抗Trop2纳米抗体及其用途的制作方法
抗trop2纳米抗体及其用途
技术领域
1.本发明涉及生物免疫治疗技术领域,具体涉及抗trop2纳米抗体及其用途。
背景技术:
2.trop2是由tacstd2基因编码表达的细胞表面糖蛋白,全称为人滋养细胞表面糖蛋白抗原2(trophoblast cell surface antigens 2)。trop2由疏水性前导肽、细胞外结构域、一个跨膜结构域和一个胞质尾部组成,是单次跨膜糖蛋白,大小为35.7kd,是一类钙离子通道信号转化器。trop2蛋白n-端为胞外域(trop2ec),该胞外域通过一个单向跨膜螺旋(tm)与胞内短尾(trop2ic)连接,从而固定于胞膜。其细胞质尾部有高度保守的磷脂酰肌醇4,5-二磷酸(pip2)结合序列,表明pip2在trop2的信号转导中起重要作用。除了pip2结合基序外,它还含有保守的酪氨酸和丝氨酸磷酸化位点。
3.正常组织中trop2不表达或者低表达,在乳腺癌、胃癌、结直肠癌、胰腺癌、前列腺癌、宫颈癌、头颈癌和卵巢癌等多种恶性肿瘤中过表达,可促进肿瘤的发生、侵袭、转移和扩散等,在肿瘤生长过程中起着关键作用,所以trop2被认为是肿瘤免疫治疗候选的一个靶点。
4.嵌合抗原受体t细胞(chimeric antigen receptor-t cell,car-t)是指经基因修饰后,能以mhc非限制性方式识别特定目的抗原,并且持续活化扩增的t细胞。2012年国际细胞治疗协会年会中指出生物免疫细胞治疗已经成为手术、放疗、化疗外的第四种治疗肿瘤的手段,是一种针对肿瘤细胞表面特异性抗原的新型免疫治疗方法。大量研究表明,car-t细胞可以有效的识别肿瘤抗原,引起特异性的抗肿瘤免疫应答,显著改善患者的生存状况。
5.嵌合抗原受体(car)是car-t的核心部件,赋予t细胞以hla非依赖的方式识别肿瘤抗原的能力,这使得经过car改造的t细胞相较于天然t细胞表面受体tcr能够识别更广泛的目标。car由三个主要的结构域组成:胞外抗原结合域、跨膜结构域与胞内信号域。抗体作为car中重要的组成部分,其对于car-t发挥特异性高效杀伤作用、降低毒性具有决定性影响。因此能够以高特异性结合目标抗原、同时具有低免疫原性的抗体是开发car-t产品的重点。
6.在羊驼外周血液中存在一种天然缺失轻链的抗体,该抗体只包含一个重链可变区(variable domain of heavy chain of heavy chain antibody,vhh)和两个常规的ch2与ch3区。克隆并表达出来的vhh 结构具有与原重链抗体相当的结构稳定性以及与抗原的结合活性,是目前已知的可结合目标抗原的最小单位。vhh晶体为2.5nm,长4nm,分子量只有15kda,因此也被称作纳米抗体(nanobody,nb)。vhh可溶性极高,易于制造和修饰,循环半衰期短,具有高组织穿透性,不易聚集,能耐高温、强酸、强碱等致变性条件,适合于原核表达和各种真核表达系统,广泛用于开发治疗性抗体药物、诊断试剂、亲和纯化基质和科学研究等领域。纳米抗体具有独特的性质如分子量小、水溶性好、稳定性强、抗原识别能力强、易于生产等,近年来作为诊断工具越来越受欢迎,也被认为是car-t和靶向药物递送的重要基础。
7.目前尚未见如本技术所记载的抗trop2纳米抗体以及含该抗trop2纳米抗体的
car-t 细胞的报道。
技术实现要素:
8.本发明提供一种trop2结合分子,包含抗trop2纳米抗体或其抗原结合片段,所述抗trop2纳米抗体的互补决定区cdr包含cdr1、cdr2和cdr3,其中cdr1包括seq id no:1所示的序列、cdr2包括seq id no:2所示的序列、和cdr3包括seq id no:3所示的序列。
9.在一个或多个实施方案中,所述抗trop2纳米抗体的重链可变区序列如seq id no:4-5中任一所示。
10.在一个或多个实施方案中,所述抗trop2纳米抗体的fr1可选自seq id no:4-5中任一所示的vhh的fr1,fr2可选自seq id no:4-5中任一所示的vhh的fr2,fr3可选自seq id no:4-5中任一所示的vhh的fr3,fr4可选自seq id no:4-5中任一所示的vhh的fr4。
11.在一个或多个实施方案中,所述trop2结合分子是包含一条、两条或多条抗trop2纳米抗体或其抗原结合片段的单价或多价纳米抗体或单域抗体、或多特异性纳米抗体或单域抗体。
12.在一个或多个实施方案中,所述多价结合分子或多特异性结合分子通过连接子连接多个抗trop2纳米抗体或其抗原结合片段。所述连接子由选自g和s的1-15个氨基酸组成。
13.在一个或多个实施方案中,所述纳米抗体是骆驼重链抗体或软骨鱼重链抗体。
14.在一个或多个实施方案中,所述纳米抗体还包含重链恒定区。
15.在一个或多个实施方案中,所述重链恒定区是骆驼重链抗体的恒定区,包含ch2和ch3。
16.在一个或多个实施方案中,所述ch2和ch3是人igg fc的ch2和ch3,例如igg1的ch2和ch3。
17.在一个或多个实施方案中,所述重链恒定区是软骨鱼重链抗体的恒定区,包含ch1、ch2、ch3、ch4和ch5。
18.在一个或多个实施方案中,本发明任一实施方案所述的trop2结合分子为嵌合抗体或完全人抗体;优选为完全人抗体。
19.本发明另一方面提供一种嵌合抗原受体,包含任选的信号肽序列、本文任一实施方案所述的trop2结合分子、铰链区、跨膜区和胞内区。
20.在一个或多个实施方案中,胞内区包括胞内共刺激域和/或胞内信号域。
21.在一个或多个实施方案中,从n端到c端,该嵌合抗原受体依次含有信号肽、本文任一实施方案所述的trop2结合分子、铰链区、跨膜区、胞内共刺激域和胞内信号域。
22.本发明还提供核酸分子,其具有选自以下任一项的序列:(1)本文任一实施方案所述trop2结合分子或嵌合抗原受体的编码序列;(2)(1)的互补序列;(3)(1)或(2)中任一序列的5-50bp的片段。
23.在一个或多个实施方案中,所述片段是引物。
24.本发明还提供一种核酸构建物,包含本文所述的核酸分子。
25.在一个或多个实施方案中,所述核酸构建物是克隆载体、表达载体或整合载体。
26.本发明还提供一种宿主细胞,选自:
(1)表达和/或分泌本文任一实施方案所述trop2结合分子或嵌合抗原受体;(2)包含本文所述的核酸分子;和/或(3)包含本文所述的核酸构建物。
27.在一个或多个实施方案中,所述宿主细胞是免疫效应细胞,优选t细胞。
28.本发明还提供一种产生本文任一实施方案trop2结合分子的方法,包括:在适合产生trop2结合分子(例如纳米抗体或其抗原结合片段,单价或多价纳米抗体或单域抗体、或多特异性纳米抗体或单域抗体)的条件下培养本文所述的宿主细胞,和任选的从培养物中纯化所述trop2结合分子。
29.本发明还提供一种药物组合物,包含本文任一实施方案所述trop2结合分子、核酸分子、核酸构建物或宿主细胞,和药学上可接受的辅料。
30.在一个或多个实施方案中,所述药物组合物用于治疗trop2表达相关的疾病或病况。
31.本发明还提供本文任一实施方案所述trop2结合分子、嵌合抗原受体、核酸分子、核酸构建物或宿主细胞在制备活化的免疫细胞(例如t细胞)中的用途。
32.本发明还提供本文任一实施方案所述trop2结合分子、嵌合抗原受体、核酸分子、核酸构建物或宿主细胞在制备用于预防或治疗trop2表达相关的疾病或病况的药物中的用途。
33.在一个或多个实施方案中,所述疾病或病况选自以下的一种或多种:乳腺癌、胃癌、结直肠癌、胰腺癌、前列腺癌、宫颈癌、头颈癌、肺癌、食道癌、肾癌、膀胱癌、子宫癌和卵巢癌。
34.本发明还提供一种治疗或预防trop2表达相关的疾病或病况的方法,所述方法包括给予需要的患者治疗有效量的本发明任一实施方案所述的trop2结合分子或宿主细胞,或本发明任一实施方案所述的药物组合物。
35.本发明还提供一种检测trop2的试剂盒,用于例如评估药物治疗效果或诊断癌症,所述的试剂盒包含本文任一实施方案所述trop2结合分子、核酸分子、核酸构建物或宿主细胞。
36.在一个或多个实施方案中,所述试剂盒还包括用于检测trop2与所述trop2结合分子的结合的试剂。例如通过酶联免疫反应法检测所述结合的试剂。
37.在一个或多个实施方案中,所述检测结合的试剂是能与trop2结合分子连接的可检测标记物,例如生物素。所述的可检测标记物被连接于所述trop2结合分子或分离地存在于试剂盒中。
38.本发明还提供一种检测样品中trop2存在情况的非诊断性方法,所述方法包括:以本文任一实施方案所述trop2结合分子与样品孵育,和检测trop2与所述trop2结合分子的结合,从而确定样品中trop2存在情况。所述检测是酶联免疫反应法检测。
39.本发明还提供本文任一实施方案所述trop2结合分子在制备用于检测样品中trop2、评估药物治疗效果或诊断癌症的试剂盒中的用途。
40.本发明具有以下有益效果:本发明提供了一种新的特异性识别trop2的纳米抗体以及含有该抗体的car修饰细胞,该抗体和细胞具有良好的靶向trop2的治疗效果,为与trop2表达相关的疾病提供了
新的治疗或改善途径。
附图说明
41.为了更清楚地说明本发明实施例的技术方案,下面将对实施例中所需要使用的附图作简单地介绍,应当理解,以下附图仅示出了本发明的某些实施例,因此不应被看作是对范围的限定,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他相关的附图。
42.图1为重组人trop2-avi-his抗原蛋白的sds电泳图。
43.图2为不同克隆trop2car的示意图。
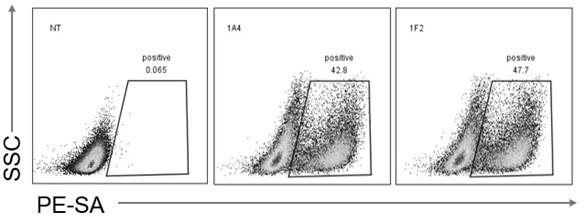
44.图3为不同克隆trop2car-t细胞的car表达阳性率。
45.图4为不同克隆trop2car-t细胞的cd107a表达。
46.图5为效靶比10:1时不同克隆trop2car-t细胞的infγ分泌。
47.图6为效靶比2:1时不同克隆trop2car-t细胞的infγ分泌。
48.图7为效靶比10:1时不同克隆trop2car-t细胞的il-2分泌。
49.图8为效靶比2:1时不同克隆trop2car-t细胞的il-2分泌。
50.图9为不同克隆trop2car-t细胞对靶细胞bxpc3-luc-gfp的杀伤实验结果。
51.图10为不同克隆trop2car-t细胞对靶细胞u251-luc-gfp的杀伤实验结果。
具体实施方式
52.本发明人经过广泛而深入地研究,经过大量的筛选,发现一类抗trop2纳米抗体及其抗原结合片段,其能够特异性识别trop2,以高亲和力与trop2结合,具有良好的功能活性。
53.具体地,本发明利用trop2蛋白免疫羊驼,获得高质量的单域抗体基因文库。然后利用噬菌体展示技术筛选抗体基因库,从而获得了trop2特异性的单域抗体基因。再将此基因转至哺乳动物细胞中,从而获得了能在哺乳动物细胞中高效表达的、且特异性高的抗体株。所述抗体或其抗原结合片段具有良好的安全性和靶向性,能够特异性结合人trop2的胞外域。
54.本发明还提供含有所述纳米抗体的嵌合抗原受体(car)。将包含该car的编码序列的载体用于感染免疫细胞,能够获得对过表达trop2的肿瘤细胞具有显著杀伤能力的免疫效应细胞,该免疫效应细胞能够应用于治疗或改善trop2表达相关的疾病,从而为trop2阳性肿瘤的治疗奠定了基础。
55.抗体本文中,“trop2结合分子”是特异性结合trop2的蛋白质,包括但不仅限于,抗体、重链抗体、纳米抗体或它们的抗原结合片段。
56.本文中,术语“抗体”包括单克隆抗体(包括全长抗体,其具有免疫球蛋白fc区),具有多表位特异性的抗体组合物,多特异性抗体(例如,双特异性抗体),双抗体和单链分子,以及抗体片段,尤其是抗原结合片段,例如,fab,f(ab’)2,fd和fv。本文中,“抗体”与“免疫球蛋白”可互换使用。
57.传统的“抗体”含有基本的4链抗体单元,是由两条相同的轻链(l)和两条相同的重
链(h)构成的异四聚体糖蛋白。每条重链在n-末端具有可变结构域(vh),接着是三个(对于每种α和γ链,ch1、ch2和ch3)和四个(对于μ和ε同种型,ch1、ch2、ch3和ch4)恒定结构域(ch)以及位于ch1结构域与ch2结构域之间的绞链区(hinge)。每条轻链在n-末端具有可变结构域(vl),接着是其另一端的恒定结构域(cl)。成对的vh和vl一起形成一个抗原结合位点。关于不同类别抗体的结构和性质,参见如basic and clinical immunology,第八版,daniel p. sties,abba i. terr 和tristram g. parsolw 编辑,appleton & lange,norwalk,ct,1994,第71 页和第6章。来自任何脊椎动物物种的轻链,根据其恒定结构域氨基酸序列,可归入两种称作κ和λ的截然不同型中的一种。根据ch序列和功能的相对较小差异,γ和α类可进一步分为亚类,例如人表达下列亚类:igg1、igg2a、igg2b、igg3、igg4、iga1和iga2。
58.本文所述“重链抗体”是源自骆驼科生物或软骨鱼科生物的抗体。相比上述4链抗体,重链抗体缺失轻链和重链恒定区1(ch1),仅包含2条由可变区(vhh)和其他恒定区组成重链,可变区通过类似铰链区结构与恒定区相连。骆驼科重链抗体的每条重链包含1个可变区(vhh)和2个恒定区(ch2和ch3),软骨鱼科重链抗体的每条重链含有1个可变区和5个恒定区(ch1-ch5)。重链抗体的抗原结合片段包括vhh和单链重链抗体。通过与人igg fc的恒定区融合,重链抗体可以具有人igg fc的ch2和ch3。
59.如本文所用,术语“单域抗体”、“抗trop2单域抗体”、“重链抗体的重链可变区结构域”、“vhh”可互换使用,均指特异性识别和结合于trop2的单域抗体。单域抗体是重链抗体的可变区。通常,单域抗体含有三个cdr和四个fr。优选地,本发明的单域抗体具有seq id no:1所示的cdr1、seq id no:2所示的cdr2、和seq id no:3所示的cdr3。单域抗体是最小的功能性抗原结合片段。通常先获得天然缺失轻链和重链恒定区1(ch1)的抗体后,再克隆抗体重链的可变区,构建仅由一个重链可变区组成的单域抗体。
60.本文中,“纳米抗体”是指含有本文所述vhh的抗体。其可以是如上所述的重链抗体,还可以是含有多条vhh的多价或多特异性抗体,也可以是将vhh和抗体fc(例如ch2和ch3或ch2、ch3和ch4)重组获得的重组抗体。包含两条或多条单域抗体的结合分子是多价单域抗体;包含两条或多条不同特异性单域抗体的结合分子是多特异性单域抗体。多价单域抗体或多特异性单域抗体通过连接子连接多个单域抗体。所述连接子通常由选自g和s的1-15个氨基酸组成。
61.本文中,重链抗体和抗体(传统四链抗体)旨在区分抗体的不同组合方式。由于二者的结构具有相似性,下述针对抗体的结构描述除涉及轻链外也均适用于重链抗体。
62.抗体的“可变区”或“可变结构域”是指抗体的重链或轻链的氨基末端结构域。重链和轻链的可变结构域可分别称为“vh”和“vl”。这些结构域通常是抗体的最可变的部分(相对于相同类型的其它抗体)并含有抗原结合位点。
63.术语“可变的”指可变结构域中的某些区段在抗体序列中差异广泛的情况。可变结构域介导抗原结合并限定特定抗体对其特定抗原的特异性。然而,变异性并非均匀分布于可变结构域跨越的全部氨基酸。相反,其集中在三个称为高变区(hvr)的区段(在轻链和重链可变结构域中均有),即分别为重链可变区的hcdr1、hcdr2、hcdr3(重链抗体中可简称为cdr1、cdr2、cdr3)以及轻链可变区的lcdr1、lcdr2和lcdr3。可变结构域中更为高度保守的部分称为骨架区(fr)。天然重链和轻链的可变结构域各自包含四个fr区(fr1、fr2、fr3和
fr4),它们大多采取β-折叠构象,通过形成环状连接且在有些情况中形成β-折叠结构一部分的三个hvr连接。每条链中的hvr通过fr区非常接近的保持在一起,并与另一条链的hvr一起促成抗体的抗原结合位点的形成。通常,轻链可变区的结构为fr1-lcdr1-fr2-lcdr2-fr3-lcdr3-fr4,重链可变区的结构为fr1-hcdr1-fr2-hcdr2-fr3-hcdr3-fr4。恒定结构域不直接参与抗体与抗原的结合,但展现出多种效应子功能,如在抗体依赖性细胞介导的细胞毒性中抗体的参与。抗体的可变区有多种标注方案,包括:chothia、kabat、imgt和contact。本文示例性使用imgt标注方案。
[0064]“fc区”(可结晶片段区域)或“fc结构域”或“fc”是指抗体重链的c-末端区域,其介导免疫球蛋白与宿主组织或因子的结合,包括与位于免疫系统的各种细胞(例如,效应细胞)上的fc受体的结合,或者与经典补体系统的第一组分(c1q)的结合。在igg,iga和igd抗体同种型中,fc区由来自抗体两条重链的ch2结构域和ch3结构域的两个相同的蛋白片段构成;igm和ige的fc区在每个多肽链中包含三个重链恒定结构域(ch结构域2-4)。虽然免疫球蛋白重链的fc区的边界可以变化,但是人igg重链fc区通常定义为从重链位置c226或p230的氨基酸残基到羧基端的序列段,其中该编号是根据eu索引,如在kabat中一样。如本文所使用的,fc区可以是天然序列fc或变体fc。
[0065]“抗体片段”包含完整抗体的一部分,优选完整抗体的抗原结合区和/或可变区。抗体片段优选为抗体的抗原结合片段。抗体片段的例子包括fab、fab’、f(ab’)、f(ab’)2、fd、和fv片段、二硫键连接的fv;双抗体;线性抗体;单链抗体分子;scfv-fc片段;由抗体片段形成的多特异性抗体;以及通过化学修饰或通过掺入脂质体中应能够增加半衰期的任何片段。抗原结合片段可以通过多种技术制备,包括但不限于将完整的抗体蛋白水解消化,以及由包含抗原结合片段的宿主细胞表达产生。
[0066]“fv”是含有完整抗原识别和结合位点的最小抗体片段。该片段由紧密、非共价结合的一个重链可变结构域和一个轻链可变结构域的二聚体组成。从这两个结构域的折叠中突出了六个高变环(重链和轻链各3个环),贡献出抗原结合的氨基酸残基并赋予抗体以抗原结合特异性。然而,即使是单个可变结构域(或只包含对抗原特异的三个hvr的半个fv)也具有识别和结合抗原的能力,尽管亲合力低于完整结合位点。“单链fv”也可缩写为“sfv”或“scfv”,是包含抗体vh和vl结构域的连接成一条多肽链的抗体片段。优选的是,sfv多肽在vh和vl结构域之间还包含多肽接头,使得sfv形成期望的抗原结合结构。对于重链抗体或纳米抗体而言,scfv即为vhh。
[0067]
本文中,术语“单克隆抗体”指从一群基本上同质的抗体中获得的抗体,即除了可能以少量存在的可能的天然出现的突变和/或翻译后修饰(例如异构化、酰胺化)之外,构成群体的各个抗体是相同的。单克隆抗体是高度特异性的,针对单个抗原位点。与多克隆抗体制剂(其典型地包括针对不同决定簇(表位)的不同抗体)相比,每个单克隆抗体针对抗原上的单个决定簇。除它们的特异性外,单克隆抗体的优势在于它们通过杂交瘤培养合成,未受到其它免疫球蛋白的污染。修饰语“单克隆”表明抗体从基本上同质的抗体群获得的特征,不应解释为要求通过任何特定方法来生产抗体。例如,将根据本发明使用的单克隆抗体可通过多种技术来生成,包括例如杂交瘤法、噬菌体展示法、重组dna法、及用于从具有部分或整个人免疫球蛋白基因座或编码人免疫球蛋白序列的基因的动物生成人或人样抗体的技术、单细胞测序法。
[0068]
单克隆抗体在本文中也包括“嵌合”抗体,其中重链和/或轻链的一部分与衍生自特定物种或属于特定抗体类别或亚类的抗体中的相应序列相同或同源,而链的剩余部分与衍生自另一物种或属于另一抗体类别或亚类的抗体中的相应序列相同或同源,以及此类抗体的片段,只要它们展现出期望的生物学活性。
[0069]
非人(例如鼠)抗体的“人源化”形式指最低限度包含衍生自非人免疫球蛋白的序列的嵌合抗体。因此,“人源化抗体”通常指可变结构域构架区与在人抗体中发现的序列交换的非人抗体。通常在人源化抗体中,整个抗体(除cdr以外)由人来源的多核苷酸编码或与这种抗体相同(除cdr以外)。cdr(其中一些或全部由源自非人生物体的核酸编码)被移植到人抗体可变区的β-折叠骨架中以产生抗体,其特异性由被移植的cdr来决定。这类抗体的产生方法本领域周知,例如使用具有基因工程免疫系统的小鼠而产生。本发明中,抗体、单域抗体、重链抗体等均包括各所述抗体的经人源化的变体。
[0070]“人抗体”指这样的抗体,其具有与由人生成的抗体的氨基酸序列对应的氨基酸序列和/或使用本文所公开的用于生成人抗体的任何技术产生。人抗体的这种定义明确排除包含非人抗原结合残基的人源化抗体。人抗体可使用本领域已知的多种技术来生成,包括噬菌体展示文库。
[0071]
在一些实施方案中,本发明还提供与本发明的任何抗trop2纳米抗体的抗原接合区结合人trop2上相同表位的纳米抗体、重链抗体、抗体或其抗原结合片段(例如单域抗体vhh),即能够与本发明的任何纳米抗体的抗原结合区交叉竞争与trop2的结合的纳米抗体、重链抗体、抗体或其抗原结合片段。
[0072]
本发明中,抗trop2单域抗体具有seq id no:1所示的cdr1、seq id no:2所示的cdr2、和seq id no:3所示的cdr3。
[0073]
本文所述的抗trop2单域抗体的fr1、fr2、fr3和fr4可分别独立选自seq id no:4-5中任一所示的单域抗体的fr1、fr2、fr3和fr4。优选地,抗trop2单域抗体的氨基酸序列如seq id no:4-5中任一所示。
[0074]
当单域抗体与重链恒定区连接时,纳米抗体是包含本文所述单域抗体的重链抗体。重链恒定区可以是骆驼重链抗体的恒定区,包含ch2和ch3。优选地,所述抗体恒定区源自:igg1、igg2、igg3、igg4、iga、igm、ige和igd中的任意一者的恒定区,更优选源自igg1、igg2、igg3、igg4中的任意一者的恒定区。在一个或多个实施方案中,所述重链恒定区是人igg fc的ch2和ch3,例如igg1的ch2和ch3。
[0075]
本文所述trop2结合分子可以是包含一条、两条或多条本文所述的抗trop2纳米抗体或单域抗体的单价或多价纳米抗体或单域抗体、或多特异性纳米抗体或单域抗体。多特异性可以是针对trop2和另一种抗原,也可以是针对trop2的两种不同表位。
[0076]
本发明还包括所述抗体衍生物和类似物。“衍生物”和“类似物”是指基本上保持本发明抗体相同的生物学功能或活性的多肽。本发明的衍生物或类似物可以是(i)在一个或多个氨基酸残基中具有取代基团的多肽,或(ii)成熟多肽与另一个化合物(比如延长多肽半衰期的化合物,例如聚乙二醇)融合所形成的多肽,或(iii)附加的氨基酸序列融合到此多肽序列而形成的多肽(如前导序列或分泌序列或用来纯化此多肽的序列或蛋白原序列,或与6his标签形成的融合蛋白)。根据本文的教导,这些衍生物和类似物属于本领域熟练技术人员公知的范围。
[0077]
在不实质性影响抗体活性的前提下,本领域技术人员可以对本发明的抗体序列改变一个或更多个(例如1、2、3、4、5、6、7、8、9或10个或更多个)氨基酸,以获得所述抗体或其功能性片段序列的变体。这些变体包括(但并不限于):一个或多个(通常为1-50个,较佳地1-30个,更佳地1-20个,最佳地1-10个)氨基酸的缺失、插入和/或取代,以及在c末端和/或n末端添加一个或数个(通常为20个以内,较佳地为10个以内,更佳地为5个以内)氨基酸。在本领域中,用性能相近或相似的氨基酸进行保守性取代时,通常不会改变蛋白质的功能。如在fr和/或fc区中将具有类似性质的氨基酸进行取代。可进行保守性取代的氨基酸残基为本领域所周知。这样的取代的氨基酸残基可以是也可以不是由遗传密码编码的。又比如,在c末端和/或n末端添加一个或数个氨基酸通常也不会改变蛋白质的功能。它们都被视为包括在本发明保护的范围内。
[0078]
本文所述抗体的变异形式包括:同源序列、保守性变异体、等位变异体、天然突变体、诱导突变体、在高或低的严紧度条件下能与本发明抗体的编码dna杂交的dna所编码的蛋白、以及利用抗本发明抗体的抗血清获得的多肽或蛋白。在一些实施方案中,本发明所述变体的序列可以与其来源序列有至少有95%、96%、97%、98%或99%的一致性。本发明所述的序列一致性可以使用序列分析软件测量。例如使用缺省参数的计算机程序blast,尤其是blastp或tblastn。本发明还包括那些具有带cdr的抗体重链可变区的分子,只要其cdr与此处鉴定的cdr具有90%以上(较佳地95%以上,最佳地98%以上)的同源性。
[0079]
可采用本领域常规的方法制备本发明的抗体,如杂交瘤技术。可采用本领域常规的方法制备本发明的纳米抗体,如本领域熟知的噬菌体展示技术。或者,本发明的抗体或纳米抗体可在其他细胞系中表达。可用编码本发明抗体的序列转化合适的哺乳动物宿主细胞,然后培养宿主细胞并纯化抗体。转化可采用任何已知的方法进行,例如包括将多核苷酸包装在病毒(或病毒载体中)并用病毒(或载体)转导宿主细胞。所用的转化程序取决于将转化的宿主。用于将异源多核苷酸引入哺乳动物细胞中的方法为本领域所熟知,包括葡聚糖介导的转染、磷酸钙沉淀、聚凝胺介导的转染、原生质体融合、电穿孔、将多核苷酸囊封在脂质体中和将dna直接微注射至核中等。可用作用于表达的宿主的哺乳动物细胞系为本领域所熟知,包括但不限于可从美国典型培养物保藏中心(atcc)获得的多种永生化细胞系,包括但不限于中国仓鼠卵巢(cho)细胞、hela细胞、幼仓鼠肾(bhk)细胞、猴肾细胞(cos)、人肝细胞癌细胞(例如,hepg2)等。
[0080]
car本发明还提供一种靶向trop2的嵌合抗原受体(car)。该car含有任选的信号肽序列、抗原识别区即本文所述的抗trop2结合分子、铰链区、跨膜区和胞内区。其中胞内区包含一个或多个胞内共刺激域和/或一个或多个胞内信号域。本文中的“铰链区”、“跨膜区”和“胞内区”均可选自已知的car-t技术中的铰链区、跨膜区和胞内区的序列。
[0081]
car上任选的信号肽可以根据需要选择。一般而言,信号肽是使多肽靶向细胞中的所需部位的肽序列。信号肽使多肽靶向细胞的分泌通路,并且将允许多肽整合和锚定至脂双层;信号肽还可以是膜定位信号肽。示例性的信号肽例如cd8信号肽、cd28信号肽、cd4信号肽或轻链信号肽,其序列在本领域技术人员的知识范围内。适用于本发明的cd8信号肽可以是本领域常用于car的各种人cd8信号肽序列。在某些实施方案中,所述cd8信号肽的氨基酸序列包含seq id no:6所示序列。
[0082]
嵌合抗原受体的铰链区位于胞外抗原结合区和跨膜区之间,铰链区是通常在蛋白质的两个域之间存在的氨基酸区段,并且可以允许蛋白质的柔性和两个域的彼此相对运动。铰链区可以是天然存在的蛋白质的铰链区或其部分。抗体(诸如igg、iga、igm、ige或igd抗体)的铰链区也可用于本文所述的嵌合抗原受体。非天然存在的肽也可用作本文所述的嵌合抗原受体的铰链区。示例性地,car的铰链区选自cd8α铰链区、igd铰链区、igg1 fc ch2ch3铰链区或igg4 fc ch2ch3铰链区,其序列在本领域技术人员的知识范围内。适用于本发明的cd8α铰链区可以是本领域常用于car的各种人cd8α铰链区序列。在某些实施方案中,所述人cd8α铰链区包含seq id no:7所示序列。
[0083]
嵌合抗原受体的跨膜区可以形成α螺旋、多于一个α螺旋的复合物、β桶或能够跨域细胞磷脂双层的任何其它稳定结构。跨膜区可以是天然或合成来源的。跨膜区可选自以下蛋白的跨膜区:cd3ε、cd4、cd5、cd8α、cd9、cd16、cd22、cd28、cd33、cd37、cd45、cd64、cd80、cd86、cd134、cd137、cd154、t细胞受体的α、β或ζ链。适用于本发明的人cd8α跨膜区可以是本领域常用于car的各种人cd8α跨膜区序列。在某些实施方案中,所述人cd8α跨膜区的氨基酸序列包含seq id no:8所示序列。
[0084]
胞内信号区(或胞内信号传导区)负责表达嵌合抗原受体的免疫效应细胞的至少一种正常效应子功能的活化。例如,t细胞的效应子功能可以是细胞裂解活性或辅助活性,包括细胞因子的分泌。虽然通常可以利用整个胞内信号传导区,但是在很多情况下,使用整个链是不必要的。就使用胞内信号传导区的截短部分而言,只要其转导效应子功能信号,就可以使用这种截短部分代替完整链。因此,胞内信号传导区包括足以转导效应子功能信号的胞内信号传导区的任何截短形式。car的胞内信号域可以根据需要选择,包括但不限于来源于cd3ζ、fcrγ (fcer1g)、fcrβ(fcεrib)、cd3γ、cd3δ、cd3ε、cd5、cd22、cd79a、cd79b和cd66d中的至少一种的胞内信号域。优选地,所述胞内信号区来源于人cd3ζ胞内信号区。进一步地,所述人cd3ζ胞内信号区具有seq id no:10所示氨基酸序列。
[0085]
在抗原特异性信号的刺激以外,很多免疫效应细胞还需要共刺激来促进细胞增殖、分化和存活,以及活化细胞的效应子功能。“共刺激域”可以是共刺激分子的胞质部分。术语“共刺激分子”是指免疫细胞(诸如t细胞)上的关联结合伴侣,该关联结合伴侣与共刺激配体特异性结合,从而由免疫细胞介导共刺激响应,诸如但不限于增殖和存活。可以根据需要选择适合的胞内共刺激域,包括具有共刺激信号分子的胞内结构域,例如源自4-1bb,card11,cd2,cd7,cd27,cd28,cd30,cd40,cd54,cd83,ox40,cd137,cd134,cd150,cd152,cd223,cd270,pd-l2,pd-l1,cd278,dap10,lat,nkd2c,slp76,trim,fcεriγ,myd88,和41bbl的胞内结构域的至少一种。在某些实施方案中,4-1bb共刺激域的氨基酸序列包含seq id no:9所示序列。
[0086]
形成本发明的嵌合抗原受体的上述各部分,如cd8信号肽、抗trop2纳米抗体、cd8α铰链区、cd8α跨膜区、cd3ζ胞内信号域、4-1bb共刺激域等,相互之间可直接连接,或者可通过接头序列连接。接头序列可以是本领域周知的适用于抗体的接头序列,例如含g和s的接头序列。通常,接头含有一个或多个前后重复的基序。例如,该基序可以是gggs、ggggs、ssssg、gsgsa和ggsgg。优选地,该基序在接头序列中是相邻的,在重复之间没有插入氨基酸残基。接头序列可以包含1、2、3、4或5个重复基序组成。接头的长度可以是3~25个氨基酸残基,例如3~15、5~15、10~20个氨基酸残基。在某些实施方案中,接头序列是多甘氨酸接头
序列。接头序列中甘氨酸的数量无特别限制,通常为2~20个,例如2~15、2~10、2~8个。除甘氨酸和丝氨酸来,接头中还可含有其它已知的氨基酸残基,例如丙氨酸(a)、亮氨酸(l)、苏氨酸(t)、谷氨酸(e)、苯丙氨酸(f)、精氨酸(r)、谷氨酰胺(q)等。在某些实施方案中,接头序列为(ggggs)n连接,其中n为1~5的整数。
[0087]
在示例性实施方案中,car从n端到c端依次含有cd8信号肽、本文所述的抗trop2纳米抗体或其抗原结合片段、cd8α铰链区、cd8α跨膜区、4-1bb共刺激域、cd3ζ胞内信号域。在具体实施例中,具有上述结构的示例性car如seq id no:11-12中任一所示。
[0088]
应理解,在基因克隆操作中,常常需要设计合适的酶切位点,这势必在所表达的氨基酸序列末端引入了一个或多个不相干的残基,而这并不影响目的序列的活性。为了构建融合蛋白、促进重组蛋白的表达、获得自动分泌到宿主细胞外的重组蛋白、或利于重组蛋白的纯化,常常需要将一些氨基酸添加至重组蛋白的n-末端、c-末端或该蛋白内的其它合适区域内,例如,包括但不限于,适合的接头肽、信号肽、前导肽、末端延伸等。因此,本发明的car的氨基端或羧基端还可含有一个或多个多肽片段,作为蛋白标签。任何合适的标签都可以用于本文。例如,所述的标签可以是flag,ha,ha1,c-myc,poly-his,poly-arg,strep-tagii,au1,ee,t7,4a6,ε,b,ge以及ty1。这些标签可用于对蛋白进行纯化。
[0089]
本发明的car中的抗原识别区可以是前述的抗trop2纳米抗体或其功能性片段序列的变体。此外,car的其他部分也可以发生序列变化,得到的突变体与该car具有至少80%,优选至少85%,优选至少90%,优选至少95%,优选至少97%的序列相同性并保留该car的生物学活性(如活化t细胞)。可采用例如ncbi的blastp计算两条比对的序列之间的序列相同性。
[0090]
突变体还包括:在任一实施方案所述的car的氨基酸序列中具有一个或数个突变(插入、缺失或取代)、同时仍保留该car的生物学活性的氨基酸序列。所述数个突变通常指1-10个以内,例如1-8个、1-5个或1-3个。取代优选是保守性取代。例如,在本领域中,用性能相近或相似的氨基酸进行保守性取代时,通常不会改变蛋白质或多肽的功能。“性能相近或相似的氨基酸”包括例如,具有相似侧链的氨基酸残基的家族,这些家族包括具有碱性侧链的氨基酸(例如赖氨酸、精氨酸、组氨酸)、具有酸性侧链的氨基酸(例如天冬氨酸、谷氨酸)、具有不带电荷的极性侧链的氨基酸(例如甘氨酸、天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸、酪氨酸、半胱氨酸)、具有非极性侧链的氨基酸(例如丙氨酸、缬氨酸、亮氨酸、异亮氨酸脯氨酸、苯丙氨酸、甲硫氨酸、色氨酸)、具有β-分支侧链的氨基酸(例如苏氨酸、缬氨酸、异亮氨酸)和具有芳香侧链的氨基酸(例如酪氨酸、苯丙氨酸、色氨酸、组氨酸)。因此,在本发明多肽中用来自同一侧链类的另一氨基酸残基替换一个或几个位点,将不会在实质上影响其活性。
[0091]
核酸本发明还提供了编码上述抗体或car的多核苷酸。本发明的多核苷酸可以是dna形式或rna形式。dna形式包括cdna、基因组dna或人工合成的dna。dna可以是单链的或是双链的。dna可以是编码链或非编码链。本发明也包括编码融合蛋白的多核苷酸序列的简并变异体,即编码相同的氨基酸序列但核苷酸序列有所不同的核苷酸序列。
[0092]
所以,本发明还涉及与上述多核苷酸序列杂交且两个序列之间具有至少50%,较佳地至少70%,更佳地至少80%相同性的多核苷酸。本发明特别涉及在严谨条件下与本发
明所述多核苷酸可杂交的多核苷酸。在本发明中,“严谨条件”是指:(1)在较低离子强度和较高温度下的杂交和洗脱,如0.2
×
ssc,0.1%sds,60℃;或(2)杂交时加有变性剂,如50%(v/v)甲酰胺,0.1%小牛血清/0.1%ficoll,42℃等;或(3)仅在两条序列之间的相同性至少在90%以上,更好是95%以上时才发生杂交。并且,可杂交的多核苷酸编码的多肽与成熟多肽有相同的生物学功能和活性。
[0093]
本发明的抗体的核苷酸全长序列或其片段通常可以用pcr扩增法、重组法或人工合成的方法获得。一种可行的方法是用人工合成的方法来合成有关序列,尤其是片段长度较短时。通常,通过先合成多个小片段,然后再进行连接可获得序列很长的片段。此外,还可将重链的编码序列和表达标签(如6his)融合在一起,形成融合蛋白。car的序列也可以如上获得。或者,可以如上得到car的各部分(信号肽、抗原识别区、铰链区、跨膜区或胞内区)的序列后再连接得到car的全长。
[0094]
一旦获得了有关的序列,就可以用重组法来大批量地获得有关序列。这通常是将其克隆入载体,再转入细胞,然后通过常规方法从增殖后的宿主细胞中分离得到有关序列。本发明所涉及的生物分子(核酸、蛋白等)包括以分离的形式存在的生物分子。目前,已经可以完全通过化学合成来得到编码本发明蛋白(或其片段,或其衍生物)的dna序列。然后可将该dna序列引入本领域中已知的各种现有的dna分子(或如载体)和细胞中。此外,还可通过化学合成将突变引入本发明蛋白序列中。car各部分可以顺序克隆到载体中或者可以整合为全长car后再克隆。
[0095]
本发明也涉及核酸构建物,该核酸构建物含有本文所述的多核苷酸序列,以及与这些序列操作性连接的一个或多个调控序列。本发明所述的多核苷酸序列可以多种方式被操作以保证所述抗体或car的表达。在将核酸构建物插入载体之前可根据表达载体的不同或要求而对核酸构建物进行操作。利用重组dna方法来改变多核苷酸序列的技术是本领域已知的。
[0096]
调控序列可以是合适的启动子序列。启动子序列通常与待表达蛋白的编码序列操作性连接。启动子可以是在所选择的宿主细胞中显示转录活性的任何核苷酸序列,包括突变的、截短的和杂合启动子,并且可以从编码与该宿主细胞同源或异源的胞外或胞内多肽的基因获得。合适的启动子的一个例子为即时早期巨细胞病毒(cmv)启动子序列。该启动子序列是能够驱动可操作地连接至其上的任何多核苷酸序列高水平表达的强组成型启动子序列。合适的启动子的另一个例子为延伸生长因子-1α(ef-1α)。然而,也可使用其他组成型启动子序列,包括但不限于类人猿病毒40(sv40)早期启动子、小鼠乳癌病毒(mmtv)、人免疫缺陷病毒(hiv)长末端重复(ltr)启动子、momulv启动子、鸟类白血病病毒启动子、eb病毒即时早期启动子、鲁斯氏肉瘤病毒启动子、以及人基因启动子,诸如但不限于肌动蛋白启动子、肌球蛋白启动子、血红素启动子和肌酸激酶启动子。进一步地,也可考虑使用诱导型启动子。诱导型启动子的使用提供了分子开关,其能够在期限表达时打开可操作地连接诱导型启动子的多核苷酸序列的表达,而在当表达是不期望的时关闭表达。诱导型启动子的例子包括但不限于金属硫蛋白启动子、糖皮质激素启动子、孕酮启动子和四环素启动子。
[0097]
调控序列也可以是合适的转录终止子序列,由宿主细胞识别以终止转录的序列。终止子序列与编码该多肽的核苷酸序列的3’末端操作性连接。在选择的宿主细胞中有功能的任何终止子都可用于本发明。调控序列也可以是合适的前导序列,对宿主细胞翻译重要
的mrna的非翻译区。前导序列与编码该多肽的核苷酸序列的5’末端可操作连接。在选择的宿主细胞中有功能的任何终止子都可用于本发明。
[0098]
在某些实施方案中,所述核酸构建物是载体,例如克隆载体、表达载体和整合载体。通常通过可操作地连接本发明的多核苷酸序列至表达载体,实现本发明多核苷酸序列的表达。典型的克隆载体包含可用于调节期望核酸序列表达的转录和翻译终止子、起始序列和启动子。整合载体含有将靶序列整合到细胞基因组上的组件。这些载体可以用于转化适当的宿主细胞,以使其能够表达蛋白质。载体通常含有用于质粒维系和用于克隆与表达外源性核苷酸序列的序列。所述序列(在某些实施方案中总称为“侧翼序列”)通常包括一个或多个以下核苷酸序列:启动子、一个或多个增强子序列、复制起点、转录终止序列、含有供体和受体剪接位点的完全内含子序列、编码用于多肽分泌的前导序列的序列、核糖体结合位点、聚腺苷酸化序列、用于插入编码将要表达的抗体的核酸的多连接子区和可选标记元件。
[0099]
此外,载体的类型不受限制,例如,质粒、噬菌粒、噬菌体衍生物、动物病毒和粘粒,可根据待导入的宿主细胞而改变。病毒载体技术在本领域中是公知的并在例如sambrook等(2001,molecular cloning: a laboratory manual,cold spring harbor laboratory,new york)和其他病毒学和分子生物学手册中进行了描述。可用作载体的病毒包括但不限于逆转录病毒、腺病毒、腺伴随病毒、疱疹病毒和慢病毒。
[0100]
为了评估car多肽或其部分的表达,被引入细胞的载体也可包含可选择的标记基因或报道基因中的任一个或两者,以便于从通过病毒载体寻求被转染或感染的细胞群中鉴定和选择表达细胞。
[0101]
细胞适用于导入本文所述核酸构建物的宿主细胞可以是原核细胞,如细菌细胞;或是低等真核细胞,如酵母细胞;或是高等真核细胞,如哺乳动物细胞,特别是免疫细胞,优选免疫效应细胞。代表性例子有:大肠杆菌,链霉菌属;鼠伤寒沙门氏菌的细菌细胞;真菌细胞如酵母;果蝇s2或sf9的昆虫细胞;cho、cos7、293细胞的动物细胞等。
[0102]“免疫效应细胞”是可执行免疫效应功能的免疫细胞。在一些实施方案中,免疫效应细胞表达至少fcγriii并执行adcc效应子功能。介导adcc的免疫效应细胞的实例包括外周血单个核细胞(pbmc)、天然杀伤(nk)细胞、单核细胞、细胞毒性t细胞、中性粒细胞和嗜酸性粒细胞。优选地,免疫效应细胞选自:由多能干细胞或胚胎干细胞培养分化的免疫细胞、t淋巴细胞、nk细胞、外周血单个核细胞(pbmc)和造血干细胞中的至少一种。更优选地,所述免疫效应细胞为t淋巴细胞(同t细胞)。在一些实施方案中,t细胞可以为cd4+/cd8-、cd4-/cd8+、cd4+/cd8+、cd4-/cd8-或它们的组合。在一些实施方案中,t细胞在表达嵌合抗原受体并结合至靶细胞时产生il-2、ifn和/或tnf。在一些实施方案中,cd8+ t细胞在表达嵌合抗原受体并结合至靶细胞时裂解抗原特异性靶细胞。
[0103]
适用于本发明的t细胞可以是各种来源的各种类型的t细胞。例如,t细胞可来源于恶性实体瘤(例如胰腺癌)患者的pbmc。在某些实施方案中,获得t细胞后,可先用适量的(例如30~80ng/ml,如50ng/ml)的cd3抗体刺激活化,然后在含有适量的(例如30~80iu/ml,如50iu/ml)的il2培养基进行培养备用。
[0104]
将核酸或载体引入哺乳动物细胞的方法是本领域已知的,所述载体可以通过物
理、化学或生物方法转入细胞。当宿主为原核生物如大肠杆菌时,能吸收dna的感受态细胞可在指数生长期后收获,用cacl2法处理,所用的步骤在本领域众所周知。当宿主是真核生物,可选用如下的dna转染方法:磷酸钙共沉淀法,常规机械方法如显微注射、电穿孔,脂质体包装等。在一些实施方案中,转导的或转染的免疫效应细胞在引入核酸或载体之后离体繁殖。
[0105]
获得的转化子可以用常规方法培养,表达本发明的基因所编码的抗体或car。根据所用的宿主细胞,培养中所用的培养基可选自各种常规培养基。在适于宿主细胞生长的条件下进行培养。当宿主细胞生长到适当的细胞密度后,用合适的方法(如温度转换或化学诱导)诱导选择的启动子,将细胞再培养一段时间。
[0106]
在上面的方法中的多肽可在细胞内、或在细胞膜上表达、或分泌到细胞外。如果需要,可利用其物理的、化学的和其它特性通过各种分离方法分离和纯化重组的蛋白。这些方法是本领域技术人员所熟知的。这些方法的例子包括但并不限于:常规的复性处理、用蛋白沉淀剂处理(盐析方法)、离心、渗透破菌、超处理、超离心、分子筛层析(凝胶过滤)、吸附层析、离子交换层析、高效液相层析(hplc)和其它各种液相层析技术及这些方法的结合。
[0107]
用途和方法通过构建纳米抗体文库,发明人筛选了可以结合trop2的纳米抗体。利用这些纳米抗体,发明人构建了car以及car-t细胞,经细胞水平实验验证,所述car-t有很强的免疫功能,更优的cd107a表达、ifn-γ和il-2分泌以及对靶细胞的特异性杀伤功能,体内药效显著。
[0108]
本文所述的抗体、car、编码序列、核酸构建物、和细胞的所有方面都可用于制备用以预防或治疗本文所述各种病况和疾病的药物,所述病况和疾病是与trop2表达相关的疾病或病况,指由trop2表达异常所直接或间接导致的疾病,通常是指由trop2过表达所导致的疾病,例如癌症,包括但不限于:乳腺癌、胃癌、结直肠癌、胰腺癌、前列腺癌、宫颈癌、头颈癌、肺癌和卵巢癌。
[0109]
本发明还包括一类细胞疗法,包括在免疫细胞(例如t细胞)中表达本文所述的car,和对需要其的接受者施用治疗有效量的细胞,细胞能够杀死接受者的肿瘤细胞。相比抗体疗法,car-t细胞能够体内复制,产生可导致持续肿瘤控制的长期持久性。由car-t细胞引起的抗肿瘤免疫应答可为主动或被动免疫应答。另外,car介导的免疫应答可为过继免疫疗法步骤的一部分,其中car-t细胞诱导对car中的抗原结合部分特异性的免疫应答。
[0110]
本发明的抗体、核酸或car-修饰的细胞可被单独施用或作为药物组合物与稀释剂和/或与其他组分诸如相关的细胞因子或细胞群结合施用。在此方面,药物组合物可以通过使具有所需纯度的活性药剂与任选的药学上可接受的载剂混合以冻干制剂或水溶液的形式制备。药学上可接受的载剂在所用的剂量和浓度下对接受者是无毒的,可包括缓冲剂(例如中性缓冲盐水、硫酸盐缓冲盐水)、抗氧化剂、防腐剂、等渗剂、稳定剂、螯合剂(例如edta或谷胱甘肽)、佐剂(例如氢氧化铝)和表面活性剂中的至少一种。此外,为了使药物组合物可用于体内施用,它们必须是无菌的。可以通过无菌过滤膜过滤使药物组合物无菌。
[0111]
在一些实施方案中,药物组合物可以含有:细胞毒性剂、化学治疗剂、细胞因子、免疫抑制剂、生长抑制剂以及待治疗的具体适应症所需的活性药剂中的至少一种添加剂。添加剂的具体添加量可根据实际需要进行调整。本发明的药物组合物可以以“免疫学上有效
量”、“抗肿瘤有效量”、“肿瘤-抑制有效量”或“治疗量”的量施用。“治疗”指向受试者采用本文所述治疗方案以达到至少一种阳性治疗效果(比如,癌症细胞数目减少、肿瘤体积减小、癌细胞浸润至周边器官的速率降低或肿瘤转移或肿瘤生长的速率降低)。当指出“免疫学上有效量”、“抗肿瘤有效量”、“肿瘤-抑制有效量”或“治疗量”时,待施用的本发明组合物的精确量可由医师确定,其考虑患者(对象)的年龄、重量、肿瘤大小、感染或转移程度和病症的个体差异。通常,包括本文描述的t细胞的药物组合物可以以104至109个细胞/kg体重的剂量,优选105至106个细胞/kg体重的剂量。t细胞组合物也可以以这些剂量多次施用。细胞可通过使用免疫疗法中公知的注入技术(见例如rosenberg等,new eng.j.of med.319:1676,1988)施用。对于具体患者的最佳剂量和治疗方案可通过监测患者的疾病迹象并因此调节治疗由医学领域技术人员容易地确定。
[0112]
组合物的施用可以以任何方便的方式进行,包括通过喷雾法、注射、吞咽、输液、植入或移植。本文描述的组合物可被皮下、皮内、瘤内、结内、脊髓内、肌肉内、通过静脉内注射或腹膜内施用给患者。在一个实施方案中,本发明的t细胞组合物通过皮内或皮下注射被施用给患者。在另一个实施方案中,本发明的t细胞组合物优选通过静脉注射施用。t细胞的组合物可被直接注入肿瘤、淋巴结或感染位置。
[0113]
在本发明的一些实施方案中,本发明的car-t细胞或其组合物可与本领域已知的其它疗法结合。所述疗法包括但不限于化疗、放疗和免疫抑制剂。例如,可结合本领域周知的治疗间皮素介导的疾病的放疗或化疗制剂进行治疗。
[0114]
本文中,“抗肿瘤效应”指一种生物学效应,其可由肿瘤体积的减少、肿瘤细胞数的减少、转移数的减少、预期寿命的增加或与癌相关的各种生理症状的改善表示。
[0115]“患者”、“对象”、“个体”等等在本文中可交换使用,指可引起免疫应答的活有机体,如哺乳动物。例子包括但不限于人、狗、猫、小鼠、大鼠和其转基因物种。
[0116]
诊断、检测和试剂盒本发明的结合分子因其与trop2的高亲合力可用于测定,例如结合测定来检测和/或定量在组织或细胞中表达的trop2。结合分子例如单域抗体可用在进一步研究trop2在疾病中的作用的研究中。检测trop2的方法大致如下:获得细胞和/或组织样本;检测样本中trop2的水平。
[0117]
本发明的trop2结合分子可用于诊断目的,用来检测、诊断或监控与trop2相关的疾病和/或病况。本发明提供使用本领域技术人员已知的经典免疫组织学方法检测样本中trop2的存在。可以体内或体外进行trop2的检测。适用于检测trop2的存在的方法实例包括elisa、facs、ria等。
[0118]
对于诊断应用来说,通常用可检测的标记基团来标记结合分子例如单域抗体。合适的标记基团包括(但不限于)以下:放射性同位素或放射性核素(例如,3h、14c、15n、35s、90y、99tc、111in、125i、131i)、荧光基团(例如,fitc、罗丹明、镧系元素磷光体)、酶促基团(例如,辣根过氧化物酶、β根半乳糖苷酶、荧光素酶、碱性磷酸酶)、化学发光基团、生物素基基团或由二级报导体识别的预定多肽表位(例如,亮氨酸拉链对序列、用于二级抗体的结合位点、金属结合结构域、表位标签)、mri(磁共振成像)或ct(电子计算机x射线断层扫描技术)造影剂。用于标记蛋白质的各种方法在本领域中已知且可用来进行本发明。
[0119]
本发明的另一方面提供检测与本发明的抗体竞争结合trop2的测试分子的存在的
方法。一种所述测定的实例将涉及在存在或不存在测试分子的情形下检测含有一定量trop2的溶液中的游离抗体的量。游离抗体(即,未结合trop2的抗体)的量增加将表示测试分子能与该抗体竞争结合trop2。在一个实施方案中,用标记基团标记抗体。或者,标记测试分子并在存在或不存在抗体的情形下监控游离测试分子的量。
[0120]
本发明还提供了用于检测trop2水平的检测试剂盒,该试剂盒包括识别trop2蛋白的抗体,用于溶解样本的裂解介质,检测所需的通用试剂和缓冲液,如各种缓冲液、检测标记、检测底物等。该检测试剂盒可以是体外诊断装置。
[0121]
下文将以具体实施例的方式阐述本发明。应理解,这些实施例仅仅是阐述性的,并非意图限制本发明的范围。实施例中所用到的方法和材料,除非另有说明,否则均为本领域常规的材料和方法。
实施例
[0122]
实施例1:重组人trop2蛋白表达载体构建与真核表达1.基因序列的合成及蛋白的表达载体的构建从uniport下载人trop2蛋白序列(https://www.uniprot.org/uniprot),经密码子在线优化工具(http://www.jcat.de/#opennewwindow)优化后,交由生工合成trop2胞外段基因序列。同时在该基因序列的3’端添加avi-tag和6
×
his-tag的核酸序列,融合基因序列编码seq id no:15所示的氨基酸序列,融合基因的核苷酸序列如seq id no:16所示。通过分子克隆,拼接产物用takara无缝克隆试剂盒克隆到ptt5中,获得表达载体。
[0123]
2.重组人trop2蛋白的表达、纯化以获得的表达载体转染293t细胞(atcc)5天后,收集培养上清,用akta explorer 100(ge)纯化重组人trop2蛋白。trop2蛋白经sds-page电泳后通过考马斯亮蓝染色显示其大小约36千道尔顿左右,结果如图1所示。
[0124]
实施例2:抗trop2vhh抗体的制备1.vhh噬菌体展示文库构建采用一步法构建纳米抗体噬菌体展示文库,即将羊驼纳米抗体vhh基因连接到噬菌体展示载体中。
[0125]
1)羊驼免疫羊驼选择:选择健康强壮、精神状态良好、体型适中的羊驼,挑选的羊驼毛色光亮,无受伤不适症状。挑选好动物,先预养1周左右以淘汰有些不合格的动物,使后期的实验能顺利进行。
[0126]
免疫方案:挑选好羊驼并确保动物适合,记录耳号后开始免疫实验。一共进行4次免疫。免疫方案如下:d0,免疫前取血10ml,作为阴性血清对照,将抗原0.5mg与cfa 1ml 混匀后皮下注射;d21,将抗原0.25mg与 cfa 1ml 混匀后皮下注射;d28,取血10 ml;d42,将抗原0.25mg与 cfa 1ml 混匀后皮下注射;d49,采50ml外周血分离淋巴细胞;d63,将抗原0.25mg与 cfa 1ml 混匀后皮下注射;d70,采50ml外周血分离淋巴细胞。
[0127]
血清检测检测:a.将抗原用0.05m 碳酸盐buffer(ph9.6)稀释至2μg/ml,按100 μl/well,4℃包被过夜;
b.弃包被液,pbst 洗涤 3 次,每孔加入300
ꢀµ
l 5% 脱脂牛奶,37℃ 封闭 1 h;c.pbst洗涤3次,加入100 μl/孔的血清稀释液(从1:2000开始倍比稀释),37 ℃,孵育45 min;d.pbst洗涤5次,分别加入辣根过氧化物酶标记的羊抗alpaca二抗(用pbs按1:1w稀释),100 μl/孔,37 ℃ 孵育45 min;e.pbst洗板5次。加入tmb显色液显色,100 μl/孔,37℃,5 min,加入终止液终止反应,50 μl/孔,于 450 nm下测光密度。
[0128]
2)cdna合成提取pbmc总rna:用trizol保存的外周血淋巴细胞冰上溶解后转移至1.5ml的离心管,加入1/5体积的氯仿震荡混匀,室温静置5分钟后4℃,12000g离心15分钟;将离心后的上清液转移到新的离心管,往新离心管中加入等体积的异丙醇,室温静置10分钟后4℃ 12000g离心10分钟,用75%乙醇清洗沉淀,4℃ 7500g离心5分钟后弃去上清,沉淀室温晾干后溶解于适量的无rna酶的水中。从a260/280来分析rna提取纯度,准备rna转录。
[0129]
cdna合成:使用superscript
™ꢀ
iv first-strand synthesis system试剂盒,反转录得cdna并-80℃保存。
[0130]
3)vhh基因扩增使用正向引物(vhh-f)和反向引物(ch2-r)以pbmc cdna为模板,pcr扩增vhh-ch2基因,pcr反应条件如下:98℃预变性45秒后进入温度循环,98℃变性15秒,58℃退火20秒,72℃延伸45秒,循环30次,72℃最终延伸7分钟。pcr产物经1.5%琼脂糖凝胶电泳后,用胶回收试剂盒(promega)回收750bp的目的条带。使用正向引物(vhh-ch2-f)和反向引物(vhj-r)以vhh-ch2为模板,pcr扩增vhh基因,pcr反应条件如下:98℃预变性45秒后进入温度循环,98℃变性15秒,60℃退火20秒,72℃延伸45秒,循环30次,72℃最终延伸7分钟。pcr产物经1.5%琼脂糖凝胶电泳后,用胶回收试剂盒(promega)回收400bp的目的条带。
[0131]
4)vhh文库构建噬菌粒载体pcomb3xtt及vhh基因用sfii dna内切酶进行单酶切,50℃酶切16h,酶切后的pcomb3xtt载体经1%琼脂糖凝胶电泳后,用胶回收试剂盒(promega)回收4000bp载体片段。酶切后的vhh基因直接用胶回收试剂盒进行过柱回收(400bp)。将vhh基因用t4 dna连接酶试剂盒(invitrogen)连接到噬菌粒载体中,16℃连接过夜,取少量连接产物琼脂糖凝胶电泳检测连接效率。连接产物用meck milliporef 微孔滤膜进行脱盐。
[0132]
将上述连接产物加入自制的tg1电转化感受态中,然后使用电转仪进行电击转化。取出50μl菌液用pbs进行梯度稀释10
2-105倍。取10μl各梯度稀释液在amp/2yt plate上流线,37℃孵育过夜,以此计数并统计噬菌体抗体文库的大小。剩余的电转化后的细菌补2yt至500ml,加入含100μg/ml氨苄青霉素,30 ℃ 220rpm培养过夜。最后得到超过9e9的vhh免疫文库。电击转化后的抗体文库经过夜扩增,离心收集文库菌体,加终浓度20%甘油-80度保存。
[0133]
取冻存的部分天然抗体噬菌体展示文库菌种接种到2yt培养集中,接种密度为0.1od,将菌液置于37℃、220rpm条件进行培养,大约1.5小时后,菌液密度达到0.6od,此时加入20倍于菌体数的m13ko7噬菌体静置30分钟侵染,然后30℃、220rpm培养过夜。第二天,10000g离心菌液,收集培养上清,向培养上清中加入1/4体积的peg/nacl溶液(20%peg8000,
2.5m nacl),混匀后冰浴1小时。冰浴结束后,8000 g离心10分钟收集沉淀,10% glycerol/pbst溶解沉淀即得到vhh噬菌体展示文库。测od268,并将其分装至1.5ml离心管中,6od/管,-80℃保存。
[0134]
2. 淘选抗trop2 vhh抗体1) 重组人trop2蛋白生物素化用生物素化试剂盒(易锦生物)按试剂盒说明书,将重组人trop2蛋白的avi-tag进行生物素修饰,得到生物素化的trop2蛋白。取10μg上述生物素修饰的重组蛋白加入到经pbs洗涤3次的100μl 链霉亲和素磁珠(dynabeads 280)中,置于旋转摇床上,速度20转每分钟,室温偶联1小时后pbs洗涤3次。
[0135]
2) 封闭噬菌体文库及阴性磁珠取vhh lib一支,室温融化后,各加入200μl 5% bsa/pbst,置于旋转摇床速度20转每分钟,室温旋转封闭1小时,这些噬菌体为input1。同时取100μl未偶联蛋白的dynabeads 280,pbs洗涤3次后,加入1ml 1% bsa/pbs,按上述条件旋转孵育1小时。
[0136]
3) 封闭阳性磁珠向上述偶联trop2的磁珠中加入1ml 1% bsa/pbs 20转每分钟,室温旋转封闭1小时。
[0137]
4)阴性淘选为去除与磁珠相互作用的抗体,有必要进行阴性淘选。将经bsa封闭的噬菌体文库和未偶联抗原的磁珠混合,按上述条件旋转孵育1小时。孵育结束后将噬菌体磁珠混合物置于磁力架上,待磁珠都贴壁之后,将上清转移至新的ep管中。
[0138]
5) 阳性淘选将上述封闭后的偶联有trop2蛋白的磁珠加入到阴性淘选后的噬菌体上清中进行阳性淘选,20转每分钟,室温旋转封闭1小时。孵育结束后用1ml pbst(0.1% tween-20 in pbs)洗涤磁珠,重复洗涤10次。洗涤结束后加入1ml 100mm甘氨酸(ph2.0),置于旋转摇床上速度设定20转每分钟,旋转洗脱10分钟。洗脱结束后将ep管置于磁力架上,待磁珠都贴壁后将洗脱液转移至新的ep管中。向洗脱液中加入0.2ml 1m tris-hcl溶液(ph 8.0)进行中和。将中和后的洗脱液加入到30ml od600约为0.6的tg1菌液中静置侵染30分钟,然后加入20倍于菌体数的m13ko7噬菌体,静置侵染30分钟,最后加入100ml 2yt培养基及终浓度为100μg/ml的氨苄霉素和卡那霉素,30℃、220rpm培养过夜。第二天按上述收获噬菌体文库的方法收获噬菌体,此时得到的噬菌体为input2。
[0139]
6) 重复阳性淘选按上述淘选方法进行2次重复,即将input2进行下一轮阴性淘选和阳性淘选得到input3。不同之处在于input3进行淘选得到的洗脱液侵染tg1后,不加m13ko7,而是取10μl菌液用pbs进行梯度稀释,取103、104、10
5 三个稀释梯度各100μl菌液涂布2yt/amp平板,30℃培养过夜,剩余的菌液30℃、220rpm培养过夜。
[0140]
7) elisa筛选阳性抗体用牙签随机挑取上述平板中的tg1单克隆到800μl 含有10
×
自诱导2yt/amp的深孔板中,深孔板粘覆透气膜,37℃、220rpm培养3小时后,30℃、220rpm培养过夜。elisa 板中包被重组人trop2蛋白,每孔100ng。次日,先在深孔板中取50μl保菌,其余的4000rpm离心10
分钟,去除孔内培养基保留菌体沉淀,每孔加入100μl tes溶液(20%蔗糖、0.1mm edta、50mm tris-hcl,ph 8.0),震荡使菌体重新悬浮后冰浴30分钟后再加入200μl超纯水震荡混匀30min,震荡结束后4000rpm离心10分钟,此时深孔板中的上清溶液即为含有抗体的周质腔提取物。用洗板机清洗elisa板三次,然后加入200μl 1% bsa/pbs,37℃封闭1小时。去除elisa板中的封闭液,加入100μl上述周质腔提取物,37℃孵育1小时,用洗板机清洗3次,加入100μl chicken anti-ha hrp(1% bsa/pbs),37℃孵育1小时,用洗板机清洗3次,加入100μl tmb显色液,37℃显色10分钟,加入100μl stop solution终止液。用酶标仪读取 od450值,将读值高于背景值3倍的克隆进行桑格测序,获得抗体的基因序列。
[0141]
8) 验证阳性克隆根据测序结果,选取抗体cdr3氨基酸序列差异较大的克隆重新接种并诱导过夜,按上述elisa方法再次验证所选克隆能否结合trop2。最终得到1a4和1f2等抗体序列。1a4的重链可变区氨基酸序列如seq id no:4所示,1f2的重链可变区氨基酸序列如seq id no:5所示。
[0142]
实施例3:含抗人trop2嵌合抗原受体元件的逆转录病毒原液制备1.靶向人trop2抗原的car的制备基因合成或克隆含抗人trop2抗原的单域抗体vhh,铰链区、跨膜区和胞内信号段的嵌合抗原受体序列,其结构如图2所示。根据装载vhh的不同,将嵌合抗原受体分别命名为1a4-bbz和1f2-bbz,其氨基酸序列分别如seq id no:11和seq id no:12所示,核苷酸序列分别如seq id no:13和seq id no:14所示。
[0143]
以逆转录载体msgv为骨架载体,构建了表达1a4和1f2克隆的嵌合抗原受体的逆转录病毒质粒。挑选测序正确的克隆,接种菌液至200ml 2yt培养基中,过夜摇菌,并按照nucleobond xtra maxi ef试剂盒说明书完成大提质粒。
[0144]
2.逆转录病毒包装用阳离子聚合物pei包装逆转录病毒,流程如下:分别用无血清600μl dmem稀释36μl pei和逆转录病毒包装质粒(病毒主质粒6μg、gag-pol3.8μg、vsvg1.5μg);然后将pei/dmem加入质粒/dmem混合物,涡旋震荡混匀,在室温下静置15分钟;将质粒-pei复合物加入预先铺板的293t细胞。转染后16h换液,在48h后收集第一次病毒上清,72h后收集第二次病毒上清,并用0.45μm滤器过滤,分装至1.5ml离心管,1ml/管,-80℃保存备用。
[0145]
实施例4:trop2car-t细胞的制备和car阳性率测定1. pbmc分离与活化领取一支pbmc,核对患者个体识别码无误后,进行复苏。用x-vivo完全培养基调整细胞密度为1
×
106/ml。将复苏后恢复过夜的pbmc,轻柔吹打,用70μm细胞筛网过滤后,转入50ml离心管中,室温离心,1500rpm,离心5min,弃上清。用适量的dpbs重悬细胞,取20μl与台盼蓝1:1混匀后计数,计算活率、cd3+细胞数,然后取所需体积的细胞,室温1500rpm,离心5min,弃上清,用于分选。按照cd3/cd28磁珠与cd3+细胞1:1比例,计算磁珠用量,磁珠量=【cd3+细胞数/4
×
105】μl。清洗磁珠:取无菌流式管一支,加入2ml的dpbs,同时加入磁珠,磁力架上静置1min后,弃上清。将流式管移除磁力架,取与细胞悬液等体积的dpbs或x-vivo15重悬,加入细胞悬液中,磁珠和细胞于15ml离心管混合后,在旋转混合仪上孵育。室温孵育30min。孵育完成后,轻柔将细胞转移至无菌流式管中,用1ml dpbs冲洗15ml离心管,冲洗液
并入同一流式管中。将无菌流式管移至磁力架上,静置1min后,吸弃未吸附的液体。将无菌流式管移出磁力架,用1ml car-t培养基重悬细胞,并用car-t培养基冲洗管壁2次,全部收集转移至同一离心管内。用car-t培养基调整细胞密度为 1
×
106ml,添加il-2 终浓度为200iu/ml,置37
˚
c,5% co2培养箱培养两天。
[0146]
2.病毒原液感染与培养将活化后的t细胞调整为5
×
105/ml,在24孔板中分别加入1ml t细胞和1ml病毒原液,每孔加2μl polybrene,32℃,2500rpm,离心1.5h。弃去上清液,每孔加入1ml t细胞培养基(含il-2 300iu/ml)。将培养板置于37℃,5% co2培养箱中培养。感染后24h,转至6孔板,每天观察细胞的密度,适时补加含il-2 300iu/ml的t细胞培养液,使t细胞的密度维持在1
×
106/ml左右,使细胞扩增。
[0147]
3.car阳性率检测逆转录病毒感染的t淋巴细胞在病毒感染72h后检测car阳性率。针对含1a4和1f2克隆的嵌合抗原受体组和阴性未感染对照组nt,分别取1
×
106个细胞,离心去除培养基,用500μl pbs洗细胞一次后用100μl重悬置于流式上样管中(bd)。加入生物素-标记的trop2抗原(1:200)四度孵育30分钟。pbs洗细胞一次后按1:100加入二抗pe-sa链霉亲合素(biolegend)四度避光孵育30分钟。500μl pbs洗细胞后用200μl pbs重悬细胞并上机检测,car-t阳性率流式分析结果如图3所示。
[0148]
实施例5:基于抗人trop2car-t细胞功能分析1.抗人trop2car-t细胞cd107a表达分析将含不同抗体克隆的car-t细胞分别与靶细胞(trop2阳性表达的人胰腺癌细胞系bxpc3)、对照靶细胞(人神经胶质细胞瘤细胞 u251)按1:1的效靶比(效应细胞和靶细胞均为3
×
105个)混合置于37℃,5% co2培养箱中共孵育4小时后,流式检测各组样品中表达cd107a的细胞分别占cd3+细胞数的比例。评价car-t细胞在受到靶细胞刺激后的脱颗粒反应。cd107a表达的流式分析结果如图4所示。
[0149]
2.抗人trop2car-t细胞细胞因子分泌能力检测将含不同抗体克隆的car-t细胞分别与靶细胞(trop2阳性表达的细胞系bxpc3)、对照靶细胞(trop2阴性表达的细胞系u251)分别按10:1、2:1的效靶比(靶细胞为3
×
104个)共孵育24小时后,收集其上清,利用elisa(酶联免疫)方法检测ifn-γ和il-2的分泌情况。ifn-γ和il-2检测使用爱必信试剂盒检测(human ifn-gamma elisa kit,human il-2 elisa kit),实验步骤依据产品说明书进行。ifn-γ 分泌的检测结果如图5,图6所示。il-2分泌的检测结果如图7,图8所示。
[0150]
3. 抗人trop2car-t细胞毒性实验car-t杀伤毒性实验通过检测car-t细胞体外对靶细胞的杀伤效果来评估car-t细胞的体外功能。以不同效靶比(以3
×
104个靶细胞为基准,效靶比分别为10:1 和2:1)将t细胞分别与稳定表达萤火虫荧光素酶的trop2阳性靶细胞bxpc3-luc-gfp,以及trop2阴性对照靶细胞u251-luc-gfp共同培养,同时设置只有靶细胞的阳性对照。37℃过夜孵育后在培养体系中加入100μl萤光素酶反应底物,检测荧光值,通过以下公式计算杀伤效率:杀伤效率=(阳性对照孔荧光值-实验孔荧光值)/ 阳性对照孔荧光值)
×
100%。实验分组及分析结果如图9,图10所示。
[0151]
以上所述仅为本发明的优选实施例而已,并不用于限制本发明,对于本领域的技术人员来说,本发明可以有各种更改和变化。凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1