水稻粒长基因GL10等位基因的应用
水稻粒长基因gl10等位基因的应用
技术领域
1.本发明属于植物基因育种应用技术领域,具体涉及水稻粒长基因gl10等位基因的应用。
背景技术:
2.水稻粒型不仅是影响水稻产量的要素,还与稻米的外观品质密切相关,是决定稻米品质的重要因素。随着水稻全基因组序列的公布、高通量测序的迅速发展、全基因组关联分析技术及生物信息学分析技术的成熟,越来越多的水稻粒型基因被相继定位和克隆。其中已克隆参与水稻粒型调控的基因就超过110个以上,但大部分基因来自突变体材料,无法在育种生产中利用。利用自然变异群体定位的主效粒型基因有gw2、gl2/osorf4、gs3、gl3.1、lgy3、gw5、tgw6、gw7/gl7、gw8和gs9等,这些基因的等位基因已在育种生产中广泛应用。目前对已克隆粒型基因参与粒型调控分子机制进行初步的探索,主要包括转录因子调控、mapk途径、泛素途径、g蛋白途径以及植物激素水平调控粒型等。
3.mads-box转录因子对植物生长发育起到至关重要的作用。这一家族不仅影响水稻花器官的形成,而且还影响种子的发育、开花时期、顶端分生组织分化、光周期反应和激素的调控等。其中,参与花器官形成的有osmads13、osmads3、osmads1、osmads5、osmads7、osmads8和osmads16;参与水稻种子发育的有osmads6、osmads29、osmads34、osmads87和osmads1。
4.定位克隆水稻粒型基因有助于丰富粒型调控分子机制网络,同时为水稻育种提供理论基础;自然变异的等位基因可有效在水稻分子设计育种中广泛利用,改良和培育新的水稻品种。这些研究对水稻基因功能研究和品种改良都具有重要的应用价值。
技术实现要素:
5.为了克服现有技术的缺点与不足,本发明的目的在于提供水稻粒长基因gl10等位基因的应用。本发明发现来自于lemont供体的等位基因缺失了gl10功能,导致水稻籽粒变短。进一步研究表明,该基因缺失会导致水稻籽粒变短,过量表达该基因会导致水稻籽粒变长。可以利用本发明接合自然变异等位基因对gl10基因进行定点改良水稻品种以提高水稻的产量。
6.本发明的目的通过下述技术方案实现:
7.本发明提供水稻粒长基因gl10等位基因的应用,所述等位基因包括等位基因gl10
hjx74
,其核苷酸序列如rap-db(https://rapdb.dna.affrc.go.jp/)数据库登录号为:os10g0536100的序列所示;和等位基因gl10
lemont
,其核苷酸序列如seq id no:3示;所述应用为如下应用中的任意一种或多种:
8.a.在调控水稻粒长中的应用;
9.b.在水稻粒形改良育种中的应用;
10.c.在培育转基因水稻中的应用。
11.gl10基因位于10号染色体上,基因座位号为os10g0536100(rap-db登录号),其全长基因组序列10448bp包括5
′
非翻译区(5
′
utr)、7个外显子、6个内含子和3
′
非翻译区(3
′
utr)(图2)。其cdna全长702bp(seq id no:1),编码234个氨基酸。gl10编码的蛋白质序列如seq id no:2所示。
12.所述的应用还包括与所述水稻粒长基因gl10等位基因相关的生物材料,所述生物材料为如下生物材料中的任意一种或多种组合:
13.1)含有所述水稻粒长基因gl10的核苷酸序列的表达盒;
14.2)含有所述水稻粒长基因gl10的核苷酸序列的重组载体,或含有1)中所述表达盒的过表达载体;
15.3)含有所述水稻粒长基因gl10的核苷酸序列的重组微生物,或含有1)中所述表达盒的重组微生物,或含有2)所述重组载体的重组微生物;
16.4)含有所述水稻粒长基因gl10的核苷酸序列的转基因植物组织、或含有1)中所述表达盒的转基因植物组织;
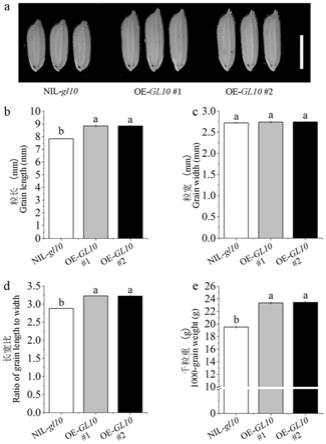
17.本发明还提供检测上述水稻粒长基因gl10等位基因的功能标记,所述功能标记包括包括如下引物对:
18.gl10-f:5'-gtccctctgatctctctttccccat-3';
19.gl10-r:5'-cggacctgctgaaaactgaaaaaat-3'。
20.所述功能标记是根据等位基因gl10
hjx74
第1外显子上的一个1019bp片段设计,是功能性分子标记;采用所述功能标记扩增水稻基因组dna时,如果有1019bp特征条带,则待测水稻含粒长等位基因gl10
hjx74
;如果不含1019bp特征条带,则待测水稻含粒长等位基因gl10
lemont
,能准确区分不同水稻资源的gl10等位基因型。
21.本发明还提供上述功能标记的应用,所述应用为如下应用中的任意一种或多种:
22.a)在鉴定水稻粒长基因gl10等位基因中的应用;
23.b)在辅助选择水稻粒形中的应用;
24.c)在水稻粒形改良育种中的应用。
25.本发明的机理是:利用来源于供体亲本lemont(莱蒙特)的单片段材料为基础,发现该片段包含有一个控制粒长的主效qtl(数量性状座位),经过与受体亲本hjx74(华粳籼74)多代回交和自交,发展出背景更纯的近等基因系nil-gl10(hjx74)和nil-gl10(lemont)。利用图位克隆的技术将目的基因定位于10号染色体长臂端上18.5kb区间内(图1)。该区间包含有一个候选基因loc_os10g39130,基因组序列分析发现nil-gl10材料中该基因第一外显子处有1019bp的片段缺失,导致该基因功能完全缺失(图2)。在hjx74背景下敲除gl10,将导致水稻籽粒粒长变短(图3);在nil-gl10背景下过量表达gl10,将导致水稻籽粒粒长明显增长(图4)。这些结果说明了水稻籽粒粒长与gl10表达量呈正相关。针对1019bp的片段缺失开发该基因的功能标记,该标记可直接鉴定水稻中gl10等位基因。利用该标记鉴定实验室21份材料,发现8类粳稻中缺失gl10功能位点,而8类籼稻和5类野生稻中gl10功能完整(图5)。将自然缺失1019bp的等位基因与携带已克隆粒形基因材料聚合,聚合系表型出明显的粒形变化(图6)。
26.本发明相对于现有技术具有如下的优点及效果:
27.本发明方法利用图位克隆的方法在自然变异群体中定位克隆一个新的粒长基因
gl10。该基因作为一个水稻粒长正调控因子,敲除gl10植株表现出粒型变短的特点,过量表达gl10植株表现出粒型明显增长。
28.本发明利用变异位点开发了一对鉴定gl10等位基因的功能标记,可有效的检测水稻中gl10基因等位变异,准确区分不同水稻资源的gl10等位基因型。
29.本发明有助于更好地了解gl10的作用机制,gl10的克隆为进一步了解水稻粒型调控网络和遗传解析奠定基础,gl10等位基因的利用将在水稻育种中具有重要的应用价值。
附图说明
30.图1是gl10基因图位克隆示意图;注:a.第10染色体上nil-gl10代换片段信息;b.利用重叠群作图将gl10定位在标记m15到np-11区间内;c.利用bc5f3群体的4000株单株筛选交换单株将gl10定位在标记nc9到nc14区间内;i-iv代表不同类型的交换单株,其中i类型的交换单株在m15和m22之间发生交换,ii类型交换单株在nc11和nc14之间发生交换,iii类型的交换单株在nc8和nc9之间发生交换,iv类型交换单株在nc9和nc10之间发生交换。
31.图2是gl10基因结构变异位点及表达谱示意图;注:a.在nil-gl10和nil-gl10两材料中gl10的基因结构;黑色方块表示基因外显子,白色方块表示utr,灰色方块表示缺失的序列,黑线表示内含子,∫∫表示为省略碱基数;b.gl10基因在nil-gl10材料中的根、茎、叶、叶鞘和幼穗不同长度相对表达量;c.rt-pcr分析gl10在nil-gl10和nil-gl10两材料中表达,actin作为内参基因;“root”表示根、“stem”表示茎、“leaf”表示叶、“1cm、2cm、3cm、4cm、5cm、6cm、8cm、10cm和15cm”分别表示相应长度的小穗。
32.图3是敲除gl10转基因粒型表型结果图;注:a.粒型表型图,标尺为0.5cm;b-e.粒型性状柱状图,分别为粒长、粒宽、长宽比和千粒重,上标不同的字母表示显著性差异p≤0.01。
33.图4是过量表达gl10转基因粒型表型结果图;注:a.粒型表型图,标尺为0.5cm;b-e.粒型性状柱状图,分别为粒长、粒宽、长宽比和千粒重,上标不同的字母表示显著性差异p≤0.01。
34.图5是gl10基因功能标记在不同水稻品种中鉴定结果示意图;注:泳道m2000为2kb dna marker;katy,nyz,irat 261,kyeema,ir65598,iapar9,tr66897b和ir66167为粳稻;tetep,amol3(sona),z4188,bg367,zihui 100,ir64,basmati385和hjx74为籼稻;hb(o.barthii),hg(o.glumaepatula),hm(o.meridionalis),hn(o.nivara)和hr(o.rufipogon)为野生稻。
35.图6是gl10等位基因与不同水稻材料聚合结果示意图;注:a.粒长表型图,标尺为0.5cm;b-g.不同聚合组合粒长表型柱状图,分别为nil-gs3/gl10、nil-gw8/gl10、nil-gl3.1/gl10、nil-gs5/gl10、nil-gw5/gl10和nil-gw7/gl10,上标不同的字母表示显著性差异p≤0.05。
具体实施方式
36.下面结合实施例及附图对本发明作进一步详细的描述,但本发明的实施方式不限于此。
37.本发明所使用的各种原料及各项设备均为常规市售产品,均能够通过市场购买直
接获得,所用引物序列均由上海捷瑞生物工程有限公司合成。
38.所述的来源于供体lemont的单片段材料由广东省植物分子育种重点实验室提供。
39.所述的基因编辑载体pc1300-cas9已在文献“increasing the efficiency of crispr-cas9-vqr precise genome editing in rice.plant biotechnology journal,2018,16(1):292-297”中公开。
40.所述的基因编辑载体pxq-35s载体已在文献“gw10,a member of p450 subfamily regulates grain size and grain number in rice”中公开。
41.实施例1水稻粒长gl10基因的图位克隆
42.1)gl10基因的初步定位
43.来源于供体lemont的单片段材料代换区间为psm344与rm147标记之间,该区间包含有一个控制粒长主效qtl(图1a)。在区间内开发indel多态标记,分别在rap-db(https://rapdb.dna.affrc.go.jp/)和hjx74 datebase(http://192.168.87.153/)网站上下载定位区间的dna序列;在bioedit软件上进行序列比对,选取相差5个碱基以上缺失的位置,在两端设计特异性引物,目的片段扩增的大小约为200bp左右;对设计的多态标记进行检查,分别跑两亲本材料和杂合型材料,电泳检测出有明显多态的标记,用于候选基因的定位。在该区间内找到了7对多态标记,分别为m4、m8、m11、m14、m15、m22和np-11。利用1200株bc5f2群体筛选该区间的交换单株;总共筛选到5类交换单株。l1株系在标记m11和m14间发生交换,粒长表型为nil-gl10型,说明基因在标记m11右侧;l2株系在标记m15和m22之间发生交换,粒长表型为nil-gl10型,说明基因在标记m15右侧;l3株系在标记m15和m22之间发生交换,粒长表型为nil-gl10型,说明基因在标记m15右侧;l4株系在标记psm344和m4之间发生交换,粒长表型为nil-gl10型,说明基因在标记psm344右侧;l5株系在标记m22和np-11之间发生交换,粒长表型为nil-gl10型,说明基因在标记np-11左侧;综合这些株系表型和基因型我们可以将gl10定位在标记m15到np-11之间,该区间大小为252kb(图1b和表1)。
44.表1不同类型交换株粒型表型
[0045][0046][0047]
注:l1、l2、l3、l4和l5代表不同交换单株发展株系;psm344、m4、m8、m11、m14、m15、m22和np-11代表的多态标记。“1”代表gl10基因型,“3”代表gl10基因型。数据为均值
±
标准误。
[0048]
所用引物及序列为:
[0049]
psm344-f:5'-ctcctaaccacctgcctaa-3';
[0050]
psm344-r:5'-atcccacagtttcctaccc-3';
[0051]
m4-f:5'-cggcgcttgatggtgattagc-3';
[0052]
m4-r:5'-agcgcgccgagaagaatagc-3';
[0053]
m8-f:5'-ctctctctccccacccaatc-3';
[0054]
m8-r:5'-gagggaggaggaggtgtagg-3';
[0055]
m11-f:5'-tggtgaagctagtaagacagatgc-3';
[0056]
m11-r:5'-ttgaccacgtgaaagctatgc-3';
[0057]
m14-f:5'-ctgggtagttgtttctcatgtgc-3';
[0058]
m14-r:5'-agtgctattgccattgtactgc-3';
[0059]
m15-f:5'-ctgaaggccgaattcatcatgg-3';
[0060]
m15-r:5'-cgtctctgtaacttccgatcaatcc-3';
[0061]
m22-f:5'-tcgtctctccgacgataaattcc-3';
[0062]
m22-r:5'-aaagcacagacaccctcttctcc-3';
[0063]
np-11-f:5'-atgcagcagtgtggtctctact-3';
[0064]
np-11-r:5'-ttctagctttcacgttgacccc-3'。
[0065]
2)gl10基因的精细定位
[0066]
基于对gl10初定位的结果,继续对m15与np-11两标记之间开发新的多态标记,开发出新的标记有nc2、nc5、nc8、nc9、nc10、nc11、nc14和nc19。利用m15与np-11标记间杂合型材料进行交换单株筛选,总共筛选了4000单株,总共筛选到11株交换单株。对这11株交换单株进行加密标记检测,最终选取4个类型交换进行精细定位,经考察交换株系的基因型和表型,最终把gl10确定在标记nc9到nc14之间的18.5kb区间内(图1c)。比较水稻粒长的表型发现,在18.5kb区间内基因型为gl10表现出长粒的特点,基因型为gl10表现出短粒的特点。
[0067]
所用引物及序列为:
[0068]
nc2-f:5'-gataaggtgggagcgatactgt-3';
[0069]
nc2-r:5'-tgtcacatccaactaaaacccc-3';
[0070]
nc5-f:5'-taagccgtcaccgtcaggagca-3';
[0071]
nc5-r:5'-ccactggaagttgacaagagat-3';
[0072]
nc8-f:5'-gattctctctcactgtcgctct-3';
[0073]
nc8-r:5'-tttcagcctgatagcaccgaca-3';
[0074]
nc9-f:5'-gctactcacagggaaaatagttggc-3';
[0075]
nc9-r:5'-cacaagagtggctcacatccgtttt-3';
[0076]
nc10-f:5'-gatctgcccaaacaacagtaca-3';
[0077]
nc10-r:5'-gcacacgataatctgatgaagt-3';
[0078]
nc11-f:5'-gacccatcaaccacaccattat-3';
[0079]
nc11-r:5'-gttgcgatgctgcagagatcaa-3';
[0080]
nc14-f:5'-tttcccaaaattggacgctccg-3';
[0081]
nc14-r:5'-gcgattcgtcgttgttagggtg-3';
[0082]
nc19-f:5'-ggaaaaatgttctacaacgccc-3';
[0083]
nc19-r:5'-tcgtgactgctttcccatttcc-3'。
[0084]
实施例2 gl10序列分析及表达模式分析
[0085]
1)gl10基因组序列分析
[0086]
根据rap-db(https://rapdb.dna.affrc.go.jp/)网站预测定位区间内仅有一个
候选基因,基因注释为loc_os10g39130(msu)。分别对nil-gl10和nil-gl10两材料的gl10候选基因进行基因组全长测序,该基因的基因组全长为10448bp,分为6段进行扩增和测序。测序结果发现该基因在两材料中存在84个变异位点,变异位点覆盖了启动子、外显子和内含子。其中,nil-gl10材料中gl10在启动子和第1外显子上的一个1019bp的片段缺失,导致基因功能的完全缺失(图2a)。
[0087]
所用基因组测序引物序列为:
[0088]
g56-1-f:5'-gtccctctgatctctctttccccat-3';
[0089]
g56-1-r:5'-cggacctgctgaaaactgaaaaaat-3';
[0090]
g56-2-f:5'-gagcgtgcgagactacaaatgg-3';
[0091]
g56-2-r:5'-tatgggtctatgggggtactatgcc-3';
[0092]
g56-3-f:5'-atcggtgtgctgtctttagtcctgt-3';
[0093]
g56-3-r:5'-ccaactcaggttcactcaaggtcaa-3';
[0094]
g56-4-f:5'-gcaataatccatcacaaaaccccta-3';
[0095]
g56-4-r:5'-gcagaacagacacaaggagcaaact-3';
[0096]
g56-5-f:5'-caacagccgcctattggcattgaaa-3';
[0097]
g56-5-r:5'-actcctcagaaagagacacgattgc-3';
[0098]
g56-6-f:5'-tcctagtatcctacactttagcacc-3';
[0099]
g56-6-r:5'-gcctaaagaaaaagacaaggacacg-3'。
[0100]
2)gl10基因表达模式分析
[0101]
a.总rna提取和第一链cdna的合成
[0102]
取水稻孕穗期根、茎、叶和不同长度的幼穗,液氮速冻保存于-80℃冰箱内。rna提取采用trizol试剂盒方法(invitrogen公司),提取的总rna用1%的琼脂糖电泳检测提取效果,紫外分光光度计测定rna浓度。反转录第一条cdna的合成,反转录按照abm公司5
×
all-in-one rt mastermix试剂盒说明进行。将2μg总rna样品加入0.2ml离心管内,加入2μl accurt reaction mix(4
×
),在用ddh2o补充至8μl;42℃孵育2min或者室温孵育5min;孵育完成后加入2μl accurt reaction stopper(5
×
),接着分别加入4μl 5
×
all-one rt mastermix和6μl ddh2o;25℃孵育10min,然后42℃孵育15min(qpcr)和50min(pcr),最后85℃孵育5min停止反应,保存-30℃冰箱内。
[0103]
b.gl10基因半定量和定量分析
[0104]
半定量反应体系:cdna模板50-100nmol,10μl 2
×
taq mix混合液,上游引物和下游引物(10μmol/l)各1μl,加水ddh2o至20μl。pcr扩增体系采用15μl的反应体系:体系中包括5.3μl mix混合液,2μl dna模板,1.2μl正反向引物,加水至15μl。pcr反应程序通常为:94℃预变性5min,循环(94℃30sec,55℃30sec,72℃45sec)38次,终延伸72℃5min。qpcr所用的反应程序参照hiefftm qpcr sybr green master mix说明书进行。反应体系:cdna模板0.5μl,hiefftm qpcr sybr green master mix 10μl,forward primer和reverse primer(10μm)各0.4μl,加水ddh2o至20μl。扩增程序:本实验全部使用两步法,95℃预变性5min;95℃变性10sec,60℃退火和延伸30sec,此过程循环40cycle;最后溶解曲线分别为95℃15sec、60℃60sec和95℃15sec。结果分析采用
‑△△
ct法。
[0105]
所用引物及序列为:
[0106]
qgl10-f:5'-gaccgctataaagcatacaca-3';
[0107]
qgl10-r:5'-tcatgtggttagccaccagc-3';
[0108]
actin-f:5'-gctatgtacgtcgccatccag-3';
[0109]
actin-r:5'-aatgagtaaccacgctccgtca-3'。
[0110]
结果如图2b和图2c所示,半定量分析发现nil-gl10材料中gl10基因不表达;对nil-gl10材料的各个组织进行gl10基因表达谱分析,定量结果表明gl10在水稻各个组织均高表达,在茎和叶中表达相对较高,在小穗中均表现出高表达的现象,其中在1-3cm的幼穗中表达较高。
[0111]
实施例3 gl10敲除载体和过量表达载体构建
[0112]
1)gl10敲除载体构建
[0113]
根据gl10基因组序列设计两对靶点引物,用水稀释至浓度100μmol/l,上游引物和下游引物各20μl混合在一起,100℃5分钟,室温冷却;sk-grna进行aari酶切(ferment公司),形成带有粘性末端的载体;利用t4连接酶将靶点引物连接上sk-grna,连接产物转化进入dh5α,长出单菌落进行菌落pcr,获得阳性菌株;pc1300-cas9载体用kpni和bamhi限制性内切酶进行酶切和回收,保存备用;将sk-grna阳性菌株抽提质粒,第一个靶点用kpni和nhei进行酶切回收,第二个靶点用xbai和bglii进行酶切回收;将酶切后两靶点到pc1300-cas9载体上,连接产物转化进入dh5α,长出单菌落进行菌落pcr,获得阳性菌株;抽取终载体质粒,测序靶点序列。所得重组质粒命名为pc1300-cas9::gl10,用于后续水稻遗传转化。
[0114]
所采用引物及序列为:
[0115]
ko-ms1-f:5'-ggcatgagaacccgacgagccggc-3';
[0116]
ko-ms1-r:5'-aaacgccggctcgtcgggttctca-3';
[0117]
ko-ms2-f:5'-ggcactccgtcctctgcgacgccg-3';
[0118]
ko-ms2-r:5'-aaaccggcgtcgcagaggacggag-3'。
[0119]
2)gl10过量表达载体构建
[0120]
首先,以华粳籼74的cdna为模板进行gl10基因编码区全长的扩增,并在5’和3’端分别加入spei和miui限制性内切酶位点,扩增产物进行纯化,酶切和回收;接着将pxq-35s载体进行酶切,使用的酶为spei和miui限制性内切酶,酶切后回收;将片段和载体酶切后的回收产物进行t4连接酶连接;最后将连接产物转化进入dh5α,长出单菌落进行菌落pcr,获得阳性菌株,测序成功后抽取质粒。所得重组质粒命名为pxq-35s::gl10,用于后续水稻遗传转化。
[0121]
所采用引物及序列为:
[0122]
oe-ms-f:5'-ggggtaccatggtgcgggggaggacggag-3';
[0123]
(下划线标记部分为限制性内切酶spei酶切位点序列);
[0124]
oe-ms-r:5'-cgggatcctcaacctgtctccgaccggtt-3';
[0125]
(下划线标记部分为限制性内切酶miui酶切位点序列)。
[0126]
3)水稻遗传转化
[0127]
1.水稻愈伤组织的诱导和培养:将成熟的水稻种子去壳,去壳后种子在70%酒精中消毒3min,转入4.5%次氯酸钠溶液浸泡45min,用无菌水冲洗干净次氯酸钠溶液,把种子均匀的播撒在n6d培养基上诱导愈伤组织,20天左右长出愈伤组织,继代到nb培养基上,每
两周继代一次,每次继代选取色泽较好的愈伤组织;
[0128]
2.农杆菌的培养:将构建好的质粒载体转化到农杆菌eha105菌株中,涂布于含50mg/l卡那霉素和25mg/l利福平的lb培养基上,28℃培养2d,挑取单菌落进行菌落pcr,鉴定出阳性的菌株。将阳性菌株在转移到lb液体培养基中,28℃,200rpm培养过夜,取500μl接入50ml的aam液体共培养基悬浮菌体至od600为0.5-0.6,用于水稻材料转化;
[0129]
3.水稻愈伤组织与农杆菌的共培养:将预培养4d的水稻愈伤组织转移到100ml锥形瓶中,倒入步骤2中的重悬液,室温静置20min,轻轻的取出愈伤组织,在无菌滤纸上吸取剩余的菌液,随即转移到浸泡有aam的无菌滤纸上,25℃黑暗培养2-3d;
[0130]
4.抗性愈伤组织筛选及植株再生:共培养后,将愈伤组织转移至含有50mg/l卡那霉素和潮霉素的选择的培养上进行筛选培养;第一轮筛选完成后,转移到第二轮选择培养基上继续筛选2周,然后将生长旺盛的抗性愈伤组织转移到分化培养基上,再生的小苗在1/2ms上生根壮苗,长至10cm移出温室;
[0131]
5.阳性植株鉴定:提取转基因水稻的总dna,以dna为模板,根据载体带的标签基因和我们目的基因上设计特异引物进行pcr扩增,根据扩增条带和测序共同来判断转基因的阳性植株,设置正负对照。
[0132]
实施例4 gl10功能标记开发和鉴定
[0133]
1)gl10功能标记开发
[0134]
根据nil-gl10和nil-gl10的基因序列差异设计功能标记,前文中已介绍在nil-gl10材料中该基因缺失了1019bp碱基,由此在缺失区域设计一对功能标记。标记序列为:
[0135]
gl10-f:5'-gtccctctgatctctctttccccat-3';
[0136]
gl10-r:5'-cggacctgctgaaaactgaaaaaat-3';
[0137]
2)gl10功能标记鉴定
[0138]
tps法用于水稻材料dna的提取。取2-3cm的水稻叶片装入2ml离心管,加入1ml tps抽提液和钢珠;在磨样机中研磨1min,吸铁石取出钢珠;75℃水浴30min,12000rpm离心10min,吸取700μl上清液转移至1.5ml离心管中;加入等体积预冷的无水乙醇,混合均匀后放入-30℃冰箱内1h;12000rpm离心10min,去上清,放入烘箱烘干酒精,加入200μl灭菌水,4℃长期保存。鉴定了21份水稻材料,结果示意图见图5。21份材料中发现8类粳稻中缺失gl10功能位点,而8类籼稻和5类野生稻中gl10功能完整。
[0139]
实施例5 gl10等位基因聚合应用
[0140]
室内杂交。1.母本的取样:分别选取nil-gs3、nil-gw8、nil-gl3.1、nil-gs5、nil-gw5和nil-gw7材料(上述材料均来源于染色体单片段代换系(single-segment substitution line,sssl)文库,详细资料如表2所示,该文库已s在文献“张桂权.基于sssl文库的水稻设计育种平台[j].遗传,2019,41(8).”中公开。)作为母本,将需杂交的母本小穗放入42℃的温水中浸泡8min,杀死花粉。2.促开花和剪颖:将去雄的小穗包裹塑料袋,加入co2气体促进开花;将开颖的小花剪去1/3的颖壳,2-3枝小穗包装上授粉袋。3.授粉:去田间剪取父本nil-gl10小穗,co2气体促进开花,将开花的父本小穗至于母本之上,抖动小穗使花粉均匀的落入母本柱头上,封好授粉袋,记录材料编号。4.获得的真杂种于田间,发展至f2代,利用案例一的多态标记进行基因型鉴定,获得纯合的聚合系材料用于粒形表型考察(每份聚合材料考察10个单株),结果示意图见图6。
[0141]
表2携带粒形基因的单片段代换系材料
[0142][0143]
注:材料由广东省植物分子育种重点实验室提供。
[0144]
上述实施例为本发明较佳的实施方式,但本发明的实施方式并不受上述实施例的限制,其他的任何未背离本发明的精神实质与原理下所作的改变、修饰、替代、组合、简化,均应为等效的置换方式,都包含在本发明的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1