一种提高橙花瑞香有性繁殖成功率的方法与流程
本发明属于生物繁殖技术领域,尤其涉及一种提高橙花瑞香有性繁殖成功率的方法。
背景技术:
座果率低是种子植物有性繁殖过程中的一个致命障碍,限制了物种的延续和扩散。植物的座果率与传粉有着密切联系,传粉是种子植物繁殖的必经阶段(吴佩纹等,2016)。通常成功的传粉需要满足以下条件:1、花粉和柱头均具有正常的结构和功能;2、拥有足够的传粉媒介及充足的访花频率保证足够数量花粉在柱头的落置;3、柱头和花粉之间能成功地相互识别(吴佩纹等,2016)。研究发现,木本植物更容易表现出座果率低的现象(,因为该类植物一般个体较大,开花较多,一方面会受到资源限制,另一方面会受到花粉限制,特别是对于自交不亲和的物种,传粉昆虫在同一植株上大量访花,直接造成大量的同株异花和自花花粉而较少或没有异株花粉在柱头落置,导致座果率和结籽率低。瑞香科(Thymelaeaceae)植物具有重要的经济和观赏价值,其中瑞香属(DaphneLinn.)约有95种,主要为落叶或常绿灌木或亚灌木,是木本植物并表现出座果率低的特征。瑞香属植物主要分布于欧洲经地中海、中亚到中国、日本,南到印度至印度尼西亚。中国资源丰富,有44种,多分布在云南、四川、贵州和甘肃等省,如橙花瑞香(DaphneaurantiacaDiels)、甘青瑞香(DaphnetanguticaMaxim)、白脉瑞香(DaphneesquiroliiLevl)和短瓣瑞香(DaphnefeddeiLevl)等。本属中多种植物具有重要的经济和药用价值,为我国民间常用中药,如最具代表性的祖师麻(瑞香属植物的根皮和茎皮)为民间治疗风湿和止痛的常用中药,具疏风通络,祛瘀止痛之功效,可用于治疗头痛,牙痛,风湿关节痛等诸多痛症,并取得了良好的疗效;瑞香提取物瑞香素(daphnetin)具有降血脂,抗溶血活性,抗肿瘤活性和强镇痛等作用。此外,瑞香属植物株形优美,花色多变,从黄色至紫色不等,管状小花,花型美观,香味浓郁,观赏性强,具有重要的观赏和利用价值。如被作为绿化树种开发利用的甘青瑞香(;被培养为名贵盆栽品种的中国传统名花金边瑞香。然而由于长期利用过程中存在着过度不合理利用,导致部分野生资源急剧减少,原材料紧缺。瑞香属植物在自然状态下主要靠种子繁殖,而研究发现,座果率低在本属中较为常见,是瑞香属的一个特征。该特征限制了对本属植物的进一步开发利用,对其原因却尚不明确,相关研究也很少,尤其是在我国,无人开展。
综上所述,现有技术存在的问题是:瑞香属植物具有重要的经济和观赏价值,其应用和市场前景广阔,而自然条件下的低座果率也就意味着其自然繁殖能力低下,如本发明对象橙花瑞香,其自然座果率仅为1.4%,严重限制了物种的繁殖扩散,也严重影响着对物种的进一步开发利用,相关问题至今基本无人研究,也没有可行的解决方案提出。瑞香属植物在自然条件下低座果率,其原因尚不明确,且研究少。
技术实现要素:
针对现有技术存在的问题,本发明提供了一种提高橙花瑞香有性繁殖成功率的方法。
本发明是这样实现的,一种提高橙花瑞香有性繁殖成功率的方法包括以下步骤:
在盛花期分别选取花苞状态的花进行套袋处理;待袋内的花苞开放后再进行人工授粉处理;包括:人工自交处理、人工异交处理、自动自花处理、对照处理。
所述提高橙花瑞香有性繁殖成功率的方法进一步包括:
去雄操作:使用针头型镊子从花苞的顶端、花瓣缝隙处垂直小心深入花苞内部,将上、下轮的各四个花药分别逐一轻轻拔出,再套袋处理。
所述提高橙花瑞香有性繁殖成功率的方法进一步包括:
授粉操作:将细毛笔用剪刀修剪,使其可自由深入到橙花瑞香的花冠管中,将准备的花粉,授在柱头上。
进一步,人工自交处理,取自花或同株异花的花粉作为花粉供体,授在柱头上,然后套袋直至花凋谢;
人工异交处理,将花苞状态的花去雄并套袋,待花开放后,随机选择20米之外10株植物的花粉至于离心管中混合,将混合的异花花粉授到柱头上,套袋直至花凋谢;
自动自花处理,一直套袋直至花凋谢;
对照处理,自然状态,不去雄,不套袋,使其授粉;一个植株内不同的处理用不同颜色的尼龙线标记;整个授粉处理植株内,同一种处理用同一种颜色的尼龙线标记。
本发明表明橙花瑞香的花部结构特殊,管状小花,花药两轮,雌雄蕊分离。花开后的花粉具有活性,柱头具有可授性,扫描电镜下,柱头和花粉的结构没有发育异常,且柱头上有花粉落置。橙花瑞香的传粉者主要是夜间访花的蛾类,访花频率低。P/O及人工授粉实验表明橙花瑞香的繁育系统为专性异交。橙花瑞香的座果率非常低,自然座果率为1.4%,人工异花授粉为23.3%,低座果率可能是受其开花量大、异花花粉限制、资源限制以及花部结构等因素的影响。
本实验以橙花瑞香(Daphne aurantiaca Diels)为实验材料,拟开展以下研究内容:1、花部特征的调查,是否存在花部结构的特殊性;2、柱头、花粉结构和功能的调查,是否存在正常的雌性和雄性功能3、繁育系统及亲和特性的调查,是否为自交不亲和或自交亲和;4、传粉者的行为和影响调查,是否存在花粉限制。通过对橙花瑞香有性繁殖及传粉特性的研究,探索其自然座果率低的原因,为瑞香属植物种质资源的利用、保存及扩繁等提供理论依据。
附图说明
图1是本发明实施提供的提高橙花瑞香有性繁殖成功率的方法流程图。
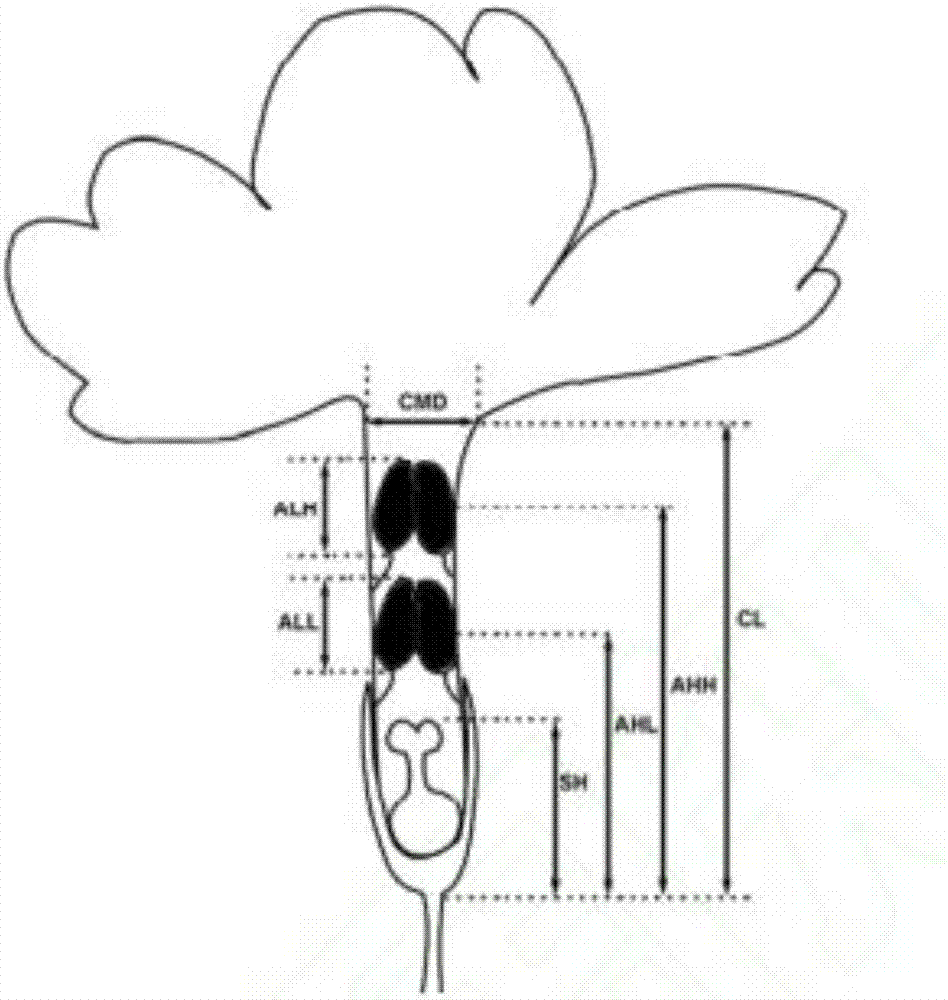
图2是本发明实施提供的花部特征测量示意图,SH,柱头高;AHL,下轮花药高;AHH,上轮花药高;CL,花冠长;ALL;下轮花药长;ALH,上轮花药长;CMD,开口大小图。
图3是本发明实施提供的MTT染色后花粉活性图。
图4是本发明实施提供的花粉、柱头的扫描电镜观察(A-D),A以单粒形式存在的花粉;B单粒花粉圆形;C圆柱状柱头;D柱头的乳突结构及乳突间的花粉粘附。
具体实施方式
为了使本发明的目的、技术方案及优点更加清楚明白,以下结合实施例,对本发明进行进一步详细说明。应当理解,此处所描述的具体实施例仅仅用以解释本发明,并不用于限定本发明。
瑞香属植物具有重要的经济和观赏价值,其应用和市场前景广阔,而自然条件下的低座果率也就意味着其自然繁殖能力低下,如本发明对象橙花瑞香,其自然座果率仅为1.4%,严重限制了物种的繁殖扩散,也严重影响着对物种的进一步开发利用,本发明通过人工授粉的方法可以提高其座果率,创造出更多的下一代资源,既增大了物种的适合度,也可以人为根据需要进行交配设计,甚至是种间杂交的尝试,这将大大提高橙花瑞香乃至整个瑞香属的利用水平,培育出更优良的下一代,应用于生产和生活。
下面结合附图对本发明的应用原理作进一步描述。
如图1所示,本发明实施例提供的提高橙花瑞香有性繁殖成功率的方法包括以下步骤:
S101,花部特征调查;
野外选择橙花瑞香20株,测量其株高、冠幅的长和宽,并统计其分支数及每支开花数。为了调查橙花瑞香的花部结构,在盛花期随机选取40株野生橙花瑞香个体,每株选取正在开放的单花一个,用数显游标卡尺测其特征值,测量指标包括:开口大小(CMD),花冠长(CL),柱头高(SH),上轮花药高(AHH),上轮花药长(ALH),下轮花药高(AHL),下轮花药长(ALL),具体测量操作见图2。所测得的平均值作为该花性状的表型值。
S102,花粉及柱头特性;
(1)花粉活力及柱头可授性测定
花粉活力测定采用MTT染色法(Dafni,1992;左丹丹等,2007;张超仪和耿兴敏,2012),随机选取25株橙花瑞香,每株上在花开之前套袋处理一个花序,待套袋内花开到盛花期以后,每株上摘取1-2朵花(开花第二至第三天),带回实验室测花粉活性。首先取少量花粉置于载玻片上,加1~2滴MTT溶液,盖上盖玻片。10min后在低倍镜下观察。选取5个视野,每个视野花粉数在30粒以上,取其平均值作为花粉的活力指标。凡是染成深红色甚至紫黑色的花粉,其生活力为强,红色的次之,无色的为不具活力的花粉或不育花粉。花粉活力比率=某视野变色的花粉粒数目/该视野花粉粒总数×100%。
柱头可授性测定采用联苯胺-过氧化氢法(王育青等,2010):随机选择25株处于盛花期的橙花瑞香(开花第二或第三天),每一株上选择1-2朵花,将柱头取下浸入凹面载玻片含有联苯胺-过氧化氢反应液(1%联苯胺:3%过氧化氢:水=4:11:22,体积比)的凹陷处。若柱头具可授性,则柱头周围呈现蓝色并有大量气泡出现。根据蓝色深浅和气泡多少表示可授性强弱。
(2)花粉及柱头扫描电镜观察
为了观察橙花瑞香的花粉及柱头的形态结构,随机收集处于盛花期的花10朵(分别来自不同植株),用FAA固定保存带回实验室,经清洗、酒精逐级脱水(70%,80%,90%,100%,100%)、用双面胶分别将柱头和花药均匀地粘附在金属台面上,在离子溅射仪上对金属台面喷金镀膜(贾子瑞等,2014),扫描电镜下观察花粉和柱头的形态。
S103,繁育;
(1)花粉、胚珠产量测定
野外收集15朵来自不同植株的花药未开裂的花,分别盛放于70%酒精的2ml离心管中保存,带回实验室。在实验室,将每朵花的上下轮共8个花药取出,放在小研钵中充分研碎,在容量瓶中稀释至5ml,混匀,用移液器随机吸取5滴(每滴50μL)放于载玻片上,在100×显微镜下数花粉的数量,将五滴的平均值乘以稀释倍数100,即为该花的花粉产量(Dafni,1992)。将子房放在体视显微镜下解剖,并将胚珠均匀散开,统计胚珠数量。
(2)人工授粉实验
在盛花期分别选取花苞状态的花进行套袋处理。待袋内的花苞开放后再进行人工授粉处理:1、人工自交处理,取自花或同株异花的花粉作为花粉供体,授在柱头上,然后套袋直至花凋谢;2、人工异交处理,将花苞状态的花去雄并套袋,待花开放后,,随机选择20米之外10株植物的花粉至于离心管中混合,将混合的异花花粉授到柱头上,套袋直至花凋谢;3、自动自花处理,一直套袋直至花凋谢;4、对照处理,自然状态,不去雄,不套袋,使其授粉。一个植株内不同的处理用不同颜色的尼龙线标记。整个授粉处理植株内,同一种处理用同一种颜色的尼龙线标记。
S104,传粉者观察;
为了观察到橙花瑞香有效的传粉者以及计算访问频率,在盛花期随机选取橙花瑞香一株,统计正在开放的花数,以30min为一个时间段,从早上9:00至下午5:00记录来访昆虫的种类,访花数量,访花行为及访问单花的时间。当访花虫体接触到花药即为一次有效的访问。晚上访花观察从19:00至21:00,用红色的布罩着手电筒以减少光线对蛾类访花的干扰(Liu&Huang,2013),同白天观察方法一样,记录夜间访花者的种类和行为。访花观察包括60个时间段,30小时。
S105,数据分析;
所有数据统计分析、作图均采用SPSS19.0软件,统计数据用平均值±标准差表示,N代表样本量。
本发明提供的实验材料为:本发明以橙花瑞香为实验材料,橙花瑞香(AphneaurantiacaDiels),别名:云南瑞香、黄花瑞香、万年青,是瑞香科、瑞香属矮小灌木,高0.6-1.2米,多分枝,枝短,幼时红褐色或褐色,老时棕褐色或褐色。生长于海拔2600-3500米的石灰岩阴坡杂木林中或灌丛中。叶小,对生或近对生,密集簇生于枝顶,纸质或近革质。花橙黄色,管状小花,芳香,2-5朵簇生于枝顶或部分腋生,偶有结实,果实为长椭圆形。产自四川西南部(乡城、会东)、云南西北部丽江-香格里拉一带。盛花期每植株的开花数可达几百甚至上千。5月上中旬始花,5月末至6月初进入盛花期,7月上中旬终花,总体花期长达近两个月,单朵花的开放时间平均为11天。本研究野外实验部分是2015年5-6月在云南省迪庆藏族自治州香格里拉县高山植物园(27°54′50″N,99°38′17″E,海拔3300m)开展,座果率统计在当年8月底至9月初完成,所选橙花瑞香植株均为野生分布,单株散生,主要生长在峨眉蔷薇、荀子等的林下碎石堆里或位置较高不易积水的石灰质斜坡地段,伴生种有报春花科刺叶点地梅、玄参科二歧马先蒿等矮小草本花卉。
1、结果与分析
1.1植株及花部特征调查
橙花瑞香的株高平均为32.2cm,冠幅长为73.8cm,冠幅宽为51.3cm平均每株分支数大于50,每分支开花数平均为12朵,因此盛花期整株开花数可达几百甚至上千,开花量大。橙花瑞香的花较小,管状结构,花冠管长度平均为6.76mm(表1),花开口较小,平均1.27mm,柱头位于花冠管的下部,花药两轮位于柱头的上方,下轮着生于花萼筒的中部(高约4.8mm,花药长1.43mm),上轮着生于花萼筒的喉部稍下面(高约7mm,花药长1.32mm),花药顶端可与花冠管顶部齐平或略有伸出,花丝短,两轮花药长度之间存在显著差异(P<0.01,两个配对样品的t检验)。
表1橙花瑞香的花部特征测量(平均值±标准差)
Table1thefloraltraitsmeasurementofD.Aurantiaca(Mean±SD).
1.2花粉及柱头特性
1.2.1花粉活力及柱头可授性测定
MTT染色观察表明,所测的25朵花的花粉活力波动范围较大,主要集中在35-75%之间,花粉活力平均值为57±18%(图3)。
联苯胺-过氧化氢法测定柱头可授性,发现柱头周围被染成蓝色近黑色并有气泡产生,说明柱头具有可授性。
1.2.2花粉及柱头扫描电镜观察
扫描电镜观察表明橙花瑞香的花粉均以单粒形式存在,圆形(图4A),大小在十几至二十微米之间。花粉外壁纹饰为穿孔型,穿孔主要为不规则多边形或圆形,穿孔的频度较大,与龙胆的花粉相似(王臣等,2005)(图4B)。扫描电镜下观察柱头为圆柱形,上面密布长长的乳突结构(图4C),且乳突之间有花粉粘附,电镜下观察,花粉已经萌发干瘪(图4D),说明花粉落置在柱头上是可以萌发的。
1.3繁育系统
1.3.1花粉、胚珠产量
橙花瑞香的单花花粉产量平均为8059粒,而其胚珠数只有1,因此其P/O平均值为8059。根据Cruden(1977)的标准:P/O为2.7~5.4时,其繁育系统为闭花受精;P/0为18.1~39.0时,繁育系统为专性自交;P/O为31.9~396.0时,繁育系统为兼性自交;P/O为244.7~2588.0时,繁育系统为兼性异交;P/O为2108.0~195525.0时,繁育系统为专性异交。橙花瑞香的的P/O值为8059,据此标准可判断其繁育系统为专性异交。
1.3.2人工授粉
无论是人工自花授粉还是自动自花授粉的座果率均为0(表2),说明橙花瑞香是严格的自交不亲和,为异花授粉,需要传粉媒介辅助授粉。人工异花授粉的座果率是23.3%,而自然授粉的座果率只有1.4%,因此与自然座果率相比,人工异花授粉提高了近17倍,说明自然状况下,橙花瑞香存在着异花花粉限制。
表2人工授粉的座果率统计
Table2fruit-setratioofhandpollination
1.4传粉者观察
长达30小时以上的访花观察发现,橙花瑞香的整体访花者较少,偶尔有食蚜蝇和蝇类访问,但在花上驻留的时间短,且它们的喙部也较短,而橙花瑞香的花管较长且柱头位于花管基部,所以食蚜蝇和蝇类不能为橙花瑞香传粉,为无效的传粉者。而夜间访花的蛾类虽然访问频率也比较低(夜间访花观察期间仅看到两只,一只访问3朵花,另一只访问2朵花),但蛾类因其喙部较长,可以同时接触到橙花瑞香的花药和柱头,并为其传粉,被视为有效的传粉者。但蛾类一般在同一株上访问数朵花,这也容易造成自花花粉在柱头的落置。
下面结效果对本发明作进一步描述。
本实验结果表明,自然条件下橙花瑞香座果率很低,只有1.4%,大部分花在花期结束后脱落,偶有结实。这和前人对瑞香属其他物种的研究结果相一致(Alonso&Herrera,2001;Alonsoetal,2007;Roccotielloetal,2009;Rodríguez-Pérez&Traveset,2011)。盛花期橙花瑞香开花量大,单花可以维持11天左右,无论是在扫描电镜下看花粉和柱头的形态,还是MTT染色法测花粉的活力以及联苯胺-过氧化氢法测柱头的可授性都可以看出柱头和花粉不存在发育异常,有正常的结构和功能。花粉在开放初期具有较高的活性,柱头则具有较强的可授性,说明橙花瑞香具有正常的雄性和雌性功能。访花观察发现,橙花瑞香的传粉者很少,白天偶尔见到蝇类和食蚜蝇活动,而且它们是无效的传粉者。晚上有蛾类访花,可以起到传粉作用,但是访问频率非常低。相似的,瑞香属D.rodriguezii的访问者也非常稀少,偶尔有小型蚂蚁(Plagiolepispygmaea)访问一次或金龟子(Tropinotahirta)啃食花药和花被片;对于D.gnidium,它的访问者多一些,大部分属于膜翅目和双翅目,然而自然状态下,D.rodriguezii和D.gnidium的座果率没有显著性差异,都比较低在5%以下(Rodríguez-Pérez&Traveset,2011)。说明访花昆虫并没有提高植物的座果率,可能是因为它们在同一株上连续访问数花,带来的都是自花花粉,而本属植物大多是自交不亲和的。扫描电镜下可以看到柱头上有较多的花粉粘附(未发表数据),且花粉存在萌发后的干瘪现象,猜测自花花粉可以在柱头上萌发,但后续果实发育不良并最终脱落。这和Alonso(2004)对D.gnidium和D.rodriguezii的研究相似,虽然花粉可以萌发,但座果率跟花粉萌发没有相关关系,因为这些花粉主要为自花花粉,果实在发育过程中大部分(约75%)会发生胚胎塌陷(embryocollapse)(75%未成熟的果实死掉,),这主要由近交衰退造成(Roccotielloetal,2009;Rodríguez-Pérez&Traveset,2011)。因此传粉者稀少,异花花粉限制是造成橙花瑞香自然座果率低的一个重要原因。
花作为被子植物的繁殖器官,具有比其它生物繁殖器官更高的变异性,多样的花部类型是植物为了适应其生境,使得物种能够顺利完成繁殖过程并延续下去而不断进化形成的(Dafni,1996;Huang&Guo,2002,钱一凡等,2015)。对橙花瑞香花部特征的调查发现橙花瑞香是典型的管状花结构,花药两轮分别位于花冠管的中部和上部,柱头位于花冠管的下部,实现了花药和柱头在空间上的完全分离,属于柱头缩入式雌雄异位(张大勇,2004),这种结构类似于经典三型花柱的短柱型结构,管状结构限制了传粉者进入花中取食花蜜或花粉的行为及花内柱头、花药与传粉者身体不同部位的接触,对于三型花柱的植物而言,这种空间结构上的分离有助于促进花粉在同一高度水平的柱头和花药间传递,既可以促进异交传粉又可以避免自花花粉浪费(Webb&Lloyd,1986;Liu,etal,2015),但对于只有一种花部形态的橙花瑞香来说却可能是一种低效的花形结构,因为限制了异花花粉在柱头的落置并最终影响生殖成功。对瑞香属D.gnidium的研究发现,其访花者主要是鳞翅目和膜翅目昆虫,而套袋和不套袋处理都产生相等数量的果实,说明传粉者对这一特殊繁殖系统的生殖成功没有起到主角作用,其花部综合特征是一种低效的结构(Herrara,1987;Roccotielloetal,2009)。
根据cruden(1977)的标准可知,橙花瑞香的繁育系统是专性异交的,这和人工授粉实验的结果相一致。然而人工授异花花粉,橙花瑞香的座果率也只有23.3%,该结果和瑞香属其它种的人工授粉结果相似,D.rodriguezii人工授异花花粉的座果率不到25%,D.gnidium的不到15%(Rodríguez-Pérez&Traveset,2011),可见仅仅近交衰退理论不能解释其低座果率(Rodríguez-Pérez&Traveset,2011)。两种假说可以解释这一现象,(1)资源限制假说(Willson&Schemske,1980;Yang,etal,2005;Rodríguez-Pérez&Traveset,2011;Arnanetal,2014)认为有足够数量有效花粉,但依然不能完全结实,这种情况通常是由于资源供应的有限性所致(McCall&Primack,1987)。资源限制是座果率低的另一重要原因,事实上,自然界中植物座果率低单一受花粉或者受资源限制的情况很少发生,大多情况下两者共同起作用(Yangetal,2005;Kleinetal,2015)。(2)性别选择假说(Bateman,1948;Bawa&Webb,1984),认为大量花在花期结束后脱落,这些脱落的过量花是为了增大雄性生殖成功,这一现象是由内部基因决定的,增大花粉扩散的选择压力以及受精卵的父系不确定性是导致潜在败育发生的主要因素。座果率和结籽率更多的取决于性别选择的强度,而不是传粉者或资源的限制(Bawa&Webb,1984)。Lloyd(1980)认为对花数量和果数量的调控是对母本投资的调控,目的是为了与可利用资源更好的匹配。
总之,橙花瑞香的低座果率不是一个偶然的存在,跟其开花量大、传粉者稀少、花部结构特殊、自交不亲和等有密切关系。虽然人工授粉可以提高其自然座果率,但该方法耗时耗力,且效率非常低,在野外大规模开展很难实现。要想提高座果率,寻找并驯化一种合适的传粉昆虫可能是一条可行的选择,如熊蜂类,具有长长的喙,能够为瑞香属花传粉(已在另一种野生瑞香中观察到),而且熊蜂作为驯化的传粉昆虫已在温室、大棚等生产中应用,并取得了不错的效果。如引用明亮熊蜂为温室草莓授粉(李继莲等,2006);熊蜂为温室茄属作物授粉(黄家兴等,2007)等。然而如何将熊蜂成功的应用在橙花瑞香的野外传粉上,还需进一步的研究和试验。
以上所述仅为本发明的较佳实施例而已,并不用以限制本发明,凡在本发明的精神和原则之内所作的任何修改、等同替换和改进等,均应包含在本发明的保护范围之内。
- 还没有人留言评论。精彩留言会获得点赞!