用于植物的除草剂耐受性的方法和组合物与流程
相关申请的引用
本申请要求2017年4月14日提交的美国临时申请序列号62/485,751的权益,所述申请以引用的方式整体并入本文。
本发明涉及生物技术领域。更具体地说,本发明涉及用于产生对生长素除草剂具有抗性的作物植物的组合物和方法。特别地,本发明涉及含有改变的ksiaa16基因和蛋白质的植物、植物组织和植物种子,所述改变的ksiaa16基因和蛋白质与未改变的ksiaa16基因和蛋白质相比提供对生长素除草剂的抗性。
序列表的并入
名称为“mons406wo_st25.txt”、在
背景技术:
杂草是在农业环境中与栽培植物竞争的植物,其使农民每年蒙受数十亿美元的作物损失并为控制杂草而付出努力。杂草还会成为作物病害和虫害的寄主。在农业生产环境中由杂草引起的损失包括:作物产量降低、作物质量下降、灌溉成本增加、收获成本增加、土地价值降低、对牲畜的伤害以及杂草藏匿的昆虫和病害对作物造成的损害。杂草引起这些影响的主要手段是:1)与作物植物竞争水分、养分、阳光和其他生长发育必需品,2)产生导致人类或动物健康问题的有毒或刺激性化学物质,3)产生大量种子或营养繁殖部位或两者,其污染农产品并且使这些杂草物种在农业用地中长期存在,以及4)在农业和非农业用地上产生大量必须要处理的植被。
化学除草剂经常用于控制杂草的生长和蔓延。化学除草剂在植物中的一个或多个靶位点处是活跃的,其中它们阻碍正常的植物功能。生长素是植物生长发育的重要调节剂,其涉及许多发育和反应途径,包括细胞分裂和伸长、植物塑造(patterning)、根系生长、维管组织发育、叶片形状、向光性和向重力性。因为这种植物激素是很多重要植物途径的关键,所以生长素水平或反应的紊乱通常导致植物死亡。这使得若干能够模仿天然存在的活性生长素吲哚-3-乙酸(iaa)功能的化合物被鉴别和商业化,使其成为有效的除草剂。这些生长素除草剂包括诸如3,6-二氯-2-甲氧基苯甲酸(麦草畏(dicamba))和2,4-二氯苯氧乙酸(2,4-d)的化合物。需要对生长素除草剂具有抗性的作物植物。
技术实现要素:
在一方面,提供一种经修饰的植物或种子,其中经修饰的植物或种子对至少一种内源生长素敏感,并且其中经修饰的植物或种子表现出对至少一种合成生长素除草剂的耐受性。内源生长素可选自由iaa和naa组成的组,并且合成生长素除草剂可选自由麦草畏、2,4-d、毒莠定(picloram)、绿草定(triclopyr)、快克草(quinclorac)和氟草烟(fluroxypyr)组成的组。在一些实施方案中,经修饰的植物或种子在编码aux/iaa蛋白的内源基因中包含至少一种修饰,其中所述至少一种修饰与没有所述至少一种修饰的对照植物相比赋予对生长素除草剂的耐受性。在进一步的实施方案中,所述修饰产生包含含有基序x1wppx2的经修饰的降解决定子(degron)结构域的aux/iaa多肽,其中x1相对于野生型aux/iaa多肽被修饰为天冬酰胺(n)、异亮氨酸(i)或亮氨酸(l)并且其中x2是异亮氨酸(i)、缬氨酸(v)、亮氨酸(l)或蛋氨酸(m)。在进一步的实施方案中,所述修饰产生包含与seqidno:219具有至少90%同一性的氨基酸序列的aux/iaa多肽,其中经修饰的植物或种子不是地肤(kochiascoparia)植物或种子。所述修饰可产生包含氨基酸序列seqidno:219的aux/iaa多肽,其中经修饰的植物或种子不是地肤植物或种子。在某些实施方案中,所述经修饰的植物或种子是作物植物或作物种子,例如选自由以下组成的组的单子叶作物植物:玉米、高粱、小麦、油菜、大豆(soy)、棉花、葡萄、番茄、马铃薯、莴苣、西兰花、黄瓜、花生、甜瓜、韭菜、洋葱、水稻和大麦,或选自由以下组成的组的双子叶作物植物:黄豆(soybean)、苜蓿、向日葵、棉花、油菜和甜菜。所述修饰可使得与没有所述修饰的对照植物或种子相比对生长素除草剂的耐受性增加。
在另一方面,提供一种产生经修饰的植物的方法,所述方法包括以下步骤:(a.)在植物细胞的aux/iaa基因中引入修饰;(b.)鉴别和选择在aux/iaa的降解决定子结构域中包含所述修饰的步骤(a.)的一种或多种植物细胞;以及c.从步骤(b)中选择的至少一种或多种细胞再生至少一种植物。
在另一方面,提供一种经修饰的dna分子,其编码经修饰的生长素/吲哚-3-乙酸(aux/iaa)多肽,其中所述aux/iaa多肽包含含有基序x1wppx2的经修饰的降解决定子结构域,其中x1相对于野生型aux/iaa多肽被修饰为天冬酰胺(n)、异亮氨酸(i)或亮氨酸(l)并且其中x2为异亮氨酸(i)、缬氨酸(v)、亮氨酸(l)或蛋氨酸(m)。在一些实施方案中,x1已从甘氨酸修饰为天冬酰胺。在进一步的实施方案中,经修饰的aux/iaa多肽包含选自由以下组成的组的多肽序列:(a.)与seqidno:2n-1,其中n=1-110,或seqidno:247-284中的任一者具有至少90%序列同一性的序列;以及(b.)(a.)中序列的片段,其具有生长素/吲哚-3-乙酸结合活性。在某些实施方案中,经修饰的aux/iaa多肽包含与seqidno:220具有至少95%序列同一性的多核苷酸序列或包含序列seqidno:220。
在另一方面,提供一种dna构建体,其包含可操作地连接至异源启动子的本文所提供的经修饰的dna分子。启动子可为诱导型启动子或植物基因组中的天然生长素/吲哚-3-乙酸基因启动子。
在另一方面,提供一种重组植物、植物部位、细胞或种子,其包含本文所提供的经修饰的dna分子。还提供一种植物基因组,其包含本文所提供的经修饰的dna分子。
在另一方面,提供一种赋予植物生长素除草剂耐受性的方法,其包括在所述植物中表达本文所述的经修饰的dna分子。所述方法可包括用经修饰的dna分子转化所述植物或其祖细胞。在一些实施方案中,生长素除草剂选自由以下组成的组:麦草畏、2,4-d、毒莠定、绿草定、快克草和氟草烟。
在另一方面,提供一种用于产生生长素除草剂耐受植物的方法,所述方法包括:修饰所述植物的aux/iaa多肽以相对于seqidno:217改变包含基序x1wppx2的降解决定子结构域中的残基x1,其中x2为异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。在某些实施方案中,x1从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸。在进一步的实施方案中,修饰包括位点特异性诱变。在一些实施方案中,降解决定子结构域被碱基编辑酶修饰。
还提供通过本文所述的方法产生的植物。
在另一方面,提供用于检测表现出对至少一种内源生长素的敏感性和对至少一种生长素除草剂的耐受性的植物的方法,其包括以下步骤:a.从所述植物获得基因组dna样品;b.使用第一引物和第二引物扩增所述dna样品以产生扩增子;c.在所述扩增子中检测编码seqidno:219内的基序x1wppx2的多核苷酸序列。在一些实施方案中,所述第一引物具有序列seqidno:221并且所述第二引物具有序列seqidno:222。检测可包括限制性片段长度多态性(rflp)分析。在一些实施方案中,所述第一引物具有序列seqidno:223并且所述第二引物具有序列seqidno:224。检测可进一步包括使用选自由以下组成的组的探针:seqidno:225、seqidno:226或其组合。
附图说明
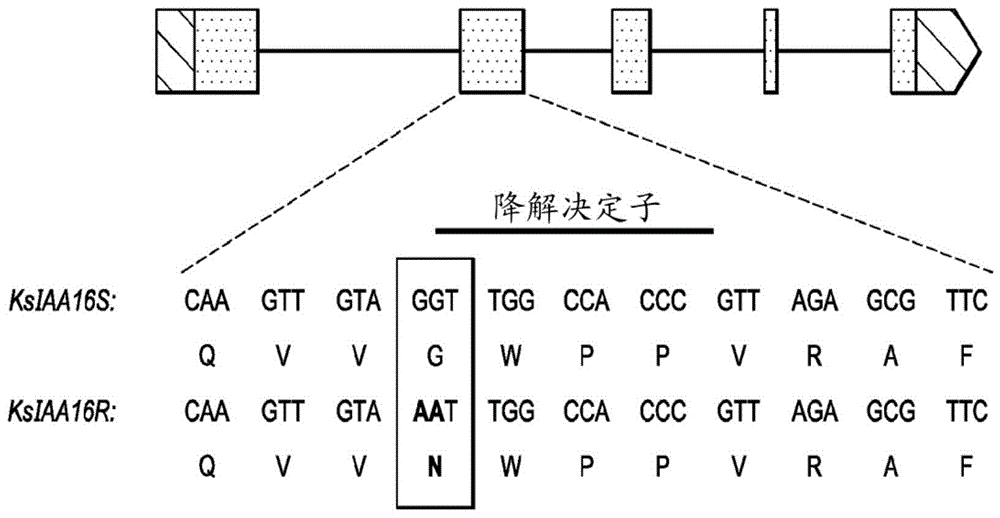
图1:示出了ksiaa16的基因结构,其中指出了敏感(s)和抗性(r)等位基因之间的核苷酸和氨基酸差异。
图2:示出了含有拟南芥(arabidopsis)aux/iaa蛋白以及在转录组序列中鉴别的推定直系同源地肤aux/iaa蛋白的系统发育树。此系统发育树的成员为:atiaa1,(seqidno:247);atiaa2,(seqidno:248);atiaa3,(seqidno:249);atiaa4,(seqidno:250);atiaa5,(seqidno:251);atiaa6,(seqidno:252);atiaa7,(seqidno:253);atiaa8,(seqidno:254);atiaa9,(seqidno:255);atiaa10,(seqidno:256);atiaa11,(seqidno:257);atiaa12,(seqidno:258);atiaa13,(seqidno:259);atiaa14,(seqidno:260);atiaa15,(seqidno:261);atiaa16,(seqidno:262);atiaa17,(seqidno:263);atiaa18,(seqidno:264);atiaa19,(seqidno:265);atiaa20,(seqidno:266);atiaa26,(seqidno:267);atiaa27,(seqidno:268);atiaa28,(seqidno:269);atiaa29,(seqidno:270);atiaa30,(seqidno:271);atiaa31,(seqidno:272);atiaa32,(seqidno:273);atiaa33,(seqidno:274);atiaa34,(seqidno:275);ksiaa1,(seqidno:276);ksiaa3,(seqidno:277);ksiaa6,(seqidno:278);ksiaa7,(seqidno:279);ksiaa8,(seqidno:280);ksiaa9,(seqidno:281);ksiaa13,(seqidno:282);ksiaa18,(seqidno:283);ksiaa31,(seqidno:284)。
图3:示出了来自以下植物的氨基酸序列的ksiaa16进化枝的直系同源物的系统发育树:欧洲油菜(brassicanapus)、栽培大豆(glycinemax)、普通小麦(triticumaestivum)、陆地棉(gossypiumhirsutum)、二色高粱(sorghumbicolor)、玉米(zeamays)、欧亚种葡萄(vitisvinifera)、厚皮甜瓜(cucumismelo)、马铃薯(solanumtuberosum)、番茄(solanumlycopersicum)、莴苣(lactucasativa)、黄瓜(cucumissativus)、蒺藜苜蓿(medicagotruncatula)和向日葵(helianthusannuus)。
图4:示出了图2中系统发育树的代表成员的多序列比对,其中框出的氨基酸表示降解决定子基序。
图5:示出了在有与没有天然和合成生长素的情况下抗性(r)和敏感(s)地肤生物型幼苗的根伸长反应。
图6:示出了麦草畏(图a)、2,4-d(图b)和氟草烟(图c)的鲜重(未处理对照的百分比)。
具体实施方式
除非本文另外定义,否则术语应根据相关领域的普通技术人员的常规用法来理解。可在alberts等人的molecularbiologyofthecell,第5版,garlandsciencepublishing,inc.中找到描述本文使用的许多与分子生物学相关的术语的资源实例:newyork,2007;rieger等人,glossaryofgenetics:classicalandmolecular,第5版,springer-verlag:newyork,1991;king等人,adictionaryofgenetics,第6版,oxforduniversitypress:newyork,2002;和lewin,genesix,oxforduniversitypress:newyork,2007。使用了在37c.f.r.第1.822节陈述的dna碱基命名法。
植物激素生长素是涉及许多植物生长、发育和反应途径的基因的重要调节剂。天然存在的活性生长素是吲哚-3-乙酸(iaa),但已发现许多其他化合物在应用于植物时可模仿iaa的功能。这使得许多起有效除草剂作用的化合物被鉴别和商业化。
玉米和其他单子叶作物对低水平的合成生长素除草剂具有天然耐受性,而双子叶作物诸如黄豆和棉花则对其高度敏感。开发生长素除草剂耐受品种的努力已集中于使生长素除草剂失活的酶的异源表达,从而使原本敏感的植物对除草剂具有耐受性。例如,已经产生了麦草畏耐受黄豆植物,其包含来自嗜麦芽寡养单胞菌(stenotrophomonasmaltophilia)的编码麦草畏单加氧酶(dmo)的基因,所述麦草畏单加氧酶是通过将麦草畏转化为无除草活性的3,6-二氯水杨酸而使其失活的一种酶。
虽然这种方法已取得成功,但基因组编辑技术的最新进展已引起人们对开发具有改良性状的作物的极大兴趣。关于生长素除草剂耐受性的性状,基因组编辑可用于产生改变植物中生长素感知机制的特定突变,从而使植物对包括生长素除草剂在内的合成生长素的影响具有抗性,但仍然对内源生长素敏感。生长素感知的这种改变可能不会导致任何重大的适应度(fitness)影响。不希望受特定理论的束缚,由于生长素感知的改变而具有耐受性的植物将不需要消耗资源来高度表达用于除草剂解毒的任何代谢蛋白,这反过来将会使植物有更多的资源用于其他生长特性。
为了利用基因组编辑技术改变耐受植物的生长素感知,必须鉴别出赋予所需生长素感知表型的特定突变。已经做了大量工作来鉴别和表征使模型植物拟南芥中生长素感知改变的突变,但是已知的突变均未在保持对内源iaa的敏感性的同时产生对合成生长素的抗性。结果,发现大多数已知的突变都会导致拟南芥中的植物生长缺陷。
生长素除草剂抗性田间分离株潜在地提供一组独特的实例,其中植物的生长素感知发生了变化,但仍保持了在比受控环境通常所提供的条件更恶劣的条件下进行繁殖的充分适应度。先前,由于抗性等位基因的频率低以及生长素除草剂抗性杂草的扩散缓慢,阻碍了在任何田间来源的杂草物种中鉴别生长素除草剂抗性的遗传基础的努力。
一种在农业上特别重要的杂草是地肤(kochiascoparial.schrad),这是一种顽强的双子叶杂草物种。尽管麦草畏抗性地肤的扎根和扩散速度极快,但尚未报道这种抗性的遗传基础。本公开内容首次证实了来自内布拉斯加州西部(westernnebraska)的地肤生物型表现出的麦草畏抗性表型的遗传基础。令人惊奇的是,本发明人已经证实这种地肤生物型中的麦草畏抗性是由于两个核苷酸碱基的变化,这种变化导致与拟南芥中的iaa16最为相似的生长素/吲哚-3-乙酸(aux/iaa)蛋白的降解决定子结构域中高度保守的"gwppv/i”基序内甘氨酸到天冬酰胺的氨基酸变化。另外,证明了来自抗性地肤生物型的突变iaa16蛋白似乎具有改变的生长素感知。
如本文所用,单数的和单数形式的术语“一个(a)”、“一种(an)”和“所述(the)”例如包括复数对象,除非上下文另外清楚地规定。因此,例如,提及“植物”、“所述植物”或“一种植物”也包括多个植物;同样,根据上下文,术语“植物”的使用还可包括与所述植物的基因相似或相同的子代;实际上,术语“核酸”的使用任选地包括所述核酸分子的许多拷贝;类似地,术语“探针”任选地(并且通常)涵盖许多相似或相同的探针分子。
如本文所用,“植物”是指整株植物、其任何部位或源自植物的细胞或组织培养物,其包含以下任何一种:整株植物、植物组成部分或器官(例如叶、茎、根等)、植物组织、种子、植物细胞和/或它们的子代。子代植物可以来自任何子代(filialgeneration),例如f1、f2、f3、f4、f5、f6、f7等。植物细胞是植物的生物细胞,其取自植物或通过培养取自植物的细胞而获得。
如本文所用,“修饰”在植物、种子、植物组成部分、植物细胞和植物基因组中是指含有从其天然或原始状态的变化或变异的状态。例如,基因的“天然转录物”是指从未修饰基因产生的rna转录物。通常,天然转录物是有义转录物。经修饰的植物或种子在其遗传物质中含有分子变化,其包括遗传或表观遗传修饰。通常,经修饰的植物或种子或其亲本或祖细胞系经历了诱变、基因组编辑(例如,但不限于通过使用位点特异性核酸酶的方法)、遗传转化(例如,但不限于通过使用农杆菌转化或微粒轰击的方法)或其组合。在一方面,本文所提供的经修饰的植物不包含非植物遗传物质或序列。在另一方面,本文所提供的经修饰的植物不包含种间遗传物质或序列。
如本文所用,“内源基因”或基因的“天然拷贝”是指来自给定生物体、细胞、组织、基因组或染色体内的基因。“内源基因”或基因的“天然拷贝”是先前未被人为修饰的基因。类似地,“内源蛋白”是指内源基因编码的蛋白。
如本文所用,“合成生长素除草剂”或“生长素除草剂”意指任何通过模仿内源植物生长素或抑制生长素化合物从细胞中移出而产生除草活性的除草剂。合成生长素除草剂的实例包括苯甲酸、苯氧基羧酸、吡啶羧酸、喹啉羧酸、半卡巴腙、氟吡草腙(diflufenzopyr)、2,4-d、2,4-db、mcpa、mcpb、mecoprop、麦草畏、氯草啶(clopyralid)、氟草烟、毒莠定、绿草定、氯氨吡啶酸(aminopyralid)、氯丙嘧啶酸(aminocyclopyrachlor)和快克草。
在一方面,本公开提供在aux/iaa蛋白的降解决定子结构域中包含合成突变的非转基因植物。在一方面,本公开提供在iaa16蛋白的降解决定子结构域中包含合成突变的非转基因植物。如本文所用,术语“合成突变”是指非自发突变,并且由于例如通过位点特异性基因组修饰酶的基因组修饰而发生。
几个实施方案涉及经修饰的生长素/吲哚-3-乙酸(aux/iaa)多肽,所述aux/iaa多肽包含含有基序x1wppx2的经修饰的降解决定子结构域,其中x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸,并且其中x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。几个实施方案涉及编码经修饰的生长素/吲哚-3-乙酸(aux/iaa)多肽的核酸序列,所述aux/iaa多肽包含含有基序x1wppx2的经修饰的降解决定子结构域,其中x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸,并且其中x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。几个实施方案涉及包含编码经修饰的生长素/吲哚-3-乙酸(aux/iaa)多肽的核酸序列和/或经修饰的生长素/吲哚-3-乙酸(aux/iaa)多肽的植物、植物部位、植物细胞或植物种子,所述aux/iaa多肽包含含有基序x1wppx2的经修饰的降解决定子结构域,其中x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸,并且其中x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。在一些实施方案中,一种植物、植物部位、植物细胞或植物种子包含经修饰的生长素/吲哚-3-乙酸(aux/iaa)多肽,所述aux/iaa多肽包含含有基序x1wppx2的经修饰的降解决定子结构域,其中x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸,并且其中x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸,所述植物、植物部位、植物细胞或植物种子对包括生长素除草剂在内的合成植物生长素的影响具有抗性,但仍然对内源生长素敏感。
在一方面,对编码蛋白质的核酸分子的修饰导致与内源蛋白相比至少1个、至少2个、至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个、至少10个、至少15个、至少20个、至少25个、至少30个、至少40个、至少50个、至少75个或至少100个错义突变。在另一方面,对编码aux/iaa蛋白的核酸分子的修饰导致与未修饰序列相比至少1个、至少2个、至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个、至少10个、至少15个、至少20个、至少25个、至少30个、至少40个、至少50个、至少75个或至少100个错义突变。如本文所用,“错义突变”是指导致产生编码不同氨基酸的密码子的单个核苷酸变化。例如,密码子“cgu”编码精氨酸氨基酸。如果错义突变将g改变为u,产生“cuu”密码子,则密码子现在编码亮氨酸氨基酸。错义突变可能是由插入、缺失、替换或倒位引起的。
在进一步的实施方案中,提供了设计和产生表现出对生长素除草剂的耐受性的作物植物的方法。在一些实施方案中,提供了如本文所述的通过修饰植物细胞的基因组以改变涉及生长素除草剂耐受性的基因而设计表现出对生长素除草剂的耐受性的植物的方法。基因组修饰可以通过如本文所述的靶向诱变来完成。
几个实施方案涉及能够为植物提供生长素抗性的经修饰的生长素/吲哚-3-乙酸(aux/iaa)多肽。几个实施方案涉及编码经修饰的生长素/吲哚-3-乙酸(aux/iaa)多肽的核酸序列,以及使用经修饰的多肽的组合物和方法。在一些实施方案中,编码经修饰的生长素/吲哚-3-乙酸(aux/iaa)多肽的核酸序列在包含重组dna分子的植物中提供对生长素除草剂的耐受性。
几个实施方案涉及通过基因工程产生的核酸分子、多核苷酸、多肽、蛋白质、细胞、种子或植物,因此它们通常不会在自然界中被发现且是通过人为干预产生。几个实施方案涉及包含不天然存在于植物基因组中且通过人为干预产生的dna序列的植物基因组。在一些实施方案中,本文提供的dna序列编码包含修饰的降解决定子结构域的修饰的aux/iaa多肽。在某些实施方案中,提供了经修饰的aux/iaa多肽,其在包含基序x1wppx2的降解决定子结构域中包含相对于ksiaa16s(seqidno:218)编码的多肽的改变。在一些实施方案中,提供了包含含有基序x1wppx2的经修饰的降解决定子结构域的重组多肽,其中x1已从甘氨酸修饰为不同的氨基酸,例如修饰为亲水性氨基酸,诸如精氨酸、天冬酰胺、天冬氨酸、谷氨酸、谷氨酰胺、赖氨酸、丝氨酸或苏氨酸,并且x2是疏水性氨基酸,诸如亮氨酸或异亮氨酸。在一些实施方案中,提供了包含含有基序x1wppx2的经修饰的降解决定子结构域的重组多肽,其中x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸,并且x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。在一些实施方案中,提供了包含以下的多肽序列的多肽:seqidno:2n-1中的任一者,其中n=1-110,或seqidno:247-284中的任一者,已被改变为具有包含基序x1wppx2的经修饰的降解决定子结构域,其中x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸,并且x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。
几个实施方案涉及编码能够赋予植物改变的植物生长素感知的多肽序列的多核苷酸,例如如下的多肽:seqidno:2n-1中的任一者,其中n=1-110,或seqidno:247-284中的任一者,已被改变为具有包含基序x1wppx2的经修饰的降解决定子结构域,其中x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸,并且x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。在一些实施方案中,多核苷酸包含多核苷酸序列seqidno:220(ksiaa16r)。在一些实施方案中,提供包含本文所述的多核苷酸或多肽序列的植物、植物部位、植物细胞或种子。
在一些实施方案中,核酸分子可完全地或部分地通过本领域已知的方法合成和修饰,其中希望提供可用于dna操纵的序列(诸如限制性酶识别位点或基于重组的克隆位点)、植物优选的序列(诸如植物密码子使用或kozak共有序列)、或可用于dna构建体设计的序列(诸如间隔区或接头序列)。在一些实施方案中,提供核酸分子,所述核酸分子与多核苷酸序列seqidno:2n-1,其中n=1-110具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%、至少99.5%或100%的序列同一性,并且能够赋予植物改变的生长素感知,从而赋予对生长素除草剂的耐受性。
几个实施方案涉及多肽序列,所述多肽序列与多核苷酸序列seqidno:2n-1,其中n=1-110具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%、至少99.5%或100%的序列同一性,并且能够赋予植物改变的生长素感知,从而赋予对生长素除草剂的耐受性。
在一些实施方案中,提供能够赋予植物改变的生长素感知、从而赋予对生长素除草剂的耐受性的多核苷酸或多肽序列的片段。在特定实施方案中,提供seqidno:2n-1中的任一者,其中n=1-110的片段,其包含seqidno:2n-1,其中n=1-110中的任一者的至少约50、95、150、250、500、750或至少约1000个连续核苷酸或更长,并且能够赋予植物改变的生长素感知,从而赋予对生长素除草剂的耐受性。
aux/iaa蛋白家族包含四个保守结构域,其中结构域ii组成一个17个氨基酸的序列作为降解决定子结构域,其赋予生长素依赖性降解。特别地,此降解决定子结构域包含基序"gwppv/i",其在所有aux/iaa蛋白中严格保守。在一些实施方案中,提供编码aux/iaa多肽的多核苷酸,所述aux/iaa多肽包含本文所提供的氨基酸序列的片段或变体并且包含含有基序x1wppx2的经修饰的降解决定子结构域,其中降解决定子结构域的修饰提供能够赋予植物改变的生长素感知的多肽,从而赋予对合成生长素除草剂的耐受性。在某些实施方案中,多核苷酸编码的多肽包含本文所提供的氨基酸序列的片段或变体并且包含含有基序x1wppx2的经修饰的降解决定子结构域,其中x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸,并且x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。
如本文所用,术语“dna”或“dna分子”是指基因组或合成来源的双链dna分子,即,脱氧核糖核苷酸碱基的聚合物或多核苷酸分子,从5′(上游)端读取到3′(下游)端。如本文使用,术语“dna序列”是指dna分子的核苷酸序列。本文使用的命名法对应于美国联邦法规第1.822节的标题37,并且在wipo标准st.25(1998),附录2,表1和表3的表格中陈述。
如本文所用,术语“分离的dna分子”是指与在其原始或天然状态中通常与其相关的其他分子至少部分地分开的dna分子。在一个实施方案中,术语“分离的”是指与在其原始或天然状态中通常侧接dna分子的一些核酸至少部分地分开的dna分子。因此,例如通过重组技术而融合到通常与之不相关的调控序列或编码序列的dna分子在本文中被认为是分离的。当整合到宿主细胞的染色体中或与其他dna分子一起存在于核酸溶液中时,因为其不是在天然状态,所以此类分子也被认为是分离的。
如本文所公开的,本领域技术人员公知的许多方法可以用于分离和操纵多核苷酸或其片段。例如,聚合酶链反应(pcr)技术可用于扩增特定的起始多核苷酸分子和/或产生原始分子的变体。多核苷酸分子或其片段也可以通过其他技术获得,诸如通过化学方法直接合成片段,如通常通过使用自动化寡核苷酸合成仪所实践的。多核苷酸可以是单链的(ss)或双链的(ds)。“双链”是指通常在生理相关条件下发生在充分互补的反平行核酸链之间以形成双链核酸结构的的碱基配对。所述方法的实施方案包括其中多核苷酸为选自由以下组成的组的至少一者的实施方案:正义单链dna(ssdna)、正义单链rna(ssrna)、双链rna(dsrna)、双链dna(dsdna)、双链dna/rna杂合体、反义ssdna或反义ssrna;可以使用任何这些类型的多核苷酸的混合物。
如本文所用,术语“序列同一性百分比”或“序列同一性%”是指当两个序列最佳地比对时(在比较窗口上适当的核苷酸或氨基酸插入、缺失、或缺口总计小于参考序列的20%的情况下)与测试(“目标”)序列(或其互补链)相比参考(“查询”)序列(或其互补链)的线性多核苷酸或多肽序列中相同的核苷酸或氨基酸的百分比。用于比对比较窗口的序列的最佳比对是本领域技术人员熟知的,并且可通过工具诸如smith和waterman的局部同源性算法、needleman和wunsch的同源性比对算法、pearson和lipman的相似性搜索方法和通过这些算法的例如使用默认参数的计算机化实现方式诸如作为
如本文所用,术语“蛋白质编码多核苷酸分子”是指包含编码蛋白质的核苷酸序列的多核苷酸分子。“蛋白质编码序列”意指编码蛋白质的多核苷酸序列。此类多核苷酸是选自由以下组成的组的至少一种:正义单链dna(ssdna)、正义单链rna(ssrna)、双链rna(dsrna)、双链dna(dsdna)、双链dna/rna杂合体、反义ssdna或反义ssrna;或任何这些类型的多核苷酸的混合物。
“序列”意指核苷酸或氨基酸的顺序布置。蛋白质编码序列的边界可由5'末端处的翻译起始密码子和3'末端处的翻译终止密码子确定。在一些实施方案中,蛋白质编码分子可包含编码蛋白质序列的dna序列。在一些实施方案中,蛋白质编码分子可包含编码蛋白质序列的rna序列。如本文所用,“转基因表达”、“表达转基因”、“蛋白质表达”、以及“表达蛋白质”意指通过将dna分子转录为信使rna(mrna)并且将mrna翻译为多肽链(其最终折叠为蛋白质)的过程产生蛋白质。
由于遗传密码的简并性,多种不同的多核苷酸序列可以编码多肽,诸如本文所公开的改变的多肽。lewin(1985)中描述了一张表格,其示出了所有可能的三联体密码子(并且其中u也替代了t)和每个密码子编码的氨基酸。另外,产生编码本文所述的相同或基本相同的改变的多肽的替代多核苷酸序列完全在本领域技术人员的能力范围内。这些变体或替代多核苷酸序列在本文所述的实施方案的范围内。如本文所用,提及“基本上相同”的序列是指编码不会实质性改变本文所述实施方案的多核苷酸编码的多肽的功能活性的氨基酸替换、缺失、添加或插入的序列。编码野生型或改变的多肽的核苷酸序列的等位基因变体也涵盖在本文所述实施方案的范围内。除了那些在野生型或改变的aux/iaa多肽中具体例举或天然存在的氨基酸以外的氨基酸的替换也考虑在本文所述实施方案的范围内,只要具有所述替换的多肽仍保留基本相同的功能活性(例如选择性生长素感知)。
i.基因组编辑
通过使用基因组编辑方法对植物基因组进行的靶向修饰可用于通过修饰植物基因组dna来产生改良的植物系。另外,基因组编辑方法可以使一种或多种目标核酸靶向插入植物基因组中。用于将供体多核苷酸引入植物基因组中或修饰植物的基因组dna的方法实例包括使用序列特异性核酸酶,诸如锌指核酸酶、工程改造的或天然的大范围核酸酶、tale核酸内切酶或rna引导的核酸内切酶(例如,规律间隔成簇短回文重复序列(crispr)/cas9系统、crispr/cpf1系统、crispr/casx系统、crispr/casy系统、crispr/cascade系统)。几个实施方案涉及基因组编辑的方法,所述方法使用单链寡核苷酸在植物基因组中引入精确的碱基对修饰,如sauer等人所述(plantphysiol.2016年4月;170(4):1917–1928)。用于修饰、缺失或将核酸序列插入基因组dna中的基因组编辑方法是本领域已知的。
几个实施方案涉及crispr/cas9系统,其用于修饰或替代植物基因组中现有的编码序列,诸如编码aux/iaa多肽的序列。几个实施方案涉及crispr/cpf1系统,其用于修饰或替代植物基因组中现有的编码序列,诸如编码aux/iaa多肽的序列。几个实施方案涉及cas9-胞苷脱氨酶,其用于修饰植物基因组中现有的编码序列,诸如编码aux/iaa多肽的序列。几个实施方案涉及cpf1-胞苷脱氨酶,其用于修饰植物基因组中现有的编码序列,诸如编码aux/iaa多肽的序列。在进一步的实施方案中,转录激活因子样效应物(tale)用于修饰或替代植物基因组中现有的编码序列,诸如编码aux/iaa多肽的序列。在一些实施方案中,通过使用序列特异性核酸酶的非模板基因组编辑来修饰植物基因组中现有的aux/iaa多肽编码序列。在一些实施方案中,通过使用序列特异性核酸酶的模板基因组编辑来修饰植物基因组中现有的aux/iaa多肽编码序列。在一些实施方案中,将序列特异性核酸酶和编码经修饰的降解决定子结构域的供体核酸引入植物细胞中。在一些实施方案中,经修饰的降解决定子结构域编码能够赋予植物对生长素除草剂的耐受性的x1wppx2基序。在一些实施方案中,x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸。在一些实施方案中,x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。根据本文所提供的方法的内源aux/iaa编码序列的修饰或替代导致包含经修饰的降解决定子结构域的多肽的表达,所述经修饰的降解决定子结构域含有基序x1wppx2,例如其中x1和x2中的一者或多者已被改变。几个实施方案因此涉及提供能够识别植物基因组中的特定目标核苷酸序列(诸如aux/iaa序列)的序列特异性核酸酶,以通过在所述位点处供体核酸的非模板编辑或整合来实现aux/iaa序列的改变。
对aux/iaa编码序列例如本文提供为seqidno:2n-1之一,其中n=1-110的序列的修饰可以产生编码包含经修饰的降解决定子结构域的多肽的序列,所述经修饰的降解决定子结构域含有如本文所述的基序x1wppx2,其能够赋予植物改变的植物生长素感知,从而赋予对生长素除草剂的耐受性。在一些实施方案中,x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸。在一些实施方案中,x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。
在一方面,“修饰”包含至少1个、至少2个、至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个、至少10个、至少15个、至少25个、至少50个、至少100个、至少200个、至少300个、至少400个、至少500个、至少750个、至少1000个、至少1500个、至少2000个、至少3000个、至少4000个、至少5000个或至少10,000个核苷酸的插入。在另一方面,“修饰”包含至少1个、至少2个、至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个、至少10个、至少15个、至少25个、至少50个、至少100个、至少200个、至少300个、至少400个、至少500个、至少750个、至少1000个、至少1500个、至少2000个、至少3000个、至少4000个、至少5000个或至少10,000个核苷酸的缺失。在另一方面,“修饰”包含至少2个、至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个、至少10个、至少15个、至少25个、至少50个、至少100个、至少200个、至少300个、至少400个、至少500个、至少750个、至少1000个、至少1500个、至少2000个、至少3000个、至少4000个、至少5000个或至少10,000个核苷酸的倒位。在另一方面,“修饰”包含至少1个、至少2个、至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个、至少10个、至少15个、至少25个、至少50个、至少100个、至少200个、至少300个、至少400个、至少500个、至少750个、至少1000个、至少1500个、至少2000个、至少3000个、至少4000个、至少5000个或至少10,000个核苷酸的替换。在一些实施方案中,“修饰”包含在核酸序列中用“a”替换“c”、“g”或“t”。在一些实施方案中,“修饰”包含在核酸序列中用“c”替换“a”、“g”或“t”。在一些实施方案中,“修饰”包含在核酸序列中用“g”替换“a”、“c”或“t”。在一些实施方案中,“修饰”包含在核酸序列中用“t”替换“a”、“c”或“g”。在一些实施方案中,“修饰”包含在核酸序列中用“c”替换“u”。在一些实施方案中,“修饰”包含在核酸序列中用“g”替换“a”。在一些实施方案中,“修饰”包含在核酸序列中用“a”替换“g”。在一些实施方案中,“修饰”包含在核酸序列中用“t”替换“c”。
在一些实施方案中,提供重组dna构建体,诸如包含编码aux/iaa蛋白的核酸序列的转基因或表达盒。在一些实施方案中,aux/iaa蛋白是iaa16同源物。在一些实施方案中,核酸序列编码包含经修饰的降解决定子结构域的aux/iaa蛋白。在一些实施方案中,经修饰的降解决定子结构域包含能够赋予植物对生长素除草剂的耐受性的x1wppx2基序。在一些实施方案中,x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸。在一些实施方案中,x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。在一些实施方案中,转基因或表达盒包含衍生自可操作地连接至aux/iaa编码序列的iaa基因的内源启动子的启动子。在一些实施方案中,可将转基因或表达盒插入或整合到植物或植物细胞的基因组内的基因座中。在一些实施方案中,可以通过定点整合将转基因或表达盒插入或整合到植物或植物细胞的基因组内的特定位点或基因座中。在一些实施方案中,重组dna构建体包含编码如下多肽的核苷酸序列:seqidno:2n-1,其中n=1-110中的任一者,已被改变为具有包含基序x1wppx2的经修饰的降解决定子结构域;或编码这种多肽的片段的序列。在一些实施方案中,重组dna构建体可进一步包含一个或两个同源臂,其侧接插入序列(即,将要插入植物基因组中的序列、转基因、盒等)。
几个实施方案涉及重组dna构建体,其包含编码位点特异性核酸酶和/或任何相关蛋白质的表达盒以进行基因组修饰。这些核酸酶表达盒可存在于与用于模板编辑的供体模板相同的分子或载体中、或包含编码如本文所述的aux/iaa蛋白的核酸序列的表达盒中(顺式)、或存在于单独的分子或载体中(反式)。用于定点整合的若干种方法是本领域已知的,其涉及切割基因组dna以在所需的基因组位点或基因座处产生双链断裂(dsb)或缺口的不同序列特异性核酸酶(或者蛋白质和/或指导rna的复合物)。如本领域所理解的,在修复由核酸酶引入的dsb或缺口的过程中,供体模板dna、转基因或表达盒可在dsb或缺口的位点处整合到基因组中。尽管插入事件可通过非同源末端连接(nhej)发生,但待整合的dna中同源臂的存在可通过同源重组来促进修复过程中插入序列在植物基因组中的采用和靶向。可使用的位点特异性核酸酶的实例包括锌指核酸酶、工程改造或天然的大范围核酸酶、tale核酸内切酶和rna引导的核酸内切酶(例如,cas9、casx、casy或cpf1)。对于使用rna引导的位点特异性核酸酶(例如,cas9、casx、casy或cpf1)的方法,重组dna构建体也可包含编码一个或多个引导rna的序列以将核酸酶引导至植物基因组内的所需位点。在一些实施方案中,可在单独的分子或载体上提供一种或多种引导rna(顺式)。
ii.位点特异性基因组修饰酶
如本文所用,术语“双链断裂诱导剂”是指可以在dna分子中诱导双链断裂(dsb)的任何试剂。在一些实施方案中,双链断裂诱导剂是位点特异性基因组修饰酶。
如本文所用,术语“位点特异性基因组修饰酶”是指任何能以序列特异性方式修饰核苷酸序列的酶。在一些实施方案中,位点特异性基因组修饰酶通过诱导单链断裂来修饰基因组。在一些实施方案中,位点特异性基因组修饰酶通过诱导双链断裂来修饰基因组。在一些实施方案中,位点特异性基因组修饰酶包含胞苷脱氨酶。在一些实施方案中,位点特异性基因组修饰酶包含腺嘌呤脱氨酶。在本公开中,位点特异性基因组修饰酶包括核酸内切酶、重组酶、转座酶、脱氨酶、解旋酶及其任何组合。在一些实施方案中,位点特异性基因组修饰酶是序列特异性核酸酶。
几个实施方案涉及通过向植物细胞提供位点特异性基因组修饰酶例如序列特异性核酸酶来促进重组。在一些实施方案中,通过提供单链断裂诱导剂来促进重组。在一些实施方案中,通过提供双链断裂诱导剂来促进重组。在一些实施方案中,通过提供链分离诱导剂来促进重组。在一方面,位点特异性基因组修饰酶选自核酸内切酶、重组酶、转座酶、脱氨酶、解旋酶或其任何组合。在一些实施方案中,b染色体之间发生重组。在一些实施方案中,b染色体与a染色体之间发生重组。
几个实施方案涉及通过提供位点特异性基因组修饰酶来促进一个或多个目标dna的整合。在一些实施方案中,通过提供单链断裂诱导剂来促进一个或多个目标dna的整合。在一些实施方案中,通过提供双链断裂诱导剂来促进一个或多个目标dna的整合。在一些实施方案中,通过提供链分离诱导剂来促进一个或多个目标dna的整合。在一方面,位点特异性基因组修饰酶选自核酸内切酶、重组酶、转座酶、解旋酶或其任何组合。
在一方面,核酸内切酶选自大范围核酸酶、锌指核酸酶(zfn)、转录激活因子样效应物核酸酶(talen)、argonaute蛋白(argonaute蛋白的非限制性实例包括:嗜热栖热菌(thermusthermophilus)来源的argonaute蛋白(ttago)、极端嗜热菌(pyrococcusfuriosus)来源的argonaute蛋白(pfago)、格氏嗜盐碱杆菌(natronobacteriumgregoryi)来源的argonaute蛋白(ngago))、rna引导的核酸酶,诸如crispr相关核酸酶(crispr相关核酸酶的非限制性实例包括cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也被称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4、cpf1、casx、casy、mad7、其同源物或其修饰形式)。
在一些实施方案中,位点特异性基因组修饰酶是dcas9-fok1融合蛋白。在另一方面,位点特异性基因组修饰酶是dcas9-重组酶融合蛋白。如本文所用,“dcas9”是指具有一个或多个氨基酸突变的cas9核酸内切酶蛋白,所述氨基酸突变导致cas9蛋白不具有核酸内切酶活性,但保留rna引导的位点特异性dna结合。如本文所用,“dcas9-重组酶融合蛋白”是蛋白质与dcas9以重组酶对dna具有催化活性的方式融合的dcas9。
在一些实施方案中,位点特异性基因组修饰酶是dcas9-胞嘧啶脱氨酶融合蛋白。在另一方面,位点特异性基因组修饰酶是dcas9-腺嘌呤脱氨酶融合蛋白。在一些实施方案中,dcas9-胞嘧啶脱氨酶融合蛋白和dcas9-腺嘌呤脱氨酶融合蛋白中的一者或多者被用于修饰编码aux/iaa蛋白的降解决定子结构域的核酸序列。在一些实施方案中,对编码降解决定子结构域的核酸序列进行修饰以编码含有基序x1wppx2的降解决定子结构域,其中x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸,并且x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。在一些实施方案中,aux/iaa蛋白是ksiaa16的直系同源物。
在一些实施方案中,位点特异性基因组修饰酶是dcpf1-胞嘧啶脱氨酶融合蛋白。在另一方面,位点特异性基因组修饰酶是dcpf1-腺嘌呤脱氨酶融合蛋白。在一些实施方案中,dcpf1-胞嘧啶脱氨酶融合蛋白和dcpf1-腺嘌呤脱氨酶融合蛋白中的一者或多者被用于修饰编码aux/iaa蛋白的降解决定子结构域的核酸序列。在一些实施方案中,对编码降解决定子结构域的核酸序列进行修饰以编码含有基序x1wppx2的降解决定子结构域,其中x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸,并且x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。在一些实施方案中,aux/iaa蛋白是ksiaa16的直系同源物。
在一些实施方案中,位点特异性基因组修饰酶是重组酶。重组酶的非限制性实例包括连接至本文所提供的dna识别基序的酪氨酸重组酶,其选自由以下组成的组:cre重组酶、gin重组酶、flp重组酶和tnp1重组酶。在一方面,本文所提供的cre重组酶或gin重组酶与锌指dna结合结构域或taledna结合结构域或cas9核酸酶连接在一起。在另一方面,连接至本文所提供的dna识别基序的丝氨酸重组酶选自由以下组成的组:phic31整合酶、r4整合酶和tp-901整合酶。在另一方面,连接至本文所提供的dna结合结构域的dna转座酶选自由以下组成的组:tale-piggybac和tale-mutator。
位点特异性基因组修饰酶,诸如大范围核酸酶、zfn、talen、argonaute蛋白(argonaute蛋白的非限制性实例包括嗜热栖热菌来源的argonaute蛋白(ttago)、极端嗜热菌来源的argonaute蛋白(pfago)、格氏嗜盐碱杆菌来源的argonaute蛋白(ngago)、其同源物或其修饰形式)、rna引导的核酸酶(rna引导的核酸酶的非限制性实例包括crispr相关核酸酶,诸如cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也被称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4、cpf1、casx、casy、mad7、其同源物或其修饰形式)以及工程改造的rna引导的核酸酶(rgn)诱导基因组修饰,诸如基因组序列靶位点的双链dna断裂(dsb)或单链dna断裂。在一些实施方案中,通过同源重组(hr)或非同源末端连接(nhej)的自然过程来修复靶dna序列中的断裂或缺口。在一些实施方案中,序列修饰发生在切割或缺口的位点或其附近,可包括导致核酸序列修饰的缺失或插入,或通过同源重组或nhej的外源核酸整合。
通过引入目标dna和所提供的位点特异性基因组修饰酶,可将本文所提供的任何目标dna整合到染色体序列的靶位点中。本文所提供的任何方法可以利用本文所提供的任何位点特异性基因组修饰酶。
几个实施方案涉及本文所提供的方法和/或组合物,其包含至少一种、至少两种、至少三种、至少四种、至少五种、至少六种、至少七种、至少八种、至少九种或至少十种位点特异性基因组修饰酶。在另一方面,本文所提供的方法和/或组合物包含编码至少一种、至少两种、至少三种、至少四种、至少五种、至少六种、至少七种、至少八种、至少九种或至少十种位点特异性基因组修饰酶的至少一种、至少两种、至少三种、至少四种、至少五种、至少六种、至少七种、至少八种、至少九种或至少十种多核苷酸。
在一方面,如本文所述的靶向基因组修饰包括使用重组酶。在一方面,连接至本文所提供的dna识别基序的酪氨酸重组酶选自由以下组成的组:cre重组酶、gin重组酶、flp重组酶和tnp1重组酶。在一方面,本文所提供的cre重组酶或gin重组酶与锌指dna结合结构域连接在一起。flp-frt定点重组系统来自面包酵母酿酒酵母(saccharomycescerevisiae)中的2μ质粒。在此系统中,flp重组酶(翻转酶)重组翻转酶识别靶标(frt)位点之间的序列。frt位点包含34个核苷酸。flp与frt位点的“臂”结合(一个臂方向相反),并在介入核酸序列的任一末端切割frt位点。切割后,flp重组两个frt位点之间的核酸序列。cre-lox是衍生自噬菌体p1的定点重组系统,其与flp-frt重组系统相似。cre-lox可用于反转核酸序列、删除核酸序列或易位核酸序列。在此系统中,cre重组酶重组一对lox核酸序列。lox位点包含34个核苷酸,其中第一个和最后13个核苷酸(臂)呈回文结构。在重组过程中,cre重组酶蛋白与不同核酸上的两个lox位点结合,并在lox位点切割。经切割的核酸被剪接在一起(相互易位)并完成重组。在另一方面,本文所提供的lox位点是loxp、lox2272、loxn、lox511、lox5171、lox71、lox66、m2、m3、m7或m11位点。
在另一方面,连接至本文所提供的dna识别基序的丝氨酸重组酶选自由以下组成的组:phic31整合酶、r4整合酶和tp-901整合酶。在另一方面,连接至本文所提供的dna结合结构域的dna转座酶选自由以下组成的组:tale-piggybac和tale-mutator。
在一些实施方案中,如本文所述的靶向基因组修饰包括使用锌指核酸酶(zfn)。zfn是合成蛋白质,其由与foki限制性核酸酶切割结构域融合的工程改造的锌指dna结合结构域组成。可以对zfn进行设计来切割几乎任何长链的双链dna以用于修饰锌指dna结合结构域。zfn从单体形成二聚体,所述单体由与经工程改造以结合靶dna序列的锌指阵列融合的foki核酸酶的非特异性dna切割结构域构成。zfn的dna结合结构域通常由3-4个锌指阵列构成。相对于锌指∞-螺旋起点而位于-1、+2、+3和+6位的、有助于与靶dna的位点特异性结合的氨基酸可以进行更改和定制以使其适应特定靶序列。其他氨基酸形成共有骨架以产生具有不同序列特异性的zfn。选择zfn的靶序列的规则是本领域已知的。foki核酸酶结构域需要二聚化才能切割dna,并且因此需要两个带有c端区域的zfn来结合切割位点的相反dna链(由5-7nt分隔开)。如果两个zf结合位点是呈回文结构,则zfn单体可以切割靶位点。如本文所用,术语zfn是宽泛的,并且包括能够切割双链dna而无需另一zfn协助的单体zfn。术语zfn也用于指一对zfn的一个或两个成员,所述一对zfn经工程改造来一起作用以在同一位点切割dna。
不受任何科学理论的限制,因为原则上可以使用多种方法中的一种重新工程改造锌指结构域的dna结合特异性,所以理论上能够构建针对几乎任何基因序列的定制zfn。用于工程改造锌指结构域的公开可用方法包括上下文依赖组装(coda)、寡聚体库工程化(open)和模块化组装。
几个实施方案涉及本文所提供的方法和/或组合物,其包含针对编码aux/iaa蛋白的基因中的靶序列的一种或多种、两种或更多种、三种或更多种、四种或更多种、或五种或更多种zfn。在另一方面,本文所提供的zfn能够产生靶向dsb。在一方面,通过本领域中已知的方法(例如,但不限于病毒转染、粒子轰击、peg介导的原生质体转染或农杆菌介导的转化)向细胞提供包含编码一种或多种、两种或更多种、三种或更多种、四种或更多种、或五种或更多种zfn的多核苷酸的载体。
几个实施方案涉及使用大范围核酸酶产生如本文所述的修饰。大范围核酸酶通常在微生物中鉴别,是独特的酶,具有高活性和长识别序列(>14nt),从而导致靶dna的位点特异性消化。天然存在的大范围核酸酶的工程设计版本通常具有扩展的dna识别序列(例如14至40nt)。大范围核酸酶的工程改造可能比zfn和talen更具挑战性,因为大范围核酸酶的dna识别和切割功能交织在单个结构域中。诱变和高通量筛选的专门方法已用于产生识别独特序列并具有改进的核酸酶活性的新型大范围核酸酶变体。
在一方面,本文所提供的方法和/或组合物包含针对编码aux/iaa蛋白的基因中的靶序列的一种或多种、两种或更多种、三种或更多种、四种或更多种、或五种或更多种大范围核酸酶。在一些实施方案中,本文所提供的大范围核酸酶能够产生靶向dsb。在一些实施方案中,通过本领域中已知的方法(例如,但不限于病毒转染、粒子轰击、peg介导的原生质体转染或农杆菌介导的转化)向细胞提供包含编码一种或多种、两种或更多种、三种或更多种、四种或更多种、或五种或更多种大范围核酸酶的多核苷酸的载体。
在一些实施方案中,如本文所述的靶向基因组修饰包括使用转录激活因子样效应物核酸酶(talen)。talen是通过将转录激活因子样效应物(tale)dna结合结构域与核酸酶结构域融合而产生的人工限制性酶。在一方面,核酸酶选自由以下组成的组:pvuii、muth、tevi和foki、alwi、mlyi、sbfi、sdai、stsi、cledorf、clo051、pept071。如本文所用,术语talen是宽泛的,并且包括能够切割双链dna而无需另一talen协助的单体talen。术语talen也用于指一对talen的一个或两个成员,所述一对talen经工程改造来一起作用以在同一位点切割dna。转录激活因子样效应物(tale)可以被工程改造来与几乎任何dna序列诸如编码aux/iaa蛋白的核酸中的靶序列结合。tale蛋白是源自黄单胞菌属(xanthomonas)的各种植物细菌病原体的dna结合结构域。x病原体在感染过程中将tale分泌到宿主植物细胞中。tale移至细胞核,其在细胞核中识别并结合宿主基因组中特定基因的启动子区域中特定dna序列的启动子区域中的特定dna序列。tale具有由33-34个氨基酸的13-28个重复单体构成的中央dna结合结构域。除了12和13位的高变氨基酸残基,每个单体的氨基酸都高度保守。这两个可变氨基酸称为重复可变双残基(rvd)。rvd的氨基酸对ni、ng、hd和nn分别优先识别腺嘌呤、胸腺嘧啶、胞嘧啶和鸟嘌呤/腺嘌呤,并且rvd的调节可以识别连续的dna碱基。氨基酸序列与dna识别之间的这种简单关系允许通过选择包含适当rvd的重复片段的组合来工程改造特定的dna结合结构域。
在一方面,本文所提供的方法和/或组合物包含一种或多种、两种或更多种、三种或更多种、四种或更多种、或五种或更多种talen。在另一方面,本文所提供的talen能够在编码aux/iaa蛋白的核酸的靶序列中产生靶向dsb。在一方面,通过本领域中已知的方法(例如,但不限于病毒转染、粒子轰击、peg介导的原生质体转染或农杆菌介导的转化)向细胞提供包含编码一种或多种、两种或更多种、三种或更多种、四种或更多种、或五种或更多种talen的多核苷酸的载体。
在一些实施方案中,如本文所述的靶向基因组修饰包括使用至少一种、至少两种、至少三种、至少四种、至少五种、至少六种、至少七种、至少八种、至少九种或至少十种位点rna引导的核酸酶。在一些实施方案中,可使用crispr/cas9系统、crispr/cpf1系统、crispr/casx系统或crispr/casy系统来产生对编码aux/iaa蛋白的核酸的修饰。crispr系统是基于rna引导的工程改造核酸酶,其使用互补碱基配对来识别靶位点的dna序列。crispr(规律间隔成簇短回文重复序列)/cas(crispr相关的)系统是合成蛋白的替代品,所述合成蛋白的dna结合结构域使其能够修饰特定序列的基因组dna(例如,zfn和talen)。crispr/cas系统是细菌和古细菌的适应性免疫系统的一部分,通过以序列依赖性方式切割外来dna来保护它们免受入侵核酸(诸如病毒)的侵害。通过在crispr基因座近端的两个相邻重复序列之间整合被称为间隔区的入侵dna的短片段来获得免疫力。crispr阵列,包括间隔区在内,在随后与侵入性dna的接触中被转录,并被加工成长度约为40nt的小干扰crisprrna(crrna),其与反式激活crisprrna(tracrrna)结合以激活和引导cas9核酸酶。这在入侵dna中切割称为前间隔区(protospacer)的同源双链dna序列。切割的先决条件是在靶dna的下游存在保守的前间隔区邻近基序(pam),其通常具有序列5'-ngg-3',但较少出现nag。特异性由pam上游约12个碱基的所谓“种子序列”提供,所述种子序列必须在rna与靶dna之间匹配。cpf1的行为与cas9相似,但是cpf1不需要tracrrna。crispr/cas系统的特异性是基于使用互补碱基配对来识别靶dna序列的rna引导。在一些实施方案中,位点特异性基因组修饰酶是crispr/cas系统。在一方面,本文所提供的位点特异性基因组修饰酶可包含任何rna引导的cas核酸酶(rna引导的核酸酶的非限制性实例包括cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4、cpf1、casx、casy、mad7、其同源物或其修饰的形式);以及任选地,靶向相应核酸酶所必需的引导rna。
几个实施方案涉及通过本文所公开的方法产生的植物细胞、植物组织、植物种子和植物。植物可能是单子叶植物或双子叶植物,并且可包括例如水稻、小麦、大麦、燕麦、黑麦、高粱、玉米、葡萄、西红柿、马铃薯、莴苣、西兰花、黄瓜、花生、甜瓜、韭菜、洋葱、黄豆、苜蓿、向日葵、棉花、油菜和甜菜植物。
iii.表达构建体
本文所述的多核苷酸可以提供在表达构建体中。表达构建体通常包括在待表达表达构建体的预期宿主细胞中起作用的调控元件。因此,本领域普通技术人员可以选择用于细菌宿主细胞、酵母宿主细胞、植物宿主细胞、昆虫宿主细胞、哺乳动物宿主细胞和人宿主细胞中的调控元件。调控元件包括启动子、转录终止序列、翻译终止序列、增强子和聚腺苷酸化元件。如本文所用,术语“表达构建体”是指提供可操作地连接的核酸序列的转录的核酸序列的组合。如本文所用,“可操作地连接”意指两个dna分子以使得一个dna分子可影响另一个的功能的方式连接。可操作地连接的dna分子可以是单一邻接的分子的一部分并且可相邻或可不相邻。例如,启动子与dna构建体中的多肽编码dna分子可操作地连接,在所述dna构建体中两个dna分子布置成使得启动子可影响dna分子的表达。
如本文所用,术语“异源性的”是指来源于不同的来源并且因此在自然界中通常不相关的两种或更多种物质之间的关系。例如,如果此组合通常不存在于自然界中,则蛋白质编码重组dna分子相对于可操作地连接的启动子是异源性的。此外,当特定重组dna分子非天然地发生在细胞、种子、或生物体中时,所述特定重组dna分子可相对于其所插入的此特定细胞、种子、或生物体是异源性的。
表达构建体可以包含与编码本文所述的经修饰的多肽的多核苷酸序列可操作地连接的启动子序列。可以使用本领域已知的标准技术将启动子结合到多核苷酸中。启动子或多个启动子的多个拷贝可用于本文所述的表达构建体。在一些实施方案中,启动子可定位在距离表达构建体的转录起始位点与其天然遗传环境中距离转录起始位点大约相同的位置上。允许该距离出现一些变化而启动子活性没有实质性降低。转录起始位点通常包括在表达构建体中。
多个实施方案涉及编码经修饰的aux/iaa多肽的重组dna分子,所述aux/iaa多肽包含含有基序x1wppx2的经修饰的降解决定子结构域,其中x1已从甘氨酸修饰为天冬酰胺,并且其中x2是异亮氨酸或缬氨酸,其中重组dna分子被进一步定义为可操作地连接至异源调控元件。在特定的实施方案中,异源调控元件是在植物细胞中起作用的启动子。在进一步的实施方案中,启动子是诱导型启动子。
如果要将表达构建体提供或引入植物细胞中,则可使用植物病毒启动子,诸如,例如花椰菜花叶病毒(cauliflowermosaicvirus,camv)35s(包括增强的camv35s启动子(参见,例如,美国专利号5,106,739))或camv19s启动子或木薯叶脉花叶病毒(cassavaveinmosaic)。可用于植物中的表达构建体的其他启动子包括,例如包括玉米zein启动子的zein启动子、prolifera启动子、ap3启动子、热休克启动子、根癌农杆菌(a.tumefaciens)的t-dna1'-或2'-启动子、多聚半乳糖醛酸酶启动子、矮牵牛的查尔酮合酶a(chs-a)启动子、烟草pr-1a启动子、泛素启动子、肌动蛋白启动子、alca基因启动子、pin2启动子(xu等人,1993)、玉米wipi启动子、玉米trpa基因启动子(美国专利号5,625,136)、玉米cdpk基因启动子和rubiscossu启动子(美国专利号5,034,322)。还考虑将组成型启动子(诸如camv、泛素、肌动蛋白或nos启动子)、发育调控型启动子和诱导型启动子(诸如受热、光、激素或化学品诱导的那些启动子)用于本文描述的多核苷酸表达构建体。
表达构建体可以任选地包含转录终止序列、翻译终止序列、编码信号肽的序列和/或增强子元件。转录终止区通常可以从真核或病毒基因序列的3'非翻译区获得。转录终止序列可以位于编码序列的下游,以提供有效的终止。信号肽序列是通常存在于蛋白质的氨基末端的短氨基酸序列,其负责将可操作地连接的成熟多肽重新定位到广泛的翻译后细胞目的地,范围从特定的细胞器区室到蛋白质作用位点和细胞外环境。通过使用可操作地连接的信号肽序列而将基因产物靶向到预期的细胞和/或细胞外目的地预期与本文所述的多肽一起使用。经典的增强子是增加基因转录的顺式作用元件并且也可以包括在表达构建体中。经典的增强子元件是本领域已知的,并且包括但不限于camv35s增强子元件、巨细胞病毒(cmv)早期启动子增强子元件和sv40增强子元件。增强基因表达的内含子介导的增强子元件也是本领域中已知的。这些元件必须存在于转录区域内并且与方向相关。实例包括玉米shrunken-1增强子元件(clancy和hannah,2002)。
iv.转化方法
几个实施方案涉及包含本文所述的重组dna的植物细胞、植物组织、植物和种子。在一些实施方案中,重组dna编码序列特异性酶。在一些实施方案中,包含重组dna分子的细胞、组织、植物和种子表现出对生长素除草剂的耐受性。在一些实施方案中,从表现出对生长素除草剂的耐受性的植物中选育出编码序列特异性酶的重组dna。
适用于转化宿主植物细胞的方法几乎包括能够将dna或rna引入细胞的任何方法(例如,将重组dna构建体稳定整合到植物染色体中,或者将重组dna构建体或rna瞬时提供给植物细胞的方法),并且所述方法是本领域众所周知的。两种有效的细胞转化方法是农杆菌介导的转化和微粒轰击介导的转化。微粒轰击方法例如在美国专利号us5,550,318;us5,538,880;us6,160,208和us6,399,861中有所说明。农杆菌介导的转化方法描述于例如美国专利号us5,591,616,其以引用方式全文并入。植物材料的转化在营养培养基(例如,允许细胞在体外生长的营养素的混合物)上的组织培养中进行实践。受体细胞靶标包括但不限于分生组织细胞、茎尖、下胚轴、愈伤组织、未成熟或成熟的胚胎、以及配子体细胞,诸如小孢子和花粉。愈伤组织可起始于包括但不限于不成熟或成熟胚胎、下胚轴、幼苗顶端分生组织、小孢子等的组织来源。含有转基因核的细胞生长成转基因植物。
转化时,通常在任一转化实验中都将dna引入仅较小百分比的靶标植物细胞中。使用标记基因来提供用于鉴别通过接受重组dna分子并将其整合至基因组中而被稳定转化的那些细胞的有效系统。优选的标记基因提供赋予对如抗生素或除草剂的选择剂的抗性的选择性标记。本公开的植物可对其具有抗性的任何除草剂是用于选择性标记的试剂。使潜在转化细胞暴露于选择剂。存活细胞群体是其中通常赋予抗性的基因被整合且在足以允许细胞存活的水平下被表达的那些细胞。可进一步测试细胞以确认外源dna的稳定整合。通常使用的选择性标记基因包括赋予对如卡那霉素和巴龙霉素(nptii)、潮霉素b(aphiv)、大观霉素(aada)和庆大霉素(aac3和aacc4)的抗生素的抗性或对如草铵膦(bar或pat)、麦草畏(dmo)和草甘膦(aroa或epsps)的除草剂的抗性的选择性标记基因。所述选择性标记的实例在美国专利号us5,550,318;us5,633,435;us5,780,708和us6,118,047中有所说明。也可采用提供目视筛选转化体的能力的标记,例如表达着色或荧光蛋白质(如荧光素酶或绿色荧光蛋白(gfp))的基因或表达β-葡萄糖醛酸酶的基因或已知其各种显色底物的uida基因(gus)。
几个实施方案涉及用具有从基因组编辑产生的经修饰的基因组dna的细胞再生植物的方法和构建体。然后可以将再生的植物用于繁殖其他植物。
几个实施方案涉及引入编码aux/iaa蛋白的基因的等位基因变体的方法,其包括:(a)在至少一个植物细胞中引入对内源aux/iaa基因的至少一种修饰,从而产生所述等位基因变体;(b)鉴别和选择步骤(a)的至少一个植物细胞,其包含表现出对生长素除草剂的耐受性的所述等位基因变体;以及(c)从步骤(b)中选择的至少一个植物细胞再生植物。几个实施方案涉及引入编码aux/iaa蛋白的基因的等位基因变体的方法,其包括:(a)在至少一个植物细胞中引入对内源aux/iaa基因的至少一种修饰,从而产生所述等位基因变体;(b)鉴别和选择步骤(a)的至少一个植物细胞,其包含表现出对生长素除草剂的耐受性的所述等位基因变体;(c)鉴别和选择步骤(b)的至少一个植物细胞,其表现出对内源生长素iaa的敏感性;以及(d)从步骤(c)中选择的至少一个植物细胞再生植物。在一方面,方法包括在编码aux/iaa蛋白的降解决定子结构域的序列中引入修饰。在一些实施方案中,经修饰的降解决定子结构域编码能够赋予植物对生长素除草剂的耐受性的x1wppx2基序。在一些实施方案中,x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸。在一些实施方案中,x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。在另一方面,引入方法包括引入重组dna载体,其包含针对编码aux/iaa蛋白的核酸中的靶序列的多核苷酸序列特异性酶。
v.生长素除草剂和具有生长素除草剂耐受性的植物
几个实施方案涉及包含含有经修饰的降解决定子结构域的aux/iaa蛋白的植物细胞、植物组织、植物和种子。在一些实施方案中,经修饰的降解决定子结构域包含能够赋予植物细胞、植物组织、植物和种子对合成生长素除草剂的耐受性的x1wppx2基序。在一些实施方案中,x1已从甘氨酸修饰为天冬酰胺、异亮氨酸或亮氨酸。在一些实施方案中,x2是异亮氨酸、缬氨酸、亮氨酸或蛋氨酸。植物可能是单子叶植物或双子叶植物,并且可包括例如水稻、小麦、大麦、燕麦、黑麦、高粱、玉米、葡萄、西红柿、马铃薯、莴苣、西兰花、黄瓜、花生、甜瓜、胡椒、胡萝卜、南瓜、洋葱、黄豆、苜蓿、向日葵、棉花、油菜和甜菜植物。
基于作用方式,合成生长素除草剂也被称为生长素类生长调节剂除草剂或第o组或第4组除草剂。合成生长素除草剂的作用方式是它们似乎影响细胞壁可塑性和核酸代谢,从而可导致细胞分裂和生长不受控制。合成生长素除草剂的组包括四个化学家族:苯氧基、羧酸(或吡啶)、苯甲酸和最新的喹啉羧酸家族。
苯氧基除草剂最为常见并且自二十世纪40年代发现(2,4-二氯苯氧基)乙酸(2,4-d)以来就被用作除草剂。其他实例包括4-(2,4-二氯苯甲基)丁酸(2,4-db)、2-(2,4-二氯苯甲基)丙酸(2,4-dp)、(2,4,5-三氯苯甲基)乙酸(2,4,5-t)、2-(2,4,5-三氯苯甲基))丙酸(2,4,5-tp)、2-(2,4-二氯-3-甲基苯氧基)-n-苯基丙酰胺(氯甲酰草胺(clomeprop))、(4-氯-2-甲基苯氧基)乙酸(mcpa)、4-(4-氯-邻甲苯氧基)丁酸(mcpb)和2-(4-氯-2-甲基苯氧基)丙酸(mcpp)。
第二大化学家族是羧酸除草剂,也被称为吡啶除草剂。实例包括3,6-二氯-2-吡啶羧酸(氯草啶)、4-氨基-3,5,6-三氯-2-吡啶羧酸(毒莠定)、(2,4,5-三氯苯氧基)乙酸(绿草定)和4-氨基-3,5-二氯-6-氟-2-吡啶氧基乙酸(氟草烟)。第三化学家族是苯甲酸类,其实例包括3,6-二氯邻茴香酸(麦草畏)和3-氨基-2,5-二氯苯甲酸(豆科威(choramben))。生长素除草剂的第四个且最新的化学家族是喹啉羧酸家族,其包括7-氯-3-甲基-8-喹啉羧酸(喹草酸(quinmerac))和3,7-二氯-8-喹啉羧酸(快克草)。后者的独特之处在于,它也会控制一些草类杂草,这与其他生长素样除草剂不同,其他生长素基本上仅控制阔叶或双子叶植物。
合成生长素除草剂可施加到包含由本文所述的组合物和方法提供的植物和种子的植物生长区域以作为控制杂草的方法。本文所述的组合物和方法提供的植物和种子包含合成生长素除草剂耐受性性状,因此可以耐受一种或多种生长素除草剂的施加。除草剂施加可以是推荐的商业比率(1x)或其任何分数或倍数,诸如推荐的商业比率的两倍(2x)。根据除草剂和剂型,生长素除草剂比率可表达为酸当量/磅/英亩(lbae/acre)或酸当量/克/公顷(gae/ha)或表达为磅活性成分/英亩(lbai/acre)或克活性成分/公顷(gai/ha)。植物生长区域在除草剂施加时可包含或可不包含杂草植物。
除草剂的施加可与一种、两种生长素除草剂、或若干种生长素除草剂的组合或任何其他相容的除草剂依序地混合或罐装混合。一种除草剂或组合地或单独地两种或更多种除草剂的多次施加可在生长季节用于包含表达如本文所述的经修饰的aux/iaa蛋白的植物的区域以用于控制广泛范围的双子叶杂草、单子叶杂草或两者,例如,两次施加(诸如种植前施加和出苗后施加或出苗前施加和出苗后施加)或三次施加(诸如种植前施加、出苗前施加和出苗后施加或出苗前施加和两次出苗后施加)。
如本文所用,“耐受性”或“除草剂耐受性”意指植物、种子、或细胞抵抗施加时的除草剂的毒性作用的能力。可以通过将植物、种子、植物组织、植物部位或细胞与合适的对照进行比较来测量植物、种子、植物组织、植物部位或细胞的除草剂耐受性。例如,可通过将除草剂施加到包含编码能赋予除草剂耐受性的经修饰的多肽的重组dna分子的植物(测试植物)和不包含编码能赋予除草剂耐受性的经修饰的多肽的重组dna分子的植物(对照植物)中、然后比较两种植物的植物伤害来测量或评估除草剂耐受性,其中与对照植物的伤害率相比伤害率降低表明测试植物的除草剂耐受性。与对照植物、种子、植物组织、植物部位或细胞相比,除草剂耐受植物、种子、植物组织、植物部位或细胞表现出对除草剂毒性作用的反应降低。如本文所用,“除草剂耐受性性状”是将与野生型植物相比的改善的除草剂耐受性赋予到植物的转基因性状。
如本文所用,术语“包括”表示“包括但不限于”。
已经详细描述了几个实施方案,将显而易见的是,可能存在修改、变化和等同实施例。此外,应理解,本公开中的实施例作为非限制性实施例提供。
实施例
实施例1:麦草畏抗性型和麦草畏敏感型地肤生物型之间的遗传差异鉴别
获得了表现出麦草畏耐受性近30倍增加的麦草畏抗性型地肤生物型,据信这一增加是由显性或半显性单一遗传基因座赋予的。从麦草畏抗性型地肤生物型(r)和麦草畏敏感型地肤生物型(s)的营养期植物的气生组织中分离的cdna的转录组测序允许鉴别出了已知涉及生长素合成、转运和调控反应的拟南芥基因的推定地肤直系同源物。比较物包括一种aux1/lax转运蛋白、一种abp同源物、几种多重耐药样蛋白(mrp)、四种tir/afb共受体和十种aux/iaa蛋白。
从温室生长的s和r地肤生物型中分别收获叶组织,并在液氮上冷冻。根据制造商的规程,使用trizol(invitrogen)从每种组织的三个生物重复物中分离出总rna。使用nanodrop8000分光光度计确定rna量,并通过rin大于6的bioanalyzer分析评估完整性。按照制造商的规程,使用illuminatruseqrnasampleprepkitv2,将2mg总rna用于测序文库制备。qpcr(sybrpcrmastermix,appliedbiosystems)用于定量测序文库。如前所述(rice等人,2014),通过hiseq2000测序仪、使用50个碱基对的读数进行测序。使用成对的末端读数为r(82,123,945对)和s(68,928,684对)生物型构建地肤转录组组装体。首先使用fastx工具包(版本0.0.13,质量分数>=30的至少50%的碱基)对读数进行质量修整,然后使用trinity(版本r2013_08_14,默认设置,最小转录长度=300)进行组装(grabherr等人,natbiotechnol.201129(7):644-52.)。
观察到r型和s型地肤转录物之间的明显差异。与来自s生物型的转录物序列相比,r生物型表现出gg到aa核苷酸变化。gg到aa核苷酸变化导致甘氨酸(g)到天冬酰胺(n)的氨基酸替换(图1)。包含此氨基酸替换(seqidno:219)的蛋白质(ksiaa16;seqidno:217)与拟南芥aux/iaa蛋白at.iaa16(seqidno:262)最为相关,后者与at.iaa7(seqidno:253)、at.iaa14(seqidno:260)和at.iaa17(seqidno:263)属于同一进化枝(图2)。观察到的氨基酸变化发生在高度保守的降解决定子区内(gwppv/i)内(图1)。已报道这一五个氨基酸的降解决定子区内发生的变化提高了aux/iaa蛋白的稳定性,并导致许多拟南芥突变体的显性生长素抗性,所述拟南芥突变体包括如表1中所示的axr5(iaa1)(yang等人,2004)、shy2(iaa3)(tian和reed,1999)、axr2(iaa7,(seqidno:247))(nagpal等人,2000)、slr(iaa14,(seqidno:260))(fukaki等人,2002)、axr3(iaa17,(seqidno:263))(rouse等人,1998)、crane(iaa18,(seqidno:264))(uehara等人,2008)、bodenlos(iaa12,(seqidno:258))(hamann等人,2002)、msg2(iaa19,(seqidno:265))(tatematsu等人,2004)、iaa16(seqidno:262)(rinaldi等人,2012)、和iaa28(seqidno:269)(rogg等人,2001)。这些发现表明,观察到的导致ksiaa16中甘氨酸(g)到天冬酰胺(n)氨基酸替换的gg到aa核苷酸变化很可能是在这种地肤生物型中观察到的麦草畏抗性的基础。
表1.拟南芥功能获得型aux/iaa突变体的降解决定子结构域内的氨基酸序列
虽然先前已报道高度保守的gwppv降解决定子序列中的突变提供对内源生长素化合物吲哚-3-乙酸(iaa)的抗性,但是尚未观察到这些突变在保持对内源生长素iaa敏感(例如仍然能够与之结合)的同时赋予对除草生长素化合物的抗性(例如,结合亲和力降低)。在拟南芥中鉴别出的降解决定子序列突变体表现出明显改变的感知内源iaa的能力,从而导致产生有害的表型和不良的劣型。
实施例2:地肤的tir/afb同源物的基于序列的鉴别
植物中已经鉴别出许多生长素感知途径。研究最深入的途径之一涉及scf泛素连接酶催化的aux/iaa转录阻遏物降解,这是由f-box蛋白的tir1/afb家族介导的。在存在生长素的情况下,scf复合物的亚基tir1充当生长素受体。一旦tir1与生长素结合,它就能以高亲和力结合aux/iaa蛋白家族的成员,从而在两种蛋白之间形成复合物,并促进aux/iaa蛋白的泛素化,为其标上降解标签。当将aux/iaa蛋白结合到结合生长素的tir1蛋白时,aux/iaa蛋白的降解决定子区中的"gwppv/i”基序被认为覆盖了整个生长素分子。已观察到,降解决定子基序中的突变导致aux/iaa蛋白的稳定,这继而导致植物模型系统中生长素反应降低和多种生长缺陷。
由于aux/iaa蛋白与tir/afb蛋白协同作为生长素共受体,因此鉴别了来自地肤的tir/afb蛋白的推定同源物,以确定在生长素存在下ksiaa16蛋白是否与tir/afb蛋白相互作用。这样的相互作用将表明ksiaa16是一种生长素共受体。
使用已知的拟南芥tir/afb蛋白作为查询序列对地肤转录组数据集进行blast分析。使用这种方法,在地肤中鉴别了四种tir/afb直系同源物。将蛋白质与拟南芥中的直系同源物tir1、afb1、afb2、afb3、afb4和afb5以及海枣(phoenixdactyliferal.,datepalm)和火炬松(pinustaedal.,loblollypine)的afb6直系同源物进行比对(parry等人,2009)。使用megalign软件(dnastar,madison,wi)中的clustalw方法进行比对。结果表明,地肤含有来自拟南芥tir/afb家族的每个已知进化枝的一种直系同源物。
实施例3:地肤生长素反应因子和生长素相互作用分析
酵母双杂交(y2h)研究使用
表2:y2h载体构建中使用的引物序列
对pcr产物进行凝胶纯化,并使用
此方法使得能够在不存在或存在100μm或500μm外源浓度的iaa、naa、麦草畏和2,4-d、毒莠定、绿草定、快克草、氟草烟或苯乙酸(paa)的情况下,研究ksiaa16的r型和s型地肤等位基因与每种地肤tir/afb同源物(kstir1(seqidno:285)和ksafb6(seqidno:287))的结合/相互作用。诱饵和猎物载体的所有组合在没有选择的情况下均显示出良好的生长,这表明转录物表达对酵母生长没有任何固有的负面影响。从y2h分析获得的结果显示在表3和表4中。
表3:测试ksiaa16s和ksiaa16r与kstir1的结合的y2h分析结果
表4:测试ksiaa16s和ksiaa16r与ksafb6的结合的y2h分析结果
在不存在生长素的情况下,几乎观察不到ksiaa16的r和s等位基因两者与kstir1和ksafb6直系同源物的生长,这表明没有生长素时蛋白质不会相互作用。ksiaa16s在存在100m和500miaa、2-(1-萘基)乙酸(naa)、麦草畏和2,4-d以及500m绿草定的情况下表现出与kstir1的相互作用,但是ksiaa16r仅在100m和500miaa和naa存在下表现出与kstir1的相互作用。在麦草畏和2,4-d存在下,ksiaa16r表现出与kstir1相互作用的能力降低。不希望受特定理论的束缚,在麦草畏和2,4-d存在下ksiaa16r与kstir1的相互作用降低、从而导致在麦草畏和2,4-d存在下ksiaa16r的稳定性增加可以解释在麦草畏抗性地肤生物型中观察到的对这些化合物的抗性。同样,在存在100μm和500μmiaa、naa和绿草定以及500μm2,4-d、毒莠定和快克草的情况下,观察到ksiaa16s与ksafb6相互作用,但仅在所测试的最高浓度的iaa和naa(500m)存在下观察到ksiaa16r与ksafb6之间的相互作用。这表明在iaa和naa存在下,ksiaa16r与ksafb6相互作用的能力降低,因为只有较高的浓度才能促进二者结合,并且在绿草定、2,4-d、快克草或毒莠定的测试浓度下未观察到ksiaa16r与ksafb6之间的相互作用。在paa或氟草烟存在的情况下,使用任何测试的蛋白质组合均未观察到相互作用。不希望受特定理论的束缚,氟草烟存在下未观察到y2h相互作用可能是由于相互作用不稳定,其阻止了酵母系统中的检测;或可能是由于缺少氟草烟甲基庚酯的新陈代谢,据报道所述植物和细菌以游离酸的形式裂解并释放。这些发现表明,ksiaa16与kstir1和/或ksafb6协同起到生长素共受体的作用,并且ksiaa16与kstir1和/或ksafb6的组合至少部分地负责对这些生长素除草剂中的每一种的感知。此外,这是首次报道的afb6同源物的表征,因为此进化枝已从多个研究过的物种中消失,包括拟南芥、水稻、高粱和玉米(hayashi,2012;mironova等人,2010;parry等人,2009)。不希望受特定理论的束缚,这些发现表明,ksafb6可以充当生长素共受体,并且与ksiaa16协同作用,ksafb6能够在几种测试的生长素除草剂存在下相互作用。
实施例4:麦草畏抗性型和麦草畏敏感型地肤中生长素介导的根伸长反应的表征
测试了来自r型和s型地肤生物型的幼苗的根伸长反应,以确定r生物型反应是否与y2h结果一致。对于根伸长研究,使用了0.5%蔗糖的植物营养培养基。将种子在含有0.1%triton-x100的30%漂白剂中进行表面灭菌10至30分钟,然后在无菌水中冲洗两次,然后重悬于0.1%琼脂中进行平板接种。测量根长为最接近的毫米数,并计算12个个体的平均根长,除非另有说明。根伸长数据如图5所示,其显示r型地肤生物型对iaa和naa的根伸长抑制作用仍然敏感,但对麦草畏和2,4-d的抑制作用具有抗性。另外,观察到在没有生长素的情况下,r生物型表现出短根表型。这些发现与观察到的y2h结果一致,其显示在所测试的浓度下,iaa和naa可以促进ksiaa16r与kstir1和/或ksafb6的一些相互作用,但麦草畏、2,4-d、毒莠定、绿草定、快克草和氟草烟不能。天然生长素paa似乎也对根伸长有影响。除了y2h结果外,在此概述的观察结果还为以下假设提供了进一步的支持:在这种地肤生物型中观察到的生长素抗性是由于ksiaa16r中的突变所致。
实施例5:ksiaa16r等位基因分子标记的开发
基于在s型和r型地肤生物型之间在ksiaa16中鉴别的碱基对替换,出于基因分型的目的,开发了限制性片段长度多态性(rflp)分析和
对于rflp分析,正向引物(seqidno:221)和反向引物(seqidno:222)被设计用于从抗性和敏感性植物的基因组dna扩增146bp的产物。发现在r生物型中鉴别的突变在ksiaa16r等位基因中产生了另外的mluci限制性位点。这使得当消化的产物在4%琼脂糖凝胶上进行电泳时可以区分r和s等位基因,因为s等位基因产物只有两个mluci限制性核酸内切酶切割位点,并在消化之后产生120、19和7个碱基对的片段,而r等位基因具有三个切割位点,并产生90、30、19和7个碱基对的片段。
对于
实施例6:地肤中麦草畏抗性性状的分离模式
为了确定ksiaa16r等位基因是否与麦草畏抗性表型共分离,对包含s型和r型植物的地肤种群进行了喷雾试验。利用s生物型作为雌性而r生物型作为雄性进行s型和r型个体之间的杂交以鉴别由异花授粉产生的个体。由于地肤繁殖器官的解剖结构,很难防止一些自花授粉,但是利用
为了确定ksiaa16r等位基因是否与麦草畏抗性表型共分离,对经过验证的杂合f1亲本植物的720f2子代进行了采样和基因分型,并且确定了对两倍于典型麦草畏田间使用率(1120g/ha)的麦草畏喷雾浓度的耐受性。在整个实验过程中保持植物身份,并确定每个f2子代的基因型和可见麦草畏伤害百分比。每个基因型的平均伤害对于s/s为约80%,对于r/s为约40%,对于r/r为约20%。基于taqman基因分型分析,鉴别出234株纯合敏感性植物(s/s),371株杂合植物(r/s)和105株纯合抗性植物(r/r)。14株植物反馈了不确定的基因型,因此代表2%的pcr反应失败率。这些基因型数量(1:1.6:0.4)与预期的孟德尔1:2:1分离模式明显不同(p<0.05),表明抗性等位基因可能影响发芽和/或幼苗形成。基因型结果与观察到的麦草畏伤害之间存在很强的相关性,s/s个体的平均伤害率为82.3%,r/s个体的平均伤害率为35.9%,r/r个体的平均伤害率为24.6%。使用anova对这些数据的统计分析表明了麦草畏抗性表型与ksiaa16r等位基因的存在有关的高确定性(p<0.0001),并且cramér-vonmises统计分析显示所述性状为半显性。
实施例7:ksiaa16在拟南芥中的表达与检测
为了确定单独ksiaa16r等位基因的存在是否必要并足以赋予麦草畏抗性,产生了几种转基因表达构建体用于在拟南芥中研究ksiaa16。通过花浸法(clough和bent,1998),使用根癌农杆菌将二元载体引入植物中。载体包含乙醇诱导型alca启动子元件(salter等人,1998)、组成型camv35s启动子、或驱动ksiaa16r(seqidno:220)或ksiaa16s(seqidno:218)的天然拟南芥iaa16启动子(atiaa16p)。gus报告基因构建体包含直接来自ksiaa16上游的2.5kb基因组dna片段,其期望包含大多数启动子元件以及驱动uida的5'utr。所有构建体在t-dna区中都包含大观霉素抗性基因(spec)或草铵膦抗性基因(bar)以作为选择性标记。对于所有构建体,收获t1种子,进行表面灭菌,然后平板接种在合适的选择性培养基上以鉴别独立的t1转化体。选择性培养基上单个种子下降(decent)和分离被用来估计转基因插入数并鉴别纯合株系。为了诱导p-alc启动子,就在倾倒板之前添加乙醇至终浓度为0.01%,并立即使用板。所有板均在22℃的percival生长室(percivalscientific,inc.)中生长,进行16小时/8小时黑暗光照循环,光强度为200μmol/min/m2。相对湿度保持在50%。根据标准方案进行gus分析。简而言之,将二甲基甲酰胺中的20mg/mlx-gluc(goldbio,st.louis,mo)储备液稀释至ph7.0的50mm磷酸盐缓冲液中,达到最终浓度为1mg/ml。将此溶液添加到植物组织中,并将样品置于真空下5分钟,以确保溶液渗透到组织中。将样品用箔纸覆盖以防止光线进入,并在37℃下放置24小时以允许显色。移除染色溶液并且通过将样品在50%乙醇中孵育数小时来清除叶绿素。对于幼苗停滞计数,表现出明显的突起但在平板接种后第8天根的长度从未超过2-3mm的幼苗被认为是停滞的。
携带35s:ksiaa16构建体和p-alc:ksiaa16构建体的株系中,很大一部分幼苗似乎在发芽后不久就停滞,这可能是由于高水平的组成型/基础表达造成的(表5)。使用0.01%乙醇诱导p-alc启动子的表达稍微增大了停滞率,但不明显。在存在或不存在乙醇的情况下,35s:ksiaa16株系没有差异。
表5:转基因拟南芥幼苗停滞数据
携带atiaa16p:ksiaa16构建体的株系似乎没有明显的幼苗停滞量,因此被用于进一步研究。如表6所示,携带atiaa16p:ksiaa16r的四个独立转化体株系在根伸长抑制分析中显示出对麦草畏的抗性,而携带atiaa16p:ksiaa16s的四个株系中只有一个显示出非常轻微的抗性。来自两种构建体的几个株系也显示出对iaa的根抑制作用的轻微抗性。综上所述,这些数据支持ksiaa16r等位基因在拟南芥中的功能与其在地肤中的功能相似的假说。此外,ksiaa16s等位基因的推定过表达似乎有时会引起对外源生长素的抗性,尽管程度较小。不希望受特定理论的束缚,这可能是由于与野生型植物相比,存在的aux/iaa蛋白总量增加,因为先前已经报道了根伸长分析中对iaa抗性的稍微增加是由于野生型aux/iaa基因ptriaa14的异源表达引起的(liu等人,2015)。总体而言,这些结果表明,ksiaa16转基因的表达方式/表达量影响了生长素感知,因为只有那些具有最接近拟南芥同源物(ataa16p)的内源启动子的植物产生了具有最少劣型的植物。
表6:植物营养素加上iaa或麦草畏下t2转基因的根长(mm)
为了了解ksiaa16的表达模式以及时空表达与观察到的地肤抗性表型相比如何,对地肤组织进行了northern分析。另外,产生了ksiaa16启动子(p):β-葡萄糖醛酸酶(gus)报告基因构建体,并在拟南芥中转化,以使ksiaa16在异源系统中的表达可视化。通过x-gluc染色对拟南芥中ksiaa16p:gus构建体的表达模式的可视化表明了报告基因不在干燥或吸胀的种子中表达,但在发芽时或刚发芽后就被强劲表达。表达在大多数组织中广泛出现,其中在维管组织和侧根原基中表达最高。在整个发育过程中均保持强劲表达,其中在所有叶片和花序组织中均可见弥漫性染色,而在维管组织中则表现最强。花朵的花粉和心皮、幼小长角果的隔膜和成熟长角果的脱落区中也明显有染色。来自敏感性和抗性株系的地肤组织的northern印迹分析显示,ksiaa16转录物的内源等位基因在所有分析的组织中都被转录。ksiaa16的这种广泛表达模式可以解释对叶面麦草畏喷雾的强抗性。
实施例8:作物植物中aux/iaa基因的修饰
基于在ksiaa16蛋白的降解决定子结构域内所鉴别的引起甘氨酸到天冬酰胺氨基酸变化的碱基对替换,可以在作物植物的iaa16直系同源物中进行对应的碱基对替换,所述作物植物诸如玉米、高粱、小麦、油菜、大豆、棉花、葡萄、番茄、马铃薯、莴苣、西兰花、黄瓜、花生、甜瓜、韭菜、洋葱、水稻、大麦或本文确定的其他直系同源物。可以使用靶向基因组编辑(例如单链寡核苷酸介导的基因组编辑、大范围核酸酶、锌指核酸酶、talen或rna引导的核酸酶(例如crispr技术)来对这些序列进行修饰。
表7:ksiaa16进化枝的直系同源物
可以按照所描述的那样(sauer,等人(2016))完成精确的基因组编辑。单链寡核苷酸用于工程改造所需植物物种的aux/iaa16直系同源物的降解决定子基序中的gg到aa核苷酸变化。通过分子分析、核苷酸变化的基因组序列确认和生长素敏感性分析来进行引入突变的选择。
或者,将一种或多种位点特异性核酸酶,例如锌指核酸酶、talen或rna引导的核酸酶(例如,crispr-cas9、crispr-cpf1、crispr-casx等)与模板一起引入植物细胞中,所述模板包含编码突变的降解决定子基序(gg至aa核苷酸变化)的外显子。位点特异性核酸酶在染色体上在编码降解决定子基序的aux/iaa16直系同源物的外显子周围诱导双链断裂(dsb)。通过同源重组,模板替代了编码野生型降解决定子基序的内源外显子。使用标准分子分析和生长素敏感性分析选择携带突变的降解决定子基序的植物。
或者,将一种或多种位点特异性核酸酶,例如锌指核酸酶、talen或rna引导的核酸酶(例如,crispr)与模板一起引入植物细胞中,所述模板包含编码突变的降解决定子基序(gg至aa核苷酸变化)的外显子。位点特异性核酸酶在染色体上诱导两个双链断裂(dsb):编码降解决定子基序的aux/iaa16直系同源物的外显子的一个dsb5'-和一个dsb3'。通过非同源末端连接,模板替代了编码野生型降解决定子基序的内源外显子。使用标准分子分析和生长素敏感性分析选择携带突变的降解决定子基序的植物。
或者,将碱基编辑酶(例如,cas9-胞苷脱氨酶、cpf1-胞苷脱氨酶等)引入植物细胞。碱基编辑酶催化编码降解决定子基序的aux/iaa16直系同源物的外显子中的c至t碱基转化,导致降解决定子基序中的甘氨酸(g)到天冬酰胺(n)氨基酸替换。使用标准分子分析和生长素敏感性分析选择携带突变的降解决定子基序的植物。
实施例9.r型地肤生物型对2,4-d和氟草烟的交叉抗性
为了阐明r型地肤生物型是否对2,4-d或氟草烟具有交叉抗性,对生长在温室中的10至15厘米高的地肤植物用履带喷雾器分别喷洒0、140、280、560、2240、4480、6720、8960或17920ga.e.ha-1的麦草畏;或0、70、140、280、560、1120、2240、4480、6720、8960ga.e.ha-1的2,4-d,或0、17.5、35、70、140、280、420、560、1120、2240ga.e.ha-1的氟草烟。在21dat确定鲜重,并使用“r”软件将其用于计算剂量反应曲线。r型地肤(实心圆)和s型地肤(空心圆)的曲线显示在图6中,a图中为麦草畏、b图中为2,4-d且c图中为氟草烟。对于所有处理均观察到r和s生物型在处理后14天(dat)的可见损伤(未示出)。
- 还没有人留言评论。精彩留言会获得点赞!