白三叶钙调素类似蛋白TrCML6基因及在抗旱中的应用
白三叶钙调素类似蛋白trcml6基因及在抗旱中的应用
技术领域
1.本发明属于基因工程领域,具体涉及一种白三叶钙调素类似蛋白trcml6基因序列,所述基因序列编码的氨基酸序列,以及所述基因序列在培育转基因抗旱植株中的应用。
背景技术:
2.干旱是最重要的环境胁迫之一,可以对植物造成严重而不可逆的伤害,进而影响植物生长发育以及作物产量。植物吸收的水分绝大多数经由保卫细胞组成的气孔以蒸腾作用散失出去,干旱胁迫可以诱导植物产生脱落酸,脱落酸可以减小气孔开度,降低水分散失。因此,发掘参与调控气孔运动的功能基因及机理分析,对改良植物的抗干旱能力具有重要意义。
3.ca
2+
是植物细胞在发育和应对外界刺激过程中最广泛的第二信使,并调节多种生理过程,如基因表达,离子平衡,以及碳水化合物,脂质和蛋白质代谢。其中,钙感受器在ca
2+
信号转导途径中占据重要的位置。钙调素类似蛋白(cmls)是近年新发现的一类植物细胞所特有的ca
2+
响应蛋白家族。目前研究表明,cmls在植物生长发育和抵抗逆境等方面发挥着重要的作用,具有很高的研究价值。
4.白三叶(trifolium repens l.)是一种多年生冷季型豆科牧草,也是温带地区建坪的主要草种。但是,其根系较短,调控蒸腾能力低,缺水时容易发病,在各种非生物和生物因素的干扰下,白三叶品质和外观表现被严重影响。因此,越来越多的学者开始关注于白三叶抗逆方面的研究,对逆境相关基因的挖掘和功能解析,也逐步成为该领域研究的一个重点。但是,目前对于白三叶抗逆基因的挖掘多数停留在表达模式分析阶段,仅少数基因转入模式植物或白三叶当中进行更深入的验证。白三叶中逆境相关的代谢途径及受哪些基因控制仍然是不清楚的,白三叶中关于cml基因功能的研究未见报道。
技术实现要素:
5.本发明通过生理性状实验和遗传分子学操作,证明白三叶钙调素类似蛋白trcml6基因编码具有钙离子结合活性的蛋白,并且可能作为ca
2+
信号响应蛋白调控气孔运动及植物干旱过程。本发明所述的白三叶钙调素类似蛋白trcml6基因,是在前期实验的基础上通过壳聚糖外施转录组数据分析筛选得到的可能具有提高白三叶耐旱性的功能基因。
6.本发明的一个目的是提供一种白三叶钙调素类似蛋白trcml6基因序列,以及白三叶钙调素类似蛋白trcml6基因编码的氨基酸序列;本发明的另一个目的是提供一种白三叶钙调素类似蛋白trcml6基因序列在提高植物抗干旱性能中的应用。
7.本发明的目的通过以下技术方案实现:
8.第一方面,本发明提供一种白三叶钙调素类似蛋白trcml6基因序列,其特征在于,所述基因序列为下述1)-3)中的任一种:
9.1)如seq id no.1所示的核苷酸序列;
10.2)具有与seq id no.1所示的核苷酸序列90%以上同源性,且编码相同功能蛋白
的核苷酸序列;
11.3)编码与seq id no.2所示氨基酸序列组成的蛋白有相同功能蛋白的核苷酸序列。
12.seq id no.1所示的核苷酸序列具体为:
13.atgggataaacacaaaaacacagcacatttcacatttcctcaaaacaaaacaaaccaaacaacatcatgtgtccttctggcagaaccctccgtccacaacctcccacaaccgatttccgaccggcattcgacattctcgacaccgattgcgacggcaaaataagccgagacgatctccgttcattctacgcaaccaccagcggcgaaggcgtctctgccgacgcaatcggtgccatgatgtcggttgcggacacaaacatggacggatttgtggaatacgaggaattcgagcgtgttgttagtggaaacaatgaaaagaaaccgttaggatgtggagccatggaagatgtgttcaaggtgatggatagagatggtgatggtaaacttagtcatgaagatttgaagaattatatgaattgggctggttttgctgcaacagatgaagagataaatgctatgattaagcttggtggtggtgatcaaaacggtggcgttagcttcgatggtttgattcgtatattagctcttgatcatttcgtccctgtttattgattcattaa attaatgatgatatattattatct
14.seq id no.2所示氨基酸序列具体为:
15.met cys pro ser gly arg thr leu arg pro gln pro pro thr thr asp phe arg pro ala phe asp ile leu asp thr asp cys asp gly lys ile ser arg asp asp leu arg ser phe tyr ala thr thr ser gly glu gly val ser ala asp ala ile gly ala met met ser val ala asp thr asn met asp gly phe val glu tyr glu glu phe glu arg val val ser gly asn asn glu lys lys pro leu gly cys gly ala met glu asp val phe lys val met asp arg asp gly asp gly lys leu ser his glu asp leu lys asn tyr met asn trp ala gly phe ala ala thr asp glu glu ile asn ala met ile lys leu gly gly gly asp gln asn gly gly val ser phe asp gly leu ile arg ile leu ala leu asp his phe val pro val tyr
16.第二方面,本发明提供一种白三叶钙调素类似蛋白trcml6基因编码的蛋白质,其特征在于,所述蛋白质为下述1)-2)中的任一种:
17.1)如seq id no.2所示氨基酸序列组成的蛋白质;
18.2)seq id no.2氨基酸序列经过1个或两个以上氨基酸残基取代和/或缺失和/或添加,且与seq id no.2氨基酸序列组成蛋白具有相同功能的蛋白质。
19.第三方面,本发明提供一种如上所述的白三叶钙调素类似蛋白trcml6基因序列在培育转基因抗旱植物中的应用。
20.所述植物包括拟南芥、白三叶。
21.第四方面,本发明提供一种转基因抗旱植株的培育方法,其特征在于,利用基因工程技术,将白三叶钙调素类似蛋白trcml6基因在植株中进行过表达,得到转基因抗旱植株。
22.所述基因工程技术可采用本领域中已记载的导入基因的技术手段,如利用农杆菌介导的花序浸染法、重组质粒、重组菌、转基因细胞系或表达盒等技术对导入基因实现过表达。
23.在本发明的具体实施方式中,所述基因工程技术为农杆菌介导的花序浸染法。
24.优选的,所述培育方法包括如下操作:
25.(1)设计特异性引物对,以白三叶品种“拉丁诺”为材料,克隆出编码钙调素类似蛋白trcml6的基因序列,并推导其编码的氨基酸序列。
26.(2)通过实时荧光定量pcr验证编码钙调素类似蛋白trcml6基因的转录本在干旱、h2o2、aba和cacl2处理下的表达模式;
27.(3)将钙调素类似蛋白trcml6的基因序列通过农杆菌介导的花序浸染法过表达到植株中,得到转基因抗旱植株。
28.具体的,所述步骤(1)包括:
29.(a)以白三叶叶片总rna进行反转录反应,获取cdna,设计简并引物,进行rt-pcr反应,获得核苷酸片段;
30.(b)进行5’race和3’race的pcr反应,获得核苷酸片段;
31.(c)将rt-pcr与race反应所得核苷酸片段进行基因全长序列拼接,获得如seq id no.1所示的核苷酸序列;
32.(d)预测开放阅读框,推导所述基因编码的氨基酸序列如seq id no.2所示。
33.优选的,所述简并引物对为:
34.trcml6 1f(seq id no.3):5
’‑
cgtccacaacctcccacaa-3’35.trcml6 1r(seq id no.4):5
’‑
cagcaaaaccagcccaat-3’,
36.所述race反应特异性引物对为:
37.cml-5’gsp(seq id no.5):5
’‑
ccgcaaccgacatcatggcacc-3’38.cml-3’gsp(seq id no.6):5
’‑
gcaaccaccagcggcgaaggcgt-3’。
39.所述步骤(3)包括:
40.(a)将钙调素类似蛋白trcml6基因重组到pbi121载体骨架上,筛选、鉴定,获得植物过表达重组载体;
41.(b)采用冻融法转化植物过表达重组载体至农杆菌,制备转染用菌液;
42.(c)采用花序侵染法将植物过表达重组载体转化至植株。
43.由以上技术方案可知,本发明通过设计特异性引物,以白三叶品种“拉丁诺”为材料,克隆出一个编码钙调素类似蛋白trcml6的基因,并推导其编码的氨基酸序列。本发明提供的白三叶钙调素类似蛋白trcml6基因具有潜在的抗干旱以及过氧化胁迫的能力。
44.通过实时荧光定量pcr验证trcml6转录本在干旱、h2o2、aba和cacl2处理下的表达模式,结果表明该编码序列在上述四种处理下,根与叶中的表达量均发生了显著的变化,trcml6基因的转录水平均有明显升高。
45.进一步的,将白三叶钙调素类似蛋白trcml6基因过表达至拟南芥中,转基因植株在干旱胁迫下表现出明显的抗旱性,且水分亏缺情况下具有更高的生物量和更长的根系,对培育抗干旱胁迫能力的植株具有重要意义。
附图说明
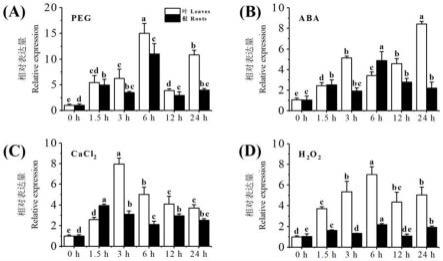
46.图1所示为干旱、h2o2、aba和cacl2四种处理下白三叶trcml6定量表达分析;其中,纵坐标为相对表达量,横坐标为处理时间(0、1.5、3、6、12、24h),每组柱形中左侧为叶片中的相对表达量,右侧为根系中的相对表达量。
47.图2所示为过表达拟南芥植株鉴定(a)及trcml6基因表达量检测(b)。
48.图3所示为过表达trcml6拟南芥幼苗和野生型在甘露醇胁迫下的表型差异。a为150mmol/甘露醇平板胁迫下拟南芥胁迫表型;b为150mmol/l甘露醇平板胁迫幼苗黄化率;c
为100mmol/l甘露醇胁迫下生长9d的拟南芥幼苗根系;d为幼苗每日自然根长。
49.图4所示为过表达trcml6拟南芥和野生型在12%peg胁迫下的表型差异。a为12%peg胁迫下拟南芥胁迫表型;b为植株地上部鲜重;c为植株地上部干重。
50.图5所示为过表达trcml6拟南芥和野生型在自然干旱胁迫下的表型差异。a为自然干旱胁迫下拟南芥表型;b为叶片相对含水量;c为离体叶片失水率。
具体实施方式
51.本发明公开了一种白三叶钙调素类似蛋白trcml6及其编码序列和应用,本领域技术人员可以借鉴本文内容,适当改进工艺参数实现。特别需要指出的是,所有类似的替换和改动对本领域技术人员来说是显而易见的,它们都被视为包括在本发明。
52.本发明所述白三叶钙调素类似蛋白trcml6及其编码序列和应用已经通过实施例进行了描述,相关人员明显能在不脱离本发明内容、精神和范围内对本文所述白三叶钙调素类似蛋白trcml6及其编码序列和应用进行改动或适当变更与组合,来实现和应用本发明技术。
53.在本发明的对比试验中,如未特别说明,各组除去人为设置的区别外(比如转入本发明所述白三叶钙调素类似蛋白trcml6的编码序列或不转入所述白三叶钙调素类似蛋白trcml6的编码序列的区别),其他试验条件均保持一致。以下就本发明所提供的一种白三叶转录因子trcml6及其编码序列和应用做进一步说明。
54.一、白三叶转录因子trcml6的基因克隆
55.1、方法
56.利用植物总rna提取试剂盒(rnaprep pure plant kit)提取培养30d的
‘
拉丁诺’白三叶叶片总rna,采用1%普通琼脂糖凝胶电泳检测rna完整性,采用nanodrop仪器测定od260/od280比值。参照反转录试剂盒(primeiscripttmⅱ1st strand cdna synthesis kit)说明书对所获得的完整、无污染的rna进行反转录反应,获取cdna。
57.利用已测转录组中白三叶trcml6的mrna序列,于genebank数据库上进行blast比对,获得同源序列。采用primer premier 5.0设计简并引物trcml6 1f(seq id no.3):5
’‑
cgtccacaacctcccacaa-3’和trcml61r(seq id no.4):5
’‑
cagcaaaaccagcccaat-3’,使用primestar max dna polymerase作为pcr反应用dna聚合酶进行pcr反应,反应体系如下:
[0058][0059]
反应程序如下:
[0060][0061]
pcr反应完成后,产物用1%的琼脂糖凝胶电泳(电泳条件:1
×
tae电泳缓冲液;120v,30min)检测。将含有单一目的条带的电泳产物于紫外灯下切胶,采用普通琼脂糖凝胶dna回收试剂盒纯化后,依据dna a-tailing kit试剂盒在纯化dna产物末端加a尾。参照pmdtm19-t vector cloning kit试剂盒将加a尾产物连接至pmdtm19-t载体上,并通过热激法转化至dh5α大肠杆菌感受态。已转化感受态于37℃、200rpm摇床活化1h后,放于离心机中室温、4000rpm离心30s,去掉上清液后,将剩余溶液均匀涂布于含50mg/l氨苄青霉素(amp)的lb固体抗性培养基中,并倒置过夜培养。用灭菌后的牙签挑取培养基上的单克隆菌株放入lb/amp液体抗性培养基中,约12-20个重复,采用菌液pcr筛选阳性单克隆并送至生工生物公司测序。
[0062]
参照race试剂盒(race 5’/3’kit user manual)说明书,以提取的白三叶总rna为模板,分别合成5’race和3’race所需cdna,利用测序所得部分序列设计race特异性引物cml-5’gsp(seq id no.5):5
’‑
ccgcaaccgacatcatggcacc-3’和cml-3’gsp(seq id no.6):5
’‑
gcaaccaccagcggcgaaggcgt-3’。进行5’race和3’race的pcr反应,反应体系如下:
[0063][0064]
反应程序如下:
[0065]
[0066][0067][0068][0069]
扩增产物经琼脂糖凝胶电泳检测后,采用试剂盒回收纯化,连接到pmdtm19-t载体上,转化至大肠杆菌感受态,挑取单克隆菌样进行菌液pcr检测,将含正确条带的菌液送至公司测序。运用dnaman软件将rt-pcr与race反应所得核酸片段进行基因全长序列拼接,利用orf finder在线软件预测trcml6开放阅读框(open reading frame,orf)。根据orf序列设计trcml6特异性引物trcml6 of(seq id no.7):5
’‑
atgtgtccttctggcagaaccctcc-3’和trcml6 or(seq id no.8):5
’‑
tcaataaacagggacgaaatgatca-3’进行orf验证。
[0070]
2、结果
[0071]
利用dnaman软件将扩增得到的片段进行拼接,得到序列全长共584bp(seq id no.1),利用orf finder预测开放阅读框,发现该基因开放阅读框长486bp,共编码161个氨基酸(seq id no.2)。再利用ncbi blast在线比对该基因与其他植物中同一基因的亲缘关系,通过mega软件采用neighbor-joining法构建trcml6与其他植物的同源钙调类似蛋白的系统发育进化树。根据系统进化树可知,白三叶trcml6与红三叶钙调类似蛋白6(tpcml6)的亲缘关系最近,聚于一个小的进化分枝,且其序列相似度达96.27%,因此将该蛋白命名为trcml6。
[0072]
将克隆得到的trcml6基因序列全长比对至白三叶基因组(
‘
crau
’‑
衍生生态型)中,发现trcml6在8o和8p染色体上各有一个拷贝,由于8p染色体基因组比对结果中存在n值较多的区域,因此将克隆所得序列与8p染色体中序列进行了比对,发现该基因无内含子序列,位于8p染色体的41306055bp-41306540bp区域。利用protparam预测可知,trcml6所编码蛋白分子式为c761h1174n206o250s11,相对分子质量(mw)为17561.58,理论等电点为4.35;含正电荷氨基酸残基(arg+lys)15个,负电荷氨基酸残基(asp+glu)29个;蛋白的不稳定系数为35.50;平均疏水系数为-0.366,介于0.5和-0.5之间。使用swiss-model软件模拟trcml6的空间结构,发现该蛋白卷曲折叠成4个螺旋-环-螺旋的ef-手单元,且ef-手单元之间有较长的柔性连接区域。
[0073]
二、trcml6基因的表达模式分析
[0074]
1、方法
[0075]
采用全营养液培养30d的“拉丁诺”白三叶为试验材料,分别用15%peg6000、100μmol/l aba(脱落酸)、10mmol/l h2o2(过氧化氢)、5mmol/lcacl2(氯化钙)对30d的白三叶幼苗全株进行处理,处理时间为0、1.5、3、6、12和24h。每处理每个时间点处理15株,处理相应时间后,随机分成3份,每份5株分别对根系和叶片进行取样编号。取样后立即用液氮冷冻,提取各处理下叶片和根系的总rna用于cdna的合成,rna保存在-80℃冰箱。
[0076]
以实验室筛选的tractin101为内参,设计特异性内参引物tractin101-f和tractin101-r(表1),根据已得trcml6基因全长序列设计特异性q-pcr引物cml-qpcr-f和cml-qpcr-r(表1),参照荧光定量试剂盒进行试验。参照荧光定量试剂盒(sybr premix ex taq ii)说明书配制反应体系,采用两步法进行扩增,扩增程序如下:95℃5min;95℃30s,58℃30s,共计40个循环。基因转录水平的采用2-δδct
的计算方法。
[0077]
表1实时荧光定量pcr引物序列
[0078][0079]
2、结果
[0080]
如图1所示:peg、aba、cacl2和h2o2处理下,trcml6的相对表达量整体呈上调趋势,且叶片表达量的变化幅度较根部明显。
[0081]
peg处理下,0-6h期间,叶片中trcml6相对表达量逐渐升高,6h表达量为对照的15倍,12h表达量有所下降,之后呈现升高的趋势;而在根系中,trcml6的相对表达量呈现多次升高后再降低的趋势,且在处理6h表达量达到顶峰,上调约11倍。
[0082]
外源aba处理可以诱导trcml6的表达,叶片trcml6相对表达量在处理后3h及24h为两个高峰,表达量分别为对照的5.1倍和8.4倍;根系中,相对表达量在6h达到高峰,而后降低,但表达量高于对照水平。
[0083]
ca
2+
能迅速诱导trcml6的表达,cacl2处理后叶片和根系的相对表达量峰值分别在3h和1.5h处,较0h上调了7.9倍和3.9倍;两时间点之后,trcml6的表达有所降低,但仍显著高于对照水平。
[0084]
h2o2处理下,叶片中trcml6的表达量先呈现逐渐升高的趋势,于处理6h达到最高值,上升约7倍,后随着处理的进行缓慢降低,24h的表达量为对照5倍;根系中trcml6相对表达量随着处理时间延长,先缓慢上升再恢复到对照水平再下降,相对表达量的峰值为对照2.1倍,表达量变幅较小,但均高于对照。以上结果表明trcml6在白三叶响应peg、aba、cacl2和h2o2处理中发挥重要作用。
[0085]
三、trcml6转拟南芥的验证
[0086]
1、方法
[0087]
(1)拟南芥转化及阳性植株鉴定
[0088]
依据pbi121载体序列,设计5’端分别引入xba i和sac i酶切位点和同源臂碱基的引物:pbi121-trcml-f(seq id no.13)和pbi121-trcml-r(seq id no.14)。以“拉丁诺”白三叶cdna为模板,用高保真酶对trcml6进行pcr扩增。xba i和sac i对pbi121载体进行双酶切,而后回收酶切片段。用无缝重组酶将trcml6的pcr片段与线性化pbi121载体片段进行连接,连接产物转化dh5α大肠杆菌感受态细胞后,将感受态细胞涂布于含50mg/l kan的lb固体抗性平板上,37℃过夜培养,挑取菌落pcr验证重组载体后,并进行测序验证,获得的植物过表达重组载体记为pbi121-trcml6。
[0089]
pbi121-trcml-f:
[0090]
agaacacgggggactctagaatgtgtccttctggcagaaccc
[0091]
pbi121-trcml-r:
[0092]
ggggaaattcgagctctcaataaacagggacgaaatga
[0093]
将测序验证后的菌落进行扩大培养,提取pbi121-trcml6过表达重组载体。采用冻融法转化重组质粒至农杆菌eha105感受态细胞中,已转化感受态细胞于28℃、200rpm震荡活化培养4h,放于离心机5000rpm离心1min收取菌液,轻轻吹打重悬菌块并且涂布于含50mg/l kan、10mg/l利福平(rif)抗生素的yeb固体培养基上,倒置放于28℃培养箱培养2-3天。挑取单菌落,摇菌扩繁后,采用菌液pcr检测阳性克隆。
[0094]
采用花序侵染法将载体转化至野生型拟南芥(col-0)。拟南芥幼苗进入生殖生长阶段后,剪去顶芽以去除顶端优势,一周后进行转化。将活化后的农杆菌以1:50比例转接至200ml yeb/kan,rif液体抗性培养基中,28℃、200rpm摇菌约36h至菌液od600=0.8~1.2。取菌液于室温、5000rpm离心15min,弃上清后将菌体重悬于5%蔗糖溶液(称取25g蔗糖于ddh2o定容至500ml,加入100μl silwet混匀)中。转化前将已经开花授粉的花和种子去除干净,将拟南芥花序浸入重悬液体中20s左右,采用黑色塑料薄膜覆盖整个侵染植株并扎数个小孔保障空气流通,于黑暗条件下培养2d后转至光照条件(光照/黑暗16h/8h)下培养,待拟南芥结种后收取t0代转基因种子。
[0095]
选取饱满的转基因t0代种子用含有0.2%的triton x-100浸泡10min,种子依次在75%酒精、0.1%naclo中消毒后,于ddh2o中润洗多次,置于含有50mg/l kan的1/2ms固体培养基上4℃暗培养2天,筛选后的种子在14h光照/10h黑暗光周期、21℃下培养。两周后选取长势正常、生长良好的阳性植株移栽至装有营养土的盆钵中,待其成熟后收种;阴性植株不萌发或萌发后不久死亡。采用上述培养方法将拟南芥植株筛选至t3代,t3代幼苗于营养土中培养20d后,提取阳性苗叶片dna,并以此为模板进行pcr验证,将电泳后与目的基因长度一致的条带切胶回收后,送至生工生物公司测序,测序结果与trcml核酸序列比对一致的认定为阳性植株。
[0096]
(2)过表达拟南芥干旱胁迫处理方法
[0097]
培养基培养:过表达t3代拟南芥与野生型拟南芥在不含抗生素的1/2ms培养基中生长5d后,使用镊子选取长势一致的幼苗移栽至含150mmol/l甘露醇的圆形培养皿中,每一株系移栽16株幼苗至同一板培养基中,重复3次,待两周后拍照并测定幼苗黄化率。同时,将
在无抗培养基中生长5d的幼苗放于含100mmol/l甘露醇的方形培养皿中,每一株系移栽4株幼苗至同一盘培养基中,且幼苗整齐排列于同一高度直线上,重复4次。将方形培养皿垂直放于拟南芥培养箱中,每日统计主根根系伸长情况。
[0098]
土壤培养:取边长为5cm的黑色正方形塑料盆钵,每盆称取相同重量的混合营养土,在托盘中倒满水,使土壤吸水饱和。分别选取长势正常且相近的转基因植株和野生型植株移栽至盆钵中,幼苗在土壤中生长20d后进行干旱胁迫。采用12%peg溶液(配制方法:100g peg溶解于1l 1/4hoagland营养液中)进行模拟干旱胁迫。将12%peg倒入装有塑料盆钵的黑色大托盘中至盆钵1/3处,摇晃均匀,每2d将托盘中剩余液体完全倒出,并倒入新的处理液。peg处理7d后测定单株鲜、干重。此外,自然干旱胁迫,以浇水至土壤饱和后为第0d,每日更换盆钵位置保证盆钵上方空气流通水平一致。于自然干旱18d后,取地上部样品测定相对含水量。
[0099]
(3)生理指标测定方法
[0100]
植株鲜、干重测定:将正常及胁迫后的拟南芥地上莲座叶小心剪下,称取每个株系地上部的鲜重,每个处理重复8次。称取完成后,将莲座叶装入牛皮纸袋中,放入烘箱,经105℃下杀青30min后,调至75℃烘干至恒重,称其干重。
[0101]
离体叶片失水率测定:剪取土壤中正常培养20d后的拟南芥植株倒数第三轮叶片,测定立即测定其鲜重,每株拟南芥取一片叶片。随后将叶片置于23℃、60%湿度环境自然风干,分别在取样后0.5、1、1.5、2.5、4、6、9h时间点测定叶片风干后重量,每个株系重复10-12次。叶片失水率=[(鲜重-风干后重量)/鲜重]
×
100%。
[0102]
叶片相对含水量测定:剪取相同部位叶片鲜重约0.2g(fw),用普通吸水纸将其包裹好,将叶片浸入装满水的50ml离心管中,于避光处静置24h使叶片组织充分吸水达到饱和。取出叶片,用吸水纸迅速吸去表面水分,称量饱和鲜重(tw)。将叶片置于鼓风烘箱中105℃下杀青45min,然后将温度调至75℃烘至恒重,称其干重(dw),重复4次,取平均值。相对含水量(%)=(鲜重fw-干重dw)/(饱和鲜重tw-干重dw)
×
100%。
[0103]
2、结果
[0104]
(1)过表达trcml6拟南芥鉴定
[0105]
对转基因t3代拟南芥植株提取总dna,以其为模板经pcr和琼脂糖凝胶电泳检测后,共有4株拟南芥的电泳条带与白三叶trcml6开放阅读框长度一致(图2a),表明这4株为转基因拟南芥阳性株系,分别编号为oe1-oe4。以atβ-actin为内参基因,进行实时荧光定量pcr来检测trcml6的相对表达量。结果表明:过表达trcml6转基因株系的trcml6表达量较高,其中,oe1与oe2株系的表达量相对较高,oe4株系的表达量稍低,而野生型拟南芥中trcml6不表达(图2b)。后续试验选择表达水平较高且表达量有差异的oe1与oe2株系来分析trcml6抗旱功能。
[0106]
(2)甘露醇模拟干旱胁迫
[0107]
如图3所示:150mmol/l的甘露醇胁迫中过表达株系幼苗与野生型之间的生长表型差异最明显,野生型植株叶片变黄、卷曲、萎焉、呈水渍状,而过表达株系叶片生长较为良好,仅部分植株出现了卷曲、轻微变黄的表现,其中oe2株系较oe1株系表现更优。同时,oe1与oe2株系的黄化干枯植株相对野生型植株更少。进一步统计植株的黄化率(以拟南芥幼苗有6片叶子变黄,认定为黄化植株),oe1株系与oe2株系植株的黄化率分别较野生型降低
38%与44%。结果表明,trcml6过表达可以增强拟南芥幼苗在甘露醇模拟干旱处理下的抗性。转移至100mmol/l甘露醇胁迫培养基中生长9d的oe1与oe1幼苗主根根系生长已接近方形平板最底端,而野生型幼苗主根系则相对较短。对1-9d的每日幼苗根长统计后发现:移苗1-3d后,幼苗根系生长差异不明显;但随着胁迫时间的增加,oe1与oe2株系的根系伸长量逐渐高于野生型拟南芥。综上结果表明,过表达trcml6可提高植株在甘露醇胁迫下的抗性,促进拟南芥根系的伸长,以获取水分维持生长发育。
[0108]
(3)peg模拟干旱胁迫
[0109]
如图4所示:peg胁迫后,野生型拟南芥叶片逐渐枯萎、由叶尖向内卷曲、叶尖变黄或整体叶片失绿,而过表达trcml6拟南芥仅有少数叶片叶尖变黄卷曲,叶片失绿情况不明显,生长受抑制程度更小(图4a)。进一步测定了正常及peg处理后植株的鲜重和干重(图4b-c),正常生长情况下,oe1、oe2与野生型拟南芥的鲜、干重差异不显著。peg处理后,过表达植株与野生型的植株鲜、干重均出现大幅度降低的现象,表明胁迫后植株正常的生长代谢受到影响,造成生物量的损失。但是,peg处理下,oe1与oe2的株系鲜重分别比野生型高出1.8和1.6倍;过表达株系的干重也显著高于野生型。结果表明,trcml6过表达可以增强拟南芥成苗在peg模拟干旱胁迫下的抗性,且在peg胁迫下过表达trcml6能够减缓地上部生物量的损失。
[0110]
(4)自然干旱胁迫
[0111]
如图5所示:自然干旱条件下野生型拟南芥叶片不再挺立,由叶尖向内逐渐萎焉、干枯;而过表达trcml6拟南芥多数叶片仍保持较为饱满挺立的状态,仅少数叶片萎焉失绿变为黄色或紫褐色。在正常条件下,过表达植株与野生型拟南芥的叶片相对含水量之间没有明显差异,而自然干旱后oe1株系与oe2株系的相对含水量显著高于野生型,分别为野生型的2.0倍和2.3倍。对各个株系的离体叶片进行失水率的测定,发现随着自然风干时间的延长,各株系叶片的失水率逐渐增加,但是过表达株系离体叶片的失水率增加程度在处理时间内(0-9h)一直低于野生型。自然风干9h后,野生型拟南芥的失水率为71.4%,而oe1株系与oe2株系的失水率分别为64.8%和59.1%。综上结果表明,过表达trcml6可以增强拟南芥对自然干旱胁迫的抗性,提高植株叶片的保水能力。
[0112]
最后应说明的是:以上各实施例仅用以说明本发明的技术方案,而非对其限制;尽管参照前述各实施例对本发明进行了详细的说明,本领域的普通技术人员应当理解:其依然可以对前述各实施例所记载的技术方案进行修改,或者对其中部分或者全部技术特征进行等同替换;而这些修改或者替换,并不使相应技术方案的本质脱离本发明各实施例技术方案的范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1