肝类器官疾病模型以及其制备和使用方法与流程
本申请要求于2016年11月4日提交的美国临时专利申请62/471,371和2016年6月9日提交的美国临时专利申请62/517,414的优先权和权益,所述每个申请的内容通过引用以其整体并入用于所有目的。
背景技术:
不可逆的上皮器官重塑是导致全球死亡和疾病的主要因素,每年使医疗保健系统损失数十亿美元(hynds和giangreco,2013)。上皮重塑疾病包含肺癌和胃肠癌以及慢性疾病,如肝硬化、慢性阻塞性肺病(copd)和炎性肠病(hynds和giangreco,2013)。可悲的是,主要针对动物进行的大多数上皮器官研究未能为这些疾病产生新疗法,并且死亡率仍然高得令人无法接受。部分原因是由于缺乏预测性人体系统以测试制药行业中大量增加的化合物库的功效,因此对开发用于模拟人类炎症和纤维化以实现临床相关治疗开发的高保真系统提出了根本性挑战。
非酒精性脂肪性肝病(nafld)是发达国家需要克服的主要挑战之一,因为发生致死性肝病的可能性增加,但缺乏有效的治疗方法。同样,医源性肠胃外营养相关肝病(pnald)是一种由肠胃外营养引起的病症,并且目前尚无有效的治疗方法。需要具有脂肪性肝炎和/或肝病的临床组织病理学特征,如脂滴积聚和膀胱骨架细丝解体、脂肪变性和肝细胞气球样变,并且可以用于解决这些和其它肝病症或疾病状态的模型。尽管使用患者干细胞的疾病模型有希望,但目前的方法在其应用中仅限于单细胞、单基因和相对简单的病理,未能捕获更普遍和复杂的疾病病理学,如上皮器官纤维化。
人体肝是一种重要的器官,为生命提供许多重要的代谢功能,如脂质代谢、铵和胆汁的产生、凝血以及外源性化合物的解毒作用。使用诱导多能干细胞(ipsc)技术,由于许多有希望的应用,包含再生疗法、药物发现和药物毒性研究,体外重建患者的肝反应对制药工业具有吸引力。为此目的,目前,常规的体外方法研究二维(2-d)和3-d分化平台以生成肝细胞。然而,大多数报道的方法主要将细胞分化成靶上皮细胞类型,完全缺乏必需的支持组分,如促纤维化和/或炎性细胞类型。替代性地,申请人和其他人已经提出通过混合上皮和支持谱系的基于共培养的方法,然而,这些测定变化很大并且经常被许多人为变化混淆,如难以选择上皮细胞培养基(ecm)和可以共同维护的培养基。因此,需要建立新的和稳健的测定系统,其中支持谱系共同开发用于疾病建模和进一步的筛选应用。
技术实现要素:
本文公开了制备和使用脂毒性类器官模型的方法。在某些方面,所述方法可以包括使肝类器官与游离脂肪酸(ffa)组合物接触的步骤。在一方面,所述ffa组合物可以包括油酸、亚油酸、棕榈酸或其组合。
附图说明
本领域中的技术人员将理解,下面描述的附图仅用于说明性目的。附图不旨在以任何方式限制本教导的范围。
专利或申请文件包含至少一幅彩色附图。具有一副或多副彩色附图的本专利或专利申请公开的副本将在请求并支付必要费用后由主管局提供。
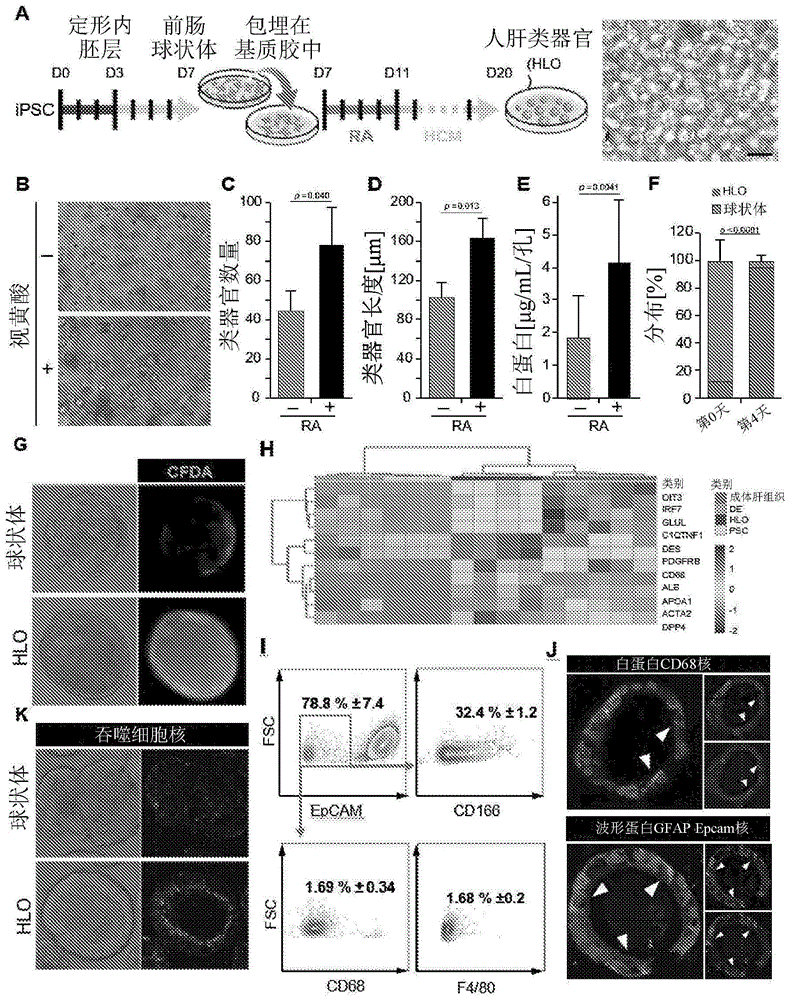
图1.人ipsc-肝类器官中纤维化前和炎症谱系的共分化a.基于视黄酸(ra)的肝类器官分化方法和第20天肝类器官的明视野图像的示意图。比例尺,100μm。b.在存在和不存在4天ra的培养之后第20天基质胶中肝类器官的明视野图像。c.在存在(黑色条)和不存在(灰色条)4天ra的培养之后第20天手动测量类器官数。d.在存在(黑色条)和不存在(灰色条)4天ra的培养之后第20天,通过图像j识别和测量类器官的直径。e.白蛋白产生是从培养之后24小时收集的培养物上清液测量的,所述培养是在第22天到第25天培养之后,在基质胶中用(黑色条)或不用(灰色条)4天ra的肝类器官。f.在存在和不存在4天ra下培养之后第25天手动计数具有内腔结构的类器官。红色和灰色条分别表示具有(hlo)/不具有(球状体)内腔结构的类器官的百分比。g.通过二乙酸荧光素(fd)监测胆汁转运活性,其在肝细胞中的酯酶水解之后,在培养基中变成荧光素(绿色)并跟踪荧光的流动。结果代表平均值±sd,n=3。通过流式细胞术确定epcam、cd166和cd68阳性群体的百分比。结果代表平均值±sd,n=2-3。h.单细胞rna测序!!由人ipsc、人ipsc衍生的定形内胚层、前肠球状体、hlo、人成体肝组织和人胎肝组织表达的肝细胞、星状细胞、库普弗细胞和内皮相关基因的fpkm(log2)值。i.通过流式细胞术确定epcam、cd166、cd68和f4/80阳性群体的百分比。结果代表平均值±sd,n=3。j.第25天hlo对白蛋白、cd68、波形蛋白、gfap和epcam的免疫荧光(if)染色。白色箭头表示cd68、gfap和波形蛋白阳性细胞的定位。结果代表平均值±sd,n=3。k.通过使用反映吞噬作用的phrodo指示剂(红色)监测细胞内ph来分析吞噬细胞活性。在共聚焦显微镜下捕获荧光表达。结果代表平均值±sd,n=6。
图2.通过脂肪酸处理生成脂肪性肝炎类器官(shlo)a.表示生成脂肪性肝炎hlo(shlo)的方法的示意图b.脂滴(绿色)、膜(红色)和核(蓝色)的活细胞成像。图像是从10个到20个z-堆叠图像的叠加采用的。以剂量依赖的方式观察到脂滴积聚的增加和脂滴和细胞的增大。c.由每个类器官大小标准化的代表性总脂质体积。条形图示出总脂质体积的平均值。脂滴以剂量依赖性方式增加0(黑色)、200μmoa(红色)、400μmoa(绿色)和800μmoa(蓝色)。d.hlo中甘油三酯的定量。从一个基质胶滴分离hlo,并在油酸(800μm)存在(蓝色条)或不存在(黑色条)下分成hcm培养基并培养3天。e.用从含有20个到30个hlo的孔获得的培养物上清液对il-6进行elisa测量,所述孔在油酸(800μm)存在或不存在下培养并培养3天。最终值通过每个孔中的类器官数量标准化。当与非处理(黑色条)相比时,il-6在800μmoa处理(蓝色条)中释放2.2倍。f.促炎细胞因子tnf-α和il-8的基因表达。它们被18s标准化。与未处理(黑色条)相比,在200μmoa(红色条)和800μmoa(蓝色条)中tnf-α和il-8基因表达两者均上调。g.将10个到20个hlo在包含0um、400um、800umoa的hcm培养基中培养3天。从每个孔收集培养的上清液,并通过使用跨膜与那些上清液测量thp-1迁移。对已经迁移的细胞进行计数,并通过每个孔中确切的类器官数量进行标准化。h.hlo(黑色条)、shlo(红色条)和chlo(蓝色条)群体上的第25天hlo的三色染色和三色染色的hlo的百分比。结果代表平均值±sd,n=8个到20个类器官。i.对于epcam和波形蛋白的第25天hlo的if染色和hlo(黑色条)、shlo(红色条)和chlo(蓝色条)群体上的epcam和波形蛋白阳性hlo的百分比。结果代表平均值±sd,n=8个到20个类器官。j.在存在或不存在油酸(800μm)下培养20个到30个hlo5天。用那些上清液通过elisa测量p3np。最终值通过每个孔中的类器官确切数量标准化。当与未处理(黑色条)相比时,p3np在800μmoa处理(蓝色条)中增加2.8倍。
图3.由afm测量脂肪性肝炎hlo的肝硬化转变。a.用afm测量hlo硬度的示意图。用afm悬臂扫描每个单个hlo的顶部区域(25μm×25μm方形中的14×14矩阵),其可以提供hlo的地形和机械信息的空间映射。比例尺,100μm。b.单个hlo的计算杨氏模量(hlo的刚度;e,kpa)的代表性直方图示出高斯样分布,并且其峰值和宽度以剂量依赖性方式0(黑色)μm、200(红色)μm、400(绿色)μm和800(蓝色)μmoa增加。c.杨氏模量(hlo的硬度)由7个到12个类器官确定,并通过点图用盒-须图绘制。以剂量依赖性方式0(黑色)μm、200(红色)μm、400(绿色)μm和800(蓝色)μmoa观察到中值的增加。
图4.chlo刚性概括了wolman病的临床表型a.hlo和从包含健康人(317d6)、nafld患者(nafld150、nafld77和nafld27)和wolman病患者(wd90、wd92和wd91)的几个ipsc系建立的chlo的明视野图像。b.衍生自几个ipsc系的单个hlo和clo的平均杨氏模量(hlo刚度:pa)。
图5.使用ipsc-shlo模拟脂肪变性的人类表型变异a.负责肝脏tg含量增加的pnpla3、gckr和tm6sf2的代表性等位基因功能。b.饼图示出基于三个风险等位基因的总多基因评分的公众可获得的2504细胞系的百分比分布:0.6<x(绿色区域),0.3<x<=0.6(黄色区域),x<=0.3(棕色区域)。c.表总结了供体特征,包含在进一步研究中指定的多基因评分。d.三个等位基因的三组总多基因评分的脂滴(绿色)和核(蓝色)的活细胞成像:0.6<x,0.3<x<0.6,x<=0.3。图像是从10个到20个z-堆叠图像的叠加采用的。e.由三个等位基因的三组总多基因评分上的每个类器官大小标准化的代表性总脂质体积:0.6<x,0.3<x<0.6,x<=0.3。红色和海军条分别表示800μmoa和200μmoa处理的shlo。f.三个等位基因的三组总多基因评分上促炎细胞因子的代表性基因表达:0.6<x(黑色条),0.3<x<=0.6(红色条),x<=0.3(蓝色条)。g.三个等位基因的三组总多基因评分上的单个clo的平均杨氏模量(hlo的刚度:pa):0.6<x(黑点),0.3<x<=0.6(红点),x<=0.3(蓝点)。h.oca反应
图6.吞噬细胞对thp-1的活性
图7.800μmoa、pa、la和sa的脂质积累比较。
图8.thp-1的实际迁移细胞数为400μm和800μmoa
图9.a.来自e-cad-mruby类器官hepg2、lx-2和thp-1的e-cad阳性和阴性分选重构球状体的照片。b.促炎细胞因子tnf-α和il-8的基因表达。c.p3np的elisa测量。
图10.白藜芦醇对肝类器官的ros产生无效
图11.通过流式细胞术确定bodipy阳性细胞(脂质)的百分比。结果代表平均值±sd,n=5。
图12.在存在和不存在400μm脂肪乳剂下,脂质在类器官中积累
图13.未处理、单独800μmoa和具有40ng/mlfgf19的800μmoa的培养物中的肝类器官的明视野图像。
具体实施方式
除非另有说明,否则术语应根据相关领域中普通技术人员的常规用法来理解。
术语“约”或“大约”意指在本领域普通技术人员确定的特定值的可接受误差范围内,这将部分取决于如何测量或确定值,例如,测量系统的局限性。例如,根据本领域的实践,“约”可以表示在1或大于1的标准偏差内。替代性地,“约”可以表示给定值的多达20%,或多达10%,或多达5%,或多达1%的范围。替代性地,特别是对于生物系统或过程,术语可以表示处于一个数量级内,优选地在值的5倍内,并且更优选地在2倍内。在申请和权利要求中描述了特定值的情况下,除非另有说明,否则应当假设术语“约”意指处于特定值的可接受误差范围内。
如本文所用,术语“全能干细胞”(也称为全能干细胞)是可以分化成胚胎和胚胎外细胞类型的干细胞。这种细胞可以构建完整的、有活力的生物体。这些细胞是由卵子和精子细胞融合而成。由受精卵的前几个分裂产生的细胞也是全能的。
如本文所用,术语“多能干细胞(psc)”涵盖可以分化成身体的几乎所有细胞类型的任何细胞,即衍生自三个胚层(生发上皮)中的任何一个的细胞,包含内胚层(内部胃壁、胃肠道、肺)、中胚层(肌肉、骨骼、血液、泌尿生殖器)和外胚层(表皮组织和神经系统)。psc可以是植入前胚泡的内细胞团细胞的后代,或者通过强迫某些基因的表达诱导非多能细胞,如成体体细胞而获得。多能干细胞可以衍生自任何合适的来源。多能干细胞来源的实例包含哺乳动物来源,包含人、啮齿动物、猪和牛。
如本文所用,术语“诱导的多能干细胞(ipsc)”,也通常缩写为ips细胞,是指通过诱导某些基因的“强制”表达,人工衍生自非正多能细胞,如成体体细胞的一类多能干细胞。hipsc是指人ipsc。
如本文所用,术语“胚胎干细胞(esc)”,也通常缩写为es细胞,是指多能的细胞并且衍生自胚泡的内细胞团,即早期胚胎。出于本发明的目的,术语“esc”有时也广泛用于涵盖胚胎生殖细胞。
如本文所用,术语“前体细胞”涵盖可以用于本文所述方法中的任何细胞,通过所述细胞,一种或多种前体细胞获得自我更新或分化成一种或多种特化细胞类型的能力。在一些实施例中,前体细胞是多能的或具有变成多能的能力。在一些实施例中,对前体细胞进行外部因子(例如生长因子)的处理以获得多能性。在一些实施例中,前体细胞可以是全能(或全能)干细胞;多能干细胞(诱导或非诱导);多能干细胞;寡能干细胞和单能干细胞。在一些实施例中,前体细胞可以来自胚胎、婴儿、儿童或成人。在一些实施例中,前体细胞可以是经受处理的体细胞,使得通过遗传操作或蛋白质/肽处理赋予多能性。
衍生自胚胎细胞的多能干细胞。
在一些实施例中,一个步骤是获得多能干细胞或可以诱导成为多能干细胞。在一些实施例中,多能干细胞衍生自胚胎干细胞,所述胚胎干细胞又衍生自早期哺乳动物胚胎的全能细胞,并且能够在体外无限制的未分化增殖。胚胎干细胞是衍生自早期胚胎的胚泡内细胞团的多能干细胞。从胚细胞衍生胚胎干细胞的方法是本领域熟知的。人胚胎干细胞h9(h9-hesc)用于本申请中描述的示例性实施例中,但是本领域技术人员将理解,本文描述的方法和系统适用于任何干细胞。
可以用于根据本发明的实施例中的额外的干细胞包含但不限于由旧金山加利福尼亚大学(ucsf)的人类胚胎干细胞研究中心的国立干细胞库(nscb)主办的数据库;wi细胞研究所的wisc细胞库;威斯康星大学干细胞与再生医学中心(uw-scrmc);novocell公司(加利福尼亚州圣地亚哥);cellartisab(瑞典哥德堡);胚胎干细胞国际公司(新加坡);以色列理工学院以色列理工学院(以色列海法);以及由普林斯顿大学和宾夕法尼亚大学主办的干细胞数据库提供或描述的那些干细胞。可以用于根据本发明的实施例中的示例性胚胎干细胞包含但不限于sa01(sa001);sa02(sa002);es01(hes-1);es02(hes-2);es03(hes-3);es04(hes-4);es05(hes-5);es06(hes-6);bg01(bgn-01);bg02(bgn-02);bg03(bgn-03);te03(13);te04(14);te06(16);uc01(hsf1);uc06(hsf6);wa01(h1);wa07(h7);wa09(h9);wa13(h13);wa14(h14)。
关于胚胎干细胞的更多细节可以在例如thomson等人,1998,“衍生自人类胚泡的胚胎干细胞系(embryonicstemcelllinesderivedfromhumanblastocysts),”《科学(science)》282(5391):1145-1147;andrews等人,2005,“胚胎干(es)细胞和胚胎癌(ec)细胞:同一枚硬币的两侧(embryonicstem(es)cellsandembryonalcarcinoma(ec)cells:oppositesidesofthesamecoin),”《生化学会会刊(biochemsoctrans)》33:1526-1530;martin1980,“畸胎癌和哺乳动物胚胎发生(teratocarcinomasandmammalianembryogenesis)”,《科学(science)》209(4458):768-776;evans和kaufman,1981,“小鼠胚胎多能细胞培养的建立(establishmentincultureofpluripotentcellsfrommouseembryos)”,《自然(nature)》292(5819):154-156;klimanskaya等人,2005,“衍生自非饲养细胞的人胚胎干细胞(humanembryonicstemcellsderivedwithoutfeedercells)”,《柳叶刀(lancet)》365(9471):1636-1641中找到;所述参考中的每一个在此以其整体并入本文。
诱导多能干细胞(ipsc)
在一些实施例中,通过将某些干细胞相关联基因转染到非多能细胞,如成体成纤维细胞中衍生ipsc。通常通过病毒载体,如逆转录病毒实现转染。转染的基因包含主转录调节因子oct-3/4(pouf51)和sox2,尽管建议其它基因提高诱导效率。3周到4周之后,少量转染细胞开始在形态学和生物化学上类似于多能干细胞,并且通常通过形态学选择、倍增时间或通过报告基因和抗生素选择分离。如本文所用,ipsc包含但不限于小鼠中的第一代ipsc、第二代ipsc和人诱导的多能干细胞。在一些实施例中,使用四种关键基因:oct3/4、sox2、klf4和c-myc,逆转录病毒系统用于将人类成纤维细胞转化成多能干细胞。在替代性实施例中,慢病毒系统用于用oct4、sox2、nanog和lin28转化体细胞。表达在ipsc中诱导的基因包含但不限于oct-3/4(例如,pou5fl);sox基因家族的某些成员(例如,sox1、sox2、sox3和sox15);klf家族的某些成员(例如,klf1、klf2、klf4和klf5),myc家族的某些成员(例如,c-myc、l-myc和n-myc),nanog和lin28。
在一些实施例中,使用基于非病毒的技术生成ipsc。在一些实施例中,腺病毒可以用于将必需的四种基因转运到小鼠的皮肤和肝细胞的dna中,产生与胚胎干细胞相同的细胞。由于腺病毒不将其任何自身基因与靶向宿主组合,因此消除了产生肿瘤的危险。在一些实施例中,重编程可以通过质粒完成,而根本没有任何病毒转染系统,尽管效率非常低。在其它实施例中,蛋白质的直接递送用于生成ipsc,因此消除了对病毒或遗传修饰的需要。在一些实施例中,使用类似的方法可以生成小鼠ipsc:用通过聚精氨酸锚定引入细胞中的某些蛋白质重复处理细胞足以诱导多能性。在一些实施例中,还可以通过在低氧条件下用fgf2处理体细胞来增加多能性诱导基因的表达。
关于胚胎干细胞的更多细节可以在例如kaji等人,2009,“病毒自由诱导多能性和随后切除重编程因子(virusfreeinductionofpluripotencyandsubsequentexcisionofreprogrammingfactors)”,《自然(nature)》458:771-775;woltjen等人,2009,“piggybac转座将成纤维细胞重编程为诱导的多能干细胞(piggybactranspositionreprogramsfibroblaststoinducedpluripotentstemcells),”《自然(nature)》458:766-770;okita等人,2008,“没有病毒载体的小鼠诱导的多能干细胞的生成(generationofmouseinducedpluripotentstemcellswithoutviralvectors)”,《科学(science)》322(5903):949-953;stadtfeld等人,2008,“无病毒整合生成的诱导多能干细胞(inducedpluripotentstemcellsgeneratedwithoutviralintegration),”《科学(science)》322(5903):945-949;和zhou等人,2009,“使用重组蛋白生成诱导的多能干细胞(generationofinducedpluripotentstemcellsusingrecombinantproteins)”,《细胞干细胞(cellstemcell)4(5):381-384中找到;所述参考中的每一个在此以其整体并入本文。
在一些实施例中,示例性ips细胞系包含但不限于ips-df19-9;ips-df19-9;ips-df4-3;ips-df6-9;ips(包皮);ips(imr90);和ips(imr90)。
关于与de发育相关的信号传导途径的功能的更多细节可以在例如zorn和wells,2009,“脊椎动物内胚层发育和器官形成(vertebrateendodermdevelopmentandorganformation),”《细胞和发育生物学的年度评论(annurevcelldevbiol)》25:221-251;dessimoz等人,2006,“fgf信号传导对于在体内沿前后轴建立肠管区域是必要的(fgfsignalingisnecessaryforestablishingguttubedomainsalongtheanterior-posterioraxisinvivo)”,《老化和发育机制(mechdev)》123:42-55;mclin等人,2007,“在前内胚层中抑制wnt/β-连环蛋白信号传导对肝和胰腺发育至关重要(repressionofwnt/β-cateninsignalingintheanteriorendodermisessentialforliverandpancreasdevelopment),《发育(development)》,”134:2207-2217;wells和melton,2000,《发育(development)》127:1563-1572;desantabarbara等人,2003,“肠上皮的发育和分化(developmentanddifferentiationoftheintestinalepithelium)”,《细胞与分子生命科学(cellmollifesci)》60(7):1322-1332中找到;所述参考中的每一个在此以其整体并入本文。
从多能细胞(例如,ipsc或esc)产生定形内胚层的任何方法都适用于本文所述的方法。示例性方法公开在例如wells等人us9719068b2的“通过定向分化将前体细胞转化为肠组织的方法和系统(methodsandsystemsforconvertingprecursorcellsintointestinaltissuesthroughdirecteddifferentiation)”和wells等人us20170240866a1的“通过定向分化将前体细胞转化为胃组织的方法和系统(methodsandsystemsforconvertingprecursorcellsintogastrictissuesthroughdirecteddifferentiation)”中。在一些实施例中,多能细胞可以衍生自桑椹胚。在一些实施例中,多能干细胞可以是干细胞。在这些方法中使用的干细胞可以包含但不限于胚胎干细胞。胚胎干细胞可以衍生自胚胎内细胞团或衍生自胚胎性腺脊。胚胎干细胞或生殖细胞可以源自多种动物物种,包含但不限于包含人的各种哺乳动物物种。在一些实施例中,人胚胎干细胞用于产生定形内胚层。在一些实施例中,人胚胎生殖细胞用于产生定形内胚层。在一些实施例中,ipsc用于产生定形内胚层。用于获得或产生可以用于本发明中的de细胞的额外方法包含但不限于d'amour等人的美国专利号7,510,876;fisk等人的美国专利号7,326,572;kubo1等人,2004,“从培养的胚胎干细胞发育定形内胚层(developmentofdefinitiveendodermfromembryonicstemcellsinculture)”,《发育(development)》131:1651-1662;d'amour等人,2005,“人胚胎干细胞对定形内胚层的有效分化(efficientdifferentiationofhumanembryonicstemcellstodefinitiveendoderm)”,《自然生物工艺学(naturebiotechnology)》23:1534-1541;和ang等人,1993,“小鼠中定形内胚层谱系的形成和维持:hnf3/叉头蛋白的参与(theformationandmaintenanceofthedefinitiveendodermlineageinthemouse:involvementofhnf3/forkheadproteins)”,《发育(development)》119:1301-1315中描述的那些。
非酒精性脂肪性肝病(nafld)是发达国家为了增加发生致命性肝病的机会而需要克服的主要挑战之一,但缺乏有效的治疗方法。类似地,医源性父母营养相关肝病(pnald)是一种目前尚无有效治疗的病症。需要具有脂肪性肝炎和/或肝病的临床组织病理学特征的模型,如脂滴积聚和骨髓基质细丝解体、脂肪变性和肝细胞气球样变。尽管使用患者干细胞的疾病模型有希望,但目前的方法在其应用中仅限于单细胞、单基因和相对简单的病理,未能捕获更普遍和复杂的疾病病理学,如上皮器官纤维化。
一方面,公开了一种制备脂毒性类器官模型的方法。所述方法可以包括使根据本文所述方法制备的肝类器官与游离脂肪酸(ffa)组合物接触的步骤。所述ffa组合物可以包括油酸、亚油酸、棕榈酸、或其组合,优选地油酸。ffa的量可以由本领域普通技术人员确定。一方面,ffa,优选地油酸,可以与肝类器官接触的量为约10μm到约10,000μm,或约20μm到约5,000μm,或约30μm到约2500μm,或约40μm到约1250μm或约50μm到约1000μm,或约75μm到约900μm,或约80μm到约800μm,或约90μm到约700μm,或约100μm到约500μm,或约200μm到约400μm。ffa可以与肝类器官接触的时间段为约1小时到约10天,或约2小时到约9天,或约3小时到约8天,或约4小时到约7天,或约5小时到约6天,或约6小时到约5天,或约7小时到约4天,或约8小时到约3天,或约9小时到约2天,或约10小时到约1天。一方面,范围是约3天到约5天,+/-24小时。
一方面,脂毒性类器官模型是脂肪肝病的模型。
一方面,脂毒性类器官模型是脂肪性肝炎的模型。
一方面,脂毒性类器官模型是肝硬化的模型。
一方面,所述脂毒性类器官模型是肠胃外营养相关肝病(pnald)的模型。
一方面,所述脂毒性类器官模型是nafld的模型。
一方面,所述脂毒性类器官模型的特征可以在于细胞骨架细丝解体、ros增加、线粒体肿胀、三酸甘油酯积累、纤维化、肝细胞气球样、il6分泌、脂肪变性、炎症、气球样和马洛里体样、组织硬化、细胞死亡和其组合。
一方面,公开了一种筛选用于治疗肝病,包含nafld和/或胆汁淤积的药物的方法。所述方法可以包括使候选药物与本文公开的脂毒性类器官模型接触的步骤。
一方面,公开了一种测定营养补充剂/tpn的有效性的方法。所述方法可以包括使所述营养补充剂/tpn与本文公开的脂毒性类器官模型接触的步骤。
一方面,公开了脂肪肝病的所述三维(3d)肝类器官模型,其中所述类器官的特征在于脂肪变性、炎症、气球样和马洛里体、ros积累和线粒体超负荷;纤维化和组织硬化以及细胞死亡。
一方面,所述三维(3d)肝类器官模型是药物诱发的肝脏毒性和炎症/纤维化的模型。
一方面,所述三维(3d)肝类器官模型是肠胃外营养相关肝病(pnald)的模型。
一方面,所述三维(3d)肝类器官模型不包括炎性细胞,例如t细胞或其它炎性分泌蛋白质。
还公开了诱导从ipsc细胞形成肝类器官的方法,其可以用于上述方法中和/或获得前述组合物。所述方法可以包括以下步骤
a)使衍生自ipsc细胞的定形内胚层(de)与fgf途径激活剂和gsk3抑制剂接触足以形成后前肠球状体的时间段,优选地约1天到约3天的时间段,以及b)在视黄酸(ra)存在下培育步骤a的所得后前肠球状体足以形成肝类器官的时间段,优选地约1天到约5天的时间段,优选地约4天。
成纤维细胞生长因子(fgf)是参与血管生成、伤口愈合和胚胎发育的生长因子家族。fgf是肝素结合蛋白,并且已经示出与细胞表面相关联的硫酸乙酰肝素蛋白多糖的相互作用对于fgf信号转导是必需的。本领域普通技术人员将容易理解合适的fgf途径激活剂。示例性fgf途径激活剂包含但不限于:选自由以下组成的组的一种或多种分子:fgf1、fgf2、fgf3、fgf4、fgf10、fgf11、fgf12、fgf13、fgf14、fgf15、fgf16、fgf17、fgf18、fgf19、fgf20、fgf21、fgf22和fgf23。在一些实施例中,靶向与fgf信号传导途径相关联的细胞成分的sirna和/或shrna可以用于激活这些途径。
在一些实施例中,用本文所述的信号传导途径中的所述一种或多种分子以10ng/ml或更高;20ng/ml或更高;50ng/ml或更高;75ng/ml或更高;100ng/ml或更高;120ng/ml或更高;150ng/ml或更高;200ng/ml或更高;500ng/ml或更高;1,000ng/ml或更高;1,200ng/ml或更高;1,500ng/ml或更高;2,000ng/ml或更高;5,000ng/ml或更高;7,000ng/ml或更高;10,000ng/ml或更高;或15,000ng/ml或更高的浓度处理de培养物。在一些实施例中,信号分子的浓度在整个治疗期间保持恒定。在其它实施例中,信号传导途径的分子浓度在治疗过程期间变化。在一些实施例中,将根据本发明的信号传导分子悬浮在包括dmem和胎牛血清丝氨酸(fbs)的培养基中。fbs浓度可以是2%及以上;5%及以上;10%或以上;15%或以上;20%或以上;30%或以上;或50%或以上。本领域技术人员将理解,本文描述的方案适用于本文所述的任何已知的单独或组合的信号传导途径分子,包含但不限于fgf信号传导途径中的任何分子。
本领域普通技术人员将容易理解合适的gsk3抑制剂。示例性gsk3抑制剂包含但不限于:例如chiron/chir99021,其抑制gsk3β。本领域普通技术人员将认识到适合于进行所公开方法的gsk3抑制剂。gsk3抑制剂可以以约1μm到约100μm,或约2μm到约50μm,或约3μm到约25μm的量施用。本领域普通技术人员将容易理解适当的量和持续时间。
一方面,干细胞可以是哺乳动物或人ipsc。
一方面,前肠球状体可以嵌入基底膜基质中,如例如以商品名称matrigel销售的市售基底膜基质。
一方面,肝类器官的特征可以在于肝类器官可以表达甲胎蛋白(afp)、白蛋白(alb)、视黄醇结合蛋白(rbp4)、细胞角蛋白19(ck19)、肝细胞核因子6(hnf6)和细胞色素p4503a4(cyp3a4)、hnf4a、e-钙粘蛋白、dapi和epcam。这种表达可以例如在第40天到第50天发生。表达水平可以类似于在人肝细胞中观察到的表达水平,例如,成人肝细胞的表达水平。
一方面,肝类器官的特征可以在于肝类器官具有胆汁转运活性。
一方面,肝类器官可以衍生自干细胞,并且可以包括腔内结构,其进一步包含内化的微绒毛和间充质细胞。腔内结构可以被极化的肝细胞和基底膜包围。肝类器官可以包括功能性星状细胞和功能性库普弗细胞。
在某些方面,肝类器官的特征可以在于具有以下中的一种或多种:胆汁生产能力、胆汁转运活性、至少50ng/ml/1xe6细胞/24小时的补体因子h表达、至少40ng/ml/1xe6细胞/24小时的补体因子b、至少1000ng/ml/1xe6细胞/24小时的c3表达;至少1000ng/ml/1xe6细胞/24小时的c4表达、至少1,000ng/ml/1xe6细胞/24小时的纤维蛋白原产量以及至少1,000ng/ml/1xe6细胞/24小时的白蛋白产量。一方面,肝类器官的特征可以在于具有至少10,000ng/ml1xe6细胞/24小时的总肝蛋白表达。肝类器官的特征可以在于它可以表达选自以下的一种或多种基因:prox1、rbp4、cyp2c9、cyp3a4、abcc11、cfh、c3、c5、alb、fbg、mrp2、alcam、cd68、cd34、cd31。一方面,肝类器官可以包括包括药物代谢细胞色素变体的细胞,如例如cy2c9*2变体。肝类器官可以包括脉管系统,如us20160177270中描述的脉管系统。
一方面,肝类器官的特征可以在于肝类器官不包括炎性细胞,例如t细胞或其它炎性分泌蛋白质。
实例
实现3d组织的高阶功能的人类器官系统与健康和疾病中的体内器官结构非常相似,但未能捕获更普遍和复杂的病理学,如炎症和纤维化。在这里,申请人开发了多细胞人肝类器官(hlo)模型,其示出了脂肪性肝炎的基本特征。培养脂肪酸暴露使得持续的脂肪变性诱导,然后逐渐激活促炎性和纤维化谱系,这在hlo中发展出大量纤维化。有趣的是,脂肪性肝炎表型的表达受临床报道的可遗传因素的强烈影响。原子力显微镜测量显示整体类器官硬化与炎症和纤维化的严重程度相关。使用三种单基因脂肪性肝炎特异性ipsc系证实了对临床表型的测量保真度。更进一步,申请人在类器官中建立了医源性父母营养相关肝病(pnald)模型。
视黄酸(ra)信号传导是早期器官规范期期间来自前肠内胚层的甲状腺、肺和胰腺的众所周知的重要特征,而对于模型生物中的肝的规范显然不是必需的(kelly和drysdale,2015)除斑马鱼外(negishi等人,2010)。实际上,非洲蟾蜍和小鸡与哺乳动物相似,因为肝规格发生在缺乏ra的胚胎中(chen等人,2004;stafford等人,2004)。相反,一些动物研究表明ra促进肝星状细胞从间充质分化,所述间充质部分受stm和星状细胞中表达的锌指转录因子wt1的控制(ijpenberg等人,2007;wang等人,2013)。另外,一些研究表明平衡的ra调节诱导人干细胞的单核细胞命运规范(purton等人,2000;ronn等人,2015)。此外,通过未知机制,ra信号传导促进了后来的肝芽生长(zorn和wells,2009)。鉴于ra对实质和非实质细胞规格两者的更广泛作用,申请人假设定时ra暴露可能影响包含促炎谱系的肝基质细胞的谱系多样化,从而促进产生用于模拟炎症和纤维化的人肝类器官。
最近,已经报道了通过来自psc的前肠球状体生成定向分化成肠类器官(mccracken等人,2017;spence等人,2011)。这些类器官不仅生成上皮细胞,还共同发育间充质细胞组分。这里,通过利用这种前肠生成方法,申请人测试了假设:进入肝类器官的及时ra脉冲优先地在肝脏规格与人类ipsc之后共同分化支持谱系。通过在4-d条件下培养上皮细胞与支持细胞系之间的紧密相互作用,其中它们可以共同发育,申请人证明肝类器官适用于模拟炎症和纤维化。申请人还建立了筛选平台,通过评估活体状态下单个类器官水平的僵硬度来确定纤维化的严重程度。此方法作为用于研究上皮器官炎症和纤维化以用于药物开发和个体化治疗的宝贵应用平台。
结果
从人ipsc生成基于ra的肝类器官模型
为了阐明ra信号传导是否对确定谱系向基质细胞的分化有影响,申请人通过瞬时诱导ra从人ipsc建立了肝类器官模型。申请人最初通过如前所述的定形内胚层(de)规范将ipsc分化为前肠球状体(spence等人,2011)。将前肠球状体与在相同孔中生长的附着细胞混合,并将混合物包埋在基质胶中。由于ra是通过背景依赖性过程的不同谱系的已知特征,申请人在肝细胞培养基(hcm)中培养肝细胞成熟过程之前设定初始ra信号传导的持续时间,并在第20天表征已建立的类器官,如图1,a中所示。由于在ra的0到5天的不同持续时间中,ra暴露4天观察到培养基中白蛋白的最高浓度,因此在0天与4天-ra暴露的两种条件之间比较已建立的类器官的进一步表征。与未处理的相比,ra处理的孔中类器官的数量增加了1.8倍,并且在ra处理之后,大小增加了1.5倍(图1、b、c和d)。当与未治疗的(0.5μg/ml到3.5μg/ml;图1,e)相比时,ra治疗组(3.0μg/ml到6.5μg/ml)中白蛋白分泌增加两倍。值得注意的是,在第20天首次检测到白蛋白之后,白蛋白分泌维持超过40天(数据未显示)。有趣的是,内部管腔结构出现在95%的ra治疗的类器官中,而仅在12%的未治疗的类器官中检测到,表明类器官的内腔化依赖于ra信号传导(图1,f)。
申请人接下来通过添加二乙酸荧光素,肝细胞中的外排转运标记物,其在肝细胞中的酯酶水解之后变成荧光素进入培养基并跟踪荧光的流动来测试胆汁转运活性。在添加二乙酸荧光素之后几分钟内荧光物质被吸收到类器官细胞中,并且随后从细胞排泄到内腔中(图1,g)。相反,在球状体中未检测到荧光。由于胆汁转运活性是肝的重要功能,因此人肝类器官,以下定义为hlo可以用作代表多种人肝细胞功能的模型,包含白蛋白分泌和胆汁转运功能。
人ipsc-肝类器官中纤维化前和炎症谱系的共分化
有趣的是,ra处理的hlo的单细胞rna测序(scrna-seq)分析示出星状细胞、肝常驻巨噬细胞库普弗细胞和内皮细胞特异性的表达特征,如由星状细胞标记物(acta2、des、pdgfrb)、库普弗细胞标记物(cd68、irf7)和内皮细胞标记物(oit3、dpp4、c1qtnf1;图1,h)所证实。(baharhalpern等人,2017;eltaghdouini等人,2015;vandegarde等人,2016)。为了证实星状细胞和库普弗细胞的存在,申请人通过facs进行了上皮细胞标记物epcam、星状细胞标记物cd166/alcam和库普弗细胞标记物cd68和f4/80的定量分析(yanagimachi等人,2013)。虽然epcam+细胞的频率为hlo的78.85±7.35%,但epcam-cd166+、epcam-cd68+和epcam-f4/80+表达细胞分别为32.4±1.2%、1.69±0.3%和1.68±0.2%(图1,i)。在免疫组织化学后,还在hlo中检测到cd68细胞的表达,并且cd68表达定位于内腔侧的细胞(图1,j)。在hlo中检测到人类肝星状细胞的新标记并且据报道是啮齿动物中星状细胞转分化过程所必需的波形蛋白和gfap(geerts等人,2001;kordes等人,2014),并且在相同细胞中共表达(图1,j),表明星状细胞驻留在hlo中。
为了检查表达cd68的细胞是否在hlo中具有与库普弗细胞类似的功能活性,申请人监测了hlo中的吞噬活性,据报道所述hlo具有活性的肝驻留库普弗细胞,具有ph敏感的基于罗丹明的活细胞染料,如在thp1人巨噬细胞中所观察到的,响应于在高吞噬作用中发生的从高ph到低ph的环境转变,荧光(红色)经历显著增加(图8)。在hlo中,活性被检测并且定位类似于表达cd68的细胞,而在球状体中很少检测到(图1,k)。这些观察结果表明,基于ra的肝类器官(hlo)是一种独特的肝模型,其自然地发展多谱系肝基质细胞,包含来自人ipsc的星状细胞和库普弗细胞。
游离脂肪酸对hlo的炎症反应和纤维化诱导
目前,游离脂肪酸(ffa)被广泛用作建立体外脂毒性肝细胞模型的起始因子;然而,脂肪堆积是唯一的表型(kanuri和bergheim,2013)。肝细胞和库普弗细胞的替代性共培养模型显示炎症反应,如il家族细胞因子、巨噬细胞相关细胞因子和mmp相关细胞因子的表达增加,但不是随后的纤维化(hassan等人,2014)。鉴于肝细胞-、星状-和库普弗样细胞的存在与脂质代谢功能相关基因通过全球转录组分析表明,申请人假设ffa暴露自然引发炎症反应并在hlo中产生纤维化反应。为了测试此假设,申请人用ffa治疗hlo3天到5天。申请人指出3天和5天oa处理的hlo为脂肪性肝炎(s)hlo和肝硬化(c)hlo(图4,a)。shlo用油酸处理hlo三天,并示出脂质积累和炎症。chlo用油酸处理hlo五天,并且除了脂质积聚和炎症外,还示出纤维化(hlo硬化)。首先比较多种ffa对脂质积累的影响,包含油酸(oa,18:1n9)、亚油酸(la,18:2)、棕榈酸(pa,16:0)和硬脂酸(sa,18:0),在ffa暴露之后3天用脂质染料bodipy进行活细胞成像。如图7所示,oa在肝细胞样细胞中诱导脂质积累最有效,并且sa效果最差。考虑到oa引发大量脂质积累,oa的浓度变化(0μm、200μm、400μm和800μm)以引发shlo中的炎症反应。活细胞成像和随后的定量显示shlo中的脂质积累以剂量依赖性方式升高(高达18倍;图4,b和c)。除了脂质积累增加之外,脂滴的大小也增大(图4,b)。肝细胞膨胀(扩大)是确定非酒精性脂肪性肝炎(nash)活性(nas评分)的病理分级指标之一,其通过细胞膜的活体成像在800μmoa治疗的shlo中得到证实(图4b)。甘油三酯,肝中积累的脂质的主要成分也在800μm治疗的shlo中检测到,但在未治疗的shlo中未检测到(图4,d)。更重要的是,培养物上清液的elisa示出,当与未处理的相比时,在800μmoa处理的shlo培养基中il-6分泌2.2倍(图4,e)。il-8和tnf-α也在200μm或800μmoa处理的条件下上调(分别增加10倍和2倍;图4,f)。此外,使用oa处理的shlo调节的培养基,在小室中培养之后24小时评估thp1的迁移,并且在oa处理的shlo中thp-1迁移细胞升高,表明ffa处理在shlo中自然引起炎症反应可能是由对肝细胞样细胞产生应激(高达2倍:图4,g和图8。)为了证实那些炎症和纤维化特征是否对hlo特异,申请人从e-cadmruby胚胎干(es)细胞衍生的类器官分离出上皮标记e-cad,重建来自e-cad阳性或阴性细胞的球状体,并应用于il6和p3np的elisa以及il8和tnf-α的rna表达。如图9所示,e-cad阳性和阴性细胞衍生的球状体均未产生p3np和il-6分泌产生并且过表达炎症标记物的基因表达。申请人还使用人肝细胞系hepg2、巨噬细胞系thp-1和肝星状细胞lx-2测试了相同的实验,并且具有类似的e-cad阳性和阴性细胞的结果。结果表明,肝类器官仅响应ffa治疗炎症和纤维化反应。
为了进一步确定hlo是否进展为纤维化样病状,申请人对shlo和chlo进行masson三色染色。在shlo上未观察到三色阳性hlo,但在chlo上显著增加(图2,h)。此外,上皮标志物epcam和纤维化标志物波形蛋白的if染色显示,在chlo中epcam阳性hlo增加和波形蛋白阳性hlo减少(图2,i),表明oa治疗诱导纤维化群体的选择性增加以及基质沉积在chlo中。此外,还在oa暴露的第5天测量了p3np的elisa。当与未处理的相比时,在800μmoa处理的hlo的培养基中p3np分泌增加2.8倍(图4,j)。总的来说,这些观察结果表明,ffa暴露不仅引起甘油三酯的积累,而且还引发hlo中的炎症反应和纤维化。
在单细胞水平通过ffa处理的炎症和纤维化的chlo谱
通过afm高通量定量纤维化
越来越多的证据已经表明,肝硬度与肝纤维化的严重程度密切相关(yoneda等人,2008),并且因此测量hlo的硬度有可能评估hlo的纤维化严重程度。申请人对平滑肌肌动蛋白免疫染色的初步定性分析表明通过oa暴露的剂量依赖性纤维化进展(图2,h、i和j)。为了以可筛选的形式获得更多的定量见解,申请人因此在活体状态中评估hlo纤维化是否可以使用原子力显微镜(afm)的微压痕以剂量依赖性方式增加刚度。如图4,a中所示,用afm悬臂扫描每个单个hlo的顶部区域(25μm×25μm方形中的14×14矩阵),其可以提供hlo的地形和机械信息的空间映射。单个hlo的计算杨氏模量(e,kpa)的代表性直方图清楚地示出高斯样分布,并且其峰值和宽度根据oa浓度增加(图4,b;0μm、200μm、400μm、800μm)。通过带有盒须图的点图(图4,c)总结了从hlo确定的杨氏模量,证明hlo的刚性从0逐渐升高到800μmoa,即根据oa浓度的升高观察到中值的逐渐变化和刚度范围的扩大。对于0μm、200μm、400μm和800μmoa处理的类器官,emedian值分别为1.2kpa、1.6kpa、2.4kpa和2.8kpa。更进一步,对于未处理的类器官,第90百分位数与第10百分位数(δp90-10)之间的差异为3.2kpa,并且对于800μmoa处理的类器官,观察到7.0kpa。如图3,f所示,tnf-α和il-8也被oa添加上调,已知其与nafld患者中的纤维化严重程度相关(ajmera等人,2017)。这些结果表明,hlo的硬度可以是纤维化严重程度的可量化测量,并且因此可以潜在地应用于纤维化的高通量筛选。
chlo的刚性概括了wolman病的临床表型
由于患者遗传影响的不确定性,与多能干细胞的纤维化和肝硬化相关的模型的临床相关性尚不清楚。在此模型中可能不会捕获纤维化的倾向。因此,申请人已经通过评估包含健康人(317d6)、nafld/nash患者(nafld150、77和27)和wolman病患者(wd90、91和92)的先天性脂肪性肝炎患者衍生的ipsc检查了对临床表型的测定保真度(图5,a)。具体地说,申请人建立了三种wolman病患者特异性ipsc系,其是一种具有致死性脂肪性肝炎的单一遗传性病症,并证实相对于正常ipsc系衍生的类器官显著的刚性增加,特别是与临床诊断时的酶活性显著相关(图5,b)。
使用ipsc-shlo模拟脂肪变性的人类表型变异。
其次,申请人扩展了这种新的类器官模型,以评估对脂肪性肝炎进展的多基因效应。三种蛋白质编码测序变体pnpla3(patatin样磷脂酶3)p.i148m、tm6sf2(跨膜6超家族成员2)p.e167k和gckr(葡萄糖激酶调节蛋白)p.p446l,已经被反复证明是通过大规模gwas的肝甘油三酯含量(htgc)的独立决定因素(xu等人,2015),(zain等人,2015)(图6,a)。因此,申请人接近大规模细胞库,并通过报告的效应大小β(snp)的总和绘制出具有多基因评分的2504细胞系(图6,b)(stender等人,2017),其中标准化htgc乘法加权剂量中每个等位基因的变化如下:∑{β(snp)×(风险等位基因剂量)}。在获得具有三个不同阈值的ipsc系之后(图6,b和c),申请人已经生成了七个ipsc类器官,并诱导了脂肪性肝炎以估计多基因对表型的影响。引人注目的是,申请人已经发现了“脂肪变性”(通过活体成像,图6,d和e)与“炎症”(qrt-pcr,图6,f)的显著相关性,但不是“纤维化”(基于afm的刚度测量,图6,g)到多基因评分。申请人还发现oca反应在细胞系中不同,并且对oca的反应取决于snp数(图6h)。
讨论
来自人psc的多细胞肝类器官
最近的一系列研究报道,通过实验性组合内皮细胞(takebe等人,2013)、间充质细胞(takebe等人,2015)和神经嵴细胞(workman等人,2017),将支持性谱系成功整合到内胚层衍生的类器官中。基于ra脉动的方法自然地使细胞以持续的细胞极性进行多样化,并且以合理的成本显著地可再现和可扩展。支持性谱系的确切性质是否与发育相关或肝特异性仍然是难以捉摸的,基质群体对已知的纤维化诱导物,包含lps和脂肪酸具有完全反应性,为模拟多细胞和复杂病理学铺就了新的方法。
用于分析炎症和纤维化的机械类器官筛查的前景
从未来的筛选角度来看,基于单一类器官的肝测量由于其稳健性、标准化和相对简单性而是非常有吸引力的读数。例如,基于类器官的活荧光成像分析,建立了囊性纤维化患者肠道类器官的功能性肿胀测定,证明了有效的药物选择(saini,2016)。来自ipsc的单个肝类器官的活力刚性评估是预测纤维化严重程度的有效方法。肝硬度测量与纤维化阶段之间的显著相关性通过弹性成像在脂肪肝病患者中临床报道(yoneda等人,2008)。此外,在具有超声波定义的nafld的受试者亚组中观察到增加的肝硬度与糖尿病(dm)的存在和/或更大的胰岛素抗性之间的强关联(koehler等人,2016)。有趣的是,人肝类器官(hlo)硬度与lps和ffa两者的比例均增加,伴有炎性细胞因子的产生和成纤维细胞的扩张。考虑一些上皮器官纤维化通过不同病理机制共享类似的表型,基于类器官刚度检测测定可以用于使用肺、肾、心脏和消化道类器官分析纤维化。
在肝类器官中,可以使用高含量成像系统对多次测量进行玻璃体内评估。事实上,脂肪滴的数量和大小的增加、胆汁转运活性、吞噬细胞和细胞形态的恶化可以在脂肪处理的肝类器官中有效地成像。因此,基于类器官的测定平台可以用于更好地理解脂肪肝病的人特异性机制,并为这些疾病的药物发现建立高含量筛选。组合化合物库和营养代谢物,基于类器官的机械筛选将是非常有吸引力的模型系统,用于暗示人类的有效治疗,否则在临床前不可测试。
营养精准医学(治疗个性化)
患者特异性ipsc衍生的类器官可以用于预测个体化药物功效和上皮反应。可以从健康和患病个体两者建立人ipsc。与建立群体ipsc小组平行,预期患者的ipsc的使用将模拟脂质毒性、药物功效和安全性问题的个体间差异(warren等人,2017a;warren等人,2017b)。在这样做的过程中,使用基于肝类器官的平台进行的表型筛选将促进针对病症的高效干预的个性化选择。例如,由于营养补充剂可能根据个体而改变,因此所公开的系统可以是用于在施用之前反映营养相关病状的高度相容的测定。具体而言,患者特异性ipsc衍生的类器官可以用于为每位患者定制pn配制品,目的是最小化肝脏脂肪变性和纤维化(pnald)的可能进展。实际上,在诊所中,pn配制品通常是定制的(mercaldi等人,2012),因为商业解决方案不能满足重症患者的热量、氨基酸和电解质需求,这些患者通常肥胖且需要液体限制,并且显示肝脏/肾功能障碍(boullata等人,2014)。除了营养需求之外,迫切需要获得有关减少与pn产品相关联的可能副作用的见解。因此,肝类器官可以用于评估安全性问题,特别是对于pnald风险评估,促进每个患者的pn配制品的定制策略。
总体而言,申请人已经证明,在4-d类器官培养物中实现上皮与基质谱系之间的串扰可以用于模拟与脂肪变性和肝纤维化相关联的临床相关病理学。此模型最终将导致对更常见的病理学分析,如nafld或nash,这是人口老龄化日益增长的问题。更广泛地说,申请人已经建立了一种可行的策略,用于模拟人文复杂病理学,结合当前正在发展的类器官技术(lancaster和knoblich,2014),为在单个类器官水平上发现针对不可恢复疾病的有效治疗铺就了新方法。
方法
hpsc维护。本研究中使用的tkda3人ipsc克隆由k.eto和h.nakauchi友情提供。如前所述维持人ipsc系(takebe等人,2015;takebe等人,2014)。在1/30稀释度、37℃、5%co2和95%空气下,将未分化的hipsc在mtesr1培养基(stemcelltechnologies,vancouver,canada)中在无饲养细胞条件下维持在涂有基质胶(corninginc.,ny,usa)的板上。
定形内胚层诱导。使用先前描述的方法稍微修改将人ipsc分化为定形内胚层(spence等人,2011)。简言之,在accutase(thermofisherscientificinc.,ma,usa)中分离人ipsc的菌落,并将150,000个细胞/ml接种在基质胶包被的组织培养板(vwrscientificproducts,westchester,pa)上。在第1天将培养基更换为含有100ng/ml激活素a(r&dsystems,mn,usa)和50ng/ml骨形态发生蛋白4(bmp4;r&dsystems)的rpmi1640培养基(lifetechnologies),在第2天将培养基更换为含有100ng/ml激活素a和0.2%胎牛血清(fcs;thermofisherscientificinc.)的培养基,并且在第3天将培养基更换为含有100ng/ml激活素a和2%fcs的培养基。在第4天到第6天,将细胞在含有500ng/ml成纤维细胞生长因子(fgf4;r&dsystems)的b27(lifetechnologies)和n2(gibco,ca,usa)和3μmchir99021(stemgent,ma,usa)的advanceddmem/f12(thermofisherscientificinc.)中培养。将细胞维持在37℃,5%co2和95%空气中,并且每天更换培养基。在分化的第7天,球状体出现在平板上。
hlo诱导。在第7天,轻轻移取球状体和附着的细胞以从培养皿分层。将它们以800rpm离心3分钟,在去除上清液之后,包埋在含有b27、n2和2μm视黄酸(ra;sigma,mo,usa)的advanceddmem/f12中的培养皿上的100%基质胶滴中,并培养4天。在ra处理之后,将嵌入基质胶滴中的球状体在具有10ng/ml肝细胞生长因子(hgf;peprotech,nj,usa),0.1μm地塞米松(dex;sigma)和20ng/ml制瘤素m(osm;r&dsystems)的肝细胞培养基(hcmlonza,md,usa)中培养。将用于hlo诱导的培养物维持在37℃,5%co2和95%空气中,并且每2天到每3天更换培养基。为了分析hlo(第20天到第30天),通过刮擦和移液从基质胶分离出类器官。
白蛋白、il-6和p3npelisa。为了测量hlo的白蛋白分泌水平,从嵌入基质胶中的hlo收集200μl培养物上清液。对于il-6和p3np,接种20个到30个类器官并在超低附着多孔板96孔板(corning)上培养。为了限定每个孔中确切的类器官数量,并且最后用数字标准化il-6和p3np的分泌水平,在keyencebz-x710荧光显微镜上捕获类器官。在培养之后24小时(对于白蛋白)、96小时(对于il-6)和120小时(p3np)时间点收集培养物上清液并储存在-80℃直到使用。将上清液以1,500rpm离心3分钟并沉淀碎片,并根据制造商的说明书,用人白蛋白elisa定量装置(bethyllaboratories,inc.,tx,usa)、人il-6elisa试剂盒(biolegend,ca,usa)和人n-末端前胶原iii前肽piiinpelisa试剂盒(mybiosource,ca,usa)测定所得上清液。通过student'st测试进行显著性测试。
胆汁转运活性。二乙酸荧光素用于评估类器官中的胆汁转运活性。将10mg/ml二乙酸荧光素(sigma)加入用hlo培养的hcm培养基中并静置5分钟并使用荧光显微镜bz-x710(keyence,osaka,japan)捕获。
吞噬细胞、脂质、ros活细胞成像。在超低附着6多孔板中培养之后,挑取5个到10个hlo并接种在microslide8wellglassbottomplate(ibidi,wi,usa)中并进行活细胞染色。使用以下抗体:用于吞噬细胞活性的
he染色和免疫组织化学。从基质胶分离hlo并在4%多聚甲醛中固定并包埋在石蜡中。切片进行he和免疫组织化学染色。使用以下一级抗体:抗α平滑肌肌动蛋白抗体(1:200稀释;abcam,cambridge,uk)、desmin抗体(预稀释;roche,basel,switzerland)和cd68抗体(1:200稀释;abcam)。
流式细胞术。从10个基质胶液滴分离hlo并用1×pbs洗涤。通过用胰蛋白酶-edta(0.05%)、酚红(gibco)处理10分钟将hlo解离成单细胞。pbs洗涤之后,用bv421缀合的epcam抗体(biolegend)、pe缀合的cd166抗体(ebioscience,ca,usa)和pe/cy7-cd68(ebioscience)对单细胞进行流式细胞术。通过碘化丙锭染色测量dna。
lps和ffa暴露以及oca和fgf19治疗。将已经从基质胶分离并用1×pbs洗涤的20个到30个hlo分成每种条件,并在超低附着6多孔板(corning)上培养。hlo与lps(sigma)、oa(sigma)、la(sigma)、sa(sigma)或pa(sigma)一起培养,并在培养之后的第1天和第3天(对于lpshlo)以及第3天和第5天(对于oa)收集。为了测试oca(int-747,medchemexpress,nj,usa)和人fgf19重组体(sigma)对hlo的抑制作用,在油酸(800μm)存在或不存在下,在hcm培养基中培养20个到30个hlo,并且将1μmoca和40ng/mlfgf19加入800μmoa条件中。在第3天收集hlo用于脂质活细胞成像,并在第5天收集用于刚度测量的hlo。
整体免疫荧光。将hlo在4%多聚甲醛中固定30分钟,并用0.5%nonidetp-40透化15分钟。用1xpbs洗涤hlo三次,并在室温下用封闭缓冲液培育1小时。然后将hlo与一级抗体培育;抗α平滑肌肌动蛋白抗体(1:50稀释;abcam)在4℃过夜。用1xpbs洗涤hlo,并在封闭缓冲液中在二级抗体中在室温下培育30分钟。使用具有dapi(abcam)的氟屏蔽封固剂洗涤和安装hlo。将染色的hlo可视化并在nikona1倒置共聚焦显微镜(日本)上使用60×水浸物镜扫描。
rna分离,rt-qpcr。使用rneasymini试剂盒(qiagen,hilden,germany)分离rna。使用superscriptiiifirst-strandsynthesissystemforrt-pcr(invitrogen,ca,usa)根据制造商的方案进行逆转录。在quantstudio3实时pcr系统(thermofisherscientificinc.)上使用taqman基因表达主混合物(appliedbiosystems)进行qpcr。每个靶基因的所有引物和探针信息均自universalprobelibraryassaydesigncenter(https://qpcr.probefinder.com/organism.jsp)获得。通过student'st测试进行显著性测试。
afm的hlo刚度测量。用0ng/ml、50200ng/ml、1400ng/ml、2800ng/mlμmlpsoa处理的hlo用于afm(nanowizardiv,jpkinstruments,germany)的硬度测量。将具有氮化硅悬臂(csc37,k=0.3n/m,mikromasch,bulgaria)的afm头安装在荧光立体显微镜(m205fa,leica,germany)上,并与z轴压电平台(jpkcellhesionmodule,jpkinstruments,germany)联接,其允许压痕测量深度达~100μm。作为类器官的底物,使用涂有纤连蛋白的培养皿。首先将组织培养皿(
thp-1细胞迁移测定。将由t.suzuki赠送的thp-1细胞维持在含有10%fbs的advanceddmem/f12(thermofisherscientificinc.)中。收集thp-1漂浮细胞,并将200,000个细胞与无血清的advanceddmem/f12一起加入到cytoselecttm96孔细胞迁移测定(5μm,fluorometricformat;cellbiolabs,ca,usa)的膜室中。将10个到20个hlo在包含0um、400um、800umoa的hcm培养基中与超低附着96多孔板(corning)一起培养三天。为了限定每个孔中确切的类器官数量,并且最后用数字标准化最终迁移的细胞,在keyencebz-x710荧光显微镜上捕获类器官。收集150μlhlo培养上清液并加入试剂盒的进料盘。将试剂盒在5%co2细胞培养箱中于37℃培育24小时。使用countessiiflautomatedcellcounter(thermofisherscientificinc.)计数已经迁移的细胞。通过student'st测试进行显著性测试。
甘油三酯测定。为了定量确定甘油三酯,从一个基质胶滴分离hlo并在油酸(800μm)存在或不存在下分成hcm培养基。将它们在超低附着6多孔板上培养三天。使用enzychrom甘油三酯测定试剂盒(bioassaysystems,ca,usa)通过酶促测定甘油三酯质量来进行肝甘油三酯积累的定量估计。
hlo存活测定。从基质胶收集hlo并用1×pbs洗涤。在超低附着6多孔板(corning)上培养30个到40个类器官。每天在keyencebz-x710荧光显微镜上捕获hlo。从照片中手动计数存活和死亡的类器官。具有圆形构造的hlo被计为存活,而变形的类器官被计为死亡。为了在相同时间点评估oa处理的hlo的存活率,使用3d细胞滴度glo测定法(promega,wi,usa)。
统计和再现性。使用未配对的双尾student'st-测试,dunn-holland-wolfe测试或welch'st测试进行统计学分析。结果示出平均值±s.e.m.;p值<0.05被认为具有统计学意义。除非另有说明,否则n值是指生物学上独立的重复。
参考文献
ajmera,v.、perito,e.r.、bass,n.m.、terrault,n.a.、yates,k.p.、gill,r.、loomba,r.、diehl,a.m.、aouizerat,b.e.和network,n.c.r.(2017).“新型血浆生物标志物与非酒精性脂肪性肝病成人肝病严重程度相关(novelplasmabiomarkersassociatedwithliverdiseaseseverityinadultswithnonalcoholicfattyliverdisease)”,《肝脏学(hepatology)》65,65-77。
baharhalpern,k.、shenhav,r.、matcovitch-natan,o.、toth,b.、lemze,d.、golan,m.、massasa,e.e.、baydatch,s.、landen,s.、moor,a.e.等人(2017).“单细胞空间重建揭示了哺乳动物肝的全球分工(single-cellspatialreconstructionrevealsglobaldivisionoflabourinthemammalianliver)”,《自然(nature)》542,352-356。
boullata,j.i.、gilbert,k.、sacks,g.、labossiere,r.j.、crill,c.、goday,p.、kumpf,v.j.、mattox,t.w.、plogsted,s.、holcombe,b.等人(2014).“a.s.p.e.n.临床指南:肠外营养订购、订单审查、复合、标签和配药(a.s.p.e.n.clinicalguidelines:parenteralnutritionordering,orderreview,compounding,labeling,anddispensing)”,《肠外和肠内营养杂志(jpenjparenterenteralnutr)》38,334-377。
chen,y.、pan,f.c.、brandes,n.、afelik,s.、solter,m.和pieler,t.(2004).“视黄酸信号传导对于胰腺发育是必需的,并且以非洲爪蟾的外分泌细胞分化为代价促进内分泌(retinoicacidsignalingisessentialforpancreasdevelopmentandpromotesendocrineattheexpenseofexocrinecelldifferentiationinxenopus)”,《发育生物学(devbiol)》,271,144-160。
dash,a.、figler,r.a.、blackman,b.r.、marukian,s.、collado,m.s.、lawson,m.j.、hoang,s.a.、mackey,a.j.、manka,d.、cole,b.k.等人(2017).“临床相关浓度的奥贝胆酸在器官型人肝细胞系统中的药理毒理学(pharmacotoxicologyofclinically-relevantconcentrationsofobeticholicacidinanorganotypichumanhepatocytesystem)”,《活体外毒理学(toxicolinvitro)》39,93-103。
elkasmi,k.c.、anderson,a.l.、devereaux,m.w.、vue,p.m.、zhang,w.、setchell,k.d.、karpen,s.j.和sokol,r.j.(2013).“植物甾醇促进肠外营养相关肝病中的肝损伤和库普弗细胞活化(phytosterolspromoteliverinjuryandkupffercellactivationinparenteralnutrition-associatedliverdisease)”,《科学转化医学(scitranslmed)》5,206ra137。
eltaghdouini,a.、najimi,m.、sancho-bru,p.、sokal,e.和vangrunsven,l.a.(2015).“活化的原代人肝星状细胞的体外逆转(invitroreversionofactivatedprimaryhumanhepaticstellatecells)”,《纤维发生组织修复(fibrogenesistissuerepair)8,14。
geerts,a.、eliasson,c.、niki,t.、wielant,a.、vaeyens,f.和pekny,m.(2001).“在小鼠肝星状细胞中形成正常的结蛋白中间丝需要波形蛋白(formationofnormaldesminintermediatefilamentsinmousehepaticstellatecellsrequiresvimentin)”,《肝脏学(hepatology)》33,177-188。
hassan,w.、rongyin,g.、daoud,a.、ding,l.、wang,l.、liu,j.和shang,j.(2014).“降低的氧化应激有助于异槲皮苷在游离脂肪酸诱导的肝细胞中的降脂作用(reducedoxidativestresscontributestothelipidloweringeffectsofisoquercitrininfreefattyacidsinducedhepatocytes)”,《氧化药物和细胞寿命(oxidmedcelllongev)》2014,313602。
hynds,r.e.和giangreco,a.(2013).“简要回顾:人类干细胞衍生的类器官模型与上皮转化医学的相关性(concisereview:therelevanceofhumanstemcell-derivedorganoidmodelsforepithelialtranslationalmedicine)”,《干细胞(stemcells)》31,417-422。
ijpenberg,a.、perez-pomares,j.m.、guadix,j.a.、carmona,r.、portillo-sanchez,v.、macias,d.、hohenstein,p.、miles,c.m.、hastie,n.d.和munoz-chapuli,r.(2007).“wt1和视黄酸信号传导对星状细胞发育和肝形态发生至关重要(wt1andretinoicacidsignalingareessentialforstellatecelldevelopmentandlivermorphogenesis)”,《发育生物学(devbiol)》,312,157-170。
ito,k.、sakuma,s.、kimura,m.、takebe,t.、kaneko,m.和arai,f.(2016).“基于力传感器探头的微流控芯片huvec/msc球体力学特性的时间转变(temporaltransitionofmechanicalcharacteristicsofhuvec/mscspheroidsusingamicrofluidicchipwithforcesensorprobes)”,《微型机械,巴塞尔(micromachines-basel)》7。
kanuri,g.和bergheim,i.(2013).“非酒精性脂肪性肝病(nafld)的体外和体内模型(invitroandinvivomodelsofnon-alcoholicfattyliverdisease(nafld))”,《国际分子科学学报(intjmolsci)》14,11963-11980。
kelly,g.m.和drysdale,t.a.(2015).“视黄酸与内胚层的发育(retinoicacidandthedevelopmentoftheendoderm)”,《发育生物学期刊(jdevbiol)》,3,25-56。
koehler,e.m.、plompen,e.p.、schouten,j.n.、hansen,b.e.、darwishmurad,s.、taimr,p.、leebeek,f.w.、hofman,a.、stricker,b.h.、castera,l.等人(2016).“糖尿病和脂肪变性的存在与一般人群的肝硬度有关:鹿特丹研究(presenceofdiabetesmellitusandsteatosisisassociatedwithliverstiffnessinageneralpopulation:therotterdamstudy)”,《肝脏学(hepatology)》63,138-147。
kordes,c.、sawitza,i.、gotze,s.、herebian,d.和haussinger,d.(2014).“肝星状细胞有助于祖细胞和肝再生(hepaticstellatecellscontributetoprogenitorcellsandliverregeneration)”,《临床研究杂志(jclininvest)》124,5503-5515。
kumar,j.a.和teckman,j.h.(2015).“全胃肠外营养诱导病理机制的争议(controversiesinthemechanismoftotalparenteralnutritioninducedpathology)”,《儿童(巴塞尔)(children(basel))》2,358-370。
lancaster,m.a.和knoblich,j.a.(2014).“培养皿中的器官发生:使用类器官技术模拟发育和疾病(organogenesisinadish:modelingdevelopmentanddiseaseusingorganoidtechnologies)”,《科学(science)》345,1247125。
mccracken,k.w.、aihara,e.、martin,b.、crawford,c.m.、broda,t.、treguier,j.、zhang,x.、shannon,j.m.、montrose,m.h.和wells,j.m.(2017).“wnt/β-连环蛋白促进小鼠和人的胃底指标(wnt/beta-cateninpromotesgastricfundusspecificationinmiceandhumans)”,《自然(nature)》541,182-187。
mercaldi,c.j.、reynolds,m.w.和turpin,r.s.(2012).“在回顾性医院索赔数据库中识别和比较医院复合和预混多腔袋的肠外营养的方法(methodstoidentifyandcompareparenteralnutritionadministeredfromhospital-compoundedandpremixedmultichamberbagsinaretrospectivehospitalclaimsdatabase)”,《肠外和肠内营养杂志(jpenjparenterenteralnutr)》36,330-336。
nandivada,p.、carlson,s.j.、chang,m.i.、cowan,e.、gura,k.m.和puder,m.(2013).“治疗肠外营养相关性肝病:脂质乳剂的作用(treatmentofparenteralnutrition-associatedliverdisease:theroleoflipidemulsions)”,《食品和营养研究进展(advnutr)》4,711-717。
negishi,t.、nagai,y.、asaoka,y.、ohno,m.、namae,m.、mitani,h.、sasaki,t.、shimizu,n.、terai,s.、sakaida,i.等人(2010).“视黄酸信号通过诱导青霉素中的wnt2bb基因表达而正调节肝规格(retinoicacidsignalingpositivelyregulatesliverspecificationbyinducingwnt2bbgeneexpressioninmedaka)”,《肝脏学(hepatology)》51,1037-1045。
neuschwander-tetri,b.a.、loomba,r.、sanyal,a.j.、lavine,j.e.、vannatta,m.l.、abdelmalek,m.f.、chalasani,n.、dasarathy,s.、diehl,a.m.、hameed,b.等人(2015).“法氏x核受体配体奥贝胆酸用于非肝硬化、非酒精性脂肪性肝炎(flint):一项多中心、随机、安慰剂对照试验(farnesoidxnuclearreceptorligandobeticholicacidfornon-cirrhotic,non-alcoholicsteatohepatitis(flint):amulticentre,randomised,placebo-controlledtrial)”,《柳叶刀(lancet)》385,956-965。
orso,g.、mandato,c.、veropalumbo,c.、cecchi,n.、garzi,a.和vajro,p.(2016).“小儿肠外营养相关性肝病和胆汁淤积:基于病理机制的预防和治疗的新进展(pediatricparenteralnutrition-associatedliverdiseaseandcholestasis:noveladvancesinpathomechanisms-basedpreventionandtreatment)”,《消化和肝疾病(digliverdis)》48,215-222。
purton,l.e.、bernstein,i.d.和collins,s.j.(2000).“全反式视黄酸增强培养的造血干细胞的长期再增殖活性(all-transretinoicacidenhancesthelong-termrepopulatingactivityofculturedhematopoieticstemcells)”,《血液(blood)》95,470-477。
ronn,r.e.、guibentif,c.、moraghebi,r.、chaves,p.、saxena,s.、garcia,b.和woods,n.b.(2015).“视黄酸调节人多能干细胞的造血发育(retinoicacidregulateshematopoieticdevelopmentfromhumanpluripotentstemcells)”,《干细胞报告(stemcellreports)》4,269-281。
saini,a.(2016).“囊性纤维化患者受益于迷你肠道(cysticfibrosispatientsbenefitfromminiguts)”,《干细胞(cellstemcell)》19,425-427。
spence,j.r.、mayhew,c.n.、rankin,s.a.、kuhar,m.f.、vallance,j.e.、tolle,k.、hoskins,e.e.、kalinichenko,v.v.、wells,s.i.、zorn,a.m.等人(2011).“人体多能干细胞体外定向分化为肠组织(directeddifferentiationofhumanpluripotentstemcellsintointestinaltissueinvitro)”,《自然(nature)》470,105-109。
stafford,d.、hornbruch,a.、mueller,p.r.和prince,v.e.(2004).“视黄酸信号传导在脊椎动物胰腺发育中的保守作用(aconservedroleforretinoidsignalinginvertebratepancreasdevelopment)”,《发育基因与进化(devgenesevol)》214,432-441。
stender,s.、kozlitina,j.、nordestgaard,b.g.、tybjaerg-hansen,a.、hobbs,h.h.和cohen,j.c.(2017).“肥胖可以放大多个位点赋予的脂肪肝病的遗传风险(adiposityamplifiesthegeneticriskoffattyliverdiseaseconferredbymultipleloci)”,《自然遗传学(natgenet)》49,842-847。
takebe,t.、enomura,m.、yoshizawa,e.、kimura,m.、koike,h.、ueno,y.、matsuzaki,t.、yamazaki,t.、toyohara,t.、osafune,k.等人(2015).“来自不同组织的血管化和复杂器官芽通过间充质细胞驱动的冷凝(vascularizedandcomplexorganbudsfromdiversetissuesviamesenchymalcell-drivencondensation)”,《干细胞(cellstemcell)》16,556-565。
takebe,t.、sekine,k.、enomura,m.、koike,h.、kimura,m.、ogaeri,t.、zhang,r.r.、ueno,y.、zheng,y.w.、koike,n.等人(2013).“来自ipsc衍生的器官芽移植物的血管化和功能性人肝(vascularizedandfunctionalhumanliverfromanipsc-derivedorganbudtransplant)”,《自然(nature)》499,481-484。
takebe,t.、zhang,r.r.、koike,h.、kimura,m.、yoshizawa,e.、enomura,m.、koike,n.、sekine,k.和taniguchi,h.(2014).“从ipsc衍生的器官芽移植物产生血管化和功能性人肝(generationofavascularizedandfunctionalhumanliverfromanipsc-derivedorganbudtransplant)”,《自然实验手册(natprotoc)》9,396-409。
tsedensodnom,o.和sadler,k.c.(2013).“ros:脂肪肝病中的还原和悖论(ros:reduxandparadoxinfattyliverdisease)”,《肝脏学(hepatology)》58,1210-1212。
vandegarde,m.d.、movita,d.、vanderheide,m.、herschke,f.、dejonghe,s.、gama,l.、boonstra,a.和vanwolleghem,t.(2016).“肝单核细胞和枯否细胞在慢性病毒感染期间保持转录不同(livermonocytesandkupffercellsremaintranscriptionallydistinctduringchronicviralinfection)”,《公共科学图书馆期刊(plosone)》11,e0166094。
wang,y.、li,j.、wang,x.、sang,m.和ho,w.(2013).“肝星状细胞、肝先天免疫和丙型肝炎病毒(hepaticstellatecells,liverinnateimmunity,andhepatitiscvirus)”,《肠胃病学与肝病期刊(jgastroenterolhepatol)》28suppl1,112-115。
warren,c.r.、jaquish,c.e.和cowan,c.a.(2017a).“nextgen遗传协会研究联盟:进军体外人口遗传学(thenextgengeneticassociationstudiesconsortium:aforayintoinvitropopulationgenetics)”,《干细胞(cellstemcell)》20,431-433。
warren,c.r.、o'sullivan,j.f.、friesen,m.、becker,c.e.、zhang,x.、liu,p.、wakabayashi,y.、morningstar,j.e.、shi,x.、choi,j.等人(2017b).“诱导多能干细胞分化使gwas变体在代谢疾病中的功能验证成为可能(inducedpluripotentstemcelldifferentiationenablesfunctionalvalidationofgwasvariantsinmetabolicdisease)”,《干细胞(cellstemcell)》20,547-557e547。
workman,m.j.、mahe,m.m.、trisno,s.、poling,h.m.、watson,c.l.、sundaram,n.、chang,c.f.、schiesser,j.、aubert,p.、stanley,e.g.等人(2017).“具有功能性肠神经系统的工程化的人多能干细胞衍生的肠组织(engineeredhumanpluripotent-stem-cell-derivedintestinaltissueswithafunctionalentericnervoussystem)”,《自然医学(natmed)》23,49-59。
xu,r.、tao,a.、zhang,s.、deng,y.和chen,g.(2015).“含有3个基因的马铃薯糖蛋白样磷脂酶结构域(pnpla3)多态性与非酒精性脂肪性肝病之间的关联:一项巨大的综述和荟萃分析(associationbetweenpatatin-likephospholipasedomaincontaining3gene(pnpla3)polymorphismsandnonalcoholicfattyliverdisease:ahugereviewandmeta-analysis)”,《科技报告(scirep)》5,9284。
yanagimachi,m.d.、niwa,a.、tanaka,t.、honda-ozaki,f.、nishimoto,s.、murata,y.、yasumi,t.、ito,j.、tomida,s.、oshima,k.等人(2013).“在血清和饲养细胞条件下,功能性单核细胞与人多能干细胞的稳健和高效分化(robustandhighly-efficientdifferentiationoffunctionalmonocyticcellsfromhumanpluripotentstemcellsunderserum-andfeedercell-freeconditions)”,《公共科学图书馆期刊(plosone)》8,e59243。
yoneda,m.、yoneda,m.、mawatari,h.、fujita,k.、endo,h.、iida,h.、nozaki,y.、yonemitsu,k.、higurashi,t.、takahashi,h.等人(2008).“通过测量非酒精性脂肪性肝病(nafld)患者的僵硬度对肝纤维化进行无创评估(noninvasiveassessmentofliverfibrosisbymeasurementofstiffnessinpatientswithnonalcoholicfattyliverdisease(nafld))”,《消化和肝疾病(digliverdis)》40,371-378。
zain,s.m.、mohamed,z.和mohamed,r.(2015).“葡萄糖激酶调节基因rs780094的常见变异和非酒精性脂肪肝疾病的风险:荟萃分析(commonvariantintheglucokinaseregulatorygeners780094andriskofnonalcoholicfattyliverdisease:ameta-analysis)”,《肠胃病学与肝病期刊(jgastroenterolhepatol)》30,21-27。
zambrano,e.、el-hennawy,m.、ehrenkranz,r.a.、zelterman,d.和reyes-mugica,m.(2004).“全肠外营养诱导的肝病理学:尸检系列24例新生儿病例(totalparenteralnutritioninducedliverpathology:anautopsyseriesof24newborncases)”,《儿童与发育病理学(pediatrdevpathol)》,7,425-432。
zorn,a.m.和wells,j.m.(2009).“脊椎动物内胚层发育和器官形成(vertebrateendodermdevelopmentandorganformation)”,《细胞与发育生物学年度回顾(annurevcelldevbiol)》25,221-251。
- 还没有人留言评论。精彩留言会获得点赞!