一种特异性调节神经铁代谢的小分子及其应用的制作方法
本发明属药物制药领域,涉及特异性调节神经铁代谢的小分子及其应用,具体涉及gsk-j4及其靶点在制备特异性调节神经铁代谢的药物中的用途。
背景技术:
资料记载了铁是人体必需微量元素之一,铁代谢平衡紊乱,会导致体内铁缺乏或铁过载。然而,铁在多种神经退行性疾病中的特定脑区显著聚集,如阿尔兹海默病,帕金森病,亨廷顿病等。有研究显示,脑铁的蓄积目前已被认为该些神经退行性疾病的主要原因之一,因此,以铁脑铁代谢为靶点,寻找特异性调节神经铁代谢,对脑铁代谢紊乱疾病的治疗将具有重要意义。
基于现有技术的基础,本申请的发明人拟提供一种特异性调节神经铁代谢的小分子及其应用,具体涉及gsk-j4及其靶点在制备特异性调节神经铁代谢的药物中的用途。
技术实现要素:
本发明目的是提供一种特异性调节神经铁代谢的小分子及其应用,尤其是小分子gsk-j4及其靶点在制备特异性调节神经铁代谢以及脑铁代谢紊乱疾病的药物中的应用,本发明将为新药筛选与制备提供基础。
本发明采用的技术方案是:
小分子gsk-j4及其靶点在特异性调节神经铁代谢及其应用。
基于现有技术的有关铁蓄积以及铁蓄积引起的氧化损伤已经成为多种神经退行性疾病的重要原因的研究基础,本发明筛选能够特异性调节神经铁代谢的小分子,进一步为解决脑铁代谢紊乱疾病提供基础。
本发明通过利用多巴胺能神经瘤细胞sh-sy5y细胞系,处理不同的小分子,结合石墨炉原子吸收光谱法gfaas检测sh-sy5y细胞铁水平,结果发现微量的小分子gsk-j4(0.1-1μm)能够显著降低神经细胞系sh-sy5y内的铁水平;通过calcein-am实验,铁补充剂(fac)能够显著降低calcein-am的信号,而gsk-j4处理却显著降低细胞内calcein-am的信号,并且铁补充剂fac能够显著抑制gsk-j4引起的calcein-am信号降低,这意味着gsk-j4能够显著降低神经细胞内游离的铁水平;
本发明通过蛋白分析,发现gsk-j4能够显著增加sh-sy5y细胞内铁摄取蛋白tfr1的水平,降低细胞内储铁蛋白ftl的水平,这进一步反映了gsk-j4处理引起了细胞内游离铁的降低;富有意义的是,本发明中发现gsk-j4能够显著且特异性降低神经细胞系sh-sy5y内的铁水平,以及特异性调节sh-sy5y的铁代谢,但是并没有影响外周细胞系的铁水平与铁代谢,如肝脏细胞系hepg2以及肾脏细胞系hek293;进一步,本发明中,gsk-j4能显著增加sh-sy5y细胞内的铁吸收蛋白dmt1,tfr1,铁外排蛋白fpn1,降低铁储存蛋白ftl,额外的铁补充剂能够显著缓解gsk-j4对这些蛋白的影响(fpn1除外),然而gsk-j4对调控fpn1水平的主要因子——铁调素hepcidin并没有显著影响,这些数据表明gsk-j4初始铁调节靶点是fpn1,并且该调节并不依赖于hepcidin-fpn1这条通路;通过检测gsk-j4已知的靶点(h3k27me3和h3k4me3),结果显示gsk-j4能够显著增加h3k27me3和h3k4me3水平,并且与铁蛋白变化一致,仅特异性出现在sh-sy5y细胞;进一步,通过添加h3k27me3抑制剂(gsk-126),结果显示,gsk-126并没有显著抑制gsk-j4对sh-sy5y细胞铁代谢的影响,然而抑制h3k4me3(通过小分子dznep)能够显著抑制gsk-j4对sh-sy5y细胞铁代谢的影响,这表明gsk-j4能够显著且特异性增加sh-sy5y细胞内h3k4me3水平,升高fpn1水平,从而特异性调节神经铁代谢;
本发明中,通过特异性升高h3k4me3水平(添加小分子cpi-455或者抑制kdm5c表达的腺相关病毒)能够显著增加神经细胞系以及原代细胞内fpn1水平,h3k4me3/kdm5是gsk-j4特异性调节神经细胞铁外排蛋白fpn1的靶点,gsk-j4已被报道能够显著通过血脑屏障;
本发明进行了体外细胞实验,结果显示gsk-j4能够显著抑制神经毒素mpp+引起的细胞毒性,进一步通过腹腔注射gsk-j4,结合mptp诱导的小鼠帕金森模型,结果显示gsk-j4能够显著改善mptp引起的小鼠帕金森疾病症状,如改善帕金森小鼠的运动能力,抑制帕金森小鼠黒质多巴胺能神经元神经元的丢失;
本发明的实验结果为小分子gsk-j4以及其靶点(kdm5/h3k4me3)特异性改善神经铁代谢紊乱疾病提供了依据,特别是对帕金森疾病,阿尔茨海默病等神经退行性疾病的药物研发提供了基础。
进一步,所述的gsk-j4及其靶点可用于制备特异性调节神经铁代谢的药物;尤其是所述小分子gsk-j4及其针对h3k4me3/kdm5的靶点药物可用于制备防治帕金森疾病,阿尔茨海默病等神经退行性疾病的饮食补充剂(如保健品)或药物。
通常,所述的饮食补充剂含0.01~50wt%,较佳地为0.1~5wt%的杨梅素;以及食品上可接受的载体。
所述的药物可添加药学上可接受的载体,制成常规的剂型,如片剂、胶囊、颗粒剂等。
本发明中,药学上可接受的载体指用于治疗剂给药的载体,包括各种赋形剂和稀释剂,它们本身并不是必要的活性成分,且施用后没有过分的毒性;合适的载体是本领域普通技术人员所熟知的,可含有液体,如水、盐水、甘油和乙醇;另外,这些载体中还可能存在辅助性的物质,如填充剂、润滑剂、助流剂、润湿剂或乳化剂、矫味剂、ph缓冲物质等。
从易于制备和给药的立场看,优选的药物是口服药物,尤其是片剂和固体填充或液体填充的胶囊。
本发明所述的药物的剂型可以是多种多样的,只要是能够使活性成分有效地到达哺乳动物体内的剂型都是可以的,比如可选自:片剂、胶囊、粉末、颗粒、糖浆、溶液、悬浮液、或气雾剂,其中杨梅素可以存在于适宜的固体或液体的载体或稀释液中,也可储存在适宜于注射或滴注的消毒器具中;通常,在制备的药物中,gsk-j4占总重量的0.01~20wt%,较佳地为0.05~5wt%,其余为药学上可接受的载体以及其它添加剂等物质。
本发明的有益效果主要体现在:
本发明提供了小分子gsk-j4及其靶点在制备特异性调节神经铁代谢药物的应用,此外已有研究表明gsk-j4能够显著通过血脑屏障,更进一步为gsk-j4在神经退行性疾病的应用提供了有利的条件,为gsk-j4改善脑铁性疾病提供了理论依据,特别是帕金森疾病,阿尔茨海默病等神经退行性疾病的药物研发提供了基础。
附图说明
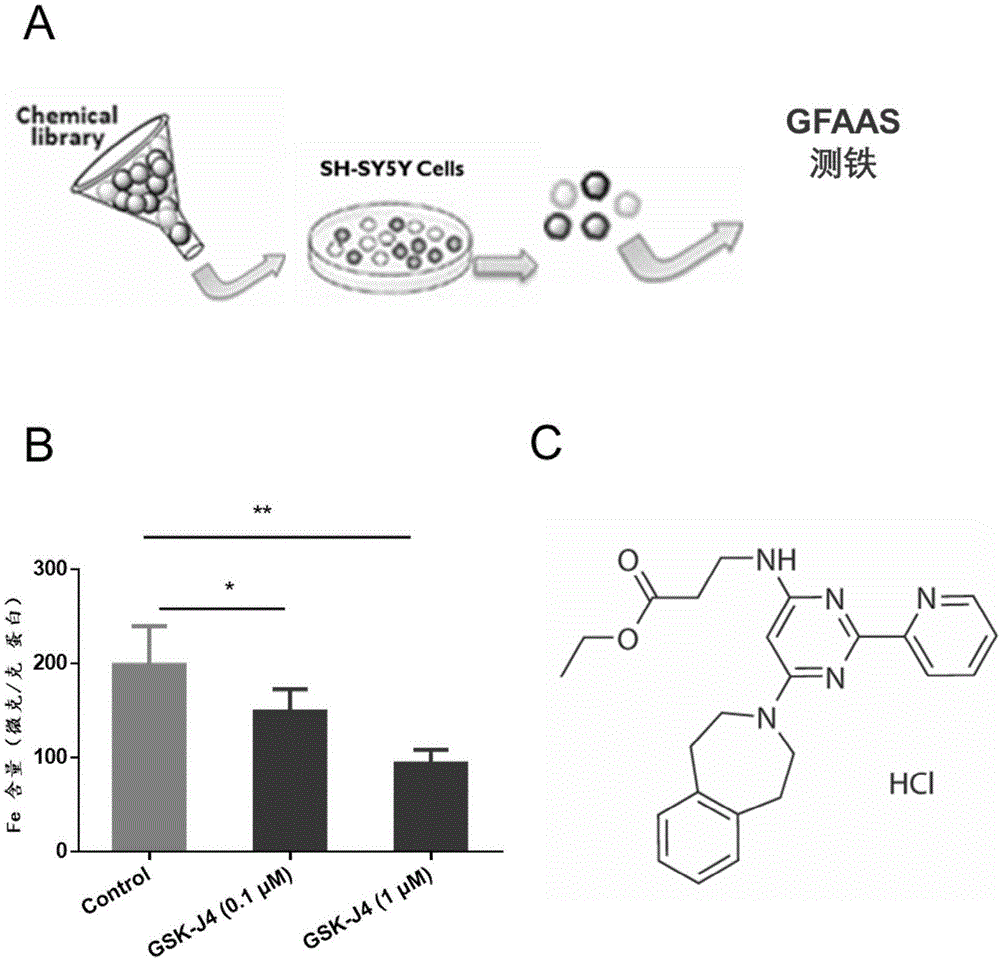
图1,筛选发现gsk-j4能够显著降低神经细胞系sh-sy5y细胞内铁;其中,
a:筛选模式图;
b:不同浓度的小分子gsk-j4(0.1-1μm)对sh-sy5y细胞铁的影响;
c:小分子gsk-j4的化学结构式。
图2为小分子gsk-j4能够显著影响神经细胞内游离铁水平及铁代谢;其中,
a:铁补充剂fac(50μg/ml)和gsk-j4(0.5μm)处理sh-sy5y细胞12小时,对sh-sy5y细胞calcein-am信号的影响;
b:calcein-am信号的定量结果;
c:gsk-j4(0.5μm)以及铁螯合剂dfo(50μm)处理sh-sy5y细胞12h,蛋白检测分析铁储存蛋白ferritin-l(ftl)和铁摄取蛋白transferritinreceptor1(tfr1);
d:定量分析铁储存蛋白ftl以及铁摄取蛋白tfr1水平;
e-f:免疫荧光实验分析gsk-j4对铁储存蛋白的影响。
图3为gsk-j4对外周细胞系铁代谢无显著影响;其中,
a:gsk-j4(0.5μm)对三种细胞系:人神经瘤细胞系sh-sy5y,人源肾细胞系hek293以及人源肝癌细胞系hepg2细胞内铁水平的影响;
b:gsk-j4(0.5μm)以及铁螯合剂dfo(50μm)处理人源肾脏细胞系hek293细胞12h,蛋白检测分析铁储存蛋白ferritin-l(ftl)和铁摄取蛋白transferritinreceptor1(tfr1);
c:定量分析gsk-j4(0.5μm)以及铁螯合剂dfo(50μm)处理人源肾脏细胞系hek293细胞对铁储存蛋白ftl以及铁摄取蛋白tfr1水平;
d:gsk-j4(0.5μm)以及铁螯合剂dfo(50μm)处理人源肝脏系hepg2细胞12h,蛋白检测分析铁储存蛋白ferritin-l(ftl)和铁摄取蛋白transferritinreceptor1(tfr1);
e:定量分析gsk-j4(0.5μm)以及铁螯合剂dfo(50μm)处理人源肝脏细胞系hepg2细胞对铁储存蛋白ftl以及铁摄取蛋白tfr1水平。
图4为gsk-j4对神经细胞系铁代谢相关蛋白的影响;其中,
a:gsk-j4(0.5μm)以及铁补充剂fac(50μg/ml)处理人源神经瘤细胞系sh-sy5y细胞12h,蛋白检测分析gsk-j4对铁摄入蛋白tfr1和dmt1的影响;
b:定量分析gsk-j4(0.5μm)以及铁补充剂fac(50μg/ml)处理人源神经瘤细胞系sh-sy5y细胞对铁储存蛋白ftl以及铁摄取蛋白tfr1水平;
c:gsk-j4(0.5μm)以及铁补充剂fac(50μg/ml)处理人源神经瘤细胞系sh-sy5y细胞12h,蛋白检测分析gsk-j4对铁外排蛋白fpn1和铁储存蛋白ftl的影响;
d:定量分析gsk-j4(0.5μm)以及铁补充剂fac(50μg/ml)处理人源神经瘤细胞系sh-sy5y细胞对铁外排蛋白fpn1和铁储存蛋白ftl的影响;
e-f:gsk-j4(0.5μm)处理人源神经瘤细胞系sh-sy5y细胞12h,蛋白检测分析gsk-j4对铁调素hepcidin的影响;
g:gsk-j4(0.5μm)处理人源神经瘤细胞系sh-sy5y细胞12h,定量pcr检测分析gsk-j4对铁外排蛋白fpn1mrna的影响。
图5为组蛋白三甲基化水平对gsk-j4特异性对神经细胞系铁代谢的影响;其中,
a:小分子gsk-j4(0.5μm)以及h3k27me3抑制剂(1μm)处理人源神经瘤细胞系sh-sy5y细胞12h,蛋白检测分析gsk-j4对h3k27me3,铁外排蛋白fpn1,铁摄入蛋白tfr1以及铁储存蛋白ftl的影响;
b:小分子gsk-j4(0.5μm)以及h4k4me3抑制剂(0.5μm)处理人源神经瘤细胞系sh-sy5y细胞12h,蛋白检测分析gsk-j4对h3k4me3,铁外排蛋白fpn1,铁摄入蛋白tfr1以及铁储存蛋白ftl的影响;
c:小分子gsk-j4(0.5μm)处理人源肾脏细胞系hek293细胞12h,蛋白检测分析gsk-j4对h3k27me3,h3k4me3的影响;
d:gsk-j4(0.5μm)处理人源神经瘤细胞系sh-sy5y细胞12h,定量分析gsk-j4对h3k27me3,h3k4me3的影响。
图6为升高组蛋白h3k4me3三甲基化水平能够增加铁外排蛋白fpn1水平;其中,
a:小分子cpi-455(1-5μm)处理人源神经瘤细胞系sh-sy5y细胞12h,蛋白检测分析gsk-j4对h3k4me3以及铁外排蛋白fpn1的影响;
b:小分子cpi-455(1-5μm)处理人源神经瘤细胞系sh-sy5y细胞12h,定量分析gsk-j4对h3k4me3以及铁外排蛋白fpn1的影响。
图7为抑制kdm5c水平显著增加铁外排蛋白fpn1水平,其中,
a-b:腺相关病毒aav-kdm5csilencevirus感染大鼠原代皮层神经元细胞72h,荧光定量pcr检测分析aav-kdm5csilencevirus对kdm5cmrna的影响;
c-d:腺相关病毒aav-kdm5csilencevirus感染大鼠原代皮层神经元细胞72h,蛋白质检测分析aav-kdm5csilencevirus对kdm5c蛋白的影响;
e-f:腺相关病毒aav-kdm5csilencevirus感染大鼠原代皮层神经元细胞72h,蛋白质检测分析aav-kdm5csilencevirus对fpn1和h3k4me3的影响。
图8gsk-j4在体外对mpp+诱导的帕金森疾病模型的保护作用,其中,
a:不同浓度的gsk-j4(0.05-20μm)对sh-sy5y细胞活性的影响;
b:gsk-j4(0.5μm)预处理1小时,对mpp+(0.5mm)诱导sh-sy5y帕金森疾病细胞模型的保护作用;
c:细胞活性的定量分析。
图9为gsk-j4在体内对mpp+诱导的帕金森疾病模型的保护作用;其中,
a:实验流程:mptp诱导的小鼠帕金森模型以及gsk-j4给药方式;
b:小鼠矿场行为分析,小鼠矿场运动距离;
c:小鼠矿场行为分析,小鼠矿场不动时间;
d:小鼠矿场行为分析,小鼠矿场运动代表轨迹以及小鼠矿场平均活动热度图;
e-g:gsk-j4能够显著抑制mptp引起的小鼠黒质以及纹状体多巴胺能神经元的丢失。
具体实施方式
下面结合具体实施例对本发明进行进一步描述,但本发明的保护范围并不仅限于此。
实施例1:
1.材料和方法
1.1实验原料制备
所有的试剂均够买于sigmachemicalcompany,st.louis,mo,usa.inaddition,除了gsk-j4(s7070,selleckchemicals),cpi-455(s8287,selleckchemicals),gsk126(15415,caymanchemical),3-deazaneplanocinahcl(dznep)(s7120,selleckchemicals),actd(11805017,thermofisherscientific),chx(239764,merckmillipore),mg-132(474790,merckmillipore).
1.2细胞株
人源神经瘤细胞系sh-sy5y由中国科学院上海生命科学院细胞库提供。培养条件:含10%fbs(gibco)的dmem高糖型培养基(gibco),37℃,5%co2饱和湿度培养箱。
1.3实验动物
6-10周龄spf级c57bl/6雄性小鼠购自上海斯莱克实验动物有限公司,spf环境饲养。基础饲料ain-76a(铁含量0.9mg/kg,researchdiets,inc)适应喂养一周后,按体重随机分组,每组6-8只,根据实验设计,对照组和实验组分别予以不同处理后,5%水合氯醛腹腔麻醉后,pfa管注,30%蔗糖脱水,oct包埋储存。
1.4细胞活性检测
mtt法又称mtt比色法,是一种检测细胞存活和生长的方法。其检测原理为活细胞线粒体中的琥珀酸脱氢酶能使外源性mtt还原为水不溶性的蓝紫色结晶甲瓒(formazan)并沉积在细胞中,而死细胞无此功能。二甲基亚砜(dmso)能溶解细胞中的甲瓒,用酶联免疫检测仪在540或720nm波长处测定其光吸收值,可间接反映活细胞数量。在一定细胞数范围内,mtt结晶形成的量与细胞数成正比。
1.5gfaas测铁含量
用pbs洗细胞三遍,洗净上清。ripa裂解细胞,12000rpm4度离心10min,取上清1-2ul用碧云天bca法试剂盒测定蛋白浓度。将裂解液充分混匀,每个样品取相同的体积到15ml离心管中,加入2倍体积的浓硝酸。用ddh2o将终体积补到3ml,沸水水浴消化过夜直至溶液澄清透明。gfaas先用铁标准品做标准曲线,然后测定样品中的铁含量。最后用蛋白浓度将铁含量normalize到单位蛋白浓度。
1.6calcein-am荧光染色测细胞内铁含量
a.细胞给予相应处理时间后,用预冷的pbs洗3遍,每遍5分钟
b.加入100ulpbs含有calcein终浓度为10um,闭光37度培养15分钟
c.用pbs洗三遍,最后用100ulpbs孵育
d.荧光显微镜下观察及拍照
1.7rna提取和realtime-pcr
trizol(lifetechnologies)法提取细胞和组织的rna,具体操作按说明书进行。nanodrop1000spectrophotometer上检测rna纯度(od260/od280≈1.9-2.1)及rna浓度(ng/μl),调整rna浓度到1μg/μl。2.0μgrna经dnase(promega)处理后,m-mlv反转录酶(promega)和oligo(dt)18primer(takarabioinc.)进行反转录。cfx96real-timesystem(bio-rad)中进行realtimepcr,检测体积为10ul,试剂采用iqsybrgreensupermix(bio-rad),引物序列如下:
kdm5cforwardprimer,5’-gaggtgaccctggatgagaa-3’,andkdm5creverseprimer,5’-caggagctgaggtctgaac-3’(steinetal.,2014);
fpn1forwardprimer5’-atgaccagggcgggagat-3’,andthereverseprimer5’-cccatccatctcggaaggta-3’(hudsonetal.,2010).
1.8aav-kdm5c-silence病毒包装
1.aav-kdm5c-silence病毒载体(paav[shrna]-egfp-u6>mkdm5c)直接购买于vectorbuildercyagenbiosciences.kdm5cshrna目标序列1491-1511bp;(agcaagctacccgggaatatactcgagtatattcccgggtagcttgct)
2.质粒扩增构建好的aav载体、包装质粒和辅助质粒需经过大量抽提,浓度大于1ug/ul,a260/280在1.7-1.8间方可用以包毒。
3.重组aav病毒的包装、鉴定、纯化和滴度测定
使用lipofectamine2000将raav-shrna-gfp、paav-rc和phelper三质粒共转染293t细胞,继续培养细胞72小时,然后收获细胞。反复冻融使细胞裂解,离心收集上清过滤得到病毒原液。同时使用paav-mcs、paav-rc和phelper三质粒共转染293t细胞包装空病毒品paav为实验阴性对照。使用氯仿-peg/nacl-氯仿法对重组病毒进行纯化。取纯化后的重组病毒和paav空病毒各200ul直接感染293t细胞,培养48-72小时后,荧光倒置显微镜观察绿色荧光蛋白gfp的表达情况。并通过实时定量pcr检测mrna表达量,并通过与空白对照组的比较计算抑制率。每个实验样品设置3个复孔,计算均值和标准差。
1.9westernblot蛋白检测
用含有pmsf(sigma-aldrich)和磷酸酶抑制剂(phosstop,roche)的ripa裂解液(碧云天生物技术研究所)裂解细胞或组织后,提取总蛋白。细胞蛋白取30ug,肝组织蛋白取100ug,经10%sds-page胶电泳后,转移至pvdf膜,该膜与特异性抗体4℃孵育过夜。洗涤三遍,与过氧化物酶标记的二抗(anti-rabbitoranti-mouseigg1:4000,proteintechgroup)室温孵育1小时后,westernblot试剂盒(eclsystem,pierce,thermoscientific)显色。
本实施例中所用一抗如下:
anti-h3k4me3(#9751,cellsignalingtechnology,1:1000),anti-h3k27me3(07-449,merckmillipore,1:1000),anti-kdm5c/smcx/jarid1c(ab190180,abcam1:1000),anti-ferritin-l(ab69090,abcam,1:1000),anti-transferrinreceptor1(13-6800,thermofisherscientific,1:1000),anti-ferroportin/slc40a1antibody(nbp1-21502,novusbiologicals,1:1000),anti-dmt1(ab123085,abcam,1:1000),anti-hepcidin(ab75883,abcam,1:1000),anti-bcl2(ab692,abcam,1:1000)andanti-cleavedcaspase-3(#9664,cellsignalingtechnology,1:1000)4c过夜.tbst洗3次,每次10分钟,然后孵育二抗(goatanti-rabbit或者anti-mouseirdye800cwsecondaryantibodies(1:5000,li-cor),室温1小时.tbst洗3次,每次10分钟。然后使用odysseyinfraredimagingsystem(li-cor)成像系统检测。多克隆兔抗anti-β-actin作为内参校准(浓度1:5000稀释)。
1.10免疫荧光染色步骤
细胞免疫荧光步骤:
1.细胞爬片;
2.4%多聚甲醛固定10min(固定细胞器用预冷的70%甲醇+30%丙酮);
3.pbs漂洗5min;
4.0.5%triton穿孔15min(丙酮固定法不用透化处理);
5.pbs漂洗2次,每次5min;
6.1%bsa封闭30min;
7.加入1%bsa稀释的一抗,于37℃杂交2h;
8.pbs漂洗2次,每次5min;
9.加入1%bsa稀释的二抗,于37℃杂交1h;
10.pbs漂洗2次,每次5min;
11.5ug/mldapi染色2min;
12.抗淬灭封片剂封片。
抗淬灭封片剂:
2.5%dabco(w/v)
50mmtris(ph8.0)
90%甘油
1.11统计方法
所用统计采用r软件分析,实验数据以mean±sd表示,实验的组间比较采用tukey’s检验(anova),两组间比较以student'st-test检验,以p<0.05认为有统计学意义。
2结果显示:
2.1筛选gsk-j4作为潜在铁鳌合剂小分子
为了筛选新型的铁螯合剂,在体外培养sh-sy5y神经瘤细胞,通过gfaas方法测量神经细胞系铁水平(如图1a所示),筛选发现,小分子gsk-j4能够显著降低神经细胞系sh-sy5y细胞系内铁水平,呈现出一定剂量依赖性。特别是,微量的gsk-j4(0.1μm)处理12小时后,能够显著降低细胞内铁水平(如图1b所示),每组重复3次。gsk-j4是一种新型的表观遗传修饰剂,化学结构如(如图1c所示),能够显著抑制组蛋白去甲基化酶的活性。
2.2gsk-j4显著降低神经细胞系内游离的自由铁水平
铁是细胞维持正常功能活动所必需的元素,但过多的铁又可损伤细胞。细胞铁水平受铁代谢相关蛋白(转铁蛋白-转铁蛋白受体,铁蛋白)的共同调节而维持平衡,其调节机制与胞浆铁调节蛋白与铁反应元件的结合有关。细胞内游离的铁是造成细胞氧化损伤的关键原因。虽然细胞内铁有不同的结合形式和不同的配体,但目前认为能够反映细胞铁状态和能够被铁调节蛋白感受的铁是胞浆中结合力较弱、分子量较小的那部分铁,即可变铁池(labileironpool,lip)。为进一步探究gsk-j4对细胞内游离铁的影响,采用钙荧光素(calcein)测定细胞lip的变化。calcein-am是一种荧光染料,可以主动穿过细胞膜,一旦进入细胞,乙酰甲基化酯被分解,calcein游离出来,脱酯的calcein滞留在细胞浆中,发出荧光,在细胞内,calcein可以与fe2+,3+结合,在生理ph条件下几乎不与其它金属离子结合,即使有很小量的结合,对添加铁螯合剂所增加的荧光几无影响,当铁与calcein结合后,荧光猝灭,铁和calcein的结合是可逆性的。在不同实验条件下,细胞内的lip水平升高时,由于铁与calcein结合,荧光减弱。而lip水平降低时,荧光就增加,因此,测定的荧光变化量可作为定量细胞lip的标志;
如(如图2a所示)铁补充剂fac(50μg/ml)和gsk-j4(0.5μm)处理sh-sy5y细胞12小时,结果显示铁补充剂fac能够显著抑制神经瘤细胞系sh-sy5y细胞内的calcein-am水平,意味着细胞内较多的游离铁lip,然而,gsk-j4能够显著增加神经瘤细胞系calcein-am水平,意味着细胞内较少的游离铁lip,并且铁补充剂fac能够一定程度抑制gsk-j4引起的calcein-am升高,进一步证实gsk-j4是通过影响细胞内游离铁水平lip,进而引起了calcein-am水平的改变,图2b显示了calcein-am定量分析结果;
进一步(如图2c所示),gsk-j4(0.5μm)以及铁螯合剂dfo(50μm)处理sh-sy5y细胞12h,蛋白检测分析铁储存蛋白ferritin-l(ftl)和铁摄取蛋白transferritinreceptor1(tfr1),间接反映细胞内游离铁水平的标志蛋白,结果如图2c所示,gsk-j4能够显著降低细胞内储铁蛋白ftl的水平,增加细胞的摄取铁蛋白(tfr1),这对细胞内铁代谢的影响类似与经典的铁鳌合剂dfo,意味着细胞处于较低的铁水平,但是gsk-j4的效率远远高于dfo,约为dfo浓度的1/100;图2d为蛋白定量分析结果;进一步,(如图2e、图2f所示)免疫荧光实验分析gsk-j4能够显著降低铁储存蛋白,由此推测,gsk-j4在神经瘤细胞系是一种高效的小分子铁螯合剂;
2.3gsk-j4对外周细胞系铁代谢无显著影响
为进一步确定gsk-j4对细胞铁水平的影响,本发明对比分析了gsk-j4(0.5μm)对三种细胞系:人神经瘤细胞系sh-sy5y,人源肾细胞系hek293以及人源肝癌细胞系hepg2细胞内铁水平的影响,与此前筛选结果一致,图3a显示gsk-j4(0.5μm)能够显著降低sh-sy5y细胞内的铁水平(p<0.01,与对照组相比),铁补充剂fac(50μg/ml)能够显著增加sh-sy5y细胞内铁水平,约为2.53倍(p<0.001),然而,gskj4(0.5μm)能够显著缓解这种铁蓄积(p<0.01);有趣的是,gsk-j4对人源肾细胞系hek293以及人源肝癌细胞系hepg2细胞内铁水平并没有显著影响,这意味着gsk-j4对细胞铁的影响具有细胞特异性,特别是gsk-j4能够显著影响神经细胞系的铁代谢。
为了进一步确定gsk-j4对细胞铁代谢的影响是否具有选择性,本发明进一步检测分析gsk-j4对人源肾细胞系hek293以及人源肝癌细胞系hepg2细胞内铁储存蛋白以及铁摄取蛋白的影响,与gsk-j4对铁水平结果一致,gsk-j4(0.5μm)在hek293和hepg2并没有引起铁储存蛋白ftl以及铁摄取蛋白tfr1的显著改变,这不同于经典的铁螯合剂dfo,dfo能够显著降低所有三种细胞系内的铁储存蛋白ftl以及增加细胞内铁的摄取蛋白(如图3b-3c和图3d-3e所示),因此,实验数据表明,gsk-j4能够选择性降低神经细胞系内的铁水平。
2.4gsk-j4对神经细胞系铁代谢相关蛋白的影响
为了进一步研究gsk-j4对神经细胞系sh-sy5y内铁代谢的影响,本发明检测了细胞内的铁摄取蛋白,铁排出蛋白,铁储存蛋白以及铁调节蛋白,结果显示,gsk-j4(0.5μm)以及铁补充剂fac(50μg/ml)处理人源神经瘤细胞系sh-sy5y细胞12h,如(如图4a-4b所示)蛋白检测分析gsk-j4显著增加铁摄入蛋白tfr1(p<0.01)和dmt1(p<0.05);如(如图4c-4d所示),gsk-j4显著增加铁排出蛋白fpn1(p<0.01);有趣的是,额外的铁补充剂fac(50μg/ml)能够显著抑制gsk-j4对铁储存蛋白ftl,铁摄入蛋白tfr1和dmt1的影响,但是并没有对fpn1产生显著改变,这意味着gsk-j4或许直接影响了铁的外排蛋白fpn1,进而对铁水平产生了影响;进一步分析铁代谢核心调节因子铁调素hepcidin,(如图4e-4f所示)gsk-j4(0.5μm)处理人源神经瘤细胞系sh-sy5y细胞12h,蛋白检测分析gsk-j4对铁调素hepcidin并无显著影响;此外,实时定量pcr检测发现,gsk-j4(0.5um,12hours)能够显著增加fpn1mrna的表达水平(如图4g所示),这进一步表明对铁外排蛋白fpn1是gsk-j4对神经细胞系sh-sy5y铁代谢影响的初始靶点。
2.5gsk-j4特异性影响神经细胞系组蛋白甲基化水平
gsk-j4是一种新型的组蛋白去甲基化酶抑制剂,特别是对组蛋白h3k4me3以及h3k27me3三甲基化水平的影响;蛋白检测分析发现(如图5a-5b所示)小分子gsk-j4(0.5μm)能够显著增加h3k27me3以及h3k4me3水平。并且,h3k27me3抑制剂gsk126(1μm)处理人源神经瘤细胞系sh-sy5y细胞12h,能够显著抑制h3k27me3三甲基化水平,但是并没有对铁代谢相关蛋白产生明显影响,而组蛋白甲基化广谱抑制剂dznep能够显著抑制h3k4me3的三甲基化水平,且显缓解了gsk-j4对铁相关蛋白的影响,这表明gsk-j4通过对组蛋白h3k4me3甲基化的影响对sh-sy5y细胞内的铁代谢产生了影响;此外,研究发现(如图5c所示)小分子gsk-j4(0.5μm)处理人源肾脏细胞系hek293细胞12h,蛋白检测分析gsk-j4对h3k27me3,h3k4me3的水平并没有影响,这进一步表明,gsk-j4对细胞铁代谢的特异性影响是通过特异性调节sh-sy5y细胞内组蛋白h3k4me3的三甲基化水平产生。
2.6组蛋白h3k4me3三甲基化参与调控铁外排蛋白fpn1表达
为了进一步证明组蛋白h3k4me3三甲基化水平对神经系sh-sy5y细胞系内铁代谢的影响,本发明通过抑制特异性升高h3k4me3水平的小分子cpi-455(1-5μm)处理人源神经瘤细胞系sh-sy5y细胞12h,(如图6a-6b所示)蛋白检测分析gsk-j4能够显著增加h3k4me3以及铁外排蛋白fpn1的影响,这进一步表明组蛋白h3k4me3的修饰参与到铁外排蛋白fpn1的调控。
2.7抑制kdm5c水平显著增加铁外排蛋白fpn1水平
基于目前已知,gsk-j4对组蛋白h3k4me3三甲基化的影响来源于对组蛋白kdm5的影响,特别是组蛋白去甲基化酶kdm5c的抑制,为了进一步探究,组蛋白h3k4me3三甲基化是否参与到铁蛋白,特别是铁外排蛋白fpn1的调节,本发明构建腺相关病毒aav-kdm5c-silence病毒,如(如图7a所示)感染大鼠原代皮层神经元细胞72h,(如图7b所示)荧光定量pcr检测分析aav-kdm5csilence病毒显著抑制kdm5cmrna的水平,并且蛋白检测分析证实(如图7c-7d所示)腺相关病毒aav-kdm5-silence病毒能够显著抑制kdm5c的蛋白水平;进一步,实验结果显示,腺相关aav-kdm5c病毒能够显著增加铁外排蛋白fpn1和h3k4me3的蛋白水平;基于此,抑制kdm5c增加h3k4me3水平,能够显著增加铁外排蛋白fpn1水平;
基于此,gsk-j4是一种能够特异性调节神经铁代谢的小分子。
2.8gsk-j4在mptp诱导的帕金森疾病模型保护作用
gsk-j4已被报道能够显著通过血脑屏障。基于铁蓄积是引起神经退行性疾病的重要原因之一,特别是在帕金森疾病中,铁鳌合剂被认为一个潜在的有效手段;本发明进行了体外细胞实验,(如图8a-8c所示)发现gsk-j4能够显著抑制神经毒素mpp+引起的细胞毒性;进一步,通过腹腔注射gsk-j4,结合mptp诱导的小鼠帕金模型(如图9a所示),发现gsk-j4能够显著改善mptp帕金森疾病小鼠的运动缺陷(如图9b-9d所示),并且能够缓解mptp引起的黒质及纹状体内的多巴胺能神经元的丢失(如图9e-g所示)。
本发明的实验结果表明,gsk-j4是一种能够特异性调节神经铁代谢的小分子,可进一步制备特异性调节神经铁代谢的药物,并且能够有效缓解脑铁蓄积的相关疾病,如帕金森疾病等。
- 还没有人留言评论。精彩留言会获得点赞!