异泽兰黄素在制备治疗或预防胶质瘤的药物中的应用的制作方法
本公开一般涉及异泽兰黄素(eupatilin)的新用途,尤其涉及异泽兰黄素在制备治疗或预防胶质瘤的药物中的应用。
背景技术:
脑胶质瘤是由于大脑和脊髓胶质细胞癌变所产生的、最常见的原发性颅脑肿瘤。年发病率约为3~8人/10万人口。如同其他肿瘤(疾病)一样,胶质瘤也是由于先天的遗传高危因素和环境的致癌因素相互作用所导致的。一些已知的遗传疾病,例如神经纤维瘤病(i型)以及结核性硬化疾病等,为脑胶质瘤的遗传易感因素。脑胶质瘤按恶性程度划分为1级(恶性程度最低、预后最好)到4级(恶性程度最高、预后最差)。其中,传统细胞病理学所谓的间变胶质瘤与who的3级相对应;胶质母细胞瘤与who的4级相对应。脑胶质瘤所导致的症状和体征,主要取决其占位效应以及所影响的脑区功能。胶质瘤由于其在空间的“占位”效应(masseffect),可以使患者产生头痛、恶心及呕吐、癫痫、视物模糊等症状。此外,由于其对局部脑组织功能的影响,还可以使患者产生其他的症状。比如,视神经胶质瘤可以导致患者视觉的丧失;脊髓胶质瘤可以使患者产生肢体的疼痛、麻木以及力弱等症状;中央区胶质瘤可以引起患者运动与感觉的障碍;语言区胶质瘤可以引起患者语言表达和理解的困难。胶质瘤由于恶性程度不同,其所产生症状的速度也不同。
脑瘤的治疗方式主要有手术切除、放射治疗和全身性的药物治疗,如果是恶性脑瘤,往往采取多种手段联合治疗的方式。外科手术切除是脑瘤的主要治疗方式,特别是良性脑瘤。放射治疗经常用于那些手术没有办法切除或者手术切除后残余的病灶,也可用于那些不可能接受手术治疗的病人。
脑胶质瘤的治疗药物效果差强人意,存在用药单一、耐药性严峻现象准确。临床上主要用替莫唑胺治疗脑胶质瘤,但发现其存在耐药情况。研究显示,o-6-甲基鸟嘌呤-dna甲基转移酶(mgmt)表达水平升高是导致u251/tr细胞对替莫唑胺耐药的主要机制,替莫唑胺可以通过自身消耗mgmt而改变u251/tr细胞的耐药特性,发挥抗耐药作用。除此之外,耐药机制尚存在非病理性原因。因此,在替莫唑胺化疗之前对非完全切除者进行mgmt检测和手术部位局部水肿程度及乳酸水平评估,来提高治疗肿瘤效果。近年来新兴以单克隆抗体贝伐单抗为代表的药物靶向治疗,但存在治疗成本高的问题。
中药抗肿瘤一直以来都是医学热点,随着科技的发展,中药中的药效成分逐渐成为抑制肿瘤生长的重要手段。柴昌等利用白花丹提取物白花丹素处理u25i胶质瘤细胞后发现白花丹素抑制胶质瘤细胞的增殖和侵袭迁移。促进其凋亡,可能与其升高mir200b、mir200c和mir-203的表达,降低sox2蛋白的表达有关。吴丽等发现黄连素可抑制u251细胞的增殖,并阻滞细胞周期于g2/m期,其抗肿瘤的机制可能与上调p21waf1/cip1、下调cyclind1(细胞周期蛋白d1)表达相关。中药姜黄素可以调控体外培养的人脑胶质瘤细胞shg44的周期进程,诱导bc1-2及caspase8的差异性表达,并且具有显著抑制肿瘤细胞增殖及促凋亡的作用。
异泽兰黄素(eupatilin)是一种有药理活性的类黄酮,从艾蒿(asteraceae)中提取,用于粘膜保护,具有抗炎特性,广泛用于治疗胃炎和消化性溃疡,它对胃粘膜损伤具有抗氧化作用,并能增强受损粘膜的再生。异泽兰黄素被鉴定为具有抗肿瘤作用:异泽兰黄素通过阻断信号转导分子和血管内皮生长因子(vegf)的表达及转录因子3的激活来抑制胃癌细胞中的血管生成。异泽兰黄素可用作人胃癌的化学预防和抗转移剂。异泽兰黄素通过上调p21使细胞周期阻滞在g2/m期来抑制人子宫内膜癌细胞的生长。异泽兰黄素还通过减少mmp-2和vegf信号传导来抑制血管生成介导的人肝细胞转移。
技术实现要素:
鉴于现有技术中脑胶质瘤的治疗药物效果差强人意,存在用药单一、耐药性严峻现象,本发明人通过体外细胞实验和体内裸鼠成瘤实验发现了异泽兰黄素对胶质瘤的治疗作用及机制。
本发明提供了异泽兰黄素或其组合物在制备用于治疗或预防胶质瘤的药物组合物中的应用。其中所述治疗或预防胶质瘤是由异泽兰黄素抑制胶质瘤细胞系的细胞活性实现,或者所述治疗或预防胶质瘤是由异泽兰黄素抑制胶质瘤细胞系的增殖实现,或者所述治疗或预防胶质瘤是由异泽兰黄素抑制肿瘤细胞的周期变化实现,或者所述治疗或预防胶质瘤是由异泽兰黄素抑制肿瘤细胞的细胞迁移实现,或者所述治疗或预防胶质瘤是由异泽兰黄素抑制肿瘤细胞的侵袭性实现。
所述异泽兰黄素或组合物可以口服或非肠道的方式使用,可将活性成分与药物学中可接受的载体一起配合并制剂化而进行使用。在制剂化的情况下,可使用一般使用的填充剂、增量剂、粘合剂、湿润剂、崩解剂及表面活性剂等稀释剂或赋形剂。用于口服使用的固形制剂包括锭剂、丸剂、散剂、颗粒剂、胶囊剂等,这种固形制剂通过在本发明的组合物中添加至少一个以上的赋形剂,例如添加淀粉、碳酸钙、蔗糖、乳糖及/或明胶等来制备。此外,还使用有镁、滑石等润滑剂。用于口服使用的液态制剂有悬浮剂、内服液剂、乳剂及糖浆剂等,除了经常使用的单纯稀释剂的水、液状石蜡以外,还可包括多种赋形剂,例如湿润剂、甜味剂、芳香剂及/或保存剂等。用于非肠道使用的制剂包括可注射的液剂、悬浮剂、乳剂、冻结干燥剂、鼻腔洗涤剂及栓剂。可注射的液剂、悬浮剂、乳剂可通过将水、无水溶剂或悬浮溶剂和活性成分混合而制备,作为无水溶剂和悬浮溶剂可以是丙二醇、聚乙二醇、橄榄油等植物油、油酸乙酯等可注射酯等。作为栓剂的基剂可使用witepsol、macrogol、吐温61(tween61)、可可脂、月桂油、甘油及/或明胶等。在非肠道使用时,可通过静脉注射或肌内注射进行使用。
在本发明的组合物中包括或在用途及方法中使用的异泽兰黄素,以成人女性为基准,可以1天0.0001~100mg/kg,优选地以0.001~10mg/kg的用量使用。可以一天使用一次或分为多次使用。但是,本发明的范围并非限定于上述使用量及使用次数。
所述异泽兰黄素可通过添加药物学上或食品学上可接受的载体、赋形剂或稀释剂等来进行制剂化,关于制剂化的内容可参照remington'spharmaceuticalscience(最新版),mackpublishingcompany,easton,pa等文献。
附图说明
通过阅读参照以下附图所作的对非限制性实施例所作的详细描述,本申请的其它特征、目的和优点将会变得更明显:
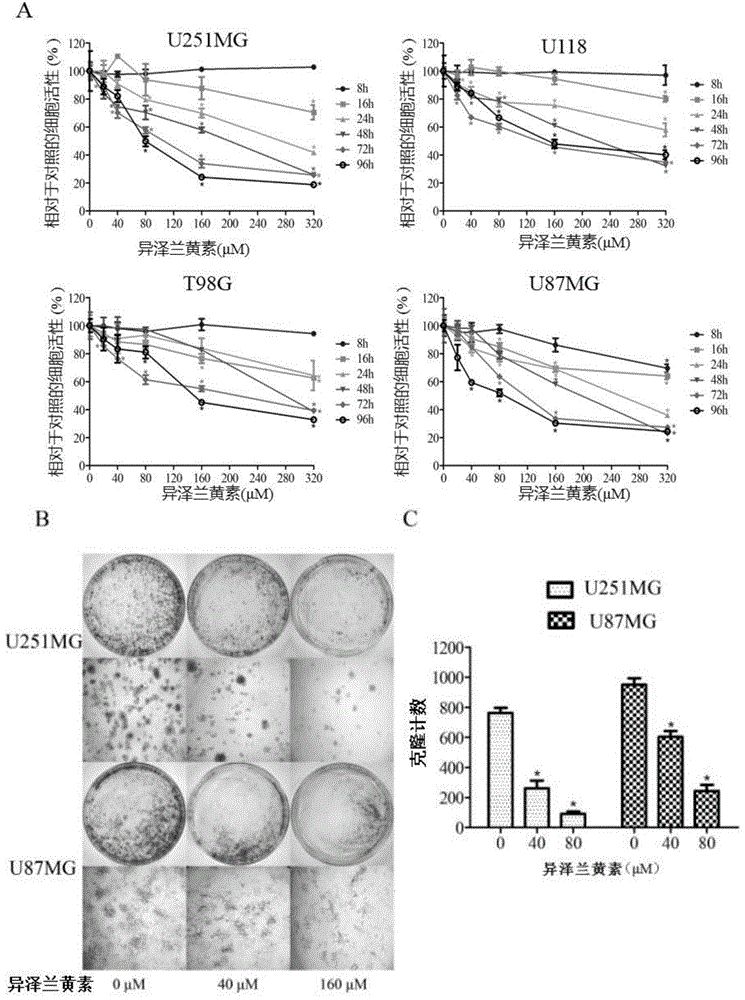
图1(cck8和单克隆)示出了异泽兰黄素抑制胶质瘤细胞系的细胞活性和增殖;
图2示出了异泽兰黄素显著抑制肿瘤细胞的周期变化,从而抑制细胞增殖,其中kinase表示激酶,cyclin表示细胞周期蛋白;
图3示出了异泽兰黄素抑制了细胞迁移(划痕实验)和侵袭(transwell实验);
图4示出了异泽兰黄素抑制了细胞迁移的机理;
图5示出了异泽兰黄素减弱胶质瘤的侵袭性的机理;
图6示出了异泽兰黄素抑制了裸鼠体内肿瘤质量的增加并显示了异泽兰黄素对主要脏器脑肝肾无药物毒性。
具体实施方式
下面结合附图和实施例对本申请作进一步的详细说明。可以理解的是,此处所描述的具体实施例仅仅用于解释相关发明,而非对该发明的限定。另外还需要说明的是,为了便于描述,附图中仅示出了与发明相关的部分。
需要说明的是,在不冲突的情况下,本申请中的实施例及实施例中的特征可以相互组合。下面将参考附图并结合实施例来详细说明本申请。
本发明人根据上述异泽兰黄素对各肿瘤治疗效果的背景,通过体外细胞实验和体内裸鼠成瘤实验发现了异泽兰黄素对胶质瘤的治疗作用及机制。
一、实验方法和试剂
异泽兰黄素和裸鼠
四个胶质瘤细胞系u251mg、u118、t98g、u87mg
1.细胞培养
人胶质母细胞瘤细胞系u251mg、u118、t98g、u87mg购自协和细胞保藏中心。将细胞系在补充有10%胎牛血清的dmem高葡萄糖培养基中培养。将细胞在37℃和5%co2下孵育。用0.05%胰蛋白酶对细胞进行胰蛋白酶消化,并在80%汇合时传代。
2.试剂和抗体
cck-8购自中国上海的东仁化学科技有限公司(目录号ck04)。异泽兰黄素购自中国成都普瑞法科技发展有限公司(目录号bp0576)。matrigel购自corning(目录号356234)。fitcannexinvapoptosisdetectionkitwithpi购自biolegend(目录号640914)。cycledetectionkit(目录号c1052)和dii(目录号c1036)购自beyotime。deoxyribonucleasei,alexafluortm594conjugate购自thermo(目录号d12372)。phalloidin-ifluor488购自abcam(目录号ab176753)。抗cofilin(目录号ab124979),抗profilin(目录号ab124904),抗calreticulin(目录号ab92516)和抗gm130(目录号ab52649)购自abcam。本文中提到的其他抗体购自cst。
3.细胞活力测定。
通过细胞计数试剂盒(cck)-8测定评估细胞活性和增殖。将细胞以每孔2×103个细胞接种到96孔板中,并在37℃下培养24小时以粘附。用不同浓度的异泽兰黄素(0、40、80、160和320μm)处理8h、16h、24h、48h和72h后,向细胞中加入10μlcck-8试剂,在37℃孵育2h。随后,用multiskango的thermoscientificskanitsoftware3.1在450nm处读取光密度(od)值。细胞的存活率=(处理组的od值/对照组的od值)×100%。实验重复三次。
4.流式细胞仪分析细胞周期和凋亡
将所有细胞系(1×106/ml)在6孔板中培养,并用0、40、80或160μm的异泽兰黄素处理24小时。然后收集细胞,用冷pbs洗涤两次,并用70%冷乙醇固定12小时。用pbs洗涤两次后,将细胞与pi染色溶液(50mg/ml碘化丙啶,100mg/mlrnasea)在37℃温育30分钟,并通过流式细胞仪分析。实验重复三次。
5.划痕实验测定
所有细胞系在6孔板中培养。将所得的细胞(70~80%汇合)用新的1ml移液管尖端轻轻地缓慢刮擦一条直线,并且在用pbs洗涤两次后除去被枪头刮离的细胞,加入异泽兰黄素(0、40、80μm)孵育24或48小时。拍摄图像,并使用imagej1.48v软件定量评估间隙距离。实验重复三次。
6.transwell细胞迁移试验
transwell小室用基质胶(用无血清培养基稀释1:8)包被,并将600ml含20%fbs和不同浓度的异泽兰黄素(0、40、80、160μm)的dmem加入下部transwell隔室。用胰蛋白酶处理,洗涤四个细胞系,并以1×105/μl(200μl/孔)的密度接种到上部隔室。将transwell板中的细胞温育24小时。用棉签轻轻除去残留在小室上侧的细胞,用4%甲醇固定小室下侧的细胞,并用0.5%结晶紫染色。用显微镜拍摄小室下侧的细胞照片。随机选择4个不同的视野,并计数细胞数。实验重复三次。
7.蛋白质印迹
用0、40、80或160μm的异泽兰黄素处理24小时后收获u251mg细胞。用冷pbs洗涤细胞两次,并用含有蛋白酶抑制剂的裂解缓冲液在4℃下裂解30分钟。离心后,将上清液在12%的sds-page凝胶(30mg总蛋白/泳道)上分离。将蛋白质转移至pvdf膜,用5%bsa封闭1小时,并与一抗(稀释倍数可在抗体的官方网站上找到)在4℃下孵育过夜。将膜用tbst洗涤三次,然后与山羊抗兔/小鼠二抗(1:10,000)一起温育2小时。将膜再次用tbst洗涤三次,扫描图像并用bio-rad凝胶成像系统分析。
8.免疫荧光
将u251mg细胞接种在12孔板中爬片,用0或160μm的异泽兰黄素处理24小时,用在1×pbs中稀释的4%甲醛固定细胞15分钟。将细胞用1×pbs洗涤3次,每次持续5分钟,并将样品在5%bsa中封闭60分钟,并将一抗在4℃温育过夜。用pbs洗涤后,在室温下用二抗在黑暗中孵育2小时,中性树脂封片后通过共聚焦显微镜观察。实验重复三次。
9.裸鼠中的异种移植肿瘤
从深圳华富康生物科技有限公司(中国深圳)购买总共30只雌性balb/c裸鼠。用pbs将u87mg细胞在对数生长期时重悬浮至2.5×105细胞/l。使用100μl显微注射器将细胞悬浮液(100μl)皮下注射到6只裸鼠的左背。1周后从肿瘤裸鼠中选择肿瘤组织,在无菌条件下麻醉后获得肿瘤组织,将肿瘤组织切成1mm3。用镊子在12只裸鼠左后方接种组织块。接种后7天,成瘤大小一致的裸鼠分为对照组(每日腹腔注射3%dmso+5%tween-80+ns),实验组(每日腹腔注射20mg/kg异泽兰黄素),每组6只小鼠,研究药物疗效。将剩余12只正常无瘤裸鼠分为对照组和双倍药物剂量组,研究药物毒性,每组各6只。每隔一天给老鼠注射,连续30天,每7天计算体重。麻醉裸鼠后,药效组均测定各组肿瘤重量,药物毒性组通过各器官he染色观察损害程度。
10.统计分析
所有数据均以平均值±标准差表示。使用spss17.0软件进行统计学分析。t检验用于两个样本比较。单因素方差分析用于多个样本比较。p值<0.05被认为具有统计学意义。
二、异泽兰黄素对四个胶质瘤细胞系的抑制作用
(一)异泽兰黄素抑制胶质瘤细胞系的细胞活性和增殖
在cck8检测细胞增殖实验中,我们发现给予不同浓度的异泽兰黄素及不同的处理时间对四个胶质瘤细胞系u251mg、u118、t98g、u87mg均有不同程度的抑制作用,在四个细胞系中,以u251mg和u87mg效果较明显(图1a)。
(二)异泽兰黄素抑制胶质瘤细胞系的单克隆能力
平板克隆实验也是测定细胞增殖能力的有效方法,并且能反应细胞群体依赖性和增殖能力两个重要性状,单个肿瘤细胞有无限增殖的能力,对u251mg和u87mg两个细胞系进行计数并取相同细胞量,加入异泽兰黄素至终浓度为160μm,常规条件培养箱孵育15天,每隔5天更换培养基并加药。15天后可见加异泽兰黄素组形成单克隆数量明显低于对照组,且差异具有统计学意义(图1bc),说明异泽兰黄素能抑制u251mg和u87mg的克隆形成能力。
(三)异泽兰黄素显著抑制肿瘤细胞的周期变化
细胞增殖伴随着细胞分裂生长,为了探究异泽兰黄素如何抑制肿瘤细胞增殖,我们检测各个细胞系的周期变化。用不同浓度异泽兰黄素处理相同时间,用流逝细胞仪计数3万个细胞,发现异泽兰黄素能显著抑制肿瘤细胞的周期变化,并且将其抑制于g1期(图2a)。为了进一步探究其机制,我们检测了细胞周期调控蛋白(cyclin)。细胞周期调节蛋白作用模式图见(图2b)。
cycline1与cycline2可联合并激活cdk2,cdk抑制剂p21waf1/cip1和p27kip1的表达上调/激活能够阻止cycline/cdk2激活,从而抑制g1/s。蛋白印迹发现异泽兰黄素能显著降低cycline2,增加p21waf/cip1的表达(图2c)。
进入g1/s检查点开始dna复制时需要周期素e,通过g2/m检查点开始有丝分裂时需要cyclinb,而s期和m期似乎均需要周期a。许多研究描述了过表达周期素a能够加速从g1期进入s期,从而导致dna复制,周期素a反义dna能够阻止dna复制。周期素a显然可用于进入有丝分裂中的一个限速步骤,并且周期素a对完成有丝分裂前期必不可少。在有丝分裂晚前期,周期素a可能不再是必需的,因为cdc2/周期素b1已被激活。组蛋白h3在ser10、ser28和thr11位点上的磷酸化与有丝分裂和减数分裂期间的染色体浓缩紧密相关,影响着dna的合成,异泽兰黄素能显著降低cyclina,p-histoneh3ser10表达(图2c)。使细胞周期停滞在g1期。
有文献报道p70s6激酶是一种丝裂原激活的ser/thr蛋白激酶,这种激酶是细胞生长和g1细胞周期进程所必需的。ser411、thr421和thr424位于假底物结构域的丝氨酸脯氨酸富集区。这些位点的磷酸化被视为可通过解除假底物的抑制而激活p70s6激酶。而p73在thr99位点的磷酸化是细胞周期g1期所必需的。我们发现异泽兰黄素能显著降低p-p70s6kinasethr421/ser424、p-p73tyr99的表达(图2c)。
在g2/m转换阶段激活cdc2/cdk1能够调控所有真核细胞进入有丝分裂的过程。这种激活是一个多步骤过程,从调节亚基和周期素b1结合cdc2/cdk1形成有丝分裂促进因子(mpf)开始。mp保持处于失活状态,直至cdk激活激酶(cak)在thr161位点磷酸化cdc2/cdk1以及cdc25c在thr14/tyr15位点cdc2/cdk1去磷酸化。在tyr15和thr14位点的磷酸化和cdc2的抑制由wee1和myt1蛋白激酶进行。尽管western显示p-wee1ser642、myt1蛋白水平下降,但mpf中的cyclinb1、p-cdc2tyr15变化并无统计学意义(图2c)。说明异泽兰黄素对g2期影响较g1期弱,并且在流式实验中未观察到g2期明显变化。
(四)异泽兰黄素抑制了细胞迁移和胶质瘤的侵袭性
迁移和侵袭是肿瘤细胞是否恶性的两大特征,本发明人研究了异泽兰黄素对细胞迁移和胶质瘤的侵袭性的影响。
1.异泽兰黄素抑制了细胞迁移
划痕实验是研究细胞迁移的经典实验,对四个胶质瘤细胞系u251mg、u118、t98g、u87mg进行划痕实验,在细胞长满六孔板后用宽度一致的枪头进行划痕,分别用40μm和80μm浓度的异泽兰黄素处理细胞,24小时和48小时时在显微镜下拍照,计算划痕两侧爬行细胞的距离,发现异泽兰黄素对细胞迁移具有明显的抑制作用,效果呈药物浓度依赖性(图3a),除u87mg细胞在24小时后迁移的距离和对照组相比无统计学意义之外,其迁移距离抑制率和对照组相比均有统计意义(图3b),考虑和u87mg的生长特性有关。在transwell侵袭实验中,加异泽兰黄素组在24小时候能显著抑制各个细胞系穿过小室的能力,图3c为u251mgtranswell小室结晶紫染色后的细胞数量图,通过显微镜下对小室四个视野进行细胞计数并行统计分析,图3d显示异泽兰黄素抑制各个细胞系的侵袭能力且呈浓度依赖性变化。
细胞迁移作为细胞的一个正常生理活动,是表征细胞生理变化的一个重要指标,也用来检测贴壁生长肿瘤细胞的侵袭转移能力。用160μm浓度的异泽兰黄素处理48小时后,观测到u251mg细胞形态变得更加细长(图4a),细胞骨架在维持细胞形态和细胞迁移运动过程中发挥重要作用,因此我们猜测异泽兰黄素是否通过影响细胞骨架蛋白来影响迁移。
利用带phalloidin-ifluor488试剂对f-actin进行染色,用共聚焦显微镜进行观察,发现正常肿瘤细胞的微丝呈现出完整的丝状,而加异泽兰黄素组的微丝断裂聚体在细胞内(图4b)。为了排除对照组共聚焦拍摄后叠加图层导致断裂的微丝片段遮挡的可能性,对两组各个0.4μm厚度的照片进行观察,对照组未发现有断裂的微丝(图4b)。
高尔基体作为细胞内重要的细胞器,承担着蛋白质的加工和分泌,细胞内物质的运输和细胞骨架密切相关,我们利用免疫荧光检测了高尔基的标志蛋白gm130,发现对照组的高尔基体在细胞内散在分布,而实验组的高尔基体从微丝上脱落在细胞内聚集(图4c)。我们也检测了内质网在细胞内的变化,未发现明显变化(结果未给出),我们猜测内质网连接着核膜,核膜对其在细胞内的定位较微丝影响大。actin在细胞内以游离单体g-actin和线性聚合物f-actin的形式存在,f-actin是微丝的主要组成部分。免疫荧光检测160μm异泽兰黄素处理24小时发现g-actin的含量在加药组上升(图4d),且两组光密度差异具有统计学意义(图4e)。蛋白印迹检测发现实验组p-limk1thr508/p-limk2thr505和p-cofilinser3含量降低(图4f)。因此,我们猜测异泽兰黄素可能影响微丝的解离和聚合。肌动蛋白丝的动态聚合和解聚过程是一个受内外信号转导活动调节的过程,对细胞运动(免疫细胞功能、迁移、浸润、转移和血管生成)、细胞分裂和黏附都非常重要。profilin是保守的肌动蛋白结合蛋白,它通过结合肌动蛋白单体并促进adp交换成atp来影响肌动蛋白的聚合速率。lim激酶(limk1和limk2)是在其氨基末端调节结构域具有两个锌指基序(称作lim基序)的丝氨酸/苏氨酸激酶。lim激酶在rho家族gtpase、pak和rock的肌动蛋白细胞骨架调节下游起作用。在活化环中的保守thr508或thr505残基位点处,pak1和rock将limk1或limk2磷酸化来提高limk的活性。被激活的lim激酶使丝切蛋白cofilin在其氨基末端ser3残基位点被磷酸化,从而来抑制它的肌动蛋白解聚活性。分别对u251mg细胞系用160μm浓度异泽兰黄素处理5分钟、10分钟、20分钟、30分钟、60分钟,发现profilin变化差异不明显,但p-limk1thr508/p-limk2thr505、p-cofilinser3、总cofilin的含量均有不同程度的下降(图4f),且相比较总cofilin的含量,p-cofilinser3下降程度更明显,统计分析cofilin/p-cofilinser3的比值增大(图4g),文献报道cofilin在ser3位点的磷酸化还能调控丝切蛋白从胞核转位到细胞浆中的过程,进一步探究了异泽兰黄素会否会影响cofilin的定位,western显示总cofilin蛋白下降,但是免疫荧光证实细胞核内cofilin的含量下降,但是胞浆内的含量确上升(图4h)。而p-cofilinser3在核内和胞浆均呈下降趋势(图4i)。说明异泽兰黄素通过影响p-limk/cofilin的途径导致细胞骨架微丝的破坏,从而影响肿瘤细胞的迁移。
2.异泽兰黄素减弱胶质瘤的侵袭性
transwell小室实验显示异泽兰黄素能抑制胶质瘤侵袭能力,dil染色细胞膜发现加药组细胞之间细胞膜连接减弱(图5a)。紧密连接或紧密闭合蛋白可在上皮细胞和内皮细胞之间形成一个连续的液体屏障,紧密连接由密封蛋白和闭合蛋白组成,并可与细胞骨架连接。claudin家族包括23个整合膜蛋白以及它们的表达,其表达在各组织类型之间各具差异,且可决定上皮细胞屏障的强度和特性,在u251mg细胞系中,claudin-1加药后48小时并未发现明显改变(图5b)。提示异泽兰黄素未影响细胞之间的紧密连接。
上皮间质转换是肿瘤由良性向恶性转化的重要经典特征之一,我们探究异泽兰黄素是否通过影响上皮间质转化抑制胶质瘤的侵袭性。经典的钙黏着蛋白亚家族包括n-、p-、r-、b-和e-钙黏着蛋白(cadherin),以及约十个在黏附连接中存在的其他成员,黏附连接是一种在极化上皮细胞顶面的细胞结构。经典钙黏着蛋白的胞质结构域可与β-联蛋白、γ-联蛋白(也称作斑珠蛋白)和p120联蛋白相互作用。β-联蛋白和γ-联蛋白与α-联蛋白结合,并且α-联蛋白将钙黏着蛋白-联蛋白复合体连接至肌动蛋白细胞骨架。n-cadherin与fgf受体协作,导致mmp-9过表达和细胞侵袭。snail是一种锌指转录因子,可抑制e-cadherin转录。slug(snai2)是一种广泛表达的转录阻遏蛋白,也是锌指转录因子snail家族的一员。与相关snail蛋白一样,slug结合e-cadherin启动子区来抑制发育期间的转录。癌细胞的上皮钙粘素(e-cadherin)丢失,神经钙粘素(n-cadherin)表达上调的这种变化被称作“钙粘素转换/上皮间质转换”。我们发现加药后转录阻遏蛋白slug和其家族成员snail表达均上调,同时e-cadherin和n-cadherin的表达下降(图5b),破坏了肿瘤细胞钙黏素转换的过程。作为转录阻遏因子,上调的slug和其家族成员snail是否通过降低了其他蛋白的表达降低肿瘤的侵袭性有待研究。
reck是一个gpi锚定膜糖蛋白,可负向调控基质金属蛋白酶(mmp)家族成员,并发挥转化抑制因子的作用,mmp是一个蛋白酶家族,可靶向许多胞外蛋白,包括其他蛋白酶、生长因子、细胞表面受体以及黏附分子。在这些家族成员中,mmp-2、mmp-3、mmp-7和mmp-9被视为胚胎发育过程、损伤修复、癌症侵袭、血管生成、致癌作用和细胞凋亡中正常组织重构的重要因子。研究已证明,mmp活性与癌症进展有关。reck负向调节mmp家族,虽然异泽兰黄素处理后reck表达下调,但是mmp-2表达也下降,而mmp-9表达未明显变化(图5b)。显示异泽兰黄素不仅影响reck本身的表达,也阻止reck的调节作用,从而减弱胶质瘤的侵袭性。
(五)异泽兰黄素对体内肿瘤体积的影响
为了进一步探究异泽兰黄素对体内肿瘤的影响,我们选择4周龄裸鼠皮下种植u87mg肿瘤,一周后选择成瘤大小一致约米粒大的裸鼠,随机分成两组,实验组腹腔注射按体重给予20mg/kg计量的异泽兰黄素。对照组于相同计量的溶解异泽兰黄素的溶剂(双蒸水+dmso+tween-80)。每两天给药,每7天称重,按重量调节给药计量,给药一个月后麻醉处死裸鼠,实验组裸鼠体内的肿瘤体积明显小于对照组(图6a),且两组在一个月内的体重均有增长,差别无统计意义(图6b)。取出体内肿瘤称重两组差异有统计学意义(图6cd)。为了验证异泽兰黄素对裸鼠有无药物毒性作用,予双倍药物计量和对照组同步给无瘤裸鼠注射,一个月后取裸鼠脑、肝、肾组织做he染色,发现无药物毒性(图6e)。
现在临床上治疗胶质瘤以手术和放疗为主。药物治疗效果很不好。其中,最常用的是替莫唑胺,但是其效果也不是很理想,而且耐药情况越来越多。本发明发现异泽兰黄素对胶质瘤的治疗和预防效果看来要优于替莫唑胺,异泽兰黄素对胶质瘤细胞的作用从实验现象来看是非常明显的,而且与其他治疗胶质瘤的药物比较,异泽兰黄素的机制更加直接,比如周期影响增殖,细胞骨架影响迁移侵袭,所以治疗和预防的效果较好。而其他的一些药物研究机制大多是一些明星间接信号通路,信号分子位于信号传导网的交叉部位,上下游的调控分子还有很多,难于研究其真正的药物作用机制。
综上所述,异泽兰黄素作为一种药理活性的类黄酮,通过阻止细胞周期至g1期抑制胶质瘤的增殖,单克隆;通过破坏细胞骨架、调节mmp家族蛋白、上皮间质转化过程而抑制胶质瘤细胞系的侵袭和迁移;并且在异位移植裸鼠体内也有较明显的抑制胶质瘤的功效,为临床上治疗胶质瘤用药单一,耐药等严峻现象展现出了新的曙光。
以上描述仅为本申请的较佳实施例以及对所运用技术原理的说明。本领域技术人员应当理解,本申请中所涉及的发明范围,并不限于上述技术特征的特定组合而成的技术方案,同时也应涵盖在不脱离所述发明构思的情况下,由上述技术特征或其等同特征进行任意组合而形成的其它技术方案。例如上述特征与本申请中公开的(但不限于)具有类似功能的技术特征进行互相替换而形成的技术方案。
- 还没有人留言评论。精彩留言会获得点赞!