转基因哺乳动物及其使用方法与流程
发明领域本发明涉及免疫球蛋白分子的产生,包括用于产生能够产生抗原特异性抗体分泌细胞以用于产生猫科动物(feline)单克隆抗体的转基因哺乳动物的方法。
背景技术:
0、发明背景
1、在以下讨论中,出于背景和介绍的目的描述了某些文章和方法。本文包含的任何内容不被解释为对现有技术的“承认”。本技术人明确地保留根据情况证明根据可适用的法定条文由本文引用的文章和方法不构成现有技术的权利。
2、抗体已经成为重要的生物药物,因为它们(i)表现出可以靶向不同分子形式的抗原的精准结合特性,(ii)是具有使其在治疗的人类和动物中良好耐受的期望药代动力学的生理分子,并且(iii)与天然抵御感染原(infectious agent)的强大免疫特性关联。此外,存在已建立的用于从实验室动物快速分离抗体的技术,所述技术可以容易地针对几乎任何不天然存在于身体中的外来物质产生特异性抗体反应。
3、在抗体的最基本形式中,抗体包括各自与相同的轻(l)链配对的两条相同的重(h)链。h链和l链二者的n-末端都包括可变结构域(分别地,vh和vl),所述可变结构域共同为配对的h-l链提供独特的抗原结合特异性。
4、编码抗体vh和vl结构域的外显子在种系dna(germ-line dna)中不存在。而是,每一个vh外显子由存在于免疫球蛋白h链基因座中的随机选择的vh、dh和jh基因区段的重组产生;同样,单独vl外显子由轻链基因座中随机选择的vl和jl基因区段的染色体重排产生。
5、在哺乳动物中,基因组通常包含两个可以表达h链的等位基因、两个可以表达kappa(κ)l链的等位基因和两个可以表达lambda(λ)l链的等位基因(每个亲本一个等位基因)。在免疫球蛋白h链基因座处存在多个vh、dh和jh基因区段,以及在免疫球蛋白κ(igk)和免疫球蛋白λ(igl)l链基因座二者处存在多个vl和jl基因区段(collins和watson(2018)immunoglobulin light chain gene rearrangements,receptor editing and thedevelopment of a self-tolerant antibody repertoire.front.immunol.9:2249.(doi:10.3389/fimmu.2018.02249))。
6、在重链基因座中,也存在用于表达不同抗体类别(同种型)的外显子。例如,在猫科动物中,编码的同种型是igm、igd、igg1a、igg2、ige和iga2。
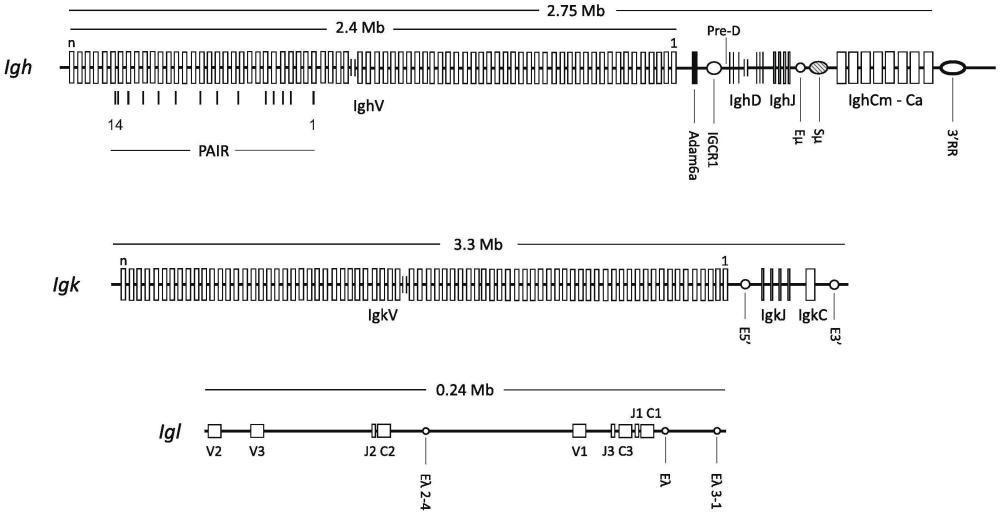
7、在b细胞发育期间,基因重排首先发生在包含h链可变基因区段的两条同源染色体中的一条上。在前b细胞中,产生的vh外显子然后在rna水平上被剪接到cμ外显子,用于igm h链(μh链)的表达。前b细胞合成的μh链大部分保留在内质网(er)中,并且最终由于部分未折叠的μh链ch1结构域与驻留的er伴侣bip之间的非共价相互作用而降解(haas和wabl,nature,306:387-9,1983;bole等人,j cell biol.102:1558,1986)。然而,一小部分μh链与替代轻链复合物缔合,该复合物包括不变的λ5和vpreb蛋白。这种缔合取代了bip,并允许μh链/λ5/vpreb复合物与igα/β信号传导分子异二聚体一起作为preb细胞受体(prebcr)离开er,并通过分泌途径运输至质膜。
8、随后,vl-jl重排一次发生在一个l链等位基因上,直到产生功能性l链,之后l链多肽可以与igm h链同源二聚体缔合,形成完全功能抗原特异性b细胞受体(bcr),其在未成熟b细胞的表面表达。
9、未成熟的b细胞迁移到次级淋巴器官,在那里它们分化为成熟的b细胞,成熟的b细胞可以对同源抗原响应,并分化成分泌抗体的浆细胞和记忆b细胞。在t细胞的帮助下,b细胞可以进行同种型转换,将抗体同种型从igm转变为igg、iga或ige,还可以进行体细胞高突变,改变vh和vl结构域的氨基酸序列。虽然这些突变被随机引入vh和vl外显子,但对免疫抗原具有较高亲和力的b细胞能够吸收更多的抗原,加工抗原并将其呈递给t滤泡性辅助细胞,并且因此与对免疫抗原具有低亲和力或没有亲和力的b细胞相比优先被激活。结果,体细胞突变在互补决定区(cdr)1、2和3中变得富集,因为这些是vh和vl结构域与抗原相互作用的区域。
10、编码各种小鼠免疫球蛋白的基因已被广泛表征。例如,blankenstein和krawinkel在eur.j.immunol.,17:1351-1357(1987)中描述了小鼠可变重链区。虽然关于猫科动物免疫球蛋白重链基因座的信息较少,但淋巴恶性肿瘤和对病毒响应的研究已经包括一些vh结构域序列[例如,rout等人,vet.clin.pat.45:48suppl.1(2019)和lu等人scientificreports 7:12713(2017)]。lu等人还表征了猫科动物igg1a、igg2和iga序列。猫科动物κ和λlc基因座已被广泛表征,并在国际免疫遗传学(imgt)信息系统中得到完全注释。
11、转基因动物诸如具有各种免疫球蛋白基因座的小鼠的产生已经允许在各种研究和开发应用中,例如在药物发现和对各种生物系统的基础研究中使用这样的转基因动物。例如,携带人类免疫球蛋白基因的转基因小鼠的产生描述于国际申请第wo 90/10077号和第wo 90/04036号中。wo 90/04036描述了具有整合的人类免疫球蛋白“微(mini)”基因座的转基因小鼠。wo 90/10077描述了用于在产生转基因动物中使用的包含免疫球蛋白显性控制区的载体。
12、已经开发了用于用例如人类免疫球蛋白序列修饰小鼠内源性免疫球蛋白可变区基因座以产生用于药物发现目的的部分或完全人类抗体的许多方法。这样的小鼠的实例包括以下中描述的那些:例如,美国专利第7,145,056号;第7,064,244号;第7,041,871号;第6,673,986号;第6,596,541号;第6,570,061号;第6,162,963号;第6,130,364号;第6,091,001号;第6,023,010号;第5,593,598号;第5,877,397号;第5,874,299号;第5,814,318号;第5,789,650号;第5,661,016号;第5,612,205号;和第5,591,669号。
13、使用发挥作为药物的功能的抗体不限于人类疾病的预防或疗法。像猫这样的家畜也会遭受与人类相似的痛苦,例如癌症、特应性皮炎和慢性疼痛。靶向神经生长因子的单克隆抗体(bedinvetmab)已经在兽医中用于治疗猫的骨关节炎,但尚未批准用于治疗癌症或特应性皮炎。然而,在临床使用之前,在小鼠体内制备的单克隆抗体必须被猫源化(felinized),即它们的氨基酸序列必须从小鼠改变为猫科动物,以防止受者猫产生不良免疫应答。
技术实现思路
0、发明概述
1、提供本概述是为了以简化的形式介绍概念选择,所述概念在下文的详述中进一步描述。本概述既不意图确定所要求保护的主题的关键或基本特征,也不意图用于限制所要求保护的主题的范围。从以下撰写的详述,包括附图中阐明的和所附的权利要求书中限定的那些方面,所要求保护的主题的其他特征、细节、效用和优点将是明显的。
2、本文描述了产生具有猫科动物免疫球蛋白可变区的小鼠抗体的方法。在一方面,提供了一种具有猫科动物可变区的抗体,该抗体可以在转基因哺乳动物中或体外细胞培养中产生。
3、在一方面,提供了具有包含异源性部分猫科动物免疫球蛋白基因座的基因组的非猫科动物哺乳动物细胞或非猫科动物哺乳动物。在一方面,异源性基因座包含猫科动物免疫球蛋白可变区基因的编码序列和基于非猫科动物哺乳动物宿主的内源性免疫球蛋白可变区基因座的非编码序列。在一方面,非猫科动物哺乳动物细胞或哺乳动物能够表达嵌合b细胞受体(bcr)或抗体,其包括非猫科动物哺乳动物宿主细胞或哺乳动物内源性的猫科动物重链(h)和轻链(l)可变区和恒定区。在一方面,转基因哺乳动物宿主细胞或哺乳动物具有部分或全部内源性免疫球蛋白可变区基因座被去除的基因组。
4、为了在非猫科动物哺乳动物宿主中产生嵌合猫科动物单克隆抗体,宿主基因组应该具有至少一个表达嵌合猫科动物免疫球蛋白h链或l链的基因座。在一方面,宿主基因组包括一个重链基因座和两个轻链基因座,分别表达嵌合的猫科动物免疫球蛋白h链和l链。
5、在一些方面,部分猫科动物免疫球蛋白基因座包括猫科动物vh编码序列和存在于非猫科动物哺乳动物宿主的内源性vh基因座的非编码序列。在一些方面,部分猫科动物免疫球蛋白基因座包括猫科动物vh编码序列和存在于非猫科动物哺乳动物宿主的内源性vh基因座的非编码调控或支架序列。在一方面,部分猫科动物免疫球蛋白基因座包含猫科动物dh和jh基因区段编码序列和存在于非猫科动物哺乳动物宿主细胞基因组的内源性dh和jh基因区段中的非编码序列。在一方面,部分猫科动物免疫球蛋白基因座包含猫科动物dh和jh基因区段编码序列和存在于非猫科动物哺乳动物宿主细胞基因组的内源性dh和jh基因区段中的非编码调控或支架序列。
6、在其他方面,部分猫科动物免疫球蛋白基因座包含猫科动物vl编码序列和存在于非猫科动物哺乳动物宿主的内源性vl基因座中的非编码序列。在其他方面,部分猫科动物免疫球蛋白基因座包含猫科动物vl编码序列和存在于非猫科动物哺乳动物宿主的内源性vl基因座中的非编码调控或支架序列。在一方面,异源性部分猫科动物免疫球蛋白基因座包含猫科动物vl编码序列和猫科动物jl基因区段编码序列以及存在于非猫科动物哺乳动物宿主细胞基因组的内源性jl基因区段中的非编码序列。在一方面,异源性部分猫科动物免疫球蛋白基因座包含猫科动物vl编码序列和猫科动物jl基因区段编码序列和存在于非猫科动物哺乳动物宿主细胞基因组的内源性jl基因区段的非编码调控或支架序列。
7、在一方面,非猫科动物哺乳动物是啮齿动物,例如,小鼠或大鼠。
8、在一方面,提供了一种用于产生包含部分猫科动物免疫球蛋白基因座的非猫科动物哺乳动物细胞的方法。在一方面,该方法包括:a)将两个或更多个重组酶靶向位点引入非猫科动物哺乳动物宿主细胞的基因组中,并整合位于包含内源性免疫球蛋白vh、dh和jh基因或内源性vl和jl基因的基因组区域的上游的至少一个位点和下游的至少一个位点;和b)通过重组酶介导的盒式交换(rmce)将异源性部分猫科动物免疫球蛋白可变基因座引入非猫科动物哺乳动物宿主细胞,该基因座包含猫科动物vh、dh和jh基因或猫科动物vl和jl基因编码序列和基于存在于非猫科动物哺乳动物宿主内源性免疫球蛋白可变区基因座中的非编码序列的非编码序列。
9、在另一方面,该方法包括在通过rmce将异源性部分猫科动物免疫球蛋白可变基因座引入非猫科动物哺乳动物宿主细胞之前,缺失宿主动物基因组中侧翼为两个异源性重组酶靶向位点的内源性免疫球蛋白可变区。
10、在一方面,异源性部分猫科动物免疫球蛋白基因座包含猫科动物vh基因区段编码序列、猫科动物dh和jh基因区段编码序列以及位于猫科动物dh基因区段(pre-d序列,图1)上游的非编码调控或支架序列,所述非编码调控或支架序列是基于非猫科动物哺乳动物宿主基因组的内源性dh基因区段上游存在的序列。在一方面,上游支架序列包含非免疫球蛋白基因,诸如adam6a(图1),其与雄性生育力有关[nishimura等人developmental biol.233(1):204-213(2011)]。在一方面,使用先前已经被引入到同一染色体上内源性免疫球蛋白vh基因座的上游和内源性jh基因座的下游的重组酶靶向位点将部分猫科动物免疫球蛋白基因座引入到宿主细胞中。
11、在一方面,支架序列包括来自另一物种的天然存在的核酸序列。在一方面,所述支架序列可以基于来自另一物种的天然存在的核酸序列来设计,例如,所述支架序列可以包括来自另一物种的天然存在的核酸序列,该序列已经被修饰过,例如,通过一个或更多个核酸置换、插入、缺失或其他修饰。在一方面,支架序列可以包括人工序列。在一方面,支架序列包括存在于猫科动物基因组的免疫球蛋白基因座中的序列与其他序列组合,所述其他序列例如来自其他物种的支架序列。
12、在另一方面,异源性部分猫科动物免疫球蛋白基因座包含猫科动物免疫球蛋白vl基因区段编码序列、猫科动物jl基因区段编码序列和基于存在于非猫科动物哺乳动物宿主细胞基因组的内源性l链基因座中的非编码序列的非编码序列。在一方面,非编码序列包括调控或支架序列。在一方面,使用先前已经被引入到同一染色体上内源性免疫球蛋白vl基因座的上游和内源性jl基因座的下游的重组酶靶向位点将异源性部分猫科动物免疫球蛋白基因座引入到宿主细胞中。
13、在一方面,异源性部分猫科动物免疫球蛋白基因座作为单个核酸合成,并且作为单个核酸区域引入到非猫科动物哺乳动物宿主细胞中。异源性部分猫科动物免疫球蛋白基因座也可以在两个或更多个连续的区段中合成,并且作为离散的区段引入哺乳动物宿主细胞。异源性部分猫科动物免疫球蛋白基因座也可以使用重组方法产生,并且在引入非猫科动物哺乳动物宿主细胞之前进行分离。在一方面,部分猫科动物免疫球蛋白重链可变区基因座可以通过计算机模拟如下产生:例如,小鼠重链免疫球蛋白基因座的基因组序列以及猫科动物vh、dh和jh编码序列从美国国家生物技术信息中心或国际免疫遗传学(imgt)信息系统获得。小鼠vh、dh和jh编码序列被猫科动物vh、dh和jh编码序列计算机模拟地替换,例如使用商业上可得的软件。有利地,vh、d和jh编码序列可以被替换,同时保持中间的小鼠非编码序列完整。相似地,部分猫科动物免疫球蛋白轻链可变区基因座可以通过计算机模拟如下产生:例如,小鼠轻链免疫球蛋白基因座的基因组序列以及猫科动物vl和jl编码序列从美国国家生物技术信息中心或国际免疫遗传学(imgt)信息系统获得。小鼠vl和jl编码序列被猫科动物vl和jl编码序列计算机模拟地替换,例如使用商业上可获得的软件。再一次,vl和jl编码序列可以被替换,同时保持中间的小鼠非编码序列完整。基于计算机模拟序列,用于合成包含部分猫科动物免疫球蛋白基因座的dna序列的方法是已知的。
14、在另一方面,提供了一种用于产生包含异源性部分猫科动物免疫球蛋白基因座的非猫科动物哺乳动物细胞的方法。在一方面,该方法包括:a)将两个或更多个不能彼此重组的序列特异性重组位点引入非猫科动物哺乳动物宿主细胞的基因组,其中至少一个重组位点被引入内源性免疫球蛋白可变区基因座的上游,并且至少一个重组位点被引入相同的内源性免疫球蛋白可变区基因座的下游;b)提供包含异源性部分猫科动物免疫球蛋白基因座的载体,该异源性部分猫科动物免疫球蛋白基因座具有i)猫科动物免疫球蛋白可变区基因编码序列和ii)基于宿主细胞基因组的内源性免疫球蛋白可变区基因座的非编码调控或支架序列,其中部分猫科动物免疫球蛋白基因座的侧翼是相同的两个序列特异性重组位点,所述相同的两个序列特异性重组位点位于宿主细胞的内源性免疫球蛋白可变区基因座的侧翼;c)将步骤b)的载体和能够识别两个重组酶位点的位点特异性重组酶引入宿主细胞;d)允许在细胞基因组和异源性部分猫科动物免疫球蛋白基因座之间发生重组事件,导致内源性免疫球蛋白可变区基因座被异源性部分猫科动物免疫球蛋白可变区基因座替换。在一方面,部分猫科动物免疫球蛋白基因座包含猫科动物vh免疫球蛋白基因区段编码序列,以及i)猫科动物dh和jh基因区段编码序列,ii)在内源性地存在于非猫科动物哺乳动物宿主的基因组中的个体vh、dh和jh基因区段侧翼的非编码调控序列或支架序列,以及iii)基于非猫科动物哺乳动物宿主细胞的内源性基因组的pre-d序列。在一方面,重组酶靶向位点被引入内源性免疫球蛋白vh基因座的上游和内源性jh基因座的下游。
15、在一方面,提供了转基因啮齿动物,所述转基因啮齿动物具有其中啮齿动物内源性免疫球蛋白可变基因座已被缺失,并被异源性部分猫科动物免疫球蛋白基因座替换的基因组,该异源性部分猫科动物免疫球蛋白基因座包含猫科动物免疫球蛋白可变基因编码序列和基于啮齿动物内源性免疫球蛋白可变基因座的非编码调控或支架序列。在一方面,转基因啮齿动物的异源性部分猫科动物免疫球蛋白基因座是功能性的,并表达包含猫科动物可变结构域和啮齿动物恒定结构域的免疫球蛋白链。在一方面,异源性部分猫科动物免疫球蛋白基因座包含猫科动物vh、dh和jh编码序列。在一方面,异源性部分猫科动物免疫球蛋白基因座包含猫科动物vl和jl编码序列。在一方面,异源性部分猫科动物免疫球蛋白基因座包含猫科动物kappa(κ)vl和jl编码序列。在一方面,异源性部分猫科动物免疫球蛋白基因座包含猫科动物lambda(λ)vl和jl编码序列。在一方面,提供了来自转基因啮齿动物的b淋巴细胞谱系的细胞。在一方面,提供了包含猫科动物可变结构域和从b淋巴细胞谱系的细胞获得的啮齿动物恒定结构域序列的部分或完整免疫球蛋白分子。在一方面,提供了衍生自b淋巴细胞谱系细胞的杂交瘤细胞。在一方面,提供了包含猫科动物可变结构域和衍生自杂交瘤细胞的啮齿动物恒定结构域的部分或完整免疫球蛋白分子。在一方面,提供了衍生自b淋巴细胞谱系细胞的永生化细胞。在一方面,提供了包含猫科动物可变结构域和衍生自永生化细胞的啮齿动物恒定结构域的部分或完整免疫球蛋白分子。在一方面,提供了转基因啮齿动物,其中异源性部分猫科动物免疫球蛋白基因座包含猫科动物vl和jl编码序列。在一方面,提供了转基因啮齿动物,其中异源性部分猫科动物免疫球蛋白基因座包含猫科动物vh、dh和jh编码序列。在一方面,异源性部分猫科动物免疫球蛋白基因座包含猫科动物kappa(κ)vl和jl编码序列。在一方面,异源性部分猫科动物免疫球蛋白基因座包含猫科动物lambda(λ)vl和jl编码序列。在一方面,啮齿动物是小鼠。在一方面,非编码调控序列包括内源性宿主的下列一个或更多个序列:每个v基因区段之前的启动子、剪接位点和用于v(d)j重组的重组信号序列。在一方面,异源性部分猫科动物免疫球蛋白基因座还包含内源性宿主的以下序列中的一个或更多个:adam6基因、pax-5激活的基因间重复序列(pair)元件以及来自重链基因间控制区1(igcr1)的ctcf结合位点。
16、在一方面,非猫科动物细胞是哺乳动物细胞。在一方面,非猫科动物哺乳动物细胞是哺乳动物胚胎干(es)细胞。
17、在一方面,选择和分离其中内源性免疫球蛋白可变区基因座被异源性部分猫科动物免疫球蛋白可变区基因座替换的非猫科动物哺乳动物细胞。在一方面,该细胞是非猫科动物哺乳动物es细胞,例如啮齿动物es细胞。在一方面,至少一个分离的非猫科动物哺乳动物细胞用于产生表达异源性部分猫科动物免疫球蛋白可变区基因座的转基因非猫科动物哺乳动物。在一方面,至少一个分离的非猫科动物哺乳动物es细胞用于产生表达异源性部分猫科动物免疫球蛋白可变区基因座的转基因非猫科动物哺乳动物。
18、在一方面,提供了一种产生转基因啮齿动物的方法。在一方面,该方法包括:a)将至少一个位点特异性重组酶靶位点整合到位于内源性免疫球蛋白可变基因座上游的啮齿动物细胞的基因组中,以及将至少一个位点特异性重组酶靶位点整合到位于内源性免疫球蛋白可变基因座下游的基因组中。在一方面,内源性免疫球蛋白可变基因座包含vh、dh和jh基因区段。在一方面,内源性免疫球蛋白可变基因座包含vκ和jκ基因区段。在一方面,内源性免疫球蛋白可变基因座包含vλ和jλ基因区段。在一方面,内源性免疫球蛋白可变基因座包含vλ、jλ基因区段和cλ基因。在一方面,该方法包括:b)提供包含异源性部分猫科动物免疫球蛋白基因座的载体。在一方面,所述异源性部分猫科动物免疫球蛋白基因座包含嵌合猫科动物免疫球蛋白基因区段。在一方面,每个部分猫科动物免疫球蛋白基因区段包含猫科动物免疫球蛋白可变基因编码序列和啮齿动物非编码调控或支架序列。在一方面,部分猫科动物免疫球蛋白可变基因座的侧翼是用于位点特异性重组酶的靶位点。在一方面,靶位点能够与引入啮齿动物细胞的靶位点重组。在一方面,该方法包括:c)将载体和能够识别靶位点的位点特异性重组酶引入啮齿动物细胞。在一方面,该方法包括:d)允许在细胞的基因组和异源性部分猫科动物免疫球蛋白基因座之间发生重组事件,其中内源性免疫球蛋白可变基因座被异源性部分猫科动物免疫球蛋白基因座替换。在一方面,该方法包括:e)选择在步骤d)中产生的包含异源性部分猫科动物免疫球蛋白可变基因座的细胞;以及使用该细胞产生包含异源性部分猫科动物免疫球蛋白可变基因座的转基因啮齿动物。在一方面,该细胞是啮齿动物胚胎干(es)细胞。在一方面,该细胞是小鼠胚胎干(es)细胞。
19、在一方面,该方法还包括在步骤a)之后和步骤b)之前,通过引入识别第一组靶位点的重组酶来缺失内源性免疫球蛋白可变基因座的步骤,其中缺失步骤在啮齿动物细胞的基因组中留下在合适的位置的至少一组不能彼此重组的靶位点。在一方面,载体包含猫科动物vh、dh和jh编码序列。在一方面,载体包含猫科动物vl和jl编码序列。在一方面,异源性部分猫科动物免疫球蛋白基因座包含猫科动物kappa(κ)vl和jl编码序列。在一方面,异源性部分猫科动物免疫球蛋白基因座包含lambda(λ)vl和jl编码序列。在一方面,载体还包括以下一个或更多个:启动子、剪接位点和重组信号序列。
20、在一方面,提供了一种用于产生转基因非猫科动物哺乳动物的方法,该转基因非猫科动物哺乳动物包含异源性部分猫科动物免疫球蛋白可变区基因座。在一方面,该方法包括:a)将一个或更多个序列特异性重组位点引入非猫科动物哺乳动物宿主细胞的基因组,该序列特异性重组位点位于内源性免疫球蛋白可变区基因座的侧翼,并且不能彼此重组。在一方面,该方法包括:b)提供包含部分猫科动物免疫球蛋白基因座的载体,该基因座具有i)猫科动物可变区基因编码序列和ii)基于内源性宿主免疫球蛋白可变区基因座的非编码调控或支架序列。在一方面,编码和非编码调控或支架序列的侧翼是与引入a)的宿主细胞基因组的序列特异性重组位点相同的序列特异性重组位点。在一方面,该方法包括:c)将步骤b)的载体和能够识别一组重组酶位点的位点特异性重组酶引入细胞。在一方面,该方法包括:d)允许在a)细胞的基因组和异源性的部分猫科动物免疫球蛋白可变区基因座之间发生重组事件。在一方面,内源性免疫球蛋白可变区基因座被部分猫科动物免疫球蛋白基因座替换。在一方面,该方法包括:e)选择包含部分猫科动物免疫球蛋白基因座的细胞;和f)使用该细胞产生包含部分猫科动物免疫球蛋白基因座的转基因哺乳动物。
21、在一方面,非猫科动物转基因哺乳动物是啮齿动物,例如,小鼠或大鼠。
22、在一方面,提供了包括至少103个文库成员的多样性的免疫球蛋白文库(也称为谱库(repertoire))。
23、在一方面,提供了包括本文描述的部分猫科动物抗体的抗体谱库。在一方面,该谱库包括多种抗体,每种抗体特异性识别相同的靶抗原。这样的谱库可以被称为具有相同抗体类型或结构的抗体文库,其中抗体的抗原结合位点不同,例如,产生识别相同表位的亲本抗体的抗体变体。在一方面,抗体文库包括亲和成熟或以其他方式优化的抗体变体。在一方面,抗体文库包括特异性识别靶抗原的抗体,但是这种靶抗原的不同表位。
24、在一方面,筛选抗体谱库,并根据期望的结构或功能特性选择单个文库成员,例如,以产生抗体产物。
25、在一方面,提供了包括本文描述的部分猫科动物抗体的抗体谱库。在一方面,该谱库包括识别不同靶抗原的多种抗体。在一方面,通过用多组分抗原免疫非猫科动物哺乳动物获得该谱库,多组分抗原包括但不限于病毒或细菌,其可以具有许多不同的靶抗原,每个靶抗原可以包含多个表位。
26、在一方面,该谱库是初始的抗体文库,也可以称为“免疫前谱库”。在一方面,免疫前谱库由最近从骨髓中离开的成熟但缺乏抗原经验的b细胞表达。
27、在一方面,抗体谱库的特征可以在于包含至少约103种抗体的多样性,例如,至少约104、约105、约106或约107,每一个的特征在于不同的抗原结合位点。
28、在一方面,提供了表达异源性免疫球蛋白可变区基因座的非猫科动物哺乳动物细胞,该异源性免疫球蛋白可变区基因座具有猫科动物可变区基因编码序列和基于宿主基因组的内源性非猫科动物免疫球蛋白基因座的非编码调控或支架序列。在一方面,非猫科动物哺乳动物细胞表达嵌合抗体,该嵌合抗体包含完全猫科动物h或l链可变结构域和它们各自的非猫科动物哺乳动物细胞或哺乳动物内源性的恒定区。
29、在一方面,提供了一种非猫科动物转基因哺乳动物,其表达异源性免疫球蛋白可变区基因座,该基因座具有猫科动物可变区基因编码序列和基于宿主基因组的内源性非猫科动物免疫球蛋白基因座的非编码调控或支架序列。在一方面,非猫科动物转基因哺乳动物表达嵌合抗体,该嵌合抗体包含完全猫科动物h或l链可变结构域和它们各自的非猫科动物哺乳动物细胞或哺乳动物内源性的恒定区。
30、在一方面,提供了来自非猫科动物转基因哺乳动物的b细胞,其能够表达具有完全猫科动物可变序列的部分猫科动物抗体。在一方面,永生化b细胞被提供作为对特定抗原特异的单克隆抗体的来源。
31、在一方面,提供了从b细胞克隆的猫科动物免疫球蛋白可变区基因序列,用于产生或优化用于诊断性、预防性和治疗性用途的抗体。
32、在一方面,提供了能够产生具有完全猫科动物免疫球蛋白可变区序列的部分猫科动物单克隆抗体的非猫科动物杂交瘤细胞。
33、在一方面,提供了从产生单克隆抗体的杂交瘤中去除编码h链和l链免疫球蛋白可变结构域的vh和vl外显子,并将vh和vl外显子修饰为包括猫科动物恒定区的方法,从而产生在注射到猫体内时不具有免疫原性的完全猫科动物抗体。
34、在一方面,提供了一种产生用于治疗性或诊断性用途的猫科动物抗体的方法。在一方面,该方法包括:
35、(i)表达从转基因啮齿动物的抗体产生细胞克隆的具有猫科动物可变结构域的抗体,所述转基因啮齿动物的基因组包含的内源性啮齿动物免疫球蛋白基因座可变区已被缺失并被异源性免疫球蛋白基因座可变区替换,所述异源性免疫球蛋白基因座可变区在免疫球蛋白重链基因座包含嵌合vh、dh和jh免疫球蛋白可变区基因区段中的每一个的至少一个、和/或在免疫球蛋白轻链基因座包含嵌合vl和jl可变基因区段中的每一个的至少一个,其中每个嵌合基因区段包含猫科动物v、d或j免疫球蛋白可变区编码序列和啮齿动物免疫球蛋白可变区非编码基因区段序列;和
36、(ii)分离具有猫科动物可变结构域的抗体,其中所述抗体适于治疗性或诊断性用途。
37、在一方面,抗体从转基因啮齿动物的b细胞克隆。在一方面,啮齿动物是小鼠。在一方面,提供了通过本文描述方法产生的治疗性或诊断性抗体。
38、在一方面,提供了一种产生具有猫科动物可变结构域的治疗性或诊断性抗体的方法。在一方面,该方法包括:
39、(i)从转基因啮齿动物克隆由抗体产生细胞表达的抗体的猫科动物可变结构域,所述转基因啮齿动物的基因组包含的内源性啮齿动物免疫球蛋白基因座可变区已被缺失并被异源性免疫球蛋白基因座可变区替换,所述异源性免疫球蛋白基因座可变区在免疫球蛋白重链基因座包含嵌合vh、dh和jh免疫球蛋白可变区基因区段中的每一个的至少一个,和/或在免疫球蛋白轻链基因座包含嵌合vl和jl可变基因区段中的每一个的至少一个,其中每个嵌合基因区段包括猫科动物v、d或j免疫球蛋白可变区编码序列和啮齿动物免疫球蛋白可变区非编码基因区段序列;和
40、(ii)产生包含由转基因啮齿动物表达的抗体的猫科动物可变结构域的治疗性或诊断性抗体。
41、在一方面,猫科动物可变结构域从转基因啮齿动物的b细胞表达的抗体克隆。在一方面,啮齿动物是小鼠。在一方面,提供了通过本文描述方法产生的治疗性或诊断性抗体。
42、在一方面,提供了一种用于产生包括猫科动物可变结构域的单克隆抗体的方法。在一方面,该方法包括:
43、(i)提供来自转基因啮齿动物的b细胞,所述转基因啮齿动物的基因组包含的内源性啮齿动物免疫球蛋白基因座可变区已被缺失并被异源性免疫球蛋白基因座可变区替换,所述异源性免疫球蛋白基因座可变区在免疫球蛋白重链基因座包含嵌合vh、dh和jh免疫球蛋白可变区基因区段中的每一个的至少一个,和/或在免疫球蛋白轻链基因座包含嵌合vl和jl可变基因区段中的每一个的至少一个,其中每个嵌合基因区段包含嵌入啮齿动物免疫球蛋白可变区非编码基因区段序列中的猫科动物v、d或j免疫球蛋白可变区编码序列;
44、(ii)使b细胞永生化;和
45、(iii)分离包含由永生化b细胞表达的猫科动物可变结构域的单克隆抗体或编码该抗体的基因。
46、在一方面,该方法包括以下步骤:
47、(iv)克隆由所述b细胞表达的所述猫科动物可变结构域;和
48、(v)产生包含从转基因啮齿动物的b细胞克隆的猫科动物可变结构域的治疗性或诊断性抗体。
49、在一方面,提供了一种用于产生包括猫科动物可变结构域的抗体的方法。在一方面,该方法包括提供转基因啮齿动物,所述转基因啮齿动物的基因组包含的内源性啮齿动物免疫球蛋白基因座可变区已被缺失并被异源性免疫球蛋白基因座可变区替换,所述异源性免疫球蛋白基因座可变区在免疫球蛋白重链基因座包含嵌合vh、dh和jh免疫球蛋白可变区基因区段中的每一个的至少一个,和/或在免疫球蛋白轻链基因座包含嵌合vl和jl可变基因区段中的每一个的至少一个,其中每个嵌合基因区段包括嵌入啮齿动物免疫球蛋白可变区非编码基因区段序列中的猫科动物v、d或j免疫球蛋白可变区编码序列,其中转基因啮齿动物的异源性免疫球蛋白基因座表达包含猫科动物可变结构域的抗体。
50、在一方面,该方法包括分离具有由转基因啮齿动物表达的猫科动物可变区的抗体或编码该抗体的基因。在一方面,该方法包括:(i)从表达针对靶抗原的特异性抗体的转基因啮齿动物获得b细胞;(ii)使b细胞永生化;和(iii)从永生化的b细胞中分离针对靶抗原的特异性抗体。
51、在一方面,该方法包括从对特定抗原特异的b细胞克隆猫科动物可变区。在一方面,啮齿动物是小鼠。在一方面,该方法包括使用从b细胞克隆的猫科动物可变区产生治疗性或诊断性抗体。在一方面,提供了通过本文描述方法产生的治疗性或诊断性抗体。
52、这些和其他方面在下面更详细地描述。
- 还没有人留言评论。精彩留言会获得点赞!