一种高光效多倍体小麦的筛选方法与流程
本发明涉及农业种植技术领域,尤其涉及一种高光效多倍体小麦的筛选方法。
背景技术:
远缘杂交和多倍体化是高等植物最重要的进化过程,在天然物种和农作物中广泛存在。研究表明,70%以上的被子植物在进化过程中至少经历了一次多倍化事件,近年来通过对芸苔属、小麦、拟南芥、棉花等植物的研究发现,异源多倍体形成早期经常伴随着与孟德尔法则不一致的基因组变化,表现在遗传、转录、表达等水平上它们有助于多倍体新物种形成,为植物的长期进化提供了可能的条件。
多倍体小麦是通过异源多倍体化方式进化和培育的典型,特别是六倍体普通小麦是世界范围种植最广泛的、最重要的粮食作物之一,其产量随着材料的进化得了显著的提高。然而现代栽培小麦由于长期的品种之间杂交育种,特别是核心种质的反复利用,导致了普通小麦群体遗传多样性的丧失,遗传基础越来越窄。作为大多数栽培小麦的起源,近缘的小麦基因型是小麦改良中获得新的遗传多样性最有价值、最易得到的来源,因此对小麦进化材料进行研究与利用可以极大的丰富育种手段,提高对环境的适应性与抗性。
光合作用是植物的物质积累与生理代谢的基本单元,光合效率的高低与作物光合产物积累、产量潜力发挥以及品质优劣密切相关。光合遗传研究是利用作物光合特性进行杂种优势评价的一种新方法,是作物遗传育种研究中的一个新领域。因此研究作物的光合性状在农业育种工作中具有重要的意义。目前,小麦的常规育种手段已经发挥到最大作用,但产量的提高不是很明显,筛选高光效材料、运用现代育种方法进行高光效育种是当今以及将来提高作物产量的有效途径。因此,亟待研究一种科学且准确的筛选高光效多倍体小麦的方法。
技术实现要素:
有鉴于此,本发明实施例提供了一种高光效多倍体小麦的筛选方法。
为达到上述目的,本发明主要提供了如下技术方案:
一方面,本发明实施例提供了一种高光效多倍体小麦的筛选方法,包括以下步骤:
在小麦发育灌浆期分别测定不同倍性小麦旗叶的光合气体交换参数、光合相关性状及光合抗逆能力;通过分析测定数据从所述不同倍性小麦中筛选出最佳高光效多倍体小麦;
其中,所述光合气体交换参数包括净光合速率、气孔导度、胞间co2浓度、蒸腾速率及水分利用效率;
所述光合相关性状包括小麦旗叶叶片叶绿素浓度、叶面积指数、旗叶面积、绿叶面积、干物质积累量及绿叶数;
所述光合抗逆能力为不同光照强度、不同co2浓度、不同温度及不同湿度条件下的光合特性;所述光合抗逆能力包括光饱和点、光补偿点、co2饱和点、co2光补偿点、最佳湿度、最佳温度、暗呼吸速率、光量子效率,羧化效率、最大净光合速率及平均净光合速率。
作为优选,所述不同倍性小麦为二倍体节节麦、四倍体硬粒小麦、六倍体普通小麦及八倍体小黑麦。
作为优选,所述不同光照强度条件下光合特性的测定方法为:选用6400-02bled红蓝光源叶室,控制叶室内温度为25℃,co2浓度控制为400μmol·ml-1,在1300μmol·m-2·s-1光强下适应15分钟后,采用自动光曲线程序进行测定;其中,计算光饱和点、光补偿点、暗呼吸速率、光量子效率和最大净光合速率。
作为优选,所述不同co2浓度条件下光合特性的测定方法为:控制叶室内温度为25℃,co2浓度设为1200μmol·ml-1,光强设为1300μmol·m-2·s-1,适应15min后采用自动co2曲线测定程序进行测定;其中,计算光饱和点、co2饱和点、co2光补偿点、光补偿点、光量子效率、羧化效率、最大净光合速率及平均净光合速。
作为优选,所述不同温度条件下光合特性的测定方法为:测定时控制光强为1300μmol·m-2·s-1,co2设为400μmol/mol,25℃下适应15min后,采用手动程序进行测定,测定时控制叶室温度,分别在外界温度的±10℃范围内每个温度点测定一次,确定最佳温度。
作为优选,所述不同湿度条件下光合特性的测定方法为:测定时控制光强为1300μmol·m-2·s-1,co2设为400μmol/mol,25℃下适应15min后,采用手动程序进行测定,测定时调节干燥管旋钮来控制湿度,从湿度为零开始,每间隔5%设置一个湿度点,在每个湿度点测定一次,确定最佳湿度。
作为优选,所述光合气体交换参数的测定结果:
所述净光合速率从大至小依次为:八倍体小黑麦﹥四倍体硬粒小麦﹥二倍体节节麦﹥六倍体普通小麦;
所述蒸腾速率从大至小依次为:八倍体小黑麦﹥二倍体节节麦﹥四倍体硬粒小麦﹥六倍体普通小麦;
所述水分利用效率从大至小依次为:八倍体小黑麦﹥六倍体普通小麦﹥四倍体硬粒小麦或二倍体节节麦;
所述胞间co2浓度:六倍体普通小麦和八倍体小黑麦均小于二倍体节节麦或四倍体硬粒小麦。
作为优选,所述光合相关性状的测定结果:
所述旗叶面积从大至小依次为:六倍体普通小麦>四倍体硬粒小麦>八倍体小黑麦>二倍体节节麦;
所述绿叶面积从大至小依次为:六倍体普通小麦>四倍体硬粒小麦>八倍体小黑麦>二倍体节节麦;
所述叶面积指数从大至小依次为:六倍体普通小麦>四倍体硬粒小麦>八倍体小黑麦>二倍体节节麦;
所述六倍体普通小麦的绿叶数为4片,其他三个小麦绿叶数为3片;
作为优选,在相同光照强度条件下,所述净光合速率从大至小依次为:
四倍体硬粒小麦>六倍体普通小麦>八倍体小黑麦或二倍体节节麦;
暗呼吸速率和光补偿点从大至小依次均为:八倍体>六倍体>四倍体>二倍体;
光饱和点从大至小依次为:二倍体>八倍体>四倍体>六倍体;
光量子效率从大至小依次为:四倍体>六倍体>二倍体>八倍体;
在相同co2浓度条件下,所述羧化效率、最大净光合速率和平均净光合速率从大至小依次均为:八倍体小黑麦>六倍体普通小麦>二倍体节节麦>四倍体硬粒小麦;
co2饱和点从大至小依次为:六倍体>八倍体>四倍体>二倍体;
羧化效率从大至小依次为:八倍体>六倍体>二倍体>四倍体。
作为优选,所述二倍体节节麦进行光合作用的最佳温度为27℃,所述四倍体硬粒小麦进行光合作用的最佳温度为30℃,所述六倍体普通小麦进行光合作用的最佳温度为28℃,所述八倍体小黑麦进行光合作用的最佳温度为26℃;
所述二倍体节节麦进行光合作用的最佳湿度为45%,所述四倍体硬粒小麦进行光合作用的最佳湿度为54%,所述六倍体普通小麦进行光合作用的最佳湿度为59%,所述八倍体小黑麦进行光合作用的最佳湿度为53%。
与现有技术相比,本发明的有益效果是:
本发明通过在小麦灌浆期测定不同倍性小麦旗叶的光合气体交换参数(净光合速率、气孔导度、胞间co2浓度、蒸腾速率及水分利用效率)、光合相关性状(小麦旗叶叶片叶绿素浓度、叶面积指数、旗叶面积、绿叶面积、干物质积累量及绿叶数)、光合抗逆能力(光饱和点、光补偿点、co2饱和点、co2光补偿点、最佳湿度、最佳温度、暗呼吸速率、光量子效率、羧化效率、最大净光合速率及平均净光合速率)以及通过分析测定数据筛选出最佳高光效多倍体小麦。采用本发明的方法可科学且准确的从多倍体小麦中筛选出高光效多倍体小麦;根据本发明研究方法也可通过人为改变环境因子,从而发挥出光合作用的最佳优势,最终达到改良增产的目的。
附图说明
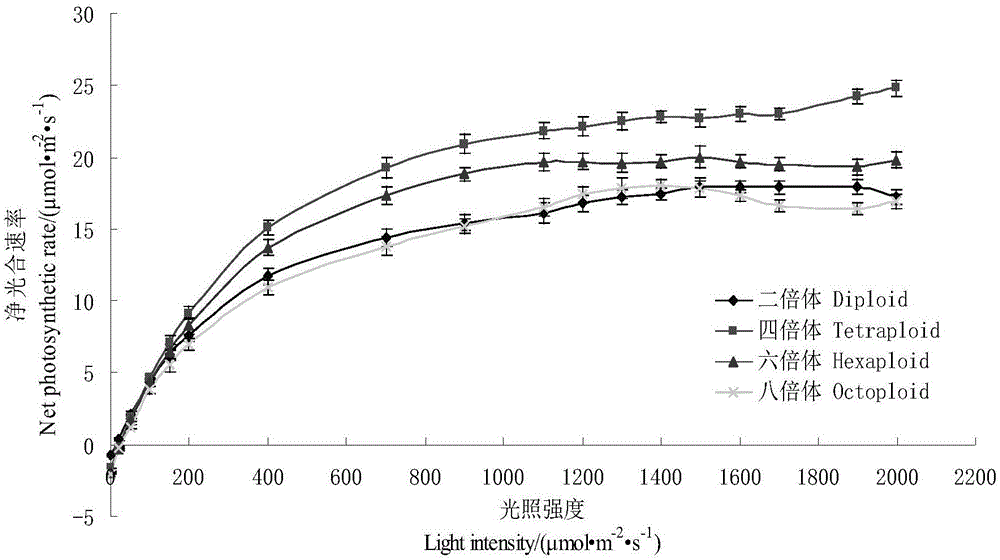
图1是本发明实施例提供的不同倍性小麦的光响应曲线;
图2是本发明实施例提供的不同倍性小麦的co2响应曲线;
图3是本发明实施例提供的不同倍性小麦的温度响应曲线;
图4是本发明实施例提供的不同倍性小麦的湿度响应曲线。
具体实施方式
为更进一步阐述本发明为达成预定发明目的所采取的技术手段及功效,以下以较佳实施例,对依据本发明申请的具体实施方式、技术方案、特征及其功效,详细说明如后。下述说明中的多个实施例中的特定特征、结构、或特点可由任何合适形式组合。
本发明涉及的专业术语解释如下:
高光效育种:是指通过提高作物本身光合能力和降低呼吸消耗的生理指标而提高作物产量的育种方法。
实施例1
试验材料:二倍体节节麦(aegilopstauschii)、四倍体硬粒小麦(t.durum)、六倍体普通小麦(t.aestivum)及八倍体小黑麦(triticale)均由陕西省杂交油菜研究中心小麦研究室提供。试验于2016-2017年在陕西省杨凌示范区小麦试验田进行,田间试验采用随机区组设计,每小区5行,行长2.0m,行距0.25m,重复3次,田间种植与管理同国家区域试验。
测定项目和方法:
(1)小麦光合气体交换参数的测定:在小麦的灌浆期,选择生长健壮、均匀一致的单株,用li-6400便携式光合作用测定仪对受光方向一致的旗叶进行测定,选用6400-02bled红蓝光源叶室,光照控制为1300μmol·m-2·s-1;温度控制为25℃,测定时间为上午9:00-11:00,每种材料测定10片叶子;叶片夹样部位选择旗叶的中段,如果叶片宽度小于2cm,不能充满整个叶室,光合速率等参数的重新计算方法参见文献(赵小光,张耀文,田建华,等.甘蓝型油菜不同发育时期光合日变化和环境因子之间的关系[j].西南农业学报,2013,26(04):1397.)
测定指标:净光合速率(pnμmol·m-2·s-1),气孔导度(gsmmol·m-2·s-1),胞间co2浓度(ciμmol·ml-1)、蒸腾速率(trmmol·m-2·s-1),水分利用效率(wue)按照wue=pn/tr进行计算。
(2)小麦光合相关性状的测定:在小麦灌浆期,每个倍体材料选取10个代表性单株,用叶绿素测定仪(spad502)均匀的在每株的旗叶上选取8个点进行测定,取平均数进行统计分析;用美国li-cor公司的li-3000c叶面积仪分别对每个单株的旗叶面积和整株绿叶面积进行测定,取平均数进行统计分析;将10个代表性单株,去掉泥土清洗干净后,按根、茎、叶、穗等部位分别烘干称重,所有部位干物质重量之和即为干物质积累量;用美国li-cor公司的lai-2200冠层分析仪在田间对每个倍体材料的小区进行测定,记录下每种材料的叶面积指数,重复三次。
(3)光胁迫下小麦光合光合能力的测定:在灌浆期选取小麦的旗叶用li-6400便携式光合作用测定系统进行测定,选用6400-02bled红蓝光源叶室,控制叶室内温度为25℃,co2浓度控制为400μmol·ml-1,在1300μmol·m-2·s-1光强下适应15分钟后,采用自动光曲线程序进行测定,每种材料测定5株。按叶子飘等的方法计算光饱和点、光补偿点、光量子效率和最大净光合速率。
(4)co2胁迫下小麦光合能力的测定:材料选择标准同上,控制叶室内温度为25℃,co2浓度设为1200μmol·ml-1,光强设为1300μmol·m-2·s-1,适应15min后采用自动co2曲线测定程序进行测定,每种材料测定5株。按照叶子飘等的方法计算co2饱和点、羧化效率、co2补偿点和最大净光合速率。
(5)温度胁迫下小麦光合能力的测定:材料选择标准同上,测定时控制光强为1300μmol·m-2·s-1,co2设为400μmol/mol,25℃下适应15min后,采用手动程序进行测定,测定时控制叶室温度,然后分别在外界温度的±10℃范围内每个温度点测定一次,每次试验测定5株,取平均数进行计算。
(6)湿度胁迫下小麦光合能力的测定:材料选择标准同上,测定时控制光强为1300μmol·m-2·s-1,co2设为400μmol/mol,25℃下适应15min后,采用手动程序进行测定,测定时调节干燥管旋钮来控制湿度,从湿度为零开始,每间隔5%设置一个湿度点,在每个湿度点测定一次,每次试验测定5株,取平均数进行计算。
数据的整理与计算:数据利用excel2003进行初步整理,用spss19.0数据统计软件进行统计分析。
结果分析:
(1)不同倍体小麦光合参数的比较:在小麦的发育后期,旗叶作为光合作用的主要功能叶,对小麦光合产物的积累和产量的贡献至关重要,因此本发明选择旗叶作为研究对象,测定了不同倍性材料的光合参数。从表1可以看出,不同材料的净光合速率之间差异比较显著,其大小依次表现为八倍体﹥四倍体﹥二倍体﹥六倍体。二倍体﹑四倍体和六倍体材料的气孔导度不存在显著差异,而八倍体材料的气孔导度分别比它们高出40.91%﹑41.61%和37.78%。不同材料的蒸腾速率差异不大,依次表现为八倍体﹥二倍体﹥四倍体﹥六倍体。二倍体和四倍体材料的胞间co2浓度不存在显著差异,六倍体和八倍体材料的胞间co2浓度也不存在显著差异。在水分利用效率上,八倍体材料最高,二倍体最低,四倍体和六倍体之间差异不大。
从上述数据分析可知,八倍体材料在光合参数上具有明显的优势,其净光合速率﹑气孔导度,蒸腾速率和水分利用效率均高于其它三种倍性材料,其胞间co2浓度低于其它三个材料也说明光合能力更强,较多的消耗掉了细胞内部的co2。
表1.不同倍性小麦的旗叶光合参数的比较
同列数值后不同小写字母表示不同倍性材料之间差异显著(p<0.05);下表同。
(2)不同倍体小麦与光合相关性状的比较:从表2可以看出,四种倍体材料的光合性状差异均比较显著。叶绿素含量随着小麦的进化而逐渐上升,二倍体最低,八倍体最高,八倍体比二倍体提高了55.38%。旗叶面积的变化则为六倍体>四倍体>八倍体>二倍体,二倍体的旗叶面积远低于其它三个倍体材料,分别比四倍体、六倍体和八倍体低63.49%、82.70%和61.96%。绿叶面积变化趋势与旗叶面积相同,也为六倍体>四倍体>八倍体>二倍体,六倍体的绿叶面积远高于其它材料,这种差异的造成主要是由于六倍体小麦在发育后期叶片叶绿素降低较慢,绿叶片数为4片,而其他三个倍体材料绿叶片数均为3片导致的。叶面积指数的变化与旗叶面积、绿叶面积变化趋势也相同,也为六倍体>四倍体>八倍体>二倍体。
通过上述分析可知,在叶面积的多个指标上,六倍体具有明显优势,可以较多的接收光能。干植物的积累量与叶面积的各指标变化相同,这主要与六倍体小麦具有叶面积的优势有关,相比其他三个倍体,可以将较多的光能转化为化学能贮存起来,这也可能是六倍体小麦目前大面积种植的一个因素。
表2.不同倍性小麦的发育后期光合相关性状
(3)不同倍性小麦受光照影响的变化规律:从图1可以看出,不同光照条件下,四种小麦材料的叶片净光合速率的变化趋势基本一致。在光强为200μmol·m-2·s-1以下时,净光合速率与光照强度均为表现为线性关系,且随着光强升高变化较快。在达到各自的光饱和点后净光合速率表现为一条近似平行的直线。在相同的光照强度下,四倍体的净光合速率最高,六倍体其次,二倍体和八倍体最低,两者的差异不大。
由表3可知,四种材料的光量子效率、最大净光合速率和平均净光合速率变化一致,均表现为四倍体>六倍体>二倍体>八倍体,说明四倍体和六倍体具有较高的光能转化和利用效率,可以将吸收光能较多的转化为化学能贮存起来。暗呼吸速率和光补偿点的变化规律一致,均表现为八倍体>六倍体>四倍体>二倍体,其中二倍体小麦的光呼吸速率和光补偿点分别为0.3291μmol·m-2·s-1和7.82μmol·m-2·s-1,远低于其它三个材料,说明二倍体对在进行光合作用的时候,通过光呼吸途径消耗的能量很低,且较低的光补偿点表明二倍体材料相对其它三个材料可以在很弱的光照条件下就能进行光合作用。四种材料的光饱和点表现为:二倍体>八倍体>四倍体>六倍体,二倍体最高,八倍体其次,表明对光合的抗逆行较强,在较强的光照下受到的光抑制较小。
表3.不同倍性小麦光响应参数的比较
(4)不同倍性小麦受co2浓度影响的变化规律:从图2可以看出,不同倍性小麦材料的co2影响曲线的变化趋势与光响应曲线基本一致,当外界co2浓度在200μmol·ml-1以下时,四种小麦材料的净光合速率与co2浓度均为表现为线性关系,并随着co2浓度增加而迅速升高。在达到各自的co2饱和点后,净光合速率的变化趋于平缓。在相同的co2浓度下,四种材料的净光合速率变化与光响应曲线上差异较大,表现为八倍体>六倍体>二倍体>四倍体。
从表4可以得到,四种小麦材料的羧化效率、最大净光合速率和平均净光合速率变化一致,均表现为八倍体>六倍体>二倍体>四倍体,说明八倍体与六倍体材料对co2的利用和转化效率较高,两者的最大净光合速率分别比自然co2浓度条件下(表1)的净光合速率增加了88.97%和135.36%,而二倍体和四倍体的最大净光合速率分别只比自然条件下增加了2.61%和35.78%。二倍体、四倍体和六倍体的co2补偿点无明显差异,八倍体材料最小。四种材料的co2饱和点表现为六倍体>八倍体>四倍体>二倍体,表明八倍体与六倍体材料对co2的抗逆性较强,在高浓度co2下其光合作用不受到抑制,具有较高的光合潜力。
表4.不同倍性小麦co2响应参数的比较
(5)不同倍性小麦受温度影响的变化规律:从图3可以看出,四种小麦材料的的净光合速率随温度的变化均表现为一条抛物线,变化比较平缓,表明小麦光合作用受温度的影响不如光照和co2明显,然而各材料的峰值、最佳温度、最大净光合速率等差异较为明显。二倍体、四倍体、六倍体和八倍体小麦的峰值各出现在27℃、30℃、28℃和26℃,这是各自进行光合作用的最佳温度,其数值大小分别为16.3μmol·m-2·s-1、15.8μmol·m-2·s-1、18.9μmol·m-2·s-1和22.4μmol·m-2·s-1。尽管八倍体小麦的最佳光合温度比较低,在夏季高温条件下光合能力会受到部分抑制,但是在整个温度曲线的范围内,各温度点的光合数值相比其他三个材料一直处在较高的水平下,其平均值分别比二倍体、四倍体和六倍体高出42.89%,41.62和17.45%。二倍体和四倍体小麦材料的净光合速率在各温度条件下都较低,对温度的抗胁迫能力较弱。
(6)不同倍性小麦受湿度影响的变化规律:从图4可以看出,不同倍性材料的湿度响应曲线各不相同,二倍体、四倍体、六倍体和八倍体小麦进行光合作用的最佳湿度分别为45%,54%,59%和53%,最大净光合速率分别为17.3μmol·m-2·s-1、17.9μmol·m-2·s-1、17.3μmol·m-2·s-1和17.1μmol·m-2·s-1,无显著性差异。在整个湿度响应曲线上的平均净光合速率分别为14.88μmol·m-2·s-1、16.67μmol·m-2·s-1、14.98μmol·m-2·s-1和14.93μmol·m-2·s-1,除了四倍体外,其它三种材料的平均净光合速率也无显著性差异。然而,不同材料净光合速率随相对湿度的变化差异较大,二倍体材料表现为明显的抛物线,较低和较高湿度下的净光合速率均比较低,只有在合适的湿度下才能充分进行光合作用,受空气水分的影响较大,抗水分胁迫能力较弱。四倍体材料表现为一条近似平行的直线,净光合速率受相对湿度的影响较小,一直处在较稳定的水平,且相比其他三个材料数值较高。六倍体和八倍体材料变化相近,在较低湿度下净光合能力较弱,但随着湿度的增加,净光合能力逐渐提高,在较高的湿度下降低较为缓慢,因此六倍体和八倍体材料适宜在降雨较多的地域种植,在雨后高湿条件下光合作用受到的影响较小。
本发明通过对不同倍性材料的光合特性分析可知,随着小麦材料的不断进化和改造,光合能力到了明显的提升。八倍体材料是目前小麦多倍体育种的最终材料,相对于其他三种材料,净光合速率、叶绿素浓度、蒸腾速率、气孔导度和水分利用效率都增加的比较明显,这是通过杂交改良,将优良的光合基因不断的聚集到一起的结果,最终提高了八倍体材料的光合效率。尽管六倍体材料的光合参数相比其他三个材料有所降低,但是六倍体小麦在绿叶面积、光合功能叶面积、叶面积指数、绿叶数等多个叶面积指标上具有明显优势,且叶片叶绿素降解缓慢,有效地延长了光合功能期,通过叶面积的优势有效的弥补了净光合能力较低的缺陷,从而使六倍体小麦可以较多的接收光照,将光能转化为化学能贮存起来,提高了干物质积累量,为小麦的高产提供了坚实的基础,这也是六倍体小麦目前大面积种植的一个重要因素。
本发明通过不同倍性小麦材料的净光合速率与光照强度、co2浓度、温度和湿度等外界环境因子的分析可知,小麦材料的光合能力既受自身遗传因素的影响,还很大程度受外界环境的制约。在相同环境因子的影响下,不同倍性小麦材料的变化规律各不相同。
在光照强度影响下,四倍体和六倍体小麦具有较高的光能转化和利用效率,而二倍体和八倍体小麦对光的的抗逆性较强,受的光抑制较小;
在co2浓度影响下,六倍体和八倍体材料对co2的利用和转化效率较高,同样对co2的抗逆性较强,在高浓度co2下其光合作用不受到抑制,具有较高的co2利用潜力;
在温度影响下,八倍体小麦的光合速率受到的影响较小,一直处在较高的水平,二倍体和四倍体小麦材料的净光合速率在各温度条件下都较低,对温度的抗胁迫能力较弱;
在湿度影响下,二倍体材料受空气水分的影响较大,抗水分胁迫能力较弱。四倍体材料受相对湿度的影响较小,一直处在较稳定的水平。六倍体和八倍体材料高湿条件下光合作用受到的影响较小。
植物的净光合速率不是固定不变的,既随着自身遗传的影响进行着日变化、光午休、光反应和暗反应等生理改变,也随着外界环境因子在不停的改变。对于光合速率的生理性改变,可通过生理或者分子手段对材料不停的改良并选育,而对于胁迫性改变,可通过大量实验摸索出净光合速率随环境因子的变化规律,通过人为改变环境因子,从而发挥出光合作用的最佳优势,最终达到改良增产的目的。
目前我国的小麦单产量已经取得了巨大的突破,但随着小麦耕地面积正逐年减少,同时伴随着环境恶化以及农村务农人数的降低,小麦的总产量呈下降。提高小麦面积单产是解决粮食问题的有效途径,但通过传统育种手段实现小麦增产已经不是很明显;因此,在传统的抗逆、品质、早熟等育种基础上,应辅助高光效育种手段,提高亲本资源的光合能力,通过杂交改良提高后代对光的截获和利用能力,降低光午休和光抑制等负效应,最大程度的将光能转化为小麦的化学能贮存起来,从而最终提高小麦的产量。
本发明实施例中未尽之处,本领域技术人员均可从现有技术中选用。
以上公开的仅为本发明的具体实施方式,但本发明的保护范围并不局限于此,任何熟悉本技术领域的技术人员在本发明揭露的技术范围内,可轻易想到变化或替换,都应涵盖在本发明的保护范围之内。因此,本发明的保护范围应以上述权利要求的保护范围为准。
- 还没有人留言评论。精彩留言会获得点赞!