乳糖降低的乳制品粉的制作方法
1.本发明涉及干或粉状乳制品组合物领域。
背景技术:
2.通常,生产乳制品粉是为了更长时间地保存乳制品的营养。与液态奶相比,乳制品粉的另一个优点是它们的运输成本大大降低。
3.乳制品粉可以从乳或乳清生产,所述乳或乳清通过分批工艺而变得不含乳糖。在此工艺中,用乳糖酶处理乳或乳清,然后进行产品的浓缩和干燥。主要问题是在经处理的液体乳制品中存在高浓度的单糖,从而导致玻璃化转变温度下降。因此,当干燥条件不适合时,此产品将导致喷雾干燥器结垢[torres等人,2017,technological aspects of lactose-hydrolyzed milk powder,food research international101,45-53]。所需的更温和的喷雾干燥条件会显著降低干燥过程的生产率,并增加成本。此外,无乳糖的乳(或乳清)粉是高度吸湿的,从而导致如果包装时不格外小心,则会在储存期间结块。由于这些挑战性的技术问题,与普通乳粉相比,无乳糖乳制品粉仍然是一个小市场。
[0004]
ep0458358 b1(snow brand)描述了一种用于通过浓缩脱脂乳并将浓缩乳与β-半乳糖苷酶一起孵育,之后进行酶灭活和喷雾干燥来生产含有10-15重量%的低聚半乳糖(galacto-oligosaccharide,gos)的脱脂乳粉的方法。所生产的产品不是不含乳糖的。
[0005]
wo2013/182686(dupont)描述了一种具有转半乳糖基化活性的多肽,所述多肽用于生产含gos的酸奶。所生产的产品不是不含乳糖的。wo2013/182686不涉及乳制品粉。
[0006]
wo 2015/086746(dupont)描述了一种用于生产含有稳定量的gos的乳产品的方法,其中所述产品需要在生产后进行热处理以灭活酶。所生产的产品不是不含乳糖的。未描述乳制品粉。
[0007]
wo2018/210821(novozymes)描述了一种用于从包含至少20重量%乳糖的乳底物开始,并且使所述乳底物与具有转半乳糖基化活性的酶接触来生产乳产品的方法。所生产的产品不是不含乳糖的。
[0008]
wo2020/117548(dupont)描述了一种用于制备具有gos纤维的低乳糖基于乳的产物的方法,所述方法具有以下步骤:提供包含乳糖的基于乳的底物;用转半乳糖基化酶处理所述基于乳的底物以提供gos纤维和剩余的乳糖;灭活所述转半乳糖基化酶,使所述具有gos纤维的基于乳的底物与乳糖酶接触以降解剩余的乳糖,从而提供所述具有gos纤维的低乳糖基于乳的产物;以及灭活所述乳糖酶。
[0009]
现有技术方法的缺点是,在添加乳糖酶之前需要灭活所述转半乳糖基化酶,和/或所述转半乳糖基化酶和所述乳糖酶的连续使用是耗时的,和/或所获得的产品不含乳糖,和/或乳底物需要包含增加水平的乳糖,和/或现有技术方法是复杂的,和/或无乳糖粉末的喷雾干燥显示出技术问题,和/或所产生的gos被分解。
附图说明
[0010]
图1:乳糖降低/无乳糖乳粉的生产的示意图
[0011]
序列表
[0012]
seq id no:1列出了奇异芽孢杆菌(sporobolomyces singularis)的β-己糖基转移酶(bht)的蛋白质序列。此序列由22个氨基酸的信号序列和572个氨基酸的推导成熟蛋白序列组成。在http://www.uniprot.org/uniref/uniref90_q564n5中还列出了奇异芽孢杆菌的bht的氨基酸序列。此序列也称为bht-132或没有任何缺失的野生型多肽。
[0013]
seq id no:2列出了bht-134的序列,bht-134是源自seq id no:1的缺失突变体,并且所述突变在第18位处开始直至且包括第54位。
[0014]
seq id no:3列出了用于在黑曲霉(aspergillus niger)中表达seq id no:1的经密码子适应的dna序列。
[0015]
seq id no:4列出了用于在黑曲霉中表达seq id no:2的经密码子适应的dna序列。
[0016]
seq id no:5列出了葡糖淀粉酶glaa启动子的经修饰的翻译起始序列。
[0017]
seq id no:6列出了含有葡糖淀粉酶启动子的a.o.部分和bht-132编码基因的dna片段,所述bht-132编码基因包含ecori和paci限制性位点。
[0018]
seq id no:7列出了含有葡糖淀粉酶启动子的a.o.部分和bht-134编码基因的dna片段,所述bht-134编码基因包含ecori和paci限制性位点。
技术实现要素:
[0019]
本发明的目的是提供一种用于生产乳糖降低(优选地无乳糖)的乳制品粉的方法,所述方法例如不会显著降低干燥设备的生产率,即,本发明旨在与用于干燥不含(或几乎不含)gos的可比的乳糖降低的乳制品粉的相同干燥设备的能力和生产时间相比,增加干燥能力和/或减少生产时间。另外或替代地,本发明的目的是生产乳糖降低(优选地无乳糖)的乳制品粉,所述乳制品粉具有改善的特性,即与不含或几乎不含gos的可比的乳糖降低的乳制品粉的吸湿性和粘性相比,吸湿性更低和/或粘性更小。另外或替代地,本发明旨在减少乳糖降低(优选地无乳糖)的乳粉生产中的质量损失。另外或替代地,本发明的目的是减少在干燥或储存期间所形成的乳糖降低(优选地无乳糖)的乳粉的褐色形成。另外或替代地,本发明旨在改进,例如简化现有技术方法。
[0020]
本发明提供了
[0021]-一种用于生产乳糖降低的乳制品粉的方法,所述方法包括
[0022]
(i)用具有转半乳糖基酶活性的第一酶和具有乳糖酶活性的第二酶处理包含乳糖的基于乳的底物,以获得乳糖降低并且包含低聚半乳糖(gos)的基于乳的产物;
[0023]
(ii)从所述乳糖降低并且包含gos的基于乳的产物制备乳糖降低的乳制品粉,
[0024]-无乳糖乳制品粉,所述无乳糖乳制品粉包含至少4%(w/w)的gos,
[0025]-无乳糖乳,所述无乳糖乳包含基于总碳水化合物至少10%(w/w)的gos并且具有》1.2的葡萄糖/半乳糖比率,
[0026]-乳,所述乳基于总(游离/)碳水化合物包含《0.1%的乳糖、至少10%的gos并且具有》1.2的葡萄糖/半乳糖比率,
[0027]-gos用于改善乳糖降低的基于乳的底物的干燥的用途,
[0028]-具有转半乳糖基酶活性的酶和具有乳糖酶活性的酶用于改善无乳糖的基于乳的底物的干燥的用途,和
[0029]-具有转半乳糖基酶活性的酶和具有乳糖酶活性的酶用于改善无乳糖的乳粉的特性的用途。
具体实施方式
[0030]
在第一实施方式中,本发明提供了一种用于生产乳糖降低的乳制品粉的方法,所述方法包括
[0031]
(i)用具有转半乳糖基酶活性的第一酶和具有乳糖酶活性的第二酶处理包含乳糖的基于乳的底物,以获得乳糖降低并且包含低聚半乳糖(gos)的基于乳的产物;
[0032]
(ii)从所述乳糖降低并且包含gos的基于乳的产物制备乳糖降低的乳制品粉。
[0033]
优选地,本发明提供了一种用于生产无乳糖的乳制品粉的方法,所述方法包括
[0034]
(i)用具有转半乳糖基酶活性的第一酶和具有乳糖酶活性的第二酶处理包含乳糖的基于乳的底物,以获得无乳糖并且包含低聚半乳糖(gos)的基于乳的产物;
[0035]
(ii)从所述无乳糖并且包含gos的基于乳的产物制备无乳糖的乳制品粉。
[0036]
短语“乳糖降低的乳制品粉”和“具有降低的乳糖的量的乳制品粉”在本文中可互换使用。
[0037]
短语“无乳糖的乳制品粉”和“不含乳糖的乳制品粉”在本文中可互换使用。
[0038]
短语“包含乳糖的基于乳的底物”和“含乳糖的基于乳的底物”在本文中可互换使用。
[0039]
短语“乳糖降低、包含低聚半乳糖(gos)的基于乳的产物”和“乳糖降低并且包含低聚半乳糖(gos)的基于乳的产物”以及“具有降低的乳糖的量并且包含低聚半乳糖(gos)的基于乳的产物”在本文中可互换使用。
[0040]
如本文所用的术语“乳制品粉”是指从基于乳的产物制备的干粉。乳制品粉可以是乳粉或乳清粉。乳粉的示例是脱脂乳粉、半脱脂乳粉、全脂乳粉、酪乳粉、乳蛋白浓缩物(milk protein concentrate,mpc)粉,并且乳清粉的示例是甜乳清粉或酸性乳清粉、乳清蛋白浓缩物(whey protein concentrate,wpc)粉和乳清渗透物粉(whey permeate powder)。
[0041]
优选地,乳制品粉是乳粉,更优选地,乳制品粉是无乳糖的乳粉,因此本发明提供了一种用于生产无乳糖的乳粉的方法,所述方法包括
[0042]
(i)用具有转半乳糖基酶活性的第一酶和具有乳糖酶活性的第二酶处理包含乳糖的基于乳的底物,以获得无乳糖并且包含低聚半乳糖(gos)的基于乳的产物;
[0043]
(ii)从所述无乳糖并且包含gos的基于乳的产物制备无乳糖的乳粉。
[0044]
如本文所用的术语“乳”是指从任何哺乳动物如奶牛、绵羊、山羊、骆驼或水牛获得的乳汁分泌物。优选地,乳是牛乳。
[0045]
如本文所用的术语“基于乳的底物”指任何原始的和/或经加工的乳材料。基于乳的底物可已经经过本领域中已知的处理例如—如—均质化、巴氏灭菌、杀菌或延长保质期(extended shelf life,esl)处理或超热处理(ultra-heat treatment,uht)。
[0046]
基于乳的底物可以是脱脂乳、全脂乳、半脱脂乳、炼乳、复原(脱脂)乳、酪乳、乳蛋白浓缩物(mpc),或乳清,例如甜乳清或酸性乳清、乳清蛋白浓缩物(wpc)、乳清蛋白分离物(wpi)或乳清渗透物。
[0047]
优选地,基于乳的底物可以是脱脂乳、全脂乳、半脱脂乳、炼乳、复原(脱脂)乳、酪乳或乳蛋白浓缩物(mpc)。
[0048]
基于乳的底物包含乳糖,并且因此所述基于乳的底物称为“包含乳糖的基于乳的底物”。通常,牛乳包含约5%的乳糖。优选地,基于乳的底物仅包含天然存在于所述基于乳的底物中的乳糖。优选地,基于乳的底物不包含添加的乳糖。优选地,基于乳的底物包含在4(w/v)%乳糖至10(w/v)%乳糖,优选地4(w/v)%乳糖至8(w/v)%乳糖,甚至更优选地4(w/v)%乳糖至6(w/v)%乳糖或4.5(w/v)%乳糖至5.5(w/v)%乳糖范围内的乳糖。
[0049]
基于乳的底物可以是浓缩的。如图1所示,乳制品粉的制备通常包括浓缩底物的蒸发步骤。如稍后将解释的,在本发明的方法中所使用的酶可以在乳制品粉生产期间的不同时间点添加,并且可以例如在蒸发期间或之后添加,在这种情况下基于乳的底物被浓缩。优选地,浓缩基于乳的底物,使得乳糖的量在4(w/v)%乳糖至10(w/v)%乳糖,优选地4(w/v)%乳糖至8(w/v)%乳糖,甚至更优选地4(w/v)%乳糖至6(w/v)%乳糖或4.5(w/v)%乳糖至5.5(w/v)%乳糖的范围内。
[0050]
将包含乳糖的基于乳的底物与具有转半乳糖基酶活性的第一酶(即从乳糖产生gos的酶)一起孵育。
[0051]
即gos是原位产生的,这意味着gos是在基于乳的底物中由所述基于乳的底物中存在的乳糖产生的。优选地,根据本发明的方法、用途或产品不包含添加的gos(即非原位产生的gos)。
[0052]
β-半乳糖苷酶是水解β-d-半乳糖苷中的末端非还原性β-d-半乳糖残基的酶,例如将乳糖水解成半乳糖和葡萄糖。这些酶属于酶ec3.2.1.23类。除了水解,此酶类还能够将半乳糖转移到其他糖上,并由此产生低聚半乳糖(gos)。ec 3.2.1.23类的不同酶对水解(β-半乳糖苷酶)活性和转半乳糖基酶活性具有不同的偏好,并且所述偏好可以例如通过转半乳糖基化活性与水解活性的比率来表示。
[0053]
如在本发明的方法中所使用的第一酶是相较于水解更偏好于转半乳糖基化的。所述第一酶主要具有转半乳糖基化活性。
[0054]
具有转半乳糖基酶活性的酶优选属于ec 3.2.1.23亚类。
[0055]
如本文所用,术语“具有转半乳糖基酶活性的酶”、“具有转半乳糖基化活性的酶”、“转半乳糖基化酶”和“转半乳糖基酶”在本文中可互换使用,并且都指能够将半乳糖从乳糖转移到例如乳糖、d-半乳糖(gal)或d-葡萄糖(glu)的羟基,由此产生低聚半乳糖的酶。
[0056]
允许第一酶的活性(即导致gos产生的酶促活性)有足够的时间来产生一定水平的gos分子。确切的时间将取决于所使用的酶、所使用的酶的量,而且还取决于所使用的温度和基于乳的底物中的乳糖浓度。技术人员能够确定酶剂量、孵育温度和孵育时间,以允许在基于乳的底物中产生gos。
[0057]
将包含乳糖的基于乳的底物与具有转半乳糖基酶活性的第一酶一起孵育,以便形成足够的gos,即基于总碳水化合物至少10%的gos。
[0058]
具有转半乳糖基酶活性的第一酶的合适的商业示例是zymstartmgos,或来自
dupont的nuricatm或来自amano的biolacta fn5。在科学文献和/或专利文献中描述了许多其他具有转半乳糖基酶活性的酶。已经从包括细菌、真菌和酵母在内的多种微生物中分离出了能够进行转半乳糖基化的酶(torres等人(2010)comprehensive reviews in food science and food safety,9:438-454)。乳糖的gos产量可取决于酶的来源、乳糖浓度、ph、酶剂量、孵育的时间和温度而变化。
[0059]
具有转半乳糖基酶活性的酶的另一个合适的示例是奇异芽孢杆菌己糖基转移酶、环状芽孢杆菌(bacillus circulans)β-半乳糖苷酶或双歧双歧杆菌(bifidobacterium bifidum)β-半乳糖苷酶。
[0060]
此外,通过使用wo2018/210820中所述的方法,可以对主要是水解性的β-半乳糖苷酶进行修饰,使其表现得更像转半乳糖基化酶。
[0061]
最合适的是产生不容易被具有乳糖酶活性的酶降解的gos的具有转半乳糖基酶活性的酶,例如作为奇异芽孢杆菌己糖转移酶的具有转半乳糖基酶活性的酶。
[0062]
本发明的方法中所使用的第二酶也属于ec 3.2.1.23亚类,并且也是β-半乳糖苷酶,但对乳糖酶活性或水解活性有偏好。
[0063]
允许第二酶的活性(即将乳糖水解为半乳糖和葡萄糖)有足够的时间来减少乳糖的量。优选地,第二酶的活性导致乳糖降低,更优选地不含乳糖的基于乳的产物。确切的时间将取决于所使用的确切的酶、所使用的酶的量以及所使用的温度。技术人员能够确定酶剂量、孵育温度和孵育时间,以使基于乳的底物中的乳糖水解为半乳糖和葡萄糖。
[0064]
对乳糖具有水解活性的酶的合适的商业示例是maxilact(dsm)、lactozyme(novozymes)、saphera(novozymes)、nolafit(chr.hansen)、ha-lactase(chr.hansen)、godo ynl-2(dupont)和一些其他被描述为乳糖酶的酶。科学文献和/或专利出版物中已经描述了其他合适的乳糖酶。
[0065]
最优选地,具有乳糖酶活性的酶偏好水解乳糖,而由具有转半乳糖基酶活性的第一酶产生的gos保持完整。
[0066]
如上所述,β-半乳糖苷酶可以具有转半乳糖基酶活性以及乳糖酶活性。主要活性取决于例如基于乳的底物中的乳糖浓度。在本文中,具有转半乳糖基酶活性的酶是(在所用条件下)以产生gos为其主要活性(但可具有次要乳糖酶活性)的酶。具有乳糖酶活性的酶是(在所用条件下)以将乳糖水解为葡萄糖和半乳糖为主要活性(但可具有导致gos产生的次要活性)的酶。换句话说,第一酶主要具有转半乳糖基酶活性,第二酶主要具有乳糖酶活性。技术人员能够选择正确的第一酶和第二酶。上文以及实施例中提供了合适的示例。
[0067]
第一酶和第二酶在孵育后通过本领域中已知的导致酶活性降低的任何方法灭活。合适的灭活方法的示例是热处理,例如巴氏灭菌,或者在喷雾干燥之前在基于乳的底物的蒸发和浓缩期间进行热处理。优选地,酶是完全灭活的(剩余《1%的初始活性)。第一酶的灭活可以在与第二酶一起孵育之前进行,但是优选在孵育之后与第二酶同时进行。
[0068]
所用的酶可以作为液体或固体(颗粒)添加到基于乳的底物中。
[0069]
第一酶和第二酶的联合作用导致乳糖降低(优选地无乳糖)并且包含gos的基于乳的产物,所述基于乳的产物可以进一步加工成乳糖降低(优选地无乳糖)的乳制品粉。
[0070]
gos可以是二糖(聚合度(dp)2,即dp2)(不是gos的乳糖除外)、三糖(dp3)、四糖(dp4)、五糖(dp5)或更长的寡糖。
[0071]
二糖的示例是半乳糖基-半乳糖(gal-gal)或异乳糖(半乳糖基-β1-6-葡萄糖)。三糖(dp3)的示例是半乳糖基-半乳糖基-乳糖(gal-gal-glu)。四糖(dp4)的示例是半乳糖基-半乳糖基-半乳糖基-乳糖(gal-gal-gal-glu),五糖(dp5)的示例是半乳糖基-半乳糖基-半乳糖基-半乳糖基-乳糖(gal-gal-gal-gal-glu)。甚至更长的寡糖的示例是(半乳糖)n-乳糖,其中n》3。
[0072]
术语“dpx+gos”意指具有dpx或更高dp的gos分子的总和。例如,术语“dp3+gos”意指具有dp3、dp4、dp5等的gos分子的总和。
[0073]
如本文所用的术语gos是指dp2+gos的组合。dp2+gos不包括乳糖(根据定义,乳糖不是gos)。
[0074]
gos分子的水平可以通过使用不同的分析技术来确定。
[0075]
例如,可以确定包含乳糖的基于乳的底物中乳糖、葡萄糖和半乳糖的量,并且在与酶一起孵育后,使用酶促分析、nmr或hpaec-pad确定乳糖、葡萄糖和半乳糖的量。单糖、二糖的定量也可以通过使用具有脉冲安培检测(hpaec-pad)分析的高性能阴离子交换色谱来进行。
[0076]
在另一个替代方法中,gos产生可以通过酶对乳糖的反应来建立,其中在给定时间内产生的半乳糖的量少于产生的葡萄糖的量。
[0077]
然后可以通过从(存在于底物中的)乳糖、葡萄糖和半乳糖的起始量的总和中减去(存在于产物中的)乳糖、葡萄糖和半乳糖的总量来确定gos的量。差值就是gos的量。在下式中:
[0078]
[gos]
产物
=([lac]+[glu]+[gal])
底物-([lac]+[glu]+[gla])
产物
[0079]
可以根据本发明的方法用该式确定gos的水平,因为技术人员有机会确定基于乳的底物中的乳糖水平。显然,如果无法确定基于乳的底物中的乳糖水平,就不能使用这种确定/式,例如,如果只提供了乳糖降低的乳制品粉(优选无乳糖的乳粉),则就是这种情况。稍后将提供另一种确定乳糖降低的乳制品粉(优选无乳糖的乳粉)中的gos水平的方法。
[0080]
使用本领域中众所周知的技术,进行从所述乳糖降低(优选无乳糖)的、包含gos的基于乳的产物制备乳糖降低(优选无乳糖)的乳制品粉的步骤。
[0081]
通常,所述乳制品粉是通过蒸发和干燥所生产的乳糖降低的、包含gos的基于乳的产物至干燥而制备的。干燥通常是通过喷雾干燥、转鼓干燥或冷冻干燥进行的。优选地,乳制品粉是通过喷雾干燥获得的。
[0082]
图1中提供了用于生产乳糖降低的乳制品粉的合适方案的示例。优选地,当乳冷藏储存时添加酶(同时或顺序地,优选顺序地)。这种储存通常已经存在于乳制品粉工厂中,并且不需要附加的设备或对乳制品粉生产过程的改变。或者,在蒸发期间或之后添加酶。
[0083]
如上所述,本发明在第一实施方式中提供了一种用于生产乳糖降低的乳制品粉的方法,所述方法包括
[0084]
(i)用具有转半乳糖基酶活性的第一酶和具有乳糖酶活性的第二酶处理包含乳糖的基于乳的底物,以获得乳糖降低并且包含低聚半乳糖(gos)的基于乳的产物;
[0085]
(ii)从所述乳糖降低并且包含gos的基于乳的产物制备乳糖降低的乳制品粉。
[0086]
本专利申请的发明人已经注意到了不同的令人惊讶的结果(当与无乳糖的乳中不存在、几乎不存在或存在低水平的gos的情况相比时),例如升高的玻璃化转变温度、提高的
(喷雾)干燥器的干燥能力、降低的(喷雾)干燥器能力损失和降低的(喷雾)干燥成本。
[0087]
因此,本发明替代地提供了
[0088]-一种用于改善(提高)用于乳糖降低(优选地无乳糖)的乳粉的(喷雾)干燥器的干燥能力的方法
[0089]-一种用于改善(减少)乳糖降低(优选地无乳糖)的乳粉的干燥时间的方法
[0090]-一种用于改善(提高)乳糖降低(优选地无乳糖)的乳粉的干燥速度的方法
[0091]-一种用于改善(降低)乳糖降低(优选地无乳糖)的乳粉的吸湿性的方法
[0092]-一种用于改善(降低)乳糖降低(优选地无乳糖)的乳粉的粘性的方法
[0093]-一种用于改善(减少)乳糖降低(优选地无乳糖)的乳粉的褐色形成的方法
[0094]
其中所述方法中的任何方法包括
[0095]
(i)用具有转半乳糖基酶活性的第一酶和具有乳糖酶活性的第二酶处理包含乳糖的基于乳的底物,以获得乳糖降低并且包含低聚半乳糖(gos)的基于乳的产物;
[0096]
(ii)从所述乳糖降低并且包含gos的基于乳的产物制备乳糖降低的乳制品粉。
[0097]
另一个不同的令人惊讶的结果是,可以制备包含至少10%(w/w)的gos(基于总碳水化合物)并且不含乳糖的基于乳的产物。因此,本发明还提供了一种用于制备包含至少10%(w/w)的gos(基于总碳水化合物)并且不含乳糖的基于乳的产物的方法,所述方法包括用具有转半乳糖基酶活性的第一酶和具有乳糖酶活性的第二酶处理包含乳糖的基于乳的底物,以获得无乳糖并且包含低聚半乳糖(gos)的基于乳的产物。
[0098]
已经提供的定义也适用于这些实施方式。
[0099]
通过所有上述方法获得的(作为中间产物的)(乳糖降低并且包含gos的)基于乳的产物包含基于总碳水化合物至少10%的gos。基于乳的底物中的术语“总碳水化合物”定义为葡萄糖、半乳糖、乳糖和gos的浓度的总和。
[0100]
如本文所用,术语“乳糖降低并且包含gos的基于乳的产物”是指乳糖降低至0.5(w/v)%或0.1(w/v)%。术语“无乳糖、包含gos的基于乳的产物”用于指乳糖降低至《0.01%(w/v)(《0.1g/l)。
[0101]
术语无乳糖的乳制品粉(优选地无乳糖的乳粉)是指乳糖降低至《0.1%(w/w)(《1g/kg)。
[0102]
因此,本发明提供了一种用于生产乳糖降低(优选地无乳糖)的乳制品粉的方法,所述方法包括
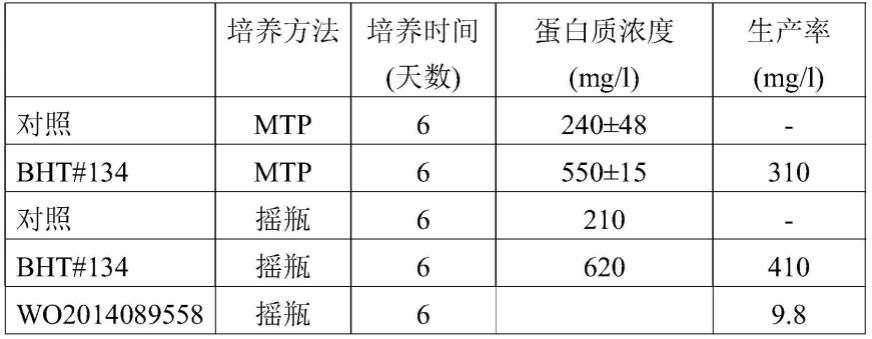
[0103]
(i)用具有转半乳糖基酶活性的第一酶和具有乳糖酶活性的第二酶处理包含乳糖的基于乳的底物,以获得乳糖降低(优选地无乳糖)并且包含低聚半乳糖(gos)的基于乳的产物;
[0104]
(ii)从所述乳糖降低(优选地无乳糖)并且包含gos的基于乳的产物制备乳糖降低(优选地无乳糖)的乳制品粉,
[0105]
其中所述基于乳的产物中的乳糖降低至0.5(w/v)%乳糖,优选地0.1(w/v)%乳糖,或者
[0106]
其中所述基于乳的产物中的乳糖降低至《0.01(w/v)%乳糖,并且其中所述乳糖降低、包含gos的基于乳的产物是无乳糖、包含gos的基于乳的产物。
[0107]
因此,本发明提供了一种用于生产无乳糖的乳制品(优选地乳)粉的方法,所述方
法包括
[0108]
(i)用具有转半乳糖基酶活性的第一酶和具有乳糖酶活性的第二酶处理包含乳糖的基于乳的底物,以获得无乳糖并且包含低聚半乳糖(gos)的基于乳的产物;
[0109]
(ii)从所述无乳糖并且包含gos的基于乳的产物制备无乳糖的乳制品(优选地乳)粉,
[0110]
其中所述无乳糖、包含gos的基于乳的产物中的乳糖降低至《0.01(w/v)%乳糖。
[0111]
由于干燥的浓缩效应,当与这些组分在所获得的乳糖降低(优选地无乳糖)、包含gos的基于乳的产物中的水平相比时,相应粉末中的gos和乳糖水平将不同。
[0112]
乳制品粉中的gos水平为至少4%(w/w)。优选地,乳制品粉中gos的水平为至少5%(w/w)、6%(w/w)、7%(w/w)、8%(w/w)、9%(w/w)或至少10%(w/w)。更优选地,乳制品粉中gos的水平为至少15%(w/w)。乳制品粉中gos的水平不仅取决于所用的特定酶,而且还取决于例如基于乳的底物中的脂肪百分比。
[0113]
乳糖降低的乳制品粉中乳糖的水平/量《5%(w/w),优选地《1%(w/w)。无乳糖乳制品(优选地乳)粉中的乳糖的水平为《0.1%乳糖。
[0114]
上文已经提供了具有转半乳糖基酶活性的第一酶的示例。具有转半乳糖基酶活性的酶的另一个示例是奇异芽孢杆菌己糖基转移酶。
[0115]
已知几种担子菌纲的酵母具有有效的转半乳糖基化活性,并且已经报道了由奇异芽孢杆菌(以前称为布勒掷孢酵母(bullera singularis))、罗伦隐球酵母(cryptococcus laurentii)、艾尔维梗孢酵母(sterigmatomyces eleviae)、小红酵母(rhodotorula minuta)和大链担耳(sirobasidium magnum)产生gos。主要困难是生产足量的这些己糖基转移酶,从而使得使用这些酶抑制性的gos的商业化生产昂贵。生产这些酶的障碍是它们的细胞附着和不良分泌,需要细胞壁提取和酶的广泛纯化,以及不良产率(例如ishikawa等人(2005)journal of bioscience and bioengineering,4:331-339)。
[0116]
为了缓解这些产生gos的酶的生产问题,确定了来自奇异芽孢杆菌的β-己糖基转移酶(bht)的基因序列(ishikawa等人(2005))。为了过表达bht,将所述基因克隆到不同的大肠杆菌(escherichia coli)菌株中,但是发现所有的酶都是不溶的,即使当其作为融合蛋白表达以增强溶解性时也是如此(dagher等人(2013)applied and environmental microbiology,79:1241-1249)。然而,在巴斯德毕赤酵母(pichia pastoris)中与α-交配因子信号序列(αmf)符合读框地表达相同基因确实导致了少量bht的分泌。使用构建体发现每升培养物最高分泌9.8mg的bht,在所述构建体中缺失了bht序列的第1-22位处的潜在分泌信号并且使用αmf信号序列代替,并且将his标签连接至bht序列的c末端(wo2014/089558a1)。wo2014/089558中的结论是“αmf和bht前导序列分泌信号都不能完全完成rbht-his的分泌,这可与氨基酸177-199之间预测的跨膜区的存在有关”。
[0117]
本技术的发明人已经能够使用乳酸克鲁维酵母(k.lactis)和黑曲霉(a.niger)作为生产宿主,从奇异芽孢杆菌生产足量的β-己糖基转移酶。两种宿主都从奇异芽孢杆菌产生了足量的己糖基转移酶。优选地,所用的来自奇异芽孢杆菌的己糖基转移酶是野生型序列的截短变体。本文在实验部分内描述了合适变体的示例。
[0118]
优选地,所用的来自奇异芽孢杆菌的己糖基转移酶(在黑曲霉或乳酸克鲁维酵母中产生)与seq id no:1或seq id no:2具有至少80%序列同一性。出于本发明的目的,在本
文中限定,为了确定两个氨基酸序列或两个核酸序列的序列同一性百分比,对序列进行比对以实现最佳比较目的。为了优化两个序列之间的比对,可以在进行比较的两个序列中的任一序列中引入空位。此类比对可以在被比较的序列的全长上进行。在描述具体位置的情况下,这些位置也应该在比较中使用。序列同一性是两个序列之间在所报告的比对区域上相同匹配的百分比。
[0119]
可以使用数学算法来完成两个序列之间的序列比较和序列同一性百分比确定。本领域技术人员将意识到事实上几种不同的计算机程序可用于比对两个序列和确定两个序列之间的同一性(kruskal,j.b.(1983),在d.sankoff和j.b.kruskal(编辑),time warps,string edits and macromolecules:the theory and practice of sequence comparison,第1-44页,addison wesley中的an overview of sequence comparison)。两个氨基酸序列之间或两个核苷酸序列之间的序列同一性百分比可以使用用于比对两个序列的needleman和wunsch算法来确定。(needleman,s.b.和wunsch,c.d.(1970)j.mol.biol.48,443-453)。该氨基酸序列和核苷酸序列都可以通过算法来进行比对。已在计算机程序needle中实现了needleman-wunsch算法。出于本发明的目的,使用来自emboss程序包的needle程序(2.8.0版或更高版本,emboss:the european molecular biology open software suite(2000)rice,p.longden,i.和bleasby,a.trends in genetics 16,(6),第276-277页,http://emboss.bioinformatics.nl/)。对于蛋白质序列,使用eblosum62来用于取代矩阵。对于核苷酸序列,使用ednafull。所使用的任选参数是为10的空位开放罚分和为0.5的空位延伸罚分。技术人员将理解,当使用不同的算法时,所有这些不同的参数将产生略微不同的结果,但是两个序列的总体同一性百分比不会显著改变。
[0120]
如上所述通过程序needle进行比对后,查询序列与本发明序列之间的序列同一性百分比计算如下:比对中对应位置(其显示两个序列中的相同氨基酸或相同核苷酸)的数目除以减去比对中的空位总数后的比对总长度。如本文所定义的同一性可以通过使用nobrief选项从needle获得,并在程序的输出中标记为“最长同一性”。
[0121]
或者,所用的来自奇异芽孢杆菌的己糖基转移酶(在黑曲霉或乳酸克鲁维酵母中产生)与seq id no:1或seq id no:2具有至少85%、90%或95%、96%、97%、98%或99%序列同一性。
[0122]
合适的基于乳的底物的示例已经在上面提供,并且可以是脱脂乳、半脱脂乳、全脂乳、巴氏灭菌乳、uht乳、具有延长保质期的乳、炼乳、由粉末制备的复原(脱脂)乳、酪乳、乳蛋白浓缩物(mpc),或乳清例如甜乳清或酸性乳清、乳清蛋白浓缩物(wpc)或乳清渗透物。
[0123]
在本发明的各方面中的一个方面中,基于乳的底物中的乳糖水平尚未降低,例如基于乳的底物尚未经受乳糖降低过滤技术。在另一方面中,在与第一酶和第二酶一起孵育之前,基于乳的底物中的乳糖水平已经降低。
[0124]
本发明的方法可能导致并非所有人都喜欢的增加的甜味。通过使用众所周知的技术减少基于乳的底物中存在的部分乳糖,可以降低甜味的增加。即,本发明的方法可以包括在用如上所述的第一酶和第二酶处理所述底物之前降低基于乳的底物中的乳糖水平的步骤。
[0125]
在用如上所述的第一酶和第二酶处理基于乳的底物之前,可以增加基于乳的底物中的乳糖水平,例如通过浓缩。用于生产乳糖降低(优选地无乳糖)的乳制品(优选地乳)粉
的典型生产方法包括浓缩步骤。如图1所示的蒸发步骤是浓缩步骤,并且本发明的方法中所使用的酶可以在蒸发期间和/或之后添加。优选地,基于乳的底物中的乳糖的量在4(w/v)%乳糖至10(w/v)%乳糖,优选地4(w/v)%乳糖至8(w/v)%乳糖,甚至更优选地4(w/v)%乳糖至6(w/v)%乳糖或4.5(w/v)%乳糖至5.5(w/v)%乳糖的范围内。
[0126]
在上述方法中的任何方法中使用的第一酶和第二酶可以同时或顺序地添加。本发明的发明人注意到通过顺序地使用第一酶和第二酶获得了最佳结果,其中首先添加具有转半乳糖基酶活性的酶。因此,首先由乳糖产生一定量的gos,并将剩余的乳糖通过具有乳糖酶(水解)活性的第二酶水解成葡萄糖和半乳糖。
[0127]
因此,本发明提供了一种用于生产乳糖降低(优选地无乳糖)的乳制品粉(优选地乳粉)的方法,所述方法包括
[0128]
(i)用具有转半乳糖基酶活性的第一酶和具有乳糖酶活性的第二酶处理包含乳糖的基于乳的底物,以获得乳糖降低(优选地无乳糖)并且包含低聚半乳糖(gos)的基于乳的产物;
[0129]
(ii)从所述乳糖降低(优选地无乳糖)并且包含gos的基于乳的产物制备乳糖降低(优选地无乳糖)的乳制品粉(优选地乳粉),
[0130]
其中所述基于乳的底物首先用具有转半乳糖基酶活性的酶处理,随后用具有乳糖酶活性的酶处理,优选地所述方法不包括在用具有转半乳糖基酶活性的酶处理与用具有乳糖酶活性的酶处理之间的酶灭活步骤。
[0131]
技术人员知道一些具有乳糖酶活性的酶能够降低gos水平,尤其是在长时段孵育时。技术人员将采取预防措施来避免具有乳糖酶活性的酶引起的gos的(过度)减少。
[0132]
本发明的发明人注意到通过同时使用第一酶和第二酶获得了非常好的结果。因此,由具有转半乳糖基酶活性的第一酶从乳糖生产gos和由具有乳糖酶(水解)活性的第二酶将乳糖水解成葡萄糖和半乳糖是同时发生的。
[0133]
因此,本发明提供了一种用于生产乳糖降低(优选地无乳糖)的乳制品粉(优选地乳粉)的方法,所述方法包括
[0134]
(i)用具有转半乳糖基酶活性的第一酶和具有乳糖酶活性的第二酶处理包含乳糖的基于乳的底物,以获得乳糖降低(优选地无乳糖)并且包含低聚半乳糖(gos)的基于乳的产物;
[0135]
(ii)从所述乳糖降低(优选地无乳糖)并且包含gos的基于乳的产物制备乳糖降低(优选地无乳糖)的乳制品粉(优选地乳粉),
[0136]
其中步骤(i)是同时用具有转半乳糖基酶活性的第一酶和具有乳糖酶活性的第二酶处理包含乳糖的基于乳的底物,以获得乳糖降低并且包含低聚半乳糖(gos)的基于乳的产物。
[0137]
上面已经描述了乳制品粉的示例:乳制品粉是乳粉或乳清粉。乳粉的示例是脱脂乳粉、半脱脂乳粉、全脂乳粉、酪乳粉、mpc粉,并且乳清粉的示例是甜乳清粉或酸性乳清粉、wpc粉和乳清渗透粉。优选地,乳制品粉是乳粉或乳清粉。更优选地,乳制品粉是乳粉,并且最优选地,乳制品粉是无乳糖的乳粉。
[0138]
本发明还提供了一种乳制品粉,优选地可通过如上所述的方法获得的乳制品粉。
[0139]
优选的乳制品粉是包含至少4%(w/w)的gos的无乳糖的乳制品粉。
[0140]
无乳糖的粉末包含至少4%(w/w)的gos和《0.1%(w/w)的乳糖。因此,本发明提供了包含至少4%(w/w)的gos和《0.1%(w/w)的乳糖的乳制品粉(优选地乳粉)。
[0141]
在用酸性乳糖酶完全水解乳制品粉后,通过从单糖(葡萄糖和半乳糖)总量中减去乳制品粉中的乳糖、葡萄糖和半乳糖浓度,可以确定乳制品粉中的gos的量(基于aoac官方方法2001.02;在本文的实验部分有更详细的描述)。
[0142]
在另一个实施方式中,本发明提供了无乳糖的乳,所述无乳糖的乳包含基于总碳水化合物至少10%(w/w)的gos并且具有》1.2的葡萄糖/半乳糖比率,或者本发明提供了乳,所述乳包含基于总碳水化合物《0.01%(w/w)的乳糖、至少10%(w/w)的gos并且具有》1.2的葡萄糖/半乳糖比率。
[0143]
优选地,(无乳糖的)乳是uht乳。现有技术的乳糖降低的uht乳包含大量的单糖,并且这些单糖的存在导致美拉德反应,所述美拉德反应随后导致uht乳褐变(特别是当储存达较长时段时)。尤其是半乳糖在美拉德反应中是反应性的,因此gos含量增加且葡萄糖/半乳糖比率增加的无乳糖uht乳将具有减少的在储存期间褐变的机会。包含基于总碳水化合物《0.01%(w/w)的乳糖、至少10%(w/w)的gos并且具有》1.2的葡萄糖/半乳糖比率的uht乳不包含大量的半乳糖,并且因此减少或没有美拉德/褐变。
[0144]
本发明的发明人在此表明,当从乳糖降低的基于乳的底物制备粉末时,所述乳糖降低的基于乳的底物中gos(优选地原位产生的gos)的存在具有优势。这些优势可以描述为干燥过程效率的优势和/或最终产品(即乳制品(优选乳)粉)的优势。这些优势的示例是(当与不包含或几乎不包含gos的乳糖降低或无乳糖的基于乳的底物相比时):改善(提高)了(喷雾)干燥器的干燥能力;改善(提高)了干燥器的干燥速度;改善(减少)了干燥过程的生产时间;改善(减少)了乳糖降低(优选地无乳糖)的乳制品粉的吸湿行为;改善(减少)了乳糖降低(优选地无乳糖)的乳制品粉的粘性;和/或改善(减少)了乳糖降低的乳制品粉在储存时的褐色形成。
[0145]
因此,在又一实施方式中,本发明因此提供了
[0146]-gos用于改善乳糖降低的基于乳的底物的干燥的用途。优选地,gos是原位产生的gos。
[0147]-gos用于改善无乳糖的基于乳的底物的干燥的用途。优选地,gos是原位产生的gos。
[0148]-gos用于提高用于乳糖降低的基于乳的底物的干燥能力的用途。优选地,gos是原位产生的gos。
[0149]-gos用于提高用于无乳糖的基于乳的底物的干燥能力的用途。优选地,gos是原位产生的gos。
[0150]-gos用于提高乳糖降低的基于乳的底物的干燥速度的用途。优选地,gos是原位产生的gos。
[0151]-gos用于提高无乳糖的基于乳的底物的干燥速度的用途。优选地,gos是原位产生的gos。
[0152]-gos用于减少从乳糖降低的基于乳的底物生产乳糖降低的乳制品粉的时间的用途。优选地,gos是原位产生的gos。
[0153]-gos用于减少从无乳糖的基于乳的底物生产无乳糖的乳制品粉的时间的用途。优
选地,gos是原位产生的gos。
[0154]-gos用于减少乳糖降低的乳制品粉的吸湿行为的用途。优选地,gos是原位产生的gos。
[0155]-gos用于减少无乳糖的乳制品粉的吸湿行为的用途。优选地,gos是原位产生的gos。
[0156]-gos用于降低乳糖降低的乳制品粉的粘性的用途。优选地,gos是原位产生的gos。
[0157]-gos用于降低无乳糖的乳制品粉的粘性的用途。优选地,gos是原位产生的gos。
[0158]-gos用于减少乳糖降低的乳粉的褐色形成的用途。优选地减少了储存时的褐色形成。优选地,gos是原位产生的gos。
[0159]-gos用于减少无乳糖的乳粉的褐色形成的用途。优选地减少了储存时的褐色形成。优选地,gos是原位产生的gos。
[0160]-具有转半乳糖基酶活性的酶和具有乳糖酶活性的酶用于改善乳糖降低的基于乳的底物的干燥的用途。
[0161]-具有转半乳糖基酶活性的酶和具有乳糖酶活性的酶用于改善乳糖降低的基于乳的底物的干燥的用途。
[0162]-具有转半乳糖基酶活性的酶和具有乳糖酶活性的酶用于改善无乳糖的乳粉的特性的用途。
[0163]
上面在方法权利要求的上下文中给出的特征和定义同样适用于用途权利要求。
[0164]
本发明还提供了附加的实施方式:
[0165]
1.一种分离的和/或重组的多肽,所述多肽具有转半乳糖基酶活性并且与seq id no:2具有至少95%的序列同一性。
[0166]
2.根据实施方式1所述的多肽,所述多肽与seq id no:2具有至少96%、97%、98%或99%的序列同一性。
[0167]
3.根据实施方式1或2所述的多肽,所述多肽由seq id no:2组成。
[0168]
4.一种用于生产乳制品的方法,所述方法包括向乳底物中添加有效量的根据实施方式1至3中任一项所述的多肽,以及使所述多肽发挥其酶促活性。
[0169]
5.一种组合物,所述组合物包含根据实施方式1至3中任一项所述的多肽和至少一种选自盐、防腐剂、多元醇或金属离子的组分。
[0170]
6.一种乳制品,所述乳制品可通过根据实施方式4所述的方法获得。
[0171]
7.一种乳制品,所述乳制品包含根据实施方式1至3中任一项所述的多肽。
[0172]
8.根据实施方式1至3中任一项所述的多肽用于产生gos,优选地原位gos的用途。
[0173]
通过以下非限制性实施例来进一步说明本发明。
[0174]
材料和方法
[0175]
本研究中使用的酶
[0176]
1.maxilact lgi5000(dsm food specialties):来自乳酸克鲁维酵母(kluyveromyces lactis)的β-半乳糖苷酶。所用的maxilact lgi5000样品具有5000nlu/ml的活性
[0177]
2.biolacta fn5(amano):来自环状芽孢杆菌的β-半乳糖苷酶。在使用前通过进行如在wo2018/210820中所述的方案,将酶糖化。为此,将biolacta fn5酶粉末以10%(w/v)溶
解在60%葡萄糖中,并于50℃孵育66小时。所使用的biolacta fn5样品的活性为225nlu/mg(糖化前)。
[0178]
3.bht:β-己糖基转移酶,一种源自奇异芽孢杆菌的转半乳糖基酶。所使用的bht样品的活性为124wbdg/ml。过去已经描述了产生这种酶的实验样品的工序(dagher等人(2013)appl.environm.microbiol.79(4)1241
–
1249)。为此,使用标准技术将编码bht酶的基因克隆到合适的生产宿主(如乳酸克鲁维酵母或黑曲霉)中的强启动子后面。在所选的重组表达菌株发酵后,从澄清的发酵液中分离出所分泌的酶,并使用超滤浓缩。下文描述了关于截短的bht酶产物的生产的更多细节。
[0179]
本研究中所使用的smp
[0180]
使用了荷兰乳品研究所(nizo,the netherlands)的nilac脱脂乳粉(smp)。典型的组成是35%的蛋白质、52%的乳糖、1%的脂肪、8%的灰分和4%的水分。
[0181]
截短的bht酶的生产
[0182]
截断的bht的设计
[0183]
为了预测野生型全长蛋白质序列可以在哪里被截短,确定了接头结构域的位置。接头结构域将推定的n末端细胞相关结构域(cell associated domain,cad)与酶中的催化结构域分开。在此接头区域中的任何地方截短可导致功能性可溶酶,因为功能结构域不会失去其结构完整性。在结构域的中间进行切割可能导致酶的结构架构出现问题,这可导致蛋白酶敏感性、错误折叠、聚集或其他妨碍形成稳定的可溶酶产物的问题。为了确定seq id no:1中的接头区域,使用如iupred(http://iupred.enzim.hu/)等在线预测服务器(设置:预测类型:结构化区域,生成曲线图)。对seq id no:1运行此工具,输出标记了在seq id no:1中从第38位延伸直至第55位的n末端接头区域。为了将cad与催化结构域分离,最优选的是对cad进行缺失,并在接头结构域中的某处终止。
[0184]
参考文献:
[0185]
the pairwise energy content estimated from amino acid composition discriminates between folded and intrinsically unstructured proteins-zsuzsanna doszt
á
nyi,veronika csizm
ó
k,p
é
ter tompa and istv
á
n simon j.mol.biol.(2005)347,827-839.
[0186]
iupred:web server for the prediction of intrinsically unstructured regions of proteins based on estimated energy content-zsuzsanna doszt
á
nyi,veronika csizm
ó
k,p
é
ter tompa and istv
á
n simon bioinformatics(2005)21,3433-3434.
[0187]
截短的bht的克隆
[0188]
设计了两种在奇异芽孢杆菌的bht中具有不同截短的构建体。
[0189]
bht#132:没有任何缺失的野生型构建体(seq id no:1)
[0190]
bht#134:缺失从第18位开始直至第54位(seq id no:2)
[0191]
用于在黑曲霉中表达蛋白质的经密码子适应的dna序列被设计成含有用于在曲霉属(aspergillus)表达载体中亚克隆的附加限制性位点。如wo2008/000632中所述进行密码子适应。用于在黑曲霉中表达这些编码seq id no:1和seq id no:2的bht#132和bht#134蛋白的基因的经密码子优化的dna序列分别在seq id no:3和seq id no:4中示出。
[0192]
葡糖淀粉酶glaa启动子的翻译起始序列被修饰成5'-caccgtcaaa atg-3'(seq id no:5),并且最佳翻译终止序列5'-taaa-3'用于表达构建体的产生(也在wo2006/077258中详细描述)。含有葡糖淀粉酶启动子的a.o.部分和bht#132和bht#134编码基因的dna片段(seq id no:6和seq id no:7)被完全合成,纯化并用ecori和paci消化。
[0193]
将pgbtop-16载体通过ecori/paci消化线性化,并随后将线性化的载体片段通过凝胶提取纯化。将dna片段克隆到pgbtop-16载体中,并将所得载体命名为pgbtopbht#132和pgbtopbht#134。随后,使用wo 2011/009700及其中的参考文献中所述的菌株和方法,在与pgbaas-4共转化的方案中用这些载体转化黑曲霉gba 306,并在含乙酰胺的培养基上选择并根据标准工序进行菌落纯化。如wo 98/46772中所述进行转化和选择。通过使用引物进行pcr,从而扩增所引入的bht#132和bht#134基因以验证bht表达盒的存在,来选择含有bht#132和bht#134基因的菌株。从每个表达bht#132基因或bht#134基因的变体中选择两个转化体,并进一步进行复制铺板以获得单一菌株接种物。
[0194]
黑曲霉菌株在微量滴定板中的发酵
[0195]
制备来自上述2个产生的菌株的新鲜黑曲霉孢子,并用于通过在含有3ml发酵培养基2(15%w/v麦芽糖、6%w/v的大豆蛋白胨、1.5%w/v的(nh4)2so4、0.1%w/v的nah2po4.h2o、0.1%w/v的mgso4.7h2o、0.1%w/v的l-精氨酸、8
‰
w/v的tween-80、2
‰
w/v的basildon、2%w/v的mes,ph 6.2)的24深孔板(axygen,union city,usa)中培养菌株来产生样品材料。将24深孔板用breathseal(greiner bio-one,frickenhausen,germany)和盖子覆盖。在34℃、550rpm和80%湿度下在microton培养摇床(infors ag,bottmingen,switzerland)中生长6天后,取1.5ml样品,通过以4000g离心30min从上清液中分离出菌丝体,并将上清液储存于-20℃直到进一步分析。
[0196]
在sds-page上分析不同培养物的少量上清液,并使用考马斯亮蓝染色。所有转化体都显示出预期大小的蛋白质条带。例如,含有构建体bht#134的两个转化体都显示出表观分子量为约70kda的蛋白质的明显分泌。此大小略小于从奇异芽孢杆菌生物质分离的野生型bht的估计大小(ishikawa等人,2005),但远小于在巴斯德毕赤酵母中表达的110kda重组bht(dagher等人,2013)。这表明bht#134在黑曲霉中的表达导致产生更类似于天然掷孢酵母属(sporobolomyces)bht的酶。
[0197]
bht#134在摇瓶中的表达
[0198]
通过在摇瓶培养物中培养黑曲霉转化体菌株,将所述菌株用于生产样品材料。wo 98/46772中描述了一种用于培养黑曲霉菌株和从培养液中分离菌丝体的有用方法。在csm-mes(每升培养基中150g的麦芽糖、60g的大豆蛋白胨(difco)、15g的(nh4)2so4、1g的nah2po4.h2o、1g的mgso4.7h2o、1g的l-精氨酸、80mg的tween-80、20mg的basildon,20g的mes,ph 6.2)中进行培养。将摇瓶在280rpm和30℃下培养。在发酵的第6天从重复培养物中取出样品,在hereaus labofuge rf中以5000rpm离心10min,并将上清液于-20℃储存直至进一步分析。
[0199]
为了获得bht#134转化体的生产率的估计值,根据制造商的使用说明(sigma-b6916)并且还如在wo2014089558中所使用的,使用bradford总蛋白分析对所有发酵上清液进行分析。对不表达或几乎不表达任何附加蛋白质的样品中的总蛋白质浓度取平均,并与bht#134转化体样品的平均值进行比较。生产率表示为bht#134样品与对照之间的差异。结
果在表1中描述。
[0200]
表1:bht#134的生产率。对照是未转化的黑曲霉cbs513.88的类似培养。wo2014089558指出了现有技术中野生型bht在巴斯德毕赤酵母中的最大生产率。
[0201][0202]
这种在mtp和摇瓶中的生产率比现有技术中所述的任何担子菌纲bht的分泌高得多。据我们所知,这种酶的最佳分泌是用奇异芽孢杆菌bht获得的,在所述奇异芽孢杆菌bht中常规信号序列被αmf信号序列取代(wo2014089558)。这种构建体在巴斯德毕赤酵母中的分泌导致摇瓶中9.8mg bht/l的生产率,比bht#134分泌低大于50倍。
[0203]
maxilact lgi5000和biolacta fn5的活性确定
[0204]
使用基本上按照如food chemical codex(fcc 6)第1124-1126页中所述进行的中性乳糖酶活性(neutral lactase activity,nlu)测定来确定maxilact lgi5000和biolacta fn5中的β-半乳糖苷酶活性。
[0205]
bht的活性确定
[0206]
bht的活性是用基本上类似于所描述的测定的测定确定的(workman和day(1982)appl.environm.microbiol.44(6)1289-1295)。稍微调整测定条件,并于37℃在对硝基苯基-对-d-吡喃葡萄糖苷(pnpg)浓度为4.7mm的醋酸钠缓冲液ph 4.40中进行。在反应终止后,测量在405nm处释放的pnp,并且将酶活性表示为每秒释放的对硝基苯酚的纳摩尔数(bhtu)。
[0207]
乳糖、葡萄糖、半乳糖和gos含量的确定
[0208]
使用基本上如由aoac(selected food products中的“aoac official method 2001.02trans-galactooligosaccharides(tgos)”,aoac international,gaithersburg,md,usa,2005)所述的hpaec-pad方法确定基于乳的样品中的乳糖、葡萄糖和半乳糖的含量。
[0209]
当如在本专利申请内所述的基于乳的底物和基于乳的产物都可用时(如实施例3中的情况),可以确定基于乳的底物和基于乳的产物中的乳糖、葡萄糖和半乳糖的浓度。基于乳的产物中的总gos浓度可以通过用式(1)从基于乳的底物中的乳糖、葡萄糖和半乳糖含量减去基于乳的产物中的乳糖、葡萄糖和半乳糖含量来计算:
[0210]
[gos]
产物
=([lac]+[glu]+[gal])
底物
ꢀ‑
([lac]+[glu]+[gla])
产物 (1)
[0211]
[lac]=乳糖浓度,单位为g/l
[0212]
[glu]=葡萄糖浓度,单位为g/l
[0213]
[gal]=半乳糖浓度,单位为g/l
[0214]
[gos]=低聚半乳糖浓度,单位为g/l
[0215]
当必须在没有可用的基于乳的底物的情况下确定基于乳的产物或基于乳的粉末的样品中的gos含量时,基于乳的样品的总低聚半乳糖(gos)含量可以用类似于标准aoac方法(selected food products中的“aoac official method 2001.02trans-galactooligosaccharides(tgos)”,aoac international,gaithersburg,md,usa,2005)的方法确定。使用所述的hpaec-pad方法,可以在用酸性乳糖酶水解样品之前(初始)和之后(最终)确定样品中半乳糖、葡萄糖和乳糖的浓度。初始样品中的gos含量可以通过用以下式(2)从糖的最终测量的量中减去糖的初始测量的量来计算:
[0216]
[gos]
初始
=([lac]+[glu]+[gal])
最终-([lac]+[glu]+[gla])
初始
ꢀꢀ
(2)
[0217]
[lac]=乳糖浓度,单位为g/l或g/kg
[0218]
[glu]=葡萄糖浓度,单位为g/l或g/kg
[0219]
[gal]=半乳糖浓度,单位为g/l或g/kg
[0220]
[gos]=低聚半乳糖浓度,单位为g/l或g/kg
[0221]
基于乳的底物、基于乳的产物或乳粉中的总碳水化合物被定义为葡萄糖、半乳糖、乳糖和gos的浓度的总和。
[0222]
通过差示扫描量热(dsc)确定玻璃化转变温度(tg)
[0223]
用精密天平称量2.5mg质量的乳粉样品,并封装在已知质量的(压接的)铝盘中。使用相同的空盘作为参考。将氮气以50ml min-1
的速度吹扫。在-80℃至120℃范围内,在dsc装置(metller dsc 3+)中以10k/min施加加热-冷却-加热-冷却-加热循环。在最高温度下选择0.5分钟的停留时间以允许水分蒸发,与此同时进行dsc信号平衡。进行三次加热-冷却循环。第一加热循环主要用于蒸发水,而第二循环和第三循环用于测量tg并建立测量的再现性。
[0224]
实施例1:用bht和maxilact处理的脱脂乳
[0225]
通过在30min内在搅动(250rpm)下将100克脱脂乳粉(smp)分散在900ml自来水(40℃)中来制备10%(w/v)的脱脂乳粉(smp)溶液。在室温下静置30分钟后,将分散体以150rpm于4℃搅动24h。将5ml这种乳的样品于8℃用bht处理24小时。bht剂量为1bhtu/ml、2bhtu/ml、5bhtu/ml或10bhtu/ml。此外,与bht同时添加1nlu/ml或2.5nlu/ml的maxilact lgi5000,或在18小时后添加5nlu/ml或10nlu/ml的maxilact lgi5000(因此与maxilact一起孵育的时间被限制为6小时)。对照样品是不添加酶的smp溶液(对照a)、与仅5nlu/ml的maxilact一起孵育(对照c)和与仅5bhtu/ml的bht一起孵育24h(对照d)。在孵育后通过于95℃加热10分钟来停止所有反应。将样品分成1ml部分用于糖分析并且4ml部分冷冻储存以供以后分析。
[0226]
表2:用bht和maxilact处理的脱脂乳样品。指示了酶的剂量和与所添加的bht一起的孵育时间(孵育时间1)或所添加的maxilact一起的孵育时间(孵育时间2)。
[0227][0228][0229]
实施例2:用糖化的biolacta和maxilact处理的脱脂乳
[0230]
通过在30min内在搅动(250rpm)下将100克脱脂乳粉(smp)分散在900ml自来水(40℃)中来制备10%(w/v)的脱脂乳粉(smp)溶液。在室温下静置30分钟后,将分散体以150rpm于4℃搅动24h。将5ml的这种乳的样品于8℃用糖化的biolacta fn5处理24小时。糖化的
biolacta fn5制备物的酶剂量为1mg/ml、2mg/ml、5mg/ml或10mg/ml的脱脂乳。此外,与糖化的biolacta同时添加1nlu/ml和2.5nlu/ml的maxilact lgi5000,或者在18小时后添加5nlu/ml和10nlu/ml的maxilact lgi5000(因此与maxilact一起孵育的时间被限制为6小时)。对照样品是不添加酶的smp溶液(对照a1)、与仅5nlu/ml的maxilact一起孵育(对照c1)和与仅10mg/ml的biolacta一起孵育24h(对照d)。在孵育24h后通过于95℃加热10分钟来停止所有反应。将样品分成1ml部分用于糖分析并且4ml部分冷冻储存以供以后分析
[0231]
表3:用糖化的biolacta和maxilact处理的脱脂乳样品。指示了酶的剂量和与所添加的糖化的biolacta一起的孵育时间(孵育时间1)或所添加的maxilact一起的孵育时间(孵育时间2)。
[0232]
[0233][0234]
实施例3:糖分析
[0235]
如材料和方法部分中所述,分析实施例1和实施例2中产生的所有样品的糖组成(葡萄糖、半乳糖和乳糖)。如本部分中所述,由所测量的糖的含量计算每个样品的gos含量。针对通过糖化的biolacta样品在样品中添加的葡萄糖量,校正实施例2的样品中所测量的葡萄糖含量。例如,在对照d1中,添加10mg/ml的具有60%葡萄糖的糖化biolacta,因此从测量中已经减去6g/l葡萄糖。
[0236]
表4:实施例1中产生的样品的乳糖、葡萄糖、半乳糖和gos的含量
[0237]
[0238][0239]
表5:实施例2中产生的样品的乳糖、葡萄糖、半乳糖和gos的含量
[0240]
通过计算每个样品中的质量平衡并与未处理的脱脂乳的初始乳糖含量进行比较,检查分析方法中所有乳糖和gos的完全转化,并且发现是令人满意的。
[0241]
结果表明,可以产生具有低乳糖和可观量(总碳水化合物的》10%(w/w))的gos的脱脂乳样品。用bht和maxilact的组合获得了最好的结果(样品编号4至8和编号13至16),但是糖化的biolacta和maxilact的组合也给出了令人满意的结果(样品编号13a至16a)。
[0242]
实施例4:乳粉样品的tg的确定
[0243]
将样品对照a、对照c、样品4和样品16(全部在实施例1中描述)冷冻干燥,并且使用如在材料和方法部分中所述的差示扫描量热法(dsc)确定粉末的玻璃化转变温度(tg)。所有的dsc首次加热曲线都产生了很可能反映水蒸发的宽吸热峰。随后的dsc加热斜升(例如,第二加热或第三加热)进行到120摄氏度的最高温度并产生热容量的可再现不连续性,所述
可再现不连续性可以解释为玻璃化转变现象;第二加热斜升与第三加热斜升之间的再现性表明,一方面水去除是成功的,并且最终的热诱导降解在实验期间没有发生。根据第二温度循环和第三温度循环的热容不连续性的起始温度计算tg,并且在表6中描述。起始温度被定义为转变前的基线与拐点切线的交点。
[0244]
表6:与对照相比,用bht和maxilact处理的冷冻干燥脱脂乳样品的tg。
[0245][0246]
该测量显示,与常规的无乳糖的脱脂乳粉样品相比,gos增加的脱脂乳粉样品的tg明显增加。tg的增加将导致干燥特性和粉末稳定性的改善。
[0247]
实施例5:乳粉样品的吸水性的确定
[0248]
完全按照实施例1中针对对照a、对照c和样品16的描述,用bht和maxilact处理大量溶解的脱脂乳粉。在实验室喷雾干燥器(labplant sd-05,keison products)上使用128.8℃的入口空气温度、78.1℃的出口空气温度、1508.9g/h的蒸发能力、1718.5g/h的脱脂乳流率和176.5g/h的粉末流率干燥每种经处理的脱脂乳的样品。于50℃在动态蒸汽吸附机器(dvs-ta instruments q5000sa)中分析用这种方法产生的3毫克脱脂乳粉。跟踪水分含量(克水/克样品)15小时。
[0249]
表7:于50℃随时间推移的吸水。吸水性被描述为克/克样品
[0250]
将所生产的脱脂乳粉储存起来,并以规律的时间间隔进行目测检查。常规smp(对照a)没有显示出任何可见的结块,而lf-smp(对照c)在储存大于一周后显示出严重的结块。与对照c相比,经bht/maxilact处理的lf-smp(样品16)显示出明显较低的结块趋势。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1