鞭毛蛋白和/或壳六糖在用于制备抑制植物光合作用的抑制剂中的应用的制作方法
[0001]
本发明属于调节植物光合作用的技术领域,具体涉及鞭毛蛋白和/或壳六糖在用于制备抑制植物光合作用的抑制剂中的应用。
背景技术:
[0002]
在自然环境中,植物难以避免地接触到各种微生物,其中有些会通过侵染植物以达到自身生长繁殖的目的。植物受侵染时会加快叶片的衰老及脱落,植物本身为防止病原微生物进一步扩散,而使侵染点周围细胞快速的程序性死亡,这种现象叫超敏反应,是植物免疫反应的常见机制。植物通过模式识别受体(prr)识别微生物保守的必要成分快速地启动免疫通路,能够被这种受体识别的病原微生物保守成分称为病程相关性分子模式(pamps),进而释放出各种抵御胁迫的信号分子,如钙离子、活性氧以及各种植物激素等。
[0003]
植物体可以利用光能,在叶绿体内将co2和h2o合成有有机物供自己的生命活动,并释放出氧气,这个过程被称为光合作用。它分为光反应和暗反应,光反应主要由ps ii和psi等复合蛋白参与将光能转化为化学能,暗反应则利用光反应产生的能量和物质将co2转化为有机物质,这个过程也就是卡尔文循环。植物光合作用负责地球上大部分氧气生成和生物量的固定。所以当植物受到胁迫时,植物细胞中则会分出大量精力抵御胁迫,这种情况下植物生理生化指标必然有所降低,这其中自然也包括光合作用。
技术实现要素:
[0004]
本发明的目的是提供鞭毛蛋白和/或壳六糖在用于制备抑制植物光合作用的抑制剂中的应用。本发明所述应用,可以有效引起植物免疫反应,从而抑制植物光合作用,具有良好的应用前景。
[0005]
为实现上述发明目的,本发明的技术方案如下:
[0006]
本发明提供了鞭毛蛋白和/或壳六糖在用于制备抑制植物光合作用的抑制剂中的应用。
[0007]
进一步的,所述鞭毛蛋白和壳六糖作用于所述植物的叶片上的模式识别受体,引起免疫反应。
[0008]
进一步的,所述鞭毛蛋白和壳六糖引起所述植物的叶片中ros爆发。
[0009]
进一步的,所述鞭毛蛋白和壳六糖诱导pr基因表达。
[0010]
进一步的,所述鞭毛蛋白和壳六糖降低ps ii反应中心密度。
[0011]
进一步的,ca
2+
信号完整的情况下,flg22触发的免疫调节上调ps ii外周蛋白基因psbo和psbp的表达。
[0012]
进一步的,所述壳六糖处理会引起psbo和psbp都会显著下调。
[0013]
进一步的,所述壳六糖诱导的免疫通路引起psbs基因下调以及vde基因上调。
[0014]
进一步的,所述鞭毛蛋白和壳六糖降低所述植物的叶片的npq。
[0015]
进一步的,所述植物为有类囊体的植物。
[0016]
本发明还提出了所述鞭毛蛋白和/或壳六糖与钙化合物的组合物在用于制备抑制植物光合作用的抑制剂中的应用。
[0017]
进一步的,所述钙化合物为四水硝酸钙。
[0018]
与现有技术相比,本发明的优点和技术效果在于:本发明所述的鞭毛蛋白和/或壳六糖明显促进植物叶片pr-4和pr-10基因的表达,同时,降低ps ii反应中心密度,降低ps i活性,降低了花生叶片的能量耗散,从而降低植物光保护能力,达到光合作用抑制剂的效果,在抑制光合作用上作用范围广、可靠性高、反应快速。
[0019]
本发明还提出了钙离子协同促进抑制光合作用,含钙化合物中的ca
2+
为植物生长过程的基本需求,而植物生长过程中所必须的钙离子可以在抑制光合作用时有促进效果。本发明提出的应用通过触发植物的超敏反应,使植物侵染点周围细胞快速的程序性死亡,加快花生叶片的衰老及脱落而实现。
[0020]
现代农业所使用的除草剂主要有光合作用抑制型、色素合成抑制型、氨基酸生物合成抑制型和脂类合成抑制型四大类。本发明所述的鞭毛蛋白和/或壳六糖作为光合作用抑制剂,在制备生物源除草剂上是很可观的。而且鞭毛蛋白和/或壳六糖资源丰富、易降解、无残毒、作用对象广、不破坏生态环境、经济效益高,具有良好的市场应用前景。
附图说明
[0021]
图1是flg22处理下花生叶片中ros含量变化,a:1μm flg22处理下花生叶片h2o2的含量变化;b:1μm flg22处理下花生叶片o
2-的含量变化,注:*:与对照相比差异显著(p<0.05)。
[0022]
图2是壳六糖处理下花生叶片中ros含量变化,a:200μg/ml壳六糖处理下花生叶片h2o2的含量变化;b:200μg/ml壳六糖处理下花生叶片o
2-的含量变化,注:*:与对照相比差异显著(p<0.05)。
[0023]
图3是flg22处理对花生叶片中病原相关基因表达的影响,a:flg22处理下花生叶片pr-4基因的表达情况;b:flg22处理下花生叶片pr-10基因的表达情况,注:*:与对照相比差异显著(p<0.05)。
[0024]
图4是壳六糖处理下花生叶片中病原相关基因的表达模式,a:壳六糖处理下花生叶片pr-4基因的表达情况;b:壳六糖处理下花生叶片pr-10基因的表达情况,注:*:与对照相比差异显著(p<0.05)。
[0025]
图5是flg22处理下花生单位反应中心的活性,a:1μm flg22处理下花生叶片abs/rc的变化;b:1μm flg22处理下花生tr
o
/rc的变化;c:1μm flg22处理下花生叶片et
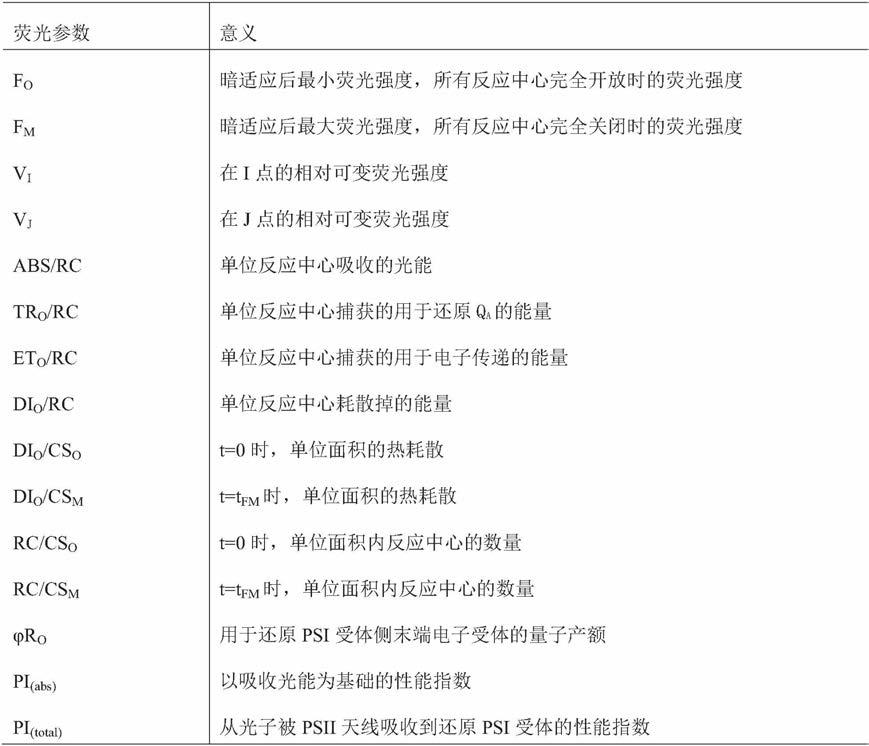
o
/rc的变化;d:1μm flg22处理下花生di
o
/rc的变化,注:*:与对照相比差异显著(p<0.05)。
[0026]
图6是壳六糖处理下花生单位反应中心的活性,a:200μg/ml壳六糖处理下花生叶片abs/rc的变化;b:200μg/ml壳六糖处理下花生tr
o
/rc的变化;c:200μg/ml壳六糖处理下花生叶片et
o
/rc的变化d:200μg/ml壳六糖处理下花生di
o
/rc的变化,注:*:与对照相比差异显著(p<0.05)。
[0027]
图7是flg22处理下花生叶片反应中心密度变化,a:1μm flg22处理下花生叶片rc/cs
o
变化;b:1μm flg22处理下花生叶片rc/cs
m
变化,注:*:与对照相比差异显著(p<0.05)。
[0028]
图8是壳六糖处理下花生叶片反应中心密度变化,a:200μg/ml壳六糖处理下花生叶片rc/cs
o
变化;b:200μg/ml壳六糖处理下花生叶片rc/cs
m
变化,注:*:与对照相比差异显著(p<0.05)。
[0029]
图9是flg22处理下花生叶片pi
(abs)
变化,注:*:与对照相比差异显著(p<0.05)。
[0030]
图10是壳六糖处理下花生叶片pi
(abs)
变化,注:*:与对照相比差异显著(p<0.05)。
[0031]
图11是flg22处理下花生叶片的变化,注:*:与对照相比差异显著(p<0.05)。
[0032]
图12是壳六糖处理下花生叶片的变化,注:*:与对照相比差异显著(p<0.05)。
[0033]
图13是flg22处理下花生叶片pi
(total)
的变化,注:*:与对照相比差异显著(p<0.05)。
[0034]
图14是壳六糖处理下花生叶片pi
(total)
的变化,注:*:与对照相比差异显著(p<0.05)。
[0035]
图15是flg22处理下花生叶片单位面积的能量耗散变化,a:1μm flg22处理下花生叶片di
o
/cs
o
变化;b:1μm flg22处理下花生叶片di
o
/cs
m
变化,注:*:与对照相比差异显著(p<0.05)。
[0036]
图16是壳六糖处理下花生叶片单位面积的热耗散变化,a:200μg/ml壳六糖处理下花生叶片di
o
/cs
o
变化;b:200μg/ml壳六糖处理下花生叶片di
o
/cs
m
变化,注:*:与对照相比差异显著(p<0.05)。
[0037]
图17是flg22处理下花生叶片npq的变化,注:*:与对照相比差异显著(p<0.05)。
[0038]
图18是壳六糖处理下花生叶片npq的变化,注:*:与对照相比差异显著(p<0.05)。
[0039]
图19是flg22处理下花生psbo和psbp相对表达水平,a:1μm flg22处理下花生叶片psbo基因相对表达水平;b:1μm flg22处理下花生叶片psbp基因相对表达水平,注:*:与对照相比差异显著(p<0.05)。
[0040]
图20是壳六糖处理下花生psbo和psbp相对表达水平,a:200μg/ml壳六糖处理下花生叶片psbo基因相对表达水平;b:200μg/ml壳六糖处理下花生叶片psbp基因相对表达水平,注:*:与对照相比差异显著(p<0.05)。
[0041]
图21是flg22处理下花生psbs和vde相对表达水平,a:1μm flg22处理下花生叶片psbs基因相对表达水平;b:1μm flg22处理下花生叶片vde基因相对表达水平,注:*:与对照相比差异显著(p<0.05)。
[0042]
图22是壳六糖处理下花生psbs和vde相对表达水平,a:200μg/ml壳六糖处理下花生叶片psbs基因相对表达水平;b:200μg/ml壳六糖处理下花生叶片vde基因相对表达水平,注:*:与对照相比差异显著(p<0.05)。
[0043]
图23是flg22处理下花生cp12相对表达水平,注:*:与对照相比差异显著(p<0.05)。
[0044]
图24是壳六糖处理下花生cp12相对表达水平,注:*:与对照相比差异显著(p<0.05)。
具体实施方式
[0045]
下面结合附图和具体实施例,对本发明的技术方案做进一步详细的说明。
[0046]
1、实验仪器
[0047]
连续激发式荧光仪(handy pea)、便携脉冲调制式荧光仪(fms-2)、紫外可见分光光度计(uv-1750)、abi 7500fast荧光定量pcr仪、冷冻离心机等等。
[0048]
2、实验试剂和试剂盒
[0049]
鞭毛蛋白flg22(sigma)、壳六糖(solarbio)、硫酸钾、七水硫酸镁、磷酸二氢钾、四水硝酸钙、乙二胺四乙酸二钠、硫酸亚铁、硼酸、五水硫酸铜、七水硫酸锌、碳酸氢铵、钼酸、丙酮等。总rna提取试剂盒购自天根生化科技有限公司、primescript
tm
rt reagent kit with gdna eraser购自takara、过氧化氢(h2o2)试剂盒(h2o2-2-y)和超氧阴离子试剂盒(sa-2-g)购自苏州科铭生物技术有限公司。
[0050]
实施例1:pamps的选取
[0051]
本发明选用鞭毛蛋白flg22(sigma)、壳六糖(solarbio)作为光合作用抑制剂的原材料,研究其引发的免疫通路对抑制光合作用的影响。
[0052]
鞭毛蛋白flg22(以下简称flg22)属于蛋白类pamps。鞭毛是细菌运动器官,它使细菌能在面对刺激时作出反应。鞭毛蛋白则是来自于各种细菌鞭毛中保守的n末端或c末端。因为这些特点,动植物细胞能够识别出鞭毛蛋白。
[0053]
壳六糖是一种壳聚糖,属于糖类pamps。壳聚糖是一种天然、生物可降解的高分子材料,在美容、食品、生物、医药、农业等领域有着广泛的应用,是脱乙酰化的几丁质。与几丁质一样,虽然是高聚合物,但是壳聚糖片段——壳寡糖一样具有生物学效应,且具有良好的水溶性。
[0054]
实施例2:叶片材料培养
[0055]
1、实验方法
[0056]
(1)选取籽粒饱满,大小相近的花生(arachis hypogaea.花育25),将花生放在去离子水中浸泡40分钟,转入铺有湿润纱布的塑料白盆中,确保花生不堆叠,再往上面再一层湿润纱布为种子提供潮湿透气的环境,将白盆置入培养箱28℃培养两天等待发芽。
[0057]
(2)发芽后的种子放入装有石英砂的小塑料盆中(上口径9cm,下口径6.5cm,高8cm),石英砂用去离子水反复清洗,将底部有孔的小塑料盆放在白色大盆中覆膜,转入温室培养。温室每天白天持续14h光照(400μmol m-2
s-1
),温度为25℃/20℃(昼/夜)。去离子水培养大概4天去膜。
[0058]
(3)将上一步得到的花生苗分为有钙(ca)和无钙(nc)两组,ca组浇完全hoagland营养液,nc组将hoagland营养液里面的四水硝酸钙去除,用碳酸氢铵平衡氮。浇灌前将各部分营养液母液混合稀释到终浓度后ph调到6.5,大塑料盆中每天更换新的营养液。
[0059]
(4)培养20天后,取花生功能叶,离体叶片放在加入去离子水的培养皿中暗适应4h,测ca和nc组的相关初始数据,再都分别加入1μm flg22和200μg/ml壳六糖处理1h、2h和4h。而测活性氧和提取rna则将材料处理后送液氮速冻保存在-70℃。
[0060]
实施例3:荧光参数测定
[0061]
1、实验方法
[0062]
(1)叶绿素快速荧光动力学参数测定:用连续激发式荧光仪(handy pea),将各组离体叶片充分暗适应后,夹在探头上的叶夹上,全程避光测量,荧光参数见表1。
[0063]
表1各种荧光参数
[0064][0065]
(2)荧光猝灭参数测定:用携脉冲调制式荧光仪(fms-2),将各组离体叶片充分暗适应后,夹扣上金属片的叶夹上,将带着叶片的叶片夹靠近探头,测之前打开金属片运行程序便可。
[0066]
实施例4:总rna提取
[0067]
1、实验方法
[0068]
花生叶片总rna提取,参考tiangen试剂盒说明书。
[0069]
(1)研钵放在液氮中预冷后,用液氮将花生叶片充分研磨,取0.1g研磨粉末放入转悠1ml裂解液rz的1.5ml离心管中,放在振荡器上振荡片刻保证混合均匀。
[0070]
(2)将样品在25℃下静置5min。
[0071]
(3)将预设并已经达到4℃的冷冻离心机调到12000rpm,5min,离心后将上清尽量全部吸入另一个空的1.5ml离心管。
[0072]
(4)加入200μl氯仿(未开封或者rna专用),盖盖后在振荡器上振荡15sec后室温静置3min。
[0073]
(5)离心10min(温度和转速同(3)),将其中的无色水相转到新管中,此过程不要吸到中间层和黄色有机相,同时尽量多吸保证后期rna纯净度和浓度。
[0074]
(6)再加入300μl无水乙醇(提前放4℃冰箱预冷)混匀,将得到的溶液和可能出现的沉淀全部转入吸附柱cr3中,4℃12000rpm离心30sec,弃废液。
[0075]
(7)向吸附柱cr3加入500μl去蛋白液rd(已经加入乙醇,并放在了4℃冰箱中预冷),离心30sec(温度和转速同上),弃废液。
[0076]
(8)向吸附柱cr3加入500μl去漂洗液rw(已经加入乙醇),然后室温静置2min,离心30sec(温度和转速同上),弃废液,并且重复操作一次。
[0077]
(9)再离心2min(温度和转速不变),弃废液后,将其放在靠近酒精灯的地方晾干片刻。
[0078]
(10)将吸附柱cr3转入新的1.5ml离心管中,加50μl无rna酶ddh2o在吸附柱薄膜上,室温静置2min后离心2min(温度和转速同上)。
[0079]
(11)将收集的rna置于冰上做后续实验,或者放在-70℃备用。
[0080]
实施例5:rna浓度和纯度检测
[0081]
将1μl实施例3得到的rna与49μl无rna酶ddh2o混匀在新的rnase free离心管中,用eppendorf biophotometer测定rna浓度和纯度。
[0082]
实施例6:rna反转录
[0083]
1、实验方法
[0084]
花生rna反转录,参考takara的primescript
tm
rt reagent kit with gdna eraser说明书。
[0085]
(1)去除基因组dna
[0086]
在冰上按照以下顺序添加试剂
[0087][0088][0089]
其中total rna是根据实施例3中得到的提取的rna浓度,以2μg为标准换算所需要的毫升数;
[0090]
混匀后42℃反应两分钟,放置在冰上。
[0091]
(2)反转录反应
[0092]
在冰上按照以下顺序添加试剂
[0093][0094]
混匀后在37℃反应15min,再在85℃反应5sec,转置冰上备用,或者放-20℃保存。
[0095]
实施例7:real time pcr反应
[0096]
1、实验方法
[0097]
表2为基因引物序列,以花生tua5为内参基因,具体步骤如下:
[0098]
(1)反应体系配置
[0099]
在冰上按照以下顺序配置
[0100][0101][0102]
用于rt pcr反应的cdna浓度是实施例5中反转录后稀释5倍的浓度。
[0103]
(2)反应程序的设置
[0104]
步骤1:预变性,95℃,30sec;
[0105]
步骤2:pcr反应,40个循环,95℃,5sec;60℃,34sec;
[0106]
表2基因引物序列
[0107][0108]
实施例8:活性氧测定
[0109]
1、实验方法
[0110]
具体参照过氧化氢(h2o2)试剂盒和超氧阴离子(orf)试剂盒说明书。
[0111]
(1)flg22对花生叶片活性氧含量的影响
[0112]
ros在植物促进组织修复和抵抗病原微生物上扮演者重要角色,在植物早期免疫反应中往往伴随着活性氧的爆发。结果由图1a可知,在flg22处理1h和2h后ca和nc花生叶片中h2o2均显著上升趋势,其中1h上升幅度相对其它时间更高,之后活性氧含量开始降低,在4h的时候恢复正常,二者趋势基本一致。图1b显示,flg22处理后花生叶片中o
2-含量均显著上升,且ca处理的含量明显低于nc处理。结果表明:flg22触发的免疫反应会引起花生叶片中ros的爆发,ca
2+
对o
2-的积累有明显的抑制作用。
[0113]
(2)壳六糖对花生叶片活性氧含量的影响
[0114]
利用壳六糖处理后,对花生叶片进行了活性氧含量的测定。结果由图2a可知,利用壳六糖处理花生叶片,在1h和2h后花生叶片中h2o2都呈显著上升趋势,在4h h2o2含量降低到大约初始水平,且ca和nc处理无明显差异。由图2b显示,花生叶片中经过壳六糖处理后,o
2-含量在开始时有所上升,然后呈现下降趋势,且ca和nc处理差异不显著。表明:壳六糖触发的免疫反应会引起花生叶片中活性氧的爆发。
[0115]
2、实验结果
[0116]
flg22和壳六糖会触发花生免疫通路,且这个过程涉及ca
2+
信号通路。活性氧(ros)是免疫过程中的信号分子,ca
2+
的存在会提高花生叶片活性氧的清除能力。
[0117]
实施例9:植物叶片病原相关基因表达
[0118]
1、鞭毛蛋白和壳六糖触发的病原相关基因表达
[0119]
植物在与pamps或者致病性病原微生物接触时,都会触发自身免疫反应,在其下游信号中,会由丝裂原活化蛋白激酶(mapk)级联信号最后引起病原相关基因pr基因表达的变化。pr基因家族在不同植物中发现大约有17种,它可以被生物或非生物胁迫诱导。在许多情况下,是对防御相关的植物激素如水杨酸、茉莉酸和乙烯作出反应的标记基因,本发明选择pr基因家族中的pr-4和pr-10检测。pr-4与编码几丁质酶有关,有抗真菌活性。pr-10基因家族编码具有胞质定位的小蛋白,这些小蛋白起着rna酶、翻译后修饰等作用,并且对非生物和生物因子都有应答。
[0120]
(1)flg22处理下花生叶片中病原相关基因的表达模式
[0121]
pr基因可以编码病原相关蛋白,当植物体被病原微生物侵染时这种蛋白被诱导,参与植物的防御反应。如图3所示,ca组花生幼苗经过flg22处理后,在1、2和4h时pr-4基因都显著上调,尤其是1和2h上调幅度最大,4h相对1和2h有所缓和。nc花生幼苗在flg22处理后,在1和2hpr-4基因都显著上调,但是上调幅度明显不如有钙,到了4h则不显著。无论有钙还是无钙幼苗的pr-10基因表达都呈显著上升,但是有钙趋势相对较为稳定。
[0122]
(2)壳六糖处理下花生叶片中病原相关基因的表达模式
[0123]
利用壳六糖处理花生幼苗叶片,对病原相关基因进行检测。由图4a我们可以看到,花生幼苗经过壳六糖处理后,ca叶片在1、2和4h pr-4和pr-10基因都显著上调,nc的花生幼苗在壳六糖处理4hpr-4基因显著上调,而pr-10基因表达在4h出现显著,nc相对ca上升幅度较小(图4b)。
[0124]
2、结果
[0125]
flg22和壳六糖诱导花生叶片pr基因的表达,且ca
2+
信号介导了壳六糖对花生叶片病原相关基因表达的调控,起到稳定免疫反应的作用。
[0126]
实施例10:反应中心活性
[0127]
1、psii反应中心的活性
[0128]
(1)flg22处理后的单位ps ii反应中心的活性
[0129]
绿色植物细胞中的天线色素将吸收的捕获的能量传递给反应中心,反应中心利用这部分能量还原q
a
,如此将电子传递给后面的光合反应通路,而abs/rc、tr
o
/rc、et
o
/rc三个参数反应的分别表示以上几个过程,di
o
/rc则表示单位反应中心的能量耗散。由图5可知abs/rc、tr
o
/rc、di
o
/rc这三个参数变化趋势相同,在flg22处理后的1h和4h,ca处理均显著
上升,而nc只在4h显著上升。et
o
/rc中的ca只在1h出现显著上升,2h、4h也有上升趋势但不显著,nc在4h显著上升。结果表明:flg22处理会增加花生叶片ps ii单位反应中心的活性。
[0130]
(2)壳六糖处理后的单位ps ii反应中心的活性。
[0131]
由图6a可知,经过壳六糖处理后,abs/rc在ca和nc的均有所上升;由图6b可知,壳六糖处理ca和nc tr
o
/rc都出现上升;图6c显示,经过壳六糖处理ca和nc的et
o
/rc上升趋势与abs/rc和tr
o
/rc相似;图6d显示壳六糖处理nc的di
o
/rc整体上升不明显,ca在2h显著上调。结果表明:壳六糖处理后,会一定程度增加花生叶片ps ii单位反应中心的活性,但与ca
2+
信号关系不大。
[0132]
2、反应中心密度
[0133]
(1)flg22处理后的反应中心密度
[0134]
rc/cs反应单位面积中反应中心的数量,rc/cs
o
和rc/cs
m
分别表示t=0和t=t
fm
对应的反应中心密度。由图7a可知,在flg22处理后1h,ca处理rc/cs
o
和显著下降,nc则是4h显著下降。而图7b中,ca处理rc/cs
m
在1h和4h显著下降,nc组4h显著下降。结果表明:flg22触发的免疫反应降低了反应中心密度,而反应中心数量的减少肯定会增加单个反应中心的负担,这是单个ps ii反应中心活性增加的原因。处理前期,ca反应中心密度明显低于nc,这是因为钙对flg22触发的免疫反应有着积极的响应,能够更快地调动资源用于免疫通路,而影响到反应中心密度。
[0135]
(2)壳六糖处理后反应中心密度
[0136]
壳六糖处理后,ca和nc的rc/cs
o
在2h和4h均出现显著下调(图8a),而rc/cs
m
(如图8b)的ca组在2h出现显著下调。结果表明:壳六糖触发的免疫反应降低了反应中心密度,而增加了单个反应中心的负担,但与ca
2+
信号途径关系不大。
[0137]
3、叶片吸收光能的性能指数
[0138]
(1)flg22处理后吸收光能的性能指数
[0139]
pi
(abs)
是可以综合反应ps ii光应中心密度、光吸收以及电子传递的重要参数。由图9可知,ca组叶片pi
(abs)
先是降低然后基本保持稳定,而nc组则基本保持不变。这表明flg22诱导的免疫通路响应过程中,ca
2+
信号传导途径会降低ps ii反应中心的还原力。钙离子是植物免疫通路的重要离子信号,所以有ca免疫通路的触发会比nc更加迅速,影响范围也会更广。
[0140]
(2)壳六糖处理后吸收光能的性能指数
[0141]
与前面flg22不同,壳六糖处理后,虽然反应中心减少,单个反应中心活性增加,但是叶片吸收光能综合性能指数pi
(abs)
并没有改变(图10)。结果表明:壳六糖诱导的免疫通路没有对pi
(abs)
产生影响。通过pr基因的相对表达情况看出,壳六糖引起的免疫反应程度flg22触发的免疫明显,这是因为壳六糖不像flg22处理后pi
(abs)
产生差异性。
[0142]
4、实验结果
[0143]
pamps触发的免疫反应会降低反应中心密度,增加花生叶片ps ii单位反应中心的活性。处理前期,由于钙对flg22触发的免疫反应有着积极的响应,能够更快地调动资源用于免疫通路,因此ca反应中心密度明显低于nc,且ca
2+
能加快这部分的响应速度。
[0144]
pamps诱导的免疫通路响应过程中,ca
2+
信号传导途径会降低ps ii反应中心的还原力。钙离子是植物免疫通路的重要离子信号,所以有ca免疫通路的触发会比nc更加迅速,
影响范围也会更广。
[0145]
实施例11、ps i的相对活性
[0146]
1、psi受体侧末端电子受体的量子产额
[0147]
是还原psi受体侧末端电子受体的量子产额,可以反应ps i的相对活性。
[0148]
由图11可知,1μm flg22处理1h和2h,ca叶片的显著降低,而nc在2h时显著降低,其余处理时间则不显著。结果表明:flg22诱导的免疫调节降低了ps i的相对活性,而ca
2+
的响应速度似乎更积极。
[0149]
图12显示在壳六糖处理前后变化ca和nc不显著。结果表明:壳六糖处理不会影响花生叶片ps i的受体侧量子产额。
[0150]
2、综合性能指数
[0151]
pi
(total)
是综合反应从ps ii的能量吸收到ps i受体侧末端的电子传递能力的参数。由图13可知,在1μmflg22处理1h、2h和4h,ca叶片的pi
(total)
显著降低,而nc在2h时pi
(total)
显著降低,其余处理时间则不显著。结果表明:flg22诱导的免疫调节降低了花生叶片的光合活性,而ca
2+
响应更为积极。
[0152]
由图14可知,壳六糖处理花生叶片前后ca和nc组的pi
(total)
不显著。结果表明:壳六糖处理不会影响综合性能指数pi
(total)
。
[0153]
实施例12能量耗散
[0154]
在光合作用过程中,捕获的光能主要通过光化学电子传递、叶绿素荧光发射和热耗散消耗。光化学电子传递与光合产物的合成有关,叶绿素荧光发射仅是光能消耗的一小部分。过剩的能量会损伤ps ii,因此,热耗散是消耗多余光能、防止光损伤的重要途径。将多余的激发能以无害的方式作为热耗散的过程称为非光化学猝灭(npq)。一般高强度的光照或者反应中心效能下降都会增加能量耗散。
[0155]
ps ii反应中心复合物的重组是npq的常见机制,包括d1蛋白的可逆失活和合成再生,ca
2+
处理植物中d1蛋白含量较高。光合反应中心蛋白组分的周转受钙调素(cam)调控,cam是ca
2+
信号转导通路的重要成分。拟南芥突变体vde活性缺失突变体npq1,在npq1中,由于vde中的点突变从而阻止了玉米黄素的积累。而拟南芥突变体psbs缺失突变体npq4,玉米黄质虽然可以积累,但是npq4同样猝灭失活,npq需要同时存在vde和psbs,这是由于vde和psbs之间的相互作用造成的,psbs作用于猝灭起始位置。紫黄质脱环氧化酶(vde)的合成受到ca
2+
的影响,ca
2+
条件下,cam可以介导vde基因的表达,促进叶黄素循环。
[0156]
1、单位面积的热耗散
[0157]
di
o
/cs
o
和di
o
/cs
m
分别代表t=0和t=t
fm
对应的单位面积热耗散。由图15可以看出只有nc在flg22处理4h时di
o
/cs
o
和di
o
/cs
m
显著下降,而ca则都不显著。结果表明:ca
2+
对flg22处理下花生叶片的单位面积热耗散影响不大。
[0158]
由图16可以看出,壳六糖处理后,ca组中,di
o
/cs
o
在2h和4h显著下调,di
o
/cs
m
在2h出现显著下调;nc组中,di
o
/cs
o
在4h出现显著下调。结果表明:壳六糖处理后会引起单位面积热耗散的降低,但与ca
2+
的作用不大。
[0159]
2、非光化学淬灭
[0160]
npq能够消耗植物体过剩的能量,它与psbs蛋白和叶黄素循环有关,由图17可知,1μm f g22处理2h和4h,ca叶片的npq显著降低,而nc的npq高于ca叶片。结果表明:ca
2+
参与降
低flg22激发的免疫响应过程中非光化学能量耗散。
[0161]
以上结果表明:flg22诱导的免疫途径降低了花生叶片的能量耗散,且npq的下调需要ca
2+
参与。
[0162]
由图18可知,壳六糖处理ca叶片的npq明显低于nc叶片。结果表明:壳六糖诱导的免疫响应反应过程,ca
2+
参与花生叶片npq的下调。
[0163]
以上结果表明:壳六糖诱导的免疫途径降低了花生叶片的能量耗散。与flg22处理时的结果基本一致,这表明免疫响应过程中,非光化学能量耗散不是唯一散热机制。
[0164]
实施例13:光合作用相关基因表达的影响
[0165]
1、花生叶片psbo和psbp基因相对表达水平
[0166]
高等植物中,ps ii是镶嵌在类囊体膜上的多亚基复合物,psii拥有自己特异的光收集系统,该系统由两个叶绿素a结合蛋白cp43(psbc)和cp47(psbb)以及几个叶绿素a/b结合蛋白组成,其中最丰富的是光捕获复合体ii(lhc-ii)。psii利用太阳能催化一系列的电子转导反应,并引起水分子裂解成氧气、质子和电子。水解反应主要由放氧复合物催化,而放氧复合物被依附于ps ii囊腔侧的外周蛋白保护,这些外周蛋白分别是psbo、psbp和psbq。psbo几乎存在于所用的光合有机体中,而psbp和psbq只在绿藻和植物中发现,而蓝藻中分别被psbv和psbu代替。psbo参与维持mn4cao5簇的稳定性,研究表明mn4cao5簇是水氧化的必要前提,放氧复合物mn4cao5作为自由辅因子在化学上不稳定,必须在利用电荷分离的获得的能量组装在光系统ii放氧复合物上。在生物发生过程中,天然的无机辅因子mn
2+
、ca
2+
和cl-结合到新翻译psii蛋白上,通过一系列明暗的步骤组装成放氧复合体。而psbq和psbp需要cl-和ca
2+
作为必要的辅因子,ca
2+
也参与了水分解相关的s态循环。水解后产生电子被运送到烟酰胺腺嘌呤二核苷酸磷酸(nadp
+
),通过线性电子流生成nadph用于卡尔文循环。
[0167]
(1)flg22处理后的psbo和psbp基因相对表达
[0168]
psbo和psbp是ps ii外周蛋白,与ps ii的功能性稳定有重要关系。由图19a,在1h和4h,ca和nc叶片的psbo基因显著上调,而nc上升幅度更大。图19b中,psbp基因具体相同趋势。结果表明:ca
2+
信号完整的情况下,flg22触发的免疫调节上调ps ii外周蛋白基因psbo和psbp的表达,而ca
2+
信号缺失的情况下,上调幅度更大,这样会更快地促进ps ii的修复。
[0169]
(2)壳六糖处理后的psbo和psbp基因相对表达
[0170]
由图20可知,壳六糖处理后1h时ca组psbo显著下降,而nc在4h显著下降;图20b中,ca组经过壳六糖处理1h和4h psbp显著下降,而nc组1h、2h和4h都显著下降。结果表明:无论有钙还是无钙,壳六糖处理会引起psbo和psbp都会显著下调。从前面的参数可以看出壳六糖触发的免疫调控不如flg22明显,部分光合参数没有出现显著差异,表示壳六糖处理并没有对ps ii外周蛋白psbo和psbp造成太多影响,所以不需要加快ps ii蛋白的修复。
[0171]
2、花生叶片psbs和vde基因相对表达水平
[0172]
(1)处理后的psbs和vde基因相对表达
[0173]
ps ii蛋白psbs和紫黄质脱环氧化酶(vde)是非光化学猝灭(npq)组分。由图21a可知nc在flg22处理1hpsbs显著上调,而ca叶片在2h显著下调。图21b,ca在2h显著下调,nc叶片在1h和2h显著下调。由于它们都与npq有关,ca组在2h两者都下调,而nc组两者基本互补,这表明ca
2+
完整的情况下,flg22触发的免疫响应中,npq会受到抑制。
[0174]
(2)壳六糖处理后的psbs和vde基因相对表达
[0175]
图22a显示,在壳六糖处理后,ca和nc组psbs基因都显著下调,且二者差异不明显。图22b则显示nc叶片1h和4h出现显著上调,而ca的vde基因表达都出现显著上升,且上升更显著。结果表明:壳六糖诱导的免疫通路引起psbs基因下调以及vde基因上调,而ca
2+
对vde基因的表达调控作用更明显。
[0176]
3、花生叶片cp12基因相对表达水平
[0177]
(1)flg22处理后的cp12基因相对表达
[0178]
cp12是核酸编码的叶绿体蛋白,它可以与甘油醛-3-磷酸脱氢酶相互作用参与卡尔文循环。由图23可知,ca在flg22处理1h和2h时cp12显著下调,nc处理在2h和4h却显著上调。结果表明:免疫响应过程中,ca
2+
存在时会下调cp12基因的表达,而ca
2+
缺乏时,会诱导cp12基因的表达。
[0179]
(2)壳六糖处理后的cp12基因相对表达
[0180]
图24可知,壳六糖处理后ca组4h cp12出现显著下调,nc叶片的2h和4h显著下调。结果表明,壳六糖引起的免疫调节抑制cp12的基因表达,而ca
2+
减缓了这种作用。
[0181]
通过上述实验,可以得到鞭毛蛋白和壳六糖对植物光合作用产生抑制的影响。作为光合作用抑制剂,鞭毛蛋白和壳六糖可以有效的与植物叶片上的识别受体相结合,诱导植物产生免疫反应,对植物光合作用相关基因表达及能量耗散产生影响,从而达到抑制光合作用的目的,在制备生物源除草剂中具有良好的市场应用前景。
[0182]
以上实施例应理解为仅用于说明本发明而不用于限制本发明的保护范围。尽管参照前述实施例对本发明进行了详细的说明,对于本领域的普通技术人员来说,依然可以对前述实施例所记载的技术方案进行修改,或者对其中部分技术特征进行等同替换;而这些修改或替换,并不使相应技术方案的本质脱离本发明所要求保护的技术方案的精神和范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1