精氨酸在制备提高桔梅奇酵母普切明酸生成能力及其生防效力的保鲜剂中的应用的制作方法
本发明属于果实采后病害生物防治技术领域,涉及精氨酸在制备提高桔梅奇酵母(metschnikowiacitriensis)普切明酸生成能力及其生防效力的保鲜剂中的应用。
背景技术:
自从1985年,wilson和pusey综述了利用拮抗菌对采后果实进行釆后生物防治的应用潜力后,关于拮抗酵母的研究报告呈现指数增长,利用拮抗酵母控制采后果实病害的研究也取得了长足的进展,其中一些综合性能良好的拮抗酵母也实现了商业化应用,如candidaoleophila(aspiretm,ecogen,langhorne,pa,us),cryptococcusalbidus(yieldplustm,lallemand,montreal,canada),candidasake(candifruittm,irta,lleida,spain),c.oleophila(nexytm,leasafre,lille,france),aureobasidiumpullulans(boniprotecttm,bio-ferm,tulln,austria),metschnikowiafructicola(shemertm,bayer,leverkusen,germany)。但是这些商业化的生防制剂以及有研究报道的具有潜在商业化价值的拮抗酵母与传统的通用型化学杀菌剂相比,其作用效果还存在着一定的差距。主要原因有:拮抗酵母在实际应用过程中与实验室研究效果差异较大;受到病原菌浓度、果实生理状态、储存环境等因素的影响导致稳定性不佳;货架期短,长期有效制剂配方开发困难;生产成本偏高等。这些问题导致拮抗酵母的大规模商业化应用进展缓慢。因此,通过安全有效的方法提高拮抗酵母的生防效力是解决酵母商业化问题的有效途径。
目前,提高拮抗酵母生防效力的方法主要有:与有机或无机化学物质结合,与热处理、气调、紫外照射等物理处理相结合,以及通过甜菜碱、维生素c、海藻糖、几丁质、适当逆境胁迫等提高拮抗酵母自身的抗逆性以最大限度地提高酵母的生防效力。
技术实现要素:
本发明的目的之一在于提供一种能够提高桔梅奇酵母的普切明酸生成能力的保鲜剂,目的之二在于提供一种能够提高桔梅奇酵母的普切明酸生成能力的方法,目的之三在于提供一种能够提高桔梅奇酵母对果实采后病害的生物防治效力的保鲜剂,目的之四在于提供一种能够提高桔梅奇酵母对果实采后病害的生物防治效力的方法,效果明确、显著。
经研究,本发明提供如下技术方案:
1.精氨酸在制备提高桔梅奇酵母(metschnikowiacitriensis)的普切明酸生成能力的保鲜剂中的应用。
2.利用精氨酸提高桔梅奇酵母(metschnikowiacitriensis)的普切明酸生成能力的方法:将桔梅奇酵母接种至含有1-10mmol·l-1精氨酸的培养基中进行培养。
更佳的,将桔梅奇酵母接种至含有5mmol·l-1精氨酸的培养基中进行培养。
3.精氨酸在制备提高桔梅奇酵母(metschnikowiacitriensis)对果实采后病害的生物防治效力的保鲜剂中的应用。
进一步,所述果实为柑橘果实。
更进一步,所述病害为酸腐病。
4.利用精氨酸提高桔梅奇酵母(metschnikowiacitriensis)对果实采后病害的生物防治效力的方法:将桔梅奇酵母接种至含有1-10mmol·l-1精氨酸的培养基中进行培养。
更佳的,将桔梅奇酵母接种至含有5mmol·l-1精氨酸的培养基中进行培养。
普切明酸(pulcherriminicacid,pa)是由cyclo(l-leu-l-leu)经氧化作用形成的环二肽,而cyclo(l-leu-l-leu)是由胞内已经活化的两分子的亮氨酸氨酰leucyl-trna经环二肽酶催化而得。桔梅奇酵母m.citriensis在单独使用时,其拮抗效力不能达到化学杀菌剂的水平。发明人的前期研究结果表明,普切明酸作为m.citriensis的次生代谢产物,其固定fe3+导致不能利用螯合铁的病原菌因铁缺乏而不能正常生长是m.citriensis拮抗果实采后病害的重要作用机制。本发明的研究结果显示,精氨酸能促进m.citriensis的生长,提高m.citriensis的普切明酸生成能力,可提高m.citriensis在果实(例如柑橘果实)伤口处的种群密度,增强对果实(例如柑橘果实)采后病害的生物防治效力,控制采后由病原菌(例如酸腐菌)引起的果实腐烂,即精氨酸可以通过提高m.citriensis的普切明酸生成能力增强m.citriensis对果实采后病害的生物防治效力,其中经5mmol·l-1精氨酸处理的m.citriensis效果最佳。
本发明的有益效果在于:本发明提供了精氨酸在制备提高桔梅奇酵母的普切明酸生成能力的保鲜剂中的应用,以及利用精氨酸提高桔梅奇酵母的普切明酸生成能力的方法,还提供了精氨酸在制备提高桔梅奇酵母对果实采后病害的生物防治效力的保鲜剂中的应用,以及利用精氨酸提高桔梅奇酵母对果实采后病害的生物防治效力的方法,效果明确、显著,能有效解决农药使用带来的农业残留和环境污染问题,在果实采后病害的生物防治方面具有良好的应用潜力,同时也为研发微生物农药和增强其效力提供了优良的方法。
附图说明
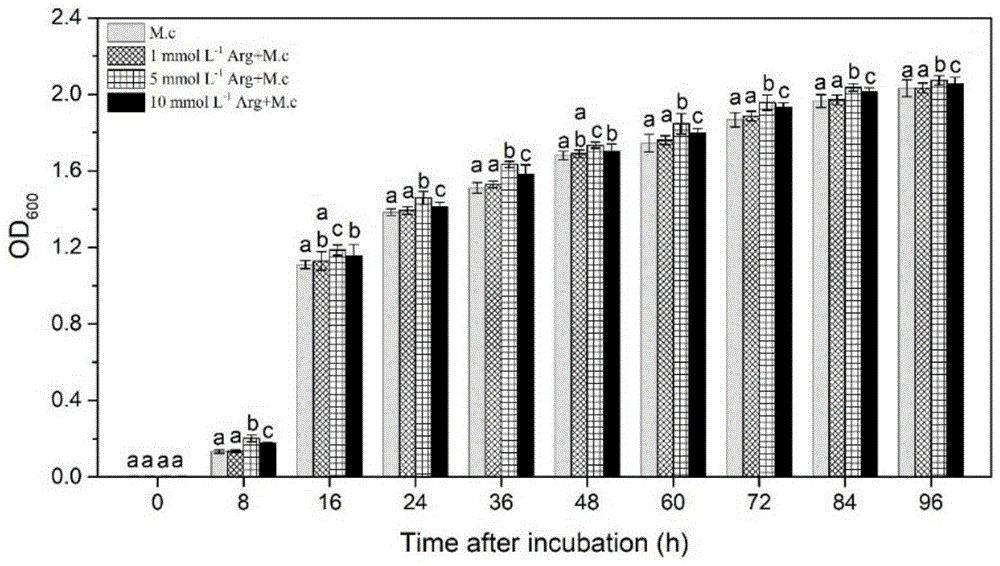
图1为精氨酸对m.citriensis生长的影响。
图2为精氨酸对m.citriensis普切明酸生成量的影响。
图3为常温(25℃)条件下各组酵母在果实伤口处的生长动态。
图4为低温(4℃)条件下各组酵母在果实伤口处的生长动态。
图5为精氨酸处理对m.citriensis防控柑橘果实在25℃贮藏3-9天时的酸腐病发病率的影响。
图6为精氨酸处理对m.citriensis防控柑橘果实在25℃贮藏3-9天时的酸腐病病斑直径的影响。
图7为各组果实在25℃贮藏9天后的酸腐病发病症状。
上述图1至图7中,“m.c”表示未经精氨酸处理的酵母(1×106cellsml-1),“1mmoll-1arg+m.c”表示1mmoll-1精氨酸处理过的酵母(1×106cellsml-1),“5mmoll-1arg+m.c”表示5mmoll-1精氨酸处理过的酵母(1×106cellsml-1),“10mmoll-1arg+m.c”表示10mmoll-1精氨酸处理过的酵母(1×106cellsml-1),control表示对照;柱和竖线分别代表生物学重复的均值和标准偏差,同一时间点不同字母表示差异显著(p<0.05)。
具体实施方式
为了使本发明的目的、技术方案和有益效果更加清楚,下面将对本发明的优选实施例进行详细的描述。
一、实验方法
1.不同氨基酸对m.citriensis普切明色素生成的影响
以含有不同氨基酸浓度(0、1、5、10、100mmol·l-1)的pda平板测试氨基酸对m.citriensis产普切明色素的影响。固定含35mg·l-1fecl3的pda培养基在每个平板上的体积为10ml。待培养基充分凝固后,取20μl1×108cells·ml-1的m.citriensis菌悬液均匀涂布在pda平板的中轴线上,涂布范围为60mm×2.6mm。将pda平板置28℃培养3d后测量普切明色素圈宽度。共测试20种氨基酸:丙氨酸(ala),缬氨酸(val),亮氨酸(leu),异亮氨酸(ile),苯丙氨酸(phe),色氨酸(trp),甲硫氨酸(met),脯氨酸(pro),甘氨酸(gly),丝氨酸(ser),苏氨酸(thr),半胱氨酸(cys),酪氨酸(tyr),天冬酰胺(asn),谷氨酰胺(gln),组氨酸(his),赖氨酸(lys),精氨酸(arg),天冬氨酸(asp)和谷氨酸(glu)。固定含不同氨基酸浓度的pda培养基在每个平板中的体积为10ml。取20μl1×108cells·ml-1的m.citriensis胞悬液均匀涂布在pda平板的中轴线上,涂布范围为60mm×2.6mm,放置10min后,将pda平板置28℃培养3d后测量普切明色素圈宽度。每组包含3个平行,每平行包含5个平板。
2.精氨酸处理对m.citriensis生长和普切明酸生成量的影响
将m.citriensis的胞悬液(1×108cells·ml-1)以2‰的接种量分别接种于含有0mmoll-1、1mmoll-1、5mmoll-1、10mmoll-1精氨酸的50mlyepd培养基中,28℃、200rpm培养。每隔8h或者12h取发酵液进行以下测定:
(1)生物量的测定:
取10ml发酵液,8000×g离心5min,弃去上清,沉淀(含有普切明和菌体)用无菌水洗涤2次,重悬于0.1mmoll-1naoh中以溶解普切明,10000×g离心2min,除去含有普切明酸的上清液,沉淀用生理盐水重悬,测定细胞密度od600。
(2)普切明酸含量的测定:
取10ml发酵液,8000×g离心5min,弃去上清,沉淀(含有普切明和菌体)用无菌水洗涤2次,加入10ml2moll-1naoh重悬,10000×g离心5min,取全部上清(含普切明酸),用紫外可见分光光度计测定od410,用于表征普切明酸的含量。
3.精氨酸处理对m.citriensis在柑橘果实伤口处的生长动态的影响
将m.citriensis分别接种于含有0mmoll-1、1mmoll-1、5mmoll-1、10mmoll-1精氨酸的100mlyepd培养基中使其细胞终浓度为2×105cellsml-1,28℃、200rpm培养16h达到对数中期后,8000×g离心5min,收集酵母细胞,无菌水洗涤2次后,用血球计数板计数,调整各组酵母胞悬液的浓度为1×106cellsml-1。
柑橘果实经表面消毒处理后,用灭菌枪头(1ml)在果实赤道部位等距离刺4个孔(深3mm,直径3mm)。在每个伤口处加入20μl以下液体:m.c组加入0mmoll-1精氨酸处理过的酵母胞悬液(1×106cells·ml-1);1mmoll-1arg+m.c组加入1mmoll-1精氨酸处理过的酵母胞悬液(1×106cells·ml-1);5mmoll-1arg+m.c组加入5mmoll-1精氨酸处理过的酵母胞悬液(1×106cells·ml-1);10mmoll-1arg+m.c组加入10mmoll-1精氨酸处理过的酵母胞悬液(1×106cells·ml-1)。静置30min后,将果实单果包装,分别放置于25℃(常温)和4℃(低温)。以接种后1h测定的酵母菌数为起始值。25℃下每2d取一次样,4℃下每5d取一次样。取样时用消毒的打孔器取伤口处直径为1cm的果皮组织,放入含10mlpbs(50mmol·l-1,ph6.8)的消毒研钵内研磨至匀浆。用稀释平板法计数,将样品匀浆液梯度稀释后涂布在ypd+fe平板上,每个浓度涂布3个平板,28℃下培养48h后计数。每组包含3个平行,每平行包含3个果实,结果以每个伤口处酵母活菌数量来表示,单位为log10cfu·wound-1。
4.精氨酸处理对m.citriensis生防效力的影响
将m.citriensis分别接种于含有0mmol·l-1、1mmol·l-1、5mmol·l-1、10mmol·l-1精氨酸的100mlyepd培养基中使其细胞终浓度为2×105cells·ml-1,28℃、200rpm培养16h达到对数中期后,8000×g离心5min,收集酵母细胞,无菌水洗涤2次后,用血球计数板计数,调整各组酵母胞悬液的浓度为1×108cellsml-1。
柑橘果实经表面消毒处理后,用灭菌枪头(1ml)在果实赤道部位等距离刺2个孔(深3mm,直径3mm)。在每个伤口处加入20μl以下液体:control组加入无菌水;m.c组加入0mmoll-1精氨酸处理过的酵母胞悬液(1×108cells·ml-1);1mmoll-1arg+m.c组加入1mmoll-1精氨酸处理过的酵母胞悬液(1×108cells·ml-1);5mmoll-1arg+m.c组加入5mmoll-1精氨酸处理过的酵母胞悬液(1×108cells·ml-1);10mmoll-1arg+m.c组加入10mmoll-1精氨酸处理过的酵母胞悬液(1×108cells·ml-1);室温放置4h后再接种10μl酸腐菌孢子悬浮液(1×106spores·ml-1)。每组包含3个平行,每平行包含10个果实。待菌悬液被果实充分吸收后用pe膜单果包装,贮藏在25℃、相对湿度85%~90%的环境下,并用pe膜覆盖保持湿度。每隔1d统计发病率和病斑直径。果实发病率(di,diseaseincidence)和病斑直径(ds,lesiondiameter)的计算公式如下:
二、实验结果
1.不同氨基酸对m.citriensis普切明色素生成的影响
不同氨基酸处理对m.citriensis普切明色素带形成大小的影响见表1。由表1可知,不同氨基酸对m.citriensis产普切明色素的影响不同,除了甲硫氨酸(met)之外,外源氨基酸处理均能诱导m.citriensis产普切明色素,但精氨酸诱导m.citriensis产生更宽的普切明色素带。
表1不同氨基酸处理对m.citriensis普切明色素带形成大小的影响
注:结果以平均值±sd表示(n=9);—表示未统计,其中100mmol·l-1的半胱氨酸(cys)和酪氨酸(tyr)在pda培养基中不溶解,不予统计,含100mmol·l-1的组氨酸(his)、天冬氨酸(asp)和谷氨酸(glu)的pda培养基无法凝固,不予测试;同一种氨基酸不同浓度处理下的色素带宽度标注不同小写字母表示有显著差异(p<0.05)。
2.精氨酸处理对m.citriensis生长和普切明酸生成量的影响
精氨酸处理对m.citriensis生长的影响如图1所示。从图中可以看出,低浓度(1mmol·l-1)的精氨酸对m.citriensis的生长没有显著的影响,而5mmol·l-1和10mmol·l-1的精氨酸均能促进m.citriensis的生长,尤其是5mmol·l-1的精氨酸,促生长作用更加明显。
精氨酸处理对m.citriensis普切明酸生成量的影响如图2所示。从图中可以看出,在m.citriensis于yepd液体培养条件下,普切明酸的产量从24h开始快速增加;当培养基中含有1mmol·l-1的精氨酸时,普切明酸的产量从36h开始显著高于未经精氨酸处理的m.c组(p<0.05),而培养基中含有5mmol·l-1和10mmol·l-1的精氨酸时,普切明酸的产量从16h开始显著高于m.c组和1mmoll-1arg+m.c组(p<0.05);总体上,培养基中含有5mmol·l-1的精氨酸时,m.citriensis产普切明酸的量最高。
3.精氨酸处理对m.citriensis在柑橘果实伤口处的生长动态的影响
精氨酸处理和无处理的m.citriensis在果实伤口处的生长动态如图3和图4所示。整体上,无论在常温(25℃)还是低温(0℃)贮藏条件下,各组酵母在果实伤口处的动态变化均呈现先急速增殖再缓慢下降的趋势,经精氨酸处理的酵母在伤口处的活菌数总体上高于未经处理的酵母,且5mmol·l-1精氨酸处理的酵母在伤口处的活菌数高于1mmol·l-1和10mmol·l-1精氨酸处理的酵母。
常温贮藏条件下,经精氨酸处理的酵母在第6d种群密度达到最高值,分别为:7.71log10cfu·wound-1(1mmoll-1arg+m.c组),7.91log10cfu·wound-1(5mmoll-1arg+m.c组)和7.81log10cfu·wound-1(10mmoll-1arg+m.c组);而未经处理的酵母则在第4d种群密度达到最高值:7.63log10cfu·wound-1;之后各组酵母的种群密度开始缓慢下降,至14d贮藏结束时,各组酵母在果实伤口处的活菌数分别为:6.57log10cfu·wound-1(m.c组),6.75log10cfu·wound-1(1mmoll-1arg+m.c组),7.42log10cfu·wound-1(5mmoll-1arg+m.c组),7.00log10cfu·wound-1(10mmoll-1arg+m.c组),精氨酸各处理组的活菌数目显著高于未经精氨酸处理的m.c组(p<0.05)。
低温贮藏条件下,经精氨酸处理的酵母在第15d种群密度达到最高值,分别为:7.73log10cfu·wound-1(1mmoll-1arg+m.c组),7.85log10cfu·wound-1(5mmoll-1arg+m.c组)和7.77log10cfu·wound-1(10mmoll-1arg+m.c组);而未经处理的酵母则在第5d种群密度达到最高值:7.66log10cfu·wound-1;之后各组酵母的种群密度开始缓慢下降,至30d贮藏结束时,各组酵母在果实伤口处的活菌数分别为:6.44log10cfu·wound-1(m.c组),6.92log10cfu·wound-1(1mmoll-1arg+m.c组),7.30log10cfu·wound-1(5mmoll-1arg+m.c组),7.11log10cfu·wound-1(10mmoll-1arg+m.c组),精氨酸各处理组的活菌数目显著高于未经精氨酸处理的m.c组(p<0.05)。
4.精氨酸处理对m.citriensis生防效力的影响
精氨酸处理对m.citriensis生防效力的影响如图5至图7所示。从图中可以看出,与未接种酵母的对照(control)组果实相比,酵母各处理组果实的酸腐病发病率和病斑直径均显著降低(p<0.05),说明酵母各处理组均能显著控制柑橘果实采后酸腐病的发展;其中,从贮藏的第4d起,精氨酸处理的各酵母组果实的发病率和病斑直径显著低于未处理酵母组果实(p<0.05);在常温贮藏9d后,与未处理酵母组相比,经1mmol·l-1、5mmol·l-1、10mmol·l-1精氨酸处理的酵母组果实的发病率分别低了16.67%、20.00%、18.33%,病斑直径分别少了8.93mm、11.99mm、11.02mm,说明精氨酸处理能显著增强m.citriensis对柑橘酸腐病的生防效力,且5mmol·l-1精氨酸处理的m.citriensis的生防效力相对最优。
最后说明的是,以上优选实施例仅用以说明本发明的技术方案而非限制,尽管通过上述优选实施例已经对本发明进行了详细的描述,但本领域技术人员应当理解,可以在形式上和细节上对其作出各种各样的改变,而不偏离本发明权利要求书所限定的范围。
- 还没有人留言评论。精彩留言会获得点赞!