加兰他敏在防治豆粕饲料引起的鱼类肠炎中的应用的制作方法
本发明属于兽药学领域,具体涉及加兰他敏在防治豆粕诱导的鱼类肠炎中的应用。
背景技术:
水产动物肠炎病是导致水产养殖经济损失的主要疾病之一。水产动物尤其是鱼虾,都需要在其饲料中添加鱼粉等动物性蛋白质,然而鱼粉资源的逐年紧缺已成为限制我国水产养殖业可持续发展的关键因素,因此使用植物蛋白源替代鱼粉并提高其利用率,是水产养殖业发展的必然选择。但是植物蛋白质源,如豆粕、棉粕等,替代鱼粉比例过大时普遍会诱发鱼类肠道炎症、肝脏病变及生长下降,使得植物蛋白源在水产动物饲料中的应用受到限制。有研究表明,豆粕在草食性鱼类草鱼饲料中替代量最大为60%,在杂食性鱼类鲤鱼和胡子鲶的饲料中替代量最大为45%。而豆粕在肉食性军曹鱼中替代量最大达到10%对健康无影响,当替代比例为20%、30%时,则对其生长、增重率、饲料系数、成活率以及肥满度影响显著。目前有关鱼类肠粘膜免疫系统对植物蛋白源的应答和免疫调节的研究主要集中在豆粕诱导的肠炎(soybeanmeal-inducedenteritis,sbmie)及其生长抑制。已经有报道sbmie症状的鱼类包括鲑鳟鱼类,如大西洋鲑和虹鳟;鲤科鱼类,如斑马鱼和鲤鱼;鲶鱼类,如黄颡鱼等。在sbmie过程中,对前炎症因子及其炎症反应研究较常见,而对缓解/抑制炎症的免疫机理及相应药理研究还处于起步阶段,并且效果不稳定。
目前,在生产中应用的多的是具有广谱治疗效果的抗生素和中药制剂,而有针对性的肠炎药物还非常缺乏。大量、长期地使用抗生素药物,极容易产生耐药菌株,导致局部地区鱼类肠炎病频频暴发,出现用药量加大和久治不愈等现象。药量加大及药残也造成水产动物潜在性地危害人体健康。因而迫切需要替代抗生素,从鱼体自身的肠粘膜免疫机理来寻找线索,进而达到无公害治疗鱼类肠炎的目的。
鱼类肠黏膜中存在肠神经系统(entericnervoussystem),其和免疫系统能够交互作用,形成调节控制消化道功能的独立整合系统。肠黏膜下层的淋巴细胞,如t、b淋巴细胞,还包括先天性淋巴细胞,与神经纤维之间是密切接触的,存在免疫和神经交互的局部环路。神经细胞和免疫细胞的相互作用依赖细胞以及可溶分子的接触,如细胞因子、趋化因子、神经肽、神经营养因子等的信号传递,而免疫细胞也可表达神经递质的受体,因而能够感受神经递质,进而调控免疫反应。
肠道神经免疫环路中胆碱能抗炎通路(cholinergicanti-inflammatorypathway,caip)发挥了重要作用,其抗炎效应反应时间非常短,它能够快速而直接地调节全身性炎症反应。迷走神经或具有胆碱乙酰转移酶(cholineacetyltransferase,chat)的免疫细胞激活后可释放乙酰胆碱,通过乙酰胆碱受体(nachr-alpha7)刺激t/b淋巴细胞、巨噬细胞、粒细胞、树突细胞等免疫细胞,抑制促炎因子的释放,从而完成免疫抑制作用;而nachr-alpha7激活后,主要通过2个信号通路来影响免疫细胞,即nf-κb及jak/stat路径。最近研究发现调节性t细胞也可表达nachr-alpha7,且该受体激活后可增强免疫抑制功能。激活nachr-alpha7可以抑制t细胞的增殖及th1和th17细胞因子的产生,促进th1向th2的漂移,从而达到免疫调节作用。对b淋巴细胞来说,胆碱受体主要参与促进b细胞共刺激分子的表达,促进前b细胞向成熟b细胞转换及b细胞的增殖等。
而作为常见的强效并可逆的乙酰胆碱酯酶抑制剂,加兰他敏可通过抑制乙酰胆碱的降解来增强乙酰胆碱通路,进而可以治疗阿尔茨海默氏病(已获得fda批准),另外也可用于重度肌无力症的治疗。关于其抗炎作用,目前只见到对浓度血症具有一定疗效。因而,我们推测在肠道炎症中,加兰他敏可能存在对胆碱能抗炎作用的加强作用,这对于水生动物肠炎的治疗是具有应用潜力的。加兰他敏在体内的清除半衰期为7-8小时,通过肝脏代谢,经排泄物清除,因而只要停止使用即可避免残留在水生动物产品中。加兰他敏常见的应用形式有氢溴酸盐形式、n末端烷基修饰形式,以及一些添加树脂合成的缓释溶液形式。
本发明首次将胆碱能抗炎通路理论用于养殖鱼类的治疗,发现乙酰胆碱酯酶抑制剂加兰他敏能够缓解豆粕饲料引起的肠炎。加兰他敏对养殖鱼类肠炎的治疗作用具体来说,能够降低炎症因子的表达水平,并恢复肠粘膜紧密连接蛋白的表达水平,同时抑制肠上皮的凋亡基因表达,从而达到抗炎及修复肠黏膜屏障的作用,同时对生长无影响。基于我们在模式生物斑马鱼和养殖鱼类草鱼中的发现以及应用经验,加兰他敏因而可用于防治豆粕饲料引起的养殖鱼类肠炎,并且从另一方面来说可以替代目前常用的抗生素类肠炎药物,减少环境污染和药物残留,在水产领域具有广阔的应用前景。
技术实现要素:
本发明的目的是在于提供了加兰他敏在防治豆粕饲料引起的养殖鱼类肠炎中的应用,加兰他敏在降低促炎因子同时提高抗炎因子的表达、提高肠道屏障功能等方面预防或治疗豆粕诱导的鱼类肠炎,其应用方法包括作为饲料添加剂和注射用针剂。
为了实现上述的目的,本发明采用以下技术措施:
加兰他敏作为饲料添加剂在缓解豆粕饲料引起的养殖鱼类肠炎中的应用:
在豆粕替代50%蛋白源的饲料中添加加兰他敏0.5~1.0ppm,连续饲喂杂食性鱼类斑马鱼3周(每天早晚各一次)后用ms-222麻醉,解剖取肠道样品通过qpcr、westernblot、h.e.染色、免疫组化检验防治效果。结果表明,饲料中添加浓度为0.5-1ppm的加兰他敏在形态水平、生长水平、基因表达(包括免疫、紧密连接、细胞凋亡)等方面均表现出具有缓解豆粕引起的肠炎的效果,并且能够促进鱼体采食量的恢复。
加兰他敏在制备预防或治疗豆粕饲料引起养殖鱼类肠炎的药物中的应用:
在患病草鱼(豆粕替代70%蛋白源的饲料饲喂7周)背部肌肉注射加兰他敏(0.05~0.1mg/kg鱼体重),每天注射2次,共注射2天,第4天取肠组织样品进行荧光定量qpcr分析,检测肠道炎症相关免疫基因的表达情况。结果表明,与注射pbs的病鱼相比,注射加兰他敏可显著降低sbmie草鱼肠组织中炎症相关免疫基因的表达水平,包括促炎因子il-1b和il17、t细胞表面标记cd4、调节性淋巴细胞转录因子foxp3。因而,在已患豆粕引起肠炎的鱼体中,注射一定剂量的加兰他敏可通过抑制炎性因子的表达,来发挥对肠炎的抑制作用,进而缓解病情。
通过对常见养殖鱼类的胆碱酯酶cholinesterase蛋白和乙酰胆碱受体nachr-alpha7蛋白的进化分析发现,鱼类中广泛存在加兰他敏的底物——胆碱酯酶,并且与哺乳类的序列相比非常保守;另一方面常见养殖鱼类都存在胆碱激活抗炎通路所必须的乙酰胆碱受体。因此对于具有乙酰胆碱受体的养殖鱼类因豆粕引起的肠炎,加兰他敏亦具有保护作用。本发明涉及的鱼类具体包括鲤科鱼类,如草鱼、青鱼、鲢、鳙、鲤、鲫、团头鲂等,鲑鳟鱼类,如大西洋鲑、虹鳟等,以及罗非鱼、斑点叉尾鮰等常见养殖鱼,并且特别适用于草食性和杂食性养殖鱼类。
与现有技术相比,本发明具有以下优点及有益效果:
本发明首次提出加兰他敏在防治豆粕饲料引起养殖鱼类肠炎中的应用,所采用的加兰他敏作为一种利用鱼体自身胆碱能抗炎通路来实现抗炎作用的药物,作为鱼类饲料添加剂使用时,避免了抗生素类药物的残留及环境安全性问题,无毒副作用,符合绿色渔药标准,可广泛应用于豆粕等含有植物蛋白源的饲料中,用于提高水生动物肠粘膜免疫力,预防饲料引起的肠炎,进而提高生长效率,同时比添加鱼粉降低成本。加兰他敏无论通过添加至鱼饲料中或是肌肉注射,都可激活养殖鱼类肠道内抗炎因子的表达,抑制肠道炎症,并维持肠道上皮屏障紧密连接,同时抑制肠上皮凋亡,因而可作为一种新型的具有抗炎效果的饲料添加剂或注射用针剂。
附图说明
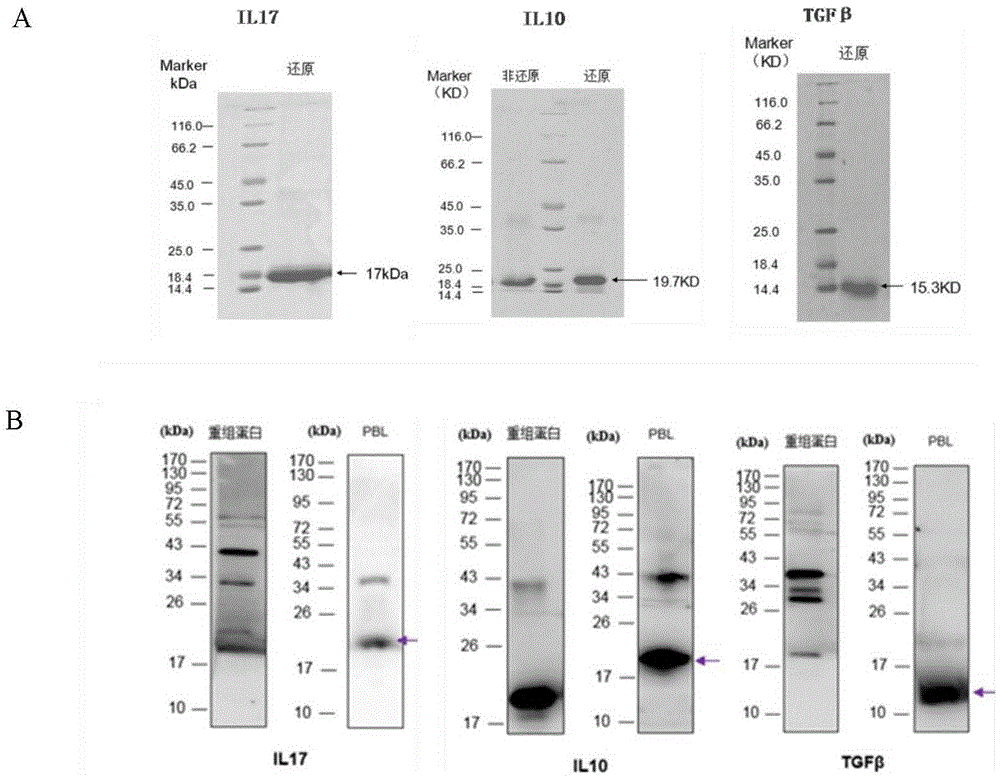
图1为il17、il10、tgf-beta的多克隆抗体特异性检测结果图。原核表达的il17、il10、tgf-b的各自抗原经过亲和层析柱得到有效纯化(a),用纯化的抗原免疫兔子得到il17、il10、tgf-b的各自的多克隆抗体,可特异识别鱼体中的相应的天然蛋白(b)。
图2为草鱼豆粕诱导肠炎各阶段后肠转录组都存在乙酰胆碱抗炎通路的激活。在神经递质与受体相互作用(neuroactiveligand-receptorinteraction)的kegg通路中,左边第一列的第一个神经递质acetylcholine即为乙酰胆碱,灰色的框为检测到转录本的基因。
图3为加兰他敏在cmap数据库中检索得到的表达谱相似药物。
图4为采用ngf+hotnet+cmap联合分析得到的与加兰他敏的表达谱相似的药物。
图5为斑马鱼豆粕饲料添加不同浓度加兰他敏后其后肠的组织形态恢复情况。fm:鱼粉饲料组(阴性对照);sbm:豆粕饲料组(阳性对照);sbm+0.5ppmgal:添加0.5ppm加兰他敏的豆粕饲料组;sbm+1ppmgal:添加1ppm加兰他敏的豆粕饲料组。
图6为斑马鱼豆粕饲料添加不同浓度加兰他敏后生长指标的恢复情况。ibm:初始体重,fbw:终体重,wg:总增重,fi:采食量,fe:饲料效率,sgr:特定生长率。豆粕为肠炎的阳性对照,而鱼粉组作为阴性对照。梅花或井号表示差异显著p<0.05,前者代表与鱼粉饲料组对比上调,后者代表与豆粕饲料组代表下调。
图7为斑马鱼豆粕饲料添加不同浓度加兰他敏后炎症相关免疫基因在后肠组织中的表达情况。a:qpcr,纵轴为相对表达量变化倍数,横轴为不同组;b:westernblot,方框中为豆粕饲料引起肠炎相关免疫蛋白的表达情况;c:免疫组化,检测cd4蛋白在后肠粘膜组织中的定位。豆粕为肠炎的阳性对照,而鱼粉组作为阴性对照。梅花号或者井号表示差异显著p<0.05,前者代表与鱼粉饲料组对比显著上调,后者代表与豆粕饲料组相比显著下调。
图8为斑马鱼豆粕饲料添加不同浓度加兰他敏后肠上皮紧密连接(a)及细胞凋亡(b)相关基因表达情况。豆粕为肠炎的阳性对照,而鱼粉组作为阴性对照。梅花或井号表示差异显著p<0.05,前者代表与fm组对比上调,后者代表与sbm组相比下调。
图9为患病草鱼(70%替代蛋白源的饲料投喂了7周)注射加兰他敏之后的免疫基因表达变化。fm:鱼粉饲料组;70%豆粕:完全替代鱼粉的豆粕饲料组。同样小写字母表示差异不显著p>0.05,不同小写字母表示差异显著p<0.05。
图10为鱼类的胆碱酯酶cholinesterase蛋白的nj树进化分析。斑马鱼cholinesterase基因(ensdarg00000031796.8)和草鱼cholinesterase基因(ci_gc_7222)采用其中的编码区序列翻译得到。ncbi下载的cholinesterase蛋白的序列号分别为:人类(np_000046.1);小鼠(np_033868.3);银鲛(xp_023648146.1);鲤鱼(ktg31068.1);青鳉(np_001188442.1);罗非鱼(xp_003441667.2);黄鳝(xp_020456087.1);大黄鱼(kkf12911.1)。而选作外类群的斑马鱼乙酰胆碱转移酶(chat,cholineacetyltransferase)的序列号为np_001124191.1。
图11为常见养殖鱼类的乙酰胆碱受体nachr-alpha7蛋白的nj树进化分析。除了草鱼的nachr-alpha7蛋白来自中国科学院水生生物研究所草鱼基因组数据库,基因号为ci_gc_10747和ci_gc_18333;其他鱼类的都来自ncbi,它们的序列号分别为:斑马鱼nachr-alpha7蛋白(aao91913.1)、斑马鱼nachr-alpha7-like蛋白(xp_005174278.4)、鲤鱼nachr-alpha7同源蛋白(ktf81060.1)、鲱鱼nachr-alpha7蛋白(xp_012674679.1)、鲃鱼nachr-alpha7蛋白(xp_016140050.1)、大西洋鲑nachr-alpha7蛋白(xp_013979444.1)、虹鳟nachr-alpha7蛋白(xp_021421301.1)、北极红点鲑nachr-alpha7蛋白(xp_023840673.1)、斑点叉尾鲴nachr-alpha7蛋白(xp_017332872.1)、黄鳝nachr-alpha7蛋白(xp_020461482.1)、大黄鱼nachr-alpha7蛋白(xp_019113632.1)、罗非鱼nachr-alpha7蛋白(xp_003450802.1)。作为外类群的人类的nachr-alpha7蛋白序列号为aaa83561.1。
具体实施方式
下面结合附图及具体实施例对本发明作进一步说明,但不应理解为对本发明的限制。在实施例中涉及的所有分子生物学操作方法若未特别说明,均为本领域技术人员所熟知的常规方法。
以下为实施例中涉及的试验材料、试剂及仪器设备:
草鱼的细胞因子il17、il10、tgf-beta的多克隆抗体为本实验室自己制备,其方法为:
(1)选择抗原片段:取去掉信号肽的其余所有的蛋白序列,草鱼il17(ncbiaccession:kc978892.1)的抗原片段为从第28个氨基酸到c末端,草鱼il10(ncbiaccession:jq768312.1)的抗原选择从第25个氨基酸到c端,草鱼tgf-beta(ncbiaccession:eu099588.1)的抗原片段从第266个到第377个氨基酸;
(2)选择表达载体:il17和tgf-beta采用gst-tag标签表达载体pgex-4t-1,il10采用his-tag标签表达载体pet-23b;
(3)获得重组蛋白:在大肠杆菌de3中进行原核表达,获得il17(可溶)、il10(包涵体复性为可溶)、tgf-beta(可溶)的重组蛋白,大小分别为17kda、19.7kda、15.3kda(图1a);
(4)免疫动物获得抗体:注射重组蛋白溶液(600μg/ml)到兔子体内,免疫3次(每次1ml,间隔2周),注射方法为第一次免疫:腘肌淋巴结和足跖部皮下,第二、三次:背部皮下多点注射,第三次免疫后1周时取血清,获得多克隆抗体;
(5)抗体特异性验证结果表明:在鱼体分离的外周血白细胞中il17抗体识别大小约为22kda的单一条带、il10抗体识别大小约为20kda的单一条带、tgf-beta抗体识别大小约为15kda的单一条带,与预测大小相当,证明我们制备的抗体能够特异识别相应的鱼体内的蛋白。
用于本发明的斑马鱼foxp3和cd4为genetex公司产品(编号分别为gtx16590和gtx16589),h.e.染液(碧云天),提取rna的试剂盒(tiangen天根生化科技(北京)有限公司),一抗(斑马鱼cd4兔源多抗购自genetex公司,斑马鱼foxp3兔源多抗购自creativediagnostics公司),trizol(invitrogen公司),
实施例1缓解肠炎的功能成分加兰他敏的药效预测
草鱼转录组的kegg通路分析暗示鱼类胆碱能抗炎通路存在:发明人已有的研究表明,40%豆粕替代鱼粉组草鱼的肠道炎症在第5周开始至7周具有可自我恢复的能力。在其病程各阶段,包括0天、3周、5周、7周,取后肠组织测转录组,其中kegg通路中分析结果表明,在豆粕诱导的肠炎发生和消退过程中乙酰胆碱(acetylcholine)通路都有激活(图2),暗示鱼类胆碱能抗炎通路可能存在。
然后,我们采用cmap数据库对胆碱酯酶抑制剂加兰他敏进行药效预测:加兰他敏作为胆碱酯酶抑制剂可能促进胆碱能抗炎通路的功能,为了进一步推定其在肠炎中可能发挥的作用,发明人采用cmap数据库进行药效预测。connectivitymap(cmap)(https://portals.broadinstitute.org/cmap/)为一个基因表达谱数据库,利用小分子药物处理人类细胞后的基因表达差异,建立一个小分子药物、基因表现与疾病相互关连的生物应用数据库。在药物开发领域上,发明人可以快速利用基因表达谱的数据比对出与疾病高关联性的药物,并能够归纳出药物分子可能作用的机制方向。在cmap库中检索galanthamine,选择mcf7细胞系,up/downthreshold设置为±0.67(2-fold)得到与galanthamine表达谱最相似的药物。如图3所示,cmap中p值排名最高的且均值为正前四种的药物分别为:1.sulfaphenazole;2.flunixin;3.fenoprofen;4.norfloxacin。它们均为抗菌抗炎药物,这说明加兰他敏也极有可能具有一定的抗炎疗效。
表1cmapp值排名前四药物简介
同时,采用联合ngf+hotnet+cmap数据库分析,对加兰他敏进行药效预测;network-guidedforests(ngf)用差异基因表达谱以及基因网络作为输入,训练得到一个能将表达谱正确划分到对应的条件下的分类模型。根据对ngf分类能力的贡献,ngf为基因网络中的每一个基因和基因互作分配一个重要性分值(importantscore,is)。hotnet方法是基于热扩散理论分别处理上调/下调数据的组合方法,可以鉴定出在基因相互作用网络中有显著差异的关键子网络。从ncbi-geodataset中下载肠炎病人芯片数据,芯片编号:gse11223(202个样本),以疾病组为正样本,健康组为负样本,通过ngf计算得到芯片中共12929个基因的重要性分值。选取重要性排名前3000的基因,以它们的重要性分值为初始热度,输入hotnet2构建肠炎疾病致病基因子网络(使用string-ppi);然后,在得到的子网络中选取p值最小为0.28的子网络,将其中的基因对应到cmap的探针,并根据芯片计算的foldchange值确定上下调的探针名称,输入cmap,检索表达谱相似的药物。如图4所示,检索得到的排名最靠前且均值为负的药物中,nortriptyline可治疗过敏性肠综合征,而trichostatina和roxarsone分别具有抗真菌和抗虫效果,提示加兰他敏可能具有抗肠道炎症的作用。
表2ngf+hotnet+cmap排名前3药物简介
实施例2加兰他敏作为饲料添加剂在缓解豆粕诱导的斑马鱼肠炎中的应用
1、斑马鱼sbmie建模及加兰他敏添加实验饲料配方、制备方法
表3斑马鱼sbmie建模及加兰他敏添加实验饲料配方
fm为对照鱼粉饲料,50sbm为本实施例所用的豆粕饲料(50%替代蛋白源),50sbm+0.5ppmgal为添加0.5ppm加兰他敏的豆粕饲料,50sbm+1ppmgal为添加1ppm加兰他敏的豆粕饲料。
制备方法如下:
(1)将鱼粉、豆粕、淀粉、面粉、纤维素、矿物质预混料、维生素预混料等过60目筛,其中鱼粉、豆粕、矿物质混合物中可能存在较大颗粒如鱼骨和豆皮等,应粉碎后过60目筛;对于含量较少的维生素、矿物质:vd3、vk3、vb12、硫胺素、vb6、叶酸、硫酸铜和亚硒酸钠需稀释20倍配预混料;加兰他敏需要稀释100倍配预混料;所述的每千克矿物质混合物含(g):硫酸镁(mgso4·2h2o)60.530g,硫酸亚铁(feso4·h2o)23.110g,硫酸铜(cuso4·5h2o)0.010g,硫酸锌(znso4·h2o)0.620g,硫酸锰(mnso4·h2o)1.640g,碘化钾(ki)0.070g,亚硒酸钠(naseo3)0.005g,用淀粉调节到1kg;所述的每千克维生素预混物中含(g):维生素b1(thiamin)0.05,维生素b2(riboflavin)0.55,维生素b6(pyridoxine)0.59,维生素b12(cyanocobalamine)0.83,泛酸(pantothenicacid)2.89,叶酸(folicacid)0.40,肌醇(inositol)19.39,烟酸(niacin)2.24,生物素(biotin)4.91,维生素c(ascorbic)7.16,维生素a(vitamina)2.40,维生素d(vitamind)0.40,维生素e(vitamine)12.55,维生素k(vitamink)0.80,用淀粉调节到1kg;
(2)根据饲料配方中各组份重量百分比的要求分别按比例称取鱼粉、豆粕、淀粉、面粉、纤维素、矿物质预混料、维生素预混料以及加兰他敏预混料;
(3)将称取的鱼粉、豆粕、淀粉、纤维素和面粉,充分搅拌混合均匀;
(4)将矿物质预混料、维生素预混料、加兰他敏预混料加入步骤(3)中的混合物继续搅拌,尽量混合均匀;
(5)将鱼油加入步骤(4)中的混合物,充分搅拌,混合均匀;
(6)用喷壶加入适量的纯净水(10%)到步骤(5)中的混合物,继续进行搅拌,混合均匀,获得一种混合物;
(7)用制粒机将步骤(6)中混合物制成粒径为0.4-0.6mm的颗粒;将制成的颗粒放到烘干机中进行烘干,温度控制在58-60℃,烘干至含水量10%左右;烘干后粉碎过15-35目筛,得到相应规格的颗粒饲料,包装放置在-20℃冰箱保存。
2、所有的试验斑马鱼(2月龄成鱼)用鱼粉组饲料饲喂2周后,按照分组分别饲喂对应组的饲料3周,每次缓缓投食,至不再抢食,采食量约为23mg每条鱼每天,试验阶段斑马鱼生长发育正常,存活率98%以上。
用ms-222麻醉(浓度0.1mg/ml),解剖取肠道样品通过qpcr、westernblot、h.e.染色、免疫组化检验防治效果,结果如下:
病理切片形态水平:在各组斑马鱼,包括鱼粉组、豆粕组、豆粕+加兰他敏0.5ppm组、豆粕+加兰他敏1ppm组,取后肠组织(n=3)固定,将后肠组织进行梯度脱水和浸蜡、石蜡包埋、切片(5微米),进行h.e.染色,在正置显微镜白光下进行形态观察,实验结果(见图5)表明在50%豆粕替代饲料中添加0.5-1ppm加兰他敏可预防豆粕引起的鱼类肠炎,且1ppm组比0.5ppm组的肠绒毛结构恢复的更好。
生长水平:生长指标包括总增重(weightgain)、初始体重(initialbodyweight)、饲料效率(feedefficiency)、终体重(finalbodyweight)、采食量(feedintake)、特定生长率(specificgrowthrate);生长指标的统计分析:采用平均数和标准差分析,然后进行t检验获得差异显著性,p<0.05为显著,p<0.01为极为显著。结果表明(见图6),从多项生长指标来看,添加加兰他敏的豆粕饲料具有帮助鱼体从豆粕饲料引起的生长抑制中恢复的作用,具体表现在0.5ppm添加量可缓解生长抑制(虽然只是显著恢复采食量,但是可缓解终体重、总增重下降),1ppm可基本恢复正常生长(显著恢复终体重、总增重、采食量、饲料效率)。
基因表达水平:
(1)转录水平
取后肠组织(n=3),提取rna,制备cdna模板,然后进行qpcr检测免疫、紧密连接、细胞凋亡基因在转录水平的表达量变化,免疫基因包括调控免疫反应的重要转录因子nk-kb和foxp3a、肠炎相关模式识别受体tlr4、促炎因子il1b、tnf-alpha、趋化因子il8,抗炎因子tgf-beta和il10(见图7a);紧密连接基因包括zo-1、occludin、claudin(1,b,c,h,12,15a)(见图8a);细胞凋亡基因包括caspase3、caspase8、caspase9(见图8b)。结果表明:从转录水平来说,对起始免疫反应的重要转录因子nf-kb、肠炎相关模式识别受体tlr4、急性炎性因子il1b和tnf-a、趋化因子il8、免疫调节相关转录因子foxp3(仅1ppm表现显著),可显著降低炎性高表达;而对抑炎效应因子tgf-b、il-10,可显著逆转炎性表达抑制。从蛋白水平,添加0.5ppm可降低豆粕饲料引起的foxp3、il17、tnf-a高表达,同时提高抗炎因子il-10表达水平,而添加量达到1ppm还能够刺激辅助性t细胞表面分子标记cd4在肠绒毛固有层(lp层)的表达,暗示了另一些th细胞类群开始起作用。从紧密连接基因的转录水平来看,添加加兰他敏可逆转tj连接基因zo-1以及tj支柱基因ocludin、claudin1、claudinb(仅1ppm表现显著)、claudinc的炎性表达抑制,同时显著降低钙离子通道蛋白claudin12和钾离子通道claudin15a的炎性高表达。从细胞凋亡基因的转录水平来看,可显著降低caspase3、8、9的炎性高表达。
(2)蛋白水平
westernblot检测蛋白水平免疫基因的表达量的变化,具体为制备后肠组织蛋白质匀浆液,采用免疫蛋白抗体,包括免疫调节相关foxp3a、辅助t细胞表面标记蛋白cd4、肠炎的主要促炎因子il17和抑炎因子tnf-a和il10,进行westernblot检测蛋白水平的表达量变化。结果表明(见图7b),与未添加加兰他敏的豆粕饲料组相比,添加加兰他敏的豆粕饲料组鱼后肠il17、cd4和foxp3显著减少,而抑炎因子tnf-a和il10显著增多。
免疫组化检测蛋白水平免疫基因的表达的组织细胞定位的变化,具体为后肠组织先在4%多聚甲醛中固定过夜(4℃),然后在-20℃采用oct包埋剂冷冻包埋,进行冰冻切片(7微米),再用cd4抗体进行免疫组化实验,检测辅助t细胞表面抗原cd4的蛋白定位的时空变化。结果表明,在后肠组织固有层,添加0.5ppm加兰他敏可显著减少cd4阳性细胞的数量,同时cd4阳性细胞亮度恢复至与鱼粉组一致;而添加1ppm加兰他敏也显著减少cd4阳性细胞,但是一些cd4阳性细胞亮度非常高,暗示可能有与炎症期不同类群的辅助性t细胞在起作用。
综合转录水平和蛋白水平的表达变化,获得饲料中添加浓度为0.5-1ppm的加兰他敏对豆粕诱导肠炎在肠组织炎症的各个方面,包括组织形态、基因表达(包括免疫、紧密连接、细胞凋亡)等方面具有缓解效果,并且能够促进鱼体一系列生长指标的恢复。
本实施例采用的斑马鱼是目前世界上公认的模式鱼类,其属于鲤科鱼类。中国人喜欢吃的四大家鱼等淡水鱼都属于鲤科鱼类,而鲤科鱼遗传背景接近,基因同源率高,因此,通过斑马鱼的豆粕引起的肠炎的疾病建模,以及在肠组织形态、生长指标、免疫和肠粘膜屏障相关基因表达的检测,可推测加兰他敏适用于至少是鲤科鱼类的食源性肠炎(主要为豆粕引起的肠炎)的预防。
实施例3肌肉注射加兰他敏降低豆粕诱导肠炎患病草鱼后的肠道炎症因子表达
1、取投喂7周70%豆粕饲料组及鱼粉组的草鱼(草鱼sbmie建模方法及饲料配方等参见integrativetranscriptomicstudyrevealsimmunemechanismfortheresiliencetosoybeanmealstressinfishgutandliver,frontierinphysiology)各10尾,在网箱中保持温度23~25℃,溶氧大于5mg/l,禁食;
2、背部肌肉注射氢溴酸加兰他敏(0.05~0.1mg/kg鱼体重),每天注射2次,共注射2天,第4天取肠组织样品;
3、进行荧光定量qpcr分析,检测肠道炎症相关免疫基因的表达情况。荧光定量数据统计分析结果表明,与注射pbs的病鱼相比,注射加兰他敏可显著降低sbmie草鱼肠组织中与急性炎症相关免疫基因,包括促炎因子il-1b、t细胞表面标记cd4、调节性淋巴细胞转录因子foxp3的表达水平(图9);同时,与慢性炎症有关的th2细胞转录因子t-bet略有上升,而与自身免疫有关stat3恢复到与鱼粉组相似水平,il17变化不显著。
实施例4常见养殖鱼类的胆碱酯酶和乙酰胆碱受体蛋白的进化分析
首先,为了论证加兰他敏对胆碱酯酶的抑制作用在鱼类中是否保守,我们对鱼类的胆碱酯酶蛋白(加兰他敏的作用底物)进行nj树进化分析。序列检索结果表明,胆碱酯酶蛋白在硬骨鱼类中广泛存在,(见图10);此外,软骨鱼也有胆碱酯酶蛋白(见图10)。其中的序列,除了斑马鱼和草鱼的蛋白序列分别来自emsembl(http://www.ensembl.org/)和草鱼基因组(http://bioinfo.ihb.ac.cn/gcgd)的基因组拼接的预测结果,其他鱼类以及哺乳类的序列均来自ncbi。进化分析结果表明,鱼类(包括斑马鱼daniorerio、鲤鱼cyprinuscarpio、青鳉oryziaslatipes、草鱼ctenopharyngodonidellus、大黄鱼larimichthyscrocea、黄鳝monopterusalbus、罗非鱼oreochromisniloticus、叶吻银鲛paramormyropskingsleyae)和哺乳类(人homosapiens和小鼠musmusculus)的胆碱酯酶蛋白序列非常保守,聚为一支。综合以上结果,可推测加兰他敏作为胆碱酯酶抑制剂的作用在进化上是保守的,不仅可作用于哺乳类,还可作用于鱼,并且可能对草食性(如草鱼)和杂食性鱼类(如鲤鱼、斑马鱼、青鳉、罗非鱼、黄鳝、大黄鱼)均作用显著。
而加兰他敏作为胆碱酯酶抑制剂的效应,表现为提高体内胆碱总量,必须通过经由乙酰胆碱受体的信号传导来加强胆碱能抗炎通路的激活。因此,为了论证养殖鱼类中乙酰胆碱抗炎通路的广泛存在,我们也对常见养殖鱼类的乙酰胆碱受体nachr-alpha7进行了蛋白序列比对和构建nj树进化分析。乙酰胆碱受体nachr-alpha7蛋白在草鱼基因组(http://bioinfo.ihb.ac.cn/gcgd)中找到2个基因,并下载其编码序列。其他的鱼类乙酰胆碱受体nachr-alpha7蛋白序列均来自ncbi。胆碱能抗炎通路中乙酰胆碱受体nachr-alpha7蛋白在鱼类的进化分析表明(见图11),常见的养殖鱼类都具有该受体,且蛋白氨基酸序列相似度很高(80%-100%)。除了草鱼nachr-alpha7基因来自中国科学院水生生物研究所草鱼基因组数据库,基因号为ci_gc_10747和ci_gc_18333,并下载预测蛋白序列;其他鱼类(包括斑马鱼、鲤鱼、鲱鱼clupeaharengus、金线鲃sinocyclocheilusgrahami、斑点叉尾鲴ictaluruspunctatus、黄鳝、大黄鱼、罗非鱼、大西洋鲑salmosalar、虹鳟oncorhynchusmykiss、北极红点鲑salvelinusalpinus、)以及人类的nachr-alpha7蛋白都来自ncbi。因而,可推测常见的各种食性的养殖鱼类都存在胆碱激活抗炎通路所必须的乙酰胆碱受体。
因而,综合以上胆碱酯酶和胆碱受体的进化分析,可推测对于具有乙酰胆碱受体的养殖鱼类(特别是草食性和杂食性鱼类)发生的因豆粕引起的肠炎,加兰他敏很可能都具有保护作用。本发明涉及的鱼类具体包括鲤科鱼类,如草鱼、青鱼、鲢、鳙、鲤、鲫、团头鲂等,鲑鳟鱼类,如大西洋鲑、虹鳟等,以及罗非鱼、斑点叉尾鮰、黄鳝、大黄鱼等。
- 还没有人留言评论。精彩留言会获得点赞!