酵母重组人源III型三螺旋胶原蛋白及其制备方法与流程
酵母重组人源iii型三螺旋胶原蛋白及其制备方法
技术领域
1.本发明涉及基因重组技术领域,尤其涉及一种酵母重组人源iii型三螺旋胶原蛋白的制备方法以及采用该制备方法得到的酵母重组人源iii型三螺旋胶原蛋白。
背景技术:
2.胶原蛋白是人和动物体内含量最多的蛋白,占体内蛋白总量的25-30%,约占体重的6%。胶原蛋白是皮肤、骨骼、牙齿、角膜、肌腱、韧带以及各种组织器官中存在的重要的结构蛋白,起支撑和保护机体组织的功能。胶原蛋白是细胞外基质的主要组分,其生物学活性主要表现在促进细胞粘附、生长分化、细胞信号通路的应答、血小板凝集、细胞和组织器官功能的维护和损伤修复等功能。胶原蛋白在空间结构上呈现出三股螺旋缠绕的结构,由三条独立的胶原α肽链主要以氢键的形式缠绕形成螺旋结构。在体内胶原蛋白以三螺旋的构象与组织和细胞结合以发挥其生物学效应,因此胶原蛋白的生物学活性很大程度上依赖其天然的三螺旋构象。胶原蛋白优良的生物学活性使其在生物医学材料,医疗美容等领域拥有巨大的应用价值和潜力。
3.目前市场上主要的胶原蛋白产品来源于动物组织提取,这种方法在提取的过程中破坏了天然的三螺旋结构,形成一种交叉螺旋结构,影响了胶原生物学活性的发挥。而且使用动物来源提取工艺得到的胶原蛋白其肽段分子量不均一,可加工性较差。由于动物胶原蛋白与人源胶原蛋白在一级序列上存在差异,因此可能会在临床上造成潜在的免疫原性,安全性较差。而且动物组织受其来源的影响需要解决动物病毒的安全隐患才能进入市场。在大肠杆菌中表达的人源胶原蛋白,由于大肠杆菌缺乏糖基化和羟基化等蛋白翻译后修饰所必须的酶类,无法形成天然的三螺旋高级结构所以不具有高的生物学活性;由于胶原蛋白在大肠杆菌系统内无法糖基化,因此会导致潜在的免疫原性;由于大肠杆菌系统自身存在内毒素和致热源,所以存在潜在的安全隐患。随着市场对胶原蛋白产品安全性、有效性要求的提高,传统的从动物组织中提取胶原蛋白的方式已经无法满足现有市场的需求。而且,以胶原蛋白作为生物材料应用于医疗场景的产品需求量也在日益增大,需要通过别的的途径来解决目前市场上存在的问题。通过基因工程等分子生物学技术来生产具有天然三螺旋结构的人源胶原蛋白在解决以上所述问题方面具有极大的潜力。
4.以具有天然胶原蛋白高级结构且更安全可靠的酵母三螺旋胶原蛋白取代动物源胶原蛋白和大肠杆菌表达的单链胶原蛋白,为下游的胶原蛋白产品如美容美妆产品,医用止血修复的填料,人工合成眼角膜等提供且更为优质安全的材料。动物源的胶原蛋白存在潜在的动物病毒隐患(如疯牛病、口蹄疫),免疫原性,质量不稳定和供应不足等问题。另外,由于宗教和习俗等原因,有些地区不能使用来自动物组织中的胶原蛋白制品。大肠杆菌表达表达的胶原蛋白无法进行蛋白翻译后修饰,对于一些具有高级构象的重组人胶原蛋白,利用大肠杆菌表达后无法糖基化。同时,由于大肠杆菌是原核生物,没有内质网(er),因此也没有能帮助胶原蛋白质折叠成高级构象的分子伴侣蛋白及一些修饰合成蛋白如羟基化、形成二硫键的酶。因此大肠杆菌系统很难表达具有天然高级结构的三螺旋胶原蛋白。
5.酵母发酵产生的人源重组胶原蛋白克服了动物源以及大肠杆菌表达的胶原蛋白的缺陷。酵母表达系统具备分子伴侣以及蛋白翻译后修饰的酶(如糖基化、羟基化、乙酰基化酶等),非常适合于具有高级结构的三螺旋胶原蛋白的形成。并且酵母重组表达系统具有安全性好、质量稳定、分子量确定、无致热源、纯化简单、重现性好等优点。酵母细胞的培养条件相对于哺乳动物细胞较低,非常适宜于大规模的工业化发酵生产。酵母系统取代动物源胶原和大肠杆菌原核表达系统源重组胶原在生物医学应用方面有重要研究价值。酵母表达的人源胶原蛋白通过基因工程的设计和改造能够具有形成天然胶原蛋白三螺旋构象的能力。三螺旋构象更接近与胶原蛋白天然的生物学结构,具有更优异的生物学活性(血小板凝聚、细胞、组织、器官的正常生理功能的维护、调节及损伤修复)、理化性质和可加工性,是更高级的生物材料。因此,研发和设计酵母重组人源三螺旋胶原蛋白作为现有胶原产品的替代和补充具有巨大的市场潜力与前景,而在国内市场,尚无获批的酵母重组三螺旋胶原蛋白生物材料。
技术实现要素:
6.本发明旨在至少解决现有技术中存在的技术问题之一。
7.为此,本发明提出一种酵母重组人源iii型三螺旋胶原蛋白及其制备方法,可以有效避免免疫原性的产生,并且提高了胶原分子间的相互作用力。
8.根据本发明第一方面实施例的酵母重组人源iii型三螺旋胶原蛋白的制备方法,包括以下步骤:
9.s1、构建含有人源iii型胶原alpha 1链的ppicz alpha b质粒;
10.s2、构建人源p4h alpha亚基和beta亚基的双启动子表达载体;
11.s3、将所述双启动子表达载体和所述ppicz alpha b质粒共转化至酵母菌中,得到共表达菌株;
12.s4、从所述共表达菌株中筛选出含有p4h alpha亚基和beta亚基的酵母重组菌株;
13.s5、对所述酵母重组菌株进行诱导表达,得到酵母重组人源iii型三螺旋胶原蛋白。
14.根据本发明实施例的酵母重组人源iii型三螺旋胶原蛋白的制备方法,利用高密度发酵技术和柱层析、膜分离等组合分离纯化技术生产,不存在致热原问题;通过具有独特生物化学性能的多糖与重组人源胶原蛋白复合,有效的避免了免疫原性的产生;通过与脯氨酸羟基化酶在酵母细胞内进行共表达,在胶原蛋白分子序列中的脯氨酸位点进行羟基化,有效的提高了胶原分子间的相互作用力,热稳定性和抗酶解水平;形成的天然三螺旋构象能够实现细胞粘附、迁移、增殖、代谢、细胞骨架与ecm的连接建立等细胞内、外多种信号传导与应答途径等多种生物学功能,满足下游产品对生物学活性的最大化的需求。
15.根据本发明的一个实施例,步骤s1包括:
16.s11、合成人源iii型胶原alpha 1链dna片段;
17.s12、设计含有strep和his亲和标签的引物,对人源iii型胶原alpha 1链dna片段进行扩增,得到两端含有ecori酶切位点的扩增片段;
18.s13、使用ecori限制性内切酶对所述扩增片段和ppicz alpha b质粒进行酶切;
19.s14、使用连接酶对所述扩增片段和所述ppicz alpha b质粒进行连接反应,并将
反应物转化至大肠杆菌中;
20.s15、对转化后的大肠杆菌进行培养,筛选出含有将人源iii型胶原alpha 1链克隆至ppicz alpha b质粒的大肠杆菌。
21.根据本发明的一个实施例,步骤s14为:使用t4连接酶对所述扩增片段和ppicz alpha b质粒在16℃下进行过夜连接反应,并将反应物采用cacl2热激法转化至感受态大肠杆菌dh5α中。
22.根据本发明的一个实施例,步骤s15为:将转化后的大肠杆菌涂布至含有博莱霉素的lb平板上培养24h,对得到的大肠杆菌菌落进行菌落pcr,以筛选含有将人源iii型胶原alpha 1链克隆至ppicz alpha b质粒的大肠杆菌菌落。
23.根据本发明的一个实施例,步骤s2包括:
24.s21、合成人源p4h alpha亚基和beta亚基的dna片段;
25.s22、设计引物,对人源p4h alpha亚基和beta亚基的dna片段进行扩增,得到5’端含bamhⅰ、3’端含ecorⅰ酶切位点的p4h alpha亚基扩增片段和5’端含xhoⅰ、3’端含xbaⅰ酶切位点的p4h beta亚基扩增片段;
26.s23、使用限制性内切酶对p4h alpha亚基扩增片段和ppic9k质粒进行酶切,对p4h beta亚基扩增片段和ppicz alpha b质粒进行酶切;
27.s24、使用连接酶对p4h alpha亚基扩增片段和ppic9k质粒进行连接反应,p4h beta亚基扩增片段和ppicz alpha b质粒进行连接反应,并分别将反应物转化至大肠杆菌中;
28.s25、对转化后的大肠杆菌分别进行培养,筛选出含有将p4h alpha亚基克隆至ppic9k质粒的大肠杆菌以及将p4h beta亚基克隆至ppicz alpha b质粒的大肠杆菌;
29.s26、设计引物对p4h beta亚基及启动子和终止子进行片段扩增,得到与ppic9k质粒经ataⅱ线性化后两端分别互补的片段;
30.s27、使用限制性内切酶对含有p4h alpha亚基扩增片段和ppic9k的质粒进行酶切;
31.s28、对步骤s26的扩增片段和含有p4h alpha亚基扩增片段和ppic9k的质粒进行连接反应,并将反应物转化至大肠杆菌中;
32.s29、对转化后的大肠杆菌进行培养,筛选含有人源p4h alpha亚基和beta亚基的双启动子表达载体。
33.根据本发明的一个实施例,步骤s3包括:
34.s31、将含有p4h alpha亚基扩增片段和ppic9k的质粒使用sali限制性内切酶进行线性化,电击转化至酵母菌中;
35.s432、对步骤s31的酵母菌进行培养,得到p4h 9k dp;
36.s33、将所述ppicz alpha b质粒使用限制性内切酶进行线性化,电击转化至含有p4h 9k dp的酵母菌中;
37.s34、对步骤s33的酵母菌进行培养,得到所述共表达菌株。
38.根据本发明的一个实施例,所述酵母菌为毕赤酵母smd1168菌株。
39.根据本发明的一个实施例,步骤s4包括:将构建好的的人源脯氨酸羟化酶载体转化至所述共表达菌株,筛选出含有p4h alpha亚基及beta亚基的酵母重组菌株。
40.根据本发明的一个实施例,步骤s5包括:将proα1(iii)胶原蛋白表达载体电转化至含有羟化酶的所述酵母重组菌株,通过转化子诱导表达,得到酵母重组人源iii型三螺旋胶原蛋白。
41.根据本发明第二方面实施例的酵母重组人源iii型三螺旋胶原蛋白,所述酵母重组人源iii型三螺旋胶原蛋白采用上述酵母重组人源iii型三螺旋胶原蛋白的制备方法制备而成。
附图说明
42.图1为根据本发明实施例中iii型胶原alpha 1链的dna序列图;
43.图2为根据本发明实施例中iii型胶原alpha 1链的蛋白序列图,其中,蛋白序列长度为1466aa,分子量为138kda;
44.图3为根据本发明实施例中p4h酶α亚基的dna序列图;
45.图4为根据本发明实施例中p4h酶α亚基的蛋白序列图,其中,蛋白序列长度为534aa;
46.图5为根据本发明实施例中p4h酶β亚基的dna序列图;
47.图6为根据本发明实施例中p4h酶β亚基的蛋白序列图,其中,蛋白序列长度为491aa;
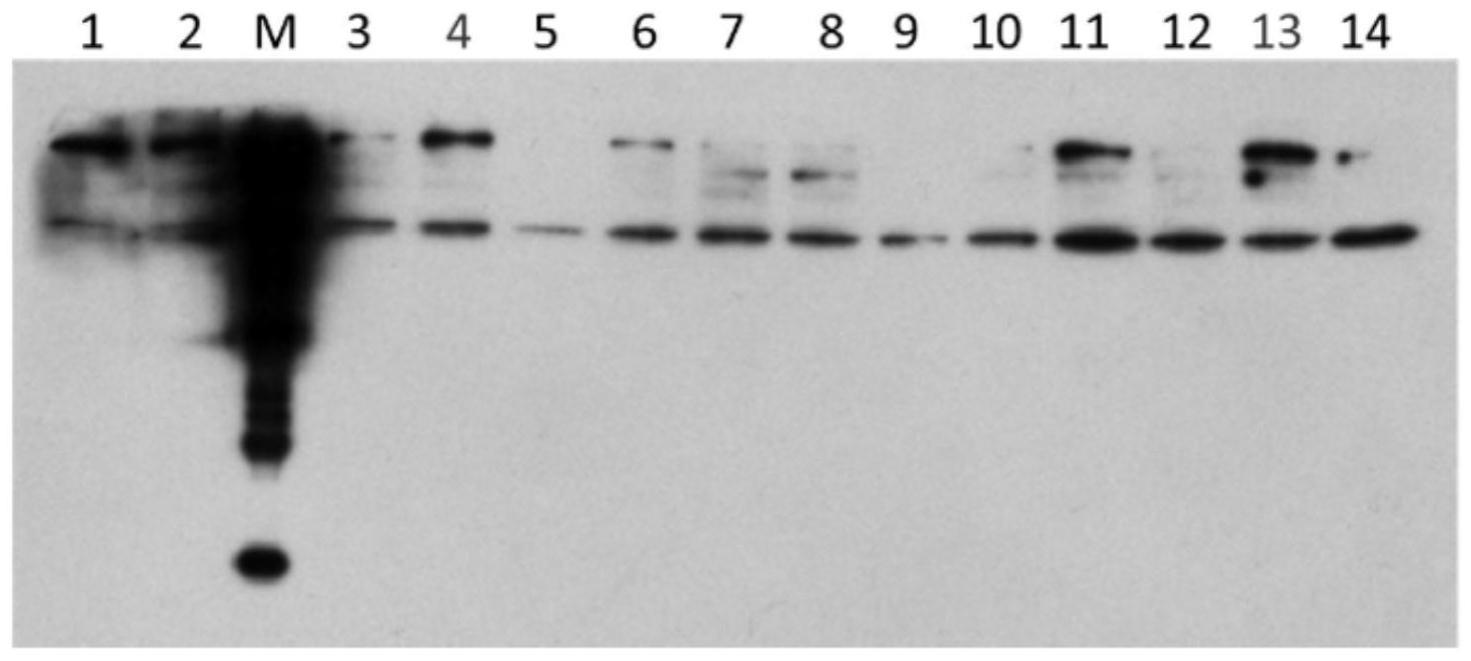
48.图7为根据本发明实施例中3a1 ab/p4h 9k dp免疫印迹法表达验证示意图,其中,1-14分别表示3a1 ab/p4h 9k dp菌株1-14;
49.图8为根据本发明实施例中3a1 ab/p4h 9k dp免疫印迹法表达验证示意图,其中,3和14分别表示3a1 ab/p4h 9k dp阳性菌株4和13;
50.图9为根据本发明实施例中3a1 ab/p4h 9k dp蛋白非变性page电泳检测示意图,其中,3a1表示非变性条件下3a1 ab/p4h 9k dp蛋白样品;
51.图10为根据本发明实施例中经胃蛋白酶处理后3a1三螺旋胶原蛋白sds-page检测示意图,其中,3a1 ab/p4h 9k dp经过胃蛋白酶降解后的3a1ab/p4h 9k dp抗降解片段为图中黄色线框所示。
具体实施方式
52.下面详细描述本发明的实施例,所述实施例的示例在附图中示出,其中自始至终相同或类似的标号表示相同或类似的元件或具有相同或类似功能的元件。下面通过参考附图描述的实施例是示例性的,仅用于解释本发明,而不能理解为对本发明的限制。
53.在本发明的描述中,需要理解的是,术语“中心”、“纵向”、“横向”、“长度”、“宽度”、“厚度”、“上”、“下”、“前”、“后”、“左”、“右”、“竖直”、“水平”、“顶”、“底”“内”、“外”、“顺时针”、“逆时针”、“轴向”、“径向”、“周向”等指示的方位或位置关系为基于附图所示的方位或位置关系,仅是为了便于描述本发明和简化描述,而不是指示或暗示所指的装置或元件必须具有特定的方位、以特定的方位构造和操作,因此不能理解为对本发明的限制。此外,限定有“第一”、“第二”的特征可以明示或者隐含地包括一个或者更多个该特征。在本发明的描述中,除非另有说明,“多个”的含义是两个或两个以上。
54.在本发明的描述中,需要说明的是,除非另有明确的规定和限定,术语“安装”、“相
连”、“连接”应做广义理解,例如,可以是固定连接,也可以是可拆卸连接,或一体地连接;可以是机械连接,也可以是电连接;可以是直接相连,也可以通过中间媒介间接相连,可以是两个元件内部的连通。对于本领域的普通技术人员而言,可以具体情况理解上述术语在本发明中的具体含义。
55.下面首先结合附图具体描述根据本发明实施例的酵母重组人源iii型三螺旋胶原蛋白的制备方法。
56.根据本发明实施例的酵母重组人源iii型三螺旋胶原蛋白的制备方法,包括以下步骤:
57.s1、构建含有人源iii型胶原alpha 1链的ppicz alpha b质粒;
58.s2、构建人源p4h alpha亚基和beta亚基的双启动子表达载体;
59.s3、将所述双启动子表达载体和所述ppicz alpha b质粒共转化至酵母菌中,得到共表达菌株;
60.s4、从所述共表达菌株中筛选出含有p4h alpha亚基和beta亚基的酵母重组菌株;
61.s5、对所述酵母重组菌株进行诱导表达,得到酵母重组人源iii型三螺旋胶原蛋白。
62.由此,根据本发明实施例的酵母重组人源iii型三螺旋胶原蛋白的制备方法,利用高密度发酵技术和柱层析、膜分离等组合分离纯化技术生产,不存在致热原问题;通过具有独特生物化学性能的多糖与重组人源胶原蛋白复合,有效的避免了免疫原性的产生;通过与脯氨酸羟基化酶在酵母细胞内进行共表达,在胶原蛋白分子序列中的脯氨酸位点进行羟基化,有效的提高了胶原分子间的相互作用力,热稳定性和抗酶解水平;形成的天然三螺旋构象能够实现细胞粘附、迁移、增殖、代谢、细胞骨架与ecm的连接建立等细胞内、外多种信号传导与应答途径等多种生物学功能,满足下游产品对生物学活性的最大化的需求。
63.可选地,在本技术的一些具体实施方式中,步骤s1包括:
64.s11、合成人源iii型胶原alpha 1链dna片段;
65.s12、设计含有strep和his亲和标签的引物,对人源iii型胶原alpha 1链dna片段进行扩增,得到两端含有ecori酶切位点的扩增片段;
66.s13、使用ecori限制性内切酶对所述扩增片段和ppicz alpha b质粒进行酶切;
67.s14、使用连接酶对所述扩增片段和所述ppicz alpha b质粒进行连接反应,并将反应物转化至大肠杆菌中;
68.s15、对转化后的大肠杆菌进行培养,筛选出含有将人源iii型胶原alpha 1链克隆至ppicz alpha b质粒的大肠杆菌。
69.其中,步骤s14为:使用t4连接酶对所述扩增片段和ppicz alpha b质粒在16℃下进行过夜连接反应,并将反应物采用cacl2热激法转化至感受态大肠杆菌dh5α中。
70.进一步地,步骤s15为:将转化后的大肠杆菌涂布至含有博莱霉素的lb平板上培养24h,对得到的大肠杆菌菌落进行菌落pcr,以筛选含有将人源iii型胶原alpha 1链克隆至ppicz alpha b质粒的大肠杆菌菌落。
71.换句话说,本技术构建含有人源iii型胶原alpha 1链的ppicz alpha b质粒(命名为3a1 ab质粒)的步骤如下:
72.人工合成一段4398bp大小的人源iii型胶原alpha 1链(命名为3a1)dna片段(如图
1所示),其蛋白质序列如图2所示。
73.设计含有strep和his亲和标签的引物:
74.3a1 forward cggaattctggagccacccgcagttcgaaaagcaatacgactcttatgacgtgaag;和
75.3a1 reverse atagtttagcggccgcttagtggtgatggtgatgatgcaaaaagcaaacaggtccaaca)。
76.对3a1片段进行扩增,得到两端含有ecori酶切位点的扩增片段。使用ecori限制性内切酶对扩增片段和ppicz alpha b质粒进行酶切,对酶切后的dna进行回收并纯化。使用t4连接酶对扩增片段和ppicz alpha b质粒在16度下进行过夜连接反应,并将样品采用cacl2热激法转化至感受态大肠杆菌dh5α中。将转化后的大肠杆菌涂布至含有博莱霉素的低盐lb平板上培养24小时,对得到的大肠杆菌菌落进行菌落pcr,以筛选含有将3a1克隆至ppicz alpha b质粒(命名为3a1 ab质粒)的大肠杆菌菌落。将阳性克隆菌落进行dna测序表明其序列为图1所示3a1序列。
77.根据本技术的一些实施例,步骤s2包括:
78.s21、合成人源p4h alpha亚基和beta亚基的dna片段;
79.s22、设计引物,对人源p4h alpha亚基和beta亚基的dna片段进行扩增,得到5’端含bamhⅰ、3’端含ecorⅰ酶切位点的p4h alpha亚基扩增片段和5’端含xhoⅰ、3’端含xbaⅰ酶切位点的p4h beta亚基扩增片段;
80.s23、使用限制性内切酶对p4h alpha亚基扩增片段和ppic9k质粒进行酶切,对p4h beta亚基扩增片段和ppicz alpha b质粒进行酶切;
81.s24、使用连接酶对p4h alpha亚基扩增片段和ppic9k质粒进行连接反应,p4h beta亚基扩增片段和ppicz alpha b质粒进行连接反应,并分别将反应物转化至大肠杆菌中;
82.s25、对转化后的大肠杆菌分别进行培养,筛选出含有将p4h alpha亚基克隆至ppic9k质粒的大肠杆菌以及将p4h beta亚基克隆至ppicz alpha b质粒的大肠杆菌;
83.s26、设计引物对p4h beta亚基及启动子和终止子进行片段扩增,得到与ppic9k质粒经ataⅱ线性化后两端分别互补的片段;
84.s27、使用限制性内切酶对含有p4h alpha亚基扩增片段和ppic9k的质粒进行酶切;
85.s28、对步骤s26的扩增片段和含有p4h alpha亚基扩增片段和ppic9k的质粒进行连接反应,并将反应物转化至大肠杆菌中;
86.s29、对转化后的大肠杆菌进行培养,筛选含有人源p4h alpha亚基和beta亚基的双启动子表达载体。
87.具体地,构建人源p4h alpha亚基(命名为p4ha)和beta亚基(命名为pdi)的双启动子(dp)表达载体的方法如下:
88.人工合成一段1633bp大小的人源p4h酶α亚基(命名为p4ha)dna片段(图3所示),其蛋白质序列如图4所示。设计引物(p4ha forward agatatcgggtccccgggat和p4ha reverse cgtgccccggaattcctatta)对p4ha片段进行扩增,得到5’端含bamhⅰ、3’端含ecorⅰ酶切位点的扩增片段。使用限制性内切酶bamhⅰ和ecorⅰ对扩增片段和ppic9k质粒进行酶切,对酶切
后的dna进行回收并纯化。使用t4连接酶对扩增片段和ppic9k质粒在16度下进行过夜连接反应,并将样品采用cacl2热激法转化至感受态大肠杆菌dh5α中。将转化后的大肠杆菌涂布至含有氨苄西林的lb平板上培养24小时,对得到的大肠杆菌菌落进行菌落pcr,以筛选含有将p4ha克隆至ppic9k质粒(p4ha 9k)的大肠杆菌菌落。菌落pcr阳性的菌株提取质粒经bamhⅰ和ecorⅰ酶切鉴定并进行dna测序表明其序列为p4ha序列。
89.接着,人工合成一段1496bp大小的人源p4h酶β亚基(命名为pdi)dna片段(图5所示),其蛋白质序列如图6所示。设计引物(pdi forward ccgctcgaggttgctattgaaaaaagagatgctccagaagaagaagatca和pdi reverse gctctagacctattattacaattcatctttaac)对pdi片段进行扩增,得到5’端含xhoⅰ、3’端含xbaⅰ酶切位点的扩增片段。使用限制性内切酶xhoⅰ和xbaⅰ对扩增片段和ppicz alpha b质粒进行酶切,对酶切后的dna进行回收并纯化。使用t4连接酶对扩增片段和ppicz alpha b质粒在16度下进行过夜连接反应,并将样品采用cacl2热激法转化至感受态大肠杆菌dh5α中。将转化后的大肠杆菌涂布至含有博莱霉素的lb平板上培养24小时,对得到的大肠杆菌菌落进行菌落pcr,以筛选含有将pdi克隆至ppicz alpha b质粒(pdi ab)的大肠杆菌菌落。菌落pcr阳性的菌株提质粒经xhoⅰ和xbaⅰ酶切鉴定,并进行dna测序表明其序列为pdi序列。
90.最后,设计引物
91.(forward ccgaaaagtgccacctgacgtagatctaacatccaaaga和reverseaggttaatgtcatgataataatggtttcttagacgtgtctcacttaatcttctgtactc)对pdi及启动子和终止子片段进行扩增,得到与ppic9k经ataⅱ线性化后两端分别互补的片段。使用限制性内切酶ataⅱ对质粒p4ha 9k进行酶切,对pcr产物和线性化质粒进行回收并纯化。使用即用型无缝克隆试剂盒对扩增片段和p4ha 9k质粒在50度下连接30min,并将样品采用cacl2热激法转化至感受态大肠杆菌dh5α中。将转化后的大肠杆菌涂布至含有氨苄西林的lb平板上培养24小时,对得到的大肠杆菌菌落进行菌落pcr,以筛选含有将pdi及相关元件克隆至p4ha 9k质粒(p4h 9k)的大肠杆菌菌落。菌落pcr阳性的菌株提质粒经bamhⅰ和bspei酶切鉴定,质粒进行dna测序表明其序列为pdi及启动子和终止子序列。将所得到的具有双启动子的p4h质粒命名为p4h 9kdp。
92.根据本技术的一些实施例,步骤s3包括:
93.s31、将含有p4h alpha亚基扩增片段和ppic9k的质粒使用sali限制性内切酶进行线性化,电击转化至酵母菌中;
94.s432、对步骤s31的酵母菌进行培养,得到p4h 9k dp;
95.s33、将所述ppicz alpha b质粒使用限制性内切酶进行线性化,电击转化至含有p4h 9k dp的酵母菌中;
96.s34、对步骤s33的酵母菌进行培养,得到所述共表达菌株。
97.可选地,所述酵母菌为毕赤酵母smd1168菌株。
98.具体地,将p4h 9k质粒使用sali限制性内切酶进行线性化,电击转化至酵母菌smd1168中,将转化后的酵母菌涂布在含有氨苄青霉素的yepd固体培养基(蛋白胨20克,酵母粉10克,葡萄糖20克,琼脂粉15克,溶于1l蒸馏水后灭菌)平板上。将平板放置在30度恒温孵育箱内培养72小时,得到长有酵母菌菌落的平板。将得到的阳性克隆命名为p4h 9k dp。
99.将3a1 ab质粒使用pmei限制性内切酶进行线性化,电击转化至含有将p4h 9k dp
的酵母菌smd1168中,将转化后的酵母菌涂布在含有博来霉素的ypd固体培养基平板上。将平板放置在30度恒温孵育箱内培养72小时,得到长有酵母菌菌落的平板。将得到的阳性克隆命名为3a1 ab/p4h 9k dp。
100.可选地,步骤s4包括:
101.将构建好的的人源脯氨酸羟化酶载体转化至所述共表达菌株,筛选出含有p4h alpha亚基及beta亚基的酵母重组菌株。
102.进一步地,步骤s5包括:
103.将proα1(iii)胶原蛋白表达载体电转化至含有羟化酶的所述酵母重组菌株,通过转化子诱导表达,得到酵母重组人源iii型三螺旋胶原蛋白。
104.下面对酵母重组人源iii型三螺旋胶原蛋白的表达和验证过程进行详细说明。
105.具体地,可以采用如下几种方式:
106.(1)3a1 ab/p4h 9k dp的摇瓶诱导表达和免疫印迹法验证。
107.在平板上挑选14个酵母菌菌落进行摇瓶诱导表达,接种至含有博来霉素的20ml bmgy液体培养基中,30度200rpm震荡培养24小时,之后加入含有0.6%甲醇的bmmy液体培养基中进行36小时的诱导表达。离心收集菌体后,加入玻璃珠和酵母破壁液高速震荡进行破壁,将酵母裂解液13500rpm转速下离心15分钟,收集上清,进行western blotting表达验证。图7为使用strep抗体检测菌落1-14的蛋白表达,结果表明菌落4和13具有相对较高的表达量。将菌落4和13使用his抗体、p4h beta抗体和p4h alpha抗体检测(图8),表明菌落4和13含有strep和his亲和标签,并且同时有p4h alpha和beta亚基的表达。得到的3a1蛋白序列为图8所示,得到的3a1胶原蛋白的预计分子量为138kda,实际表达后得到的分子量大于150kda,推测是由于蛋白翻译后修饰的作用使得3a1胶原蛋白的某些氨基酸位点发生了糖基化和羟基化。
108.(2)3a1 ab/p4h dp非变性电泳验证。
109.1a1胶原蛋白菌株4经过摇瓶表达后,使用桨质匀浆机进行破壁处理,破壁后的样品13500rpm转速下离心20分钟,取上清进行非变性电泳验证其在天然状态下的结构和分子量大小。如图9所示,3a1 ab/p4h dp在天然状态下以三聚体的形式存在,分子量大于480kda。
110.(3)3a1胶原蛋白的胃蛋白酶酶切验证
111.三螺旋胶原蛋白的形成导致其具有抗胃蛋白酶降解的特点,因此也可以被用于验证胶原蛋白三螺旋结构的形成。使用0.1m盐酸作为裂解液在高压均质机中对1a1胶原蛋白进行破壁,13500rpm离心20分钟后得到上清液。在上清液中加入0.2mg/ml的胃蛋白酶在37度恒温孵育箱内反应1小时,之后加入100mm naoh溶液终止胃蛋白酶酶切反应。将处理后的样品用超速离心管进行浓缩40倍后得到的3a1胶原蛋白经过还原性sds-page凝胶电泳验证。如图10所示,经过胃蛋白酶处理后,40kda处的条带为胃蛋白酶本身,3a1三螺旋胶原蛋白c端前肽被胃蛋白酶被降解,产生分子量大小为约120kda的抗胃蛋白酶降解的片段。
112.由此,通过上述方法可以制备得到酵母重组人源iii型三螺旋胶原蛋白。
113.总而言之,本技术以开发基因重组三螺旋胶原蛋白技术应用于生物材料和美容美妆领域,对领域内现有的技术进行替代和补充,实现产业化革新和技术工艺路线的提升。对于解决应用于生物材料和医疗美容领域的材料存在的生物活性水平较低,潜在安全性隐患
等问题具有十分重要的现实意义。
114.本技术采用基于人iii型胶原蛋白α1链gly-x-y(甘氨酸-x-y)的三肽重复序列结构特征,利用蛋白质工程、分子生物学及基因重组原理,通过dna设计、密码子优化、基因合成人源胶原蛋白表达载体proα1(iii)和脯氨酸羟基化酶p4h alpha及beta亚基表达载体。首先优化人源脯氨酸羟基化酶p4hα亚基和β亚基的碱基序列,并将两个基因分别克隆到ppic9k载体的两个不同位点,每个位点各带有一个独立的启动子。将构建好的的脯氨酸羟化酶载体转化至毕赤酵母smd1168,筛选出含有p4halpha及beta亚基的毕赤酵母重组菌株。优化人源iii型胶原alpha 1链的碱基序列,并克隆至ppicz alpha b载体,对载体进行酶切线性化后转导入含有p4hα亚基和β亚基的毕赤酵母smd1168菌株,得到共表达菌株。对共表达菌株诱导蛋白表达,收菌后破碎细胞提取胞内蛋白,利用sds-page分析确定iii型胶原蛋白分子量大小,得到目的蛋白大小为180kda,经胃蛋白酶水解后的抗降解片段大小为120kda。
115.本技术利用高密度发酵技术和柱层析、膜分离等组合分离纯化技术生产,不存在致热原问题,安全性高。本技术利用具有独特生物化学性能的多糖与重组人源胶原蛋白复合,有效的避免了免疫原性的产生。在胶原蛋白分子序列中的脯氨酸位点进行羟基化,有效的提高了胶原分子间的相互作用力,热稳定性和抗酶解水平。形成的天然三螺旋构象能够实现细胞粘附、迁移、增殖、代谢、细胞骨架与ecm的连接建立等细胞内、外多种信号传导与应答途径等多种生物学功能,满足下游产品对生物学活性的最大化的需求。
116.以上所述是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明所述原理的前提下,还可以作出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1