经修饰的甾醇酰基转移酶的制作方法
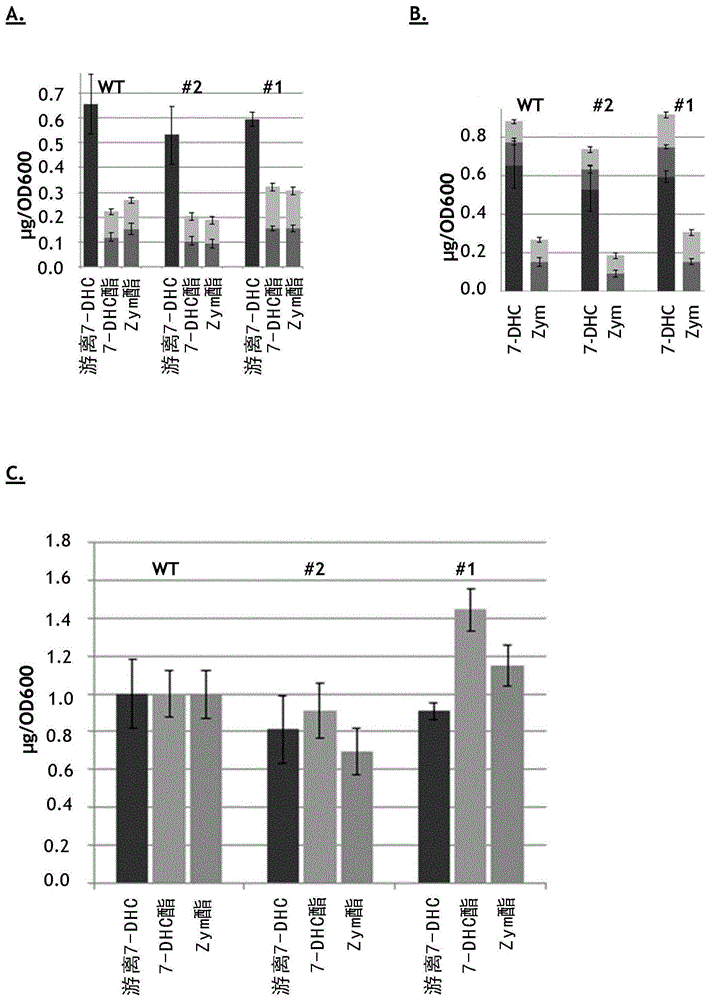
本发明涉及经修饰的甾醇酰基转移酶,所述经修饰的甾醇酰基转移酶具有朝向要在维生素d3的生物技术生产中使用的维生素d3前体7-脱氢胆固醇(7-dhc)的酰化的改善的活性和/或特异性。本发明还涉及一种表达所述经修饰的酶的宿主菌株,以及所述宿主菌株在用于产生维生素d3或其衍生物和/或代谢产物的方法中的用途。维生素d3(也称为胆钙化醇或胆骨化醇)可以在哺乳动物皮肤中由前维生素d3(也称为7-脱氢胆固醇或7-dhc)合成,前维生素d3是在暴露于紫外光时胆固醇生物合成的产物,由此7-dhc被光化学转化为前维生素d3,前维生素d3在体温下异构化为生物活性形式维生素d3。在肝脏中,维生素d3转化为无生物活性的25-羟基维生素d3(也称为骨化二醇(calcidiol/calcifediol)、25-羟基胆钙化醇、25-oh-d3或hyd),这是维生素d3的主要循环形式。进一步的羟基化发生在肾脏中。对于维生素d3的工业生产,(原则上)化学和生物技术合成两者都是可行的。化学合成开始于从例如羊毛脂中分离出胆固醇,然后将所述胆固醇脱氢成7-dhc,7-dhc是化学和生物技术合成两者中的重要中间体。通过暴露于紫外光和进一步的纯化/萃取步骤,将7-dhc转化为维生素d3。经修饰的酵母菌株可用于7-dhc的生物合成,其中乙酰辅酶a在多步酶促过程中转换为7-dhc。所述酶促转换发生在酵母的内质网中。细胞膜中不需要的过量的甾醇,包括7-dhc及其前体,过量的甾醇对酵母有毒并且因此作为甾醇酯存储在细胞内的细胞器(所谓的脂质体)中,它们可以从所述细胞器中进一步分离出。游离甾醇与脂质体中存储的甾醇(主要以甾醇酯形式)之间的平衡是通过几种蛋白质(酶)的作用,包括甾醇酰基转移酶的作用触发的。在酵母,特别是酿酒酵母(saccharomycescerevisiae)中,甾醇的酯形成主要由两种甾醇酰基转移酶are1p和are2p进行。由于所述甾醇酰基转移酶的非特异性作用,存储在脂质体内的甾醇酯库是相对多样的,包括但不限于例如麦角甾醇、酵母甾醇、羊毛甾醇、烯胆甾烷醇、胆甾-5,7,24(25)-三烯醇、或7-dhc的酯。由于只有7-dhc的前体胆甾-5,7,24(25)-三烯醇,而不是酵母甾醇可用于维生素d3合成,因此需要将特定的酯(例如7-dhc的酯)选择性存储到脂质体中和/或需要增加由此类酵母菌株产生的7-dhc的中间体的周转,所述7-dhc的中间体被进一步转换为维生素d3和/或其衍生物或代谢物。同样也是本发明的焦点的特定代谢物是25-羟基维生素d3。因此,持续性的任务是产生能够产生甾醇的宿主细胞(例如酵母),所述宿主细胞具有高7-dhc生产率/对7-dhc的特异性和/或减少的副产物/中间体(包括酵母甾醇、羊毛甾醇或烯胆甾烷醇),特别是存储在脂质体中的此类中间体的酯的积累。令人惊讶地,我们现在发现,可经由在are2和/或are1的序列中引入某些氨基酸取代来使宿主细胞中甾醇酰基转移酶活性的特异性和/或活性偏移,这将导致宿主细胞朝向作为维生素d3生产中的重要中间体的7-dhc的更高生产率。因此,本发明涉及具有甾醇酰基转移酶活性的经修饰的酶,即经修饰的甾醇酰基转移酶,特别是甾醇酰基转移酶同种型are1p和/或are2p的活性,所述经修饰的酶包含在与根据seqidno:1的多肽中的选自由11、281、366、442、551、554、572、624、626、627、636位以及它们的组合组成的组的残基对应的一个或多个位置处的一个或多个氨基酸取代,所述经修饰的酶具有相对于包括酵母甾醇在内的副产物/中间体增加的对7-dhc的特异性和/或增加的朝向酯形成(包括7-dhc的酯)的活性。。已经从酿酒酵母中分离出了显示are2活性的根据seqidno:1的多肽,包括编码所述多肽的多核苷酸。已经从酿酒酵母中分离出了显示are1活性的根据seqidno:3的多肽,包括编码所述多肽的多核苷酸。术语“甾醇酰基转移酶”、“酰基转移酶”、“are”、“具有酰基转移酶活性的酶”或仅“酶”在本文中可互换使用,并且是指酶[ec2.3.1.26],即将脂肪酰基基团从一个分子转移至另一个分子的酰基转移酶。此类转移或酶促活性可以通过技术人员已知的手段来测量。已从不同来源,包括哺乳动物、酵母或植物分离出了甾醇酰基转移酶。are1和are2都能够将甾醇(例如酵母甾醇和/或7-dhc)酰化成相应的酯。在本文中,“经修饰的”酶,即经修饰的酰基转移酶,与例如酵母甾醇的酯化和/或包含例如7-dhc或酵母甾醇的总甾醇酯的改进形成相比,具有优选的朝向7-dhc的酯化的活性和/或特异性。优选的酰基转移酶同种型(isoform)是are2p或are1p。在本文中,“未修饰的”甾醇酰基转移酶,特别是are1和are2,是指不携带如本文所定义的一个或多个氨基酸取代的相应内源酶。在本文中,携带如本文所定义的经修饰的甾醇酰基转移酶活性,特别是包含如本文所定义的一个或多个氨基酸取代的are2和/或are1的宿主细胞被称为“经修饰的”宿主细胞。携带未修饰的甾醇酰基转移酶活性,即编码野生型are2和/或are1基因的相应宿主细胞被称为“未修饰的”宿主细胞。在本文中,指定维生素d3中间体的术语“酵母甾醇”、“羊毛甾醇”、“烯胆甾烷醇”、“胆甾-5,8,24(25)-三烯醇”、“胆甾-5,7,24(25)-三烯醇”或“7-dhc”包括所述化合物的游离形式和酯形式两者。在本文中,甾醇混合物包含7-dhc和“副产物”或中间体,包括但不限于酵母甾醇、羊毛甾醇、烯胆甾烷醇、胆甾-5,8,24(25)-三烯醇或胆甾-5,7,24(25)-三烯醇。在本文中,“产生胆固醇的酵母”不再能够产生麦角甾醇,而是产生包括但不限于以下的胆固醇产物:胆甾-5,7,24(25)-三烯醇、胆甾-5,8,24(25)-三烯醇、胆甾-7,24(25)-二烯醇、7-dhc或酵母甾醇。具体地,这可以通过引入erg5erg6双敲除来实现。特别地,所述修饰对应于甾醇酰基转移酶2和/或1的经修饰的活性,即are2和/或are1活性,所述修饰包括一个或多个氨基酸取代,其中优选地在与根据seqidno:1的多肽中的选自由11、281、366、442、551、554、572、624、626、627、636位以及它们的组合组成的组的残基对应的一个或多个位置处的至少一个氨基酸取代对应于e11和/或l281和/或d366和/或i442和/或h551和/或h554和/或f572和/或f624和/或l626和/或g627和/或c636的一个或多个氨基酸取代。更优选地,所述修饰对应于经修饰的are2活性,甚至更优选地根据seqidno:1的经修饰的多肽,其中已经引入了选自由11、281、366、442、551、554、572、624、626、627、636位以及它们的组合组成的组的残基对应的一个或多个位置处的至少一个氨基酸取代。优选地,具有经修饰的are2和/或are1活性的酶源自酵母属(saccharomyces),例如酿酒酵母。在一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的are2和/或are1活性,包含在与根据seqidno:1的多肽中的残基11对应的位置处的氨基酸取代,优选地由甘氨酸取代谷氨酸(e11g)。可以将与seqidno:1中的残基e11g对应的位置处的所述氨基酸取代与如本文所定义的其他取代(即,如本文所述的与根据seqidno:1的多肽中的氨基酸残基281、366、442、551、554、572、624、626、627和/或636对应的一个或多个位置上的取代)进行组合。优选地,可以将与seqidno:1中的残基e11g对应的位置处的氨基酸取代与其他取代(例如在与seqidno:1中的f624l、g627d、d366v、c636s和/或i442v对应的一个或多个位置处的一个或多个氨基酸取代)组合,更优选地是与seqidno:1中的残基e11g-f624l、e11g-g627d、e11g-d366v-c636s、e11g-d366v-g627d-c636s、e11g-d366v-f624l-c636s、e11g-d366v-i442v-f624l-c636s或e11g-d366v-i442v-g627d-c636s对应的取代的组合,其中最优选的组合选自e11g-d366v-c636s、e11g-d366v-g627d-c636s、e11g-d366v-f624l-c636s、e11g-d366v-i442v-g627d-c636s或e11g-d366v-i442v-f624l-c636s。通过使用携带此类经修饰的酶,特别是包含所述一个或多个氨基酸取代中的一个氨基酸取代的根据seqidno:1的酶的产生胆固醇的酵母,与使用如本文所定义的未修饰的酵母相比,酶特异性可提高在至少约3倍至5倍的范围内。在一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的are2和/或are1活性,包含在与根据seqidno:1的多肽中的残基281对应的位置处的氨基酸取代,优选地由异亮氨酸取代亮氨酸(l281i)。可以将与seqidno:1中的残基l281i对应的位置处的所述氨基酸取代与如本文所定义的其他取代(即,如本文所述的与根据seqidno:1的多肽中的氨基酸残基11、366、442、551、554、572、624、626、627和/或636对应的一个或多个位置上的取代)进行组合。通过使用携带此类经修饰的酶,特别是包含所述一个或多个氨基酸取代中的一个氨基酸取代的根据seqidno:1的酶的产生胆固醇的酵母,与使用如本文所定义的未修饰的酵母相比,相对酯形成可提高在至少约1.5倍的范围内。在一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的are2和/或are1活性,包含在与根据seqidno:1的多肽中的残基366对应的位置处的氨基酸取代,优选地由缬氨酸取代天冬氨酸(d366v)。可以将与seqidno:1中的残基d366v对应的位置处的所述氨基酸取代与如本文所定义的其他取代(即,如本文所述的与根据seqidno:1的多肽中的氨基酸残基11、281、366、442、551、554、572、624、626、627和/或636对应的一个或多个位置上的取代)进行组合。通过使用携带此类经修饰的酶,特别是包含所述一个或多个氨基酸取代中的一个氨基酸取代的根据seqidno:1的酶的产生胆固醇的酵母,与使用如本文所定义的未修饰的酵母相比,酶特异性可提高在至少约3倍的范围内。在另一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的are2和/或are1活性,包含在与根据seqidno:1的多肽中的残基442对应的位置处的氨基酸取代,优选地由缬氨酸取代异亮氨酸(i442v)。可以将与seqidno:1中的残基i442v对应的位置处的所述氨基酸取代与如本文所定义的其他取代(即,如本文所述的与根据seqidno:1的多肽中的氨基酸残基11、281、366、551、554、572、624、626、627和/或636对应的一个或多个位置上的取代)进行组合。优选地,可以将与seqidno:1中的残基i442v对应的位置处的氨基酸取代与其他取代(例如在与seqidno:1中的f624l、l626f和/或g627d对应的一个或多个位置处的氨基酸取代)组合,更优选地是与seqidno:1中的残基i442v-l626f、i442v-g627d、i442v-f624l-l626f或i442v-l626f-g627d对应的取代的组合,其中最优选的组合选自i442v-g627d、i442v-f624l-l626f或i442v-l626f-g627d。通过使用携带此类经修饰的酶,特别是包含所述一个或多个氨基酸取代中的一个氨基酸取代的根据seqidno:1的酶的产生胆固醇的酵母,与使用如本文所定义的未修饰的酵母相比,酶特异性可提高在至少约2.2倍至4.5倍的范围内。在又一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的are2和/或are1活性,包含在与根据seqidno:1的多肽中的残基551对应的位置处的氨基酸取代,优选地由酪氨酸取代组氨酸(h551y)。可以将与seqidno:1中的残基h551y对应的位置处的所述氨基酸取代与如本文所定义的其他取代(即,如本文所述的与根据seqidno:1的多肽中的氨基酸残基11、281、366、442、554、572、624、626、627和/或636对应的一个或多个位置上的取代)进行组合。通过使用携带此类经修饰的酶,特别是包含所述一个或多个氨基酸取代中的一个氨基酸取代的根据seqidno:1的酶的产生胆固醇的酵母,与使用如本文所定义的未修饰的酵母相比,酶特异性可提高在至少约1.7倍的范围内。在一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的are2和/或are1活性,包含在与根据seqidno:1的多肽中的残基554对应的位置处的氨基酸取代,优选地由谷氨酰胺取代组氨酸(h554q)。可以将与seqidno:1中的残基h554q对应的位置处的所述氨基酸取代与如本文所定义的其他取代(即,如本文所述的与根据seqidno:1的多肽中的氨基酸残基11、281、366、442、551、572、624、626、627和/或636对应的一个或多个位置上的取代)进行组合。优选地,可以将与seqidno:1中的残基h554q对应的位置处的氨基酸取代与其他取代(例如在与seqidno:1中的f624l、f572l和/或g627d对应的一个或多个位置处的氨基酸取代)组合,更优选地是与seqidno:1中的残基h554q-f572l-f624l或h554q-f572l-g627d对应的取代的组合。通过使用携带此类经修饰的酶,特别是包含所述一个或多个氨基酸取代中的一个氨基酸取代的根据seqidno:1的酶的产生胆固醇的酵母,与使用如本文所定义的未修饰的酵母相比,酶特异性可提高在至少约1.4倍至超过12倍的范围内。在另一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的are2和/或are1活性,包含在与根据seqidno:1的多肽中的残基572对应的位置处的氨基酸取代,优选地由亮氨酸取代苯丙氨酸(f572l)。可以将与seqidno:1中的残基f572l对应的位置处的所述氨基酸取代与如本文所定义的其他取代(即,如本文所述的与根据seqidno:1的多肽中的氨基酸残基11、281、366、442、551、554、624、626、627和/或636对应的一个或多个位置上的取代)进行组合。通过使用携带此类经修饰的酶,特别是包含如上面所定义的所述一个或多个氨基酸取代中的一个氨基酸取代的根据seqidno:1的酶的产生胆固醇的酵母,与使用如所定义的未修饰的酵母相比,酶特异性可提高在至少约1.7倍的范围内。在一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的are2和/或are1活性,包含在与根据seqidno:1的多肽中的残基624对应的位置处的氨基酸取代,优选地由亮氨酸取代苯丙氨酸(f624l),所述取代对应于在根据seqidno:3的多肽中的f592l取代。可以将与seqidno:1中的残基f624l对应的位置处的所述氨基酸取代与如本文所定义的其他取代(即,如本文所述的与根据seqidno:1的多肽中的氨基酸残基11、281、366、442、551、554、572、626、627和/或636对应的一个或多个位置上的取代)进行组合。通过使用携带此类经修饰的酶,特别是包含所述一个或多个氨基酸取代中的一个氨基酸取代的根据seqidno:1的酶的产生胆固醇的酵母,与使用如本文所定义的未修饰的酵母相比,酶特异性可提高在至少约3.1倍的范围内。在一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的are2和/或are1活性,包含在与根据seqidno:1的多肽中的残基626对应的位置处的氨基酸取代,优选地由苯丙氨酸取代亮氨酸(l626f)。可以将与seqidno:1中的残基l626f对应的位置处的所述氨基酸取代与如本文所定义的其他取代(即,如本文所述的与根据seqidno:1的多肽中的氨基酸残基11、281、366、442、551、554、572、624、627和/或636对应的一个或多个位置上的取代)进行组合。通过使用携带此类经修饰的酶,特别是包含如上面所定义的所述一个或多个氨基酸取代中的一个氨基酸取代的根据seqidno:1的酶的产生胆固醇的酵母,与使用如本文所定义的未修饰的酵母相比,酶特异性可提高在至少约2.2倍的范围内。在一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的are2和/或are1活性,包含在与根据seqidno:1的多肽中的残基627对应的位置处的氨基酸取代,优选地由天冬氨酸取代甘氨酸(g627d),所述取代对应于在根据seqidno:3的多肽中的g595d取代。可以将与seqidno:1中的残基g627d对应的位置处的所述氨基酸取代与如本文所定义的其他取代(即,如本文所述的与根据seqidno:1的多肽中的氨基酸残基11、281、366、442、551、554、572、624、626和/或636对应的一个或多个位置上的取代)进行组合。通过使用携带此类经修饰的酶,特别是包含如上面所定义的所述一个或多个氨基酸取代中的一个氨基酸取代的根据seqidno:1的酶的产生胆固醇的酵母,与使用如本文所定义的未修饰的酵母相比,酶特异性可提高在至少约2.2倍的范围内。在一个实施方式中,如本文所定义的经修饰的酶,特别是经修饰的are2和/或are1活性,包含在与根据seqidno:1的多肽中的残基636对应的位置处的氨基酸取代,优选地由丝氨酸取代胱氨酸(c636s)。可以将与seqidno:1中的残基c636s对应的位置处的所述氨基酸取代与如本文所定义的其他取代(即,如本文所述的与根据seqidno:1的多肽中的氨基酸残基11、281、366、442、551、554、572、624、626和/或627对应的一个或多个位置上的取代)进行组合。通过使用携带此类经修饰的酶,特别是包含所述一个或多个氨基酸取代中的一个氨基酸取代的根据seqidno:1的酶的产生胆固醇的酵母,与使用如本文所定义的未修饰的酵母相比,酶特异性可提高在至少约3倍的范围内。根据本发明的经修饰的甾醇酰基转移酶,例如are2和/或are1,优选地包含在与根据seqidno:1的多肽中的f624l对应的位置上的至少一个氨基酸取代,从而导致酶特异性为约4。这可以通过以下方式来增大:引入一个或多个其他氨基酸取代(例如,在与f572l和/或h554q对应的位置上的氨基酸取代),从而导致与使用如本文所定义的未修饰的酵母相比,酶特异性增加在超过12倍的范围内。根据本发明的另一个实施方式的经修饰的甾醇酰基转移酶,例如are2和/或are1,优选地包含在与根据seqidno:1的多肽中的g627d对应的位置上的至少一个氨基酸取代,从而导致酶特异性大于4。这可以通过以下方式来增大:引入一个或多个其他氨基酸取代(例如,在与f572l和/或h554q对应的位置上的氨基酸取代),从而导致与使用如本文所定义的未修饰的酵母相比,酶特异性增加为约5倍。在本文中,are2和/或are1的活性被修饰。这可以通过以下方式来实现:例如将一个或多个突变引入编码are2和/或are1的一个或多个内源基因中,即在如本文所述的一个或多个位置上的一个或多个氨基酸取代。技术人员知道如何遗传操纵酵母细胞,从而导致are2和/或are1活性的改变。这些遗传操纵包括但不限于例如使用质粒、病毒或其他载体进行基因置换、基因扩增、基因破坏、转染、转化。可以以不同的方式来执行核酸或氨基酸中的突变产生,即诱变,诸如通过随机化或定点诱变,由诸如辐射等试剂引起物理损伤、化学处理,或插入遗传元件。技术人员知道如何引入突变。本发明特别地涉及如本文所定义的此类经修饰的are2和/或are1酶在用于产生维生素d3的中间体7-dhc的方法中的用途。优选地,本发明的经修饰的酶被引入合适的宿主细胞和/或在合适的宿主细胞中表达,所述合适的宿主细胞为例如酵母,优选地产生甾醇的酵母,特别是产生胆固醇的酵母细胞,例如选自酿酒酵母、裂殖酵母属(schizosaccharomycesspp.)、毕赤酵母属(pichiaspp.)、克鲁维酵母属(klyuveromycesspp.)、汉逊酵母属(hansenulaspp.)或解脂耶氏酵母(yarrowialipolytica),优选地酿酒酵母。将经修饰的宿主用于生产7-dhc,所述7-dhc可以进一步转化为维生素d3和/或25-羟基维生素d3。可以进一步修饰合适的宿主细胞,以进一步增加7-dhc的产量和/或减少副产物的积累,7-dhc是朝向维生素d3生物合成的重要中间体。因此,在一个实施方式中,本发明涉及一种酵母菌株,所述酵母菌株具有改变的are2和/或are1活性,并且此外其中erg5和erg6被灭活。可以通过以下方式来进一步修饰酵母细胞:表达具有c24还原酶活性的异源酶,所述异源酶特别地选自ec1.3.1.72,例如作用于胆甾-7,24-二烯醇、酵母甾醇或三烯醇(例如胆甾-5,7,25-三烯醇)的异源c24还原酶,优选地植物或脊椎动物甾醇δ24还原酶,更优选地来自脊椎动物来源,甚至更优选地来自人、猪、狗、小鼠、大鼠、马、斑马鱼(daniorerio)或任何已知来源,只要所述酶可以在所述酵母细胞内表达即可。最优选地,甾醇δ24-还原酶选自斑马鱼、大鼠或人。表达所述甾醇δ24还原酶的序列是可公开可得的,包括但不限于uniprotkb/swiss-prot参考q15392、q60hc5、q8vch6、q5bqe6、q39085或p93472(参见例如wo2003064650)。在另一个实施方式中,可以通过以下方式来进一步修饰根据本发明的宿主细胞:引入参与7-dhc的生物合成的内源酶的同源物,例如c5-甾醇去饱和酶(erg3)和/或c8-甾醇异构酶(erg2),导致对7-dhc的特异性和/或7-dhc生产率提高并且副产物或维生素d3中间体(包括但不限于酵母甾醇、羊毛甾醇和/或烯胆甾烷醇)的积累减少。在一个特定的实施方式中,本发明涉及一种用于将宿主细胞改善为朝向7-dhc生产的方法,其中所述经修饰的宿主细胞如本文所定义,即经由在如本文所定义的甾醇酰基转移酶are2和/或are1中引入一个或多个氨基酸取代而被修饰,特别是产生胆固醇的酵母细胞,优选地其中erg5和erg6被灭活并且其中任选地表达具有如本文所定义的c-24还原酶活性的异源酶,和/或其中任选地表达内源性erg2和/或erg3的同源物的酵母细胞,其中所述宿主细胞被改进为使得与如本文所定义的未修饰的宿主细胞相比,由所述宿主细胞产生的甾醇总量中7-dhc的百分比增加和/或宿主细胞朝向产生甾醇的活性增加。在一个实施方式中,本发明涉及经修饰的甾醇酰基转移酶,特别是经修饰的are2和/或are1,所述经修饰的甾醇酰基转移酶包含至少一个或多个如本文所定义的氨基酸取代,其中与未修饰的酶的特异性相比,所述酶的特异性增加,从而导致由携带此类经修饰的甾醇酰基转移酶的合适宿主细胞产生的甾醇混合物中的7-dhc与包括例如酵母甾醇在内的副产物的比率较高。该比率可增加至少约1.4倍,例如经由在相应的are2和/或are1序列中引入与h554q对应的氨基酸取代;例如增加至少3倍,例如经由在相应的are2和/或are1序列中引入与e11g-f624l、i442v-g627d或i442v-f624l-l626f对应的氨基酸取代;例如增加至少4倍,例如经由在相应的are2和/或are1序列中引入与f624l或g627d对应的氨基酸取代;例如增加至少5倍、10倍或甚至12倍或更多倍,例如经由在相应的are2和/或are1序列中引入与h554q-f572l-g627d或e11g-d366v-i442v-g627d-c636s对应的氨基酸取代。在一个实施方式中,本发明涉及经修饰的甾醇酰基转移酶,特别是经修饰的are2和/或are1,所述经修饰的甾醇酰基转移酶包含至少一个或多个如本文所定义的氨基酸取代,其中与未修饰的酶的活性相比,所述酶的活性增加,从而导致更高的甾醇和/或甾醇酯的总产量,包括更高量的由携带此类经修饰的甾醇酰基转移酶的合适宿主细胞产生的7-dhc。因此,相对的酯形成,即所有酯与游离7-dhc的比率可增加至少1.2倍,例如经由在相应的are2和/或are1序列中引入与h55q或h554q-f572l对应的氨基酸取代;例如增加至少2倍或3倍,例如经由在相应的are2和/或are1序列中引入与i442v-g627d或i442v-l626f-g627d对应的氨基酸取代。如本文所述,经由引入一个或多个氨基酸取代,可以使总7-dhc产量增加至少约1.2倍至1.4倍。通过使用如本文所述的经修饰的宿主细胞,例如酵母,例如产生甾醇的酵母,特别是产生胆固醇的酵母,甾醇混合物中7-dhc的百分比可基于甾醇的总量,增加至少40%,例如50%、60%、70%、80%、90%的7-dhc。在本发明的一个方面中,将包含如本文所定义的经修饰的are2和/或are1活性的宿主细胞用于生产维生素d3前体7-dhc的方法中。经修饰的宿主细胞可以如技术人员关于相应产生胆固醇的宿主细胞所已知的那样在需氧或厌氧条件下在补充有适当营养物的水性培养基中培养。任选地,如本领域已知的,所述培养是在参与电子转移的蛋白质和/或辅因子存在下进行的。宿主细胞的培养/生长可以以分批、补料分批、半连续或连续模式进行。如本领域技术人员所已知的,取决于宿主细胞,优选地,维生素d3及其前体(例如7-dhc)的生产可以变化。维生素d3生产中7-dhc和其他中间体的培养和分离描述于例如wo2011067144或wo2017108799中。7-dhc可以从甾醇混合物中分离和/或任选地进一步纯化,并且可以使用本领域已知的方法进一步转换成维生素d3和/或25-羟基维生素d3。术语“are1”和“are1p”、“are2”和“are2p”、“erg5”和“erg5p”、“erg6”和“erg6p”在本文中可互换使用,并且是指由相应基因are1、are2、erg5和erg6编码的多肽。出于本发明的目的,产生胆固醇的酵母细胞被修饰为使得其确实显示出are2和/或are1的经修饰的活性,例如在内源性are2、are1或两者中携带修饰,从而导致are2和/或are1的经修饰的特异性,其中所述修饰包括引入如本文所定义的一个或多个氨基酸取代。编码erg5、erg6、are1、are2、erg2、erg3或甾醇δ24还原酶(erg4)的基因,如本文所用的酵母细胞的培养和遗传工程改造是已知的并且描述于例如us7608421中。在本文中,术语“c-24-还原酶”或“δ24-还原酶”在本文中可互换地使用。在酵母中,该酶由erg4编码,并且对24位上的碳原子的甲基有活性。因此,在所述位置上不显示此类甲基的三烯醇不是酵母erg4的可接受底物。术语“c-8甾醇异构酶”、“δ8,7-异构酶”或“具有c-8甾醇异构酶活性的酶”在本文中可互换地使用,并且是指能够催化胆甾-8-烯醇转换为胆甾-7-烯醇和/或酵母甾醇转换为胆甾-7,24-二烯醇的酶。在酵母中,该酶由erg2编码。在酵母中,该酶由erg2编码。要在根据本发明的经修饰的宿主细胞中使用的优选的erg2同源物是与seqidno:5具有至少41%,例如至少44%、45%、48%、49%、53%、56%、60%、70%、80%、90%、92%、95%、98%或至多100%同一性的多肽,所述多肽显示出c-8甾醇异构酶活性并且包含编码此类多肽的多核苷酸,可从玉米黑粉菌获得。具体地,在如本文所定义的经修饰的宿主细胞中表达所述erg2同源物的1个或更多个拷贝,例如至少1个、2个、3个、5个拷贝。术语“c-5甾醇去饱和酶”、“具有c-5甾醇去饱和酶活性的酶”、“去饱和酶”或“erg3-同源物”在本文中可互换使用,并且是指能够催化胆甾-8-烯醇转换为胆甾-7,24-二烯醇和/或胆甾-7-烯醇转换为胆甾-5,7,24-三烯醇和/或7-dhc的酶。在酵母中,该酶由erg3编码。要在根据本发明的经修饰的宿主细胞中使用的优选的erg3同源物是与seqidno:7具有至少45%,例如至少50%、52%、60%、70%、80%、90%、92%、95%、98%或至多100%的同一性的多肽,所述多肽显示出c-5甾醇去饱和酶活性并且包含编码此类多肽的多核苷酸,可从巴斯德毕赤酵母(pichiapastoris)或粟酒裂殖酵母(schizosaccharomycespombe)获得。具体地,在如本文所定义的经修饰的宿主细胞中表达所述erg3同源物的1个或更多个拷贝,例如至少1个、2个、3个、5个拷贝。如本文所定义的“相对的酯形成”是比率(全部酯/游离7-dhc)突变体/(全部酯/游离7-dhc)野生型。如本文所定义的酶“特异性”是比率(7-dhc酯/酵母甾醇酯)突变体/(7-dhc酯/酵母甾醇酯)野生型。如本文所定义的“总7-dhc产量”是比率(总7-dhc)突变体/(7-dhc)野生型。在本文中,关于酶的术语“比活性”或“活性”是指其催化活性,即其催化从给定底物形成产物的能力。比活性定义了在给定时间段内和在限定温度下每限定量的蛋白质消耗的底物和/或产生的产物的量。通常,比活性表示为每mg蛋白质每分钟消耗的底物或形成的产物的μmol数。通常,μmol/min缩写为u(=unit)。因此,在本文件全文中可互换地使用μmol/min/(mg蛋白质)或u/(mg蛋白质)的比活性单位定义。如果酶在体内(即在如本文所限定的宿主细胞内)或在合适的底物存在下的合适的(无细胞)系统内执行其催化活性,则所述酶是有活性的。技术人员知道如何测量酶活性,例如通过hplc。关于本发明,应当理解的是,生物(诸如微生物、真菌、藻类或植物)也包括具有相同生理性质的此类物种的同义词或基名(basonyms),如由国际原核生物命名法(internationalcodeofnomenclatureofprokaryotes)或关于藻类、真菌和植物的国际命名法(internationalcodeofnomenclature)(melbourne法)所限定的。特别地,本发明的特征在于以下实施方式:(1)一种如本文所定义的具有甾醇酰基转移酶活性的经修饰的酶,所述经修饰的酶包含在与根据seqidno:1的多肽中的选自由11、281、366、442、551、554、572、624、626、627、636位以及它们的组合组成的组的残基对应的一个或多个位置处的一个或多个氨基酸取代,优选地与e11g和/或l281i和/或d366v和/或i442v和/或h551y和/或h554q和/或f572l和/或f624l和/或l626f和/或g627d和/或c636s和/或它们的组合对应的一个或多个氨基酸取代。(2)如本文所定义并且根据实施方式(1)所述的经修饰的酶,所述经修饰的酶催化包括7-脱氢胆固醇和酵母甾醇在内的甾醇的酯化,其中与使用相应的未修饰酶的催化中的7-dhc与酵母甾醇的比率相比,所述甾醇酯中7-dhc与酵母甾醇的比率增加了至少约1.4倍。(3)如本文所定义并且根据实施方式(1)或(2)所述的经修饰的酶,其中所述一个或多个氨基酸取代选自f624l、g627d、e11g、h554q、i442v,以及它们的组合。(4)一种宿主细胞,优选地酵母,更优选地产生甾醇的酵母,甚至更优选地产生胆固醇的酵母,所述宿主细胞包含如本文所定义并且根据实施方式(1)、(2)、(3)中任一项所述的经修饰的酶,所述宿主细胞任选地还包含在与根据seqidno:3的多肽中的选自592和/或595位的残基对应的一个或多个位置处的一个或多个氨基酸取代,优选地与f592l和/或g595d对应的取代。(5)如本文所定义并且根据实施方式(4)所述的宿主细胞,所述宿主细胞用于产生包含7-dhc和酵母甾醇的甾醇混合物,其中与表达未修饰的酶的宿主细胞相比,7-dhc与酵母甾醇的比率增加了至少约1.4倍。(6)如本文所定义并且根据实施方式(4)或(5)所述的宿主细胞,所述宿主细胞任选地还包含:-erg5和erg6的灭活,和/或-选自具有甾醇δ24-还原酶活性的ec1.3.1.72的异源酶的表达,优选地其中所述异源酶源自植物或脊椎动物,更优选地源自人、猪、狗、小鼠、大鼠、马或斑马鱼。(7)一种用于降低包含酵母甾醇和7-dhc的甾醇混合物中的酵母甾醇的百分比的方法,所述方法包括在合适的条件下培养如本文所定义并且根据实施方式(4)、(5)或(6)中的一项所述的宿主细胞,以及任选地从所述甾醇混合物中分离和/或纯化7-dhc,或者一种用于增加包含7-dhc和酵母甾醇的甾醇混合物中7-dhc的百分比的方法,所述方法包括在合适的条件下培养如本文所定义并且根据实施方式(4)、(5)或(6)中的一项所述的宿主细胞,以及任选地从所述甾醇混合物中分离和/或纯化7-dhc。(8)一种用于产生7-dhc的方法,所述方法包括使用如本文所定义并且根据实施方式(4)、(5)或(6)中的一项所述的宿主细胞将乙酰辅酶a酶促转换成包含酵母甾醇和7-dhc的甾醇混合物,其中所述甾醇混合物中7-dhc的百分比为至少40%,并且任选地还将7-dhc转换成维生素d3和/或25-羟基维生素d3。(9)如本文所定义并且根据实施方式(1)、(2)、(3)中任一项所述的经修饰的酶或如本文所定义并且根据实施方式(4)、(5)或(6)中的一项所述的宿主细胞在用于产生7-dhc的方法中的用途,其中所述7-dhc是从包含酵母甾醇和7-dhc的甾醇混合物中分离出的,并且其中与分别使用相应的未修饰的酶和宿主细胞的方法相比,7-dhc与酵母甾醇的比率增加了至少约1.4倍。附图说明图1.are2野生型菌株(“wt”)和are2变体#2和#1的脂质提取物的hplc分析(参见表1)。关于7-dhc的酯(“7-dhc酯”)和酵母甾醇的酯(“zym酯”),如由相应柱中的黑色和浅灰色所示检测到了两种酯形式,游离7-dhc以黑色表示。(a)7-dhc与酯形成的比率,(b)总7-dhc(包括游离形式和酯形式)与总酵母甾醇酯的比率,(c)游离7-dhc、7-dhc酯和酵母甾醇酯的形成由不同的柱示出。将菌株在没有挡板的烧瓶中用两次葡萄糖进料培养两天。数据是3个独立的转化体的平均值,每个转化体培养一次。图2.are2野生型菌株(“wt”)和are2变体#2和#1的脂质提取物的hplc分析(参见表1)。有关进一步详细信息,请参见图1的图例。将菌株在无挡板的烧瓶中用1次葡萄糖进料培养四天。数据是2个独立的转化体的平均值,每个转化体培养一次。图3.are2野生型菌株(“wt”)和are2变体#9和#1的脂质提取物的hplc分析(参见表1)。有关进一步详细信息,请参见图1的图例。根据标准工序进行hplc分析。有关进一步详细信息,请参见文本。图4.are2野生型菌株(“wt”)和are2变体#24和#20的脂质提取物的hplc分析(参见表1)。有关进一步详细信息,请参见图1的图例。根据标准工序进行hplc分析。有关进一步详细信息,请参见文本。图5.are2野生型菌株(“wt”)和are2变体#22和#27的脂质提取物的hplc分析(参见表1)。有关进一步详细信息,请参见图1的图例。根据标准工序进行hplc分析。有关进一步详细信息,请参见文本。图6.are2野生型菌株(“wt”)和are2变体#20、#22、#27和#22的脂质提取物的hplc分析(参见表1)。有关进一步详细信息,请参见图1的图例。根据标准工序进行hplc分析。有关进一步详细信息,请参见文本。图7.are2野生型菌株(“wt”)和are2变体#34、#32、#43、#24、#20、#22和#27的脂质提取物的hplc分析(参见表1)。有关进一步详细信息,请参见图1的图例。根据标准工序进行hplc分析。有关进一步详细信息,请参见文本。图8.are2野生型菌株(“wt”)和are2变体#9、#35、#36、#39和#40的脂质提取物的hplc分析(参见表1)。有关进一步详细信息,请参见图1的图例。根据标准工序进行hplc分析。有关进一步详细信息,请参见文本。图9.are2野生型菌株(“wt”)和are2变体#27、#33、#34、#37和#38的脂质提取物的hplc分析(参见表1)。有关进一步详细信息,请参见图1的图例。根据标准工序进行hplc分析。有关进一步详细信息,请参见文本。图10.are2野生型菌株(“wt”)和are2变体#41、#42、#43、#24和#20的脂质提取物的hplc分析(参见表1)。有关进一步详细信息,请参见图1的图例。根据标准工序进行hplc分析。有关进一步详细信息,请参见文本。以下实施例仅是说明性的,并不意图以任何方式限制本发明的范围。实施例实施例1:are2突变体的产生和筛选通过薄层色谱法(tlc)筛选具有10,000个表达酿酒酵母酰基转移酶2(scare2)变体的酵母克隆的易错文库以获得甾醇酯级分中提高的7-dhc含量(相对于are1和are2的野生型序列(参见序列表))。筛选方法包括从具有少量经消化的细胞壁的细胞中同时提取和分离甾醇。将经处理的生物质直接施加在tlc板上,并浸入溶剂中,这以一个步骤进行了含甾醇的级分的提取和分离。在甾醇酯级分中,通过利用具有共轭双键的化合物的不同分光光度特性(例如,猝灭荧光的能力、紫外检测),将具有共轭双键的甾醇(如例如7-dhc)的比率设置为相对于没有共轭双键的甾醇。将最佳变体一式五份地重新筛选,测序,在摇瓶中培养,并通过hplc-uv以生物学一式三份进行分析,以确定甾醇和甾醇酯的组成。分离出含有最佳变体的质粒,并将其重新转化为产生胆甾-5,7,24-三烯醇的酿酒酵母菌株10a(are1are2erg5erg6::24r;关于构建参见wo2017108799中的实施例1)。通过以下方式分离出具有多个氨基酸交换的突变变体:通过定点诱变(不考虑沉默突变)将相应的突变引入are2,从而找出引起所需效应的突变。将菌株进行培养并通过hplc分析。实施例2:hplc-uv分析标准工序预培养—用我们感兴趣的菌株(每个are2变体3个转化体)接种10ml含有遗传霉素(100μg/ml)的ypd,并在30℃下生长至合适的密度(24小时至48小时)。为了更好地进行比较,还接种了具有野生型are2质粒的三种不同的转化体,它们是与变体同时转化的。在250ml的无挡板摇瓶中将50ml含遗传霉素的ypd的主要培养物接种至od600为0.1,并在30℃下使用200rpm和80%的湿度,使用3次葡萄糖进料培养3天(在约30小时、45小时和60小时后将葡萄糖添加至2%的终浓度)。收获200od单位的生物质(用1600xg离心5分钟并去除上清液)在15mlgreiner管中,并存储在-20℃下直至分析。为了进行提取,将200od细胞沉淀物解冻,重悬于1ml溶细胞酶溶液(5mg/ml溶细胞酶20t在50mmkpi中的溶液,ph7,含有1md-山梨糖醇)中,并在37℃下孵育15分钟(在热混合器上以750rpm孵育)。在离心(2500xg,5分钟)后去除溶细胞酶溶液,并向沉淀物中添加3.73ml的无水etoh(通过小心地上下吹吸而用1ml重悬,然后添加额外的2.73ml)。添加267μl内标(胆固醇乙酸酯,在etoh中1mg/ml的溶液),将细胞悬浮液涡旋,并在混合(在热混合器上以750rpm)下加热至70℃持续1小时。在移开试管几分钟以冷却至室温后,将细胞碎片沉淀(2500xg,在室温下10分钟),并将3ml上清液转移到pyrex试管中,使所述pyrex试管在n2下干燥。将脂质吸收在200μl的乙酸乙酯中(涡旋并在40℃的热混合器上以750rpm混合15分钟)。将溶液再次离心(2500xg,5分钟),然后转移至带有嵌体的玻璃小瓶中以供进行后续hplc-uv分析。通过hplc在两个波长(210nm和280nm)下用紫外检测来分析脂质提取物。在210nm处检测到了酵母甾醇化合物,在280nm处对7-dhc化合物进行定量。溶剂:80%etoh、20%meoh、0.1%tfa柱:ymc-packproc18rs方法:进样量:10μl注射器恒温器:40℃流量:0.6ml/min柱温箱:20℃紫外检测:210nm、280nm(具有共轭双键的甾醇)还分析3种不同浓度(每种物质0.5mg/ml、1.0mg/ml和2.0mg/ml)的7-dhc、酵母甾醇、胆甾醇乙酸酯和角鲨烯的标准混合物,并生成每种物质的标准曲线,以计算以μg/μl或μg/od600为单位的提取物中的甾醇的浓度。实施例3:对are2变体的活性和/或特异性的评估为进行直接比较,将野生型are2质粒与表达are2变体的质粒(参见实施例1)一起重新转化到菌株10a中,并在同一轮次中分析所得的菌株(参见实施例2)。hplc分析的结果汇总在表1和图1至图10中。表中的值给出了与野生型相比,表达突变体/变体的菌株之间的倍数变化。第一个值指示关于酯级分与游离7-dhc级分之间的比率的改善,而第二个值显示关于酯级分中的7-dhc和酵母甾醇的比率的改善。第三个值是突变体和野生型的生物质中总7-dhc含量的比较。列出的变体中的一些变体尤其显示出酯水平的改善,而其他变体显示出酯级分中7-dhc/酵母甾醇比率的改善。表1a:对基于几个独立实验的are2变体的相对酯形成的汇总。“相对酯形成”是指基于使用所示氨基酸交换而不是野生型are2产生的甾醇的总量,7-dhc百分比的x倍增加;“7-dhc酯/zym酯”是指来自7-dhc的酯与酵母甾醇(“zym”)的酯的比率;“总7-dhc/总zym”是总7-dhc(游离和酯)与总酵母甾醇(游离和酯)的比率。有关更多说明,请参见文本。表1b:对基于几个独立实验的are2变体的特异性的汇总。数值表示在使用指定的氨基酸交换而不是野生型are2生成的甾醇mx中,与酵母甾醇百分比相比,7-dhc百分比增加了x倍。有关更多说明,请参见文本。编号#一个或多个氨基酸交换特异性1h554q1.42v286v-h551y-f572l-s633s1.79h554q-f572l1.820f624l3.922g627d4.432e11g-d366v-f624l-c636s3.133e11g-d366v-f624l-c636s3.834e11g-d366v-g627d-c636s4.435e11g-f624l3.236e11g-g627d4.237e11g-d366v-i442v-f624l-c636s3.438e11g-d366v-i442v-g627d-c636s4.539h554q-f572l-f624l12.240h554q-f572l-g627d5.041i442v-f624l-l626f3.142i442v-l626f-g627d2.243i442v-g627d3.5表1c:对基于几个独立实验的are2变体的总7-dhc产量的汇总。数值表示使用指定的氨基酸交换而不是野生型are2生成的所产生7-dhc的总量增加了x倍。有关更多说明,请参见文本。编号#一个或多个氨基酸交换总7-dhc产量1h554q1.020f624l1.022g627d1.024i442v-l626f1.327e11g-d366v-c636s1.232e11g-d366v-f624l-c636s1.433e11g-d366v-f624l-c636s1.434e11g-d366v-g627d-c636s1.335e11g-f624l1.136e11g-g627d1.137e11g-d366v-i442v-f624l-c636s1.438e11g-d366v-1442v-g627d-c636s1.139h554q-f572l-f624l1.040h554q-f572l-g627d1.341i442v-f624l-l626f1.442i442v-l626f-g627d1.443i442v-g627d1.4序列表<110>帝斯曼知识产权资产管理有限公司<120>经修饰的甾醇酰基转移酶<130>32958-wo-pct<160>8<170>patentinversion3.5<210>1<211>642<212>prt<213>酿酒酵母(saccharomycescerevisiae)<400>1metasplyslyslysaspleuleugluasngluglnpheleuargile151015glnlysleuasnalaalaaspalaglylysargglnserilethrval202530aspaspgluglygluleutyrglyleuaspthrserglyasnserpro354045alaasngluhisthralathrthrilethrglnasnhisservalval505560alaserasnglyaspvalalapheileproglythralathrglugly65707580asnthrgluilevalthrglugluvalilegluthraspaspasnmet859095phelysthrhisvallysthrleuserserlysglulysalaargtyr100105110argglnglyserserasnpheilesertyrpheaspaspmetserphe115120125gluhisargproserileleuaspglyservalasngluprophelys130135140thrlysphevalglyprothrleuglulysgluileargargargglu145150155160lysgluleumetalametarglysasnleuhishisarglysserser165170175proaspalavalaspservalglylysasnaspglyalaalaprothr180185190thrvalprothralaalathrsergluthrvalvalthrvalgluthr195200205thrileileserserasnpheserglyleutyrvalalaphetrpmet210215220alailealapheglyalavallysalaleuileasptyrtyrtyrgln225230235240hisasnglyserphelysaspsergluileleulysphemetthrthr245250255asnleuphethrvalalaservalaspleuleumettyrleuserthr260265270tyrphevalvalglyileglntyrleucyslystrpglyvalleulys275280285trpglythrthrglytrpilephethrseriletyrglupheleuphe290295300valilephetyrmettyrleuthrgluasnileleulysleuhistrp305310315320leuserlysilepheleupheleuhisserleuvalleuleumetlys325330335methisserphealaphetyrasnglytyrleutrpglyilelysglu340345350gluleuglnpheserlysseralaleualalystyrlysaspserile355360365asnaspprolysvalileglyalaleuglulyssercysgluphecys370375380serphegluleuserserglnserleuseraspglnthrglnlysphe385390395400proasnasnileseralalysserphephetrpphethrmetphepro405410415thrleuiletyrglnileglutyrproargthrlysgluileargtrp420425430sertyrvalleuglulysilecysalailepheglythrilepheleu435440445metmetileaspalaglnileleumettyrprovalalametargala450455460leualavalargasnserglutrpthrglyileleuaspargleuleu465470475480lystrpvalglyleuleuvalaspilevalproglypheilevalmet485490495tyrileleuaspphetyrleuiletrpaspalaileleuasncysval500505510alagluleuthrargpheglyaspargtyrphetyrglyasptrptrp515520525asncysvalsertrpalaasppheserargiletrpasnileproval530535540hislyspheleuleuarghisvaltyrhissersermetserserphe545550555560lysleuasnlysserglnalathrleumetthrphepheleuserser565570575valvalhisgluleualamettyrvalilephelyslysleuargphe580585590tyrleuphephepheglnmetleuglnmetproleuvalalaleuthr595600605asnthrlysphemetargasnargthrileileglyasnvalilephe610615620trpleuglyilecysmetglyproservalmetcysthrleutyrleu625630635640thrphe<210>2<211>1929<212>dna<213>酿酒酵母(saccharomycescerevisiae)<400>2atggacaagaagaaggatctactggagaacgaacaatttctccgcatccaaaagctcaac60gctgccgatgcgggcaaaagacaatctataacagtggacgacgagggcgaactatatggg120ttagacacctccggcaactcaccagccaatgaacacacagctaccacaattacacagaat180cacagcgtggtggcctcaaacggagacgtcgcattcatcccaggaactgctaccgaaggc240aatacagagattgtaactgaagaagtgattgagaccgatgataacatgttcaagacccat300gtgaagactttaagctccaaagagaaggcacggtataggcaagggtcctctaactttata360tcgtatttcgatgatatgtcatttgaacacaggcccagtatattagatgggtcagttaac420gagcccttcaagaccaaattcgtgggacctactttagaaaaggagatcagaagaagggag480aaagagctaatggccatgcgcaaaaatttacaccaccgcaagtcctccccagatgctgtc540gactcagtagggaaaaatgatggcgccgccccaactactgttccaactgccgccacctca600gaaacggtggtcaccgttgaaaccaccataatttcatccaatttctccgggttgtacgtg660gcgttttggatggctattgcatttggtgctgtcaaggctttaatagactattattaccag720cataatggtagcttcaaggattcggagatcttgaaatttatgactacgaatttgttcact780gtggcatccgtagatcttttgatgtatttgagcacttattttgtcgttggaatacaatac840ttatgcaagtggggggtcttgaaatggggcactaccggctggatcttcacctcaatttac900gagtttttgtttgttatcttctacatgtatttaacagaaaacatcctaaaactacactgg960ctgtccaagatcttcctttttttgcattctttagttttattgatgaaaatgcattctttc1020gccttctacaatggctatctatggggtataaaggaagaactacaattttccaaaagcgct1080cttgccaaatacaaggattctataaatgatccaaaagttattggtgctcttgagaaaagc1140tgtgagttttgtagttttgaattgagctctcagtctttaagcgaccaaactcaaaaattc1200cccaacaatatcagtgcaaaaagctttttttggttcaccatgtttccaaccctaatttac1260caaattgaatatccaagaactaaggaaatcagatggagctacgtattagaaaagatctgc1320gccatcttcggtaccattttcttaatgatgatagatgctcaaatcttgatgtatcctgta1380gcaatgagagcattggctgtgcgcaattctgaatggactggtatattggatagattattg1440aaatgggttggattgctcgttgatatcgtcccagggtttatcgtgatgtacatcttggac1500ttctatttgatttgggatgccattttgaactgtgtggctgaattgacaagatttggcgac1560agatatttctacggtgactggtggaattgtgttagttgggcagacttcagtagaatttgg1620aacatcccagtgcataagtttttgttaagacatgtttaccatagttcaatgagttcattc1680aaattgaacaagagtcaagcaactttgatgacctttttcttaagttccgtcgttcatgaa1740ttagcaatgtacgttatcttcaagaaattgaggttttacttgttcttcttccaaatgctg1800caaatgccattagtagctttaacaaatactaaattcatgaggaacagaaccataatcgga1860aatgttattttctggctcggtatctgcatgggaccaagtgtcatgtgtacgttgtacttg1920acattctaa1929<210>3<211>610<212>prt<213>酿酒酵母(saccharomycescerevisiae)<400>3metthrgluthrlysaspleuleuglnaspgluglupheleulysile151015argargleuasnseralaglualaasnlysarghisservalthrtyr202530aspasnvalileleuproglnglusermetgluvalserproargser354045serthrthrserleuvalgluprovalgluserthrgluglyvalglu505560serthrglualagluargvalalaglylysglngluglnglugluglu65707580tyrprovalaspalahismetglnlystyrleuserhisleulysser859095lysserargserargphehisarglysaspalaserlystyrvalser100105110phepheglyaspvalserpheaspproargprothrleuleuaspser115120125alaileasnvalpropheglnthrthrphelysglyprovalleuglu130135140lysglnleulysasnleuglnleuthrlysthrlysthrlysalathr145150155160vallysthrthrvallysthrthrglulysthrasplysalaaspala165170175proproglyglulysleugluserasnpheserglyiletyrvalphe180185190alatrpmetpheleuglytrpilealaileargcyscysthrasptyr195200205tyralasertyrglyseralatrpasnlysleugluilevalglntyr210215220metthrthraspleuphethrilealametleuaspleualametphe225230235240leucysthrphephevalvalphevalhistrpleuvallyslysarg245250255ileileasntrplystrpthrglyphevalalavalserilepheglu260265270leualapheileprovalthrpheproiletyrvaltyrtyrpheasp275280285pheasntrpvalthrargilepheleupheleuhisservalvalphe290295300valmetlysserhisserphealaphetyrasnglytyrleutrpasp305310315320ilelysglngluleuglutyrserserlysglnleuglnlystyrlys325330335gluserleuserprogluthrarggluileleuglnlyssercysasp340345350phecysleuphegluleuasntyrglnthrlysaspasnaspphepro355360365asnasnilesercysserasnphephemetphecysleupheproval370375380leuvaltyrglnileasntyrproargthrserargileargtrparg385390395400tyrvalleuglulysvalcysalaileileglythrilepheleumet405410415metvalthralaglnphephemethisprovalalametargcysile420425430glnphehisasnthrprothrpheglyglytrpileproalathrgln435440445glutrpphehisleuleupheaspmetileproglyphethrvalleu450455460tyrmetleuthrphetyrmetiletrpaspalaleuleuasncysval465470475480alagluleuthrargphealaaspargtyrphetyrglyasptrptrp485490495asncysvalserphegluglupheserargiletrpasnvalproval500505510hislyspheleuleuarghisvaltyrhissersermetglyalaleu515520525hisleuserlysserglnalathrleuphethrphepheleuserala530535540valphehisglumetalametphealailepheargargvalarggly545550555560tyrleuphemetpheglnleuserglnphevaltrpthralaleuser565570575asnthrlyspheleuargalaargproglnleuserasnvalvalphe580585590serpheglyvalcysserglyproserileilemetthrleutyrleu595600605thrleu610<210>4<211>1833<212>dna<213>酿酒酵母(saccharomycescerevisiae)<400>4atgacggagactaaggatttgttgcaagacgaagagtttcttaagatccgcagactcaat60tccgcagaagccaacaaacggcattcggtcacgtacgataacgtgatcctgccacaggag120tccatggaggtttcgccacggtcgtctaccacgtcgctggtggagccagtggagtcgact180gaaggagtggagtcgactgaggcggaacgtgtggcagggaagcaggagcaggaggaggag240taccctgtggacgcccacatgcaaaagtacctttcacacctgaagagcaagtctcggtcg300aggttccaccgaaaggatgctagcaagtatgtgtcgttttttggggacgtgagttttgat360cctcgccccacgctcctggacagcgccatcaacgtgcccttccagacgactttcaaaggt420ccggtgctggagaaacagctcaaaaatttacagttgacaaagaccaagaccaaggccacg480gtgaagactacggtgaagactacggagaaaacggacaaggcagatgcccccccaggagaa540aaactggagtcgaacttttcagggatctacgtgttcgcatggatgttcttgggctggata600gccatcaggtgctgcacagattactatgcgtcgtacggcagtgcatggaataagctggaa660atcgtgcagtacatgacaacggacttgttcacgatcgcaatgttggacttggcaatgttc720ctgtgcactttcttcgtggttttcgtgcactggctggtgaaaaagcggatcatcaactgg780aagtggactgggttcgttgcagtgagcatcttcgagttggctttcatccccgtgacgttc840cccatttacgtctactactttgatttcaactgggtcacgagaatcttcctgttcctgcac900tccgtggtgtttgttatgaagagccactcgtttgccttttacaacgggtatctttgggac960ataaagcaggaactcgagtactcttccaaacagttgcaaaaatacaaggaatctttgtcc1020ccagagacccgcgagattctgcaaaaaagttgcgacttttgccttttcgaattgaactac1080cagaccaaggataacgacttccccaacaacatcagttgcagcaatttcttcatgttctgt1140ttgttccccgtcctcgtgtaccagatcaactacccaagaacgtcgcgcatcagatggagg1200tatgtgttggagaaggtgtgcgccatcattggcaccatcttcctcatgatggtcacggca1260cagttcttcatgcacccggtggccatgcgctgtatccagttccacaacacgcccaccttc1320ggcggctggatccccgccacgcaagagtggttccacctgctcttcgacatgattccgggc1380ttcactgttctgtacatgctcacgttttacatgatatgggacgctttattgaattgcgtg1440gcggagttgaccaggtttgcggacagatatttctacggcgactggtggaattgcgtttcg1500tttgaagagtttagcagaatctggaacgtccccgttcacaaatttttactaagacacgtg1560taccacagctccatgggcgcattgcatttgagcaagagccaagctacattatttactttt1620ttcttgagtgccgtgttccacgaaatggccatgttcgccattttcagaagggttagagga1680tatctgttcatgttccaactgtcgcagtttgtgtggactgctttgagcaacaccaagttt1740ctacgggcaagaccgcagttgtccaacgttgtcttttcgtttggtgtctgttcagggccc1800agtatcattatgacgttgtacctgaccttatga1833<210>5<211>241<212>prt<213>玉米黑粉菌(ustilagomaydis)<400>5metalaserhisargproargserasnlysalaalaasnglyalaser151015thrserprolysargsertrpileilevalseralaalaleuvalgly202530phecysalaleuilealaalaleuaspserileargserserphetyr354045ilepheasphislysalailetyrlysilealaserthralavalala505560asnhisproglyasnalathralailepheaspaspvalleuaspasn65707580leuargalaaspprolysleualaprotyrileasnlysasnhisphe859095seraspgluserglutrpmetpheasnasnalaglyglyalametgly100105110sermetpheileilehisalaservalthrglutyrleuilephephe115120125glythrprovalglythrgluglyhisthrglyarghisthralaasp130135140asptyrpheasnileleuthrglyasnglntyralapheproalagly145150155160alaleulysalagluhistyrproalaglyservalhishisleuarg165170175argglythrvallysglntyrmetmetprogluaspglycystrpala180185190leugluleualaglnglytrpileproprometleupropheglyleu195200205alaaspvalleuserserthrleuaspleuprothrpheglyilethr210215220valtrpilethralaargglumetvalglyasnleuleuileglylys225230235240phe<210>6<211>726<212>dna<213>玉米黑粉菌(ustilagomaydis)<400>6atggcatcgcatagaccacgcagcaacaaggctgccaatggtgcttcgacttcacccaaa60cgcagctggataattgtctcagctgcgctcgttggcttctgcgctctcatcgccgctctc120gattcgatccgatccagcttctacatctttgaccacaaggcaatctacaagatcgcatcg180actgcggtcgccaaccatccaggcaatgcgacggccatctttgatgatgtcctcgacaac240cttcgtgccgaccccaagctcgcgccttacatcaacaagaatcatttcagcgacgagtca300gaatggatgttcaacaatgccggtggtgctatgggtagcatgttcatcattcatgcttcc360gtcaccgagtacctgatcttctttggcactcccgtcggaaccgagggtcacactggtcgt420cacacagccgatgactacttcaacatccttaccggtaaccaatacgctttcccagctggt480gcgctcaaggcggagcactaccctgccggatcagtgcaccatcttcgccgcggaacggtc540aagcagtacatgatgcctgaagacggctgctgggcgctcgagcttgctcagggctggatc600ccacccatgcttccctttggtctcgccgatgtgctcagctcgacgctcgacctgcccacc660tttggtatcactgtctggatcactgcacgagaaatggttggcaatctgctcatcggcaag720ttttga726<210>7<211>384<212>prt<213>巴斯德毕赤酵母(pichiapastoris)<400>7metaspilealaleugluileleuaspthrphevalpheasplysval151015tyralalysleuleuproileserleuvalglnhisleuproaspgly202530tyrleulysthrleuglyhisleuthrglyalaasnasnthrmetglu354045serleupheglyilealaproasnvalaspglnalaserlysasnhis505560trpleuargthrvalasnaspserilealaleualaargproglyglu65707580argleuvaltyrglyvalasnalaproleuhisphepheaspgluthr859095alatyrthrtyralaserileleuglyargserasnileilearggln100105110phethrthrleumetileleumetileleupheglytrpglyleutyr115120125leuservalalaserphesertyrtyrphevalpheasplysalaile130135140pheasnhisproargtyrleulysasnglnmetserleugluilehis145150155160glnalaleuthralaileprothrmetvalleuleuthrvalprotrp165170175pheleuilegluleuargglytyrserlysleutyrpheaspvalasn180185190gluserthrglyglytrplysalaileiletrpglnileprocysphe195200205ilemetphethraspcyscysiletyrpheilehisargtrpleuhis210215220trpproservaltyrlysargleuhislysprohishislystrpile225230235240valcysthrprophealaserhisalaphehisprovalaspglytyr245250255alaglnserleuprotyrhisleutyrglymetleupheproleuhis260265270lysvalsertyrleuileleupheglyleuvalasnphetrpthrval275280285metilehisaspglyglutyrleuserargaspproilevalasngly290295300alaalacyshisthrvalhishisleutyrpheasntyrasntyrgly305310315320glnphethrthrleutrpaspargleuglyglysertyrargmetpro325330335asplysgluleupheasplysasnlyslyslysaspvallysthrtrp340345350argserglnvallysglnalaaspserilearggluaspleuglugly355360365lysgluasppheargglutyrglythrgluglulysleulysserthr370375380<210>8<211>1155<212>dna<213>巴斯德毕赤酵母(pichiapastoris)<400>8atggacattgctttggagattctagacacttttgtctttgacaaagtctatgcaaaacta60ctgcccatttctctggtgcaacatttgccagatggctatttgaagactttgggacatttg120actggtgccaacaacaccatggaatcactgttcggaatagctccaaacgttgaccaagcg180tctaagaaccactggctgagaacagtgaatgactctattgccttagcccgtcccggtgag240cgtctggtctacggtgtcaacgctcctttacacttttttgacgaaacagcgtatacatac300gcatcgatcttgggacgctccaatatcattcgacaattcacaactttgatgattctgatg360attctttttggctggggtttgtatttatctgtggcttcattttcatactactttgttttt420gataaagccattttcaatcacccaagatacctcaaaaaccagatgtctctggagatccat480caagcgttgactgctatacctacgatggttttgcttacagttccatggtttttgattgag540ttgcgtggatactctaaattatactttgatgtaaatgagtctactggaggatggaaggct600attatttggcaaattccttgcttcattatgtttaccgattgttgtatctactttattcat660cgttggttgcactggccatccgtgtataagcgtttgcacaagcctcaccacaagtggatt720gtttgtacaccttttgctagtcatgccttccatccagttgatggttatgcacaatcacta780ccttaccatttgtatggaatgttgtttccactacacaaggtgagctatctgatcttattt840gggcttgtgaacttttggactgttatgatccatgatggagaatacctgtccagagaccct900atagtcaatggagctgcttgtcatacagtgcatcacctatacttcaactacaattacggc960cagttcacaacactttgggaccgtcttggtggatcatacagaatgccagacaaggaactc1020tttgataagaacaagaagaaagatgtaaagacatggcgttcacaagtcaagcaggccgat1080tcgataagagaagacttagagggaaaagaagatttccgtgagtatggaactgaggaaaaa1140cttaaaagcacatag1155当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1