一种鸭源CCCH型锌指抗病毒蛋白及其应用
一种鸭源ccch型锌指抗病毒蛋白及其应用
技术领域
1.本发明属于医学或兽医学技术领域,具体涉及一种鸭源ccch型锌指抗病毒蛋白(duck ccch
‑
type zinc finger antiviral protein,duzap)及其应用。
背景技术:
2.鸭在我国广泛被养殖,是重要的经济动物。鸭因天性喜水,不适合完全集约化笼养,多采用半散养模式,其特殊的养殖模式导致长期以来,存在着老病不断,新病层出等突出问题。因此,深入挖掘、利用与鸭天然免疫调控相关的抗病毒基因不仅有助于了解水禽天然免疫分子的调控机理,也可以为鸭抗病毒药物的研发和提高鸭存活率等方面提供理论依据和应用指导。
3.天然免疫系统是宿主抵抗病原感染的第一道防线,病毒感染机体后通过一系列级联反应诱导i型干扰素和其他先天性免疫基因的表达。不论体外还是体内,干扰素(interferon,ifn)通路都发挥着主要的抗病毒作用,是目前免疫学的研究热点。而与人源和鼠源等脊椎动物相比,禽类含有法氏囊等特有的免疫器官,免疫系统也具有显著的差异,尤其是鸭作为一种经济动物,相关研究相对比较滞后,目前有关鸭的天然免疫应答反应、信号转导过程和干扰素下游isg的表达及具体调节机制尚不十分清楚。
4.本发明所述的duzap也叫parp
‑
13,属于poly(adp
‑
ribose)polymerases (parps)超家族,含4个保守的ccch锌指模体的蛋白家族,目前已经发现了至少 17个成员,它们参与细胞增殖、凋亡等细胞进程。近年的研究表明,parp家族成员主要通过保守的4个ccch锌指模体直接结合rna或dna病毒的基因组导致病毒降解从而达到抗病毒的作用。最新的研究显示,根据c端结构域的不同,人源zap鉴定出四个结构和功能不同的剪切变异体,但是关于鸭源zap的基因结构特征和细胞功能方面的信息知之甚少。
技术实现要素:
5.本发明所要解决的技术问题是提供一种鸭源ccch型锌指抗病毒蛋白及其应用。
6.为解决上述技术问题,本发明提供的技术方案是:
7.提供一种鸭源ccch型锌指抗病毒蛋白(duck ccch
‑
type zinc finger antiviralprotein,duzap)基因,所述鸭源ccch型锌指抗病毒蛋白基因的碱基序列如seqid no.1所示。所述duzap基因的n端具有4个重复的ccch锌指模体结构,c 端包含1个wwe和parp功能域。
8.本发明还包括上述鸭源ccch型锌指抗病毒蛋白基因的编码蛋白,所述鸭源 ccch型锌指抗病毒蛋白基因的编码蛋白的氨基酸序列如seq id no.2所示。
9.本发明的又一个方面是一种带有上述鸭源ccch型锌指抗病毒蛋白基因的真核表达载体。
10.本发明的又一个方面是上述鸭源ccch型锌指抗病毒蛋白的应用,所述鸭源 ccch型锌指抗病毒蛋白在制备鸭抗病毒感染药物或疫苗佐剂中的应用。
11.本发明的又一个方面是上述真核表达载体的应用,所述真核表达载体在制备鸭抗病毒感染药物或疫苗佐剂中的应用。
12.按上述方案,所述真核表达载体及鸭源ccch型锌指抗病毒蛋白在鸭胚成纤维细胞(duck embryo fibroblast cells,defs)中过表达,通过激活转录因子nf
‑
κb 和irf1诱导促进鸭i型干扰素β(interferonβ,ifn
‑
β)分泌。
13.本发明扩增得到duzap基因全长,并对其进行了生物信息学分析,明确了 duzap的结构特征及同其它物种zap之间的进化关系。duzap的n端具有4个重复的ccch锌指模体结构,c端包含1个wwe和parp功能域。
14.本发明分析了duzap的鸭组织分布、细胞定位及其结构域功能。并通过双链 rna类似物poly(i:c)和鸭坦布苏病毒(duck tembusu virus,dtmuv)来研究duzap 在鸭源天然免疫信号通路中的作用。本发明证明了在鸭胚成纤维细胞(duck embryofibroblast cells,defs)中,duzap能通过激活转录因子nf
‑
κb和irf1诱导促进鸭 i型干扰素β(interferonβ,ifn
‑
β)分泌。通过构建duzap不同结构域的截短突变体的真核表达质粒发现,确定其c端的wwe和parp结构域对激活干扰素信号通路是必需的。
15.本发明所述的鸭源ccch型锌指抗病毒蛋白为申请人首次从鸭组织中扩增解析,相关序列信息已上传至美国国家生物信息中心ncbi(http:// www.ncbi.nlm.nih.gov/),登录号为mn736630,全长cdna为2154bp。
16.发明效果:
17.本发明的积极效果在于细胞内证明了duzap表达能显著促进鸭ifn
‑
β分泌,干扰duzap的表达,poly(i:c)和dtmuv激活的ifn
‑
β分泌则会明显被抑制。duzap 有望成为增强鸭体自身免疫的新靶点,在作为鸭抗病毒感染药物及疫苗增强剂等方面具有较大的应用潜力。
附图说明
18.图1a为鸭源duzap表达载体的双酶切鉴定图,图1b为duzap基因全长的结构域模式图;
19.图2为健康鸭各组织的duzap分布图;
20.图3为duzap的细胞定位图;
21.图4为duzap对于鸭源ifn
‑
β启动子活性的影响图;
22.图5为干扰def细胞内源性duzap的表达与ifn
‑
β的启动子活性关系图;
23.图6为duzap过表达对鸭坦布苏病毒(duck tembusu virus,dtmuv)复制的影响图。
具体实施方式
24.本发明更详细的实施方法可参见实施例,本实施例是用于解释、而不是以任何方式限制本发明。实验所涉及的药品购自上海罗氏制药有限公司、普洛麦格公司、生工生物工程(上海)股份有限公司,β
‑
actin鼠单克隆抗体、抗flag标签的鼠单克隆抗体购自mbl公司;异硫氰酸荧光素(fitc)标记的羊抗鼠igg二抗均购自上海碧云天生物技术有限公司。具体实验操作依据《分子克隆实验指南》。本发明实施例中%未说明的情况下指的是体积百分比。
25.实施例1
26.鸭源ccch锌指抗病毒蛋白duzap的分子克隆
27.1.1鸭源zap基因的克隆和真核表达质粒pzap
‑
flag的构建
28.1.1.1取健康鸭脾脏,利用trizol法提取总rna,方法如下:
29.取黄豆大小的鸭脾脏组织,用生理盐水研磨后12,000rpm离心5min,取200 μl上清,加入1mltrizol,室温放置5min;加入200μl氯仿,剧烈振荡15s后室温放置3min,使其自然分相;4℃、12,000rpm离心10min,将上清小心移至新的eppendorf管(无rnaase)中,加入等体积的异丙醇,混匀后室温放置10min;4℃、12,000rpm离心10min,小心弃上清;向rna沉淀中加入1ml 75%乙醇(用 rnase
‑
free水配制),温和悬浮沉淀;4℃、8,000rpm离心5min,小心弃上清,室温干燥沉淀;向沉淀中加入50μl rnase
‑
free水,置于55℃下静置10min,以充分溶解rna,测定浓度,分装于
‑
80℃保存备用。
30.1.1.2逆转录反应(rt)
31.用商品化的试剂盒,参考罗氏反转录试剂盒说明书进行。对不同样品的rna 进行等质量的反转录。1μl oligo(dt)18引物、1μg rna样品,再补加depc水至 13μl,置于pcr仪65℃下处理10min,反应结束后将产物置于冰上,随后加入4μl 的5
×
rt缓冲液、2μl的dntps、0.5μl的rna酶抑制剂、0.5μl的反转录酶,混匀后,置于pcr仪,55℃处理30分钟、85℃处理5min,得到cdna样品。
32.1.1.3目的基因的获得及真核质粒的构建
33.用设计的两端带有酶切位点(ecorⅰ和xhoⅰ)目的片段的上下游引物pcr 扩增duzap基因的全长cdna,最后将其连入pcaggs
‑
flag真核表达载体中送往测序。扩增条件为:95℃5min预变性;95℃40s,74℃90s,35个循环;72℃10min 片段延伸。上下游扩增引物如下:
34.pduzap
‑
f:cggaattcatgtgcgattcggtggtgtgcag
35.pduzap
‑
r:ccgctcgagttaagatataatacatttttttcctg
36.以步骤1.1.2中制备的脾脏cdna为模板,用上述带有酶切位点的特异性引物进行pcr扩增,得到大小在2000bp左右的扩增产物。根据《分子克隆实验指南》的常规试验技术,将pcr扩增得到的目的片段和空载pcaggs
‑
flag同时进行双酶切回收,16℃连接过夜,转化至dh5a感受态细胞,涂布抗性平板培养过夜,挑菌扩大培养提取质粒进行双酶切鉴定,双酶切鉴定图详见图1a,泳道1
‑
3为duzap 的真核表达质粒pzap
‑
flag/ecori+xhoi双酶切电泳图,将双酶切正确的质粒送往生工生物工程(上海)股份有限公司进行测序,经测序验证序列正确性、无移码突变,证明pcaggs
‑
duzap
‑
flag载体(简写为pzap
‑
flag)构建完成。
37.鸭源duzap的相关序列信息已提交至美国国家生物信息中心ncbi(http:// www.ncbi.nlm.nih.gov/),登录号为mn736630,全长cdna为2154bp。
38.duzap基因的碱基序列seq id no.1:
39.atgtgcgattcggtggtgtgcagcttcctcaccaagaccctgtgtgcccagggcggccgg ctggggctcacagagctccgggagcacatcgacctgtcggagcagcagctgatggagac gctgcgggcagcggggccccggcggttcctgctaactgcagatgacggcctccccaccg aggtgctggccgtgtcggacgtgcgggtctgtgtcctcaaggagtgcccgggctgcgac cgcctgcacctctgcaagctgcacctcgggggcaagtgcaacctgggtcccagagcttg taggtactctcatgacatcagcagcactgagaacaaaaaagtcctaaagacccatgagt tgtctggcctc
agtgaggatgagctgcgagtcctgcttctccaaaatgaccctttcttcc ttcctgatgtctgccaattttacaacagaaagggtggtgcctgcaaccagaaaaacaat tgcagcaggcttcatatttgccgacactatctcaaaggaaattgtaaattttttgtatgc aagagatcccataagctcttggatgaacactcactgagagtgttggaaactgcaggcat tgatgcaaagatagttgcaaacttccaaattatatacgatcacaagcatgtggagttca acaaggaacagaacaaggagaaaactaaaccccatcatcaccgtgtttactactgtaag ccagtaaaaacttcgatgaaaaaacaggcagatacaatgagcgaagaacagaagctac ctttagaaacaacagtgggctccggggtgcatgtaccaccttcaaaagcttcaaaagat cccagtagcaacgaacctcctcagagccaactccagctgccagcaggtgccagaggtaa agatgaaggtaaaaaagacgattcctctgcgaacattttgaaggacaagaaggaagat aacactgaagagatctgcttgttctatgtctggaagtactgcaaacataatgacaaatg cagatcgattcattaccatttgccatatcgatggcaagtatttaatgggttcacctggaa tgacctttccatgatggaggaaattgaaaaggcctattgtgacccaaaaatgaacagta tagcagataagaacattaatttccagacaatgacctgctcctcttcattgcttcgacgtc tctctacaccatcatctgtcacacaacccatgtttgtattgactacaaagtggatttggt attggcagaatgaccaaggccagtgggttgaatatggagaacagggagaaggtgatgt tgtgaactcgccatcttctgacatacttgagaatttgtatctggcagatccagatgccac cgtacctttccaggctggcttgtatggttaccagctcaattttaaagaaatgacccagac aaacatttattctaaaactcaaagacgggtctgcaggcgaccaaagtttgtgtcttatga agaagtgcagaagataaagcaaagcagtcagagggattcttctattccaaatcaagcct gtcctccccactgggatccatttgacttgtctgacttaaaatacaaggcagtggagatca gtaacacatcctctgaatacaaagaaataaagaagcagtttgagcagactatgaaaacc tacaacatcgttagagtacaaaggattcagaatccatcactctggaaagtatttcagtg gaaaaaggagcaaatgaagaaggaagctggaaagaaggaagtaaacgaaaggctcct gttccatggaaccaaggatgcccgcctggaagatatctgcataaacaactttgactgga gaacttgtggaagcaatggagccaactatggaaagggaagttactttgctagagatgct tcatattcccatgagtactgtcagtctgcagtgaaagcaaacgtcatgttcatggctcgt gtattggttggagattatgttcgaggcagggaagactacgttcgccccccaccaaagtct gttgatgggctttgtttttatgacagttgtgtggacaacaagttaaatccctccatttttg ttatctttgaaaaaaatcaaatttacccagagtatctgatagaatataaggaggcagga aaaaaatgtattatatcttaa
40.duzap蛋白的氨基酸序列seq id no.2:
41.mcdsvvcsfltktlcaqggrlgltelrehidlseqqlmetlraagprrflltad dglptevlavsdvrvcvlkecpgcdrlhlcklhlggkcnlgpracryshdisst enkkvlkthelsglsedelrvlllqndpfflpdvcqfynrkggacnqknncsrl hicrhylkgnckffvckrshklldehslrvletagidakivanfqiiydhkhvef nkeqnkektkphhhrvyyckpvktsmkkqadtmseeqklplettvgsgvhvpp skaskdpssneppqsqlqlpagargkdegkkddssanilkdkkednteeiclfyv wkyckhndkcrsihyhlpyrwqvfngftwndlsmmeeiekaycdpkmnsiad kninfqtmtcsssllrrlstpssvtqpmfvlttkwiwywqndqgqwveygeqg egdvvnspssdilenlyladpdatvpfqaglygyqlnfkemtqtniysktqrrv crrpkfvsyeevqkikqssqrdssipnqacpphwdpfdlsdlkykaveisntssey keikkqfeqtmktynivrvqriqnpslwkvfqwkkeqmkkeagkkevnerllf hgtkdarledicinnfdwrtcgsnganygkgsyfardasysheycqsavkanv mfmarvlvgdyvrgredyvrpppksvdglcfydscvdnklnpsifvifeknqiy peylieykeagkkciis
42.1.2.鸭源duzap基因全长的序列信息与特性分析
43.序列分析表明,duzap基因全长2154bp,编码717个氨基酸多肽,预测蛋白质分子量约为79kda。duzap的c端有四个重复的ccch保守的锌指结构域 (ccch
‑
zinc fingers),n端含有一个wwe和parp结构域,duzap基因全长的结构域模式图见图1b。利用mega6软件对鸭源
duzap与已报道的其它脊椎动物zap进行序列比对和进化分析,结果表明,duzap与鸡源的zap有82.1%同源性,与人源的zap仅有44.4%同源性,这说明各物种之间zap蛋白的差异较大。
44.1.3.鸭源duzap基因在健康鸭体内的表达模式分析
45.取3只7日龄健康雏鸭为试验动物,分别取心脏(heart)、肝脏(liver)、脾脏(spleen)、肺脏(lung)、肾脏(kidney)、腺胃、胰脏(pancreas)、肠道(small instestine)、法氏囊(bursa of fabricius)、胸腺(thymus)、大脑(brain)、腿肌(muscle)十二个组织,﹣80℃冻存备用。
46.1.3.1每份组织约为1g,用生理盐水研磨后取200μl上清用trizol法按实施例1中步骤1.1.1提取组织中rna。
47.1.3.2将提取的rna反转录成cdna,用商品化的试剂盒,参考罗氏反转录试剂盒说明书进行,具体操作按照实施例1中的操作步骤1.1.2进行。
48.1.3.3duzap实时荧光定量pcr(rt
‑
qpcr)
49.用荧光定量pcr方法检测目的基因的mrna表达。
50.目的基因duzap特异性引物如下
51.上游引物qduzap
‑
f:tgtgcagcttcctcaccaag
52.下游引物qduzap
‑
r:tctgcagttagcaggaaccg
53.内参基因gapdh
54.上游引物qgapdh
‑
f:atgagaagtatgacaagtcc
55.下游引物qgapdh
‑
r:actgtcttcgtgtgtggct
56.使用sybr green试剂盒(toyobo公司)检测样品中duzap的mrna表达。反应体系如下:10.0μl的sybr green i master、0.5μl的上游引物、0.5μl的下游引物、8μl的灭菌水、1μl的cdna模板。同时设置gapdh作为内参。pcr 扩增条件为:94℃10min进行预变性;94℃15s,56℃30s,72℃40s进行40个循环的扩增并收集荧光信号。每个样品3个重复,应用δδct法计算每个基因的相对mrna表达水平。如图2所示为健康鸭各组织的duzap分布图,结果表明duzap 在鸭体各组织中广泛存在,其中在免疫器官脾脏和胸腺中的含量最高。
57.实施例2
58.鸭源ccch锌指抗病毒蛋白duzap的应用
59.2.1脂质体介导转染鸭胚成纤维细胞及细胞定位
60.用脂质体介导法将构建的真核表达质粒pzap
‑
flag转染进defs(实验室保存的传代细胞系)中,转染后24h,弃去细胞上清,用间接免疫荧光法对细胞进行固定,孵育相应抗体,用激光共聚焦显微镜观察,如图3所示为duzap的细胞定位图,分别将空载体pcaggs
‑
flag和pzap
‑
flag转染def细胞,24
‑
36小时后弃去细胞上清进行间接免疫荧光试验,图中vector为空载组(空白对照组),duzap 为目的基因真核表达载体pzap
‑
flag组,fitc表示目的基因duzap荧光染色(绿色),dapi表示细胞核染色(蓝色),merge为细胞核与目的基因共定位,结果表明,目的基因duzap大量表达在def细胞的胞浆中。
61.2.1.1真核细胞的质粒转染
62.转染试剂用jettransfection reagent,按照说明书进行。
63.以24孔板为例。将适量的def细胞接种到24孔板中,次日细胞待长至60%~ 70%
时即可转染。转染前换为无血清和无抗生素的培养液。将真核表达质粒 pzap
‑
flag(对照组用空载体质粒代替),质粒总量按1μg/孔的量,依次加入到jet transfection reagent试剂盒配套的转染buffer中,涡旋混匀后加入2倍质粒剂量的转染试剂,涡旋混匀后瞬时离心,室温静止10min即可加入到24孔板中, 4h后换上含2%胎牛血清(fetal bovine serum,fbs)的无双抗细胞维持液。转染后 24h,即可弃去细胞上清用于后续试验。
64.2.1.2间接免疫荧光法检测duzap细胞定位
65.接种细胞前,将细胞爬片置于24孔板中。收样时,弃去细胞板中的培养基上清,将预冷的pbs沿细胞孔边缘缓慢加入,置于摇床,缓慢摇转5min。如此反复洗2次。随后加入4%多聚甲醛300μl进行固定10min,弃去多聚甲醛后,加入 800μl预冷甲醇。透化10min后,pbs洗3次。加入300μl 5%(质量百分浓度) 浓度的牛血清蛋白封闭1h。弃去封闭液后,加入150μl稀释的抗flag标签的鼠单克隆抗体的一抗,室温孵育1h,pbs洗3次。弃去上清后,再加入150μl稀释的fitc标记的羊抗鼠igg二抗,避光孵育40min,弃上清后加入稀释的dapi染料,孵育15min后,pbs缓冲液洗3次。取出爬片,用50%浓度(体积百分浓度) 的甘油将爬片密封于载玻片,使用lsm 900meta激光共聚焦显微镜观察目的蛋白的表达、定位。
66.2.2鸭源duzap对ifn
‑
β启动子活性的影响
67.通过荧光素酶检测系统,将pzap
‑
flag分别与鸭干扰素β启动子荧光素酶报告质粒pgl
‑
ifnb
‑
luc及其启动子转录因子irf1和nf
‑
κb结合位点的荧光素酶报告质粒pgl
‑
irf1
‑
luc和pgl
‑
nf
‑
κb
‑
luc(实验室构建,具体方法参照步骤1.1.3) 共转染def细胞,ptk作为内参同时转染,24小时后,使用poly(i:c)刺激,刺激 16小时后收集细胞,使用双荧光素酶报告基因检测试剂盒进行检测,检测荧光素酶活化情况。duzap对于鸭源ifn
‑
β启动子活性的影响图见图4,图中纵坐标显示值为萤火虫荧光素酶与海肾荧光素酶的比值。western blotting检测转染质粒蛋白的表达。图在a表明不同剂量(0.25μg、0.5μg和1μg)pzap
‑
flag激活鸭干扰素β启动子呈剂量依赖性,b和c表明pzap
‑
flag能显著激活干扰素β启动子的转录因子irf1和nf
‑
κb结合位点,通过荧光素酶产生的变化检测发现,duzap与ifn
‑
β启动子的激活呈剂量依赖性,说明超表达zap可以激活ifn
‑
β启动子的活化(图 4a)。转录因子irf1和nf
‑
κb是激活ifn
‑
β信号通路的两个关键因子,图4b和 c说明,duzap能通过转录因子irf1和nf
‑
κb激活ifn
‑
β的表达。
68.2.2.1真核细胞的基因转染
69.转染试剂用jettransfection reagent,按照说明书进行。具体参照实施例2中的步骤2.1.1。
70.用无血清和无抗生素的培养基,将适量的def细胞接种到24孔板中,待细胞待长至50%~60%时即用于转染。将pzap
‑
flag分别与鸭源ifn
‑
β荧光素酶报告质粒pgl
‑
ifnb
‑
luc、pgl
‑
irf1
‑
luc和pgl
‑
nf
‑
κb
‑
luc,以及内参海参荧光素酶 tk质粒等真核表达质粒(对照组用空载体质粒代替)共转染细胞,具体转染质粒剂量如下:pgl
‑
ifnb
‑
luc、pgl
‑
irf1
‑
luc和pgl
‑
nf
‑
κb
‑
luc均为0.1μg/孔,内参海参荧光素酶tk质粒(购自promega公司)0.01μg/孔,pzap
‑
flag等真核表达质粒(对照组用空载体质粒代替)总量1.0μg/孔。按上述剂量依次将质粒加入到试剂盒配套的转染buffer中,转染的质粒总量相等(不足的用空载体质粒补足),随后按步骤加入2倍剂量的转染试剂,将质粒转染进defs,4h后换上含2%fbs的无双抗细胞维持液。每组试验设立3个重复。
71.2.2.2报告基因转录活性检测
72.双荧光素酶检测实验按照检测试剂盒说明书进行操作,其过程如下:
73.第一步裂解细胞:转染24
‑
36h后,移去24孔板中的培养基,预冷的pbs缓冲液洗涤两次,加入试剂盒中的5倍稀释好的裂解液150μl,放在摇床上摇动30 min。
74.第二步收集细胞上清:收集细胞裂解液,12,000r/min离心2min,收集上清。
75.第三步荧光素酶活性的测定:将20μl上清液加入100μl试剂盒中预先配好的lar ii中,td
‑
20/20luminometer上检测萤火虫荧光素酶(firefly luciferase)活性。随后加入100μl试剂盒中预先配好的stop&reagent中检测海肾荧光素酶(ranilla luciferase)的活性。并由仪器计算出其相对活性。
76.2.3、下调duzap的表达促进内源性ifn
‑
β的表达
77.用rna干扰的方法,针对目的基因duzap合成了三对rna小分子,def 细胞转染duzap的sirna 36h后,用trizol收样通过荧光定量pcr检测了内源性zap的mrna水平,结果发现干扰分子siduzap
‑
944的干扰效果最显著。进一步将筛选出的干扰分子siduzap
‑
944和阴性对照sinegative control分别与鸭源 ifn
‑
β启动子的荧光素酶报告质粒和内参tk质粒共转染defs,通过双荧光素酶报告试验表明,内源性zap的表达降低后可以使poly(i:c)和dtmuv刺激的ifn
‑
β表达显著降低。
78.sirna干扰duzap
79.duzap的sirna(smallinterferencerna,sirna),三对靶序列由吉玛公司合成),
[0080][0081]
将sirna配成20μm的母液
‑
20℃下储存备用,工作母液为2μm,全程严格无菌,用无rna酶的枪头进行操作。
[0082]
2.3.1sirna小分子干扰效率的检测
[0083]
def细胞中分别转染duzap干扰分子阴性对照siduzap
‑
944、siduzap
‑
1576 和siduzap
‑
2283 36小时后,提取细胞总rna,进行荧光定量pcr检测内源性 duzap的转录水平,筛选出干扰效率最高的干扰分子。转染试剂用jet transfection reagent,按照说明书进行。分别将siduzap
‑
944、siduzap
‑
1576和siduzap
‑
2283以及阴性对照sinegative control按推荐的最佳浓度加入到试剂盒配套的转染buffer中,涡旋混匀后加入2倍sirna剂量的转染试剂,每个干扰分子做3个重复。4h后换2%维持液,36h后在无菌操作台中用trizol吹打收样,提取细胞中的总rna反转成cdna后,用荧光定量pcr技术检测细
胞中内源性 zap的表达情况,从而筛选出干扰效率最高的干扰分子。结果表明siduzap
‑
944 的干扰效率最高(见图5a)。
[0084]
2.3.2内源duzap的rna干扰试验
[0085]
def细胞中分别将干扰分子阴性对照nc和干扰效率最高的siduzap
‑
944与 pgl
‑
ifnb
‑
luc和内参prl
‑
tk共转染def细胞,24小时后用poly(i:c)或dtmuv 刺激,12小时后收集细胞,使用双荧光素酶报告基因检测试剂盒进行检测。转染试剂用jettransfection reagent,按照说明书进行。详细转染步骤参照步骤2.2.1,将鸭源ifn
‑
β启动子的荧光素酶报告质粒、内参海参荧光素酶tk质粒依次加入到jettransfection reagent试剂盒配套的转染buffer中,涡旋混匀后分别加入筛选出的干扰分子siduzap
‑
944和阴性对照sinegative control,按步骤用转染试剂将目的基因转染进def细胞,4h后换上含2%fbs的无双抗细胞维持液。每组3个重复。24h后用poly(i:c)刺激或感染dtmuv,过夜培养后收集用细胞裂解液收集样品,然后按照双荧光素酶报告基因检测试剂盒说明书进行检测。如图5b所示,结果表明内源性duzap的表达被降低后可以使poly(i:c)和dtmuv 刺激的ifn
‑
β表达显著降低。图5说明干扰def细胞内源性duzap的表达可以抑制ifn
‑
β的启动子活性。
[0086]
2.4过表达duzap对鸭坦布苏病毒(duck tembusu virus,dtmuv)复制的影响
[0087]
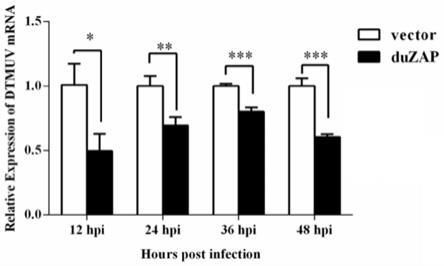
在def细胞中转染pzap
‑
flag 24后感染dtmuv,12h、24h、36h和48h后用trizol法收集细胞,提取细胞总rna进行dtmuv mrna水平的检测。表达 duzap的载体和空载体分别转染def细胞,转染24h后,感染dtmuv(moi=1),感染后12h、24h、36h、48h,用trizol法收集细胞,按实施例1中步骤1.1.1 提取细胞中总rna。然后按实施例1中步骤1.3.3进行duzap实时荧光定量pcr 检测细胞中的dtmuv表达。如图6所示为duzap过表达对鸭坦布苏病毒(ducktembusu virus,dtmuv)的复制影响图,与vector组(空载组)相比,超表达zap 12h后即可以抑制dtmuv复制,随着时间的增加,超表达24h、36h、48h抑制 dtmuv的复制效果越发显著,说明过表达duzap可以抑制dtmuv的复制。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1