工程细胞和使用方法与流程
相关申请的交叉引用
本申请要求2017年2月2日提交的美国临时专利申请序列第62/453922号和2017年10月30日提交的美国临时专利申请序列第62/578721号的优先权权益,其全部内容通过引用并入本文。
政府权利
本发明部分地在美国国立卫生研究院(nih)授予的授权号gm093282的政府支持下完成。美国政府拥有本发明的特定权利。
本公开涉及工程细胞和经修饰的细胞,特别是在医学领域。
背景技术:
本文中的所有出版物均以引用并入,其程度如同具体和单独地指出每个单独的出版物或专利申请通过引用并入。以下描述包括可用于理解本发明的信息。本文提供的任何信息均不承认为是现有技术或与当前要求保护的发明相关,或者具体或暗含地引用的任何出版物是现有技术。
癌症是美国和全世界的死亡的主要原因。最近,除了用于治疗癌症的手术、放射和化疗之外,免疫治疗已成为第四种武器。癌症免疫治疗旨在利用身体自身的免疫系统来对抗癌症。虽然已经在过继细胞治疗(act)和检查点阻断治疗方面取得了良好的效果,但输注到患者体内的t细胞的抗癌特性仍有待改善,包括其迁移潜力和抵抗肿瘤微环境中免疫抑制信号的能力。自然杀伤(nk)细胞在对抗恶性细胞的先天免疫中起关键作用,并且还被开发为癌症免疫治疗中有效的癌症杀手。然而,基于nk细胞的治疗的一个挑战是难以从患者的血液中获得足够数量的活性nk细胞。nk-92是从具有克隆性nk细胞淋巴瘤的患者建立的高细胞毒性自然杀伤(nk)细胞系,其可用于以gmp级产生更多数量的细胞毒性nk细胞。特异性靶向是良好癌症治疗的黄金标准。然而,nk-92细胞不表达靶向特定细胞的用于adcc作用的fc受体,这限制了它的广泛应用。
同样,树突状细胞(dc)长期以来一直被认为是癌症免疫治疗的核心参与者。dc的迁移能力对基于dc的免疫治疗的结果具有深远的影响。然而,显示只有一小部分(1%至2%)总施用的dc到达次级淋巴器官以活化t细胞,这限制了该疫苗接种方法的实际应用。
因此,本领域仍然需要能够有效地将抗原特异性抗体标记在细胞表面上,这反过来又引导这些细胞消灭患病细胞中的特异性抗原表达并有效地治疗疾病的新组合物和方法。
技术实现要素:
本文公开的各种实施方案包括工程细胞,其包含与细胞表面上存在的细胞表面聚糖共价结合的化学或生物部分,其中化学或生物部分选自小分子、多核苷酸、多肽和抗体。在一个实施方案中,工程细胞是免疫细胞。在一个实施方案中,工程细胞是t细胞或自然杀伤(nk)细胞。在一个实施方案中,t细胞是cd8+t细胞或cd4+t细胞。在一个实施方案中,细胞是树突状细胞(dc)。在一个实施方案中,小分子是小药物分子或其药学上可接受的盐或共晶体。在一个实施方案中,抗体是单链可变片段(scfv)、片段抗原结合(fab)片段或全长抗体。在一个实施方案中,抗体是免疫球蛋白g(igg)抗体。在一个实施方案中,igg是全长igg。在一个实施方案中,与工程细胞连接的化部分学或生物部分是生物标志物和/或探针。在一个实施方案中,化学部分或生物部分是生物素探针、荧光探针、生物正交反应柄(biorthogonalreactionhandle)和/或染料标记的单链dna。在一个实施方案中,染料是fam。在一个实施方案中,荧光探针是cy3。在一个实施方案中,生物正交反应柄是四嗪。在一个实施方案中,岩藻糖衍生物是gdp-岩藻糖。在一个实施方案中,工程细胞是嵌合抗原受体(car)-t细胞。在一个实施方案中,car-t细胞包含基因修饰的t细胞,其细胞表面glcnac与携带新基序的gdp-岩藻糖共价结合。在一个实施方案中,car包含三个结构域:scfv、fab和/或与同源受体结合的成熟配体。在一个实施方案中,细胞表面聚糖是n-乙酰葡糖胺(glcnac)。在一个实施方案中,化学部分或生物部分通过岩藻糖衍生物与glcnac共价结合。在一个实施方案中,细胞表面聚糖是唾液酸(neuac)。
本文公开的各种实施方案还包括组合物,其包含:抗体-细胞缀合物,其中一种或多于一种抗体与细胞表面上的一个或多于一个聚糖部分共价结合。在一个实施方案中,细胞是免疫细胞。在一个实施方案中,细胞是原代人t细胞、自然杀伤(nk)细胞、cd4+细胞和/或原代cd8+ot-1t细胞。在一个实施方案中,nk细胞是nk-92mi细胞。在一个实施方案中,细胞是树突状细胞(dc)。在一个实施方案中,抗体是曲妥珠单抗。在一个实施方案中,细胞是nk-92mi,并且至少一种抗体是曲妥珠单抗。在一个实施方案中,多于一种类型的抗体缀合在细胞表面上。在一个实施方案中,聚糖部分是n-乙酰葡糖胺(glcnac)。在一个实施方案中,抗体通过岩藻糖衍生物与glcnac共价结合。在一个实施方案中,聚糖部分是唾液酸(neuac)。在一个实施方案中,组合物是药物组合物。在一个实施方案中,药物组合物还包含药学上可接受的载体或赋形剂。在一个实施方案中,药物组合物用于治疗对象的疾病。在一个实施方案中,抗体-细胞缀合物能够牢固地结合在炎症位点上(抗e-选择素)、靶向特异性癌细胞(抗her2)、和/或阻断免疫检查点(抗pd-l1)。
本文公开的各种实施方案还包括治疗对象的疾病的方法,其包括:提供包含工程细胞和药学上可接受的载体的药物组合物;和通过向对象施用治疗有效剂量的药物组合物来治疗疾病,其中工程细胞包含与其表面上存在的聚糖部分共价结合的化学部分或生物部分,并且其中化学部分或生物部分选自小分子、多核苷酸、多肽和抗体。在一个实施方案中,抗体是免疫球蛋白g(igg)抗体。在一个实施方案中,抗体是抗pd-l1抗体。在一个实施方案中,抗体是抗e-选择素抗体。在一个实施方案中,抗体是抗her2抗体。在一个实施方案中,抗体是曲妥珠单抗。在一个实施方案中,多于一种化学部分或生物部分与细胞表面共价结合以同时靶向多于一种疾病。在一个实施方案中,细胞是树突状细胞。在一个实施方案中,细胞是免疫细胞。在一个实施方案中,细胞是原代人t细胞、自然杀伤(nk)细胞、cd4+细胞和/或原代cd8+ot-1t细胞。在一个实施方案中,nk细胞是nk-92mi细胞。在一个实施方案中,聚糖部分是n-乙酰葡糖胺(glcnac)。在一个实施方案中,化学部分或生物部分通过岩藻糖衍生物与glcnac共价结合。在一个实施方案中,岩藻糖衍生物包含岩藻糖-炔烃。在一个实施方案中,聚糖部分是唾液酸(neuac)。在一个实施方案中,疾病是癌症。在一个实施方案中,治疗疾病包括减少对象的癌性肿瘤的大小。在一个实施方案中,癌症是乳腺癌。
本公开的实施方案还包括治疗、减少、抑制或减轻对象的癌症的方法,其包括:向对象施用治疗有效剂量的药物组合物,所述药物组合物包含工程细胞,该工程细胞具有与其表面上的聚糖部分共价结合的抗体。在一个实施方案中,癌症是乳腺癌。在一个实施方案中,抗体是曲妥珠单抗。在一个实施方案中,细胞是nk-92mi。在一个实施方案中,多于一种抗体缀合在细胞表面上以同时靶向多于一种癌症。
本公开的实施方案还包括制备工程细胞的方法,其包括:制备包含与岩藻糖或gdp-岩藻糖共价结合的化学部分或生物部分的岩藻糖衍生物或gdp-岩藻糖衍生物;通过将细胞与包含(a)岩藻糖衍生物和/或gdp-岩藻糖衍生物和(b)岩藻糖基转移酶的组合物一起孵育来制备工程细胞。在另一个实施方案中,本文公开了制备工程细胞的方法,其包括:通过使携带tco部分的抗体与cs-az-tz或cs-poc-tz反应来生成与抗体缀合的cmp-唾液酸(cs-igg);和通过将天然存在的细胞与包含(a)cs-igg和(b)唾液酸转移酶的组合物一起孵育来制备工程细胞。在一个实施方案中,岩藻糖基转移酶是α-1,3-岩藻糖基转移酶。在一个实施方案中,α-1,3-岩藻糖基转移酶是幽门螺杆菌(h.pylori)α-1,3-岩藻糖基转移酶。在一个实施方案中,重组制备α-1,3-岩藻糖基转移酶。在一个实施方案中,唾液酸转移酶选自st6gal1、多杀巴斯德氏菌(pasteurellamultocida)α(2,3)唾液酸转移酶m144d突变体(pm2,3st-m144d)和美人鱼发光杆菌(photobacteriumdamsel)α(2,6)唾液酸转移酶(pd2,6st)。在一个实施方案中,化学部分或生物部分选自小药物分子、生物分子、探针分子、荧光团、多核苷酸、多肽和完整igg、或其组合。在一个实施方案中,生物分子是曲妥珠单抗。在一个实施方案中,细胞是免疫细胞。在一个实施方案中,细胞是原代人t细胞或自然杀伤(nk)细胞。在一个实施方案中,nk细胞是nk-92mi细胞。在一个实施方案中,t细胞是cd4+细胞或cd8+t细胞。在一个实施方案中,化学部分或生物部分通过岩藻糖衍生物与细胞表面上的n-聚糖通用单元lacnac共价连接。在一个实施方案中,岩藻糖修饰的生物分子和/或gdp-岩藻糖修饰的生物分子包括岩藻糖-炔烃和/或gdp-岩藻糖叠氮化物。在一个实施方案中,使用配体加速和生物相容的铜(i)催化的炔烃-叠氮化物环加成(cuaac)反应,用化学部分或生物部分进一步修饰生物分子-细胞缀合物。
本文公开的各种实施方案还包括制备适于将分子连接在工程细胞表面上的工程细胞的一锅法,其包括:通过将岩藻糖类似物与包含atp、gtp、l-岩藻糖激酶/gdp-岩藻糖焦磷酸化酶(fkp)、mg2+或mn2+、和无机焦磷酸酶(pp酶)的混合物组合来制备gdp-岩藻糖类似物;和通过将天然存在的细胞添加到包含来自步骤(a)的粗产物和幽门螺杆菌α-1,3-岩藻糖基转移酶的组合物中来制备工程细胞。在一个实施方案中,本文公开了制备适于将分子连接在工程细胞表面上的工程细胞的一锅一步法,其包括:将细胞与包含岩藻糖类似物、atp、gtp、l-岩藻糖激酶/gdp-岩藻糖焦磷酸化酶(fkp)、mg2+或mn2+、无机焦磷酸酶(pp酶)和幽门螺杆菌α-1,3-岩藻糖基转移酶的组合物一起孵育。在一个实施方案中,岩藻糖类似物包含与化学部分或生物部分缀合的岩藻糖。在一个实施方案中,岩藻糖类似物包含岩藻糖-炔烃。在一个实施方案中,工程细胞包含与细胞表面上的聚糖连接的化学部分或生物部分。在一个实施方案中,来自步骤(a)的产物通过铜催化的叠氮化物-炔烃环加成(cuacc)反应被进一步修饰,以产生与小分子、多核苷酸、多肽和/或抗体缀合的gdp-岩藻糖类似物。
本公开的其他实施方案包括将细胞表面lacnac或slacnac转化为lex或slex的一锅原位岩藻糖基化策略,其包括:通过将岩藻糖类似物与包含atp、gtp、fkp、mg2+或mn2+和无机焦磷酸酶(pp酶)的混合物组合来制备gdp-岩藻糖类似物;和通过将天然存在的细胞加入到包含来自步骤(a)的粗产物和幽门螺杆菌α-1,3-岩藻糖基转移酶的组合物中,将细胞表面lacnac或slacnac转化为lex或slex。
本文公开的各种实施方案还包括试剂盒,其包含:gdp-岩藻糖衍生物和幽门螺杆菌α-1,3-岩藻糖基转移酶。
通过以下结合附图的详细描述,本发明的其他特征和优点将变得明显,附图通过举例的方式示出了本发明的各种实施方案。
附图说明
示例性实施方案在参考图中示出。本文公开的实施方案和附图旨在被认为是说明性的而非限制性的。
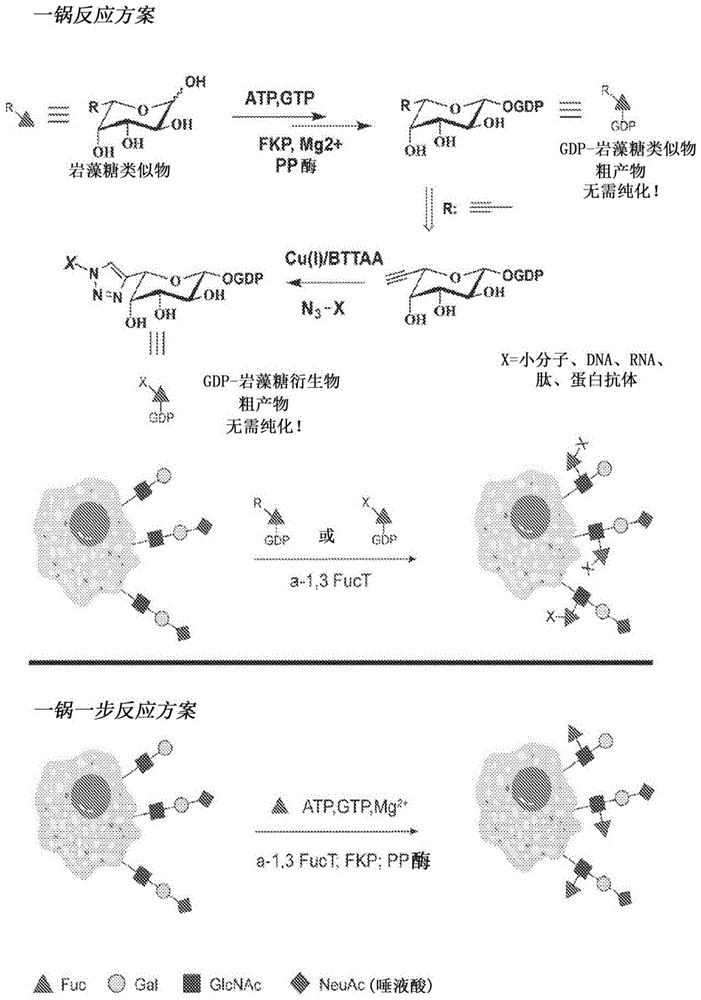
图1描绘了根据本文的实施方案,原位一锅法岩藻糖基化试剂制备和细胞表面反应方案。
图2描绘了根据本文的实施方案,使用粗制一锅法gdp-岩藻糖对培养的中国仓鼠卵巢(cho)细胞进行原位一锅岩藻糖基化。
图3描绘了根据本文的实施方案,使用粗制的一锅法gdp-岩藻糖-生物素修饰培养的lec2cho细胞表面。
图4描绘了根据本文的实施方案,使用粗制的一锅法gdp-岩藻糖-cy3修饰培养的lec2cho细胞表面。
图5描绘了根据本文的实施方案,使用粗制的一锅法gdp-岩藻糖-四嗪修饰培养的lec2cho细胞表面。
图6描绘了根据本文的实施方案,使用粗制的一锅法gdp-岩藻糖-bsa修饰培养的lec2cho细胞表面。
图7描绘了根据本文的实施方案,使用粗制的一锅法gdp-岩藻糖-抗体修饰培养的lec2cho细胞表面。
图8描绘了根据本文的实施方案,使用粗制一锅法gdp-岩藻糖-tz和随后的tco-抗体修饰培养的lec2cho细胞表面。
图9描绘了根据本文的实施方案,使用粗制的一锅法gdp-岩藻糖-生物素修饰培养的野生型cho细胞表面。
图10描绘了根据本文的实施方案,使用粗制的一锅法gdp-岩藻糖-生物素修饰活化的小鼠cd8+细胞表面。
图11描绘了根据本文的实施方案,使用粗制的一锅法gdp-岩藻糖-生物素修饰小鼠树突状细胞(dc)表面。
图12描绘了根据本文的实施方案,使用粗制的一锅法gdp-岩藻糖-生物素修饰初始或活化的人cd8+t细胞表面。
图13描绘了根据本文的实施方案,使用粗制的一锅法gdp-岩藻糖-tz修饰活化的小鼠cd8+t细胞表面,然后与tco-小鼠igg缀合。
图14描绘了根据本文的实施方案,使用粗制的一锅法gdp-岩藻糖-tz修饰活化的小鼠cd8+t细胞表面,然后与tco-小鼠igg和tco-大鼠igg一起缀合。
图15描绘了根据本文的实施方案,在ot-1t细胞表面上配置抗pd-l1抗体。
图16描绘了根据本文的实施方案,在ot-1t细胞表面上配置抗pd-l1抗体促进了过继细胞治疗(act)。
图17描绘了根据本文的实施方案,体内扩增的小鼠cd8+t细胞的原位一锅法岩藻糖基化增加了e-选择素结合并抑制了gal-1结合。
图18描绘了根据本文的实施方案,诱导的小鼠调节性t细胞(itreg)的原位一锅法岩藻糖基化增加了e-选择素结合。
图19描绘了根据本文的实施方案,人t细胞的原位一锅法岩藻糖基化增加了e-选择素结合。
图20描绘了根据本文的实施方案,人天然调节性t(ntreg)细胞的原位一锅法岩藻糖基化增加了e-选择素结合并抑制了gal-1结合。
图21描绘了根据本文的实施方案,her2特异性人car-t细胞的原位一锅法岩藻糖基化增加了e-选择素结合并抑制了gal-1结合。
图22描绘了根据本文的实施方案,体外分化的小鼠cd8t细胞的原位一锅法岩藻糖基化显著增强了e-选择素结合并降低了gal-1结合。
图23描绘了根据本文的实施方案,体外分化的小鼠ot-1cd8t细胞的原位一锅法岩藻糖基化对细胞表面标志物的表达几乎没有影响。
图24描绘了根据本文的实施方案,用体外分化的聚糖修饰的ot-1t细胞处理b16-ova包埋小鼠显著降低了肿瘤生长。
图25描绘了根据本文的实施方案,用体内分化的聚糖修饰的ot-1t细胞处理b16-ova包埋小鼠显著地降低了肿瘤生长并延长肿瘤包埋小鼠的寿命。
图26描绘了根据本文的实施方案,体外分化的小鼠树突状细胞(dc)的原位一锅法岩藻糖基化显著增强了e-选择素结合并降低了半乳凝素-1结合。
图27描绘了根据本文的实施方案,负载抗原的dc的岩藻糖基化诱导了显著更强的抗肿瘤免疫应答。
图28描绘了根据本文的实施方案,使用粗制的一锅法gdp-岩藻糖-dna-fam修饰培养的lec2cho细胞表面。
图29描绘了根据本文的实施方案,配置在培养的lec2cho细胞表面上的抗人e-选择素可以结合人e-选择素。
图30描绘了根据本文的实施方案,配置在培养的lec2cho细胞表面上的抗小鼠pd-l1可以结合小鼠pd-l1。
图31描绘了根据本文的实施方案,配置在培养的cho细胞表面上的抗小鼠ctla4可以结合小鼠ctla4蛋白。
图32描绘了根据本文的实施方案,配置在小鼠树突状细胞表面上的抗小鼠ctla4可以结合小鼠ctla4蛋白。
图33描绘了根据本文的实施方案,配置在ot-1cd8t细胞表面上的小鼠igg可以在表面上停留多于24小时。
图34描绘了根据本文的实施方案,ot-1cd8t细胞与小鼠igg抗体缀合不影响经修饰的细胞的增殖速率。
图35描绘了根据本文的实施方案,配置在ot-1cd8t细胞表面上的抗小鼠pd-l1可以在细胞表面上停留多于24小时。
图36描绘了根据本文的实施方案,ot-1cd8t细胞与抗pd-l1抗体缀合不影响经修饰的细胞的增殖速率。
图37描绘了根据本文的实施方案,配置在ot-1cd8t细胞表面上的抗小鼠pd-l1可以结合小鼠pd-l1。
图38描绘了根据本文的实施方案,配置在ot-1cd8t细胞表面上的抗小鼠pd-l1可以增强t细胞对癌细胞的杀伤功能。
图39描绘了根据本文的实施方案,基于一步法酶促岩藻糖基化的细胞表面工程策略。(a)两种常规使用的细胞表面工程方法。代谢工程用于将反应柄(x)配置到细胞表面上,它可以与目标分子上的互补柄(y)反应。基因工程强大且稳健,能够在细胞表面配置功能性分子和反应柄。这两种方法通常具有多个步骤,并持续数天。(b)酶促糖工程方法,其能够一步将多种功能性分子转移至细胞表面。通过来自幽门螺杆菌的α1,3fuct实现细胞表面lacnac/唾液酸lacnac和gdp-fuc衍生物之间的反应,使得可以在岩藻糖的c6位置处耐受与完整igg一样大的修饰。(c)描述了gf-al和gf-az衍生物的一锅合成方案。显示的功能性(z)包括生物正交柄(四嗪,tz)、生物物理探针(生物素,cy3)和生物材料(ssdna)。
图40描绘了根据本文的实施方案,igg向lec2cho细胞的细胞表面的酶促转移。(a)gdp-fuc修饰的igg(gf-igg)合成的示意图。显示了生物正交柄和接头的化学结构。(b)使用fuct将gf-igg转移至cholec2细胞表面的方案。(c)用酶ft、底物gf-rigg或两者处理的lec2细胞的流式细胞术分析。(d)在酶促转移中gf-rigg浓度(0.005mg/ml至0.2mg/ml)的滴定。每个反应使用相同量的ft并在室温下放置30分钟。平均值±sd(误差条),来自三个独立实验的代表性图。(e)gf-rigg在冰上酶促转移至lec2细胞的时间过程。在37℃下的反应是最大标记对照。(f)igg标记之前和之后lec2细胞活力的流式细胞术分析。(g)在酶促igg转移中将没有lacnac表达的cholec8细胞作为阴性对照与lec2细胞进行比较。平均值±sd(误差条),来自三个独立实验的代表性图。(h)当与alexafluor647标记的gf-rigg一起孵育时,使用或不使用ft处理的lec2细胞的共聚焦显微镜图像。细胞核用hoechst33342染色。比例尺:2μm。(i)同时用rigg和migg标记的lec2细胞的流式细胞术分析。
图41描绘了根据本文的实施方案,α-e-选择素酶促转移至原代人t细胞。(a)修饰的(1)和天然的(2)人t细胞在huvec细胞上结合和跨膜迁移过程的示意图。(b)ft介导的来自三个不同供体的原代人t细胞上的gf-α-he-sel标记。平均值±sd(误差条),来自三个独立实验的代表性图。(c)人e-选择素-fc嵌合体在未标记的t细胞和经migg或α-he-sel标记的t细胞上的结合。平均值±sd(误差条)。(d)在剪切应力条件下对人e-选择素-fc包被的载玻片的流动室测定分析。将用migg或α-he-sel标记的t细胞与未标记的t细胞进行比较。在imagej中定量细胞数(细胞/mm2)。平均值±sd(误差条)。(e、f)结合huvec的人t细胞的定量分析和荧光显微镜图像。用cfse(绿色)对人t细胞进行染色,用hoechst33342(蓝色)和did(红色)对huvec细胞进行染色。如果需要,用tnf-α预处理huvec。平均值±sd(误差条),来自三个独立实验的代表性图。比例尺:50μm。(g)huvec的跨膜迁移测定分析。通过荧光信号定量底层中迁移的t细胞(leukotracker,绿色)。在所有图中:ns,p>0.05;*p<0.05;**p<0.01;****p<0.0001。
图42描绘了根据本文的实施方案,用于her2+癌症治疗的赫赛汀和nk-92mi细胞缀合物的构建。(a)nk-92mi-赫赛汀缀合物可特异性结合her2+癌细胞,因此由于接近而具有增强的杀伤效果。(b)分析特异性her2抗原与nk-92mi-赫赛汀缀合物的结合。平均值±sd(误差条)。(c、d)nk-92mi-赫赛汀和bt474(her2+)之间特异性结合的流式细胞术分析和荧光显微镜图像。用celltrackerorange(红色)对nk-92mi细胞进行染色,用celltrackergreen(绿色)对bt474细胞进行染色。显示的是荧光和相差的合并通道。绿色区域是bt474细胞的簇。比例尺:50μm。(e)定量细胞介导的nk-92mi细胞对bt474细胞的细胞毒作用的ldh释放测定。nk-92mi-赫赛汀与亲本nk-92mi进行比较,其中亲本nk-92mi具有或不具有额外添加的游离赫赛汀(5μg/ml)。nk-92mi-higg用作阴性对照。平均值±sd(误差条),来自三个独立实验的代表性图。(f)nk-92mi和nk-92mi-赫赛汀在杀伤具有或不具有her2表达的不同癌细胞系中的比较。平均值±sd(误差条),来自三个独立实验的代表性图。(g)nk-92mi-赫赛汀在不同的用于酶促转移的gf-赫赛汀浓度下的杀灭活性。平均值±sd(误差条)。(h)nk-92mi和nk-92mi-赫赛汀在不同效应物与靶细胞比例下杀伤bt474的比较。平均值±sd(误差条)。(i)将赫赛汀和α-egfr双重标记的nk-92mi细胞与nk-92mi-赫赛汀以及nk-92mi-α-egfr在杀伤her2+/egfr+skov3癌细胞方面进行比较。平均值±sd(误差条)。(j)未照射和经照射的(6gy)nk-92mi细胞杀伤bt474细胞的比较。(k)nk-92mi-赫赛汀的体内抗肿瘤活性。给nsg小鼠静脉注射50万个mda-mb-435/her2+/f-luc细胞。然后,在肿瘤细胞注射后第1天通过静脉注射500万个nk-92mi或nk-92mi-赫赛汀细胞一次处理动物。对照小鼠接受hbss。肿瘤攻击后六天,向小鼠腹腔注射d-荧光素,通过ivis系统进行成像。显示个体小鼠的肿瘤大小和平均值±sd;n=10。还显示了代表性图。在所有图中:ns,p>0.05;*p<0.05;**p<0.01;***p<0.001;****p<0.0001。
图43描绘了根据本文的实施方案,将α-pd-l1酶促转移至ot-1t细胞以增强t细胞在特异性杀伤方面的活化。(a)ot-1t细胞(tcr-t)和b16-ova黑素瘤细胞之间相互作用的方案。b16-ova上的mhc复合体将ova抗原呈递给ot-1t细胞上的特异性tcr以诱导活化,而b16-ova上的pd-l1与ot-1t细胞上的pd-1相互作用抑制活化信号。(b)通过ot-1t细胞表面上结合的α-pd-l1阻断pd-1/pd-l1途径的方案。原位阻断可以增强t细胞的活化和对癌细胞的杀伤。(c)在不同处理下对ot-1t细胞上的pd-l1抗原结合的分析。平均值±sd(误差条)。(d)分析pd-l1抗原在酶促转移中不同gf-α-pd-l1浓度下的结合。平均值±sd(误差条)。(e)定量细胞介导的ot-1t细胞-α-pd-l1缀合物对b16-ova细胞的细胞毒作用的测定。平均值±sd(误差条),来自三个独立实验的代表性图。与rigg缀合的ot-1t细胞和与α-pd-l1缀合的p14t细胞显示为阴性对照。(f)ot-1t细胞和ot-1t细胞-α-pd-l1在不同的效应物与靶细胞比例下杀伤b16-ova的比较。平均值±sd(误差条)。(g)用于酶促转移的不同gf-α-pd-l1浓度的ot-1t细胞和ot-1t细胞-α-pd-l1的杀伤活性。平均值±sd(误差条)。(h)在不同处理下与b16-ova混合的ot-1t细胞的ifn-γelisa。与α-pd-l1缀合的p14t细胞显示为阴性对照。仅b16-ova是背景。平均值±sd(误差条),来自三个独立实验的代表性图。(i)在有或没有α-pd-l1标记的情况下杀伤b16-ova的ot-1t细胞的显微镜图像。蓝色箭头表示较少的癌细胞,紫色箭头表示较大的t细胞簇。比例尺:50μm。(j)通过cfse稀释由b16-ova介导的活化后ot-1t细胞增殖的分析。平均值±sd(误差条)。(k)比较用或不用α-pd-l1标记的血液、引流淋巴结和肿瘤中的t细胞频率的竞争测定。用α-pd-l1标记具有cd45.1或thy1.1同类系标志物的ot-1t细胞,随后与未标记的具有另一种同类系标志物的ot-1t细胞混合。ot-1t细胞混合物被静脉注射到b16-ova攻击的小鼠中并在扩增48小时后进行分析。显示了个体小鼠的相对比例和平均值±sd;n=3。在所有图中:ns,p>0.05;**p<0.01;***p<0.001;****p<0.0001。
图44描绘了根据本文的实施方案,一步法酶促标记比两步法酶促标记更有效并且更加生物相容。(a)一步法和两步法标记系统方案。在一步法标记系统中,使用α1,3-fuct将gf-al-生物素的一锅法产物直接转移至lec2细胞表面上的lacnac表位。相反,两步法标记涉及第一步中gf-a1的酶促转移和随后步骤中表面炔烃和叠氮化物-生物素探针之间的cuaac反应。(b)在一步法或两步法标记过程中的生物相容性的流式细胞术分析。反应后的lec2细胞用dapi染色。(c)在(a)中描述的一步法和两步法标记方法的效率比较。细胞用链霉抗生物素蛋白-apc染色。
图45描绘了根据本文实施方案的gdp-fuc衍生物的化学结构,其包括gf-al-生物素、gf-al-cy3、gf-al-tz和gf-az-tz。
图46描绘了根据本文的实施方案,将一锅法gdp-fuc衍生物一步酶促转移至lec2细胞。(a)使用α1,3-fuct将一锅法gdp-fuc衍生物转移至lec2cho细胞表面上的lacnac表位的方案。(b)lec2细胞用ft和gf-al-cy3、仅ft或仅gf-al-cy3处理,或未处理。然后通过流式细胞术分析细胞,仅当用ft和gf-al-cy3两者处理时,显示成功的标记。(c)lec2细胞用ft和gf-az-tz、仅ft或仅gf-az-tz处理,或未处理。然后将细胞与tco-cy5反应并通过流式细胞术分析。仅在用ft和gf-az-tz处理的组中显示清晰标记。(d)在ft介导的lec2细胞上的转移反应中比较gf-al-tz和gf-az-tz。酶促反应后细胞与tco-cy5反应,并通过流式细胞术分析。gf-az-tz组显示出比gf-al-tz更强的信号。(e)lec2细胞用ft和gf-az-ssdna-fam,或仅gf-az-ssdna-fam处理,或未处理。然后通过流式细胞术分析细胞。用ft和gf-az-ssdna-fam处理的组具有最强的fam信号。
图47描绘了根据本文的实施方案,在α1,3-fuct的活性位点中的gdp-岩藻糖(a)和gf-az-tz(b)的分子模型。图像来自先前pymol中报道的hp-fuct和gdp-岩藻糖复合物的晶体结构(pdb:2nzy)。
图48描绘了根据本文的实施方案,lacnac表位的阻断抑制了ft介导的gf-rigg向lec2细胞表面的转移。lec2细胞首先用ft和gdp-fuc处理,这可能占据几乎所有的lacnac表位。之后,用ft和gf-rigg处理经封闭的细胞。通过流式细胞术分析rigg标记。与直接gf-rigg转移相比,阻断的lec2具有显著降低的rigg信号。误差条为平均值±sd。****p<0.0001。
图49描绘了根据本文的实施方案,lec2细胞上两种不同igg分子的双重标记。用gf-migg、gf-rigg和ft处理lec2细胞。经gf-migg和gf-rigg处理的显示为阴性对照。标记的细胞用抗migg和抗rigg荧光抗体染色,并通过流式细胞术分析。
图50描绘了根据本文的实施方案,migg标记之前和之后的人t细胞的表型。(a)人pbmc被抗cd3/cd28抗体活化并在体外扩增约两周。用抗cd3、抗cd45、抗cd4和抗cd8荧光抗体对细胞进行染色,并通过流式细胞术分析。(b)用gf-igg和ft处理活化的人t细胞。标记的或未标记的细胞用抗cd4、抗cd8、抗cd25、抗cd45ro、抗cd44和抗cd62l荧光抗体染色,并通过流式细胞术分析。初始t细胞用作对照。
图51描绘了根据本文的实施方案,将igg分子转移到人t细胞上。(a)将人pbmc上的lacnac在活化后的不同天数用gf-生物素标记。标记的细胞用链霉抗生物素蛋白-apc、抗cd4和抗cd8荧光抗体染色,并通过流式细胞术分析。(b)人t细胞用ft和gf-α-he-sel,或单独的gf-α-he-sel处理,或未处理。用抗migg荧光抗体对细胞进行染色,并通过流式细胞术分析。(c)人t细胞用ft和gf-migg或单独的gf-migg处理,或未处理。用抗migg荧光抗体对细胞进行染色,并通过流式细胞术分析。(d)用抗migg荧光抗体对用α-he-sel标记的人t细胞进行染色,并在标记后的不同时间点通过流式细胞术分析。(e)在t细胞培养基中培养具有或不具有migg标记的人t细胞,并在标记后的不同天数对每组中的活细胞进行计数。(f)将具有不同修饰的人t细胞与e-选择素-fc蛋白一起孵育,然后用抗hfc荧光抗体染色用于facs分析。误差条为平均值±sd。在所有图中:****p<0.0001。
图52描绘了根据本文的实施方案,将igg分子转移到nk-92mi细胞上。(a)nk-92mi细胞用gf-higg和ft,或仅gf-higg处理,或未经处理。用抗hfc荧光抗体对细胞进行染色并通过流式细胞术分析。(b)nk-92mi细胞用gf-赫赛汀和ft或单独的gf-赫赛汀处理,或未经处理。用抗hfc荧光抗体对细胞进行染色并通过流式细胞术分析。(c)与赫赛汀缀合的nk-92mi细胞用抗hfc荧光抗体染色,并在标记后的不同时间点通过流式细胞术分析。(d)然后将具有不同修饰的nk-92mi细胞与her2-his6蛋白一起孵育,然后用抗his6荧光抗体染色用于facs分析。(e)与赫赛汀缀合或不与赫赛汀缀合的nk-92mi细胞与bt474以5:1(效应物:靶标)的比例共培养4小时。通过elisa分析培养上清液中的颗粒酶b分泌。误差条为平均值±sd。在所有图中:ns,p>0.05;****p<0.0001。
图53描绘了根据本文的实施方案,酶促转移中滴定的gf-赫赛汀浓度。在标准标记条件下用不同浓度的gf-赫赛汀处理nk-92mi细胞。在流式细胞术分析前,标记的细胞用抗hfc荧光抗体直接染色(a),或用her2-his6孵育,然后用抗his6荧光抗体染色(b)。
图54描绘了根据本文的实施方案,在nk-92mi细胞上的双重抗体标记。nk-92mi细胞用ft和gf-赫赛汀处理,然后用ft和gf-α-egfr处理。在流式细胞术分析前,标记的细胞用抗hfc荧光抗体染色(a),或用抗migg荧光抗体染色(b)。
图55描绘了根据本文的实施方案,nk-92mi细胞在γ-辐射后的生长和细胞毒活性。在酶促反应之前,在有或没有6gyγ辐射的情况下接受nk-92mi细胞。将标记的和未标记的nk-92mi细胞再培养3天。(a)在照射和标记后的不同时间点通过流式细胞术计数活细胞。(b)赫赛汀标记的细胞还用抗hfc荧光抗体染色,并在标记后的不同时间点通过流式细胞术分析。被照射的细胞具有较慢的赫赛汀标记衰减。误差条为平均值±sd。在所有图中:ns,p>0.05;***p<0.001;****p<0.0001。
图56描绘了根据本文的实施方案,显示在小鼠中抗肿瘤活性增强的nk-92mi-赫赛汀的图像。显示了三组中30只小鼠的图像。数据在图4k中进行了总结和分析。
图57描绘了根据本文的实施方案,rigg酶促转移至ot-1cd8+t细胞。(a-b)来自ot-1小鼠的脾细胞被ova肽活化,并通过加入il2或il7/il15在体外扩增。扩增4天后,用抗cd3和抗cd8荧光抗体对细胞进行染色,并通过流式细胞术分析(a)。还用gf-al-生物素和ft处理细胞,然后用链霉抗生物素蛋白-apc染色,并通过流式细胞术分析以跟踪t细胞活化后不同天数的lacnac水平(b)。(c)ot-1t细胞用gf-rigg和ft,或单独的gf-rigg处理,或未经处理。然后用抗rigg荧光抗体对细胞进行染色并通过流式细胞术分析。(d)ot-1+/-cd45.1+/-t细胞首先用cfse标记,然后用gf-rigg和fuct处理,或未经处理。岩藻糖基化后,将细胞与ova肽脉冲的wtb6脾细胞共培养48小时,并通过流式细胞术分析t细胞增殖。误差条为平均值±sd。在所有图中:ns,p>0.05;****p<0.0001。
图58描绘了根据本文的实施方案,α-pd-l1转移至ot-1cd8+t细胞上。(a)ot-1cd8+t细胞用gf-α-pd-l1和ft,或单独的gf-α-pd-l1处理,或未经处理。然后用抗rigg荧光抗体对细胞进行染色并通过流式细胞术分析。(b-c)来自p14小鼠的cd8+t细胞用gf-α-pd-l1和ft,或单独的gf-α-pd-l1处理,或未经处理。细胞用抗rigg荧光抗体染色(b),或首先与pd-l1-hfc一起孵育,然后用抗hfc荧光抗体染色(c)。染色后,通过流式细胞术分析这些细胞。(d)在有或没有ft的情况下,在不同浓度的gf-α-pd-l1下标记ot-1t细胞。标记后,用抗rigg荧光抗体对细胞进行染色,并通过流式细胞术分析。(e)ot-1t细胞用gf-α-pd-l1和ft,或单独的gf-α-pd-l1处理,或未经处理。然后用抗rigg荧光抗体对细胞进行染色,并在反应后的不同时间点通过流式细胞术分析。误差条为平均值±sd。在所有图中:ns,p>0.05;****p<0.0001。
图59描绘了根据本文的实施方案,将α-pd-l1转移到ot-1t细胞表面上增强了杀伤癌细胞期间的细胞因子分泌。(a)ot-1t细胞和p14t细胞用gf-α-pd-l1和ft,或gf-rigg和ft处理,或未经处理。然后将t细胞与b16-ova细胞共培养9小时。通过elisa试剂盒定量培养上清液中的tnf-α浓度。(b-c)ot-1t细胞在有或没有ft的不同浓度的gf-α-pd-l1下处理,然后与b16-ova细胞共培养9小时。通过elisa分析培养上清液中的tnf-α和ifn-γ。误差条为平均值±sd。在所有图中:ns,p>0.05;***p<0.001;****p<0.0001。
图60描绘了根据本文的实施方案,将α-pd-l1转移到t细胞表面上增加了杀伤期间t细胞簇的大小和t细胞增殖。(a)ot-1t细胞和p14t细胞用gf-α-pd-l1和ft,或gf-rigg和ft处理,或未经处理。然后将细胞与b16-ova细胞共培养20小时。杀伤期间的t细胞簇用显微镜成像。比例尺:50μm。(b)ot-1t细胞用cfse染色并用gf-α-pd-l1和ft,或gf-rigg和ft处理,或未经处理。然后将细胞与或不与b16-ova细胞一起培养72小时。通过cfse信号稀释分析ot-1t细胞的增殖。
图61描绘了根据本文的实施方案,nk-92mi-赫赛汀在腹膜内癌模型中的体内抗肿瘤活性。向nsg小鼠腹膜内注射100万个mda-mb-435/her2+/f-luc细胞。然后,在肿瘤细胞注射后第5天和第10天通过静脉注射hbss、500万个nk-92mi或nk-92mi-赫赛汀细胞一次处理动物。在肿瘤攻击后第4天、第12天、第17天,通过腹腔注射向小鼠注射d-萤光素,通过ivis系统成像。(a)肿瘤接种后第12天的代表性图像显示赫赛汀标记组小鼠的抗肿瘤活性增强。(b)分析和计算第4天、第12天和第17天的荧光素信号。(c)分析并比较第12天的荧光素信号。(d)分析并比较第17天的荧光素信号。显示个体小鼠的肿瘤大小和平均值±sd;n=8。还显示了代表性图像。在所有图中:ns,p>0.05;*p<0.05;**p<0.01;***p<0.001;****p<0.0001。
图62描绘了根据本文的实施方案,另一种抗体细胞缀合物(acc)策略。(a),用gf-16a(抗muc1)或用gf-16a和ft处理的nk-92细胞的流式细胞术分析。(b),在反应缓冲液中滴定gf-16a浓度范围为0.05mg/ml至0.2mg/ml;每个反应使用相同量的ft并在室温下进行30分钟。(c)和(d),mda-mb-435和skbr3癌细胞上muc1表达的流式细胞术分析。(e)和(f),萤光素酶测定法定量nk-92mi细胞对skbr3-luc和mda-mb-435-luc细胞的细胞介导的细胞毒作用;将16a-nk-92mi缀合物与具有或不具有另外添加的游离16a(5μg/ml)的亲本nk-92mi进行比较。16a和在标记中仅用gf-16a处理的nk-92mi细胞用作阴性对照。平均值±sd(误差条),来自三个独立实验的代表性图。ns,p>0.05;**p<0.01。
图63描绘了根据本文的实施方案,使用唾液酸化策略的酶促生物大分子转移。(a),唾液酸转移酶使得能够在一步中将各种功能性分子酶促转移到细胞表面。(b),cmp-唾液酸-叠氮化物-四嗪(cs-az-tz)和cmp-唾液酸-氨基甲酸炔丙酯-四嗪(cs-poc-tz)的化学结构。(c),cmp-唾液酸缀合的igg(cs-igg)的合成的示意图。(d),st催化的cs-igg向nk92-mi细胞表面转移的流程图(这里测试了三种不同的唾液酸转移酶)。(e),用酶st、底物cs-az-migg或两者处理的nk-92mi细胞的流式细胞术分析。(f),用酶st、底物cs-poc-migg或两者处理的nk-92mi细胞的流式细胞术分析。
具体实施方式
本文引用的所有参考文献、出版物和专利均通过引用整体并入,如同完全阐述了它们一样。除非另外定义,否则本文使用的技术和科学术语具有与本发明所属领域的普通技术人员通常理解的含义相同的含义。hornyak等人,introductiontonanoscienceandnanotechnology,crcpress(2008);singleton等人,dictionaryofmicrobiologyandmolecularbiology第3版,j.wiley&sons(newyork,ny2001);march,advancedorganicchemistryreactions,mechanismsandstructure第7版,j.wiley&sons(newyork,ny2013);和sambrook和russel,molecularcloning:alaboratorymanual第4版,coldspringharborlaboratorypress(coldspringharbor,ny2012),为本领域技术人员提供了本申请中使用的许多术语的一般指导。本领域技术人员将认识到可用于本发明的实践中的许多类似于或等同于本文所述的方法和材料。实际上,本发明决不限于所描述的方法和材料。
如本文所用,术语“岩藻糖基转移酶”是指将岩藻糖从gdp-岩藻糖(鸟苷二磷酸-岩藻糖)供体底物转移至受体底物的酶。受体底物可以是另一种糖,例如在n-连接的糖基化的情况下将岩藻糖转移至核心glcnac(n-乙酰葡糖胺)糖,或如在o-连接的糖基化的情况下由o-岩藻糖基转移酶将岩藻糖转移至蛋白质。
术语“肽”、“多肽”和“蛋白质”可互换使用,是指分离的氨基酸残基聚合物,除非另有定义,否则不限于最小长度。肽、寡肽、二聚体、多聚体等也由通过肽键连接的线性排列的氨基酸组成,无论是生物学上产生并从天然环境中分离,使用重组技术产生,或通常使用天然存在的氨基酸合成产生。在一些方面,多肽或蛋白质是包含非天然存在的氨基酸的“经修饰的多肽”。在一些方面,多肽包含天然存在的和非天然存在的氨基酸的组合,并且在一些实施方案中,肽仅包含非天然存在的氨基酸。如本文所用的术语“肽”包括天然肽(降解产物、合成的肽或重组肽)和肽模拟物(通常是合成的肽),以及作为肽类似物的类肽和半类肽,例如其可具有修饰使得肽在体内更稳定或更能穿透到细胞中。此类修饰包括但不限于n-末端修饰、c-末端修饰、肽键修饰、主链修饰和/或侧链修饰。
如本文所用的术语“抗体”包括特异性结合抗原表位或其模拟表位的多肽或蛋白质复合物。抗体包括完整抗体或其与完整抗体竞争特异性结合的结合片段,并包括嵌合抗体、人源化抗体、完全人抗体和双特异性抗体。结合片段包括但不限于fab、fab'、f(ab')2、fv和单链抗体。在一些实施方案中,抗体被称为免疫球蛋白并且包括各种类别和同种型,例如iga(iga1和iga2)、igd、ige、igm和igg(igg1、igg3和igg4)等。在一些实施方案中,本文所用的术语“抗体”是指多克隆抗体和单克隆抗体及其功能片段。抗体包括经修饰的或衍生的抗体变体,其保留特异性结合表位的能力。抗体能够选择性地结合靶抗原或表位。抗体可包括但不限于多克隆抗体、单克隆抗体(mab)、人源化抗体和其他嵌合抗体、单链抗体(scfv)、fab片段、f(ab')2片段和二硫键连接的fv(sdfv)片段。在一些实施方案中,抗体来自任何来源,例如小鼠或人,包括其嵌合抗体。在一些实施方案中,抗体是人源化的。
抗体的实例包括但不限于3f8、8h9、阿巴伏单抗(abagovomab)、阿昔单抗(reopro)、abituzumab、abrilumab、actoxumab、阿达木单抗(修美乐)、阿德木单抗(adecatumumab)、aducanumab、afasevikumab、阿非莫单抗、阿夫土珠、alacizumabpegol(培阿赛珠单抗)、ald518、阿仑单抗(lemtrada、坎帕斯)、阿利库单抗(alirocumab,praluent)、阿妥莫单抗喷替酸盐(hybri-ceaker)、amatuximab、麻安莫单抗(anatumomabmafenatox)、anetumabravtansine、安弗露单抗、安芦珠单抗、阿泊珠单抗、阿西莫单抗(cea-scan)、ascrinvacumab、阿塞珠单抗、阿特朱单抗(特善奇)、安妥明单抗、阿托木单抗、阿维拉姆单抗(阿维单抗)、巴匹珠单抗、巴利昔单抗(舒莱)、巴维昔单抗、贝妥莫单抗(lymphoscan)、贝格洛姆单抗、贝利木单抗(benlysta)、贝拉珠单抗、柏替木单抗、贝索单抗(斯克提木)、贝伐单抗(安维汀)、bezlotoxumab(zinplava)、比西单抗(fibriscint)、双马单抗、贝美珠单抗、莫比瓦妥单抗、比勒斯单抗、博纳吐单抗(blincyto)、布隆妥维单抗(blontress)、布洛唑单抗、博科齐单抗、布拉兹库单抗、维本妥昔单抗(adcentris)、布里亚明单抗、溴达拉单抗(siliz)、溴化丙单抗、支舒巴单抗、burosumab、cabiralizumab、康纳单抗(易来力)、莫坎妥珠单抗、坎妥单抗瑞伐坦、卡普利单抗、卡罗单抗喷地肽(prostascint)、卡伦单抗、carotuximab、卡妥索单抗(removab)、cbr96-阿霉素免疫偶联物、西利珠单抗、克果图珠单抗、peg化的塞妥珠单抗(cimzia)、西妥昔单抗(爱必妥)、伯格西他土珠单抗、西妥木单抗、克拉扎珠单抗、克立昔单抗、clivatuzumabtetraxetan(hpam4-cide)、钴妥珠单抗、克托西单抗(coltuximabravtansine)、西他土珠、康昔单抗、克仁珠单抗、克洛特单抗、cr6261、达塞珠单抗、达利珠单抗(赛尼哌)、达妥珠单抗、peg化的达匹罗单抗、达雷木单抗(darzalex)、德克特单抗、地昔单抗、地宁图珠单抗(denintuzumabmafodotin)、地诺单抗(prolia)、地帕西珠单抗(depatuxizumabmafodotin)、地洛昔单抗-生物素、地莫单抗、丁妥昔单抗(unituxin)、地利达莫单抗、冬麻罗单抗、阿托度单抗(dorlimomabaritox)、多兹图单抗、度立凸单抗、度丕卢单抗(dupixent)、度夫芦单抗(imfinzi)、度司荼单抗、伊克麦昔单抗、依库珠单抗(舒立瑞)、埃巴单抗、依屈洛单抗(panorex)、依法珠单抗(raptiva)、依芬古单抗(mycograb)、埃尔德单抗、埃格曼图姆单抗、艾洛珠单抗(立控醇)、艾西莫单抗、艾马珠单抗、依米妥珠单抗、依米西珠单抗、恩凡珠单抗、恩夫图单抗(enfortumabvedotin)、peg化恩莫单抗、恩波妥珠单抗、恩诺珠单抗、恩提库单抗、恩昔妥昔单抗、恩皮莫单抗(epitumomabcituxetan)、依帕珠单抗、厄恩诺单抗、厄利珠单抗、艾土马单抗(rexomun)、伊塔木单抗(abegrin)、依曲利珠单抗、依伐单抗、依福单抗(立控醇)、艾韦单抗、法索单抗(neutrospec)、法拉莫单抗、法鲁珠单抗、法辛单抗、fbta05(lymphomun)、泛维珠单抗、费扎尼单抗、菲巴珠单抗、菲拉特珠单抗、弗吉妥姆单抗、弗瑞单抗、弗兰单抗、弗乐提单抗、冯特利珠单抗(huzaf)、芙拉单抗、夫瑞单抗、弗瑞索单抗、弗兰单抗、富图希单抗、加坎珠单抗、加利昔单抗、加尼单抗、甘腾纳单抗、加维莫单抗、吉妥单抗-奥佐米星偶联物(麦罗塔)、格伐克单抗、吉妥昔单抗(rencarex)、维格列巴单抗(glembatumumabvedotin)、哥利木单抗(simponi)、高密昔单抗、古塞尔库姆单抗、伊巴珠单抗、替伊莫单抗(泽瓦林)、伊库克单抗、伊达瑞珠单抗(泰毕安)、伊戈伏单抗(indimacis-125)、imab362、伊马单抗、英西单抗(myoscint)、伊马曲单抗、依克拉单抗、尹达西单抗、英杜斯妥单抗、英彼利珠单抗、英夫利昔单抗(类克)、英特单抗、伊诺莫单抗、奥英妥珠单抗(inotuzumabozogamicin)、易普利姆玛、伊拉土木单抗、伊撒西单抗、伊利珠单抗、伊希珠单抗(taltz)、克里希单抗、拉贝珠单抗(cea-cide)、兰波利珠单抗、兰纳德单抗、兰德珠单抗、拉普特西单抗(laprituximabemtansine)、利勃珠单抗、利马索单抗、棱德里珠单抗、棱西单抗、乐德木单抗、莱卡单抗、利比维单抗、维利伐妥单抗(lifastuzumabvedotin)、利格珠单抗、立罗妥单抗(lilotomabsatetraxetan)、林妥珠单抗、利鲁单抗、洛德昔单抗、洛克韦单抗、莫洛伐妥单抗(lorvotuzumabmertansine)、卢卡珠单抗、peg化的卢利珠单抗(lulizumabpegol)、鲁米昔单抗、鲁曲单抗、mabp1(xilonix)、马塔单抗、马妥昔单抗、马司莫单抗、马维里单抗、马妥单抗、美泊利单抗(bosatria)、迈他珠单抗、米拉珠单抗、明妥莫单抗、米弗西单抗(mirvetuximabsoravtansine)、米托单抗、莫加利单抗、莫纳利珠单抗、莫洛利单抗、莫他珠单抗(numax)、莫昔妥木单抗(moxetumomabpasudotox)、莫罗单抗-cd3(orthocloneokt3)、纳考洛单抗(nacolomabtafenatox)、纳米洛单抗、纳布妥木单抗(naptumomabestafenatox)、纳鲁西单抗(naratuximabemtansine)、纳鲁特单抗、那他珠单抗(tysabri)、那西珠单抗、纳弗单抗、奈巴库单抗、奈米单抗(portrazza)、奈莫珠单抗、奈利莫单抗、奈西伐单抗、尼妥珠单抗(theracim、theraloc)、纳武单抗(欧狄沃)、巯诺莫单抗-美培坦偶联物(verluma)、奥比妥昔单抗、奥匹妥珠单抗(gazyva)、奥卡鲁单抗、奥克里昔单抗(ocrevus)、奥杜莫单抗、奥法木单抗(arzerra)、欧妥单抗(lartruvo)、欧罗单抗、奥马珠单抗(xolair)、欧那妥珠单抗、欧特西珠单抗、欧皮西单抗、莫奥珠单抗(oportuzumabmonatox)、奥戈伏单抗(ovarex)、奥替单抗、奥立昔单抗、奥特鲁单抗、奥索单抗、奥扎尼单抗、奥索利单抗、帕吉巴昔单抗、帕利珠单抗(synagis、abbosynagis)、pamrevlumab、帕尼单抗(维克替比)、潘可单抗、帕诺库单抗、帕萨妥珠单抗、帕昔单抗、帕索妥昔单抗、帕替利单抗、帕特利单抗、派姆单抗(可瑞达)、排姆妥姆单抗(theragyn)、排乐珠单抗、帕妥珠单抗(奥密塔克)、培昔单抗、匹利珠单抗、维匹那妥单抗、品妥莫单抗、普鲁单抗、帕洛利珠单抗、泊加丽珠单抗、波拉妥珠单抗(polatuzumabvedotin)、泊纳珠单抗、泊洛利珠单抗、普立昔单抗、普特夏西单抗、普林木单抗、pro140、秋丽珠单抗、雷克妥木单抗、雷德妥单抗、瑞非单抗、拉潘西珠单抗、雷莫芦单抗(cyramza)、雷珠单抗(诺适得)、拉克昔单抗、瑞伐珠单抗、瑞加韦单抗、瑞利珠单抗、利妥珠单抗、利努单抗、risankizumab、利妥昔单抗(美罗华、rituxan)、利巴珠单抗(rivabazumabpegol)、罗妥木单抗、洛乐杜单抗、罗姆珠单抗、荣塔里珠单抗、罗弗皮珠单抗(rovalpituzumabtesirine)、罗维珠单抗(leukarrest)、卢利珠单抗(antova)、沙斯妥珠单抗(sacituzumabgovitecan)、萨摩利珠单抗、萨皮利珠单抗、萨里单抗、沙妥莫单抗喷地肽、苏金单抗(cosentyx)、塞巴坦单抗、塞妥昔单抗、赛韦单抗、西布曲单抗、sgn-cd19a、sgn-cd33a、西伐木单抗、西妥昔单抗(sylvant)、西妥珠单抗、西普利单抗、西鲁库单抗、维索菲妥单抗、索冷珠单抗、索利单抗、索普昔单抗、索土珠单抗、丝塔单抗、舒乐单抗(leukoscan)、苏维珠单抗、它巴单抗、它卡斯珠单抗(afp-cide)、他度珠单抗、他利珠单抗、它木弗单抗(tactress)、它尼珠单抗、泰普利莫单抗(taplitumomabpaptox)、它瑞妥单抗、太妃珠单抗(aurexis)、阿替莫单抗、特纳妥姆单抗、替奈昔单抗、替普珠单抗、四丙珠单抗、司他洛单抗、替妥洛明单抗、特则培单抗、tgn1412、替西木单抗(tremelimumab)、替卓珠单抗、替加妥单抗、替莫单抗、替索托单抗(tisotumabvedotin)、tnx-650、托珠单抗(actemra,roactemra)、托利珠单抗、托萨托单抗、托西莫单抗(bexxar)、托韦妥单抗、川隆单抗、曲妥珠单抗(赫赛汀)、曲妥珠单抗埃姆他辛(kadcyla)、trbs07(ektomab)、曲加利珠单抗、曲美木单抗、曲弗单抗、西莫白介素单抗、妥韦单抗、乌立妥昔单抗、乌克普鲁单抗、乌鲁单抗、乌托珠单抗、优特金单抗(stelara)、乌托米单抗、维达西单抗(vadastuximabtalirine)、凡多珠单抗(vandortuzumabvedotin)、凡提单抗、凡诺西珠单抗、伐利昔单抗、瓦利芦单抗、瓦特里珠单抗、维多珠单抗(entyvio)、维妥珠单抗、维帕莫单抗、维森单抗、维西珠单抗(nuvion)、弗巴利珠单抗、弗洛昔单抗、沃斯妥珠单抗(vorsetuzumabmafodotin)、伏妥莫单抗(humaspect)、疝托珠单抗、扎鲁木单抗(humax-egfr)、扎木单抗(humax-cd4)、扎昔单抗、齐拉木单抗和阿佐莫单抗。在一个优选实施方案中,抗体是曲妥珠单抗(赫赛汀)。
如本文所用的术语“抗体”还包括本文所述的单克隆抗体的生物类似物或第二代形式。
本文所用的术语“多核苷酸”或“核苷酸”通常是指天然或经修饰的核苷,包括脱氧核糖核苷、核糖核苷、其α-异头物形式等的线性聚合物,通常通过磷酸二酯键或其类似形式连接成几个单体单元,例如2个至4个,到几百个单体单元的大小。当多核苷酸由字母的序列例如“atgcctg”表示时,应理解核苷酸从左至右的顺序为5'->3'。如本文所用的多核苷酸还包括无碱基的糖-磷酸酯或糖-硫代磷酸酯聚合物。
如本文所用的术语“小分子”通常是指低分子量化合物,通常小于900道尔顿。
本文所用的术语“点击化学”或“铜催化的叠氮化物-炔烃环加成反应”或“cuacc反应”是指铜(i)催化的[3+2]-huisgen1,3-偶极环加成末端炔烃和叠氮化物产生1,2,3-三唑。它也可以指还可以使用的该反应的无铜变型(j.m.baskin,j.a.prescher,s.t.laughlin,n.j.agard,p.v.chang,i.a.miller,a.lo,j.a.codelli,c.r.bertozzi,proc.natl.acad.sci.u.s.a.2007,104,16793.)。
如本文所述,根据本文的各种实施方案,发明人开发了一种新的细胞工程技术,通过该技术,分子共价连接在细胞表面上存在的聚糖上。此类分子可以是小化学部分,或更大的生物部分,例如蛋白质、多核苷酸和/或抗体。本文公开的工程细胞的应用广泛,其说明性实例包括增强免疫细胞用于过继细胞治疗(act)的活性;和/或将小分子、蛋白质或抗体与细胞表面缀合,用于新的免疫治疗策略。
在一个实施方案中,本文公开了非天然存在的工程细胞,其包含:通过岩藻糖衍生物与存在于细胞表面上的聚糖共价结合的化学部分或生物部分。如本公开内容所公开的,多种化学和生物部分可以连接到细胞表面的聚糖上。实例包括但不限于药物(例如药品)、催化剂、抗生素、抗体、抗真菌剂、碳水化合物、细胞因子、酶、糖蛋白、脂质、核酸、核苷酸、寡核苷酸、多核苷酸、蛋白质、肽、配体、细胞、核酶、探针分子或其组合。类似地,化学或生物部分可以连接到存在于细胞表面上的不同类型的聚糖,其非限制性实例包括gal(半乳凝素)、glcnac(n-乙酰葡糖胺)、lacnac和/或neuac(唾液酸)。在一个实施方案中,通过将分子共价连接到细胞表面上存在的聚糖上来制备工程细胞的这种技术用一锅法完成,无需纯化。在另一个实施方案中,该技术用一锅法一步原位完成。
在一个方面,本公开提供了工程细胞,其包含与细胞表面上存在的细胞表面聚糖共价结合的化学部分或生物部分,其中化学部分或生物部分选自小分子、多核苷酸、多肽和抗体。在一个实施方案中,工程细胞是免疫细胞。在一个实施方案中,工程细胞是t细胞、树突状细胞(dc)、或自然杀伤(nk)细胞。在一个实施方案中,t细胞是cd8+t细胞或cd4+t细胞。在一个实施方案中,小分子是小药物分子或其药学上可接受的盐或其共晶体。在一个实施方案中,抗体是单链可变片段(scfv)、片段抗原结合(fab)片段或全长抗体,例如全长免疫球蛋白g(igg)抗体。在一个实施方案中,与工程细胞连接的化学部分或生物部分是生物标志物和/或探针。在一个实施方案中,化学部分或生物部分是生物素探针、荧光探针、生物正交反应柄和/或染料标记的单链dna。在一个实施方案中,染料是fam。在一个实施方案中,荧光探针是cy3。在一个实施方案中,生物正交反应柄是四嗪。在一个实施方案中,岩藻糖衍生物是gdp-岩藻糖。在一个实施方案中,工程细胞是嵌合的抗原受体(car)-t细胞。在一个实施方案中,car-t细胞包含基因修饰的t细胞,其中细胞表面glcnac与携带新基序的gdp-岩藻糖共价结合。在一个实施方案中,car包含三个结构域:scfv、fab和/或与同源受体结合的成熟配体。在一个实施方案中,细胞表面聚糖是n-乙酰葡糖胺(glcnac)。在一个实施方案中,化学部分或生物部分通过岩藻糖衍生物与glcnac共价结合。在一个实施方案中,细胞表面聚糖是唾液酸(neuac)。
在一个实施方案中,本公开提供了组合物,其包含:抗体-细胞缀合物,其中一种或多于一种抗体与细胞表面上的一个或多于一个聚糖部分共价结合。在一个实施方案中,细胞是免疫细胞。在一个实施方案中,细胞是原代人t细胞、自然杀伤(nk)细胞、cd4+细胞和/或原代cd8+ot-1t细胞。在一个实施方案中,nk细胞是nk-92mi细胞。在一个实施方案中,细胞是树突状细胞(dc)。在一个实施方案中,抗体是曲妥珠单抗。在一个实施方案中,细胞是nk-92mi,并且至少一种抗体是曲妥珠单抗。在一个实施方案中,多于一种类型的抗体缀合在细胞表面上。在一个实施方案中,聚糖部分是n-乙酰葡糖胺(glcnac)。在一个实施方案中,抗体通过岩藻糖衍生物与glcnac共价结合。在一个实施方案中,聚糖部分是唾液酸(neuac)。在一个实施方案中,组合物是药物组合物。在一个实施方案中,药物组合物还包含药学上可接受的载体或赋形剂。在一个实施方案中,药物组合物用于治疗对象的疾病。在一个实施方案中,抗体-细胞缀合物能够牢固地结合在炎症位点上(抗e-选择素)、靶向特异性癌细胞(抗her2)、和/或阻断免疫检查点(抗pd-l1)。
除了手术、放射和化疗之外,癌症免疫治疗已经成为治疗癌症的第四种方法,它利用身体自身的免疫系统来对抗癌症。虽然在过继细胞治疗(act)和检查点阻断治疗中已经取得了良好的结果,但是输注到患者体内的t细胞的抗癌特性仍有待改善。同样,树突状细胞(dc)长期以来一直被认为是癌症免疫治疗的核心参与者。然而,显示只有一小部分(1%至2%)的总施用dc到达次级淋巴器官以活化t细胞,这限制了该疫苗接种方法的实际应用。
在一个实施方案中,本文公开的工程细胞和方法提供了有吸引力的克服这些限制的方式。在一个实施方案中,本公开提供了治疗对象的疾病的方法,其包括:提供包含工程细胞和药学上可接受的载体的药物组合物;通过向对象施用治疗有效剂量的药物组合物来治疗疾病,其中工程细胞包含与其表面上存在的聚糖部分共价结合的化学部分或生物部分,并且其中化学部分或生物部分选自小分子、多核苷酸、多肽和抗体。在一个实施方案中,抗体是免疫球蛋白g(igg)抗体。在一个实施方案中,抗体是抗pd-l1抗体。在一个实施方案中,抗体是抗e-选择素抗体。在一个实施方案中,抗体是抗her2抗体。在一个实施方案中,抗体是曲妥珠单抗。在一个实施方案中,多于一种化学部分或生物部分与细胞表面共价结合以同时靶向多于一种疾病。在一个实施方案中,细胞是树突状细胞。在一个实施方案中,细胞是免疫细胞。在一个实施方案中,细胞是原代人t细胞、自然杀伤(nk)细胞、cd4+细胞和/或原代cd8+ot-1t细胞。在一个实施方案中,nk细胞是nk-92mi细胞。在一个实施方案中,聚糖部分是n-乙酰葡糖胺(glcnac)。在一个实施方案中,化学部分或生物部分通过岩藻糖衍生物与glcnac共价结合。在一个实施方案中,岩藻糖衍生物包含岩藻糖-炔烃。在一个实施方案中,聚糖部分是唾液酸(neuac)。在一个实施方案中,疾病是癌症。在一个实施方案中,治疗疾病包括减少对象的癌性肿瘤的大小。在一个实施方案中,癌症是乳腺癌。
在另一个实施方案中,本公开提供了治疗、减少、抑制或减轻对象的癌症的方法,其包括:向对象施用治疗有效剂量的药物组合物,所述药物组合物包含工程细胞,该工程细胞具有与其表面上的聚糖部分共价结合的抗体。在一个实施方案中,癌症是乳腺癌。在一个实施方案中,抗体是曲妥珠单抗。在一个实施方案中,细胞是nk-92mi。在一个实施方案中,多于一种抗体结合在细胞表面上以同时靶向多于一种癌症。
本公开还提供了用于制备本文公开的工程细胞的各种方法。在本公开的一个实施方案中,提供了制备工程细胞的方法,该方法包括:制备岩藻糖衍生物或gdp-岩藻糖衍生物,其包含与岩藻糖或gdp-岩藻糖共价结合的化学部分或生物部分;通过将细胞与包含(a)岩藻糖衍生物和/或gdp-岩藻糖衍生物和(b)岩藻糖基转移酶的组合物一起孵育来制备工程细胞。在本公开的另一个实施方案中,提供了制备工程细胞的方法,其包括:通过使携带tco部分的抗体与cs-az-tz或cs-poc-tz反应,产生与抗体缀合的cmp-唾液酸(cs-igg);通过将天然存在的细胞与包含(a)cs-igg和(b)唾液酸转移酶的组合物一起孵育来制备工程细胞。在一个实施方案中,岩藻糖基转移酶是α-1,3-岩藻糖基转移酶。在一个实施方案中,α-1,3-岩藻糖基转移酶是幽门螺杆菌α-1,3-岩藻糖基转移酶。在一个实施方案中,重组制备α-1,3-岩藻糖基转移酶。在一个实施方案中,唾液酸转移酶选自st6gal1、多杀巴斯德氏菌α(2,3)唾液酸转移酶m144d突变体(pm2,3st-m144d)和美人鱼发光杆菌α(2,6)唾液酸转移酶(pd2,6st)。在一个实施方案中,化学部分或生物部分选自小药物分子、生物分子、探针分子、荧光团、多核苷酸、多肽和完整igg、或其组合。在一个实施方案中,生物分子是曲妥珠单抗。在一个实施方案中,细胞是免疫细胞。在一个实施方案中,细胞是原代人t细胞或自然杀伤(nk)细胞。在一个实施方案中,nk细胞是nk-92mi细胞。在一个实施方案中,t细胞是cd4+细胞或cd8+t细胞。在一个实施方案中,化学部分或生物部分通过岩藻糖衍生物共价连接到细胞表面上的n-聚糖的通用单元lacnac。在一个实施方案中,岩藻糖修饰的生物分子和/或gdp-岩藻糖修饰的生物分子包含岩藻糖-炔烃和/或gdp-岩藻糖叠氮化物。在一个实施方案中,使用配体加速和生物相容的铜(i)催化的炔-叠氮化物环加成(cuaac)反应,用化学部分或生物部分进一步修饰生物分子-细胞缀合物。
在一个实施方案中,本文公开了制备适于将分子连接到工程细胞表面上的工程细胞的一锅法,其包括:通过将岩藻糖类似物与包含atp、gtp、l-岩藻糖激酶/gdp-岩藻糖焦磷酸化酶(fkp)、mg2+或mn2+、和无机焦磷酸酶(pp酶)的混合物组合来制备gdp-岩藻糖类似物;通过将天然存在的细胞添加到包含来自步骤(a)的粗产物和幽门螺杆菌α-1,3-岩藻糖基转移酶的组合物中来制备工程细胞。在一个实施方案中,本文公开了制备适于将分子连接到工程细胞表面上的工程细胞的一锅一步法,其包括:将细胞与包含岩藻糖类似物、atp、gtp、l-岩藻糖激酶/gdp-岩藻糖焦磷酸化酶(fkp)、mg2+或mn2+、无机焦磷酸酶(pp酶)和幽门螺杆菌α-1,3-岩藻糖基转移酶的组合物一起孵育。在一个实施方案中,岩藻糖类似物包含与化学部分或生物部分缀合的岩藻糖。在一个实施方案中,岩藻糖类似物包含岩藻糖-炔烃。在一个实施方案中,工程细胞包含与细胞表面上的聚糖连接的化学部分或生物部分。在一个实施方案中,来自步骤(a)的产物通过铜催化的叠氮-炔烃环加成(cuacc)反应被进一步修饰,以产生与小分子、多核苷酸、多肽和/或抗体缀合的gdp-岩藻糖类似物。
在一个实施方案中,本文公开了将细胞表面lacnac或slacnac转化为lex或slex的一锅原位岩藻糖基化策略,其包括:通过将岩藻糖类似物与包含atp、gtp、fkp、mg2+或mn2+和无机焦磷酸酶(pp酶)的混合物组合来制备gdp-岩藻糖类似物;通过将天然存在的细胞添加到包含来自步骤(a)的粗产物和幽门螺杆菌α-1,3-岩藻糖基转移酶的组合物中,而将细胞表面lacnac或slacnac转化为lex或slex。
在一个实施方案中,本文公开了包含工程细胞的组合物,所述工程细胞适于将分子连接到其表面上。在一个实施方案中,工程细胞是免疫细胞。在一个实施方案中,工程细胞是t细胞。在一个实施方案中,连接到细胞表面的分子是小分子药物、抗体、治疗细胞、多肽和/或多核苷酸。在一个实施方案中,抗体是全长抗体。根据本公开内容使用的抗体可以是多克隆抗体、单克隆抗体、嵌合抗体、小鼠抗体、人源化抗体或完全人抗体。在一个实施方案中,与工程细胞连接的分子是单链可变片段(scfv)。在一个实施方案中,与工程细胞连接的分子是片段抗原结合(fab)片段。在一个实施方案中,与工程细胞连接的分子是生物标志物和/或探针。在一个实施方案中,生物标志物和/或探针是荧光染料。在一个实施方案中,工程细胞是嵌合抗原受体(car)-t细胞,其中car-t细胞包含基因修饰的t细胞,其中细胞表面聚糖与携带新基序的gdp-岩藻糖共价结合。在一个实施方案中,car分子包含3个结构域:scfv、fab和/或与同源受体结合的成熟配体。
本公开描述了用于修饰细胞表面聚糖的一锅原位岩藻糖基化策略。其应用包括(1)增强用于过继细胞治疗(act)的免疫细胞的活性;(2)将小分子、蛋白质或抗体缀合到细胞表面以获得新的免疫治疗策略。
几乎所有涉及先天和适应性免疫系统的关键细胞表面分子都是糖基化的。糖基化分子在免疫细胞分化和运输中起重要作用。聚糖配体唾液酸lewisx(slex)与聚糖结合蛋白的选择素家族(p-选择素和e-选择素)之间的相互作用介导血管细胞壁上循环白细胞的束缚和滚动,这促进了白细胞通过内皮进入周围组织的随后的外渗和迁移。然而,发明人已经发现许多具有治疗潜力的活化免疫细胞如淋巴细胞或髓系细胞具有非常低水平的slex。
本发明人通过在此公开一锅原位岩藻糖基化策略将细胞表面n-乙酰乳糖胺(lacnac、galβ1,4glcnac)或其唾液酸化形式(slacnac)转化为lewisx(lex)或唾液酸lewisx(slex)表位解决了该问题。通过该策略将细胞表面lacnac和slacnac残基转化为免疫细胞表面的离体slex,作为“一石二鸟”方法,以提高基于细胞的免疫治疗的功效。在免疫细胞表面上产生的slex表位引导治疗细胞向最需要它们的病变组织的归巢、植入和保留。此外,通过将lacnac和slacnac残基转化为slex,抑制了半乳凝素-lacnac相互作用,从而抑制半乳凝素介导的免疫细胞凋亡。使用这种一锅原位岩藻糖基化方法,可以将诸如抗体的生物大分子直接连接到免疫细胞的细胞表面上用于免疫治疗。
因此,在一个实施方案中,本文公开了制备适于将分子连接到工程细胞表面上的工程细胞的一锅法,其包括:通过将岩藻糖类似物与包含atp、gtp、fkp、mgso4、无机焦磷酸酶(pp酶)的混合物组合来制备gdp-岩藻糖类似物,和通过向细胞添加来自步骤(a)的粗产物和α-1,3-fuct来制备工程细胞。在一个实施方案中,本文公开了制备适于将分子连接到工程细胞表面上的工程细胞的一锅一步法,其包括:将细胞与岩藻糖类似物、atp、gtp、fkp、mgso4、无机焦磷酸酶(pp酶)和α-1,3-fuct一起孵育。在一个实施方案中,工程细胞包含与细胞表面上的聚糖连接的分子。在一个实施方案中,岩藻糖类似物包含岩藻糖-炔烃。在一个实施方案中,来自步骤(a)的产物通过cuacc反应被进一步修饰,以产生与小分子、多核苷酸、多肽和/或抗体缀合的gdp-岩藻糖类似物。在一个实施方案中,本文公开了一锅原位岩藻糖基化策略,用于将细胞表面lacnac或slacnac转化为lex或slex,该策略包括:通过将岩藻糖类似物与包含atp、gtp、fkp、mgso4和无机焦磷酸酶(pp酶)的混合物组合来制备gdp-岩藻糖类似物;和通过向细胞中加入包含来自步骤(a)的粗产物和幽门螺杆菌α-1,3-fuct的组合物来制备工程细胞。尽管本发明人通常使用来自生物体幽门螺杆菌的α-1,3-岩藻糖基转移酶,但对于本领域技术人员明显的是任何岩藻糖基转移酶,无论来源如何,都可用于本文所述的方法、组合物和试剂盒。
在一个实施方案中,本文提供了治疗对象疾病的方法,其包括提供适于将分子连接到工程细胞表面上的工程细胞,和通过向对象施用工程细胞来治疗疾病。在一个实施方案中,本文提供了治疗对象的癌症的方法,其包括:提供工程细胞,其中抗pd-l1抗体与免疫细胞的细胞表面lacnac分子共价连接;向对象施用工程细胞,其中工程细胞中的抗体识别癌细胞上的特异性抗原;和通过使工程细胞与癌细胞接触来治疗对象的癌症。在一个实施方案中,工程细胞包含与细胞表面上的聚糖连接的分子。在一个实施方案中,岩藻糖包含岩藻糖-炔烃。在一个实施方案中,来自第一步骤的产物通过cuacc反应被进一步修饰,以产生与小分子、多核苷酸、多肽和/或抗体缀合的gdp-岩藻糖类似物。在一个实施方案中,在向患者施用工程细胞后,患者的癌性肿瘤的大小减小。
从外部设计免疫细胞表面是改善基于细胞的治疗的有效方法,该方法由于淋巴细胞转化和基因组突变的低风险,而可以与基因操作竞争,或者至少可以用作替代方法。细胞表面聚糖位于细胞表面的最外面,因此是优良的设计靶标。它们可以通过代谢工程或化学酶工程进行有效修饰。如本公开全文所讨论的,发明人开发了一锅原位岩藻糖基化策略以将细胞表面lacnac或其唾液酸化形式的slacnac转化为lex或slex表位。该方法已被广泛用于提高细胞治疗效率,例如基于干细胞、调节细胞和脐带血细胞的治疗。发明人扩展了一锅岩藻糖基化策略以将从小分子到生物大分子(如完整的igg抗体)的不同功能性配置到细胞表面上。幽门螺杆菌α-1,3岩藻糖基转移酶催化的过程快速有效,即使使用大的底物,如与gdp-岩藻糖缀合的蛋白质也是如此。总之,发明人开发了一种全新的将免疫细胞工程化的方法,该方法是生物相容的且基于天然表位模拟物,并且还打开了用小分子药物、抗体和治疗细胞的不同组合进行操作的大门。
例如,可以对t细胞进行基因工程改造,以在它的表面上表达称为嵌合抗原受体(car)的特殊受体。car是允许t细胞识别肿瘤细胞上的特定蛋白质(抗原)的蛋白质。然后将这些工程化的cart细胞离体培养直至它们的数量达到十亿级。然后将扩增的cart细胞群输注到患者中。输注后,如果一切按计划进行,t细胞在患者体内繁殖,并且在其工程化受体的指导下,识别并破坏表面上含有抗原的癌细胞。car可以由三个结构域组成,即抗原结合结构域、跨膜结构域和胞质信号活化结构域。抗原结合结构域分为3大类:(a)源自抗体的单链可变片段(scfv);(b)选自文库的fab抗原结合片段(fab);或(c)与天然配体的同源受体结合的天然配体。car-t在治疗血癌方面显示出有希望的功效,然而,由于肿瘤微环境中存在抑制机制,在实体瘤的治疗中观察到令人失望的结果。
一锅法策略可用于将靶向检查点抑制剂的小分子、scfv、fab或全长抗体配置到car-t、离体扩增的肿瘤浸润淋巴细胞或其他类型的免疫细胞的表面以增强它们的活性。例如,使用该策略,发明人可以将阻断检查点抑制剂的抗体,例如程序性死亡配体1(pd-l1)和细胞毒性t淋巴细胞相关抗原4(ctal-4)引入到car-t细胞(或离体扩增的肿瘤浸润淋巴细胞)的表面上。类似地,发明人可以使用相同的策略将抗ctal-4的抗体引入离体成熟的负载抗原的树突状细胞的表面上以增强其抗肿瘤或抗病毒活性。此外,使用该策略,发明人可以将两种或多于两种不同的分子配置到细胞表面上,例如,抗pd-l1和抗tim3以抑制多于一种检查点抑制剂。可以使用相同的策略将除抗体之外的生物分子配置到细胞表面上。例如,程序性细胞死亡蛋白1可与调节性t细胞缀合以增强其免疫抑制功能。
将抗体如抗pd-l1缀合至t细胞的细胞表面以进行联合免疫治疗可以显著减少待施用的抗体的量。在临床试验中,向患者静脉内施用抗-pd-l1抗体(逐渐增加的剂量为每千克体重0.3mg至10mg),在6周的周期中每14天施用一次,最多16个周期。通过施用缀合抗pd-l1的t细胞,将使用低10倍至100倍的抗pd-l1。
因此,如本文所公开的,发明人开发了通过构建抗体-细胞缀合物来改造细胞以通过一步酶促糖工程来增强细胞疗法的新方法。如本文所述,根据本文的各种实施方案,发明人描述了一锅原位岩藻糖基化的新策略,以将抗原特异性抗体标记到自然杀伤(nk)细胞表面上,从而指导nk细胞杀伤表达特异性抗原的癌细胞。使用表达her2+的癌症模型,发明人已经证明,赫赛汀与nk细胞的酶促缀合在体外和体内增强了对表达her2+的癌细胞的杀伤。该方法比基因方法更容易且更有效。
在一个实施方案中,本公开提供了组合物,其包含:抗体-细胞缀合物,其中一种或多于一种抗体与细胞表面上的一个或多于一个聚糖分子共价结合。在一个实施方案中,细胞是免疫细胞。在一个实施方案中,细胞是原代人t细胞、自然杀伤(nk)细胞系,和/或原代cd8+ot-1t细胞。在一个实施方案中,nk细胞系是nk-92mi。在一个实施方案中,抗体-细胞缀合物如图1b中所述。在一个实施方案中,抗体是曲妥珠单抗(赫赛汀)。在一个实施方案中,多于一种类型的抗体缀合在细胞表面上。在一个实施方案中,细胞是nk-92mi,并且至少一种抗体是曲妥珠单抗(赫赛汀)。在一个实施方案中,抗体-细胞缀合物能够牢固地结合在炎症位点上(抗e-选择素)、靶向特异性癌细胞(抗her2)、和/或阻断免疫检查点(抗pd-l1)。
在一个实施方案中,本公开提供了药物组合物,其包含:如上所述的抗体-细胞缀合物,和药学上可接受的载体或赋形剂。在一个实施方案中,药物组合物用于治疗对象的疾病。在一个实施方案中,本文提供了治疗、减少、抑制或减轻对象的癌症的方法,其包括:向对象施用治疗有效剂量的药物组合物,所述药物组合物包含如本文公开的抗体-细胞缀合物。在一个实施方案中,癌症是乳腺癌。在一个实施方案中,多于一种抗体缀合到细胞表面上以同时靶向多于一种癌症。
在一个实施方案中,本文公开了制备生物分子-细胞缀合物的方法,其包括:将细胞与包含(a)一种或多于一种岩藻糖修饰的生物分子和/或gdp-岩藻糖修饰的生物分子和(b)岩藻糖基转移酶的组合物一起孵育。在一个实施方案中,岩藻糖基转移酶是α-1,3-岩藻糖基转移酶。在一个实施方案中,α-1,3-岩藻糖基转移酶获自幽门螺杆菌。在一个实施方案中,重组制备α-1,3-岩藻糖基转移酶。在一个实施方案中,生物分子是探针分子、荧光团、多核苷酸、多肽和完整igg、或其组合。在一个实施方案中,生物分子是曲妥珠单抗(赫赛汀)。在一个实施方案中,细胞是免疫细胞。在一个实施方案中,细胞是原代人t细胞、自然杀伤(nk)细胞系,和/或原代cd8+ot-1t细胞。在一个实施方案中,细胞是nk-92mi。在一个实施方案中,生物分子共价连接到细胞表面上的n-聚糖的通用单元lacnac。在一个实施方案中,岩藻糖修饰的生物分子和/或gdp-岩藻糖修饰的生物分子包含岩藻糖-炔烃和/或gdp-岩藻糖叠氮化物。在一个实施方案中,使用配体加速和生物相容的铜(i)催化的炔-叠氮化物环加成(cuaac)反应,用化学部分或生物部分进一步修饰生物分子-细胞缀合物。在一个实施方案中,化学部分或生物部分是生物素探针、荧光探针cy3、生物正交反应柄四嗪和/或染料(fam)标记的单链dna。
在一个实施方案中,使用化学酶促方法标记针对nk-92细胞表面的抗体提供了显著的经济优势和节省时间的程序。nk-92细胞不表达靶向特定细胞的用于adcc作用的fc受体,这限制了其广泛应用。基因方法用于在nk细胞上配置高亲和力cd16,从而可结合大剂量的抗体注射用于体内治疗。使用化学酶促标记方法,可以在数小时内产生与抗体缀合的nk细胞,并且仅消耗少量的抗体。
在一个实施方案中,将与岩藻糖连接的抗体转移至细胞表面的一锅法提供了同时标记两种或多于两种抗体的独特、特异性和生物相容的方式。使用化学酶促标记系统,可以在同一nk细胞上配置两种或多于两种抗原特异性抗体,这有助于nk细胞同时靶向不同的癌症,而同一细胞中两种或多于两种抗体的基因表达非常困难。
在一个实施方案中,本文公开的组合物是药物组合物。在各种实施方案中,根据本公开的药物组合物可以配制用于通过任何给药途径来递送。“给药途径”可以指本领域已知的任何给药途径,包括但不限于气溶胶、鼻、口、经黏膜、经皮或肠胃外。“肠胃外”是指通常与注射相关的给药途径,包括眼眶内、输注、动脉内、囊内、心内、皮内、肌肉内、腹膜内、肺内、脊柱内、胸骨内、鞘内、子宫内、静脉内、蛛网膜下、被膜下、皮下、经黏膜或经气管。通过肠胃外途径,组合物可以是用于输注或用于注射的溶液或混悬剂的形式,或者作为冻干粉末。
根据本发明的药物组合物还可含有任何药学上可接受的载体。如本文所用的“药学上可接受的载体”是指药学上可接受的材料、组合物或载剂,其涉及将目标化合物从身体的一个组织、器官或部分携带或运输到身体的另一个组织、器官或部分。例如,载体可以是液体或固体填充剂、稀释剂、赋形剂、溶剂或包封材料、或其组合。载体的每种组分必须是“药学上可接受的”,因为它必须与制剂的其他成分兼容。它也必须适合与任何与之接触的组织或器官接触,这意味着它不得引起毒性、刺激、过敏反应、免疫原性或任何其他过度超过其治疗益处的并发症的风险。
根据本发明的药物组合物还可以被包封、压片或制备成乳剂或糖浆剂用于经口施用。可加入药学上可接受的固体或液体载体以增强或稳定组合物,或促进组合物的制备。液体载体包括糖浆、花生油、橄榄油、甘油、盐水、醇和水。固体载体包括淀粉、乳糖、硫酸钙、二水合物、石膏粉、硬脂酸镁或硬脂酸、滑石、果胶、阿拉伯胶、琼脂或明胶。载体还可包括缓释材料。例如单独的或与蜡一起的单硬脂酸甘油酯或二硬脂酸甘油酯。
药物制剂按照常规药学技术制备,包括必要时针对片剂形式进行研磨、混合、造粒和压制;或对于硬明胶胶囊形式进行研磨、混合和填充。当使用液体载体时,制剂将呈糖浆剂、酏剂、乳剂或水性或非水性混悬剂的形式。这种液体制剂可以直接经口(p.o.)施用或填充进软明胶胶囊中。
根据本发明的药物组合物可以以治疗有效量递送。精确的治疗有效量是在给定对象的治疗功效方面产生最有效结果的组合物的量。该量将根据多种因素而变化,包括但不限于治疗化合物的特征(包括活性、药代动力学、药效学和生物利用度)、对象的生理状况(包括年龄、性别、疾病类型和阶段、一般身体状况、对给定剂量和药物类型的反应性)、制剂中药学上可接受的载体的性质和施用途径。临床和药理学领域的技术人员将能够通过常规实验确定治疗有效量,例如,通过监测对象对化合物施用的反应并相应地调整剂量。有关其他指导,请参阅remington:thescienceandpracticeofpharmacy(gennaro第21版,williams&wilkinspa,usa)(2005)。
有效组合物的典型剂量可以在制造商推荐的范围内,其中使用已知的治疗化合物,并且还如本领域技术人员通过体外响应或动物模型响应所指示的。这些剂量通常可以减少高达约一个数量级的浓度或量而不丧失相关的生物活性。因此,如前所述,实际剂量将取决于医生的判断、患者的状况、以及治疗方法的有效性,该治疗方法基于例如相关原代培养细胞或组织培养组织样品的体外反应性,例如活组织检查的恶性肿瘤,或在适当的动物模型中观察到的反应。
在一个实施方案中,本文公开了包含gdp-岩藻糖衍生物和幽门螺杆菌α-1,3-岩藻糖基转移酶的试剂盒。该试剂盒可用于实施制备适于将分子连接到工程细胞表面上的工程细胞的本发明方法。试剂盒是材料或组分的组合,包括至少一种本发明的组合物。因此,如上所述,在一些实施方案中,试剂盒含有包含gdp-岩藻糖衍生物和幽门螺杆菌α-1,3-岩藻糖基转移酶的组合物。
在本发明的试剂盒中配置的组分的确切性质取决于其预期目的。例如,一些实施方案被配置用于治疗癌症。在一个实施方案中,试剂盒被特别配置用于治疗哺乳动物对象。在另一个实施方案中,试剂盒被特别配置用于治疗人类对象。在其他实施方案中,试剂盒被配置用于兽医应用、治疗对象,例如但不限于农场动物、家畜和实验室动物。
试剂盒中可以包括使用说明。“使用说明”通常包括有形表达,其描述用于使用试剂盒的组分以实现期望结果,例如制备工程细胞的技术。任选地,试剂盒还包含其他有用的组分,例如稀释剂、缓冲剂、药学上可接受的载体、注射器、导管、涂药器、移液或测量工具、包扎材料或如本领域技术人员容易认识到的其他有用的物品。
装配在试剂盒中的材料或组分可以以保持它们的可操作性和实用性的任何方便和合适的方式提供给从业者。例如,组分可以是溶解的、脱水的或冻干的形式;它们可以在室温、冷藏或冷冻温度下提供。组分通常包含在合适的包装材料中。如本文所用,短语“包装材料”是指用于容纳试剂盒内容物,例如本发明的组合物等的一种或多于一种物理结构。包装材料通过众所周知的方法构造,其优选地提供无菌、无污染的环境。试剂盒中使用的包装材料是通常用于化学或生物技术领域的包装材料。如本文所用,术语“包装”是指能够容纳各个试剂盒组分的合适的固体基质或材料,例如玻璃、塑料、纸、箔等。因此,例如,包装可以是玻璃小瓶,其用于容纳适量的含有gdp-岩藻糖衍生物和幽门螺杆菌α-1,3-岩藻糖基转移酶的本发明组合物。包装材料通常具有外部标签,其指示试剂盒和/或其组分的内容物和/或目的。
在以下实施例中进一步描述本公开的实施方案。这些实施例仅是说明性的,并不以任何方式限制所要求保护的本发明的范围。
实施例
实施例1
细胞工程师在不断努力培养作为活跃治疗剂的细胞。过去几年在用过继细胞转移(act)治疗的患者中见证了惊人的结果。最值得注意的是,最近嵌合抗原受体t细胞(car-t)治疗
在临床实践中制备治疗细胞通常需要离体分离、活化和特异性细胞的大规模扩增。基因工程和代谢工程都在细胞制备的早期阶段(例如在活化期间)开始并且可能干扰这些程序。此外,这些方法通常在数天内通过多个步骤完成,有时然后进行随后的化学反应,这需要更多的细胞处理和体外质量控制(图39a)。它们在程序中的复杂性可能会减慢临床转化速度。相反,如本文所公开的细胞表面聚糖的酶工程可以在输注到人体前容易地在一步中实现。酶工程的一个令人印象深刻的实例是用gdp-岩藻糖(gdp-fuc或gf)和外源α(1,3)-岩藻糖基转移酶(fuct或ft)进行的细胞(例如干细胞和调节性t细胞)的离体岩藻糖基化可以促进或改善过继转移中的黏附、归巢或植入过程,目前正在针对其进行一些临床试验。然而,在糖工程中将天然供体底物转移至细胞表面的应用范围较窄。为了丰富细胞表面酶促糖工程的应用,非常需要扩展底物范围以配置多种功能。
此前,palcic和同事描述了利用可以识别gdp-fuc衍生物的人α1,3fuct的松弛底物特异性,用血型b三糖抗原修饰血型o红细胞的方法(参见srivastava,g.;kaur,k.j.;hindsgaul,o.;palcic,m.m.j.biol.chem.1992,267,22356.)。虽然该方案提供了重要的发展,其最值得注意的是将多样化的非天然功能分子酶促转移到细胞表面;但仍然存在几个关键缺点,这使得该方法无法用于治疗应用。
目前公开的原位岩藻糖基化或唾液酸化策略提供了有吸引力的替代方案。本文公开的反应系统比先前公开的方法更有效、更具生物相容性,并且可用于将大分子例如蛋白质、多肽、多核苷酸和抗体转移至细胞。
在一个实施方案中,本公开内容提供了唾液酸转移酶和岩藻糖基转移酶,其有效且位点特异性地在活细胞表面上配置如生物正交柄、荧光探针或预装配的寡糖的功能性,用于聚糖检测、糖蛋白分析或聚糖结构重塑。虽然已经出现了新的应用,但是可以酶促转移的部分仍然是化学合成的小分子或寡聚体。此外,如本文所公开的,复杂且生物学上重要的大分子可以配置在细胞表面上,并且所得的工程细胞可广泛应用于细胞治疗。例如,单克隆抗体(mab)在治疗癌症或自身免疫等疾病方面显示出很大的功效。制备抗体-药物缀合物(adc)将进一步增强mab在癌症治疗中的效力。在一个实施方案中,mab向治疗性细胞表面的酶促转移将通过重塑细胞-细胞相互作用、阻断信号传导途径或产生协同治疗效果而瞬时改变细胞的功能。
在一个实施方案中,本文公开了使用来自幽门螺杆菌的α1,3fuct构建抗体-细胞缀合物(acc)的新策略,其可以在几分钟内将用gdp-fuc衍生物修饰的完整igg抗体转移至活细胞表面上的n-聚糖通用单元lacnac(galβ1,4glcnac)。同时,还发明了将化学酶促反应与生物正交化学偶联的一锅法方案,以由易合成的岩藻糖类似物产生gdp-fuc衍生物修饰的生物材料,这使得该工程方法更加普遍适用且具有成本效益。在一个实施方案中,使用该方法,建立了三种acc在免疫细胞上的实例,包括原代人t细胞、自然杀伤细胞系(nk-92mi)和原代cd8+ot-1t细胞。缀合在细胞表面上的单克隆抗体有助于这些免疫细胞增强在炎症位点的结合(抗e-选择素),靶向特异性癌细胞(抗her2)或阻断免疫检查点(抗pd-l1),从而可以改善不同阶段的过继细胞治疗。
实施例2
图1说明了两步一锅法反应和一步一锅法反应的一个实施方案。
两步一锅法反应:反应通常在含有1.0ml缓冲液(ph7.5)的1.5mleppendorf管中进行,该缓冲液含有l-岩藻糖或其c-5经取代的类似物(最终浓度10mm)、atp(10mm)、gtp(10mm)、mgso4(10mm)、无机焦磷酸酶(90单位,无内毒素)和fkp(9单位,无内毒素)。将反应混合物在37℃下振荡(225rpm)孵育5至6小时。通过tlc监测反应进程,使用10mm四丁基氢氧化铵的80%乙腈水溶液作为展开溶剂(对茴香醛糖染色)。在岩藻糖或岩藻糖类似物消失后,粗产物可直接用于细胞表面岩藻糖基化。如果使用岩藻糖-炔烃或叠氮化物,可以通过cuaac反应进一步修饰粗产物以产生gdp-岩藻糖缀合物。例如,在cu/bttaa(100μm)和抗坏血酸钠(2mm)的存在下,使粗制gdp-岩藻糖-炔烃样品(5mm)与叠氮化物探针(5mm)在37℃下反应1小时。反应完成后,加入bcs(1mm)以淬灭反应。在点击反应之前或之后,粗制的一锅法产物可以直接用于细胞表面上的后续岩藻糖基化反应而无需纯化。用pbs洗涤106个活细胞3次,并重悬于含有20mmmgso4、3mmhepes、0.1%fbs、200μmgdp-fuc(或gdp-fuc-缀合物)、30muα(1,3)fuct的100μlhbss缓冲液中。在37℃下孵育20分钟后,将细胞洗涤3次,然后在含有2%fbs的pbs中封闭30分钟,以进一步应用或分析。
一步一锅法反应:在含有100ul缓冲液(ph7.8)的1.5mleppendorf管中制备试剂,该缓冲液含有l-岩藻糖或其c-5经取代的类似物(最终浓度,1mm)、atp(10mm)、gtp(10mm)、mgso4(20mm)、无机焦磷酸酶(90单位,无内毒素)、fkp(9单位,无内毒素)和30mu幽门螺杆菌α(1,3)fuct(无内毒素)。然后将活细胞(1×106)加入到反应混合物中。在37℃下孵育60分钟后,将细胞洗涤三次,然后在含有2%fbs的pbs中封闭30分钟,以进一步应用或分析。
实施例3
如图2所示,在一个实施方案中,对培养的cho细胞进行一锅岩藻糖基化反应。岩藻糖基化后,将细胞表面slacnac转化为slex,其可通过apc缀合的抗cla抗体进行检测(图2a)。将活cho细胞用pbs洗涤3次,并重悬于100μl岩藻糖基化缓冲液(含有幽门螺杆菌α-1,3-fuct)中,该缓冲液含有纯化的gdp-岩藻糖(在图中表示为一般反应)或一锅法gdp-岩藻糖。在37℃下孵育20分钟后,洗涤细胞,封闭,然后用apc-抗-cla染色。洗涤后,通过流式细胞术分析样品。如图2b和图2c所示,岩藻糖基化反应与一锅法gdp-岩藻糖一起运行良好。浓度滴定表明,使用100μm一锅法gdp-岩藻糖可以使反应饱和,根据本文公开的结果,这与纯化的gdp-岩藻糖相同。对照实验的特征还在于没有幽门螺杆菌α-1,3-fuct或gdp-岩藻糖(图2b),这表明反应需要这两种试剂。对照抗体染色显示在图2d中。(图2c,单向anova检验,柱间,p<0.0001)
实施例4
如图3所示,在一个实施方案中,使用一锅法岩藻糖基化反应用生物素探针来修饰lec2cho细胞(具有高水平的lacnac表达)。根据实施例1的方法在一锅法中合成gdp-岩藻糖-生物素衍生物。使用一锅法岩藻糖基化反应用生物素探针来修饰lec2cho细胞(具有高水平的lacnac表达)。使用gdp-岩藻糖-生物素作为底物,细胞表面用生物素分子进行岩藻糖基化,该生物素分子特异性地位于lacnac上(图3a)。将活lec2细胞用pbs洗涤3次,并重悬于100μl含有一锅法gdp-岩藻糖-生物素的岩藻糖基化缓冲液(含有幽门螺杆菌α-1,3-fuct)中。在37℃孵育20分钟后,洗涤细胞,封闭,然后用apc-链霉抗生物素蛋白(apc-sa)染色。洗涤后,通过流式细胞术分析样品。如图3b和图3c所示,使用一锅法合成的gdp-岩藻糖-生物素,在岩藻糖基化反应后用生物素有效地标记lec2细胞。浓度滴定表明,当使用50um一锅法gdp-岩藻糖-生物素时达到饱和浓度。对照实验的特征还在于没有幽门螺杆菌α-1,3-fuct或gdp-岩藻糖-生物素(图3b的3和4),这表明反应需要这两种试剂。对照抗体染色显示在图3d中。也可以合成其他gdp-岩藻糖衍生物并用于使用相同的方案修饰细胞。这是在lacnac上特异性修饰细胞表面的一般方法。(图3c,单向anova检验,柱间,p<0.0001)。生物素化细胞还可以用sa缀合的部分进行官能化。
实施例5
如图4至图8所示,可以使用本文公开的一锅法岩藻糖基化反应策略修饰细胞表面。
在一个实施方案中,图4说明使用粗制一锅法gdp-岩藻糖-cy3对培养的lec2cho细胞表面进行的修饰。在一个实施方案中,一锅法岩藻糖基化反应用于用荧光染料cy3来修饰lec2cho细胞。通过本文公开的一锅法合成gdp-岩藻糖-cy3衍生物。使用gdp-岩藻糖-cy3作为底物,特异性地在lacnac上用cy3探针修饰细胞表面,该探针可以直接检测(图4a)。岩藻糖基化后,用流式细胞术分析样品。如图4b和4c所示,lec2细胞用cy3探针有效标记。对照实验的特征还在于没有幽门螺杆菌α-1,3-fuct或gdp-岩藻糖-cy3(图4b),这表明反应需要这两种试剂。可以使用共聚焦显微镜原位跟踪细胞表面上的cy3标记的lacnac。
实施例6
在一个实施方案中,图5说明使用粗制一锅法gdp-岩藻糖-四嗪对培养的lec2cho细胞表面进行的修饰。使用一锅法岩藻糖基化反应,用有效的生物正交基团四嗪(tz)来修饰cho-lec2细胞。根据实施例1的方法,通过一锅法合成gdp-岩藻糖-四嗪衍生物。使用gdp-岩藻糖-tz作为底物,在lacnac上特异性地用四嗪基团对细胞表面进行官能化,其可以通过反电子需求diels-alder反应(invda)进一步有效地结合反式环辛烯(tco)(图5a)。岩藻糖基化方案与实施例3中的相同。岩藻糖基化后,样品与tco-cy5探针结合用于检测四嗪基团,然后用流式细胞术分析。如图5b和5c所示,lec2细胞用cy5探针有效标记。对照实验的特征还在于没有α-1,3-fuct或gdp-岩藻糖-tz(图5b),这表明反应需要这两种试剂。
实施例7
在一个实施方案中,图6说明使用粗制一锅法gdp-岩藻糖-bsa对培养的lec2cho细胞表面进行的修饰。使用一锅法岩藻糖基化反应,用模式bsa蛋白来修饰cho-lec2细胞。首先使bsa蛋白与nhs-生物素和nhs-叠氮化物探针反应。然后通过cuaac反应使bsa蛋白上的叠氮基与gdp-岩藻糖-炔烃探针反应。反应后,将混合物用bcs淬灭,然后脱盐。使用gdp-岩藻糖-bsa作为底物,细胞表面特异性地在lacnac上与bsa蛋白结合(图6a)。在该实施例中,gdp-岩藻糖-bsa浓度为0.4mg/ml。岩藻糖基化后,样品用apc-sa染色以检测bsa蛋白上的生物素探针,然后用流式细胞术分析。如图6b和图6c所示,lec2细胞用bsa蛋白有效标记。对照实验的特征还在于没有幽门螺杆菌α-1,3-fuct或gdp-岩藻糖-bsa(图6b),这表明反应需要这两种试剂。
实施例8
在一个实施方案中,图7说明使用粗制一锅法gdp-岩藻糖-抗体对培养的lec2cho细胞表面进行的修饰。使用一锅法岩藻糖基化反应,用抗体来修饰lec2cho细胞。首先使大鼠igg与nhs-叠氮化物探针反应。然后通过cuaac反应使大鼠igg上的叠氮基与gdp-岩藻糖-炔烃探针反应。反应后,将混合物用bcs淬灭,然后脱盐。使用gdp-岩藻糖-igg作为底物,细胞表面特异性地在lacnac上与大鼠igg结合(图7a)。在该实施例中,gdp-岩藻糖-igg浓度为0.2mg/ml。岩藻糖基化后,样品用apc-抗大鼠igg染色以检测大鼠igg,然后用流式细胞术分析。如图7b和图7c所示,lec2细胞用大鼠-igg有效标记。对照实验的特征还在于没有幽门螺杆菌α-1,3-fuct(图6b)。
实施例9
在一个实施方案中,图8说明使用粗制一锅法gdp-岩藻糖-tz和随后的tco-抗体对培养的lec2cho细胞表面进行的修饰。如上文实施例5中所述,在cho细胞表面的lacnac上特异性地用四嗪基团官能化。岩藻糖基化后,使样品与0.1mg/mltco-小鼠igg在室温下反应20分钟(图8a)。首先通过将igg用nhs-tco基团标记来制备tco-igg蛋白。与小鼠igg抗体结合后,用apc-抗小鼠igg对细胞进行染色,然后通过facs分析。如图8b和8c所示,lec2细胞用小鼠igg有效标记。对照实验的特征还在于没有幽门螺杆菌α-1,3-fuct或gdp-岩藻糖-tz(图5b),这表明反应需要这两种试剂。该实施例表明,用四嗪官能化的细胞可以容易地被tco-部分修饰。
实施例10
如图9至图14所示,可以使用本文公开的一锅法岩藻糖基化反应策略修饰细胞表面。
在一个实施方案中,图9说明使用粗制的一锅法gdp-岩藻糖-生物素对培养的野生型cho细胞表面进行的修饰。为了说明一般适用性,使用实施例3中公开的方案,通过一锅法岩藻糖基化反应,用生物素探针修饰野生型cho细胞。岩藻糖基化和用sa-apc染色后,用流式细胞术分析样品。如图9所示,cho细胞用生物素有效标记。
实施例11
在一个实施方案中,图10说明使用粗制一锅法gdp-岩藻糖-生物素对活化的小鼠cd8+细胞表面进行的修饰。为了进一步说明一般适用性,使用一锅法岩藻糖基化反应,用生物素探针修饰活化的小鼠cd8+t细胞。首先表征细胞的表型(图10)。大多数细胞是cd3+cd8+细胞。cd44高表达表明t细胞的活化。将细胞岩藻糖基化并用sa-apc染色,随后用流式细胞术分析样品。如图10所示,小鼠cd8+t细胞用生物素有效标记。
实施例12
在一个实施方案中,图11说明使用粗制一锅法gdp-岩藻糖-生物素对小鼠树突状细胞(dc)表面进行的修饰。为了进一步说明一般适用性,使用一锅法岩藻糖基化反应,用生物素探针修饰小鼠dc细胞。首先表征细胞的表型(图11)。大多数细胞是cd11c+细胞。岩藻糖基化和用sa-apc染色后,用流式细胞术分析样品。如图11所示,dc细胞用生物素有效标记。
实施例13
在一个实施方案中,图12说明使用粗制一锅法gdp-岩藻糖-生物素对初始或活化的人cd8+t细胞表面进行的修饰。为了进一步说明一般适用性,使用一锅法岩藻糖基化反应,用生物素探针修饰人cd8+t细胞。首先表征细胞的表型(图12)。大多数细胞是cd3+cd8+细胞。cd45ro高表达表明t细胞的活化。岩藻糖基化和用sa-apc染色后,用流式细胞术分析样品。如图12所示,初始和活化的人cd8+t细胞都用生物素标记。活化的人cd8+t细胞具有更高的lacnac水平并且被更有效地标记。
实施例14
图13说明使用粗制一锅法gdp-岩藻糖-tz对活化的小鼠cd8+t细胞表面进行修饰然后与tco-小鼠igg结合的一个实施方案。为了说明一般适用性,使用一锅法岩藻糖基化反应将igg配置到活化的小鼠cd8+细胞表面上。表征细胞的表型,然后进行岩藻糖基化。在用gdp-岩藻糖-tz进行岩藻糖基化后,然后将细胞与tco-小鼠igg结合。之后,用apc-抗小鼠igg对细胞进行染色,然后用流式细胞术分析。如图13所示,cd8+t细胞用小鼠igg有效标记。对照实验的特征还在于没有α-1,3fuct(图13),这表明标记是基于非天然的岩藻糖基化。tco-igg不具有非特异性结合(图13)。
实施例15
图14说明使用粗制一锅法gdp-岩藻糖-tz对活化的小鼠cd8+t细胞表面进行修饰,然后与tco-小鼠igg和tco-大鼠igg结合的一个实施方案。在小鼠t细胞表面的lacnac上特异性地用四嗪基团官能化。岩藻糖基化后,使样品与0.1mg/mltco-小鼠igg和0.1mg/mltco-大鼠igg一起在室温下反应20分钟(图14)。首先通过将igg用nhs-tco基团标记来制备tco-igg蛋白。与igg抗体结合后,用apc-抗小鼠igg、fitc-抗大鼠igg对细胞进行染色,然后通过facs分析。如图14所示,小鼠cd8+t细胞用小鼠igg和大鼠igg有效地标记。对照实验的特征还在于仅使用tco-ab(大鼠igg对小鼠t细胞具有一些非特异性结合)(图14)。该实施例表明,用四嗪官能化的细胞可以容易地被两种不同的带有tco基团的igg抗体修饰。
实施例16
图15说明了在ot-1t细胞表面上配置抗pd-l1抗体的一个实施方案。如上文实施例8和实施例13中所述,使用gdp-岩藻糖-tz作为底物,可以用四嗪基团修饰细胞表面,然后通过tz和tco基团之间的invda反应将抗体配置到细胞表面上。在该实施例中,抑制免疫检查点(pd-1/pd-l1途径)的抗pd-l1抗体通过该方法与细胞表面lacnac和slacnac共价连接。将ot-1cd8+t细胞(样品1)用gdp-岩藻糖-tz(样品3)岩藻糖基化,然后与抗pd-l1抗体一起孵育以封闭t细胞表面上的pd-l1蛋白(样品4)。之后,用fitc-抗大鼠igg对细胞进行染色,以标记与天然表达的pd-l1结合的抗体。然后,通过invda反应使细胞与tco-抗pd-l1抗体结合(样品5)。最后,用apc-抗大鼠igg对细胞进行染色,以显示细胞表面上共价连接的抗pd-l1。进行对照实验:将样品4与抗pd-l1混合,或者用pd-l1直接封闭样品1(样品2),然后与tco-抗pd-l1混合。如图15所示,只有带有tz的细胞与抗pd-l1共价结合(apc通道,对照样品与空白细胞相比具有少许背景)。并且所有对照样品仅在fitc通道中显示阳性信号,这表明抗pd-l1与天然表达的pd-l1蛋白结合。该实施例表明该方法可用于采用功能性抗体修饰t细胞表面。
实施例17
图16说明了在ot-1cd8+t细胞表面上配置抗pd-l1抗体以促进过继细胞治疗(act)的一个实施方案。如实施例14中所示,抗pd-l1抗体可以与ot-1细胞表面共价连接。在该实施例中,在天然表达的pd-l1蛋白被封闭后,还用抗pd-l1抗体修饰ot-1cd8+t细胞(在该实施例中,样品在pd-l1抗体处理后未染色)。将鼠b16-ova黑素瘤细胞(5×105)皮下接种到8周龄c57bl/6雌性wt小鼠的剃毛侧腹中。10天后,将105个ot-1cd8+t细胞(具有不同修饰,样品1、样品4和样品5)过继转移至那些携带b16-ova肿瘤的小鼠(第0天,每组n=2)。随后,将105个(在第5天)相同处理的ot-1cd8+t细胞转移到每只实验小鼠中。通过卡尺测量来确定一天或两天的肿瘤大小。在第11天,样品1组中的一只小鼠死亡,并且对所有其他小鼠实施安乐死并称重每只小鼠的肿瘤(图16)。肿瘤质量和生长曲线均表明在癌症治疗中具有共价连接的抗pd-l1抗体的t细胞的过继转移具有显著改善。
实施例18
图17说明体内扩增的小鼠cd8+t细胞的原位一锅法岩藻糖基化增加e-选择素结合并抑制gal-1结合的一个实施方案。如图17a所示,收集来自ot-1+/-cd45.1+/-小鼠的脾细胞并分选以获得纯cd8+初始t细胞。将105个这些新鲜t细胞眶后注射到野生型b6小鼠中。半天后,还向这些小鼠眶后注射104个李斯特氏菌-ova细胞,以激活那些初始的ot-1cd8+t细胞。扩增数天后,分选来自wt受体小鼠的cd8+t细胞,然后通过一锅法反应进行岩藻糖基化。岩藻糖基化后,用e-选择素或gal-1蛋白对细胞进行染色,然后洗涤,用第二抗体染色。染色过程后,用流式细胞术分析样品。cd45.1同类系标志物用于追踪ova-抗原活化的cd8+t细胞。如图17b所示,在每个时间点岩藻糖基化后,ova特异性cd8+t细胞上的e-选择素结合显著增加。在第14天,岩藻糖基化和非岩藻糖基化之间的e-选择素结合的差异高达10倍。相反,在岩藻糖基化后gal-1结合减少(图17c)。
实施例19
图18说明诱导的小鼠调节性t细胞(itreg)的原位一锅法岩藻糖基化增加e-选择素结合的一个实施方案。从samp或akr小鼠脾细胞分选初始cd4+t细胞。然后将这些细胞重悬于t细胞培养基中,并在补充有2μg/mla-cd28、10ng/mlil2、20ng/mltgf-β、10nmra的预包被板中诱导。通过添加具有il2和tgf-β的培养基将细胞浓度保持在1^106/ml。扩增5天后,通过流式细胞术测定多于80%的活细胞是cd4+cd25+t细胞。cd4+cd25+中foxp3+细胞的比例高于90%,这表明这些细胞中的大多数是itreg细胞(图18a)。然后在第6天,用一锅发岩藻糖基化处理itreg细胞。岩藻糖基化后,e-选择素结合水平明显提高(图18b)。
实施例20
图19说明了人t细胞的原位一锅法岩藻糖基化使e-选择素结合增加的一个实施方案。通过ficoll密度梯度离心从人血液中分离人pbmc,然后用平板结合的抗cd3(5ug/ml)和可溶性抗cd28(2ug/ml)活化。活化后二十四小时,更换培养基并将pbmc在含有il2的新鲜t细胞培养基中进行培养并扩增约2周(保持在1^106个细胞/ml)。扩增14天后,通过流式细胞术测定,多于95%的总培养的pbmc是cd45+cd3+t细胞,并且约60%的t细胞表达cd8+表型。用一锅法岩藻糖基化试剂对扩增的人t细胞进行岩藻糖基化。岩藻糖基化后,e-选择素结合水平增加约8倍(橙色对蓝色,图19)。
实施例21
图20说明人天然调节性t(ntreg)细胞的原位一锅法岩藻糖基化增加e-选择素结合并抑制gal-1结合的一个实施方案。通过ficoll密度梯度离心从人血液的分离人pbmc。人ntreg细胞分类为cd4+cd127低cd25+细胞。分选后,使ntreg细胞在含有il2的t细胞培养基中保持1^106个细胞/ml。扩增14天后,约90%的细胞是foxp3+细胞(图20a)。在一锅法培养后,e-选择素结合增加并且gal-1结合略微降低(图20b和图20c)。
实施例22
图21说明了her2特异性人car-t细胞的原位一锅法岩藻糖基化增加e-选择素结合并抑制gal-1结合的一个实施方案。通过ficoll密度梯度离心从人血液中分离人pbmc,然后用平板结合的抗cd3(5ug/ml)和可溶性抗cd28(2ug/ml)活化。活化后二十四小时,用感染复数为5的慢病毒载体转导pbmc,然后在含有il2的新鲜t细胞培养基中培养并扩增约2周(保持1^106个细胞/ml)。扩增14天后,通过流式细胞术测定,多于95%的总培养的pbmc是cd45+cd3+t细胞,并且约90%的t细胞表达her2特异性car-t细胞(图21a)。然后将这些her2特异性car-t细胞接种到her2+skbr3细胞或her2-mda-mb-468细胞的培养物中。在t细胞接种24小时后,仅在her2+skbr3细胞中看到t细胞簇的形成和癌细胞单层的消除(图21b、图21c)。这些结果表明,her2特异性t细胞以her2特异性方式识别并杀死her2+细胞。然后用一锅法岩藻糖基化试剂对扩增的人car-t细胞进行岩藻糖基化。岩藻糖基化后,e-选择素结合水平增加约4倍(图21d),而gal-1结合仅略微降低(图21e)。
实施例23
图22说明了体外分化的小鼠cd8t细胞的原位一锅法岩藻糖基化显著增强e-选择素结合并降低gal-1结合的一个实施方案。(a)将ot-1+/+rag-/-小鼠的脾细胞与1nmsiinfekl肽在生长培养基中一起孵育。三天后,将细胞在补充有20ng/mlil-15和10ng/mlil-7的rpmi培养基中培养。更换培养基并每2天添加细胞因子。细胞因子处理3天后,通过流式细胞术测定,多于95%的活细胞是cd8+t细胞。将相同数量的cd8+t细胞在含有岩藻糖、gtp、atp、fkp和幽门螺杆菌α-1,3-岩藻糖基转移酶的原位岩藻糖基化反应缓冲液(一锅法反应)或含有gtp、atp、fkp和幽门螺杆菌α-1,3岩藻糖基转移酶而没有岩藻糖的原位岩藻糖基化反应缓冲液(对照)中在37℃下孵育30分钟。(b)原位岩藻糖基化和非岩藻糖基化(作为对照)细胞分别用生物素化的半乳凝素-1和人fc缀合的e-选择素染色,洗涤,然后分别用链霉抗生物素蛋白-pe和抗人fc-pe染色,并用流式细胞术分析。
实施例24
图23说明了体外分化的小鼠ot-1cd8t细胞的原位一锅法岩藻糖基化对细胞表面标志物的表达几乎没有影响的一个实施方案。将ot-1+/+rag-/-小鼠的脾细胞与1nmsiinfekl肽在生长培养基中一起孵育。两天后,将细胞在补充有il-2或il-15或il-15+il-7的rpmi培养基中培养。细胞因子处理3天后,通过流式细胞术测定,多于95%的活细胞是cd8+t细胞。将相同数量的cd8+t细胞在含有岩藻糖、gtp、atp、fkp和α1,3岩藻糖基转移酶的原位岩藻糖基化反应缓冲液(一锅法反应)中于37℃孵育30分钟或不进行处理。用相应的抗体对细胞表面标志物进行染色并使用流式细胞术分析。
实施例25
图24说明用体外分化的聚糖修饰的ot-1t细胞处理b16-ova包埋小鼠显著减少肿瘤生长的一个实施方案。将鼠b16-ova黑素瘤细胞(6×105)皮下接种到8周龄c57bl/6雌性wt小鼠的剃毛侧腹中。8天后,将105个原位岩藻糖基化或非岩藻糖基化的il-15+il-7体外分化的ot-1cd8+t细胞过继转移至携带b16-ova肿瘤的小鼠(第0天,每组n=10)。随后,将2×105个(第6天)和5×105个(第9天)相同处理的ot-1cd8+t细胞转移到每只实验小鼠中。通过卡尺测量每天确定肿瘤大小。*p<0.05(通过非参数t检验确定)。
实施例26
图25说明用体内分化的聚糖修饰的ot-1t细胞处理b16-ova包埋小鼠显著减少肿瘤生长并延长肿瘤包埋小鼠的寿命的一个实施方案。(a)为了在体内获得ot-1记忆cd8+t细胞,将1×104个ot-1初始cd8t细胞转移到cd45.1+/+小鼠中,然后在3小时后,用1×104cfu的表达ova的单核球增多性李斯特菌菌株(lm-ova)感染。7天后,用1×105个编码卵清蛋白的囊泡状口炎病毒(vsv-ova)感染受体小鼠,以增强ova-特异性免疫应答。21天后,可通过阴性选择从cd45.1小鼠脾细胞中收获含有约3%至5%ova特异性ot-1记忆cd8t细胞的cd8t细胞。在原位岩藻糖基化或未处理后,含有1×105个ot1记忆cd8t细胞的cd8t细胞静脉注射到携带b16-ova肿瘤的wt小鼠侧腹,每只小鼠在10天前被皮下植入5×105个b16-ova肿瘤。在第0天、第5天和第10天注射记忆t细胞三次。(b)将如上所述用原位岩藻糖基化处理的或未处理的ot-1记忆cd8+t细胞过继转移到携带b16-ova肿瘤的小鼠(每组n=5)中。每隔一天通过卡尺测量监测肿瘤大小,当肿瘤大小达到1.8cm时处死小鼠。数据(平均值+/-s.e.m.)代表三个独立实验。*p<0.05,**p<0.005(通过非参数t检验分析)。(c)随时间监测小鼠存活率。通过kaplan-meier对数秩检验分析存活率。
实施例27
图26说明了体外分化的小鼠树突状细胞(dc)的原位一锅法岩藻糖基化显著增强e-选择素结合并降低半乳凝素-1结合的一个实施方案。根据本文公开的方法,使用一锅法岩藻糖基化反应来修饰小鼠骨髓来源的树突状细胞。通过用gm-csf培养骨髓祖细胞并用lps刺激来制备成熟bmdc,其中cd11c和mhcii高表达(图26a和图26b)。岩藻糖基化后,细胞表面slacnac转化为slex,其可通过与alexafluor647结合的抗cla抗体进行检测(图26c)。e-选择素结合也增加(图26d)。相反,在岩藻糖基化后,半乳凝素-1结合降低(图26e)。(a).骨髓来源的树突状细胞(bmdc)的分化和成熟方案。简而言之,收集骨髓细胞,通过加入红细胞裂解缓冲液悬浮于pbs中以消除红细胞,然后在含有5%co2的湿润培养箱中、在37℃下以1×106个细胞/ml接种到含有重组gm-csf(20ng/ml,peprotech)的rpmi1640培养基中。用相同剂量的重组gm-csf以3天的间隔喂养细胞一次。在第6天收获未成熟的树突状细胞,其中cd11c+细胞的纯度高于90%。然后用lps刺激细胞进行成熟。(b).用lps(100ng/ml)刺激未成熟的dc12小时,通过流式细胞术分析cd11c和mhcii的表达。(c-e).使用一锅法策略使成熟dc岩藻糖基化,然后用抗cla抗体、e-选择素或gal-1蛋白对细胞进行染色,然后洗涤,用第二抗体染色。染色过程后,用流式细胞术分析样品。m.f.i表示“荧光强度中值”。数据表示为平均值±标准差,并通过prism(graphpad)软件中的双尾学生t检验分析统计显著性。p值<0.05的差异被认为是统计学上显著的。****表示p<0.05。
实施例28
图27说明了负载抗原的dc的岩藻糖基化诱导显著更强的抗肿瘤免疫应答的一个实施方案。为了评估岩藻糖基化dc在诱导抗肿瘤免疫应答中的作用,使用b16-ova黑素瘤肿瘤模型(图27a)。与未岩藻糖基化dc疫苗组相比,发现岩藻糖基化dc疫苗更有效地抑制肿瘤生长。(a).岩藻糖基化树突状细胞疫苗抑制体内肿瘤生长的方案。c57bl/6野生型受体小鼠在右侧腹皮下接种5x105个b16-ova黑素瘤细胞,在肿瘤细胞接种后第6天通过眼球后注射将5×103个初始ot-it细胞接种到小鼠中。在第8天,将成熟dc与200nmova257-264(siinfekl)肽一起孵育1.5小时,洗涤,然后使用一锅法策略使其岩藻糖基化。岩藻糖基化后,通过右侧腹皮下注射将4×105个dc接种到携带肿瘤的小鼠中。然后记录肿瘤生长。(b).通过游标卡尺测量肿瘤大小并计算。每个实验组中有8只小鼠。数据表示为平均值±标准差,并在prism(graphpad)软件中用mann-whitney非参数检验分析统计学显著性。p值<0.05的差异被认为是统计学上显著的。****表示p<0.05。
实施例29
图28说明使用粗制一锅法gdp-岩藻糖-dna-fam来修饰培养的lec2cho细胞表面的一个实施方案。一锅法岩藻糖基化反应用于用多核苷酸(单链dna)来修饰cho-lec2细胞。根据实施例1的方法,通过一锅法合成gdp-岩藻糖-dna-fam。dna底物是具有5'炔基和3'fam荧光团的单链dna。如本文所用的术语“fam”或“6-fam”是指荧光染料6-羧基荧光素。使用gdp-岩藻糖-dna-fam作为底物,细胞表面的lacnac特异性地被单链dna官能化,这可以通过dna上的荧光信号证实(图28)。岩藻糖基化方案与实施例3中的相同。如图28(b)所示,用gdp-岩藻糖-dna-fam岩藻糖基化的lec2细胞显示fam荧光的显著增加。对照实验的特征还在于没有1,3fuct(图28(b)),这表明信号来自岩藻糖基化反应。
实施例30
图29说明了配置在培养的lec2cho细胞表面上的抗人e-选择素可与人e-选择素结合的一个实施方案。通过实施例8中的方案将抗人e-选择素抗体配置在lec2细胞表面上。之后,将用抗人e-选择素或其同种型抗体标记的细胞与人e-选择素fc蛋白一起孵育。结果表明只有用抗人e-选择素标记的细胞可以与e-选择素蛋白结合(图29),这通过用apc-抗-fc抗体染色进行检测。
实施例31
图30说明了配置在培养的lec2cho细胞表面上的抗小鼠pd-l1可以结合小鼠pd-l1的一个实施方案。使用实施例8中描述的方案将抗小鼠pd-l1配置在lec2细胞表面上。之后,将用抗小鼠pd-l1或其同种型抗体标记的细胞与小鼠pd-l1fc融合蛋白一起孵育。结果表明只有用抗小鼠pd-l1标记的细胞才能与小鼠pd-l1蛋白结合(图30),这通过用apc-抗-fc抗体染色进行检测。
实施例32
图31说明了配置在培养的cho细胞表面上的抗小鼠ctla4可以结合小鼠ctla4的一个实施方案。使用实施例8中描述的方案将抗小鼠ctla4配置在cho细胞表面上。之后,将用抗小鼠ctla4或其同种型抗体标记的细胞与小鼠ctla4fc蛋白一起孵育。结果表明只有用抗小鼠ctla4标记的细胞才能与小鼠ctla4蛋白结合(图31),这通过用apc-抗-fc抗体染色进行检测。
实施例33
图32说明配置在小鼠树突状细胞表面上的抗小鼠ctla4可与小鼠ctla4蛋白结合的一个实施方案。首先将小鼠dc细胞与抗cd16/32一起孵育以封闭fc结合。然后使用抗ctla4抗体封闭dc细胞表面上的ctla4抗原。之后,使用实施例8中描述的方案将tco-抗-ctla4抗体配置在dc表面上。然后将用抗小鼠ctla4或其同种型抗体标记的细胞与小鼠ctla4fc蛋白一起孵育。结果表明,与结合同种型的细胞相比,用抗小鼠ctla4标记的细胞可以结合更多的ctla4蛋白,这通过用apc-抗-fc抗体染色进行检测。
实施例34
图33说明配置在ot-1cd8t细胞表面上的小鼠igg可在表面上停留多于24小时的一个实施方案。如实施例13中所述,将小鼠igg抗体与ot-1cd8t细胞结合。随后,使用apc-抗小鼠igg追踪t细胞表面共价连接的小鼠igg分子。如图33所示,24小时后可以明显检测到来自小鼠igg的信号。
实施例35
图34说明了ot-1cd8t细胞与小鼠igg抗体结合不影响增殖的一个实施方案。如实施例13中所述,将小鼠igg抗体与ot-1cd8t细胞结合。然后将细胞在含有10ng/mlil2的t细胞培养基中培养。在三天内计数细胞数量。数据显示,经修饰细胞的增殖率与未经修饰细胞的增殖率相似。
实施例36
图35说明配置在ot-1cd8t细胞表面上的抗小鼠pd-l1可在细胞表面上停留多于24小时的一个实施方案。如实施例15中所述,将抗pd-l1抗体与ot-1cd8t细胞结合(在用抗pd-l1抗体封闭后,在该实施例中细胞未用fitc-抗大鼠igg染色)。之后,apc-抗大鼠igg用于追踪t细胞表面共价连接的抗pd-l1抗体。如图35所示,24小时后可以明显检测到来自结合的抗pd-l1的信号。
实施例37
图36说明了ot-1cd8t细胞与抗pd-l1抗体结合不影响增殖的一个实施方案。如实施例16中所述,将抗pd-l1抗体与ot-1cd8t细胞结合。然后将细胞在含有10ng/mlil2的t细胞培养基中培养。在三天内计数细胞数量。数据显示,经修饰细胞的增殖率与未经修饰细胞的增殖率相似。
实施例38
图37说明了配置在ot-1cd8t细胞表面上的抗小鼠pd-l1可以结合小鼠pd-l1的一个实施方案。通过实施例16中的方案将抗小鼠pd-l1或大鼠igg同种型对照配置在ot-1t细胞表面上(在与tco-抗体共价反应之前,细胞被抗pd-l1封闭)。之后,将用抗小鼠pd-l1或其同种型抗体标记的细胞与小鼠pd-l1fc蛋白一起孵育。结果显示,与结合同种型的细胞相比,用抗小鼠pd-l1标记的细胞可以结合更多的pd-l1蛋白(图37)。同种型对照组中的pd-l1结合来自t细胞表面上的封闭的抗pd-l1抗体。
实施例39
图38说明了配置在ot-1cd8t细胞表面上的抗小鼠pd-l1可以增强t细胞对癌细胞的杀伤功能的一个实施方案。通过实施例16中的方案将抗小鼠pd-l1或大鼠igg同种型对照配置在ot-1t细胞表面上。之后,将具有或不具有修饰的ot-1cd8t细胞与b16-ova-luc细胞(b16黑素瘤细胞,其具有过表达的ova抗原和荧光素酶标记物)混合并孵育2天。根据成像和荧光素酶活性测定,与ot-1cd8t细胞或同种型标记的t细胞相比,具有共价连接的抗pd-l1抗体的ot-1细胞可以诱导更多的t簇并杀死更多的癌细胞。
实施例40
在一个实施方案中,本文公开了一种用于有效且成本效益好地产生gdp-fuc衍生物的一锅法。在制备规模中缺乏用于合成和纯化通用供体底物的简便方法阻碍了酶促糖工程的应用。事实上,本发明人先前已经表明,来自幽门螺旋杆菌的fuct酶可以接受具有比岩藻糖c6位置处的叠氮化物大的官能团(例如荧光素)的gdp-fuc衍生物。然而,在纯化一步改造底物中产物的大量损失使得在细胞表面更优选两步法。为扩大fuct实现的一步改造的范围和应用,gdp-fuc的各种衍生物应易于大规模制备。由于gdp-fuc类似物合成的方法几乎是定量的(>90%),如果反应后的所有组分都是生物相容的,那么随后的细胞表面反应似乎不需要产物的纯化。为了测试该假设,本发明人使用生物相容性mg2+制备了gdp-fuc的一锅法粗制产物。一锅法产物无需纯化即可直接使用,并且效率与细胞表面上的纯gdp-fuc相同(图44)。为了使这种一锅法与通用gdp-fuc衍生物兼容,使用先前开发的配体加速和生物相容的铜(i)催化的炔烃-叠氮化物环加成(cuaac)用不同的部分直接修饰gdp-fuc炔烃(gf-al)或gdp-fuc叠氮化物(gf-az)的一锅法产物(图39c)。参见,例如,wang,w.等人,proc.natl.acad.sci.usa2009,106,16096;besanceney-webler,c.等人angew.chem.int.ed.2011,50,8051,其全部内容通过引用整体并入,包括附图。过夜后所有反应均以高产率(>95%)进行,然后通过铜螯合剂(浴铜灵磺酸盐,bcs)淬灭,使得一锅法产物更具生物相容性,可直接用于细胞。与用于细胞表面lacnac标记的常规两步方案(酶促转移随后进行细胞表面点击化学)相比,使用一锅法gf-al-生物素的一步酶标记更有效且更加生物相容(图45)。除生物素探针外,荧光探针cy3、生物正交反应柄四嗪(tz)和染料(fam)标记的单链dna(ssdna)也通过此高效的一锅法方案与gf-al或gf-az结合(图39c和图46)。随后使用fuct通过一步酶促反应将所有这些一锅法产物有效地转移到细胞表面(图47)。这些结果证明这种一锅法是可靠的,该产物一般适用于随后的细胞表面反应。另外,由gf-az制成的一锅法tz衍生物比由gf-a1制备的一锅法tz衍生物更有效(图46d)。与发明人先前的结果一致,gf-az是fuct的更有利的底物。
实施例41
在一个实施方案中,本文公开了使用fuct将完整igg分子转移至细胞表面的酶促转移方法。通过一锅法方案,gdp-fuc衍生物可以轻松实现多元化。其中,gf-az-tz是具有非常强大的生物正交柄(tz)的目标衍生物,可以与携带有应变的亲双烯体的生物大分子进一步反应。这种类型的反应是所谓的反电子需求狄尔斯-阿尔德反应(iedda),具有极高的效率和优异的生物正交性。发明人考察了来自完整igg的fuct的底物限制,因为它是最大的治疗性生物分子之一。通过标准胺偶联程序将具有peg接头的反式环辛烯(tco)生物正交柄配置到mab或其同种型对照上。tco-peg4-nhs酯的标记试剂可商购获得,并且对方案进行了充分研究。之后,携带tco部分的mab直接与一锅法gf-az-tz反应以产生gdp-fuc修饰的igg分子(gf-igg)(图40a)。gdp-fuc标记的大鼠igg(gf-rigg)的一锅法产物首次在细胞表面上通过fuct应用于酶促转移中(图40b)。由于lec2cho细胞具有明确定义的聚糖补体,选择具有主要终止于lacnac单元的复合/杂合n-聚糖的lec2cho细胞作为模式细胞。令人惊讶的是,gf-rigg向lec2表面的转移在正常离体岩藻糖基化条件下是有效且特异的:通过流式细胞仪分析的细胞表面上rigg的非常阳性信号仅在添加fuct(60mu)和gf-rigg(0.1mg/ml)两者时检测到(图40c)。同时,标记强度依赖于gf-rigg浓度,其在0.1mg/ml时达到饱和(图40d)。最值得注意的是,即使在冰上,反应也可在10分钟内完成(图40e)。rigg标记的细胞的活力与未标记细胞相似,这进一步证实了反应和一锅法底物的生物相容性(图40f)。将不表达lacnac的lec8cho细胞用作阴性对照以确认转移依赖于igg分子上的gdp-fuc部分与细胞表面上的lacnac单元之间的反应。如所预期的,与lec2细胞相比,反应后lec8细胞显示的荧光没有增加(图40g)。还进行了使用被fuct的天然底物gdp-fuc封闭的细胞的竞争实验,其中随后的gf-rigg标记几乎被消除(图48)。这些结果进一步证实了gf-rigg的酶促转移的反应位点与细胞表面上的gdp-fuc几乎相同,已经证明该反应位点为复合/杂合n-聚糖中的lacnac单元。除流式细胞术分析外,还拍摄了共聚焦显微镜图像以确认大部分标记的rigg停留在细胞表面(图40h)。为了显示酶工程在引入多种功能方面的优势,在酶促反应中以1:1的比例混合gf-rigg和gf-migg(gdp-fuc修饰的小鼠igg)。清楚地检测到lec2细胞表面上两种不同igg分子的双重标记,这在基因工程中很难实现(图40i和图49)。
实施例42
在一个实施方案中,本文公开了用于将α-e-选择素转移至t细胞的酶促转移方法,使得能够牢固地结合至发炎部位。细胞归巢到患病组织是act成功的第一个重要问题。简而言之,聚糖配体唾液酸路易斯x(slex)和选择素家族(例如e-选择素)之间的相互作用介导血管细胞壁上循环白细胞的束缚和滚动,这促进了随后的白细胞通过内皮进入周围组织的外渗和迁移(图41a)。在患者中,来自发炎部位如肿瘤、感染或受损组织的细胞因子(如tnf-α)诱导附近血管内皮细胞上e-选择素的过度表达,这有助于募集免疫细胞进行治疗。因此,靶向e-选择素诱导牢固结合可以帮助过继转移细胞归巢到患病组织以进行增强治疗。与增加细胞表面上slex的量的常规方法不同,在一个实施方案中,发明人已经提出在t细胞表面上配置α-e-选择素可以直接介导t细胞与具有高e-选择素表达水平的内皮细胞的结合,从而可以绕过束缚、滚动和牢固黏附的自然过程(图41a)。
为了证明这种可能性,本发明人使用了临床相关的原代人t细胞(在car-t的相似培养条件下)。通过抗cd3/cd28处理来活化新鲜制备的人外周血单核细胞(pbmc)并离体扩增约两周(图50中表型的表征)。在培养过程中,通过具有gf-al-生物素的酶标记追踪人t细胞的lacnac水平(图51a)。cd4+t细胞和cd8+t细胞在活化后大量增加了lacnac水平,其在两周后保持在中等水平(cd8+高于cd4+)。在确认离体扩增的人t细胞具有足够的lacnac用于标记后,使用fuct将gdp-fuc修饰的抗人e-选择素(gf-α-he-sel)转移至人t细胞以及同种型对照gf-migg。这两种抗体都特异性地标记于人t细胞,而三种不同供体之间标记效率的变化可忽略(图41b、图51b、图51c)。之后,使用几种表面标志物对与migg结合的细胞进行染色,并与未标记的细胞进行比较。结果表明igg分子向细胞表面的酶促转移不影响表面标志物(图50b)。结合在人t细胞表面上的完整igg保持多于24小时(24小时后剩余约50%)并且对细胞增殖没有影响(图51d和图51e)。然后将标记的或未标记的人t细胞与he-选择素/人fc嵌合体蛋白(he-选择素-hfc)混合。与未标记的细胞或migg组相比,用α-he-sel标记的细胞与其抗原he-选择素-hfc蛋白强烈结合,这表明与细胞表面结合的mab仍然具有功能(图41c、图51f)。同时,在流动室测定中,将具有不同修饰的t细胞分别与未标记的细胞混合,然后在一定的剪切应力下流过he-选择素-hfc包被的载玻片。只有当t细胞与α-he-sel结合时才能实现与载玻片上e-选择素的牢固结合(图41d)。为了进一步证实对细胞表面的结果,用抗体修饰cfse标记的人t细胞(绿色),然后加入用或不用tnf-α刺激的人脐静脉内皮细胞(huvec)。洗去未结合的细胞后,使用平板读数器通过cfse的荧光强度对结合huvec的t细胞进行定量。当将与α-he-sel结合的t细胞加入tnf-α刺激的huvec时,显示出最高信号(图41e)。图41f中所示的并行成像实验具有类似的结果。此外,进行迁移(transwell)测定以显示人t细胞相对于huvec单层的跨内皮迁移能力不受抗体标记的影响(图41g)。这些结果表明,通过配置α-he-sel对人t细胞进行改造使得t细胞在tnf-α刺激的内皮细胞上牢固结合而不影响跨膜迁移能力,这表明该改造可能潜在地促进t细胞有效归巢到患病组织。
实施例43
在一个实施方案中,本文公开了用于酶促构建nk-92mi细胞与赫赛汀的缀合物以增强癌症杀伤的方法。特异性杀伤是act在基于细胞的癌症免疫治疗中的关键步骤。人类自然杀伤(nk)细胞在对抗恶性细胞的先天免疫中起关键作用,并且还被开发为癌症免疫治疗中的有效消灭癌症的方式。nk细胞介导靶细胞的特异性杀伤涉及几种机制,其中抗体依赖性细胞介导的细胞毒作用(adcc)是最重要的机制之一。然而,基于nk细胞的治疗的一个挑战是难以从患者的血液中获得足够数量的活性nk细胞。nk-92是一种从患有克隆性nk细胞淋巴瘤患者建立的高细胞毒性自然杀伤(nk)细胞系,可用于产生gmp级的更多数量的细胞毒性nk细胞,这就是所谓用于基于nk的过继性癌症免疫治疗的“现成的治疗”。目前,在美国、加拿大和德国进行使用正常nk-92细胞管理不同恶性肿瘤的四项i期临床试验。然而,特异性靶向是良好癌症治疗的黄金标准。nk-92细胞不表达针对特定细胞的用于adcc作用的fc受体,这限制了它的广泛的应用。在一个实施方案中,如本文所述,mab可以比基因方法更容易且更有效地酶促转移至细胞表面。因此,在一个实施方案中,nk-92细胞-mab缀合物的构建将是在治疗中实现特异性靶向的一种很好的技术。
nk-92mi细胞是不断表达高水平lacnac的nk-92细胞系的il-2独立变体,这对于基于fuct的改造是有益的。预期,在正常的离体岩藻糖基化条件下,将gdp-fuc修饰的人igg(gf-higg)转移至nk-92mi细胞表面是有效和特异的(图52a)。赫赛汀也称为曲妥珠单抗,是用于治疗人表皮生长因子受体2阳性(her2+)乳腺癌的单克隆抗体。在一个实施方案中,与赫赛汀结合的nk-92mi细胞可以靶向her2+癌细胞以使得通过接近而增强癌症杀伤(图42a)。使用fuct将gf-赫赛汀成功转移至nk-92mi细胞表面(图52b)。显示在细胞表面上的赫赛汀的半衰期约为20小时(图52c)。与nk-92mi细胞结合的赫赛汀仍然仅与her2抗原结合(图42b、图52d、图52e)。另外,与赫赛汀结合的nk-92mi细胞在共培养系统中与her2+乳腺癌细胞bt474强烈结合,而未修饰的nk-92mi细胞与bt474弱结合(图42c和图42d)。此外,赫赛汀标记的nk-92mi细胞在离体中比未标记的nk-92mi细胞更有效地杀伤bt474细胞(图42e)。对照higg标记和游离赫赛汀共处理都不能增强nk-92mi对bt474的杀伤活性,这表明需要赫赛汀和nk-92mi细胞表面之间的结合(图42e)。nk92-mi-赫赛汀的增强杀伤作用后来在包括skbr3和mda-mb-435/her2+在内的其他her2+癌细胞中得到证实,但在her2-癌细胞如mda-mb-435和mda-mb-468中未被证实(图42f)。该特异性进一步证实与赫赛汀结合的nk-92mi细胞通过与癌细胞表面上的her2抗原的强结合而增强杀伤。有趣的是,颗粒酶b的总分泌不会增加以增强杀伤效果(图52e)。该结果告诉我们nk-92mi细胞和靶细胞之间的接近可以充分诱导nk-92mi细胞的增强的杀伤活性和特异性。酶促反应中高浓度的gf-赫赛汀使nk-92mi细胞表面上发生更多的赫赛汀结合和抗原结合(图53a和53b)。因此,nk-92mi细胞的杀伤效率也取决于标记反应中的gf-赫赛汀浓度(图42g)。与其他细胞介导的细胞毒作用相似,较高的效应物与靶细胞比例也具有更好的杀伤结果,但仅在赫赛汀标记的nk-92mi组中,其在5:1时达到饱和(图42h)。酶工程的另一个重要优点是人们可以同时将几种抗体结合到细胞表面上。为了显示该应用,用赫赛汀和α-egfr两者标记nk-92mi细胞(图54a和54b),这对skov3细胞(her2+egfr+)的杀伤效率比单个mab标记更好(图42i)。
如从淋巴瘤患者中开发的,应当辐照nk-92用于临床应用。用6gy辐照nk-92mi细胞阻止其增殖(图55a)。辐照nk-92mi细胞与未辐照的细胞的杀伤效率相当(图42j)。在一个实施方案中,发现与未辐照的组相比,与经辐照的nk-92mi结合的赫赛汀的半衰期略长(图57b)。增强离体杀伤效果的有希望的结果使我们尝试在体内测试该系统。在一个实施方案中,发明人选择实验性肺转移模型用来证明构思,其中nsg小鼠接受静脉内注射mda-mb-435/her2+/f-luc细胞(用萤火虫萤光素酶稳定转导)。在肿瘤细胞接种一天后,通过静脉内注射hbss缓冲液、亲本nk-92mi或nk-92mi-赫赛汀细胞(经辐照的nk-92mi细胞)处理小鼠。在肿瘤攻击后6天,通过纵向无创生物发光成像确定小鼠的肺肿瘤大小。虽然用亲本nk-92mi细胞处理可适度减少肺肿瘤形成(比hbss组小约48%),但赫赛汀标记的nk-92mi细胞具有显著增强的体内肿瘤杀伤活性(比hbss组小约83%)(图42j和图56)。总之,nk-92mi-赫赛汀缀合物在离体和体内均比亲本nk-92mi细胞显著增强了抗肿瘤效果,这显示出可以转化为临床试验的很大潜力。
实施例44
在一个实施方案中,如本文所公开的,结合在细胞表面上的α-pd-l1能够阻断pd-1/pd-l1途径并在体外增强t细胞增殖。除了增强或诱导细胞-细胞相互作用的应用之外,本公开内容显示结合至细胞表面的mab可特异性地影响信号传导途径以增强治疗。首先,因为ot-1cd8+t细胞的转基因t细胞受体(tcr)仅识别由mhci分子呈递的ova257-264肽,所以选择它作为模式细胞,这是研究cd8+t细胞应答特异性抗原的良好模型。ot-1脾细胞被ova肽活化,并通过加入il2或il7/il15离体扩增。三天后,大多数细胞是cd8+t细胞(图57a),而它们的lacnac水平足够高以在活化后进行标记(图57b)。为了排除细胞表面上的大igg分子非特异性地阻断tcr信号传导的可能性,使用ova肽脉冲的野生型b6脾细胞再次刺激与rigg结合的ot-1+/-cd45.1+/-t细胞(图57c)。根据cfse稀释的结果,igg分子不阻断tcr和mhci之间的相互作用,因为标记的t细胞与未标记的细胞具有相似的增殖速率(图57d)。
虽然t细胞活化依赖于tcr信号传导,但额外的共刺激信号也会微调这种应答。例如,程序性死亡1(pd-1)受体和pd-配体(pd-l)途径之间的相互作用抑制t细胞活化(图43a),这是癌症免疫逃避的原因。因此,在一个实施方案中,在t细胞表面上配置α-pd-l1可以特异性地阻断pd-1/pd-l1相互作用以增强t细胞的活化和随后的癌症杀伤作用(图43b)。使用fuct和gf-α-pd-l1有效标记tcr-t细胞(ot-1或p14)(图58a和58b)。之后,进一步证实了α-pd-l1在t细胞表面上的抗原结合能力(图43c和图58c)。如所预期的,标记强度和抗原结合均依赖于gf-α-pd-l1浓度(图43d和图58d)。然后对具有不同修饰的ot-1cd8+t细胞进行离体杀伤测定,其中使用表达卵清蛋白的b6衍生的黑素瘤细胞系b16f10(b16-ova)作为抗原特异性癌细胞。孵育20小时后,与α-pd-l1结合的ot-1t细胞对b16-ova细胞显示出显著增强的细胞毒活性,而同种型rigg标记具有与未标记的细胞相似的结果(图43e)。值得注意的是,另一种不相关特异性的tcr-转基因cd8+t细胞(p14)也与α-pd-l1结合作为阴性对照(图43e),这表明在没有特异性tcr-t的情况下α-pd-l1本身不能诱导杀伤。这种协同杀伤作用表明,与cd8+t细胞结合的α-pd-l1通过阻断pd-1/pd-l1途径改善了tcr特异性细胞毒作用。此外,在不同的效应细胞与靶细胞比例(>1:1)下,可以看到增强的杀伤效果(图43f)。与在约0.1mg/mlgf-α-pd-l1时饱和的抗原结合不同(图43d),增强的杀伤效果在约0.05mg/mlgf-α-pd-l1时达到最大值(图43g),这表明最大标记的仅一半可以有效阻断pd-1/pd-l1途径。
为了进一步证实这些α-pd-l1/cd8+t细胞缀合物可以逃避pd-1/pd-l1共抑制信号传导,发明人测量了当与b16-ova细胞混合时ot-1t细胞的细胞因子分泌。仅在与α-pd-l1结合的t细胞中观察到增强的ifn-γ和tnf-α分泌,其也是α-pd-l1标记强度依赖的(图43h和图59)。阻断共抑制信号传导促进t细胞活化,从而产生更多的细胞因子分泌。这种增强的t细胞活化也可以在显微镜中直接观察到,因为在α-pd-l1标记组中t细胞簇比其他细胞簇大得多(图43i和图60a)。此外,增强的t细胞活化还促进t细胞增殖,这通过用b16-ova再刺激ot-1t细胞的cfse稀释测定证实(图43j和图60b)。在确认α-pd-l1标记可持续多于24小时后(图58e),设计竞争测定以体内测试α-pd-l1/cd8+t细胞缀合物是否比未标记的细胞具有更好的增殖速率。具有同类系标志物(ot-1+/-cd45.1+/-)的ot-1cd8+t细胞与α-pd-l1结合,然后与另一种同类系标志物(ot-1+/-thy1.1+/-)标记的ot-1cd8+t细胞以约为1:1的比例混合。类似地,ot-1+/-thy1.1+/-t细胞与α-pd-l1结合,然后与未修饰的ot-1+/-cd45.1+/-t细胞混合。将这两组ot-1t细胞分别通过尾静脉注射到b16-ova攻击的b6小鼠中。扩增48小时后,分析两组中两种不同同类系标志物标记的ot-1细胞的比例。然而,两组在血液、引流淋巴结(dln)和肿瘤中的比例没有显著差异(图43k),这表明肿瘤微环境中的共抑制信号传导比离体更复杂。
实施例45
在一个实施方案中,如本文所公开的,酶促构建nk-92mi细胞和赫赛汀的缀合物在体内腹膜内癌模型中增强nk细胞的癌杀伤。通过腹膜内注射mda-mb-435/her2+/f-luc细胞建立腹膜内肿瘤模型。在肿瘤接种后第7天和第10天,通过腹膜内注射hbss缓冲液、亲本nk-92mi或nk-92mi-赫赛汀细胞(经辐照的nk-92mi细胞)处理小鼠。通过纵向非侵入性生物发光成像监测肿瘤生长(图61a)。与nk-92mi细胞处理组相比,赫赛汀标记的nk-92mi细胞显著增强了肿瘤的杀伤(图61b、图61c和图61d)。总之,nk-92mi-赫赛汀缀合物在离体和体内均比亲本nk-92mi细胞显著增强了抗肿瘤效果,这显示出可以转化为临床试验的很大潜力。
实施例46
在一个实施方案中,如本文所公开的,癌症相关抗原muc1是一种大的(>250kda)高度糖基化的1型跨膜蛋白,在超过一半的所有癌症病例中被肿瘤细胞过表达和修饰。16a抗体(小鼠)与人muc1反应。在一个实施方案中,通过本文公开的一锅岩藻糖基化方法构建16a-nk-92mi缀合物。仅当用gf-16a和ft处理时,16a与nk-92mi细胞结合,这也是gf-16a浓度依赖性的(图62a和图62b)。乳腺癌细胞系skbr3在细胞表面上具有比mda-mb-435更多的muc-1抗原(图62c和图62d)。用16a修饰的nk-92mi细胞比未修饰的nk-92mi细胞更有效地诱导skbr3细胞的裂解(图62e)。在没有ft的情况下用gf-16a处理或用过量游离的16a(5μg/ml)共处理均不会增强nk-92mi对skbr3细胞的杀伤活性,这表明需要赫赛汀与nk-92mi细胞表面的共价结合(图62e)。相比之下,16a-nk-92mi缀合物对mda-mb-435细胞没有增强的杀伤作用,因为该细胞muc1表达低。在所有杀伤实验中,用0.2mg/mlgf-16a处理nk-92mi细胞。将癌细胞与nk-92mi细胞或其缀合物以1/2的比例混合4小时。该实验证明,用于增强nk-92mi细胞杀伤活性的acc策略可以扩展到除her2之外的其他抗原。
实施例47
在一个实施方案中,如本文所公开的酶促生物大分子转移从岩藻糖基化延伸至唾液酸化(图63a)。其他组已经证明了几种唾液酸转移酶可以将cmp-唾液酸衍生物转移到细胞表面。然而,与如完整igg分子的蛋白质结合的cmp-唾液酸不在这些酶的先前底物范围内。我们首先使用一锅法点击化学合成cmp-唾液酸-叠氮化物-四嗪(cs-az-tz)和cmp-唾液酸-氨基甲酸炔丙酯-四嗪(cs-poc-tz)(图63b)。然后,通过反电子需求狄尔斯-阿尔德反应(iedda)使携带tco部分的小鼠igg2a抗体与cs-az-tz或cs-poc-tz反应,以产生与cmp-唾液酸结合的igg分子(cs-igg)(图63c)。然后将cmp-唾液酸结合的小鼠igg(cs-migg)的一锅法产物在唾液酸转移酶(st)存在下与表达丰富的末端lacnac单元的nk-92mi细胞一起孵育(图63d)。我们测试了三种st,包括重组st6gal1、多杀巴斯德氏菌α(2,3)唾液酸转移酶m144d突变体(pm2,3st-m144d)、美人鱼发光杆菌α(2,6)唾液酸转移酶(pd2,6st)。根据结果,所有这三种酶都可以将cs-migg转移到细胞表面(图63e和图63f)。通常,与cs-poc-igg相比,cs-az-igg是这些st的更有利的底物。st6gal1是这三种酶中最有效的st(图63e和图63f)。总之,st6gal1和cs-az-igg是通过唾液酸化将igg转移至细胞表面的最佳配对(图63e)。该实施例证明了通过聚糖进行细胞表面工程以配置生物大分子也可以使用唾液酸转移酶来实现。使用fuct的本公开中的所有应用将通过st类似地实现。
实施例48
在一个实施方案中,本公开提供了通过针对lacnac的细胞表面糖工程的一步酶促acc构建方法,其快速且易于使用。由于如本文所公开的gdp-fuc衍生物的一锅法合成方案,该方法也是成本有效的。该方法在至少三种不同的免疫细胞中得到证实,包括原代细胞和细胞系,这显示了其一般适用性。此外,该方法中使用的fuct和gdp-fuc修饰的生物分子(例如gf-igg)的试剂在4℃下稳定多于一个月。在一个实施方案中,由于这些有吸引力的性质,该方法将广泛用于细胞表面工程中。在一个实施方案中,在一个步骤中用完整igg分子对细胞表面进行酶促改造是突破性的,因为其显示使用生物大分子(例如酶、蛋白质、病毒甚至细胞)进行细胞表面非基因工程的更多可能性,尤其是在治疗应用中。
尽管由于增殖和内化(半衰期:实验中8小时至24小时),细胞表面结合的mab会被稀释,但这些瞬时修饰可能有很多应用,并且可作为永久性基因工程方法的补充。其他优点,例如同时配置多个mab,使得即时酶促方法在许多情况下比基因工程更具吸引力。在一个实施方案中,发明人已应用本文公开的新方法容易地构建三种acc,其在三个不同阶段:归巢、靶向和信号传导具有增强act的潜力。其中,nk-92mi-赫赛汀缀合物因其在离体和体内杀伤her2+癌细胞而被证明是突出的。由于nk-92mi细胞处于临床试验中并且赫赛汀是商业药物,因此本文公开的nk-92mi-赫赛汀缀合物在临床转化中更容易。此外,nk-92mi细胞可以通过该方法容易地与其他治疗性mab结合,这可以在癌症治疗中广泛地应用。在一个实施方案中,可以使用本文公开的方法容易地组合增加的mab和基于细胞的治疗以获得协同效应。
实施例49
材料和试剂:除非另有说明,否则所有化学试剂和溶剂均购自sigma-aldrich并且无需进一步纯化即可使用。所有细胞培养材料都列在细胞培养方法中。immunocult人cd3/cd28t细胞活化剂购自stemcelltechnologies,inc。重组小鼠ifn-γ、重组小鼠pd-l1-人fc嵌合体(pd-l1-hfc)、cfse细胞分裂示踪试剂盒、apc链霉抗生物素蛋白、抗-小鼠cd3(17a2)-fitc、抗小鼠cd8(53-6.7)-pe、抗小鼠cd45.1(a20)-太平洋蓝、抗小鼠thy1.1(ox7)-alexafluor700、抗小鼠igg(poly4060)-apc、抗人fc(hp6017)-apc、抗大鼠igg(poly4050)-apc、抗his标签(j095g46)-pe、抗人cd3(hit3a)-apc、抗人cd45(hi30)-fitc、抗人cd4(rpa-t4)-pe/cy7、抗人cd8(sk1)-太平洋蓝、抗人cd25(m-a251)-percp/cy5.5、抗人cd44(bj18)-fitc、抗人cd45ro(uchl1)-alexafluor700、抗人cd62l(dreg-56)-pe和人fc受体阻断溶液购自biolegend。包括抗人e-选择素(α-he-sel,cl2)、同种型小鼠igg2a(c1.18.4)、抗小鼠pd-l1(α-pd-l1,10f.9g2)、同种型大鼠igg2b(ltf-2)和抗人egfr(528)的大体积单克隆抗体购自bioxcell。治疗性赫赛汀来自genentech。对照人igg获自athensresearchandtechnology。重组人e-选择素/cd62efc嵌合蛋白(he-选择素-hfc)和重组人tnf-α蛋白购自r&dsystems,inc。具有his标签的重组人her2/erbb2蛋白(her2-his)购自sinobiological,inc。点击试剂,包括生物素-peg3-叠氮化物(az-104,分子量:444.5)、甲基四嗪-peg4-叠氮化物(1014,分子量:389.40)、甲基四嗪-peg4-炔烃(1013-老,分子量:487.5,在它们网站中停产的产品)、tco-peg4-nhs酯(a137,mw:514.6)和cy5-tco(1089,mw:959.20)购自clickchemistrytoolsllc。cy3-叠氮化物(mw:712.8)由xingchen教授实验室(中国北京大学)赠送。(5octdu)-5'-cagtcagtcagtcagtcagt-3'(6-fam)从integrateddnatechnologies,inc订购。d-萤光素(一钠盐)、tnf-α小鼠elisa试剂盒、ifn-γ小鼠elisa试剂盒、颗粒酶b人elisa试剂盒、did'固体、hoechst33342、dapi、alexafluor647nhs酯、细胞示踪剂绿色(cmfda)和橙色(cmtmr)购自thermofisherscientific。cytoselecttm白细胞跨膜迁移测定试剂盒和5×ripa缓冲试剂盒购自cellbiolabs,inc。
设备:所有流式细胞术分析均在attunenxt流式细胞仪上进行。在chemidocxrs+(bio-rad)上拍摄蛋白质凝胶的图像,包括考马斯sds-page凝胶和蛋白质印迹膜。在多模式酶标仪(synergytmh4,bio-tek)中监测吸光度、荧光强度和发光。共聚焦图像在尼康旋转盘共聚焦显微镜(te2000)上拍摄。在all-inone荧光显微镜(keyence,bz-x700)上拍摄荧光和相差显微镜图像。使用ivis光谱系统进行活小鼠的生物发光成像。
细胞:除非另有说明,否则细胞系全部来自atcc。cho细胞系(wt、lec2和lec8,来自pamelastanley教授实验室)在补充有10%胎牛血清(fbs)(omegascientific,inc)的α-最低基础培养基(α-mem)(gibco)中以单层生长。huvec细胞系在含有20%热灭活的fbs(omegascientific,inc)、40μg/ml内皮细胞生长补充剂(corning)、100μg/ml肝素(sigma-aldrich)、50μmβ-me(gibco)和10mmhepes(gibco)的rpmi1640(glutamax,gibco)中培养。将huvec培养皿预先用0.1%明胶溶液(atcc)在4℃下包被过夜。nk-92mi细胞系在myeloculttmh5100(stemcelltechnologies,inc)中生长。包括bt474、skbr3、mda-mb-435(her2+和her2-)、mda-mb-468、skov3和小鼠b16-ova(来自gregoirelauvau教授实验室)的癌细胞系都在补充有10%fbs的dmem(dulbecco改良的eagle培养基,glutamax,gibco)中生长。根据tsri正常献血者服务计划(#irb15-6710)从健康供体收集人血样品。通过ficoll(ficoll-paqueplus,ge)密度离心获得外周血单核细胞(pbmc)。pbmc、活化的人t细胞和小鼠t细胞均在含有10%热灭活的fbs、1mm丙酮酸钠、50μmβ-me、10mmhepes和1×memneaa(gibco)的rpmi1640(glutamax)(下文称为t细胞培养基)中培养。如所指示添加t细胞培养基中的细胞因子(rhil2、rhil7和rhil15均来自nih项目)。将所有细胞培养物在37℃、5%co2下孵育。
小鼠:在无特定病原体(spf)条件下培养或饲养所有小鼠。所有动物实验均经tsri动物护理和使用委员会批准。ot-1小鼠购自taconicbiosciences。来自c57bl/6j(b6)基因背景的cd45.1和thy1.1小鼠购自jackson实验室。通过杂交育种产生ot-1+/-/cd45.1+/-和ot-1+/-/thy1.1+/-的品系。六只雌性b6背景p14小鼠由johnteijaro教授实验室赠送。176只雌性nsg小鼠由philippea.gallay教授实验室赠送。8周龄至12周龄的雄性和雌性小鼠都用于大多数实验。
用于产生gdp-fuc衍生物的一锅法:反应通常在含有5ml100mmhepes缓冲液(ph7.5)的15ml康宁管中进行,该缓冲液含有l-岩藻糖类似物(终浓度10mm)、atp(10mm)、gtp(10mm)、mgso4(10mm)、kcl(50mm)、无机焦磷酸酶(90单位,约0.17g/l,无内毒素)和fkp(9单位,约0.6g/l,无内毒素)。将反应混合物在37℃下振荡(225rpm)孵育5小时至6小时。在反应完成后(通过tlc分析监测),通过向粗产物中加入5ml冷etoh来沉淀酶。通过离心(8000×g,5分钟)除去沉淀物后,可以直接使用粗产物(含有约10mmgdp-fuc类似物)。对于gf-al和gf-az,可以通过cuacc反应进一步修饰。在cu/bttp(1/2,500μm)和抗坏血酸钠(2mm)存在下,粗制gf-al/gf-az样品(5mm,在hepes缓冲液中)与叠氮化物/炔烃探针(5mm)在30℃反应6小时(对于tz底物,在反应混合物中加入一体积的meoh)。反应完成后(通过tlc和lc-ms分析监测),加入bcs(浴铜灵磺酸盐,2mm)以淬灭反应。按照该方案,制备gf-al-生物素、gf-al-cy3、gf-al-tz和gf-az-tz的一锅法产物。它们的结构如图45所示。
将gdp-fuc衍生物酶促转移至细胞表面的一般程序:将活细胞(1000万至2000万)重悬浮于含有20mmmgso4、3mmhepes、0.5%fbs、100μmgdp-fuc衍生物和30mufuct(约0.02mg/ml)的100μlhbss缓冲液中。孵育20分钟后(从在冰上升至37℃),用pbs洗涤细胞,准备进一步应用或分析。对于生物素检测,反应后用apc链霉抗生物素蛋白对细胞进行染色。对于tz检测,将细胞与20μmtco-cy5在冰上反应30分钟并洗涤三次。
gdp-fuc修饰的抗体的制备:首先将用于结合的所有抗体(全长igg,mw:约150kda)脱盐至pbs中并浓缩至6mg/ml溶液。根据tco-peg4-nhs酯的标准标记方案和先前关于它在igg标记上的应用的报道,首先将tco基团引入到抗体上。简而言之,制备新鲜的10mm在dmso中的tco-peg4-nhs试剂储备液,并将其加入到igg样品中(终浓度6mg/ml),终浓度为0.5mm。将反应物在室温下孵育30分钟,并通过加入tris缓冲液(ph8.0)至tris终浓度为50mm来淬灭反应。将淬灭的反应混合物在室温下孵育5分钟,然后使用脱盐柱(ge)脱盐至pbs中。脱盐的tco-igg的浓度为约4.5mg/ml(每个igg约4个tco)。之后,将约10mm的gf-az-tz一锅法产物加入到tco-igg中,终浓度为0.15mm(5当量的igg)。在室温下孵育30分钟后,这些一锅法gf-igg产物即可使用,并可在4℃下保存长达2个月。对于用于共聚焦成像的alexafluor647标记的gf-rigg,首先根据手册用alexifluor647探针标记rigg分子,然后进行该方案。根据参考文献,这些由nhs酯和随后的iedda反应修饰的mab具有与未修饰的mab相似的抗原结合亲和力。
将gf-igg酶促转移至细胞表面的一般程序:将活细胞(1000万至2000万)重悬浮于含有20mmmgso4、3mmhepes、0.5%fbs、0.1mg/ml一锅法gf-igg(与滴定中使用不同浓度)和60mufuct(约0.04mg/ml)的100μlhbss缓冲液中。孵育20分钟后(从在冰上升至37℃),用pbs洗涤细胞,准备进一步应用或分析。对于标记检测,将igg标记的细胞用dapi和针对标记的igg的荧光二抗(1/50至1/200稀释)在冰上染色30分钟。对于共聚焦成像,将在室盖玻片(nunc)上铺展的lec2细胞用0.1mg/mlalexafluor647标记的gf-rigg在与上述相同的岩藻糖基化条件下处理。缀合后,洗涤活细胞并在冰上用hoechst33342染色30分钟,然后洗涤用于成像。为了确认细胞表面缀合抗体的结合活性,在结合缓冲液(含有5mmhepes、2mmcacl2、和1mmmgcl2的hbss)中使标记的细胞与10μg/ml抗原(he-sel-hfc、pd-l1-hfc或her2-his)在冰上结合30分钟。结合后,用pbs洗涤细胞两次,并用dapi和apc-抗人fc(1/50稀释)或pe-抗his(1/100稀释)染色。
原代人t细胞制备及其表面上的igg标记:如上所述新鲜制备新鲜的pbmc。在含有15ng/mlrhil2的t细胞培养基中培养4百万每mlpbmc,并用人cd3/cd28t细胞活化剂活化2天。之后,使活化的人t细胞在t细胞培养基中保持在4*106个细胞/ml(每两天加入含有细胞因子的新鲜培养基)。在扩增两周后表征表型(>95%是人t细胞)。在第0天(初始t细胞)、第2天、第4天、第7天、第11天和第13天通过用gf-生物素的岩藻糖基化追踪cd4+t细胞和cd8+t细胞上的lacnac水平。使用酶促gf-igg转移的一般程序用α-he-sel和migg对照标记活化的人t细胞。之后,标记检测和抗原(he-sel-hfc)结合均得到证实。然后将标记的人t细胞在t细胞培养基中以0.5*106个细胞/ml的起始细胞密度培养。标记后24小时跟踪细胞表面migg分子的衰减(抗小鼠igg染色)。在三天内将标记的t细胞与未标记的人t细胞的细胞增殖率进行比较(活细胞计数)。
e-选择素包被载玻片的流动室测定:将无菌载玻片(目录号#12-545m,fisherscientific)用重组人e-选择素-fc(2μg/ml)在室温下包被2小时,然后用酪蛋白封闭至少30分钟。将ibidi黏性玻片i0.1luer(高:150μm,宽:5mm,ibidi)安置在经包被的载玻片上并与入口管和出口管连接。将两组人t细胞混合(用细胞示踪剂绿色染色的未修饰的人t细胞和用细胞示踪剂橙色染色的migg或α-he-sel缀合的人t细胞),然后通过入口管以2x106个细胞/ml的浓度灌注。出口管连接到注射泵(harvardapparatus)。如前所述测定壁剪切应力(abadier等人,2015,coisne等人,2013)。首先使细胞以1dyn/cm2稳定1分钟以在视场(fov)中积累并开始成像获取。然后将剪切应力增强至10dyn/cm2,持续3分钟。为了研究t细胞的行为,用倒置的zeissaxiovert200m(zeiss,feldbach,瑞士)以10x放大率进行图像采集。通过imagej软件(美国国立卫生研究院,bethesda,md)对牢固结合的细胞(剪切应力增加为10dyn/cm2后2分钟期间未移动的)的数量进行定量。
huvec表面上结合的人t细胞的分析:将2000个huvec接种在明胶包被的96孔板的每个孔中并生长至汇合。huvec未经处理或用200u/mltnf-α刺激4小时。使用通用gf-igg转移方案,用migg或α-he-sel标记经cfse染色的人t细胞。用pbs洗涤标记的细胞并在无血清rpmi1640中重悬至106个细胞/ml。在除去huvec的培养基后,将三组(未标记的、migg标记的和α-he-sel标记的)100μlcfse染色的人t细胞(106个细胞/ml)加入未处理组或tnf-α处理组。使细胞在37℃下附着20分钟。除去未结合的人t细胞,并用温热的rpmi轻柔洗涤每个孔两次。使用ripa缓冲液裂解附着的cfse染色的人t细胞。使用平板读数器通过cfse荧光信号(激发492nm/发射517nm)对每个孔中附着的t细胞数进行定量。对于成像分析,huvec在t细胞结合之前被染色(5μg/mldid和2μg/mlhoechst33342在培养基中,37℃持续20分钟)。在加入cfse染色的t细胞之前,用温pbs洗涤huvec三次。结合后,在活细胞成像溶液中补充附着的细胞,并用荧光显微镜成像。
人t细胞的跨膜迁移测定:按照提供的方案,使用白细胞跨膜迁移测定试剂盒进行迁移(transwell)测定。简而言之,将300μlhuvec生长培养基中的50000个huvec细胞添加到底部具有500μl生长培养基的24孔板的插入物中。使细胞生长至汇合并用200u/mltnf-α处理4小时。将106个/ml人t细胞用leukotracker在无血清培养基(含有0.5%bsa、2mmcacl2和2mmmgcl2的rpmi)中于37℃染色1小时,并用无血清培养基洗涤三次。用migg或α-he-sel标记染色的t细胞,并以106个细胞/ml重悬于无血清培养基中。将transwell插入物移至底部含有500μl新鲜t细胞培养基的新孔中,并用300μlt细胞悬浮液替换插入物中的培养基。在37℃孵育4小时后,裂解底部迁移的t细胞,用平板读数器(激发480nm/发射520nm)测量所得荧光信号。
nk-92mi细胞上的igg标记:根据一般方案,用赫赛汀或对照人igg标记nk-92mi细胞或经辐照的nk-92mi细胞(6gy)。标记后,标记检测和抗原(her2-his)结合均得到证实。然后将有或没有igg结合的nk-92mi细胞在t细胞培养基中以0.5*106个细胞/ml的起始细胞密度培养。在第0天、第1天和第2天追踪细胞表面缀合的赫赛汀的衰减(抗人fc染色)。在第0天、第1天、第2天和第3天追踪细胞的增殖率(dapi染色和facs计数)。对于双抗体标记,首先将nk-92mi细胞与gf-α-egfr缀合,然后在洗涤后与gf-赫赛汀缀合。
nk-92mi和bt474之间结合的流动和成像分析:对于流式细胞术分析,用cfse对nk-92mi细胞进行染色,然后根据一般方案用赫赛汀标记或不用赫赛汀标记。首先用did对bt474细胞进行染色,然后以1:1的比例与经赫赛汀标记的或未经赫赛汀标记的nk-92mi细胞混合。两小时后,通过流式细胞术分析细胞混合物。对于荧光成像,将bt474细胞用cfse染色并在玻璃底培养皿中培养过夜。然后用细胞示踪剂橙色对nk-92mi细胞进行染色,然后用赫赛汀或不用赫赛汀标记。将nk-92mi细胞以1:1的比例加入bt474培养物中。两小时后,在pbs洗涤之前和之后通过荧光显微镜对共培养的细胞成像。
nk-92mi细胞介导的针对her2+癌细胞的细胞毒作用的分析:在指定效应物/靶标比例下,将标记的或未标记的nk-92mi细胞与不同类型的癌细胞在96孔板中共培养4小时。在大多数实验中,效应物/靶标比例是5/1。如果指示,则加入游离赫赛汀,最终浓度为5μg/ml。通过上清液中的ldh分泌检测特异性癌细胞裂解(cytotox96,promega)。根据制造商的说明,进行对照组的设置和特异性裂解的计算。还收集每组的上清液并进行颗粒酶belisa试剂盒测试用于定量。
nk-92mi介导的her2+癌症杀伤的小鼠模型:通过尾静脉注射用5*105个mda-mb-435her2+/f-luc细胞接种的30只雌性nsg小鼠(6周龄至8周龄)。一天后,将小鼠随机分成三组(每组10只小鼠)。通过尾静脉注射(每只小鼠3*106个nk细胞)用hbss、nk-92mi或赫赛汀标记的nk-92mi细胞处理每组。在肿瘤攻击后六天,通过腹膜内注射给小鼠注射200μld-萤光素(15mg/ml)。十二分钟后,通过perkinelmerivis系统分析小鼠的生物发光信号。通过ivis软件对指示肿瘤小鼠的总光子进行定量。
原代ot-1cd8+t细胞制备及其表面上的igg标记:将来自ot-1小鼠的脾细胞用1nmova肽在t细胞培养基中活化两天。之后,活化的细胞在含有15ng/mlrhil2或10ng/mlrhil7和20ng/mlrhil15的新鲜t细胞培养基中体外扩增数天(保持8*106个细胞/ml,每两天添加一次含有细胞因子的新鲜培养基)。表征表型(>95%是ot-1cd8+t细胞)。在第0天(初始t细胞)、第2天、第4天、第7天、第11天和第13天通过用gf-生物素的岩藻糖基化追踪ot-1t细胞(用两种不同的细胞因子培养)的lacnac水平。使用酶促gf-igg转移的一般程序用α-pd-l1和migg对照标记活化的ot-1t细胞。之后,标记检测和抗原(pd-l1-hfc)结合均得到证实。然后将标记的ot-1t细胞在t细胞培养基中以0.5*106个细胞/ml的起始细胞密度培养。标记后24小时跟踪细胞表面α-pd-l1分子的衰减(抗大鼠igg染色)。
使用ova脉冲的脾细胞重新刺激ot-1t细胞:用siinfekl(ova)肽(1μg/ml,在t细胞培养基中)脉冲处理来自b6wt小鼠的脾细胞2小时。之后,在使用前将细胞洗涤三次。具有cd45.1同类系标志物的ot-1t细胞用cfse染色,然后用rigg标记。将105个ot-1t细胞(标记的或未标记的)与106个ova脉冲的脾细胞在500μlt细胞培养基中混合。对照组没有脾细胞。将细胞混合物培养3天。之后,用apc抗cd45.1和dapi对细胞进行染色。针对活cd45.1阳性细胞分析cfse稀释信号。
分析ot-1cd8+t细胞介导的对b16-ova的细胞毒作用:将b16-ova细胞(用萤火虫萤光素酶稳定转导)接种于96孔板中,并用10ng/mlifn-γ过夜处理(在这项工作中,用ifn-γ处理所有b16-ova细胞以诱导高表达水平的pd-1)。在指定效应物/靶标比例下,将标记的或未标记的ot-1细胞与b16-ova癌细胞在96孔板中共培养20小时。在大多数实验中,效应物/靶标比例是5/1。在细胞数量化之前对t细胞簇的表型进行成像。根据参考文献,通过萤光素酶活性对b16-ova细胞数进行定量。根据制造商的手册(bright-glo,promega)将检测试剂直接添加到每个孔中的培养基中。对于细胞因子分泌定量,将细胞培养9小时,收集上清液并进行tnf-α和ifn-γelisa试剂盒测定。对于ot-1t细胞在杀伤b16-ova中的增殖,在igg标记之前,通过cfse对ot-1细胞进行染色。将经修饰的细胞与b16-ova细胞以效应物/靶标比例2/1混合。72小时后,将细胞混合物用apc-抗cd8和dapi染色。针对cd8+细胞分析cfse稀释信号。
用于分析ot-1t细胞针对b16肿瘤的体内增殖的竞争测定:将来自ot-1+/-cd45.1+/-和ot-1+/-thy1.1+/-小鼠的脾细胞用1nmova肽活化,通过加入10ng/mlil7和20ng/mlil15进行体外扩增4天。用或不用gf-α-pd-l1和ft处理具有不同同类系标志物的ot-1t细胞。将用α-pd-l1标记的cd45.1+ot-1t细胞与thy1.1+ot-1t细胞以1:1的比例混合。类似地,将未标记的cd45.1+ot-1t细胞与用α-pd-l1标记的thy1.1+ot-1t细胞以1:1的比例混合。通过流式细胞术分析两种不同群体的比例以及α-pd-l1的成功标记。将混合t细胞(10*106个)通过尾静脉注射到携带b16-ova黑素瘤(肿瘤大小:约5mm*5mm)的小鼠中,其中10天前小鼠通过皮下注射在右侧腹接种106个b16-ova细胞。注射t细胞48小时后,收集血液、引流淋巴结(dln)和受体小鼠的肿瘤组织。获得来自每个组织的单细胞,用抗cd3、抗cd8、抗thy1.1、抗cd45.1荧光抗体染色,并通过流式细胞术分析。计算这些组织中thy1.1+/-ot1t细胞和cd45.1+/-ot-1t细胞的比例。
上述各种方法和技术提供了许多实现本发明的方法。当然,应该理解,根据本文描述的任何特定实施方案,无需实现所描述的所有目标或优点。因此,例如,本领域技术人员将认识到,可以以实现或优化本文所教导的一个优点或一组优点的方式执行所述方法,而不必实现本文可能教导或建议的其他目的或优点。本文提到了各种有利和不利的替代方案。应当理解,一些优选实施方案具体包括一个、另一个或几个有利特征,而其他优选实施方案具体地排除一个、另一个或几个不利特征,而另一些优选实施方案通过包含一个、另一个或几个有利的特征明确地减轻当前的不利特征。
此外,技术人员将认识到来自不同实施方案的各种特征的适用性。类似地,本领域普通技术人员可以混合和匹配上面讨论的各种元素、特征和步骤,以及每个这样的元素、特征或步骤的其他已知等同物,以执行根据本文描述的原理的方法。在各种元素、特征和步骤中,在不同的实施方案中将特别包括其中一些,并且特别排除其他元素、特征和步骤。
尽管已经在某些实施方案和实施例的上下文中公开了本发明,但是本领域技术人员将理解,本发明的实施方案超出了具体公开的实施方案,延伸到其他替换实施方案和/或其用途和修改及其等同物。
在本发明的实施方案中已经公开了许多变型和替代元素。更进一步的变型和替代元素对于本领域技术人员来说是显而易见的。在这些变型中,但不限于这些变型,选择本发明组合物的组成模块,以及可以用本发明组合物诊断、预后或治疗的疾病和其他临床病症。本发明的各种实施方案可以具体包括或排除这些变型或元素中的任何一个。
在一些实施方案中,用于描述和要求保护本发明一些实施方案的表示成分、性质的量如浓度、反应条件等的数字应理解为在一些情况下被术语“约”修饰。因此,在一些实施方案中,书面描述和所附权利要求书中列出的数值参数是近似值,其可以根据特定实施方案旨在获得的所需性质而变化。在一些实施方案中,数值参数应根据报告的有效数字的数和通过应用普通的舍入技术来解释。尽管本发明的一些实施方案阐述的宽范围的数值范围和参数是近似值,但尽可能精确地报告了具体实施例中列出的数值。在本发明的一些实施方案中呈现的数值可能包含必然由于在其各自的测试测量中发现的标准差引起的某些误差。
在一些实施方案中,在描述本发明的特定实施方案的背景以及类似的参考文献中(特别是在以下一些权利要求的上下文中)使用的术语“一种”、“一个”和“该”或不使用数量词可以解释为涵盖单数和复数两者。本文中对数值范围的描述仅旨在用作单独提及落入该范围内的每个单独值的简写方法。除非本文另有说明,否则将各个单独的值并入本说明书中,如同其在本文中单独引用一样。除非本文另有说明或上下文明显矛盾,否则本文所述的所有方法均可以任何合适的顺序进行。关于本文的某些实施方案提供的任何和所有实例或示例性语言(例如“例如”)的使用仅旨在更好地说明本发明,而不是对要求保护的本发明的范围构成限制。说明书中的任何语言都不应被解释为表示对于本发明的实践必不可少的任何未要求保护的要素。
本文公开的本发明的替代元素或实施方案的分组不应解释为限制。每个组成员可以单独地或与该组中的其他成员或本文中找到的其他元素任意组合地提及和要求保护。出于方便和/或可专利性的原因,可以将一个或多于一个组成员包括在组中或从组中删除。当发生任何这样的包含或删除时,本说明书在此被认为包含经修改的组,从而实现所附权利要求中使用的所有马库什组的书面描述。
本文描述了本发明的优选实施方案,包括发明人已知的实施本发明的最佳方式。在阅读前面的描述后,那些优选实施方案的变化对于本领域普通技术人员来说将变得显而易见。预期技术人员可以适当地采用这些变化,并且本发明可以以不同于本文具体描述的方式实施。因此,本发明的许多实施方案包括适用法律所允许的所附权利要求中所述主题的所有修改和等同物。此外,除非本文另有说明或上下文明显矛盾,否则本发明涵盖上述元素的所有可能变体的任何组合。
此外,在整个说明书中已经参考了许多专利和印刷出版物。上文引用的每篇参考文献和印刷出版物均通过引用整体并入本文。
最后,应该理解这里公开的本发明的实施方案是对本发明原理的说明。可以进行在本发明的范围内可以采用的其他修改。因此,作为实施例而非限制,可以根据本文的教导利用本发明的替代配置。因此,本发明的实施例不限于精确地如所示和所述的那些。
- 还没有人留言评论。精彩留言会获得点赞!