是通过好氧反硝化作用转化成含氮气体。假单胞菌xn2(pseudomonasstutzerixn2),保藏于广东省微生物菌种保藏中心(简称:gdmcc),地址为广州市先烈中路100号大院59号楼5楼,广东省微生物研究所,保藏编号为gdmccno:60531,保藏时间为2019年1月29日。附图说明图1为菌株xn2平板培养菌落形态图。图2为菌株xn2形态透射电镜图,其中,(a)为菌落群体,(b)为单个菌落。图3为菌株xn216srdnapcr电泳图。图4为菌株xn216srdna系统发育树图。图5为菌株xn2功能基因napa、nirs、nosz电泳图图6为菌株xn2功能基因系统发育树图,其中,上图为napa、中图为nirs、下图为nosz。图7为菌株xn2异养硝化过程生长与脱氮曲线图。图8为菌株xn2好氧反硝化过程的生长与脱氮曲线图。图9为菌株xn2好氧亚硝化过程的生长与脱氮曲线图。图10为不同碳源对菌株xn2生长与异养硝化脱氮性能影响图。图11为不同c/n对菌株xn2生长与异养硝化脱氮性能影响图。图12为不同ph对菌株xn2生长与异养硝化脱氮性能影响图。图13为不同温度对菌株xn2生长及异养硝化脱氮性能影响图。图14为不同溶氧量对菌株xn2生长及异养硝化脱氮性能影响图。图15为不同接种量对菌株xn2生长异养硝化脱氮性能影响图。图16为不同c/n对菌株xn2生长及好氧反硝化脱氮性能影响图。图17为不同c/n对菌株xn2生长及好氧亚硝化脱氮性能影响图。图18为不同nh4+-n浓度对菌株xn2的生长曲线图。图19为不同nh4+-n浓度对菌株xn2的生长及脱氮性能影响图。图20为不同no3--n浓度对菌株xn2的生长曲线图。图21为不同no3--n浓度对菌株xn2的生长及脱氮性能影响图。图22为不同no2--n浓度对菌株xn2的生长曲线图。图23为不同no2--n浓度对菌株xn2生长及脱氮性能影响图。具体实施方式以下结合具体优选实施例对本发明作进一步描述,但并不因此而限制本发明的保护范围。实验材料和方法所分离的菌种来源于龙岩市龙津河沉积物。培养基配方如表1所示:表1培养基配方注:以上培养基均需在121℃条件下灭菌20min后方可使用;本实验过程中装量为100ml的各式培养基均装于250ml三角瓶中;装量为200ml的各式培养基均装于500ml三角瓶中。主要实验仪器如表2所示:表2主要实验仪器常规指标的检测项目及分析方法如表3所示:表3常规指标的检测项目及分析方法检测项目分析方法nh4+-n纳氏试剂分光光度法no3--n麝香草酚光光度法no2--nn-(1-萘基)-乙二胺光度法tn碱性过硫酸钾消解分光光度计od600超微量分光光度计phub-7精密ph计好氧反硝化菌初筛与分离纯化:将稀释菌液涂布于btb平板培养基上,30℃培养1-2d后,挑取培养基变蓝部位的单菌落划线培养2-3次,接种到牛肉膏蛋白胨试管斜面保藏,4℃冰箱保存,以备复筛。通过初筛得到10株异养硝化-好氧反硝化菌;通过复筛,根据各个菌株的nh4+-n和no3--n的去除率,确定一株nh4+-n和no3--n去除率最高的菌株,编号为xn2。种子液制备:将纯菌按3%接种量接入装量为100mllb培养基,30℃、150rmp培养至对数期,取菌液4000rpm离心10min,弃上清后沉淀用无菌水洗涤2-3次,随后用无菌水调洗涤后的菌悬液浓度,使其od600=0.8左右,作为种子液。异养硝化-好氧反硝化菌复筛:将种子液分别以3%接种量接入装有100ml的异养硝化培养基或好氧反硝化培养基的250ml三角锥瓶,30℃,150rpm振荡培养2d,分别测定培养前后培养液中nh4+-n或no3--n的浓度,并计算nh4+-n或者no3--n的去除率。挑取nh4+-n和no3--n去除率较高的菌株,甘油管-20℃低温保存,以备进一步鉴定。注:nh4+-n去除率=[c(初始氨氮)-c(培养后剩余的氨氮)]/c(初始氨氮)×100%n03--n去除率=[c(初始硝酸盐氮)-c(培养后剩余的硝酸盐氮)]/c(初始硝酸盐氮)×100%对于分离纯化得到的目的菌株,本实验采用两种方法保藏如表4所示。表4菌种保藏方法菌株鉴定形态学鉴定:通过平板观察初步鉴定菌落形态;采用扫描电镜观察菌体形态与特征;将菌株xn2接种于牛肉膏蛋白胨琼脂培养基上,30℃培养2d后观察菌落形态特征如图1所示:菌株xn2形成的菌落为透明橘黄色、圆形、表面湿润、易挑起,随着培养时间的延长,菌落的透明度减小、体积增大、颜色变深。菌种形态如图2透射电镜图所示:菌体呈杆状,大小为4.5×18μm,有鞭毛、无芽孢。生理生化鉴定:菌株生理生化特性鉴定参照《常见细菌系统鉴定手册》和《伯杰氏细菌鉴定手册》进行,主要包括革兰氏染色、葡萄糖发酵、淀粉水解、接触酶和硝酸盐还原试验等。结果如下:菌株xn2经革兰氏染色鉴定该菌为革兰氏阳性,菌株xn2是需氧型的,4℃、41℃条件下可生长,氧化酶、接触酶、吲哚、产硫化氢、柠檬酸盐、绿脓菌素、尿素水解、淀粉水解、油脂水解、硝酸盐还原、明胶液化实验均为阳性,说明该菌株对尿素、淀粉、油脂等大分子底物均能进行水解利用。mr、vp石蕊牛奶、乙酸氧化实验均为阴性,葡萄糖、乳糖氧化发酵中不产酸不产气。以上结果表明:菌株xn2的生理生化指标与假单胞菌的生理生化特性相似。分子生物学鉴定(1)16srdna扩增和测序:采用sanprep柱式dna胶回收试剂盒(生工生物工程上海股份有限公司)提取菌株的dna,以其为模板进行16srdna片段的pcr扩增,pcr反应体系、引物序列及反应条件如下表5、6和7所示。表5pcr反应体系反应体系体积(μl)双蒸水40.510×pcrbuffer5dntp1f1r1taq酶0.5模板dna2pcr产物用1.0%的琼脂糖凝胶电泳进行检测,条带单一、大小正确后,将扩增产物送至苏州金唯智公司完成菌株16srdna序列测定,测序结果递交至ncbi数据库进行比对,并将序列提交到genbank数据库,选取同源性较高的序列利用mega7.0软件,采用neighbor-joining法构建系统发育树。(2)菌株功能基因的扩增:反硝化过程一般有四步反应:no3--n→no2--n→no→n2o→n2,通过四步反应将no3--n转化为n2,反应分别由硝酸盐还原酶(nar)、亚硝酸盐还原酶(nir)、一氧化氮还原酶(nor)和一氧化二氮还原酶(nos)催化实现生物脱氮,相对应的功能基因分别为nar/nap、nir、nor和nos,其中nar一般分为两种,一种是膜质硝酸盐还原酶(nar)只在厌氧条件下才表达,另一种是周质硝酸盐还原酶(nap),在好氧条件下占主导地位,在厌氧和好氧条件下都可以表达。以目标菌株16srdna为模板进行napa、nirs、norb、nosz功能基因pcr扩增,功能基因引物序列和pcr反应条件分别如表6和表7所示:表6napa、nirs、nosz、norb功能基因引物序列表7pcr反应条件(1)菌株xn216srdna扩增及测序结果:菌株xn216srdnapcr扩增产物经琼脂糖凝胶电泳结果如图3所示:目标条带在1000-2000bp之间且条带明亮无杂带,菌株xn216srdna测序获得长度为1375bp序列,将所得的菌株序列提交至genbank中通过blast检索,应用mega7软件,以neighbor-joining法绘制系统发育树如图4所示,菌株xn2与pseudomonasstutzeri1w1-1a(cp027664)相似度最高达99%,结合菌株的形态特征和生理生化特性,可初步确定菌株xn2属于施氏假单胞菌(pseudomonasstutzeri)。(2)菌株xn2硝酸盐还原酶基因napa扩增结果:硝酸盐还原酶催化反硝化反应的第一步,该酶主要有周质硝酸还原酶和膜结合硝酸还原酶两种存在形式。周质硝酸还原酶基因与膜结合硝酸还原酶基因没有关联,即相互独立,互不影响。周质硝酸盐还原酶是一种二聚体类物质,由napa和napb两个亚单位结构组成。napa基因负责编码大亚基napa,而napb基因负责编码小亚基napb;周质硝酸还原酶基因操纵子除了napa和napb基因之外,还存在1个napc基因。有氧或者无氧条件下,由基因napa编码的nap催化中心部位均可实现其功能,但有氧条件下优先表达。本试验利用引物napaf-napar,对菌株xn2的dna样品进行pcr扩增、琼脂糖凝胶电泳,结果如图5所示:目标条带在750-1000bp之间且条带明亮,经测序获得长度为849bp的napa基因序列,将所得的基因序列提交至genbank中通过blast比对,挑选最相似的序列进行多重序列比对分析后构建系统发育树,结果如图6的上图所示:菌株xn2与pseudomonasstutzerinctc10450(lr134319)的napa基因相似度最高98.93%,这与xn216srdna测序结果一致。综上所述,菌株xn2存在周质硝酸还原酶,推测菌株xn2可以在有氧条件下具有还原硝酸盐的功能。(3)菌株xn2亚硝酸还原酶基因nirs扩增结果:由亚硝酸盐还原酶催化,亚硝酸盐可转化为no,该反应是反硝化作用的标志性反应,是反硝化过程中最重要的限速步骤。亚硝酸盐还原酶的存在方式主要有2种:细胞色素cd1型亚硝酸盐还原酶和含铜型亚硝酸还原酶,分别由nirs基因和nirk基因编码。普遍的观点认为同一株菌中不会同时出现nirs和nirk基因,但也有人发现了能同时含有nirs和nirk这2种基因的菌株。本试验利用两对引物nirsf-nirsr和nirkf-nirkr对xn2的dna样品进行pcr扩增,琼脂糖凝胶电泳结果如图5所示:nirs基因在750-000bp之间存在明亮条带,而nirk基因无条带,说明菌株xn2具有细胞色素cd1型亚硝酸盐还原酶。经测序获得长度为852bp的nirs基因序列,将所得的基因序列提交至genbank中,通过blast比对,挑选最相似的序列进行多重序列比对分析后构建系统发育树,结果如图6的中图所示:菌株xn2与多株pseudomonasstutzeri的nirs基因基因相似性均达99%,其中与pseudomonasstutzerilys-68(gu474546)相似度最高99.28%。综上所述,菌株xn2存在细胞色素cd1型亚硝酸盐还原酶,推测菌株xn2具有还原亚硝酸盐的功能。(4)菌株xn2一氧化氮还原酶基因norb扩增结果:一氧化氮还原酶位于细胞膜上,能够将no还原为n2o。该酶主要分为3种类型:一种是单体酶qnor,由qnorb基因编码;一种既可以是细胞色素c,也可以是苯二酚的qcuanor型一氧化氮还原酶的电子供体,该酶的小亚基中含2个cua,大亚基中含2个血红素b和feb;还有一种是细胞色素bc型酶cnor,由norb和norc两个亚基组成,其中norb基因编码大亚基norb,norc基因编码小亚基norc。本试验利用引物norbf-norbr对xn2的dna样品进行pcr扩增,琼脂糖凝胶电泳。电泳结果显示norb基因无条带,说明菌株xn2没有一氧化氮还原酶,推测菌株xn2不能将no还原为n2o,可能直接将no还原为n2。(5)菌株xn2一氧化二氮还原酶基因nosz扩增结果:nosz被广泛用作研究环境样品中反硝化细菌的功能标志基因,只有nosz基因编码将n2o还原成n2的一氧化二氮还原酶,这是反硝化作用最后一步反应的关键酶。该酶是一种可溶性含铜离子蛋白酶,位于膜外周质中,其编码基因由3个转录单元组成:nosz、nosr和nosdfyl,其中,nosz基因的结构最大,常用来编码催化亚基。本试验利用引物noszf-noszr对xn2的dna样品进行pcr扩增,琼脂糖凝胶电泳结果如图5所示:250-500bp之间出现明亮条带,经测序获得长度为262bp的序列,将其提交至genbank中,通过blast比对,挑选最相似的序列进行多重序列比对分析后构建系统发育树,结果如图6的下图所示:菌株xn2与多株pseudomonasstutzeri的nosz基因基因相似度均达98%,其中与pseudomonasstutzeridw2-1(cp027543)相似度最高。综上所述,推测菌株xn2存在一氧化二氮还原酶,能够将no还原成n2。综上,菌株xn2具有napa、nirs和nosz功能基因,这表明了该菌具有nar、nir和nos酶,从分子水平上证明了菌株xn2具有好氧反硝化能力,也从酶学的角度进一步证明了菌株xn2具有将no3--n和no2--n还原为n2的能力,推测菌株xn2的脱氮途径为no3--n→no2--n→no→n2。菌株xn2硝化性能研究本实验探究菌株xn2对不同n源脱除规律,3%接种量,初始氮浓度为100mg·l-1,以乙酸钠为c源,c/n=10,将xn2接入装量为200ml的不同培养基,30℃、150rpm培养48h,每6h取样一次测定。n源、培养基、培养条件及测定指标等如表8所示:表8硝化性能研究菌株xn2异养硝化性能研究结果:以(nh4)2so4为唯一氮源的异养硝化脱氮过程如图7所示:菌株xn2在0-12h内处于适应期,在12-36h内处于对数生长阶段,在36h到达稳定期,之后进入衰退期,od600开始出现下降的趋势;12h前菌株xn2的nh4+-n、tn去除率较低,其主要发生在对数生长期,xn2培养至36hnh4+-n、tn去除率均达到最大,分别为99.36%和93.96%;整个异养硝化过程没有检测到no2--n,但在18h开始出现少量no3--n的积累,随后no3--n的积累量逐渐减少至零;菌株xn2在整个脱氮过程中ph由7.44逐渐先升高至8.27,36h后下降为7.85,说明在此期间伴随有反硝化反应,硝化产生的酸和反硝化产生的碱发生中和反应,使培养基中的ph较为稳定。综上,菌株xn2可同时进行硝化和反硝化反应,这与已报道的研究结果相似,如pseudomonassp.y3,以nh4cl为唯一氮源时,0-10h内处于适应期,对nh4+-n的去除率较低,nh4+-n浓度由142.99mg·l-1降至139.89mg·l-1,在10h后进入对数生长期,最高nh4+-n去除率达98.69%,在整个nh4+-n降解过程中无明显no2--n和no3--n的积累;alteromonasmacleodii8d异养硝化过程中,经过48h培养,对nh4+-n去除率仅58.64%,在整个脱氮过程中,无明显no3--n和no2--n的积累。菌株xn2的好氧反硝化性能研究结果:以kno3为唯一氮源的好氧反硝化脱氮过程如图8所示:菌株xn2在0-6h内处于适应期,在6-18h内处于对数生长期,迅速生长,od600达到0.261,在18h后生长达到稳定期;菌株xn2在0-12h内对no3--n和tn的去除较为缓慢,18hno3--n去除率达100%,tn去除率达到94.90%,说明xn2脱氮主要发生在对数生长期;菌株xn2好氧反硝化过程中,no2--n积累量很小,对数生长期时达到8.13mg·l-1,之后逐渐降低至0mg·l-1,可能是因为no2--n替代no3--n作为主要的电子受体,同时会诱导亚硝酸盐还原酶的产生,从而将no3--n还原为反硝化气体;菌株xn2在培养过程中ph逐渐升高,说明好氧反硝化是个产碱的过程。综上,菌株xn2具有较强的好氧反硝化性能,能够将no3--n和no2--n通过反硝化过程转化为气态产物,推测菌株xn2的反硝化途径可能是no3--n→no2--n→气态产物。菌株xn2在18hno3--n去除率达100%,平均反硝化速率达到5.56mg·(l·h)-1,与目前已报道的菌株相比,xn2具有较强的好氧反硝化性能,例如,acinetobacterbaumanniiwj6,以kno3为唯一氮源,初始no3--n浓度为50mg·l-1,培养32h的最大no3--n去除率81.70%,tn去除率68.5%,其中最大反硝化速率2.99mg·(l·h)-1;pseudomonassp.lhj-1,以kno3为唯一氮源,初始no3--n浓度为6mg·l-1,在0-6h内,no3--n去除最快,平均反硝化速率0.817mg·(l·h)-1,培养30hno3--n去除率最高92.46%,平均反硝化速率0.18mg·(l·h)-1,在整个好氧反硝化脱氮过程中,伴随有少量no2--n的积累。菌株xn2的好氧亚硝化性能研究结果:以nano2为唯一氮源的好氧亚硝化脱氮过程如图9所示:菌株xn2在0-6h内处于适应期,在6-18h内处于对数生长期,迅速生长,od600达到0.226,在18h后生长达到稳定期;菌株xn2在0-6h内对no2--n和tn的去除率较低,12hno2--n去除率基本达到100%,tn去除率达到92.65%,说明xn2亚硝化脱氮主要发生在对数生长期;菌株xn2好氧亚硝化过程中几乎没有no3--n的积累,nh4+-n积累量很小,在对数生长期时,达到5.68mg·l-1,之后逐渐降低至0mg·l-1。菌株xn2在好氧亚硝化脱氮过程中ph由7.48逐渐升高至8.46。综上,菌株具有较好的好氧亚硝化性能,能将no2--n转化为气态产物。菌株xn2在12hno2--n去除率达100%,平均反硝化速率达到8.33mg·(l·h)-1,与目前已报道的菌株相比,xn2具有更强的好氧亚硝化性能,例如,rhizobiumsp.zb612,以nano2为唯一氮源,初始no2--n浓度为100mg·l-1时,经过36h培养最大no2--n去除率60.34%,平均反硝化速率1.68mg·(l·h)-1;acinetobactersp.tn-14,在初始no2--n浓度为100mg·l-1时,经过30h培养,菌株tn-14的最大no2--n去除率78.22%,平均反硝化速率2.55mg·(l·h)-1,在整个好氧亚硝化脱氮过程中,有少量no3--n的积累14.26mg·l-1。不同环境因素对菌株xn2异养硝化脱氮性能影响本实验设置不同c源、c/n、温度、ph、溶解氧、接种量因素确定菌株xn2最适培养条件。接种量3%,以乙酸钠为c源,硫酸铵为n源,初始nh4+-n浓度为100mg·l-1,c/n=10(本实验只改变研究的单个环境因素,其他培养条件均按以上要求设置不变),将xn2接入装量为100ml异养培养基,30℃、150rmp培养24h,取样测od600、ph、tn和nh4+-n含量,计算其tn和nh4+-n去除率。不同环境因素下其设置参数如表9所示:表9不同环境因素的设置参数不同碳源对菌株xn2异养硝化脱氮性能的影响:碳源是微生物细胞代谢必不可少的营养物质,不仅是组成微生物细胞组织结构的重要物质,还能为机体提供维持生命活动所需的能量,是影响微生物生长的主要因素。目前研究中好氧反硝化细菌所利用的碳源主要有葡萄糖、蔗糖、柠檬酸钠、琥珀酸钠和乙酸钠等。不同碳源对菌株xn2的生长状况及脱氮有重要的影响,结果如图10所示:菌株xn2对9种碳源的利用情况由高到低依次为:乙酸钠、柠檬酸钠、琥珀酸钠、乙醇、丙酮、乳糖、蔗糖、草酸钠、碳酸氢钠。以柠檬酸钠、乙酸钠和琥珀酸钠为碳源时,xn2的od600均在0.400左右,nh4+-n去除率分别达到100.00%、100.00%、94.00%,tn去除率在70.00%-80.00%之间,其中以乙酸钠为碳源时菌株xn2的od600最高为0.421,这表明以这3种有机酸为碳源时,菌株xn2的生长和脱氮性能较好;但以草酸钠为碳源时,xn2的od600几乎不增长,nh4+-n去除率仅4.16%,说明菌株xn2基本不能利用草酸钠为碳源生长和脱氮;菌株xn2以乳糖为碳源时nh4+-n去除率为11.54%,略高于蔗糖;以乙醇为碳源时,ph从7.10下降到6.00,说明菌株xn2在生长过程中产生了酸,可能影响其生长,但菌株xn2在以乙醇和丙酮为碳源时od600、nh4+-n和tn去除率均高于以糖类为碳源的情况;以不添加碳源的对照组情况下,菌体几乎不生长且nh4+-n基本没有去除,说明菌株xn2需要碳源才能生长和脱氮;以碳酸氢钠为碳源时,菌体几乎不生长且nh4+-n基本没有去除,说明菌株xn2不可以利用无机碳源生长和脱氮。综上,菌株xn2以乙酸钠为碳源时,od600、nh4+-n和tn去除率均最高,因此,选择乙酸钠作为菌株xn2生长和脱氮的最佳碳源。菌株xn2碳源利用情况与已报道的研究结果相似,如raoultellasp.sari01以柠檬酸钠和乙酸钠为碳源时具有较好的脱氮特性,24hnh4+-n去除率均达到94.00%以上;pseudomonasaeruginosayl以乙酸钠、琥珀酸钠和柠檬酸钠为碳源时,24hnh4+-n去除率均达到90.00%以上。不同c/n对菌株xn2异养硝化脱氮性能的影响结果:脱氮细菌的生长代谢需要适量的碳源和氮源,当有机碳源缺乏时,菌株的生长就会受到抑制或产生大量中间代谢产物,而有机碳源的量过多,超过菌体生长所需的量时,会降低异养硝化速率,且容易造成资源浪费。因此c/n是影响菌株生长状况和脱氮特性的重要因素之一。不同c/n对菌株xn2生长及异养硝化脱氮性能的影响结果如图11所示:当c/n=10时,菌株xn2的od600最高达0.421,nh4+-n去除率达到100%,tn去除率达94.42%;当c/n为30和50时,菌株xn2的od600分别为0.322和0.300,nh4+-n去除率分别下降至83.81%和70.57%,tn去除率分别下降至81.4%、67.54%;当c/n高于80以上时,菌株xn2的od600均小于0.075,nh4+-n去除率也均小于15.76%。综上c/n=10时,即可满足菌株xn2在异养硝化过程中对碳源的需求,随着c/n的增加,菌株xn2的od600明显降低,nh4+-n和tn去除率呈下降趋势,这说明c/n提高抑制了菌株xn2的生长和脱氮性能,这可能是由于碳源量过高,部分有机物会直接嵌入酶结构,影响了酶活性,从而抑制了菌株的生长和异养硝化脱氮性能。总体来说,菌株xn2在c/n=10时,其生长最好和异养硝化性能较好,在低c/n条件下,异养硝化脱氮性能较好,优于目前报道的大多数异养硝化-好氧反硝化细菌,例如pseudomonasadn-42c/n为9时,其nh4+-n去除率65.70%,最佳c/n=12时,其nh4+-n去除率为75.4%;acinetobactersp.y1在c/n为10的条件下,nh4+-n去除率为88.90%;bacilluspumilusbp-171c/n为6时,其nh4+-n去除率约28.00%,随着c/n增加,其nh4+-n的脱氮效率呈上升趋势,c/n为12时,其nh4+-n去除率约60.00%,c/n为18时,其nh4+-n去除率最高76.88%。总之,菌株xn2对营养的需求较低,在处理含氮污水方面可节约资源、能源,同时菌株xn2的脱氮效率高,在处理低碳浓度的含氮污水方面具有潜在优势。不同ph对菌株xn2异养硝化脱氮性能的影响结果:ph能够通过影响酶的活性和微生物细胞膜电位的变化,从而影响微生物摄取营养物质的能力,ph过低或过高都会影响菌株的生长和脱氮性能。不同ph对菌株xn2生长及异养硝化脱氮性能的影响结果如图12所示:当ph<7时,菌株xn2几乎不生长,且nh4+-n去除率小于5%,说明此菌不适合在酸性条件下生长和脱氮;当ph在7-11范围内,菌株xn2的od600均高于0.340,nh4+-n去除率均高于70.00%,tn去除率均高于60.00%;当ph=9时,菌株xn2的生长和脱氮效果最好,od600最大达到0.424,nh4+-n去除率达98.12%,tn去除率为94.17%;当ph在10-11范围内,随着ph的增加,菌株xn2的od600、nh4+-n、tn去除率都呈下降趋势,但其od600、nh4+-n、tn仍能达到0.377、89.77%、79.08%以上,菌株xn2仍具有较强的异养硝化脱氮性能,说明该菌在碱性环境下能较好地生长并发挥异养硝化作用,具有嗜碱特性。综上,菌株xn2对ph的适应范围较广,在中性到碱性条件下更适合生长,同时对nh4+-n、tn的降解率较高,说明菌株xn2具有较强的耐碱特性,优于目前分离出的大多数异养硝化-好氧反硝化细菌,例如,pseudoxanthomonassp.c2的最佳ph为9,其nh4+-n去除率最高达到94.89%,ph升高到10时,其生长和脱氮能力均受到抑制,nh4+-n去除率为60.00%;rhizobiumsp.zb612的最佳ph为7,其nh4+-n去除率最高达到85.99%,ph>7时,其生长和脱氮能力均受到抑制,当ph=8.4时,nh4+-n去除率为80.00%;klebsiellasp.y5的最适ph为7,其nh4+-n去除率最高达到65.24%,随着ph升高,其生长和脱氮能力均受到抑制。不同温度对菌株xn2异养硝化脱氮性能的影响结果:温度过高会导致蛋白质或核酸的变性失活,而温度过低会使酶活性受到抑制,微生物的生长代谢减弱,从而影响脱氮效果。不同温度对菌株xn2生长及异养硝化脱氮性能的影响结果如图13所示:当温度为20℃和25℃时,菌株xn2od600分别为0.260和0.300,nh4+-n去除率分别升高至55.00%和58.97%,tn去除率均在55.55%左右,说明菌株xn2可以在低温条件下生长,但脱氮性能较弱,这可能是因为低温条件下,酶活性会受到抑制;当温度为30℃时,菌株xn2od600最高达0.386,nh4+-n去除率达到86.35%,tn去除率达74.43%;当温度在35-40℃之间时,菌株xn2的od600均高于0.360,nh4+-n去除率均在86.00%左右,tn去除率均在74.00%左右。当温度为45℃时,菌株xn2仍表现出了很强的生长和脱氮能力,od600为0.353,nh4+-n去除率下降至60.67%,tn去除率下降至55.38%。综上,菌株xn2生长的温度范围较广,即菌株xn2在高温和低温环境下均生长良好,其较宽的温度适应范围有利于其在实际应用中脱氮潜力的发挥。此外,菌株xn2具有较强的耐高温能力,其生长及脱氮最适合温度范围30-40℃,35-40℃仍具有较强的脱氮能力,优于目前分离出的大多数异养硝化-好氧反硝化细菌,例如,acinetobactercalcoaceticu.n730℃时,其nh4+-n去除率达到最高,高于30℃时,其生长和脱氮能力均受到抑制,37℃时,nh4+-n去除率为76.30%;45℃时,zobellellasp.b307,基本失去脱氮能力;alicaligenefaecalisni3-135℃时,其nh4+-n去除率达到最高,高于35℃时,其生长和脱氮能力均受到抑制,37℃时,nh4+-n去除率为80.00%。不同溶解氧浓度对菌株xn2异养硝化脱氮性能的影响结果:溶解氧浓度是影响菌体异养硝化过程的重要影响因素之一,其浓度高低直接影响异养硝化-好氧反硝化菌的生长状况及脱氮效率。本实验通过调节摇床转速调节培养过程中的溶解氧浓度。不同摇床转速对菌株xn2生长及异养硝化脱氮性能的影响结果如图14所示:当转速为0,即静置培养时,菌株xn2可以生长,od600为0.100,但对nh4+-n和tn的去除率较低,分别为24.79%和20.19%,说明静置培养溶解氧不足,异养硝化活性低;当转速为150rmp时,菌株xn2od600达到最高为0.393,同时nh4+-n去除率达到94.09%,tn去除率达到90.34%。随着转速的升高,能够促进菌株xn2的生长和异养硝化脱氮能力,但当转速提高到一定程度时,反而会抑制菌株的生长和脱氮效率。当转速为200rmp时,菌株xn2od600下降至0.313,nh4+-n去除率下降至86.65%,tn去除率下降至64.11%,可能是因为在较高溶解氧条件下,菌株生长较快而提前进入衰亡期。综上,菌株xn2生长的溶解氧浓度范围较广,可以在静置培养即较低溶解氧的条件下生长,也可以在较高溶解氧的条件下生长,菌株xn2在摇床转速为150rmp时达到最佳脱氮效果,将摇床转速提高到高于150rmp时,抑制了菌株xn2的生长和脱氮性能。不同菌种和菌株之间,适宜的溶解氧浓度差异较大。例如,alcaligenesfaecaliswy-01摇床转速低于120rmp时,nh4+-n去除率随着摇床转速的增加去除效果显著,菌株wy-01的最佳摇床转速约120rmp,高于120rmp时,其nh4+-n去除率基本不变;bacillussp.jb4的最佳溶解氧浓度为摇床转速180rmp,nh4+-n去除率达到最高81.46%,当转速为210rmp时,其生长和脱氮能力均受到抑制,nh4+-n去除率为70.00%;acinetobacteryb摇床转速80rmp时,其nh4+-n去除率较低53.92%,随着摇床转速的增加,nh4+-n去除率逐步增大,菌株yb的最佳溶解氧浓度为摇床转速200rmp。不同接种量对菌株xn2异养硝化脱氮性能的影响结果:接种量不足则不能充分发挥微生物生长繁殖的潜力而导致脱氮效果差,接种量过大会引起生物竞争营养物而致使部分细菌死亡,适量的接种对提高脱氮效果及实际应用有重要意义[52]。不同接种量对菌株xn2生长及异养硝化脱氮性能的影响结果如图15所示:当接种量为3%时,菌株xn2od600为0.275,对nh4+-n、tn去除率分别为91.03%、76.10%;当接种量为5%和10%时,菌株xn2od600增长到0.310左右,nh4+-n、tn去除率分别为94.00%和84.70%左右;当接种量增大到15%时,菌株xn2od600增长到0.328,nh4+-n、tn去除率下降,分别为92.38%、81.11%。据报道接种量过低菌体较少,脱氮的能力较弱,但接种量过高则无法满足微生物生长所需的营养物质,菌体会发生自溶现象而死亡。综上所述,接种量为5%和10%的条件下,对菌株xn2的异养硝化能力影响不大,为了节省成本,本实验后续研究选用5%的接种量较为合适。目前分离出的大多数hn-ad菌接种量最适范围在3-10%,菌株xn2接种量与已报道的研究结果相似,例如,pseudonomassp.hn接种量为5%,其no3--n去除率97.20%,增大接种量对菌株的脱氮性能影响不大,接种量为10%的条件下,其对no3--n去除率达到最高为97.70%;acinetobacterpittii.a14在接种量为5%的条件下,对no3--n的去除率接近80.00%,增大接种量至10%的条件,其no3--n去除率达到最大值86.62%;bacillussp.jb4接种量为3%时,其nh4+-n去除率最高在75.00%左右,但当继续增大接种量,其nh4+-n去除率呈下降趋势,接种量为5%时,nh4+-n去除率下降至50.00%左右。c/n对菌株xn2好氧反硝化脱氮性能影响:以乙酸钠为碳源,以kno3为氮源,初始no3--n浓度为100mg·l-1,调节乙酸钠含量,使c/n为5、10、20、30、50、100。菌株xn2种子液以3%的接种量分别接入装有100ml的好氧反硝化培养基的250ml三角锥瓶中,30℃、150rpm震荡培养24h,取样测od600、ph、no3--n和tn含量,计算其no3--n和tn去除率。c/n是影响菌株生长状况及好氧反硝化脱氮能力的重要影响因素之一。适宜的c/n对微生物的脱氮效果有重大影响。当c/n过低时,不能提供足够的碳源来满足菌体的正常生长代谢,但c/n过高时,则会抑制菌体生长使菌体对碳源的利用率较低且浪费原料,影响环境。不同c/n对菌株xn2好氧反硝化脱氮性能的影响结果如图16所示:当c/n=5时,菌株xn2od600仅为0.197,no3--n、tn去除率分别为58.90%和49.38%,这可能是由于碳源不充足,菌体的生长和反硝化活性都受到一定程度的抑制,no3--n不能完全去除;在c/n10-50范围内,即c/n分别为10、20、30和50,菌株xn2od600分别为0.233、0.235、0.322、0.325,no3--n去除率均达100%,tn去除率也在99%左右,即菌株xn2的生长随着c/n增加呈增长趋势,具有较强的好氧反硝化脱氮能力;但c/n增加到100时,菌株xn2od600减少至0.280,no3--n、tn去除率也分别下降到41.35%和25.12%,这可能是因为c/n过高,对菌株xn2生长和反硝化脱氮作用产生了抑制。综上所述,菌株xn2在相对低的c/n条件下具有较强的好氧反硝化脱氮能力,在c/n10-50范围内生长良好,能将no3--n完全转化去除,综合考虑节约原料和降低成本,菌株xn2后续脱氮实验研究选用c/n=10较为合理。在c/n范围较宽条件下(10-50),菌株xn2均表现出较高的好氧反硝化脱氮性能,优于目前报道的大多数异养硝化-好氧反硝化细菌,例如,pseudomonassphn,在c/n7-12范围内,no3--n去除率随着c/n的升高而下降,c/n为7时,其no3--n的脱氮效率最高99.50%,c/n高于7时,其生长和脱氮能力均受到抑制,c/n为12时,no3--n去除率下降至90.00%;pseudomonasmonteil.h97,在c/n0-25范围内,随着c/n升高,其no3--n去除率先升高后下降,c/n为l5时,其no3--n去除率最高99.13%,tn去除率61.42%。总体来说,菌株xn2的最适c/n范围较广,相比于其他对c/n需求范围较小的菌株,xn2显然更适合处理c/n宽范围的含氮污水。c/n对菌株xn2好氧亚硝化脱氮性能影响:以乙酸钠为碳源,以nano2为氮源,初始no2--n浓度为100mg·l-1,调节乙酸钠含量,使c/n为5、10、20、30、50、100。菌株xn2种子液以3%的接种量分别接入装有100ml的好氧亚硝化培养基的250ml三角锥瓶中,30℃、150rpm震荡培养24h,取样测od600、ph、no2--n和tn含量,计算其no2--n和tn去除率。不同c/n对菌株xn2好氧亚硝化脱氮性能的影响结果如图17所示:当c/n=5时,菌株xn2od600仅为0.174,no2--n、tn去除率分别为35.75%和26.86%;当c/n在10-50范围内时,菌株xn2的生长随着c/n增加呈增长趋势,c/n为10、20、30和50,其od600分别为0.296、0.247、0.257、0.373,no2--n去除率均达100%,具有较强的好氧亚硝化脱氮性能;但随着c/n继续升高,xn2对tn的去除率呈下降趋势,c/n增加到100时,菌株xn2的od600减少至0.196,氮去除能力也减弱,其中no2--n、tn去除率分别下降至12.06%和2.90%,说明此时碳源已达到饱和,远大于菌体需求,过高的碳源对菌株xn2的生长和亚硝化脱氮作用产生了抑制。综上所述,在c/n10-50范围内,菌株xn2能将no2--n完全转化去除,综合考虑节约原料和降低成本,菌株xn2后续脱氮实验研究选用c/n=10较为合理。在c/n范围较宽条件下(10-50),菌株xn2均表现出较强的好氧亚硝化脱氮性能,优于目前报道的异养硝化-好氧反硝化细菌,例如,pantoeadispersacpf-1,在c/n5-20范围内,随着c/n升高,其no3--n去除率先升高后下降,c/n为20时,其no2--n去除率最高99.00%,c/n高于20时,其no2--n的脱氮效率呈下降趋势。菌株xn2耐高氮特性研究菌株xn2耐高nh4+-n特性研究:碳源为乙酸钠、氮源为(nh4)2so4、c/n=10,nh4+-n浓度分别设为100、300、500、800、1000、1500、2000和2200mg·l-1,菌株xn2种子液按5%的接种量接入装有100ml异养硝化培养基的250ml三角锥瓶中,30℃、150rmp培养5d,每天取样测定od600、ph、tn、nh4+-n、no3--n和no2--n含量,并分别计算tn和nh4+-n去除率以及no3--n和no2--n积累量。不同初始nh4+-n浓度对菌株xn2的生长影响结果如图18所示:当初始nh4+-n浓度100-300mg·l-1范围内,菌株xn2od600均随培养时间延长呈增长趋势,培养至3d达到最大值,随后进入平稳期;当初始nh4+-n浓度300mg·l-1时,经过3d培养,菌株xn2的od600达到最高0.717,培养4d后菌株xn2开始衰退,od600下降至0.662,这可能是因为在高浓度的nh4+-n条件下,菌株xn2刚开始要先适应高浓度的环境,适应后菌株开始迅速生长,随后进入衰退期,菌株的生长就会受到抑制或产生大量中间代谢产物;当初始nh4+-n浓度由300mg·l-1增大到500mg·l-1时,菌株xn2培养至第5d,其od600呈增长趋势;当将初始nh4+-n继续增大到800-2200mg·l-1范围内,菌株xn2几乎不生长,这可能是因为过高的nh4+-n浓度对菌株产生抑制作用,抑制了菌株的生长代谢。因此,菌株xn2能够耐受nh4+-n的最高浓度为500mg·l-1。不同初始nh4+-n浓度对菌株xn2异养硝化脱氮性能的影响结果如图19所示:当初始nh4+-n浓度100mg·l-1时,菌株xn2对nh4+-n、tn去除率最高,分别为90.36%、82.23%;随着初始nh4+-n浓度逐渐增加,该菌对nh4+-n、tn去除率逐渐降低,当初始nh4+-n浓度增大到500mg·l-1时,菌株xn2的nh4+-n、tn去除率下降至69.77%、63.49%;当nh4+-n浓度继续增大到800-2200mg·l-1时,菌株xn2几乎无异养硝化脱氮性能,这可能是因为过量的底物与酶分子结合形成中间产物,抑制菌株的酶活性,从而抑制菌株的生长,影响其异养硝化脱氮性能。菌株xn2在整个异养硝化过程中几乎没有出现no3--n和no2--n的积累,这可能是因为以(nh4)2so4为氮源时,在代谢过程中作为中间产物的no3--n和no2--n被迅速转化并未产生积累,还有可能是菌株将nh4+-n直接转化为气态氮,并未转化为no3-n或no2-n等中间产物。综上,随着初始nh4+-n浓度逐渐增加,菌株xn2对nh4+-n、tn去除率呈下降趋势,菌株xn2对nh4+-n耐受最高浓度为500mg·l-1,与目前报道的许多异养硝化-好氧反硝化菌株相比,xn2具有较好的异养硝化能力和耐受较高浓度nh4+-n的能力,例如,pseudomonasputidaly1和acinetobactesp.hy2,当初始nh4+-n浓度50mg·l-1时,对nh4+-n去除率均达到最高,分别为64.00%和74.00%,随着初始nh4+-n浓度的升高,菌株y1、hy2脱氮率均逐渐下降;acinetobacter.spyn3,在初始nh4+-n浓度150mg·l-1时,培养1d,nh4+-n去除率仅18.7%,当将初始nh4+-n浓度提高到200mg·l-1时,菌株yn3几乎不发生脱氮反应,因此菌株yn3对nh4+-n耐受的最高浓度为150mg·l-1。菌株xn2耐高no3--n特性研究:碳源为乙酸钠、氮源为kno3、c/n=10,no3--n浓度分别设为100、300、500、800、1000、1200、1500和2000mg·l-1,菌株xn2种子液按5%的接种量接入装有100ml好氧反硝化培养基的250ml三角锥瓶中,30℃、150rmp培养8d,每天取样测定od600、ph、tn和no3--n含量,并分别计算tn和no3--n去除率。水体中氮磷含量过多会导致水体富营养化,目前地下水和地表水的no3--n污染已经成为严重的环境问题,水中no3--n含量过高会引发高铁血红蛋白症。因此,去除水体中氮素污染研究意义重大。不同初始no3--n浓度对菌株xn2的生长和好氧反硝化性能影响结果如图20所示:当初始no3--n浓度100-500mg·l-1范围内,菌株xn2经过1d培养均达到对数生长期,当初始no3--n浓度为500mg·l-1时,od600由0.045迅速增长到0.562;当初始no3--n浓度500-1200mg·l-1范围内,菌株xn2od600均随培养时间延长呈增长趋势,培养至3d达到最大值,随后进入平稳期;当将初始no3--n浓度继续增大到1500mg·l-1时,经过4d培养,菌株xn2的od600达到0.935;当将初始no3--n浓度提高到2000mg·l-1时,经过8d培养,菌株xn2的od600达到最高1.275,说明菌株xn2能够耐受的no3--n浓度可达2000mg·l-1。不同初始no3--n浓度对菌株xn2的生长及好氧反硝化脱氮性能的影响结果如图21所示:菌株xn2在不同初始no3--n浓度中均能生长,经过较短时间的适应期后进入对数生长期,与此同时培养基中的no3--n浓度培养初期减少较快速。当初始no3--n浓度100-500mg·l-1范围内,菌株xn2经过1d培养,no3--n均完全去除,tn去除率均在95.00%以上,no3--n最大平均去除速率20.83mg·(l·h)-1;当初始no3--n浓度500-1200mg·l-1范围内,菌株xn2经过3d培养,no3--n均完全去除,tn去除率均在94.00%左右,no3--n最大平均去除速率16.67mg·(l·h)-1;随着no3--n浓度的增大,菌株xn2降解no3--n所需的时间不断延长,当初始no3--n浓度为1500mg·l-1时,菌株经过4d培养,no3--n完全去除、tn去除率达到93.20%,no3--n平均去除速率15.63mg·(l·h)-1。当将初始no3--n浓度继续提高到2000mg·l-1时,菌株xn2经过8d培养,no3--n完全去除、tn去除率逐渐下降至89.16%,no3--n平均去除速率10.42mg·(l·h)-1。综上,菌株xn2可以耐受的no3--n高达2000mg·l-1,转化no3--n的速度较快,耐受高浓度no3--n,具有非常强的好氧反硝化能力,优于目前报道的大多数异养硝化-好氧反硝化细菌,例如,bacillushwajinpoensisslwx2在初始no3--n浓度300mg·l-1时,经过4d培养,no3--n去除率达53.00%,当初始no3--n浓度增大到500mg·l-1时,经过4d培养,no3--n去除率下降至24.00%;enterobactercloacaehnr在初始no3--n浓度为300mg·l-1时,培养2d,no3--n去除率仅74.50%,no3--n去除速率4.69mg·(l·h)-1;paracoccusversutusad-4在初始no3--n浓度10-500mg·l-1范围内,培养2d,no3--n去除率均达到99%,但当初始no3--n浓度大于500mg·l-1时,no3--n去除率明显下降,可能是较高的no3--n抑制了反硝化酶;pseudomonascitronellolisad-9在初始no3--n100-1000mg·l-1范围内,均能达到较好脱氮效果,对no3--n的耐受浓度可达到1000mg·l-1。菌株xn2耐高no2--n特性研究:碳源为乙酸钠、氮源为nano2、c/n=10,no2--n浓度分别设为100、150、200、300、500、800和1000mg·l-1,菌株xn2种子液按5%的接种量接入装有100ml好氧亚硝化培养基的250ml三角锥瓶中,30℃、150rmp培养5d,每天取样测定od600、ph、tn和no2--n含量,并分别计算tn和no2--n去除率。no2--n浓度过低不能满足微生物生长代谢所需的氮源,而过高可能会对微生物产生毒害作用也可能具有抑制作用,但目前有关高浓度no2--n对异养硝化-好氧反硝化菌脱氮能力鲜见报道。不同初始no2--n浓度对菌株xn2的生长情况影响如图22所示:当初始no2--n浓度100-200mg·l-1范围内,菌株xn2经过1d培养达到对数生长期,当初始no2--n浓度为200mg·l-1时,od600由0.045迅速增长到0.375;当初始no2--n浓度200-400mg·l-1范围内,菌株xn2od600随着培养时间延长呈增长趋势,培养至2d达到最大值,随后进入平稳期;当将初始no2--n浓度继续增大到500mg·l-1时,经过3d培养,菌株xn2的od600由0.045增长到0.811;当将初始no2--n浓度提高到1000mg·l-1时,经过6d培养,菌株xn2的od600达到最高1.254。以上结果说明菌株xn2在高浓度的no2--n条件下生长良好,且能够耐受较高浓度的no2--n。不同初始no2--n浓度对菌株xn2好氧亚硝化脱氮性能的影响结果如图23所示:当初始no2--n浓度在100-400mg·l-1范围内,菌株xn2经过2d培养,no2--n均完全去除,tn去除率均在90.00%以上;当将初始no2--n浓度提高到500mg·l-1时,菌株xn2经过3d培养,no2--n完全去除,tn去除率达89.15%,no2--n平均去除速率6.95mg·(l·h)-1;当将初始no2--n浓度继续提高到1000mg·l-1时,菌株xn2经过5d培养,no2--n完全去除,tn去除率下降至85.41%,no2--n平均去除速率达8.34mg·(l·h)-1。以上结果说明菌株xn2能够耐受高浓度的no2--n,且具有高效的no2--n去除能力,优于目前报道的大多数异养硝化-好氧反硝化细菌,例如pantoeadispersacpf-1最大no2--n耐受浓度为200mg·l-1;pseudomonasputiday-12和pseudomonastolaasiiy-11对高浓度的no2--n具有一定的耐受能力,但no2--n浓度100mg·l-1时,no2--n和tn脱除率均在40.00%左右;a.xylosoxidansx10no2--n浓度在25-300mg·l-1范围内,48h其no2--n去除率均达99.80%,no2--n浓度在300-700mg·l-1范围内,其no2--n去除率逐渐下降,no2--n浓度700mg·l-1时,其no3--n去除率仅为20.00%左右;pseudomonassp.hg-7no2--n浓度为48.1mg·l-1,24h其no2--n去除率达到100.00%,no2--n浓度提高到91.4mg·l-1时,24h其no2--n去除率下降至40.00%。菌株xn2好氧反硝化过程氮平衡计算:反硝化过程的氮平衡分析,是探讨菌株利用no3-n进行好氧反硝化机理的前提和基础。氮平衡计算的前提是将整个系统看成一个封闭系统,进入这个系统的氮等于排出这个系统氮的总量。菌株利用no3-n过程中,一部分氮转变为氮气或其他氮氧化物逸出水体,还有一部分氮被微生物自身的生长繁殖所利用转化为蛋白质、有机胺等参与菌体合成。为确保最终总氮测定包括此部分菌体内部的氮,本实验使用scientz-iid超声波细胞破碎仪器,在400w下每个样品进行80个周期(一个周期包括5s脉冲、6s暂停)超生破碎菌体,使菌株体内的氮释放出来。本次实验确定c/n=10,以乙酸钠为碳源,kno3为唯一氮源,设置初始no3--n浓度分别为100mg·l-1和800mg·l-1。菌株xn2种子液按5%的接种量接入装有100ml好氧反硝化培养基的250ml三角锥瓶中,30℃、150rpm培养3d,每天取样,测定以下指标:初始氮含量:空白培养基中的tn、no3--n含量;最终总氮:超声波细胞破碎的tn含量;最终溶解性氮:经过培养,菌体超声波细胞破碎后,10000rpm离心5min上清液经过0.22μm醋酸纤维滤膜过滤,滤液中tn、nh4+-n、no3--n、no2--n含量。通过建立的氮平衡方式,根据质量守恒对反应前后的氮总量进行衡算,计算方法如下:(1)胞内氮=最终总氮-最终溶解性总氮;(2)最终溶解性有机氮=最终溶解性总氮-最终溶解性无机氮(氨氮+硝氮+亚硝氮);(3)转化为细胞体内氮的比率=胞内氮/初始总氮值×100%;(4)氮去除率=(初始总氮-最终总氮)/初始总氮×100%。本研究在高浓度或低浓度no3--n条件下,通过不同培养时间测定菌株xn2对氮素同化作用和好氧反硝化转化比率。不同no3--n浓度对菌株xn2好氧反硝化过程的氮平衡分析结果如下:当初始no3--n浓度为100mg·l-1左右时,菌株xn2培养24h,no3--n下降至1.84±0.15mg·l-1,此外培养基中还有少量溶解性no2--n和有机氮,分别为0.02±0.03mg·l-1、16.05±0.79mg·l-1,通过建立的氮平衡方式,对反应前后的氮总量衡算,约有49.44±0.50%的氮被菌株同化所利用,约有32.43±0.88%的氮转化为含氮气体;菌株xn2培养48h,剩余的溶解性no3--n为1.53±0.14mg·l-1,几乎没有溶解性no2--n、nh4+-n的积累,溶解性有机氮下降到13.61±0.65mg·l-1,此时约有25.73±1.27%的氮被菌体同化,约有58.93±0.50%的氮转化成含氮气体;菌株xn2培养72h,剩余的溶解性no3--n为1.39±0.17mg·l-1,几乎没有溶解性no2--n、nh4+-n的积累,溶解性有机氮下降至5.65±0.55mg·l-1,此时约有24.31±1.10%的氮同化为胞内氮,68.55±0.47%的氮转化为含氮气体。综上所述,细胞同化作用和好氧反硝化作用是菌株xn2脱氮的主要形式,初始no3--n浓度较低时,菌株xn2在培养初期主要转化成胞内氮,少部分转化成含氮气体去除,随着培养时间的延长,细胞内的氮逐渐减少,而转化为含氮气体的总氮量逐渐增加。当初始no3--n浓度为800mg·l-1左右时,菌株xn2培养24h,剩余的溶解性no3--n为398.37±5.93mg·l-1,剩余的溶解性no2--n为16.49±0.88mg·l-1,没有溶解性nh4+-n的积累,还有部分溶解性有机氮102.59±0.28mg·l-1,此时约有12.98±0.84%的氮被菌体同化,22.94±0.76%的氮转化为含氮气体;菌株xn2培养48h,溶解性no3--n减少至44.42±0.16mg·l-1,几乎没有溶解性no2--n、nh4+-n的积累,仍有92.92±1.78mg·l-1溶解性有机氮,此时约有13.41±0.47%的氮被菌体同化所利用,大约有64.73±0.88%的氮转化为含氮气体2培养72h,剩余的溶解性no3--n仅为1.04±0.30mg·l-1,几乎没有溶解性no2--n、nh4+-n的积累,溶解性有机氮100.65±1.95mg·l-1,此时约有15.17±0.41%的氮被菌体同化,大约有72.01±0.50%的氮转化为含氮气体。综上所述,初始no3--n浓度较高时,随着培养时间的延长,同化作用转化成细胞内的氮增加量不明显,而转化为含氮气体的总氮量迅速增加,菌株xn2好氧反硝化过程中的no3--n主要通过好氧反硝化作用转化成含氮气体,而不是同化作用转化为胞内氮。综上,菌株xn2在初始no3--n浓度为800mg·l-1时,相比在初始no3--n浓度为100mg·l-1时转化为n2的比例更高,表明在高浓度no3--n条件下,菌株xn2进行好氧反硝化作用的效果更好,可用于处理较高浓度的no3--n污水,在生物脱氮方面具有潜在优势。菌株xn2在初始no3--n浓度为100mg·l-1时,具有较高的含氮气体生产率68.55%,优于目前报道的大部分异养硝化-好氧反硝化菌株,例如,acinetobacter.spyn3,当初始no3--n浓度为88.3mg·l-1时,仅35.40%的no3--n通过反硝化作用转化为气体,47.00%的氮同化为细胞内的生物量;diaphorobactersp.pdb3,初始nh4+-n浓度为109.88mg·l-1时,36h同化到细胞内的氮占nh4+-n去除量的49.41%,以气态氮的形式达到的氮损失占nh4+-n去除量的42.74%。目前在高浓度no3--n条件下,分析异养硝化-好氧反硝化细菌的氮平衡过程还未见报道,菌株xn2在此方面具有较强的反硝化氮转化能力,在初始no3--n浓度为800mg·l-1时,培养72h菌株xn2具有较高的n2生产率约72.01%和较低的胞内氮积累约15.17%,在含高浓度no3--n污水处理中更具优势,可以减少二次污染,降低处理费用。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1