合成的免疫受体及其使用方法与流程
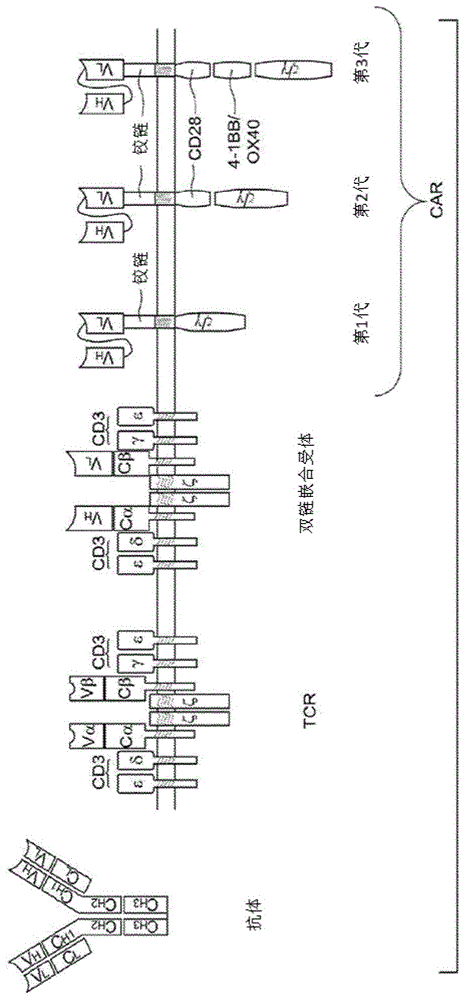
本申请依据35u.s.c.§119要求于2016年12月2日提交的美国临时专利申请序列号62/429,619和2016年12月2日提交的美国临时专利申请号62/429,597的优先权,这些临时专利申请的公开内容通过引用整体并入本文。本发明涉及合成免疫受体(sir)多肽、多核苷酸、表达构建体和工程化以表达合成免疫受体(sir)的免疫效应细胞(例如t细胞、nkt细胞)和干细胞的用途。本公开还提供了使用此类多肽、多核苷酸、表达构建体和重组细胞来治疗疾病和病症的方法,包括但不限于癌症、感染疾病、过敏性疾病、自身免疫疾病、退行性疾病或上述的组合。通过引用序列表并入本文件随附名称为“sequence_st25.txt”的序列表,该序列表在2017年12月2日创建,并具有77,085,242字节的数据,在ibm-pc、ms-windows操作系统上机器格式化。出于所有目的,该序列表通过引用整体并入本文。
背景技术:
:活化免疫细胞以选择性识别并破坏肿瘤的策略,即癌症免疫疗法提供了癌症疗法的有力方案。使用肿瘤特异性t细胞和嵌合抗原受体(car)修饰t细胞(car-t细胞)的过继转移的免疫疗法在一些患有转移癌症的患者中介导了持久且完全的疾病消退。尽管使用car-t细胞有所成功,但是此方法存在若干限制。在响应于工程化cart细胞的大多数患者中,促炎性细胞因子的过量释放引起包括发烧、血压过低、血氧过少、心功能障碍、肾衰竭和电解质异常情况(总称为“细胞因子释放综合征”(crs))的症状。在一些情况下,car疗法可导致神经性症状,包括震颤、癫痫,并且可能为致命的。抵消crs的策略包括使用免疫抑制剂和细胞因子的抗体阻断细胞因子释放来进行治疗。另外,成功癌症免疫疗法的最重要挑战之一是遗传修饰t细胞以在转移之后持续超过若干个月。这已证明对于用car基因修饰的t细胞而言为更大挑战。对car构建体进行最新分子工程化以包括共刺激结构域cd28或41bb,导致持久性得到改善。然而,在car构建体中包含共刺激结构域导致通过受体进行非生理信号传导。一些car显示紧张性抗原独立性信号传导,其导致不受限的细胞活化,最终导致细胞凋亡、独立于同源抗原的过量细胞因子释放、以及免疫学耗尽。一些含有cd28和cd3z串联信号传导结构域的car的表达导致组成性活化和转导原代人类t细胞的增殖,该增殖与低级体内功效相关(frigault等人,2015)。发现导致具有连续t细胞增殖的car的表型的一种机制是在细胞表面处的高密度car(frigault等人,2015)。在t细胞表面上表达t细胞受体(tcr)。在人类中,这些受体识别在人类白细胞抗原(hla)分子与抗原肽之间形成的复合物。这些肽的识别导致t细胞免疫功能活化。在大部分t细胞中,tcr是阿尔法(α)和贝塔(β)链的异二聚体。β链具有两种亚型:cβ2(在80%人类t细胞中)和cβ1(在20%人类t细胞中)。tcr的每条链包含n末端免疫球蛋白(ig)样可变(v)结构域和ig样恒定(c)结构域,该结构域进而包含跨膜区和c末端处的短胞质尾部。技术实现要素:本公开提供了至少一种重组多核苷酸,其编码至少一种合成免疫受体(sir),该至少一种sir包含(a)具有选自由以下组成的组中的氨基酸序列的t细胞受体(tcr)恒定链:(i)与seqidno:3010至少98%一致并在位置48、61、91、92、93和/或94处具有一个或多个突变并且可以包含任选辅助模块的氨基酸序列;(ii)与seqidno:3024至少98%一致并在位置18、22、57、79、133、136和/或139处具有一个或多个突变并且可以包含任选辅助模块的氨基酸序列;(iii)与seqidno:3025至少98%一致并在位置18、22、57、79、133、136和/或139处具有一个或多个突变并且可以包含任选辅助模块的氨基酸序列;(iv)与seqidno:3046、3047或3048至少98%一致并且可以包含任选辅助模块的氨基酸序列;(v)与seqidno:3049至少98%一致并且可以包含任选辅助模块的氨基酸序列;(vi)与seqidno:3051或3052至少98%一致并且可以包含任选辅助模块的氨基酸序列;以及(vii)选自(i)和(ii)、(i)和(iii)、(iv)和(ii)、(iv)和(iii)、以及(v)和(vi)的两条tcr恒定链的二聚体组合;(b)任选的接头;以及(c)连接至(a)的选自由以下组成的组中的一个或多个非天然tcr抗原结合结构域:(1)抗体;(2)抗体片段(例如fv、fab、(fab')2);(3)抗体的重链可变区(vh结构域)或其片段;(4)抗体的轻链可变区(vl结构域)或其片段;(5)单链可变片段(scfv)或其片段;(6)单结构域抗体(sdab)或其片段;(7)骆驼科vhh结构域或其片段;(8)抗体的单体可变区;(9)非免疫球蛋白抗原结合支架,诸如darpin、亲合体(affibody)、affilin、adnectin、affitin、obodies、repebody、fynomer、alphabody、avimer、atrimer、centyrin、pronectin、anticalin、kunitz型结构域、armadillo重复蛋白或其片段;(10)受体或其片段;(11)配体或其片段;(12)双特异性-抗体、-抗体片段、-scfv、-vhh、-sdab、-非免疫球蛋白抗原结合支架、-受体或–配体;以及(13)自身抗原或其片段,其中(a)(i)–(a)(iii)的突变和(a)(vii)的二聚体提供了对抗原结合结构域的靶抗原的不同结合亲和力并且比具有相同结合结构域的ctcr大至少5%,并且当所述合成免疫受体在淋巴细胞中表达时,在淋巴细胞表面上表达在一条或多条连续链中的所述抗原结合结构域和所述t细胞受体恒定链两者,使得淋巴细胞被触发以活化、增殖、分泌细胞因子和/或调节(诱导或抑制)靶细胞的杀伤,并且当所述表达的抗原结合结构域结合至其抗原时具有mhc-限制性和mhc-非限制性抗体型特异性。在一个实施方式中,包含(a)(vii)的tcr恒定链,非天然tcr结合结构域选自由以下组成的组:对预定义的靶抗原特异性的抗体或其片段的重链和轻链的可变区,使得当表达时,该抗体或其片段的所述重链和轻链中的一条附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且该抗体或其片段的所述重链和轻链中的另一条附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个单链可变片段(scfv),使得当表达时,所述scfv中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述scfv中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个抗体片段,使得当表达时,所述抗体片段中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述抗体片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个单结构域抗体(sdab)片段,使得当表达时,所述sdab片段中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述sdab片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个骆驼科vhh结构域,使得当表达时,所述vhh结构域中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述vhh结构域中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个非免疫球蛋白抗原结合支架,使得当表达时,所述非免疫球蛋白抗原结合支架中的一个附接至所述t细胞恒定区的所述两条链的(a)(vii)中的一条,并且所述非免疫球蛋白抗原结合支架中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个受体或其片段,使得当表达时,所述受体或其片段中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述受体或其片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个配体或其片段,使得当表达时,所述配体或其片段中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述配体或其片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个结构不同的抗原结合片段,使得当表达时,所述抗原结合片段中的一个附接至所述t细胞恒定区的所述两条链的(a)(vii)中的一条,并且所述抗原结合片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;两个结合片段,其中的一者或两者有双特异性或多特异性,使得当表达时,所述抗原结合片段中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述抗原结合片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;两个自身抗原或其片段,使得当表达时,所述自身抗原或其片段中的一个附接至所述t细胞恒定区的所述两条链的(a)(vii)中的一条,并且所述自身抗原或其片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;以及两个vl或其片段,使得当表达时,所述vl或其片段中的一个附接至所述t细胞恒定区的所述两条链的(a)(vii)中的一条,并且所述vl或其片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;以及两个vh或其片段,使得当表达时,所述vh或其片段中的一个附接至所述t细胞恒定区的所述两条链的(a)(vii)中的一条,并且所述vh或其片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条。在前述任一者的另一个实施方式中,(a)(iv)的tcr恒定链具有选自由以下组成的组中的非天然tcr结合结构域:对预定义的靶抗原特异性的抗体或其片段的重链(vh)的可变区;对预定义的靶抗原特异性的抗体或其片段的轻链(vl)的可变区;对预定义的靶抗原特异性的单链可变片段(scfv)或其片段;对预定义的靶抗原特异性的抗体片段(例如fv、fab、(fab')2);对预定义的靶抗原特异性的单结构域抗体(sdab)片段;对预定义的靶抗原特异性的骆驼科vhh结构域;对预定义的靶抗原特异性的非免疫球蛋白抗原结合支架;对预定义的靶抗原特异性的受体或其片段;对预定义的靶抗原特异性的配体或其片段;对一个或多个预定义的靶抗原特异性的双特异性-抗体、-抗体片段、-scfv、-vhh、-sdab、-非免疫球蛋白抗原结合支架、-受体或-配体;以及自身抗原或其片段。在又一个实施方式中,一种编码(i)、(ii)、(iii)、(iv)、(v)或(vi)的多核苷酸,非天然tcr结合结构域选自由以下组成的组:对预定义的靶抗原特异性的抗体的重链(vh)的可变区;对预定义的靶抗原特异性的抗体的轻链(vl)的可变区;对预定义的靶抗原特异性的单链可变片段(scfv);对预定义的靶抗原特异性的抗体片段(例如fv、fab、(fab')2);对预定义的靶抗原特异性的单结构域抗体(sdab)片段;对预定义的靶抗原特异性的骆驼科vhh结构域;对预定义的靶抗原特异性的非免疫球蛋白抗原结合支架;对预定义的靶抗原或其片段特异性的受体;对预定义的靶抗原或其片段特异性的配体;对一个或多个预定义的靶抗原特异性的双特异性-抗体、-抗体片段、-scfv、-vhh、-sdab、-非免疫球蛋白抗原结合支架、-受体或-配体;以及自身抗原或其片段。在前述任一者的另一个实施方式中,编码tcr恒定链的多核苷酸是密码子优化序列。在前述任一者的另一个实施方式中,编码(a)的tcr恒定链的多核苷酸编码包含增强tcr恒定链表达和/或配对并减少其与内源性t细胞受体链配对的突变的tcr恒定链。在前述任一者的另一个实施方式中,编码(a)的tcr恒定链的多核苷酸包含核酸序列seqidno:730至743或与核酸序列seqidno:730至743具有至少70%一致性的序列的1-40个修饰的核酸序列,并且能够与tcrβ1或tcrβ2链进行二聚合。在前述任一者的另一个实施方式中,编码(b)或(c)的tcr恒定链的多核苷酸包含核酸序列seqidno:744至765或与核酸序列seqidno:744至765具有至少70%一致性的序列的1-40个修饰的核酸序列,并且能够与tcrα链进行二聚合。在前述任一者的另一个实施方式中,编码(v)的tcr恒定链的多核苷酸包含核酸序列seqidno:769至770或与核酸序列seqidno:769至770具有至少70%一致性的序列的1-40个修饰的核酸序列,并且能够与tcrδ链配对。在前述任一者的另一个实施方式中,编码(v)的tcr恒定链的多核苷酸包含核酸序列seqidno:771至772或与核酸序列seqidno:771至772具有至少70%一致性的序列的1-40个修饰的核酸序列,并且能够与tcrγ链进行二聚合。在前述任一者的另一个实施方式中,编码(iv)的tcr恒定链的多核苷酸包含核酸序列seqidno:766至768或与核酸序列seqidno:766至768具有至少70%一致性的序列的1-40个修饰的核酸序列,并且能够与tcrβ1或tcrβ2链进行二聚合。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至一个或多个疾病相关抗原,这些疾病相关抗原选自由以下组成的组:cd19;cd123;cd22;cd30;cd171;cs-1(也称为cd2亚单位1、cracc、slamf7、cd319和19a24);c型凝集素样分子-1(cll-1或clecl1);cd33;表皮生长因子受体变体iii(egfrviii);神经节苷脂g2(gd2);神经节苷脂gd3(aneu5ac(2-8)aneu5ac(2-3)bdgalp(l-4)bdglcp(l-l)cer);tnf受体家族成员b细胞成熟(bcma);tn抗原((tnag)或(galnacα-ser/thr));前列腺特异性膜抗原(psma);受体酪氨酸激酶样孤儿受体1(ror1);fms样酪氨酸激酶3(flt3);肿瘤相关糖蛋白72(tag72);cd38;cd44v6;在急性白血病或淋巴瘤上而不在造血祖细胞上表达的糖基化cd43表位、在非造血癌症上表达的糖基化cd43表位、癌胚抗原(cea);上皮细胞粘附分子(epcam);b7h3(cd276);kit(cd117);白介素-13受体亚基α-2(il-13ra2或cd213a2);间皮素;白介素11受体α(il-llra);前列腺干细胞抗原(psca);蛋白酶丝氨酸21(睾蛋白或prss21);血管内皮生长因子受体2(vegfr2);路易斯(lewis)(y)抗原;cd24;血小板源性生长因子受体β(pdgfr-β);阶段特异性胚胎抗原-4(ssea-4);cd20;叶酸受体α;受体酪氨酸蛋白激酶erbb2(her2/neu);细胞表面相关粘蛋白1(muc1);表皮生长因子受体(egfr);神经细胞粘附分子(ncam);前列腺酶;前列腺酸性磷酸酶(pap);突变的延长因子2(elf2m);肝配蛋白b2;成纤维细胞活化蛋白α(fap);胰岛素样生长因子1受体(igf-i受体),碳酸酐酶ix(calx);蛋白酶体(前体,巨蛋白因子)亚基,β型,9(lmp2);糖蛋白100(gpl00);由裂点簇区(bcr)和阿贝尔森(abelson)鼠类白血病病毒癌基因同源物1(abl)组成的癌基因融合蛋白(bcr-abl);酪氨酸酶;肝配蛋白a型受体2(epha2);岩藻糖基gm1;唾液酸化路易斯粘附分子(sle);神经节苷脂gm3(aneu5ac(2-3)bdclalp(l-4)bdglcp(l-1)cer);转谷氨酰胺酶5(tgs5);高分子量-黑素瘤相关抗原(hmwmaa);o-乙酰基-gd2神经节苷脂(oacgd2);叶酸受体β;肿瘤内皮标记1(tem1/cd248);肿瘤内皮标记7相关(tem7r);密封蛋白(claudin)6(cldn6);甲状腺刺激性激素受体(tshr);g蛋白偶联受体c类5组成员d(gprc5d);x染色体开放阅读框61(cxorf61);cd97;cd179a;间变性淋巴瘤激酶(alk);聚唾液酸;胎盘特异性1(plac1);globoh糖神经酰胺六糖部分(globoh);乳腺分化抗原(ny-br-1);尿空斑蛋白(uroplakin)2(upk2);甲型肝炎病毒细胞受体1(havcr1);肾上腺受体β3(adrb3);泛连接蛋白3(panx3);g蛋白偶联受体20(gpr20);淋巴细胞抗原6复合物,基因座k9(ly6k);嗅觉受体51e2(or51e2);tcrγ替代性阅读框蛋白质(tarp);威尔姆氏瘤蛋白质(wt1);癌症/睾丸抗原1(ny-es0-1);癌症/睾丸抗原2(lage-1a);黑素瘤相关抗原1(mage-a1);位于染色体12p上的ets易位变体基因6(etv6-aml);精子蛋白17(spa17);x抗原家族成员la(xagel);血管生成素结合细胞表面受体2(tie2);黑素瘤癌症睾丸抗原-1(mad-ct-1);黑素瘤癌症睾丸抗原-2(mad-ct-2);fos相关抗原1;肿瘤蛋白p53(p53);p53突变体;前列腺特异性蛋白;存活素(surviving);端粒酶;前列腺癌肿瘤抗原-1(pcta-1或半乳凝素8)、由t细胞识别的黑素瘤抗原1(melana或marti);大鼠肉瘤(ratsarcoma,ras)突变体;人类端粒酶逆转录酶(htert);肉瘤易位裂点;黑素瘤细胞凋亡抑制剂(ml-iap);erg(跨膜蛋白酶,丝氨酸2(tmprss2)ets融合基因);n-乙酰基葡萄糖胺基-转移酶v(na17);配对框蛋白质pax-3(pax3);雄激素受体;周期素bl;v-myc禽类骨髓细胞瘤病病毒癌基因神经母细胞瘤衍生性同源物(mycn);ras同源家族成员c(rhoc);酪氨酸酶相关蛋白2(trp-2);细胞色素p450lb1(cyplb1);ccctc-结合因子(锌指蛋白)样(boris或印迹位点调节剂同级物)、由t细胞识别的鳞状细胞癌抗原3(sart3);配对框蛋白质pax-5(pax5);前顶体蛋白结合蛋白sp32(oy-tesl);淋巴细胞特异性蛋白质酪氨酸激酶(lck);a激酶锚定蛋白4(akap-4);滑膜肉瘤,x裂点2(ssx2);晚期糖基化终产物受体(rage-1);肾泛素1(rul);肾泛素2(ru2);豆荚蛋白;人乳头状瘤病毒e6(hpve6);人乳头状瘤病毒e7(hpve7);肠道羧基酯酶;突变的热休克蛋白70-2(muthsp70-2);cd79a;cd79b;cd72;白细胞相关免疫球蛋白样受体1(lairl);iga受体的fc片段(fcar或cd89);白细胞免疫球蛋白样受体子家族a成员2(lilra2);cd300分子样家族成员f(cd300lf);c型凝集素结构域家族12成员a(clec12a);骨髓基质细胞抗原2(bst2);egf样含模块粘蛋白样激素受体样2(emr2);淋巴细胞抗原75(ly75);磷脂酰肌醇聚糖-3(gpc3);fc受体样5(fcrl5);以及免疫球蛋白λ样多肽1(iglll)、mpl、、生物素、c-myc表位标签、cd34、lamp1trop2、gfrα54、cdh17、cdh6、nybr1、cdh19、cd200r、slea(ca19.9;唾液酸化路易斯抗原)岩藻糖基-gm1、ptk7、gpnmb、cdh1-cd324、dll3、cd276/b7h3、il11ra、il13ra2、cd179b-igll1、alktcrγ-δ、nkg2d、cd32(fcgr2a)、tnag、cspg4-hmw-maa、tim1-/hvcr1、csf2ra(gm-csfr-α)、tgfβr2、vegfr2/kdr、lewsag、tcr-β1链、tcr-β2链、tcr-γ链、tcr-δ链、fitc、促黄体生成激素受体(lhr)、卵泡刺激素受体(fshr)、绒膜促性腺激素激素受体(cghr)、ccr4、gd3、slamf6、slamf4、hiv1包膜糖蛋白、htlv1-tax、cmvpp65、ebv-ebna3c、甲型流感凝集素(ha)、gad、pdl1、鸟苷酸环化酶c(gcc)、kshv-k8.1蛋白、kshv-gh蛋白、桥粒芯蛋白自身抗体3(dsg3)、桥粒芯蛋白自身抗体1(dsg1)、hla、hla-a、hla-a2、hla-b、hla-c、hla-dp、hla-dm、hla-doa、hla-dob、hla-dq、hla-dr、hla-g、ige、cd99、rasg12v、组织因子1(tf1)、afp、gprc5d、密封蛋白18.2(cld18a2或cldn18a.2))、p-糖蛋白、steap1、liv1、粘连蛋白-4、cripto、gpa33、bst1/cd157、低电导率氯离子通道和由tnt抗体识别的抗原。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含抗体、抗体片段、scfv、fv、fab、(fab')2、单结构域抗体(sdab)、vh或vl结构域、骆驼科vhh结构域、非免疫球蛋白抗原结合支架,例如darpin、亲合体、affilin、adnectin、affitin、obodies、repebody、fynomer、alphabody、avimer、atrimer、centyrin、pronectin、anticalin、kunitz型结构域、armadillo重复蛋白、受体或配体。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域选自由以下组成的组:(i)由具有seqidno226至400或10203至10321中任一个的序列或与其有至少98%一致性的序列并编码结合至其抗原的多肽的多核苷酸编码的重链可变区(vh);(ii)由具有seqidno16至191或10085至10202中任一个的序列或与其有至少98%一致性的序列并编码结合至其抗原的多肽的多核苷酸编码的轻链可变区(vl);(iii)由具有seqidno488至657、10346至10400或18098至18160中任一个的序列或与其有至少98%一致性的序列并编码结合至其抗原的多肽的多核苷酸编码的单链可变片段(scfv);(iv)由具有seqidno421至445或10322至10337中任一个的序列或与其有至少98%一致性的序列并编码结合至其抗原的多肽的多核苷酸编码的骆驼科vhh结构域;(v)由具有seqidno439至443中任一个的序列或与其有至少98%一致性的序列并编码结合至其抗原的多肽的多核苷酸编码的非免疫球蛋白支架;(vi)由具有seqidno456至468中任一个的序列或与其有至少98%一致性的序列并编码结合至其同源物的多肽的多核苷酸编码的受体;以及(vii)由具有seqidno476至486或10402至10404中任一个的序列或与其有至少98%一致性的序列并编码结合至其同源物的多肽的多核苷酸编码的配体。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含seqidno:13999至14879或14880中任一个所示的所选靶抗原的轻链互补决定区中的一个或多个和/或seqidno:14881至15761或15762中任一个所示的所选靶抗原的重链互补决定区中的一个或多个。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含含有seqidno:2307至2482或12042至12159中任一个的序列并具有至多10个保守性氨基酸取代的可变轻链(vl)结构域和/或含有seqidno:2506至2680或12160至12278中任一个的序列并具有至多10个保守性氨基酸取代的可变重链(vh)结构域。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含seqidno:2701至2725或12279至12294中任一个所示并具有至多10个保守性氨基酸取代的所选抗原的骆驼科vhh互补决定区中的一个或多个。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含具有seqidno:2728-2732或12296至12301中任一个所示的序列并具有至多10个保守性氨基酸取代的非免疫球蛋白抗原结合结构域。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含scfv结构域,该结构域包含含有seqidno:2307至2482或12042至12159中任一个的序列的可变轻链(vl)结构域的一个或多个轻链互补决定区和含有seqidno:2506至2680或12160至12278中任一个的序列的可变重链(vh)结构域的一个或多个重链互补决定区。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含具有选自由seqidno:2770至2939、12303至12357或18162至18224组成的组中的序列的scfv片段,其各自具有至多10个保守性氨基酸取代。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含含有seqidno:2736至2748中任一个的氨基酸序列并具有至多10个保守性氨基酸取代的一个或多个受体。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含含有seqidno:2758-2768或12359至12361中任一个的序列并具有至多10个保守性氨基酸取代的一个或多个配体。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含cd16a、nkg2d、cd4、pd1、桥粒芯蛋白3(dsg3)或cd4-dc-sign的细胞外结构域。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含htpo、mtpo、cghα链、cghβ链、fhβ链、lhβ链、tshβ链、april或其组合中的一个或多个的细胞外结构域的细胞外结构域。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含含有seqidno:2770至2939、12303至12357或18162至18224中任一个的序列并具有至多10个保守性氨基酸取代的任何单链可变片段(scfv),以及a)seqidno:2701至2725或12279至12294中任一个所示的并具有至多10个保守性氨基酸取代的任何骆驼科vhh,或b)具有seqidno:2728-2732或12296至12301中任一个所示的序列并具有至多10个保守性氨基酸取代的任何非免疫球蛋白抗原结合结构域;或c)包含seqidno:2736至2748中任一个的氨基酸序列并具有至多10个保守性氨基酸取代的受体的任何细胞外结构域;或d)包含seqidno:2758-2768或12359至12361中任一个的序列并具有至多10个保守性氨基酸取代的配体的任何细胞外结构域。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含含有seqidno:2701至2725或12279至12294中任一个所示并具有至多10个保守性氨基酸取代的骆驼科vhh,以及a)包含seqidno:2770至2939、12303至12357或18162至18224中任一个的序列并具有至多10个保守性氨基酸取代的任何单链可变片段(scfv),或b)具有seqidno:2728-2732或12296至12301中任一个所示的序列并具有至多10个保守性氨基酸取代的任何非免疫球蛋白抗原结合结构域;或c)包含seqidno:2736至2748中任一个的氨基酸序列并具有至多10个保守性氨基酸取代的受体的任何细胞外结构域;或d)包含seqidno:2758-2768或12359至12361中任一个的序列并具有至多10个保守性氨基酸取代的配体的任何细胞外结构域。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域通过接头区任选地连接至tcr恒定区链的每一条,其中所述接头区核酸编码选自由seqidno:2981至2992及其任何组合或与其有至少98%一致性的序列组成的组中的氨基酸序列;或者所述接头由选自由seqidno:701至714或与其有至少98%一致性的序列组成的组中的核酸序列编码。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域与其靶抗原的结合亲和力比从中获得该结合结构域的抗体小至少5倍。在前述任一者的另一个实施方式中,编码sir的多核苷酸还包含存在于每条链的n末端处并包含选自由seqidno:1-9和10组成的组中的序列的前导序列或信号肽。在前述任一者的另一个实施方式中,该至少一个多核苷酸编码两个sir。在前述任一者的另一个实施方式中,该多核苷酸编码通过编码可裂解接头的核苷酸序列连接的两个sir。在前述任一者的另一个实施方式中,该可裂解接头是自身裂解的可裂解接头。在前述任一者的另一个实施方式中,可裂解接头是2a接头、2a样接头或其功能等效物中的任一个或多个。在前述任一者的另一个实施方式中,可裂解接头是t2a接头、p2a、f2a、e2a接头或其功能等效物中的任一个或多个。在前述任一者的另一个实施方式中,可裂解接头包含seqidno:780至785中任一个或多个的序列。在前述任一者的另一个实施方式中,编码可裂解接头的多核苷酸序列任选地在编码弗林蛋白酶裂解位点或弗林蛋白酶样裂解位点或其功能等效物的核苷酸序列后面。在前述任一者的另一个实施方式中,在可裂解接头前面的弗林蛋白酶样裂解位点包含seqidno:788至790中任一个或多个的序列。在前述任一者的另一个实施方式中,编码可裂解接头的多核苷酸序列在编码柔性接头的核苷酸序列后面。在前述任一者的另一个实施方式中,在可裂解接头前面的柔性接头编码ser-gly接头、ser-gly-ser-gly接头或其功能等效物中的一个或多个。在前述任一者的另一个实施方式中,在可裂解接头前面的柔性接头包含序列seqidno:786或787。在前述任一者的另一个实施方式中,编码弗林蛋白酶裂解位点的多核苷酸序列之后为编码柔性接头的多核苷酸,在编码柔性接头的多核苷酸之后为编码可裂解接头的多核苷酸,使得该次序为弗林蛋白酶裂解位点-柔性接头-可裂解接头。在前述任一者的另一个实施方式中,编码可裂解接头的多核苷酸存在于编码前导序列(信号肽)的序列之前,所述序列编码第二sir。在前述任一者的另一个实施方式中,sir可以设计为对所选抗原具有不同结合亲和力。在前述任一者的另一个实施方式中,sir包含辅助模块。在前述任一者的另一个实施方式中,辅助模块包含cd3z结构域。在前述任一者的另一个实施方式中,tcr恒定链选自由以下组成的组:(viii)与seqidno:12401或12402或12403或12408或12409至少98%一致的氨基酸序列;(ix)与seqidno:12421或12422或12423或12427或12428至少98%一致的氨基酸序列;以及(x)(viii)和(ix)的两条tcr恒定链的二聚体组合。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至cd19。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与seqidno:2318-2324、12060-12068、12108、12127、或12156中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2517-2523、12178-12186、1227、12246或12275中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:12288至少98%一致的序列的多肽;以及包含与seqidno:2770-2774、12325、12308、18162-18170或12354中任一个至少98%一致的序列的多肽;在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3135-3235、3250-3346、3396、3401-3403、3406、3429-3432、3435-3439、3540、3855-3859、12431-12489、12491-12493、12495-12530、12534、13195-13203、13250、13267、13289、13429-13437、13483、13501和13523组成的组中的序列的多肽。在另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至cd20。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与seqidno:2325-2326、12069-12077或12078中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2524-2525、12187-12195或12196中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:12289或12290至少98%一致的序列的多肽;以及包含与seqidno:2787-2788、18177-18186或18187中任一个至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3263、3348、3456-3457、3876-3877、12464-12465、12477-12482、12492、12534、13204-13213、13438-13446和13447组成的组中的序列的多肽。在另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至cd22。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与seqidno:2327-2329、12122-12126或12132中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2526-2528、12241-12245或12251中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);以及包含与seqidno:2789-2791、12320-12330或18188中任一个至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3332、3433、3458-3460、3878-3880、12483、12485、12488-12490、13241-13245、13268、13475-13479和13502组成的组中的序列的多肽。在另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至bcma。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与seqidno:2310-2313、12046-12048、12118-12119、12139-12145或12146中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2509-2512、12164-12166、12237-12238、12258-12264或12265中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:12279-12281、12283-12285、12287、12291-12292、12293或12294中任一个至少98%一致的序列的多肽;以及包含与seqidno:2780-2783、12237-12344、18174-18175或18176中任一个至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3445-3449、3866-3869、12463、12533、12535-12536、13181-13183、13261-13262、13277-13284、13415-13417、13495-13496、13511-13517和13518组成的组中的序列的多肽。在另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至mpl。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与seqidno:2414-2421、12120、12128或12129中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2611-2618、12239、12247或12248中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);以及包含与seqidno:2871-2878、12326-12327以及12318中任一个至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3347、3373、3427-3428、3495、3556-3562、3979-3985、4025、12454、12456、12458、12462、12532、13259、13265-13266、13493、13499和13500组成的组中的序列的多肽。在另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至cs1。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与seqidno:2355-2358、12090-12094或12095中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2553-2555、12209-12213或12214中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);以及包含与seqidno:2817-2819、18211-18215或18216中任一个至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3376、3487-3489、3907-3909、12455、12457、12459、12461、12476、13226-13231、13460-13464和13465组成的组中的序列的多肽。在另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至cd33。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与seqidno:2336-2337、12079-12084或12085中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2535-2536、12197-12202或12203中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);以及包含与seqidno:2795-2796、18189-18193或18194中任一个至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3464-3465、3884-3885、12460、12473、12479、13214-13220、13448-13453和13454组成的组中的序列的多肽。在另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至cd123。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与seqidno:2315、2472、12049-12058或12059中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2514、2670、12167-12176或12177中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2716或2717至少98%一致的序列的多肽;以及包含与seqidno:2801、2929、18196-18205或18206中任一个至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3266-3267、3366-3368、3375、3378、3405、3409、3434、3470、3492-3497、3617、3890、3912-3913、4041、12480、13184-13194、13418-13427和13428组成的组中的序列的多肽。在另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至叶酸受体1。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与2373至少98%一致的序列的多肽或其中所含有的任何互补决定区(cdr);包含与seqidno:2570中任一个至少98%一致的序列的多肽或其中所含有的任何互补决定区(cdr);以及包含与seqidno:2833中任一个至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3511和3928组成的组中的序列的多肽。在另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至间皮素。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与seqidno:2413、12154或12155中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2609-2610、12273或12274中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2713-2714或2725至少98%一致的序列的多肽;以及包含与seqidno:2870、2899、12352或12353中任一个至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3414、3419、3554、3585、3976、4008、13287-13288、13521和13522组成的组中的序列的多肽。在另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至il13ra2。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:-包含与seqidno:2399或2400中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2595或2596中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);以及包含与seqidno:2858或2859中任一个至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3541-3542、3963和3964组成的组中的序列的多肽。在另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至cd138。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与seqidno:2316至少98%一致的序列的多肽或其中所含有的任何互补决定区(cdr);包含与seqidno:2515至少98%一致的序列的多肽或其中所含有的任何互补决定区(cdr);以及包含与seqidno:2802至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3268、3374、3404、3471和3891组成的组中的序列的多肽。在另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至tcrgd。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与seqidno:2449至少98%一致的序列的多肽或其中所含有的任何互补决定区(cdr);包含与seqidno:2646至少98%一致的序列的多肽或其中所含有的任何互补决定区(cdr);以及包含与seqidno:2907至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3594和4017组成的组中的序列的多肽。在又一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至tcrb1。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与seqidno:2445或2446中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2642或2643中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);以及包含与seqidno:2903或2904中任一个至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3590-3591、4013和4014组成的组中的序列的多肽。在另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域结合至tcrb2。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:包含与seqidno:2447或2448中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);包含与seqidno:2644或2645中任一个至少98%一致的序列的多肽或前述多肽中任一个所含有的任何互补决定区(cdr);以及包含与seqidno:2905或2906中任一个至少98%一致的序列的多肽。在前述任一者的另一个实施方式中,重组多核苷酸编码包含选自由seqidno:3353-3364、3592-3593、4015和4016组成的组中的序列的多肽。本公开还提供一种重组表达系统,其包含与治疗控制剂共同表达的上文所述的任一重组多核苷酸,其中该治疗控制剂选自由以下组成的组:截短的表皮生长因子受体(tegfr)、截短的表皮生长因子受体viii(tegfrviii)、截短的cd30(tcd30)、截短的bcma(tbcma)、截短的cd19(tcd19)、cd34、胸苷激酶、胞嘧啶脱氨酶、硝基还原酶、黄嘌呤鸟嘌呤磷酸核糖转移酶、人类半胱天冬酶8、人类半胱天冬酶9、诱导型半胱天冬酶9(icaspase9)、嘌呤核苷磷酸化酶、亚麻苦苷酶/亚麻苦苷/葡萄糖氧化酶、脱氧核苷激酶、辣根过氧化物酶(hrp)/吲哚-3-乙酸(iaa)、γ-谷氨酰半胱氨酸合成酶、cd20/αcd20、cd34/胸苷激酶嵌合物、dox依赖性半胱天冬酶-2、突变体胸苷激酶(hsv-tksr39)、ap1903/fas系统、嵌合细胞因子受体(ccr)、选择标记物及其组合。在一个实施方式中,tegfr和tegfrviii结合egfr特异性sirna、小分子、抗-egfr抗体或其片段、或其组合中的任一个或多个。在另一个实施方式中,tcd30结合cd30特异性sirna、小分子、抗-cd30抗体或其片段、或其组合中的任一个或多个。在另一个实施方式中,tcd19结合cd19特异性sirna、小分子、抗-cd19抗体或其片段、或其组合中的任一个或多个。在另一个实施方式中,cd34结合cd34特异性sirna、小分子、抗-cd34抗体或其片段、或其组合中的任一个或多个。在另一个实施方式中,选择标记物包含二羟基叶酸受体(dhfr)、突变体dhfr、甲基化dna-蛋白质-半胱氨酸甲基转移酶、次黄嘌呤一磷酸盐脱氢酶ii(imdhp2)、嘌呤霉素乙酰转移酶(pac)、杀稻瘟菌素抗性基因、突变体钙调磷酸酶a/b(can/b)、cna12、cnb30或其组合中的任一个或多个。在另一个实施方式中,ccr包含(i)il-7细胞因子接头-il7ra,(ii)il2rβ的il-7ra-胞浆结构域的il-7ra-跨膜结构域的il-7细胞因子接头细胞外结构域,(iii)il-7细胞因子接头-il2rβ,以及(iv)其组合中的任一个或多个。在前述任一者的另一个实施方式中,重组表达系统包含本公开的与辅助模块共同表达的重组多核苷酸,其中该辅助模块选自由以下组成的组:41bbl、cd40l、k13、mc159、cflip-l/mritα、cflip-p22、htlv1tax、htlv2tax、htlv2tax-rs突变体、fkbpx2-k13、fkbpx2-htlv2-tax、fkbpx2-htlv2-tax-rs、il6r-304-vhh-alb8-vhh、il12f、pd1-4h1scfv、pd1-5c4scfv、pd1-4h1-alb8-vhh、pd1-5c4-alb8-vhh、ctla4-伊匹单抗-scfv、ctla4-伊匹单抗-alb8-vhh、il6-19a-scfv、il6-19a-scfv-alb8-vhh、shvem、shvem-alb8-vhh、htert、fx06、cd3z、cd3z-gggs-41bb、cd3-bbz、cd3-cd28z、cd3-cd28-lck融合蛋白、shrna靶向brd4、嵌合抗原受体(car)、htert、肝素酶、car、抑制性car及其任何组合。在前述任一者的另一个实施方式中,编码sir的重组多核苷酸和一个或多个治疗控制剂和/或一个或多个辅助模块通过编码可裂解接头的核苷酸序列连接。在另一个实施方式中,该可裂解接头是自身裂解的可裂解接头。在前述任一者的另一个实施方式中,编码可裂解接头的多核苷酸序列在编码弗林蛋白酶裂解位点或弗林蛋白酶样裂解位点或其功能等效物的核苷酸序列后面。在前述任一者的另一个实施方式中,编码可裂解接头的多核苷酸序列任选地在编码柔性接头的核苷酸序列后面。本公开还提供至少一种载体,该载体包含如本文上文所述的重组多核苷酸,其中该载体选自由dna载体、rna载体、质粒、慢病毒载体、腺病毒载体、逆转录病毒载体、杆状病毒载体、睡美人转座子载体和piggybac转座子载体组成的组。在一个实施方式中,载体骨架具有选自由seqidno:870至875和876组成的组中的序列。在一些实施方式中,该载体包含选自ef-1启动子、cmvie基因启动子、ef-1α启动子、泛素c启动子、mscvltr启动子或磷酸甘油酸激酶(pgk)启动子的启动子。在另一个实施方式中,ef-1启动子包含seqidno:877的序列或与其有80-99%一致性的序列。在前述任一者的另一个实施方式中,载体是体外转染的载体,或者载体还包含聚(a)尾部或3'utr。本公开提供至少一种多肽,该多肽由本公开的至少一种重组多核苷酸编码。本公开还提供重组细胞,该重组细胞表达至少一种如本文上文所述的重组多核苷酸。本公开还提供一种分离的合成免疫受体(sir)多肽或多肽异二聚体,其包含:(a)具有选自由以下组成的组中的氨基酸序列的t细胞受体(tcr)恒定链:(i)与seqidno:3010至少98%一致并在位置48、61、91、92、93和/或94处具有一个或多个突变并且可以包含任选辅助模块的氨基酸序列;(ii)与seqidno:3024至少98%一致并在位置18、22、57、79、133、136和/或139处具有一个或多个突变并且可以包含任选辅助模块的氨基酸序列;(iii)与seqidno:3025至少98%一致并在位置18、22、57、79、133、136和/或139处具有一个或多个突变并且可以包含任选辅助模块的氨基酸序列;(iv)与seqidno:3046、3047或3048至少98%一致并且可以包含任选辅助模块的氨基酸序列;(v)与seqidno:3049至少98%一致并且可以包含任选辅助模块的氨基酸序列;(vi)与seqidno:3051或3052至少98%一致并且可以包含任选辅助模块的氨基酸序列;以及(vii)选自(i)和(ii)、(i)和(iii)、(iv)和(ii)、(iv)和(iii)、或者(v)和(vi)的两条tcr恒定链的二聚体组合;(b)任选的接头;以及(c)连接至(a)的选自由以下组成的组中的一个或多个非天然tcr抗原结合结构域:(1)抗体;(2)抗体片段(例如fv、fab、(fab')2);(3)抗体的重链可变区(vh结构域)或其片段;(4)抗体的轻链可变区(vl结构域)或其片段;(5)单链可变片段(scfv)或其片段;(6)单结构域抗体(sdab)或其片段;(7)骆驼科vhh结构域或其片段;(8)抗体的单体可变区;(9)非免疫球蛋白抗原结合支架,例如darpin、亲合体、affilin、adnectin、affitin、obodies、repebody、fynomer、alphabody、avimer、atrimer、centyrin、pronectin、anticalin、kunitz型结构域、armadillo重复蛋白或其片段;(10)受体或其片段;(11)配体或其片段;(12)双特异性-抗体、-抗体片段、-scfv、-vhh、-sdab、-非免疫球蛋白抗原结合支架、-受体或-配体;以及(13)自身抗原或其片段,其中(a)(i)-(a)(iii)的突变提供对抗原结合结构域的靶抗原的不同结合亲和力,并且该合成免疫受体在淋巴细胞中表达时,在淋巴细胞表面上表达一条或多条连续链中的所述抗原结合结构域和所述t细胞受体恒定链两者,使得淋巴细胞被触发以活化、增殖、分泌细胞因子和/或调节(诱导或抑制)靶细胞的杀伤并且当所述表达的抗原结合结构域结合至其抗原时具有mhc限制性或mhc非限制性抗体型特异性。在前述任一者的另一个实施方式中,(a)(vii)的tcr恒定链具有选自由以下组成的组中的非天然tcr结合结构域:对预定义的靶抗原特异性的抗体或其片段的重链和轻链的可变区,使得当表达时,该抗体或其片段的所述重链和轻链中的一条附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且该抗体或其片段的所述重链和轻链中的另一条附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个单链可变片段(scfv),使得当表达时,所述scfv中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述scfv中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个抗体片段,使得当表达时,所述抗体片段中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述抗体片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个单结构域抗体(sdab)片段,使得当表达时,所述sdab片段中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述sdab片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个骆驼科vhh结构域,使得当表达时,所述vhh结构域中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述vhh结构域中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个非免疫球蛋白抗原结合支架,使得当表达时,所述非免疫球蛋白抗原结合支架中的一个附接至所述t细胞恒定区的所述两条链的(a)(vii)中的一条,并且所述非免疫球蛋白抗原结合支架中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个受体或其片段,使得当表达时,所述受体或其片段中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述受体或其片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个配体或其片段,使得当表达时,所述配体或其片段中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述配体或其片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;对一个或多个预定义的靶抗原特异性的两个结构不同的抗原结合片段,使得当表达时,所述抗原结合片段中的一个附接至所述t细胞恒定区的所述两条链的(a)(vii)中的一条,并且所述抗原结合片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;两个结合片段,其中的一者或两者有双特异性或多特异性,使得当表达时,所述抗原结合片段中的一个附接至所述t细胞恒定区的(a)(vii)的所述两条链中的一条,并且所述抗原结合片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;两个自身抗原或其片段,使得当表达时,所述自身抗原或其片段中的一个附接至所述t细胞恒定区的所述两条链的(a)(vii)中的一条,并且所述自身抗原或其片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;以及两个vl或其片段,使得当表达时,所述vl或其片段中的一个附接至所述t细胞恒定区的所述两条链的(a)(vii)中的一条,并且所述vl或其片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条;两个vh或其片段,使得当表达时,所述vh或其片段中的一个附接至所述t细胞恒定区的所述两条链的(a)(vii)中的一条,并且所述vh或其片段中的另一个附接至所述t细胞恒定区的所述两条链中的另一条。在前述任一者的另一个实施方式中,(a)(iv)的tcr恒定链具有选自由以下组成的组中的非天然tcr结合结构域:对预定义的靶抗原特异性的抗体或其片段的重链(vh)的可变区;对预定义的靶抗原特异性的抗体或其片段的轻链(vl)的可变区;对预定义的靶抗原特异性的单链可变片段(scfv)或其片段;对预定义的靶抗原特异性的抗体片段(例如fv、fab、(fab')2);对预定义的靶抗原特异性的单结构域抗体(sdab)片段;对预定义的靶抗原特异性的骆驼科vhh结构域;对预定义的靶抗原特异性的非免疫球蛋白抗原结合支架;对预定义的靶抗原特异性的受体或其片段;对预定义的靶抗原特异性的配体或其片段;对一个或多个预定义的靶抗原特异性的双特异性-抗体、-抗体片段、-scfv、-vhh、-sdab、-非免疫球蛋白抗原结合支架、-受体或-配体;以及自身抗原或其片段。在前述任一者的另一个实施方式中,(i)、(ii)、(iii)、(iv)、(v)或(vi)的tcr恒定结构域包含选自由以下组成的组中的非天然tcr结合结构域:对预定义的靶抗原特异性的抗体的重链(vh)的可变区;对预定义的靶抗原特异性的抗体的轻链(vl)的可变区;对预定义的靶抗原特异性的单链可变片段(scfv);对预定义的靶抗原特异性的抗体片段(例如fv、fab、(fab')2);对预定义的靶抗原特异性的单结构域抗体(sdab)片段;对预定义的靶抗原特异性的骆驼科vhh结构域;对预定义的靶抗原特异性的非免疫球蛋白抗原结合支架;对预定义的靶抗原或其片段特异性的受体;对预定义的靶抗原或其片段特异性的配体;对一个或多个预定义的靶抗原特异性的双特异性-抗体、-抗体片段、-scfv、-vhh、-sdab、-非免疫球蛋白抗原结合支架、-受体或-配体;以及自身抗原或其片。在前述任一者的另一个实施方式中,tcr恒定链包含增强tcr恒定链表达和/或配对并减少其与内源性t细胞受体链配对的突变。在前述任一者的另一个实施方式中,tcr的恒定区是包含具有选自由seqidno:3010至3023组成的组中的序列或与选自由seqidno:3010至3023组成的组中的氨基酸序列至少98%一致的序列的1-40个氨基酸取代或突变的氨基酸序列的tcr受体α链(cα)。在前述任一者的另一个实施方式中,tcr的恒定区是包含具有选自由seqidno:3024至3044组成的组中的序列或与选自由seqidno:3024至3044组成的组中的氨基酸序列至少98%一致的序列的1-40个氨基酸取代或突变的氨基酸序列的tcr受体β链(cβ)。在前述任一者的另一个实施方式中,tcr的恒定区是包含具有选自由seqidno:3049至3050组成的组中的序列或与选自由seqidno:3049至3050组成的组中的氨基酸序列至少98%一致的序列的1-40个氨基酸取代或突变的氨基酸序列的tcr受体γ链(cγ)。在前述任一者的另一个实施方式中,tcr的恒定区是包含具有选自由seqidno:3051至3052组成的组中的氨基酸序列或与选自由seqidno:3051至3052组成的组中的氨基酸序列至少98%一致的序列的1-40个氨基酸取代或突变的氨基酸序列的tcr受体δ链(cδ)。在前述任一者的另一个实施方式中,tcr的恒定区是包含具有选自由seqidno:3046至3048组成的组中的氨基酸序列或与选自由seqidno:3046至3048组成的组中的氨基酸序列至少98%一致的序列的1-40个氨基酸取代或突变的氨基酸序列的前体tcr受体α链(前体cα)。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域结合至一个或多个疾病相关抗原,这些疾病相关抗原选自由以下组成的组:cd19;cd123;cd22;cd30;cd171;cs-1(也称为cd2亚单位1、cracc、slamf7、cd319和19a24);c型凝集素样分子-1(cll-1或clecl1);cd33;表皮生长因子受体变体iii(egfrviii);神经节苷脂g2(gd2);神经节苷脂gd3(aneu5ac(2-8)aneu5ac(2-3)bdgalp(l-4)bdglcp(l-l)cer);tnf受体家族成员b细胞成熟(bcma);tn抗原((tnag)或(galnacα-ser/thr));前列腺特异性膜抗原(psma);受体酪氨酸激酶样孤儿受体1(ror1);fms样酪氨酸激酶3(flt3);肿瘤相关性糖蛋白72(tag72);cd38;cd44v6;在急性白血病或淋巴瘤上而不在造血祖细胞上表达的糖基化cd43表位、在非造血癌症上表达的糖基化cd43表位、癌胚抗原(cea);表皮细胞粘附分子(epcam);b7h3(cd276);kit(cd117);白介素-13受体亚基α-2(il-13ra2或cd213a2);间皮素;白介素11受体α(il-llra);前列腺干细胞抗原(psca);蛋白酶丝氨酸21(睾蛋白或prss21);血管内皮生长因子受体2(vegfr2);路易斯(y)抗原;cd24;血小板源性生长因子受体β(pdgfr-β);阶段特异性胚胎抗原-4(ssea-4);cd20;叶酸受体α;受体酪氨酸蛋白激酶erbb2(her2/neu);细胞表面相关粘蛋白1(muc1);表皮生长因子受体(egfr);神经细胞粘附分子(ncam);前列腺酶;前列腺酸性磷酸酶(pap);突变的延长因子2(elf2m);肝配蛋白b2;成纤维细胞活化蛋白α(fap);胰岛素样生长因子1受体(igf-i受体)、碳酸酐酶ix(calx);蛋白酶体(前体、巨蛋白因子)亚基,β型,9(lmp2);糖蛋白100(gpl00);由裂点簇区(bcr)和阿贝尔森(abelson)鼠类白血病病毒癌基因同源物1(abl)组成的癌基因融合蛋白(bcr-abl);酪氨酸酶;肝配蛋白a型受体2(epha2);岩藻糖基gm1;唾液酸化路易斯粘附分子(sle);神经节苷脂gm3(aneu5ac(2-3)bdclalp(l-4)bdglcp(l-1)cer);转谷氨酰胺酶5(tgs5);高分子量-黑素瘤相关抗原(hmwmaa);o-乙酰基-gd2神经节苷脂(oacgd2);叶酸受体β;肿瘤内皮标记1(tem1/cd248);肿瘤内皮标记7相关(tem7r);密封蛋白6(cldn6);甲状腺刺激性激素受体(tshr);g蛋白偶联受体c类5组成员d(gprc5d);x染色体开放阅读框61(cxorf61);cd97;cd179a;间变性淋巴瘤激酶(alk);聚唾液酸;胎盘特异性1(plac1);globoh糖神经酰胺六糖部分(globoh);乳腺分化抗原(ny-br-1);尿空斑蛋白2(upk2);甲型肝炎病毒细胞受体1(havcr1);肾上腺受体β3(adrb3);泛连接蛋白3(panx3);g蛋白偶联受体20(gpr20);淋巴细胞抗原6复合物,基因座k9(ly6k);嗅觉受体51e2(or51e2);tcrγ替代性阅读框蛋白质(tarp);威尔姆氏瘤蛋白质(wt1);癌症/睾丸抗原1(ny-es0-1);癌症/睾丸抗原2(lage-1a);黑素瘤相关抗原1(mage-a1);位于染色体12p上的ets易位变体基因6(etv6-aml);精子蛋白17(spa17);x抗原家族成员la(xagel);血管生成素结合细胞表面受体2(tie2);黑素瘤癌症睾丸抗原-1(mad-ct-1);黑素瘤癌症睾丸抗原-2(mad-ct-2);fos相关抗原1;肿瘤蛋白p53(p53);p53突变体;前列腺特异性蛋白;存活素;端粒酶;前列腺癌肿瘤抗原-1(pcta-1或半乳凝素8)、由t细胞识别的黑素瘤抗原1(melana或marti);大鼠肉瘤(ras)突变体;人类端粒酶逆转录酶(htert);肉瘤易位裂点;黑素瘤细胞凋亡抑制剂(ml-iap);erg(跨膜蛋白酶,丝氨酸2(tmprss2)ets融合基因);n-乙酰基葡萄糖胺基-转移酶v(na17);配对框蛋白质pax-3(pax3);雄激素受体;周期素bl;v-myc禽类骨髓细胞瘤病病毒癌基因神经母细胞瘤衍生性同源物(mycn);ras同源家族成员c(rhoc);酪氨酸酶相关蛋白2(trp-2);细胞色素p450lb1(cyplb1);ccctc-结合因子(锌指蛋白)样(boris或印迹位点调节剂同级物)、由t细胞识别的鳞状细胞癌抗原3(sart3);配对框蛋白质pax-5(pax5);前顶体蛋白结合蛋白sp32(oy-tesl);淋巴细胞特异性蛋白质酪氨酸激酶(lck);a激酶锚定蛋白4(akap-4);滑膜肉瘤,x裂点2(ssx2);晚期糖基化终产物受体(rage-1);肾泛素1(rul);肾泛素2(ru2);豆荚蛋白;人乳头状瘤病毒e6(hpve6);人乳头状瘤病毒e7(hpve7);肠道羧基酯酶;突变的热休克蛋白70-2(muthsp70-2);cd79a;cd79b;cd72;白细胞相关免疫球蛋白样受体1(lairl);iga受体的fc片段(fcar或cd89);白细胞免疫球蛋白样受体子家族a成员2(lilra2);cd300分子样家族成员f(cd300lf);c型凝集素结构域家族12成员a(clec12a);骨髓基质细胞抗原2(bst2);egf样含模块粘蛋白样激素受体样2(emr2);淋巴细胞抗原75(ly75);磷脂酰肌醇聚糖-3(gpc3);fc受体样5(fcrl5);以及免疫球蛋白λ样多肽1(iglll)、mpl、、生物素、c-myc表位标签、cd34、lamp1trop2、gfrα54、cdh17、cdh6、nybr1、cdh19、cd200r、slea(ca19.9;唾液酸化路易斯抗原)岩藻糖基-gm1、ptk7、gpnmb、cdh1-cd324、dll3、cd276/b7h3、il11ra、il13ra2、cd179b-igll1、alktcrγ-δ、nkg2d、cd32(fcgr2a)、tnag、cspg4-hmw-maa、tim1-/hvcr1、csf2ra(gm-csfr-α)、tgfβr2、vegfr2/kdr、lewsag、tcr-β1链、tcr-β2链、tcr-γ链、tcr-δ链、fitc、促黄体生成激素受体(lhr)、卵泡刺激素受体(fshr)、绒膜促性腺激素激素受体(cghr)、ccr4、gd3、slamf6、slamf4、hiv1包膜糖蛋白、htlv1-tax、cmvpp65、ebv-ebna3c、甲型流感凝集素(ha)、gad、pdl1、鸟苷酸环化酶c(gcc)、kshv-k8.1蛋白、kshv-gh蛋白、桥粒芯蛋白自身抗体3(dsg3)、桥粒芯蛋白自身抗体1(dsg1)、hla、hla-a、hla-a2、hla-b、hla-c、hla-dp、hla-dm、hla-doa、hla-dob、hla-dq、hla-dr、hla-g、ige、cd99、rasg12v、组织因子1(tf1)、afp、gprc5d、密封蛋白18.2(cld18a2或cldn18a.2))、p-糖蛋白、steap1、liv1、粘连蛋白-4、cripto、gpa33、bst1/cd157、低电导率氯离子通道和由tnt抗体识别的抗原。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域包含抗体、抗体片段、scfv、fv、fab、(fab')2、单结构域抗体(sdab)、vh或vl结构域、骆驼科vhh结构域、非免疫球蛋白抗原结合支架,例如darpin、亲合体、affilin、adnectin、affitin、obodies、repebody、fynomer、alphabody、avimer、atrimer、centyrin、pronectin、anticalin、kunitz型结构域、armadillo重复蛋白、受体或配体。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域选自由以下组成的组:(i)包含seqidno2506至2680或12160至12278中任一个所示的序列或与其有至少98%一致性的序列并编码结合至其抗原的多肽的重链可变区(vh);(ii)包含seqidno2307至2482或12042至12159中任一个所示的序列或与其有至少98%一致性的序列并编码结合至其抗原的多肽的轻链可变区(vl);(iii)包含seqidno:2770至2939、12303至12357或18162至18224中任一个所示的序列或与其有至少98%一致性的序列并编码结合至其抗原的多肽的单链可变片段(scfv);(iv)包含seqidno2701至2725或12279至12294中任一个所示的序列或与其有至少98%一致性的序列并编码结合至其抗原的多肽的骆驼科vhh结构域;(v)由seqidno439至443中任一个或与其有至少98%一致性的序列并编码结合至其抗原的多肽的多核苷酸编码的非免疫球蛋白支架;(vi)包含seqidno2736至2748中任一个所示的序列或与其有至少98%一致性的序列并编码结合至其同源物的多肽的受体;以及(vii)包含seqidno2758至2768或12359至12361中任一个所示的序列或与其有至少98%一致性的序列并编码结合至其同源物的多肽的配体。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域包含seqidno:13999至14879或14880中任一个所示的所选靶抗原的轻链互补决定区中的一个或多个和/或seqidno:14881至15761或15762中任一个所示的所选靶抗原的重链互补决定区中的一个或多个。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域包含含有seqidno:2307至2482或12042至12159中任一个的序列并具有至多10个保守性氨基酸取代的可变轻链(vl)结构域和/或含有seqidno:2506至2680或12160至12278中任一个的序列并具有至多10个保守性氨基酸取代的可变重链(vh)结构域。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域包含seqidno:2701至2725或12279至12294中任一个所示并具有至多10个保守性氨基酸取代的所选抗原的骆驼科vhh互补决定区中的一个或多个。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域包含具有seqidno:2728-2732或12296至12301中任一个所示的序列并具有至多10个保守性氨基酸取代的非免疫球蛋白抗原结合结构域。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域包含scfv结构域,该结构域包含含有seqidno:2307至2482或12042至12159中任一个的序列的可变轻链(vl)结构域的一个或多个轻链互补决定区和含有seqidno:2506至2680或12160至12278中任一个的序列的可变重链(vh)结构域的一个或多个重链互补决定区。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域包含具有选自由seqidno:2770至2939、12303至12357或18162至18224组成的组中的序列的scfv片段,其各自具有至多10个保守性氨基酸取代。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域包含含有seqidno:2736至2748中任一个的氨基酸序列并具有至多10个保守性氨基酸取代的一个或多个受体。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域包含含有seqidno:2758-2768或12359至12361中任一个的序列并具有至多10个保守性氨基酸取代的一个或多个配体。在前述任一者的另一个实施方式中,该一个或多个非天然tcr抗原结合结构域包含cd16a、nkg2d、cd4、pd1、桥粒芯蛋白3(dsg3)或cd4-dc-sign的细胞外结构域。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含htpo、mtpo、cghα链、cghβ链、fhβ链、lhβ链、tshβ链、april或其组合中的一个或多个的细胞外结构域的细胞外结构域。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含含有seqidno:2770至2939、12303至12357或18162至18224中任一个的序列并具有至多10个保守性氨基酸取代的任何单链可变片段(scfv),以及a)seqidno:2701至2725或12279至12294中任一个所示的并具有至多10个保守性氨基酸取代的任何骆驼科vhh,或b)具有seqidno:2728-2732或12296至12301中任一个所示的序列并具有至多10个保守性氨基酸取代的任何非免疫球蛋白抗原结合结构域;或c)包含seqidno:2736至2748中任一个的氨基酸序列并具有至多10个保守性氨基酸取代的受体的任何细胞外结构域;或d)包含seqidno:2758-2768或12359至12361中任一个的序列并具有至多10个保守性氨基酸取代的配体的任何细胞外结构域。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域包含含有seqidno:2701至2725或12279至12294中任一个所示并具有至多10个保守性氨基酸取代的骆驼科vhh,以及a)包含seqidno:2770至2939、12303至12357或18162至18224中任一个的序列并具有至多10个保守性氨基酸取代的任何单链可变片段(scfv),或b)具有seqidno:2728-2732或12296至12301中任一个所示的序列并具有至多10个保守性氨基酸取代的任何非免疫球蛋白抗原结合结构域;或c)包含seqidno:2736至2748中任一个的氨基酸序列并具有至多10个保守性氨基酸取代的受体的任何细胞外结构域;或d)包含seqidno:2758-2768或12359至12361中任一个的序列并具有至多10个保守性氨基酸取代的配体的任何细胞外结构域。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域通过接头区任选地连接至每条tcr恒定区链,其中所述接头区核酸编码选自由seqidno:2981至2992及其任何组合或与其有至少98%一致性的序列组成的组中的氨基酸序列;或者所述接头由选自由seqidno:701至714或与其有至少98%一致性的序列组成的组中的核酸序列编码。在前述任一者的另一个实施方式中,所述一个或多个非天然tcr抗原结合结构域与其靶抗原的结合亲和力比从中获得该结合结构域的抗体小至少5倍。在前述任一者的另一个实施方式中,编码sir的多核苷酸还包含存在于每条链的n末端处并包含选自由seqidno:1-9和10组成的组中的序列的前导序列或信号肽。在前述任一者的另一个实施方式中,sir包含sir异二聚体。在前述任一者的另一个实施方式中,该多肽包含通过可裂解接头连接的两个sir。在另一个实施方式中,该可裂解接头是自身裂解的可裂解接头。在另一个实施方式中,可裂解接头是2a接头、2a样接头或其功能等效物中的任一个或多个。在又一个实施方式中,可裂解接头是t2a接头、p2a、f2a、e2a接头或其功能等效物中的任一个或多个。在又一个实施方式中,可裂解接头包含seqidno:780至785中任一个或多个的序列。在另一个实施方式中,可裂解接头任选地在弗林蛋白酶裂解位点或弗林蛋白酶样裂解位点或其功能等效物后面。在又一个实施方式中,在可裂解接头前面的弗林蛋白酶样裂解位点包含seqidno:788至790中任一个或多个的序列。在任一前述实施方式中,可裂解接头在柔性接头后面。在另一个实施方式中,在可裂解接头前面的柔性接头编码ser-gly接头、ser-gly-ser-gly接头或其功能等效物中的一个或多个。在另一个实施方式中,在可裂解接头前面的柔性接头包含序列seqidno:786或787。在又一个实施方式中,弗林蛋白酶裂解位点之后为柔性接头,在其之后为可裂解接头,使得该次序为弗林蛋白酶裂解位点-柔性接头-可裂解接头。在前述任一者的另一个实施方式中,sir设计为对所选抗原具有希望的结合亲和力。本公开还提供一种免疫效应细胞或干细胞,该细胞包含至少一种如本文上文所述的多肽或异二聚体。本公开还提供一种免疫效应细胞或干细胞,该细胞包含至少一种如本文上文所述的重组多核苷酸。本公开还提供一种免疫效应细胞或干细胞,该细胞包含至少一种如本文上文所述的本公开的载体。在前述任一者的另一个实施方式中,任何免疫细胞或干细胞包含多种sir多肽。在前述任一者的另一个实施方式中,多种sir多肽中的至少一种sir多肽靶向不同于至少另一种sir多肽的抗原。在前述任一者的另一个实施方式中,多种sir多肽中的至少一种sir多肽靶向相同抗原。在前述任一者的另一个实施方式中,多种sir多肽中的至少一种sir多肽包含不同于至少另一种sir多肽的对抗原的结合亲和力。在前述任一者的另一个实施方式中,免疫细胞还包含至少一种嵌合抗原受体(car)多肽。在前述任一者的另一个实施方式中,sir多肽的抗原结合结构域靶向不同于car多肽的抗原结合结构域的抗原。在前述任一者的另一个实施方式中,car多肽包含含有共刺激信号传导结构域的细胞内信号传导结构域,但不包含初级信号传导结构域,或者包含含有初级信号传导结构域的细胞内信号传导结构域,但不包含共刺激信号传导结构域。在前述任一者的另一个实施方式中,car多肽包含含有选自由4-lbb、cd28、cd27或ox-40组成的组中的蛋白质的功能性信号传导结构域的共刺激信号传导结构域,或者car分子包含含有cd3ζ的功能性信号传导结构域的初级信号传导结构域。在前述任一者的另一个实施方式中,car多肽为抑制性car多肽,其中该抑制性car多肽包含抑制性分子的抗原结合结构域、跨膜结构域和细胞内结构域,其中该抑制性分子选自由pdl、pd-ll、ctla4、tim3、lag3、vista、btla、tigit、lairl、cd160、2b4、tgfrβ、ceacam-1、ceacam-3和ceacam-5组成的组。在前述任一者的另一个实施方式中,car多肽还包含含有初级信号传导结构域的细胞内信号传导结构域和/或细胞内信号传导结构域,其中该细胞内信号传导结构域包含含有cd3ζ的功能结构域的初级信号传导结构域和含有4-lbb或cd28或两者的功能结构域的共刺激信号传导结构域。在前述任一者的另一个实施方式中,car多肽包含氨基酸序列seqidno:3077至seqidno:3083。在前述任一者的另一个实施方式中,免疫效应细胞为人类t细胞、人类nk细胞或可产生免疫效应细胞的干细胞,任选地其中该t细胞是二甘油(diaglycerol)激酶(dgk)和/或伊卡洛斯(ikaros)缺陷的和/或brd4缺陷的。本公开提供一种制备表达sir的免疫效应细胞的方法,该方法包括在使得sir多肽表达的条件下将至少一种本公开的载体或至少一种本公开的重组多核苷酸引入到免疫效应细胞或可产生免疫效应细胞的造血干细胞或祖细胞中。在前述任一者的另一个实施方式中,该方法还包括a)提供免疫效应细胞群体;并且b)从该群体中去除t调控细胞,从而提供t调控耗尽的细胞群体;其中步骤a)和b)在将编码sir的载体或重组多核苷酸引入群体之前进行。在前述任一者的另一个实施方式中,t调控细胞使用抗-cd25抗体或抗-gitr抗体从细胞群体中去除。在前述任一者的另一个实施方式中,该方法还包括:a)提供免疫效应细胞群体;并且b)将来自群体的p-糖蛋白(p-gp或pgp;mdr1、abcb1、cd243)阳性细胞富集,从而提供富含p-糖蛋白(p-gp或pgp;mdr1、abcb1、cd243)的细胞;其中步骤a)和b)在引入编码sir的载体或重组多核苷酸之前或之后进行。在前述任一者的另一个实施方式中,p-糖蛋白阳性细胞使用选自由以下组成的组中的任一种或多种细胞富集:i)使用一种p-糖蛋白特异性抗体或其混合物进行的免疫选择,ii)在p-糖蛋白作为泵起作用的条件下使用一种或多种作为p-糖蛋白底物的荧光染料(四甲基若丹明甲酯(tmrm)、阿霉素和放线菌素-d)进行染色并富集用该染料染色较少的细胞,iii)选择对作为p-糖蛋白底物的光毒性化合物有抗性的细胞,这些化合物诸如th9402、2-(4,5-二溴-6-氨基-3-亚氨基-3h-氧杂蒽-9-基)-苯甲酸甲酯盐酸盐、2-(4,5-二溴-6-氨基-3-亚氨基-3h-氧杂蒽-9-基)-苯甲酸乙酯盐酸盐、2-(4,5-二溴-6-氨基-3-亚氨基-3h-氧杂蒽-9-基)-苯甲酸辛酯盐酸盐、2-(4,5-二溴-6-氨基-3-亚氨基-3h-氧杂蒽-9-基)-苯甲酸正丁酯盐酸盐、2-(6-乙基氨基-3-乙基亚氨基-3h-氧杂蒽-9-基)-苯甲酸正丁酯盐酸盐或其衍生物、或其组合中的任一种或多种,以及iv)选择对作为p-糖蛋白底物的细胞毒性化合物有抗性的细胞,这些化合物诸如长春新碱、长春花碱、紫杉酚、紫杉醇、米托蒽醌、依托泊苷、阿霉素、红比霉素和放线菌素-d。本公开还提供一种生成rna工程化细胞群体的方法,该方法包括将体外转录的一个或多个rna或合成的一个或多个rna引入细胞或细胞群体中,其中该一个或多个rna包含本文上文所述的一个或多个重组多核苷酸。本公开提供一种提供受试者抗疾病免疫力的方法,该方法包括向该受试者给予有效量的本公开的免疫效应细胞或可产生免疫效应细胞的干细胞,其中该细胞是自体t细胞或同种异体t细胞、或自体nk细胞或同种异体nk细胞、或可产生免疫效应细胞的自体或同种异体造血干细胞。在一个实施方式中,同种异体t细胞或同种异体nk细胞缺乏功能性tcr或功能性hla的表达或具有其低表达。本公开还提供一种包含免疫效应细胞或可生成免疫效应细胞的干细胞的组合物,该细胞包含一种或多种合成免疫受体(sir)分子,该组合物用于与增加免疫效应细胞的功效的药剂组合以供在治疗患有与疾病相关抗原表达相关的疾病的受试者或预防具有增加的与疾病相关抗原表达相关的疾病风险的受试者的疾病中使用,其中:(i)该sir分子包含经由任选接头连接至一个或多个抗原结合结构域的t细胞受体恒定链中的一条或多条,该一个或多个抗原结合结构域结合至与该疾病相关的该疾病相关抗原,并且所述疾病相关抗原选自由以下组成的组:cd5、cd19;cd123;cd22;cd30;cd171;cs-1(也称为cd2亚单位1、cracc、slamf7、cd319和19a24);c型凝集素样分子-1(cll-1或clecl1);cd33;表皮生长因子受体变体iii(egfrviii);神经节苷脂g2(gd2);神经节苷脂gd3(aneu5ac(2-8)aneu5ac(2-3)bdgalp(l-4)bdglcp(l-l)cer);tnf受体家族成员b细胞成熟(bcma);tn抗原((tnag)或(galnacα-ser/thr));前列腺特异性膜抗原(psma);受体酪氨酸激酶样孤儿受体1(ror1);fms样酪氨酸激酶3(flt3);肿瘤相关性糖蛋白72(tag72);cd38;cd44v6;在急性白血病或淋巴瘤上而不在造血祖细胞上表达的糖基化cd43表位、在非造血癌症上表达的糖基化cd43表位、癌胚抗原(cea);表皮细胞粘附分子(epcam);b7h3(cd276);kit(cd117);白介素-13受体亚基α-2(il-13ra2或cd213a2);间皮素;白介素11受体α(il-llra);前列腺干细胞抗原(psca);蛋白酶丝氨酸21(睾蛋白或prss21);血管内皮生长因子受体2(vegfr2);路易斯(y)抗原;cd24;血小板源性生长因子受体β(pdgfr-β);阶段特异性胚胎抗原-4(ssea-4);cd20;叶酸受体α;受体酪氨酸蛋白激酶erbb2(her2/neu);细胞表面相关粘蛋白1(muc1);表皮生长因子受体(egfr);神经细胞粘附分子(ncam);前列腺酶;前列腺酸性磷酸酶(pap);突变的延长因子2(elf2m);肝配蛋白b2;成纤维细胞活化蛋白α(fap);胰岛素样生长因子1受体(igf-i受体)、碳酸酐酶ix(calx);蛋白酶体(前体、巨蛋白因子)亚基,β型,9(lmp2);糖蛋白100(gpl00);由裂点簇区(bcr)和阿贝尔森(abelson)鼠类白血病病毒癌基因同源物1(abl)组成的癌基因融合蛋白(bcr-abl);酪氨酸酶;肝配蛋白a型受体2(epha2);岩藻糖基gm1;唾液酸化路易斯粘附分子(sle);神经节苷脂gm3(aneu5ac(2-3)bdclalp(l-4)bdglcp(l-1)cer);转谷氨酰胺酶5(tgs5);高分子量-黑素瘤相关抗原(hmwmaa);o-乙酰基-gd2神经节苷脂(oacgd2);肿瘤内皮标记1(tem1/cd248);肿瘤内皮标记7相关(tem7r);密封蛋白6(cldn6);甲状腺刺激性激素受体(tshr);g蛋白偶联受体c类5组成员d(gprc5d);x染色体开放阅读框61(cxorf61);cd97;cd179a;间变性淋巴瘤激酶(alk);聚唾液酸;胎盘特异性1(plac1);globoh糖神经酰胺六糖部分(globoh);乳腺分化抗原(ny-br-1);尿空斑蛋白2(upk2);甲型肝炎病毒细胞受体1(havcr1);肾上腺受体β3(adrb3);泛连接蛋白3(panx3);g蛋白偶联受体20(gpr20);淋巴细胞抗原6复合物,基因座k9(ly6k);嗅觉受体51e2(or51e2);tcrγ替代性阅读框蛋白质(tarp);威尔姆氏瘤蛋白质(wt1);癌症/睾丸抗原1(ny-es0-1);癌症/睾丸抗原2(lage-1a);黑素瘤相关抗原1(mage-a1);位于染色体12p上的ets易位变体基因6(etv6-aml);精子蛋白17(spa17);x抗原家族成员la(xagel);血管生成素结合细胞表面受体2(tie2);黑素瘤癌症睾丸抗原-1(mad-ct-1);黑素瘤癌症睾丸抗原-2(mad-ct-2);fos相关抗原1;肿瘤蛋白p53(p53);p53突变体;前列腺特异性蛋白;存活素;端粒酶;前列腺癌肿瘤抗原-1(pcta-1或半乳凝素8)、由t细胞识别的黑素瘤抗原1(melana或marti);大鼠肉瘤(ras)突变体;人类端粒酶逆转录酶(htert);肉瘤易位裂点;黑素瘤细胞凋亡抑制剂(ml-iap);erg(跨膜蛋白酶,丝氨酸2(tmprss2)ets融合基因);n-乙酰基葡萄糖胺基-转移酶v(na17);配对框蛋白质pax-3(pax3);雄激素受体;周期素bl;v-myc禽类骨髓细胞瘤病病毒癌基因神经母细胞瘤衍生性同源物(mycn);ras同源家族成员c(rhoc);酪氨酸酶相关蛋白2(trp-2);细胞色素p450lb1(cyplb1);ccctc-结合因子(锌指蛋白)样(boris或印迹位点调节剂同级物)、由t细胞识别的鳞状细胞癌抗原3(sart3);配对框蛋白质pax-5(pax5);前顶体蛋白结合蛋白sp32(oy-tesl);淋巴细胞特异性蛋白质酪氨酸激酶(lck);a激酶锚定蛋白4(akap-4);滑膜肉瘤,x裂点2(ssx2);晚期糖基化终产物受体(rage-1);肾泛素1(rul);肾泛素2(ru2);豆荚蛋白;人乳头状瘤病毒e6(hpve6);人乳头状瘤病毒e7(hpve7);肠道羧基酯酶;突变的热休克蛋白70-2(muthsp70-2);cd79a;cd79b;cd72;白细胞相关免疫球蛋白样受体1(lairl);iga受体的fc片段(fcar或cd89);白细胞免疫球蛋白样受体子家族a成员2(lilra2);cd300分子样家族成员f(cd300lf);c型凝集素结构域家族12成员a(clec12a);骨髓基质细胞抗原2(bst2);egf样含模块粘蛋白样激素受体样2(emr2);淋巴细胞抗原75(ly75);磷脂酰肌醇聚糖-3(gpc3);fc受体样5(fcrl5);以及免疫球蛋白λ样多肽1(iglll)、mpl、、生物素、c-myc表位标签、cd34、lamp1trop2、gfrα54、cdh17、cdh6、nybr1、cdh19、cd200r、slea(ca19.9;唾液酸化路易斯抗原)岩藻糖基-gm1、ptk7、gpnmb、cdh1-cd324、dll3、cd276/b7h3、il11ra、il13ra2、cd179b-igll1、alktcrγ-δ、nkg2d、cd32(fcgr2a)、cspg4-hmw-maa、tim1-/hvcr1、csf2ra(gm-csfr-α)、tgfβr2、vegfr2/kdr、lewsag、tcr-β1链、tcr-β2链、tcr-γ链、tcr-δ链、fitc、促黄体生成激素受体(lhr)、卵泡刺激素受体(fshr)、绒膜促性腺激素激素受体(cghr)、ccr4、gd3、slamf6、slamf4、hiv1包膜糖蛋白、htlv1-tax、cmvpp65、ebv-ebna3c、甲型流感凝集素(ha)、gad、pdl1、鸟苷酸环化酶c(gcc)、kshv-k8.1蛋白、kshv-gh蛋白、桥粒芯蛋白自身抗体3(dsg3)、桥粒芯蛋白自身抗体1(dsg1)、hla、hla-a、hla-a2、hla-b、hla-c、hla-dp、hla-dm、hla-doa、hla-dob、hla-dq、hla-dr、hla-g、ige、cd99、rasg12v、组织因子1(tf1)、afp、gprc5d、密封蛋白18.2(cld18a2或cldn18a.2))、p-糖蛋白、steap1、liv1、粘连蛋白-4、cripto、gpa33、bst1/cd157、低电导率氯离子通道和由tnt抗体识别的抗原,(ii)增加免疫细胞功效的药剂选自以下一种或多种:蛋白磷酸酶抑制剂;激酶抑制剂(例如pi3k/akt抑制剂或mtor抑制剂);细胞因子;免疫抑制分子抑制剂;降低treg细胞水平或活性的药剂;增加sir修饰细胞的增殖和/或持久性的药剂;趋化因子;增加sir表达的药剂;允许调控sir表达或活性的药剂;允许控制sir修饰细胞的存活和/或持久性的药剂;控制sir修饰细胞的副作用的药剂;brd4抑制剂;将治疗剂(例如shvem)或预防剂递送至疾病位点的药剂;增加sir所针对的靶抗原的表达的药剂;以及腺苷a2a受体拮抗剂。本公开提供一种治疗或预防受试者中与疾病相关抗原的表达相关的疾病的方法,该方法包括向该受试者给予有效量的包含合成免疫受体(sir)分子的免疫效应细胞,与增加免疫细胞功效的药剂的组合,其中:(i)该sir分子包含经由任选接头连接至一个或多个抗原结合结构域的t细胞受体恒定链中的一条或多条,该一个或多个抗原结合结构域结合至与该疾病相关的该疾病相关抗原,并且所述疾病相关抗原选自由以下组成的组:cd5、cd19;cd123;cd22;cd30;cd171;cs-1(也称为cd2亚单位1、cracc、slamf7、cd319和19a24);c型凝集素样分子-1(cll-1或clecl1);cd33;表皮生长因子受体变体iii(egfrviii);神经节苷脂g2(gd2);神经节苷脂gd3(aneu5ac(2-8)aneu5ac(2-3)bdgalp(l-4)bdglcp(l-l)cer);tnf受体家族成员b细胞成熟(bcma);tn抗原((tnag)或(galnacα-ser/thr));前列腺特异性膜抗原(psma);受体酪氨酸激酶样孤儿受体1(ror1);fms样酪氨酸激酶3(flt3);肿瘤相关性糖蛋白72(tag72);cd38;cd44v6;在急性白血病或淋巴瘤上而不在造血祖细胞上表达的糖基化cd43表位、在非造血癌症上表达的糖基化cd43表位、癌胚抗原(cea);表皮细胞粘附分子(epcam);b7h3(cd276);kit(cd117);白介素-13受体亚基α-2(il-13ra2或cd213a2);间皮素;白介素11受体α(il-llra);前列腺干细胞抗原(psca);蛋白酶丝氨酸21(睾蛋白或prss21);血管内皮生长因子受体2(vegfr2);路易斯(y)抗原;cd24;血小板源性生长因子受体β(pdgfr-β);阶段特异性胚胎抗原-4(ssea-4);cd20;叶酸受体α;受体酪氨酸蛋白激酶erbb2(her2/neu);细胞表面相关粘蛋白1(muc1);表皮生长因子受体(egfr);神经细胞粘附分子(ncam);前列腺酶;前列腺酸性磷酸酶(pap);突变的延长因子2(elf2m);肝配蛋白b2;成纤维细胞活化蛋白α(fap);胰岛素样生长因子1受体(igf-i受体)、碳酸酐酶ix(calx);蛋白酶体(前体、巨蛋白因子)亚基,β型,9(lmp2);糖蛋白100(gpl00);由裂点簇区(bcr)和阿贝尔森(abelson)鼠类白血病病毒癌基因同源物1(abl)组成的癌基因融合蛋白(bcr-abl);酪氨酸酶;肝配蛋白a型受体2(epha2);岩藻糖基gm1;唾液酸化路易斯粘附分子(sle);神经节苷脂gm3(aneu5ac(2-3)bdclalp(l-4)bdglcp(l-1)cer);转谷氨酰胺酶5(tgs5);高分子量-黑素瘤相关抗原(hmwmaa);o-乙酰基-gd2神经节苷脂(oacgd2);叶酸受体β;肿瘤内皮标记1(tem1/cd248);肿瘤内皮标记7相关(tem7r);密封蛋白6(cldn6);甲状腺刺激性激素受体(tshr);g蛋白偶联受体c类5组成员d(gprc5d);x染色体开放阅读框61(cxorf61);cd97;cd179a;间变性淋巴瘤激酶(alk);聚唾液酸;胎盘特异性1(plac1);globoh糖神经酰胺六糖部分(globoh);乳腺分化抗原(ny-br-1);尿空斑蛋白2(upk2);甲型肝炎病毒细胞受体1(havcr1);肾上腺受体β3(adrb3);泛连接蛋白3(panx3);g蛋白偶联受体20(gpr20);淋巴细胞抗原6复合物,基因座k9(ly6k);嗅觉受体51e2(or51e2);tcrγ替代性阅读框蛋白质(tarp);威尔姆氏瘤蛋白质(wt1);癌症/睾丸抗原1(ny-es0-1);癌症/睾丸抗原2(lage-1a);黑素瘤相关抗原1(mage-a1);位于染色体12p上的ets易位变体基因6(etv6-aml);精子蛋白17(spa17);x抗原家族成员la(xagel);血管生成素结合细胞表面受体2(tie2);黑素瘤癌症睾丸抗原-1(mad-ct-1);黑素瘤癌症睾丸抗原-2(mad-ct-2);fos相关抗原1;肿瘤蛋白p53(p53);p53突变体;前列腺特异性蛋白;存活素;端粒酶;前列腺癌肿瘤抗原-1(pcta-1或半乳凝素8)、由t细胞识别的黑素瘤抗原1(melana或marti);大鼠肉瘤(ras)突变体;人类端粒酶逆转录酶(htert);肉瘤易位裂点;黑素瘤细胞凋亡抑制剂(ml-iap);erg(跨膜蛋白酶,丝氨酸2(tmprss2)ets融合基因);n-乙酰基葡萄糖胺基-转移酶v(na17);配对框蛋白质pax-3(pax3);雄激素受体;周期素bl;v-myc禽类骨髓细胞瘤病病毒癌基因神经母细胞瘤衍生性同源物(mycn);ras同源家族成员c(rhoc);酪氨酸酶相关蛋白2(trp-2);细胞色素p450lb1(cyplb1);ccctc-结合因子(锌指蛋白)样(boris或印迹位点调节剂同级物)、由t细胞识别的鳞状细胞癌抗原3(sart3);配对框蛋白质pax-5(pax5);前顶体蛋白结合蛋白sp32(oy-tesl);淋巴细胞特异性蛋白质酪氨酸激酶(lck);a激酶锚定蛋白4(akap-4);滑膜肉瘤,x裂点2(ssx2);晚期糖基化终产物受体(rage-1);肾泛素1(rul);肾泛素2(ru2);豆荚蛋白;人乳头状瘤病毒e6(hpve6);人乳头状瘤病毒e7(hpve7);肠道羧基酯酶;突变的热休克蛋白70-2(muthsp70-2);cd79a;cd79b;cd72;白细胞相关免疫球蛋白样受体1(lairl);iga受体的fc片段(fcar或cd89);白细胞免疫球蛋白样受体子家族a成员2(lilra2);cd300分子样家族成员f(cd300lf);c型凝集素结构域家族12成员a(clec12a);骨髓基质细胞抗原2(bst2);egf样含模块粘蛋白样激素受体样2(emr2);淋巴细胞抗原75(ly75);磷脂酰肌醇聚糖-3(gpc3);fc受体样5(fcrl5);以及免疫球蛋白λ样多肽1(iglll)、mpl、、生物素、c-myc表位标签、cd34、lamp1trop2、gfrα54、cdh17、cdh6、nybr1、cdh19、cd200r、slea(ca19.9;唾液酸化路易斯抗原)岩藻糖基-gm1、ptk7、gpnmb、cdh1-cd324、dll3、cd276/b7h3、il11ra、il13ra2、cd179b-igll1、alktcrγ-δ、nkg2d、cd32(fcgr2a)、cspg4-hmw-maa、tim1-/hvcr1、csf2ra(gm-csfr-α)、tgfβr2、vegfr2/kdr、lewsag、tcr-β1链、tcr-β2链、tcr-γ链、tcr-δ链、fitc、促黄体生成激素受体(lhr)、卵泡刺激素受体(fshr)、绒膜促性腺激素激素受体(cghr)、ccr4、gd3、slamf6、slamf4、hiv1包膜糖蛋白、htlv1-tax、cmvpp65、ebv-ebna3c、甲型流感凝集素(ha)、gad、pdl1、鸟苷酸环化酶c(gcc)、kshv-k8.1蛋白、kshv-gh蛋白、桥粒芯蛋白自身抗体3(dsg3)、桥粒芯蛋白自身抗体1(dsg1)、hla、hla-a、hla-a2、hla-b、hla-c、hla-dp、hla-dm、hla-doa、hla-dob、hla-dq、hla-dr、hla-g、ige、cd99、rasg12v、组织因子1(tf1)、afp、gprc5d、密封蛋白18.2(cld18a2或cldn18a.2))、p-糖蛋白、steap1、liv1、粘连蛋白-4、cripto、gpa33、bst1/cd157、低电导率氯离子通道和由tnt抗体识别的抗原,(ii)增加免疫细胞功效的药剂选自以下一种或多种:蛋白磷酸酶抑制剂;激酶抑制剂(例如pi3k/akt抑制剂或mtor抑制剂);细胞因子;免疫抑制分子抑制剂;降低treg细胞水平或活性的药剂;增加sir修饰细胞的增殖和/或持久性的药剂;趋化因子;增加sir表达的药剂;允许调控sir表达或活性的药剂;允许控制sir修饰细胞的存活和/或持久性的药剂;控制sir修饰细胞的副作用的药剂;brd4抑制剂;将治疗剂(例如shvem)或预防剂递送至疾病位点的药剂;增加sir所针对的靶抗原的表达的药剂;以及腺苷a2a受体拮抗剂,从而治疗受试者或预防受试者的疾病。本公开提供一种治疗或预防受试者中与疾病相关抗原的表达相关的疾病的方法,该方法包括向该受试者给予有效量的包含合成免疫受体(sir)分子的免疫效应细胞,其中:(i)该sir分子包含经由任选接头连接至一个或多个抗原结合结构域的t细胞受体恒定链中的一条或多条,该一个或多个抗原结合结构域结合至与该疾病相关的该疾病相关抗原,并且所述疾病相关抗原选自由以下组成的组:cd5、cd19;cd123;cd22;cd30;cd171;cs-1(也称为cd2亚单位1、cracc、slamf7、cd319和19a24);c型凝集素样分子-1(cll-1或clecl1);cd33;表皮生长因子受体变体iii(egfrviii);神经节苷脂g2(gd2);神经节苷脂gd3(aneu5ac(2-8)aneu5ac(2-3)bdgalp(l-4)bdglcp(l-l)cer);tnf受体家族成员b细胞成熟(bcma);tn抗原((tnag)或(galnacα-ser/thr));前列腺特异性膜抗原(psma);受体酪氨酸激酶样孤儿受体1(ror1);fms样酪氨酸激酶3(flt3);肿瘤相关性糖蛋白72(tag72);cd38;cd44v6;在急性白血病或淋巴瘤上而不在造血祖细胞上表达的糖基化cd43表位、在非造血癌症上表达的糖基化cd43表位、癌胚抗原(cea);表皮细胞粘附分子(epcam);b7h3(cd276);kit(cd117);白介素-13受体亚基α-2(il-13ra2或cd213a2);间皮素;白介素11受体α(il-llra);前列腺干细胞抗原(psca);蛋白酶丝氨酸21(睾蛋白或prss21);血管内皮生长因子受体2(vegfr2);路易斯(y)抗原;cd24;血小板源性生长因子受体β(pdgfr-β);阶段特异性胚胎抗原-4(ssea-4);cd20;叶酸受体α(fra或fr1);叶酸受体β(frb);受体酪氨酸蛋白激酶erbb2(her2/neu);细胞表面相关粘蛋白1(muc1);表皮生长因子受体(egfr);神经细胞粘附分子(ncam);前列腺酶;前列腺酸性磷酸酶(pap);突变的延长因子2(elf2m);肝配蛋白b2;成纤维细胞活化蛋白α(fap);胰岛素样生长因子1受体(igf-i受体)、碳酸酐酶ix(calx);蛋白酶体(前体、巨蛋白因子)亚基,β型,9(lmp2);糖蛋白100(gpl00);由裂点簇区(bcr)和阿贝尔森(abelson)鼠类白血病病毒癌基因同源物1(abl)组成的癌基因融合蛋白(bcr-abl);酪氨酸酶;肝配蛋白a型受体2(epha2);岩藻糖基gm1;唾液酸化路易斯粘附分子(sle);神经节苷脂gm3(aneu5ac(2-3)bdclalp(l-4)bdglcp(l-1)cer);转谷氨酰胺酶5(tgs5);高分子量-黑素瘤相关抗原(hmwmaa);o-乙酰基-gd2神经节苷脂(oacgd2);肿瘤内皮标记1(tem1/cd248);肿瘤内皮标记7相关(tem7r);密封蛋白6(cldn6);甲状腺刺激性激素受体(tshr);g蛋白偶联受体c类5组成员d(gprc5d);x染色体开放阅读框61(cxorf61);cd97;cd179a;间变性淋巴瘤激酶(alk);聚唾液酸;胎盘特异性1(plac1);globoh糖神经酰胺六糖部分(globoh);乳腺分化抗原(ny-br-1);尿空斑蛋白2(upk2);甲型肝炎病毒细胞受体1(havcr1);肾上腺受体β3(adrb3);泛连接蛋白3(panx3);g蛋白偶联受体20(gpr20);淋巴细胞抗原6复合物,基因座k9(ly6k);嗅觉受体51e2(or51e2);tcrγ替代性阅读框蛋白质(tarp);威尔姆氏瘤蛋白质(wt1);癌症/睾丸抗原1(ny-es0-1);癌症/睾丸抗原2(lage-1a);黑素瘤相关抗原1(mage-a1);位于染色体12p上的ets易位变体基因6(etv6-aml);精子蛋白17(spa17);x抗原家族成员la(xagel);血管生成素结合细胞表面受体2(tie2);黑素瘤癌症睾丸抗原-1(mad-ct-1);黑素瘤癌症睾丸抗原-2(mad-ct-2);fos相关抗原1;肿瘤蛋白p53(p53);p53突变体;前列腺特异性蛋白;存活素;端粒酶;前列腺癌肿瘤抗原-1(pcta-1或半乳凝素8)、由t细胞识别的黑素瘤抗原1(melana或marti);大鼠肉瘤(ras)突变体;人类端粒酶逆转录酶(htert);肉瘤易位裂点;黑素瘤细胞凋亡抑制剂(ml-iap);erg(跨膜蛋白酶,丝氨酸2(tmprss2)ets融合基因);n-乙酰基葡萄糖胺基-转移酶v(na17);配对框蛋白质pax-3(pax3);雄激素受体;周期素bl;v-myc禽类骨髓细胞瘤病病毒癌基因神经母细胞瘤衍生性同源物(mycn);ras同源家族成员c(rhoc);酪氨酸酶相关蛋白2(trp-2);细胞色素p450lb1(cyplb1);ccctc-结合因子(锌指蛋白)样(boris或印迹位点调节剂同级物)、由t细胞识别的鳞状细胞癌抗原3(sart3);配对框蛋白质pax-5(pax5);前顶体蛋白结合蛋白sp32(oy-tesl);淋巴细胞特异性蛋白质酪氨酸激酶(lck);a激酶锚定蛋白4(akap-4);滑膜肉瘤,x裂点2(ssx2);晚期糖基化终产物受体(rage-1);肾泛素1(rul);肾泛素2(ru2);豆荚蛋白;人乳头状瘤病毒e6(hpve6);人乳头状瘤病毒e7(hpve7);肠道羧基酯酶;突变的热休克蛋白70-2(muthsp70-2);cd79a;cd79b;cd72;白细胞相关免疫球蛋白样受体1(lairl);iga受体的fc片段(fcar或cd89);白细胞免疫球蛋白样受体子家族a成员2(lilra2);cd300分子样家族成员f(cd300lf);c型凝集素结构域家族12成员a(clec12a);骨髓基质细胞抗原2(bst2);egf样含模块粘蛋白样激素受体样2(emr2);淋巴细胞抗原75(ly75);磷脂酰肌醇聚糖-3(gpc3);fc受体样5(fcrl5);以及免疫球蛋白λ样多肽1(iglll)、mpl、、生物素、c-myc表位标签、cd34、lamp1trop2、gfrα54、cdh17、cdh6、nybr1、cdh19、cd200r、slea(ca19.9;唾液酸化路易斯抗原)岩藻糖基-gm1、ptk7、gpnmb、cdh1-cd324、dll3、cd276/b7h3、il11ra、il13ra2、cd179b-igll1、alktcrγ-δ、nkg2d、cd32(fcgr2a)、cspg4-hmw-maa、tim1-/hvcr1、csf2ra(gm-csfr-α)、tgfβr2、vegfr2/kdr、lewsag、tcr-β1链、tcr-β2链、tcr-γ链、tcr-δ链、fitc、促黄体生成激素受体(lhr)、卵泡刺激素受体(fshr)、绒膜促性腺激素激素受体(cghr)、ccr4、gd3、slamf6、slamf4、hiv1包膜糖蛋白、htlv1-tax、cmvpp65、ebv-ebna3c、甲型流感凝集素(ha)、gad、pdl1、鸟苷酸环化酶c(gcc)、kshv-k8.1蛋白、kshv-gh蛋白、桥粒芯蛋白自身抗体3(dsg3)、桥粒芯蛋白自身抗体1(dsg1)、hla、hla-a、hla-a2、hla-b、hla-c、hla-dp、hla-dm、hla-doa、hla-dob、hla-dq、hla-dr、hla-g、ige、cd99、rasg12v、组织因子1(tf1)、afp、gprc5d、密封蛋白18.2(cld18a2或cldn18a.2))、p-糖蛋白、steap1、liv1、粘连蛋白-4、cripto、gpa33、bst1/cd157、低电导率氯离子通道和由tnt抗体识别的抗原;以及(ii)sir分子的结合结构域具有的结合亲和力比该抗原结合结构域所来源的抗体小至少5倍。在前述方法或用途任一者的另一个实施方式中,与疾病相关抗原的表达相关的疾病选自由增生性疾病、癌前病状、癌症以及与该疾病相关抗原的表达相关的非癌症相关适应症组成的组。在前述方法或用途任一者的另一个实施方式中,该癌症是选自慢性淋巴细胞性白血病(cll)、急性白血病、急性淋巴细胞白血病(all)、b细胞急性淋巴细胞白血病(b-all)、t细胞急性淋巴细胞白血病(t-all)、慢性髓细胞性白血病(cml)、b细胞幼淋巴细胞性白血病、母细胞性浆细胞样树突细胞肿瘤、伯基特淋巴瘤、弥漫性大b细胞淋巴瘤、原发性渗出性淋巴瘤、滤泡性淋巴瘤、毛细胞白血病、小细胞或大细胞滤泡性淋巴瘤、恶性淋巴组织增生病状、malt淋巴瘤、套细胞淋巴瘤、边缘区淋巴瘤、多发性骨髓瘤、骨髓增生异常和骨髓增生异常综合征、非霍奇金淋巴瘤、霍奇金淋巴瘤、浆母细胞性淋巴瘤、浆细胞样树突细胞肿瘤、瓦尔登斯特伦巨球蛋白血症或白血病前期中一种或多种的血液癌症。在前述方法或用途任一者的另一个实施方式中,该癌症选自由结肠癌、直肠癌、肾细胞癌、肝癌、非小细胞肺癌、小肠癌、食道癌、黑素瘤、骨癌、胰腺癌、皮肤癌、头颈癌、皮肤或眼内恶性黑素瘤、子宫癌、卵巢癌、直肠癌、肛区癌、胃癌、睾丸癌、子宫癌、输卵管癌、子宫内膜癌、子宫颈癌、阴道癌、外阴癌、霍奇金病、非霍奇金淋巴瘤、内分泌系统癌症、甲状腺癌、甲状旁腺癌、肾上腺癌、软组织肉瘤、尿道癌、阴茎癌、儿童实体瘤、膀胱癌、肾或输尿管癌、肾盂癌、中枢神经系统肿瘤(cns)、原发性cns淋巴瘤、肿瘤血管生成、脊轴肿瘤、脑干胶质瘤、垂体腺瘤、卡波济肉瘤、默克尔细胞癌、表皮样癌、鳞状细胞癌、t细胞淋巴瘤、环境诱导的癌症、所述癌症的组合和所述癌症的转移性病灶组成的组。在前述方法或用途任一者的另一个实施方式中,该疾病与病毒的感染相关,该病毒包括但不限于hiv1、hiv2、htlv1、爱泼斯坦巴尔病毒(epsteinbarrvirus,ebv)、巨细胞病毒(cmv)、腺病毒、腺相关病毒、bk病毒(ebv)、人类疱疹病毒6、人类疱疹病毒8流感病毒、副流感病毒、禽流感病毒、mers和sars冠状病毒、克里米亚刚果出血热病毒、鼻病毒、肠道病毒、登革病毒、西尼罗河病毒、埃博拉病毒、马尔堡病毒、拉沙热病毒、塞卡病毒、rsv、麻疹病毒、腮腺炎病毒、鼻病毒、水痘病毒、单纯疱疹病毒1和2、水痘-带状疱疹病毒、hiv-1、htlv1、肝炎病毒、肠道病毒、乙型肝炎病毒、丙型肝炎病毒、尼帕病毒和裂谷热病毒、日本脑炎病毒、梅克尔细胞多瘤病毒,或者与结核分枝杆菌、非典型分支杆菌种类、耶氏肺孢子虫、弓形体病、立克次氏体、诺卡氏菌属、曲霉属、毛霉菌或假丝酵母的感染有关。在前述方法或用途任一者的另一个实施方式中,该疾病是免疫或退行性疾病,包括但不限于糖尿病、多发性硬化症、风湿性关节炎、寻常性天疱疮、强直性脊柱炎、下本氏甲状腺炎(hoshimoto’sthyroiditis)、sle、结节病、硬皮病、混合型结缔组织病、移植物抗宿主病或阿尔茨海默病。在前述方法或用途任一者的另一个实施方式中,(i)该蛋白磷酸酶抑制剂是shp-1抑制剂和/或shp-2抑制剂;(ii)该激酶抑制剂选自cdk4抑制剂、cdk4/6抑制剂、mtor抑制剂、mnk抑制剂或双重p13k/mtor抑制剂中的一种或多种;(iii)抑制该免疫抑制性分子的该药剂包括抗体或抗体片段、抑制性核酸、聚集的规则间隔短回文重复序列(crispr)、转录活化因子样效应核酸酶(talen)或抑制抑制性分子表达的锌指核酸内切酶(zfn);(iv)降低这些treg细胞的水平或活性的该药剂选自环磷酰胺、抗gitr抗体、cd25耗尽或其组合;和/或(v)该brd4抑制剂选自如us20140256706a1中所述的jq1、ms417、otxo15、ly303511和brd4抑制剂或其衍生物。在前述方法或用途任一者的另一个实施方式中,免疫抑制性分子选自由pd1、pd-l1、ctla-4、tim-3、lag-3、vista、btla、tigit、lair1、cd160、2b4、tgfβ、ceacam-1、ceacam-3和ceacam-5组成的组。在前述方法或用途任一者的另一个实施方式中,抑制抑制性分子的药剂包含含有抑制性分子的第一多肽或其片段以及向细胞提供阳性信号的第二多肽,并且其中该第一多肽和第二多肽在含有car的免疫细胞上表达,其中(i)该第一多肽包含pd1、pd-l1、ctla-4、tim-3、lag-3、vista、btla、tigit、lair1、cd160、2b4、tgfβ、ceacam-1、ceacam-3和ceacam-5或其片段;和/或(ii)该第二多肽包含细胞内信号传导结构域,该细胞内信号传导结构域包含初级信号传导结构域和/或共刺激信号传导结构域。在前述方法或用途任一者的另一个实施方式中,初级信号传导结构域包含cd3ζ的功能结构域,和/或共刺激信号传导结构域包含选自41bb、cd27和cd28的蛋白质的功能结构域。在前述方法或用途任一者的另一个实施方式中,细胞因子选自il-15或il-21或两者。在前述方法或用途任一者的另一个实施方式中,包含一个或多个sir分子的免疫效应细胞和增加免疫效应细胞的功效的药剂基本上同时或依次给予。在前述方法或用途任一者的另一个实施方式中,包含sir分子的免疫细胞与靶向gitr和/或调节gitr功能的分子组合给予。在前述方法或用途任一者的另一个实施方式中,靶向gitr和/或调节gitr功能的分子在表达sir的细胞或细胞群体之前或在单采血液成分术之前给予。在前述方法或用途任一者的另一个实施方式中,受试者是人类。本公开还提供一种组合物,该组合物包含至少一种本公开的多核苷酸、本公开的sir多肽分子、本公开的载体或本公开的细胞和药学上可接受的赋形剂。本公开还提供一种试剂盒,该试剂盒包含至少一种本公开的多核苷酸、本公开的sir多肽分子、本公开的载体或本公开的细胞和/或本公开的组合物。本公开还提供一种编码合成免疫受体的多核苷酸,该多核苷酸包含选自由seqidno:900至2264、seqidno:4531至6013、seqidno:7519至8160、seqidno:8803至9230、seqidno:9659至9856、seqidno:10474至12041、seqidno:15786至16011、seqidno:16240至16465、seqidno:16694至16926、seqidno:17162至seqidno:17394、seqidno:17864至17979、seqidno:18321至18322、seqidno:18242至18259、seqidno:18280至18588、seqidno:18899、seqidno:18915至18916组成的组中的序列,或者与seqidno:900至2264、seqidno:4531至6013、seqidno:7519至8160、seqidno:8803至9230、seqidno:9659至9856、seqidno:10474至12041、seqidno:15786至16011、seqidno:16240至16465、seqidno:16694至16926、seqidno:17162至seqidno:17394、seqidno:17864至17979、seqidno:18321至18322、seqidno:18242至18259、seqidno:18280至18588、seqidno:18899和seqidno:18915至18916中任一个所示的编码合成免疫受体的核苷酸序列具有至少75%一致性的序列。本公开还提供一种编码合成免疫受体多肽的氨基酸序列,该氨基酸序列选自由seqidno:3135至4498、seqidno:6044至7518、seqidno:8161至8802、seqidno:9231至9658、seqidno:9873至10070、seqidno:12431至13998、seqidno:16013至16238、seqidno:16467至16692、seqidno:16928至17160、seqidno:17396至17628、seqidno:17981至18096、seqidno:18239至18240、seqidno:18261至18278、seqidno:18590至18898、seqidno:18900和seqidno:18919至18920组成的组,或者与编码seqidno:3135至4498、seqidno:6044至7518、seqidno:8161至8802、seqidno:9231至9658、seqidno:9873至10070、seqidno:12431至13998、seqidno:16013至16238、seqidno:16467至16692、seqidno:16928至17160、seqidno:17396至17628、seqidno:17981至18096、seqidno:18239至18240、seqidno:18261至18278、seqidno:18590至18898、seqidno:18900和seqidno:18919至18920中任一个所示的合成免疫受体多肽的氨基酸序列具有至少75%一致性的序列。本发明的一个或多个实施方式的细节在附图和以下描述中阐明。根据说明书和附图并且根据权利要求书,本发明的其它特征、目标和优点将是显而易见的。附图说明图1示出抗体、tcr、双链嵌合受体和不同代的car的示意性描述。图2示出本公开的示例性载体构建体。图3a--q示出本公开的sir在表达时可以具有的各种格式的描绘。在这些描绘中,tcr对的每个tcr连接至vh或vl结合结构域。在其它实施方式中,每个tcr可以结合至vhh结构域,而非vh或vl。图4a--q示出本公开的sir在表达时可以具有的各种格式的描绘。在这些描绘中,tcr对的一个tcr连接至vh或vl并且vh或vl然后连接至相反vh或vl。尽管在(a)中显示vl连接至tcrb链,但是将认识到的是在全部(a)-(q)描绘中的取向可以交换,以使得在(a)中vl连接至tcra等。图5a--q示出本公开的sir在表达时可以具有的各种格式的描绘。在这些构建体中,sir是基于单结构域抗体(vhh),该抗体结合至两条tcr恒定链中的仅一条恒定链并且另一条链保持为空。示例性此构建体是cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-her3-17b05so-vhh-myc-[htcra-csdvp]-f-f2a-pac[seqidno:1715]。可以制成基于其它非免疫球蛋白结合结构域而非vhh结构域的类似构建体。例如,此非免疫球蛋白结合结构域可以是基于亲合体、darpin、自身抗原(例如dsg3)、配体(例如mpl或trail)或受体(例如cd16)。图6a--q示出本公开的sir在表达时可以具有的各种格式的描绘。在这些构建体中,sir含有两种不同类型的结构不同的抗原结合结构域。在一个实例中,一个抗原结合结构域包含单结构域抗体(vhh),而另一个抗原结合结构域包含vl-接头-vh取向或vh-接头-vl取向的scfv片段。在另一个实例中,一个抗原结合结构域包含单结构域抗体(vhh),而另一个抗原结合结构域包含受体(例如cd16)。在另一个实例中,一个抗原结合结构域包含单结构域抗体(vhh),而另一个抗原结合结构域包含亲合体。使用两种不同类型的抗原结合结构域的优点在于它们不太可能彼此干扰。第一抗原结合结构域(例如vhh)可以针对一种抗原并且第二抗原结合结构域(例如scfv片段)可以靶向另一种抗原。或者,它们可以针对相同抗原,以增加亲合力。图7a--q示出本公开的sir在表达时可以具有的各种格式的描绘。在这些构建体中,sir是基于两个scfv片段。两个scfv片段可以针对两种不同抗原(例如cd8sp-cd19bu12-scfv-v5-[htcrb-kaciah]-f-p2a-sp-cd20-2f2-scfv-myc-[htcra-csdvp]-f-f2a-pac(040716-b04)[seqidno:1028])。或者,它们两者可以针对相同抗原以增加亲合力(例如cd8sp-cd19bu12-scfv-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-scfv-myc-[htcra-csdvp]-f-f2a-pac(020216-b07)[seqidno:1026])。scfv的格式可以是vh-接头-vl或vl-接头-vh。图8a--t示出本公开的sir在表达时可以具有的各种格式的描绘。在一些sir构建体中,由与vl区、gly-ser接头(ggggsx3)和vh区同框融合的信号肽(例如来源于人类cd8信号肽)组成的scfv片段融合至cα、cβ、pre-cα、cδ或cγ链并且没有互补链。图9a--b示出测量car在293ft细胞中的表达的nluc测定。将未转染293ft细胞和用cd19(fmc63-bbz-pac)和161-bbz-paccar转染的那些细胞与cd19-ggsg-nluc-acv5和mpl-ggsg-nluc-acv5上清液一起孵育,随后用pbs洗涤并通过以pbs稀释的coeleoentrazine(ctz;nanolight)来测量nluc活性。使用biotek板读取器定量荧光。数据表示三个重复孔的平均值+/-标准偏差(sd)。图10示出表达161(vl+vh)-myc-bbz-pacr07、175(vl+vh)-myc-28z-pacq04、vb22(vl+vh)-myc-28z-pacb06car的t细胞与mpl-ggsg-nlucacv5上清液的强结合和表达161(vl+vh)-myc-28z-pacz07、ab317(vl+vh)-myc-28z-pact04和12e10(vl+vh)-myc-28z-pacb06的t细胞与该上清液的适度结合,而在未感染t细胞或表达4c3(vl+vh)-myc-28z-pac对照car的那些细胞上未观察到显著结合。类似地,在任何mplcar-t细胞上未观察到与cd19-ggsg-nluc-acv5上清液的特异性结合,从而证明该测定的特异性。图11示出证明由野生型核苷酸序列编码的含有sir的tcrα和tcrβ恒定区未能在人类原代t细胞中有效表达的图。相反,有效表达了携带促进链间二硫键的额外半胱氨酸残基的含有密码子优化的人类tcra/b链的sir。如(081415-d06)[seqidno:992]sir中可见的人类tcrα/β恒定链的鼠类化导致sir表达进一步增加。另外,如(082815-g07)[seqidno:1620]和(082815-e05)[seqidno:1622]构建体中可见,scfv片段在与tcrb恒定链共同表达时可以被表达为融合至tcra恒定区,即使tcrb不携带任何抗原结合部分也如此。图12示出一种生成具有希望的或不同的结合亲和力的sir集合的方法。图13a--b示出代表性facs分析。(a)将对照jurkat-nfat-gfp细胞或表达靶向cd19(克隆id051716-i08)、mpl(克隆id:040716-a07)和bcma(克隆id:011116-a07)的sir的那些细胞分别与raji(顶部)、hel(中间)或u266(底部)细胞一起孵育。在将表达sir的jurkat-nfat-gfp细胞与其对应靶细胞共同培养时,gfp表达的诱导是明显的。(b)将表达靶向cdh6(克隆id:051716-j05)、cd276(克隆id:050516-q06)和her2/neu(克隆id:050516-i03)的sir的jurkat细胞分别与skov3(顶部)和mc7(中间和底部)细胞一起孵育。在将表达sir的jurkat-nfat-gfp细胞与其对应靶细胞共同培养时,gfp表达的诱导是明显的。图14示出用于表达本公开的sir的逆转录病毒载体的示例性结果。图15示出使用睡美人转座子载体的结果,用构建体psbbi-puro-fmc63vl-v5-[tcrb-kaciah]-f-p2a-fmc63vh-myc-[tcra-csdvp]-f-f2a[010616-b01](seqidno:875)转染的jurkat-nfat-gfp细胞在与对应raji靶细胞系共同培养时显示gfp诱导。图16示出在t细胞上与bu12carcd8sp-cd19bu12-(vl-vh)-myc-bbz-t2a-pac(082815-p08)[seqidno:4503]相比bu12sir(cd8sp-cd19bu12-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-cd19bu12-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(070215-m03)[seqidno:1021]的更低细胞表面表达,如通过分别使用apc-myc-apc和生物素-蛋白质-l加上apc-链霉亲和素进行染色来确定的。图17示出与针对mpl的对照sircd8sp-mpl-161-vl-v5-[htcrb-s57c-opt1]-f-p2a-mpl-161-vh-myc-[htcra-t48c-opt1]-f-f2a-pac(040315-u02)[seqid1112]的t细胞相比用raji细胞注射并接受表达针对cd19的sircd8sp-fmc63-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-fmc63-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(050515-l05)[seqidno:900]和cd8sp-cd19bu12-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-cd19bu12-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(070215-m03)[seqidno:1021]的t细胞的小鼠的存活。图18示出通过将表达本公开的构建体的jurkat-nfat-gfp细胞与蛋白l珠粒共同培养所进行的gfp诱导。图19a--d示出与293-蛋白l-ii细胞共同培养导致在表达以下各项的jurkat-nfat-gfp细胞中gfp表达的强诱导:(b)cd8sp-fmc63(vl-vh)-myc-bbz-t2a-pac(112014-a13)[seqidno:4501]car、(c)cd8sp-huluc64-vl-v5-[htcrb-kaciah]-f-p2a-sp-huluc64-vh-myc-[htcra-csdvp]-f-f2a-pac(092916-e07)[seqidno:1253]和(d)cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-cd19bu12-vl-gly-ser-接头-cd19bu12-vh-myc-[htcra-csdvp]-f-f2a-pac(082815-e05)[seqidno:1622]sir构建体。具体实施方式除非上下文明确地指示其它的情况,否则如本文和附属权利要求中使用的,单数形式“一个/种(a/an)”和“该(the)”包括复数指代物。因此,例如提及“细胞”包括多种这类细胞,并且提及“多核苷酸”包括提及一种或多种多核苷酸等。而且,除非另外指示,否则使用“或”意指“和/或”。类似地,“包含(comprise/comprises/comprising)”和“包括(include/includes/including)”是可互换的并且不旨在为限制性的。还应理解,在使用术语“包含”描述不同实施方式时,本领域技术人员将理解,在一些特定情况下,一个实施方式可以可替代地使用语言“基本上由...组成”或“由...组成”来描述。除非另外定义,否则本文所用的所有技术和科学术语均具有与本发明所属领域的普通技术人员通常所理解的相同的含义。allen等人,remington:thescienceandpracticeofpharmacy[雷明顿:药物科学与实践]第22版,pharmaceuticalpress[药物出版社](2012年9月15日);hornyak等人,introductiontonanoscienceandnanotechnology[纳米科学和纳米技术介绍],crcpress[crc出版社](2008);singleton和sainsbury,dictionaryofmicrobiologyandmolecularbiology[微生物学和分子生物学字典]第3版,修订版,j.wiley&sons[j.威利父子公司](纽约,ny2006);smith,march’sadvancedorganicchemistryreactions,mechanismsandstructure[马奇高级有机化学反应、机制和结构]第7版,j.wiley&sons[j.威利父子公司](纽约,ny2013);singleton,dictionaryofdnaandgenometechnology[dna和基因组技术字典]第3版.,wiley-blackwell[布莱克威尔公司](2012年11月28日);以及green和sambrook,molecularcloning:alaboratorymanual[分子克隆:实验室手册]第4版,coldspringharborlaboratorypress[冷泉港实验室出版社](冷泉港,ny2012),为本领域技术人员提供了本申请中所用的许多术语的一般指导。关于如何制备抗体的参考文献,参见greenfield,antibodiesalaboratorymanual[抗体实验室手册]第2版,coldspringharborpress[冷泉港出版社](冷泉港ny,2013);和milstein,derivationofspecificantibody-producingtissuecultureandtumorlinesbycellfusion[通过细胞融合获得特异性抗体产生组织培养物和肿瘤系],eur.j.immunol.[欧洲免疫学杂志]1976年7月,6(7):511-9;queen和selick,humanizedimmunoglobulins[人源化免疫球蛋白],美国专利号5,585,089(1996年12月);以及riechmann等人,reshapinghumanantibodiesfortherapy[再成形用于治疗的人类抗体],nature[自然]1988年3月24,332(6162):323-7all,本文提供的标题和子标题仅为了便于阅读并且不应被理解为限制本发明。虽然与本文所述的那些方法和材料类似或等效的方法和材料可以用于本发明的实践或测试,但是以下描述了合适的方法和材料。本文提及的所有公布、专利申请、专利以及其它参考文献均以引用的方式整体并入。在有矛盾的情况下,将以本说明书(包括定义)为准。此外,材料、方法和具体实例仅是说明性的而不旨在为限制性。本公开的合成免疫受体(sir)包含可以例如以mhc依赖性或mhc独立性方式结合至抗原的抗原结合结构域(例如,抗体或抗体片段)。.通常,源自内源性蛋白质的肽填充主要组织相容性复合物(mhc)i类分子的口袋,并且被cd8+t淋巴细胞上的t细胞受体(tcr)识别。mhci类复合物由所有有核细胞组成型表达。在癌症中,病毒特异性和/或肿瘤特异性肽/mhc复合物代表用于免疫疗法的独特类别的细胞表面靶标。已描述了在人类白血病抗原(hla)-a1或hla-a2背景下源自病毒或肿瘤抗原的靶向tcr样抗体的肽(参见例如sastry等人,jviral.[病毒杂志]201185(5):1935-1942;sergeeva等人,blood[血液],2011,117(16):4262-4272;verma等人,jlmmunol[免疫学杂志],2010,184(4):2156-2165;willemsen等人,genether.[基因治疗],2001,8(21):1601-1608;dao等人,scitranslmed.[科学转译医学],2013,5(176):176ra33;tassev等人,cancergenether.[癌症基因疗法],2012,19(2):84-100)。例如,可以从筛选文库(如人scfv噬菌体展示文库)鉴定tcr样抗体。本公开大体上提供合成免疫受体(sir),该合成免疫受体包含可操作连接至t细胞受体结构域的抗原结合位点。本公开进一步提供一种或多种包含编码sir的序列的重组核酸构建体,其中该sir包含结合至抗原或靶分子(如本文下文进一步描述)并连接至一个或多个t细胞受体恒定链(包括其突变体或变体)的一个或多个抗原结合结构域(例如,抗体、抗体片段、非免疫球蛋白抗原结合结构域、自身抗原、配体或受体)。sir的一个或多个抗原结合结构域特异性结合至一个或多个本文所述的疾病相关抗原或同源物,其中每个抗原结合结构域的编码序列与编码它所连接的每条t细胞受体恒定链可操作地连接,使得该抗原结合结构域与t细胞恒定链可操作地表达。在一些实施方式中,sir可包含连接至单一t细胞受体恒定链的单一抗原结合结构域。在一些实施方式中,sir包含各自连接至单独t细胞受体恒定链的两个抗原结合结构域。例如,抗原结合结构域1连接至t细胞受体α(tcrα)的恒定链以构造“功能单元1”并且抗原结合结构域2连接至t细胞受体β(tcrβ)的恒定链以构造“功能单元2”。此类sir的两个功能单元在同一细胞中共同表达以变成具有功能活性(例如,异二聚合)。在一些实施方式中,sir包含与一个t细胞受体恒定链同框连接(功能单位1)但与第二t细胞受体恒定链共同表达的抗原结合结构域。在此类sir中第二t细胞受体恒定链的目的在于促进功能单元1(例如连接至t细胞受体恒定链的抗原结合结构域1)的细胞表面表达。这样,第二t细胞受体恒定链可以自身表达或表达为携带表位标签(例如myc、v5、acv5、g4sx2、streptagii等)的融合蛋白或表达为携带不干扰功能单元1的组装和功能的任何不相关蛋白片段(例如vl或vh片段)的融合蛋白。作为一个实例,sir可以包含与t细胞受体α(tcrα)的恒定链和t细胞受体β(tcrβ)的空白(即缺乏抗原结合结构域)恒定链可操作地同框连接的抗原结合结构域1。此类sir的两个功能单元在同一细胞中共同表达以变成具有功能活性。在一些实施方式中,sir的两个功能单元通过转染编码两个功能单元的单个多核苷酸来共同表达,而在其它实施方式中,两个功能单元通过转染各自编码一个功能单元的两个不同多核苷酸来共同表达。在一些实施方式中,sir的两个功能单元被插入在单一基因组基因座处,而在其它实施方式中,两个功能单元被插入在两个基因组基因座处。例如,在一些实施方式中,两个功能单元可以插入在tcrα恒定链(trac)基因座处并且表达为单一多核苷酸。在其它实施方式中,功能单元1可以被插入在tcrα恒定链(trac)基因座处,而功能单元2可以被插入在tcr恒定链β1(trbc1)基因座处。在一些实施方式中,sir的两个功能单元通过转染编码两个功能单元的单个多核苷酸来共同表达,而在其它实施方式中,两个功能单元通过转染各自编码一个功能单元的两个不同多核苷酸来共同表达。在一些实施方式中,sir包含与一个t细胞受体恒定链同框连接(功能单位1)但与第二t细胞受体恒定链共同表达的抗原结合结构域。在此类sir中第二t细胞受体恒定链的目的在于促进功能单元1(例如连接至t细胞受体恒定链的抗原结合结构域1)的细胞表面表达。这样,第二t细胞受体恒定链可以自身表达或表达为携带表位标签(例如myc、v5、acv5、g4sx2、streptagii等)的融合蛋白或表达为携带不干扰功能单元1的组装和功能的任何不相关蛋白片段(例如vl或vh片段)的融合蛋白。作为一个实例,sir可以包含与t细胞受体α(tcrα)的恒定链和t细胞受体β(tcrβ)的空白(即缺乏抗原结合结构域)恒定链可操作地同框连接的抗原结合结构域1。此类sir的两个功能单元在同一细胞中共同表达以变成具有功能活性。在一些实施方式中,sir的两个功能单元使用单一载体共同表达,而在其它实施方式中,两个功能单元在同一细胞中使用不同载体共同表达。在一些实施方式中,sir的两个功能单元通过转染编码两个功能单元的单个多核苷酸来共同表达,而在其它实施方式中,两个功能单元通过转染各自编码一个功能单元的两个不同多核苷酸来共同表达。本公开的sir的不同构型提供于图3--8中。本公开提供一类嵌合t细胞受体(合成免疫受体(sir)),其可以用于过继细胞治疗以治疗癌症、感染、自身免疫和退行性疾病。与嵌合抗原受体(car)相反,本公开的sir接合生理学t细胞受体信号传导途径的全力并且因此不太可能导致与car相关的并发症,诸如细胞因子释放综合征、神经毒性和体内持久性缺乏。与car相反,本公开的sir具有较小抗原结合结构域自身聚集趋势、较小补剂信号传导机会和较小早期t细胞耗尽机会。本公开的sir含有融合至tcrα(cα)、tcrβ(cβ)、tcrδ(cδ)、tcrγ(cγ)或pretcrα(cα)(包括前述任一者的变体和突变体)的恒定链的一个或多个抗原结合结构域。抗原结合结构域可以包含抗体或抗体片段、抗体的vl或/和vh片段、衍生自抗体的scfv片段、单结构域抗体、亲合体、darpin、任何抗原结合配体或受体、自身抗原、或任何其它非免疫球蛋白抗原结合片段。抗原结合结构域可以靶向单一抗原或多个抗原(双特异性或多特异性sir)。sir的tcr恒定结构域可以单独表达,但是通常成对表达(例如,cα与cβ、或precα与cβ、或cδ与cγ等),以便于最佳细胞表面表达。tcr恒定链片段通常被密码子优化以允许最佳细胞表面表达。tcr恒定片段可以携带额外突变或取代,以便于其最佳表达并与互补链配对和/或减少与内源性tcr链的配对和/或稳定抗原结合结构域之间的相互作用。sir也可以将一个或多个额外结构域(例如myc、streptag、v5、flag、ritx标签等)表达为融合蛋白。本公开的sir可以使用许多技术引入到细胞中,这些技术包括但不限于使用慢病毒载体、逆转录病毒载体、腺相关病毒载体、杆状病毒载体、睡美人转座子、piggybac转座子或通过mrna转染、或使用以上方法的组合。还公开了用于递送sir的优化载体。可以表达本公开的sir,以使得它们处于内源性启动子(例如tcrα或tcrβ启动子)的控制下。在一些实施方式中,本公开的sir使用外来启动子(例如cmv启动子)表达。本公开的sir可以共同表达额外模块,诸如促进sir表达或功能(例如cd3z、cd3ε、cd3δ、cd3z-41bb融合蛋白等)、促进t细胞增殖、持久、扩增和活化(例如41bbl、cd40l、il12f、k13、tax、tax2、mc159l、cflip、scfv靶向pd1、shrna靶向brd4等)、减小毒性(例如vhh或scfv靶向il6r、il6、tnfα等)的cdna编码分子、选择标记(例如tegfr、tegfrviii、tbcma、tcd19等)和/或自杀基因(例如icaspase9、hsv-胸苷激酶)。本公开的sir可以在免疫效应细胞(例如t细胞)中或在可产生免疫效应细胞的干细胞(包括诱导型多能干细胞)中表达。本公开还提供用于表达sir的免疫效应细胞子集和用于活化并扩增表达sir的免疫效应细胞的方法。本公开还描述可以用于增强表达sir的免疫效应细胞的活性和持久性或减小其毒性的药剂。本公开描述可以用于鉴别适用于不同应用的sir的体外和体内测定。当指可测量的值诸如量、时距等时,术语“约”意在涵盖与指定值±20%、或在一些情况下±10%、或在一些情况下±5%、或在一些情况下±1%、或在一些情况下±0.1%的变化,因为此类变化适于执行所公开的方法或描述本文的组合物。此外,任何值或范围(例如,小于20或类似术语明确包括此类值之间的任何整数或至多该值。因此,例如,“一个至五个突变”明确包括1、2、3、4和/或5个突变。术语“辅助模块”是指可以与sir共同表达的以下任一个或多个:41bbl、cd40l、k13、mc159、cflip-l/mritα、cflip-p22、htlv1tax、htlv2tax、htlv2tax-rs突变体、fkbpx2-k13、fkbpx2-htlv2-tax、fkbpx2-htlv2-tax-rs、il6r-304-vhh-alb8-vhh、il12f、pd1-4h1scfv、pd1-5c4scfv、pd1-4h1-alb8-vhh、pd1-5c4-alb8-vhh、ctla4-伊匹单抗-scfv、ctla4-伊匹单抗-alb8-vhh、il6-19a-scfv、il6-19a-scfv-alb8-vhh、shvem、shvem-alb8-vhh、htert、fx06、cd3z、cd3z-gggs-41bb、cd3-bbz、cd3-cd28z、cd3-cd28-lck融合蛋白、shrna靶向brd4及其组合。辅助模块可以使用单个载体或使用两个或更多个不同载体与sir共同表达。在一个实施方式中,辅助模块包含seqidno:3087至3117(dna编码序列seqidno:812-842)的氨基酸序列或与其具有80-99%一致性的序列。在其它实施方式中,编码辅助模块的核酸序列包括seqidno:800或seqidno:842的序列或与其具有80-99%一致性的序列。“自身抗体”是指通过对自身抗原特异性的b细胞产生的抗体。如本文所用,术语“抗体”是指源自与抗原特异性地结合的免疫球蛋白分子的蛋白质或多肽序列。抗体可以是单克隆或多克隆、多链或单链、或完整免疫球蛋白,并且可以源自天然来源或源自重组来源。抗体可以是免疫球蛋白分子的四聚体。抗体可以是‘人源化’、‘嵌合的’或非人类的。术语“抗体片段”是指抗体的至少一部分,该至少一部分保留与抗原表位特异性地相互作用(例如,通过结合、空间位阻、稳定/去稳定、空间分布)的能力。抗体片段的实例包括但不限于fab、fab'、f(ab')2、fv片段、scfv抗体片段、二硫键连接的fv(sdfv)、由vh和ch1结构域组成的fd片段、线性抗体、单结构域抗体诸如sdab(vl或vh)、骆驼科vhh结构域、由抗体片段(诸如包含在铰链区通过二硫桥连接的两个fab片段的二价片段)形成的多特异性抗体、以及分离的cdr、或抗体的其它表位结合片段。抗原结合片段还可以掺入到单结构域抗体、大型抗体(maxibodies)、微型抗体(minibodies)、纳米抗体、胞内抗体、双体抗体、三体抗体、四体抗体、v-nar和bis-scfv中(参见例如,hollinger和hudson,naturebiotechnology[自然生物技术]23:1126-1136,2005)。还可以将抗原结合片段接枝到基于多肽的支架,例如纤连蛋白iii型(fn3)中(参见美国专利号6,703,199,其描述了纤连蛋白多肽微型抗体)。术语“抗体重链”是指抗体分子中以其天然存在的构象存在的两种类型多肽链中较大的一种,并且通常决定抗体所属的类别。术语“抗体轻链”是指抗体分子中以其天然存在的构象存在的两种类型多肽链中较小的一种。kappa(κ)和lambda(λ)轻链是指两种主要的抗体轻链同种型。术语“抗癌作用”是指可以通过各种方式表现的生物学效应,包括但不限于肿瘤体积减少、癌细胞数目减少、转移瘤数目减少、预期寿命增加、癌细胞增殖减少、癌细胞存活降低、或与癌性病症相关的各种生理症状改善。“抗癌作用”还可以通过sir首先预防癌症发生的能力来表现。“抗癌剂”是指抑制异常细胞分裂和生长、抑制肿瘤细胞迁移、抑制侵入或预防癌症生长和转移的药剂。该术语包括化学治疗剂、生物剂(例如sirna、病毒载体诸如工程化mlv、腺病毒、递送细胞毒性基因的疱疹病毒)、抗体等。术语“抗原”或“ag”是指引起免疫反应的分子。此免疫反应可以涉及抗体产生或特定免疫活性细胞的活化或两者。本领域技术人员将理解实际上包括所有蛋白质或肽的任何大分子都可以充当抗原。此外,抗原可以衍生自重组或基因组dna。技术人员将理解包含编码引发免疫反应的蛋白质的核苷酸序列或部分核苷酸序列的任何dna都因此编码“抗原”(当该术语在本文中使用时)。此外,本领域技术人员将理解抗原不需要仅由基因的全长核苷酸序列编码。显而易见的是,本公开包括但不限于使用多于一种基因的部分核苷酸序列,并且这些核苷酸序列以各种组合排列以编码引发所希望的免疫反应的多肽。此外,本领域技术人员将理解抗原根本不需要由“基因”编码。显而易见的是,抗原可以合成产生或可以源自生物样品,或者可以是除多肽外的大分子。这种生物样品可以包括但不限于组织样品、肿瘤样品、细胞或具有其它生物组分的流体。靶抗原的非限制性实例包括:cd5、cd19;cd123;cd22;cd30;cd171;cs-1(也称为cd2亚单位1、cracc、slamf7、cd319和19a24);c型凝集素样分子-1(cll-1或clecl1);cd33;表皮生长因子受体变体iii(egfrviii);神经节苷脂g2(gd2);神经节苷脂gd3(aneu5ac(2-8)aneu5ac(2-3)bdgalp(l-4)bdglcp(l-l)cer);tnf受体家族成员b细胞成熟(bcma);tn抗原((tnag)或(galnacα-ser/thr));前列腺特异性膜抗原(psma);受体酪氨酸激酶样孤儿受体1(ror1);fms样酪氨酸激酶3(flt3);肿瘤相关性糖蛋白72(tag72);cd38;cd44v6;在急性白血病或淋巴瘤上而不在造血祖细胞上表达的糖基化cd43表位、在非造血癌症上表达的糖基化cd43表位、癌胚抗原(cea);上皮细胞粘附分子(epcam);b7h3(cd276);kit(cd117);白介素-13受体亚基α-2(il-13ra2或cd213a2);间皮素;白介素11受体α(il-llra);前列腺干细胞抗原(psca);蛋白酶丝氨酸21(睾蛋白或prss21);血管内皮生长因子受体2(vegfr2);路易斯(lewis)(y)抗原;cd24;血小板源性生长因子受体β(pdgfr-β);阶段特异性胚胎抗原-4(ssea-4);cd20;叶酸受体α(fra或fr1);叶酸受体β(frb);受体酪氨酸蛋白激酶erbb2(her2/neu);细胞表面相关粘蛋白1(muc1);表皮生长因子受体(egfr);神经细胞粘附分子(ncam);前列腺酶;前列腺酸性磷酸酶(pap);突变的延长因子2(elf2m);肝配蛋白b2;成纤维细胞活化蛋白α(fap);胰岛素样生长因子1受体(igf-i受体),碳酸酐酶ix(calx);蛋白酶体(前体,巨蛋白因子)亚基,β型,9(lmp2);糖蛋白100(gpl00);由裂点簇区(bcr)和阿贝尔森(abelson)鼠类白血病病毒癌基因同源物1(abl)组成的癌基因融合蛋白(bcr-abl);酪氨酸酶;肝配蛋白a型受体2(epha2);唾液酸化路易斯粘附分子(sle);神经节苷脂gm3(aneu5ac(2-3)bdclalp(l-4)bdglcp(l-1)cer);转谷氨酰胺酶5(tgs5);高分子量-黑素瘤相关抗原(hmwmaa);o-乙酰基-gd2神经节苷脂(oacgd2);肿瘤内皮标记1(tem1/cd248);肿瘤内皮标记7相关(tem7r);密封蛋白(claudin)6(cldn6);甲状腺刺激性激素受体(tshr);g蛋白偶联受体c类5组成员d(gprc5d);x染色体开放阅读框61(cxorf61);cd97;cd179a;间变性淋巴瘤激酶(alk);聚唾液酸;胎盘特异性1(plac1);globoh糖神经酰胺六糖部分(globoh);乳腺分化抗原(ny-br-1);尿空斑蛋白(uroplakin)2(upk2);甲型肝炎病毒细胞受体1(havcr1);肾上腺受体β3(adrb3);泛连接蛋白3(panx3);g蛋白偶联受体20(gpr20);淋巴细胞抗原6复合物,基因座k9(ly6k);嗅觉受体51e2(or51e2);tcrγ替代性阅读框蛋白质(tarp);威尔姆氏瘤蛋白质(wt1);癌症/睾丸抗原1(ny-es0-1);癌症/睾丸抗原2(lage-1a);黑素瘤相关抗原1(mage-a1);位于染色体12p上的ets易位变体基因6(etv6-aml);精子蛋白17(spa17);x抗原家族成员la(xagel);血管生成素结合细胞表面受体2(tie2);黑素瘤癌症睾丸抗原-1(mad-ct-1);黑素瘤癌症睾丸抗原-2(mad-ct-2);fos相关抗原1;肿瘤蛋白p53(p53);p53突变体;前列腺特异性蛋白;存活素(surviving);端粒酶;前列腺癌肿瘤抗原-1(pcta-1或半乳凝素8)、由t细胞识别的黑素瘤抗原1(melana或marti);大鼠肉瘤(ratsarcoma,ras)突变体;人类端粒酶逆转录酶(htert);肉瘤易位裂点;黑素瘤细胞凋亡抑制剂(ml-iap);erg(跨膜蛋白酶,丝氨酸2(tmprss2)ets融合基因);n-乙酰基葡萄糖胺基-转移酶v(na17);配对框蛋白质pax-3(pax3);雄激素受体;周期素bl;v-myc禽类骨髓细胞瘤病病毒癌基因神经母细胞瘤衍生性同源物(mycn);ras同源家族成员c(rhoc);酪氨酸酶相关蛋白2(trp-2);细胞色素p450lb1(cyplb1);ccctc-结合因子(锌指蛋白)样(boris或印迹位点调节剂同级物)、由t细胞识别的鳞状细胞癌抗原3(sart3);配对框蛋白质pax-5(pax5);前顶体蛋白结合蛋白sp32(oy-tesl);淋巴细胞特异性蛋白质酪氨酸激酶(lck);a激酶锚定蛋白4(akap-4);滑膜肉瘤,x裂点2(ssx2);晚期糖基化终产物受体(rage-1);肾泛素1(rul);肾泛素2(ru2);豆荚蛋白;人乳头状瘤病毒e6(hpve6);人乳头状瘤病毒e7(hpve7);肠道羧基酯酶;突变的热休克蛋白70-2(muthsp70-2);cd79a;cd79b;cd72;白细胞相关免疫球蛋白样受体1(lairl);iga受体的fc片段(fcar或cd89);白细胞免疫球蛋白样受体子家族a成员2(lilra2);cd300分子样家族成员f(cd300lf);c型凝集素结构域家族12成员a(clec12a);骨髓基质细胞抗原2(bst2);egf样含模块粘蛋白样激素受体样2(emr2);淋巴细胞抗原75(ly75);磷脂酰肌醇聚糖-3(gpc3);fc受体样5(fcrl5);以及免疫球蛋白λ样多肽1(iglll)、mpl、、生物素、c-myc表位标签、cd34、lamp1trop2、gfrα54、cdh17、cdh6、nybr1、cdh19、cd200r、slea(ca19.9;唾液酸化路易斯抗原)岩藻糖基-gm1、ptk7、gpnmb、cdh1-cd324、dll3、cd276/b7h3、il11ra、il13ra2、cd179b-igll1、tcrγ-δ、nkg2d、cd32(fcgr2a)、tnag、tim1-/hvcr1、csf2ra(gm-csfr-α)、tgfβr2、lewsag、tcr-β1链、tcr-β2链、tcr-γ链、tcr-δ链、fitc、促黄体生成激素受体(lhr)、卵泡刺激素受体(fshr)、促性腺激素激素受体(cghr或gr)、ccr4、gd3、slamf6、slamf4、hiv1包膜糖蛋白、htlv1-tax、cmvpp65、ebv-ebna3c、kshvk8.1、kshv-gh、甲型流感凝集素(ha)、gad、pdl1、鸟苷酸环化酶c(gcc)、桥粒芯蛋白自身抗体3(dsg3)、桥粒芯蛋白自身抗体1(dsg1)、hla、hla-a、hla-a2、hla-b、hla-c、hla-dp、hla-dm、hla-doa、hla-dob、hla-dq、hla-dr、hla-g、ige、cd99、rasg12v、组织因子1(tf1)、afp、gprc5d、密封蛋白18.2(cld18a2或cldn18a.2))、p-糖蛋白、steap1、liv1、粘连蛋白-4、cripto、gpa33、bst1/cd157、低电导率氯离子通道和由tnt抗体识别的抗原。术语“抗原呈递细胞”或“apc”是指免疫系统细胞,诸如辅助细胞(例如,b细胞、树突细胞等),该免疫系统细胞在其表面上展示与主要组织相容性复合物(mhc)复合的外来抗原。t细胞可以使用它们的t细胞受体(tcr)识别这些复合物。apc加工抗原并将抗原呈递给t细胞。术语“抗感染效应”是指可以通过各种方式表现的生物效应,这些方式包括但不限于例如感染剂效价减小、感染剂菌落计数减少、与感染病状相关的各种生理症状改善。“抗感染效应”还可以通过肽、多核苷酸、细胞和抗体首先预防癌症发生的能力来表现。术语“抗肿瘤效应”或“抗癌效应”是指可以通过各种方式表现的生物学效应,包括但不限于例如肿瘤体积减少、肿瘤细胞数目减少、肿瘤细胞增殖减少、或肿瘤细胞存活降低。如本文所用,“亲和力”意指描述结合强度的量度。在一些情况下,亲和力取决于结合剂与其靶标之间(例如抗体与抗原(包括对结合结构域特异性的表位)之间)的立体化学配合接近度、它们之间的接触面积大小以及带电荷和疏水性基团的分布。亲和力通常是指结合剂结合其靶标的“能力”。本领域存在用于测量“亲和力”的许多方式。例如,用于计算抗体对抗原的亲和力的方法为本领域中已知的,包括使用计算亲和力的结合实验。结合亲和力可以使用本领域已知的各种技术来测定,这些技术例如表面等离子体共振、生物层干涉、双重极性干涉、静态光散射、动态光散射、等温滴定量热法、elisa、分析超速离心以及流式细胞术。用于确定结合亲和力的示例性方法采用表面等离子体共振。表面等离子体共振是一种光学现象,该光学现象允许例如使用biacore系统通过检测生物传感器基质内蛋白质浓度的变化来分析实时生物特异性相互作用(瑞典乌普萨拉和新泽西州皮斯卡塔韦的玛西亚生物传感器公司(pharmaciabiosensorab,uppsala,swedenandpiscataway,n.j.))。“抗原结合结构域”或“抗原结合模块”或“抗原结合区段”是指由于其初级、二级或三级序列和或翻译后修饰和/或电荷而以高特异性程度结合至抗原的多肽或肽。抗原结合结构域可以源自不同来源,例如抗体、非免疫球蛋白结合蛋白、配体或受体。本公开提供包含结合至一个或多个靶抗原的抗原结合结构域的sir。本公开还提供包含不源自抗体的抗原结合结构域的sir。“亲合力”是指结合剂与靶标之间的相互作用强度(例如,抗体与其抗原靶标、受体及其同源物等之间的相互作用强度)。亲合力可以是弱的或强的。用于计算抗体对抗原的亲和力的方法为本领域中已知的,包括使用计算亲和力的结合实验。在功能测定(例如流式细胞术测定)中的抗体活性也反映了抗体亲和力。抗体和亲和力可以在表型方面表征并且使用功能测定(例如流式细胞术测定)比较。术语“缔合常数(ka)”被定义为受体与配体缔合的平衡常数。术语“自身抗原”是指刺激自身免疫反应产生,诸如自身抗体产生的内源性抗原。自身抗原也包括自体抗原或来自正常组织的抗原,其为可导致自身免疫疾病发展的细胞介导的或抗体介导的免疫反应的靶标。自身抗原的实例包括但不限于桥粒芯蛋白1、桥粒芯蛋白3及其片段。如本文所用,“有利结果”可以包括但决不限于减轻或缓和疾病病状的严重性、预防疾病病状恶化、治愈疾病病状、预防疾病病状发展、降低患者发展疾病病状的机会以及延长患者的生命或预期寿命。如本文所用,术语“结合结构域”或“抗体分子”是指包含至少一个结构域,例如可以高于非特异性结构域的亲和力结合至靶标的免疫球蛋白可变结构域序列的蛋白质,例如免疫球蛋白链或其片段。该术语涵盖抗体和抗体片段。在另一个实施方式中,抗体分子是多特异性抗体分子,例如它包含多个免疫球蛋白可变结构域序列,其中该多个中的第一免疫球蛋白可变结构域序列对第一表位具有结合特异性并且该多个中的第二免疫球蛋白可变结构域序列对第二表位具有结合特异性。在另一个实施方式中,多特异性抗体分子是双特异性抗体分子。双特异性抗体对两种抗原具有特异性。双特异性抗体分子的特征在于对第一表位具有结合特异性的第一免疫球蛋白可变结构域序列和对第二表位具有结合特异性的第二免疫球蛋白可变结构域序列。“结合与...相同的表位”意指抗体、scfv或其它抗原结合结构域结合至靶抗原的能力并且其具有与所列举抗体、scfv或其它抗原结合结构域相同的表位。作为实例,所列举抗体、scfv或其它结合剂和其它抗体的表位可以使用标准表位作图技术来确定。本领域熟知的表位作图技术包括epitopemappingprotocolsinmethodsinmolecularbiology[分子生物学方法的表位作图方案],第66卷(glenne.morris编辑,1996)humanapress,totowa,newjersey[新泽西州托托瓦的胡马纳出版社]。例如,线性表位可以通过例如同时合成固体载体上的大量肽、对应于蛋白质分子部分的肽并且在这些肽仍连接至载体时使这些肽与抗体反应来确定。此类技术是本领域已知的并且描述于例如美国专利号4,708,871;geysen等人,(1984)proc.natl.acad.sci.usa[美国国家科学院院刊]8:3998-4002;geysen等人,(1985)proc.natl.acad.sci.usa[美国国家科学院院刊]82:78-182;geysen等人,(1986)mol.lmmunol.[分子免疫学]23:709-715。由sir的抗原结合结构域结合的表位也可以通过表位分箱测定来确定。表位分箱是用于表征并且然后分选针对靶蛋白的单克隆抗体文库的竞争性免疫测定。以成对方式针对文库中的所有其它抗体测试针对类似靶标的抗体,以查看抗体是否阻断另一种抗体与抗原表位的结合。在每种抗体具有针对文库中的所有其它抗体创建的特征曲线之后,对文库中的每种抗体相对于其它抗体创建竞争性阻断特征曲线。紧密相关分箱特征曲线指示抗体具有相同或紧密相关的表位,并且一起“分箱”。类似地,构象表位容易通过确定氨基酸的空间构象来鉴定,诸如通过例如氢/氘交换、x射线结晶学和二维核磁共振。参见例如epitopemappingprotocols[表位作图方案],同上。蛋白质的抗原区域也可以使用标准抗原性和亲水性图鉴别,诸如使用例如可得自牛津大学分子组(oxfordmoleculargroup)的omiga1.0版软件程序计算的那些图。此计算机程序采用hopp/woods方法,hopp等人,(1981)proc.natl.acad.sciusa[美国国家科学院院刊]78:3824-3828;用于确定抗原性图,并且采用kyte-doolittle技术,kyte等人,(1982)j.mol.bioi.[分子生物学杂志]157:105-132;用于亲水性图。为了确定针对靶标(例如cd19)的所选单克隆抗体是否结合至独特表位,每种抗体可以使用可商购获得的试剂(pierce,rockford,ill.)进行生物素化。可以使用cd19细胞外结构域包被的elisa平板来进行使用未标记单克隆抗体和生物素化单克隆抗体的竞争研究。可以用链霉亲和素-碱性磷酸酯酶探针检测生物素化mab结合。如本文所用,术语“cdr”或“互补决定区”旨在意指重链和轻链多肽两者的可变区内可见的不连续抗原组合位点。这些特定区域已由kabat等人,j.bioi.chern.[生物化学杂志]252:6609-6616(1977);kabat等人,u.s.dept.ofhealthandhumanservices[美国卫生与公众服务部],"sequencesofproteinsofimmunologicalinterest[具有免疫学意义的蛋白质序列]"(1991);chothia等人,j.mol.bioi.[分子生物学杂志]196:901-917(1987);以及maccallum等人,j.mol.bioi.[分子生物学杂志]25262:732-745(1996)进行描述,其中定义当彼此相比较时包括氨基酸残基的重复部分或子集合。然而,任一定义用于是指抗体或接枝抗体或其变体的cdr,旨在处于如本文所定义和使用的术语的范围内。如本文所用,抗体的不同cdr也可以通过不同定义的组合来定义。例如,vhcdr1可以基于kabat来定义并且vhcdr2可以基于chothia来定义。涵盖如每一份上文引用的参考文献所定义的cdr的氨基酸残基如下:cdr定义(残基编号对应于鉴别的参考)。表5中提供了构成本公开的sir的靶向不同抗原的抗原结合结构域的不同vl和vh区段的cdr的seqid。在一些实施方式中,对特异性结合至靶抗原的抗原结合模块(诸如fab样或fv样抗原结合模块)的提及意指抗原结合模块结合至靶抗原的(a)亲和力是其与其它分子的结合亲和力的至少10(例如,约10、20、30、40、50、75、100、200、300、400、500、750、1000或更多)倍;或者(b)kd是其与其它分子结合的kd的不多于1/10(例如,1/10、1/20、1/30、1/40、1/50、1175、1/100、1/200、1/300、1/400、1/500、1/750、1/1000或更少)倍。结合亲和力可以通过本领域已知的方法确定,诸如elisa、荧光活化细胞分选(facs)分选或放射性免疫沉淀测定(ria)。kd可以通过本领域已知的方法确定,诸如利用例如biacore仪器的表面等离子体共振(spr)或利用例如sapidyne仪器的动力学排除测定(kinexa)。术语“癌症”和“癌性的”是指或描述典型地特征为细胞生长失控的哺乳动物的生理病状。癌症的实例包括但不限于b细胞淋巴瘤(霍奇金淋巴瘤和/或非霍奇金淋巴瘤)、t细胞淋巴瘤、骨髓瘤、骨髓发育异常综合征、皮肤癌、脑肿瘤、乳腺癌、结肠癌、直肠癌、食道癌、肛门癌、不明原发灶癌、内分泌癌、睾丸癌、肺癌、肝细胞癌、胃癌、胰腺癌、宫颈癌、卵巢癌、肝癌、膀胱癌、泌尿道癌、生殖器癌、甲状腺癌、肾癌、瘤、黑素瘤、头颈癌、脑癌(例如,多形性成胶质细胞瘤)、前列腺癌(包括但不限于雄激素依赖性前列腺癌和雄激素非依赖性前列腺癌)、以及白血病。其它癌症和细胞增生性病症在本领域中将容易识别。术语“肿瘤”和“癌症”在本文中可互换使用,例如,这两个术语包括实体和液体,例如弥散或循环肿瘤。如本文所用,术语“癌症”或“肿瘤”包括恶化前以及恶性癌症和肿瘤。“化学治疗剂”是已知用于在癌症化学疗法中使用的化合物。化学治疗剂的非限制性实例可以包括烷基化剂诸如塞替派和环磷酰胺;烷基磺酸盐,诸如白消安、英丙舒凡(improsulfan)和哌泊舒凡(piposulfan);氮丙啶,诸如苯佐替派(benzodopa)、卡波醌(carboquone)、美妥替哌(meturedopa)和乌瑞替哌(uredopa);乙撑亚胺和甲基蜜胺,包括六甲蜜胺、三乙撑蜜胺、三乙撑磷酰胺(trietylenephosphoramide)、三乙撑硫代磷酰胺(triethylenethiophosphoramide)和三羟甲蜜胺(trimethylolomelamine);番荔枝内酯(尤其是布拉它辛(bullatacin)和布拉它辛酮(bullatacinone));喜树碱(包括合成类似物拓扑替康);苔藓抑素(bryostatin);凯利他汀(callystatin);cc-1065(包括其阿多来新(adozelesin)、卡折来新(carzelesin)和比折来新(bizelesin)合成类似物);念珠藻素(cryptophycin)(特别是念珠藻素1和念珠藻素8);多拉司他丁(dolastatin);倍癌霉素(duocarmycin)(包括合成类似物kw-2189和cb1-tm1);软珊瑚醇(eleutherobin);潘卡他汀(pancratistatin);匍枝珊瑚醇(sarcodictyin);海绵抑素(spongistatin);氮芥,诸如苯丁酸氮芥、萘氮芥(chlornaphazine)、胆磷酰胺(cholophosphamide)、雌莫司汀、异环磷酰胺(ifosfamide)、二氯甲基二乙胺(mechlorethamine)、二氯甲基二乙胺氧化物盐酸盐、美法仑(melphalan)、新氮芥(novembichin)、苯芥胆甾醇(phenesterine)、泼尼莫司汀(prednimustine)、曲磷胺(trofosfamide)、尿嘧啶氮芥;亚硝基脲,诸如卡莫司汀(carmustine)、氯脲菌素(chlorozotocin)、福莫司汀(fotemustine)、洛莫司汀(lomustine)、尼莫司汀(nimustine)和雷莫司汀(ranimustine);抗生素,诸如烯二炔抗生素(例如卡里奇霉素,尤其是卡里奇霉素γ1i和卡里奇霉素ωi1)(参见例如agnew,chem.intl.ed.engl.[英文版国际化学反应],33:183-186(1994));达内霉素(dynemicin),包括达内霉素a;二膦酸盐(bisphosphonate),诸如氯膦酸盐;埃斯培拉霉素(esperamicin);以及新抑癌菌素(neocarzinostatin)发色团和相关色蛋白烯二炔类抗生素发色团、阿克拉霉素(aclacinomysin)、放线菌素(actinomycin)、安曲霉素(authramycin)、重氮丝氨酸(azaserine)、博来霉素(bleomycin)、放线菌素c(cactinomycin)、卡拉比星(carabicin)、洋红霉素(carminomycin)、嗜癌菌素(carzinophilin)、色霉素(chromomycin)、放线菌素d(dactinomycin)、柔红霉素(daunorubicin)、地托比星(detorubicin)、6-重氮基-5-氧-l-正亮氨酸、多柔比星(doxorubicin)(包括吗啉代-多柔比星、氰基吗啉代-多柔比星、2-吡咯啉并-多柔比星和脱氧多柔比星)、表柔比星(epirubicin)、依索比星(esorubicin)、伊达比星(idarubicin)、麻西罗霉素(marcellomycin)、丝裂霉素(mitomycin)(诸如丝裂霉素c)、霉酚酸(mycophenolicacid)、诺加霉素(nogalamycin)、橄榄霉素(olivomycin)、培洛霉素(peplomycin)、泊非霉素(potfiromycin)、嘌呤霉素(puromycin)、三铁阿霉素(quelamycin)、罗多比星(rodorubicin)、链黑菌素(streptonigrin)、链脲霉素(streptozocin)、杀结核菌素(tubercidin)、乌苯美司(ubenimex)、净司他丁(zinostatin)、佐柔比星(zorubicin);抗代谢物,诸如甲氨喋呤和5-氟尿嘧啶(5-fu);叶酸类似物,诸如二甲叶酸(denopterin)、甲氨蝶呤、蝶罗呤(pteropterin)、三甲曲沙(trimetrexate);嘌呤类似物,诸如氟达拉滨(fludarabine)、6-巯基嘌呤、硫咪嘌呤、硫鸟嘌呤;嘧啶类似物,诸如安西他滨(ancitabine)、阿扎胞苷(azacitidine)、6-氮尿苷、卡莫氟(carmofur)、阿糖胞苷(cytarabine)、双脱氧尿苷(dideoxyuridine)、去氧氟尿啶(doxifluridine)、依诺他滨(enocitabine)、氟尿苷(floxuridine);雄激素,诸如卡鲁睾酮(calusterone)、屈他雄酮丙酸盐(dromostanolonepropionate)、环硫雄醇(epitiostanol)、美雄烷(mepitiostane)、睾内酯(testolactone);抗肾上腺剂,诸如氨鲁米特(aminoglutethimide)、米托坦(mitotane)、曲洛司坦(trilostane);叶酸补充剂,诸如亚叶酸(frolinicacid);醋葡内酯(aceglaone);醛磷酰胺糖苷(aldophosphamideglycoside);氨基乙酰丙酸;恩尿嘧啶(eniluracil);安吖啶(amsacrine);倍曲布西(bestrabucil);比生群(bisantrene);依达曲沙(edatraxate);地佛法明(defofamine);地美可辛(demecolcine);地吖醌(diaziquone);依洛尼塞(elfornithine);依利醋铵(elliptiniumacetate);埃博霉素(epothilone);依托格鲁(etoglucid);硝酸镓;羟脲;香菇多糖(lentinan);洛尼代宁(lonidamine);美登木素生物碱,诸如美登素和安丝菌素(ansamitocin));米托胍腙(mitoguazone);米托蒽醌(mitoxantrone);莫哌达醇(mopidamol);二胺硝吖啶(nitracrine);喷司他丁(pentostatin);蛋氨氮芥(phenamet);吡柔比星(pirarubicin);洛索蒽醌(losoxantrone);鬼臼酸(podophyllinicacid);2-乙基酰肼;丙卡巴肼(procarbazine);多糖复合物(jhsnaturalproducts,eugene,oreg.);雷佐生(razoxane);根霉素(rhizoxin);西佐喃(sizofiran);锗螺胺(spirogermanium);细交链孢菌酮酸(tenuazonicacid);三亚胺醌(triaziquone);2,2',2”-三氯三乙胺;单端孢霉烯族毒素(trichothecene)(尤其是t-2毒素、verracurina、杆孢菌素a和蛇形菌素(anguidine));乌拉坦(urethan);长春地辛(vindesine);达卡巴嗪(dacarbazine);甘露醇氮芥(mannomustine);二溴甘露醇(mitobronitol);二溴卫矛醇(mitolactol);哌泊溴烷(pipobroman);gacytosine;阿拉伯糖苷(arabinoside)(“ara-c”);环磷酰胺;紫杉烷(taxoid),例如紫杉醇(bristol-myerssquibboncology,princeton,n.j.)、无氢化蓖麻油白蛋白工程化纳米颗粒紫杉醇制剂(americanpharmaceuticalpartners,schaumberg,ill.)和多西他赛(doxetaxel)(rhone-poulencrorer,antony,france);苯丁酸氮芥(chloranbucil);吉西他滨(gemcitabine);6-硫鸟嘌呤;巯基嘌呤;甲氨蝶呤;铂类似物,诸如顺铂、奥沙利铂(oxabplatin)和卡铂;长春碱(vinblastine);铂;依托泊苷(etoposide)(vp-16);异环磷酰胺;米托蒽醌(mitoxantrone);长春新碱(vincristine);去甲长春碱(navelbine);长春瑞宾(vinorelbine);诺消灵(novantrone);替尼泊苷(teniposide);依达曲沙(edatrexate);道诺霉素(daunomycin);氨基蝶呤;希罗达(xeloda);伊班膦酸盐(ibandronate);伊立替康(camptosar,cpt-11)(包括伊立替康与5-fu和亚叶酸的治疗方案);拓扑异构酶抑制剂rfs2000;二氟甲基鸟氨酸(dmfo);类维生素a,诸如视黄酸;卡培他滨(capecitabine);康普瑞汀(combretastatin);亚叶酸(leucovorin)(lv);奥沙利铂,包括奥沙利铂治疗方案(folfox);拉帕替尼(lapatinib)(tykerb);减少细胞增殖的pkc-α、raf、h-ras、egfr(例如埃罗替尼(erlotinib))和vegf-a的抑制剂以及上述任何物质的药学上可接受的盐、酸或衍生物或其组合。“嵌合抗原受体”(car)是预期用作使用称为过继性细胞转移的技术的癌症疗法的人工t细胞受体。复合物的必需抗原结合、信号传导和刺激性功能已通过基因重组方法减少至单一多肽链,其通常称为嵌合抗原受体(car)。参见例如eshhar,美国专利号7,741,465;eshhar,美国专利申请公开号2012/0093842。特异性构建car以响应于car所结合的特异性抗原而刺激t细胞活化和增殖。术语“嵌合抗原受体”或可替代地“car”是指一组多肽,在最简单的实施方式中典型地是两种多肽,当在免疫效应细胞中表达时,它为该细胞提供针对靶细胞(典型地针对癌细胞)的特异性,并且提供细胞内信号生成。在一些实施方式中,car至少包含细胞外抗原结合结构域、跨膜结构域和细胞质信号传导结构域(本文中也称为“细胞内信号传导结构域”),该细胞质信号传导结构域包含源自刺激分子和/或共刺激分子的功能性信号传导结构域。在一些方面,该组多肽彼此邻接。在一个方面,刺激分子是与t细胞受体复合物相关的ζ链。在一个方面,细胞质信号传导结构域还包含源自如下定义的至少一种共刺激分子的一个或多个功能性信号传导结构域。在一个方面,共刺激分子选自本文描述的共刺激分子,例如4-1bb(即cd137)、cd27和/或cd28。在一个方面,car包含car融合蛋白的氨基末端(n-ter)处的任选前导序列。在一个方面,car在细胞外抗原结合结构域的n-末端还包含前导序列,其中该前导序列任选地在细胞加工和car定位至细胞膜的过程中从抗原结合结构域(例如,scfv)切割。典型地使用“car-t细胞”,其实质已被工程化以含有嵌合抗原受体的t细胞。因此,携带此类car的t淋巴细胞通常被称为car-t淋巴细胞。“密码子优化”或“控制物种密码子偏爱”是指特定宿主细胞的优选密码子使用。如本领域普通技术员将理解的,有利的是修饰编码序列以增强其在特定宿主中的表达。基因密码冗余有64个可能的密码子,但是大部分生物体通常使用这些密码子的子集。最常在物种中使用的密码子被称为最佳密码子,并且不常使用的那些密码子被分类为罕见或低使用密码子。可以制备含有特定原核细胞或真核细胞优选的密码子的最佳化编码序列(还参见murray等人(1989)nucl.acidsres.[核酸研究]17:477-508),例如以与由非最佳序列产生的转录物相比增加翻译速率或产生具有希望特性(诸如更长半衰期)的重组rna转录物。翻译终止密码子也可以被修饰来反映宿主偏好。本领域技术人员将认识到,由于遗传密码的简并性质,可以使用许多核苷酸序列不同的dna化合物来编码本公开的给定多肽。如本文所用,“共同表达”是指两种或更多种基因的表达。基因可以是例如编码作为单一多肽链的单一蛋白质或嵌合蛋白质的核酸。本文所述的sir可以由单个多核苷酸链编码并且被合成为单一多肽链,该多肽链随后裂解成不同多肽,每个多肽表示不同功能单元。在一些实施方式中,在sir由两个或更多个功能多肽单元组成时,不能功能单元使用一条或多条多核苷酸链共同表达。在另一个实施方式中,不同多核苷酸链通过编码可裂解接头(例如,t2a、f2a、p2a、e2a等)的核酸序列连接。在另一个实施方式中,ser-gly-ser-gly(sgsg)基序(seqidno:3065)也被添加在可裂解接头序列上游,以增强裂解的效率。可裂解接头的潜在缺点为位于n末端蛋白质的末端处的小2a标签可影响蛋白质功能或引起蛋白质抗原性的可能性。为了克服此缺点,在一些实施方式中,将弗林蛋白酶裂解位点(rakr)(seqidno:3066)添加在sgsg基序的上游,以便于在翻译之后裂解残余2a肽。编码sir的不同单元的多核苷酸可以通过ires(内部核糖体进入位点)序列连接。或者,sir的不同功能单元通过并非经由接头连接而是由例如两个不同载体编码的两个不同多核苷酸编码。可裂解接头和弗林蛋白酶裂解位点的核酸序列被提供于seqidno:780至seqidno:790中。“保守性取代”或“保守性序列修饰”是指不会显著影响或改变编码蛋白质的结合特征或功能的氨基酸修饰。例如,“保守性序列修饰”是指不会显著影响或改变tcr恒定链、抗体、抗体片段或非免疫球蛋白结合结构域的结合特征或功能的氨基酸修饰。此类保守修饰包括氨基酸取代、添加和缺失。修饰可以通过本领域已知的标准技术,诸如定点诱变和ppcr介导的诱变来引入到本公开的tcr恒定链、抗体或抗体片段、非免疫球蛋白结合结构域或其它蛋白质或多肽。保守氨基酸取代是其中氨基酸残基被具有类似侧链的氨基酸残基替代的取代。在本领域中已经定义了具有类似侧链的氨基酸残基家族。这些家族包括具有以下侧链的氨基酸:碱性侧链(例如赖氨酸、精氨酸、组氨酸)、酸性侧链(例如天冬氨酸、谷氨酸)、不带电极性侧链(例如甘氨酸、天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸、酪氨酸、半胱氨酸、色氨酸)、非极性侧链(例如丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、甲硫氨酸)、β-分支侧链(例如苏氨酸、缬氨酸、异亮氨酸)以及芳香族侧链(例如酪氨酸、苯丙氨酸、色氨酸、组氨酸)。因此,本公开的sir内的一个或多个氨基酸残基可以被来自相同侧链家族的其它氨基酸残基替代,并且可以使用本文描述的结合和/或功能测定法测试改变的sir。术语“t细胞受体α的恒定区”或“t细胞受体α的恒定链”或“tcrα”或“cα”被定义为来自非人类物种,例如小鼠、啮齿动物、猴、猿等的作为seqidno:3010或等效残基(即同源物)提供的蛋白质。本公开还提供tcrα多肽的某些突变。例如,cα中证明本公开的合成免疫受体(sir)的表达增加且错配减少的突变位点位于seqidno3010的位置91、92、93和94。在cα中具有thr48cys(t48c)突变并且在cβ1或cβ2链中具有ser-57-cys(s57c)突变的sir(本文别处更充分描述)在两条tcr恒定链(α和β)之间形成额外二硫键。这进而导致与免疫细胞中的内源性tcr链的错配减少并且功能性增强。类似地,在cα中具有ser61arg(s61r)突变并且在cβ1或cβ2链中具有arg79gly(r79g)突变的sir(本文别处更充分描述)由于配对的“钮与孔(knobandhole)”设计而导致与内源性tcr链的错配减少并且功能性增强。本公开提供了具有根据以下表1的一个或多个或全部突变的cα多肽。人类基因组编码两条高度同源的tcrβ恒定链;tcrβ1(tcrβ1或tcrb1或cβ1)和tcrβ2(tcrβ2或tcrb2或cβ2)。本公开的sir可以包含这两条链中的任一条。类似地,其它哺乳动物物种的tcrβ1或tcrβ2链可以用于本公开的制备sir的方法中。术语“t细胞受体β1的恒定链”或“t细胞受体β1的恒定区”(tcr-β1或tcrβ1或tcrb1或htcrβ-β1或cβ1)被定义为来自非人类物种,例如小鼠、啮齿动物、猴、猿等的作为seqidno:3024或等效残基(即同源物)提供的蛋白质。术语“t细胞受体β2的恒定链”或“t细胞受体β2的恒定区”(tcr-β2或tcrβ2或tcrb2或cβ2)被定义为来自非人类物种,例如小鼠、啮齿动物、猴、猿等的作为seqidno:3025或等效残基(即同源物)提供的蛋白质。术语“t细胞受体β的恒定链”或“t细胞受体β的恒定区”(tcr-β或tcrβ或tcrb或cβ)被定义为来自非人类物种,例如小鼠、啮齿动物、猴、猿等的作为seqidno:3024或seqidno:3025或等效残基(即同源物)提供的蛋白质。已知cβ2(seqidno:3025)和cβ1(seqidno:3024)两者的蛋白质序列。cβ2与β1的序列之间的差异容易通过使用本领域中的典型和普通技术比对序列来鉴别。本公开还提供tcrβ的某些突变。例如,本文提供了cβ中证明合成免疫受体(sir)的表达增加且其与内源性tcrα链的错配降低的突变位点。在cβ1和cβ2中的这些突变位点位于seqidno3025和3024的位置18、22、57、79133、136和139处并且概括于以下表2和3中。cβ1和cβ2中的突变位点在其位置中为相同的。两个序列之间的唯一差别是在位置136处的突变。在此位置处,谷氨酸(e)存在于cβ2中,而缬氨酸存在于cβ1中。术语“pretcra的恒定链”(pretcr-α或pretcrα或pretcra或precα)或“pretcra的恒定区”被定义为来自非人类物种,例如小鼠、啮齿动物、猴、猿等的作为seqidno:3046或seqidno:3047或等效残基(即同源物)提供的蛋白质。术语“pretcra-del48的恒定链”(pretcr-α-del48或pretcrα-del48或pretcra-del48或precα-del48)或“pretcra-del48的恒定区”被定义为来自非人类物种,例如小鼠、啮齿动物、猴、猿等的作为seqidno:3048或等效残基(即同源物)提供的蛋白质。术语“tcr-γ的恒定链”或“tcr-γ的恒定区”(tcr-γ或tcrγ或tcrg或tcr-γ1或tcrγ1或tcrg1或cγ)被定义为来自非人类物种,例如小鼠、啮齿动物、猴、猿等的作为seqidno:3049或等效残基(即同源物)提供的蛋白质。术语“tcr-δ的恒定链”或“tcr-δ的恒定区”(tcr-δ或tcrδ或tcrd或cδ)被定义为来自非人类物种,例如小鼠、啮齿动物、猴、猿等的作为seqidno:3052或等效残基(即同源物)提供的蛋白质。将认识到的是,蛋白质可以彼此具有一致性或同源性并且保留类似或相同功能。本公开包括与本文所述的任一序列具有85%、90%、95%、97%、98%、98.5%、99%或99.9%一致性同时保留生物活性的tcr恒定区。因此,本公开提供具有选自由以下组成的组中的序列的t细胞受体恒定链:(a)与seqidno:3010至少98%一致并且可在位置61、91、92、93和/或94处具有一个或多个突变的氨基酸序列;(b)与seqidno:3024至少98%一致并且可在位置18、22、57、79、133、136和/或139处具有一个或多个突变的氨基酸序列;(c)与seqidno:3025至少98%一致并且可在位置18、22、57、79、133、136和/或139处具有一个或多个突变的氨基酸序列;(d)与seqidno:3046或3047至少98%一致的氨基酸序列;(e)与seqidno:3048至少98%一致的氨基酸序列;(f)与seqidno:3049至少98%一致的氨基酸序列;以及(g)与seqidno:3051或3052至少98%一致的氨基酸序列。(a)-(g)中任一个的t细胞受体恒定链保留与其具有一致性或同源性的野生型t细胞受体恒定链的至少一种生物活性。术语“共刺激分子”是指t细胞上与共刺激配体特异性结合从而介导t细胞的共刺激反应(诸如但不限于增殖)的同源结合配偶体。共刺激分子是除抗原受体或其配体之外的细胞表面分子,其有助于高效的免疫反应。共刺激分子包括但不限于mhci类分子、btla和toll配体受体、以及ox40、cd27、cd28、cd8、icam-1、lfa-1(cd11a/cd18)、icos(cd278)、以及4-1bb(cd137)。此类共刺激分子的其它实例包括cd8、icam-1、gitr、baffr、hvem(lightr)、slamf7、nkp80(klrf1)、nkp44、nkp30、nkp46、cd160、cd19、cd4、cd8α、cd8β、il2rβ、il2rγ、il7rα、itga4、vla1、cd49a、itga4、ia4、cd49d、itga6、vla-6、cd49f、itgad、cd11d、itgae、cd103、itgal、cd11a、lfa-1、itgam、cd11b、itgax、cd11c、itgb1、cd29、itgb2、cd18、lfa-1、itgb7、nkg2d、nkg2c、tnfr2、trance/rankl、dnam1(cd226)、slamf4(cd244、2b4)、cd84、cd96(tactile)、ceacam1、crtam、ly9(cd229)、cd160(by55)、psgl1、cd100(sema4d)、cd69、slamf6(ntb-a、ly108)、slam(slamf1、cd150、ip0-3)、blame(slamf8)、selplg(cd162)、ltbr、lat、gads、slp-76、pag/cbp、cd19a、以及与cd83特异性地结合的配体。共刺激细胞内信号传导结构域可以是共刺激分子的细胞内部分。共刺激分子可以在以下蛋白质家族中表示:tnf受体蛋白、免疫球蛋白样蛋白、细胞因子受体、整联蛋白、信号传导淋巴细胞活化分子(slam蛋白)和活化型nk细胞受体。此类分子的实例包括cd27、cd28、4-1bb(cd137)、ox40、gitr、cd30、cd40、icos、baffr、hvem、icam-1、淋巴细胞功能相关抗原-1(lfa-1)、cd2、cs8、cd7、cd287、light、nkg2c、nkg2d、slamf7、nkp80、nkp30、nkp44、nkp46、cd160、b7-h3、以及与cd83特异性地结合的配体等。细胞内信号传导结构域可以包括衍生出它的分子的整个细胞内部分或整个天然细胞内信号传导结构域,或其功能片段或衍生物。术语“ctcr”是指野生型tcr核酸编码序列和连接至抗原结合结构域的对应野生型tcr蛋白质。在一些实施方式中使用ctcr并且以参考为准。例如,具有cd19结合结构域的ctcr和cd19-sir(包含突变tcr链和cd19结合结构域)将具有不同的表达和/或不同的对靶抗原的结合亲和力。术语“退行性病症”是指基于影响组织或器官的退化细胞变化的连续过程的结果的疾病,该疾病将随着时间推移而逐渐恶化,无论是由于正常身体磨损还是由于生活方式选择诸如锻炼或饮食习惯。示例性退行性疾病包括阿尔兹海默病、腓骨肌萎缩症(charcot–marie–toothdisease)、克罗伊茨费尔特-雅各布病(creutzfeldt–jakobdisease)、弗里德希氏共济失调(friedreich'sataxia)、糖尿病(ii型)和动脉粥样硬化。“源自”(当该术语在本文中使用时)表示第一分子与第二分子之间的关系。它通常是指第一分子和第二分子之间的结构相似性,并不暗示或包括对源自第二分子的第一分子的过程或来源的限制。例如,在源自cd3ζ分子的抗原结合结构域的情况下,抗原结合结构域保留足够的抗体结构,这样使得其具有所需的功能,即结合至抗原的能力。它没有暗示或包括对产生抗体的特定过程的限制,例如,它并不意指为了提供抗原结合结构域,必须从抗体序列开始并删除不需要的序列,或强加突变,以到达抗原结合结构域。短语“与靶抗原的表达相关的疾病”或“如本文描述的疾病相关抗原”包括但不限于与如本文描述的靶抗原的表达相关的疾病或与表达如本文描述的靶抗原的细胞相关的病状,包括例如增生性疾病(诸如癌症或恶性肿瘤)或癌前病状(诸如骨髓增生异常、骨髓发育异常综合征或白血病前期);或与表达如本文描述的靶抗原的细胞相关联的非癌相关适应症。在一个方面,与如本文描述的肿瘤抗原的表达相关的癌症是血液癌症。在一个方面,与如本文描述的肿瘤抗原的表达相关的癌症是实体癌。与本文描述的肿瘤抗原的表达相关的其它疾病包括但不限于非典型和/或非经典癌症、恶性肿瘤、癌前病状或与如本文描述的肿瘤抗原的表达相关的增生性疾病。与如本文描述的靶抗原的表达相关的非癌症相关适应症包括但不限于例如自身免疫性疾病(例如狼疮)、炎性病症(过敏症和哮喘)和移植。在一些实施方式中,表达靶抗原的细胞表达或在任何时间表达编码靶抗原的mrna。在另一个实施方式中,表达靶抗原的细胞产生靶抗原蛋白(例如野生型或突变体),并且靶抗原蛋白可以以正常水平或降低的水平存在。在另一个实施方式中,表达靶抗原的细胞在一个时间点产生可检测水平的靶抗原蛋白,并且随后基本上不产生可检测的靶抗原蛋白。如本文所用的“由遗传修饰细胞靶向的疾病”包括通过磨砺疾病或靶组织或细胞类型的遗传修饰细胞靶向以任何方式参与任何疾病的任何细胞,无论遗传修饰细胞是靶向患病细胞或是靶向健康细胞以完成治疗有利的结果。术语“解离常数(kd)”被定义为受体-配体相互作用解离的平衡常数。如本文所用,“不同的sir集合”或“不同的合成免疫受体集合”是指具有连接至不同t细胞受体恒定链集合的相同结合结构域的多个sir,其中包含结合结构域和不同t细胞恒定链的每种构建体提供了不同的与靶抗原结合的范围和/或改变的表达水平。例如,根据恒定结构域(例如突变体tcra+tcrb)的突变组成,结合结构域与其靶标的结合亲和力发生改变。在一些实施方式中,本公开的sir(单链或异二聚体)包含的结合亲和力大于野生型tcr(例如ctcr)与相同结合结构域的结合亲和力。在一个实施方式中,sir的表达水平比ctcr高至少1.25倍至约10000倍(以及在此之间的任何数值),其中sir和ctcr的不同之处仅在于tcr结构域中的突变。在另一个实施方式中,sir对靶标具有的结合亲和力比具有结合结构域的ctcr对相同抗原的结合亲和力高至少1.5倍至约10000倍。在另一个实施方式中,sir具有的结合亲和力高于ctcr对相同抗原的结合亲和力,但小于具有相同结合结构域的嵌合抗原受体(car)。在一些实施方式中,表达sir的效应细胞与靶抗原的结合比对应表达ctcr的效应细胞的结合大至少1.25倍,但比对应ctcr小100000倍。在一些实施方式中,抗原结合结构域具有约10-4m至10-8m之间的解离常数(kd,反映了其结合亲和力)。在一些实施方式中,抗原结合结构域结合至一种或多种上文所述的抗原。在一些实施方式中,抗原结合结构域对于靶抗原具有的kd为在约10-4m至10-8m之间,例如在约10-5m至10-7m之间,例如在约10-5m至10-6m之间。在一个实施方式中,该抗原结合结构域的结合亲和力比参考抗体小至少五倍、10倍、20倍、30倍、50倍、100倍或1000倍。在一个实施方式中,编码的抗原结合结构域具有的结合亲和力比参考抗体小至少5倍。在一些实施方式中,参考抗体是抗原结合结构域所来源的抗体。如本文所用,“表位”被定义为能够引发免疫反应的抗原部分或者结合至抗体或抗体片段的抗原部分。表位可以是蛋白质序列或子序列。术语“表达载体”是指包含重组多核苷酸的载体,该重组多核苷酸包含与有待表达的核苷酸序列可操作地连接的表达控制序列。表达载体包含足够用于表达的顺式作用元件;其它用于表达的元件可以由宿主细胞供应或在体外表达系统中供应。表达载体包括本领域已知的所有表达载体,包括并入重组多核苷酸的粘粒、质粒(例如,裸露的或含有在脂质体中)和病毒(例如,慢病毒、逆转录病毒、腺病毒和腺相关病毒)。术语sir的“功能多肽单元(fpu)”是指包含在功能上连接至tcr恒定链的氨基末端信号序列的多肽。在一些实施方式中,fpu含有位于信号序列与tcr恒定链之间的抗原结合结构域。在其它实施方式中,fpu缺乏位于信号序列与tcr恒定链之间的抗原结合结构域。fpu可以含有额外序列,诸如接头。作为实例,fpu可以含有抗原结合结构域与tcr恒定链之间的myc2-tag(eqkliseedlgsg)接头。fpu也可以含有可裂解接头(例如p2a、f2a)、ser-gly(sgsg)接头和弗林蛋白酶裂解位点(rakr)。术语“功能部分”当参考sir使用时是指sir的任何部分或片段,该部分或片段保留该部分或片段为其一部分的sir(亲本sir)的生物活性。功能蛋白包括例如sir的在与亲本sir类似的程度、相同的程度或更高程度上保留识别靶细胞或检测、治疗或预防疾病的能力的那些部分。参考亲本sir,功能部分可以包括例如亲本sir的约10%、25%、30%、50%、68%、80%、90%、95%或更大。如本文所用的“遗传修饰细胞”、“重定向细胞”、“遗传工程化细胞”或“修饰细胞”是指已被修饰以表达合成免疫受体(sir)并且可以任选地包含嵌合抗原受体的细胞。例如,表达sir的遗传修饰t淋巴细胞是遗传修饰细胞。术语“免疫病症”是指特征在于免疫系统紊乱的疾病。自身免疫疾病是由对正常身体部分的异常免疫反应引起的病状。自身免疫疾病存在至少80种类型。“免疫效应细胞”(当该术语在本文中使用时)是指参与免疫反应(例如促进免疫效应反应)的细胞。免疫效应细胞的实例包括t细胞,例如α/βt细胞和γ/δt细胞、b细胞、天然杀伤(nk)细胞、天然杀伤t(nkt)细胞、肥大细胞和骨髓衍生性吞噬细胞。术语“免疫效应功能或免疫效应反应”(当该术语在本文中使用时)是指增强或促进对靶细胞的免疫攻击的(例如免疫效应细胞的)功能或反应。例如,免疫效应功能或反应是指t细胞或nk细胞促进靶细胞的杀伤或生长抑制或增殖的特性。在t细胞的情况下,初级刺激和共刺激是免疫效应子功能或响应的实例。术语“细胞内信号传导结构域”(当该术语在本文中使用时)是指分子的细胞内信号传导部分。细胞内信号传导结构域产生信号,该信号促进含有tcr的细胞的免疫效应功能。免疫效应功能的实例包括细胞溶解活性和辅助活性(包括分泌细胞因子)。tcrα/β/γ/δ链不具有其本身的细胞内信号传导结构域,但通过与tcr信号传导复合物(例如cd3z、cd3e、cd3d和cd3g)的具有信号传导结构域的其它链缔合来传输信号。在另一个实施方式中,细胞内信号传导结构域可以包含初级细胞内信号传导结构域。示例性初级细胞内信号传导结构域包括衍生自负责初级刺激或抗原依赖性模拟的分子的那些。在另一个实施方式中,细胞内信号传导结构域可以包含共刺激细胞内结构域。示例性共刺激细胞内信号传导结构域包括衍生自负责共刺激信号或抗原非依赖性刺激的分子的那些。例如,初级细胞内信号传导结构域可以包含cd3z的细胞质序列,并且共刺激细胞内信号传导结构域可以包含来自共受体或共刺激分子诸如cd28或41bb的细胞质序列。初级细胞内信号传导结构域可以包含信号传导基序,该信号传导基序被称为基于免疫受体酪氨酸的活化基序或itam。含有itam的初级细胞质信号传导序列的实例包括但不限于衍生自cd3ζ、常见fcrγ(fcer1g)、fcγriia、fcrβ(fcεr1b)、cd3γ、cd3δ、cd3ε、cd79a、cd79b、dap10和dap12的那些。如本文所用,术语“接头”(也是“接头结构域”或“接头区”)是指将本文所公开的sir的两个或更多个结构域或区域接合在一起的寡肽或多肽。无论何处,该接头的长度可以是1至500个氨基酸。在一些实施方式中,“接头”是可裂解或不可裂解的。除非另外指定,否则本文所用的术语“接头”意指不可裂解接头。可以用于生成sir的示例性不可裂解接头提供于表6d中。所述不可裂解接头可以由允许相邻蛋白质结构域相对于彼此自由运动的柔性残基组成。此类残基的非限制性实例包括甘氨酸和丝氨酸。在一些实施方式中,接头包含非柔性残基。具有非柔性接头的接头的示例性实施方式是eaaak(seqidno:18933)、e-coil(seqidno:18931)、k-coil(seqidno:18932)或pg4sp(18929)。靶向cd19并含有衍生自fmc63抗体的抗原结合结构域的sir显示当在抗原结合结构域与tcr恒定链之间用非柔性接头(例如eaaak(seqidno:18933)、e-coil(seqidno:18931)、k-coil(seqidno:18932)或pg4sp(18929))构建时对靶抗原的结合亲和力与不含有接头的sir相比高约1.5倍。因此,在一些实施方式中,非柔性接头(例如eaaak(seqidno:18933)、e-coil(seqidno:18931)、k-coil(seqidno:18932)、或pg4sp(18929)表示用于构建sir的优选接头。在其它实施方式中,将双链sir的抗原结合结构域与tcr恒定链接合在一起的两个接头共享类似长度。在其它实施方式中,将双链sir的抗原结合结构域和tcr恒定链接合在一起的两个接头的长度相差不多于20个氨基酸,通常不多于10个氨基酸,优选地不多于5个氨基酸,更优选地不多于2个氨基酸。在以下实施方式中,将双链sir的抗原结合结构域和tcr恒定链接合在一起的两个接头具有相同或类似的氨基酸组成。具有相同组成的示例性接头是pg4sp(seqidno:18922)和pg4sp-v2(seqidno:18923)。在一些实施方式中,将双链sir的抗原结合结构域和tcr恒定链接合在一起的两个接头是pg4sp(dnaseqidno:18922;prtseqidno:18929)和pg4sp-v2(dnaseqidno:18923;prtseqidno:18929或18930)。在一些实施方式中,将双链sir的抗原结合结构域和tcr恒定链接合在一起的两个接头是eaaak(seqidno:18926;prtseqidno:18933和18934)和eaaak-v2(dnaseqidno:18927)。在一些实施方式中,将双链sir的抗原结合结构域和tcr恒定链接合在一起的两个接头是e-coil(dnaseqidno:18924)和k-coil(dnaseqidno:18925)。在一些实施方式中,接头可以包括表位标签。在一些实施方式中,表位标签选自由myc标签、v5标签、acv5标签、streptagii、flag标签或ha组成的组。在一些实施方式中,不可裂解接头的长度足以确保两个相邻结构域在空间上不会彼此干扰。在本公开的一个实施方式中,将三个氨基酸残基(gly-ser-gly)添加到接头(例如myc标签或v5标签)的羧基末端,该接头位于sir的抗原结合结构域与tcr恒定链之间。在某些实施方式中,接头可以携带额外序列,诸如限制性酶位点。若干示例性接头的核酸序列提供于seqidno:701至seqidno:725中并且若干示例性接头的氨基酸序列提供于seqidno:2981至seqidno:3003中。如所使用的术语“柔性多肽接头”是指由单独或组合使用的氨基酸(诸如甘氨酸和/或丝氨酸)残基组成的肽接头,以将多肽链连接在一起(例如,将可变重链区和可变氢链区连接在一起)。在一个实施方式中,柔性多肽接头是gly/ser接头并且包含氨基酸序列(gly-gly-gly-ser)n,其中n是等于或大于1的正整数。例如,n=1,n=2,n=3,n=4,n=5,以及n=6,n=7,n=8,n=9,以及n=10。在一个实施方式中,柔性多肽接头包括但不限于(gly4ser)4或(gly4ser)3(seqidno:2500)。在另一个实施方式中,接头包括(gly2ser)、(glyser)或(gly3ser)(seqidno:2501和2502)的多个重复。描述于wo2012/138475中的接头也包括在本公开的范围内,该专利通过引用结合在本文中。可裂解接头的非限制性实例包括2a接头(例如t2a)、小核糖核酸病毒2a样接头、猪捷申病毒的chysel序列(p2a)、thoseaasigna病毒(t2a)、2a样接头或其功能等效物以及其组合。在一些实施方式中,接头序列可以包含引起在2a甘氨酸与2b脯氨酸之间裂解的基序(参见例如t2a序列、seqidno:3061、c末端gly-pro)。若干示例性可裂解接头的核酸序列提供于seqidno:780至seqidno:785中并且若干示例性接头的氨基酸序列提供于seqidno:3060至seqidno:3064中。本文中可以使用的其它可裂解接头是本领域技术人员容易了解的。术语“慢病毒”是指逆转录病毒科的一个属。在逆转录病毒科中慢病毒的独特之处在于它能够感染非分裂细胞;它们可以将显著量的遗传信息递送到宿主细胞的dna中,因此它们是基因递送载体的最有效方法中的一种。hiv、siv和fiv全部都是慢病毒的实例。术语“慢病毒载体”是指源自慢病毒基因组的至少一部分的载体,尤其包括如milone等人,mol.ther.[分子疗法]17(8):1453–1464(2009)中提供的自灭活慢病毒载体。可以在临床中使用的慢病毒载体的其它实例包括但不限于例如来自牛津生物医药公司(oxfordbiomedica)的基因递送技术、来自lentigen的lentimaxtm载体系统等。非临床类型的慢病毒载体也是可获得的并且是本领域技术人员已知的。慢病毒载体的其它实例是plenti-ef1α(seqidno:870)和plenti-ef1α-dwpre(seqidno:871)。如本文所用的“哺乳动物”是指哺乳类的任何成员,包括但不限于,人类和非人灵长类,诸如猩猩和其它猿类和猴物种;农场动物,诸如牛、绵羊、猪、山羊和马;家养哺乳动物,诸如狗和猫;实验室动物,包括啮齿动物诸如小鼠、大鼠和豚鼠等。该术语不指示特定年龄或性别。因此,成人和婴儿受试者以及男性或女性胎儿旨在包括在该术语的范围内。如本文所用,“非天然存在的tcr抗原结合结构域”是指可操作地连接至相对于存在于自然界中的tcr为嵌合和非天然存在的tcr恒定区的结合结构域。换言之,非天然存在的tcr抗原结合结构域使用重组分子生物学技术“工程化”以可操作地连接至tcr并且此外,该抗原结合结构域得自或源自不同于自然界中可见的tcr的分子。不同于自然界中可见的tcr的抗原结合结构域包括抗体vh和vl片段、人源化抗体片段、嵌合抗体片段、受体配体等。术语“可操作地连接”是指在第一组分与第二组分之间功能性连接或缔合,以使得每种组分可以是功能性的。例如,可操作地连接包括调控序列与异源核酸序列之间的导致后者表达的缔合。例如,当第一核酸序列被放置成与第二核酸序列有功能关系时,该第一核酸序列与该第二核酸序列可操作地连接。在可操作地连接的两种多肽的背景下,第一多肽以其将独立于任何连接的方式起作用并且第二多肽在两者之间不存在连接时起作用。在两个或更多个核酸或多肽序列的背景下,“一致性百分比”是指两个或更多个相同的序列。当在比较窗口上(或使用以下序列比较算法之一或通过手动校准和目视检查测量的指定区域)进行比较和比对以获得最大对应性时,如果两个序列具有指定百分比的相同的氨基酸残基或核苷酸(例如,在规定区域,或者当未规定时,在整个序列中,60%一致性,任选地70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%一致性),则两个序列是“基本上相同的”。任选地,一致性存在于长度为至少约50个核苷酸(或10个氨基酸)的区域上,或更优选地在长度为100至500或1000或更多个核苷酸(或20、50、200或更多个氨基酸)的区域上。对于序列比较,通常一个序列充当参考序列,将测试序列与该参考序列比较。当使用序列比较算法时,将测试序列和参考序列输入到计算机中,必要时指定子序列坐标,并且指定序列算法程序参数。可以使用默认程序参数,或者可以指定替代参数。然后,序列比较算法将基于程序参数计算测试序列相对于参考序列的序列一致性百分比。用于比较的序列比对方法在本领域中是熟知的。例如通过smith和waterman,(1970)adv.appl.math.[应用数学进展]2:482c的局部同源性算法,通过needleman和wunsch,(1970)j.mol.biol.[分子生物学期刊]48:443的同源性比对算法,通过pearson和lipman,(1988)proc.nat’l.acad.sci.usa[美国国家科学院院刊]85:2444的相似性方法研究,通过这些算法(威斯康星州麦迪逊的科学大道575号遗传学计算机小组(geneticscomputergroup,575sciencedr.,madison,wi)的威斯康星遗传学软件包中的gap、bestfit、fasta和tfasta)的计算机化实现方式,或通过手动比对和目测检查(参见例如,brent等人,(2003)currentprotocolsinmolecularbiology[分子生物学实验指南]),可以进行用于比较的序列的最佳比对。可以用于确定序列一致性百分比和序列相似性的两个算法实例为blast和blast2.0算法,这两个算法分别描述于altschul等人,(1977)nuc.acidsres.[核酸研究]25:3389-3402;以及altschul等人,(1990)j..mol.bioi.[分子生物学杂志]215:403-410。用于执行blast分析的软件可通过美国国家生物技术信息中心(nationalcenterforbiotechnologyinformation)公开地获得。也可以使用已并入align程序(2.0版)中的e.meyers和w.miller,(1988)comput.appl.biosci.[生物科学中的计算机应用]4:11-17)的算法、使用pam120权重残基表、12的空位长度罚分和4的空位罚分来确定两个氨基酸序列之间的一致性百分比。此外,可以使用已并入gcg软件包(在www.gcg.com可获得)中的gap程序中的needleman和wunsch(1970)j.mol.biol.[分子生物学杂志]48:444-453)算法,使用blossom62矩阵或pam250矩阵以及16、14、12、10、8、6或4的空位权重和1、2、3、4、5或6的长度权重来确定两个氨基酸序列之间的一致性百分比。术语“多核苷酸”、“核酸”或“重组核酸”是指核苷酸的聚合物,诸如脱氧核糖核酸(dna)以及在适当时的核糖核酸(rna)。“蛋白质”或“多肽”两个术语在本文中可互换使用,包含称为氨基酸的化学构建嵌段的一条或多条链,这些化学构建嵌段通过称为肽键的化学键连接在一起。术语“逆转录病毒载体”是指源自逆转录病毒基因组的至少一部分的载体。逆转录病毒载体的实例包括可获自addgene或clontech的mscvneo、mscv-pac(或mscv-puro)、mscv-hygro。逆转录病毒载体的其它实例是mscv-bgl2-avrii-bam-ecor1-xho-bstb1-mlu-sal-clai.i03(seqidno:872)。术语“睡美人转座子”或“睡美人转座子载体”是指源自睡美人转座子基因组的至少一部分的载体。睡美人转座子载体的实例是psbbi-pur(seqidno:874)。编码sir的睡美人转座子载体的其它实例提供于seqidno:875和seqidno:876中。术语“scfv”是指融合蛋白,该融合蛋白包含至少一个包含轻链可变区的抗体片段和至少一个包含重链可变区的抗体片段,其中轻链可变区和重链可变区例如经由合成接头(例如短柔性多肽接头)连续连接并且能够表达为单链多肽,并且其中scfv保留了衍生出它的完整抗体的特异性。除非另有说明,否则如本文所用,scfv可以按任何顺序具有vl和vh可变区,例如相对于多肽的n末端和c末端,scfv可以包括vl-接头-vh或可以包括vh-接头-vl。在本发明中,scfv也描述为vl-gly-ser-接头-vh。例如,fmc63-vl-gly-ser-接头-fmc63-vh是指含有fmc63单克隆抗体的经由由gly和ser组成的接头连接的vl和vh片段的scfv。示例性gly-ser接头的氨基酸序列提供于seqidno:2500中。或者,scfv被描述为(vl+vh)。例如,fmc6-(vl+vh)是指含有fmc63抗体的经由接头连接的vl和vh片段的scfv,其中vl片段位于n末端处。术语“信号传导结构域”是指蛋白质的功能部分,该功能部分在细胞内传递信息,以经由限定的信号传导途径通过生成第二信使或通过响应于此类信使而起到效应子的作用来调节细胞活性。术语“合成免疫受体”或可替代地“sir”是指一组多肽,在一些实施方式中典型地是两种多肽,当在效应细胞中表达时,它为该细胞提供针对靶细胞(典型地针对癌细胞)的特异性,并且提供细胞内信号生成。在一个典型实施方式中,sir包含结合至如本文描述的抗原并且经由任选接头连接至一个或多个t细胞受体恒定链的一个或多个抗原结合结构域(例如,抗体或抗体片段、配体或受体)。在一些实施方式中,该组多肽彼此邻接。在一些实施方式中,sir包含两组或更多组的两个或更多个多肽。每组sir的多肽彼此(功能多肽单元1)邻接,但与另一组(功能多肽单元2)不邻接。在一些方面中,sir的t细胞受体恒定链(或区)选自以下各项的恒定链:人类t细胞受体α(tcr-α或tcrα或tcra或htcr-α或htcrα或htcra或cα)、人类t细胞受体-β1(tcr-β1或tcrβ1或tcrb1或htcr-β1或htcrβ1或htcrb1或cβ1)、人类t细胞受体-β2(tcr-β2或tcrβ2或tcrb2或htcr-β2或htcrβ2或htcrb2或cβ2,也称为tcr-β、tcrβ或tcrb或cβ)、人类pre-t细胞受体α((pretcr-α或pretcrα或pretcra或precα)、人类t细胞受体-γ(tcr-γ或tcrγ或tcrg或或htcr-γ或htcrγ或htcrg或htcrγ1或htcrγ1或cγ)、或人类t细胞受体δ(tcr-δ或tcrd或tcrδ或htcr-δ或htcrd或htcrδ或cδ)。在一些实施方式中,sir的tcr恒定链由其野生型核苷酸序列编码,而在其它方面中,sir的tcr恒定链由非野生型核苷酸序列编码。在一些实施方式中,sir的tcr恒定链由其密码子优化的序列编码。在一些实施方式中,sir的tcr恒定链编码野生型多肽序列,而在其它实施方式中,sir的tcr恒定链编码携带一个或多个突变的多肽。在一些实施方式中,sir的tcr恒定链由其携带一个或多个突变的密码子优化序列编码。包含靶向特定肿瘤标志物x的抗原结合结构域(例如,scfv或tcr)的sir(诸如本文描述的那些)也被称为x-sir或xsir。例如,包含靶向cd19的抗原结合结构域的sir被称为cd19-sir或cd19sir。sir的tcr恒定链/结构域可以来源于最终将使用sir的相同物种中。例如,为了在人类中使用,对于sir的tcr恒定链而言可能有利的是来源于或包含人类tcr恒定链。然而,在一些情况下,对于tcr恒定链而言可能有利的是来源于最终将sir的相同物种,但是被修饰来携带增强tcr恒定链表达的氨基酸取代。例如,为了在人类中使用,对于sir的tcr恒定链而言可能有利的是来源于或包含人类tcr恒定链,但是其中某些氨基酸被来自鼠类tcr恒定链的对应氨基酸置换。此类鼠类化tcr恒定链提供了增加的sir表达。示例性鼠类化tcr恒定链的氨基酸序列提供于seqidno:3017、seqidno:3033至3039(还参见表1--3)。sir或其功能部分可以在氨基末端或羧基末端或两个末端处包含额外氨基酸,这些额外氨基酸不可见于tcr或构成sir的抗原结合结构域的氨基酸序列中。希望的是,额外氨基酸不干扰sir或功能部分的生物功能,例如识别靶细胞、检测癌症、治疗或预防癌症等。更希望的是,与亲本sir的生物活性相比,额外氨基酸增强了生物活性。术语“刺激”是指通过刺激分子(例如,tcr/cd3复合物或car)与其同源配体(或sir情况下的靶抗原)结合而诱导的初级反应,从而介导信号转导事件,诸如但不限于,经由tcr/cd3进行信号转导。刺激可以介导某些分子的改变的表达。术语“刺激分子”是指由免疫细胞(例如,t细胞、nk细胞、b细胞)表达的分子,该分子提供以针对免疫细胞信号传导途径的至少一些方面的刺激方式调节免疫细胞活化的一个或多个细胞质信号传导序列。在一个方面,信号是初级信号,该初级信号通过例如tcr/cd3复合物与载有肽的mhc分子的结合而启动,并且导致了介导t细胞反应,包括但不限于增殖、活化、分化等。以刺激方式起作用的初级细胞质信号传导序列(也称为“初级信号传导结构域”)可含有信号传导基序,该信号传导基序称为基于免疫受体酪氨酸的活化基序或itam。含有itam的细胞质信号传导序列的实例包括但不限于衍生自cd3ζ、常见fcrγ(fcer1g)、fcγriia、fcrβ(fcεr1b)、cd3γ、cd3δ、cd3ε、cd79a、cd79b、dap10和dap12的那些。术语“受试者”旨在包括可以在其中引发免疫反应的活生物体(例如,任何家养的哺乳动物或人)。术语“t细胞”和“t淋巴细胞”在本文中是可互换的并且同义使用。实例包括但不限于原初t细胞(“淋巴细胞祖细胞”)、中枢性记忆t细胞、效应性记忆t细胞、记忆t干细胞(tscm)、ipsc衍生性t细胞、合成t细胞或其组合。术语“治疗作用”是指可以通过各种方式展现的生物学效应,包括但不限于例如肿瘤体积减少、癌细胞数目减少、转移瘤数目减少、预期寿命增加、癌细胞增殖减少、癌细胞存活降低、感染剂效价减小、感染剂菌落计数减少、与疾病病状相关的各种生理症状改善。“治疗作用”还可以通过肽、多核苷酸、细胞和抗体首先预防疾病发生或预防疾病复发的能力来展现。术语“转移载体”是指包含分离的核酸并且可以用于向细胞内部递送该分离的核酸的物质组合物。许多载体在本领域中是已知的,包括但不限于线性多核苷酸、与离子化合物或两亲化合物相关的多核苷酸、质粒、以及病毒。因此,术语“转移载体”包括自主复制的质粒或病毒。该术语还应当解释为还包括便于将核酸转移到细胞中的非质粒和非病毒化合物,例如像聚赖氨酸化合物、脂质体等。病毒转移载体的实例包括但不限于腺病毒载体、腺相关病毒载体、逆转录病毒载体、慢病毒载体等。如本文所用,“治疗(treatment/treating)”是指治疗性治疗或防范性或预防性措施,其中目标在于预防或减慢(减轻)靶向病理学病状,预防病理学病状,得到或获得有益结果,或即使在治疗最终不成功也降低个体发展病状的机会。需要治疗的那些包括已患有病状的那些以及易于患上病状的那些或有待预防病状的那些。如本文所用,“肿瘤”是指所有赘生性细胞生长和增殖(无论恶性或良性)以及所有癌前和癌性细胞和组织。术语“ζ”或可替代地“ζ链”、“cd3-ζ”或“tcr-ζ”被定义为作为以genban登录号bag36664.1提供的蛋白质或来自非人物种例如小鼠、啮齿动物、猴子、猿等的等效残基,并且“ζ刺激结构域”或可替代地“cd3-ζ刺激结构域”或“tcr-ζ刺激结构域”被定义为来自ζ链的细胞质结构域的氨基酸残基或其功能衍生物,这些氨基酸残基或其功能衍生物足以在功能上传递t细胞活化所必需的初始信号。在一个方面,ζ的细胞质结构域包含genbank登录号bag36664.1的残基52至54或来自非人物种(例如,小鼠、啮齿动物、猴、猿等)的为其功能直向同源物的等效残基。在一个方面,“ζ刺激结构域”或“cd3-ζ刺激结构域”是作为seqidno:18提供的序列。在一个方面,“ζ刺激结构域”或“cd3-ζ刺激结构域”是作为seqidno:20提供的序列。如上文和本文别处所述,本公开的sir的每条链具有一般结构:信号肽-(结合结构域)-(任选的接头)-(t细胞受体恒定区)-(任选的辅助分子)。sir的t细胞受体恒定区包含t细胞受体恒定链与具有任选共刺激结构域的cd3信号传导链之间的融合物。示例性tcrβ恒定链和cd3z融合蛋白提供于seqidno:12401-12407中。示例性tcrβ恒定链和具有额外共刺激结构域的cd3z融合蛋白提供于seqidno:12408-12409中。示例性tcrα恒定链和cd3z融合蛋白提供于seqidno:12422-12426中。示例性tcrα恒定链和具有额外共刺激结构的cd3z融合蛋白提供于seqidno:12427-12428中。下文描述了本公开的sir的每个“结构域”或“区段”。本领域技术人员将认识到各种tcr可以被改组并且与不同结合结构域等组合。本公开提供编码本公开的sir的多核苷酸序列、sir多肽、表达构建体、包含本公开的sir或构建体的重组工程化细胞、以及制备和使用此类多肽、多核苷酸和细胞的方法。本公开提供编码合成免疫受体(sir)的分离的核酸分子,其中该sir包含结合至如本文描述的抗原并且连接至一个或多个t细胞受体恒定链的一个或多个抗原结合结构域(例如,抗体或抗体片段、自身抗原、配体或受体)。在一些实施方式中,sir可以包含连接至单一t细胞受体恒定链的单一抗原结合结构域或由该单一抗原结合结构域组成。在一些实施方式中,sir可以包含经由接头连接并且进而连接至单一t细胞受体恒定链(例如seqidno:1169、seqidno:1182、seqidno:10497-10508;10524-10538)的多于一个抗原结合片段(例如vl和vh片段或两个vhh片段)或者由该抗原结合片段组成。在另一个实施方式中,sir包含各自与单独t细胞受体恒定链(例如seqidno:1200)同框连接的两个抗原结合结构域或由该两个抗原结合结构域组成。例如,抗原结合结构域1连接至tcrα的恒定链(cα)以构造功能单元1并且抗原结合结构域2连接至tcrβ的恒定链(cβ)以构造功能单元2。此类sir的两个功能单元在同一细胞中共同表达以变成具有功能活性。在一些实施方式中,sir的两个功能单元使用单一载体共同表达,而在其它实施方式中,两个功能单元在同一细胞中使用不同载体共同表达。在一些实施方式中,sir的两个功能单元通过转染编码两个功能单元的单一mrna序列来共同表达,而在其它实施方式中,两个功能单元通过转染各自编码一个功能单元的两个不同mrna序列来共同表达。在另一个实施方式中,sir包含连接至一个t细胞受体恒定链(功能单元1)但与第二t细胞受体恒定链(例如seqidno:1620)共同表达的抗原结合结构域或者由该抗原结合结构域组成。在此类sir中第二t细胞受体恒定链的目的在于促进功能单元1(例如,连接至t细胞受体恒定链的抗原结合结构域1)的细胞表面表达。这样,第二t细胞受体恒定链可以自身表达或者表达为携带表位标签(例如myc、v5、acv5、g4sx2、streptagii等)的融合蛋白或者表达为携带任何不相关蛋白质片段(例如vl或vh片段)的融合蛋白,该蛋白质片段不会干扰功能单元1的组装和功能。作为实例,sir可以包含连接至cα的抗原结合结构域1或空白(缺乏抗原结合结构域)cβ(例如,seqidno:1620)。此类sir的两个功能单元在同一细胞中共同表达以变成具有功能活性。在一些实施方式中,sir的两个功能单元使用单一载体共同表达,而在其它实施方式中,两个功能单元在同一细胞中使用不同载体共同表达。在一些实施方式中,sir的两个功能单元通过转染编码两个功能单元的单一mrna序列来共同表达,而在其它实施方式中,两个功能单元通过转染各自编码一个功能单元的两个不同mrna序列来共同表达。本文描述的sir可以由单一多核苷酸链编码并且翻译成单一多肽链,该多肽链随后裂解成不同蛋白质。编码sir的核酸分子可以包含一个或多个前导序列(也称为信号肽)。在一个实施方式中,sir的每个功能单元(例如,连接至t细胞受体恒定链的抗原结合结构域加上弗林蛋白酶-sgsg-可裂解接头或t细胞受体恒定链加上弗林蛋白酶-sgsg-可裂解接头)可以在将sir作为i型跨膜蛋白引导至细胞表面的前导序列后面。在一个实施方式中,sir的抗原结合结构域是面向细胞外。在一些实施方式中,前导序列包含seqidno:1至9中任一个的核酸序列和seqidno:2300至seqidno:2302的氨基酸序列。在一些实施方式中,包含限制性酶位点的短核酸序列(3-9个核酸)位于sir的不同亚基之间,例如在信号序列与sir的抗原结合结构域之间或者在抗原结合链与tcr链之间。可以生成具有不同tcr恒定链的合成免疫受体(sir)。tcr恒定链可以由其野生型序列、非野生型序列或密码子优化序列编码。另外,tcr恒定链可以携带特定突变(例如具有表2或3中所示的一个或多个突变的tcrβ恒定链和具有表1中的一个或多个突变的tcrα恒定链),以增强其细胞表面表达和/或与彼此的配对并且减少与内源性tcr连的配对。sir的tcr结构域中的突变分别改变了sir与靶标或细胞的结合亲和力和/或表达。例如,本公开涵盖针对特定抗原靶标的sir的不同群体,其可以基于核酸序列密码子优化和/或tcr链中的突变来设计和筛选以促进结合结构域与tcr结构域之间的配对或表达和/或接头的使用。在一些实施方式中,与表达来自含有相同结合结构域(例如,源自与存在于测试sir中的相同的结合结构域)的集合的另一种sir的可比较免疫效应细胞相比,当在类似条件下测定时,表达来自集合的sir的免疫效应细胞在选自由以下组成的组中的一种或多种特征中显示大于2倍、大于5倍、大于10倍以及甚至大于100倍差异:抗原结合亲和力、细胞表面表达、细胞信号传导、nfat报告基因活性、细胞毒性、细胞因子分泌、增殖、体内持久性、耗尽标记物的表达以及体内活性。本公开涵盖x-sir分子的文库,其中x是抗原结合结构域靶标,使得文库或“集合”提供具有改变的结合亲和力、表达水平和功能特征(例如,细胞毒性、细胞因子产生和长期持久性)的sir。在一些实施方式中,与含有相同结合结构域(例如,源自与存在于测试sir中的相同的scfv的结合结构域)的集合中的另一种sir相比,当在类似条件下测定时,集合中的sir当在免疫效应细胞中表达时在一种或多种选自由以下组成的组中的特征中具有大于2倍、优选地大于5倍、甚至更优选地大于10倍以及甚至更优选地大于100倍差异:抗原结合亲和力、细胞表面表达、细胞毒性、细胞因子分泌、t细胞增殖、t细胞持久性、t细胞耗尽以及体内活性。集合中的不同sir可以用不同dna条形码标记以允许其通过下一代测序或本领域已知的其它技术鉴别。示例性条形码由seqidno:864至869呈现。条形码可以插入在编码sir的载体的便利位置处,以使得它们不会干扰sir的表达。在一个示例性实施方式中,条形码可以插入在紧邻sir的终止密码子的下游。本领域技术人员可以使用一种或多种本文所述的测定筛选此类集合以鉴别具有希望的结合亲和力、表达水平或功能特征的x-sir。不同sir或不同的sir集合可以适用于不同的疾病和疾病病状并且可以被组合来产生不同的和多克隆的免疫反应。因此,表达对靶标具有高亲和力的sir的t细胞可以更有效于短期杀伤肿瘤细胞,但是快速耗尽和/或具有短期体内持久性。表达高亲和力sir的此类t细胞可以与表达低亲和力sir的t细胞组合,后者不能有效于短期杀伤肿瘤细胞,但是不含快速耗尽和/或在体内持续更长时间。本公开的sir(包括不同sir集合)也可以与其它遗传工程化t细胞诸如car-t细胞组合,以生成不同的免疫反应。因此,本公开提供了x-sir的文库。如上文和本文所述,sir包含可操作地连接至一个或多个t细胞受体(tcr)恒定链区域的一个或多个抗原结合结构域。本公开的sir可以包含人类β1链恒定区(cβ1)或人类β2链恒定区(cβ2)。在一个实施方式中,sir的人类恒定β区(1或2)在位置18处包含碱性氨基酸。此碱性氨基酸选自由精氨酸(r)和赖氨酸(k)组成的组。术语“位置18”是指在序列seqidno:3025(对于cβ2)或seqidno:3024(对于cβ1)中的第18个氨基酸残基。除了在位置18处的该突变之外,sir的cβ1或cβ2可以包含其它突变,只要sir的生物功能保持完整。如果虽然不同但也类似地执行该生物功能(例如,更好或更差),则生物活性是完整的。术语“sir的功能”意指是指sir例如以特定亲和力特异性结合至给定抗原和/或通过活化导致t细胞功能活化的细胞信号传导来响应于该抗原的能力,这些t细胞功能诸如活化、增殖、细胞因子分泌和/或细胞毒性。在一个实施方式中,sir除了在位置18处具有碱性氨基酸之外,在cβ1或cβ2链中包含至少一个额外突变。该突变选自由位置22的丙氨酸(a)、位置133的异亮氨酸(i)、位置136的丙氨酸(a)和位置139的组氨酸(h)组成的组,其中所提及的位置是在序列seqidno:3025(对于cβ2)或seqidno:3024(对于cβ1)中的那些位置。在另一个实施方式中,sir包含在cβ1或cβ2链的位置18处的碱性氨基酸和选自由位置22处的丙氨酸(a)、位置133处的异亮氨酸(i)、位置136处的丙氨酸(a)和位置139处的组氨酸(h)组成的组中的两个或更多个额外突变,其中所提及的位置是序列seqidno:3025(对于cβ2)或seqidno:3024(对于cβ1)中的那些。本公开证明在α和β链的恒定区中含有额外半胱氨酸的sir可以促进彼此优选配对,增加所引入sir的总表面表达,改进对靶抗原的结合亲和力并且改进功能性。例如,在cα中具有thr48cys(t48c)并且在cβ1或cβ2链中具有ser-57-cys(s57c)突变的sir在两条链之间具有额外二硫键,该二硫键减少了与内源性tcr链的错配并且增强了功能性。其它二硫键位置(或其组合)如下:cα-t48c和cβ1或cβ2-s57c、cα-s15c和cβ1或cβ2-e15c、cα-t45c和cβ1或cβ2-d59c、cα-t45c和cβ1或cβ2-s77c、cα-y10c和cβ1或cβ2-s17c。克服所引入tcrα-链和tcrβ-链与内源性tcr链的不希望的配对的问题的另一种方法包括使用“钮入孔(knob-into-hole)”或“孔入钮(hole-into-knob)”配置和静电环境。本公开证明在cα中具有ser61arg(s61r)突变并且在cβ1或cβ2突变中具有arg79gly(r79g)突变的sir也导致与内源性tcr链的错配减少和功能性增强。增强所引入tcr链的表达的其它方法(诸如去除n-糖基化位点)也可以在本公开的方法中用于生成具有增加的表面表达和功能性的sir。克服sir的所引入tcrα-链和tcrβ-链与内源性tcr链的不希望的配对的问题的另一种方法包括使用遗传靶向敲除内源性tcrα或/和tcrβ链的表达。内源性tcrα或/和tcrβ链的敲除可以使用本领域已知的许多技术来实现,诸如使用crisp/cas9和zn指核酸酶。seqidno:897和898提供了靶向tcrα和tcrβ基因座的grna的序列。这些grna可以连同cas9mrna一起引入到t细胞或ipsc或干细胞中,以敲除内源性tcrα和tcrβ链的表达。可以使用此类tcrα/β敲除细胞来引入本公开的sir。在一个替代性实施方式中,可以使用相同方法来增强ctcr的表达和链配对,包括表7a中所示的本公开的ctcr。当在内源性tcr链的表达减少或消除的t细胞中表达时,ctcr需要sir的一些功能特性。在本公开的一个替代性实施方式中,sir或ctcr可以首先引入在t细胞或ipsc或干细胞中,随后敲除tcrα和tcrβ链。假如希望在tcrγδ细胞中表达sir或ctcr,则可以使用基本上类似的方法来减少或消除内源性tcrγ或/和tcrδ链的表达。表1--3提供了cα、cβ1和cβ2多肽链的适合取代的非限制性实例。其它取代也是可能的。例如,可以在相同“突变”位置中使用等效氨基酸(即,保守性取代)。在tcrα、tcrβ1和tcrβ2的恒定区中导致所引入sir链的表达增加和与内源性tcr链的错配减少的额外突变可以并入在本公开的sir的设计中。另外,在文献中已知鼠类tcr与人类tcr相比更好地表达。本公开证明含有鼠类化人类tcrα和β链的sir(即,其中人类tcrα和β恒定区的某些氨基酸残基被小鼠tcrα和β链的对应氨基酸置换)与含有人类tcrα和β恒定链的野生型氨基酸序列的sir相比更好地表达。人类tcrα和tcrβ链的额外突变可以基于小鼠tcrα和tcrβ链的序列类似地生成。含有此类鼠类化tcrα和tcrβ链的sir可以在本文所述的测定中容易地测试(例如,使用nluc结合测定的与靶抗原的结合、细胞因子分泌、细胞杀伤、jurkatnfat-gfp测定等),以鉴别导致sir的表达和/或功能活性增加的变体。编码本公开的sir的核酸编码一个或多个t细胞受体恒定链或区域。示例性t细胞受体恒定链或区域的可以用于制备sir的核酸序列提供于seqidno:730至775、10427-10452和10464-10471中。对应氨基酸序列提供于seqidno:3010至3055、12384-12409和12421-12428(表4)。在一些实施方式中,编码所编码sir分子的t细胞受体恒定链的核酸序列包含人类t细胞受体α(tcr-α或tcrα或tcra或cα;seqidno:730和731)、人类t细胞受体β1(tcr-β1或tcrβ1或tcrb1或cβ1;seqidno:744)、人类t细胞受体β2(tcr-β2或tcrβ2或tcrb2或cβ2,也称为tcr-β、tcrβ或tcrb或cβ;seqidno:745和746)、人类pre-t细胞受体α(pretcr-α或pretcrα或pretcra或precα)、人类t细胞受体γ(tcr-γ或tcrγ或tcrg或cγ;seqidno:769)、或人类t细胞受体δ(tcr-δ或tcrd或tcrδ或cδ)的恒定链的野生型序列。表4:在一些实施方式中,编码所编码sir分子的t细胞受体恒定链的核酸序列包含人类t细胞受体-α(tcr-α或tcrα或tcra或cα)、人类t细胞受体-β1(tcr-β1或tcrβ1或tcrb1或cβ1)、人类t细胞受体-β2(tcr-β2或tcrβ2或tcrb2或cβ2,也称为tcr-β、tcrβ或tcrb或cβ)、人类pre-t细胞受体α((pretcr-α或pretcrα或pretcra或precα)、人类t细胞受体-γ(tcr-γ或tcrγ或tcrg或cγ)、或人类t细胞受体-δ(tcr-δ或tcrd或tcrδ或cδ)的非野生型核酸序列。例如,非野生型序列可以是密码子优化的序列和/或包含导致编码多肽突变的一个或多个突变的序列。在一些实施方式中,编码所编码sir分子的t细胞受体恒定链的核酸序列包含人类t细胞受体-α(tcr-α或tcrα或tcra或cα)、人类t细胞受体-β1(tcr-β1或tcrβ1或tcrb1或cβ1)、人类t细胞受体-β2(tcr-β2或tcrβ2或tcrb2或cβ2,也称为tcr-β、tcrβ或tcrb或cβ)、人类pre-t细胞受体α(pretcr-α或pretcrα或pretcra或precα)、人类t细胞受体-γ(tcr-γ或tcrγ或tcrg或cγ)、或人类t细胞受体-δ(tcr-δ或tcrd或tcrδ或cδ)的密码子优化序列。示例性密码子优化的人类tcrβ1恒定区核酸序列提供于seqidno:752中。示例性密码子优化的人类tcrβ2恒定区核酸序列提供于seqidno:749和750。在一些实施方式中,编码所编码sir分子的t细胞受体恒定链的核酸序列包含人类t细胞受体-α(tcr-α或tcrα或tcra或cα)、人类t细胞受体-β1(tcr-β1或tcrβ1或tcrb1或cβ1)、人类t细胞受体-β2(tcr-β2或tcrβ2或tcrb2或cβ2或cβ2,也称为tcr-β、tcrβ或tcrb或cβ)、人类pre-t细胞受体α(pretcr-α或pretcrα或pretcra或precα)、人类t细胞受体-γ(tcr-γ或tcrγ或tcrg或cγ)、或人类t细胞受体-δ(tcr-δ或tcrd或tcrδ或cδ)的恒定链,这些恒定链携带增强sir的链的表达和配对并减少其与内源性t细胞受体链的配对的特异性突变(点突变或缺失或两者)。在一些实施方式中,编码所编码sir分子的t细胞受体恒定链的核酸序列包含人类t细胞受体-α(tcr-α或tcrα或tcra或cα)、人类t细胞受体-β1(tcr-β1或tcrβ1或tcrb1或cβ1)、人类t细胞受体-β2(tcr-β2或tcrβ2或tcrb2或cβ2或cβ2,也称为tcr-β、tcrβ或tcrb或cβ)、人类pre-t细胞受体α(pretcr-α或pretcrα或pretcra或precα)、人类t细胞受体-γ(tcr-γ或tcrγ或tcrg或cγ)、或人类t细胞受体-δ(tcr-δ或tcrd或tcrδ或cδ)的恒定链,这些恒定链被密码子优化并且携带增强sir的链的表达和配对并减少其与内源性t细胞受体链的配对的特异性突变(点突变或缺失或两者)。在一些实施方式中,编码所编码sir分子的t细胞受体恒定链的核酸序列包含犬类t细胞受体-α(tcr-α或tcrα或tcra或cα)、犬类t细胞受体-β(tcr-β或tcrβ或tcrb或cβ)、犬类pre-t细胞受体α((pretcr-α或pretcrα或pretcra或precα)、犬类t细胞受体-γ(tcr-γ或tcrγ或tcrg或cγ)、或犬类t细胞受体-δ(tcr-δ或tcrd或tcrδ或cδ)的野生型、非野生型或密码子优化的恒定链。在一些实施方式中,编码所编码sir分子的t细胞受体恒定链的核酸序列包含犬类t细胞受体-α(tcr-α或tcrα或tcra或cα)、犬类t细胞受体-β(tcr-β或tcrβ或tcrb或cβ)、犬类pre-t细胞受体α(pretcr-α或pretcrα或pretcra或precα)、犬类t细胞受体-γ(tcr-γ或tcrγ或tcrg或cγ)、或犬类t细胞受体-δ(tcr-δ或tcrd或tcrδ或cδ)的恒定链,这些恒定链被密码子优化并且携带增强sir的链的表达和配对并减少其与内源性t细胞受体链的配对的特异性突变(点突变或缺失或两者)。在一些实施方式中,编码所编码sir分子的t细胞受体恒定链的核酸序列包含鼠类t细胞受体-α(tcr-α或tcrα或tcra或cα)、鼠类t细胞受体-β(tcr-β或tcrβ或tcrb或cβ)、鼠类pre-t细胞受体α(pretcr-α或pretcrα或pretcra或precα)、鼠类t细胞受体-γ(tcr-γ或tcrγ或tcrg或cγ)、或鼠类t细胞受体-δ(tcr-δ或tcrd或tcrδ或cδ)的野生型、非野生型或密码子优化的恒定链,这些恒定链可以携带或不携带增强sir的链的表达和配对并减少其与内源性t细胞受体链的配对的特异性突变(点突变或缺失或两者)。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体α(tcr-α或tcrα或tcra或htcra或htcrα或cα)恒定链的如seqidno:730、seqidno:731或seqidno:733所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码人类t细胞受体α的恒定链的氨基酸序列,该氨基酸序列具有氨基酸序列seqidno:3010或seqidno:3011或与氨基酸序列seqidno:3010或seqidno:3011具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰。在某些实施方式中,sir分子的核酸序列编码人类tcra的包含序列seqidno:3010或seqidno:3011的恒定链。在某些实施方式中,sir分子的核酸序列包含编码氨基酸序列seqidno:3010但该序列携带包括以下各项的一个或多个突变的人类t细胞受体α恒定区(链)的核酸序列:在位置91处的丝氨酸(s)、在位置92处的(d)、在位置93处的缬氨酸(v)、在位置94处的脯氨酸(p)、在位置48处的半胱氨酸(c)以及在位置61处的精氨酸(r)(例如,seqidno:732、735、736、737、738、739或740)。在某些实施方式中,sir分子的核酸序列包含编码氨基酸序列seqidno:3010但是其中一个或多个氨基酸被小鼠tcrα恒定链(seqidno:3022)的对应氨基酸置换的人类t细胞受体α恒定区(链)的核酸序列。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体α(tcr-α或tcrα或tcra或htcra或htcrα或cα)恒定链的如seqidno:732所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码携带增强所编码多肽的表达及其与sir的互补tcrb恒定链的链配对并减少与内源性tcrβ链的链配对的氨基酸取代(csdvp)的人类tcra的恒定链。在某些实施方式中,sir的核酸序列包含编码人类t细胞受体α的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3012或与氨基酸序列seqidno:3012具有80-99%一致性的序列的至少一个、五个或十个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcra的恒定链包含氨基酸序列seqidno:3012。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体α(tcr-α或tcrα或tcra或htcra或htcrα或cα)恒定链的如seqidno:737所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码人类tcra的携带增强所编码多肽的表达的氨基酸取代(sdvp)的恒定链。在某些实施方式中,sir的核苷酸序列包含编码人类t细胞受体α(tcr-α或tcrα或tcra或cα)的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3017或与氨基酸序列seqidno:3017具有80-99%一致性的序列的至少一个、五个或十个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcra的恒定链包含氨基酸序列seqidno:3017。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体α(tcr-α或tcrα或tcra或htcra或htcrα或cα)恒定链的如seqidno:740所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码人类tcra的携带增强所编码多肽的表达的氨基酸取代(sd)的恒定链。在某些实施方式中,sir的核酸序列包含编码人类t细胞受体α(tcr-α或tcrα或tcra或cα)的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3020或与氨基酸序列seqidno:3020具有80-99%一致性的序列的至少一个、五个或十个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcra的恒定链包含氨基酸序列seqidno:3020。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体α(tcr-α或tcrα或tcra或htcra或htcrα或cα)恒定链的如seqidno:736所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码人类tcrα的具有thr48cys(t48c)取代的恒定链,该恒定链当与携带s57c(ser57cys)取代的所引入人类突变体tcrβ恒定链共同表达时促进额外链间二硫键形成并且减少与内源性tcrβ链的链配对。在某些实施方式中,sir的核酸序列包含编码人类t细胞受体α(tcr-α或tcrα或tcra或cα)的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3016或与氨基酸序列seqidno:3016具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcra的恒定链包含氨基酸序列seqidno:3016。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体α(tcr-α或tcrα或tcra或htcra或htcrα或cα)恒定链的如seqidno:738所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码人类tcrα的具有ser61arg(s61r)取代的恒定链,该恒定链当与携带r79g(arg79gly)取代的所引入人类突变体tcrβ恒定链共同表达时促进链配对并且减少与内源性tcrβ链的链配对。在某些实施方式中,sir的核酸序列包含编码人类t细胞受体α(tcr-α或tcrα或tcra或cα)的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3018或与氨基酸序列seqidno:3018具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcra的恒定链包含氨基酸序列seqidno:3018。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体α(tcr-α或tcrα或tcra或htcra或htcrα或cα)恒定链的如seqidno:739所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码携带增强所编码多肽的表达及其与互补tcrβ恒定链的配对并减少与内源性tcrβ链的链配对的氨基酸取代(sdvpr)的人类tcra的恒定链。在某些实施方式中,sir的核苷酸序列包含编码人类t细胞受体α(tcr-α或tcrα或tcra或cα)的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3019或与氨基酸序列seqidno:3019具有80-99%一致性的序列的至少一个、五个或十个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcra的恒定链包含氨基酸序列seqidno:3019。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体β(tcr-β或tcrb或tcrβ或htcr-β或htcrb或htcrβ或cβ)恒定链的如seqidno:744或seqidno:745所示的核酸序列。在某些实施方式中,sir的核苷酸序列编码人类t细胞受体β的恒定链的氨基酸序列,该氨基酸序列具有氨基酸序列seqidno:3024或seqidno:3025或与氨基酸序列seqidno:seqidno:3024或seqidno:3025具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcrb的恒定链包含氨基酸序列seqidno:seqidno:3024或seqidno:3025。在某些实施方式中,sir分子的核酸序列包含编码氨基酸序列seqidno:3024或seqidno:3025但携带包括以下各项的一个或多个突变的人类t细胞受体β(tcr-β或tcrb或tcrβ或htcr-β或htcrb或htcrβ或cβ)恒定链的氨基酸序列:在位置18处的碱性氨基酸(arg或lys)、在位置22处的丙氨酸(a)、在位置22处的异亮氨酸(i)、在位置136处的丙氨酸(a)、在位置139处的组氨酸(h)、在位置57处的半胱氨酸(c)和/或在位置79处的甘氨酸(g)。在某些实施方式中,sir分子的核酸序列包含编码人类tcrβ1(seqidno:3024)或tcrβ2(seqidno;3025)的恒定链的氨基酸序列但是其中一个或多个氨基酸被小鼠tcrβ恒定链(seqidno:3047)的对应氨基酸置换的人类t细胞受体β(tcr-β或tcrb或tcrβ或htcr-β或htcrb或htcrβ或cβ)恒定区(链)的核酸序列。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体β(tcr-β或tcrb或tcrβ或htcr-β或htcrb或htcrβ或cβ)恒定链的如seqidno:748所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码携带增强所编码多肽的表达及其与sir的互补tcra恒定链的链配对并减少与内源性tcrβ链的链配对的如seqidno:3028所示的氨基酸取代(kaciah)的人类tcrb的恒定链。在某些实施方式中,sir的核酸序列包含编码人类tcrb的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3028或与氨基酸序列seqidno:3028具有80-99%一致性的序列的至少一个、五个或十个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcrb的恒定链包含氨基酸序列seqidno:3028。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体β(tcr-β或tcrb或tcrβ或htcr-β或htcrb或htcrβ或cβ)恒定链的如seqidno:747所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码人类tcrb的携带ser57cys(s57c)取代的如seqidno:3027所示的恒定链,该恒定链当与携带t48c(thr57cys)取代的人类突变体tcrα恒定链共同表达时促进额外链间二硫键形成并且减少与内源性tcrα链的链配对。在某些实施方式中,sir的核酸序列包含编码人类tcrb的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3027或与氨基酸序列seqidno:3027具有80-99%一致性的序列的至少一个、五个或十个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcrb2的恒定链包含氨基酸序列seqidno:3027。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体β(tcr-β或tcrb或tcrβ或htcr-β或htcrb或htcrβ或cβ)恒定链的如seqidno:762所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码人类tcrb的携带arg79gly(r79g)取代的如seqidno:3042所示的恒定链,该恒定链促进与携带ser61arg(s61r)取代的所引入人类突变体tcrα恒定链的链配对并且减少与内源性tcrα链的链配对。在某些实施方式中,sir的核酸序列包含编码人类t细胞受体β的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3042或与氨基酸序列seqidno:3042具有80-99%一致性的序列的至少一个、五个或十个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcrb的恒定链包含氨基酸序列seqidno:3042。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体β(tcr-β或tcrb或tcrβ或htcr-β或htcrb或htcrβ或cβ)恒定链的如seqidno:753至759所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码人类tcrb的携带增强其表达的氨基酸取代的如seqidno:3034、3035、3036、3037、3038或3039所示的恒定链。在某些实施方式中,sir的核酸序列包含编码人类t细胞受体β的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3034至3038或3039或与氨基酸序列seqidno:3034、3035、3036、3037、3038或3039具有80-99%一致性的序列的至少一个、五个或十个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcrb的恒定链包含氨基酸序列seqidno:3034、3035、3036、3037、3038或3039。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体β(tcr-β或tcrb或tcrβ或htcr-β或htcrb或htcrβ或cβ)恒定链的如seqidno:753所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码人类tcrb的携带增强其表达的氨基酸取代(kaiah)的如seqidno:3033所示的恒定链。在某些实施方式中,sir的核酸序列包含编码人类t细胞受体β的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3033或与氨基酸序列seqidno:3033具有80-99%一致性的序列的至少一个、五个或十个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcrb的恒定链包含氨基酸序列seqidno:3033。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体β(tcr-β或tcrb或tcrβ或htcr-β或htcrb或htcrβ或cβ)恒定链的如seqidno:761所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码人类tcrb的携带增强其表达以及与所引入tcra恒定链的配对的氨基酸取代(kag)的如seqidno:3041所示的恒定链。在某些实施方式中,sir的核苷酸序列包含编码人类t细胞受体β的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3041或与氨基酸序列seqidno:3041具有80-99%一致性的序列的至少一个、五个或十个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcrb的恒定链包含氨基酸序列seqidno:3041。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体β(tcr-β或tcrb或tcrβ或htcr-β或htcrb或htcrβ或cβ)恒定链的如seqidno:760所示的核酸序列。在某些实施方式中,sir分子的核酸序列编码人类tcrb的携带增强其表达以及与所引入tcra恒定链的配对的氨基酸取代(kaihag)的如seqidno:3040所示的恒定链。在某些实施方式中,sir的核苷酸序列包含编码人类t细胞受体β的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3040或与氨基酸序列seqidno:3040具有80-99%一致性的序列的至少一个、五个或十个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcrb的恒定链包含氨基酸序列seqidno:3040。在某些实施方式中,sir分子的核酸序列包含序列seqidno:755、756、757、758或759,该序列编码人类tcrb2的携带增强其表达以及与所引入tcra恒定链的配对的氨基酸取代(k18i133或k18a136或k18a136或k18h139或r18a22或r18)的恒定链。在某些实施方式中,sir的核苷酸序列包含编码人类t细胞受体β2(tcr-β2或tcrβ2或tcrb2或cβ2;也称为tcr-β、tcrβ或tcrb或cβ)的恒定链的氨基酸序列的序列,该氨基酸序列具有氨基酸序列seqidno:3035、3036、3037、3038或3039或与氨基酸序列seqidno:3035、3036、3037、3038或3039具有80-99%一致性的序列的至少一个、五个或十个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcrb2的恒定链包含氨基酸序列seqidno:3035、3036、3037、3038或3039。在某些实施方式中,sir的核酸序列编码错失c末端48个氨基酸的人类pret细胞受体α(pre-tcr-α-del48或pre-tcrα-del48或pre-tcra-del48或precα-del48)的恒定链的氨基酸序列并且具有如seqidno:768所示的核酸序列。在某些实施方式中,sir的核酸序列编码precα-del48恒定链的氨基酸序列,该氨基酸序列具有氨基酸序列seqidno:3048或与氨基酸序列seqidno:3048具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类pre-tcra-del48的恒定链包含氨基酸序列seqidno:3048。在某些实施方式中,sir分子的核酸序列包含人类pret细胞受体α(pre-tcr-α或pre-tcrα或pre-tcra或precα)恒定链的如seqidno:766或767所示的核酸序列。在某些实施方式中,sir的核酸序列编码人类pret细胞受体α的恒定链的氨基酸序列,该氨基酸序列如seqidno:3046或3047所示并且具有氨基酸序列seqidno:3046或3047或与氨基酸序列seqidno:3046或3047具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类pre-tcra的恒定链包含氨基酸序列seqidno:3046或3047。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体γ(tcr-γ或tcrγ或tcrg或htcr-γ、或htcrγ或htcrg或cγ)恒定链的如seqidno:769或770所示的核酸序列。在某些实施方式中,sir的核酸序列编码人类t细胞受体γ的恒定链的氨基酸序列的序列,并且该氨基酸序列具有氨基酸序列seqidno:3049或3050或与氨基酸序列seqidno:3049或3050具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcrg的恒定链包含氨基酸序列seqidno:3049或3050。在某些实施方式中,sir分子的核酸序列包含人类t细胞受体δ(tcr-δ或tcrδ或tcrd或htcr-δ、或htcrδ、或htcrd或cδ)恒定链的如seqidno:771或772所示的核酸序列。在某些实施方式中,sir的核酸序列编码人类t细胞受体δ的恒定链的氨基酸序列,该氨基酸序列如seqidno:3052所示并且具有氨基酸序列seqidno:3052或与氨基酸序列seqidno:3052具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的人类tcr-δ的恒定链包含氨基酸序列seqidno:3052。在某些实施方式中,sir分子的核酸序列包含犬类t细胞受体α(犬类tcr-α或犬类tcrα或犬类tcra或犬类或ctcrα、或ctcrα或ctcra或ccα)恒定链的如seqidno:743所示的核酸序列。在某些实施方式中,sir的核酸序列编码犬类t细胞受体α的恒定链的氨基酸序列,该氨基酸序列如seqidno:3023所示并且具有氨基酸序列seqidno:3023或与氨基酸序列seqidno:3023具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的犬类tcr-α的恒定链包含氨基酸序列seqidno:3023。在某些实施方式中,sir分子的核酸序列包含犬类t细胞受体β(犬类tcr-β或犬类tcrβ或犬类tcrb或犬类或犬类-cβ或ctcrβ、或ctcrβ或ctcrb或ccβ)恒定链的如seqidno:764所示的核酸序列。在某些实施方式中,sir的核酸序列编码犬类t细胞受体β的恒定链的氨基酸序列,该氨基酸序列如seqidno:3044所示并且具有氨基酸序列seqidno:3044或与氨基酸序列seqidno:3044具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的犬类tcr-β的恒定链包含氨基酸序列seqidno:3044。在某些实施方式中,sir分子的核酸序列包含鼠类t细胞受体α(鼠类tcr-α或鼠类tcrα或鼠类tcra或鼠类-cα或mtcrα、或mtcrα或mtcra或mcα)恒定链的如seqidno:742所示的核酸序列。在某些实施方式中,sir的核酸序列编码鼠类t细胞受体α的恒定链的氨基酸序列,该氨基酸序列如seqidno:3022所示并且具有氨基酸序列seqidno:3022或与氨基酸序列seqidno:3022具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的鼠类tcr-α的恒定链包含氨基酸序列seqidno:3022。在某些实施方式中,sir分子的核酸序列包含鼠类t细胞受体β(鼠类tcr-β或鼠类tcrβ或鼠类tcrβ或鼠类-cβ或mtcrβ、或mtcrβ或mtcrb或mcβ)恒定链的如seqidno:763所示的核酸序列。在某些实施方式中,sir的核酸序列编码鼠类t细胞受体β的恒定链的氨基酸序列,该氨基酸序列如seqidno:3043所示并且具有氨基酸序列seqidno:3043或与氨基酸序列seqidno:3043具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰。在某些实施方式中,由sir分子编码的鼠类tcr-β的恒定链包含氨基酸序列seqidno:3043。在某些实施方式中,sir分子的核酸序列包含人类tcra恒定链细胞外结构域与如seqidno:741所示的人类cd3ζ(cd3ζ)链的细胞外结构域、跨膜结构域和细胞质结构域融合的核酸序列。在某些实施方式中,sir的核酸序列编码如seqidno:3021所示并且具有氨基酸序列seqidno:3021或与氨基酸序列seqidno:3021具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰的氨基酸序列。在某些实施方式中,人类tcra恒定链细胞外结构域与由sir分子编码的人类cd3ζ(cd3ζ)链的细胞外结构域、跨膜结构域和细胞质结构域融合的恒定链包含氨基酸序列seqidno:3021。在某些实施方式中,sir分子的核酸序列包含人类tcrb恒定链细胞外结构域与如seqidno:765所示的人类cd3ζ(cd3ζ)链的细胞外结构域、跨膜结构域和细胞质结构域融合的恒定链的核酸序列。在某些实施方式中,sir的核酸序列编码如seqidno:3045所示并且具有氨基酸序列seqidno:3045或与氨基酸序列seqidno:3045具有80-99%一致性的序列的至少一个、五个或九个修饰,但是不超过20个修饰的氨基酸序列。在某些实施方式中,人类tcra恒定链细胞外结构域与由sir分子编码的人类cd3ζ(cd3ζ)链的细胞外结构域、跨膜结构域和细胞质结构域融合的恒定链包含氨基酸序列seqidno:3045。在某些实施方式中,编码本公开的sir的核酸编码包含或源自人类、小鼠或犬类来源的tcra、tcrb、pre-tcra、tcr-γ或tcr-δ链的恒定链的单一t细胞受体恒定链。具有单一tcr恒定链的示例性sir由克隆id:051216-f04表示,其核酸和氨基酸序列分别以seqidno:1023和3258给出。在某些实施方式中,编码本公开的sir的核酸编码包含或源自人类、小鼠或犬类来源的tcra、tcrb、pre-tcra、tcr-γ或tcr-δ链的两条t细胞受体恒定链。具有两条tcr恒定链的示例性sir由克隆id:102615-c08表示,其核酸和氨基酸序列分别以seqidno:1200和3435给出。在某些实施方式中,sir的两条t细胞受体恒定链可以具有相同类型(例如tcra/tcra;tcrb/tcrb;pretcra/pretcra;tcrγ/tcrγ;以及tcr-δ/tcr-δ)。具有相同类型的两条tcr恒定链的示例性sir是克隆id:021116-e08(seqidno:905)、克隆id:012216-p08(seqidno:906)、克隆idno:012216-q05(seqidno:907)、克隆idno:012216-r04(seqidno:908)和克隆idno:012216-s02(seqidno:909)。在另一个实施方式中,sir的两条t细胞受体恒定链具有不同类型(例如tcra/tcrb;pretcra/tcrb;tcrγ/tcr-δ等)。具有不同类型的两条tcr恒定链的示例性sir由克隆id:102615-c08表示,其核酸和氨基酸序列分别以seqidno:1200和3435给出。如上文所述,本公开的sir包含连接至抗原结合结构域的tcr结构域。因此,本公开的sir可以包含一个或多个抗原结合结构域(例如,抗体或抗体片段、配体或受体)和一个或多个t细胞受体恒定链(如本文上文所述),其中所述一个或多个抗原结合结构域结合至靶抗原。非限制性示例性靶抗原包括:cd19;cd123;cd22;cd23、cd30;cd171;cs-1(也称为cd2亚单位1、cracc、slamf7、cd319和19a24);c型凝集素样分子-1(cll-1或clecl1);cd33;表皮生长因子受体变体iii(egfrviii);神经节苷脂g2(gd2);神经节苷脂gd3(aneu5ac(2-8)aneu5ac(2-3)bdgalp(l-4)bdglcp(l-l)cer);tnf受体家族成员b细胞成熟(bcma);tn抗原((tnag)或(galnacα-ser/thr));前列腺特异性膜抗原(psma);受体酪氨酸激酶样孤儿受体1(ror1);fms样酪氨酸激酶3(flt3);肿瘤相关性糖蛋白72(tag72);cd38;cd44v6;在急性白血病或淋巴瘤上而不在造血祖细胞上表达的糖基化cd43表位、在非造血癌症上表达的糖基化cd43表位、癌胚抗原(cea);表皮细胞粘附分子(epcam);b7h3(cd276);kit(cd117);白介素-13受体亚基α-2(il-13ra2或cd213a2);间皮素;白介素11受体α(il-llra);前列腺干细胞抗原(psca);蛋白酶丝氨酸21(睾蛋白或prss21);血管内皮生长因子受体2(vegfr2);路易斯(y)抗原;cd24;血小板源性生长因子受体β(pdgfr-β);阶段特异性胚胎抗原-4(ssea-4);cd20;叶酸受体α(fra或fr1);叶酸受体β(frb);受体酪氨酸蛋白激酶erbb2(her2/neu);细胞表面相关粘蛋白1(muc1);表皮生长因子受体(egfr);神经细胞粘附分子(ncam);前列腺酶;前列腺酸性磷酸酶(pap);突变的延长因子2(elf2m);肝配蛋白b2;成纤维细胞活化蛋白α(fap);胰岛素样生长因子1受体(igf-i受体)、碳酸酐酶ix(calx);蛋白酶体(前体、巨蛋白因子)亚基,β型,9(lmp2);糖蛋白100(gpl00);由裂点簇区(bcr)和阿贝尔森(abelson)鼠类白血病病毒癌基因同源物1(abl)组成的癌基因融合蛋白(bcr-abl);酪氨酸酶;肝配蛋白a型受体2(epha2);唾液酸化路易斯粘附分子(sle);神经节苷脂gm3(aneu5ac(2-3)bdclalp(l-4)bdglcp(l-1)cer);转谷氨酰胺酶5(tgs5);高分子量-黑素瘤相关抗原(hmwmaa);o-乙酰基-gd2神经节苷脂(oacgd2);\肿瘤内皮标记1(tem1/cd248);肿瘤内皮标记7相关(tem7r);密封蛋白6(cldn6);甲状腺刺激性激素受体(tshr);g蛋白偶联受体c类5组成员d(gprc5d);x染色体开放阅读框61(cxorf61);cd97;cd179a;间变性淋巴瘤激酶(alk);聚唾液酸;胎盘特异性1(plac1);globoh糖神经酰胺六糖部分(globoh);乳腺分化抗原(ny-br-1);尿空斑蛋白2(upk2);甲型肝炎病毒细胞受体1(havcr1);肾上腺受体β3(adrb3);泛连接蛋白3(panx3);g蛋白偶联受体20(gpr20);淋巴细胞抗原6复合物,基因座k9(ly6k);嗅觉受体51e2(or51e2);tcrγ替代性阅读框蛋白质(tarp);威尔姆氏瘤蛋白质(wt1);癌症/睾丸抗原1(ny-es0-1);癌症/睾丸抗原2(lage-1a);黑素瘤相关抗原1(mage-a1);位于染色体12p上的ets易位变体基因6(etv6-aml);精子蛋白17(spa17);x抗原家族成员la(xagel);血管生成素结合细胞表面受体2(tie2);黑素瘤癌症睾丸抗原-1(mad-ct-1);黑素瘤癌症睾丸抗原-2(mad-ct-2);fos相关抗原1;肿瘤蛋白p53(p53);p53突变体;前列腺特异性蛋白;存活素;端粒酶;前列腺癌肿瘤抗原-1(pcta-1或半乳凝素8)、由t细胞识别的黑素瘤抗原1(melana或marti);大鼠肉瘤(ras)突变体;人类端粒酶逆转录酶(htert);肉瘤易位裂点;黑素瘤细胞凋亡抑制剂(ml-iap);erg(跨膜蛋白酶,丝氨酸2(tmprss2)ets融合基因);n-乙酰基葡萄糖胺基-转移酶v(na17);配对框蛋白质pax-3(pax3);雄激素受体;周期素bl;v-myc禽类骨髓细胞瘤病病毒癌基因神经母细胞瘤衍生性同源物(mycn);ras同源家族成员c(rhoc);酪氨酸酶相关蛋白2(trp-2);细胞色素p450lb1(cyplb1);ccctc-结合因子(锌指蛋白)样(boris或印迹位点调节剂同级物)、由t细胞识别的鳞状细胞癌抗原3(sart3);配对框蛋白质pax-5(pax5);前顶体蛋白结合蛋白sp32(oy-tesl);淋巴细胞特异性蛋白质酪氨酸激酶(lck);a激酶锚定蛋白4(akap-4);滑膜肉瘤,x裂点2(ssx2);晚期糖基化终产物受体(rage-1);肾泛素1(rul);肾泛素2(ru2);豆荚蛋白;人乳头状瘤病毒e6(hpve6);人乳头状瘤病毒e7(hpve7);肠道羧基酯酶;突变的热休克蛋白70-2(muthsp70-2);cd79a;cd79b;cd72;白细胞相关免疫球蛋白样受体1(lairl);iga受体的fc片段(fcar或cd89);白细胞免疫球蛋白样受体子家族a成员2(lilra2);cd300分子样家族成员f(cd300lf);c型凝集素结构域家族12成员a(clec12a);骨髓基质细胞抗原2(bst2);egf样含模块粘蛋白样激素受体样2(emr2);淋巴细胞抗原75(ly75);磷脂酰肌醇聚糖-3(gpc3);fc受体样5(fcrl5);以及免疫球蛋白λ样多肽1(iglll)、mpl、生物素、c-myc表位标签、cd34、lamp1trop2、gfrα4、cdh17、cdh6、nybr1、cdh19、cd200r、slea(ca19.9;唾液酸化路易斯抗原)岩藻糖基-gm1、ptk7、gpnmb、cdh1-cd324、dll3、cd276/b7h3、il11ra、il13ra2、cd179b-igll1、tcrγ-δ、nkg2d、cd32(fcgr2a)、tnag、tim1-/hvcr1、csf2ra(gm-csfr-α)、tgfβr2、lewsag、tcr-β1链、tcr-β2链、tcr-γ链、tcr-δ链、fitc、促黄体生成激素受体(lhr)、卵泡刺激素受体(fshr)、绒膜促性腺激素激素受体(cghr)、ccr4、gd3、slamf6、slamf4、hiv1包膜糖蛋白、htlv1-tax、cmvpp65、ebv-ebna3c、kshvk8.1、kshv-gh、甲型流感凝集素(ha)、gad、pdl1、鸟苷酸环化酶c(gcc)、桥粒芯蛋白自身抗体3(dsg3)、桥粒芯蛋白自身抗体1(dsg1)、hla、hla-a、hla-a2、hla-b、hla-c、hla-dp、hla-dm、hla-doa、hla-dob、hla-dq、hla-dr、hla-g、ige、cd99、rasg12v、组织因子1(tf1)、afp、gprc5d、密封蛋白18.2(cld18a2或cldn18a.2))、p-糖蛋白、steap1、liv1、粘连蛋白-4、cripto、gpa33、bst1/cd157、低电导率氯离子通道和由tnt抗体识别的抗原。在一些实施方式中,sir多肽分子的抗原结合结构域结合至肿瘤抗原。可以由sir多肽靶向的肿瘤抗原的非限制性实例包括tshr、cd171、cs-1、cll-1、gd3、tnag、flt3、cd38、cd44v6、b7h3、kit、il-13ra2、il-11ra、psca、prss21、vegfr2、lewisy、cd24、pdgfr-β、ssea-4、muc1、egfr、ncam、calx、lmp2、epha2、岩藻糖基gm1、sle、gm3、tgs5、hmwmaa、o-乙酰基-gd2、叶酸受体β、tem1/cd248、tem7r、cldn6、gprc5d、cxorf61、cd97、cd179a、alk、聚唾液酸、plac1、globoh、ny-br-1、upk2、havcr1、adrb3、panx3、gpr20、ly6k、or51e2、tarp、wt1、etv6-aml、精子蛋白17、xage1、tie2、mad-ct-1、mad-ct-2、fos相关抗原1、p53突变体、htert、肉瘤易位裂点、ml-iap、erg(tmprss2ets融合基因)、na17、pax3、雄激素受体、周期素b1、mycn、rhoc、cyp1b1、boris、sart3、pax5、oy-tes1、lck、akap-4、ssx2、cd79a、cd79b、cd72、lair1、fcar、lilra2、cd300lf、clec12a、bst2、emr2、ly75、gpc3、fcrl5以及iglll。在一些实施方式中,sir多肽分子的抗原结合结构域结合至与hla-a2相关的抗原。与hla-a2相关地识别的抗原的非限制性实例包括tarp、wt1、htert、gp100、酪氨酸酶、mart1、ny-eso1、cmvpp65、ebvebna3c、hiv1gag、htlv1-tax、pr1、cmvpp65、ebv-ebna3c、rasg12v突变体以及gad。在一些实施方式中,sir多肽分子的抗原结合结构域包含结合至自身抗体的自身抗原或其片段。自身抗原的非限制性实例包括dsg1和dsg3。在一些实施方式中,sir多肽分子的抗原结合结构域源自或包含抗体、抗体片段、scfv、fv、fab、(fab')2、单结构域抗体(sdab)、vh或vl结构域、骆驼科vhh结构域、或非免疫球蛋白抗原结合支架,例如darpin、亲合体、affilin、adnectin、affitin、obodies、repebody、fynomer、alphabody、avimer、atrimer、centyrin、pronectin、anticalin、kunitz型结构域、armadillo重复蛋白、自身抗原、受体或配体的野生型或非野生型序列。在一些实施方式中,所编码sir多肽含有多于一个抗原结合结构域。在实施方式中,抗原结合结构域直接或经由任选接头可操作地连接至tcr结构域的nh2末端(即,tcr-α、tcr-β1、tcr-β2、pretcr-α、pre-tcr-α-del48、tcr-γ或tcr-δ的恒定链)。若干示例性接头的核酸和氨基酸序列提供于seqidno:701-725、18922-18927和2981-3003、18929-18934中。编码示例性此类sir的构建体提供于克隆idno:082815-g07中。所编码sir多肽的氨基酸序列对应于seqidno:3855。在一些实施方式中,sir多肽分子的抗原结合结构域源自或构成抗体的vl和vh结构域,这些结构域单独连接至t细胞受体的两条恒定链(即,tcr-α、tcr-β1、tcr-β2、pretcr-α、pre-tcr-α-del48、tcr-γ或tcr-δ或其突变体或变体的恒定链,如本文描述的)的nh2末端,以共同构成单一抗原结合结构域。靶向cd19的示例性此类sir提供于克隆idno:102615-c08中。该sir的氨基酸序列对应于seqidno:3435。在该sir中,源自cd19单克隆抗体fmc63的vl片段经由接头连接至突变体(kaciah)人类tcrb链的恒定区,而源自fmc63单克隆抗体的vh片段经由接头连接至突变体(csdvp)人类tcrα链的恒定区。在一些实施方式中,sir多肽具有源自或构成抗体的两个或更多个抗原结合结构域,这些抗体被表达为单链可变片段(scfv)并且单独连接至t细胞受体的两条恒定链(即,tcr-α、tcr-β1、tcr-β2、pretcr-α、pre-tcr-α-del48、tcr-γ、或tcr-δ或其变体或突变体的恒定链)的nh2末端。在一些实施方式中,所编码sir分子的两个(或更多个)抗原结合结构域由编码两个单链可变片段(scfv)的核苷酸序列编码,该两个单链可变片段同框融合至源自t细胞受体的两条恒定链(即,tcr-α、tcr-β1、tcr-β2、pretcr-α、pre-tcr-α-del48、tcr-γ、或tcr-δ的恒定链)。所编码两个scfv片段可以靶向相同抗原(即非特异性sir)或不同抗原(即,双特异性或多特异性sir)。在非特异性sir的情况下,两个scfv可以编码具有相同氨基酸序列或不同氨基酸序列的多肽。另外,在非特异性sir的情况下,在两个scfv由具有相同氨基酸序列的多肽编码时,编码两个相同scfv多肽的核苷酸序列可以是相同或不相同的。具有两个scfv的示例性非特异性sir由seqidno:1026表示。该sir的两个抗原结合结构域构成源自两种不同单克隆抗体cd19bu12和fmc63的scfv,该两种抗体靶向人类cd19抗原。具有两个scfv的示例性多特异性sir由seqidno:1028表示。该sir的两个抗原结合结构域构成源自两种不同单克隆抗体cd19bu12和cd20-2f2的scfv,该两种抗体分别靶向人类cd19和cd20抗原。此sir的实例由040716-b04(cd8sp-cd19bu12-scfv-v5-[htcrb-kaciah]-f-p2a-sp-cd20-2f2-scfv-myc-[htcra-csdvp]-f-f2a-pac)表示,其靶向cd19和cd20,并且对应的氨基酸序列由seqidno:1028表示。具有两个结合结构域的示例性sir由040716-b04(cd8sp-cd19bu12-scfv-v5-[htcrb-kaciah]-f-p2a-sp-cd20-2f2-scfv-myc-[htcra-csdvp]-f-f2a-pac)表示,其靶向cd19和cd20,对应氨基酸序列由seqidno:1028表示。两个scfv多肽片段可以靶向相同抗原(即非特异性sir)或不同抗原(即,双特异性或多特异性sir)。在非特异性sir的情况下,两个scfv可以具有相同氨基酸序列或不同氨基酸序列。靶向两种不同抗原的示例性sir由seqidno:1028和1163表示。在某些实施方式中,两种sir多肽的抗原结合结构域的结构类似(例如,两个抗原结合结构域是scfv或骆驼科vhh结构域或亲合体或vl或vh)。例如,第一sir多肽的抗原结合结构域包含靶向her2的骆驼科vhh结构域并且第二sir多肽的抗原结合结构域包含靶向her3的vhh结构域。两个抗原结合结构域均由vl链组成的sir是cd8sp-fmc63-11-vl-v5-[tcrb-kaciah]-f-p2a-fmc63vl-myc-[tcra-csdvp]-f-f2a-pac并且由seqidno:10474表示。在一个实施方式中,两种sir多肽的抗原结合结构域的结构不类似(例如,第一抗原结合结构域是scfv并且第二抗原结合结构域是骆驼科vhh)。示例性此类sir是cd8sp-il6r-304-vhh-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vl-gly-ser-gly-接头-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1166)。在某些实施方式中,第一sir多肽的抗原结合结构域(功能多肽单元1)包含靶向cd123的骆驼科vhh结构域并且第二sir多肽的抗原结合结构域(功能多肽单元2)包含靶向mpl的scfv。在一些实施方式中,所编码sir多肽的抗原结合结构域由对应野生型序列或非野生型序列抗体、单结构域抗体(sdab)、vh结构域、vl结构域、骆驼科vhh结构域、或非免疫球蛋白支架,例如darpin、亲合体、affilin、adnectin、affitin、obodies、repebody、fynomer、alphabody、avimer、atrimer、centyrin、pronectin、anticalin、kunitz型结构域、armadillo重复蛋白、自身抗原、受体或配体的密码子优化的核苷酸序列编码。在一些实施方式中,sir多肽的一个或多个编码的抗原结合结构域包含seqidno2307至2482和12042至12159的轻链可变结构域(vl或vl)氨基酸序列(其中至多9个氨基酸残基但不多于10个氨基酸被任何其它氨基酸残基置换),或与氨基酸序列seqidno2307至2482和12042至12159具有80-100%一致性的序列,或与seqidno2307至2482和12042至12159的互补决定区(cdr)具有98-100%一致性的序列中的任一个或多个。表5示出本公开中所用的示例性vl结构域的cdr1-3的靶抗原、名称、seqidno(dna)、seqidno(prt)、seqidno(prt)。在一些实施方式中,sir多肽的一个或多个编码的抗原结合结构域包含seqidno2506至2680和12160至12278的重链可变结构域(vh或vh)氨基酸序列(其中至多9个氨基酸残基但不多于10个氨基酸被任何其它氨基酸残基置换),或与氨基酸序列seqidno2506至2680和12160至12278具有80-100%一致性的序列,或与seqidno2506至2680和12160至12278的互补决定区(cdr)具有98-100%一致性的序列中的任一个或多个。表5示出本公开中所用的示例性vh结构域的cdr1-3的靶抗原,示出靶抗原、名称、seqidno(dna)、seqidno(prt)、seqidno(prt)。vh片段的名称可以用于基于后者的名称鉴别对应vl片段。例如,vh片段alk-48-vh(seqidno:226)源自与vl片段alk-48-vl(seqidno:16)相同的抗体或scfv,并且两种组分可以一起用于制成靶向alk的scfv或sir。在某些情况下,表5列出了具有相同名称后跟编号的两个或更多个vl或vh片段,诸如fmc63(seqidno:30)、fmc63-[2]-vl(seqidno:31)和fmc63-[3]-vl(seqidno:32)。在此类情况下,任一条以上fmc63vl链可以连接至任一条fmc63-vh链(seqidno:241和242),以基于fmc63基结合结构域产生对应sir。在一些实施方式中,sir多肽的一个或多个编码的抗原结合结构域包含seqidno2701至2725和12279-12294的骆驼科单结构域抗体(vhh或vhh)氨基酸序列(其中至多9个氨基酸残基但不多于10个氨基酸被任何其它氨基酸残基置换),或与氨基酸序列seqidno2701至2725和12279-12294具有80-100%一致性的序列,或在seqidno2701至2725和12279-12294的三个互补决定区(cdr)中具有98-100%一致性的序列中的任一个或多个。表5示出本公开中所用的示例性vhh结构域的靶抗原、名称、seqidno(dna)和seqidno(prt)。在一些实施方式中,sir多肽的一个或多个编码的抗原结合结构域包含seqidno2728至2732和12296-12301的非免疫球蛋白抗原结合支架氨基酸序列(其中至多9个氨基酸残基但不多于10个氨基酸被任何其它氨基酸残基置换),或与氨基酸序列seqidno2728至2732和12296-12301具有80-100%一致性的序列中的任一个或多个。表6a示出本公开中所用的示例性非免疫球蛋白抗原结合支架的靶抗原、名称、seqidno(dna)、seqidno(prt)、名称。在一些实施方式中,sir多肽的一个或多个编码的抗原结合结构域包含seqidno2736至2747的受体氨基酸序列(其中至多19个氨基酸残基但不多于20个氨基酸被任何其它氨基酸残基置换),或与氨基酸序列seqidno2736至2747具有80-100%一致性的序列中的任一个或多个。表6a示出靶抗原、seqidno(dna)、seqidno(prt)和名称。在一些实施方式中,sir多肽的一个或多个编码的抗原结合结构域包含seqidno2748的自身抗原氨基酸序列(其中至多19个氨基酸残基但不多于20个氨基酸被任何其它氨基酸残基置换),或与氨基酸序列seqidno2748具有80-100%一致性的序列中的任一个或多个。表6a示出靶抗原、seqidno(dna)、seqidno(prt)和名称。在一些实施方式中,sir分子的一个或多个编码的抗原结合结构域包含seqidno2758至2768和12359-12361和18918的配体氨基酸序列(其中至多19个氨基酸残基但不多于20个氨基酸被任何其它氨基酸残基置换),或与氨基酸序列seqidno2758至2768和12359-12361和18918具有80-100%一致性的序列中的任一个或多个。表6a示出靶抗原、seqidno(dna)、seqidno(prt)和名称。在一些实施方式中,sir多肽的一个或多个编码的抗原结合结构域包含seqidno2770至2939、12303-12357和18162-18224的scfv氨基酸序列(其中至多18个氨基酸残基但不多于20个氨基酸被任何其它氨基酸残基置换),或与氨基酸序列seqidno2770至2939、12303-12357和18162-18224具有80-100%一致性的序列,或在seqidno2770至2939、12303-12357和18162-182244的六个互补决定区(cdr)中具有98-100%一致性的序列中的任一个或多个。表5示出本公开中所用的示例性scfv的靶抗原、seqidno(dna)、seqidno(prt)、名称和氨基酸序列。在一些实施方式中,sir多肽的一个或多个编码的抗原结合结构域包含靶向该抗原的vl和vh片段的抗原结合部分(例如cdr)中的任一个或多个。表5中列出了靶向不同抗原的vl和vh片段的cdr1-3的seqidno。在一些实施方式中,sir多肽的一个或多个编码的抗原结合结构域包含含有sir多肽的scfv的vl和vh片段的抗原结合部分(例如cdr)中的任一个或多个。表5中列出了包含靶向不同抗原的scfv片段的vl和vh片段的cdr1-3的seqidno。表6a中列出了靶向不同抗原的scfv片段的seqidno(dna)和seqidno(prt)并且其对应cdr1-3的序列可以通过本领域已知的方法或者由其在表5中列出的组分vl和vh片段的cdr1-3的seqidno确定。在一些实施方式中,sir多肽的一个或多个编码的抗原结合结构域包含靶向该抗原的vhh片段的抗原结合部分(例如cdr)中的任一个或多个。表5中列出了靶向不同抗原的vhh片段的seqidno并且其对应cdr1-3的序列可以通过本领域已知的方法来确定。在一个实施方式中,sir的抗原结合结构域是已知结合此靶抗原的受体的抗原结合部分。在一些实施方式中,sir多肽的一个或多个编码的抗原结合结构域包含含有sir多肽的受体的抗原结合部分中的任一个或多个。在一些实施方式中,sir多肽的一个或多个编码的抗原结合结构域包含含有sir多肽的配体的抗原结合部分中的任一个或多个。在一些实施方式中,sir多肽的一个或多个编码的抗原结合结构域包含含有sir多肽的非免疫球蛋白支架的抗原结合部分中的任一个或多个。在另一个实施方式中,本公开提供结合至表7a--7h中所述的不同靶标上与本公开的任一sir相同的表位的sir(即,具有与本公开的任一sir交叉竞争结合不同靶标的能力的sir)。在一些实施方式中,这些sir的抗原特异性结构域可以由用作sir的组分的抗体的vl片段、vh片段和/或scfv片段确定。在一些实施方式中,用于交叉竞争研究中确定由表7a--7h中所述的本公开的sir识别的靶标表位的参考抗体是具有如seqidno:seqidno2770至2939、12303-12357和18162-18224(表6b)所示的序列的scfv。在一个示例性实施方式中,由seqidno:2770表示的参考scfvfmc63可以在交叉竞争研究中用于确定由表7a--7h中所述的本公开的fmc63基sir识别的靶标表位。在一些实施方式中,用于交叉竞争研究中确定由表7a--7h中所述的本公开的sir识别的靶标表位的参考vhh片段是具有如seqidno:2701至2725和12279-12294(表5)所示的序列的vhh片段。在一些实施方式中,用于交叉竞争研究中用于交叉竞争研究中确定由表7a--7h中所述的本公开的sir识别的靶标表位的参考非免疫球蛋白抗原结合支架是具有如seqidno:2728至2732和12296-12301(表6a)所示的序列的非免疫球蛋白抗原结合支架。在一些实施方式中,用于交叉竞争研究中确定由表7a--7h中所述的本公开的sir识别的靶标表位的参考配体是具有如seqidno:2758至2768和12359-12361(表6a)所示的序列的配体。在一些实施方式中,用于针对靶向不同靶标的sir的交叉竞争研究的参考sir是具有如seqidno:3435-3634、13184-13292(表7d)和seqidno:3855-4051和13411-13526(表7e)所示的序列的sir。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向mpl的sir(例如,seqidno:3566-3562、13259和13265-13266)所识别的靶标表位的参考scfv是表6b中所示的对应scfv(例如,seqidno:2871-2878、12318、12326-12327)。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向mpl的sir识别的靶标表位的参考scfv由seqidno:2871-2874表示。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向mpl的sir识别的靶标表位的参考配体是表6a中所示的对应配体(例如,seqidno:2758-2759)。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向mpl的sir识别的靶标表位的参考sir是表7d和7e中所示的mpl-sir(例如,seqidno:3566-3562、13259和13265-13266)。在另一个实施方式中,本公开的靶向mpl的sir结合至与肽序列–pwqdgpk-(seqidno:15784)相对应或重叠的mpl-表位。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd19的sir(例如,seqidno:3645-3649、13195-13203、13249和13267)所识别的靶标表位的参考scfv是表6b中所示的对应scfv(例如,seqidno:2770-2774、12308、12325、18162-18170)。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd19的sir识别的靶标表位的参考scfv由seqidno:2771、2772、12308和18169表示。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd19的sir识别的靶标表位的参考sir是表7a--7h中所示的靶向cd19的sir(例如,seqidno:3645-3649、13195-13203、13249和13267)。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd20的sir(例如,seqidno:3456-3457、13204-13213)所识别的靶标表位的参考scfv是表6b中所示的对应scfv(例如,seqidno:2787-2788、18177-18187)。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd20的sir识别的靶标表位的参考scfv由seqidno:18182、18185和2787表示。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd20的sir识别的靶标表位的参考sir是表7a--7h中所示的cd20-sir(例如,seqidno:3456-3457、13204-13213)。在优选的实施方式中,本公开的靶向cd20的sir结合至与以下一个或多个序列相对应的表位::–pagiyapi–(seqidno:18902)、–flkmeslnfirahtp–(seqidno:18903)、–hflkmeslnfirahtpy–(seqidno:18904)、–ynaepanpseknspstqy–(seqidno:18905)、–ynaepanpseknspst–(seqidno:18906)和–yncepanpseknsp–(seqidno:18907)。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向bcma的sir(例如,seqidno:3446-3449、3632-3634、13277-13284)所识别的靶标表位的参考scfv是表6b中所示的对应scfv(例如,seqidno:2780-2783、12337-12344和18174-18176)。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向bcma的sir识别的靶标表位的参考scfv由seqidno:2780-2781、18175-18176表示。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向bcma的sir识别的靶标表位的参考sir是表7a--7h中所示的bcma-sir(例如,seqidno:3446-3449、3632-3634、13277-13284)。在优选实施方式中,本公开的靶向bcma的sir结合至与seqidno:18908-18912中列出的一个或多个序列相对应的表位。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd22的sir(例如,seqidno:3458-3460、13241-13245、13268)所识别的靶标表位的参考scfv是表6b中所示的对应scfv(例如,seqidno:2789-2791、12320-12324、12330、18188)。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd22的sir识别的靶标表位的参考scfv由seqidno:18188、12330和12320表示。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd22的sir识别的靶标表位的参考sir是表7a--7h中所示的cd22-sir(例如,seqidno:3458-3460、13241-13245、13268)。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd123的sir(例如,seqidno:2929、3470、13184-13194)所识别的靶标表位的参考scfv是表6b中所示的对应scfv(例如,seqidno:2801、18196-18206)。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd123的sir识别的靶标表位的参考scfv由seqidno:2929、18196、18197、18200、18202和18205表示。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd123的sir识别的靶标表位的参考sir是表7a--7h中所示的cd123-sir(例如,seqidno:3470、13184-13194)。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd33的sir(例如,seqidno:3464-3465、13214-13220)所识别的靶标表位的参考scfv是表6b中所示的对应scfv(例如,seqidno:2795-2796、18189-18194)。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd33的sir识别的靶标表位的参考scfv由seqidno:2795、2796和18127表示。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd33的sir识别的靶标表位的参考sir是表7a--7h中所示的cd33-sir(例如,seqidno:3464-3465、13214-13220)。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cs1的sir(例如,seqidno:3487-3489、13226-1323)所识别的靶标表位的参考scfv是表6b中所示的对应scfv(例如,seqidno:2817-2819、18211-18216)。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cs1的sir识别的靶标表位的参考scfv由seqidno:2818、18212、18213、18215和18216表示。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cs1的sir识别的靶标表位的参考sir是表7a--7h中所示的cs1-sir(例如,seqidno:3487-3489、13226-1323)。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cll1的sir(例如,seqidno:3484-3485、13222-13225)所识别的靶标表位的参考scfv是表6b中所示的对应scfv(例如,seqidno:2814-2815、18207-18210、12345-12346)。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cll1的sir识别的靶标表位的参考sir是表7a--7h中所示的cll1-sir(例如,seqidno:3484-3485、13222-13225)。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向间皮素的sir(例如,seqidno:3554、13287-13288)所识别的靶标表位的参考scfv是表6b中所示的对应scfv(例如,seqidno:2870、12352-12353)。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向间皮素的sir识别的靶标表位的参考sir是表7a--7h中所示的间皮素-sir(例如,seqidno:3554、13287-13288)。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向bst1/cd157的sir识别的靶标表位的参考scfv是表6b中所示的对应配体。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向bst1/cd157的sir识别的靶标表位的参考sir是表7a--7h中所示的bst1/cd157-sir。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向dll3的sir识别的靶标表位的参考scfv是表6b中所示的对应scfv。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向dll3的sir识别的靶标表位的参考sir是表7a--7h中所示的dll3-sir。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向ptk7的sir识别的靶标表位的参考scfv是表6b中所示的对应scfv。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向ptk7的sir识别的靶标表位的参考sir是表7a--7h中所示的ptk7-sir。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向il13ra2的sir识别的靶标表位的参考scfv是表6b中所示的对应scfv。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向il13ra2的sir识别的靶标表位的参考sir是表7a--7h中所示的il13ra2-sir。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向ror1的sir识别的靶标表位的参考scfv是表6b中所示的对应scfv。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向ror1的sir识别的靶标表位的参考sir是表7a--7h中所示的ror1-sir。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向tcrgd的sir识别的靶标表位的参考scfv是表6b中所示的对应scfv。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向tcrgd的sir识别的靶标表位的参考sir是表7a--7h中所示的tcrgd-sir。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向tcrb1的sir识别的靶标表位的参考scfv是表6b中所示的对应scfv。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向tcrb1的sir识别的靶标表位的参考sir是表7a--7h中所示的tcrb1-sir。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向tcrb2的sir识别的靶标表位的参考scfv是表6b中所示的对应scfv。在另一个实施方式中,用于交叉竞争研究中确定由本公开的靶向tcrb2的sir识别的靶标表位的参考sir是表7a--7h中所示的tcrb2-sir。在一些实施方式中,靶向gp100、mart、酪氨酸酶、htert、muc1、cmv-pp65、htlv1-tax、hiv1-gag、ny-eso、wt1、afp、hpv-16-e7、pr1和rasg12v的sir结合至表7i中所示的与mhci类(例如,hla-a201)复合的靶向肽。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向afp/mhci的sir识别的靶标表位的参考scfv由seqidno:18171和18173表示。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向wt1/mhci的sir识别的靶标表位的参考scfv由seqidno:2926-2928表示。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向alk的sir识别的靶标表位的参考scfv由seqidno:2777表示。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向b7h4的sir识别的靶标表位的参考scfv由seqidno:2934-2935表示。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd30的sir识别的靶标表位的参考scfv由seqidno:2792-2793表示。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向cd138的sir识别的靶标表位的参考scfv由seqidno:2802表示。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向egfrviii的sir识别的靶标表位的参考scfv由seqidno:2826表示。在一个实施方式中,用于交叉竞争研究中确定由本公开的靶向fr1(叶酸受体1)的sir识别的靶标表位的参考scfv由seqidno:2833表示。在另一个实施方式中,用于交叉竞争性研究中确定由本公开的靶向trop2、lamp1、cdh19、cdh17、cd70、cd79b、cdh6、tshr、alk、wt1/mhc1、ny-eso-1/mhci、hiv1envgp、nybr1、lym1、lym2、tslrp、叶酸受体α、b7h4、cd200r、igk-轻链、cd179a、cd179b、cripto、steap1、hliv1、ilrap、粘连蛋白-4、gpa33、psca、psma、muc1/mhci、gfra4、egfrviii、egfr、her2、csf2ra、clec5a、gprc5d、tn-muc1、flt3、pr1/mhci、afp/mhci和hpv16-e7/mhci的sir所识别的靶标表位的参考scfv是表6b中列出的对应scfv。在另一个实施方式中,用于交叉竞争性研究中确定由本公开的靶向trop2、lamp1、cdh19、cdh17、cd70、cd79b、cdh6、tshr、alk、wt1/mhc1、ny-eso-1/mhci、hiv1包被糖蛋白、nybr1、lym1、lym2、tslrp、叶酸受体α、b7h4、cd200r、igk-轻链、cd179a、cd179b、cripto、steap1、hliv1、ilrap、粘连蛋白-4、gpa33、psca、psma、muc1/mhci、gfra4、egfrviii、egfr、her2、csf2ra、clec5a、gprc5d、tn-muc1、flt3、pr1/mhci、afp/mhci和hpv16-e7/mhci的sir识别的靶标表位的参考sir是表7a--7h中列出的对应sir。表6a:表6b:表6c:用于生成sir的mhci(hla-a2)限制性肽蛋白质片段名称缩氨酸seqseqidgp100g9-209imdqvpfsv15764gp100g9-280ylepgpvtv15765gp100g9-154ktwgqywqv15766muc1-a7(130-138)a7nltisdvsv15767muc1-d6(13-21)d6llltvltvv15768tax(11-19)llfgypvyv15769htert(540-548)t540ilakflhwl15770htert(865-873)t865rlvddfllv15771hiv1gag(77-85)sl9slyntvatl15772cmv-pp65(495-503)nlvpmvatv15773mart(26-35)eaagigiltv15774ebna-3a(596-604)svrdrlarl15775ebna-3clldfvrfmgv15776wt1rmfpnapyl15777pr1vlqelnvtv15778rasras9-g12vlvwgavgv15779hpvhpv16-e7ymldlqpet15780ny-eso-1ny-eso-1-(155-163)qlsllmwit15781ny-eso-1ny-eso-1-(157-165)sllmwitqc15782ny-eso-1ny-eso-1-(157-167)sllmwitqcfl15783表6d:用于生成sir的示例性接头在本公开的一个实施方式中,sir构建体包含scfv结构域,其中scfv可以在任选前导序列诸如seqidno:2300、2301或2302中任一个所提供的序列后面,并且之后为任选的接头序列诸如seqidno:2981-2986中任一个所提供的序列以及t细胞受体恒定链诸如seqidno:3010至3020、seqidno:3022至3044、seqidno:3045至3052中任一个所提供的恒定链(包括如本文描述的突变体和变体),其中这些结构域邻接并处于同一阅读框中以形成单一融合蛋白。接头序列可以存在或不存在于sir构建体中。在一个实施方式中,sir含有两个功能多肽单元,在此情况下两个单元由可裂解接头诸如seqidno:3060至3064中所提供的接头分开。可裂解接头可以在短柔性接头(例如sgsg)诸如seqidno:3065和弗林蛋白酶裂解位点(例如rakr)诸如seqidno:3066后面。sir的两个功能多肽单元也可以由通过ires序列分开的两个不同多核苷酸编码。或者,编码sir的两个功能多肽单元的两个不同多核苷酸可以由两个不同载体编码。在一个实施方式中,示例性sir构建体包含前导序列(例如,本文所述的前导序列)、细胞外抗原结合结构域(例如,本文所述的抗原结合结构域)、任选接头(例如,本文所述的接头区)和t细胞受体恒定链(例如,本文所述的t细胞受体恒定链,包括突变体和变体)。例如,本公开的sir包含选自seqidno:2300、2301和seqidno:2302且连接至抗原结合结构域诸如表5和表6a--b中识别的任何抗原结合结构域的示例性前导序列;选自seqidno:2981至2985和2986(还参见表6d)且连接至包含选自由seqidno:3010至3020、3022至3044、3045至3051和3052组成的组中的序列的t细胞受体恒定链结构域的任选接头序列(以及其如本文描述的突变体和变体)。sir可还包含t细胞受体恒定链的细胞外结构域和cd3z(诸如seqidno:3021和seqidno:3045中提供的)的细胞外跨膜细胞溶质结构域的融合物以及突变体和变体。在本文所述的任一实施方式中,当在类似条件下比较时,表达sir的效应细胞与对应表达ctcr的效应细胞(诸如在其表面上呈递包含sir的抗原结合结构域的ctcr,诸如包含含有sir的抗原结合结构域的scfv、vl和/或vh片段的ctcr的效应细胞)相比显示更高的对靶抗原的结合。示例性靶向cd19的sir由cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1200)表示。对应靶向cd19的ctcr由cd8sp-fmc63-vl-[htcrb-wt]-f-p2a-sp-fmc63-vh-[htcra-wt]-f-f2a-pac(seqidno:18280)表示。本公开的若干示例性ctcr的核苷酸和氨基酸seqidno提供于表7a--7b中。例如,在一些实施方式中,靶向cd19的表达sir的效应细胞在类似条件下与对应表达ctcr的效应细胞相比,在4℃孵育60分钟之后具有更高的对cd19-ecd-ggsg-nluc-acv5融合蛋白的结合。在一些实施方式中,表达sir的效应细胞在类似条件下与对应表达ctcr的效应细胞相比具有更高的对靶抗原的特异性结合。靶抗原对表达sir的效应细胞的特异性结合通过从使用表达sir的效应细胞获得的结合值中减去使用对照效应细胞获得的值来测量。在一些实施方式中,对照效应细胞是不表达任何sir的亲代效应细胞,例如未转导t细胞。在一个替代性实施方式中,对照效应细胞是表达靶向除由测试sir靶向的抗原之外的抗原的对照sir。比较cd19-sir的结合活性的示例性对照sir是cd8sp-mpl-161-vl-v5-[htcrb-kaciah]-f-p2a-sp-161-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1322)。在一些实施方式中,表达sir的效应细胞与靶抗原在4℃下孵育60分钟之后的结合比对应表达ctcr的效应细胞的结合大至少5%、10%、20%、30%、40%、50%或100%。在一些实施方式中,表达sir的效应细胞与靶抗原的结合比对应表达ctcr的效应细胞的结合大至少1.25倍(例如,1.5倍、2倍、5倍或10倍)。在一些实施方式中,表达sir的效应细胞与靶抗原的结合比对应表达ctcr的效应细胞的结合高不超过100000倍(例如,5000倍、10000倍或50000倍)。在一些实施方式中,表达sir的效应细胞与靶抗原的结合比与对应表达ctcr的效应细胞的结合高约1.25倍至约100000倍(例如,约5倍至约50000倍、约10倍至约10000倍、或约100倍至约1000倍、以及任一前述范围之间的任何值)。在一些实施方式中,在类似条件下,表达sir的效应细胞与靶抗原在4℃下孵育60分钟之后的结合比对应表达ctcr的效应细胞的结合大至少1.25倍至小于约100000倍。在一些实施方式中,该表达sir的效应细胞是sirt细胞。在一些实施方式中,该表达sir的效应细胞是表达sir的jurkatt细胞。在本文所述的其它实施方式中,当在类似条件下比较时,表达sir的效应细胞与对应表达car的效应细胞(诸如在其表面上呈递包含sir的抗原结合结构域的car,诸如包含含有sir的抗原结合结构域的scfv的car的效应细胞)相比显示更低的对靶抗原的结合。示例性靶向cd19的sir由cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1200)表示。对应靶向cd19的car由cd8sp-fmc63(vl-vh)-myc-bbz-t2a-pac(seqidno:4501)表示。本文提供了若干示例性car的核苷酸和氨基酸seqidno。例如,在一些实施方式中,靶向cd19的表达sir的效应细胞在类似条件下与对应表达car的效应细胞相比,具有更低的对cd19-ecd-ggsg-nluc-acv5的结合。在一些实施方式中,在类似条件下,表达sir的效应细胞与靶抗原在4℃下孵育60分钟之后的结合比对应表达car的效应细胞的结合小至少5%(例如,10%、20%、30%、40%或50%、或任何前述整数之间的任何值)。在一些实施方式中,表达sir的效应细胞与靶抗原的结合比对应表达car的效应细胞的结合小至少约1.5倍(例如,2倍、5倍、10倍、20-100倍、100-500倍、500-1000倍、1000-10000倍、10000-50000倍或50000-100000倍、或其之间的任何整数)。在一些实施方式中,在类似条件下,表达sir的效应细胞与靶抗原在4℃下孵育60分钟之后的结合比对应表达car的效应细胞的结合小至少1.25倍至100000倍(或其之间的任何值)。在一些实施方式中,该表达sir的效应细胞是sirt细胞。在一些实施方式中,该表达sir的效应细胞是表达sir的jurkatt细胞。在本文所述的其它实施方式中,当在类似条件下比较时,表达sir的效应细胞与对应表达ctcr的效应细胞相比显示更高的对靶抗原的结合,但与对应表达car的效应细胞相比显示更低的对靶抗原的结合。例如,在一些实施方式中,在类似条件下,靶向cd19的表达sir的效应细胞与对应表达ctcr的效应细胞相比具有更高的对cd19-ecd-ggsg-nluc-acv5的结合,但与对应表达car的效应细胞相比具有更低的结合。在一些实施方式中,在类似条件下,表达sir的效应细胞与靶抗原在4℃下孵育60分钟之后的结合比对应表达ctcr的效应细胞的结合大至少5%(例如,10%、20%、30%、40%、50%或100%、或其之间的任何值),但比对应表达car的效应细胞的结合小至少5%(例如,10%、20%、30%、40%、50%、60%、70%、80%或90%、或其之间的任何值)。在一些实施方式中,表达sir的效应细胞与靶抗原在4℃下孵育60分钟之后的结合在类似条件下比对应表达ctcr的效应细胞的结合大至少1.25倍(例如,1.5倍、2倍、5倍或10倍、或其之间的任何值),但比对应表达car的效应细胞的结合小至少1.5倍(例如,2倍、5倍或10倍、或其之间的任何值)。在一些实施方式中,该表达sir的效应细胞是sirt细胞。在一些实施方式中,该表达sir的效应细胞是表达sir的jurkatt细胞。在本文所述的其它实施方式中,sir与对应ctcr相比当在效应细胞中表达并在类似条件下比较时显示更高的细胞表面表达。示例性靶向cd19的sir由cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1200)表示。对应靶向cd19的ctcr由cd8sp-fmc63-vl-[htcrb-wt]-f-p2a-sp-fmc63-vh-[htcra-wt]-f-f2a-pac(seqidno:18280)表示。例如,在一些实施方式中,当在类似条件下检查时,sir的细胞表面表达(如通过与apl缀合的蛋白质l的结合测量的)高于对应ctcr的细胞表面表达。在一些实施方式中,表达sir的效应细胞与对应表达ctcr的效应细胞相比在孵育60分钟之后具有大于约5%(直到大于约10%、15%、20%、25%、30%、35%、40%、45%或50%中的任一者,包括这些值之间的任何范围)的与apc缀合蛋白质l的结合。在一些实施方式中,该表达sir的效应细胞是sirt细胞。sir和对应ctcr在效应细胞表面上的表达可以通过替代性方法测量,包括与cd8sp-蛋白质l-ggsg-nluc-4xflag-x2strep-8xhis融合蛋白的结合或使用插入在sir和ctcr的细胞外结构域内的表位标签(例如myc标签)的染色。在本文所述的其它实施方式中,sir与对应car相比当在效应细胞中表达并在类似条件下比较时显示更低的细胞表面表达。示例性靶向cd19的sir由cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1200)表示。对应靶向cd19的car由cd8sp-fmc63(vl-vh)-myc-bbz-t2a-pac(seqidno:4501)表示。例如,在一些实施方式中,当在类似条件下检查时,sir的细胞表面表达(如通过与apl缀合的蛋白质l的结合测量的)低于对应car的细胞表面表达。在一些实施方式中,在类似条件下,表达sir的效应细胞与apc-蛋白质l的结合比对应表达car的效应细胞的结合小至少5%(例如,10%、20%、30%、40%或50%、或其之间的任何值)。在一些实施方式中,表达sir的效应细胞与apc-蛋白质l的结合比对应表达car的效应细胞的结合小至少1.5倍至约1000倍(或其之间的任何值)。在一个实施方式中,表达sir的效应细胞与apc-蛋白质l的结合比对应表达car的效应细胞的结合小至少2倍至约100倍(或其之间的任何值)。在一些实施方式中,该表达sir的效应细胞是sirt细胞。在一些实施方式中,该表达sir的效应细胞是表达sir的jurkatt细胞。sir和对应car在效应细胞表面上的表达可以通过替代性方法测量,包括与cd8sp-蛋白质l-ggsg-nluc-4xflag-x2strep-8xhis融合蛋白的结合或使用插入在sir和car的细胞外结构域内的可比较位置(例如n末端区域)中的表位标签(例如myc标签)的染色。在本文所述的一些此类实施方式的任一实施方式中,当在效应细胞中表达并在类似条件下比较时,sir与对应ctcr相比显示更高的细胞表面表达,但与对应car相比显示更低表达。例如,在一些实施方式中,当在类似条件下检查时,sir的细胞表面表达(如通过与apl缀合的蛋白质l的结合测量的)高于对应ctcr的细胞表面表达,但低于对应car的细胞表面表达。在一些实施方式中,表达sir的效应细胞与对应表达car的效应细胞相比在孵育60分钟之后具有大于约5%(诸如大于约5%、10%、15%、20%、25%、30%、35%、40%、或45%,包括这些值之间的任何范围)的与apc缀合蛋白l的结合,但与对应表达car的效应细胞相比在孵育60分钟之后具有小于约50%(诸如小于约45%、40%、35%、30%、25%、20%、15%、10%、5%、4%、3%、2%、1%,包括这些值之间的任何范围)的与apc缀合蛋白l的结合。在一些实施方式中,该表达sir的效应细胞是sirt细胞。在本文所述的其它实施方式中,当在类似条件下比较时,表达sir的效应细胞与对应表达ctcr的效应细胞(诸如在其表面上呈递包含sir的抗原结合结构域的ctcr,诸如包含含有sir的抗原结合结构域的scfv、vl和/或vh片段的ctcr的效应细胞)相比显示更高的对表达靶抗原的细胞的细胞毒性。示例性靶向cd19的sir由cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1200)表示。对应靶向cd19的ctcr由cd8sp-fmc63-vl-[htcrb-wt]-f-p2a-sp-fmc63-vh-[htcra-wt]-f-f2a-pac(seqidno:18280)表示。本公开的若干示例性sir和ctcr的核苷酸和氨基酸seqidno提供于表7a和7d中。例如,在一些实施方式中,靶向cd19的表达sir的效应细胞在类似条件下与对应表达ctcr的效应细胞相比,在37℃孵育4小时至96小时之后显示更高的针对raji-gluc细胞的细胞毒性。在一些实施方式中,表达sir的效应细胞在类似条件下与对应表达ctcr的效应细胞相比具有更高的针对表达其靶抗原的细胞的特异性细胞毒性。表达sir的效应细胞的特异性细胞毒性通过从使用表达sir的效应细胞获得的细胞毒性值中减去使用对照效应细胞获得的值来测量。在一些实施方式中,对照效应细胞是不表达任何sir的亲代效应细胞,例如未转导t细胞。在一个替代性实施方式中,对照效应细胞是表达靶向除测试sir之外的抗原的对照sir。例如,比较cd19sir的细胞毒性活性的示例性对照sir是cd8sp-mpl-161-vl-v5-[htcrb-kaciah]-f-p2a-sp-161-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1322)。在一些实施方式中,表达sir的效应细胞对表达靶抗原的细胞(即靶细胞)的细胞毒性在37℃下共同培养4小时至96小时之后比对应表达ctcr的效应细胞的细胞毒性大至少5%、10%、20%、30%、40%、50%或100%。在一些实施方式中,表达sir的效应细胞对靶细胞的细胞毒性比对应表达ctcr的效应细胞的细胞毒性大至少1.25倍、1.5倍、2倍、5倍或10倍。在一些实施方式中,该表达sir的效应细胞是sirt细胞。在本文所述的任一前述实施方式的另一个或其它实施方式中,当在类似条件下比较时,表达sir的效应细胞与对应表达ctcr的效应细胞(诸如在其表面上呈递包含sir的抗原结合结构域的ctcr,诸如包含含有sir的抗原结合结构域的scfv、vl和/或vh片段的ctcr的效应细胞)相比显示更高的针对表达靶抗原的细胞的体内活性。示例性靶向cd19的sir由cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1200)表示。对应靶向cd19的ctcr由cd8sp-fmc63-vl-[htcrb-wt]-f-p2a-sp-fmc63-vh-[htcra-wt]-f-f2a-pac(seqidno:18280)表示。本公开的若干示例性sir和ctcr的核苷酸和氨基酸seqidno提供于表7a和7d中。例如,在一些实施方式中,靶向cd19的表达sir的效应细胞在类似条件下与对应表达ctcr的效应细胞相比,在nsg小鼠异种移植物模型中显示更高的针对raji-fluc细胞的体内活性。在一些实施方式中,表达sir的效应细胞在类似条件下与对应表达ctcr的效应细胞相比具有更高的针对表达其靶抗原的细胞的体内活性。表达sir的效应细胞的特异性体内活性通过从使用表达sir的效应细胞获得的体内活性减去使用对照效应细胞获得的值(例如从表达fluc的肿瘤中获得的肿瘤减小或生物荧光值减小)来测量。在一些实施方式中,对照效应细胞是不表达任何sir的亲代效应细胞,例如未转导t细胞。在一个替代性实施方式中,对照效应细胞是表达靶向除由测试sir靶向的抗原之外的抗原的对照sir。例如,比较cd19sir的体内活性的示例性对照sir是cd8sp-mpl-161-vl-v5-[htcrb-kaciah]-f-p2a-sp-161-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1322)。在一些实施方式中,表达sir的效应细胞对表达靶抗原的细胞(即靶细胞)的体内活性在nsg小鼠异种移植物模型中比对应表达ctcr的效应细胞的体内活性大至少5%、10%、20%、30%、40%、50%或100%。在一些实施方式中,表达sir的效应细胞对靶细胞的体内活性比对应表达ctcr的效应细胞的体内活性大至少1.25倍、1.5倍、2倍、5倍或10倍。在一些实施方式中,该表达sir的效应细胞是sirt细胞。在一些实施方式中,表达sir的效应细胞的体内活性通过其它方法,诸如存活改善或使用卡尺测量的肿瘤体积减小来测量。在本文所述的任一前述实施方式的另一个或其它实施方式中,当在类似条件下比较时,表达sir的效应细胞与对应表达ctcr的效应细胞(诸如在其表面上呈递包含sir的抗原结合结构域的ctcr,诸如包含含有sir的抗原结合结构域的scfv、vl和/或vh片段的ctcr的效应细胞)相比当与其靶细胞(例如,表达其靶抗原的细胞)共同培养时显示更高的tnfα产生。示例性靶向cd19的sir由cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1200)表示。对应靶向cd19的ctcr由cd8sp-fmc63-vl-[htcrb-wt]-f-p2a-sp-fmc63-vh-[htcra-wt]-f-f2a-pac(seqidno:18280)表示。靶向不同抗原的若干示例性sir和ctcr的核苷酸和氨基酸seqidno提供于表7a和7d中。例如,在一些实施方式中,靶向cd19的表达sir的效应细胞在类似条件下与对应表达ctcr的效应细胞相比,当与nalm6靶细胞在37℃下共同培养4小时至96小时时具有更高的tnfα产生,如通过elisa所测量。在一些实施方式中,表达sir的效应细胞在类似条件下与对应表达ctcr的效应细胞相比具有更高的成倍诱导的tnfα产生。表达sir的效应细胞的成倍诱导的tnfα产生通过从在单独培养表达sir的效应细胞时获得的tnfα值除以在表达sir的细胞与其靶细胞共同培养时获得的tnfα水平来测量。在一些实施方式中,表达sir的效应细胞在类似条件下与对应表达ctcr的效应细胞相比当与其靶细胞共同培养时具有更高的特异性tnfα产生。表达sir的效应细胞在暴露于靶细胞时的特异性tnfα产生当在类似条件下与靶细胞在37℃下共同培养4小时至96小时时通过从使用表达sir的效应细胞获得的tnfα值减去使用对照效应细胞获得的值来测量。在一些实施方式中,对照效应细胞是不表达任何sir的亲代效应细胞,例如未转导t细胞。在一个替代性实施方式中,对照效应细胞是表达靶向除由测试sir靶向的抗原之外的抗原的对照sir。比较cd19sir的结合活性的示例性对照sir是cd8sp-mpl-161-vl-v5-[htcrb-kaciah]-f-p2a-sp-161-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1322)。在一些实施方式中,表达sir的效应细胞对靶抗原的特异性tnfα产生在37℃下孵育24小时之后比对应表达ctcr的效应细胞的特异性tnfα产生大至少5%、10%、20%、30%、40%、50%或100%。在一些实施方式中,表达sir的效应细胞的特异性tnfα产生比对应表达ctcr的效应细胞的特异性tnfα产生大至少1.25倍、1.5倍、2倍、5倍或10倍。在一些实施方式中,表达sir的效应细胞的特异性tnfα产生比对应表达ctcr的效应细胞的特异性tnfα产生大了小于100000倍。在一些实施方式中,表达sir的效应细胞的特异性tnfα产生在类似条件下比对应表达ctcr的效应细胞的特异性tnfα产生大至少1.25倍、1.5倍、2倍、或5倍或10倍,但在类似条件下比对应表达ctcr的效应细胞的特异性tnfα产生大小于100000倍(例如,小于50000倍、10000倍、或1000倍)。在一些实施方式中,该表达sir的效应细胞是sirt细胞。在本文所述的任一前述实施方式的另一个或其它实施方式中,当在类似条件下比较时,表达sir的效应细胞与对应表达ctcr的效应细胞(诸如在其表面上呈递包含sir的抗原结合结构域的ctcr,诸如包含含有sir的抗原结合结构域的scfv、vl和/或vh片段的ctcr的效应细胞)相比当与其靶细胞(例如,表达其靶抗原的细胞)共同培养时显示更高的il2产生。示例性靶向cd19的sir由cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1200)表示。对应靶向cd19的ctcr由cd8sp-fmc63-vl-[htcrb-wt]-f-p2a-sp-fmc63-vh-[htcra-wt]-f-f2a-pac(seqidno:18280)表示。靶向不同抗原的若干示例性sir和ctcr的核苷酸和氨基酸seqidno提供于表7a和7d中。例如,在一些实施方式中,靶向cd19的表达sir的效应细胞在类似条件下与对应表达ctcr的效应细胞相比,当与nalm6靶细胞在37℃下共同培养4小时至96小时时具有更高的il2产生,如通过elisa所测量。在一些实施方式中,表达sir的效应细胞在类似条件下与对应表达ctcr的效应细胞相比具有更高的成倍诱导的il2产生。表达sir的效应细胞的成倍诱导的il2产生通过从在单独培养表达sir的效应细胞时获得的il2值除以在表达sir的细胞与其靶细胞共同培养时获得的il2水平来测量。在一些实施方式中,表达sir的效应细胞在类似条件下与对应表达ctcr的效应细胞相比当与其靶细胞共同培养时具有更高的特异性il2产生。表达sir的效应细胞在暴露于靶细胞时的特异性il2产生当在类似条件下与靶细胞在37℃下共同培养4小时至96小时时通过从使用表达sir的效应细胞获得的il2值减去使用对照效应细胞获得的值来测量。在一些实施方式中,对照效应细胞是不表达任何sir的亲代效应细胞,例如未转导t细胞。在一个替代性实施方式中,对照效应细胞是表达靶向除由测试sir靶向的抗原之外的抗原的对照sir。比较cd19sir的结合活性的示例性对照sir是cd8sp-mpl-161-vl-v5-[htcrb-kaciah]-f-p2a-sp-161-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1322)。在一些实施方式中,表达sir的效应细胞对靶抗原的特异性il2产生在37℃下孵育24小时之后比对应表达ctcr的效应细胞的特异性il2产生大至少5%、10%、20%、30%、40%、50%或100%。在一些实施方式中,表达sir的效应细胞的特异性il2产生比对应表达ctcr的效应细胞的特异性il2产生大至少1.25倍、1.5倍、2倍、5倍或10倍。在一些实施方式中,表达sir的效应细胞的特异性il2产生比对应表达ctcr的效应细胞的特异性il2产生大了小于100000倍。在一些实施方式中,表达sir的效应细胞的特异性il2产生在类似条件下比对应表达ctcr的效应细胞的特异性il2产生大至少1.25倍、1.5倍、2倍、或5倍或10倍,但在类似条件下比对应表达ctcr的效应细胞的特异性il2产生大小于100000倍(例如,小于50000倍、10000倍、或1000倍)。在一些实施方式中,该表达sir的效应细胞是sirt细胞。在一些实施方式中,该表达sir的效应细胞是表达sir的jurkatt细胞。在本文所述的任一前述实施方式的另一个或其它实施方式中,当在类似条件下比较时,表达sir的效应细胞与对应表达car的效应细胞(诸如在其表面上呈递包含sir的抗原结合结构域的car,诸如包含含有sir的抗原结合结构域的scfv、vl和/或vh片段的car的效应细胞)相比当与其靶细胞(例如,表达其靶抗原的细胞)共同培养时显示更低的tnfα和/或il2产生。示例性靶向cd19的sir由cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(seqidno:1200)表示。例如,在一些实施方式中,靶向cd19的表达sir的效应细胞在类似条件下与对应表达ctcr的效应细胞相比,当与nalm6靶细胞在37℃下共同培养4小时至96小时时具有更高的tnfα和/或il2产生,如通过elisa所测量,但在类似条件下与对应表达car的效应细胞相比具有更低更高的tnfα和/或il2产生。在一些实施方式中,在类似条件下,表达sir的效应细胞与对应表达ctcr的效应细胞相比具有更高的成倍诱导的tnfα和/或il2产生,但与对应表达car的效应细胞相比具有更低的成倍诱导的tnfα和/或il2产生。在一些实施方式中,在类似条件下,表达sir的效应细胞与对应表达ctcr的效应细胞相比当与其靶细胞共同培养时具有更高的特异性tnfα和/或il2产生,但与表达car的效应细胞相比具有更低的特异性tnfα和/或il2产生。在一些实施方式中,表达sir的效应细胞对靶抗原的特异性tnfα和/或il2产生在37℃下孵育24小时之后比对应表达ctcr的效应细胞的特异性tnfα和/或il2产生大至少5%、10%、20%、30%、40%、50%或100%,但比对应表达car的效应细胞的特异性tnfα和/或il2产生小至少5%、10%、20%、30%、40%、50%或100%。在一些实施方式中,在类似条件下,表达sir的效应细胞的特异性tnfα和/或il2产生比对应表达ctcr的效应细胞的特异性tnfα和/或il2产生大至少1.25倍、1.5倍、2倍、5倍或10倍,但比对应表达car的效应细胞的特异性tnfα和/或il2产生小至少1.25倍、1.5倍、2倍、5倍或10倍。在一些实施方式中,该表达sir的效应细胞是sirt细胞。在一些实施方式中,该表达sir的效应细胞是表达sir的jurkatt细胞。在本文所述的任一实施方式中,当在类似条件下比较时,表达一种类型的sir的效应细胞与表达不同类型的sir的效应细胞(诸如在其表面上呈递包含第一sir的抗原结合结构域但具有不同tcr链的sir,例如包含含有第一sir的抗原结合结构域但具有不同tcr链的scfv、vl和/或vh片段的sir的效应细胞)相比显示不同的特性。表7a-7c提供了不同类型的示例性sir的seqid。在sir在设计中为模块时,本领域技术人员可以通过将一个模块用另一个模块置换来生成额外sir类型。不同类型的sir当在免疫效应细胞中表达时可显示多样性的示例性特征包括但不限于结合亲和力、细胞表面表达、细胞毒性、细胞因子产生、细胞增殖、终末分化、耗尽和体内生物活性。在示例性实施方式中,当在类似条件下检查时并且当两种sir类型均靶向trac(tcrα恒定链)基因组基因座以排除由于不同sir构建体的无规整合位点而引起的任何表达变化时,表达含有基于fmc63的靶向cd19的结构域的sir1(seqidno:1200)的效应细胞与表达靶向cd19的sir2(seqidno:1410)或sir3(seqidno:4531)的对应效应细胞相比在4℃下孵育60分钟之后具有更高的与cd19-ecd-ggsg-nluc-acv5融合蛋白的结合。在一些实施方式中,当在类似条件下检查时并且当两种sir类型靶向trac(tcrα恒定链)基因组基因座时,表达一种类型的sir(例如sir1)的效应细胞的靶抗原结合在4℃下孵育60分钟之后比表达含有相同结合结构域的另一种类型的sir(例如sir2)的效应细胞的靶抗原结合大至少5%、10%、20%、30%、40%、50%或100%。在一些实施方式中,当在类似条件下检查时并且当两种sir类型靶向trac(tcrα恒定链)基因组基因座时,表达含有相同结合结构域的不同类型的sir(例如sir1、sir2、sir3等等)的效应细胞的靶抗原结合在4℃下孵育60分钟之后改变超过5倍、10倍、20倍、50倍或100倍。将基因组插入物靶向特异性基因组基因座的技术为本领域中已知的。在一些实施方式中,当在类似条件下检查时并且当两种sir类型靶向trac(tcrα恒定链)基因组基因座时,表达含有相同结合结构域的不同类型的sir(例如sir1、sir2、sir3等等)的效应细胞的靶抗原结合在4℃下孵育60分钟之后改变超过5倍、10倍、20倍、50倍或100倍。在一些实施方式中,当在类似条件下检查时并且当不同sir类型和ctcr靶向trac基因座时,表达含有相同结合结构域的不同类型的sir(例如sir1、sir2、sir3等等)的效应细胞的靶抗原结合的标准偏差与独立地分离的表达对应ctcr的效应细胞群体的靶抗原结合的标准偏差相比在4℃下孵育60分钟之后是超过2倍、5倍、10倍、20倍、50倍或100倍。在其它实施方式中,当每一种sir类型和ctcr插入在trac基因座处时,表达含有相同结合结构域的不同类型的sir(例如sir1、sir2、sir3等等)的效应细胞的细胞毒性的标准偏差与独立地分离的表达对应ctcr的效应细胞群体的细胞毒性的标准偏差相比在37℃下与靶细胞一起孵育4小时之后是超过2倍、5倍、10倍、20倍、50倍或100倍。标准偏差是方差的平方根并且可以通过本领域已知的方法测量。在一些实施方式中,该表达sir的效应细胞是sirt细胞。在一些实施方式中,该表达sir的效应细胞是表达sir的jurkatt细胞。在本文所述的任何或一些此类实施方式中,sir包含由人类密码子优化的多核苷酸编码的野生型和变体tcra(例如,seqidno:732-740)和tcrb(例如,seqidno:747-762)恒定链,而对应ctcr包含由野生型多核苷酸序列编码的野生型tcra(seqidno:730-731)和tcrb恒定链(seqidno:744-746)。在一些实施方式中,sir还含有将一个或多个抗原结合结构域连接至tcra和tcrb恒定链的任选接头。在一些实施方式中,sir包含由人类密码子优化的多核苷酸编码的野生型和变体pre-tcra(例如,seqidno:767-768)和tcrb恒定链(例如,seqidno:747-762),而对应ctcr包含由野生型多核苷酸序列编码的野生型tcra和tcrb恒定链。在一些实施方式中,sir还含有将一个或多个抗原结合结构域连接至pre-tcra和tcrb链的任选接头。在本文所述的任何或一些此类实施方式中,sir包含由人类密码子优化的多核苷酸编码的野生型和变体tcrg(例如,seqidno:770)和tcrd(例如,seqidno:772)恒定链,而对应ctcr包含由野生型多核苷酸序列编码的野生型tcrg(seqidno:769)和tcrd恒定链(seqidno:771)。在一些实施方式中,sir还含有将一个或多个抗原结合结构域连接至tcra和tcrb恒定链的任选接头。在一些实施方式中,sir包含野生型和变体htcrbecd-bam-cd3zecdtmcp(seqidno:10444-10452)和htcraecd-kpn-cd3zecdtmcp-opt2(seqidno:10464-10471)恒定链,而对应ctcr包含由野生型多核苷酸序列编码的野生型tcra和tcrb恒定链。在一些实施方式中,sir还含有将一个或多个抗原结合结构域连接至htcrbecd-bam-cd3zecdtmcp(seqidno:10444-10452)和htcraecd-kpn-cd3zecdtmcp-opt2(seqidno:10464-10471)恒定链的任选接头。cd19-ecd-ggsg-nluc-acv5在一些实施方式中,抗原结合结构域具有约10-4m至10-8m之间的解离常数(kd,反映了其结合亲和力)。在一些实施方式中,抗原结合结构域结合至一种或多种上文所述的抗原。在一些实施方式中,抗原结合结构域对于靶抗原具有的kd为在约10-4m至10-8m之间,例如在约10-5m至10-7m之间,例如在约10-5m至10-6m之间。在一个实施方式中,该抗原结合结构域的结合亲和力比参考抗体小至少五倍、10倍、20倍、30倍、50倍、100倍或1000倍。在一个实施方式中,编码的抗原结合结构域具有的结合亲和力比参考抗体小至少5倍。在一些实施方式中,参考抗体是抗原结合结构域所来源的抗体。在一些实施方式中,当存在于细胞表面上时,双链sir的所述第一链的抗原结合结构域与其同源抗原的结合基本上不会因sir的所述第二链的存在或car的存在而降低。在一些实施方式中,在sir(或car)的所述第二链存在下sir的所述第一链的抗原结合结构域与其同源抗原的结合是在与其同源抗原的sir(或car)的所述第二链不存在下sir的所述第一链的抗原结合结构域与其同源抗原的结合的70%、80%、85%、90%、95%、96%、97%、98%或99%。例如,如果细胞表达双链sir(其中第一链包含靶向cd19的连接至tcrα的scfv并且第二链包含靶向cd123的连接至tcrβ2的骆驼科vhh片段),那么在sir的所述第二链存在下sir的所述第一链的抗原结合结构域与其同源抗原(即cd19)的结合是在sir的所述第二链与其同源抗原(即cd123)不存在下sir的所述第一链的抗原结合结构域与其同源抗原(即cd19)的结合的70%、80%、85%、90%、95%、96%、97%、98%或99%。例如,如果细胞表达双链sir(其中第一链包含靶向cd19的连接至tcrα的fmc63抗体的vl片段并且第二链包含靶向cd19的连接至tcrβ2的fmc63抗体的vh片段)连同包含靶向cd123的vhh片段的car,那么在所述car存在下双链sir的所述第一链和所述第二链的抗原结合结构域与其同源抗原(即cd19)的结合是在所述car与其同源抗原(即cd123)不存在下sir双链的所述第一链和所述第二链的抗原结合结构域与其同源抗原(即cd19)的结合的70%、80%、85%、90%、95%、96%、97%、98%或99%。在一些实施方式中,当存在于细胞表面上时,双链sir的所述第一链和所述第二链的抗原结合结构域彼此的缔合小于两者都是scfv抗原结合结构域的情况下的缔合。在一些实施方式中,双链sir的所述第一链和所述第二链的抗原结合结构域彼此的缔合比两者都是scfv抗原结合结构域的情况下的缔合少5%、10%、20%、30%、40%、50%、60%、70%、80%、90%或99%。本文描述的sir功能多肽单元可以由单一多核苷酸链编码并且合成为单一多肽链,该多肽链随后裂解成不同多肽。最初可以合成sir多肽,这些多肽包含随后从成熟多肽移除的一个或多个前导序列(也称为信号肽)。在一个优选实施方式中,sir多肽的每个功能多肽单元(即,与t细胞受体恒定链同框连接的抗原结合结构域加上弗林蛋白酶-sgsg-可裂解接头或t细胞受体恒定链加上弗林蛋白酶-sgsg-可裂解接头)是在将sir的功能多肽单元作为i型跨膜蛋白引导至细胞表面的前导序列后面。在优选实施方式中,sir的抗原结合结构域是面向细胞外。在一些实施方式中,sir多肽的前导序列包含seqidno:2300至seqidno:2302的序列。在某些实施方式中,sir的两条t细胞受体恒定链可以具有相同类型(即tcra和tcra;tcrb和tcrb;pretcra和pretcra;tcrγ和tcrγ;以及tcr-δ和tcr-δ)。具有相同类型的两条tcr恒定链的一些示例性sir是克隆id:021116-e08(seqidno:905)、克隆id:012216-p08(seqidno:906)、克隆idno:012216-q05(seqidno:907)、克隆idno:012216-r04(seqidno:908)和克隆idno:012216-s02(seqidno:909)。在优选实施方式中,sir的两条t细胞受体恒定链具有不同类型(例如tcra和tcrb;pretcra和tcrb;tcrγ和tcr-δ等)。具有不同类型的两条tcr恒定链的示例性sir由克隆id:102615-c08表示,其核酸和氨基酸序列分别以seqidno:1200和seqidno:3435给出。在某些实施方式中,sir多肽包含含有或源自人类、小鼠或犬类来源的tcra、tcrb、pre-tcra、tcr-γ或tcr-δ链的单一t细胞受体恒定链。具有单一tcr恒定链的示例性sir由克隆id:051216-f04表示,其氨基酸序列是seqidno:3258。在某些实施方式中,本公开的sir多肽编码包含或源自人类、小鼠或犬类来源的tcra、tcrb、pre-tcra、tcr-γ或tcr-δ链的两条t细胞受体恒定链。具有两条tcr恒定链的示例性sir由克隆id:102615-c08表示,其核酸和氨基酸序列分别以seqidno:1200和seqidno:3435给出。在某些实施方式中,sir多肽的两条t细胞受体恒定链具有相同类型(即tcra和tcra;tcrb和tcrb;pretcra和pretcra;tcrγ和tcrγ;以及tcr-δ和tcr-δ)。具有相同类型的两条tcr恒定链的示例性sir是克隆id:021116-e08,其氨基酸序列以seqidno:3140给出。在优选实施方式中,sir多肽的两条t细胞受体恒定链具有不同类型(例如tcra和tcrb;pretcra和tcrb;tcrγ和tcr-δ等)。具有不同类型的两条tcr恒定链的示例性sir由克隆id:102615-c08表示,其核酸和氨基酸序列分别以seqidno:1200和seqidno:3435给出。在一些实施方式中,sir多肽的两条t细胞受体恒定链均不是野生型tcra或野生型tcrb或野生型tcrg或野生型tcrd或野生型pretcra。下表汇总了本公开中所述的若干种示例性sir的靶抗原、克隆id、seqid(dna)、seqid(prt)和名称。这些构建体通常通过将表5--6中所述的抗原结合片段与本文所述的tcr恒定链的示例性变体(包括表1--3中提供的变体)组合来制成。基于其骨架,即存在于其中的tcr恒定链的类型,将sir分成不同类型(例如sir1-sir18)。然而,应理解,sir在设计中为模块化的并且本公开的范围不限于下表中所述的sir并且可以通过转换不同模块来生成不同sir。因此,可以将抗原结合结构域与tcr恒定链的其它变体组合,但是在下表7中所述的sir中不包括它们。可以使用表5--6中未列出的抗原结合结构域设计sir。还可以将不同治疗和辅助模块添加到sir中或置换或移除这些模块。因此,虽然下表含有具有抗生素抗性基因(例如pac)的若干sir,但是可移除此模块。除了本公开的sir之外,表7也描述了含有cd3z初级信号传导结构域和41bb共刺激结构域的若干种嵌合抗原受体(car)。应理解,类似car可以使用其它初级和共刺激结构域(例如来自cd28)生成。这些car可以与本文所述的sir组合表达。在表7a--c中,sir类型是指在“示例性sir”下列出的构建体骨架,每个序列具有相同“骨架”但具有不同抗原结合结构域。表7a--c可以用于确定含有特定结合结构域并属于特定sir类型ctcr或car的构建体的dna和prtseqidno。因此,seqidno:1200具有fmc63结合结构域,而seqidno:1201具有相同sir骨架但具有hufmc63抗原结合结构域。表7d中列出了若干种示例性sir1型构建体的靶抗原、dna和prtseqidno以及名称(包括结合结构域)。表7a--b中呈现了在骨架sir2-9和ctcr上参考sir1型dna和prtseqidno的不同结合结构域的dna和prtseqidno的顺序。因此,通过使用表7a和7d,可以确定sir1-sir6型骨架和ctcr上任何抗原结合结构域的dna和prtseqidno。类似地,通过使用表7b和7d,可以确定sir7-sir9型骨架上任何抗原结合结构域的dna和prtseqidno。表7e中列出了若干种示例性sir10型构建体的靶抗原、dna和prtseqidno以及名称(包括结合结构域)。表7c中呈现了在骨架sir10-18和car上参考sir10型dna和prtseqidno的不同结合结构域的dna和prtseqidno的顺序。因此,通过使用表7e和7c,可以确定sir10-sir18型骨架和car上任何抗原结合结构域的dna和prtseqidno。或者,本公开的含有特定抗原结合结构域的sir的序列可以通过随附本公开的seq列表文件的同源搜索来确定。最后,由于sir在设计中是模块化的,所以含有特定模块的sir的dna和氨基酸序列可以通过用新模块简单取代存在于sir1和sir10型中的模块来生成。表7a--sir1--sir6和ctcr参考sir1的序列识别的指导表7b--sir1和sir7--9型的序列识别的指导,其中sir1用作参考。表7c--sir10--sir18型和car的序列识别的指导,其中sir10用作参考。表7d--靶向sir1型骨架上的不同抗原的sir表7e:靶向sir10型骨架上的不同抗原的sir表7f:靶向具有基于fmc63的结合结构域的cd19的示例性sir构建体表7g:靶向基于bu12结合结构域的cd19的示例性sir表7h:靶向不同抗原的不同类型的sir表7i--抗原luc融合构建体在另一方面,本公开提供一种分离的sir多肽分子,该分子包含结合至如本文描述的抗原并且连接至一个或多个t细胞受体恒定链的一个或多个抗原结合结构域(例如,抗体或抗体片段、配体或受体)。在一些实施方式中,sir可以包含含有连接至单一t细胞受体恒定链(i类)的nh2末端的单一抗原结合结构域的单一多肽或由该单一多肽组成。编码示例性1类sir的构建体提供于克隆idno:051216-f04中。编码sir的核酸序列呈现于seqidno:1023中并且编码sir的氨基酸序列对应于seqidno:3258。在一些实施方式中,sir包含组装形成功能性sir(2类)的两种多肽或由该两种多肽组成。此双链2类sir的每个多肽含有t细胞受体恒定链并且含有(如在2a类中)或不含有(如在2b类中)抗原结合结构域。在2a类sir中,每个抗原结合结构域连接至单独t细胞受体恒定链的n末端。例如,抗原结合结构域1(例如抗体的vl片段)连接至t细胞受体β(tcrβ)的恒定链以构造功能多肽单元1并且抗原结合结构域2(抗体的vh片段)连接至t细胞受体α(tcrα)的恒定链以构造功能多肽单元2。此类sir的两个功能多肽单元在同一细胞中共同表达并且彼此配对以变成具有功能活性。应注意,每个抗原结合结构域可进而是双特异性的或多特异性的,从而使得2类sir靶向多于2种抗原。靶向cd19的示例性2a类sir提供于cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(102615-c08)[seqidno:1200]中。此sir的核酸序列呈现于seqidno:1200中并且其氨基酸序列对应于seqidno:3435。在该sir中,源自cd19单克隆抗体fmc63的vl片段经由接头连接至突变体(kaciah)形式的人类tcrb链的恒定区,而源自fmc63单克隆抗体的vh片段经由接头连接至突变体(csdvp)人类tcrα链的恒定区。在一些实施方式中,多肽双链sir包含连接至仅一个t细胞受体恒定链(功能多肽单位1)的nh2末端但与第二t细胞受体恒定链共同表达的抗原结合结构域或由该抗原结合结构域组成。此类sir被指定为2b类。在此类2bsir中第二t细胞受体恒定链的目的在于促进功能多肽单元1(即,连接至t细胞受体恒定链的抗原结合结构域1)的细胞表面表达。这样,2b类sir中的第二t细胞受体恒定链可以自身表达或者表达为携带表位标签(例如myc、v5、acv5、g4sx2、streptagii等)的融合蛋白或者表达为携带任何不相关蛋白质片段(例如vl或vh片段)的融合蛋白,只要不相关蛋白质不会干扰功能单元1的组装和功能。作为实例,2b类sir可以包含连接至t细胞受体α(tcrα)的恒定链以构成功能多肽单元1的抗原结合结构域1和构成功能多肽单元2的t细胞受体β(tcrβ)的空(即缺乏抗原结合结构域)恒定链或由其组成。此类sir的两个功能多肽单元在同一细胞中共同表达。编码示例性2b类sir的构建体提供于cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-fmc63-vl-gly-ser-接头-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(082815-g07)[seqidno:1620]中。编码sir的核酸和氨基酸序列分别对应于seqidno:1620和seqidno:3855。在一些实施方式中,2类sir的两个功能多肽单元在细胞中使用不同载体共同表达。在一些实施方式中,2类sir的两个功能多肽单元在一个细胞中使用单一载体共同表达,该单一载体采用两个单独调控元件(例如启动子)编码对2类sir的两个功能多肽单元进行编码的两个多核苷酸。在一些实施方式中,2类sir的两个功能多肽单元在一个细胞中使用单一载体共同表达,该单一载体采用单一启动子表达含有分开编码sir的两个多肽的核苷酸片段的ires序列的多核苷酸。在一些实施方式中,2类sir的两个功能多肽单元在一个细胞中使用单一载体共同表达,该单一载体采用单一启动子表达编码含有可裂解接头(例如f2a、t2a、e2a、p2a、弗林蛋白酶-sgsg-f2a、弗林蛋白酶-sgsg-t2a、弗林蛋白酶-sgsg-e2a、弗林蛋白酶-sgsg-p2a等)的单一多肽的多核苷酸。所得mrna编码单一多肽,该单一多肽随后生成sir的两个功能多肽单元。在一些实施方式中,2类sir的两个功能多肽单元通过转染编码两个功能多肽单元的单一mrna序列来共同表达,而在其它实施方式中,两个功能多肽单元通过转染各自编码一个功能多肽单元的两个不同mrna序列来共同表达。在一些实施方式中,编码sir的载体或mrna可以编码额外基因/蛋白质(治疗性控制、抑制性分子、辅助模块等),其可通过ires或可裂解接头或其组合与sir编码序列分开。在另一个实施方式中,治疗性控制或辅助模块或两者可以在使用单独载体或mrna表达sir的细胞中表达。示例性治疗性控制提供于表8(seqidnos:3070至3076)中。应理解,治疗性控制或辅助模块不是sir的功能所必需的并且可以使用该实施方式的任何sir而无需治疗性控制或辅助模块。例如,抗生素抗性盒诸如pac(嘌呤霉素抗性基因)可以从本公开的sir编码载体移除而不会损害sir的功能。还提供本文所述的sir的功能性变体,这些变体具有与亲本sir的大量或显著序列一致性或相似性,该功能变体保留其所突变的sir的生物活性。功能变体包括例如本文描述的sir(亲本sir)的在与亲本sir类似的程度、相同的程度或更高程度上保留识别靶细胞的能力的那些变体。参考亲本sir,功能变体的氨基酸序列可以例如与亲本sir至少约30%、或50%、或75%、或80%、或85%、或90%、或91%、或92%、或93%、或94%、或95%、或96%)、或97%、或98%、或99%或更大一致。功能变体可以例如包含亲本sir的具有至少一个保守性氨基酸取代的氨基酸序列。可替代地或另外地,功能变体可以包含亲本sir的具有至少一个非保守性氨基酸取代的氨基酸序列。在此情况下,非保守性氨基酸取代不干扰或抑制功能变体的生物活性是优选的。非保守性氨基酸取代可以增强功能变体的生物活性,使得该功能变体的生物活性与亲本car相比有所增加。sir(包括功能部分和功能变体)可以具有任何长度,即可以包含任何数目的氨基酸,条件是sir(或其功能部分或功能变体)保留其生物活性,例如特异性结合抗原、检测受试者中的患病细胞、或治疗或预防哺乳动物中的疾病的能力等。例如,sir的长度可以是约300至约5000个氨基酸,诸如长度为300、400、500、600、700、800、900、1000或更多个氨基酸。sir(包括本公开的功能部分和功能变体)可以包含替代一个或多个天然存在的氨基酸的合成氨基酸。此类合成氨基酸为本领域中已知的并且包括例如氨基环己烷羧酸、正亮氨酸、a-氨基n-癸酸、高丝氨酸、s-乙酰基氨基甲基-半胱氨酸、反式-3-和反式-4-羟基脯氨酸、4-氨基苯丙氨酸、4-硝基苯丙氨酸、4-氯苯丙氨酸、4-羧基苯丙氨酸、β-苯基丝氨酸、β-羟基苯丙氨酸、苯基甘氨酸、α-萘基丙氨酸、环己基丙氨酸、环己基甘氨酸、二氢吲哚-2-羧酸、1,2,3,4-四氢异喹啉-3-羧酸、氨基丙二酸、氨基丙二酸单酰胺、n'-苄基-n'-甲基-赖氨酸、n',n'-二苄基-赖氨酸、6-羟基赖氨酸、鸟氨酸、α氨基环戊烷羧酸、α-氨基环己烷羧酸、oc-氨基环庚烷羧酸、-(2-氨基-2-降莰烷)-羧酸、γ-二氨基丁酸、α,β-二氨基丙酸、高苯丙氨酸以及α-叔丁基甘氨酸。sir(包括功能部分和功能变体)可以被糖基化、酰胺化、羧酸化、磷酸化、酯化、n-酰基化、经由例如二硫桥环化、或者转化成酸加成盐和/或任选地二聚化或聚合或缀合。本公开提供一种重组核酸构建体,其包含编码sir的核酸分子,其中该核酸分子包含编码一个或多个抗原结合结构域的核酸序列,其中编码每个抗原结合结构域的核苷酸序列与编码t细胞受体恒定链的核酸序列邻接并且与其处于相同阅读框中。可以用于构建sir的示例性t细胞受体恒定链包括但不限于tcrα、tcrβ1、tcrβ2、tcrγ、tcrδ、pretcrα及其变体和突变体的恒定链(参见例如表1--3)。在一些情况下,sir可以包含tcrα、tcrβ1、tcrβ2、tcrγ、tcrδ、pretcrα等的恒定链。本公开提供包含选自以下的tcr恒定链对的sir:tcrα和tcrβ1、tcrα和tcrβ2、pretcrα和tcrβ1、pretcrα和tcrβ2、以及tcrγ和tcrδ。本公开提供tcr恒定链与可在sir的构建中取代tcr恒定链的cd3z链的融合物。另外,sir中的人类pretcrα恒定链缺乏野生型人类pretcrα恒定链的羧基端48个氨基酸。缺乏羧基端48个氨基酸的pretcrα的氨基酸序列提供于seqidno:3048中。本公开还提供一种或多种载体,其包含核酸序列或编码本文所述的sir的序列。在一个实施方式中,sir由单一载体编码。在另一个实施方式中,sir由多于一个载体编码。在另一个实施方式中,sir的两个功能多肽单元各自由单独载体或由单独核酸编码。在一个实施方式中,sir的两个功能多肽单元各自由单一载体或单一核酸编码。在一个实施方式中,一个或多个载体选自一个或多个dna载体、一个或多个rna载体、一个或多个质粒、一个或多个慢病毒载体、一个或多个腺病毒载体、一个或多个逆转录载体、一个或多个杆状病毒载体、一个或多个睡美人转位子载体、或一个或多个背负式(piggyback)转座子。在一个实施方式中,该载体是慢病毒载体或逆转录病毒载体。在另一个实施方式中,载体是睡美人转座子载体。若干示例性载体的核酸序列提供于seqidno:870至876中。载体plenti-ef1α(seqidno:870)和plenti-ef1α-dwpre(seqidno:871)是与plenti-ef1α-dwpre缺乏wpre区域的事实不同的空慢病毒载体。本公开的sir编码序列可以克隆在这些载体的nhei与sali位点之间。载体mscv-bgl2-avrii-bam-ecor1-xho-bstb1-mlu-sal-clai.i03(seqidno:872)是逆转录病毒载体并且本公开的sir编码序列可以克隆在此载体的多克隆位点之间。载体mscv-fmc63vl-v5-[tcrb-kaciah]-f-p2a-2-spe-fmc63vh-myc-[tcrα-csdvp]-f-f2a-pac.n01(seqidno:873)也是逆转录病毒载体,其中已经存在sir编码序列。本公开的sir编码序列可以通过移除现有sir并插入编码新sir的核酸来克隆在此载体中。载体psbbi-pur(seqidno:874)是睡美人转座子载体。载体psbbi-pur-ef1-fmc63vl-v5-[tcrb-kaciah]-f-p2a-fmc63vh-myc-[tcra-csdvp]-f-f2a-xba.b01(seqidno:875)和psbbi-pur-ef1-nhe-fmc63vl-xho-v5-[tcrb-s57c-opt]-f-p2a-spe-fmc63vh-mlu-myc-[tcra-t48c-opt]-f2a-mcs-i01(seqidno:876)是含有含有sir核酸的睡美人转座子载体,这些载体可以用于在使用本领域已知的标准重组dna技术去除现有sir之后亚克隆本公开的sir。本公开还包括可以直接转染到细胞中的rna构建体。用于生成用于转染的mrna的方法涉及用特别设计的引物对模板进行体外转录(ivt)、然后添加聚a以产生含有3'和5'非翻译序列(“utr”)(例如,本文描述的3’和/或5’utr)、5’帽(例如,本文描述的5’帽)和/或内部核糖体进入位点(ires)(例如,本文描述的ires)、待表达的核酸和聚a尾的构建体,典型地长度为50-2000个碱基(seqidno:860和861)。如此产生的rna可以高效地转染不同种类的细胞。在一个实施方式中,模板包含针对sir的序列。在一个实施方式中,通过电穿孔将rnasir载体转导到细胞(例如,t细胞或nk细胞)中。在另一个实施方式中,通过在细胞膜中使用微流体装置引起瞬时扰动来将rnasir载体转导到细胞(例如,t细胞或nk细胞)中。sir的不同链(或功能多肽单元)也可以使用一个或多于一个载体、不同载体或技术的组合来引入细胞中。作为实例,载体cloneidno:050216-t02和cloneidno:050216-s08编码seqidno:913(cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-pac),其包含sir的功能多肽单元1,其中fmc63-vl链经由v5接头连接至htcrb-kaciah链。这些载体也表达嘌呤霉素抗性基因(pac)。载体cloneidno:041916-a02和041916-b03编码seqidno:997(cd8sp-fmc63-vh-myc-[tcrα-csdvp]-f-f2a-blastr),其包含sir的功能多肽单元2,其中fmc63-vh链经由myc接头连接至htcra-csdvp链。这些载体也表达杀稻瘟菌素抗性基因(blastr)。细胞可以同时或依次用编码seqidno:913和seqidno:997的慢病毒感染并且然后任选地针对嘌呤霉素和杀稻瘟菌素两者的抗性进行选择以富集双重感染细胞。用两种病毒感染的细胞两者均表达功能多肽单元,该功能多肽单元然后将组装以在细胞表面上表达功能sir。在另一个实施方式中,sir的一条链或功能多肽单元可以使用逆转录病毒载体引入,而另一个功能多肽单元使用慢病毒载体引入。在另一方面,sir的一个功能多肽单元可以使用慢病毒载体引入,而另一个功能多肽单元使用睡美人转座子引入。在另一方面,sir的一个功能多肽单元可以使用慢病毒载体引入,而另一个功能多肽单元使用rna转染引入。在另一方面,一个功能多肽单元在细胞中通过使用本领域已知的基因靶向技术在内源性tcr链基因座处进行遗传重组来产生,而另一个功能多肽单元使用慢病毒或逆转录病毒载体引入。可以使用许多不同方法中的任一种将rna引入到靶细胞中,这些不同方法例如可商购的方法,包括但不限于:电穿孔(amaxanucleofector-ii(德国科隆的艾玛科萨生物系统公司(amaxabiosystems,cologne,germany)),(ecm830(btx)(马萨诸塞州波士顿的哈佛仪器公司(harvardinstruments,boston,mass.))或genepulserii(科罗拉多州丹佛的伯乐公司(biorad,denver,colo.)),multiporator(德国汉堡的艾本德公司(eppendort,hamburggermany));阳离子脂质体介导的转染(使用脂质转染);聚合物包封;肽介导的转染;或生物射弹颗粒递送系统诸如“基因枪”(参见,例如nishikawa等人humgenether.[人类基因疗法],12(8):861-70(2001))或通过在细胞膜中使用微流体装置引起瞬时扰动(参见例如专利申请wo2013/059343a1和pct/us2012/060646)。在一些实施方式中,非病毒方法包括使用转座子(也称为可转座元件)。在一些实施方式中,转座子是可以将自身插入基因组中的位置处的dna片段,例如能够自我复制并将其拷贝插入基因组中的dna片段,或可以从较长核酸中剪接并插入基因组中的另一个位置中的dna片段。例如,转座子包含由侧接基因的用于转座的反向重复序列构成的dna序列。使用转座子的核酸递送的示例性方法包括睡美人转座子系统(sbts)和piggybac(pb)转座子系统。参见例如aronovich等人.hum.mol.genet.[人类分子遗传学]20.r1(2011):r14-20;singh等人.cancerres.[癌症研究]15(2008):2961-2971;huang等人.mol.ther.[分子疗法]16(2008):580-589;grabundzija等人.mol.ther.[分子疗法]18(2010):1200-1209;kebriaei等人.blood.[血液]122.21(2013):166;williams.moleculartherapy[分子疗法]16.9(2008):1515-16;bell等人.nat.protoc.[自然实验手册]2.12(2007):3153-65;以及ding等人.cell.[细胞]122.3(2005):473-83,所有这些文献均通过引用结合在本文中。sbts包括两个组分:1)含有转基因的转座子和2)转座酶的来源。转座酶可以将转座子从载体质粒(或其它供体dna)转移到靶标dna,如宿主细胞染色体/基因组。例如,转座酶与载体质粒/供体dna结合,从质粒中切出转座子(包括一个或多个转基因),并将它插入到宿主细胞的基因组中。参见例如aronovich等人同上。示例性转座子包括基于pt2的转座子。参见例如grabundzija等人.nucleicacidsres.[核酸研究]41.3(2013):1829-47;以及singh等人.cancerres.[癌症研究]68.8(2008):2961-2971,所有文献均通过引用结合在本文中。示例性转座子的核酸序列提供于seqidno:874和seqidno:875中。示例性转座酶包括tc1/海员型转座酶(mariner-typetransposase),例如sb10转座酶或sb11转座酶(可以例如从巨细胞病毒启动子表达的过度活跃的转座酶)。参见例如aronovich等人;kebriaei等人;以及grabundzija等人,所有文献均通过引用结合在本文中。sbts的使用允许转基因(例如,编码本文所述sir的核酸)的高效整合和表达。本文提供了生成细胞(例如,t细胞或nkt细胞或干细胞或ipsc或合成t细胞)的方法,该细胞例如使用转座子系统诸如sbts稳定地表达本文所述的sir。根据本文所述的方法,在一些实施方式中,将含有sbts组分的一种或多种核酸(例如质粒)递送至细胞(例如,t或nkt细胞或干细胞或ipsc或合成t细胞)。例如,通过核酸(例如,质粒dna)递送的标准方法递送一种或多种核酸,这些标准方法例如本文描述的方法,例如电穿孔、转染或脂质转染。在一些实施方式中,核酸含有包含转基因(例如,编码本文所述sir的核酸)的转座子。在一些实施方式中,核酸含有包含转基因(例如,编码本文所述sir的核酸)以及编码转座酶的核酸序列的转座子。在一些实施方式中,提供了具有两种核酸的系统,例如双质粒系统,例如,其中第一质粒含有包含转基因的转座子,并且第二质粒含有编码转座酶的核酸序列。例如,将第一核酸和第二核酸共同递送至宿主细胞中。在一些实施方式中,通过使用基因插入(使用sbts)和基因编辑(使用核酸酶(例如,锌指核酸酶(zfn)、转录活化因子样效应子核酸酶(talen)、crispr/cas系统、或工程化大范围核酸酶重新工程化的归巢内切核酸酶))的组合生成表达本文所述sir的细胞,例如t或nkt或干细胞或ipsc或合成t细胞。在一些实施方式中,使用非病毒递送方法允许细胞例如t细胞或nkt细胞或干细胞或ipsc或合成t细胞的重编程,并且将这些细胞直接输注到受试者体内。非病毒载体的优点包括但不限于容易且相对低成本地产生满足患者群体所需的足够量、储存期间的稳定性和缺乏免疫原性。在一些实施方式,包含编码sir的核酸序列的载体还可包含编码一种或多种抑制性分子的核酸序列。本文包括的抑制性分子的非限制性实例包括例如inhkir细胞质结构域;跨膜结构域,例如kir跨膜结构域;以及抑制剂细胞质结构域,例如itim结构域,例如inhkiritim结构域。在一个实施方式中,抑制性分子是野生型inhkir,或与野生型inhkir共享至少50%、60%、70%、80%、85%、90%、95%或99%同源性或相差不超过1、2、3、4、5、6、7、8、9、10、15或20个残基的序列;slam家族细胞质结构域;跨膜结构域,例如slam家族跨膜结构域;以及抑制剂细胞质结构域,例如slam家族结构域,例如slam家族itim结构域。在另一个实施方式中,抑制性分子是野生型slam家族成员,或与野生型slam家族成员共享至少50%、60%、70%、80%、85%、90%、95%或99%同源性或相差不超过1、2、3、4、5、6、7、8、9、10、15或20个残基的序列。在一些实施方式中,本公开的载体还可以包含启动子。启动子的非限制性实例包括例如ef-1启动子、cmvie基因启动子、ef-1α启动子、泛素c启动子、核心启动子或磷酸甘油酸激酶(pgk)启动子。在一些实施方式中,启动子是ef-1启动子。在另一个实施方式中,ef-1启动子包含seqidno:877。在一些实施方式中,该载体是rna核酸。在一些实施方式中,载体包含聚(a)尾。例如,本文考虑包含约150个腺苷碱基的聚(a)尾(seqidno:860至seqidno:864)。在一些实施方式中,该载体包含3'utr。在另一个方面,本公开提供一种制备细胞(例如,免疫效应细胞或其群体)的方法,该方法包括将包含编码sir(例如本文描述的sir)的核酸的载体或编码sir分子(例如本文描述的sir)的核酸引入到(例如转导到)细胞(例如,本文描述的t细胞、nkt细胞或干细胞或ipsc或合成t细胞)中。该细胞可以是免疫效应细胞(例如t细胞或nkt细胞或其组合)或可产生免疫效应细胞或合成t细胞的干细胞/祖细胞。在一些实施方式中,这些方法中的细胞是甘油二酯激酶(dgk)和/或ikaros缺陷型的。在一些实施方式中,这些方法中的细胞缺乏内源性t细胞受体α、β1、β2、pre-tcrα、γ或δ或其组合的恒定链。在一些实施方式中,这些方法中的细胞缺乏hla抗原。在一些实施方式中,这些方法中的细胞缺乏β2微球蛋白。在一些实施方式中,这些方法中的细胞缺乏sir的靶抗原的表达。例如,表达sir的t细胞在sir针对cd5的情况下缺乏内源性cd5,或者在sir针对tcr-β1恒定链的情况下缺乏tcr-β1恒定链,或者在sir针对tcr-β2的情况下缺乏tcr-β2恒定链,或者在sir针对cs1的情况下缺乏cs1。在一些实施方式中,引入编码sir的核酸分子包括转导包含编码sir的核酸分子的载体,或转染编码sir的核酸分子,其中该核酸分子是体外转录的rna。在一些实施方式中,核酸分子编码sir的两个或更多个组分,通过用超过一个载体转导细胞或用编码sir的不同亚基的两个或更多个核酸分子转染来引入。例如,细胞可以用各自编码sir的两个功能多肽单元之一的两个单独载体转导。由两个单独载体编码的示例性sir构建体由sir慢病毒构建体050216-s08提供,该构建体含有对应于seqidno:913且编码sir片段cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-pac的sir序列,其中源自cd19单克隆抗体fmc63的vl片段经由v5接头连接至具有kaciah突变的htcrb的恒定链。此sirfpu经由f-p2a可裂解接头连接至pac(嘌呤霉素抗性)基因。050216-s08构建体的载体是plenti-ef1α(seqidno:870)。sir慢病毒构建体041916-a02含有对应于seqidno:997且编码sir片段cd8sp-fmc63-vh-myc-[tcrα-csdvp]-f-f2a-blastr的sir序列,其中源自cd19单克隆抗体fmc63的vh片段经由myc接头连接至具有csdvp突变的htcra的恒定链。此sirfpu经由f-f2a可裂解接头连接至杀稻瘟菌素抗性基因。041916-a02构建体的载体是plenti-ef1α-dwpre(seqidno:871)。示例性选择标记物呈现于seqidno:795至seqidno:801中。类似地,细胞可以用各自编码sir的两个功能多肽单元之一的两个单独的体外转录的rna转导。除了sir的功能多肽单元之外,每个rna可以携带不同选择标记物或报告基因(例如tegfr或cd34或cnb30或突变体dhfr),其可以用于选择用两个rna转导并因此表达sir的两个功能多肽单元的细胞。在一些实施方式中,该方法还包括:a)提供免疫效应细胞(例如t细胞或nk细胞)群体;并且b)从该群体中去除t调控细胞,从而提供t调控耗尽细胞群体;其中步骤a)和b)在将编码sir的核酸引入该群体之前进行。在这些方法的实施方式中,t调控细胞包含cd25+t细胞,并且使用抗cd25抗体或其片段从细胞群体中去除。抗cd25抗体或其片段可以连接至底物(例如珠)。在其它实施方式中,从步骤(b)提供的t调控细胞被耗尽的免疫效应细胞群体含有少于30%、25%、20%、15%、10%、5%、4%、3%、2%、1%的cd25+细胞。在其它实施方式中,该方法还包括:从该群体中去除表达不包含cd25的疾病相关抗原的细胞,以提供t调控耗尽和肿瘤抗原耗尽的细胞群体,之后将编码sir的核酸引入该群体。疾病相关抗原可以选自cd19、cd30,、cd123、cd20、cd22、cd33、cd138、bcma、lym1、lym2、cd79b、cd170、cd179b、cd14或cd11b、或其组合。在其它实施方式中,该方法还包括从该群体中耗尽表达检查点抑制剂的细胞,以提供t调控耗尽和抑制性分子耗尽的细胞群体,之后将编码sir的核酸引入该群体。检查点抑制剂可以选自ctla-4、pd-1、lag-3、tim3、b7-h1、cd160、p1h、2b4、ceacam(例如ceacam-1、ceacam-3和/或ceacam-5)、tigit、btla和lairl。本公开还提供包含如本文描述的核酸分子、sir多肽分子或载体的重组细胞,例如免疫效应细胞(例如,细胞群体,例如,免疫效应细胞群体)和/或干细胞(例如造血干细胞、外周血干细胞、骨髓源性干细胞、免疫干细胞、诱导型多能干细胞或ipsc)。在一些实施方式中,该细胞是免疫细胞。免疫细胞的非限制性实例包括t细胞和nk细胞。另外,t细胞的非限制性实例包括treg、cd8+t细胞和cd4+t细胞。在一个实施方式中,细胞是人类t细胞。在一些实施方式中,细胞是人类细胞。在一些实施方式中,细胞是狗细胞。在一个实施方式中,人类t细胞是表达p-糖蛋白((p-gp或pgp;mdr1、abcb1、cd243)的t细胞。在一个实施方式中,人类t细胞是用为p-糖蛋白介导流出物的底物的染料迟钝染色的t细胞。在一个实施方式中,细胞是如申请号pct/us2017/042248中所述的t细胞,该申请通过引用结合在本文中。在一些实施方式中,从群体去除缺乏p-gp表达或p-gp活性的细胞。在一些实施方式中,细胞是甘油二酯激酶(dgk)和/或ikaros缺陷型的t细胞。在一个实施方式中,细胞是t细胞并且t细胞缺乏一条或多条内源性t细胞受体链。稳定缺乏根据本公开的功能tcr的表达的t细胞可以使用多种方法产生,这些方法诸如使用zn指核酸酶(zfn)、crisp/cas9和靶向内源性t细胞受体链的shrna。与shrna相关的非限制性示例性方法描述于us2012/0321667a1中,该专利通过引用结合在本文中。与使用靶向tcr的α和β链的恒定区的zfn消除内源性tcr表达相关的另一种非限制性示例性方法描述于torikaih等人(blood[血液],119(24),2012年6月14日)中。应注意在一些实施方式中,本公开的sir包含被密码子优化或设计为核苷酸序列不同于内源性tcr恒定链的tcr的恒定链并且因此逃避靶向内源性tcr恒定链的crisp/cas9、zfn和/或shrna所进行的靶向。缺乏功能性内源性tcr的t细胞可以例如被工程化以使其在其表面上不表达任何功能性内源性tcr,被工程化以使其不表达包含功能性内源性tcr的一个或多个亚基(例如内源性tcrα、tcrβ1、tcrβ2、tcrγ、tcrδ或pre-tcrα的恒定链),或者被工程化以使其在其表面上产生非常少的功能性内源性tcr。可替代地,t细胞可以例如通过表达tcr的一个或多个亚基的突变或截短形式来表达严重受损的内源性tcr。术语“严重受损的tcr”意指该tcr不会在宿主中引发不利的免疫反应。未修饰tcr当异位表达(例如使用逆转录病毒或慢病毒载体)时通常在原代人类t细胞上较差表达,这表明它们无效率地与内源性tcr链竞争细胞表面表达。然而,已显示,优化tcr链以用于在人类细胞中有效翻译,使得所引入tcr得到更好表达。更重要的是,此类主导tcr的异位表达防止在人类t细胞中发生大比例内源性tcr库的表面表达。在一个实施方式中,细胞是干细胞并且干细胞缺乏一条或多条内源性t细胞受体链。在另一个实施方式中,细胞是干细胞,其中sir的一个或多个靶抗原(例如mpl、cd33、cd123、cd19等)已缺失或突变成不再被sir识别的形式。作为实例,靶向cd19的sir在干细胞中表达,这些干细胞已使用crisp/cas9或zn指核酸酶制成缺乏cd19,使得由此类干细胞产生的b细胞未能通过表达靶向cd19的sir的t细胞消除。可替代地,靶向cd19的sir在干细胞中表达,在这些干细胞中内源性cd19已使用crisp/cas9或zn指核酸酶突变成sir不能触发的形式,使得由此类干细胞产生的b细胞未能通过表达靶向cd19的sir的t细胞消除。在另一个实施方式中,sir在免疫效应细胞中表达并且来自自体或同种异体供体的干细胞被遗传工程化以缺乏sir靶抗原的表达或以表达sir靶抗原的sir不能识别的突变形式。例如,靶向cd19的sir在连同自体或同种异体造血干细胞一起输注到患者体内的t细胞中表达,这些干细胞已使用crisp/cas9或zn指核酸酶制成缺乏cd19,使得由此类干细胞产生的b细胞未能通过表达靶向cd19的sir的t细胞消除。可替代地,靶向cd19的sir在连同自体或同种异体造血干细胞一起输注到患者体内的t细胞中表达,在这些干细胞中内源性cd19已使用crisp/cas9或zn指核酸酶突变成sir不能触发的形式,使得由此类干细胞产生的b细胞未能通过表达靶向cd19的sir的t细胞消除。可以使用类似方法,在接受靶向这些抗原的sir-t细胞的受试者中使用shrna、crisp/cas9或zn指核酸酶突变或消除干细胞中的其它内源性抗原(例如mpl、cd33、cd123等),以用于治疗这些抗原在疾病相关细胞或引起疾病细胞上表达的特定疾病。t细胞或天然杀伤细胞(nk)或干细胞可以从受试者中获得。术语“受试者”旨在包括可以在其中引发免疫响应的活生物体(例如,哺乳动物)。受试者的实例包括人、猴、黑猩猩、狗、猫、小鼠、大鼠及其转基因物种。t细胞可以从许多来源获得,包括外周血单核细胞、骨髓、淋巴结组织、脐带血、胸腺组织、来自感染部位的组织、腹水、胸膜积液、脾组织和肿瘤。t细胞可以是组织留置γ-δt细胞,其可以在体外培养并扩增,之后表达sir。表达sir和/或car和/或ab-tcr和/或truc和/或ctcr的免疫效应细胞可以通过用蛋白质l刺激来扩增。在一个方面,蛋白质l固定于珠或另一个表面诸如板上。在一个方面,蛋白质l固定于与cd3抗体相同的珠上。在一个方面,蛋白质l固定于与cd28抗体相同的珠上。在一个方面,蛋白质l固定于cd3抗体和cd28抗体所固定的珠上。在一个方面,蛋白质l在人工抗原呈递细胞的表面上表达。在一个方面,蛋白质l与一个或多个共刺激分子结合在人工抗原呈递细胞的表面上表达。在一个方面,共刺激分子包括cd28、41bb或ox40中的一种或多种。在一个方面,在其表面上表达蛋白质l的细胞是哺乳动物细胞。在一个方面,细胞是人类细胞。在一个方面,细胞是293ft细胞。在一个方面,细胞是k562细胞。在一个方面中,蛋白质l在细胞中稳定表达。在另一个方面中,蛋白质l在细胞中瞬时表达。蛋白质l可以通过本领域已知的任何方法在细胞中表达。在一个方面,表达sir或car或ab-tcr或truc或ctcr的免疫效应细胞通过与涂覆蛋白质l的珠或apc共培养10min至若干天或周的时间段(或其之间的任何时间段)来扩增。在本公开的某些方面,可以使用任何数量的本领域技术人员已知的技术(诸如ficolltm分离)从自受试者收集的血液单位获得免疫效应细胞,例如t细胞。在一个优选的方面,通过单采血液成分术获得来自个体的循环血液的细胞。单采血液成分术产物典型地含有淋巴细胞(包括t细胞)、单核细胞、粒细胞、b细胞、其它有核白细胞、红细胞和血小板。在一个方面,可以洗涤通过单采血液成分术收集的细胞以去除血浆部分并且任选地将细胞置于适当的缓冲液或培养基中用于后续加工步骤。在本发明的一个实施方式中,使用磷酸盐缓冲盐水(pbs)洗涤这些细胞。在一个替代实施方式中,洗涤溶液缺少钙并且可能缺少镁,或者可能缺少许多(如果不是全部)二价阳离子。在不存在钙的情况下的初始活化步骤可以导致放大的活化。如本领域普通技术人员易于理解的,洗涤步骤可以通过本领域技术人员已知的方法完成,例如通过根据制造商的说明书使用半自动“流通”离心机(例如,cobe2991细胞处理器、百特(baxter)cytomate、或汉尼帝克(haemonetics)cellsaver5)。洗涤后,可将细胞重悬于各种生物相容性缓冲液(例如无ca、无mg的pbs,plasmalytea,或具有或不具有缓冲液的其它盐溶液)中。可替代地,可以去除单采血液成分术的样品中不需要的组分,并将细胞直接重悬于培养基中。应认识到,本申请的方法可以利用包含5%或更少(例如2%)人类ab血清的培养基条件,并采用已知的培养基条件和组合物,例如smith等人,“exvivoexpansionofhumantcellsforadoptiveimmunotherapyusingthenovelxeno-freectsimmunecellserumreplacement[使用新型无xenocts免疫细胞血清替代物进行过继性免疫疗法的人t细胞的离体扩增]”clinical&translationalimmunology[临床和转化免疫学](2015)4,e31;doi:10.1038/cti.2014.31中所述的那些。在一方面,通过裂解红细胞并耗尽单核细胞(例如,通过逆流离心淘洗或通过percolltm梯度离心)从外周血淋巴细胞分离t细胞。在另一个实施方式中,本文描述的表达sir的免疫效应细胞可以进一步表达增强表达sir的细胞的活性的药剂。在一些实施方式中,该药剂是抑制抑制性分子的药剂。抑制性分子的非限制性实例包括pd-1、pd-l1、ctla-4、tim-3、ceacam(例如,ceacam-1、ceacam-3和/或ceacam-5)、lag-3、vista、btla、tigit、lair1、cd160、2b4和tgfβ。抑制这些抑制性分子的药剂的非限制性实例提供于seqidno:3102至3107中(编码序列seqidno:827-832)(参见表8)。在一个实施方式中,抑制抑制性分子的药剂包括结合抑制性分子的第一多肽,例如scfv或vhh或受体或配体片段,其与向细胞提供正信号的第二多肽缔合,该第二多肽例如细胞内信号传导结构域,诸如41bb、cd27、ox40、cd28、dap10、cd2、cd5、icam-1、lfa-1、lck、tnfr-1、tnfr-ii、fas、cd30、cd40或其组合)和/或初级信号传导结构域(例如cd3ζ信号传导结构域)。表达此类多肽的示例性sir呈现于seqidno:3217至3219和seqidno:3221和3222中。在一个实施方式中,抑制抑制性分子的药剂包括结合抑制性分子的第一多肽,例如scfv或vhh片段或受体或配体片段,其与本文所述t细胞受体恒定链(例如tcra、tcrb1、tcrb2、pre-tcra、pre-tcra-del48、tcr-γ或tcr-δ的恒定链)缔合。结合抑制性分子的示例性sir呈现于seqidno:3572、3573、3574和3575中。在另一个实施方式中,本文所述的表达sir的细胞可以进一步表达辅助模块,例如增强表达sir的细胞的活性的药剂。包括可增强表达sir的细胞的活性的药剂的辅助模块的若干实例提供于seqidno:3087至3116(表8)。例如,在一个实施方式中,该药剂可以是增加sir链(例如cd3ζ、cd3δ、cd3ε、cd3γ或其组合)的表达和/或活性的药剂。在另一个实施方式中,该药剂可以是向表达sir的细胞提供共刺激信号的药剂(例如vflipk13、vflipmc159、cflip-l、cflip-p22、htlv1tax、htlv2tax、41bb或cd28)。在另一个实施方式中,该药剂可以是以可诱导方式向表达sir的细胞提供共刺激信号的药剂(例如fkbpx2-k13、fkbpx2-mc159、fkbpx2-cflip、fkbpx2-cflip-l、fkbpx2-cflip-p22、fkbpx2-htlv1tax、fkbpx2-htlv2tax、fkbpx2-41bb或fkbpx2-cd28、myr-myd88-cd40-fv'-fv等)。在另一个实施方式中,该药剂可以是促进表达sir的细胞的增殖或持久性的细胞因子或趋化因子(例如cd40l、il2、il-7、il-15、il12f或il-21)。在另一个实施方式中,该药剂可以是促进表达sir细胞的活性和/或与表达sir的细胞协同作用的可溶性受体(例如shvem或shvem-alb8-vhh)。在一个实施方式中,该药剂可以是抑制抑制性分子的药剂。在一些实施方式中,抑制性分子(例如,pd1)可降低表达sir的细胞产生免疫效应子反应的能力。在另一个实施方式中,该药剂可以是靶向pd1或ctla4的scfv。示例性靶向pd1和ctla4的scfv提供于seqidno:3102至3107中。在一个实施方式中,该药剂包含例如抑制性分子的第一多肽,该抑制性分子诸如pd1、pd-l1、ctla4、tim3、ceacam(例如,ceacam-1、ceacam-3和/或ceacam-5)、lag3、vista、btla、tigit、lair1、cd160、2b4或tgfβ、或这些中的任一个的片段(例如,这些中的任一个的细胞外结构域的至少一部分),以及第二多肽,该第二多肽是本文所述的细胞内信号传导结构域(例如,包含共刺激结构域(例如,41bb、cd27或cd28,例如,如本文描述的)和/或初级信号传导结构域(例如,本文所述的cd3ζ信号传导结构域)。在一个实施方式中,该药剂包含pd1的第一多肽或其片段(例如,pd1的细胞外结构域的至少一部分)以及本文所述的细胞内信号传导结构域(例如,本文所述的cd28信号传导结构域和/或本文所述的cd3ζ信号传导结构域)的第二多肽。表8:在一个实施方式中,该药剂包含pd-1的第一多肽或其片段以及本文描述的细胞内信号传导结构域(例如,cd28、cd27、ox40或4-ibb信号传导结构域和/或cd3ζ信号传导结构域)的第二多肽。在另一个实施方式中,该药剂包含pd-1的第一多肽或其片段以及第二多肽,该第二多肽是本文所述的t细胞受体恒定链(例如tcra、tcrb1、tcrb2、pre-tcra、pre-tcra-del48、tcr-γ或tcr-δ的恒定链)。在一个实施方式中,本文所述的表达sir的效应细胞可以还包含可诱导对相同或不同靶标的不同抗原结合结构域的第二sir。在一些实施方式中,第二sir可靶向与第一sir相同或不同的细胞类。在一个实施方式中,本文所述的表达sir的效应细胞可以还包含任选地对相同或不同靶标具有相同或不同抗原结合结构域的car。在一些实施方式中,car可靶向与第一sir相同或不同的细胞类。若干示例性car的核酸和氨基酸序列分别呈现于seqidno:9659至9854和seqidno:9873至10068中。在一个实施方式中,car包含针对在与疾病相关抗原相同的疾病细胞类型(例如癌症)上表达的靶标的抗原结合结构域。在一个实施方式中,表达sir的细胞包含靶向第一抗原的sir和靶向第二不同抗原的car并且包含不具有初级信号传导结构域但具有共刺激信号传导结构域的细胞内信号传导结构域。虽然不希望受到理论约束,但将共刺激信号传导结构域例如4-1bb、cd28、cd27或ox-40置于car上,可以调节sir对表达两种靶标的细胞的活性。在一个实施方式中,表达sir的细胞包含i)第一疾病相关抗原sir,该第一疾病相关抗原sir包含结合本文描述的靶抗原的一个或多个抗原结合结构域、以及一条或两条tcr恒定链;以及ii)car,该car靶向不同靶抗原(例如,在与第一靶抗原相同的疾病相关(例如癌症)细胞类型上表达的抗原)并且包含抗原结合结构域、跨膜结构域和初级信号传导结构域以及共刺激结构域。具有此配置的示例性构建体的核酸和氨基酸序列分别呈现于seqidno:983和seqidno:3218中。在此构建体中sir的抗原结合结构域包含源自靶向cd19的fmc63单克隆抗体的vl和vh片段,而car的抗原结合结构域包含pd1的细胞外结构域,在此构建体中car的初级信号传导结构域包含cd3z细胞溶质结构域,而共刺激结构域包含4-1bb细胞溶质结构域。在另一个实施方式中,表达sir的细胞包含i)sir,该sir包含结合本文描述的靶抗原的抗原结合结构域和一条或两条tcr恒定链;以及ii)car,该car靶向除第一靶抗原以外的抗原(例如,在与第一靶抗原相同的癌细胞类型上表达的抗原)并且包含针对该抗原的抗原结合结构域、跨膜结构域和共刺激信号传导结构域。具有此配置的示例性构建体的核酸和氨基酸序列分别呈现于seqidno:982和seqidno:3217中。此构建体类似于seqidno:983中所示的构建体,除了car缺乏cd3z结构域。在另一个实施方式中,表达sir的细胞包含i)第一疾病相关抗原sir,该第一疾病相关抗原sir包含结合本文描述的靶抗原的一个或多个抗原结合结构域、以及一条或两条tcr恒定链;以及ii)car,该car靶向不同靶抗原(例如,在与第一靶抗原相同的疾病相关(例如癌症)细胞类型上表达的抗原)并且包含抗原结合结构域、跨膜结构域和初级信号传导结构域,但不具有共刺激结构域。在一个实施方式中,car包含抗原结合结构域、跨膜结构域和细胞内信号传导结构域(诸如但不限于来自41bb、cd27、ox40、cd28、dap10、cd2、cd5、icam-1、lfa-1、lck、tnfr-1、tnfr-ii、fas、cd30、cd40或其组合的一个或多个细胞内信号传导结构域)和/或初级信号传导结构域(诸如但不限于cd3ζ信号传导结构域)。与car共同表达的示例性sir呈现于seqidno:3217至3219和seqidno:3221和3222中。在一个实施方式中,表达sir的效应细胞包含本文所述的sir和抑制性car。在一个实施方式中,抑制性car包含结合正常细胞而非癌细胞上发现的抗原的抗原结合结构域。在一个实施方式中,抑制性car包含抑制性分子的抗原结合结构域、跨膜结构域和细胞内结构域。例如,抑制性car的细胞内结构域可以是pd1、pd-l1、ctla-4、tim-3、ceacam(例如,ceacam-1、ceacam-3和/或ceacam-5)、lag-3、vista、btla、tigit、lair1、cd160、2b4或tgfβ中任一个的细胞内结构域。与抑制性car共同表达的示例性sir多肽呈现于seqidno:3220中。在此多肽中抑制性car表达与lair1的跨膜结构域和细胞溶质结构域融合的靶向cxcr4的vhh。在某些实施方式中,sir分子的抗原结合结构域包含scfv,并且car分子的抗原结合结构域不包含scfv。例如,sir分子的抗原结合结构域包含scfv,并且car分子的抗原结合结构域包含骆驼科vhh结构域。在一个实施方式中,本公开提供一种免疫效应细胞(例如,t细胞、nk细胞),该细胞表达sir,该sir包含结合至如本文描述的肿瘤抗原的抗原结合结构域;和car,该car包含pd1细胞外结构域或其片段。在一些实施方式中,该细胞还包含抑制性分子,该抑制性分子包含inhkir细胞质结构域;跨膜结构域,例如kir跨膜结构域;以及抑制剂细胞质结构域,例如itim结构域,例如inhkiritim结构域。在一个实施方式中,抑制性分子是天然存在的inhkir,或与天然存在的inhkir共享至少50%、60%、70%、80%、85%、90%、95%或99%同源性或相差不超过1、2、3、4、5、6、7、8、9、10、15或20个残基的序列;或slam家族细胞质结构域;跨膜结构域,例如slam家族跨膜结构域;以及抑制剂细胞质结构域,例如slam家族结构域,例如slam家族itim结构域。在另一个实施方式中,抑制性分子是天然存在的slam家族成员,或与天然存在的slam家族成员共享至少50%、60%、70%、80%、85%、90%、95%或99%同源性或相差不超过1、2、3、4、5、6、7、8、9、10、15或20个残基的序列。本公开还提供一种方法,该方法包括向受试者给予sir分子、表达sir分子的细胞或包含编码sir分子的核酸的细胞。在一个实施方式中,受试者患有本文描述的病症,例如受试者患有癌症、感染疾病、过敏性疾病、退行性疾病或自身免疫疾病,该病症表达本文所述的靶抗原。在一个实施方式中,受试者具有增加的本文描述的病症的风险,例如受试者具有增加的癌症、感染疾病、过敏性疾病、退行性疾病或自身免疫疾病的风险,该病症表达本文所述的靶抗原。在一个实施方式中,受试者是人类。在另一个实施方式中,受试者是成人。在另一个实施方式中,受试者是伴侣动物诸如狗。本公开提供了用于治疗或预防与本文描述的疾病相关抗原的表达相关的疾病的方法。在一个实施方式中,本公开提供治疗或预防疾病的方法,这些方法通过向有需要的受试者提供被工程化以表达靶向x-sir的免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行,其中x表示如本文描述的疾病相关抗原,并且其中该引起疾病细胞或疾病相关细胞表达所述x抗原。表9提供了不同抗原和可以使用表达靶向这些抗原的sir的免疫效应细胞预防、抑制或治疗的示例性疾病的列表。表9:在另一个实施方式中,本公开提供治疗或预防癌症的方法,这些方法通过向有需要的受试者提供被工程化以表达本文所述的xsir(或x-sir)的免疫效应细胞(例如t细胞)来进行,其中癌症细胞表达抗原靶标“x”。在一个实施方式中,x在正常细胞和癌症细胞两者上表达,但在正常细胞上的表达水平较低。在一个实施方式中,该方法还包括选择sir,该sir结合x的亲和力允许xsir结合并杀伤表达x的癌细胞,但少于30%、25%、20%、15%、10%、5%或更少的表达x的正常细胞被杀伤,例如,如通过本文所述的测定来测定的。例如,本文所述的gluc释放细胞毒性测定可以用于识别靶向例如癌细胞的xsir。在一个实施方式中,所选sir具有如下抗原结合结构域,该抗原结合结构域对靶抗原的结合亲和力kd为约10-4m至10-8m,更通常为约10-5m至10-7m,以及通常为约10-6m或10-7m。在一个实施方式中,所选抗原结合结构域的结合亲和力比参考抗体(例如本文描述的抗体)低至少2倍、至少5倍、10倍、20倍、30倍、50倍、100倍或1000倍并且sir的结合结构域来源于该参考抗体。在另一个实施方式中,本公开提供治疗或预防疾病的方法,这些方法通过向有需要的受试者提供被工程化以表达tcrb1-sir的免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行,其中该引起疾病细胞或疾病相关细胞表达tcrb1(t细胞受体β1链)。在一个实施方式中,有待治疗或预防的疾病是癌症或免疫疾病。在一个实施方式中,有待治疗或预防的癌症是t细胞白血病或t细胞淋巴瘤。在一个实施方式中,有待治疗或预防的免疫病症是多发性硬化、类风湿关节炎、强直性脊柱炎、炎性肠病、糖尿病、移植物抗宿主病或自身免疫性甲状腺炎。在另一个实施方式中,本公开提供治疗或预防疾病的方法,这些方法通过向有需要的受试者提供被工程化以表达tcrb2-sir的免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行,其中该引起疾病细胞或疾病相关细胞表达tcrb2(t细胞受体β2sir)。在一个实施方式中,有待治疗或预防的疾病是癌症或免疫病症。在一个实施方式中,有待治疗或预防的癌症是t细胞白血病或t细胞淋巴瘤。在一个实施方式中,有待治疗或预防的免疫病症是多发性硬化、类风湿关节炎、强直性脊柱炎、炎性肠病、糖尿病、移植物抗宿主病或自身免疫性甲状腺炎。在另一个实施方式中,本公开提供治疗或预防疾病的方法,这些方法通过向有需要的受试者提供被工程化以表达t细胞受体γ-δ-sir的免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行,其中该引起疾病细胞或疾病相关细胞表达t细胞受体γ-δ。在一个实施方式中,有待治疗或预防的疾病是癌症或免疫病症。在一个实施方式中,有待治疗或预防的癌症是t细胞白血病或t细胞淋巴瘤。在一个实施方式中,有待治疗或预防的免疫病症是多发性硬化、类风湿关节炎、强直性脊柱炎、炎性肠病、糖尿病、移植物抗宿主病或自身免疫性甲状腺炎。在另一个实施方式中,本公开提供治疗或预防疾病的方法,这些方法通过向有需要的受试者提供被工程化以表达编码cd4-dc-sign的sir的免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行。在一个实施方式中,有待治疗或预防的疾病是hiv1/aids。在另一个实施方式中,本公开提供治疗或预防自身免疫疾病的方法,这些方法通过向有需要的受试者提供被工程化以表达编码自身抗原或其片段的sir的免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行。在一个实施方式中,自身免疫疾病是糖尿病、类风湿关节炎、多发性硬化、寻常天疱疮、副肿瘤性天疱、肾小球肾炎、强直性脊柱炎、溃疡性结肠炎或克罗恩病(crohn’sdisease)。在一个方面,该疾病是寻常天疱疮,并且sir的抗原结合结构域包含桥粒芯蛋白3(dsg3)的细胞外结构域。在另一个实施方式中,本公开提供治疗或预防癌症、感染、自身免疫或过敏性疾病的方法,这些方法通过向有需要的受试者提供免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行,这些免疫效应细胞被工程化以表达编码cd16或其缺失或点突变体片段的通用sir以及结合至sir的cd16结构域和疾病相关细胞上表达的抗原的抗体或抗体片段。在一个方面,疾病相关细胞是癌细胞、受感染细胞、或浆细胞或b细胞或t细胞。在另一个实施方式中,本公开提供治疗或预防癌症、感染、自身免疫或过敏性疾病的方法,这些方法通过向有需要的受试者提供被工程化以表达编码免疫球蛋白结合受体或其缺失或点突变体片段的通用sir的免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行。向患者给予表达sir的免疫效应细胞以及结合至sir受体的免疫球蛋白结合结构域的一种或多种抗体或抗体片段和在疾病相关细胞上表达的一种或多种抗原。在一个方面,疾病相关细胞是癌细胞、受感染细胞、或浆细胞或b细胞或t细胞。在另一个实施方式中,本公开提供治疗或预防癌症、感染、自身免疫或过敏性疾病的方法,这些方法通过向有需要的受试者提供免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞,这些免疫效应细胞被工程化以表达连接至t细胞受体恒定链(例如tcrα的恒定链)的编码免疫球蛋白结合受体或其缺失或点突变体片段的通用sir和连接至t细胞受体恒定链(例如tcrβ的恒定链)的抗原结合结构域(例如,scfv、vhh、vl、vh或非免疫球蛋白抗原结合结构域)。向患者给予表达sir的免疫效应细胞以及结合至第一sir受体的免疫球蛋白结合结构域的一种或多种抗体或抗体片段和在疾病相关细胞上表达的一种或多种抗原。在一个方面,疾病相关细胞是癌细胞、受感染细胞、或浆细胞或b细胞或t细胞。在另一个实施方式中,本公开提供治疗或预防癌症、感染、自身免疫或过敏性疾病的方法,这些方法通过向有需要的受试者提供免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行,这些免疫效应细胞被工程化以表达编码免疫球蛋白受体或其缺失或点突变体片段的通用sir连同结合至以上受体和疾病相关细胞上表达的抗原的抗体或抗体片段。在另一个实施方式中,本公开提供治疗或预防癌症、感染、自身免疫或过敏性疾病的方法,这些方法通过向有需要的受试者提供免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞,这些免疫效应细胞被工程化以表达连接至t细胞受体恒定链的编码cd16或其缺失或点突变体(例如v158突变体)片段的通用sir和连接至t细胞受体恒定链的编码抗原结合结构域(例如,scfv、vhh、vl、vh或非免疫球蛋白抗原结合结构域)的sir。向患者给予表达sir的免疫效应细胞以及结合至sir的cd16结构域的一种或多种抗体或抗体片段和在疾病相关细胞上表达的一种或多种抗原。在一个方面,疾病相关细胞是癌细胞、受感染细胞、或浆细胞或b细胞或t细胞。在另一个实施方式中,本公开提供治疗或预防癌症、感染、自身免疫或过敏性疾病的方法,这些方法通过向有需要的受试者提供免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行,这些免疫效应细胞被工程化以表达编码cd16或其缺失或点突变体片段(例如v158突变体)的通用sir以及结合至sir的cd16结构域和疾病相关细胞上表达的一种或多种抗原的一种或多种抗体或抗体片段。在一个方面,疾病相关细胞是癌细胞、受感染细胞、或浆细胞或b细胞或t细胞。在另一个实施方式中,本公开提供治疗或预防癌症、感染、自身免疫或过敏性疾病的方法,这些方法通过向有需要的受试者提供被工程化以表达编码nkg2d或其缺失或点突变体片段的sir的免疫效应细胞(例如t细胞)。在一个方面,疾病相关细胞是癌细胞、受感染细胞、或浆细胞或b细胞或t细胞。在另一个实施方式中,本公开提供治疗或预防疾病的方法,这些方法通过向有需要的受试者提供被工程化以表达cd19sir的免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行。在一个方面,该疾病是免疫或过敏性疾病。在另一个实施方式中,本公开提供治疗或预防疾病的方法,这些方法通过向有需要的受试者提供被工程化以表达cd20sir的免疫效应细胞(例如t细胞)来进行。在一个方面,该疾病是免疫或过敏性疾病。在另一个实施方式中,本公开提供治疗或预防疾病的方法,这些方法通过向有需要的受试者提供被工程化以表达cd22sir的免疫效应细胞(例如t细胞)来进行。在一个方面,该疾病是免疫或过敏性疾病。在另一个实施方式中,本公开提供治疗或预防癌症、感染、自身免疫或过敏性疾病的方法,这些方法通过向有需要的受试者提供免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行,这些免疫效应细胞被工程化以表达fitc-sir以及结合至疾病相关细胞上表达的抗原的fitc标记抗体或抗体片段或抗体片段或受体或配体或非免疫球蛋白支架。在一个方面,疾病相关细胞是癌细胞、受感染细胞、或浆细胞或b细胞或t细胞。在另一个实施方式中,本公开提供治疗或预防癌症、感染、自身免疫或过敏性疾病的方法,这些方法通过向有需要的受试者提供免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行,这些免疫效应细胞被工程化以表达抗生物素蛋白-sir以及结合至疾病相关细胞上表达的抗原的生物素标记抗体或抗体片段或抗体片段或受体或配体或非免疫球蛋白支架。在一个方面,疾病相关细胞是癌细胞、受感染细胞、或浆细胞。在另一个实施方式中,本公开提供治疗或预防癌症、感染、自身免疫或过敏性疾病的方法,这些方法通过向有需要的受试者提供免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞来进行,这些免疫效应细胞被工程化以表达streptag-sir以及结合至疾病相关细胞上表达的抗原的含streptag抗体或抗体片段或抗体片段或受体或配体或非免疫球蛋白支架。在一个方面,疾病相关细胞是癌细胞、受感染细胞、或浆细胞。在另一个实施方式中,本公开提供治疗或预防疾病的方法,这些方法通过向有需要的受试者提供被工程化以表达ige-sir的免疫效应细胞(例如t细胞)来进行,该ige-sir的抗原结合结构域包含结合至ige的抗体或抗体片段。在一个方面,该疾病是免疫或过敏性疾病。在另一个实施方式中,本公开涉及在体内使用pd1sir(即,含有pd1的细胞外结构域作为其抗原结合结构域的sir)治疗受试者,使得癌性肿瘤的生长得到抑制。示例性pd1-sir的核酸序列提供于seqidno:1337中。pd1sir可以单独用于抑制癌性肿瘤的生长。可替代地,pd1sir可以与其它sir、car、免疫原性药剂、标准癌症治疗或其它抗体结合使用。在一个实施方式中,用本文描述的pd1sir和xsir治疗受试者。在另一个实施方式中,pd1sir与另一种sir或car(例如,本文描述的sir或car)和激酶抑制剂(例如,本文描述的激酶抑制剂)结合使用。在另一个实施方式中,本公开涉及在体内使用xsir和pd1-car或ctl4-car治疗受试者,使得癌性肿瘤的生长得到抑制。在一个实施方式中,用本文描述的pd1-car或ctla4-car和xsir治疗受试者。编码pd1-car和xsir(例如cd19-sir)的示例性构建体的核酸序列提供于seqidno:982-984中。编码ctla4-car和xsir(例如cd19-sir)的示例性构建体的核酸序列提供于seqidno:986-987中。在另一个实施方式中,pd1-car与另一种sir或car(例如,本文描述的sir或car)和激酶抑制剂(例如,本文描述的激酶抑制剂)结合使用。在一个实施方式中,xsir与pd1-car或ctl4-car结合使用。在另一个方面,提供了一种治疗受试者,例如减轻或改善受试者的过度增生性病症或病状(例如,癌症)、例如实体肿瘤、软组织肿瘤、血液癌症或转移性病灶的方法。如本文使用的术语“癌症”意欲包括所有类型的癌性生长或致癌性过程、转移性组织或恶性转化的细胞、组织或器官,而不考虑组织病理学类型或侵袭的阶段。实体肿瘤的实例包括各种器官系统的恶性肿瘤,例如腺癌、肉瘤和癌,这些器官系统诸如影响乳腺、肝、肺、脑、淋巴、胃肠(例如结肠)、泌尿生殖道(例如,肾、尿路上皮细胞)、前列腺和咽的那些。腺癌包括癌症,例如大多数结肠癌、直肠癌、肾细胞癌、肝癌、非小细胞肺癌、小肠癌和食管癌。在一个实施方式中,癌症是黑素瘤,例如晚期黑素瘤。还可以使用本公开的方法和组合物治疗或预防前述癌症的转移性病灶。可以治疗或预防的其它癌症的实例包括胰腺癌、骨癌、皮肤癌、皮肤或眼内恶性黑素瘤、子宫癌、卵巢癌、直肠癌、头颈癌、肛区癌、胃癌、睾丸癌、子宫癌、输卵管癌、子宫内膜癌、子宫颈癌、阴道癌、外阴癌、霍奇金病、非霍奇金淋巴瘤、食道癌、小肠癌、内分泌系统癌症、甲状腺癌、甲状旁腺癌、肾上腺癌、软组织肉瘤、尿道癌、阴茎癌、慢性或急性白血病(包括急性骨髓性白血病、慢性骨髓性白血病、急性成淋巴细胞白血病、慢性成淋巴细胞性白血病)、儿童实体瘤、淋巴细胞性淋巴瘤、膀胱癌、肾或输尿管癌、肾盂癌、中枢神经系统(cns)肿瘤、原发性cns淋巴瘤、肿瘤血管生成、脊轴肿瘤、脑干胶质瘤、垂体腺瘤、卡波济氏肉瘤、表皮样癌、鳞状细胞癌、t细胞淋巴瘤、环境诱导的癌症(包括由石棉诱导的那些)、以及所述癌症的组合。可以使用本文描述的抗体分子来实现转移性癌症,例如表达pd-l1的转移性癌症的治疗(iwai等人(2005)int.immunol.[国际免疫学]17:133-144)。生长可能受到抑制的示例性癌症包括典型地对免疫疗法有反应的癌症。治疗的癌症的非限制性实例包括肾癌(例如清晰细胞癌)、黑素瘤(例如转移性恶性黑素瘤)、乳腺癌、前列腺癌(例如激素难治性前列腺腺癌)、结肠癌和肺癌(例如非小细胞肺癌)。另外,可以使用本文描述的分子来治疗复发性或难治性恶性肿瘤。在一个实施方式中,本公开涉及一种载体,该载体包含可操作地连接至启动子以在哺乳动物免疫效应细胞(例如t细胞)或可产生免疫效应细胞的干细胞中表达的sir。在一个方面,本发明提供了表达本发明sir的重组免疫效应细胞,其用于治疗或预防表达如本文描述的癌症相关抗原的癌症。在一个方面,本公开的表达sir的细胞能够使肿瘤细胞与在其表面上表达的至少一种癌症相关抗原接触,以使得表达sir的细胞靶向癌细胞并抑制癌症的生长。在一个方面,本公开提供了表达本发明sir的重组免疫效应细胞,其用于治疗或预防表达如本文描述的疾病相关抗原的疾病。在一个方面,本公开的表达sir的细胞能够使引起疾病细胞或疾病相关细胞与在其表面上表达的至少一种疾病相关抗原接触,以使得表达sir的细胞靶向引起疾病细胞或疾病相关细胞并抑制疾病的生长。在一个实施方式中,本公开涉及一种抑制疾病(例如癌症、自身免疫疾病、感染疾病或过敏性疾病或退行性疾病)的生长的方法,该方法包括使引起疾病细胞或疾病相关细胞与本发明的表达sir的细胞接触,使得sirt响应于该抗原而活化并且靶向引起疾病细胞或疾病相关细胞,其中该引起疾病细胞或疾病相关细胞的生长受到抑制。在一个方面,本公开涉及一种预防疾病的方法,该方法包括向处于疾病风险下的患者给予表达sir的细胞或能够生成本发明的表达sir的细胞的细胞,使得sirt响应于该抗原而活化并且靶向引起疾病细胞或疾病相关细胞,其中该引起疾病细胞或疾病相关细胞的生长得到预防。在一个方面,该疾病是癌症、感染疾病、免疫疾病、过敏性疾病或退行性疾病。在另一个实施方式中,本公开涉及一种治疗受试者的癌症的方法。该方法包括向受试者给予本发明的表达sir的细胞,以使得治疗受试者的癌症。在一个方面,与如本文描述的癌症相关抗原的表达相关的癌症是血液癌症或血液学癌症。在一个方面,血液学癌症是白血病或淋巴瘤。在一个方面,与如本文描述的癌症相关抗原的表达相关的癌症包括癌症和恶性肿瘤,包括但不限于例如一种或多种急性白血病,包括但不限于例如b细胞急性淋巴性白血病(“ball”)、pre-b细胞急性淋巴细胞性白血病、t细胞急性淋巴性白血病(“tall”)、急性淋巴性白血病(all);一种或多种慢性白血病,包括但不限于例如慢性髓细胞性白血病(cml)、慢性淋巴性白血病(cll)。与如本文描述的癌症相关抗原的表达相关的另外癌症或血液病状包括但不限于例如b细胞幼淋巴细胞性白血病、母细胞性浆细胞样树突细胞肿瘤、伯基特淋巴瘤、弥漫性大b细胞淋巴瘤、滤泡性淋巴瘤、毛细胞白血病、小细胞或大细胞滤泡性淋巴瘤、恶性淋巴组织增生性病状、malt淋巴瘤、套细胞淋巴瘤、边缘区淋巴瘤、多发性骨髓瘤、骨髓增生异常和骨髓增生异常综合征、非霍奇金淋巴瘤、浆母细胞性淋巴瘤、浆细胞样树突细胞肿瘤、瓦尔登斯特伦巨球蛋白血症、以及为由骨髓血细胞的无效产生(或发育异常)联合在一起的各种血液病状集合的“白血病前期”等。与如本文描述的癌症相关抗原表达相关的其它疾病包括但不限于例如与如本文描述的癌症相关抗原的表达相关的非典型和/或非经典癌症、恶性肿瘤、癌前病状或增生性疾病。在另一个方面,本公开涉及一种治疗受试者的疾病的方法。该方法包括向受试者给予本发明的表达sir的细胞,以使得治疗受试者的疾病。在一个方面,与如本文描述的疾病相关抗原的表达相关的疾病是感染疾病。在一个方面,该感染疾病是与以下病毒的感染相关的疾病:hiv1、hiv2、htlv1、爱泼斯坦巴尔病毒(ebv)、巨细胞病毒(cmv)、腺病毒、腺相关病毒、bk病毒(ebv)、人类疱疹病毒6、人类疱疹病毒8、甲型流感病毒、乙型流感病毒、副流感病毒、禽流感病毒、mers和sars冠状病毒、克里米亚刚果出血热病毒、鼻病毒、肠道病毒、登革病毒、西尼罗河病毒、埃博拉病毒、马尔堡病毒、拉沙热病毒、塞卡病毒、rsv、麻疹病毒、腮腺炎病毒、鼻病毒、水痘病毒、单纯疱疹病毒1和2、水痘-带状疱疹病毒、hiv-1、htlv1、肝炎病毒、肠道病毒、乙型肝炎病毒、丙型肝炎病毒、尼帕病毒和裂谷热病毒、日本脑炎病毒、结核分枝杆菌、非典型分支杆菌种类、耶氏肺孢子虫、弓形体病、立克次氏体、诺卡氏菌属、曲霉属、毛霉菌或假丝酵母。在另一个方面,本公开涉及一种治疗受试者的疾病的方法。该方法包括向受试者给予本发明的表达sir的细胞,以使得治疗受试者的疾病。在一个方面,与如本文描述的疾病相关抗原的表达相关的疾病是免疫或过敏性或生殖性疾病。在一个方面,免疫疾病或退行性疾病是糖尿病、多发性硬化、类风湿关节炎、寻常天疱疮、强直性脊柱炎、下本氏甲状腺炎(hoshimoto’sthyroiditis)、sle、结节病、硬皮病、混合型结缔组织病、移植物抗宿主病、花生过敏、慢性自发性荨麻疹、食物过敏、枯草热、季节性过敏、花粉过敏、hlh(噬血细胞淋巴组织细胞增多症)、淀粉样变性或阿尔茨海默病。在一些实施方式中,可以用本发明的表达sir的细胞治疗或预防的癌症是多发性骨髓瘤。多发性骨髓瘤是特征在于骨髓中的浆细胞克隆的积累的癌症。用于多发性骨髓瘤的当前疗法包括但不限于使用萨力多胺类似物来那度胺进行的治疗。来那度胺具有以下活性,包括抗肿瘤活性、血管生成抑制和免疫调节。通常,通过流式细胞术,骨髓瘤细胞被认为对于如本文描述的癌症相关抗原cd19呈阴性。因此,在一些实施方式中,例如如本文描述的cd19sir可以用于靶向骨髓瘤细胞。在一些实施方式中,本发明的sir疗法可以与一种或多种另外的疗法(例如来那度胺治疗)组合使用。本发明描述的其它sir(例如bcma-sir、cd138-sir、csi-sir、gprc5d-sir等)也可以用于治疗或预防多发性骨髓瘤。本公开包括一种类型的细胞疗法,其中免疫效应细胞(例如t细胞或可引起t细胞的干细胞)被遗传修饰以表达合成抗原受体(sir),并且将表达sir的t细胞或干细胞输注给有需要的受体。输注的细胞能够杀伤受体中的疾病相关细胞(例如肿瘤细胞或病毒感染细胞)。与抗体疗法不同,sir修饰的免疫效应细胞(例如t细胞、干细胞)能够在体内复制,从而产生可以导致持续的肿瘤控制的长期持久性。在各个方面,在向患者给予t细胞或干细胞之后,向患者给予的免疫效应细胞(例如t细胞或可引起t细胞的干细胞)或其后代在患者中持续至少四个月、五个月、六个月、七个月、八个月、九个月、十个月、十一个月、十二个月、十三个月、十四个月、十五个月、十六个月、十七个月、十八个月、十九个月、二十个月、二十一个月、二十二个月、二十三个月、两年、三年、四年、或五年。本公开还包括一种类型的细胞疗法,其中例如通过体外转录的rna修饰免疫效应细胞(例如t细胞)以瞬时表达合成抗原受体(sir),并且将sirt细胞输注给有需要的受体。输注的细胞能够杀伤受体中的疾病相关细胞(例如肿瘤细胞或病毒感染细胞)。因此,在不同方面,在向患者给予t细胞之后,向患者给予的免疫效应细胞(例如t细胞)存在少于一个月,例如三周、两周、一周。本公开还包括一种类型的细胞疗法,其中能够产生免疫效应细胞(例如t细胞)的干细胞(例如造血干细胞或淋巴样干细胞或胚胎干细胞或诱导型多能干细胞)被修饰以表达合成抗原受体(sir)并且向有需要的受体给予这些干细胞。所给予的干细胞在植入到受体中之后产生免疫效应细胞(例如t细胞),其(即免疫效应细胞)能够杀伤受体中的疾病相关细胞。因此,在各个方面,在向患者给予t细胞或干细胞之后,在给予表达sir的干细胞之后在患者中产生的免疫效应细胞(例如t细胞)在患者中持续至少一周、两周、三周、一个月、两个月、三个月、四个月、五个月、六个月、七个月、八个月、九个月、十个月、十一个月、十二个月、十三个月、十四个月、十五个月、十六个月、十七个月、十八个月、十九个月、二十个月、二十一个月、二十二个月、二十三个月、两年、三年、四年、五年、十年或二十年。本公开还包括一种类型的细胞疗法,其中能够产生免疫效应细胞(例如t细胞)的干细胞被修饰以表达合成抗原受体(sir)并且在体外分化以生成向有需要的受体输注的免疫效应细胞。输注的免疫效应细胞(例如t细胞)在输注到受体之后能够杀伤受体中的疾病相关细胞。因此,在各个方面,向患者给予的中产生的免疫效应细胞(例如t细胞)在患者中持续至少1天、2天、3天、4天、5天、6天、一周、两周、三周、一个月、两个月、三个月、四个月、五个月、六个月、七个月、八个月、九个月、十个月、十一个月、十二个月、十三个月、十四个月、十五个月、十六个月、十七个月、十八个月、十九个月、二十个月、二十一个月、二十二个月、二十三个月、两年、三年、四年、五年、十年或二十年。本公开还包括一种类型的细胞疗法,其中免疫效应细胞(例如t细胞)被修饰以表达编码自身抗原(例如dsg3或dsg1)的sir。本公开的此类表达自身抗原的sir可以用于消除表达针对自身抗原的自身抗体的b细胞和浆细胞。此类自身抗原-sir可以用于治疗和预防自身免疫病症,诸如寻常天疱疮。本公开还包括一种类型的细胞疗法,其中调控性免疫效应细胞(例如treg或cd25+t细胞)被修饰以表达靶向特异性抗原的sir。向患者给予此类sir-treg以抑制针对特异性抗原的免疫反应。sir-treg可以用于预防并治疗自身免疫疾病并增强免疫耐受性。在不希望受到任何特定理论约束的情况下,由sir修饰的免疫效应细胞(例如t细胞)引发的抗肿瘤免疫力反应可以是主动或被动免疫反应,或者可替代地可以是由于直接对比间接免疫反应。在一个方面,sir转导的免疫效应细胞(例如,t细胞)表现出响应于表达如本文描述的疾病相关抗原的人类患病细胞(例如癌症或患病细胞)的特异性促炎性细胞因子分泌和高效细胞溶解活性,抵抗如本文描述的可溶性疾病相关抗原,介导旁观者(bystander)杀伤并介导已确立的人类肿瘤(包括癌症)的消退。例如,表达如本文描述的癌症相关抗原的肿瘤的异质区域内的少抗原肿瘤细胞可能易受先前已经针对邻近的抗原阳性癌细胞反应的如本文描述的癌症相关抗原重定向的免疫效应细胞(例如,t细胞)的间接破坏。在一个方面,本公开的完全人类sir修饰的免疫效应细胞(例如,t细胞)可以是用于哺乳动物的离体免疫和/或体内疗法的一类疫苗。在一个方面,哺乳动物是人。在一个方面,哺乳动物是狗。关于离体免疫,在体外在向哺乳动物给予该细胞之前发生以下至少一种:i)扩增细胞、ii)将编码sir的核酸引入细胞或iii)将这些细胞低温保存。离体程序在本领域中是熟知的,并在下文更全面地讨论。简言之,从哺乳动物(例如,人类)分离细胞,并用表达本文公开的sir的载体进行遗传修饰(即体外转导或转染)。可以将sir修饰的细胞给予至哺乳动物受体以提供治疗益处。哺乳动物受体可以是人类,并且sir修饰的细胞可以相对于受体是自体的。可替代地,相对于接受者,细胞可以是同种异体的、同基因的或异基因的。在另一个实施方式中,离体使用sir修饰的细胞以清除疾病相关细胞(例如癌症细胞)的骨髓或外周血造血干细胞。作为实例,表达cd19-sir的t细胞与从患有急性淋巴细胞白血病或非霍奇金淋巴瘤的患者获取的骨髓或外周血干细胞样品共同培养,以便杀伤掉存在于骨髓或外周血干细胞制备物中的白血病或淋巴瘤。在体外(离体)培养适合持续时间(该持续时间范围可为从6小时至若干天)之后,将清除的骨髓和外周血样品用于在患者中自体移植。造血干细胞和祖细胞的离体扩增程序描述于美国专利号5,199,942(该专利通过引用结合在本文中)中,其可以应用于本发明的细胞。其它合适的方法在本领域中是已知的,因此本发明不限于离体扩增细胞的任何具体方法。简言之,免疫效应细胞(例如t细胞)的离体培养和扩增包括:(1)从哺乳动物外周血收获物或骨髓外植体收集cd34+造血干细胞和祖细胞;以及(2)离体扩增此类细胞。除了美国专利号5,199,942中描述的细胞生长因子外,其它因子诸如flt3-l、il-1、il-3和c-kit配体也可以用于培养和扩增细胞。除了在离体免疫方面使用基于细胞的疫苗之外,本发明还提供了用于体内免疫以引发针对患者中的抗原的免疫反应的组合物和方法。通常,如本文描述活化和扩增的细胞可用于治疗和预防免疫受损个体中出现的疾病。特别地,本公开的sir修饰的免疫效应细胞(例如,t细胞)用于治疗与如本文描述的疾病相关抗原(例如癌症抗原或病毒抗原)的表达相关的疾病、病症和病状。在某些方面,本公开的细胞用于治疗处于发展与如本文描述的疾病相关抗原的表达相关的疾病、病症和病状的风险下的患者。因此,本公开提供了用于治疗或预防与如本文描述的疾病相关抗原的表达相关的疾病、病症和病状的方法,这些方法包括向有需要的受试者给予治疗有效量的本公开的sir修饰的免疫效应细胞(例如,t细胞)或能够生成免疫效应细胞的干细胞。在一个方面,本公开的表达sir的细胞可用于治疗增生性疾病,诸如癌症或恶性肿瘤、或是癌前病状(诸如骨髓增生异常、骨髓增生异常综合征或白血病前期)。与如本文描述的癌症相关抗原表达相关的其它疾病包括但不限于例如非典型和/或非经典癌症、恶性肿瘤、癌前病状或表达如本文描述的癌症相关抗原的增生性疾病。与如本文描述的疾病相关抗原的表达相关的非癌症相关适应症包括但不限于例如自身免疫性疾病(例如狼疮)、炎性病症(过敏症和哮喘)、感染性病状(例如hiv1、cmv、ebv、流感)和移植。本公开的sir修饰的免疫效应细胞(例如,t细胞)可以单独给予或作为药物组合物与稀释剂和/或与其它组分(诸如il-2或其它细胞因子或细胞群体)组合给予。血液学癌症或血液癌症病状是癌症的类型,诸如白血病、淋巴瘤以及影响血液、骨髓和淋巴系统的恶性淋巴组织增生性病状。白血病可分为急性白血病和慢性白血病。急性白血病可进一步分类为急性骨髓性白血病(aml)和急性淋巴性白血病(all)。慢性白血病包括慢性骨髓性白血病(cml)和慢性淋巴性白血病(cll)。其它相关病症包括骨髓增生异常综合征(mds,以前称为“白血病前期”),这是由髓系血细胞的无效产生(或发育不良)联合的血液学病症的多样化集合,并且具有转化为aml的风险。淋巴瘤是一组从淋巴细胞发展的血细胞肿瘤。示例性淋巴瘤包括非霍奇金淋巴瘤和霍奇金淋巴瘤。本发明提供用于治疗和预防癌症的组合物和方法。在一个方面,癌症是血液学癌症或血液癌症,包括但不限于血液学癌症,其为白血病或淋巴瘤。在一个方面,本公开的表达sir的细胞可用于治疗癌症和恶性肿瘤,诸如但不限于例如急性白血病,包括但不限于例如b细胞急性淋巴性白血病(“ball”)、t细胞急性淋巴性白血病(“tall”)、急性淋巴性白血病(all);一种或多种慢性白血病,包括但不限于例如慢性骨髓性白血病(cml)、慢性淋巴细胞白血病(cll);额外血液学癌症或血液学病状,包括但不限于例如b细胞幼淋巴细胞白血病、母细胞性浆细胞样树突细胞肿瘤、伯基特氏淋巴瘤、弥漫性大b细胞淋巴瘤、滤泡性淋巴瘤、毛细胞白血病、小细胞或大细胞滤泡性淋巴瘤、恶性淋巴组织增生性病状、malt淋巴瘤、套细胞淋巴瘤、边缘区淋巴瘤、多发性骨髓瘤、骨髓增生异常和骨髓增生异常综合征、非霍奇金淋巴瘤、浆母细胞性淋巴瘤、浆细胞样树突状细胞瘤、瓦尔登斯特伦巨球蛋白血症以及为由骨髓血细胞的无效产生(或发育异常)联合在一起的各种血液病状集合的“白血病前期”等。与如本文描述的癌症相关抗原表达相关的其它疾病包括但不限于例如非典型和/或非经典癌症、恶性肿瘤、癌前病状或表达如本文描述的癌症相关抗原的增生性疾病。本公开还提供了用于抑制表达如本文描述的疾病相关抗原的细胞群体的增殖或减少该细胞群体的方法,这些方法包括使包含表达如本文描述的疾病相关抗原的细胞的细胞群体与本公开的表达sir的t细胞接触,该表达sir的t细胞结合至表达如本文描述的疾病相关抗原的细胞。在一个具体方面,本发明提供了用于抑制表达如本文描述的疾病相关抗原的患病细胞群体的增殖或减少该患病细胞群体的方法,这些方法包括使表达如本文描述的疾病相关抗原的癌细胞群体与本公开的表达sir的t细胞接触,该表达sir的t细胞结合至表达如本文描述的疾病相关抗原的细胞。在一个方面,本发明提供了用于抑制表达如本文描述的疾病相关抗原的患病细胞群体的增殖或减少该患病细胞群体的方法,这些方法包括使表达如本文描述的疾病相关抗原的患病细胞群体与本公开的表达sir的t细胞接触,该表达sir的t细胞结合至表达如本文描述的疾病相关抗原的细胞。在某些方面,在患有骨髓性白血病或与表达如本文描述的疾病相关抗原的细胞相关的另一种疾病的受试者或骨髓性白血病或与表达如本文描述的疾病相关抗原的细胞相关的另一种疾病的动物模型中,相对于阴性对照,本公开的表达sir的t细胞使细胞和/或患病细胞的数量(quantity)、数目(number)、量(amount)或百分比减少至少25%、至少30%、至少40%、至少50%、至少65%、至少75%、至少85%、至少95%、或至少99%。在一方面,受试者是人类。在一个方面,该疾病是癌症、感染疾病、免疫疾病、过敏症或退行性疾病。本发明还提供了用于预防、治疗和/或管理与表达如本文描述的疾病相关抗原的细胞相关的疾病(例如,表达如本文描述的疾病相关抗原的血液癌症或非典型癌症或感染疾病或免疫疾病或过敏性疾病或退行性疾病)的方法,这些方法包括向有需要的受试者给予本公开的sirt细胞,该sirt细胞结合至表达如本文描述的疾病相关抗原的细胞。在一方面,受试者是人类。与表达如本文描述的疾病相关抗原的细胞相关的病症的非限制性实例包括自身免疫性病症(诸如狼疮)、炎性病症(诸如过敏症和哮喘)、感染(诸如hiv1、htlv1、流感病毒、cmv、腺病毒、ebv和hhv8)和癌症(诸如表达如本文描述的癌症相关抗原的血液学癌症或非典型癌症)。本公开还提供了用于预防、治疗和/或管理与表达如本文描述的疾病相关抗原的细胞相关的疾病的方法,这些方法包括向有需要的受试者给予本公开的sirt细胞,该sirt细胞结合至表达如本文描述的疾病相关抗原的细胞。在一方面,受试者是人类。本公开提供了用于预防与表达如本文描述的疾病相关抗原的细胞相关的癌症的复发的方法,这些方法包括向有需要的受试者给予本公开的sirt细胞,该sirt细胞结合至表达如本文描述的疾病相关抗原的细胞。在一个方面,这些方法包括向有需要的受试者给予有效量的本文描述的表达sir的t细胞与有效量的另一种疗法的组合,该表达sir的t细胞结合至表达如本文描述的疾病相关抗原的细胞。本公开还提供一种治疗或预防患有与靶抗原的表达相关的疾病或具有增加的该疾病的风险的受试者的疾病的方法,该方法包括向该受试者给予有效量的包含sir分子的细胞。本公开还提供一种治疗患有与靶抗原的表达相关的疾病或具有增加的该疾病的风险的受试者的疾病或预防该受试者的疾病的方法,该方法包括向该受试者给予有效量的包含sir分子的细胞,例如免疫效应细胞(例如免疫效应细胞群体),其中sir分子包含一个或多个抗原结合结构和一条或多条t细胞受体恒定链,其中所述抗原结合结构域结合至与该疾病相关的靶抗原。本文上文公开了靶抗原的非限制性实例。本公开提供了一种向受试者给予细胞例如免疫效应细胞或其群体(每个细胞包含sir分子)任选地与增加免疫细胞的功效和/或安全性的组合的方法。在不同实施方式中,增加免疫细胞的功效和/或安全性的药剂选自由以下组成的组:(i)蛋白质磷酸酶抑制剂;(ii)激酶抑制剂;(iii)细胞因子;(iv)免疫抑制分子的抑制剂;(v)降低treg细胞的水平或活性的药剂;(vi)增加sir修饰细胞的增殖和/或持久性的药剂;(vii)趋化因子;(viii)增加sir的表达的药剂;(ix)允许调控sir的表达或活性的药剂;(x)允许控制sir修饰细胞的存活和/或持久性的药剂;(xi)控制sir修饰细胞的副作用的药剂;(xii)brd4抑制剂;(xiii)将治疗剂(例如shvem)或预防剂递送至疾病位点的药剂;(xiv)增加sir所针对的靶抗原的表达的药剂;(xv)腺苷a2a受体拮抗剂;以及(xvi)(i)-(xv)的任何组合。在一些实施方式中,有待治疗或预防的疾病是血液学癌症。在其它实施方式中,血液学癌症为白血病。急性白血病的非限制性实例包括但不限于b细胞急性淋巴性白血病(“ball”)、t细胞急性淋巴性白血病(“tall”)、急性淋巴性白血病(all);一种或多种慢性白血病,包括但不限于慢性骨髓性白血病(cml)、慢性淋巴细胞白血病(cll);额外血液学癌症或血液学病状,包括但不限于b细胞幼淋巴细胞性白血病、母细胞性浆细胞样树突细胞肿瘤、伯基特淋巴瘤、弥漫性大b细胞淋巴瘤、滤泡性淋巴瘤、毛细胞白血病、小细胞或大细胞滤泡性淋巴瘤、恶性淋巴组织增生性病状、malt淋巴瘤、套细胞淋巴瘤、边缘区淋巴瘤、多发性骨髓瘤、骨髓增生异常和骨髓增生异常综合征、非霍奇金淋巴瘤、霍奇金淋巴瘤、浆母细胞性淋巴瘤、浆细胞样树突细胞肿瘤、瓦尔登斯特伦巨球蛋白血症、以及为由骨髓血细胞的无效产生(或发育异常)联合在一起的各种血液病状集合的“白血病前期”,并且与本文描述的肿瘤抗原的表达相关的疾病包括但不限于表达如本文描述的肿瘤抗原的非典型和/或非经典癌症、恶性肿瘤、癌前病状或增生性疾病;以及其任何组合。在另一个实施方式中,与本文描述的肿瘤抗原相关的疾病是实体瘤。在一些实施方式中,与该疾病相关的肿瘤抗原选自cd5、cd19、cd123、cd22、cd23、cd30、cd171、cs-1、cll-1(clecl1)、cd33、egfrviii、gd2、gd3、bcma、tnag、psma、ror1、flt3、tag72、cd38、cd44v6、cea、epcam、b7h3、kit、il-13ra2、间皮素、il-11ra、psca、prss21、vegfr2、lewisy、cd24、pdgfr-β、ssea-4、cd20、叶酸受体α、erbb2(her2/neu)、muc1、egfr、ncam、蛋白酶、pap、elf2m、肝配蛋白b2、fap、igf-i受体、calx、lmp2、gpl00、bcr-abl、酪氨酸酶、epha2、fucosylgm1、sle、gm3、tgs5、hmwmaa、o-乙酰基-gd2、叶酸受体β、tem1/cd248、tem7r、cldn6、tshr、gprc5d、cxorf61、cd97、cd179a、alk、聚唾液酸、plac1、globoh、ny-br-1、upk2、havcr1、adrb3、panx3、gpr20、ly6k、or51e2、tarp、wt1、ny-es0-1、lage-1a、mage-a1、magea1、etv6-aml、精子蛋白17、xage1、tie2、mad-ct-1、mad-ct-2、fos相关抗原1、p53、p53突变体、蛋白质(前列腺特异性蛋白)、存活素和端粒酶、pcta-1/半乳凝素8、melana/mart1、ras突变体、htert、肉瘤易位裂点、ml-iap、erg(tmprss2ets融合基因)、na17、pax3、雄激素受体、周期素b1、mycn、rhoc、trp-2、cyp1b1、boris、sart3、pax5、oy-tes1、lck、akap-4、ssx2、rage-1、人类端粒酶逆转录酶、rul、ru2、豆荚蛋白、hpve6、e7、肠羧基脂酶、muthsp70-2、cd79a、cd79b、cd72、lair1、fcar、lilra2、cd300lf、clec12a、bst2、emr2、ly75、fcrl5、iglli、mpl、、fitc、生物素、c-myc表位标签、cd34、lamp1、trop2、gfrα4、cdh17、cdh6、nybr1、cdh19、cd200r、slea(ca19.9;唾液酸路易斯抗原)、ptk7、gpnmb、cdh1-cd324、dll3、cd276/b7h3、il11ra、il13ra2、cd179b-igll1、alk、tcrγ-δ、nkg2d、cd32(fcgr2a)、tnag、cspg4-hmw-maa、tim1-/hvcr1、csf2ra(gm-csfr-α)、tgfβr2、vegfr2/kdr、lewsag、tcr-β1链、tcr-β2链、tcr-γ链、tcr-δ链、fitc、促黄体生成激素受体(lhr)、ccr4、gd3、glypican-3(gpc3)、slamf6、slamf4、htlv1-tax、ebv-ebna3c、hla、hla-a、hla-a2、hla-b、hla-c、hla-dp、hla-dm、hla-doa、hla-dob、hla-dq、hla-dr、hla-g、ige、cd99、rasg12v、组织因子1(tf1)、afp、gprc5d、密封蛋白18.2(cld18a2或cldn18a.2))、p-糖蛋白、steap1、liv1、粘连蛋白-4、cripto、gpa33、bst1/cd157、低电导率氯离子通道、由tnt抗体识别的抗原、tshr、cd171、cs-1、cll-1、gd3、tnag、flt3、cd38、cd44v6、在急性白血病或淋巴瘤上表达但不在造血祖细胞上表达的糖基化cd43表位、在非造血癌症上表达的糖基化cd43表位、b7h3、kit、il-13ra2、il-llra、psca、prss21、vegfr2、lewisy、cd24、pdgfr-β、ssea-4、muc1、egfr、ncam、calx、lmp2、epha2、岩藻糖基gm1、sle、gm3、tgs5、hmwmaa、o-乙酰基-gd2、叶酸受体β、tem1/cd248、tem7r、cldn6、gprc5d、cxorf61、cd97、cd179a、alk、聚唾液酸、plac1、globoh、ny-br-1、upk2、havcr1、adrb3、panx3、gpr20、ly6k、or51e2、tarp、wt1、etv6-aml、精子蛋白17、xage1、tie2、mad-ct-1、mad-ct-2、fos相关抗原1、p53突变体、htert、肉瘤易位裂点、ml-iap、erg(tmprss2ets融合基因)、na17、pax3、雄激素受体、周期素b1、mycn、rhoc、cyp1b1、boris、sart3、pax5、oy-tes1、lck、akap-4、ssx2、cd79a、cd79b、cd72、lair1、fcar、lilra2、cd300lf、clec12a、bst2、emr2、ly75、gpc3、fcrl5、iglli、tshr、cldn6、gprc5d、cxorf61、cd97、cd179a、alk、聚唾液酸、plac1、globoh、ny-br-1、upk2、havcr1、adrb3、panx3、gpr20、ly6k、以及or51e2。在一些实施方式中,有待治疗的疾病是感染疾病,包括但不限于以下各项的感染:hiv1、hiv2、htlv1、爱泼斯坦巴尔病毒(ebv)、巨细胞病毒(cmv)、腺病毒、腺相关病毒、bk病毒(ebv)、人类疱疹病毒6、人类疱疹病毒8、流感病毒、副流感病毒、禽流感病毒、mers和sars冠状病毒、克里米亚刚果出血热病毒、鼻病毒、肠道病毒、登革病毒、西尼罗河病毒、埃博拉病毒、马尔堡病毒、拉沙热病毒、塞卡病毒、rsv、麻疹病毒、腮腺炎病毒、鼻病毒、水痘病毒、单纯疱疹病毒1和2、水痘带状疱疹病毒、hiv-1、htlv1、肝炎病毒、肠道病毒、乙型肝炎病毒、丙型肝炎病毒、尼帕病毒和裂谷热病毒、日本脑炎病毒、结核分枝杆菌、非典型分支杆菌种类、耶氏肺孢子虫、弓形体病、立克次氏体、诺卡氏菌属、曲霉属、毛霉菌或假丝酵母。在此类疾病中,与该疾病相关的靶抗原选自hiv1包膜糖蛋白、hiv1-gag、htlv1-tax、cmvpp65、ebv-ebna3c、甲型流感血球凝集素(ha)和gad。有待通过本公开的方法和组合物治疗或预防的疾病可以是免疫或退行性疾病,例如糖尿病、多发性硬化症、风湿性关节炎、寻常性天疱疮、强直性脊柱炎、下本氏甲状腺炎、sle、结节病、硬皮病、混合型结缔组织病、移植物抗宿主病或阿尔茨海默病。在此类实施方式中,与该疾病相关的靶抗原是自身抗体。为sir的适合靶标的示例性自身抗体是针对dsg3或dsg1的自身抗体。与靶抗原的表达相关的疾病的其它非限制性实例包括以下癌症或相关病状中的任一种:结肠癌、直肠癌、肾细胞癌、肝癌、非小细胞肺癌、小肠癌、食道癌、黑素瘤、骨癌、胰腺癌、皮肤癌、头颈癌、皮肤或眼内恶性黑素瘤、子宫癌、卵巢癌、直肠癌、肛区癌、胃癌、睾丸癌、子宫癌、输卵管癌、子宫内膜癌、子宫颈癌、阴道癌、外阴癌、霍奇金病、非霍奇金淋巴瘤、内分泌系统癌症、甲状腺癌、甲状旁腺癌、肾上腺癌、软组织肉瘤、尿道癌、阴茎癌、儿童实体瘤、膀胱癌、肾或输尿管癌、肾盂癌、中枢神经系统肿瘤(cns)、原发性cns淋巴瘤、肿瘤血管生成、脊轴肿瘤、脑干胶质瘤、垂体腺瘤、卡波济肉瘤、表皮样癌、鳞状细胞癌、t细胞淋巴瘤、环境诱导的癌症、所述癌症的组合、以及所述癌症的转移性病灶。在本文所述的方法或用途的某些实施方式中,sir分子与增加免疫效应细胞的功效的药剂组合给予,该药剂例如以下一种或多种:蛋白磷酸酶抑制剂、激酶抑制剂、细胞因子、趋化因子、scfv片段、双特异性抗体、免疫抑制性分子的抑制剂;细胞信号传导蛋白、病毒信号传导到、或降低treg细胞的水平或活性的药剂。蛋白磷酸酶抑制剂的非限制性实例包括shp-1抑制剂和/或shp-2抑制剂。激酶抑制剂的非限制性实例包括:cdk4抑制剂、cdk4/6抑制剂(例如,帕博西尼)、btk抑制剂(例如,依鲁替尼或rn-486)、mtor抑制剂(例如,雷帕霉素或依维莫司(rad001))、mnk抑制剂或双重p13k/mtor抑制剂。在一个实施方式中,btk抑制剂不会降低或抑制白细胞介素2诱导型激酶(itk)的激酶活性。a2a受体拮抗剂的非限制性实例包括vipadenant。在一些实施方式中,抑制免疫抑制性分子的药剂可以是以下一种或多种:抗体或抗体片段、抑制性核酸、聚集的规则间隔短回文重复序列(crispr)、转录活化因子样效应核酸酶(talen)或抑制抑制性分子的表达的锌指核酸内切酶(zfn)。在本文描述的方法或用途的其它实施方式中,降低treg细胞的水平或活性的药剂选自环磷酰胺、抗gitr抗体、cd25耗尽或其组合。在本文描述的方法或用途的某些实施方式中,免疫抑制性分子选自由以下组成的组:pd1、pd-l1、ctla-4、tim-3、lag-3、vista、btla、tigit、lair1、cd160、2b4、tgfβ、ceacam-1、ceacam-3、以及ceacam-5。在其它实施方式中,细胞因子选自il2、il-7、il-15或il-21、或两者。在其它实施方式中,包含sir分子的免疫效应细胞和第二例如本文公开的任何组合疗法(例如,增加免疫效应细胞的功效的药剂)基本上同时或依次给予。在其它实施方式中,抑制抑制性分子的药剂包含含有抑制性分子的第一多肽或其片段以及向细胞提供阳性信号的第二多肽,并且其中该第一多肽和第二多肽在含有sir的免疫细胞上表达,其中(i)该第一多肽包含pd1、pd-l1、ctla-4、tim-3、lag3、vista、btla、tigit、lair1、cd160、2b4、tgfβ、ceacam-1、ceacam-3、以及ceacam-5或其片段;和/或(ii)该第二多肽包含含有初级信号传导结构域和/或共刺激信号传导结构域的细胞内信号传导结构域。在一个实施方式中,初级信号传导结构域包含cd3ζ的功能结构域,和/或共刺激信号传导结构域包含选自41bb、cd27和cd28的蛋白质的功能结构域。在一个实施方式中,淋巴细胞输注(例如同种异体淋巴细胞输注)用于治疗癌症、感染或免疫疾病,其中淋巴细胞输注包含至少一种本公开的表达sir的细胞。在一个实施方式中,自体淋巴细胞输注用于治疗癌症、感染或免疫疾病,其中自体淋巴细胞输注包含至少一种本文描述的表达sir的细胞。在一个实施方式中,该方法包括给予如本文描述的表达sir分子的细胞与增强表达sir的细胞的活性的药剂的组合,其中该药剂是细胞因子,例如il-2、il-7、il-15、il-21或其组合。可以与给予表达sir的细胞组合,例如同时或在给予该细胞之后不久递送细胞因子。可替代地,可以在给予表达sir的细胞后延长的时间段后,例如在评估受试者对表达sir的细胞的应答之后递送细胞因子。在一个实施方式中,将细胞因子与给予根据权利要求143至161中任一项所述的细胞或细胞群体同时(例如,在同一天给予)或在给予该细胞或细胞群体之后不久(例如,在给予该细胞或细胞群体之后给予1天、2天、3天、4天、5天、6天或7天)给予至受试者。在其它实施方式中,将细胞因子在细胞或细胞群体之后或在评估受试者对细胞的反应之后延长的时间段(例如,例如,至少2周、3周、4周、6周、8周、10周或更长时间)后给予至受试者。在其它实施方式中,将表达sir分子的细胞与改善与表达sir分子的细胞的给予相关的一种或多种副作用的药剂组合给予。与表达sir的细胞相关的副作用可以选自细胞因子释放综合征(crs)、噬血细胞性淋巴组织细胞增多症(hlh)或神经学并发症。此类药剂的实例包括类固醇(例如强的松、地塞米松)、il6r拮抗剂(例如托珠单抗)、src激酶抑制剂(例如达沙替尼)、激酶抑制剂(例如依鲁替尼)、钙调磷酸酶抑制剂(例如他克莫司或环孢霉素a)或化学疗法药物(例如环磷酰胺、甲氨蝶呤或长春碱)。在任何前述方法或用途的实施方式中,表达sir分子的细胞与治疗与肿瘤抗原的表达相关的疾病的药剂(例如本文公开的第二或第三疗法中的任一种)组合给予。另外的示例性组合包括以下中的一种或多种。在另一个实施方式中,表示sir分子的细胞(例如,如本文描述)可以与增加sir所针对的靶抗原的表达的另一种药剂组合给予。例如,经典霍奇金淋巴瘤的特征在于在b细胞中表达的基因的实际缺乏。据报道通过启动子超甲基化和组蛋白去乙酰化以及b细胞定型转录因子表达减少导致该疾病中丧失的b细胞表型。du,j等人鉴定促进b细胞表型在经典霍奇金淋巴瘤细胞中重新表达的化合物(化合物27、40、49)(blood[血液];在2016年10月12日在线重新公布)。还据报道抗白血病药物三氧化二砷和atra当单独使用或与鉴定的化合物27、40和49组合使用时促进b细胞表型在经典霍奇金淋巴瘤中重新表达。在一个实施方式中,表达靶向b细胞标记物诸如cd19、cd20、cd22等的sir的细胞可以与化合物27、40、49、三氧化二砷和atra中的一种或多种组合给予。在另一个实施方式中,本公开的表达sir的免疫效应细胞(例如t细胞、nk细胞或造血干细胞)连同破坏某些癌症肿瘤环境中的免疫抑制途径的药剂一起给予至受试者。在一个实施方式中,破坏癌症肿瘤微环境中的免疫抑制途径的药剂是腺苷a2a受体拮抗剂。可以连同本公开的表达sir的免疫效应细胞一起给予的示例性腺苷a2a受体拮抗剂是vipadenant。在一个实施方式中,将本公开的表达sir的免疫效应细胞(例如,t细胞、nk细胞或造血干细胞)给予至已接受先前干细胞移植(例如自体干细胞移植或同种异体干细胞移植)的受试者。在一个实施方式中,将本公开的表达sir的免疫效应细胞(例如,t细胞、nk细胞或造血干细胞)给予至已接受先前剂量的化学疗法(诸如美法仑、氟达拉滨或环磷酰胺)的受试者。在一个实施方式中,将本公开的表达sir的免疫效应细胞(例如t细胞、nk细胞或造血干细胞)给予至已接受先前剂量增强sir靶抗原的表达的药剂的受试者,所述药剂诸如化合物27、40和49(du,j等人,blood[血液],在2016年10月12日在线重新公布)、三氧化二砷或atra。在一个实施方式中,将表达sir分子(例如,本文描述的sir分子)的细胞与增加表达sir分子的细胞的功效的药剂(例如本文描述的药剂)组合给予。在一个实施方式中,将表达sir分子(例如,本文所述的sir分子)的细胞与低免疫增强剂量的mtor抑制剂组合给予。虽然不希望受到理论约束,但是据信使用低免疫增强剂量(例如,剂量不会完全抑制免疫系统但足以改善免疫功能)的治疗伴随pd-1阳性t细胞减少或pd-1阴性细胞增加。pd-1阳性t细胞而非pd-1阴性t细胞可以通过与表达pd-1配体(例如,pd-l1或pd-l2)的细胞接合而耗尽。动物模型也可用于测量sir活性。例如,可以使用异种移植模型,该异种移植模型使用本文描述的人类癌症相关抗原特异性sir+t细胞来治疗免疫缺陷小鼠中的初级人类pre-b-all。参见例如,milone等人,moleculartherapy[分子疗法]17(s):1453-1464(2009)。简而言之,在建立all后,将小鼠随机分配至治疗组。将不同数量的癌症相关抗原特异性sir(例如cd19-sir)工程化的t细胞以1:1比率共注射到携带b-all的nod-scid-γ-/-小鼠中。在t细胞注射后的不同时间,评价来自小鼠的脾dna中的癌症相关抗原特异性sir载体的拷贝的数量。以每周间隔评估动物的白血病。在注射了b-all-sir+t细胞或模拟转导的t细胞的小鼠中测量外周血母细胞计数。使用时序检验比较组的存活曲线。此外,还可以分析在nod-scid-γ-!-小鼠中t细胞注射后4周的绝对外周血cd4+和cd8+t细胞计数。给小鼠注射白血病细胞,并在3周后注射t细胞,这些t细胞经工程化以通过编码与egfp连接的sir的双顺反子慢病毒载体表达sir。通过在注射前与模拟物转导的细胞混合将t细胞标准化为45%-50%输入的gfp+t细胞,并通过流式细胞术确认。以1周为间隔评估动物的白血病。使用时序检验比较sir+t细胞组的存活曲线。可以评价剂量依赖性sir治疗应答。参见例如,milone等人,moleculartherapy[分子疗法]17(s):1453-1464(2009)。例如,在第21天用sirt细胞、相同数目的模拟物转导的t细胞、或无t细胞注射的小鼠中建立白血病之后35-70天获得外周血。将每组小鼠随机放血以确定外周血b+all母细胞计数,并且然后在第35和49天处死。在第57天和第70天评价剩余的动物。先前已经描述了对细胞增殖和细胞因子产生的评估,例如在milone等人,moleculartherapy[分子疗法]17(s):1453-1464(2009)中。简言之,通过将洗涤的t细胞与表达本文描述的疾病相关抗原(k19)或cd32和cd137(kt32-bbl)的k562细胞混合(最终t细胞:k562比率为2:1),在微量滴定板中进行sir介导的增殖的评估。在使用前用γ射线照射k562细胞。将抗cd3(克隆okt3)和抗cd28(克隆9.3)单克隆抗体添加至具有kt32-bbl细胞的培养物中以充当用于刺激t细胞增殖的阳性对照,因为这些信号支持长期cd8+t细胞离体扩增。使用countbrighttm荧光珠(加利福尼亚州卡尔斯巴德英杰公司(invitrogen,carlsbad,ca))和流式细胞术(如由制造商描述的)在培养物中计数t细胞。使用用egfp-2a连接的表达sir的慢病毒载体进行工程化的t细胞,通过gfp表达来鉴定sir+t细胞。对于不表达gfp的sir+t细胞,用生物素化的重组如本文描述的癌症相关抗原和次级抗生物素蛋白-pe缀合物检测sir+t细胞。还用特异性单克隆抗体(bd生物科学公司(bdbiosciences))同时检测t细胞上的cd4+和cd8+表达。根据制造商的说明书,使用人th1/th2细胞因子细胞计数珠阵列试剂盒(加利福尼亚州圣地牙哥bd生物科学公司(bdbiosciences,sandiego,ca))对再刺激后24小时收集的上清液进行细胞因子测量。使用facscalibur流式细胞仪评估荧光,并根据制造商的说明书分析数据。可通过标准51cr-释放测定来评估细胞毒性。参见例如,milone等人,moleculartherapy[分子疗法]17(8):1453-1464(2009)。简言之,在37℃将靶细胞(k562系和初级原b-all细胞)负载51cr(如nacro4,马萨诸塞州波士顿新英格兰核公司(newenglandnuclear,boston,ma))2小时伴随频繁搅拌,在完全rpmi中洗涤两次并铺板到微量滴定板中。将效应t细胞与完全rpmi的孔中的靶细胞以不同比例的效应细胞:靶细胞(e:t)混合。还制备了仅含有培养基(自发释放,sr)或1%triton-x100洗涤剂溶液(总释放,tr)的另外的孔。在37℃下孵育4小时后,收获来自每个孔的上清液。然后使用γ颗粒计数器(马萨诸塞州沃尔瑟姆帕卡德仪器公司(packardinstrumentco.,waltham,ma))测量释放的51cr。每种条件进行至少一式三份,并且使用下式计算溶解百分比:细胞溶解%=(er-sr)/(tr–sr),其中er表示每种实验条件释放的平均51cr。此测定或类似测定可以用于检测任何群体中sir细胞的存在。此测定也可以用于测量任何群体中sir细胞的扩增和持久性。成像技术可以用于评价携带肿瘤的动物模型中sir的特异性运输和增殖。例如barrett等人,humangenetherapy[人类基因疗法]22:1575-1586(2011)中已经描述了此类测定。简言之,nod/scid/γ-!-(nsg)小鼠iv注射nalm-6细胞,7天后在用编码sir的mrna电穿孔后4小时用t细胞注射。将t细胞用慢病毒构建体稳定转染以表达萤火虫荧光素酶,并将小鼠针对生物发光成像。可替代地,单次注射sir+t细胞在nalm-6异种移植模型中的治疗效果和特异性可以如下测量:向nsg小鼠注射被转导以稳定表达萤火虫荧光素酶的nalm-6,然后在7天后单次尾静脉注射用本公开的sir(例如cd19-sir;seqidno:1200)电穿孔的t细胞。在注射后的不同时间点对动物成像。例如,可以产生在第5天(治疗前2天)和第8天(sir+pbl后24小时)的代表性小鼠中萤火虫荧光素酶阳性白血病的光子密度热图。类似方法可以用于评价靶向其它癌症或其它疾病的sir。其它测定,包括本文实例部分中描述的那些测定以及本领域中已知的那些测定,也可以用于评价本文描述的sir。本公开的药物组合物可以包含表达sir的细胞(例如如本文描述的多种表达sir的细胞)与一种或多种药学上或生理学上可接受的载体、稀释剂或赋形剂的组合。此类组合物可以包含缓冲液,诸如中性缓冲盐水、磷酸盐缓冲盐水等;碳水化合物,诸如葡萄糖、甘露糖、蔗糖或葡聚糖、山梨醇;蛋白质;多肽或氨基酸,诸如甘氨酸;抗氧化剂;螯合剂,诸如edta或谷胱甘肽;佐剂(例如氢氧化铝);以及防腐剂。在一方面,本公开的组合物被配制用于静脉内给予。该组合物可以还包含第二活性剂(例如抗癌剂、抗病毒剂或抗生素剂)。本公开的药物组合物可以适于待治疗(或预防)的疾病的方式给予。给予的数量和频率将由例如患者的状况以及患者的疾病的类型和严重性等因素来确定。当指示“免疫有效量”、“抗肿瘤有效量”、“肿瘤抑制有效量”或“治疗量”或“抗感染”时,医生可以考虑到年龄、体重、肿瘤大小、感染或转移的程度以及患者(受试者)的状况的个体差异来确定待给予的本公开组合物的量。通常可以说,可以将包含本文描述的免疫效应细胞(例如,t细胞、nk细胞)的药物组合物以104至109个细胞/kg体重、在一些情况下105至106个细胞/kg体重(包括那些范围内的所有整数值)的剂量给予。还可以将t细胞组合物以这些剂量多次给予。可以通过使用免疫疗法中通常已知的输注技术来给予细胞(参见例如,rosenberg等人,neweng.j.ofmed.[新英格兰医学杂志]319:1676,1988)。在某些方面,可能希望向受试者给予活化的免疫效应细胞(例如,t细胞、nk细胞),并且然后随后重抽血液(或进行单采血液成分术),根据本公开从其活化免疫效应细胞(例如,t细胞、nk细胞),并用这些活化和扩增的免疫效应细胞(例如,t细胞、nk细胞)回输患者。该过程可以每隔几周进行多次。在某些方面,可以将来自从10cc至400cc抽血的免疫效应细胞(例如,t细胞、nk细胞)活化。在某些方面,将来自20cc、30cc、40cc、50cc、60cc、70cc、80cc、90cc、或100cc抽血的免疫效应细胞(例如,t细胞、nk细胞)活化。在一些实施方式中,受试者可以经历白细胞单采法,其中离体收集、富集、或耗尽白细胞以选择和/或分离感兴趣的细胞(例如t细胞)。这些t细胞分离物可以通过本领域已知的方法扩增并处理和/或转化,使得可以引入本发明的一个或多个sir构建体,从而产生本发明的sirt细胞。有需要的受试者随后可以接受高剂量化学疗法的标准治疗,然后进行外周血干细胞移植。在某些方面,在移植之后或与移植同时,受试者接受输注本发明的扩增的sirt细胞。在另一方面,在手术之前或之后给予扩增的细胞。还提供了实践本公开的试剂盒。例如,用于治疗受试者的癌症或制备表达一种或多种本文公开的sir的sirt细胞的试剂盒。试剂盒可包括编码sir的核酸分子或多肽分子或编码sir的载体以及将该核酸引入免疫效应细胞的方法。该试剂盒可包括包含编码sir的核酸和化学物质诸如聚凝胺的病毒以增强病毒转导。该试剂盒可含有用于分离表达sir的t细胞的组分。可替代地,该试剂盒可含有表达sir的免疫效应细胞(例如t细胞或nk细胞)或干细胞。试剂盒中包含多于一种公开的sir。该试剂盒可以包含容器以及在该容器上或与该容器相关联的标签或包装说明书。合适的容器包括例如瓶子、小瓶、注射器等。该容器可以由各种材料(诸如玻璃或塑料)形成。容器通常容纳包含核酸分子、病毒、载体、表达sir的t细胞中的一种或多种的组合物。在若干实施方式中,该容器可以具有无菌存取口(例如,该容器可以是静脉输液袋或具有可由皮下注射针刺穿的塞子的小瓶)。该标签或包装说明书指示该组合物用于治疗个体的特定病状。该标签或包装说明书通常将还包括关于所公开核酸分子、sir或表达sir的t细胞在治疗或预防肿瘤或制备sirt细胞的方法中使用的说明书。包装说明书是指在治疗产品的商业包装中通常包括的说明书,其含有关于使用此类治疗产品的适应症、用法、剂量、给药、禁忌症和/或警告的信息。说明性材料可以是书面的,呈电子形式(诸如计算机磁盘或高密度磁盘),或者可以是可视的(诸如视频文件)。该试剂盒还可包括另外组分以促进设计试剂盒针对的特定应用。因此,例如,该试剂盒可另外含有用于测量t细胞上的sir表达的手段或测量表达sir的t细胞的数目或百分比的手段或测定sirt细胞功能性的手段。所述试剂盒可另外包括缓冲剂和常规用于实施特定方法的其它试剂。此类试剂盒和适当内容物是本领域技术人员熟知的。通过参考以下实验实例进一步描述本公开。提供这些实施仅出于说明的目的,并且不旨在进行限制,除非另外说明。因此,本公开决不应被解释为限于以下实例,而是应该被解释为涵盖由于本文提供的传授而变得明显的任何和所有变化。实施例sir的活性可以通过若干种本文和下文所述的体外和体内测定来测试。下文提供了用于生成、选择和使用适合sir的一般方案。鉴定用于sir生成的靶标基于文库或基因表达数据库的搜索来选择设计sir所针对的适合靶标。一般而言,与正常健康细胞相比,sir的适合靶标在引起疾病细胞或疾病相关细胞上显示更高表达。sir的生成.一旦鉴定sir的候选靶抗原,就基于文库中可用的信息设计sir的抗原结合结构域。一般而言,sir的抗原结合结构域典型地是基于抗体、抗体片段、scfv、或骆驼科vhh结构域。抗体、骆驼科vhh结构域和各种受体和配体的重链(vh)和轻链(vl)的可变链的序列可以通过测序或通过可公开获得的数据库获得并且可以用于使用本文描述的方法合成sir,如不同实例中所示。将包含sir的抗原结合结构域的序列密码子优化并且使用可公开获得的软件(例如thermofisher或idt)和商业供应商(例如idt)来人工合成。将所得片段pcr扩增并且使用标准分子生物学技术在含有不同sir骨架的不同载体中克隆。一般而言,sir构建体典型地在慢病毒载体中克隆。使用自动化测序确认sir构建体的序列。示例性sir构建体是plenti-ef1a-cd8sp-myc3-wt1-ab13-vl-v5-[htcrb-kaciah]-f-p2a-sp-wt1-ab13-vh-myc4-[htcra-csdvp]-f-f2a-pac-dwpre[cloneid:071516-j04](seqidno:1380)。此构建体具有许多便利的限制性位点,使得抗原结合结构域片段(例如vl和vh结构域)可被切割并用靶向其它抗原的抗原结合结构域置换。例如,vl结构域可以使用ecori和xhoi限制性酶位点从该载体中切割并且用含有ecori和xhoi位点并编码靶向另一种抗原的vl结构域的新dna片段置换。载体在cd8信号肽(cd8sp)上游携带nhei位点,该位点也可以连同xhoi位点一起用于在携带5’信号肽的新vl片段中克隆。bstbi和mlui位点可以用于置换vh片段。xhoi和spei位点可以用于置换编码v5-[htcrb-kaciah]-f-p2a的模块。类似地,mlui和xba位点可以用于置换含有myc4-[htcra-csdvp]-f-f2a的模块。编码pac的辅助模块可以使用xbai(或ndei)和sali限制性位点置换。因此,本领域普通技术人员可以使用此载体和抗原结合结构域的序列(例如抗体的vl和vh结构域)生成靶向任何其它新抗原的sir。可以用于治疗sir的另一种示例性构建体是plenti-ef1a-cd8sp-myc-[htcra-t48c-opt1]-f-f2a-fmc63-vh-v5-[htcrb-s57c-opt1]-f-p2a-pac-dwpre[042616-b03](seqidno896),在该sir中该抗原结合结构域连接至tcrb恒定链,而tcra恒定链保持为空。抗原结合结构域可以在此载体中在bstbi与mlui限制性酶位点之间克隆。可替代地,具有其自身信号序列的抗原结合结构域可以在此载体中在spei与mlui限制性酶位点之间克隆。可以用于治疗sir的另一种示例性构建体是plenti-ef1a-cd8sp-v5-[htcrb-kaciah]-f-p2a-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac-dwpre[081415-f04](seqidno895),在该sir中该抗原结合结构域连接至tcra恒定链,而tcrb恒定链保持为空。抗原结合结构域可以在此载体中在bstbi与mlui限制性酶位点之间克隆。可替代地,具有其自身信号序列的抗原结合结构域可以在此载体中在spei与mlui限制性酶位点之间克隆。分泌抗原--nluc融合蛋白的生成..(任选步骤).为了测量sir在免疫效应细胞上的表达和结合活性,sir的抗原靶标的细胞外结构域经由短gly-ser接头融合至荧光素酶,诸如nluc或gluc或mluc7或turboluc16或paluc。例如,cd19的细胞外结构域经由gly-ser-接头与nluc同框融合。融合蛋白携带n-末端信号序列。所得构建体在293ft细胞中瞬时转染并且在48-72小时之后收集含有分泌性融合蛋白的上清液。分泌性抗原结合结构域(abd)--nluc融合蛋白的生成和使用..(任选步骤).为了鉴定用于测试sir活性的适合细胞系,sir的抗原结合结构域经由短gly-ser接头融合至荧光素酶,诸如nluc或gluc或mluc7或turboluc16或paluc。例如,在sir是基于靶向cd19的fmc63单克隆抗体的情况下,fmc63的scfv片段(vl-gly-ser-接头-vh)经由gly-ser-接头与nluc同框融合。融合蛋白携带n-末端信号序列。所得构建体在293ft细胞中瞬时转染并且在48-72小时之后收集含有分泌性融合蛋白的上清液。测试一组细胞系与abd-nluc融合蛋白的结合以鉴定表达高水平sir靶标并因此可用于测试sir活性的细胞系。表a提供了表达不同抗原靶标的细胞系的示例性列表,这些抗原靶标可以用于测定本公开的sir的活性。也可以使用替代性方法鉴定表达sir的靶标的细胞系,这些替代性方法诸如文库搜索、使用可商购获得的抗体进行的免疫染色或通过搜索可公开获得的基因表达数据库。在以下测定中测试表达sir的免疫效应细胞以鉴别功能性sir。(a)nluc结合测定:用靶抗原-nluc融合蛋白(如上文描述)染色表达对照载体和sir的jurkat-nfat-gfp或t细胞并且通过测量nluc活性测定其与靶抗原结合的能力。例如,与表达对照载体的jurkat-nfat-gfp细胞或亲本jurkat-nfat-gfp细胞相比,表达基于fmc63的靶向cd19的sir的jurkat-nfat-gfp细胞显示增加的与cd19-nluc融合蛋白的结合。(b)诱导nfat启动子驱动的gfp表达.将表达对照载体和sir的jurkat-nfat-gfp细胞与表达靶抗原的细胞系(如上文描述)共同培养4-24小时并且通过使用流式细胞术测量gfp表达的诱导来测定其与靶抗原结合的能力。收集细胞上清液并且测定细胞因子(例如il2)的诱导。(c)测定细胞因子产生:将表达对照载体和sir的jurkat-nfat-gfp或t细胞与靶细胞系共同培养4-96小时并且使用elisa检查上清液中细胞因子(例如il2、ifnγ、tnfα等)表达的诱导。(d)测定体外和体内细胞毒性活性:将未感染t细胞或表达对照载体或sir的那些细胞与表达非分泌形式的荧光素酶(诸如gluc、nluc、turboluc16等)共同培养4-96小时并且通过测量荧光素酶活性来检查细胞裂解的诱导,如pct/us17/52344中所述。也可以使用用于测量细胞毒性活性的替代性方法(例如51cr释放测定或ldh释放测定)。还可以在体内使用免疫缺陷型小鼠的适当异种移植模型测定表达sir的t细胞的活性。基于以上方法,本领域普通技术人员可以容易设计、构建、测试并选择针对任何抗原的适当功能化sir或sir库。该sir或sir库可以用于人类临床试验和用于预防和治疗各种疾病病状的临床用途。表9提供了可使用本公开的sir治疗的人类疾病病状的示例性列表。可能不同sir或sir亚集根据各种因素最佳适用于不同疾病病状,这些因素包括但不限于靶抗原在引起疾病细胞和疾病相关细胞上的表达流行性和水平、疾病负荷和疾病进展速率。根据其功效和毒性概括以及患者的病状,不同sir可以甚至最佳适用于不同患者的单一疾病病状。本公开提供生成不同过继免疫反应的显著技术和逻辑障碍的解决方案。通过基因重排产生正常tcr多样性。胸腺中的严格阳性和阴性选择过程确保唯有表达局限于识别低亲和力范围内的自身肽/mhc的αβtcr的t细胞可以居住于外周。因此,胸腺环境允许生成自身限制性而非自身反应性αβt细胞库。由不同抗原结合结构域生成不同sir库受限于生成并测试多个抗原结合结构域的技术和财政障碍。更重要的是,由于每个抗原结合结构域(例如抗体的vl和vh片段)具有结合其它抗原并引起脱靶毒性的可能性,所以仅基于多个抗原结合结构域的不同sir库可能具有增加的毒性风险。因此,此库的潜在多样性将必须局限于减少脱靶毒性。本公开通过经由将单一或几个抗原结合结构域附接至tcr链的不同变体来由该单一或几个抗原结合结构域生成不同sir库来克服此问题。sir库的多样性通过使用不同接头进一步增加。表达该库的t细胞的多样性可以通过使用本公开所述的不同辅助模块和治疗控制来进一步增加。此不同sir库可以用于提供针对表达所述抗原的引起疾病细胞或疾病相关细胞的不同免疫反应。可替代地,不同sir库可以任选地使用本领域已知的技术进行dna条形码化并且随后用于选择具有最佳生物和临床特征的单一sir或sir子组。这些特征可包括但不限于体外生物测定的性能(例如细胞毒性、细胞因子分泌、结合亲和力、细胞表面表达、脱靶效应、t细胞增殖、耗尽标记物表达和终末分化等)、体内测定的性能(例如存活、肿瘤减小、t细胞持久性、t细胞扩增等)以及临床经验(例如疾病消退、复发率、毒性等)。本公开的sir可以单独使用或者与本领域已知的其它sir、car、ctcr和其它天然和合成免疫受体组合使用,以生成不同免疫效应细胞库,以预防并治疗由表达其靶抗原的细胞造成或与这些细胞相关的不同疾病病状。本公开的sir是嵌合t细胞受体的修饰形式,其不仅已针对多样性进行优化,而且还针对结合亲和力、细胞表面表达、链配对、信号传导和细胞递送进行优化。为了生成适用于临床应用的双链sir,解决了若干技术和概念障碍。双链ctcr平台的主要限制为递送两种不同质粒构建体的技术性难题。尽管可能用两个单独载体或具有两个单独启动子的单一载体中的双链sir的两条链转导t细胞,这两种方法具有不平衡的引入sir链的表达的风险,从而增加过量表达的链将与内源性tcr链错配对的可能性。两种载体方法的额外限制是与单一载体方法相比更低的靶细胞感染效率。在单一载体中使用两个单独启动子具有增加插入物大小并因此增加载体大小超过大部分逆转录病毒和慢病毒载体的最佳包装限制的缺点,进而导致病毒效价降低。另外,内部启动子可以在逆转录病毒载体中通过启动子干扰来沉默。尽管脑心肌炎病毒的内部核糖体进入位点(ires)序列已广泛用于构建双顺反子病毒载体,但是ires介导的翻译相对无效。另外,ires的大小相对大并且使用ires元件可导致竞争翻译因子和/或同源重组。出于这种考虑,选择衍生自小核糖核酸病毒或猪捷申病毒的‘自身裂解’2a肽序列,以结合在两条sir链之间,以实现两条引入的sir链的等摩尔表达。通过核糖体跳跃机制,这些序列诱导从单一mrna转录物翻译两个单独肽,从而实现每种肽的大约化学计量产生。可裂解“2a”肽的实例提供于seqidno:3060-3062和3064中。在一些实施方式中,(sg)2也添加在2a序列上游以增强裂解效率。另外,为了避免连接至sir的任何剩余2a肽序列,将弗林蛋白酶裂解位点添加在(sg)2继续上游以促进在翻译之后残余2a肽的裂解。最初选择慢病毒和逆转录病毒载体以开发适用于临床应用的双链sir系统。随后,实验使用睡美人转座子和mrna转染来在t细胞中成功表达sir。plenti-blast载体通过去除lacz基因而衍生自plenti6v5gw_lacz载体(英杰公司;赛默飞世尔科技公司(thermofisherscientific))。plenti-mp2由pantelistsoulfas惠赠(爱迪基因公司(addgene)质粒编号36097;enomoto等人.exp.neurol.[实验神经学]248:170-82,2013)并且用于使用标准分子生物学技术通过用人类ef1α启动子置换cmv启动子来生成plenti-ef1α慢病毒载体(seqidno:870)。pspax2由didiertrono惠赠(爱迪基因公司质粒编号12260)。plp/vsvg包膜质粒和293ft细胞获得自迎接公司(赛默飞世尔科技公司)。逆转录病毒转移载体mscvneo、mscvhygro和mscvpac以及包装载体pkat获得自robertillaria博士的实验室。phrgtk海肾荧光素酶质粒来自普洛麦格公司(promega)。使用商业供应商(idt)以单个或多个片段人工合成编码不同信号肽、抗体结合结构域、接头、tcr恒定链、可裂解接头和选择标记物(例如pac、egfp、cnb30等)的基因片段并且在使用含有适当限制性酶的引物的pcr反应中用作模板。将扩增的片段用适当限制性酶消化并且然后在plenti-ef1α(seqidno:870)、plenti-ef1α-dwpre(seqidno:871)或mscv-bgl2-avrii-bam-ecor1-xho-bstb1-mlu-sal-clai.i03(seqidno:872)载体中使用标准分子生物学技术来克隆。plenti-ef1α-dwpre载体与plenti-ef1α的不同之处在于缺乏wpre区域。可替代地,可以人工合成编码整个sir盒的基因片段(例如cd8信号肽-161-vl-tcrb-f-2a-igh-信号肽-161-vh-tcra-f-2a-pac)。然后可以在使用含有适当限制性酶的引物的pcr反应中将所得片段用作模板。可以用适当限制性酶消化扩增的片段并且然后在适当载体中使用分子生物学技术克隆。制成许多不同慢病毒构建体,其含有衍生自fmc63单克隆抗体的vl和vh片段,这些片段分别与人类tcr-β(tcrb)链和人类tcr-α(tcra)链的恒定区同框融合。大部分构建体也携带通过编码弗林蛋白酶-sgsg-2a的接头融合至sir表达盒的不同选择标记物(例如嘌呤霉素n-乙酰基-转移酶(pac)、增强的绿色荧光蛋白(egfp)和分泌性nluc)。示例性构建体是在plenti-ef1a载体(seqidno:870)中克隆的cd8sp-fmc63-vl-v5-[htcrb-wt]-f-p2a-sp-fmc63-vh-myc-[htcra-wt]-f-f2a-pac(080815-f02)[seqidno:922]。在此载体中的sir表达盒由人类ef1α(延长1α)启动子驱动。sir表达盒包含从编码人类cd8信号肽(cd8sp)、密码子优化的fmc63vl片段、在c-末端处含有gly-ser-gly氨基酸的v5接头、人类tcr-β2(tcrb)链的恒定区(c区域)、弗林蛋白酶裂解位点(rakr)、sgsg接头、p2a核糖体跳跃序列、人类igh信号肽、密码子优化的fmc63vh片段、衍生自常用myc表位标签并在c末端处含有gly-ser-gly氨基酸的myc接头、人类tcr-α(tcra)链的恒定区(c区域)、弗林蛋白酶裂解位点(rakr)、sgsg接头、f2a核糖体跳跃序列的5’-末端-核苷酸和嘌呤霉素抗性基因(pac)的变体形式。在vl片段与v5表位标签之间存在xhoi限制性酶位点,在人类igh信号肽之前存在spei限制性位点,在myc标签之前存在mlui位点,并且在pac基因之前存在含有xbai和ndei位点的短接头。在plenti-ef1a载体(seqidno:870)中在nhei与sali位点之间克隆整个表达盒。此载体可以用于通过经由分别用nhei和xhoi和spei和mlui酶消化来去除fmc63-vl和-vh片段并使用标准分子生物学技术用靶向其它抗原的抗原结合结构域(例如vl、vh、vhh、scfv、受体或配体)的dna片段置换它们来克隆靶向其它抗原的抗原结合结构域。也创建以上构建体的若干变体:1.plenti-ef1-fmc63vl-v5-[mtcrb-wt]-f-p2a-fmc63-vh-myc-[mtcra-wt]-f-f2a-pac-b06:此构建体类似于plenti-ef1-fmc63vl-v5-[htcrb-wt]-f-p2a-fmc63-vh-myc-[htcra-wt]-f-f2a-pac-f02构建体,除了他具有小鼠tcr-β(tcrb)和小鼠tcrα链而非对应人类链的恒定区(c区域)。2.plenti-ef1-fmc63-vl-v5-[tcrb-s57c-opt1]-f-p2a-fmc63-vh-myc-[tcr-ca-t48c-opt1]-f-f2a-pac-l05(050515-l05):此构建体在以下方面不同于plenti-ef1-fmc63vl-v5-[htcrb-wt]-f-p2a-fmc63-vh-myc-[htcra-wt]-f-f2a-pac-f02:a.编码tcrb和tcra链的恒定区的核苷酸序列被密码子优化以增加sir表达。b.为了减少所引入tcr链与内源性tcr链的错配,将额外半胱氨酸残基添加到每条链以促进形成额外链间二硫键。因此,tcrb序列携带ser57至半胱氨酸突变并且tcra序列携带苏氨酸48至半胱氨酸突变。3.plenti-ef1-fmc63-vl-v5-[tcrb-kaciah]-f-p2a-fmc63-vh-myc-[tcra-csdvp]-f-f2a-pac-d06(081415-d06):此构建体在以下方面不同于plenti-ef1-fmc63vl-v5-[htcrb-wt]-f-p2a-fmc63-vh-myc-[htcra-wt]-f-f2a-pac-f02:a.编码tcrb和tcra链的恒定区的核苷酸序列被密码子优化以增加sir表达。b.为了减少所引入tcr链与内源性tcr链的错配,将额外半胱氨酸残基添加到每条链以促进形成额外链间二硫键。因此,tcrb序列携带ser57至半胱氨酸突变并且tcra序列携带苏氨酸48至半胱氨酸突变。c.鼠类tcr已显示表达好于其人类相对物并且人类tcr的鼠类化已显示改进其表达。在此构建体中,人类tcrβ(tcrb)恒定区的五个氨基酸被对应鼠类氨基酸置换。这些鼠类氨基酸是k-18、a-22、i-133、a-136和h-139。另外,在人类tcrα(tcra)恒定链(seqidno:3010)中,含有四种氨基酸丝氨酸(s-91)、天冬氨酸(d-92)、缬氨酸(v-93)和脯氨酸(p-94)的鼠类tcrα(tcra)恒定链的区域被取代。除了慢病毒载体之外,sir表达盒被克隆到逆转录病毒载体中。出于此目的,pmscv-puro逆转录病毒载体通过用bglii和clai酶消化并且连接含有avrii、bamhi、ecori、xhoi、bstbi、mlui和sali的限制性酶位点的bglii和clai切割的双链寡核苷酸来修饰。所得载体的序列在seqidno:872中被称为mscv-bgl2-avrii-bam-ecor1-xho-bstbi-mlu-sal-clai.i03。使用nhei和sali酶从plenti-ef1载体中消化不同sir构建体的表达盒并且在已用avrii和sali消化的以上载体中克隆。示例性sir构建体mscv-fmc63vl-v5-[tcrb-kaciah]-f-p2a-2-spe-fmc63vh-myc-[tcra-csdvp]-f-f2a-pac.n01[cloneid:032216-n01]的序列呈现于seqidno:873中。基于mscv的逆转录病毒载体中的其它sir构建体可以容易通过从其对应plenti-ef1克隆nhei至sali片段来构建。也将sir表达盒克隆到睡美人载体psbbi-pur(seqidno:874;爱迪基因公司;质粒编号60523)和psbbi-gp(爱迪基因公司;质粒编号60511)。出于此目的,使用agei和xbai酶从plenti-ef1载体中消化不同sir构建体的表达盒并且在已用agei和xbai消化的psbbi-pur和psbbi-gp载体中克隆。编码靶向cd19的sir的示例性载体的序列提供于seqidno:875中。具有延长多克隆位点的另一种示例性睡美人载体的序列提供于seqidno:876中。被工程化以表达荧光素酶(例如gluc或nluc)的细胞系提供于表a中,该这些荧光素酶用于测量靶向不同细胞表面和细胞内抗原的不同构建体的细胞毒性。用于此实验中的细胞系、这些细胞系上的靶抗原及其生长介质示出于以下表a中。在37℃下在5%co2润湿培养器中培养细胞。从atcc获得细胞系,nihaids试剂程序或可在实验室中获得。表a:用nfat依赖性gfp报道基因工程化的jurkat细胞系(克隆e6–1)由ucsf的arthurweiss博士惠赠。将jurkat细胞保持在补充有10%fbs、青霉素和链霉素的rpmi-1640培养基中。生成慢病毒和逆转录病毒.在基本上如先前所述的293ft细胞中通过基于磷酸钙的转染生成慢病毒(matta,hozayev,tomar,chugh,&chaudhary,2003)。使293ft细胞在具有10%fcs4mml-谷氨酸盐、0.1mmmem非必需氨基酸和1mmmem丙酮酸钠的dmem(在此称为dmem-10)中生长为了生成慢病毒,在组织培养板中在10ml无抗生素的dmem-10培养基中接种293ft细胞,使得它们将在转染那天为约80汇流。次日,使用10μg编码不同基因的慢病毒表达质粒、7.5μgpspax2质粒和2μgplp/vsvg质粒通过磷酸钙转染方法来转染细胞。在转染后大约15-16小时,去除9ml培养基并且用5ml新鲜培养基置换。在转染后大约48小时,收集5ml上清液(第一次收集)并且用新鲜5ml培养基置换。在转染后大约72小时,收集全部培养基(第二次收集,通常约6ml)。将所收集上清液合并并且在1000rpm下离心1分钟以去除任何细胞碎片和非粘附细胞。通过0.45μm注射过滤器过滤无细胞上清液。在一些情况下,通过在18500rpm下在4℃下超离心2小时来进一步浓缩该上清液。将病毒团块重新悬浮于1/10初始体积的xvivo培养基中。然后新鲜使用病毒感染靶细胞或在-80℃下以等分试样冷冻保存。感染t细胞和pbmc.从洛杉矶儿童医院的血库的健康未鉴定成人供体获得血沉棕黄层细胞并且用于通过泛影葡胺(ficoll-hypaque)梯度离心分离外周血单核细胞(pbmc)。使用cd3磁性微珠(美天旎生物技术公司(miltenyibiotech))并且遵循制造商说明书,如此使用pbmc或或将其用于分离t细胞。将pbmc或分离的t细胞重新悬浮于补充有10ng/mlcd3抗体、10ng/mlcd28抗体和100iu重组人类-il2的xvivo培养基(龙沙公司(lonza))。在37℃下在5%co2润湿培养器中培养细胞。将细胞在以上培养基中活化1天,之后用慢病毒载体感染。一般而言,在早晨通过自旋感染(在37℃下1800rpm持续90分钟)使用已重新悬浮于存在8μg/mlof(西格玛公司(sigma),目录号h9268)的xvivo培养基中的300μl浓缩病毒感染初代细胞(例如t细胞)。在晚上更换培养基并且重复多两天感染,总计3次感染。在第3次感染之后,使细胞成团并将其重新悬浮于含有10ng/mlcd3抗体、10ng/mlcd28抗体和100iu重组人类-il2并补充有相应抗生素(在指示时)的新鲜xvivo培养基中并且置于在细胞培养烧瓶中以进行选择,除非另外指示。在以上培养基中培养细胞,在未使用药物选择的情况下持续10-15天并且在使用药物选择的情况下持续20-30天。假如在用表达egfp的慢病毒感染细胞时,在未进行药物选择的情况下扩增这些细胞或者流式分选这些细胞以富集表达egfp的细胞。为了感染癌细胞系,在具有(西格玛公司,目录号h9268)的3ml总体积中使用2ml未浓缩病毒上清液感染大约500000个细胞。然后次日早上,使细胞成团并且将其重新悬浮于具有对应抗生素的培养基中并且置于细胞培养烧瓶中进行选择。将基本上与上文对于慢病毒载体产生所述程序类似的程序用于生成逆转录病毒,除了通常使用10μg逆转录病毒构建体、4μgpkat和2μgvsvg质粒在10cm组织培养板的10mldmem-10培养基中转染293ft细胞。基本上如上文对于慢病毒载体所述进行病毒收集和靶细胞感染。使用睡美人载体的jurkat细胞电穿孔..为了电穿孔,将5×106个jurkat细胞在90g下离心10分钟,将其重新悬浮于100μl缓冲液中并且将其与20μg编码睡美人sir的质粒和5μgsb100x转座酶质粒混合。向电穿孔缓冲液和比色皿中提供来自龙沙公司的amaxacelllinenucleofector试剂盒v(vca-1003)。将重新悬浮的细胞转移至比色皿并且使用程序x-001电穿孔。在电穿孔之后,将细胞在比色皿中在室温下孵育10分钟并且然后将1ml补充有20%fbs的预温热rpmi培养基添加到比色皿的细胞中。将细胞转移至每孔含有1ml预温热培养基的6孔板中并且在37℃下孵育过夜。次日,将细胞离心并且通过补充有10%fbs和250ng/ml嘌呤霉素的rpmi置换培养基以选择表达睡美人-sir的jurkat细胞。抗体和药物.洋地黄皂苷购自西格玛公司(目录号d141)并且以dmso制成100mg/ml储备溶液。以pbs制备稀释的1mg/ml储备液。用于细胞裂解的洋地黄皂苷的最终浓度为30μg/ml,除非另外指示。il2elisa.在表达sir的jurkat-nfat-gfp效应细胞或t细胞的细胞培养上清液中测量人类il2,这些效应细胞或t细胞已使用来自r&d系统公司(明尼苏达州明尼阿波利斯市(minneapolis,mn))的il2-elisa试剂盒并遵循制造商的建议与特定靶细胞系共同培养24至96小时。facs分析.小鼠抗人类c-mycapc-缀合的单克隆抗体(目录号ic3696a)来自r&d系统公司(明尼苏达州明尼阿波利斯市)。生物素化蛋白质l购自genescript公司(新泽西州皮斯卡塔韦(piscataway,nj)),以磷酸盐缓冲盐水(pbs)以1mg/ml重构并且在4℃保存。链霉亲和素-apc(sa1005)购自赛默飞世尔科技公司。为了使用myc染色检测car和sir,收获1×106个细胞并且用3ml含有4%牛血清白蛋白(bsa)洗涤缓冲液的冰冷1×pbs洗涤三次。在洗涤之后,将细胞重新悬浮于含有10μlapc缀合的myc抗体的0.1ml冰冷洗涤缓冲液并且在黑暗中孵育1小时,随后用冰冷洗涤缓冲液进行两次洗涤。为了使用蛋白质l染色检测car和sir,收获1×106个细胞并且用3ml含有4%牛血清白蛋白(bsa)洗涤缓冲液的冰冷1×pbs洗涤三次。在洗涤之后,将细胞重新悬浮于含有1μg蛋白质l的0.1ml冰冷洗涤缓冲液中,在4℃下持续1小时。将细胞用冰冷洗涤缓冲液洗涤三次,并且然后用0.1ml洗涤缓冲液中的10μlapc缀合链霉亲和素孵育(在黑暗中)30分钟,随后用冰冷洗涤缓冲液进行两次洗涤。使用来自bd生物科学公司(bdbiosciences)的facsverse分析仪分析facs。细胞死亡测定.为了测量细胞死亡,利用基于gluc或nluc的异位细胞溶质表达的新型测定,如pct/us17/52344“anon-radioactivecytotoxicityassay[非放射性细胞毒性测定]”所述。该方法涉及以一种方式在靶细胞中表达报道基因,使得其优选保留在健康细胞内,而从死亡和将死细胞中释放或者其活性可优选在死亡和将死细胞中测量。用于此测定的优选报道基因是1)来自桡足动物的非分泌形式的荧光素酶,诸如gaussiaprinceps、腹突乳点水蚤(pleuromammaabdominalis)、太平洋长腹水蚤(metridiapacifica)、残尾长腹水蚤(metridiacurticauda)、非对称长腹水蚤(metridiaasymmetrica)、metridiaokhotensis、细长长腹水蚤(metridialonga)、lucicutiaovaliformis、heterorhabdustanneri和盾片状乳点水蚤(pleuromammascutullata),2)来自深海虾的工程化荧光素酶报道基因,诸如nanoluc。若干此类报道基因载体的序列提供于seqidno:881至seqidno:887中。以上载体用于生成逆转录病毒和慢病毒,这些病毒进而用于生成在用适当抗生素选择之后稳定表达gluc、nluc、turboluc或mluc7的若干靶细胞系的多克隆群体。除非另外指示,否则将稳定表达不同荧光素酶(gluc、nluc、mluc7或turboluc16)的靶细胞重复三次接种在384孔板的用于生长靶细胞的培养基中。将悬浮液中生长的靶细胞通常以每孔2-3x104的浓度接种,而生长为粘附单层的靶细胞以每孔1-2x104的浓度接种。除非另外指示,否则将靶细胞与遗传修饰t细胞(即表达sir或car的那些细胞)共同培养,其效应:靶标(e:t)比率在1:1至10:1之间变化,持续4小时至96小时。在靶细胞生长为粘附细胞(例如hela细胞)的情况下,使它们附接至孔底部过夜,之后添加t细胞。通过直接注射含有天然腔肠素(coeloentrazine)(nanaolight)的0.5xctz测定缓冲液来增加荧光素酶活性(如通过bioteksynergy板读取器测量),测定靶细胞裂解的t细胞介导的诱导。ctz测定.通过将1mg冻干ct粉末溶解于补充有30μl6nhcl的1.1ml100%甲醇中以避免ctz随时间氧化来制成天然腔肠素(ctz;nanolight,目录号303)的100x储备溶液。为了制成ctz测定缓冲液,将100xctz储备溶液以pbs稀释至0.5x浓度。除非另外指示,否则将总体积15μl的ctz测定缓冲液(如上文制备)添加至384孔白板(格林纳公司(greiner),384孔白板目录号781075)的每个孔中,这些孔在约50-60μl培养基体积中含有表达非分泌形式的荧光素酶的细胞,并且使用bioteksynergyh4板读取器以端点模式读取板的荧光。对于96孔板,将细胞接种在200μl培养基中并且添加大约50μl0.5xctz测定缓冲液。除非另外指示,否则将0.5xctz测定缓冲液用于测定gluc、turboluc16和mluc7的活性。在一些实验中也将ctz测定缓冲液(稀释至0.125x浓度)用于测量nluc活性(参见下文)。一般而言,除非另外指示,否则所添加的0.5xctz测定缓冲液的体积为含有细胞的孔中的液体体积的大约1/4,尽管在将0.5xctz测定以1:1体积添加到含有细胞的培养基中时该测定也可用。在指示时,将含有单独培养基(med)的孔和靶细胞与未用任何sir构建体(t-ui)感染的t细胞孵育的孔中的gluc活性用作对照。在无先前细胞裂解的情况下使用bioteksynergyh4板读取器以端点模式读取板的荧光。在一些实验中,使用ctz测定缓冲液测量nluc活性,但在此将该缓冲液稀释至0.125x的最终浓度。当将ctz测定缓冲液用于测量nluc活性时,通过注射器将大约15μl总体积(除非另外指示)的0.125xctz测定缓冲液添加至384孔白板(格林纳公司,384孔白板目录号781075)的含有大约50-60μl体积培养基中的细胞的每个孔中并且使用bioteksynergyh4板读取器以端点模式读取板的荧光。对于96孔板,通常将细胞接种在200μl培养基中并且添加大约50μl0.125xctz测定缓冲液。开发检测cd19和mpl(促血小板生成素受体)抗原的表达的测定..为了检测sir及其靶抗原的表达,利用基于荧光素酶的报道基因测定,如pct/us2017/025602“ahighlysensitiveandspecificluciferasebasedreporterassayforantigendetection[用于抗原检测的高度敏感且具特异性的基于荧光素酶的报道基因测定]”。.cd19和mpl(也称为促血小板生成素受体或tpo-r)两者在造血细胞上表达,但在不同品系的细胞中显示不同表达。fmc63是特异性识别人类cd19的良好表征的小鼠单克隆抗体。类似地,161是识别人类mpl的单克隆抗体。基于fmc63vl和vh片段的已知序列,生成fmc63单链fv(scfv)片段。编码fmc63scfv片段的cdna由5’至3’端编码衍生自人类cd8分子的信号肽、fmc63vl片段、(gly4ser)x3接头和fmc63-vh片段的核苷酸序列组成。编码fmc63scfv片段的cdna然后在其3’端处通过gly-gly-ser-gly接头与编码acv5标记的nluc的cdna同框融合,以生成fmc63-ggsg-nluc,然后将其克隆在慢病毒载体plenti-ef1的人类ef1α启动子下游,以制成构建体plenti-ef1a-fmc63(vl-vh)-ggsg-nluc-acv5-u09(seqidno:880)。插入物片段序列提供于seqidno:4516中。使用针对mpl的161单克隆抗体的vl和vh片段类似地生成编码161-ggsg-nluc的构建体。插入物片段的核酸序列提供于seqidno:4517中。通过磷酸钙共沉淀方法将plenti-ef1-fmc63-ggsg-nluc-acv5和plenti-ef1-161-ggsg-nluc-acv5质粒转染到293ft细胞中。转染后大约20h,用xvivo培养基置换细胞培养基。48-72h后收集含有分泌的fmc63-ggsg-nluc-acv5和161-ggsg-nluc-acv5蛋白质的条件培养基。将含有fmc63-ggsg-nluc-acv5和161-ggsg-nluc-acv5蛋白质的上清液用于检测cd19和mpl在jurkat、k562、raji、rs-4-11(rs411)和hel.92.1.7(hel)细胞表面上的表达,这些细胞已通过用表达人类c-mplcdna的慢病毒载体或空载体转导这些细胞来工程化以表达c-mplcdna。这些细胞还表达缺乏其信号肽的人源化gluccdna。将表达载体和mpl的jurkat-gluc、k562-gluc、hel.92.1.7-gluc、raji-gluc和rs411-gluc细胞与fmc63-ggsg-nluc-acv5和161-ggsg-nluc-acv5上清液在4℃下孵育1h,随后用补充有0.1%bsa的冷pbs进行充分洗涤。将细胞重新悬浮于冷pbs中并且在平底384孔板(格林纳公司,384孔白板目录号781075)中每孔接种30μl细胞上清液,重复三次。通过自动分配器使用bioteksynergyh4板读取器以孔模式将含有作为nluc底物的天然腔肠素(ctz)(以pbs稀释的30μl/孔的天然腔肠素)的nluc测定缓冲液添加到每个孔中并且测量作为nluc活性的量度的光发射。在hel.92.1.7-gluc-载体细胞上观察到用161-ggsg-nluc-acv5观察的强结合,这表明mpl的内源性显著表达。mpl在hel.92.1.7-gluc-mpl细胞中的异位表达引起161-ggsg-nluc-acv5结合的轻微增加。相反,在表达载体的jurkat、raji和rs411细胞上观察到与161-ggsg-nluc-acv5的极弱结合,并且在异位表达mpl时仅适当增加。在k562-载体细胞上观察到161-ggsg-nluc-acv5的结合并且该结合在k562-mpl细胞上显著增加。与161-ggsg-nluc-acv5相比,fmc63-ggsg-nluc-acv5上清液在表达载体和mpl的raji细胞上显示最强结合,在rs411细胞上显示适当强的结合,并且在其它细胞上显示极弱至可忽略的结合。开发用于定量测量靶向cd19和mpl(促血小板生成素受体)的嵌合抗原受体(car)和嵌合tcr(sir)与其受体的结合亲和力的测定..过继性细胞疗法领域的常见问题是缺乏可以检测表达嵌合抗原受体和sir的细胞的敏感且特异性的测定。尽管使用蛋白质-l的染色可以用于检测含有car和sir的scfv的细胞表面表达,但是它未能测量car和sir与其靶抗原的相互作用。为了检测靶向cd19和mpl的car的结合亲和力,使用高敏感性基于荧光素酶报道基因的抗原检测测定,如pct/us2017/025602所述,该专利通过引用整体结合。人类cd19和人类mpl(包括其信号肽)的细胞外结构域与编码gly-gly-ser-gly接头、nluc(无分泌信号)和acv5表位标签的核苷酸序列同框融合。在cd19构建体的情况下,将flag标签插入在信号肽与细胞外结构域开始部分之间。将整个盒克隆到慢病毒载体plenti-ef1的人类ef1α启动子下游,以分别制成构建体plenti-ef1-cd19-ggsg-nluc-acv5和plenti-ef1-mpl-ggsg-nluc-acv5。插入物片段的核酸序列分别提供于seqidno:4518和4519中。通过磷酸钙共沉淀方法将构建体转染到293ft细胞中。转染后大约20h,用新鲜培养基置换细胞培养基。48-72h后收集含有分泌的flag-cd19-ggsg-nluc-acv5和mpl-ggsg-nluc-acv5蛋白质的条件培养基。使用磷酸钙共转染方法用表达靶向cd19(fmc63-bbz-pac;seqidno:4501)、mpl(161-bbz-pac-r07;seqidno:4502或161-28z-pac-z07)的嵌合抗原受体的慢病毒构建体瞬时转染293ft-细胞(在24孔板中,500ul体积)或使其未转染。次日早上,在转染后大约18小时,通过在1.5ml管中上下吸取来收集细胞。将管在1500rpm下旋转5分钟。然后用洗涤缓冲液(pbs中的1%fbs)将细胞洗涤一次,随后与100μl指示的分泌形式的ggsnluc上清液一起孵育。将细胞在4℃下孵育1小时。在孵育之后,将细胞用洗涤缓冲液洗涤5次(每次洗涤1ml)。最终,使团块重新悬浮于200μl洗涤缓冲液中。将重新悬浮的细胞置于384孔板中,重复三次(每次25μl)。在将含有天然腔肠素的nluc测定缓冲液(普洛麦格公司)(每孔25μl)直接添加到每个孔中(一次一孔)之后,使用bioteksynergyh4板读取器测量荧光素酶活性。如图9a--b所示,表达cd19(fmc63-bbz-pac)-car的293ft细胞证实与flag-cd19-ggsg-nluc-acv5的强结合,如通过nluc测定所测量,而在未感染t细胞(ui)或表达对照161-bbz-paccar的那些细胞上可见极少结合。类似地,与未转染293ft细胞或用fmc63-bbz-paccar转染的那些细胞相比,表达161-cd28z-paccar的293ft细胞显示与mpl-ggsg-nluc-acv5上清液的强结合。结果证实了ggsg-nluc测定(或nluc结合测定)以敏感且定量的方式测量细胞表面表达的嵌合受体与其抗原靶标的结合的能力。构建许多含有car和sir的不同潜在靶标的细胞外结构域的其它nluc融合蛋白并且使用表达其对应car的293t或t细胞验证。这些构建体的名称、dna和氨基酸seqidno提供于表7i中。可以通过经由短柔性接头将为car/sir靶标的细胞外结构域融合至nluc来针对sir的其它抗原靶标生成类似构建体。cd20是具有两个细胞外环的iii型膜蛋白。成功生成cd20-ecx2-ecd-ggsg-turboluc16-4xflag-2xstreptag-8xhis-t2a-pac(060816-i04)融合构建体并且针对表达cd20car的细胞进行验证。此构建体的氨基酸序列由seqidno:12374表示。因此,可以使用car或sir的任何蛋白质抗原靶标生成nluc(或turboluc16)融合蛋白,该抗原靶标可以用于检测表达car/sir的细胞的表达和结合亲和力。已知蛋白质l结合至κ轻链。为了检测含有κ轻链的car和sir的表达,构建在cd8信号肽下游含有n末端蛋白质l编码区的两个nluc融合构建体。两个构建体是相同的,除了构建体cd8sp-protein-l-2-ggsg-nluc-4xflag-x2strep-8xhis-t2a-pac(101916-p03)[seqidno:12382]在蛋白质l编码区中缺乏单一氨基酸,该氨基酸存在于构建体cd8sp-蛋白质-l-ggsg-nluc-4xflag-x2strep-8xhis-t2a-pac(112316-q02)[seqidno:12381]。通过在293ft细胞中转染构建体来生成含有融合蛋白的条件上清液并且其显示结合至含有κ轻链的car构建体,其中该κ轻链结合至蛋白质l。抗原-nluc融合蛋白诸如cd19-ecd-ggsg-nluc-acv5结合至car或sir的抗原结合结构域并且因此可以用于测量表达car或sir的免疫效应细胞的结合亲和力。相比之下,蛋白质l-ggsg-nluc融合蛋白结合至car或sir的κ轻链组分。这样,这些试剂的主要效用是用于检测car或sir的表达并且它们不能用于测量car或sir与其靶抗原的结合亲和力。表达mplcar的t细胞能够结合mpl--ggsg--nluc融合蛋白。将表达不同mplcar构建体或对照car(4c3)的jurkatt细胞与mpl-ggsg-nluc-acv5和cd19-ggsg-nluc-acv5上清液一起孵育并且在充分洗涤之后测定nluc活性,基本上如先前实例中所述。结果呈现于图10中并且证实表达不同mplcar构建体的jurkat细胞显示不同水平的与mpl-ggsg-nluc-acv5融合蛋白的结合。此不同可反应不同构建体的表达水平的差异或其相应scfv片段与mpl或两者的结合亲和力的差异。sir在其初代人类t细胞上的表达和通过nluc测定进行的检测..用表达靶向cd19的指示sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。将编码靶向mpl(161-sir-u01)的sir的慢病毒载体用作阴性对照。在感染之后,使细胞在含有10ng/ml可溶抗-cd3、10ng/ml可溶抗-cd28和100iu重组人类-il2的xvivo培养基中扩增并且用嘌呤霉素选择,除非另外指示。将表达不同sir构建体的t细胞与cd19-ggsg-nluc-acv5上清液一起孵育并且在充分洗涤之后测定nluc活性,基本上如先前实例中所述。数据显示cd19-ggsg-nluc-acv5与表达不同sir的t细胞的结合。与表达阴性对照sir161-sir-u01的t细胞(平均nluc值=1580)相比,表达cd8sp-fmc63-vl-v5-[htcrb-wt]-f-p2a-sp-fmc63-vh-myc-[htcra-wt]-f-f2a-pac(080815-f02)[seqidno:922]的t细胞不会显示与cd19-ggsg-nluc-acv5的任何显著结合(平均nluc值=1190),该抗体含有由野生型人类tcrα和人类tcrβ2编码的tcrα和tcrβ恒定区。相比之下,表达cd8sp-fmc63-vl-v5-[mtcrb-opt]-f-p2a-sp-fmc63-vh-myc-[mtcra-opt]-f-f2a-pac(080815-b06)[seqidno:953]的t细胞显示与cd19-ggsg-nluc-acv5的显著结合(平均nluc值=5359),该抗体含有由密码子优化的鼠类tcrα和鼠类tcrβ核苷酸序列编码的tcrα和tcrβ恒定区。表达cd8sp-fmc63-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-fmc63-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(050515-l05)[seqidno:900]的t细胞显示与cd19-ggsg-nluc-acv5的牢固结合(平均nluc值=19178)。此构建体含有由密码子优化的人类tcrα和人类tcrβ2核苷酸序列编码的tcrα和tcrβ恒定区并且携带tcrβ恒定链中的s57c突变和tcrα恒定链中的t48c突变。cd8sp-fmc63-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-fmc63-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(050515-l05)[seqidno:900]sir构建体也携带fmc63-vl区与tcrβ链之间的v5表位标签和fmc63-vh区与tcrα链之间的myc标签。表达cd8sp-cd19bu12-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-cd19bu12-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(070215-m03)[seqidno:1021]sir的t细胞显示与cd19-ggsg-nluc-acv5的更强结合(平均nluc值=39575)。此构建体类似于基于fmc63的050515-l05构建体,除了它具有源自hcd19-bu12抗体的vl和vh片段,该抗体是针对人类cd19的人源化单克隆抗体。表达cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(081415-d06)[seqidno:992]的t细胞显示与cd19-ggsg-nluc-acv5的最强结合(平均nluc值=107077)。此构建体含有由密码子优化的人类tcrα和人类tcrβ2核苷酸序列编码的tcrα(tcra)和tcrβ(tcrb)恒定区并且携带tcrβ恒定链中的s57c突变和tcrα恒定链中的t48c突变。fmc63-sir-d06构建体也携带fmc63-vl区与tcrβ链之间的v5表位标签和fmc63-vh区与tcrα链之间的myc标签。最后,在此构建体中,将tcrα和tcrβ恒定链鼠类化。因此,人类tcr-b恒定区的五个氨基酸被对应鼠类氨基酸置换。这些鼠类氨基酸是k-18、a-22、i-133、a-136和h-139。另外,在人类tcra恒定链中,含有四种氨基酸丝氨酸(s-90)、天冬氨酸(d-91)、缬氨酸(v-92)和脯氨酸(p-93)的鼠类tcra恒定链的区域被取代。表达cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-fmc63-vl-gly-ser-接头-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(082815-g07)[seqidno:1620]和cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-cd19bu12-vl-gly-ser-接头-cd19bu12-vh-myc-[htcra-csdvp]-f-f2a-pac(082815-e05)[seqidno:1622]sir构建体的t细胞也显示与cd19-ggsg-nluc-acv5的显著结合(分别为平均nluc值=29262和4671.5)。在082815-g07构建体中,fmc63-scfv片段[即fmc63(vl+vh)]融合至tcra-csdvp片段,而tcrb-kaciah恒定区片段被表达而无任何抗原结合部分。082815-e05构建体类似于082815-g07,除了cd19-bu12scfv片段代替存在于082815-g07构建体中的fmc63-scfv。总之,这些结果表明含有由野生型核苷酸序列编码的tcrα和tcrβ恒定区的sir未能显示与cd19的显著结合,这可能是由于此构建体在人类原代t细胞中的不良表达。相反,携带促进链间二硫键的额外半胱氨酸残基的含有密码子优化的人类tcra/b链的sir显示有效cd19结合和功能表达。如(081415-d06)[seqidno:992]sir中可见的人类tcrα/β恒定链的鼠类化导致cd19结合进一步增加。另外,如(082815-g07)[seqidno:1620]和(082815-e05)[seqidno:1622]构建体中可见,scfv片段在与tcrb恒定链共同表达时可以在功能上表达为融合至tcra恒定区,即使tcrb不携带任何抗原结合部分也如此(参见图11)。可替代地,此类构建体中的tcrb链可表达不相关vl或vh部分,只要它不干扰存在于scfv片段中的vl和vh链的功能组装。用不同构建体(seqidno:922、953、900、992、1110和1021)转导jurkat-nfat-luc细胞并且在嘌呤霉素中选择。将细胞与cd19-ggsg-nluc-acv5上清液一起孵育并且在充分洗涤之后测定nluc活性,基本上如先前所述。亲本jurkat和表达不同构建体的那些细胞的nluc值分别是996、3606、12128、37216、503043、101958和128996。实验显示表达含有具有野生型核苷酸序列的tcra/b恒定链的cd8sp-fmc63-vl-v5-[htcrb-wt]-f-p2a-sp-fmc63-vh-myc-[htcra-wt]-f-f2a-pac(080815-f02)[seqidno:922]构建体的jurkat与cd19-ggsg-nlucacv5上清液无显著结合,而与含有具有密码子优化的核苷酸序列的tcra/b恒定链且/或携带特异性氨基酸取代的其它构建体的不同水平的结合增强了链配对和/或表达。具体地说,具有seqidno:900和992的构建体分别显示cd19-ggsg-nluc结合的超过10倍和15倍增加。用不同sir构建体稳定转导jurkat-nfat-luc细胞并且在嘌呤霉素中选择。将细胞与cd19-ggsg-nluc-acv5上清液一起孵育并且在充分洗涤之后测定nluc活性,基本上如先前所述。不同sir构建体显示与cd19-ggsg-nluc-acv5融合蛋白的不同水平的结合。具体地说,含有源自tcr-γ和tcr-δ的tcr恒定链的构建体cd8sp-fmc63-vl-v5-[htcrg-opt]-f-p2a-sp-fmc63-vh-myc-[htcrd-opt]-f-f2a-pac(091015-a06)[seqidno:949]也显示与cd19-ggsg-nluc-acv5融合蛋白的结合。亲本jurkat和表达具有seqidno:1620、1623、1622、926、949、1112的构建体的那些细胞的nluc值分别是1515、27594、6357、10254、693、2176和179。因此,具有seqid:926的构建体再次显示与可溶性cd19的最低结合。用指示的构建体稳定转导jurkat-nfat-luc细胞并且在嘌呤霉素中选择。将细胞与cd19-ggsg-nluc-acv5上清液一起孵育并且在充分洗涤之后测定nluc活性,基本上如先前所述。不同sir构建体显示与cd19-ggsg-nluc-acv5融合蛋白的不同水平的结合。表达cd19sir的t细胞诱导表达cd19的raji淋巴瘤细胞中的细胞毒性。用表达靶向cd19的指示sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。用嘌呤霉素选择细胞并扩增。将细胞内稳定表达hgluc的raji细胞与表达sir的t细胞共同培养,其效应物:靶标(e:t)比率为10:1,持续4小时。通过直接注射含有天然腔肠素(nanaolight)的0.5xctz测定缓冲液来增加gluc活性(如通过bioteksynergy板读取器测量),测定靶细胞裂解的sir-t细胞介导的诱导。数据表明与靶向mpl的对照sircd8sp-mpl-161-vl-v5-[htcrb-s57c-opt1]-f-p2a-mpl-161-vh-myc-[htcra-t48c-opt1]-f-f2a-pac(040315-u02)[seqidno:1112]的t细胞相比在与表达cd19特异性sircd8sp-fmc63-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-fmc63-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(050515-l05)[seqidno:900]和cd8sp-cd19bu12-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-cd19bu12-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(070215-m03)[seqidno:1021]的t细胞共同培养之后gluc活性增加,这指示靶细胞裂解。将使用洋地黄皂苷的治疗用于显示最大细胞死亡。暴露于表达构建体(040315-u02)[seqidno:1112]、(050515-l05)[seqidno:900]、(070215-m03)[seqidno:1021]的t细胞和在洋地黄皂苷治疗之后的raji细胞的平均gluc值分别是119、3042、2547和3869。表达cd19sir的t细胞诱导表达cd19的raji淋巴瘤中的细胞毒性。用表达靶向cd19的指示sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞或使其未感染(t-ui)。用嘌呤霉素选择细胞并扩增。将稳定表达hgluc的raji细胞与表达sir的t细胞共同培养,其效应物:靶标(e:t)比率为10:1,持续4小时。通过直接注射含有天然腔肠素(nanaolight)的0.5xctz测定缓冲液来增加gluc活性(如通过bioteksynergy板读取器测量),测定靶细胞裂解的sir-t细胞介导的诱导。在指示时,在此实验和随后的实验中将含有单独培养基(med)的孔和靶细胞与未用任何sir构建体(t-ui)感染的t细胞孵育的孔中的gluc活性用作对照。数据表明与靶向mpl的对照sircd8sp-mpl-161-vl-v5-[htcrb-s57c-opt1]-f-p2a-mpl-161-vh-myc-[htcra-t48c-opt1]-f-f2a-pac(040315-u02)[seqidno:1112]的t细胞或未感染t细胞(t-ui)相比在与表达cd19特异性sircd8sp-fmc63-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-fmc63-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(050515-l05)[seqidno:900]、cd8sp-fmc63-vl-v5-[htcrb-wt]-f-p2a-sp-fmc63-vh-myc-[pretcra-del48]-f-f2a-pac(091015-y08)[seqidno:926]和cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(081415-d06)[seqidno:992]的t细胞共同培养之后gluc活性增加,这指示靶细胞裂解。将使用洋地黄皂苷的治疗用于显示最大细胞死亡。使用(091015-y08)[seqidno:926]构建体的结果显示pretcrα-del48恒定链片段可以取代tcrα恒定链区,以在与tcrβ的恒定链共同表达时制成功能性双链sir。请注意med指示单独的培养基,而t-ui表示未用任何sir构建体感染的t细胞。表达其中抗原结合结构域连接至tcr恒定链中的一条并缺乏互补tcr链的cd19单链sir的t细胞未能诱导表达cd19的raji淋巴瘤中的显著细胞毒性。生成sir构建体,其中源自针对人类cd19的cd19bu12和fmc63单克隆抗体的scfv片段融合至tcrb恒定链并且未表达互补tcra链。在构建体cd8sp-cd19bu12-vl-gly-ser-接头-cd19bu12-vh-v5-[htcrb-wt]-f-p2a-pac(051216-d08)[seqidno:1022]中,cd19bu12的scfv片段(表示为cd19bu12-vl-gly-ser-接头-cd19bu12-vh)经由v5接头连接至含有野生型核苷酸序列的tcrb恒定链。在构建体cd8sp-fmc63-vl-gly-ser-接头-fmc63-vh-v5-[htcrb-s57c-opt]-f-p2a-pac(051216-g01)[seqidno:912]中,fmc63的scfv片段(表示为fmc63-vl-gly-ser-接头-fmc63-vh)经由v5接头连接至含有其密码子优化的核苷酸序列并携带s57c突变的tcrb恒定链。在构建体cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-fmc63-vl-gly-ser-接头-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(082815-g07)[seqidno:1620]中,fmc63的scfv片段经由myc接头连接至含有其密码子优化的核苷酸序列并携带csdvp突变的tcra恒定链,该恒定链经由v5接头与含有其密码子优化的核苷酸序列并携带kaciah突变的tcrb恒定链共同表达。构建体cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(081415-d06)[seqidno:992]和cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-k13-flag-f-t2a-pac(051216-k04)[seqidno:918]是其中源自fmc63的vl和vh片段分别附接至htcrb-kaciah和htcra-csdvp链的双链片段。不同构建体在t细胞中表达并且仍测试其裂解raji-gluc细胞的能力。数据显示与未感染t细胞(t-ui)或含有单独培养基的孔相比,表达缺乏互补tcra恒定链的(051216-d08)[seqidno:1022]和(051216-g01)[seqidno:912]sir的t细胞未能诱导raji细胞裂解。暴露于表达构建体(082815-g07)[seqidno:1620]、(051216-k04)[seqidno:918]和(081415-d06))[seqidno:992]的t细胞的raji-gluc细胞显示与暴露于t-ui细胞的那些相比gluc活性增加超过2倍、超过2倍和超过4倍。表达其中抗原结合结构域连接至tcr恒定链中的一条并缺乏互补tcr链的cd19单链sir(scsir)的t细胞未能诱导表达cd19的raji淋巴瘤中的细胞毒性。用表达靶向cd19的指示sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。用嘌呤霉素选择细胞并扩增。将稳定表达hgluc的raji细胞与表达sir的t细胞共同培养,其效应物:靶标(e:t)比率为10:1,持续4小时。通过直接注射含有天然腔肠素(nanaolight)的0.5xctz测定缓冲液来增加gluc活性(如通过bioteksynergy板读取器测量),测定靶细胞裂解的sir-t细胞介导的诱导。数据显示与未感染t细胞(t-ui)或含有单独培养基的孔相比,表达sir构建体(051216-d08)[seqidno:1022]、(051216-f03)[seqidno:1023]和(051216-g01)[seqidno:912]sir的t细胞未能诱导raji细胞裂解,在该构建体中抗原结合结构域连接至tcrb恒定链并且缺乏互补tcra恒定链。表达cd19sir的t细胞诱导表达cd19的raji淋巴瘤中的细胞毒性。用表达靶向cd19的不同sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。将靶向tcrβ2链(cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-tcrb2-cp01-e05-vl-gly-ser-接头-tcrb2-cp01-e05-vh-myc-[htcra-csdvp]-f-f2a-pac(030816-c05)[seqidno:1781])的构建体用作阴性对照。使细胞未得到选择(对于构建体022216-a04和031416-a18)或用嘌呤霉素选择(对于其它构建体)并且扩增。测试细胞裂解raji-gluc细胞的能力。结果显示与未感染t细胞(t-ui)或具有单独培养基的孔(med)相比通过表达所有构建体的t细胞进行有效靶细胞裂解。具体而言,通过构建体(031616-b05)[seqidno:1019]、(031616-c05)[seqidno:1020]、(021816-n02)[seqidno:1016]观察到有效靶细胞裂解,在这些构建体中cd19bu12scfv片段附接至tcrb恒定链并且与空htcra恒定链共同表达。还通过构建体cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-2-cd19mm-vl-gly-ser-接头-cd19mm-vh-myc-[htcra-csdvp]-f-f2a-pac(031616-a050[seqidno:1623]观察到有效靶细胞裂解,在该构建体中cd19mmscfv经由myc接头附接至htcra-csdvp恒定链并且与携带v5接头的空htcrb-kaciah恒定链共同表达。还通过构建体cd8sp-cd19bu12-scfv-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-scfv-myc-[htcra-csdvp]-f-f2a-pac(020216-b07)[seqidno:1026]观察到有效靶细胞裂解,从而证实可以构建含有两个不同scfv片段的功能性sir,在该构建体中cd19bu12scfv附接至htcrb-kaciah链并且fmc63-scfv附接至htcra-csdvp链。最后,还通过构建体cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-k13-flag-f-t2a-cnb30(022216-a04)[seqidno:920]和cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63vh-myc-[htcra-csdvp]-f-p2a-cd3z-41bb-t2a-cnb30(031416-a18)[seqidno:998]观察到有效靶细胞裂解,在这些构建体中fmc63源性vl和vh片段分别附接至htcrb-kaciah和htcra-csdvp恒定链。然而,这些构建体分别共同表达卡波济氏肉瘤相关疱疹病毒编码的病毒flice抑制性蛋白质(vflip)k13和融合蛋白cd3z-41bb。已知k13蛋白通过结合至细胞蛋白质nemo来选择性活化nf-κb途径,而cd3z-41bb融合蛋白含有连接至tcr复合物的cd3z链的共刺激分子41bb的细胞溶质信号传导结构域。通过提供共刺激信号,k13和cd3z-41bb蛋白将增强表达sir的细胞的活化和增殖,从而产生更好的功能性和长期持久性。022216-a04和031416-a18构建体还表达钙调磷酸酶b链的cnb30突变体,该链赋予对钙调磷酸酶抑制剂诸如fk506(他克莫司)的抗性。表达靶向cd19的双链(dc)和一条半链(oah)sir诱导表达cd19的raji淋巴瘤中的细胞毒性。用表达靶向cd19的不同sir构建体的慢病毒感染t细胞并且测试针对raji-gluc细胞的细胞毒性。数据显示与靶向mpl的阴性对照构建体cd8sp-mpl-161-vl-v5-[htcrb-s57c-opt1]-f-p2a-mpl-161-vh-myc-[htcra-t48c-opt1]-f-f2a-pac(040315-u02)[seqidno:1112]相比,含有由其野生型核苷酸序列(即分别为seqidno:746和seqidno:731)编码的tcrβ和tcrα恒定链的cd8sp-fmc63-vl-v5-[htcrb-wt]-f-p2a-sp-fmc63-vh-myc-[htcra-wt]-f-f2a-pac(080815-f02)[seqidno:922]sir构建体未能引起显著靶细胞裂解。相比之下,所有其它sir构建体(例如(050515-l05)[seqidno:900]、(070215-m03)[seqidno:1021]、(081415-d06)[seqidno:992]、(080815-b06)[seqidno:953]、(082815-g07)[seqidno:1620]和(082815-e05)[seqidno:1622])显示gluc活性增加,从而指示靶细胞裂解。(082815-g07)[seqidno:1620]和(082815-e05)[seqidno:1622]构建体表达分别融合至fmc63和cd19bu12scfv片段的空htcrb-kaciah(seqidno:748)恒定链连同htcra-csdvp(seqidno:732)恒定链片段。因此,其中抗原结合结构域融合至tcra恒定链的sir在与空互补tcrb恒定链共同表达时可以诱导有效靶细胞裂解。此类sir构建体中的空tcrb恒定链可能有助于携带tcra恒定链的抗原结合结构域的细胞表面表达。表达cd19bu12sir且具有tcra和tcrb野生型核苷酸序列的t细胞未能诱导表达cd19的raji淋巴瘤中的细胞毒性。将raji-gluc细胞与表达靶向cd19的不同sir的t细胞共同培养,其效应物:靶标(e:t)比率为10:1,持续4小时。像之前一样测定靶细胞裂解的sir-t细胞介导的诱导。数据显示与可能是由于021916-q03sir构建体在人类原代t细胞中表达较差而引起的阴性对照构建体111815-o05或未感染t细胞相比,表达基于cd19bu12的cd8sp-cd19bu12-vl-v5-[htcrb-wt]-f-p2a-cd19bu12-vh-myc-[htcra-wt]-f-f2a-pac(021916-q03)[seqidno:1038]sir构建体且含有由其野生型核苷酸序列编码的tcrb和tcra恒定链的t细胞未能引起显著靶细胞裂解,。因此,类似于cd8sp-fmc63-vl-v5-[htcrb-wt]-f-p2a-sp-fmc63-vh-myc-[htcra-wt]-f-f2a-pac(080815-f02)[seqidno:922]构建体,另一种基于tcra和tcrb的野生型核苷酸序列的sir未能诱导针对靶细胞系的显著毒性。表达靶向cd19的一条半链(oahsir)的t细胞诱导表达cd19的raji淋巴瘤中的细胞毒性。用编码靶向cd19的不同sir的慢病毒感染人类外周血t细胞并且使用raji-gluc细胞测试细胞毒性,其效应物:靶标(e:t)比率为1:1,持续96小时。数据显示与阴性对照构建体040315-u02相比,含有由野生型核苷酸序列编码的tcrb和tcra恒定链的cd8sp-fmc63-vl-v5-[htcrb-wt]-f-p2a-sp-fmc63-vh-myc-[htcra-wt]-f-f2a-pac(080815-f02)[seqidno:922]sir构建体未能引起显著靶细胞裂解,这可能是由于此(即080815-f02)构建体在人类初代t细胞中表达较差。相比之下,所有其它sir构建体(例如050515-l05、070215-m03、081415-d06、080815-b06、082815-g07和082815-e05)显示gluc活性增加,这指示靶细胞裂解。082815-g07和082815-e05构建体表达tcrb恒定链(kaciah版本)连同分别与fmc63和cd19bu12scfv片段融合的tcra(csdvp)片段。因此,基于融合至tcra恒定链的抗原结合结构域的sir在与空tcrb恒定链共同表达时可以诱导有效细胞裂解。此sir被指定为一条半(oahsir)。因此,oahsir比单链sir(scsir)更有效。这可能是因为tcra和tcrb需要互补链在t细胞上有效表达。表达靶向cd19的双链(dcsir)的t细胞诱导表达cd19的raji淋巴瘤中的细胞毒性。将raji-gluc细胞与表达靶向cd19的不同sir的t细胞共同培养,其效应物:靶标(e:t)比率为10:1,持续4小时,随后测量gluc活性。数据显示与未感染t细胞或表达靶向cd4或kshv蛋白质的阴性对照car的t细胞相比,表达cd8sp-fmc63-vl-myc-[htcra-t48c-opt1]-f-f2a-fmc63-vh-v5-[htcrb-c57c-opt1]-f-p2a-pac(100515-e03)[seqidno:902]构建体的t细胞可以诱导有效靶细胞裂解,在该构建体中fmc63-vl链连接至htcra-t48c-opt1恒定链并且fmc63-vh连接至htcrb-c57c-opt1恒定链。类似地,表达cd8sp-fmc63-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-fmc63-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(050515-l05)[seqidno:900]和cd8sp-fmc63-vl-v5-[htcrb-s57c-opt]-f-p2a-sp-fmc63-vh-myc-[htcra-t48c-opt]-f-f2a-pac(100815-b04)[seqidno:951]构建体的t细胞诱导了有效靶细胞裂解,在这些构建体中fmc63-vl链连接至tcrb-s57c-opt1或htcrb-s57c-opt恒定链并且fmc63-vh连接至tcra-t48c-opt1或htcra-t48c-opt恒定链。最后,表达ighsp-fmc63-vh-[htcrb-c57c-opt]-f-p2a-cd8sp-fmc63-vl-myc-[htcra-t48c-opt]-f-f2a-pac(101415-m05)[seqidno:901]构建体的t细胞也可以诱导有效靶细胞裂解,在该构建体中fmc63-vh链连接至htcrb-c57c-opt恒定链并且fmc63-vl连接至htcra-t48c-opt恒定链。因此,在双链sir构建体中,抗体的vl片段可以连接至tcra或tcrb恒定链并且vh片段连接至互补tcrb或tcra恒定链中的任一者。另外,在双链sir中,在两个功能多肽单元(fpu)由同一载体表达时,含有fpu的tcrb或tcra链可以是第一(或5’)fpu。表达cd20sir的t细胞诱导表达cd20的raji细胞中的细胞毒性。将raji-gluc细胞与表达靶向cd19和cd20的不同sir的t细胞共同培养,其效应物:靶标(e:t)比率为10:1,持续4小时,随后测量gluc活性。数据显示与未感染t细胞或含有单独培养基(med)的孔相比表达cd8sp-cd20-2f2-vl-v5-[htcrb-kaciah]-f-p2a-sp-cd20-2f2-vh-myc-[htcra-csdvp]-f-f2a-pac(100615-d05)[seqidno:1221]sir的t细胞有效诱导靶细胞裂解。使用靶向cd19的sir构建体(cd8sp-fmc63-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-fmc63-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(050515-l05)[seqidno:900]和cd8sp-fmc63-vl-v5-[htcrb-s57c-opt]-f-p2a-sp-fmc63-vh-myc-[htcra-t48c-opt]-f-f2a-pac(100815-b04)[seqidno:951])也观察到适度细胞毒性。表达含有犬类tcra和犬类tcrb恒定链的t细胞诱导表达cd20的raji细胞的细胞毒性。使用表达靶向cd20的sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞,在该sir构建体中单克隆抗体针对人类cd20的vl和vh片段(2f2)连接至密码子优化的犬类tcra和tcrb恒定链。在以10:1的e:t比率共同培养4小时之后,测试细胞针对raji-gluc细胞的细胞毒性。.数据显示与未感染t细胞相比,表达基于犬类tcrb和tcra恒定链的cd8sp-cd20-2f2-vl-[犬类-tcrb-opt]-f-p2a-cd20-2f2-vh-[犬类-tcra-opt]-f-f2a-pac(051716-e02)[seqidno:1113]的t细胞有效诱导靶细胞裂解。表达lym1sir的t细胞诱导表达lym1的kasumi--1细胞中的细胞毒性。用编码cd8sp-lym1-vl-[htcrb-opt2]-f-p2a-sp-lym1-vh-[htcra-opt2-del]-f-f2a-pac(012716-b01)[seqidno:1185]sir的慢病毒感染人类t细胞并且在用嘌呤霉素选择之后在10:1的效应物:靶标(e:t)比率下持续4小时后测试针对kasumi-1-gluc细胞的细胞毒性或者以未感染t细胞(t-ui)作为对照。通过增加gluc活性来测定细胞毒性。数据显示与未感染t细胞或含有单独培养基的孔相比,表达(012716-b01)[seqidno:1185]sir的t细胞有效诱导靶细胞裂解,且gluc活性增加将近9倍。表达lym1和lym2sir的t细胞诱导表达lym1和lym2的raji细胞中的细胞毒性。在以10:1的e:t比率共同培养4小时之后,测试表达靶向lym1和lym2的sir的t细胞针对raji-gluc细胞的细胞毒性。来自gluc-细胞毒性测定的数据显示与表达对照sir(seqidno:4639)的t细胞、未感染t细胞或单独培养基相比,cd8sp-lym1-vl-v5-[htcrb-kaciah]-f-p2a-sp-lym1-vh-myc-[htcra-csdvp]-f-f2a-pac(021216-h02)[seqidno:1314]和cd8sp-lym2-vl-v5-[htcrb-kaciah]-f-p2a-sp-lym2-vh-myc-[htcra-csdvp]-f-f2a-pac(100615-b07)[seqidno:1315]sir构建体有效诱导靶细胞裂解,且gluc值分别高将近超过20倍和10倍。082815-p08是含有cd19bu12scfv并靶向cd19抗原的常规car。表达基于cd19bu12的(021916-q03)[seqidno:1038]sir构建体的t细胞再次未能引起显著靶细胞裂解,该构建体含有由其野生型核苷酸序列编码的tcrb和tcra恒定链。表达针对cs1(slamf7或cd319)的sir的t细胞诱导表达cs1的l363和u266细胞中的细胞毒性。用表达靶向cs1(slamf7)的sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。使用嘌呤霉素选择细胞并且在10:1的e:t比率下持续4小时后测试其针对l363-gluc和u266-gluc细胞的细胞毒性。gluc-细胞毒性测定显示与未感染t细胞或含有单独培养基的孔相比,表达cd8sp-cs1-huluc90-vl-v5-[htcrb-kaciah]-f-p2a-sp-huluc90-vh-myc-[htcra-csdvp]-f-f2a-pac(012716-a02)[seqidno:1254]sir的t细胞有效诱导l363-gluc和u266-gluc裂解,如由gluc值将近超过15倍和10倍增加所证明。表达针对bcma(b细胞成熟抗原或tnfrsf17或cd269)和csi的sir的t细胞诱导表达bcma的l363和u266细胞中的细胞毒性。用表达靶向cs1(slamf7)和bcma的sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。使用嘌呤霉素选择细胞并且在10:1的e:t比率下持续4小时后测试针对l363-gluc和u266-gluc细胞的细胞毒性。gluc细胞毒性测定显示与未感染t细胞、表达对照sircd8sp-kshv-4c3-vl-v5-[htcrb-s57c-opt]-f-p2a-sp-4c3-vh-myc-[htcra-t48c-opt]-f-f2a-pac(111815-o05)[seqidno:4639]的t细胞或含有单独培养基的孔相比,表达cd8sp-bcma-huc12a3-l3h3-vl-v5-[htcrb-kaciah]-f-p2a-sp-bcma-huc12a3-l3h3-vh-myc-[htcra-csdvp]-f-f2a-pac(011116-a07)[seqidno:1212]和cd8sp-cs1-huluc90-vl-v5-[htcrb-kaciah]-f-p2a-sp-huluc90-vh-myc-[htcra-csdvp]-f-f2a-pac(012716-a02)[seqidno:1254]sir的t细胞有效诱导靶细胞裂解。通过表达靶向lamp1的sircd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-lamp1-mb4-vl-gly-ser-接头-lamp1-mb4-vh-myc-[htcra-csdvp]-f-f2a-pac(050216-f05)[seqidno:1732]和靶向cmet和her3的双特异性car041316-f06的t细胞也观察到轻度至适度靶细胞裂解。表达针对cd138、cs1、gprc5d和wt1的sir的t细胞诱导u266和l363细胞中的细胞毒性。使用嘌呤霉素选择表达靶向cd138、cs1、gprc5d和wt1的sir构建体的t细胞并且在2:1的e:t下持续72小时后针对稳定表达gluc的u266和l363细胞进行测试。gluc-细胞毒性测定显示与未感染t细胞或含有单独培养基(med)的孔相比,表达cd8sp-cd138-vl-v5-[htcrb-kaciah]-f-p2a-sp-cd138-vh-myc-[htcra-csdvp]-f-f2a-pac(100815-a05)[seqidno:1236]sir的t细胞有效诱导靶细胞裂解。表达含有野生型tcra和tcrb恒定链的sircd8sp-cd138-vl-v5-[htcrb-wt]-f-p2a-sp-cd138-vh-myc-[htcra-wt]-f-f2a-pac(021916-r04)[seqidno:1139]的t细胞在u266细胞中仅具最低有效性并且在l363细胞中为无效的。单链sircd8sp-cd138-vl-gly-ser-接头-cd138-vh-myc-[htcra-csdvp]-f-f2a-pac(030316-g03)[seqidno:1169]仅在u266细胞中有效,在该单链sir中cd138scfv附接至htcra-csdvp恒定链。双特异性构建体lamp1-humab1-2-vl-v5-[tcrb-kaciah]-f-p2a-gprc5d-et150-18-vh-myc-[tcra-csdvp]-f-f2a-pac-e05(092916-e05-vn)(seqidno:1163)也有效地诱导两种细胞类型中的细胞毒性,该双特异性构建体携带针对lamp1的抗体vl片段和针对gprc5d的抗体vh片段。最后,使用靶向cs1和wt1的构建体观察到轻度至适度细胞毒性,尤其是在u266细胞中。构建体针对cs1(seqidno:1674、1253)和wt1(seqidno:1804和1805)的有限细胞毒性可能是由于在此实验中使用较低e:t比率。表达针对cll1的sir的t细胞诱导表达cll1的hl60细胞中的细胞毒性。使用嘌呤霉素选择表达靶向cll1的sircd8sp-cll1-m26-vl-[htcrb-opt2]-f-p2a-sp-cll1-m26-vh-[htcra-opt2]-f-f2a-pac(012616-a05)[seqidno:4790]的t细胞并且在10:1的e:t比率下持续4小时后针对hl60-gluc细胞进行测试。gluc细胞毒性测定显示与未感染t细胞或含有单独培养基的孔相比,表达(012616-a05)[seqidno:4790]的t细胞有效诱导靶细胞裂解。表达针对clec5a和cll1的sir的t细胞诱导表达clec5a和cll1的hl60细胞中的细胞毒性。使用表达针对clec5a和cll1的sir的t细胞重复该实验。结果显示与未感染t细胞或含有单独培养基的孔相比,表达cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-clec5a-8h8f5-vl-gly-ser-接头-clec5a-8h8f5-vh-myc-[htcra-csdvp]-f-f2a-pac(042816-e05)[seqidno:1666]、cd8sp-cll1-m26-vl-[htcrb-opt2]-f-p2a-sp-cll1-m26-vh-[htcra-opt2]-f-f2a-pac(012616-a05)[seqidno:4790]和cd8sp-cll1-m32-vl-v5-[htcrb-kaciah]-f-p2a-sp-cll1-m32-vh-myc-[htcra-csdvp]-f-f2a-pac(021216-i03)[seqidno:1250]sir的t细胞有效诱导hl60-gluc细胞裂解。在用表达三种sir的t细胞处理的细胞中,gluc值分别高将近9倍、4倍和7倍。表达靶向csf2ra、lamp1和cll1的sir的t细胞诱导表达csf2ra、lamp1和cll1的thp1细胞中的细胞毒性。将表达不同sir的t细胞与thp-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定来测试。结果显示与未感染t细胞或含有单独培养基的孔相比,表达分别靶向csf2ra、lamp1和cll1的cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-csf2ra-ab1-vl-gly-ser-接头-csf2ra-ab1-vh-myc-[htcra-csdvp]-f-f2a-pac(050216-b02)[seqidno:1676]、cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-lamp1-mb4-vl-gly-ser-接头-lamp1-mb4-vh-myc-[htcra-csdvp]-f-f2a-pac(050216-f05)[seqidno:1732]和cd8sp-cll1-m32-vl-v5-[htcrb-kaciah]-f-p2a-sp-cll1-m32-vh-myc-[htcra-csdvp]-f-f2a-pac(021216-i03)[seqidno:1250]sir的t细胞有效诱导靶细胞裂解。表达靶向csf2ra的sir的t细胞诱导表达csf2ra的molm13细胞中的细胞毒性。将表达靶向csf2ra的sir的t细胞与molm13-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定来测试。结果显示与未感染t细胞或表达靶向kshv蛋白质(111815-o05)的sir的t细胞或含有单独培养基的孔相比,表达靶向csf2ra的sircd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-csf2ra-ab1-vl-gly-ser-接头-csf2ra-ab1-vh-myc-[htcra-csdvp]-f-f2a-pac(050216-b02)[seqidno:1676]的t细胞有效诱导靶细胞裂解。表达靶向tshr(甲状腺刺激激素受体)和tnag的sir的t细胞诱导表达tshr和tnag的jurkat和peer细胞中的细胞毒性。将表达靶向tshr和tnag的不同sir的t细胞与jurkat-gluc和peer1-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定来测试。结果显示与未感染t细胞、表达阴性对照sir(111815-o05、042816-h07、031516-j07)的t细胞或含有单独培养基的孔相比,分别表达靶向tshr和tnag的sircd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-tshr-kb1-vl-gly-ser-接头-tshr-kb1-vh-myc-[htcra-csdvp]-f-f2a-pac(042916-e03)[seqidno:1795]和cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-tnag-vl-gly-ser-接头-tnag-vh-myc-[htcra-csdvp]-f-f2a-pac(050216-a04)[seqidno:1788]的t细胞有效诱导靶细胞裂解。表达靶向tnag的sir的t细胞诱导表达tnag的jurkat和peer细胞中的细胞毒性。将表达靶向tnag的sir的t细胞与jurkat-gluc和peer1-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定来测试。结果显示与未感染t细胞或含有单独培养基的孔相比,表达靶向tnag的sircd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-tnag-vl-gly-ser-接头-tnag-vh-myc4-[pretcra-del48]-f-f2a-pac(080816-h06)[seqidno:2003]的t细胞有效诱导靶细胞裂解。表达靶向mpl(tpo受体)的sir的t细胞诱导表达mpl的细胞中的细胞毒性。将表达靶向mpl的sir的t细胞与hel-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定来测试。结果显示与未感染t细胞或含有单独培养基的孔相比,表达靶向mpl的sircd8sp-mpl-175-vl-[htcrb-opt2]-f-p2a-sp-175-vh-[htcra-opt2]-f-f2a-pac(042116-g01)[seqidno:4862]的t细胞有效诱导靶细胞裂解。表达靶向flt3的sir的t细胞诱导表达flt3的rs:411(或rs411)细胞中的细胞毒性。将表达靶向flt3的sir的t细胞与rs:411-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定来测试。数据显示与未感染t细胞、表达阴性对照sir(111815-o05)的t细胞或含有单独培养基的孔相比,表达靶向flt3的sircd8sp-flt3-nc7-vl-v5-[htcrb-kaciah]-f-p2a-sp-flt3-nc7-vh-myc-[htcra-csdvp]-f-f2a-pac(050316-c01)[seqidno:1273]的t细胞有效诱导靶细胞裂解。表达表达nkg2d细胞外结构域的sir和靶向flt3和csf2ra的sir的t细胞诱导mv411靶细胞中的细胞毒性。在效应物:靶标(e:t)比率10:1下持续4小时后针对mv411-gluc细胞测试表达nkg2d的细胞外结构域(经由ggggs-ggggd-myc接头连接至htcr-csdvp恒定链)的人类外周血t细胞和靶向flt3和csf2ra的那些细胞。gluc细胞毒性测定显示与未感染t细胞、表达对照sircd8sp-kshv-4c3-vl-v5-[htcrb-s57c-opt]-f-p2a-sp-4c3-vh-myc-[htcra-t48c-opt]-f-f2a-pac(111815-o05)[seqidno:4639]sir的t细胞或含有单独培养基的孔相比,表达cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-nkg2d-(ggggs-ggggd)-myc-[htcra-csdvp]-f-f2a-pac(042916-a06)[seqidno:1755]和cd8sp-flt3-nc7-vl-v5-[htcrb-kaciah]-f-p2a-sp-flt3-nc7-vh-myc-[htcra-csdvp]-f-f2a-pac(050316-c01)[seqidno:1273]和cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-csf2ra-ab1-vl-gly-ser-接头-csf2ra-ab1-vh-myc-[htcra-csdvp]-f-f2a-pac(050216-b02)[seqidno:1676]sir的t细胞有效诱导靶细胞裂解。以上结果证明其中抗原结合结构域包含受体(即nkg2d)的sir具有功能活性。表达cd30和wt1sir的t细胞诱导表达u266和l363靶细胞中的细胞毒性。可以生成针对细胞内肽的sir,该细胞内肽可与特异性hla抗原联合识别。用表达靶向cd30或wt1的指示sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。wt1sir识别源自wt1的肽(rmfpnapyl)连同hla-a2分子。用嘌呤霉素选择细胞并扩增。将稳定表达hgluc的u266(wt1+/hla-a2+)和l-363(wt1+/hla-a2+)细胞与表达sir的t细胞共同培养,其效应物:靶标(e:t)比率为10:1,持续4小时。gluc细胞毒性测定显示与未感染t细胞(t-ui)相比,虽然表达sircd8sp-wt1-ab1-vl-v5-[htcrb-s57c-opt]-f-p2a-sp-wt1-ab1-vh-myc-[htcra-t48c-opt]-f-f2a-pac(012816-g01)[seqidno:4709]的t细胞是最低效的,但是表达sircd8sp-wt1-ab5-vl-v5-[htcrb-s57c-opt]-f-p2a-sp-wt1-ab5-vh-myc-[htcra-t48c-opt]-f-f2a-pac(111815-c04)[seqidno:4710]的t细胞有效杀伤靶细胞。另外,靶向cd30的sircd8sp-cd30-ac10-vl-v5-[htcrb-kaciah]-f-p2a-sp-cd30-ac10-vh-myc-[htcra-csdvp]-f-f2a-pac(010716-k01)[seqidno:1227]有效杀伤表达cd30的靶细胞。表达靶向cd33和cd179b的sir的t细胞诱导表达cd33和cd179b的hl60和molm13细胞中的细胞毒性。将表达靶向cd33和cd179b的sir的t细胞与hl60-gluc和molm13-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定测试。数据显示与未感染t细胞或含有单独培养基的孔相比,分别表达靶向cd33和cd179b的sircd8sp-cd33-af5-vl-v5-[tcrβ-kaciah]-f-p2a-sp-cd33-af5vh-myc-[tcrα-csdvp]-f-f2a-pac(052416-k05)[seqidno:1229]和cd179b-vl-v5-{tcrβ-kaciah}-f-p2a-sp-cd179b-vh-myc-[tcrα-csdvp]-f-f2a-pac(063016-y06)[seqidno:1237]的t细胞有效诱导靶细胞裂解。表达靶向cd33的sir的t细胞诱导表达cd33的hl60细胞中的细胞毒性。将表达靶向cd33的sir的t细胞与hl60-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定来测试。实验显示与未感染t细胞或含有单独培养基的孔相比,表达靶向cd33的sircd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-cd33-humyc9-vl-gly-ser-接头-cd33-humyc9-vh-myc-[htcra-csdvp]-f-f2a-pac(090116-c02)[seqidno:1650]和cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-cd33-af5-vl-gly-ser-接头-cd33-af5-vh-myc4-[pretcra-del48]-f-f2a-pac(083116-e02)[seqidno:1864]和cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-cd33-humyc9-vl-gly-ser-接头-cd33-humyc9-vh-myc4-[pretcra-del48]-f-f2a-pac(083116-c06)[seqidno:1865]的t细胞有效诱导靶细胞裂解。表达靶向cxcr4的sir的t细胞诱导表达cxcr4的thp细胞中的细胞毒性。用表达靶向cxcr4的指定双特异性sir构建体cd8sp-cxcr4-1-vhh-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdv]-f-f2a-pac(101415-v01)[seqidno:1171]的慢病毒感染用cd3磁珠分离的人类外周血t细胞。该sir构建体还表达fmc63抗体针对cd19的vh片段。将细胞与hl60-gluc细胞以5:1的e:t比率培养4小时并且使用gluc测定测量细胞毒性。结果显示与未感染t细胞或含有单独培养基的孔相比,表达靶向cxcr4的sir的t细胞有效诱导靶细胞裂解。表达针对il11ra的sir的t细胞诱导表达il11ra的bv173细胞中的细胞毒性。将表达靶向il11ra的sir的t细胞与thp-1-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定来测试。结果显示与t-ui细胞或单独培养基相比,表达cd8sp-il11ra-8e2-ts107-vl-v5-[htcrb-kaciah]-f-p2a-sp-il11ra-8e2-ts107-vh-myc-[htcra-csdvp]-f-f2a-pac(050516-r06)[seqidno:1304]sir的t细胞有效诱导bv173靶细胞裂解。表达cd16sir的t细胞与cd20单克隆抗体利妥昔单抗结合诱导表达cd20的raji淋巴瘤中的细胞毒性。用表达sir构建体cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp2-cd16a-v158-v2-myc-[htcra-t48c-opt]-f-f2a-pac(020416-a08)[seqidno:1186的慢病毒感染用cd3磁珠分离的外周血t细胞,该构建体表达对于人类igg具有高亲和力的cd16a变体v158a。将针对kshv蛋白的4c3sir用作阴性对照。用嘌呤霉素选择细胞并扩增。将稳定表达hgluc的raji细胞与表达sir的t细胞共同培养,其效应物:靶标(e:t)比率为10:1,持续4小时,这是在利妥昔单抗(1μg/ml)存在和不存在的情况下进行的。通过直接注射含有天然腔肠素(nanaolight)的0.5xctz测定缓冲液来增加gluc活性(如通过bioteksynergy板读取器测量),测定靶细胞裂解的sir-t细胞介导的诱导。结果显示仅在利妥昔单抗存在下cd16a-v158sir构建体进行有效靶细胞裂解。因此,cd16av158sir可以用作通用sir,其与任何单克隆抗体一起使用,因此避免了对制成针对不同抗原靶标的个别sir的需要。表达cd123--161双特异性sir的t细胞诱导表达mpl的bv173和hel细胞中的细胞毒性。用表达靶向cd123和mpl(161)的ighsp-cd123-2-vhh-v5-[htcrb-s57c-opt]-f-p2a-cd8sp-mpl-161-hl-myc-[htcra-t48c-opt]-f-f2a-pac(022516-m08)[seqidno:4591]sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。用嘌呤霉素选择细胞并扩增。将稳定表达hgluc的bv173和hel细胞与表达sir的t细胞共同培养,其效应物:靶标(e:t)比率为10:1,持续4小时。gluc细胞毒性测定显示双特异性sir可以诱导表达其一种或多种靶抗原的靶细胞的有效裂解。表达针对cd123的sir的t细胞诱导表达cd123的l428细胞中的细胞毒性。将表达靶向cd123的sir的t细胞与l428-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定来测试。实验显示与t-ui细胞或单独培养基相比,表达cd8sp-cd123-csl362-vl-v5-[htcrb-kaciah]-f-p2a-sp-cd123-csl362-vh-myc-[pretcra-del48]-f-f2a-pac(041416-k04)[seqidno:1445]sir的t细胞有效诱导l428靶细胞裂解。表达针对cd123的sir的t细胞诱导表达cd123的bv173细胞中的细胞毒性。将表达靶向cd123的sir的t细胞与bv173-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定来测试。结果显示与t-ui细胞或单独培养基(med)相比,表达ighsp-cd123-2-vhh-v5-[htcrb-kaciah]-f-p2a-sp-cd123-1-vhh-myc-[pretcra-del48]-f-f2a-pac(041416-v03)[seqidno:1467]的t细胞有效诱导bv173靶细胞裂解。此sir含有分别附接至htcrb-kaciah和pretcra-del48恒定链的两个不同骆驼科vhh(cd123-2和cd123-1)。这些结果表明含有两个不同抗原结合结构域的sir均具有功能活性。表达靶向cd79b和cd138的sir的t细胞诱导表达cd79b和cd138的raji和l363细胞中的细胞毒性。将表达靶向cd79b和cd138的sir的t细胞与raji-gluc和l363-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定来测试。结果显示与未感染t细胞或含有单独培养基的孔相比,分别表达靶向cd79b和cd138的cd8sp-cd79b-2f2-vl-v5-[htcrb-s57c-opt]-f-p2a-sp-cd79b-2f2-vh-myc-[pretcra-del48]-f-f2a-pac(041216-h05)[seqidno:1130]和cd8sp-cd138-vl-v5-[htcrb-kaciah]-f-p2a-sp-cd138-vh-myc-[pretcra-del48]-f-f2a-pac(041416-i03)[seqidno:1446]sir的t细胞有效诱导靶细胞裂解。表达tcrβ1sir的t细胞诱导表达tcrβ1的jurkat细胞中的细胞毒性。用表达靶向tcrb1(tcrβ1)恒定链的cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-tcrb1-cp01-e09-vl-gly-ser-接头-tcrb1-cp01-e09-vh-myc-[htcra-csdvp]-f-f2a-pac(030816-d04)[seqidno:1778]和cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-tcrb1-jovi1-vl-gly-ser-接头-tcrb1-jovi1-vh-myc-[htcra-csdvp]-f-f2a-pac(030816-b07)[seqidno:1779]sir构建体和靶向cd30的cd8sp-cd30-ac10-vl-v5-[htcrb-kaciah]-f-p2a-sp-cd30-ac10-vh-myc-[htcra-csdvp]-f-f2a-pac(063016-k02)[seqidno:1227]sir的慢病毒感染用cd3磁珠分离的人类外周血t细胞。用嘌呤霉素选择细胞并扩增。将稳定表达hgluc的jurkat细胞与表达sir的t细胞共同培养,其效应物:靶标(e:t)比率为10:1,持续4小时。通过增加gluc活性来测定靶细胞裂解的sir-t细胞介导的诱导。结果显示cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-tcrb1-jovi1-vl-gly-ser-接头-tcrb1-jovi1-vh-myc-[htcra-csdvp]-f-f2a-pac(030816-b07)[seqidno:1779]和cd8sp-cd30-ac10-vl-v5-[htcrb-kaciah]-f-p2a-sp-cd30-ac10-vh-myc-[htcra-csdvp]-f-f2a-pac(063016-k02)[seqidno:1227]诱导有效靶细胞裂解,而cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-tcrb1-cp01-e09-vl-gly-ser-接头-tcrb1-cp01-e09-vh-myc-[htcra-csdvp]-f-f2a-pac(030816-d04)[seqidno:1778]不是那么有效。表达tcrβ2sir的t细胞诱导表达含有tcrβ2恒定链的sir的jurkat细胞中的细胞毒性。在初始实验中,在细胞毒性测定中表达针对tcrβ2链的sir的t细胞是无功能活性的。原因在于,这可能是由于以下事实:这些sir中的tcrβ恒定链和因此表达sir的t细胞将具有自杀死亡或自相残杀者(即杀伤掉相邻的表达sir的t细胞)。为了避免此问题,使用基于tcrβ1链的tcrβ1-opt4恒定链(核酸seqidno:752和氨基酸seqidno:3032)生成靶向tcrβ2的sir。用表达靶向tcrb2(tcrβ2)恒定链的指定sir(cd8sp-tcrb2-d05-vl-[htcrb-opt4]-f-p2a-sp-tcrb2-d05-vh-myc-[htcra-csdvp]-f-f2a-pac-k06(072816-k06)[seqidno:1129]和cd8sp-tcrb2-cp01-e05-vl-[htcrb-opt4]-f-p2a-sp-tcrb2-cp01-e05-vh-myc-[htcra-csdvp]-f-f2a-pac(072816-l06)[seqidno:1128])构建体感染用cd3磁珠分离的人类外周血t细胞。用嘌呤霉素选择细胞并扩增。对于靶细胞系,使用稳定表达靶向psma的含有tcrb2恒定链的sir的jurkat细胞。jurkat细胞还共同表达turboluc的信号肽缺陷型版本作为报道基因。将jurkat细胞与表达sir的t细胞共同培养,其效应物:靶标(e:t)比率为10:1,持续4小时。通过直接注射含有天然腔肠素(nanaolight)的0.5xctz测定缓冲液来增加turboluc活性(如通过bioteksynergy板读取器测量),测定靶细胞裂解的sir-t细胞介导的诱导。结果显示当与未感染t细胞(t-ui)或单独培养基相比时,表达cd8sp-tcrb2-d05-vl-[tcrb-opt4]-f-p2a-sp-tcrb2-d05-vh-myc-[tcrα-csdvp]-f-f2a-pac-k06(072816-k06)[seqidno:1129]和cd8sp-tcrb2-cp01-e05-vl-[htcrb-opt4]-f-p2a-sp-tcrb2-cp01-e05-vh-myc-[htcra-csdvp]-f-f2a-pac(072816-l06)[seqidno:1128]sir的t细胞诱导靶细胞有效裂解,如通过turboluc活性增加所显示的。表达叶酸受体1(fr1)sir的t细胞诱导表达fr1的skov3、pc3和lncap细胞中的细胞毒性。用表达靶向fr1的指定sir构建体(cd8sp-fr1-humov19-vl-v5-[htcrb-kaciah]-f-p2a-sp-fr1-humov19-vh-myc-[htcra-csdvp]-f-f2a-pac(102915-p07)[seqidno:1276])的慢病毒感染用cd3磁珠分离的人类外周血t细胞。用嘌呤霉素选择细胞并扩增。将稳定表达hgluc的skov3、pc3和lncap细胞与表达sir的t细胞共同培养,其效应物:靶标(e:t)比率为10:1,持续4小时。通过增加gluc活性来测定靶细胞裂解的sir-t细胞介导的诱导。结果显示与未感染t细胞(t-ui)相比,表达cd8sp-fr1-humov19-vl-v5-[htcrb-kaciah]-f-p2a-sp-fr1-humov19-vh-myc-[htcra-csdvp]-f-f2a-pac(102915-p07)[seqidno:1276]sir的t细胞有效诱导skov3和pc3细胞死亡。表达靶向epcam1和l1cam的sir的t细胞也显示与未感染t细胞相比在skov3和pc3细胞系中的弱细胞死亡诱导。表达针对细胞内抗原tert、mart1、muc1、gp100、酪氨酸酶和nyeso的sir的t细胞诱导靶细胞中的细胞毒性。用表达指定sir构建体(cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-tert-3g3-t865-vl-gly-ser-接头-tert-3g3-t865-vh-myc-[htcra-csdvp]-f-f2a-pac(021216-l07)[seqidno:1784]、cd8sp-tert-3g3-t865-vl-[htcrb-opt2]-f-p2a-sp-tert-3g3-t865-vh-[htcra-opt2]-f-f2a-pac(050316-a01)[seqidno:4902]、cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-mart1-cag10-vl-gly-ser-接头-mart1-cag10-vh-myc-[htcra-csdvp]-f-f2a-pac(021216-n03)[seqidno:1739]、cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-muc1-d6-m3b8-vl-gly-ser-接头-muc1-d6-m3b8-vh-myc-[htcra-csdvp]-f-f2a-pac(021616-b05)[seqidno:1751]、cd8sp-muc1-d6-m3a1-vl-[htcrb-opt2]-f-p2a-sp-muc1-d6-m3a1-vh-[htcra-opt2]-f-f2a-pac(050316-b01)[seqidno:4870]、cd8sp-nyeso-t1-vl-[htcrb-opt2]-f-p2a-sp-nyeso-t2-vh-[htcra-opt2]-f-f2a-pac(040416-d01)[seqidno:4877]、cd8sp-gp100-vl-[htcrb-opt2]-f-p2a-sp-gp100-vh-[htcra-opt2]-f-f2a-pac(031516-b03)[seqidno:4828]和cd8sp-tyros-b2-vl-[htcrb-opt2]-f-p2a-sp-tyros-b2-vh-[htcra-opt2]-f-f2a-pac(032816-b03)[seqidno:4915]的慢病毒转染用cd3磁珠分离的人类外周血t细胞。靶向tert、mart1、muc1、gp100、酪氨酸酶和nyeso的sir识别源自这些细胞内蛋白质的肽连同如先前所述的hla-a2分子。用嘌呤霉素选择细胞并扩增。将稳定表达hgluc的指定靶(hla-a2)细胞与表达sir的t细胞共同培养,其效应物:靶标(e:t)比率为10:1,持续4小时。通过增加gluc活性来测定靶细胞裂解的sir-t细胞介导的诱导。结果显示靶向tert、mart1、muc1、gp100、酪氨酸酶和nyeso的sir显示表达这些细胞内抗原的hla-a2阳性靶细胞系的裂解。另外,表达靶向gd3的sircd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-gd3-km-641-vl-gly-ser-接头-gd3-km-641-vh-myc-[htcra-csdvp]-f-f2a-pac(042816-a04)[seqidno:1697]的t细胞显示表达gd3的mel624靶细胞的裂解。表达针对egfr的sir的t细胞诱导表达egfr的hela细胞中的细胞毒性。用表达靶向egfr的sir构建体的慢病毒感染人类外周血t细胞。用嘌呤霉素选择细胞并扩增。将稳定表达hgluc的hela细胞与表达sir的t细胞或未感染t细胞(t-ui)共同培养,其效应物:靶标(e:t)比率为5:1,持续24小时。通过增加gluc活性来测定靶细胞裂解的sir-t细胞介导的诱导。结果显示与t-ui细胞或单独培养基相比,表达cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-西妥昔单抗-vl-gly-ser-接头-西妥昔单抗-vh-myc4-[pretcra-del48]-f-f2a-pac(062916-g04)[seqidno:1880]sir的t细胞有效诱导hela靶细胞裂解。表达针对cd324的sir的t细胞诱导表达cd324的mda--mb--231细胞中的细胞毒性。用表达靶向cd324的指定sir构建体的慢病毒感染所分离的人类外周血t细胞。用嘌呤霉素选择细胞并扩增。将稳定表达hgluc的mda-mb-231细胞与表达sir的t细胞或未感染t细胞(t-ui)共同培养,其效应物:靶标(e:t)比率为5:1,持续24小时。通过增加gluc活性来测定靶细胞裂解的sir-t细胞介导的诱导。数据显示与t-ui细胞或单独培养基相比,表达cd8sp-cd324-sc10-6-vl-v5-[htcrb-kaciah]-f-p2a-sp-cd324-sc10-6-vh-myc4-[htcra-csdvp]-f-f2a-pac(071516-l04)[seqidno:1239]sir的t细胞有效诱导cd324靶细胞裂解。表达靶向cd276和il13ra2的sir的t细胞诱导表达这些抗原的u87--mg靶细胞中的细胞毒性。用表达靶向cd276和il13ra2的sir构建体的慢病毒感染t细胞。用嘌呤霉素选择细胞并扩增。将u87-mg-gluc细胞与表达sir的t细胞或未感染t细胞(t-ui)以10:1的e:t比率共同培养4小时。通过增加gluc活性来测定靶细胞裂解的sir-t细胞介导的诱导。数据显示与t-ui细胞或单独培养基相比,表达cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-cd276-17-vl-gly-ser-接头-cd276-17-vh-myc-[htcra-csdvp]-f-f2a-pac(042816-c03)[seqidno:1658]sir的t细胞有效诱导靶细胞裂解。表达靶向gd2的sir的t细胞诱导表达这些抗原的skmel--31和skmel--37靶细胞中的细胞毒性。用表达靶向gd2的sir构建体的慢病毒感染t细胞。用嘌呤霉素选择细胞并扩增。将稳定表达gluc的skmel-31和skmel-37细胞与表达sir的t细胞或未感染t细胞(t-ui)以10:1的e:t比率共同培养4小时。通过增加gluc活性来测定靶细胞裂解的sir-t细胞介导的诱导。数据显示与t-ui细胞或单独培养基相比,表达cd8sp-gd2-hu3f8-vl-v5-[htcrb-kaciah]-f-p2a-sp-gd2-hu3f8-vh-myc-[pretcra-del48]-f-f2a-pac(041816-e01)[seqidno:1489]sir的t细胞有效诱导靶细胞裂解。表达针对l1cam的sir的t细胞诱导表达l1cam的skov3细胞中的细胞毒性。将表达靶向l1cam的sir的t细胞与skov3-gluc细胞以10:1的e:t比率培养24小时并且使用gluc-细胞毒性测定来测试。数据显示与t-ui细胞或单独培养基相比,表达cd8sp-myc-[htcra-t48c-opt1]-f-f2a-cd8sp-l1cam-9-3-hu3-v5-[htcrb-t57c-opt1]-f-p2a-pac(080316-t02)[seqidno:1136]sir的t细胞有效诱导skov3靶细胞裂解。表达针对cdh6的sir的t细胞诱导表达cdh6的skov3细胞中的细胞毒性。将表达靶向cdh6的sir的t细胞与skov3-gluc细胞以10:1的e:t比率培养24小时并且使用gluc-细胞毒性测定进行测试。数据显示与t-ui细胞或单独培养基相比,表达cd8sp-cdh6-nov712-vl-v5-[htcrb-kaciah]-f-p2a-sp-cdh6-nov712-vh-myc-[htcra-csdvp]-f-f2a-pac(062816-u01)[seqidno:1242]和cd8sp-cdh6-nov710-vl-v5-[htcrb-kaciah]-f-p2a-sp-cdh6-nov710-vh-myc-[htcra-csdvp]-f-f2a-pac(063016-t05)[seqidno:1241]sir的t细胞有效诱导skov3靶细胞裂解。表达针对trop2的sir的t细胞诱导表达trop2的pc3细胞中的细胞毒性。将表达靶向trop2的sir的t细胞与pc3-gluc细胞以10:1的e:t比率培养24小时并且使用gluc-细胞毒性测定来测试。数据显示与t-ui细胞或单独培养基相比,表达cd8sp-trop2-ara47-hv3kv3-vl-v5-[htcrb-kaciah]-f-p2a-sp-trop2-ara47-hv3kv3-vh-myc-[htcra-csdvp]-f-f2a-pac(062816-s01)[seqidno:1367]sir的t细胞有效诱导pc3靶细胞裂解。表达针对gfra4(gdnf家族受体α4)的sir的t细胞诱导表达gfra4的tt细胞中的细胞毒性。将表达靶向gfra4的sir的t细胞与tt-gluc细胞以10:1的e:t比率培养24小时并且使用gluc-细胞毒性测定来测试。数据显示与t-ui细胞或单独培养基相比,表达cd8sp-gfra4-p4-10-vl-v5-[htcrb-kaciah]-f-p2a-sp-gfra4-p4-10-vh-myc-[htcra-csdvp]-f-f2a-pac(062816-w05)[seqidno:1282]sir的t细胞有效诱导tt靶细胞裂解。表达靶向mpl(促血小板生成素受体)的sir的t细胞诱导表达mpl的hel--92..1..7细胞中的细胞毒性。将表达靶向mpl的sir的t细胞与hel-92.1.7-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定来测试。结果显示与t-ui细胞或表达cd8sp-fmc63-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-fmc63-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(050515-l05)[seqidno:900]sir的t细胞或单独培养基相比,表达cd8sp-mpl-161-vl-v5-[htcrb-s57c-opt1]-f-p2a-mpl-161-vh-myc-[htcra-t48c-opt1]-f-f2a-pac(040315-u02)[seqidno:1112]sir的t细胞有效诱导hel-92.1.7靶细胞裂解。表达靶向mpl(促血小板生成素受体)的sir的t细胞诱导表达mpl的hel--92..1..7细胞中的细胞毒性。将表达靶向mpl的sir的t细胞与hel-92.1.7-gluc细胞以10:1的e:t比率培养4小时并且使用gluc-细胞毒性测定来测试。结果显示与t-ui细胞或表达cd8sp-fmc63(vl-vh)-myc-bbz-t2a-pac(112014-a13)[seqidno:4501]car的t细胞或单独培养基相比,表达cd8sp-mpl-161-vl-v5-[htcrb-s57c-opt1]-f-p2a-mpl-161-vh-myc-[htcra-t48c-opt1]-f-f2a-pac(040315-u02)[seqidno:1112]sir的t细胞有效诱导hel-92.1.7靶细胞裂解。当将表达单链sircd8sp-mpl-161-vl-ser-gly-接头-mpl-161-vh-myc-[htcra-t48c-opt1]-f-t2a-pac(040915-x03)[seqidno:1192]和cd8sp-mpl-161-vl-ser-gly-接头-mpl-161-vh-v5-[htcrb-s57c-opt1]-t2a-pac(032415-e07)[seqidno:1193]与hel-92.1.7靶细胞共同培养时,也观察到靶细胞裂解的诱导。最后,表达单链sircd8sp-mpl-161-vl-ser-gly-接头-mpl-161-vh-myc-[htcra-t48c-opt1]-f-t2a-pac(040915-x03)[seqidno:1192]的t细胞诱导jurkat-mpl细胞的裂解。其它示例性单链sir由dnaseqidno:7519至7715和16694至16809以及prtseqidno:8161至8357和16928至17043表示,在该单链sir中本公开的不同scfv附接至突变体tcrα链。优选地在内源性tcrα链的表达已减少或消除的细胞中,在没有互补外源性tcrβ链的情况下表达这些单链sir。单链sir由dnaseqidno:7733至7929和16811至16926以及prtseqidno:8375至8571和17045至17160表示,在该单链sir中本公开的不同scfv附接至突变体tcrβ链。优选地在内源性tcrβ1和tcrβ2链的表达已减少或消除的细胞中,在没有互补外源性tcrα链的情况下表达这些单链sir。表达针对tslpr(胸腺基质淋巴生成素受体)的sir的t细胞诱导表达tslpr的jurkat细胞中的细胞毒性。将表达靶向tslpr的sir的t细胞与jurkat-gluc细胞以10:1的e:t比率培养24小时并且使用gluc-细胞毒性测定来测试。数据显示与t-ui细胞或单独培养基相比,表达cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-tslpr-vl-gly-ser-接头-tslpr-vh-myc-[htcra-csdvp]-f-f2a-pac(091216-c03)[seqidno:1797]sir的t细胞有效诱导jurkat靶细胞裂解。表达针对ssea4的sir的t细胞诱导表达ssea4的p19和f9胚胎性癌细胞中的细胞毒性。将表达靶向ssea4的sir的t细胞与hel-92.1.7-gluc细胞以10:1的e:t比率培养24小时并且使用gluc-细胞毒性测定来测试。结果显示与t-ui细胞相比,表达cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-ssea4-vl-gly-ser-接头-ssea4-vh-myc-[htcra-csdvp]-f-f2a-pac(091516-i06)[seqidno:1776]和cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-ssea4-vl-gly-ser-接头-ssea4-vh-myc4-[pretcra-del48]-f-f2a-pac(091516-k06)[seqidno:1991]sir的t细胞有效诱导ssea4靶细胞裂解。表达针对cdh17的sir的t细胞诱导表达cdh17的lovo细胞中的细胞毒性。将表达靶向cdh7的sir的t细胞与lovo-gluc细胞以10:1的e:t比率培养24小时并且使用gluc-细胞毒性测定来测试。结果显示与t-ui细胞或单独培养基相比,表达cd8sp-cdh17-pta001a4-vl-v5-[htcrb-kaciah]-f-p2a-sp-cdh17-pta001a4-vh-myc-[htcra-csdvp]-f-f2a-pac(062816-x02)[seqidno:1243]sir的t细胞有效诱导lovo靶细胞裂解。表达针对间皮素的sir的t细胞诱导表达间皮素的skov3卵巢癌细胞中的细胞毒性。将表达靶向间皮素的sir的t细胞与skov3-gluc细胞以10:1的e:t比率培养24小时并且使用gluc-细胞毒性测定来测试。数据显示与t-ui细胞或单独培养基相比,表达cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-间皮素-m912-vl-gly-ser-接头-m912-vh-myc4-[pretcra-del48]-f-f2a-pac(090616-e04)[seqidno:1956]sir的t细胞有效诱导skov3靶细胞裂解。表达针对fshr(卵泡刺激素受体)的sir的t细胞诱导表达fshr的mdamb--231人类乳腺癌细胞中的细胞毒性。用表达靶向fshr的sir构建体的慢病毒感染t细胞。sir的抗原结合结构域包含经由gly-ser接头连接至cgha(绒毛膜促性腺激素α链)的fshb(卵泡刺激素β链)。用嘌呤霉素选择细胞并扩增。将稳定表达hgluc的mdamb-231细胞与表达sir的t细胞或未感染t细胞(t-ui)共同培养,其效应物:靶标(e:t)比率为10:1,持续24小时。通过增加gluc活性来测定靶细胞裂解的sir-t细胞介导的诱导。结果显示与t-ui细胞或单独培养基相比,表达cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-fshb-gly-ser-接头-cgha-myc4-[pretcra-del48]-f-f2a-pac(091516-n06)[seqidno:1909]sir的t细胞有效诱导mdamb-231靶细胞裂解。结果证明其中抗原结合结构域包含异二聚体细胞因子的sir具有功能活性。表达针对lhr(促黄体生成激素受体)的sir的t细胞诱导表达lhr的mcf7人类乳腺癌细胞中的细胞毒性。用表达靶向lhr的sir构建体的慢病毒感染t细胞。此sir的抗原结合结构域包含经由gly-ser接头连接至cgha(绒毛膜促性腺激素α链)的lhb(黄体生成素β链)。用嘌呤霉素选择细胞并扩增。将稳定表达hgluc的mcf7细胞与表达sir的t细胞或未感染t细胞(t-ui)共同培养,其效应物:靶标(e:t)比率为2:1,持续24小时。通过增加gluc活性来测定靶细胞裂解的sir-t细胞介导的诱导。结果显示与t-ui细胞或单独培养基相比,表达cd8sp-v5-[htcrb-kaciah]-f-p2a-sp-lhb-gly-ser-接头-cgha-myc-[htcra-csdvp]-f-f2a-pac(091616-r03)[seqidno:1735]的t细胞适度诱导mdamb-231靶细胞裂解。结果证明其中抗原结合结构域包含异二聚体细胞因子的sir具有功能活性。表达针对dll3(δ样3)的sir的t细胞诱导表达dll3的skmel31和skmel37黑素瘤细胞中的细胞毒性。将表达靶向dll3的sir的t细胞与skmel31-gluc和skmel37-gluc细胞以10:1的e:t比率培养24小时并且使用gluc-细胞毒性测定来测试。数据显示与t-ui细胞或单独培养基相比,表达cd8sp-dll3-hsc16-13-vl-v5-[htcrb-kaciah]-f-p2a-sp-dll3-hsc16-13-vh-myc4-[htcra-csdvp]-f-f2a-pac(071516-n04)[seqidno:1263]sir的t细胞有效诱导skmel31和skmel37靶细胞裂解。表达针对egfrviii突变体的sir的t细胞诱导表达egfrviii的hela细胞中的细胞毒性。用表达靶向egfrviii的sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。用嘌呤霉素选择细胞并扩增。通过用逆转录病毒载体(mscv-egfrviii)和表达hgluc的逆转录病毒载体感染来将hela细胞工程化以表达egfrviii和hgluc。将稳定表达hgluc的hela-egfrviii-hgluc细胞与表达sir的t细胞或未感染t细胞(t-ui)共同培养,其效应物:靶标(e:t)比率为2:1,持续24小时。通过增加gluc活性来测定靶细胞裂解的sir-t细胞介导的诱导。实验显示与t-ui细胞或单独培养基相比,表达cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-egfrviii-2173-vh-gly-ser-接头-egfrviii-2173-vh-myc4-[pretcra-del48]-f-f2a-pac(090616-d03)[seqidno:1902]sir的t细胞有效诱导靶细胞裂解。表达针对egfr的sir的t细胞诱导表达egfr的hela细胞中的细胞毒性。将表达靶向egfr的sir的t细胞与hela-gluc细胞以2:1的e:t比率培养24小时并且使用gluc-细胞毒性测定来测试。数据显示与t-ui细胞或单独培养基相比,表达cd8sp-西妥昔单抗-vl-v5-[htcrb-kaciah]-f-p2a-sp-西妥昔单抗-vh-myc4-[htcra-csdvp]-f-f2a-pac(071516-h04)[seqidno:1245]sir的t细胞诱导egfr靶细胞裂解。表达针对hiv1包膜糖蛋白的sir的t细胞诱导表达hiv1包膜糖蛋白的hl2/3细胞中的细胞毒性。用表达靶向hiv1包膜糖蛋白的指示sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。用嘌呤霉素选择细胞并扩增。将表达hiv1包膜糖蛋白并被工程化以稳定表达hgluc的hl2/3细胞与表达sir的t细胞或未感染t细胞(t-ui)共同培养,其效应物:靶标(e:t)比率为2:1,持续24小时。通过增加gluc活性来测定靶细胞裂解的sir-t细胞介导的诱导。结果显示与t-ui细胞或单独培养基相比,表达cd8sp-hiv1-3bnc117-vl-myc2-[htcrb-kaciah]-f-p2a-sp-hiv1-3bnc117-vh-myc4-[htcra-csdvp]-f-f2a-pac(091616-y01-rp)[seqidno:1297]sir的t细胞诱导ehiv1包膜靶细胞裂解。靶向bst1/cd157的sir发挥针对molm13白血病细胞的细胞毒性。将t细胞工程化以表达具有seqidno:11333至11335的靶向bst1的sir,并且在以10:1的e:t比率共同培养24小时之后,测试针对molm13-gluc细胞的细胞毒性。通过gluc测定测量细胞毒性。靶向bst1的sir-t细胞显示引起显著细胞死亡,如通过gluc活性增加所测量的。靶向il1rap的sir发挥针对molm13白血病细胞的细胞毒性。将t细胞工程化以表达具有seqidno:18242、18248和18254的靶向il1rap的sir并且在以10:1的e:t比率共同培养24小时之后测试针对molm13-gluc细胞的细胞毒性。通过gluc测定测量细胞毒性。靶向il1rap的sir-t细胞显示引起显著细胞死亡,如通过gluc活性增加所测量的。sir--treg细胞阻断了sir--t细胞的细胞毒性活性。使用来自美天旎公司(miltenyi)的t-reg分离试剂盒(130-091-301)并遵循制造商的建议分离t-reg细胞。在补充有雷帕霉素100ng/mlt的t细胞培养基中培养t-reg细胞。用指定的car和sir慢病毒感染总t细胞和t-reg细胞。用嘌呤霉素400ng/ml选择细胞并扩增。将表达靶向cd19的car和sir的t细胞和t-reg细胞与raji-gluc细胞单独或组合地以10:1的e:t比率共同培养4小时并且通过直接注射含有天然腔肠素(nanaolight)的0.5xctz测定缓冲液来测量细胞毒性。结果显示表达靶向cd19的sir的t-reg细胞可以部分阻断表达sir的t细胞的细胞毒性活性。用于合成免疫受体的jurkatnfat--gfp测定..以一种方式将jurkat-nfat-gfp细胞工程化,使得携带nfat结合位点的il-2启动子克隆在gfp基因上游。这些细胞已用于研究经由tcr和car进行的信号传导。通过慢病毒介导的基因转移,随后用嘌呤霉素进行选择来在jurkatnfat-gfp细胞中稳定表达不同sir。将表达sir的jurkat-nfat-gfp细胞与靶细胞以大约1:2的e:t比率共同培养大约4小时至18小时。当sir与其靶抗原之间的相互作用引起nfat途径活化时,诱导gfp表达。因此,表达sir的jurkat-nfat-gfp细胞当与表达sir的受体的靶细胞系相互作用时显示gfp表达水平增加。研究通过将表达不同sir构建体的jurkat-nfat-gfp细胞与不同靶细胞共同培养来诱导gfp表达,基本上如先前所述的(wu,roybal,puchner,onuffer,&lim,2015)。通过facs分析监测gfp表达。此测定的若干代表性实例示出于图13a--b中。在子图a中,将对照jurkat-nfat-gfp细胞或表达靶向cd19(克隆id051716-i08)、mpl(克隆id:040716-a07)和bcma(克隆id:011116-a07)的sir的那些细胞分别与raji(顶部)、hel(中间)或u266(底部)细胞一起孵育。在将表达sir的jurkat-nfat-gfp细胞与其对应靶细胞共同培养时,gfp表达的诱导是明显的。在子图b中,将表达靶向cdh6(克隆id:051716-j05)、cd276(克隆id:050516-q06)和her2/neu(克隆id:050516-i03)的sir的jurkat细胞分别与skov3(顶部)和mc7(中间和底部)细胞一起孵育。在将表达sir的jurkat-nfat-gfp细胞与其对应靶细胞共同培养时,gfp表达的诱导是明显的。将jurkat-nfat-gfp细胞工程化以表达靶向其它抗原的不同sir并且在与表达其对应靶抗原的靶细胞共同培养时重复实验。将jurkat-nfat-gfp(亲代)细胞用作对照。使用不同sir的结果汇总在以下总结表10a中。不同sir的名称、其seqidno、组分抗原结合结构域和tcr链可以通过参考表7a--7h来确定。在表达sir的jurkat-nfat-gfp细胞当与靶细胞系培养时与亲代jurkat-nfat-gfp细胞相比显示更大%gfp阳性细胞的情况下,sir在测定中被认为是阳性的。因此,表达由seqidno:1207表示的sir的jurkat-nfat-gfp细胞当与lan5细胞共同培养时与亲代jurkat-nfat-gfp细胞相比显示更大gfp表达诱导,但是当与karpass299、sudhl-1或h460细胞系共同培养时显示更大gfp诱导。在细胞系名称之后的符号+/-、+、2+等指示在jurkat-nfat-gfp测定中的相对阳性程度,如通过在培养表达sir的jurkat-nfat-gfp细胞与该细胞系之后由%gfp阳性细胞测量。结果证明存在含有源自相同抗体(例如fmc63)的结合结构域的不同sir,使用此测定当暴露于相同细胞系时根据其结构中所用的tcr链和接头而显示其活化nfat信号传导的能力的更大多样性。另外,使用表达含有靶向相同抗原的不同抗原结合结构域的sir(例如具有源自不同cd19抗体的抗原结合结构域的sir)的jurkat细胞观察到针对相同靶细胞系的反应的更大多样性,甚至当sir共享相同tcr链和接头时也如此。最后,表达靶向不同抗原(例如cd19对比cd20)的sir的jurkat细胞当暴露于相同靶细胞系时显示反应多样性。因此,通过将具有不同tcr链、接头、抗原结合结构域和靶标特异性的sir组合来生成针对单一靶细胞的不同免疫反应。表10a还汇总了使用不同sir当暴露于其靶细胞系时观察到的基于gluc的t细胞细胞毒性测定的结果。符号+/-、+和2+等指示当在类似条件下执行测定时,与对照t细胞,即不表达sir或表达不相关sir(靶向特定靶细胞系上不表达的抗原的sir)的t细胞相比,在将靶细胞系与表达sir的t细胞共同培养4-96小时之后使用gluc细胞毒性测定观察到细胞毒性程度。而且,与使用jurkat-nfat-gfp细胞获得的结果类似,表达不同sir的t细胞当暴露于表达其靶抗原的细胞时根据其tcr链、接头、抗原结合结构域、靶特异性和靶细胞系,显示其发挥细胞毒性的能力的更大多样性。在表达sir的t细胞内当它们在相当条件下暴露于靶细胞系时根据其tcr链/接头/抗原结合结构域/靶特异性和靶细胞系观察到诱导细胞因子产生(例如il2、tnfα和ifnγ)的能力的类似多样性。然而,当在类似条件下执行测定时,sir-t细胞当暴露于其靶细胞时进行的细胞因子产生通常低于使用相当car-t细胞观察到的细胞因子产生。表达含有相同抗原结合结构域但含有不同tcr链和接头的不同sir的t细胞当在类似条件下测定时也观察到增殖和耗尽标记的表达的差异。表10a还汇总了使用含有tcrα和tcrβ链的细胞外结构域与具有和不具有源自41bb或cd28的任选共刺激结构域的cd3z链的细胞外、跨膜和细胞溶质结构域的融合的sir获得的结果。基于fmc63结合结构域的这种类似的示例性sir是cd8sp-fmc63-vl-tcrb-kac-ecd-bam-cd3zecdtmcp-opt-f-p2a-sp-fmc63-vh-htcra-csdvp-ecdn-cd3zecdtmcp-opt2-f-f2a-pac(seqidno:10554)。在此构建体中,fmc63的vl片段附接至tcrb-kac-ecd-bam-cd3zecdtmcp-opt链(seqidno:12402),而fmc63的vh片段附接至htcra-csdvp-ecdn-cd3zecdtmcp-opt2链(seqidno:12422)。此sir在jurkat-nfat-gfp测定中为强阳性的,但是显示低抗原结合活性,如通过与可溶性cd19-nluc融合蛋白的结合所测量的。具有类似设计但具有不同tcra和tcrb链变体的若干其它构建体由seqidno:10552至10557表示并且在jurkat-nfat-gfp测定和t细胞细胞毒性测定中显示强但不同的水平的活性。在sir构建体cd8sp-fmc63-vl-htcraecdn-cd3zecdtmcp-opt2-f-f2a-sp-fmc63-vh-tcrbecd-bam-cd3zecdtmcp-opt-f-p2a-pac(seqidno:10564)中,fmc63vl片段附接至tcraecdn-cd3zecdtmcp-opt2链(seqidno:12421),而fmc63vh片段附接至tcrbecd-bam-cd3zecdtmcp-opt(seqidno:12401)。此构建体在jurkat-nfat-gfp测定中也显示强阳性。含有来自插入在两条链各自的cd3z细胞质结构域中的41bb的共刺激结构域的构建体cd8sp-fmc63-vl-tcrbecd-bam-cd3zecdtmcp-bbz-opt-f-p2a-sp-fmc63-vh-myc4-htcraecdn-cd3zecdtmcp-bbz-opt2-f-f2a-pac(seqidno:10565)在jurkat-nfat-gfp测定中也为有活性的。构建体(seqidno:10563)的设计类似于seqidno:10565,但其仅一条链含有41bb共刺激结构域。此构建体在jurkat-nfat-gfp测定中也为有活性的。最后,在一条链上含有cd28共刺激结构域的构建体cd8sp-fmc63-vl-v5-tcrbecd-bam-cd3zecdtmcp-opt-f-p2a-sp-fmc63-vh-myc-htcraecdn-cd3zecdtm-28z-opt2-f-f2a-pac(seqidno:10557)在jurkat-nfat-gfp测定中也是有活性的。表10a.使用靶向不同抗原的不同sir文库进行nfat--gfp和t细胞细胞毒性测定的汇总为了证明生成含有相同抗原结合结构域的不同sir库,比较稳定表达含有基于fmc63的抗原结合结构域但含有不同tcr链和接头的sir的jurkat-nfat-gfp细胞的以下能力:1)结合cd19-ggsg-nluc-acv5融合蛋白(即抗原结合测定);2)在与raji细胞共同培养18小时后诱导gfp表达(即细胞信号传导测定);3)在与raji细胞共同培养18小时后诱导il2产生(即细胞因子产生测定);4)用apc缀合蛋白质-l染色(即细胞表面表达的测量)。结果汇总于表10b中并且在以上测定中证实具有相同抗原结合结构域的sir中的巨大多样性。含有由野生型核苷酸序列编码的tcrα和tcrβ链的构建体seqidno:992显示最低水平的cd19-ggsg-nluc结合、gfp诱导和蛋白质l染色。包括稳定表达cd8sp-fmc63(vl-vh)-myc-bbzt2a-pac(seqidno:4501)car的jurkat-nfat-gfp细胞以用于在il2产生和蛋白质l染色的测定中进行比较。引人注意的是,car与所有sir相比显示显著更高的il2产生(1159pg/ml)和蛋白质-l染色(61%)。表10b:靶向cd19并含有fmc63结合结构域的sir中的多样性在靶向相同抗原但含有不同抗原结合结构域的sir中的多样性。为了生成具有甚至更大多样性的特定靶标的sir,使用源自不同抗体的抗原结合结构域生成靶向cd19的若干种sir。这些sir的不同之处也在于其tcr链、接头和抗原结合结构域格式。相应car(即还生成含有与存在于sir中的相同的抗原结合结构域的car)。将不同sir和car稳定转导到jurkat-nfat-gfp细胞中并且比较所得细胞结合至cd19-ggsg-nluc-acv5并在与raji细胞共同培养过夜后活化nfat信号传导的能力。表9c证实含有不同cd19抗原结合结构域的不同sir和car中对于结合至cd19并在暴露于靶细胞后活化nfat信号传导的能力的显著多样性。含有tcrα和tcrβ链的野生型核苷酸序列的基于bu12抗体的sircd8sp-cd19bu12-vl-v5-[htcrb-wt]-f-p2a-cd19bu12-vh-myc-[htcra-wt]-f-f2a-pac(seqidno:1038)显示极少cd19结合和最低nfat信号传导。在不同sir中其结合cd19并活化nfat信号传导的能力也存在显著差异并且当比较含有不同抗原结合结构域的sir时这两种参数并不直接相关。因此,由seqidno:11240表示的sir显示与由seqidno:10815和11245表示的sir相比高将近10倍的cd19-nluc-acv5结合,但具有更低nfat-信号传导。结果证实了生成表达具有不同特性的sir库的免疫效应细胞以用于生成不同过继性免疫反应的能力。表达sir的免疫效应细胞可以与表达car和其它嵌合免疫受体的免疫细胞组合以用于生成甚至更不同的免疫反应。表10c:在靶向cd19并含有不同抗原结合结构域的sir和car中的多样性sir显示与car相比更低的抗原结合但显示相当的nfat信号传导。将表达sircd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(010616-c01)[seqidno:1200]和carcd8sp-fmc63(vl-vh)-myc-bbz-t2a-pac(112014-a13)[seqidno:4501]的jurkat-nfat-gfp细胞连同亲代jurkat-nfat-gfp细胞一起测试在暴露于raji细胞过夜之后nfat诱导型gfp表达的诱导。表达sir(seqidno:1200)和car(seqidno:4501)的细胞显示与亲代细胞相比诱导相当和显著更高的gfp诱导,证明sir和car有效诱导nfat信号传导。随后重复三次使用cd19-ggsg-nluc结合测定测试细胞的抗原结合。与亲代jurkat细胞的平均nluc结合活性是93,而表达sir(seqidno:1200)和car(seqidno:4501)的jurkat细胞分别显示16422和186567的平均nluc值。因此,表达sirseqidno:1200并当两者均暴露于靶抗原时具有与相应car(seqidnno:4501)几乎等效nfat信号传导活性的jurkat细胞显示低将近10倍的抗原结合亲和力。基于bu12的sir与相当的bu12car相比显示显著更低的细胞表面表达。已知car在t细胞表面上的表达增加导致自身聚集、补剂信号传导和早期细胞耗尽。用编码靶向cd19的基于bu12的sir(cd8sp-cd19bu12-(vl-vh)-myc-bbz-t2a-pac(seqidno:4503)和相当的car(cd8sp-cd19bu12-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-cd19bu12-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(070215-m03)[seqidno:1021]的慢病毒感染人类外周t细胞。用嘌呤霉素选择细胞并扩增。表达sir和car的t细胞显示针对raji-gluc细胞的有效细胞毒性。还使用利用针对myc标签的抗体和生物素化蛋白质l随后用apc缀合的链霉亲和素蛋白质l进行的染色检查t细胞的sir和car的细胞表面表达。图16显示表达sir(seqidno:1021)的t细胞当通过任一种方法测量时显示与表达对应car(seqidno:4503)的t细胞相比显著更低的细胞表面表达。sir--t细胞显示与对应car--t细胞相比更低的tnfα产生。t细胞被工程化以表达carcd8sp-fmc63-vl-gly-ser-接头-fmc63-vh-myc-cd8tm-bbz(seqidno:9659)和对应sir(cd8sp-fmc63-vl-[htcra-csdvp]-f-f2a-sp-fmc63-vh-[htcrb-kaciah]-f-p2a-pac(seqidno:10596)。将这些细胞与nalm6和bv173靶细胞系在37℃下共同培养24小时并且通过elisa测量tnfα产生的诱导。表达car的t细胞显示与sir-t细胞相比引起更大tnfα产生的成倍增加。sir-t和car-t细胞两者显示诱导针对靶细胞系的显著细胞毒性。两种载体表达sir的用途。在前述实验中,使用单一载体表达sir的两个功能多肽单元(fpu)。其次,如果可以使用两种不同载体表达sir的两个fpu,则测试该载体。sir慢病毒构建体050216-t02和050216-s08含有对应于seqidno:913且编码sir片段cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-pac的sir序列,其中源自cd19单克隆抗体fmc63的vl片段经由v5接头连接至具有kaciah突变的htcra的恒定链。此sirfpu经由f-p2a可裂解接头连接至pac(嘌呤霉素抗性)基因。050216-t02构建体的载体是plenti-ef1α-dwpre(seqidno:871),而050216-s08的载体是plenti-ef1α(seqidno:870)。sir慢病毒构建体041916-a02和041916-b03含有对应于seqidno:997且编码sir片段cd8sp-fmc63-vh-myc-[tcrα-csdvp]-f-f2a-blastr的sir序列,其中源自cd19单克隆抗体fmc63的vh片段经由myc接头连接至具有csdvp突变的htcra的恒定链。此sirfpu经由f-f2a可裂解接头连接至杀稻瘟菌素抗性基因。041916-a02构建体的载体是plenti-ef1α-dwpre(seqidno:871),而041916-b03的载体是plenti-ef1α(seqidno:870)。将jurkat-nfat-gfp细胞用050216-s08感染并用嘌呤霉素选择。还将jurkat-nfat-gfp细胞用041916-a02和041916-b03构建体感染并用杀稻瘟菌素选择。最后,将jurkat-nfat-gfp细胞用050216-s08感染并且在用嘌呤霉素选择时,用041916-a02或041916-b03构建体感染并且用杀稻瘟菌素选择以选择表达sir的两种fpu的细胞。使用nluc-结合测定测试单次感染或双重感染细胞与cd19-ggs-nluc的结合。结果显示用单独构建体050216-s08[seqidno:913]、041916-a02[seqidno:997]或041916-b03[seqidno:997]构建体感染的jurkat-nfat-gfp细胞不显示与cd19-ggs-nluc的任何显著结合。相比之下,用构建体050216-s08加上041916-a02[seqidno:997]以及050216-s08[seqidno:913]加上041916-b03[seqidno:997]感染的jurkat-nfat-gfp细胞显示与cd19-ggs-nluc的强结合。用构建体050216-s08[seqidno:913]加上041916-a02[seqidno:997]以及050216-s08[seqidno:913]加上041916-b03[seqidno:997]感染的jurkat-nfat-gfp细胞当与raji靶细胞共同培养时也显示gfp表达的强诱导,而用这些构建体逐一感染的jurkat-nfat-gfp细胞未能如此。这些结果证明sir的两个fpu可以使用两种不同载体表达并且可以聚集以在双重感染的细胞中形成功能性受体。表达本公开的不同vl片段的示例性sir构建体由dnaseqidno:8803至8978和17162至17277以及prtseqidno:9231至-9406和17396至17511表示,这些vl片段经由v5接头连接至tcrb链(htcrb-s57c-opt)。表达本公开的不同vh片段的示例性对应sir构建体由dnaseqidno:9017至9191和17279-17394以及prtseqidno:9445至9619和17513至17628表示,这些vh片段经由myc接头连接至tcra链(htcra-t48c-opt)。逆转录病毒载体用于表达sir的用途..通过用nhei和sali酶消化将许多sir插入物从慢病毒载体切割出来并且将其亚克隆到avrii和sali消化的逆转录病毒载体mscv-bgl2-avrii-bam-ecor1-xho-bstb1-mlu-sal-clai.i03(seqidno:872)中。在下表11中示出插入物的克隆id、seqid和名称。表11:使用以上构建体生成对应逆转录病毒,进而使用这些逆转录病毒感染jurkat-nfat-gfp细胞。使用嘌呤霉素选择这些感染的细胞并且在不同测定中用于测试sir表达和活性。用以上构建体感染的jurkat-nfat-gfp细胞显示在与对应靶细胞系共同培养之后的gfp诱导。这些结果证明逆转录病毒载体可以用于表达本公开的sir。使用示例性构建体(seqidno:1212)的结果示出在图14中。睡美人转座子载体用于表达sir的用途.通过用agei和xbai酶消化将许多sir插入物从慢病毒载体切割出来并且将其亚克隆到agei和xbai消化的睡美人转座子载体psbbi-pur(seqidno:874)中。将所得构建体连同编码转座酶的载体pcmv/sb10(爱迪基因公司质粒编号24551)一起转染到jurkat-nfat-gfp细胞中。用如上嘌呤霉素选择细胞并扩增。如以下图15中所示,用构建体psbbi-puro-fmc63vl-v5-[tcrb-kaciah]-f-p2a-fmc63vh-myc-[tcra-csdvp]-f-f2a[010616-b01](seqidno:875)转染的jurkat-nfat-gfp细胞在与对应raji靶细胞系共同培养时显示gfp诱导。这些结果证明睡美人转座可以用于表达本公开的sir。体外转录的(ivt)rna用于表达sir的用途..执行生成编码sir的rna的ivt,基本上如(zhaoy等人,moleculartherapy[分子疗法]第13卷,第1期,2006)所述。利用mmessagemmachine高产率加盖的rna转录试剂盒(英杰公司)生成ivtrna。使用rneasy微型试剂盒(美国加利福尼亚州巴伦西亚的凯杰公司(qiagen,inc.,valencia,ca,usa))纯化ivtrna并且在无rna酶水中以1-0.5μg/ml洗脱纯化的rna。对于未刺激的pbmc的电穿孔,用5μg编码靶向cd19的sircd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(102615-c08)[seqidno:1200]的rna电穿孔细胞(0.1ml)。通过将细胞和比色皿置于冰上5分钟来预先冷却它们,之后电穿孔。随后,将0.05至0.2ml细胞与2μg/1106个ivtrnat细胞(如所指示)混合并且在2-mm比色皿(美国马萨诸塞州霍利斯顿的哈佛仪器btx公司(harvardapparatusbtx,holliston,ma,usa))中使用ecm830方波电穿孔器(哈佛仪器btx公司)电穿孔。紧接着在电穿孔之后,将细胞转移至具有300iu/mlil-2的新鲜cm中并且在37℃下孵育。在48-72h后测定以编码sir的ivtrna转录的细胞。sir转染的细胞显示与cd19-ggs-nluc融合蛋白的结合增加和raji-gluc靶细胞的裂解增加。靶向cd19的sir的体内功效..用表达cd8sp-fmc63-vl-v5-[htcrb-wt]-f-p2a-sp-fmc63-vh-myc-[htcra-wt]-f-f2a-pac(080815-f02)[seqidno:922]、cd8sp-fmc63-vl-v5-[mtcrb-opt]-f-p2a-sp-fmc63-vh-myc-[mtcra-opt]-f-f2a-pac(080815-b06)[seqidno:953]、cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(081415-d06)[seqidno:992]sir构建体和编码非分泌形式guassia荧光素酶的gluc-pac-g07对照构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。用嘌呤霉素选择大约一半细胞,而在无嘌呤霉素选择的情况下扩增另一半。以175cgy剂量亚致死地辐照nsg小鼠(杰克逊实验室(jacksonlab))。在辐照后大约24小时(第2天),经由尾静脉向小鼠注射2.5x104个raji细胞。在第3天,用5百万t细胞(50%嘌呤霉素选择的+50%未选择的)处理小鼠(每组n=5),这些细胞已用编码指定sir的慢病毒或gluc-pac-g07构建体感染。对照小鼠(n=5)未接受t细胞或接受未感染t细胞。隔日向小鼠给予人类il2(腹膜内400iu),直到对照组的所有小鼠均死亡。表12示出每一组中的小鼠存活。其中tcrb和tcra恒定链由其野生型核苷酸序列编码的基于fmc63的sir(080815-f02)未能赋予存活性,而含有密码子优化的小鼠tcrb和tcra恒定链序列的sir构建体(080815-b06)[seqidno:953]赋予了存活优点。类似地,含有密码子优化的鼠类化人类tcrb和tcra恒定链的sir构建体(081415-d06)[seqidno:992]赋予存活优点。表12:用表达指定sir构建体(040315-u02)[seqidno:1112]、(050515-l05)[seqidno:900]、(082815-g07)[seqidno:1620]、(082815-e05)[seqidno:1622]和(091015-y08)[seqidno:926]的慢病毒感染用cd3磁珠分离的人类外周血t细胞。用嘌呤霉素选择大约一半细胞,而在无嘌呤霉素选择的情况下扩增另一半。以175cgy剂量亚致死地辐照nsg小鼠(杰克逊实验室(jacksonlab))。在辐照后大约24小时(第2天),经由尾静脉向小鼠注射2.5x104个raji细胞。在第3天,用5百万t细胞(50%嘌呤霉素选择的+50%未选择的)处理小鼠(每组n=5),这些细胞已用编码指定sir的慢病毒感染。对照小鼠(n=5)未接受t细胞或接受未感染t细胞。隔日向小鼠给予人类il2(腹膜内400iu),直到对照组的所有小鼠均死亡。表13示出每一组中的小鼠存活。双链基于fmc63的sir(050515-l05)[seqidno:900]赋予存活优点,在该sir中tcrb和tcra恒定链由密码子优化的核苷酸序列编码并且携带氨基酸取代以促进链配对。重要的是,含有融合至野生型tcrb恒定链的fmc63-vl和融合至pretcra-del48链的fmc63-vh的双链sir构建体(091015-y08)[seqidno:926]赋予甚至更大的存活优点(中值存活=31天)。(082815-g07)[seqidno:1620]构建体赋予甚至更大的存活优点(中值存活=34天),在该构建体中tcrb-kaciah恒定链不具有与其融合的任何vl(或vh)片段并且fmc63来源的scfv片段(vl-接头-vh)被表达为融合至tcra-csdvp。最后,(082815-e05)[seqidno:1622]构建体赋予最大存活优点(中值存活=36天),在该构建体中tcrb-kaciah恒定链不具有与其融合的任何vl(或vh)片段并且cd19-bu12来源的scfv片段(vl-接头-vh)被表达为融合至tcra-csdvp。表13:用表达cd8sp-fmc63-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-fmc63-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(050515-l05)[seqidno:900]sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞以175cgy剂量亚致死地辐照nsg小鼠(杰克逊实验室(jacksonlab))。在辐照后大约24小时(第2天),经由尾静脉向小鼠注射2.5x104个raji细胞。在第3天,用5百万t细胞处理小鼠(每组n=5),这些t细胞已用编码指定cd8sp-fmc63-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-fmc63-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(050515-l05)[seqidno:900]sir的慢病毒感染。向对照小鼠(n=5)注射5百万未感染t细胞或不给予t细胞。表示出每一组中的小鼠存活。给予单独的raji细胞和未感染t细胞的小鼠的中值存活是22天。相比之下,接受(050515-l05)[seqidno:900]-sirt细胞的小鼠的中值存活是27天,该存活显著增加。因此,输注表达(050515-l05)[seqidno:900]-sir的t细胞导致在此raji异种移植淋巴瘤模型中与未给予t细胞的小鼠或给予未感染t细胞的小鼠相比小鼠存活显著改善。用表达cd8sp-fmc63-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-fmc63-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(050515-l05)[seqidno:900]和cd8sp-cd19bu12-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-cd19bu12-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(070215-m03)[seqidno:1021]sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞包括表达对照sircd8sp-mpl-161-vl-v5-[tcrb-s57c-opt1]-f-p2a-mpl-161-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(040315-u02)[seqidno:1112]sir的t细胞作为对照。以175cgy剂量亚致死地辐照nsg小鼠(n=5)。在辐照后24小时(第2天),经由尾静脉向小鼠注射2.5x104个raji细胞。在第3天,用5百万t细胞处理小鼠(每组n=5),这些细胞已用编码指定sir的慢病毒感染。隔日向小鼠给予人类il2(400iui.p.),直到对照组的所有小鼠均死亡。图17示出每一组中的小鼠存活。给予raji细胞和表达对照sir的t细胞(seqidno:1112)的小鼠的中值存活是12天。相比之下,接受(050515-l05)[seqidno:900]-sir-t和(070215-m03)[seqidno:1021]sir-t细胞的小鼠的中值存活是28.0天和51天,该存活显著增加(p=.0004)。用表达靶向叶酸受体1(fr1)、l1cam和epcam1的指示sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。在第1天,以175cgy剂量亚致死地辐照nsg小鼠。在辐照后24小时(第2天),向小鼠腹膜内注射1x106个skov-3细胞。在第3天,用5百万指定的car-t细胞腹膜内处理小鼠。从第4天开始,在注射car-t细胞之后每隔一日向小鼠i.p.注射400iu/小鼠的人类il-2,直至对照组中的所有小鼠死亡。用表达靶向cs1的指示sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。在第1天,以175cgy剂量亚致死地辐照nsg小鼠。在辐照后24小时(第2天),向小鼠静脉内注射小鼠0.5x106个l363细胞。在第3天,用10百万指定的car-t细胞腹膜内处理小鼠。从第4天开始,在注射car-t细胞之后每隔一日向小鼠i.p.注射400iu/小鼠的人类il-2,直至对照组中的所有小鼠死亡。用表达靶向lym1和lym2的指示sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。在第1天,以175cgy剂量亚致死地辐照nsg小鼠。在辐照后24小时(第2天),向小鼠静脉内注射小鼠25x103个raji细胞。在第3天,用10百万指定的car-t细胞腹膜内处理小鼠。从第4天开始,在注射car-t细胞之后每隔一日向小鼠i.p.注射400iu/小鼠的人类il-2,直至对照组中的所有小鼠死亡。如先前实例中所述的基本上类似的实验计划将用于使用nsg小鼠和表达sir的靶标的适当细胞系测试本发明所述的其它sir构建体的体内功效。蛋白质l微珠和表达蛋白质l的细胞用于刺激表达sir和car的细胞的用途..已描述了用于癌症免疫疗法的许多基于人类细胞的人工抗原呈递细胞。一种用于离体扩增car+和sir+t细胞的方法涉及使用表达靶向肿瘤相关抗原(taa)的辐照的人工抗原呈递细胞(aapc)的用途。然而,此方法需要aapc被工程化以表达每种个别taa。为了克服此问题,开发一种通用抗原呈递细胞。蛋白质l是由厌氧细菌种类大消化链球菌表达的免疫球蛋白(ig)氢链结合蛋白。蛋白质l结合至κ氢链vl结构域的框架区并且不干扰抗原结合结构域。作为在aapc上表达个别taa的替代方案,执行实验以确定蛋白质l是否可以不依赖于特异性而活化car+t和sir+t细胞。在初始实验中,测试表达不同car和sir构建体的jurkat-nfat-gfp细胞的共同培养以确定蛋白质l磁珠是否将引起gfp表达的诱导。皮尔斯(pierce)蛋白质l磁珠(目录号88849;浓度10mg/ml)购自赛默飞世尔公司(thermofisher)并且以pbs1:10稀释。将大约10μl稀释的珠粒添加到u形底部96孔板的每个孔中。将jurkat-nfat-gfp亲代细胞或表达cd8sp-fmc63(vl-vh)-myc-bbz-t2a-pac(112014-a13)[seqidno:4501]car的jurkat-nfat-gfp细胞添加到每个孔中并且将其与珠粒共同培养18小时。通过流式细胞术检查gfp表达的诱导。下图18示出通过将表达cd8sp-fmc63(vl-vh)-myc-bbz-t2a-pac(112014-a13)[seqidno:4501]car的jurkat-nfat-gfp细胞与蛋白质l珠粒共同培养来对gfp表达进行的适度诱导。在细胞表面上表达蛋白质l的人类aapc的生成..生成慢病毒嵌合抗原受体(car),该受体表达蛋白质l的不同区域与人类cd8、41bb共刺激结构域和cd3z链的铰链和跨膜区域的融合物。对蛋白质l的核苷酸序列进行密码子优化以用于在人类细胞中最佳表达。需要注意,出于本发明的目的,41bb共刺激结构域和cd3z链不是蛋白质l的功能所必需的。载体还携带源于人类cd8或igh的n-末端信号肽,以允许细胞表面转运蛋白质l。载体的完全核酸和氨基酸序列提供于seqidno:888和seqidno:889。将不同蛋白质l构建体瞬时转染到293ft细胞中。通过将细胞与scfv-ggsg-nluc融合蛋白上清液孵育来检查细胞表面表达的蛋白质l结合至不同scfv片段的能力。表达不同蛋白质l区域的293ft细胞显示与scfv-ggsg-nluc上清液的结合增加,因此证明蛋白质l可以成功地在哺乳动物细胞中表达为细胞表面蛋白质。接着,用编码蛋白质l-ii构建体(072716-k01)的慢病毒载体感染293细胞并且在用750ng/ml嘌呤霉素选择之后生成稳定表达蛋白质l的多克隆细胞群体。将这些293-蛋白质l-ii细胞用作抗原呈递细胞并且测试其活化表达不同sir和car构建体的jurkat-nfat-gfp细胞的能力。出于此目的,将293-蛋白质l-ii细胞接种在24孔板中并且在12-24h后用表达不同sir和car构建体的jurkat-nfat-gfp细胞覆盖,在共同培养大约18小时后,通过流式细胞术检查gfp表达的诱导。如图19a--d所示出,与293-蛋白l-ii细胞共同培养导致在表达以下各项的jurkat-nfat-gfp细胞中gfp表达的强诱导:cd8sp-fmc63(vl-vh)-myc-bbz-t2a-pac(112014-a13)[seqidno:4501]car和cd8sp-huluc64-vl-v5-[htcrb-kaciah]-f-p2a-sp-huluc64-vh-myc-[htcra-csdvp]-f-f2a-pac(092916-e07)[seqidno:1253]、cd8sp-v5-[htcrb-kaciah]-f-p2a-cd8sp-cd19bu12-vl-gly-ser-接头-cd19bu12-vh-myc-[htcra-csdvp]-f-f2a-pac(082815-e05)[seqidno:1622]sir构建体。这些结果证明在其表面上表达蛋白质l的哺乳动物细胞可以用作通用抗原呈递细胞以用于刺激表达sir和car的免疫效应细胞。然而,表达蛋白质l的aapc的使用不限于以上类型的细胞。这些细胞可以用作任何免疫效应细胞的通用aapc,其携带源自含有适合κ链的抗体的可结合至蛋白质l的抗原结合结构域。sir在pgp阳性淋巴细胞中的表达..使用泛影葡胺分离获得外周血单核细胞(1千万个细胞)。将细胞离心并且将细胞团用200μl人类ab血清在4℃下封闭1h。将细胞用含有1%fcs的冰冷pbs洗涤,并且用针对p-糖蛋白(pgp)的三种单克隆抗体uic2(200μg/ml;圣克鲁斯生物技术公司(santacruzbiotechnology);sc-73354)、mrk16和4e3在4℃下染色1h。使用浓度为0.5μg/百万个细胞)的每种抗体增加测定的敏感度。在用含有1%fcs的pbs充分洗涤之后,用5μl(2.5μg)fitc缀合的山羊f(ab)2抗小鼠igg(h+l)人类吸收的抗体(南方生物技术公司(southernbiotechnology);目录号1032-02)染色细胞。在2次洗涤之后,将细胞用pe缀合人类cd3抗体在4℃下标记1h。将细胞洗涤并且通过流式细胞术分析。将cd3阳性t淋巴细胞分选到pgp+和pgp-级份中。用编码cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(081415-d06)[seqidno:992]sir或阴性对照sircd8sp-kshv-4c3-vl-v5-[htcrb-s57c-opt]-f-p2a-sp-4c3-vh-myc-[htcra-t48c-opt]-f-f2a-pac(111815-o05)[seqidno:4639]的慢病毒载体感染每种级份中的细胞。以175cgy剂量亚致死地辐照nsg小鼠(杰克逊实验室)。在辐照后大约24小时(第2天),经由尾静脉向小鼠注射2.5x104个raji细胞。在第3天,用5百万个淋巴细胞处理小鼠(每组n=6),这些细胞已用编码指定sir的慢病毒感染。对照小鼠(n=6)未接受t细胞或接受未感染t细胞。隔日向小鼠给药人类il2。给与用cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(081415-d06)[seqidno:992]sir感染的pgp+t细胞的小鼠与给与用cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(081415-d06)[seqidno:992]sir感染的pgp-t细胞或用阴性对照sircd8sp-kshv-4c3-vl-v5-[htcrb-s57c-opt]-f-p2a-sp-4c3-vh-myc-[htcra-t48c-opt]-f-f2a-pac(111815-o05)[seqidno:4639]感染的pgp+或pgp-t细胞的小鼠相比存活更长时间。给与用cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(081415-d06)[seqidno:992]sir感染的pgp+t细胞的小鼠的更长存活与sir-t细胞的更长体内持久性相关。通过如国际申请号pct/us2017/042248(该申请的公开内容通过引用结合在此)所述的其它方法(包括macs(磁性活化细胞分选))并且通过暴露于th9402随后暴露于光来富集表达pgp的t淋巴细胞来重复以上实验。再次,给与用cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[htcra-csdvp]-f-f2a-pac(081415-d06)[seqidno:992]sir感染的pgp+t细胞的小鼠存活更长时间,这与sir-t细胞的更长体内持久性相关。达沙替尼(dasatiinib)阻断sir--t细胞的活性的用途..将表达sircd8sp-cd19bu12-vl-v5-[tcrb-s57c-opt1]-f-p2a-sp-cd19bu12-vh-myc-[tcra-t48c-opt1]-f-f2a-pac(070215-m03)[seqidno:1021]和cd8sp-cd20-2f2-vl-v5-[htcrb-kaciah]-f-p2a-sp-cd20-2f2-vh-myc-[htcra-csdvp]-f-f2a-pac(100615-d05)[seqidno:1221]的t细胞与100nm达沙替尼在37℃下预先孵育大约30min或者使其未处理。将药物处理的和未处理的t细胞接种在白色384孔板中。将稳定表达gluc的raji细胞添加到含有浓度为30k个细胞/孔的t细胞的孔中以给出5:1的e:t比率。还将达沙替尼添加到孔中以维持100nm的最终浓度。在共同培养4-24h之后,通过直接注射含有天然腔肠素的0.5xctz测定缓冲液来增加gluc活性,从而测定sir-t细胞介导的靶细胞裂解的诱导。结果显示100nm达沙替尼显著抑制sir诱导的细胞死亡。发现在将sir-t细胞与靶细胞系共同培养之后,使用达沙替尼的治疗显著抑制ifnγ和tnfα产生。结果证明达沙替尼阻断细胞毒性和由sirt细胞进行的细胞因子产生的能力。由于达沙替尼是fda批准的药物并且可以穿过血脑屏障,所以可以使用它来在向患者给药之后控制sir-t细胞诱导的细胞毒性,包括神经毒性,并且控制sir-t细胞的活性。pi3k抑制剂扩增sir--t细胞的用途..用表达靶向cd19的指示sir构建体的慢病毒感染用cd3磁珠分离的人类外周血t细胞。使细胞未选择或用嘌呤霉素选择细胞并使用如先前所述的标准方案使用cd3/cd28珠粒和il2但是在双重pi3k/mtor抑制剂pf-04691502(0.10μm至0.5μm)存在或不存在下扩增这些细胞。以175cgy剂量亚致死地辐照nsg小鼠(杰克逊实验室)。在辐照后24小时(第2天),经由尾静脉向小鼠注射2.5x104个raji细胞。在第3天,用5百万t细胞处理小鼠(每组n=5),这些细胞已用编码指定sir的慢病毒感染并且在pf-04691502存在或不存在下扩增。向对照小鼠(n=5)注射5百万未感染t细胞。隔日向小鼠给予人类il2(400iui.p.),直到对照组的所有小鼠均死亡。接受在pi3k/akt抑制剂存在下扩增的(050515-l05)[seqidno:900]-sir-t的小鼠的中值存活高于接受在pi3k/akt抑制剂不存在下扩增的sir-t细胞的小鼠。表达cd16的通用sir--t细胞作为抗原结合结构域的用途..在nsg小鼠(杰克逊实验室)中腹膜内注射(i.p.;0.3x106个细胞/小鼠)表达荧光素酶的daudi细胞。一些小鼠i.p.接受利妥昔单抗(150mg)。在daudi接种之后四天注射表达cd8sp-myc-[htcra-t48c-opt1]-f-f2a-sp-cd16a-v158-ecd-v2-v5-[htcrb-s57c-opt1]-f-p2a-pac(seqidno:2069)sir的人类t细胞(1x107)。对照小鼠接受组织培养基,而非利妥昔单抗或t细胞。每周重复利妥昔单抗注射,持续4周,无进一步t淋巴细胞注射。所有小鼠每周两次接受1000–2000iuil-2的腹膜内注射,持续4周。接受利妥昔单抗加上sir-t细胞的小鼠显示肿瘤生长减小。体外和体内选择用于选择具有所需特性的sir的用途..使用tracgrna(seqidno:896)和本领域已知的技术将表7a--7h中所示的靶向cd19的一系列sir靶向t细胞中的trac基因座。靶向载体还携带位于sir插入物的终止密码子下游的dna条形码。t细胞可以源自外周血。在一个替代性实施方式中,t细胞使用本领域已知的技术衍生自ipsc或造血干细胞的单一克隆。将表达该系列sir的t细胞与raji细胞在体外共同培养1至21天。收集sir-t细胞库的等分试样,之后将其与靶细胞培养并且持续共同培养之后的不同天数。使样品经历下一代测序以确定在暴露于靶细胞之后不同sir的相对频率。使用生物信息学分析确定在与靶细胞共同培养之后与更好增殖反应相关的sir。使用基本上类似的方法确定与屈服于肿瘤挑战的动物相比在体内对t细胞赋予更高增殖潜能和/或在体内长期存留和/或当在存活动物的起始t细胞群体中针对其频率进行标准化时以更高频率存在。在本公开的替代性实施方式中,对于人类临床样品使用基本上类似的方法以鉴别与不同特性和/或结果相关的sir,这些特性或结果包括但不限于更长期的存活、更低细胞因子释放综合征发生率、更低神经毒素和/或更高长期持久性。随后可以单一或以不同组合使用此类sir,以开发不同sir子库,其含有靶向相同或不同抗原结合结构域的sir,具有不同特性以用于治疗不同疾病病状和不同患者。在其它实施方式中,将sir-t细胞暴露于其靶细胞系并且然后基于细胞内ifnγ程度(如通过流式细胞术测定)分选到不同组中。在低对比高ifnγ群体中不同sir的频率通过下一代测序确定并且将其标准化至其在对照sir-t细胞群体,即尚未暴露于靶细胞系或暴露于未表达sir靶标的细胞系的sir-t细胞中的频率。由此分析,可以确定与不同水平的ifnγ产生相关的sir。将类似方法用于筛选并选择具有任何所需特性或属性或其组合的sir,这些特性或属性包括但不限于较低tnfα产生、较低耗尽标记物的表达、较低终末分化标记物的表达和/或更高细胞毒性标记物的表达。il7连同sir--t细胞的用途..如先前实例所述进行研究,除了在注射sir–t细胞之后开始1天,通过腹膜内注射向小鼠给与三次剂量为200ng/小鼠的外源性重组人类il-7,而对照小鼠接受生理盐水。接受il-7的小鼠显示排斥其肿瘤并且比对照小鼠存活更长时间。靶向brd4的shrna连同sir--t细胞的用途..如先前实例所述进行研究,除了使用表达由seqidno:893(plenti-ef1α-cd8sp-fmc63-vl-v5-[htcrb-kaciah]-f-p2a-sp-fmc63-vh-myc-[pretcra-del48]-f-f2a-pac-shrna-brd4-dwpre)表示的sir构建体的t细胞,该构建体共同表达针对brd4的h1启动子驱动的shrna。将此构建体与表达由seqidno:1200表示但缺乏靶向brd4的shrna的sir的构建体相比较。接受共同表达brd4-shrna(即seqidno:893)的shrna构建体的小鼠显示更长sir-t细胞持久性并且存活更长时间。使用接受表达由seqidno:1200表示的sir的t细胞的小鼠重复该实验,除了一半小鼠(n=6)接受每日两次brd4抑制剂jq1(+)(50mg/kg)的腹膜内注射,而对照小鼠接受媒介物对照(10%β-环糊精,西格玛公司)。接受jq1(+)的小鼠显示与对照小鼠相比更长sir-t细胞持久性并且存活更长时间。自体sir--t细胞用于过继性细胞疗法的用途..本批露的sir-t细胞可以用于过继性细胞疗法。作为实例,患有复发性急性淋巴细胞白血病(all)、慢性淋巴细胞白血病(cll)或高风险中级b细胞淋巴瘤的患者可以接受使用靶向cd19的过继性转移的自体sir-t细胞的免疫疗法。使用来自美天旎生物技术公司(miltenyibiotec)的clinimacs系统并且根据制造商的建议,使从每位患者收集的白细胞去除术产品经历cd3阳性t淋巴细胞的选择。任选地在用pgp抗体染色之后用流式分选、在用pgp抗体染色之后用macs、或者在暴露于th9402加上光之后进行光动力学选择,对于pgp阳性t细胞富集t淋巴细胞。将细胞用临床级别cd19-sir病毒(例如cd8sp-cd19bu12-vl-v5-[htcrb-kaciah]-f-p2a-cd19bu12-vh-myc-[htcra-csdvp]-f-f2a-gmcsf-sp-tegfr()[seqidno:1087]转导并且然后在闭合系统中发生sir-t细胞的选择和扩增。在所得细胞产物经历品质控制测试(包括无菌和肿瘤特异性细胞毒性测定)之后,将它们低温保存。同时,在白细胞去除术之后,研究参与者以淋巴细胞消耗(lymphodepletive)化学疗法(30mg/m2/日氟达拉滨加上500mg/m2/日环磷酰胺x3日)开始。在完成其淋巴细胞消耗方案之后一日,将先前保存的sir-t细胞产品运输、解冻并在患者床边输注。研究参与者接受静脉内输注的sir转导的淋巴细胞,随后每隔8小时接受高剂量(720000iu/kg)il-2(阿地白介素;加利福尼亚州圣地亚哥的普罗米修斯公司(prometheus,sandiego,ca)),获得耐受性。根据研究方案,sir-t产品的剂量从1x104sir+cd3细胞/kg至5x109sir+cd3细胞/kg之间改变。可以单次输注或分次输注给与sir-t产品。在t细胞输注与15mg/kg对乙酰氨基酚p.o.(最大值650mg)和苯海拉明0.5-1mg/kgi.v.(最大剂量50mg)之前至少30分钟向研究参与者预先给药。研究参与者可以任选地接受人类il-2的每日注射。然后可以根据医生的判断进行临床和实验室相关随访研究,并且这些研究可以包含关于表达cd19的all/淋巴瘤细胞和/或过继性转移的t细胞的存在的定量rt-pcr研究;fdg-pet和/或ct扫描;用于疾病特异性病理评估的骨髓检查;和/或根据适用于基因转移研究的fda生物反应调节剂咨询委员会(fda'sbiologicresponsemodifiersadvisorycommittee)所示的指南进行的长期随访。可以使用基本上类似的方法使用已被工程化以表达本批露的sir的自体免疫细胞(例如t细胞)治疗其它疾病,在这些自体免疫细胞中sir靶向引起疾病细胞或疾病相关细胞上表达的一种或多种抗原。靶向多种抗原的自体sir--t细胞用于过继性细胞疗法的用途..在irb批准的第i期临床实验中登记患有许多不同疾病的患者并且服用使用靶向不同引起疾病抗原或疾病相关抗原的过继性转移的自体sir-t细胞进行的免疫疗法,这些疾病包括感染疾病(例如hiv1、ebv、cmv、htlv1等)、退行性疾病(例如阿尔兹海默病)、自身免疫疾病(例如寻常天疱疮)、过敏疾病(例如慢性特发性荨麻疹)和多种癌症。基于靶抗原在引起疾病细胞或疾病相关细胞中的已知表达选择用于不同疾病的sir。在可能时,sir靶标在引起疾病细胞或疾病相关细胞上的表达通过abd-ggs-nluc融合蛋白的结合来确认,其中sir的抗原结合结构域经由柔性接头融合至非分泌形式的nluc蛋白质。可替代地,使用以可商购获得的抗体进行的免疫组织化学或流式细胞术来确认sir靶标在引起疾病细胞或疾病相关细胞上的表达。使用白细胞去除术从受试者中收集t细胞,用编码适当sir的慢病毒载体转导这些细胞并且在闭合系统中使用cd3/cd28珠粒离体扩增。在所得细胞产物经历品质控制测试(包括无菌和肿瘤特异性细胞毒性测定)之后,将它们低温保存。同时,研究参与者以淋巴细胞消耗化学疗法(30mg/m2/日氟达拉滨加上500mg/m2/日环磷酰胺x3日)开始。在完成其淋巴细胞消耗方案之后一日,研究参与者接受静脉内输注的转导的淋巴细胞,随后每隔8小时接受高剂量(720000iu/kg)il-2(阿地白介素;加利福尼亚州圣地亚哥的普罗米修斯公司),获得耐受性。将先前保存的sir-t细胞产品运输、解冻并在患者床边输注。根据研究方案,sir-t产品的剂量从1x104sir+cd3细胞/kg至5x109sir+cd3细胞/kg之间改变。可以单次输注或分次输注给与sir-t产品。在t细胞输注与15mg/kg对乙酰氨基酚p.o.(最大值650mg)和苯海拉明0.5-1mg/kgi.v.(最大剂量50mg)之前至少30分钟向研究参与者预先给药。研究参与者可以任选地接受人类il-2的每日注射。然后可以根据医生的判断进行临床和实验室相关随访研究。mtor抑制剂rad001与sir--t细胞的组合的用途..如先前实例所述进行研究,除了在输注sir–t细胞之后开始1日,向研究参与者给予mtor抑制剂,例如变构抑制剂,例如rad001,其剂量提供目标低谷水平0.1至3ng/ml,其中低谷水平”是指正好在下一个剂量之前血浆中的药物浓度或者两个剂量之间的最小药物浓度。依鲁替尼与sir--t细胞的组合的用途..如先前实例所述进行研究,除了在输注sir–t细胞之后开始1日,向研究参与者给予剂量为140mg/d至420mg/d的口服依鲁替尼。应注意,接受依鲁替尼的研究参与者与接受sir-t细胞而无依鲁替尼的参与者相比具有严重细胞因子释放综合征的更小发生率。同种异体sir--t细胞用于过继性细胞疗法的用途..患有复发性急性淋巴细胞白血病或高风险中级b细胞淋巴瘤且经历同种异体骨髓移植的患者可接受使用过继性转移的异体sir-t细胞的免疫疗法。使用来自美天旎生物技术公司的clinimacs系统并且根据制造商的建议,使从每位供体(如对于同种异体移植所使用的相同供体)收集的白细胞去除术产品经历cd3阳性t淋巴细胞的选择。任选地在用pgp抗体染色之后用流式分选、在用pgp抗体染色之后用macs、或者在暴露于th9402加上光之后进行光动力学选择,对于pgp阳性t细胞富集t淋巴细胞。使用基于cd3和cd28磁珠的人工抗原呈递细胞活化细胞并且用临床级别cd19-sir病毒(例如cd8sp-cd19bu12-vl-v5-[htcrb-kaciah]-f-p2a-cd19bu12-vh-myc-[htcra-csdvp]-f-f2a-icasapase9[seqidno:1080]转导。在闭合系统中将细胞扩增9-12天。在所得细胞产物经历品质控制测试(包括无菌和肿瘤特异性细胞毒性测定)之后,将它们低温保存。同时,研究参与者以淋巴细胞消耗化学疗法(30mg/m2/日氟达拉滨加上500mg/m2/日环磷酰胺x3日)开始。在完成其淋巴细胞消耗方案之后一日,研究参与者接受静脉内输注的转导的淋巴细胞,随后每隔8小时接受高剂量(720000iu/kg)il-2(阿地白介素;加利福尼亚州圣地亚哥的普罗米修斯公司),获得耐受性。将sir-t细胞产品运输、解冻并在患者床边输注。根据研究方案,sir-t产品的剂量可从1x104sir+cd3细胞/kg至5x109sir+cd3细胞/kg之间改变。可以单次输注或分次输注给与sir产品。在sir-t细胞输注与15mg/kg对乙酰氨基酚p.o.(最大值650mg)和苯海拉明0.5-1mg/kgi.v.(最大剂量50mg)之前至少30分钟向研究参与者预先给药。然后可以根据医生的判断进行临床和实验室相关随访研究,并且这些研究可以包含关于表达cd19的all/淋巴瘤细胞和/或过继性转移的t细胞的存在的定量rt-pcr研究;fdg-pet和/或ct扫描;用于疾病特异性病理评估的骨髓检查;和/或根据适用于基因转移研究的fda生物反应调节剂咨询委员会(fda'sbiologicresponsemodifiersadvisorycommittee)所示的指南进行的长期随访。也根据医生的判断使用免疫抑制药物。可以使用基本上类似的方法使用表达本批露的sir的同种异体免疫细胞(例如t细胞)治疗其它疾病,在这些同种异体免疫细胞中sir靶向引起疾病细胞或疾病相关细胞上表达的一种或多种抗原。sir--t细胞肝动脉输注..除了静脉内输注之外,可以动脉内输注sir-t细胞以在涉及疾病的局部区域或器官中提供高浓度sir-t细胞。在以下实例中,在患有来自表达叶酸受体α(fr1)的胃肠癌的肝转移癌的患者的情况下使用此方法。可以使用基本上类似的方法来动脉内输注靶向其它肿瘤抗原的sir-t细胞。在基线时经由右股总动脉方法进行映射血管造影。除了其它潜在来源的肝外灌注之外,用微线圈栓塞胃与十二指肠和胃右动脉。进行相同动脉出入程序以用于给予表达cd8sp-fr1-humov19-vl-v5-[htcrb-kaciah]-f-p2a-sp-fr1-humov19-vh-myc-[htcra-csdvp]-f-f2a-pac(102915-p07)[seqidno:1276]sir的t细胞。在第0天从患者收集t细胞并且用编码sir的慢病毒感染这些t细胞并如先前实例所述扩增。在第14天(108个sir-t细胞)、第28天(109个sir-t细胞)和第44天(1010个sir-t细胞)以剂量递增方式给予sir-t细胞。经由60cc注射器以<2cc/s的速率手动注射sir-t细胞。输注的总体积是大约100cc。在第一次输注50cc之后并且在完成sir-t输注后进行具有校准的比对率的血管造影术,以确认保留的动脉血流。当可能时将输注物递送到适合的肝动脉。某些患者具有异常肝动脉解剖结构,其中右肝动脉或左肝动脉不是从适当肝动脉开始。在此类情况下,基于肝叶体积计算分割sir-t细胞的剂量。在此类情况下,将分割的剂量单独递送至右肝动脉和左肝动脉以确保成比例的sir-t递送至两个肝叶。在基线时、在输注日、以及在输注后1日、2日、4日和7日进行临床评估。在第一次输注之前一个月内并且然后在最后一次sir-t输注之后一个月内安排使用肝mri和pet检查的计划的成像评定。研究放射学家(bs)根据改良recist(mrecist)和免疫相关反应标准(wolchok等人,2009,clincancerres[临床癌症研究],15:7412-7420)将反应分级。在处理之前以及最终剂量之后三周进行经皮活检。盲性病理学家对活检样本上的肿瘤坏死和纤维化进行评分。根据方案进行安全性评估。使用国家癌症研究所通用术语标准不良事件版本3.0(nationalcancerinstitutecommonterminologycriteriaforadverseeventsversion3.0)对不良事件的严重性进行分级。sir--t细胞的腹膜内给予..还可以腹膜内给予sir-t细胞,基本上如konerum等人(journaloftranslationalmedicine[转化医学杂志];2015;13:102)所述。在以下实例中,在患有表达叶酸受体α(fr1)的腹膜参与的卵巢癌的患者的情况下使用此方法。可以使用基本上类似的方法来腹膜内输注靶向其它肿瘤抗原的sir-t细胞。将向患有复发性高级严重卵巢癌的患者提供筛查知情同意书以测试其表达fr1的癌症。在通过免疫组织化学确认fr1的表达的情况下,那么患者将具有从外周血中获得的白细胞去除术产品。从白细胞去除术产品和冷冻产品中去除过量血小板和红血细胞污染。在研究的治疗期中,将解冻并洗涤白细胞去除术产品。随后,将通过使用cd3/cd28珠粒进行磁性分离来从解冻的白细胞去除术产品分离cd3+t细胞。将用cd8sp-fr1-humov19-vl-v5-[htcrb-kaciah]-f-p2a-sp-fr1-humov19-vh-myc-[htcra-csdvp]-f-f2a-pac(102915-p07)[seqidno:1276]sir慢病毒转导活化的t细胞并且使用cd3/cd28珠粒扩增方案进一步扩增。这是i期临床试验,该试验在患有复发性fr1+卵巢癌、输卵管或原发性腹膜癌的患者中测试静脉内(iv)和腹膜内(ip)输注(使用或不使用先前环磷酰胺化学疗法)遗传修饰的自体t细胞的安全性。这些自体t细胞将被遗传工程化以表达cd8sp-fr1-humov19-vl-v5-[htcrb-kaciah]-f-p2a-sp-fr1-humov19-vh-myc-[htcra-csdvp]-f-f2a-pac(102915-p07)[seqidno:1276]sir。患有复发性高级严重卵巢癌、原发性腹膜癌或输卵管癌的患者对于该研究而言将是合格的,这些癌症已显示表达通过免疫组织化学(ihc)分析存放的(banked)(石蜡包埋)或新鲜活检的肿瘤来确认的fr1抗原。仅中度至强免疫反应评分(3–5)将被认为是阳性的,其中评分3被描述为51%-75%强染色或51%-100%弱染色,4被描述为76%-99%强染色,并且5被描述为100%强染色。所有患者将均接受现有化学疗法用于复发病症,其中最多允许五种现有级别的化学疗法。患有其它活动性恶性肿瘤、寿命<3个月、或在计划治疗时卡氏体力状况(karnofskyperformancestatus,kps)评分<70%的患者将是不合格的。在该试验中将使用i期剂量递增给药。将向3–6位患者的组输注递增剂量的修饰t细胞以建立最大耐受剂量(mtd)。存在四个计划的剂量水平:3×105、1×106、3×106和1×107cd8sp-fr1-humov19-vl-v5-[htcrb-kaciah]-f-p2a-sp-fr1-humov19-vh-myc-[htcra-csdvp]-f-f2a-pac(102915-p07)[seqidno:1276]sir-t细胞/kg。将用3×105cd8sp-fr1-humov19-vl-v5-[htcrb-kaciah]-f-p2a-sp-fr1-humov19-vh-myc-[htcra-csdvp]-f-f2a-pac(102915-p07)[seqidno:1276]sir-t细胞/kg治疗i组和ii组,但是ii组患者也将接受淋巴细胞消耗环磷酰胺。ii-v组将在用环磷酰胺预先治疗之后接受递增剂量的修饰t细胞。以750mg/m2给药的淋巴细胞消耗环磷酰胺将在初始t细胞输注之前2–4天给与。将遵循标准3+3剂量递增方案。如果第一剂量水平超过mtd,则后续3–6位患者的组将以-1剂量水平1×105cd8sp-fr1-humov19-vl-v5-[htcrb-kaciah]-f-p2a-sp-fr1-humov19-vh-myc-[htcra-csdvp]-f-f2a-pac(102915-p07)[seqidno:1276]sir-t细胞/kg治疗而无需添加淋巴细胞消耗的环磷酰胺(-i组)。将放置ip导管,之后进行t细胞输注。将在修饰t细胞准备好给与时放置导管。将准许患者进入医院的住院病人病房,之后首次输注sirt细胞,并且将一直住院,直到第二次输注sirt细胞之后至少3天。将准许第一组有待治疗的患者和每个后续组中治疗的第一位患者进入特护病房(icu);可以准许后续患者进入内科肿瘤学住院服务(经过治疗医生的临床判断)。患者将在开始用sir修饰的t细胞治疗之前2至4天接受单一剂量的淋巴细胞消耗的环磷酰胺(750mg/m2iv)化学疗法。将在输注之前对转导的t细胞进行数量、纯度、生存力和无菌性的品质测试。必须满足以下t细胞释放标准,包括生存力>80%、cd3+≥95%并且基于扩增的t细胞群体的流式细胞术分析,输注的t细胞群体必须具有>20%的转导级份。另外,转导的t细胞的平均载体拷贝数目将在输注之前通过实时pcr测定并且将需要处于0.3至5个拷贝/细胞的范围内,并且将使用pcr来确保复制能力的慢病毒不存在于转导的t细胞中。所有患者均将静脉内接受50%遗传修饰的t细胞剂量。将密切监测患者的毒性。在一至3天后,将以ip输注方式给与其余剂量的t细胞。将以剂量水平1治疗至少3位患者,其中在每个剂量水平内每个月不超过2位患者有自然增长。在登记的每位患者的治疗之间的时间将为至少一周。在先前组中治疗的所有患者从初始t细胞输注那天开始观察最少4周,之后下一个组发生递增。根据在环磷酰胺疗法之后中性粒细胞减少(anc≤1000/mm3)的显著风险,用环磷酰胺治疗的患者可以根据研究者的判断用生长因子支持物治疗(单次皮下注射乙二醇化非格司亭或3个连续日的皮下非格司亭)。血液样品将在评定遗传修饰的t细胞的毒性、治疗效果和存活之前和之后从所有患者获得。将在t细胞输注之后大约1h、1天以及1、2、3、4、5、6、7、8和12周收集治疗后血液样品,然后之后每月收集直至1年,然后之后每年收集,直至t细胞输注之后15年。如果技术上可行的话,也可以在环磷酰胺或t细胞疗法(首先进行)之前以及随访期间对患者的腹水进行取样。在t细胞输注之后大约6周、3个月、6个月、9个月和12个月以及在临床上指示时在此之后,患者将进行ct扫描。sir--t细胞用于肿瘤内注射的用途,也可以肿瘤内给与sir-t细胞,基本上如brownce等人,clincancerres.[临床癌症研究]2015年9月15日;21(18):4062–4072所述。在以下实例中,在患有表达il13ra2的复发性恶性胶质瘤(gbm)的患者的情况下使用此方法。可以使用基本上类似的方法来肿瘤内输注靶向其它肿瘤抗原的sir-t细胞。将进行初步安全性和可行性研究以在复发性gbm中测试表达cd8sp-il13ra2-hu107-vl-v5-[htcrb-kaciah]-f-p2a-sp-il13ra2-hu107vh-myc-[htcra-csdvp]-f-f2a-pac(051816-y03)[seqidno:1306]sir的t细胞。将要求所有参与患者签署知情同意书。该临床方案将由南加州大学机构审查委员会(universityofsoutherncaliforniainstitutionalreviewboard)批准并且根据研究性新药申请(investigationalnewdrugapplication)进行,并且在clinicaltrials.gov.上登记。合格患者将包括患有复发性或难治性单病灶幕上iii或iv级神经胶质瘤且其肿瘤不显示与心室/csf途径通信并且易于切除的成人(18-70岁)。参与此试验将独立于il13rα2(或il13ra2)肿瘤抗原状态。患者将在初始诊断高级神经胶质瘤(whoiii或iv级)之后登记,此时他们将经历白细胞去除术,以收集外周血单核细胞(pbmc)。将使用这些细胞以在用如先前实例所述的对应慢病毒载体感染之后使t细胞工程化以表达含有嘌呤霉素抗性基因(pac)的cd8sp-il13ra2-hu107-vl-v5-[htcrb-kaciah]-f-p2a-sp-il13ra2-hu107vh-myc-[htcra-csdvp]-f-f2a-pac(051816-y03)[seqidno:1306]sir。可替代地,可以在用逆转录病毒感染之后或者使用睡美人转座子或通过转染ivtmrna来生成sir-t细胞。随后,将低温保存释放测试的治疗性sir-t细胞并且将其保存用于随后使用。在肿瘤首次复发时,研究参与者将经历肿瘤切除以及瑞克曼储器(rickhamreservoir)/导管的放置。同时,将治疗性sir-t细胞解冻,在体外使用基于cd3/cd28珠粒的快速扩增方案重新扩增。在从手术恢复之后并在基线mr成像之后,将经由内置导管将cd8sp-il13ra2-hu107-vl-v5-[htcrb-kaciah]-f-p2a-sp-il13ra2-hu107vh-myc-[htcra-csdvp]-f-f2a-pac(051816-y03)[seqidno:1306]sir直接给予至切除腔,基本上如(brownce等人,clincancerres.[临床癌症研究]2015年9月15日;21(18):4062–4072)所述。将使用21号蝴蝶针将细胞手动注射到瑞克曼储器中,以在5-10分钟内递送2ml体积,随后在5分钟内用无防腐剂生理盐水进行2ml冲洗。方案治疗计划将指定患者内剂量递增安排表,其目标在于在包括每周治疗周期的5周时间段内颅内给予12个cart细胞剂量。在第1、2、4和5个周期内,将在周期周的第1、3和5天进行t细胞输注,并且第3周将是休息周期。为了安全,在第1周期内,我们将利用患者内递增策略,其中在第1、3和5天分别给予107、5×107和108个细胞/输注的sirt细胞剂量,并且这之后将是在4周内9次额外sirt细胞输注108个细胞。在第3周休息周期内和第5周之后将进行成像以评定反应。将遵循nct常见毒性标准2.0版(ncicommontoxicitycriteriaversion2.0,https://ctep.ifo.nih.gov/l)中所提供的指南,以用于监测毒性和不良事件报道sir--t细胞用于在移植之前离体净化骨髓或外周血干细胞制备物的用途..可以使用sirt细胞在干细胞移植之前净化癌细胞的骨髓或外周血干细胞制备物。在以下实例中,在自体干细胞(或骨髓)移植之前使用表达cd8sp-huluc64-vl-v5-[htcrb-kaciah]-f-p2a-sp-huluc64-vh-myc-[htcra-csdvp]-f-f2a-pac(092916-e07)[seqidno:1253]的sir-t细胞净化从患有多发性骨髓瘤的患者获得骨髓或外周血干细胞。患者将经历白细胞去除术以收集外周血单核细胞(pbmc)。将使用cd3珠粒纯化t细胞。将使用这些细胞以在用如先前实例所述的对应慢病毒载体感染之后使t细胞工程化以表达含有嘌呤霉素抗性基因(pac)的cd8sp-huluc64-vl-v5-[htcrb-kaciah]-f-p2a-sp-huluc64-vh-myc-[htcra-csdvp]-f-f2a-pac(092916-e07)[seqidno:1253]sir。此sir靶向骨髓瘤细胞上表达的抗原cs1。表达cd8sp-cs1-huluc90-vl-v5-[htcrb-kaciah]-f-p2a-sp-huluc90-vh-myc-[htcra-csdvp]-f-f2a-pac(012716-a02)[seqidno:1254]、cd8sp-cd138-vl-v5-[htcrb-kaciah]-f-p2a-sp-cd138-vh-myc-[htcra-csdvp]-f-f2a-pac(100815-a05)[seqidno:1236]或cd8sp-gprc5d-et150-5-vl-myc2-[htcrb-kaciah]-f-p2a-sp-gprc5d-et150-5-vh-myc4-[htcra-csdvp]-f-f2a-pac(100616-c03)[seqidno:1285]的sir-t将作为替代方案使用或者与以上靶向cs1的sir-t细胞组合使用。可替代地,可以在用逆转录病毒感染之后或者使用睡美人转座子或通过转染ivtmrna来生成sir-t细胞。随后,将低温保存释放测试的治疗性sir-t细胞并且将其保存用于随后使用或者新鲜使用。骨髓细胞和外周血祖细胞产品将在标准程序之后从患有多发性骨髓瘤的患者中收集。对于外周血干细胞的调动,患者将接受3gm/m2环磷酰胺,随后在环磷酰胺之后24h开始每天皮下接受10μg/kgg-csf,直至完成提取法。一旦外周血cd34+-细胞计数为15个细胞/μl,就将收集外周血干细胞。收集目标将是每天处理三个血液体积,直至在处理之后达到最少2.0倍的106cd34+细胞/kg。使用来自美天旎生物技术公司的clinimacs系统并且根据制造商的建议,将任选地耗尽骨髓和外周血干细胞产品中的红血细胞和/或富集表达cd34的细胞。将使用这些产品以离体净化新鲜或低温保存产品。为了净化,将骨髓或外周血干细胞产品与解冻的sir-t细胞在补充有100iu重组人类il2的xvivo培养基(龙沙公司)中以从5:1至30:1的效应物与靶标比率范围共同培养4至24小时。将在37℃下在5%co2润湿培养器中培养细胞。在共同培养时间段结束后,获取细胞等份试样以用于无菌和品质测试(包括测量cfu-gm以及cd34和cd138阳性细胞的流式细胞术)。将向先前已接受脊髓抑制性化学疗法(例如,高剂量美法仑,两个分剂量为70mg/m2,总剂量为140mg/m2)静脉内给予其余样品。自身sir用于抑制自身免疫疾病的用途..在临床试验中将登记患有寻常天疱疮的患者以测试含有dsg3细胞外结构域的sir的安全性和功效。患者将在诊断对治疗有抗性的寻常天疱疮之后登记,此时他们将经历白细胞去除术,以收集外周血单核细胞(pbmc)。将使用这些细胞以在用如先前实例所述的对应慢病毒载体感染之后使t细胞工程化以表达sircd8sp-myc-[htcra-t48c-opt1]-f-f2a-sp-dsg3-ecd-v5-[htcrb-s57c-opt1]-f-p2a-pac(seqidno:1144)。可替代地,将在用逆转录病毒感染之后或者使用睡美人转座子或通过转染ivtmrna来生成sir-t细胞。随后,将低温保存释放测试的治疗性sir-t细胞并且将其保存用于随后使用。患者将在开始用sir修饰的t细胞治疗之前2至4天接受单一剂量的淋巴细胞消耗的环磷酰胺(750mg/m2iv)化学疗法。将在输注之前对转导的t细胞进行数量、纯度、生存力和无菌性的品质测试。根据研究方案,sir-t产品的剂量可从1x104sir+cd3细胞/kg至5x109sir+cd3细胞/kg之间改变。可以单次输注或分次输注给与sir产品。在sir-t细胞输注与15mg/kg对乙酰氨基酚p.o.(最大值650mg)和苯海拉明0.5-1mg/kgi.v.(最大剂量50mg)之前至少30分钟向研究参与者预先给药。然后可以根据医生的判断进行临床和实验室相关随访研究。上文已陈述许多实施方式来说明本公开。以下权利要求将进一步陈述申请人的
发明内容。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 金黄色葡萄球菌肠毒素C2的核酸适配体C203及其筛选方法和应用与制造工艺
- 一种全人源化EGFRvIII嵌合抗原受体T细胞及其制备方法与制造工艺
- 嵌合抗原受体及制备方法与制造工艺
- 检测外周血CAR‑T细胞的TaqMan实时荧光定量PCR试剂盒的制造方法与工艺
- CD20特异性嵌合抗原受体及其应用的制造方法与工艺
- 一种用于辣根过氧化物酶染色的染色剂及其制备方法、染色组合、应用及标记酶染色试剂盒与制造工艺
- 对SSEA4抗原具有特异性的嵌合抗原受体的制造方法与工艺
- 具有针对CD79和CD22的特异性的分子的制造方法与工艺
- 具有针对CD45和CD79的特异性的分子的制造方法与工艺
- 一种检测嵌合抗原受体T细胞对肝癌细胞抑制作用的模型和方法与制造工艺