一种内生蜡样芽孢杆菌、应用及其分离方法与流程
[0001]
本发明涉及的是在农业领域中,一种可以控制小白菜根肿病侵染并有效改善植 物生长的微生物接种剂。具体是指本发明从植物根际中分离出了新的蜡状芽孢杆菌 mz-12菌株,该菌株可以抑制芸苔属根肿菌的感染和提高植物生物量。
背景技术:
[0002]
根肿病是鞭毛菌亚门芸薹根肿菌(plasmodiophora brassicae woron)侵染引起的 一种土传真菌病害,严重危害十字花科植物根部,是世界范围内的毁灭性土传植物 病害。由于根肿病具有传染性强、传播速度快、传播途径多、防治困难等特点,使 得该病在我国迅速蔓延,已造成十字花科作物生产在几乎所有省份都受到了极大的 影响。小青菜(brassica campestris sp.chinensis l.)十字花科芸薹属,又名小白菜, 因根肿病的蔓延,我国小青菜的生产受到了严重制约。
[0003]
芸薹根肿菌是严格的专性寄生菌,该病原体的生命周期复杂,大体可以分为两 个阶段:第一阶段为根毛感染阶段,病菌休眠孢子释放出初级游动孢子,游动孢子 穿过细胞壁抵达根毛的表面;第二阶段为孢子囊内生成数个次生游动孢子,穿透根 部的皮层组织并在皮质组织内细胞发育数周,导致根异常肿大。根瘤的形成阻碍了 水分和养分向上运输,地上部变得枯萎和发育迟缓,造成产量和品质的下降。
[0004]
目前,十字花科根肿病的防治主要通过拔除田间野生寄主、使用诱饵作物、轮 作及使用石灰、曝晒土壤、使用杀菌剂等处理方式减少病原菌的传播及降低土壤中 病原菌含量来控制根肿病的发生,但都不能完全消除根肿病菌。抗性品种可能是一 个很好的选择,但由于物种特异性,抗药性可能会由于病原小种的改变而受到影响。 各种化学药剂对根肿病的防治效果不一,并且农药的残留量大、毒性大等原因而并 未在上海地区广泛使用。
[0005]
由于十字花科蔬菜根肿病病原菌具有很强的抗逆性,在土壤中存活时间长。因 此,采用土壤中的生防菌对其进行防治成为根肿病防治新的思路。有益的内生或根 际微生物的应用是管理根部合理、经济和生态的方法。已有大量文献说明了植物相 关微生物的潜在应用,这些微生物可以促进植物生长并管理土壤生态系统。例如枯 草芽孢杆菌(bacillus subtilis qst713)对根肿病有较好的防效,可以达到60%以上。 灰红链霉菌(streptomyces griseorube)可以有效控制大白菜根肿病,在温室条件下 防效可以达到72.8%,在大田条件下防效可以达到68.5%。cheah等从室内和田间试 验结果发现,1个链霉菌菌株(599)和3个木霉菌株(tc32,tc45,tc63)可有 效控制十字花科根肿病的发生。由于土壤微生物可以在根部或根际定殖,因此生物 防治根肿病是非常有效的策略。
[0006]
从芥菜植物中分离出的蜡状芽孢杆菌菌株能够减少棉花幼苗中由茄状枯萎病 引起的根腐病的发生。此外,蜡状芽孢杆菌菌株bt8和bp24已被用于马铃薯、番 茄和山核桃等几种作物的病害生物防治剂。在另一项研究中,从番茄植物中分离出 的蜡状芽孢杆菌通过快速的根际定殖成功地控制了南方根结线虫的发生,从而显著 减少了病害的危害。
技术实现要素:
[0007]
基于现有技术的不足,本发明提出了一种内生蜡样芽孢杆菌、应用及其分离方 法。
[0008]
为了实现上述目的,本发明提出了一种内生蜡样芽孢杆菌,所述内生蜡样芽孢杆 菌保藏于中国微生物菌种保藏管理委员会普通生物中心,保藏编号为cgmcc no.21141。
[0009]
第二方面,本发明还提出了一种内生蜡样芽孢杆菌的分离方法,其包括如下步骤:
[0010]
步骤s1:从土壤中分离细菌菌株;
[0011]
步骤s2:将步骤s1中分离出的细菌菌株进行筛选,得到所述需要的内生蜡样芽孢 杆菌。
[0012]
优选地,在所述步骤s1中,所述土壤为健康小白菜根际土壤;
[0013]
在所述步骤s2中,通过评估芸苔假单胞菌休眠孢子萌发的抑制率和活力测定的技 术方案来选择菌株。
[0014]
此外,本发明还提出了一种含有上述的内生蜡样芽孢杆菌和/或上述的内生蜡样芽 孢杆菌的代谢产物的微生物菌剂。
[0015]
另外,本发明还提出了一种病原菌抑制剂,所述病原菌抑制剂包括上述的内生蜡样 芽孢杆菌和/或上述的内生蜡样芽孢杆菌的代谢产物。
[0016]
优选地,所述病原菌抑制剂对下述全部或部分病原菌具有抑制作用:
[0017]
芸苔假单胞菌或芸薹根肿病菌(plasmodiophora brassicae woronin)。
[0018]
此外,本发明还提出了一种病害抑制剂,所述病害抑制剂包括如上述的内生蜡样芽 孢杆菌和/或上述的内生蜡样芽孢菌的代谢产物。
[0019]
优选地,所述病害包括根肿病。
[0020]
再者,本发明提出一种促进小白菜生长的方法,所述方法包括向小白菜表面和/或 周围土壤中喷施上述内生蜡样芽孢菌和/或上述内生蜡样芽孢菌的代谢产物。
[0021]
此外,本发明上述内生蜡样芽孢菌或/和上述的内生蜡样芽孢菌的代谢物的下述任 一种应用:
[0022]
1)所述内生蜡样芽孢菌或/和上述的内生蜡样芽孢菌的代谢物在抑制病原菌中的 应用;
[0023]
2)所述内生蜡样芽孢菌或/和上述的内生蜡样芽孢菌的代谢物在制备病原菌抑制 剂中的应用;
[0024]
3)所述内生蜡样芽孢菌或/和所述内生蜡样芽孢菌的代谢物在制备病害抑制剂中 的应用;
[0025]
4)所述内生蜡样芽孢菌或/和上述内生蜡样芽孢菌的代谢物在抑制病害中的应用。
[0026]
上述方案中,本发明是在中国上海市松江区从健康小白菜根际中分离出118株 细菌。
[0027]
首先通过评估芸苔根肿病菌休眠孢子萌发的抑制率和活力测定的技术方案来 选择菌株。
[0028]
只有八种细菌分离物对引起根瘤的休眠孢子具有抑制作用。其中,mz-12的抑 制作用最为显著(73.4%)。与未接种mz-12的植物以及仅受根肿病菌感染的植物 相比,接种
mz-12可以提高作物生物量。如感病指数所示,与仅感染根肿病菌的植 物相比,mz-12共同接种可使植株根瘤减少64%。同时,选择植株在感染根肿病 菌7、14和21天三个时间点再接种细菌菌株mz-12。发现三个时间点重复接种mz-12 菌株比只在任何一个时间点接种可以更有效地抑制根毛感染。
[0029]
采用定量聚合酶链反应(qpcr)的技术评估根肿病菌相关基因在侵染过程中的 相对表达差异,并对病原体的侵染过程进行了验证。
[0030]
根据16s rdna序列分析,菌株mz-12(mt889679)与蜡状芽孢杆菌 (mt598023.1)型菌株具有100%的序列相似性。这也是蜡状芽孢杆菌可作为生物 防治剂防御根肿病的首次应用。
[0031]
本发明公开了一种可从生物学角度控制或抑制根肿病发生的方法。目前的研究 评估了分离的内生细菌蜡状芽孢杆菌(mz-12)在抑制小白菜根肿病发生的作用。 蜡状芽孢杆菌(mz-12)不仅对白菜的植物生长和营养品质具有有益的作用,还可 以通过直接抑制根肿病菌游动孢子来抑制感染。
附图说明
[0032]
通过阅读参照以下附图对非限制性实施例所作的详细描述,本发明的其它特征、 目的和优点将会变得更明显:
[0033]
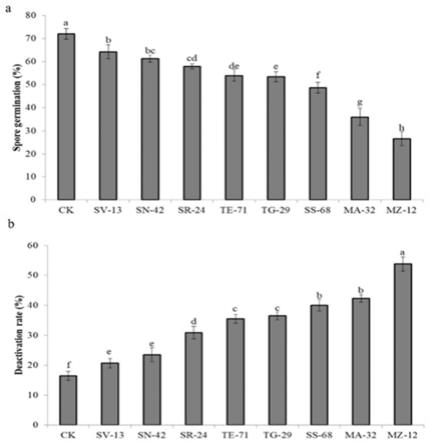
图1显示了拮抗菌株对休眠孢子囊发芽和活力的抑制作用;其中,图(a)表示 接种不同细菌分离物后对休眠孢子囊的发芽率;图(b)表示接种了不同细菌分离 物后的休眠孢子囊的失活率;实验数据采用三次生物学重复,通过lsd测试确定, 具有不同字母的表示p<0.05时有显著差异(下列各图均采用相同表示方法);
[0034]
图2为评估分离细菌菌株(mz-12)对根肿病抑制作用的时程试验示意图;
[0035]
图3(a)为小白菜的表型;;图3(b)为小白菜根瘤的表型;其中,c表示未 经处理;mz-12表示仅用细菌菌株处理;pb:表示仅用根肿病菌;pb.mz表示用细 菌菌株和根肿病菌共接种处理(下列各图均采用相同表示方法);
[0036]
图4显示了mz-12接种对根肿病菌引起的小白菜根肿病的影响;其中,图4 (a)为感病指数;图4(b)为健康根部和感染根的新重;图4(c)为不同处理下 根的鲜重;
[0037]
图5显示了根肿病菌和mz-12接种对植株形态属性的影响;其中,图5(a) 鲜重(单位:g);图5(b)干重(单位:g);图5(c)茎直径(毫米);图5(d) 叶片数;图5(e)为叶面积(cm2);
[0038]
图6显示了不同处理的根在56天的相对电导率;
[0039]
图7显示了不同处理后根部的超氧化物歧化酶(superoxide dismutase,sod)、 过氧化物酶(peroxidase,pod)和过氧化氢酶(catalase,cat)的活性;
[0040]
图8显示了接种细菌菌株mz-12在不同时间(7、14和21天)对根肿病菌感 染小白菜根毛的影响;
[0041]
图9显示了小白菜根中mz-12菌株在用mz-12培养悬浮液对土壤进行接种第7 天和第14天)后的定量表达;
[0042]
图10显示了芸苔假单胞菌和细菌菌株不同处理第28天和第56天后的白菜根 部中10个根肿病菌的表达分析;
[0043]
图11显示了基于16s rdna序列的蜡状芽孢杆菌mz-12和其他相关物种的系 统发
育树。
具体实施方式
[0044]
下面结合具体实施例对本发明进行详细说明。以下实施例将有助于本领域的技术人 员进一步理解本发明,但不以任何形式限制本发明。应当指出的是,对本领域的普通技 术人员来说,在不脱离本发明构思的前提下,还可以做出若干变化和改进。这些都属于 本发明的保护范围。
[0045]
在拟南芥和小白菜上的各种实验证明,与有益细菌或真菌共同接种可减轻根肿病的 症状。
[0046]
从无症状小白菜根际土壤中分离出的118株细菌菌株。基于它们对根肿病菌休眠孢 子囊的萌发率和活力的抑制作用,我们仅筛选出8种细菌菌株(实验结果如图1所示)。 这8株菌株包括sv-13,sn-42,sr-24,te-71,tg-29,ss-68,ma-32和mz-12,其中 mz-12对休眠孢子囊的萌发表现出最高的抑制作用,该mz-12菌株为本案所述的内生蜡 样芽孢杆菌,其保藏于中国微生物菌种保藏管理委员会普通生物中心,保藏编号为cgmccno.21141,分类命名为蜡样芽孢杆菌(bacillus cereus),保藏日期为2020年11月9 日。(发芽率为26.6%,对照组为72%)(图1a)。我们还评估了这8种菌株对根肿病菌 孢子存活力的抑制作用,三种细菌菌株ss-68(40%),ma-32(42.8%)和mz-12(53.8%) 表现出较高的休眠孢子失活率(对照组为16.5%)(图1b)。拮抗细菌菌株可以通过抑 制休眠孢子萌发来减少根肿病的发生。因此,mz-12可被选作潜在的生物防治剂。
[0047]
我们在温室条件下检测了细菌分离株mz-12对根肿病菌的抑制作用。按照图2定 时检测接种了mz-12菌株的芸薹根肿病菌(plasmodiophora brassicae woronin)(以下 简称根肿病菌)的最大抑制作用。与仅感染根肿病菌的植物相比,细菌分离株mz-12 是抗根肿病菌的有效生物防治剂,并通过减少根瘤形成而有效抑制了根肿病(参考图3a, b)。仅接种根肿病原体的小白菜的染病指数为74.3%,而用mz-12菌株治疗可有效减轻 根肿病,其染病指数为36%(参考图4c)。
[0048]
与单独接种菌株mz-12比较,单独接种根肿病菌或细菌菌株mz-12和根肿病菌共接 种都明显增加了根的鲜重(图4a)。根肿病菌感染引起根瘤形成,因此增加了根的鲜重。 把每个处理下的健康根和感染根做进一步分析,发现与仅感染根肿病菌的植物相比,细 菌分离株mz-12和根肿病菌共接种的根几乎都能正常生长(图4b)。
[0049]
在四种不同处理下(所述四种处理方式包括:对照组,即未做任何处理;仅用细菌 菌株mz-12处理的;仅用根肿病菌pb处理;以及用根肿病菌和细菌菌株pb和mz-12共 同处理)得到了白菜的不同形态学参数,数据见图5(a-e)。细菌分离株mz-12的接种 通过抑制根肿病菌的致病作用,在正常和患病条件下均可以改善植物的生物量。与对照 植物相比,仅用菌株mz-12接种的植株干鲜重分别增加了42.18%和25.1%。同样,细 菌株mz-12和根肿病菌共接种后的植株鲜重也比仅用根肿病菌接种后的植株干鲜重高 51.11%和31%。同时,在正常和染病条件下接种了拮抗菌株mz-12的植株显著地增加 了相应植株地叶片数量、叶面积和植株高度(图5a-e)。这种由细菌分离物增强的生物 量特性表明它是一种促进植物生长的根际细菌(pgpr)。此外,这种植物生长的正调控 也可能与土壤养分吸收的提高有关。
[0050]
四种处理下对植物的pod,sod和cat等抗氧化酶活性进行了分析,结果如图7(a-c) 所示。在正常和染病的条件下,抗氧化酶的活性可以反映细菌菌株mz-12的作用。由 于拮抗性分离株mz-12的作用,植物中的抗氧化酶(pod,sod和cat)的活性分别提高 了15%,30%和23%。同时,与对照相比,根肿病菌感染的植物的pod(48.4%),sod (59.01%)和cat(33.8%)活性也都提高了。但是,细菌株mz-12和根肿病菌共接 种后的植物的pod(82.0%),sod(86.0%)和cat(52.0%)的活性进一步增加。抗 氧化酶可以正向调节植物对病原体的防御作用。已发现pod,sod和cat的酶活性与植 物对病原体感染的抗性相关,并且这些酶的活性变化可以反映出病原体和植物之间的关 系。在我们的研究中,随着染病程度的增加,这三种酶的活性显著下降。与仅接种根肿 病菌的植物相比,同时接种mz-12和根肿病菌的植物的pod,sod和cat的活性更高。 从本案的实验结果表明,增强酶的活性可以增加植物对根肿病菌的抗性。
[0051]
根据不同实验处理的主成分分析,酶活性分析和植物生长指标数据结果显示,细菌 菌株mz-12对接种了根肿病菌的小白菜呈阳性反应。植物生长指数之间的生物图也显示 出显著正相关,而根鲜重,pod,sod和cat酶活性与所有其他参数负相关。
[0052]
mz-12菌株在缓解疾病方面显示出积极的结果,根毛感染(rhi)实验接种的第7、 14和21日采样发现,与仅接种根肿病菌的植物相比,由于接种mz-12延缓或抑制了游 动孢子的感染从而大大降低了rhi的患病率(如图8所示)。同时,随着时间点的延长, rhi总体呈上升趋势。
[0053]
mz-12菌株可以减少小白菜根系根肿病的患病率。休眠孢子萌发和rhi是影响植物 中根肿病菌感染的两个主要方面。当在7、14和21天施用生物防治剂时,其效率得到 了提高。mz-12在三种时间点同时应用比一种更有效地抑制根毛感染。细菌分离株mz-12 的施用主要集中在根毛感染和根或皮层组织上。游动孢子通常会在7到21个das之间 达到最高水平,这是由于在此时间段内三次应用mz-12菌株有助于维持其高浓度。这些 结果与7天,14天和21天在根中检测到的mz-12dna的含量高度相关。
[0054]
使用qpcr评估白菜中mz-12在根中定植。在感染病原体的根中观察到细菌dna, 但在用水处理过的根中未观察到细菌dna(如图9所示)。同时,细菌菌株mz-12的表达 在7、14和21天之间增加。
[0055]
rhi的结果与7天,14天和21天在根中检测到的mz-12dna数量密切相关。在此 阶段,根或皮质感染可能仍处于初始阶段,而rhi可能已达到最大。用细菌分离株接种 的白菜根中的mz-12菌株的种群始终较高(参考图3)。细菌分离株mz-12并未完全消除 rhi,但最终几乎没有出现严重的根肿症状。在接种mz-12菌株的植物中,可能还有其 他一些机制可以抑制白菜根肿病菌感染或根部发育。
[0056]
通过在28天后和收获时(56天)使用qrt-pcr对疾病期间差异表达的11个根肿病 菌的表达分析,证实了通过接种拮抗菌株mz-12对根肿发育的抑制(如图10所示)。28 天后基因分析,例如crac,pro1,tps,brip 9,cc240,cc249,pbstkl1和cc243在 mz-12共接种植物中表现出较低的表达,相反,y10和psa表达水平在共培养56天后降 低-接种(图10a,b)。在两个时间点(28天和56天)的表达水平的确定表明,与brip 和cc243一起接种的植物相比,mbp-12与brip和cc243共同接种时,与m.-12一起降 低了11个选择基因的表达水平。因此,可以提出mz-12对根肿病菌的发病具有抑制作 用。
[0057]
感病指数和分子生物学的数据表明,细菌拮抗剂mz-12除了延缓根肿病菌的生命周 期外,还特别延迟了休眠孢子的产生。mz-12的生物防治机制可能是通过细菌菌株产生 的次级代谢产物来抑制休眠孢子的萌发。结果表明,mz-12菌株能够在小白菜根部定殖 并减少病害。
[0058]
进行16s rdna扩增和测序以鉴定细菌分离株,使用blast搜索程序分析16s rdna 序列,mz-12菌株(登录号:mt889679)的分子特征证实其为芽孢杆菌。表现出与蜡 状芽孢杆菌的相似性为100%(登录号:mt598023.1)。
[0059]
实施例一
[0060]
2019年,在中国上海松江区土壤染病比较严重的基地,从无症状小白菜的根际土壤 中收集样品。为了进行细菌分离,将10g根际土壤悬浮于100ml无菌蒸馏水(sdw) 中,并用sdw连续稀释至10-3
、10-4
、10-5
和10-6
。取每种稀释液50μl涂抹在lb培养 基中(10g/l胰蛋白、10g/l nacl、5g/l酵母提取物和20g/l琼脂,ph 7.0),并在 28℃下培养。3天后接种,挑取菌落于lb液体培养基中,28℃下以180rpm的振荡 培养。24小时后收集细菌悬浮液,并稀释至107cfu
·
ml-1
。
[0061]
从种植在中国上海松江区的染病小白菜的根瘤中分离出根肿病菌的休眠孢子。首先 用搅拌机把小白菜的根瘤研磨均匀,然后将所得的浆液通过孔径为25μm的纱布过滤后, 5000rpm离心滤液10分钟得到孢子。最后用血细胞计数器测定休眠孢子浓度,用无菌 水稀释至1
×
107·
ml-1
以接种土壤。
[0062]
小白菜“xinxiaqing”种子是由中国寿光仁和种业有限公司提供。种子用1%次氯 酸钠表面消毒5分钟,然后用sdw洗涤三次。将小白菜种子播种在装有高压灭菌珍珠岩 的塑料托盘中,并用murashige skoog营养液浇灌。然后将其放置温室内,生长条件为: 相对湿度60%,光照/黑暗16h/8h,28℃(白天)/15℃(夜间)。7d后,将生长一 致的幼苗移栽到30孔的穴盘中(2cm
×
2cm)。第一个实验包括以下四种处理:(1)对 照组(c);(2)细菌分离株(mz-12);(3)根肿病菌(pb);(4)根肿病菌+mz-12(pb.mz)。 在每个穴盘浇灌600ml休眠孢子悬浮液和300ml细菌培养液,而对照组的幼苗则以600 ml无菌水处理。在初次接种后的第7天和第14天,将幼苗用300ml细菌培养物再次接 种,同时将对照组和患病幼苗用等体积的去离子水处理。实验独立重复三次。种子发芽 后56天进行病害指数评估。
[0063]
实施例二
[0064]
为了获得根分泌物,将生长五天的小白菜幼苗分别转移到装有hoagland溶液的容 器(直径4厘米
×
高5.5厘米)中。并在24℃下培养7天。用0.22μm的硝酸纤维素 过滤器过滤后,收集根分泌物并将其置于4℃下保存。为了制备分离菌株培养物,挑选 细菌分离株的单个菌落于液体lb培养基中,在28℃下180rpm培养3天,然后8000rpm 离心细菌培养物15分钟,并将上清液通过0.22μm硝酸纤维素过滤器。
[0065]
为了进行孢子萌发抑制试验,用根系分泌液(ph 6.3)将休眠孢子悬浮液稀释至 1
×
107·
ml-1
。将0.5ml细菌液体和4.5ml休眠孢子悬浮液放于一个50ml的烧瓶,另 一个烧杯装有以lb培养液为对照的休眠孢子悬浮液,并都放在黑暗中25℃培养。在bs203光学显微镜下,用乙霉素(1%)染色法观察到休眠孢子的萌发。每个实验重复三 次。采用zhu等人所述的公式计算休眠孢子的发芽抑制率。
[0066]
所述公式具体如下:
[0067]
n=[(a-b)/a]
×
100
[0068]
上式中,n表示休眠孢子萌发抑制率;a表示未处理下的休眠孢子萌发率;b表示处 理下的休眠孢子萌发率。
[0069]
为了测定休眠孢子的活力,将0.5ml细菌液体与0.5ml芸苔假孢子悬浮液(4
×
107·
ml-1
)在无菌试管中混合,并在25℃下培养2天。将每种处理分别添加到等体积的磷 酸盐缓冲液(ph 7.4)中,然后用1ml的evans blue溶液(淡红色)混合。之后放在 室温下30分钟,并用无菌蒸馏水洗涤四次。在bs203显微镜下观察休眠孢子的活力, 没有活力的孢子显示为深蓝色。无菌lb培养基作为对照。处理重复3次。采用zhu等 人所述的公式计算非存活休眠孢子的校正频率。
[0070]
所述公式具体如下:
[0071]
n=[(a-b)/a]
×
100
[0072]
上式中,n表示休眠孢子萌发抑制率;a表示未处理下的休眠孢子萌发率;b表示处 理下的休眠孢子萌发率。
[0073]
实施例三
[0074]
发芽56天后,收获植物以表现形态特征,例如鲜重的根/茎,株高,叶数和叶面积, 在收获时手动测量形态学指标,而将地上部在75℃下干燥72h用于干生物量测定。
[0075]
在测完根的鲜重后,使用手术刀将处理过的根的健康和感染部位分离,并分别测量 两个分离组织的鲜重。播种56天后,如huang和siemens所述,使用病害指数(di) 对每种植物的病害程度进行定性评估,并使用五类量表方法分析了根肿病病症。每个处 理约一个穴盘(25株植物),每个实验三个穴盘。比较均值来分析差异性。
[0076]
实施例四
[0077]
萌发56天后,分析了根系中过氧化物酶(pod),超氧化物歧化酶(sod)和过氧化 氢酶(cat)等抗氧化酶的活性。使用试剂盒(南京建城生物技术研究所的pod-a084-3 sod-a001-1和cat-a007-1)分别用分光光度法在420、550和405nm处分析pod,sod 和cat的活性。
[0078]
实施例五
[0079]
如先前所述进行电导率的测量。首先用sdw冲洗根的样品,并保存15ml去离子水 的封闭的小瓶中,然后在25℃下培养1h。测量初始电导率(e1)后,将根样品高压 灭菌20分钟,以获得最终电导率(e2)。使用以下公式评估每个样品的相对电导率(rec):
[0080]
rec(%)=(e1
÷
e2)
×
100。
[0081]
实施例六
[0082]
为了观察根毛感染情况,将生长7天的小白菜幼苗分别转移到装有沙土的容器中(直 径4厘米
×
高5.5厘米)。用黑纸覆盖进行黑暗处理,从而有助于休眠孢子发芽并感染。 移植后,立即用1ml拮抗菌株mz-12的细菌液和1
×
107的根肿病菌悬浮液处理幼苗,使 用静止的孢子悬浮液作为对照。接种后第7、14和21天在显微镜下观察根肿病菌的根 毛侵染情况。将处理组和对照组的根切成1厘米的小块,浸入固定液(70%酒精)中10 分钟,然后用乙酰-大黄素(1%)染色过夜。利用显微镜观察根毛中的游动孢子。每株 植物监测35根根毛,并记录了感染的发生率。
[0083]
实施例七
[0084]
为了通过定量实时pcr(qpcr)确定内生菌的定殖,在接种后7、14和21天采集 根,
冲洗根后提取细菌gdna,并在以下条件下进行qpcr定量实验:第一步,95℃预变 性10min;第二步,95℃变性15s,60℃退火1min和72℃延伸15s,共执行40个循环。
[0085]
特异性引物f为:(5'-gcc ctg gta tgt ata ttg gat cta c-3',其序列如seq id no.1所示;
[0086]
引物r(5
′-
ggt cat aat aac ttc tac agc agg a-3
′
,其序列如seq id no.2 所示。校准曲线估算出的蜡状芽孢杆菌(mz-12)dna量是根据基因组细菌dna样品稀释 5倍得到的。
[0087]
实施例八
[0088]
利用rna提取试剂盒(tiangen,rnaprep纯植物试剂盒)在28天和56天从染病样 品中提取总rna。使用nanodrop 1000分光光度计(thermo scientific,美国)测量 rna的浓度和质量。使用primescript
tm
rt试剂盒将总rna反转录成cdna。
[0089]
表1中列出的引物用于检查根肿病菌基因的相对表达。使用sybr1 premix ex taq tm
(takara)进行扩增,程序为:第一步,95℃预变性10min;第二步,95℃变性15s, 60℃退火1min和72℃延伸15s,共执行40个循环。使用根肿病菌的肌动蛋白表达作为 参考基因来计算相对表达量。
[0090]
实施例九
[0091]
选择具有生物防治效力的细菌分离株mz-12用于16s核糖体dna(rdna)序列的扩 增。用qiagen genomic dna kit提取细菌分离物的基因组dna。通过takara taq聚 合酶(takara biotechnology)扩增16s rdna。使用通用引物27f (5'-agagtttgatcctggctcag-3',其序列如seq id no.3所示)和1492r(5'-ggttaccttgttacgactt-3',其序列如seq id no.4所示)鉴定mz-12菌株。按照以下步骤进行 聚合酶链反应(pcr):94℃预变形5min,然后95℃变形40s,55℃退火40s,72℃ 延伸90s进行35个循环,最后72℃延伸10min。pcr产物的测序由tsingke biologicaltechnology进行,并使用blast搜索程序(https://blast.ncbi.nlm.nih.gov/blast) 进行分析。与利用mega 6.0将细菌分离物的核苷酸序列进行比对,包括进行聚类分析, 构建系统发育树,如图11所示。
[0092]
需要说明的是,本发明的保护范围中现有技术部分并不局限于本申请文件所给出的 实施例,所有不与本发明的方案相矛盾的现有技术,包括但不局限于在先专利文献、在 先公开出版物,在先公开使用等等,都可纳入本发明的保护范围。
[0093]
此外,本案中各技术特征的组合方式并不限本案权利要求中所记载的组合方式或是 具体实施例所记载的组合方式,本案记载的所有技术特征可以以任何方式进行自由组合 或结合,除非相互之间产生矛盾。
[0094]
以上对本发明的具体实施例进行了描述。需要理解的是,本发明并不局限于上 述特定实施方式,本领域技术人员可以在权利要求的范围内做出各种变化或修改, 这并不影响本发明的实质内容。在不冲突的情况下,本申请的实施例和实施例中的 特征可以任意相互组合。
[0095]
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1