抗病原性装置及其用于抗真菌应用的方法与流程
1.本公开总体上涉及用于抗病原性装置的系统和方法,并且具体而言,涉及一种具有抗葡萄霜霉病菌病原体的抗真菌特性,对人类没有毒性或对环境没有不利影响的氮化硅生物陶瓷。
背景技术:
2.施用农用化学品是防止葡萄藤感染和改善收获产率的最常见方法。然而,由卵菌葡萄霜霉病菌引起的白粉病和霜霉病需要频繁施用大量的抗真菌剂。在欧盟,所有所施用的近三分之二的合成杀真菌剂用于控制这些类型的植物病原体。原产于北美,且在上世纪末偶然引入欧洲,葡萄霜霉病菌仅可通过多次气候调节的年度施用来控制。为了将健康风险和它们对环境的影响降至最低,仅使用最低浓度的有限数量的杀真菌剂,但这两个因素都增加了病原体将产生抗性的风险。由于这些原因,寻找化学治疗替代物的努力已经获得了相当大的关注,包括开发生态友好的抗真菌产品。一些新概念包括赋予植物病原体系统抗性的微生物。使用群体遗传学通过操纵宿主-病原体相互作用来培育对感染的抗性是另一种受欢迎的技术。这通常与化学控制同时进行。后一种方法起源于转录组学和分析方法。鉴于公众对健康、安全和环境问题的敏感性,寻找农用化学品的替代物已成为我们现代社会的优先事项。
3.葡萄霜霉病菌侵袭植物的所有部分,包括叶和幼果。在温和潮湿的天气的帮助下,无性孢子囊释放游动孢子,这些游动孢子最终附接至气孔并被包裹在气孔中,以形成向下延伸到气孔下腔的贯穿芽管。芽管最终转化为感染囊泡。初生菌丝出现并迅速发展出分枝,这些分枝的吸器穿透植物组织以吸取养分。经过几天的感染培养后,孢子囊柄出现,形成新的孢子囊。在生长季节期间施用大量的杀真菌剂,但是它们的类型、数量和时机取决于疾病的性质和葡萄藤的品种。在大多数地理区域,霜霉病的管理需要多种应用,从生长周期的早期开始。杀真菌剂的频繁使用、它们的高成本以及它们对环境的长期危害要求开发更有效、持久和生态友好的替代物。鉴于这些新概念和新物质的潜力,传统的杀真菌化合物应该限制在真正需要它们的条件下。
4.其中,正是考虑到这些观察,构思和开发了本公开的各个方面。
技术实现要素:
5.本文公开了用于治疗或预防植物中的病原体的方法,该方法包含使植物与包含氮化硅的组合物接触。该组合物可以包含氮化硅颗粒和水溶剂的浆料。在一些实施例中,溶剂可以包括水。在一些实施例中,组合物可包含约0.5vol.%至约20vol.%的氮化硅。接触步骤可以包括喷雾、雾化或浸渍。植物可以包括农业植物、树或藤。在一些具体实施例中,植物可以是谷物、豆类、块茎、草、油籽、蔬菜或水果。在一些其他具体实施例中,树可以是果树、景观树或林木。在又其他具体实施例中,藤可以是葡萄藤。在一个实例中,植物可以是葡萄,包括赤霞珠、卡诺娜或苏丹娜。病原体可以包括引起植物疾病的病原体,植物疾病包括霜霉
病、白粉病、葡萄孢腐病、镰孢腐病、锈病、丝核菌腐病、核盘菌腐病或菌核腐病。病原体可以是真菌,如葡萄霜霉病菌。
6.下文更彻底地描述了本发明的其他方面和迭代。
附图说明
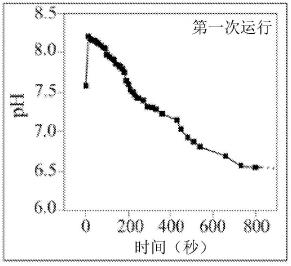
7.图1a-1c是显示在si3n4水悬浮液中随时间变化的ph测量值的图示,并且图1d是si3n4粉末分散后不久产生的气泡的图像。
8.图2a-2c是在悬浮液中带有si3n4粉末颗粒的水环境中孢子囊的显微照片;图2d-2f显示在si3n4粉末不存在的情况下包埋在纯水中的孢子囊的显微照片;并且图2g-2k显示说明那些逐渐被si3n4颗粒完全覆盖并且不释放游动孢子的孢子囊的显微照片。
9.图3a是显示孢子囊膜和si3n4颗粒之间随时间的相互作用的图像,并且图3b是图3a的放大图。
10.图4a和4b是含si3n4颗粒(图4a)和不含si3n4颗粒(图4b)的浓度为3.0x 104ml-1
的活孢子囊在水中悬浮0至2小时的荧光图像;并且图4c是分数图的图示,其显示在荧光图像上直接计数时检测到的孢子囊的总分数和活孢子囊的同时定量。
11.图5a-5c是葡萄藤物种赤霞珠的图像,其中显示对照组(图5a)、预处理的组(图5b)和共处理的组(图5c)。
12.图6a-6c是葡萄藤物种卡诺娜的图像,其中显示对照组(图6a)、预处理的组(图6b)和共处理的组(图6c)。
13.图7a和7b是室温下在纯水中(图7a)和在含有1.5vol.%的si3n4粉末的水悬浮液中(图7b)浸没10分钟后,葡萄霜霉病菌的平均拉曼光谱的图示。
14.图8a是显示nh3和nh
4+
的相对浓度的图示,并且图8b是显示水中洗脱的氮物种随ph变化的定量图的图示。
15.图9a是暴露于水中的孢子囊的dna核碱基的原始结构,观察到主要的振动模式;图9b是孢子囊的dna核碱基,显示由于被动渗透的nh3的存在而导致基因组完整性的丧失;图9c显示质子化咪唑环,其中环中的n原子都被质子化并且处于中性形式;并且图9d显示失去一个质子的去质子化咪唑环。
16.图10a和10b是显示来自光谱区1050-1150cm-1
的高分辨率拉曼信号的图示,其分别与图9a和19所示结构的带电咪唑环和中性咪唑环的振动相关。
17.图11a是显示截留si3n4颗粒的气孔中的缓冲作用和相关的氨洗脱的示意图,并且图11b是显示si3n4颗粒对孢子囊的电荷吸引的示意图,其中形成铵和氨作为在病原体/si3n4界面处有活性的抗真菌剂。
18.图12a显示接种了未处理的葡萄霜霉病菌的赤霞珠叶。图12b显示接种了用1.5vol.%的si3n4粉末处理1分钟的葡萄霜霉病菌的赤霞珠叶。
19.图13是对照和处理的葡萄霜霉病菌的赤霞珠和卡诺娜叶的受感染叶面积图。
20.图14a显示未处理的孢子囊。图14b显示si3n4存在情况下的孢子囊。
21.附图标记在附图的各图中指示对应元件。图中所使用的标题不限制权利要求书的范围。
具体实施方式
22.在下文详细论述本公开的各种实施例。虽然论述特定实施例,但是应当理解,这仅仅是出于说明的目的进行的。相关领域的技术人员将认识到,在不脱离本公开的精神和范围的情况下,可以使用其它组件和配置。因此,以下描述和附图是说明性的并且不应被解释为限制性的。描述了许多具体细节以提供对本公开的透彻理解。然而,在某些情况下,未对众所周知的或常规的细节进行描述,以避免使描述内容模糊不清。
23.在本公开的上下文内以及在使用每一术语的具体上下文中,本说明书中使用的术语通常具有它们在本领域中的普通含义。对于本文所论述的术语中的任何一个或多个术语,可以使用替代性语言和同义词,并且不管术语是否在本文详细说明或论述,都不应加以特殊意义。在一些情况下,提供某些术语的同义词。对一个或多个同义词的详述并不排除其它同义词的使用。本说明书中任何地方使用的实例(包括本文所论述的任何术语的实例)仅是说明性的,并且不希望进一步限制本公开或任何实例术语的范围和含义。同样地,本公开不限于在本说明书中给出的各种实施例。
24.对“一个实施例”或“实施例”的引用意味着结合该实施例描述的特定特征、结构或特性包括在本公开的至少一个实施例中。在说明书中各个地方出现的短语“在一个实施例中”不一定全部指代同一个实施例,也不是与其他实施例相互排斥的单独实施例或替代性实施例。此外,描述了可以由一些实施例但不由其他实施例展现的各种特征。
25.如本文所用,术语“包含(comprising)”、“具有(having)”和“包括(including)”以其开放的非限制性的意义使用。术语“一个/一种(a和an)”和“该”被理解为涵盖复数以及单数。因此,术语“其混合物(a mixture thereof)”也涉及“其混合物(mixtures thereof)”。
26.如本文所用,“约”是指数值,包括整数、分数、百分比等,无论是否明确指出。术语“约”通常是指会认为等同于所列举的值(例如,具有相同的功能或结果)的数值范围,例如,所列举的值的
±
0.5%-1%、
±
1%-5%或
±
5%-10%。
27.如本文所用,术语“氮化硅”包括si3n4、α相或β相si3n4、siyalon、siyon、sialon或这些相或材料的组合。
28.本公开涉及用于治疗或预防植物中的病原体的方法。该方法包括使植物与包含氮化硅的组合物接触。在一些实施例中,接触步骤可以包括喷雾、雾化或浸渍。
29.si3n4的表面在水环境中经历si-n共价键的均裂解离。氮和硅的释放是si3n4的生物有效性的化学来源。水解过程引发si3n4表面处氨基的质子化。相邻硅位点处的亲电性增加,这进而导致这些位点受到水的亲核攻击的倾向。当水与硅位点相互作用时,亚稳定的五配位络合物首先形成,但随后迅速衰变,释放出氨/铵离子,其比率取决于环境ph。氨(nh3)或铵(nh
4+
)的ph相关洗脱与固体表面上二氧化硅(sio2)的形成一起发生。后一种物种的水解产生原硅酸si(oh)4。si3n4在水中的基本化学反应如下:
30.si3n4+6h2o
→
3sio2+4nh3ꢀꢀꢀ
(1)
31.h
+
+nh3→
nh
4+
ꢀꢀꢀ
(2)
32.sio2+2h2o
→
si(oh)4ꢀꢀꢀ
(3)
33.在室温下,nh3的分数根据s形关系随ph变化:
34.[nh3][h
+
]/[nh
4+
]=5.7x10-10
ꢀꢀꢀ
(4)
[0035]
这表明在生理ph下游离nh3的相对分数相当低(即,洗脱的铵物种总量的1%至
2%)。然而,在ph约8.3时,它增加到约7%。图8a提供方程(4)的曲线图。它显示nh3和nh
4+
的相对浓度。在图8b中,显示它们在纯水中的浓度随ph变化的定量图。这些值是使用方程(4)从图1a的数据计算出来的,并根据公布的比色氨测定进行校准。氨可以容易地穿透病原体的细胞膜,其中它切割磷酸脱氧核糖dna主链(称为通过碱性酯交换进行的基因组切割)。然而,si3n4在水中的另一个重要方面是自由基的形成。这涉及一系列瞬态非化学计量反应,这些反应从si-n键断裂、自由电子释放和氧自由基形成开始,如下所示:
[0036]
si
–n→
si
+
+n
·
+e-ꢀꢀꢀ
(5)
[0037]
h2o+e-→h·
+oh-ꢀꢀꢀ
(6)
[0038]1o2+e-→
o2·-ꢀꢀꢀ
(7)
[0039]
o2·-+h
+
→
ho2·-ꢀꢀꢀ
(8)
[0040]
si
–
oh
2+(s)
→
si
–
oh
(s)
+h
+(aq)
ꢀꢀꢀ
(9)
[0041]
si
–
oh
(s)
→
si
–
o-(s)
+h
+(aq)
ꢀꢀꢀ
(10)
[0042]
si
–
nh
3+(s)
→
si
–
nh
2(s)
+h
+(aq)
ꢀꢀꢀ
(11)
[0043]
这些反应,方程(5)至(11),代表包括自由电子释放(方程(5))、水分子的分裂(方程(6))以及自由基氧阴离子和高氧化性质子化物种的形成(方程式(7)和(8))在内的一系列化学事件。后面这些物种有助于表面硅烷醇的解离(方程(9)至(11)),进而导致另外的氧自由基的形成,即(≡si
–o·
)和(≡si
–
o2·-)。自由电子还将氨(nh3)氧化成羟胺(nh2oh,即氨单加氧酶),并随后与水反应形成亚硝酸hno2,且产生另外的自由电子和质子。
[0044]
nh3+2e-+2h+o2→
nh2oh+h2o
[0045]
→
hno2+4e-+4h
+
ꢀꢀꢀ
(12)
[0046]
nh2oh
→
no+3h
+
3e-ꢀꢀꢀ
(13)
[0047]
2hno2→
no+no
2-+h2o
ꢀꢀꢀ
(14)
[0048]
方程(12)(即氨单加氧酶)提供催化nh3氧化所需要的自由电子,以及亚硝酸、另外的自由电子和氢质子的形成。方程(13)(即羟胺氧化还原酶)产生一氧化氮(no)、另外的自由电子和氢质子。根据方程(14)形成另外的no和亚硝酸盐(no
2-)以及来自方程(7)的氧自由基(o2·-)导致形成过氧亚硝酸根onoo-,如下所示:
[0049]o2-·
+
·
no
→-oo-n=o
ꢀꢀꢀ
(15)
[0050]
这最终导致一氧化氮(no)和过氧亚硝酸根(oono-)自由基的形成。它们是病原体最致命的药剂。在最近的si3n4和白色念珠菌相互作用的研究中,使用受激发射损耗显微镜和针对过氧亚硝酸根的硝化应力传感的特定荧光染色试剂盒,通过实验证实了过氧亚硝酸根的形成。相反,过氧亚硝酸根对植物细胞没有毒性,且no是诱导植物抵抗病原体感染的关键信号,因此对植物防御相关基因的表达产生积极的间接影响。
[0051]
i.组合物
[0052]
本公开的组合物包含氮化硅。
[0053]
在一些实施例中,可以将氮化硅粉末掺入包括但不限于浆料、悬浮液、凝胶、喷雾剂或膏剂的组合物中。在至少一个实例中,组合物可以包含分散在溶剂中的氮化硅颗粒的浆料。在一些方面,溶剂可以是水。例如,氮化硅颗粒可以与水以及任何合适的分散剂和浆料稳定剂混合,且然后通过将浆料喷洒到各种农业植物、果树、藤、谷类作物等上来施用。在至少一个实例中,可以将氮化硅浆料喷洒在受真菌感染的葡萄叶上。
[0054]
在实例中,抗病原性组合物可以是氮化硅粉末和水的浆料。氮化硅粉末可以以约0.1vol.%至约20vol.%的浓度存在于浆料中。在各种实施例中,浆料可以包括约0.1vol.%、0.5vol.%、1vol.%、1.5vol.%、2vol.%、5vol.%、10vol.%、15vol.%或20vol.%氮化硅。
[0055]
该组合物可以包括约0.5vol.%至约20vol.%的氮化硅。在一些实施例中,组合物可包括约0.5vol.%、1vol.%、2vol.%、3vol.%、4vol.%、5vol.%、6vol.%、7vol.%、8vol.%、9vol.%、10vol.%、11vol.%、12vol.%、13vol.%、14vol.%、15vol.%、16vol.%、17vol.%、18vol.%、19vol.%或约20vol.%的氮化硅。在一些另外的实施例中,该组合物可以包括约0.5vol.%至约3vol.%、约3vol.%至约6vol.%、约6vol.%至约9vol.%、约9vol.%至约12vol.%、约12vol.%至约15vol.%、约15vol.%至约18vol.%或约18vol.%至约20vol.%的氮化硅。在一个示例性实施例中,该组合物包括约1vol.%至约3vol.%的氮化硅。
[0056]
氮化硅粉末可以具有约1μm至约5μm的颗粒大小。在至少一个实例中,氮化硅粉末可以具有约2μm的颗粒大小。
[0057]
ii.病原体
[0058]
本公开的方法可用于治疗或预防植物中的许多已知的病原体。在一些实施例中,病原体可引起一种或多种植物疾病,包括霜霉病、白粉病、葡萄孢腐病、镰孢腐病、锈病、丝核菌腐病、核盘菌腐病、菌核腐病和本领域已知的其他病原性植物疾病。在一些另外的实施例中,病原体可以是真菌,包括葡萄霜霉病菌、葡萄球座菌、葡萄钩丝壳菌、富氏葡萄孢盘菌和本领域已知的其他真菌。
[0059]
iii.植物
[0060]
本公开的第i部分中公开的组合物可以施用于植物,其中植物是农业植物、树或藤。在一些实施例中,农业植物可以包括谷物、豆类、块茎、草、油籽、蔬菜或水果。
[0061]
(a)谷物
[0062]
在一些方面,谷物可以包括画眉草、小麦、燕麦、大米、玉米、大麦、高粱、黑麦、小米、黑小麦、苋菜、荞麦、藜麦、干小麦、法老小麦、翡麦或本领域已知的其他谷物。
[0063]
在一些另外的方面,豆类可以包括花生、鹰嘴豆、黄豆、豌豆、扁豆、羽扇豆、苜蓿、三叶草、豆科灌木、角豆、大豆、罗望子和本领域已知的其他豆类。
[0064]
在又另外的方面,块茎可以包括甜菜、胡萝卜、辣根、欧洲防风草、马铃薯、萝卜、甘薯、芜菁、芜菁甘蓝、芋头、荸荠、山药和本领域已知的其他块茎。
[0065]
在进一步另外的方面,草可以包括竹子、马拉姆草、草甸草、芦苇、黑麦草、甘蔗和本领域已知的其他草。
[0066]
在再其他方面,油籽可以包括棕榈、大豆、油菜籽、棕榈仁、棉籽、花生、橄榄、椰子、玉米、芝麻、亚麻子、红花、向日葵、麻风树、亚麻子、刺菜蓟、菥蓂和本领域已知的其他油籽。
[0067]
在更进一步的方面,蔬菜可以包括朝鲜蓟、芦笋、甜菜根、西兰花、抱子甘蓝、卷心菜、胡萝卜、花椰菜、块根芹、芹菜、茴香、大蒜、生姜、羽衣甘蓝、韭菜、莴苣、欧洲防风草、萝卜、沙拉菜、葱、菠菜、大葱、姜黄、芜菁、豆瓣菜和本领域已知的其他蔬菜。
[0068]
在又进一步的方面,水果可以包括苹果、鳄梨、杏、香蕉、黑莓、蓝莓、面包果、哈密瓜、樱桃、柠檬、椰子、蔓越橘、枣、无花果、葡萄柚、番石榴、蜜瓜、菠萝蜜、猕猴桃、金橘、柠
檬、酸橙、橘子、芒果、油桃、橙子、木瓜、西番莲、桃、梨、菠萝、大蕉、李子、石榴、树莓、大黄、草莓、橘子、西瓜或本领域已知的任何其他水果。
[0069]
(b)树
[0070]
在一些实施例中,树可以包括果树、景观树或林木。
[0071]
在一些方面,果树可以包括扁桃树、苹果树、杏树、鳄梨树、腰果树、樱桃树、椰子树、无花果树、葡萄柚树、番石榴树、菠萝蜜树、柠檬树、酸橙树、芒果树、橄榄树、桔树、桃树、梨树、山核桃树、李树、石榴树、核桃树或本领域已知的任何其他树。
[0072]
在一些另外的方面,景观树可以包括木兰树、苹果树、山茱萸树、枫树、白果树、桂树、云杉树、侧柏树、桦树、棕榈树、樱桃树、冬青树、山毛榉树和本领域已知的其他景观树。
[0073]
在又另外的方面,林木可以包括白蜡树、桦树、白杨树、椴树、山毛榉树、樱桃树、栗树、棉白杨、榆树、冷杉树、山核桃树、刺槐树、枫树、橡树、松树、雪松、云杉树、梧桐树、柳树和本领域已知的其他林木。
[0074]
(c)藤
[0075]
在一些实施例中,藤可以是葡萄藤、西瓜藤、黄瓜藤、常春藤、爬山虎、啤酒花、茉莉或本领域已知的其他藤。在一些方面,藤是葡萄藤葡萄。在一些实例中,葡萄可以包括赤霞珠、卡诺娜、苏丹娜、霞多丽、白雷司令、白比诺、灰比诺、琼瑶浆、麝香葡萄、白苏维浓、黑比诺、莫尼耶比诺、品丽珠、梅乐、琳贝格、黑佳美、特罗灵格、味而多、比内洛、歌海娜、席拉、阿依仑、丹魄和本领域已知的其他葡萄品种。
[0076]
iv.方法
[0077]
本文还提供了通过使病原体与包含氮化硅的组合物接触来灭活病原体的方法。病原体可以是真菌或基于植物的病原体。组合物可以是包含氮化硅颗粒和水的浆料。
[0078]
在进一步的实施例中,该方法可以包括使氮化硅浆料与活的农业植物、树、谷物等的感染了植物性病原体的表面接触。在实施例中,受感染的叶可以用约1vol.%至约40vol.%的氮化硅在水中的浆料喷洒。叶可以暴露于氮化硅浆料至少1分钟、至少5分钟、至少10分钟、至少20分钟、至少30分钟、至少1小时、至少2小时、至少5小时或至少1天。
[0079]
在各种实例中,叶的受感染面积可减少至少50%、至少60%、至少70%、至少80%、至少90%、至少95%或至少99%。在实例中,在暴露1分钟后,叶的受感染面积可减少约95%。令人惊讶地发现,氮化硅颗粒可以被电吸引并且附接至病原体的孢子。
[0080]
实例
[0081]
实例1:
[0082]
葡萄藤中的霜霉病感染通常通过密集施用杀真菌剂(包括铜和硫(接触杀真菌剂))或通过合成的广谱内吸杀真菌剂如苯并咪唑或三唑来预防。然而,它们的使用会对环境和人类健康产生负面影响。杀真菌剂残留物是长期的土壤污染物,且在葡萄酒中可以发现不可忽略的量的这些化合物。虽然严格的法规试图将其有害后果降至最低,但形势要求开发替代的杀真菌策略。这些实例展示生物陶瓷氮化硅的前所未有的情况,它具有抗葡萄霜霉病菌的抗真菌特性,但对人类没有毒性或对环境没有不利影响。活孢子囊的拉曼光谱评估从机理上表明,生物陶瓷表面的氮化学是抑制宿主感染的原因。
[0083]
这些实例使用氮化硅(si3n4)在感染周期早期击倒葡萄霜霉病菌。这种陶瓷的选择是基于其在水环境中独特的表面化学性质。它具有抗菌、抗病毒和抗真菌的特性,同时仍然
对真核细胞友好和支持。由于这些原因,si3n4可以被认为是保护葡萄藤的环境友好的替代物。利用原位拉曼光谱研究影响葡萄霜霉病菌在葡萄藤叶上的致病性及其被si3n4灭活的分子机制。拉曼光谱是非侵入性的方法,其可以在没有标记的情况下应用于活的病原体,从而允许延时实验来揭示它们的代谢变化。该方法监测病原体的结构及其在与抗病原性药剂的化学相互作用期间的演变。
[0084]
实例2:
[0085]
为了显示氮化硅对农业真菌灭活的影响,用浓度为3x 104个孢子囊/毫升的葡萄霜霉病菌感染赤霞珠叶。将处理的葡萄霜霉病菌暴露于1.5vol.%的氮化硅浆料中1分钟。
[0086]
图12a显示赤霞珠叶上未处理的葡萄霜霉病菌。图12b显示赤霞珠叶上处理的葡萄霜霉病菌。可以看出,接种了用1.5vol.%si3n4粉末处理1分钟的葡萄霜霉病菌的叶在叶表面上具有较少的真菌。图13进一步证明了这一点,该图描绘接种了对照和处理的葡萄霜霉病菌的赤霞珠叶和卡诺娜叶的受感染叶面积的百分比。图13清楚地显示对照和处理的真菌之间受感染叶面积的统计学显著性差异。
[0087]
如图14b所示,氮化硅颗粒似乎被电吸引并且附接至病原体的孢子上。图14a显示未处理的葡萄霜霉病菌孢子囊的显微图像,而图14b显示在si3n4存在的情况下葡萄霜霉病菌孢子囊的显微图像。
[0088]
实例3:
[0089]
2018年在田地中收获的葡萄霜霉病菌(p.viticola)分离物如polesani等人“在易感性(葡萄)和抗性(葡萄)葡萄藤物种中对霜霉病感染的一般和物种特异性转录反应(general and species-specific transcriptional responses to downy mildew infection in a susceptible(vitis vinifera)and a resistant(v.riparia)grapevine species)”bmc基因组学11:117(2010)中所描述进行无病菌生长。为了评估si3n4可能的植物毒性,使用两种不同的葡萄品种赤霞珠和卡诺娜进行处理。赤霞珠叶取自3年生植物,而卡诺娜叶取自在受控条件下(16小时光照/8小时黑暗,温度范围18℃-28℃)在温室中生长的幼苗。
[0090]
使用具有约2μm的颗粒大小的si3n4粉末。它是通过研磨标称组成为90重量%的α-si3n4、6重量%的氧化钇(y2o3)和4重量%的氧化铝(al2o3)的烧结β-si3n4粉末来获得。这些成分在约1700℃下烧结》3小时,并在约1600℃下热等静压2小时。制备后,在悬浮于无菌蒸馏水中之前,将其在180℃下加热灭菌2小时。
[0091]
为了评价预防效果,从每个葡萄品种的无菌叶上切下三批,每批五个圆片。将一个批次通过完全浸没1.5vol.%的si3n4水悬浮液中1分钟进行处理,并在24小时后接种40-μl萌发的孢子囊悬浮液(3x104/ml)(预处理的样品)。第二批暴露于与1.5vol.%si3n4悬浮液混合的孢子囊。在这种情况下,si3n4颗粒在萌发期间保持与孢子囊直接接触(共处理的样品)。将第三批接种葡萄霜霉病菌并作为感染对照组。将所有的圆片在21℃-24℃的生长室中分别培养16小时和8小时的日/夜光周期,并监测6天,直到对照上出现孢子形成。
[0092]
为了评估潜在的疗效,从高度易感性苏丹娜品种的无菌葡萄藤叶上切下三批,每批六个圆片。使用40μl孢子囊悬浮液(3x104/ml)对所有三批接种葡萄霜霉病菌,并在21℃-24℃在生长室中培养16/8小时的日/夜光周期,以允许感染开始。24小时后,用前面讨论的相同程序除去液滴。出现感染后三天,将两批通过完全浸没1.5vol.%的si3n4水悬浮液中1
分钟进行处理。然后,将两批中的一批在蒸馏水中洗涤一分钟,以除去si3n4残余物。第三批未处理,作为对照组。
[0093]
显微镜观察
[0094]
悬浮在水中或1.5vol.%的si3n4悬浮液(3x104个孢子囊/ml)的孢子囊在落射荧光显微镜(激发滤光器bp 340-380nm;分色镜400nm;抑制过滤器lp》430nm)观察或用荧光素二乙酸盐(fda)染色,并使用荧光显微镜观察,以检查3小时时间过程内的孢子囊存活力。在细胞计数b
ü
rker室中进行观察,以计算与水处理的对照相比的活孢子囊的百分比。
[0095]
ph测量
[0096]
加入15vol.%的si3n4粉末后,用ph计测量无菌双蒸水的ph。当在室温下搅拌时,以10秒的时间间隔随时间变化进行测量至多800秒,直到最终ph稳定。为了检查ph趋势是否可再现,通过离心(13x10
3 rpm持续3分钟)分离测试的粉末样品,并在空气中于180℃干燥2小时。冷却至室温后,将粉末在相同的水浓度(即1.5vol.%)下重新悬浮,用于另外的ph测量。用相同的粉末重复该过程三个后续循环。
[0097]
原位拉曼光谱
[0098]
在含有和不含si3n4粉末的水溶液中悬浮的孢子囊样品上收集原位拉曼光谱。光谱是使用用具有50x光学透镜以微探针模式操作的专用仪器获得的。分光镜配有全息陷波滤波器,其同时获得高效率和高分辨率的光谱。用功率为15mw的785nm激光源进行激发。使用与气冷电荷耦合装置(ccd)检测器连接的单个单色仪监测拉曼散射光。一个光谱的采集时间通常为60秒。将在约10个不同的收集位置的不同孢子囊样品的光谱求平均。使用商业可获得软件(例如labspec 4.02)将拉曼光谱解卷积成高斯-洛伦兹叉积子带分量。根据已发表的文献进行光谱带指定。
[0099]
实例4:
[0100]
si3n4粉末在水悬浮液中的ph分析
[0101]
对于15vol.%的si3n4水悬浮液,ph随时间的变化示于图1a-1c中。这一ph实验被设想为模拟在已经喷洒了一定剂量的si3n4粉末之后在葡萄藤田周期性下雨的影响。三个连续的重复试验,包括悬浮、测量和干燥,分别在图1a、图1b和1c中给出。与运行顺序无关,这些图显示ph从最初的中性值(ph约7.5)突然(几秒钟内)增加到最大值(ph约8.3)。第一次运行和第二次运行的曲线非常相似,而第三次运行显示出随时间的急剧下降,然而所有试验的平稳期(ph约6.3-6.7)相似。这种现象在先前使用ph显微镜和比色氨测定的研究中被表征,与si3n4的表面处si-n键的断裂和洗脱的氮与氢反应形成氨(nh3)或铵(nh
4+
)相关联。在开放系统中,ph值在约5分钟内逐渐降至6.3至6.7。由于溶液中nh3的分数与ph值成反相关,时间相关的数据表明,增加的高挥发性nh3的分数离开水体系。这通过直接观察si3n4粉末分散后不久产生的气泡(图1d)以及典型的刺激性氨气味得到证实。在ph约8.3时,nh3的分数计算为7%至10%,而在酸性溶液中几乎为零。这些结果表明,假设si3n4粉末的一部分保持附接至叶片皱纹或截留在叶片气孔腔中,相同的si3n4粉末可以在连续的降雨事件期间提供延长的ph缓冲。
[0102]
孢子囊/si3n4颗粒相互作用的原位显微监测
[0103]
图2a-2c显示与si3n4粉末颗粒相互作用的水悬浮液中孢子囊的显微照片。在图2d-2f中给出了纯水中孢子囊的类似显微照片。si3n4似乎被静电吸引至孢子囊的外壁。接触几
乎是在孢子囊进入悬浮液后立即发生的。这导致壁在不到一分钟的时间内过早破裂(参见图2a-2c)。相反,纯水中的孢子囊仅在约110分钟后才释放出成熟的游动孢子。(参见图2d-2f)。有趣的是,也有一部分孢子囊在与si3n4颗粒接触时没有破裂。它们逐渐被si3n4颗粒覆盖,但直到几个小时后才释放游动孢子(参见图2g-2k中的延时显微照片)。总的来说,这些显微照片揭示在孢子囊附近形成了侵蚀性的环境。氨气泡的存在伴随着ph值的增加,如图1a-1c所示,与气态氨的产生有关,气态氨是容易穿透孢子囊细胞壁的挥发性分子。类似地,真菌细胞对氨是可渗透的,氨通过未解离分子的自由扩散进入细胞。
[0104]
图3a显示在接触时间《1分钟时孢子囊膜和si3n4颗粒之间相互作用的详细观察。图3b显示插图中正方形区域的放大图。这一放大显示膜的局部肿胀(即凸形弯曲),表明未成熟游动孢子即将破裂和释放。通过光学和荧光显微镜监测与si3n4颗粒接触的孢子囊的存活,以对陶瓷的抗真菌效果进行定量。图4a和4b分别显示在含有和不含1.5vol.%的si3n4颗粒情况下在水中悬浮3小时然后用二乙酸荧光素染色的浓度为105ml-1
的活孢子囊的荧光图像。孢子囊的总分数(通过光学显微镜)和通过荧光图像的直接计数检测到的活孢子囊的同时定量提供了图4c所示的分数图。当与纯水相比时,观察到含有si3n4颗粒的悬浮液具有明显的抗真菌效果。
[0105]
监测si3n4对葡萄霜霉病菌的预防和治疗效果
[0106]
来自两种葡萄藤物种赤霞珠和卡诺娜的叶圆片的实验结果示于图5a-5c和图6a-6c中。在立体显微镜下对处理的未感染的叶进行视觉检查,在实验的5天中没有揭示植物毒性的迹象。在这两种情况下,测试了两次实验运行中每一次运行的五个叶圆片样品(n=10),两次实验运行显示了相似的结果。同时检查对照样品。所有对照叶圆片显示了感染。接种后5天,在赤霞珠叶圆片上出现病原体孢子形成(图5a)。对于卡诺娜,对照叶圆片上的感染更严重(图6a),在大多数情况下导致感染斑坏死。这通常在高度易感性栽培品种或幼嫩叶组织中观察到。在整个5天的实验中没有观察到植物毒性。对于用si3n4预处理的或共处理的样品,观察到完全的感染保护作用(分别示于图5a和5c的图中)。对于卡诺娜物种,在预处理的样品中仅在5个叶圆片中的3个中观察到完全保护(图6b),而在共处理的圆片中没有孢子形成的迹象(图6c)。
[0107]
si3n4的潜在疗效通过在感染后3天处理高度易感性苏丹娜品种来进一步评估。由于残余的si3n4颗粒可以保持在气孔中,这一测试研究了在相对晚期的感染状态下是否可以停止发病。换句话说,该实验试图确定si3n4如何影响叶组织内的菌丝体或阻止新孢子囊的释放。观察到葡萄霜霉病菌已经在细胞间隙定殖并在叶肉细胞中产生吸器,但没有检测到孢子形成。这通常会在5-6天内发生。在这种情况下,似乎湿si3n4颗粒部分地穿透气孔。随后从颗粒中洗脱的氮很可能扩散到葡萄霜霉病菌生长菌丝的胞内空间。这种抑制机制类似于碳酸氢铵的抗真菌作用,其中碳酸氢根阴离子提供了建立足以杀死病原体的nh3浓度所必需的碱性。
[0108]
活孢子囊的原位拉曼光谱监测
[0109]
在室温下在纯水和含有1.5vol.%的si3n4粉末的悬浮液中浸没10分钟后收集的葡萄霜霉病菌的拉曼光谱分别示于图7a和7b中。光谱针对在424cm-1
观察到的散射强度进行归一化(未示出)。这种代表骨骼振动的信号是所有葡聚糖共有的非特异性标记。在含有和不含si3n4粉末的情况下,在暴露于水时它在强度上没有显示出明显的差异。然后将光谱解卷
积成伏格特(voigtian)子带并进行比较。这两个光谱之间的明显差异是由于卵菌结构的变化。每个光谱的解卷积拉曼谱带显示在图7a和7b中,且它们的相关指定和参考文献列于下面的表1中。
[0110]
表1
[0111]
[0112]
[0113][0114][0115]
暴露于纯水的孢子囊光谱
[0116]
根据更新的分类,卵菌最近被重新分类到不等鞭毛门中。主要结构特征包括:细胞壁中存在纤维素,昆布多糖代替糖原作为碳基能源,明显缺乏壳多糖。最近对卵菌寄生疫霉
(与葡萄霜霉病菌密切相关)中碳水化合物含量的分析揭示,细胞壁完全没有壳多糖,且由约85%的β-葡聚糖组成,其中约40%由纤维素表示。还存在低聚合度的1,3β-葡聚糖和1,3,6β-葡聚糖,以及较低分数的葡萄糖醛酸和甘露聚糖。不可获得葡萄霜霉病菌的此类详细信息,但以前的证据表明,这种病原体可能与霜霉病菌中其他熟知的生物略有不同。事实上,葡萄霜霉病菌可以表达至少两种不同的壳多糖合成酶,并且在植物体内生长期间,在孢子囊、孢子囊柄和菌丝细胞壁的表面检测到壳多糖。
[0117]
当卵孢子处于休眠状态时(就像在这种情况下),孢子囊的结构由分布在细胞质中的大脂质小球组成——充满了整个细胞腔。它们作为卵孢子萌发的贮藏物质。线粒体位于脂质小球之间的小间隙中。小球(或液泡)大小不同,且被容纳在相对较薄的空隙中。卵菌的整个外壁很复杂,且分为两层——卵孢子外壁和内壁(分别为oow和iow)。oow和iow被薄的略波状质膜隔开。iow主要由β-1,3-连接的葡聚糖(约80%;包括壳多糖,一种由n-乙酰氨基葡萄糖形成的均聚物)、纤维素(约10%)和蛋白质(分为细胞壁相关酶和结构蛋白)组成。葡聚糖是iow结构中的优势化学物种。它们含有以直线平行阵列取向的纤维素性质的原纤维,以及少量的甘露糖和葡糖胺。壳多糖具有重要的结构功能,因为它有助于壁的刚性和强度。oow主要由甘露聚糖和用β-1,6-葡聚糖将其连接到内壁的蛋白质构成,但它也含有脂质。卵原细胞壁是位于外侧上且通过周质空间与卵孢子分离的较厚的纤维壁,含有相对高含量的脂质和蛋白质。脂质赋予该结构疏水性,这是在休眠期间保持病原体安全所必需的。negrel等人最近寻找轴霜霉特异性代谢物,并鉴定了三种类型的非典型脂质——神经酰胺、花生四烯酸和二十碳五烯酸的衍生物。据报道,这些脂质在葡萄霜霉病菌发育的非常早期就存在。
[0118]
这些结构特征在图7a的拉曼光谱中观察到(也参见表1)。对于真菌结构,葡萄霜霉病菌的细胞壁主要由多糖组成,包括含支链聚合葡萄糖的β-葡聚糖、含非支链聚合n-乙酰基-d-氨基葡萄糖的壳多糖和与糖/甘露糖蛋白共价结合的聚合甘露糖。与多糖相比,蛋白质和脂质仅占总量的一小部分。因此,预计碳水化合物的振动模式在拉曼光谱中占主导地位。在482cm-1
处的带1中发现了来自主链葡萄糖环的累积信号。在643cm-1
、649cm-1
、710cm-1
和893cm-1
处发现了由壁中的壳多糖贡献的带,并分别标记为带14、15、19(=c-h弯曲)和33(c-h环拉伸)。这些带可能都与壳多糖有关,然而壳多糖实际上被认为是所研究的卵菌样品的碳水化合物中的次要组分。更可能指定给带19和33的是纤维素,而带14和15也可能被指定给线性聚合物纤维素中的β-d-葡萄糖。在583cm-1
处发现来自纤维素和支链淀粉的指纹信号(带9;c-c-o弯曲和c-o扭转振动)。正如从壁的结构中所预期的,在893cm-1
处发现了标记信号(赤道c-h弯曲振动),该信号用作β-葡聚糖的标记。在550cm-1
处没有拉曼信号(糖苷键的c-o-c弯曲),这是α-葡聚糖的指纹振动,表明这种多糖异构体不是真菌细胞壁的主要组分。由于这个原因,从图7a的光谱中排除了α-葡聚糖对942cm-1
拉曼信号(反对称环振动)的任何贡献。类似的推理适用于葡聚糖结构的环振动,其应该发生在922cm-1
处,且其糖苷信号发生在550cm-1
处。在暴露于水的葡萄霜霉病菌的光谱中没有检测到这些。这些观察排除了这种复合葡聚糖在当前卵菌结构中的任何优势存在。在图7a中490cm-1
和560cm-1
之间的窄光谱区中,带2和3(分别在500cm-1
和558cm-1
处)是来自多糖中的c-c主链拉伸、d(+)-甘露糖的信号,而带4、5和7(分别在510cm-1
、535cm-1
和558cm-1
处)分别被指定给纤维素、海藻糖(环变形)和β-d-葡萄糖(参见表s1)。二糖海藻糖是在603cm-1
处带11的主要贡献者,且它也对带
6和30有贡献(分别在544cm-1
和837cm-1
处)。海藻糖对带31和34(分别在846和906处)的贡献与其他碳水化合物结构相比权重可能更低(参见表s1)。更具体地,带31代表来自葡萄糖和葡聚糖的强累积信号,但它也含有来自甘油三酯的几个中等/强信号(参见表s1)。海藻糖是许多真菌物种代谢中的重要分子,因为它是一种能量来源和对抗环境胁迫的保护分子。例如,白色念珠菌促进非还原性海藻糖二糖的合成,并在热或氧化应激反应中积累。在葡萄藤中,已知葡萄霜霉病菌诱导不可逆的气孔开放,这反过来有利于宿主被游动孢子感染,并且这种失调与海藻糖积累相关联,外源施加的海藻糖拮抗气孔关闭。因此,孢子囊中存在的海藻糖水平的升高可能表示促进葡萄藤叶感染的信号。
[0119]
从磷酸二酯和嘌呤键上都发现了与核酸相关的信号。dna中c'5-o-p-o-c'3磷酸二酯键对称拉伸(在782cm-1
处的带25)是在暴露于纯水的葡萄霜霉病菌的低频光谱中检测到的最强信号(图7a)。与这个单独的信号不同,在827cm-1
处对应的反对称拉伸带29与来自甾醇、真菌膜中的典型分子的几个信号重叠(参见表1)。还观察到来自嘌呤的振动带,其与腺嘌呤(分别在535cm-1
、623cm-1
、731cm-1
和942cm-1
处的带5、12、21和36)、胞嘧啶(分别在544cm-1
、558cm-1
、594cm-1
、603cm-1
、710cm-1
和795cm-1
处的带6、7、10、11、19和26)、鸟嘌呤(在570cm-1
、643cm-1
、681cm-1
和846cm-1
处的带8、14、17和31)、胸腺嘧啶(分别在623cm-1
、746cm-1
和753cm-1
处的带12、22和23)和尿嘧啶(在807cm-1
处的带27)相关。846cm-1
处的带31,是在所研究的频率范围内检测到的第二最强信号(参见图7a和7b),主要是由鸟嘌呤环的c4-n9-c8+n1-c2-n3和n2-c2-n3面内变形造成的。
[0120]
寻找来自通常存在于细胞膜中的脂质的特殊信号,磷脂酰丝氨酸在所研究的光谱区域中的强发射预计在约734cm-1
处。在暴露于纯水的孢子囊的光谱中观察到731cm-1
处的带21(图7a)。然而,对该带的贡献也可能来自dna腺嘌呤和海藻糖中咪唑环的环呼吸。低频光谱中磷脂酰胆碱的主带预计在约719cm-1
和875cm-1
处。然而,在图7a中,在暴露于纯水的孢子囊中没有观察到这些信号。由于磷脂酰肌醇导致的在519cm-1
和868cm-1
处的主要低频带也在暴露于水的孢子囊的光谱中缺失。相反,分别在558cm-1
(带7)和681cm-1
(带17)检测到可能来自甾醇和神经酰胺的清晰信号。不幸的是,这些信号与来自dna嘌呤的信号强烈重叠。真菌细胞膜中最丰富的甾醇麦角甾醇提供了清晰的信号。这一分子的特征在于复杂的拉曼光谱,其包括在594cm-1
、710cm-1
、725cm-1
、827cm-1
和942cm-1
处(即,分别为带10、19、20、29和36)的标记低频信号。
[0121]
甾醇的特征在于复杂的拉曼光谱,其包括清晰的低频信号(参见表1)。然而,精确的筛选揭示这些低频信号中没有一个不含来自其他膜分子的重叠信号。甾醇是调节细胞膜的流动性、渗透性和完整性的重要组分。与真正的真菌不同,霜霉菌不能合成甾醇,然而它们在有性繁殖和无性繁殖中都需要甾醇。在疫霉属中,来自植物宿主的植物甾醇不经任何进一步修饰就被吸收和使用。
[0122]
关于其他脂质化合物,花生四烯酸是植物中卵菌释放的一种众所周知的激发子,且最近的发现表明葡萄霜霉病菌中的神经酰胺以及花生四烯酸和二十碳五烯酸的衍生物是在感染过程的非常早期阶段产生的。带32(在861cm-1
处)和35(在931cm-1
处)被指定给α-亚麻酸的c-o振动和花生四烯酸中的c-h弯曲。前一个带作为葡萄霜霉病菌特有的脂肪酸的指纹,而后者不幸地与来自葡萄糖和组氨酸的带重叠(如下所述)。脂肪酸通常在受到卵菌病原体感染时被释放到植物中。甘油磷脂卵磷脂在低频的最强信号(被指定给c-n拉伸)也
可以对带20(在715cm-1
)有贡献。卵磷脂的另外的带出现在764cm-1
和827cm-1
处(分别为带24和29)。这些分别归因于o-p-o对称和反对称拉伸。表1给出了图7a所示光谱的完整标记的尝试。
[0123]
暴露于1.5vol.%的si3n4水悬浮液的孢子囊的光谱
[0124]
由si3n4在水悬浮液中的存在引起的葡萄霜霉病菌孢子囊细胞结构的变化通过图7a和7b之间的光谱变化示出。总的趋势是,当与纯水中孢子囊的对应带相比时,暴露于si3n4悬浮液的孢子囊的所有拉曼带显示出相对较低的强度。主要变化如下:
[0125]
在si3n4悬浮液中孢子囊的光谱中,几个高或中等强度的带消失或仅在强度显著降低情况下出现。其中包括:来自腺嘌呤的带5和12(分别在535cm-1
和623cm-1
处);来自胞嘧啶的带10(在594cm-1
处);来自dna中c'5-o-p-o-c'3磷酸二酯对称拉伸的带25(在782cm-1
处);来自鸟嘌呤的带31(在846cm-1
处);以及来自纤维素(也可能由壳多糖提供)的带33(在893cm-1
处)。
[0126]
暴露于si3n4悬浮液的孢子囊谱中出现三个新的带。它们是:带11*(在613cm-1
处)、带15*(在654cm-1
处)和带32*(在872cm-1
处)。这些拉曼信号的来源是由于预先存在的分子的化学修饰或孢子囊响应于环境压力而产生的新的化学物种(如后面所述)。
[0127]
在si3n4存在情况下的另外的光谱变化是:来自多糖中的c-c主链拉伸的带2和来自d(+)-甘露糖的带3(分别在490cm-1
和500cm-1
处)。这些信号经历了强度趋势反转,前者变得比后者更强;并且来自纤维素的带9(在583cm-1
处),其也显示出相对高的强度。在带17(在681cm-1
处)中观察到类似的趋势,该带被指定给神经酰胺中的o=cn和cco弯曲,但也具有来自鸟嘌呤环的贡献。代表胸腺嘧啶的带23(在753cm-1
处)、来自组氨酸的带35(在931cm-1
处)和来自腺嘌呤的带36(在942cm-1
处)经历了强度的显著降低。
[0128]
暴露于纯水的孢子囊和si3n
4-粉末水悬浮液之间的显著光谱差异的原因是孢子囊和si3n4颗粒之间发生化学反应的结果。
[0129]
实例5:
[0130]
葡萄霜霉病菌和si3n4之间的化学相互作用
[0131]
通过直接观察,该研究证实了si3n4在水悬浮液中的强ph缓冲和气态氨的释放(参见图1a-1c)。观察到的ph缓冲是一种瞬时现象,因为si3n4表面被原硅酸逐渐覆盖,并且由于气态氮离开开放系统(参见图1d中观察到的气泡)。
[0132]
在图2g-2k中,si3n4颗粒似乎被静电吸引至孢子囊壁。霜霉病菌的细胞壁仅由有限量的壳多糖组成,并且主要由葡聚糖复合物和甘露糖蛋白组成。后一种成分通过含有五个甘露糖残基的糖磷酸基团连接到β-葡聚糖。磷酸化的甘露糖侧链赋予细胞壁负电荷。此外,孢子囊表面的官能团(即磷酸、羧基和氨基)在高碱性环境中会去质子化。它们与si3n4表面的正电荷位点相互作用,这些位点包括氮空位(带3+电荷)和n-n键(带+电荷)。一旦接触,孢子囊和si3n4颗粒之间的相互作用受到局部形成的高碱性ph和高挥发性nh3分子穿过细胞壁的被动扩散的强烈影响。仅在与si3n4颗粒接触1分钟后,这些离子相互作用导致膜破裂和观察到的未成熟游动孢子的败育(图2a-2c)。
[0133]
拉曼分析的解释
[0134]
氨对核酸预期的主要化学反应是水解。核酸首先分解成两个二核苷酸,一个含有腺嘌呤和尿嘧啶基团,另一个保留鸟嘌呤和胞嘧啶基团。虽然腺嘌呤-尿嘧啶二核苷酸比鸟
嘌呤-胞嘧啶相对更稳定,但两者在ph值》8时都分解成单核苷酸。在nh3、腺嘌呤和鸟嘌呤存在的情况下,磷酸二酯键被去质子化并强烈不稳定。在任何碱性ph下,由于氮环的弱碱性,胸腺嘧啶中n(3)上的氢也被除去。暴露于si3n4后,最显著的光谱变化是两个最强信号的消失,即带25和31(即,分别与dna中c'5-o-p-o-c'3磷酸二酯对称拉伸和鸟嘌呤环的c4-n9-c8+n1-c2-n3面内变形有关)。注意到与腺嘌呤(带5、12和36)和胞嘧啶(带6、10、11、19和26)相关的几个带的强度显著降低,如果不消失的话(参见图7a-7b)。这些观察与先前关于si3n4和病原体之间相互作用的研究一致。在由nh3扩散和氮自由基反应产生的氧化效应之前和之后的dna核碱基的示意图分别提供在图9a和9b中。最显著的特征是dna中磷酸二酯键的断裂,拉伸消失(带25),鸟嘌呤环(g
→
gh)由于与过氧亚硝酸根自由基相互作用而打开,以及其最强的环振动消失(带31)。带14的显著强度降低,也是由鸟嘌呤中的环呼吸模式造成的,这证实了图9b中所示的gh机制。随着其结构(a')的氧化,还观察到腺嘌呤单元(a)的修饰。腺苷氧化被认为是两个环变形带5和12消失的原因,因为形成了两个氧双键,如图9b所示。胞嘧啶核碱基(c)修饰成5-oh-u结构可以解释带6和10的消失。前者在原始胞嘧啶结构中代表n3=c4-n4和c-c=c弯曲模式,消失是因为这些键在氧化的5-oh-u单元中分别被n3-c=o和c-c-oh键取代,而后者来自原始胞嘧啶结构中的c2=o弯曲。它被氧化的5-oh-u结构中对称的c4=o键的出现所抵消。来自胸腺嘧啶和尿嘧啶(u)的带似乎更稳定,因为它们只有一个c=c双键。最后,注意一些与氧化的核碱基相关的带显示出较低的强度降低(即分别在558cm-1
、681cm-1
和731cm-1
处的带7、17和21)。然而,这些带都具有由脂质(分别为胆固醇、神经酰胺和磷脂酰丝氨酸)所贡献的特征。
[0135]
sonois等人通过实验和理论计算描述了几种氨基酸的拉曼行为。在组氨酸的情况下,ph增加的环境导致在约613cm-1
、656cm-1
和860cm-1
处出现新的拉曼带。这三个带对应于在暴露于si3n4的孢子囊中检测到的新带(参见图7b),且标记为带11*、15*和32*。组氨酸分子的侧链是含有6个π
–
电子的芳香咪唑环。根据环境ph值,可以存在组氨酸的不同互变异构体和离子形式。在ph《6时,质子化环中的两个n原子都是带正电荷的形式(图9c)。相反,在增加的ph值下,组氨酸在其咪唑环上失去一个质子,逐渐形成中性形式(图9d)。valery等人研究了自组装含组氨酸肽的构象变化及其在高ph下稳定的球状构象。振动光谱评估揭示了组氨酸-丝氨酸氢键和组氨酸-芳香相互作用。在ph=8.5时,组氨酸去质子化发生,且改变了拉曼光谱中1050至1150cm-1
频率范围内的c-n环拉伸带。
[0136]
为了加强对新形成的带11*、15*和32*的组氨酸解释,监测了咪唑环在1,050至1,150cm-1
频率范围内的c-n拉伸光谱区域。当比较纯水(ph=6.5)中的孢子囊与si3n4悬浮液(ph=8.3,分别参见图10a和10b)中的孢子囊上收集的拉曼光谱时,观察到不同的趋势。根据pogostin等人最近的一篇论文并考虑图9c和9d所示的环结构,组氨酸环去质子化的光谱指纹如下:
[0137]
在约1075cm-1
处出现了一个新的强带。这是最初在约1090cm-1
处观察到的强度明显减弱的带之外的带。这些带分别与(c2-n3)
+
和(c2-n3)键构型的拉伸振动有关。前一种构型涉及更强的键(即,由于n1-c2-n3电子共享)。它的振动能量更大,并且出现在更高的频率处。
[0138]
在原始(c4-n3)
+
和去质子化(c4-n3)键构型中观察到拉伸键的类似趋势,它们分别出现在约1125-1
(肩带)和约1118cm-1
(原始带)处。这种趋势可以使用上一段中给出的相
同推理来解释,然而向较低频率的频率偏移没有上一段中的情况明显。这种情况与去质子化环内的键强度平衡有关。c2-n3键比c4-n3键弱,因为其相邻的双键n1=c2比c4-n3旁边的双键c4=c5强(即,由于n比c的电负性高)。
[0139]
当环构型去质子化时,没有观察到与(c2-n1(h))
+
键(在约1100cm-1
处)相关的拉伸带的显著位移或强度变化,而只是轻微的增宽。
[0140]
还应注意,在约1600cm-1
处的c=c和c=n光谱区中组氨酸残基的咪唑环的另外的拉曼分析(未显示)提供了与图10a和10b所示结果一致的特征。基于上述拉曼分析,拉曼光谱特征与在高碱性环境中发生的去质子化有关。在高ph值下,这些反应对于含组氨酸的肽是常见的。
[0141]
组氨酸激酶蛋白存在于大多数原核和真核生物中。它们调节对各种环境因子的几种适应性转录反应。在卵菌中,组氨酸激酶的功能分析缺失,而组氨酸位点处的磷酸化是真菌对渗透应力的常见代谢反应。例如,响应于将渗透应力感知为环境条件的变化,保守的组氨酸残基被来自三磷酸腺苷的磷酸基团磷酸化这与图7b中检测到的腺苷拉曼带的减少一致。磷酰基向保守的天冬氨酸残基的连续转移导致介导信号转移至信号传导通路的调节。由于si3n4颗粒和孢子囊壁之间的高度局部碱性,相当一部分nh3渗透入胞吞空间(endocytotic space)并严重改变孢子囊的渗透平衡。因此,由于si3n4暴露引起的拉曼数据的改变表明孢子囊对渗透应力作出反应;且组氨酸激酶感受到渗透应力的增加,类似于对酿酒酵母的假设。应注意的是,在模型卵菌致病疫霉中,已经发现蛋白激酶与萌发过程中的孢子囊分裂有关。
[0142]
在本研究中,壳多糖主要信号(即在893cm-1
处的带33)的消失和与细胞壁结构中的纤维素(和/或壳多糖)和其他线性碳水化合物相关的所有其他信号(即500cm-1
、643cm-1
、649cm-1
和710cm-1
处的谱带3、14、15和19)强度的显著降低证实了这一假设。纤维素中的线性聚合物链通过β-糖苷键连接在一起。这些键不受si3n4诱导的碱性ph水平的影响,也不受与nh3的任何直接相互作用的影响。另一方面,水解酶可以分解壳多糖的糖苷键,且从而改变植物病原体的细胞壁。鉴于拉曼实验是如何进行的,酶促反应只能是孢子囊本身固有的。众所周知,藻菌能够通过酶来控制其细胞壁的可塑性。真菌壁被“软化”,且必须扩张才能萌发和随后生长。它们也在假菌丝和孢子壁的形成过程中被改造,且形成酚交联。由β-葡聚糖和壳多糖构成的细胞壁内部基质提供了拉伸强度和硬度。然而,壁的组成可以通过经由促分裂原活化蛋白激酶通路响应于环境变化而重塑。细胞壁的弹性受到快速结构重组的调节,这使得病原体能够在渗透休克中存活。如图3a和3b所示,在si3n4颗粒附近观察到芽的出现,这种现象在许多情况下导致过早的孢子形成(参见图2a-2c)。
[0143]
turchini等人测量到,与在低渗透压培养基中生长的真菌细胞相比,在高渗透压培养基中生长的真菌细胞的壳多糖含量减少了50%,这与先前的数据一致,该数据显示,在低渗透压培养基中生长的细胞比在高渗透压培养基中生长的细胞的壳多糖合酶活性更高。这些研究人员解释,在高渗透压介质中观察到的真菌壁的弱化是一种拯救机制,能够使膜拉伸并提高保持细胞完整性的可能性。基于这些研究,暴露于si3n4的孢子囊的主要壳多糖带33的消失是真菌细胞试图抵抗渗透应力而活化的酶指纹。最后,还应注意的是,在与si3n4接触仅1分钟后就令人惊讶地观察到的过早萌发(图2a-2c)可能是孢子囊不能产生维持萌发至成熟所需的壳多糖壁的从头合成的结果。
[0144]
si3n4与其他生态友好方法的比较的效果
[0145]
葡萄霜霉病菌通过气孔进入寄主叶组织,并保持在气孔下的空气空间中,在那里它缓慢发育12-15小时,直到形成第一个吸器。这一初始阶段被认为是整个感染过程中最关键的阶段。在这个时期开始之前,可以预防性地将si3n4颗粒喷洒在葡萄藤叶上。一旦进入气孔,它们就长时间滞留在里面(例如,可能一个季节)。在下雨事件期间,水将重复活化铵部分的洗脱和ph值的快速上升,从而为孢子囊创造不利的环境(图11a)。相比之下,目前可获得的基于氢氧化铜和硫酸铜的杀真菌剂只能对葡萄藤提供预防性保护。它们不是全身性的,因此不能根除预先存在的感染。此外,它们很容易被雨水冲走。因此,像si3n4这样的固态杀真菌剂的一个优点是它在下雨事件期间可以重复活化(参见图1a-1c),和其即使在真菌穿透叶组织后对抗病原体的功效。在病原体/si3n4界面处有活性的抗真菌机制示意性地总结在图11b中。
[0146]
已经实施了三种策略来取代对生态不友好的农用化学品的使用:(i)开发新的环境友好的抗真菌产品;(ii)使用微生物诱导对植物病原体的系统抗性;和(iii)通过控制群体遗传学来操纵宿主-病原体相互作用。
[0147]
可以替代农用化学品的有效的生态友好分子是多糖。例如,低聚壳寡糖是植物防御的有效促进剂,具有诱导抑制寄生虫生长的分子积累的能力(即植物抗毒素和有效的抗氧化剂,如反式和顺式白藜芦醇及其衍生物)。壳聚糖还引发葡萄叶中酶分子(例如壳多糖酶和α-1,3-葡聚糖酶)的产生,该酶分子能够溶解病原体,从而显著降低霜霉病感染的可能性。低分子量壳聚糖还具有穿透真菌分生孢子的能力,导致膜解体和细胞内容物损失。它与真菌质膜的外部阴离子组分相互作用,导致膜破裂。另一种能够控制葡萄霜霉病菌感染的多糖是水溶性β-1,3-葡聚糖昆布多糖,其可以从褐藻掌状海带中获得。其抗病原性作用的来源在于有效地引发葡萄藤细胞的防御反应。然而,尽管壳聚糖和昆布多糖具有公认的抗真菌效率,但有报道显示这些多糖对葡萄藤的氨基酸组成有不良影响,会改变葡萄藤中的氮浓度,这是最终葡萄品质的关键参数。
[0148]
香草醛和大蒜提取物也被归类为生态友好的抗真菌物质。前者的化学结构中含有醛基,而后者包括大蒜素的强大抗真菌活性,阻断真菌酵母中的脂质、蛋白质和核酸合成。然而,这些化合物的主要缺点是它们容易与水反应。例如,大蒜素会迅速形成二烯丙基二硫化物,这是一种抗菌活性不太明显的化合物。
[0149]
测试的其他天然衍生化合物包括水解蛋白、植物提取物和无机盐。在这种情况下,最近提出了一种有希望的策略,该策略基于使用所选择的适体肽,特异性抑制葡萄霜霉病菌纤维素合酶2,并因此预防感染且对非靶生物没有副作用。
[0150]
在无机盐中,实例主要包括碳酸氢盐、磷酸盐、硅酸盐、氯化物和亚磷酸盐。据报道,它们的活性主要针对包括葡萄藤在内的不同作物的白粉病,而只有碳酸氢钠对葡萄藤霜霉病表现出有限的效力。基于氮化硅的植物检疫产品的开发属于最后一类。在本上下文中,硅酸盐值得特别提及:几种可溶性硅酸盐对不同的真菌感染具有直接和间接的活性,通过刺激植物的天然防御机制和增强植物细胞壁而起作用。从si3n4产生sio2和si(oh)4(如反应(1)-(3)中所述),因此可以通过诱导植物抗性来补充氨对葡萄霜霉病菌孢子囊和游动孢子的直接作用,这可以至少部分地对我们实验中观察到的几乎完全抑制感染过程负责。此外,与容易被雨水冲走的可溶性盐相比,si3n4可以通过铵部分从不溶性粉末中的不同洗脱
循环以及活性氮物质的产生来提供更持久的保护,这与先前对人类病原体的研究一致。事实上,在处理后,si3n4颗粒可以滞留在气孔内(图11a),并且在降雨事件期间,水可以重复产生不利的化学环境(图11b),可能削弱继发感染循环中的孢子囊散发,从而减少进一步化学处理的需要。一些微生物可以提供对植物病原体的系统抗性,从而降低疾病的严重程度。抗性是宿主植物生理学改变的结果,从而产生代谢反应,导致保护性酶分子的合成。哈茨木霉菌株t39,一种商业生物防治剂,可能是用于针对霜霉病的系统抗性的合适微生物。已经研究了三种不同的生物激发子(即哈茨木霉、褶皱链霉菌和荧光假单胞菌)在诱导系统抗性中的效率。哈茨木霉对霜霉病的抗性最强,其次是褶皱链霉菌。然而,处理后观察到叶绿素和胡萝卜素都增加了;并且在三种生物疗法之间可以发现蛋白质表达水平的相当大的差异。一般来说,使用微生物诱导系统抗性的缺点是仅对单一形式的抗性有效。它们的多样性最终会导致病原体突变。
[0151]
从更一般的观点来看,卵菌葡萄霜霉病菌致病机理背后的分子机制在很大程度上是未知的。为了引发感染,卵菌分泌细胞质和质外体效应蛋白,抑制免疫并增强植物易感性。已经发现在测序的卵菌基因组中的效应子快速进化并获得新的功能,从而抵消植物抗性基因并抑制植物rna沉默机制。由于基因组测序的复杂性,抵消效应蛋白的作用传统上是困难的。它强制要求多组学方法。利用比较基因组学,brilli等人最近报道了在葡萄霜霉病菌基因组中发现了一个缺失的代谢特征,这可以解释其生物营养模式。他们发现了一种在有抵抗力的葡萄藤中引发免疫力的蛋白质效应器。german等人开发了一种并行分析rna末端的新方法,该方法结合了小rna和全基因组降解测序,揭示了感染期间靶向基因的小rna的复杂网络。因此,提出一种新的双向rna沉默策略。尽管病理测量技术最近已经达到了很高的先进水平,但它们也揭示了病原体-宿主相互作用的复杂性。所研究的系统只能对有限数量的基因进行编码和解释(预计会有许多基因对基因的相互作用),即使只识别出“抗性”和“易感性”类型的反应。
[0152]
si3n4表现出耐人寻味的多机制抗病原性行为,有可能解决环境友好的农业化学替代方法的若干缺点。si3n4的广谱抗病原性效果是由于它的氮化学。水充当释放氮的触发器,导致引起病原体溶菌的级联反应。在si3n4表面处产生的氮物种改变病原体的蛋白质,诱导对dna的亚硝化损伤,并刺激改变病原体膜结构的代谢酶。这些发现与之前对人类病原体的研究一致。由于nh3穿过细胞壁的扩散而在病原体的细胞质中发生的多机制溶菌反应降低了病原体突变的可能性。此外,因为si3n4用作可植入的生物材料,所以它对真核细胞没有毒性。它只包含地球史前固有的环境友好元素。一些植物物种受益于硅施肥,特别是在缓解生物和非生物应力时。由si3n4分解产生的铵是参与有机氮合成的主要无机物质。土壤中的铵和硝酸根离子通过根特异性转运蛋白直接被吸收并有效利用。在葡萄藤的情况下,nh
4+
在发酵前占总氮的80%,而在成熟后下降到5%-10%,且在葡萄汁发酵后甚至更低。从si3n4中洗脱的氮可能有助于改善浆果质量和发酵条件。作为一个限制因素,应该注意平衡来自肥料和si3n4的氮的量,因为过量的氮可能会改变酚类化合物的产生以及葡萄和葡萄酒的味道或质量。一些植物物种受益于硅施肥,特别是在缓解生物和非生物应力时。
[0153]
实例6:
[0154]
这些实例为si3n4对抗葡萄霜霉病菌葡萄藤感染的效果提供了新的见解。作为一种无机环境友好剂,它有可能取代重金属农药和新的生态友好抗病原性分子。si3n4的使用也
符合当前旨在减少葡萄栽培中重金属使用的管理趋势。si3n4的独特化学性质在孢子囊中诱导渗透应力,并引发其未成熟游动孢子的败育,甚至在低至1.5vol.%的浓度下也是如此,该浓度在农业应用中用于其他无机盐如碳酸氢盐的摩尔浓度范围内。拉曼实验提供了关于化学机制的重要信息,包括磷酸脱氧核糖主链的断裂和鸟嘌呤环的破坏。对来自不同葡萄藤物种的叶片的实验表明,si3n4在非常早期阶段有效地严重减少或阻断感染过程,影响孢子囊萌发和游动孢子存活力,如显微观察所揭示的。si3n4的使用对高氮需求的葡萄藤最为有利,其中铜基制剂不仅对环境有害,而且对葡萄酒质量也不利。由于si3n4仅含有环境友好的元素,这种陶瓷也将是接触杀真菌剂的合适替代物,接触杀真菌剂包括有毒的铜和硫元素。si3n4可被视为一种有前途的生物杀虫剂,与常规合成产品相比具有多种益处,且与其他无机盐相比具有技术优势,并且可能是综合疾病管理中的有用组分。
[0155]
从前面的描述中应该理解,尽管已经示出和描述了特定的实施例,但是对于本领域技术人员来说显而易见的是,在不脱离本发明的精神和范围的情况下,可以对其进行各种修改此类变化和修改在所附权利要求中定义的本发明的范围和教导内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1