用于抑制衰老、预防、改善或治疗衰老相关疾病或症状、或者延长寿命的组合物的制作方法
本发明涉及以rubicon为靶标的用于抑制衰老、预防、改善或治疗衰老相关疾病或症状、或者延长寿命的组合物。另外,本发明涉及受检化合物具有抑制衰老的活性、预防、改善或治疗衰老相关疾病或症状的活性、或者延长寿命的活性的可能性的评价方法。进而,本发明涉及检查衰老的程度、衰老相关疾病或症状的发病风险、或者寿命缩短的风险的方法、以及用于该检查的分子。
背景技术:
巨自噬(以下仅简称为“自噬”)是进化上保守的细胞内膜运输过程,其中,被称为自噬体的双层膜结构将细胞质材料隔离并与溶酶体融合,其内容物被分解。自噬最初被描述为本体分解系统,但已经清楚凝集蛋白质、脂质、受损细胞器以及侵入细菌也选择性地成为靶标。通过广泛分解靶标,自噬维持细胞稳态。其结果是,自噬的功能障碍与包括癌症、神经变性疾病和代谢病症在内的许多人类疾病相关。
最近的证据表明,自噬也与动物的衰老有关。在许多物种中,自噬活性随着衰老而降低(非专利文献1~3)。秀丽隐杆线虫(c.elegans)的研究引导发现了包括胰岛素/igf-1信号传导的轻度减少、热量限制、生殖细胞系列的去除、线粒体呼吸的减少和tor信号传导的减少在内的多个保守的延长寿命途径。重要的是,所有这些干预都激活自噬,并且以依赖于该激活的自噬的方式延长动物的寿命。并且,这提示自噬是许多延长寿命途径的趋同性机制(非专利文献4、5)。
然而,关于这种自噬活性与衰老的关系通过何种机制成立,其详细情况尚不明确。
现有技术文件
非专利文献
非专利文献1:chang,j.t.etal.,elife6,18459(2017).
非专利文献2:uddin,m.n.etal.,age(dordr)34,75-85(2012).
非专利文献3:donati,a.etal.,jgerontolabiolscimedsci56,b375-383(2001)
非专利文献4:toth,m.l.etal.autophagy4,330-338(2008)
非专利文献5:madeo,f.etal,jclininvest125,85-93(2015)
非专利文献6:matsunaga,k.etal.,naturecellbiology11,385-396(2009)
非专利文献7:zhong,y.etal.,natcellbiol11,468-476(2009)
技术实现要素:
本发明要解决的课题
本发明是鉴于上述情况而完成的,其目的在于鉴定成为自噬活性与衰老的关系的基础的分子。另外,本发明的目的在于以该分子为靶标,进行衰老的抑制、衰老相关疾病或症状的预防、改善或治疗、或者寿命的延长。本发明的进一步目的在于以该分子为靶标,进行衰老的程度、衰老相关疾病或症状的发病风险、或寿命缩短的风险的检查。
用于解决课题的手段
本发明者们迄今为止鉴定了抑制自噬体-溶酶体融合过程的、作为自噬的负的控制因子的rubicon(rundomainbeclin-1interactingandcysteine-richcontainingprotein)(非专利文献6、7)。因此,本发明者们验证了rubicon是否参与以自噬活性与衰老的关系为基础的机制。
结果发现,rubicon表达在蠕虫和小鼠组织中随着衰老而增加。这表明通过rubicon年龄依赖性地增加而自噬经时地减弱,从而缩短动物的健康寿命。这种想法从通过rubicon的敲低(knockdown)而蠕虫和苍蝇的寿命延长、多种年龄相关表型改善得到了证实。通过组织特异性实验还判明,神经元中rubicon的敲低对寿命具有最大的效果。此外,在rubicon敲除小鼠中,均为主要的按年龄发病的表型的肾间质性纤维化症和α-突触核蛋白的脑内积聚都减少。此外,rubicon表达在蠕虫和小鼠两者中均被包括热量限制在内的多种寿命延长条件抑,。
此外,本发明人发现,rubicon作用于mml-1/mondo的下游,mml-1/mondo是涉及多个寿命范式的趋同转录因子。此外,上位分析表明,两种主要的自噬的负调节因子mtor和rubicon彼此部分独立地控制寿命。
由以上本发明人发现,rubicon是通过抑制自噬活性而成为衰老的关键的分子,以rubicon为靶标,能够抑制衰老、预防、改善或治疗衰老相关疾病或症状,或者延长寿命,能够检查衰老的程度、衰老相关疾病或症状的发病风险、或寿命缩短的风险,从而完成了本发明。
本发明更详细地提供以下内容。
[1]一种用于抑制对象的衰老、预防、改善或治疗衰老相关疾病或症状、或者延长寿命的组合物,以抑制rubicon基因的功能的分子作为有效成分。
[2]根据[1]所述的组合物,其中,抑制rubicon基因的功能的分子是下述(a)~(c)的任一项所记载的分子,
(a)与rubicon基因的转录产物结合的rna或编码该rna的dna,
(b)与rubicon蛋白质结合的rna或dna,
(c)与rubicon蛋白质结合的抗体。
[3]根据[1]或[2]所述的组合物,其中,衰老相关疾病或症状是代谢性骨病或其症状、应激耐受性的提高或器官的纤维化。
[4]根据[1]~[3]的任一项所述的组合物,其中,对象是脊椎动物。
[5]一种评价受检化合物具有抑制对象的衰老的活性、预防、改善或治疗衰老相关疾病或症状的活性、或者延长寿命的活性的可能性的方法,包括
使受检化合物与rubicon蛋白质接触的工序、和
检测受检化合物与rubicon蛋白质是否结合的工序,
其中,
当受检化合物与所述rubicon蛋白质结合时,评价为存在受检化合物具有抑制衰老的活性、预防、改善或治疗衰老相关疾病或症状的活性、或者延长寿命的活性的可能性。
[6]一种评价受检化合物具有抑制对象的衰老的活性、预防、改善或治疗衰老相关疾病或症状的活性、或者延长寿命的活性的可能性的方法,包括
使受检化合物与表达rubicon基因的细胞接触的工序、和
检测该细胞内的rubicon基因的表达的工序,
其中,
当与不使受检化合物接触的情况相比,受检化合物使rubicon基因的表达减少时,评价为存在受检化合物具有抑制衰老的活性、预防、改善或治疗衰老相关疾病或症状的活性、或者延长寿命的活性的可能性。
[7]一种评价受检化合物具有抑制对象的衰老的活性、预防、改善或治疗衰老相关疾病或症状的活性、或者延长寿命的活性的可能性的方法,包括
使受检化合物与具有在rubicon基因的启动子区的下游功能性地结合了报道基因的dna的细胞接触的工序、和
检测该细胞内的所述报道基因的表达的工序,
其中,
当与不使受检化合物接触的情况相比,受检化合物使所述报道基因的表达减少时,评价为存在受检化合物具有抑制衰老的活性、预防、改善或治疗衰老相关疾病或症状的活性、或者延长寿命的活性的可能性。
[8]一种检查对象的衰老的程度、衰老相关疾病或症状的发病风险、或者寿命缩短的风险的方法,
包括检测受检试样中的rubicon基因的表达产物的工序,
当rubicon基因的表达产物的量与对照相比高时,评价为衰老的程度、衰老相关疾病或症状的发病风险、或者寿命缩短的风险高。
[9]根据[5]~[8]的任一项所述的方法,其中,衰老相关疾病或症状是代谢性骨病或其症状、应激耐受性的提高或器官的纤维化。
[10]根据[5]~[9]的任一项所述的方法,其中,对象是脊椎动物。
[11]在[8]~[10]的任一项所述的方法中用于检测rubicon基因的表达产物的组合物,包含下述(a)或(b),
(a)与rubicon基因的转录产物结合的寡核苷酸引物或寡核苷酸探针,
(b)与rubicon蛋白质结合的抗体。
发明的效果
根据本发明,通过抑制rubicon基因的功能,能够介由自噬的激活而抑制衰老、预防、改善或治疗衰老相关疾病或症状、或者延长寿命。特别是,极其令人惊讶的是,仅通过抑制单一基因的功能,就能够带来改善成为代谢性骨病的原因的骨密度或骨体积/骨组织体积比的降低和骨形态的变化、减少成为神经系统变性疾病原因的凝集性蛋白质、减少眼睛的变性、以及改善运动功能降低、提高应激耐受性、减轻脏器的纤维化、延长寿命等多种效果。另外,根据本发明,通过检测rubicon基因的表达产物,还能够检查衰老的程度、上述疾病或症状的发病风险、或者寿命缩短的风险。
附图说明
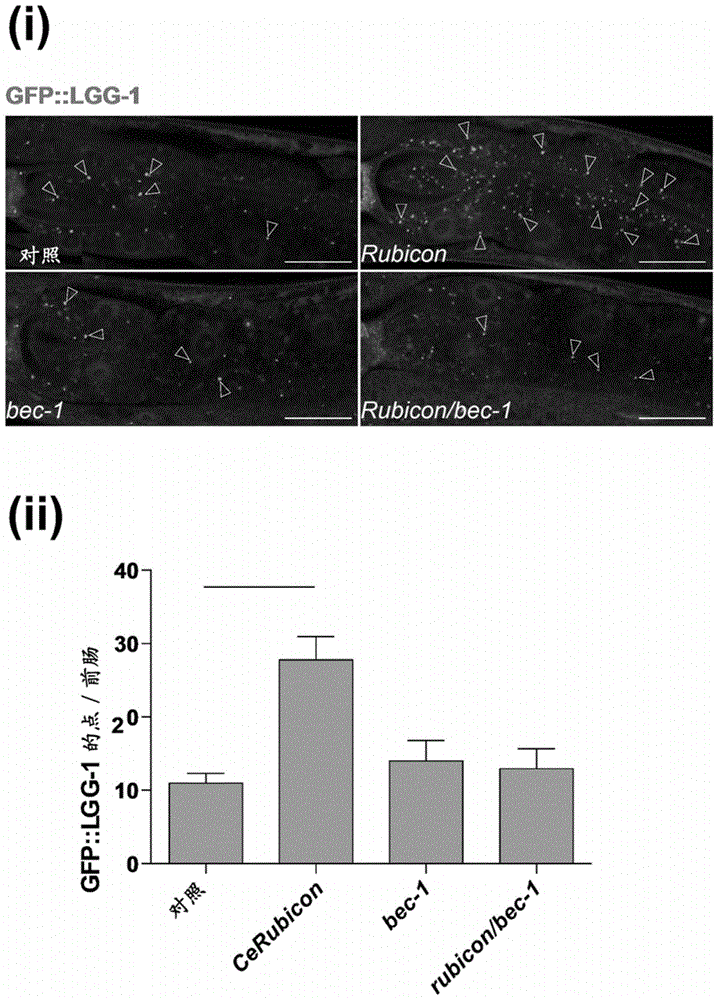
图1a:是显示秀丽隐杆线虫(c.elegans)的rubicon通过调节自噬来控制寿命的图。(i)是l4阶段的前肠的照片。显示了各rnai。被gfp::lgg-1标记的自噬体(轮廓线箭头)在cerubicon的敲低中更丰富,但通过bec-1/beclin1的同时敲低而停止增加。比例尺为20μm。(ii)是将各敲低条件下的前肠中gfp::lgg-1的点定量化而得的图。值表示标本平均±标准误差(n=3)。p值(**p<0.01)通过使用tukey检验的单因素方差分析相对于对照而确定。
图1b:(i)是对照或进行了cerubiconarnai的mah215(mcherry::gfp::lgg-1)株中的肠的代表性照片。自噬流(autophagyflux)通过cerubicon的敲低而增加。通过cerubicon的敲低,自噬体(ap,gfp+mcherry+,由轮廓线箭头表示)和自噬溶酶体(al,gfp-mcherry+,由洋红色轮廓线箭头表示)两者增加。(ii)是将各条件下的ap和al定量化而得的图。p值(***p<0.001,****p<0.0001)通过t检验确定。比例尺为10μm。
图1c:(i)是显示与第1天相比在第7天总荧光增加的、表达cerubicon::egfp的转基因蠕虫的照片。比例尺为50μm。(ii)是显示野生型蠕虫的第1天和第7天的cerubicon表达的qrt-pcr分析的图。显示了来自三个独立的实验的标本平均±标准误差,并相对于野生型n2的第1天的样品进行了归一化。p值(***p<0.001)通过t检验来确定。
图1d:被cerubicon付与的寿命通过(i)bec-1/beclin1或(ii)unc-51/ulk1的同时敲低而消失。作为统计分析进行时序检验(log-ranktest)。
图1e:(i)是显示通过qrt-pcr而在转基因蠕虫中确认了cerubicon的表达增加的结果的图。(ii)是显示通过cerubicon的过表达而野生型蠕虫的寿命缩短的图。
图2a:是显示全身的rubicon敲低改善依赖于年龄的表型的图。在体壁肌中表达polyq35::yfp的转基因蠕虫中,在第5天的阶段,polyq包含体(轮廓线箭头)依赖于年龄而蓄积。通过cerubicon的敲低而包含体的蓄积减少,通过bec-1的同时敲低表型复原。显示了各rnai。比例尺为50μm。
图2b(i)是显示图2a中的每1只蠕虫的polyq包含体的数的图。在各实验中分析了超过20只蠕虫,并且将实验重复3次。p值(*p<0.05,**p<0.01)通过t检验确定。(ii)是显示显示在第5天由多蠕虫追踪分析计算出的运动速度的图。cerubicon的敲低蠕虫维持高运动量。p值(*p<0.05)通过t检验确定。(iii)是显示由cerubicon的敲低引起的自噬依赖性的氧化应激耐受性增加的图。是显示在4.4mmh2o2处理2小时后,进行了指定的rnai的野生型第1天的蠕虫存活率的图。p值(**p<0.01)通过t检验确定。
图2c:是显示drubicon整体大幅延长雌蝇的寿命的图。
图2d:(i)是显示与对照相比,复眼的变性通过表达膨胀polyq蛋白质mjdtr-q78s(gmr-gal4>mjdtr-q78蝇)的蝇的drubicon敲低(drubicon-ir)而减弱的照片。轮廓线箭头表示坏死的小眼。比例尺表示100μm。(ii)是将(i)的gmr-gal4>mjdtr-q78蝇中的坏死的小眼的数定量化而得的图。对于各基因型分析了超过20只眼。p值(***p<0.001)通过t检验确定。
图2e:(i)是显示与2月龄的小鼠相比,20月龄的小鼠肾脏中的rubicon蛋白质水平上升的western印迹(蛋白质印迹)图像。通过来自wt小鼠和rubiconko(ko)小鼠的western印迹(蛋白质印迹)显示rubicon特定的条带。(ii)是将(i)中所示的western印迹(蛋白质印迹)定量化而得的图。
图2f:(i)是显示20月龄的小鼠的肾脏中的lc-3和p62的western印迹(蛋白质印迹)图像。(ii)是显示与wt相比,在rubicon敲除(rubiconko)肾脏中i型胶原阳性区域减少的20月龄的肾脏的免疫组织化学图像。p值(*p<0.05)通过t检验确定。比例尺为100μm。
图2g(i)是将图2f(ii)中的i型胶原阳性区域定量化而得的图。值表示标本平均±标本平均的标准误差(wt:n=5;rubiconko:n=6)。p值(*p<0.05)通过t检验确定。(ii)是显示20月龄的肾脏中的多个纤维化标志物(tgf-b1,col1a1)的qrt-pcr分析结果的图。值表示标本平均±标本平均的标准误差(wt:n=5;rubiconko:n=6)。p值(*p<0.05)通过t检验确定。
图3a:是显示通过降低神经元的rubicon水平而延缓衰老的图。(i)是显示通过神经元cerubicon敲低有效地延长了蠕虫寿命的图。(ii)是显示通过drubicon的神经元敲低而延长了雌蝇的寿命的图。drubicon-ir的转基因表达通过泛神经元elav-gal4驱动基因(driver)诱导。
图3b:(i)是显示通过drubicon的神经元敲低而雌蝇的脑中的kenyon(凯尼恩)细胞层内的polyq(mjdtrq78w)包含体减少了的照片。比例尺是10μm。(ii)是将(i)中的polyq强度定量化而得的图。分析了来自5只动物的kenyon细胞层的双侧层10片。p值(***p<0.001)通过t检验确定。(iii)是显示通过神经元中的drubicon的敲低而雌蝇中的由polyq表达所致的运动功能障碍被改善的图。运动功能通过爬升测定法评价。数据表示标本平均±标准误差。通过使用tukey(图基)检验的双因素方差分析(elav_mjdtrq78w对elav_mjdtrq78w/rubicon-ir)确定了p值(*p<0.05,**p<0.01,***p<0.0001)。
图3c:(i)是显示向小鼠注射10个月后的包含磷酸化α-syn的路易体和路易神经突状包含体(空心箭头)的照片。与对照(rubiconflox/+:nestin-cre)相比,它们在rubicon敲除(rubiconflox/flox:nestin-cre)小鼠中数量较少。比例尺为500μm。(ii)是显示磷酸化α-syn阳性信号的数较少(少于100)、中等(100至200)、或较多(超过200)的小鼠的比例的图。通过卡方检验(rubiconflox/+:nestin-cre,n=7;rubiconflox/flox:nestin-cre,n=6)确定了p值(*p<0.05)。
图4a:是显示rubicon被多种寿命延长条件下调的图。(i)是显示包括daf-2(e1370)、eat-2(ad465)和glp-1(e2141)的多种长寿株中的cerubicon表达的qrt-pcr分析的图。显示来自三个独立实验的标本平均±标本平均的标准误差,并且相对于野生型n2的第1天的样品进行归一化。通过暴露于高温(25℃)两天,诱导不具有生殖细胞谱系的glp-1的表型。p值(*p<0.05,**p<0.01)通过使用tukey检验的单因素方差分析来确定。(ii)是显示与自由(al)进食相比,在进行了9个月的热量限制(cr)的情况下,肾脏中的rubicon蛋白质水平降低的western印迹(蛋白质印迹)图像。
图4b:(i)是显示在进行了指定的rnai处理的指定株的蠕虫中发生了cerubicon表达的qrt-pcr分析的图。如果mml-1或mxl-2被rnai敲低,则glp-1不具有生殖细胞谱系的蠕虫中的cerubicon的抑制消失。相对于野生型n2的第1天的样品,显示了来自三个独立实验的标本平均±标准误差。p值(***p<0.001)通过使用tukey检验的单因素方差分析来确定。(ii)是其显示野生型(n2)和mml-1oe(mml-1过表达株)中的cerubicon的qrt-pcr分析的图。相对于野生型n2的第1天的样品,显示了来自三个独立实验的标本平均±标本平均的标准误差。p值(*p<0.05)通过t检验来确定。
图4c:(i)是显示mondoasirna敲低使rpe-1细胞中的rubicon蛋白质水平显著增加的western印迹(蛋白质印迹)图像。(ii)是显示用po-s归一化后的rubicon的相对值的图(左)。显示使用了指定的sirna的敲低后的mondoa表达的qrt-pcr分析的图(右)。值表示相对于siluc的对照的来自三个独立实验的标本平均±标准误差。p值(***p<0.0001)通过t检验来确定。
图4d:是显示如果进行cerubicon和let-363/mtor的双重敲低,则将它们各自敲低的情况相比,寿命进一步延长的图。
图5a:是显示秀丽隐杆线虫(c.elegans)的rubicon同源物(y56a3a.16)与人kiaa00226(rubicon)、人kiaa0226l和果蝇rubicon(cg12772)类似的、由clustalw分析生成的系统树。
图5b:(i)是显示自噬流通过cerubicon的敲低而增加的、对照或进行了cerubiconrnai的mah215(mcherry::gfp::lgg-1)株的咽肌的代表性照片。通过cerubicon的敲低,自噬体(ap,gfp+mcherry+,由轮廓线白色箭头表示)和自噬溶酶体(al,gfp-mcherry+,由轮廓线洋红色箭头表示)双方都增加。比例尺为20μm。(ii)是将各条件下的ap和al定量化而得的图。p值(**p<0.01)通过t检验确定。
图5c:(i)是显示通过cerubicon的敲低而寿命延长,但通过atg-18的同时敲低而寿命消失的图。(ii)是显示利用rnai的cerubicon和atg-18的敲低效率的qrt-pcr分析的图。显示了相对于具有对照rnai的野生型n2的、来自三次独立实验的标本平均±标本平均的标准误差。
图6a:是显示由rubicon敲低产生的与年龄相关的变化的、多蠕虫追踪分析的快照。将指定的rnai处理下的各个蠕虫的踪迹和速度。
图6b:(i)是显示cerubicon的敲低不改变咽抽吸速率的图。(ii)是显示热应激耐受性的结果的图。判明了cerubicon的敲低不改变耐受性。
图6c:(i)是显示drubicon通过敲低而被大幅下调的qrt-pcr分析的图。p值(****p<0.0001)通过t检验确定(wt:n=3;rubicon-ir:n=4)。(ii)是雄蝇中的drubicon的敲低的个体数统计分析的图。
图6d(i)是显示脑的kc层中的自噬流测定的结果的照片和图。显示drubicon敲低使自噬体(ap,白色箭头)和自噬溶酶体(al,红色箭头)增加了。数值表示标本平均±标本平均的标准误差。p值(**p<0.01)通过t检验(对于对照n=6,对于drub-ir1n=8)确定。比例尺为10μm。(ii)是显示rubicon蛋白质水平在小鼠肝脏中随着衰老而增加(2个月对比20个月)的western印迹(蛋白质印迹)图像。
图7a:是显示通过rubicon的组织特异性作用而有助于寿命控制的图。显示了使用(i)肌肉(nr350)、(ii)皮下组织(nr222)和(iii)肠(vp303)这样的多个组织特异性rnai敏感株的个体统计学分析的图。肠和皮下组织的cerubicon敲低尽管微小,但有意义地延长了寿命。
图7b:(i)神经元atg-18敲低消除了cerubicon的神经元敲低所付与的寿命;(ii)雄性苍蝇中神经元drubicon敲低的个体数统计分析。
图8a:是显示cerubicon的敲低与多个长寿途径之间的上位的关系的图。即使进行了cerubicon的敲低,也没有进一步延长(i)eat-2(热量限制)、(ii)isp-1(线粒体功能障碍)、(iii)daf-2(胰岛素/igf-1信号传导降低)和(iv)glp-1(生殖细胞谱系寿命)的寿命。
图8b:(i)是显示通过6个月的热量限制,肾脏中的rubicon蛋白质水平降低的western印迹(蛋白质印迹)图像。(ii)是显示cerubicon表达的qrt-pcr分析的图。即使敲低mml-1和mxl-2,cerubicon表达也不恢复到野生型(n2)水平。
图8c:(i)是显示即使敲低cerubicon,作为mtor的直接靶标的荧光体s6激酶的水平也不变化的western印迹(蛋白质印迹)图像。(ii)是显示cerubicon不因top敲低而变化的、cerubicon表达的qrt-pcr分析的图。
图9a:(i)显示rubicon缺陷小鼠的股骨的ct图像。比例尺表示500μm。(ii)是显示rubicon缺陷小鼠的股骨中的骨密度(bmd)、骨体积/骨组织体积比(bv/tv)和骨矿物质含量(bmc)的图。野生型(wt)n=6,敲除小鼠(ko)n=5。
图9b:(i)是通过trap染色检测rubicon缺陷小鼠中的破骨细胞的分化的照片。比例尺表示200μm。(ii)是显示trap阳性细胞数的测定结果的图。
图9c:(i)在窝形成测定(pitformationassay)中评价了rubicon缺陷小鼠中的破骨细胞的骨吸收功能。(i)是trap染色图像,(ii)是显示孔中的相对吸收区域的比例的图。
图9d:评价了rubicon缺陷小鼠中的成骨细胞的分化能力。(i)是alp染色图像,(ii)是显示孔中的alp染色区域的比例的图。
图9e:显示rubicon缺陷小鼠中的成骨细胞的矿化能力的评价结果。(i)是茜素染色图像,(ii)是显示孔中的茜素染色区域的比例的图。
图10:是显示通过lc3流测定确认rubicon缺陷小鼠的成骨细胞中的自噬的增强而得的结果的图和电泳图像。
图11a:显示rubicon缺陷小鼠中的骨密度的增加的自噬依赖性的评价结果。(i)是显示对rubicon缺陷小鼠、atg5缺陷小鼠、和rubicon/atg5缺陷小鼠中的骨形态进行ct分析的结果的照片。(ii)是显示各小鼠中的骨密度的图。
图11b:显示rubicon缺陷骨质疏松症模型中的骨密度和骨体积/骨组织体积比的评价结果。(i)是股骨整体的ct图像,(ii)是骨小梁的ct图像,(iii)是显示骨密度和骨体积/骨组织体积比的图。
图12a:是显示为了评价rubicon缺陷小鼠的成骨细胞中的自噬与notch信号通路的关系,而检测notch下游的转录因子和成骨性转录因子的mrna的表达的结果的图。
图12b:是显示rubicon缺陷小鼠的成骨细胞中的notch的分解的检测结果的电泳图像和图。
图13a:显示使rubicon缺陷小鼠中的成骨细胞超表达nicd,通过alp染色来评价其分化能力的结果。(i)是alp染色图像,(ii)是显示孔中的alp染色区域的比例的图。
图13b:是显示通过免疫沉淀法检测nicd与lc3的结合的结果的照片。
具体实施方式
<用于衰老的抑制等的组合物>
本发明提供以抑制rubicon基因的功能的分子作为有效成分、用于抑制对象的衰老、预防、改善或治疗衰老相关疾病或症状、或者延长寿命的组合物。
本发明中的“rubicon”是阻碍自噬体-溶酶体融合过程的自噬的负的控制因子(非专利文献6、7)。本发明中的rubicon与其来源的生物无关(参照图5a)。例如,当本发明的组合物以人为对象时,人的rubicon成为靶标。人的rubicon蛋白质的典型氨基酸序列示于序列号2,编码该蛋白质的cdna的典型碱基序列示于序列号1(数据库登录号:nm_014687.3)。另外,相应的小鼠、鸡、斑马鱼、果蝇、线虫的各rubicon蛋白质的典型氨基酸序列依次示于序列号3~7。
另外,rubicon的序列中可能产生个体差异或突变。因此,本发明中的rubicon蛋白质中,包含由在序列号2~7中任一项所述的氨基酸序列中置换、缺失、插入或添加了一个或多个氨基酸的氨基酸序列构成,并且功能上等同的蛋白质。这里所谓“一个或多个”,通常为20个氨基酸以内(例如,10个氨基酸以内、5个氨基酸以内、3个氨基酸以内、1个氨基酸)。另外,所谓“功能上等同”是指具有负地控制自噬的功能(非专利文献6、7)。
另外,在blastb2.8.0+(altschuls.fnucleicacidsres.25:3389-3402(1997);altschuls.f.etal.,febslj.272:5101-5109(2005))的默认设置下的比较(max分数)中,不同物种之间的rubicon的同一性(相同氨基酸比例)和类似性(类似氨基酸比例)在小鼠和人之间分别为85%和89%(gaps2%),在人和鸡之间分别为73%和81%(gaps2%),在鸡和斑马鱼之间分别为56%和66%(gaps10%),在斑马鱼和果蝇之间的分别为39%和57%(gaps6%),在果蝇和线虫之间分别为29%和47%(gaps6%)。因此,在除上述以外的物种中的rubicon的氨基酸序列与上述物种中的任一rubicon的氨基酸序列之间,通常认为同一性为25%以上(例如30%以上、35%以上、40%以上、45%以上、50%以上、55%以上、60%以上、65%以上、70%以上、75%以上、80%以上、85%以上、90%以上、95%以上),类似性为45%以上(例如50%以上、55%以上、60%以上、65%以上、70%以上、75%以上、80%以上、85%以上、90%以上、95%以上)。因此,本发明中的rubicon蛋白质包含具有与序列号2~7中任一项所示的氨基酸序列具有上述同一性或类似性的氨基酸序列、并且功能等同的蛋白质。在此,“功能上等同”是指具有负地控制自噬的功能(非专利文献6、7)。
本发明中的“衰老”是指随着年龄的增加,在生物的个体中发生的变化。个体可因衰老而发病各种疾病、症状。作为成为本发明的对象的“衰老相关疾病或症状”,只要是起因于rubicon对自噬的负控制,就没有特别限制。例如,可列举出骨质疏松症等代谢性骨病、帕金森病、阿尔茨海默病、多聚谷氨酰胺病(例如,亨廷顿病、脊髓小脑变性症)、肌萎缩性侧索硬化症(als)等神经变性疾病、脏器的纤维化(例如,肾脏的纤维化、心肌的纤维化)、运动功能障碍(例如肌肉力量降低、耐力降低、反应时间延长、运动速度降低、巧妙性降低、深部感觉降低、平衡能力降低)、认知功能降低(例如记忆障碍、失语、失用、失认、执行功能障碍)、骨量或骨密度·骨体积/骨组织体积比降低、骨形态变化、眼睛变性、应激耐受性降低等,但并不限于此。另外,本发明中的“寿命”是指生物从出生到死亡的时间。将日常的、不依赖于持续的医疗、护理、且以自己的身心维持生命、能够自立生活的生存时间特别称为“健康寿命”,本发明中的“寿命延长”包括由健康寿命延长带来的寿命延长。
实际上,在本实施例中,确认了通过抑制rubicon基因的功能而寿命延长,同时,也确认了改善成为代谢性骨病的原因的骨量或骨密度·骨体积/骨组织体积比的降低和骨形态的变化、减少成为神经系统变性疾病的原因的凝集性蛋白质、减少眼睛的变性、以及改善运动功能的降低、提高应激耐受性、减轻脏器的纤维化等多种衰老相关疾病或症状的抑制效果。
关于本发明中的“基因的功能的抑制”,其机制可以是基因的表达的抑制(例如,转录的抑制、翻译的抑制),也可以是基因的翻译产物(蛋白质)的活性的抑制。
作为“抑制rubicon基因功能的分子”,例如,可列举出与rubicon基因的转录产物结合的rna或编码该rna的dna、与rubicon蛋白质结合的rna或dna、与rubicon蛋白质结合的抗体或低分子化合物、编辑rubicon基因或其表达控制区的dna的碱基序列或rubicon基因的转录产物的rna序列的位点特异性核酸酶等。
本发明中的“与rubicon基因的转录产物结合的rna或编码该rna的dna”的一个状态是,与编码rubicon蛋白质的基因的转录产物互补的dsrna(双链rna)或编码该dsrna的dna。
作为本发明中使用的dsrna,优选sirna、shrna(短发夹rna)或mirna。sirna是由在细胞内不显示毒性的范围的短链构成的双链rna。另外,shrna是由正义rna和反义rna配置了间隔序列的单链rna,是指在细胞内等在正义rna与反义rna之间产生氢键,间隔序列形成发夹结构,能够通过该发夹结构在细胞内被切断而形成sirna的rna。mirna一般是指单链rna的形式,但作为具体的实施方式,可以采用双链rna的形式。
dsrna能够抑制靶标基因的表达,并且只要不显示毒性,对其链长就没有特别限制,但通常为15~50个碱基对,优选为15~35个碱基对,进一步优选为20~25个碱基对。通过进一步延长链长,可以提高与mrna的亲和性,减少脱靶效应。
dsrna或编码该dsrna的dna不必与靶标基因的碱基序列完全相同,但与靶标基因的序列具有至少70%以上、优选80%以上、更优选90%以上(例如95%、96%、97%、98%、99%以上)的序列的同一性。
dsrna可以根据rubicon基因的序列信息(例如,序列号1),例如通过将各个链化学合成来制备。编码dsrna的dna包含反义dna和正义dna,其中反义dna编码针对靶标基因的转录产物(mrna)的任一区域的反义rna,正义dna编码mrna任一区域的正义rna,可以通过该反义dna和该正义dna分别使反义rna和正义rna表达。可以由这些反义rna和正义rna制作dsrna。
作为在载体等中保持dsrna的表达系统时的构成,有由同一载体表达反义rna和正义rna的情况,和由不同载体表达反义rna和正义rna的情况。在由同一载体表达反义rna和正义rna的构成中,例如,分别构建在反义dna和正义dna的上游分别连接了能够表达像poliii系统那样的短rna的启动子的反义rna和正义rna表达盒,将这些盒沿相同方向或相反方向插入到载体中。
还可以构建将反义dna和正义dna以在不同链上相对的方式反方向地配置的表达系统。在该构成中,具有反义rna编码链和正义rna编码链成对的一条双链dna,在其两侧以能够由各个链表达反义rna和正义rna的方式对向地具备启动子。在这种情况下,为了避免在正义rna和反义rna的下游添加额外的序列,优选在各个链(反义rna编码链、正义rna编码链)的3’末端分别具备终止子。另外,在该回文型的表达系统中,优选两个启动子的种类不同。
另外,在由不同载体表达反义rna和正义rna的构成中,例如,分别构建在反义dna和正义dna的上游分别连接了能够表达像poliii系统那样的短rna的启动子的反义rna和正义rna表达盒,使不同的载体保持这些盒。
作为本发明中的“与rubicon基因的转录产物结合的rna或编码该rna的dna”的其他状态,可列举与rubicon基因的转录产物互补的反义rna或编码该rna的dna(反义dna)。
反义rna或dna阻碍转录、剪接或翻译过程,从而抑制靶标基因的表达(平岛和井上,“新生化学実験講座2核酸iv遺伝子の複製と発現”,日本生化学会编,东京化学同人,319-347(1993)),本发明中使用的反义rna或反义dna可以通过任一作用抑制靶标基因的表达。
反义rna或反义dna的序列优选为与rubicon的转录产物互补的序列,但只要能有效地阻碍基因的表达即可,可以不完全互补。反义rna或反义dna具有相对于成为靶标的基因的转录产物优选为90%以上(例如,95%、96%、97%、98%、99%以上)的互补性。只要能够抑制靶标基因的表达、且不显示毒性,对其链长就没有特别限制。反义rna或反义dna的长度通常为10~50个碱基,优选12~30个碱基。虽然制造成本增加,但通过使链长更长,能够提高与mrna的亲和性,减少脱靶效应。
反义rna或反义dna可以基于rubicon基因的序列信息,例如通过化学合成来制备。
另外,作为本发明中的“与rubicon基因的转录产物结合的rna或编码该rna的dna”的进一步的其他状态,可以举出具有特异性地切割rubicon基因的转录产物的核酶活性的rna、或编码该rna的dna。核酶中有像i型内含子型、如rnasep所含的m1rna那样400个核苷酸以上的大小的核酶,但也有被称为锤头型、发夹型的具有40个核苷酸左右的活性结构域的核酶(小泉誠和大塚栄子、蛋白質核酸酵素,1990,35卷,2191页)。在本发明中,可以利用任何形式的核酶。
本发明中的“与rubicon蛋白质结合的rna或dna”典型地为核酸适配体。作为核酸适配体,从在生物体内的稳定性高这样的观点出发,优选dna。
作为结构特征,核酸适配体可以包含至少一个环(loop)结构、包含大量脱氧鸟苷(包括鸟苷、鸟苷类似物)的一级结构、或脱氧鸟苷四聚体簇集结构(所谓的“g-四链体”)。此外,核酸适配体可以是由双链构成的核酸和由单链构成的核酸的任一种,优选由单链构成的核酸。
核酸适配体的长度只要具有能够与rubicon蛋白质特异性地结合的碱基数,就没有特别限制,通常为10~100个碱基,优选15~70个碱基,更优选20~50个碱基。虽然制造成本增加,但也可以使用链长更长的核酸适配体。
本发明中使用的核酸适配体,只要是本领域技术人员,就可以适当选择公知的方法来制造。作为公知的方法,例如可以举出体外筛选法(selex法)(tuerkc.&goldl.,science.3;249(4968):505-510(1990)、greenr.etal.,methodscompanmethodsenzymol.2:75-86(1991)、goldl.etal.,annurevbiochem64:763-97(1995)、uphoffk.w.etal.,curr.opin.struct.biol.6:281-288(1996))。
对于用作本发明组合物的有效成分的核酸分子,从提高生物体内的稳定性、增强与靶标(mrna、蛋白质)的亲和性、或抑制脱靶效应等观点出发,可以适当地在其一部分或全部中使用修饰型核酸。例如,在反义核酸中,根据需要,也可以利用仅在两端使用修饰型核酸的gapmer。在双链rna中,例如,也可以将一条链作为gapmer而制成异源双链rna。
核酸的化学修饰包括例如磷酸部分的修饰、糖部分的修饰和碱基部分的修饰。作为这样的修饰,可列举例如。胞嘧啶的5-甲基化、5-氟化、5-溴化、5-碘化、n4-甲基化、胸苷的5-去甲基化、5-氟化、5-溴化、5-碘化、腺嘌呤的n6-甲基化、8-溴化、鸟嘌呤的n2-甲基化、8-溴化、硫代磷酸酯化、甲基膦酸酯化、甲基硫代膦酸酯化、手性甲基膦酸酯化、二硫代磷酸酯化、氨基磷酸酯化、2’-o-甲基化、2’-moe化、2’-ap化、2’-氟化等。另外,还可以利用己糖醇核酸(hna)、环己烯核酸(cena)、肽核酸(pna)、二醇核酸(gna)、苏糖核酸(tna)、吗啉代核酸、三环dna(tcdna)、2’-o-甲基化核酸、2’-moe(2’-o-甲氧基乙基)化核酸、2’-ap(2’-o-氨基丙基)化核酸、2’-氟化核酸、2’f-阿拉伯糖核酸(2’-f-ana)、bna(桥核酸,bridgednucleicacid)这样的修饰核酸。
作为bna,可列举例如,也称为lna(锁核酸,2’,4’-bna)的α-l-亚甲基氧基(4’-ch2-o-2’)bna或β-d-亚甲基氧基(4’-ch2-o-2’)bna、也称为ena的亚乙基氧基(4’-(ch2)2-o-2’)bna、β-d-硫代(4’-ch2-s-2’)bna、氨基氧基(4’-ch2-o-n(r3)-2’)bna、也称为2’,4’-bnanc的氧基氨基(4’-ch2-n(r3)-o-2’)bna、2’,4’-bnacoc、3’-氨基-2’,4’-bna等。
关于核酸的化学修饰,例如参照文献(例如小比贺聪等,日本药理志,148,100~104(2016)、井上贵雄,drugdeliverysystem31-1,10~22(2016))、专利公报(例如,日本特开平10-304889号公报、国际公开第2005/021570号、日本特开平10-195098号公报、日本特表2002-521310号公报、国际公开第2007/143315号、国际公开第2008/043753号、国际公开第2008/029619号、国际公开第2008/049085号)。
本发明中使用的“与rubicon蛋白质结合的抗体”可以是多克隆抗体,也可以是单克隆抗体,还可以是抗体的功能性片段。另外,“抗体”包括免疫球蛋白的所有类和亚类。抗体的“功能性片段”是指特异性地识别rubicon蛋白质的抗体的一部分(部分片段)。具体而言,可以举出fab、fab’、f(ab’)2、可变区片段(fv)、二硫键连接的fv、单链fv(scfv)、sc(fv)2、双特异性抗体、多特异性抗体及它们的聚合物等。
如果抗体是多克隆抗体,则可用抗原(靶标蛋白、其部分肽或表达它们的细胞)免疫动物,并通过常规手段(例如,盐析、离心、透析、柱层析等)从其抗血清中抗体来获得。另外,单克隆抗体可通过杂交瘤法(kohler&milstein,nature,256:495(1975))或重组dna法(例如,p.j.delves,antibodyproduction:essentialtechniques,(1997)、wiley,p.shepherd&c.deanmonoclonalantibodies,oxforduniversitypress(2000)、vandammea.m.etal.,eur.j.biochem.192:767-775(1990))来制作。
与rubicon蛋白质结合的抗体包括嵌合抗体、人源化抗体、人抗体。当将抗体作为治疗剂施用于人时,从减少副作用的观点来看,嵌合抗体、人源化抗体或人抗体是理想的。
“嵌合抗体”是将某种抗体的可变区和与其异源的抗体的恒定区连接而得的抗体。嵌合抗体可以如下获得:例如,将抗原免疫小鼠,从该小鼠单克隆抗体的基因切下与抗原结合的抗体可变区(可变区),与源自人骨髓的抗体恒定区(恒定区)基因结合,将其组装于表达载体中导入宿主中使嵌合抗体产生(例如,日本特开平8-280387号公报、美国专利第4816397号公报、美国专利第4816567号公报、美国专利第5807115号公报)。
“人源化抗体”是将非人来源抗体的抗原结合位点(cdr)的基因序列移植(cdr移植)到人抗体基因中而得抗体,其制备方法是已知的(参照例如,ep239400、ep125023、wo90/07861、wo96/02576)。
“人抗体”是所有区域都源自人的抗体。在人抗体的制作中,可以使用能够通过免疫而产生人抗体库的转基因动物(例如小鼠)。人抗体的制作方法是已知的(例如jakobovits,a.etal.,nature362,255-258(1993)、lonberg,n.&huszar,d.,intern.rev.immunol.,13,65-93(1995)、marksj.d.etal.,jmolbiol.5;222(3):581-97(1991)、mendezm.j.etal.,natgenet.15(2):146-156(1997)、tomizukak.etal.,procnatlacadsciusa.jan18;97(2):722-727(2000)、日本特开平10-146194号公报、日本特开平10-155492号公报、日本特许2938569号公报、日本特开平11-206387号公报、日本特表平8-509612号公报、日本特表平11-505107号公报)。
作为本发明中使用的“用于编辑rubicon基因或其表达控制区的dna的碱基序列或rubicon基因的转录产物的rna序列的位点特异性核酸酶”,只要能够位点特异性地编辑核酸,就不限制,优选包含cas蛋白质作为构成要素的crispr-cas系统。作为crispr-cas系统,可以利用例如,crispr-cas9、crispr-cpf1(cas12a)、crispr-cas12b、crispr-casx(cas12e)、crispr-cas14等。作为位点特异性核酸酶,此外还可以利用talens、zfns、或ppr(与碱基修饰酶融合的形式)等人工核酸酶。
本发明中使用的“与rubicon蛋白质结合的低分子化合物”可以是公知的化合物,也可以是通过后述的本发明的评价方法、筛选方法鉴定的化合物。
本发明的组合物可以是药物组合物、饮食品(包括动物饲料)、或用于研究目的(例如体外或体内实验)的试剂的形式。
药物组合物可以用公知的制剂学方法进行制剂化。例如,可以制成注射剂、栓剂、胶囊剂、片剂、丸剂、液剂、散剂、颗粒剂、细粒剂、薄膜包衣剂、湾(pellet)剂、糖锭(troche)剂、舌下剂、咀嚼剂、口腔剂、糊剂、糖浆剂、悬浮剂、酏剂、乳剂、涂布剂、软膏剂、硬膏剂、敷剂、经皮吸收型制剂、洗剂、吸入剂、气雾剂等,非经口或经口地使用。在这些制剂化中,可以适当地与药理学上允许的载体、具体为灭菌水或生理盐水、植物油、溶剂、基质、乳化剂、悬浮剂、表面活性剂、稳定剂、ph调节剂、调味剂、芳香剂、赋形剂、载体、防腐剂、粘合剂、稀释剂、等张剂、镇痛剂、增量剂、崩解剂、缓冲剂、包衣剂、润滑剂、着色剂、甜味剂、粘稠剂、矫味剂、溶解助剂或其他添加剂等组合。另外,也可以以脂质体递送体系等的形式进行制备。
作为本发明的药物组合物的施用形式没有特别限制,可列举例如,静脉内施用、动脉内施用、腹膜内施用、皮下施用、皮内施用、呼吸道内施用、直肠施用和肌肉内施用、利用输液的施用、直接施用(例如对患部的局部施用)等。
本发明的药物组合物可以与增进医疗上有用的特性的辅助部分结合。作为代表性的有用特性,可列举例如,促进化合物向靶标区域(例如,患部)的递送、在靶标区域维持化合物的治疗浓度、改变化合物的药代动力学和药效学性质、或改善化合物的治疗指数或整体安全特性(safetyprofile)。
作为用于向靶标区域递送的分子,例如,在靶标区域是脑的情况下,可列举脑屏障透过物质。作为脑屏障透过物质,已知例如,抗云铁蛋白受体抗体(例如,国际公开第2016/208695号公报)、源自狂犬病病毒的由29个氨基酸组成的糖蛋白质(参照kumarp.etal.,nature.20075;448(7149):39-43(2007))。此外,根据肾脏、肝脏等靶标区域,可以使用公知的递送用的分子。
另外,本发明的药物组合物可以与用于抑制衰老、预防、改善或治疗与衰老相关疾病或症状、或者延长寿命的其他组合物联合使用。
将本发明的组合物用作饮食品时,该饮食品可以为例如健康食品、功能性食品、特定保健用食品、营养辅助食品、病人用食品、食品添加物或动物用饲料。饮食品除了可以作为如上所述的组合物摄取以外,也可以作为各种饮食品摄取。作为饮食品的具体例,可列举出食用油、调味品、蛋黄酱、人造黄油等含有油分的制品;汤类、乳饮料、清凉饮料水、茶饮料、酒精饮料、饮剂、果冻状饮料、功能性饮料等液状食品;米饭类、面条类、面包类等含碳水化合物的食品;火腿、香肠等畜产加工食品;鱼糕(かまぼこ)、干物(由生的或者盐渍后的鱼、虾、贝类等晒干制成)、盐辛(将鱼、虾、贝类等腌咸制成)等水产加工食品;腌菜等蔬菜加工食品;果冻、酸奶等半固体状食品;酱、发酵饮料等发酵食品;西式糕点类、日式糕点类、糖果类、口香糖类、软糖、冷点、冰点等各种糕点类;咖喱、浇汁、中式煲汤等蒸煮制品;即食汤、即食酱汤等即食食品或微波炉对应食品等。进而,还可列举出调制成粉末、颗粒、片剂、胶囊剂、液状、糊状或胶冻状的健康饮食品。
饮食品的制造可通过该技术领域公知的制造技术来实施。在该饮食品中,可添加对抑制衰老、预防、改善或治疗衰老相关疾病或症状、或者延长寿命有效的1种或2种以上的成分。另外,也可通过与发挥其他功能的成分或其他功能性食品组合,制成多功能性的饮食品。
本发明的组合物可以以包括人在内的动物作为对象而使用。作为除人之外的动物没有特别限制,可以以各种家畜、家禽、宠物、实验用动物等作为对象。动物优选为脊椎动物,可列举例如,猪、牛、马、绵羊、山羊、狗、猫、兔、仓鼠、小鼠、大鼠、猴等哺乳类、鸡、野鸭、鸵鸟、家鸭等鸟类等,但不限于这些。如果是研究目的,则也可以是各种实验用生物。
当施用或摄取本发明的组合物时,可以根据组合物的种类(药物、饮食品等)、对象的类型、年龄、体重、症状、健康状况等适当地选择其施用或摄取的量。此外,本发明的组合物的施用量(有效成分换算量)也根据有效成分的种类不同而不同,例如,当有效成分是rna或dna时,通常为0.1μg~10mg/kg体重,当有效成分是抗体时,通常为1~10mg/kg体重。此外,作为每天施用或摄取的次数,没有特别限制,可以考虑如上所述的各种因素来适当地选择。当有效成分是编码与rubicon基因的转录产物结合的rna的dna时,可以制作并施用表达构建体使得适当量的rna表达。
这样,本发明还提供了抑制对象的衰老、预防、改善或治疗衰老相关疾病或症状、或者延长的寿命的方法,其特征在于,向对象施用或使对象摄取本发明的组合物。
本发明的组合物(药物、饮食品、试剂等)或其说明书可附有用于抑制衰老、预防、改善或治疗衰老相关疾病或症状、或者延长寿命的含义的标示。在此,“在产品或说明书上附有标示”是指在产品的主体、容器、包装等上附有标示,或在公开产品信息的说明书、附件、宣传材料、其它印刷材料等上附有标示。上述标示可包括关于本发明组合物的作用机制的信息。作为这样的机制,可列举例如,通过抑制rubicon基因的功能来促进自噬等。
<受检化合物具有抑制衰老的活性等的可能性的评价>
-以对rubicon蛋白质的结合作为指标的方法-
如下述实施例所示,判明了通过抑制rubicon基因的功能,能够实现改善成为代谢性骨病的原因的骨量或骨密度·骨体积/骨组织体积比的降低及骨形态变化、减少成为神经系统变性疾病的原因的凝集性蛋白质、减少眼睛的变性、和改善运动功能的降低、提高应激耐受性、减轻脏器的纤维化等各种衰老相关疾病或症状改善,延长寿命。因此,作为开发用于抑制衰老、预防、改善或治疗衰老相关疾病或症状、或者延长寿命的组合物的一个步骤,鉴定与rubicon蛋白质结合的化合物是有效的。即,本发明提供评价受检化合物具有抑制对象的衰老的活性、预防、改善或治疗衰老相关疾病或症状的活性、或者延长寿命的活性的可能性的方法作为本发明的评价方法的第一状态,该方法包括:(a)使受检化合物与rubicon蛋白质接触的工序;以及(b)检测受检化合物与rubicon蛋白质是否结合的工序。
作为成为本发明的评价方法的对象的“受检化合物”,没有特别限制,可以列举例如,基因文库的表达产物、合成低分子化合物文库、肽文库、多核苷酸文库、抗体、细菌释放物质、细胞(微生物、植物细胞、动物细胞)的提取液和培养上清、纯化或部分纯化多肽、从海洋生物、植物或动物采集的提取物等。受检化合物也可以是以rubicon蛋白质的立体结构为基础基于生物信息学(insilico)的设计而合成的。
在本发明的评价方法中使用的“rubicon蛋白质”,在人蛋白质的情况下,例如可以举出由序列号2所记载的氨基酸序列组成的蛋白质或其部分肽。rubicon蛋白质除了上述天然型的蛋白质、其部分肽以外,根据需要可以使用其改变体、修饰体等。例如,为了容易检测或纯化,可以使用与其他蛋白质(例如碱性磷酸酶(seap)、β-半乳糖苷酶等酶、绿色荧光蛋白质(gfp)等荧光蛋白质、谷胱甘肽-s-转移酶(gst)等标签)的融合蛋白质。受检化合物与rubicon蛋白质的“接触”可以通过向该评价体系中添加受检化合物等来进行。
“检测受检化合物与rubicon蛋白质是否结合”可适当采用已知的方法,例如,使固定后的rubicon蛋白质与这些受检化合物接触,从而鉴定与rubicon蛋白质结合的化合物的方法。作为检测受检化合物与rubicon蛋白质结合的方法,可利用各种公知的方法,但作为合适的方法的一例,可以举出利用了表面等离子共振现象的生物传感器。
当受检化合物是合成低分子化合物文库时,可以使用例如基于组合化学技术的高通量法(wrightonn.c.etal.,science.26;273(5274)458-464(1996)、verdine,g.l.nature38411-13(1996)、hoganj.c.,jrdirectedcombinatorialchemistry.nature.384:17-19(1996))。此外,当受检化合物是多核苷酸文库时,可以使用例如上述体外筛选方法,当受检化合物是基因文库时,可以使用例如使rubicon蛋白质作为诱饵蛋白质表达并利用的酵母双杂交系统。
根据以上结果,当受检化合物与rubicon蛋白质结合时,评价为存在具有抑制衰老、预防、改善或治疗衰老相关疾病或症状、或者延长寿命的活性的可能性。
-以rubicon基因的表达量的减少作为指标的方法-
如下述的实施例所示,rubicon基因的表达年龄依赖性地上调,提示这是各种衰老相关疾病或症状的发病原因。因此,作为用于开发抑制衰老、预防、改善或治疗衰老相关疾病或症状、或者延长寿命的组合物的一个步骤,鉴定使rubicon基因的表达量减少的化合物是有效的。即,本发明提供评价受检化合物具有抑制对象的衰老、预防、改善或治疗衰老相关疾病或症状。或者延长寿命活性的可能性的方法,作为本发明评价方法的第二状态,所述方法包括:(a)使表达rubicon基因的细胞与受检化合物接触的工序;和(b)检测所述细胞内的rubicon基因的表达的工序。
第二状态中使用的“受检化合物”与第一状态相同。作为成为检测的对象的“rubicon基因”,不特别考虑其来源的生物,但在以人作为对象的组合物的开发中,人rubicon基因是合适的。作为人基因,例如,可以以编码由序列号2所示的氨基酸序列组成的蛋白质的基因(例如,由序列号1所示的碱基序列组成的基因)。此外,作为“表达rubicon基因的细胞”,如果是人细胞,可列举例如rpe-1细胞、hek293细胞、hela细胞等,但不限于此。受检化合物与表达rubicon基因的细胞的“接触”例如,可通过将受检化合物添加到上述细胞的培养液中等来进行。
“rubicon基因表达的检测”可以采用公知的方法。“基因表达的检测”可以在转录水平(mrna水平)检测,也可以在翻译水平(蛋白质水平)检测。作为在转录水平进行检测的方法,可列举例如rt-pcr、rna印迹(northernblotting)、原位(insitu)杂交、斑点印迹(dotblot)法、rna酶保护测定法等。另外,作为在翻译水平进行检测的方法,可列举例如,western印迹(蛋白质印迹)法、放射免疫测定法、化学发光免疫测定法、化学发光酶联免疫测定法、酶联免疫测定法、免疫沉淀法、免疫层析法、免疫组织化学染色法、成像细胞术、流式细胞术、胶乳凝集法、质量分析法(ms)等。
根据以上结果,当与不接触受检化合物时相比,受检化合物使rubicon基因的表达减少时,评价为存在具有抑制衰老、预防、改善或治疗衰老相关疾病或症状、或者延长寿命的活性的可能性。
例如,在采用western印迹(蛋白质印迹)法的情况下,可以通过比较受检化合物存在下检测的rubicon蛋白质的量(例如,来自rubicon蛋白质的条带的强度)和受检化合物不存在下检测的rubicon蛋白质的量(对照值)来评价rubicon基因表达的减少。即,受检化合物存在下的rubicon蛋白质的量与受检化合物不存在下的量相比低(例如,对照值的80%以下、50%以下、30%以下、10%以下的情况)时,可以评价为存在该受检化合物具有抑制衰老的活性、预防、改善或治疗衰老相关疾病或症状的活性、或者延长寿命的活性的可能性。在上述表达的检测中,采用western印迹(蛋白质印迹)法以外的方法时也同样可以施用受检化合物不存在下的上述表达量作为对照值来进行评价。
-报道系统-
受检化合物是否使rubicon基因表达水平减少的评价,也可以在使用报道基因的系统中进行。即,本发明提供评价受检化合物具有抑制对象的衰老、预防、改善或治疗衰老相关疾病或症状、或者延长寿命的活性的可能性的方法,作为本发明评价方法的第三状态,该方法包括:(a)使受检化合物与具有在rubicon基因的启动子区的下游功能性地结合了报道基因的dna的细胞接触的工序;和(b)检测该细胞内的所述报道基因的表达的工序。
“rubicon基因的启动子区”是控制rubicon基因转录的区域,是指转录因子能够结合的、rubicon基因编码区的上游区域。关于“rubicon基因的启动子区”,只要是本领域技术人员,就能够通过以序列号1所示的碱基序列或其一部分作为探针的基因组dna文库的筛选获得。如果在制备在获得的基因组dna的下游连接有报道基因的载体,并导入到适当的细胞内的情况下,能够诱导该报道基因的表达,则可以将该基因组dna鉴定为rubicon基因的启动子区。
作为“报道基因”,只要其表达是可检测的,就没有特别限制,可列举例如,本领域技术人员一般使用的cat基因、lacz基因、萤光素酶基因、β-葡糖醛酸糖苷酶基因(gus)和gfp基因。
在此,在rubicon基因的启动子区下游的“功能性地结合了”报告基因,是指rubicon基因的启动子区与报道基因结合,以使得通过转录因子与rubicon基因的启动子区结合,从而诱导报道基因的表达。
另外,“rubicon基因”、“受检化合物”、和“接触”与上述第二状态相同。作为在第三状态中使用的“细胞”,只要是在导入了该报道基因系统时,通过转录因子与rubicon基因的启动子区结合,从而报道基因的表达被诱导的细胞即可,例如可以举出rpe-1细胞、hek293细胞、hela细胞等,但并不限于这些。
报道基因的表达可以根据所用报道基因的种类,通过本领域技术人员公知的方法检测。例如,如果报道基因是cat基因,则可通过检测由该基因产物产生的氯霉素的乙酰化来检测报道基因的表达。如果报道基因是lacz基因,则可通过检测在该基因表达产物的催化作用下的色素化合物的显色来检测报道基因的表达;如果报道基因是萤光素酶基因,则可通过检测在该基因表达产物的催化作用下的化学发光来检测报道基因的表达;如果报道基因是gus基因,则可通过检测在该基因表达产物的催化作用下的葡萄糖醛酸的发光或5-溴-4-氯-3-吲哚基-β-葡萄糖苷(x-gluc)的显色来检测报道基因的表达;如果报道基因是gfp基因,则可通过检测gfp蛋白产生的荧光来检测报道基因的表达。
根据以上结果,在与不接触受检化合物时相比,受检化合物使上述报道基因的表达减少的情况下,评价为具有抑制衰老的活性、预防、改善或治疗衰老相关疾病或症状的活性、或者延长寿命的活性的可能性。
报道基因的表达的减少,例如,在使用萤光素酶基因作为报道基因的情况下,可以通过比较在受检化合物存在下检测到的化学发光的强度和在受检化合物不存在下检测到的化学发光的强度(对照值)来评价。即,在受检化合物存在下的发光强度比在受检化合物不存在下的发光强度低的情况(例如,对照值的80%以下、50%以下、30%以下、10%以下的情况)下,可以评价存在该受检化合物具有抑制衰老的活性、预防、改善或治疗衰老相关疾病或症状的活性、或者延长寿命的活性的可能性。在前述表达的检测中,使用萤光素酶基因以外的报道基因的情况也同样,可以使用在受检化合物不存在下检测到的前述表达量作为对照值来进行评价。
以上对于本发明的评价方法进行了说明,如果对多个受检化合物实施本发明的评价方法,并以上述的与rubicon蛋白质的结合、rubicon基因的表达的减少、或报道基因的表达的减少为指标来选择化合物,则能够筛选出具有抑制衰老的活性、预防、改善或治疗衰老相关疾病或症状的活性、或者延长寿命的活性的化合物。因此,本发明还提供这样的筛选方法。
对于通过本发明的评价方法、筛选方法而鉴定出的化合物,进一步优选评价细胞中的自噬的促进活性,或在动物实验、临床试验中实际检验是否具有上述活性。作为动物实验,例如,如后述的实施例所示,可列举出在衰老相关疾病或症状的模型动物中的该疾病或症状的改善(例如,成为代谢性骨病的原因的骨量或骨密度·骨体积/骨组织体积比的降低和骨形态的变化的改善、成为神经系统变性疾病的原因的凝集性蛋白质的减少、眼睛的变性的减少、和运动功能的降低的改善、应激耐受性的提高、脏器的纤维化的减轻等)或寿命的延长等。
<衰老的程度等的检查>
如后述实施例所示,rubicon基因的表达年龄依赖性地上调,提示这是各种衰老相关疾病或症状发病、寿命缩短的原因。因此,可以以rubicon基因的表达量为指标,检查衰老的程度、衰老相关疾病或症状的发病风险、或寿命缩短的风险。即,本发明提供检查对象的衰老的程度、衰老相关疾病或症状的发病风险、或寿命缩短的风险的方法,该方法包括检测受检试样中的rubicon基因的表达产物的工序,当rubicon基因的表达产物的量与对照相比高时,评价为衰老的程度、衰老相关疾病或症状的发病风险、或者寿命缩短的风险高。本发明的检查方法也可以表现为提供用于由医生、兽医等的诊断的信息的方法,或者辅助由医生、兽医等的诊断的方法。
本发明的检查方法可以以包括人在内的动物作为对象。作为除人之外的动物没有特别限制,可以以各种家畜、家禽、宠物、实验用动物等作为对象。具体可列举猪、牛、马、绵羊、山羊、狗、猫、兔、仓鼠、小鼠、大鼠、猴等哺乳类、鸡、野鸭、鸵鸟、家鸭等鸟类等,但不限于这些。如果是研究目的,则也可以是各种实验用生物。
本发明的检查方法中的“受检试样”是从检查衰老的程度、衰老相关疾病或症状的发病风险、或者寿命缩短的风险的对象中分离出的生物学试样。生物学试样只要包含rubicon基因的表达产物(转录和/或翻译产物),就没有特别限制,可列举例如,肝脏。肾脏的细胞等。
“rubicon基因的表达产物的检测”可以使用公知的方法。“基因的表达产物的检测”既可以以转录产物(mrna)为对象进行检测,也可以以翻译产物(蛋白质)为对象进行检测。作为以转录产物为对象进行检测的方法,可列举例如,rt-pcr、rna印迹(northernblotting)、原位(insitu)杂交、斑点印迹(dotblot)法、rna酶保护测定法等。作为以翻译产物(蛋白质)为对象进行检测的方法,可列举例如,western印迹(蛋白质印迹)法、放射免疫测定法、化学发光免疫测定法、化学发光酶联免疫测定法、酶联免疫测定法、免疫沉淀法、免疫层析法、免疫组织化学染色法、成像细胞术、流式细胞术、胶乳凝集法、质量分析法(ms)等。
在这样检测rubicon基因的表达产物的结果是受检试样中的rubicon基因的表达产物的量比对照高的情况下,评价为对象衰老的程度、衰老相关疾病或症状的发病风险、或者寿命缩短的风险高。其中“发病风险”不仅包括涉及发病的有无的风险,还包括发病的早期化的风险。作为对照,可以利用例如从未衰老的个体、未发病衰老相关疾病或症状的个体中分离出的试样中的rubicon基因的表达产物的量。
此外,本发明提供用于在上述方法中检测rubicon基因的表达产物的组合物,包含下述(a)或(b),
(a)与rubicon基因的转录产物结合的寡核苷酸引物或寡核苷酸探针,
(b)与rubicon蛋白质结合的抗体。
上述寡核苷酸引物(以下仅称为“引物”),可以基于编码rubicon的cdna的碱基序列信息(例如序列号1),以使得除了rubicon基因的转录产物之外的转录产物尽可能不被扩增的方式设计。这样的引物设计只要是本领域技术人员就能够通过常规方法进行。引物的长度通常为15~50个碱基长,优选15~30个碱基长,但根据方法和目的不同也可以比这更长。
上述寡核苷酸探针(以下仅称为“探针”),可以基于编码rubicon的cdna的碱基序列信息(例如序列号1),以使其尽可能地不与除了rubicon基因的转录产物以外的转录产物结合的方式设计。这样的探针设计只要是本领域技术人员就能够通过常规方法进行。探针的长度通常为15~200个碱基长,优选15~100个碱基长,更优选15~50个碱基长,但根据方法和目的不同也可以更长。另外,引物和探针可以适当标记而使用。
引物和探针例如可以通过市售的寡核苷酸合成仪来制作。探针也可以作为通过限制性酶处理等而获得的双链dna片段而制作。另外,引物和探针也可以仅由天然的核苷酸(脱氧核糖核酸(dna)、核糖核酸(rna))构成,但根据需要,也可以在一部分或全部中使用上述的经化学修饰的核酸。另外,引物和探针可以适当标记而使用。
与rubicon蛋白质结合的抗体在通过直接方法检测rubicon蛋白质的情况下,通常使用结合有标记物质的抗体。另一方面,在通过间接方法检测rubicon蛋白质的情况下,可以与rubicon蛋白质结合的抗体不结合标记物质,而利用结合有标记物质的等进行检测。其中,“二抗”是指对与rubicon蛋白质结合的抗体显示反应性的抗体。例如,在作为小鼠抗体制备与rubicon蛋白质结合的抗体的情况下,可以使用抗小鼠igg抗体作为二抗。针对来自兔、山羊和小鼠等各种物种的抗体,能够使用的标记二抗都有市售,可以根据与rubicon蛋白质结合的抗体所来自的物种来选择和使用适当的二抗。也可以使用结合有标记物质的蛋白g、蛋白a等代替二抗。
作为标记物质,只要能够检测,就没有特别限制,例如可以举出125i、131i、3h、14c、32p、33p、35s等放射性同位素;碱性磷酸酶(alp)、辣根过氧化物酶(hrp)、β-半乳糖苷酶(β-gal)等酶;异硫氰酸荧光素(fitc)、异硫氰酸罗丹明(ritc)等荧光色素;别藻蓝蛋白(apc)、藻红蛋白(r-pe)等荧光蛋白质;亲和素、生物素、金属粒子、胶乳等。
在本发明的组合物中,除了作为有效成分的上述分子以外,根据需要,还可以含有灭菌水、生理盐水、缓冲剂、保存剂等作为试剂所允许的其他成分。
该组合物还可以该检查所需的其他制剂组合而制成试剂盒。作为其他制剂,可列举例如,检测标记所需的底物、阳性对照、阴性对照、或用于样品的稀释或洗涤的缓冲液。另外,在以未标记的抗体作为制剂的情况下,试剂盒可以包括将该抗体结合的物质(例如,二抗、蛋白g、蛋白a等)标记化的试剂。进而,试剂盒可以包含其使用说明书。
实施例
下面,基于实施例更具体地说明本发明,但本发明不限于以下实施例。
[材料和方法]
(1)秀丽隐杆线虫(c.elegans)的饲养条件和株
只要不特别说明,线虫在含有大肠杆菌株op50的线虫饲养培养基(ngm)琼脂平板上,用标准技术于20℃培养。在本实施例中使用下列蠕虫株。
n2(wt);da2123,adis2122[lgg-1p::gfp::lgg-1+rol-6(su1006)];am140,rmis132[unc-54p::q35::yfp];tu3401,sid-1(pk3321)v;uis69[pcfj90(myo-2p::mcherry)+unc-119p::sid-1];vp303,rde-1(ne219)v;kbis7[nhx-2p::rde-1+rol-6(su1006)].nr350,rde-1(ne219)v;kzis20[hlh-1p::rde-1+sur-5p::nls::gfp];nr222,rde-1(ne219)v;kzis9[(pkk1260)lin-26p::nls::gfp+(pkk1253)lin-26p::rde-1+rol-6(su1006)];cb1370,daf-2(e1370)iii;da465,eat-2(ad465)ii;cb4037,glp-1(e2141ts)iii;op198,unc-119(ed3)iii;wgis198[mml-1::ty1::egfp::3xflag(92c12)+unc-119(+)];mq887,isp-1(qm150)iv;mah215,sqis11[lgg-1p::mcherry::gfp::lgg-1+rol-6]。
(2)质粒构建和转基因
对于cerubicon::egfp翻译融合构建物,将cerubicon4kb内源性启动子以及编码序列克隆到含有egfp标签的pdc4载体中。将构建物的显微注射与共注入标记物myo-2p::mcherry一起实施,以产生cerubicon::egfp。
(3)小鼠
cag-cre小鼠和nestin-cre小鼠分别获自ジャクソン研究所和宫崎纯一博士的研究所(大阪大学)。将cag-cre小鼠与rubiconflox小鼠(tanaka,s.etal.,hepatology64,1994-2014(2016))交配而制作rubicon全部缺陷小鼠。将所制作的具有rubicon等位基因的小鼠与c57bl/6j野生型株回交5次,然后使其与rubicon+/-小鼠之间杂交而制作rubicon-/-小鼠和野生型对照。使nestin-cre小鼠与rubiconflox小鼠交配而制作在脑中特异性地具有rubicon的纯合缺陷的小鼠。除了热量限制小鼠之外,本实施例中使用的所有小鼠均保持在c57bl/6j背景下。这些实验中使用了由turturro等人(turturro,a.etal.,jgerontola-biol54,b492-b501(1999))开发的成人发病的40%热量限制方案。将雌性bdf1小鼠单独饲养在笼中。cr方案持续至9个月龄。随意饮水。
(4)蝇原种和培养条件
将蝇在标准的玉米粉-琼脂-酵母基的培养基中于25℃饲养。具有uas-mjdtr-q27(#8149)、uas-mjdtr-q78s(#8150,用于眼变性)、uas-mjdtr-q78w(#8141)、uas-gfp-ir(#9330)、uas-drubicon(cg12772)-ir(#43276)、uas-gfp-mcherry-atg8a(#37749)、da-gal4(#55849)和elav-gal4c155(#458)的蝇获自bloomingtonstockcenter。目前已经报道了具有gmr-gal4的转基因蝇系统(yamaguchi,m.etal.,oncogene18,6767-6775(1999))。
(5)个体数统计分析
对于蠕虫,在rnai板上从4-6小时的产卵获得同步的卵。从第1天的成体,以每板15~20只的密度建立寿命测试,并在20℃下进行。蠕虫每隔一天转移到新的板。也每隔一天计数存活率。所有rnai寿命从卵开始实施。死亡作为在用铂线刺激后没有任何动作而记录。排除自内部孵化、外阴部破裂或从板爬出的蠕虫。torrnai实验中,由成体进行敲低。使用mantel-cox对数秩检验法在excel(microsoft)中进行统计分析。将实验用蝇用标准培养基饲养,在出生后48小时使其交配,然后使用co2麻醉进行分选。不进行麻醉,每2-3天将瓶用新鲜食物交换,记录死亡直至所有蝇死亡。
(6)rna干扰
rna干扰(rnai)通过提供被针对靶标基因产生dsrna的载体l4440转化的ht115(de3)细菌来实施。将同步的卵置于指定的包含iptg和氨苄青霉素的rnai板上。rnai克隆获自ahringer或vidal的rnai文库。let-363/torrnai克隆由hansen博士(sanford-burnhammedicalresearchinstitute)提供。在双敲低实验中,将等量的细菌与各rnai混合并接种于板。使用空载体(l4440)和萤光素酶(l4440:luc)rnai作为非靶标对照。
(7)rna提取和qrt-pcr
在trizol(invitrogen社)或qiazol(qiagen社)中采集蠕虫、蝇和小鼠组织样品。使用rneasy试剂盒(qiagen社)提取总rna。使用iscript(bio-rad社)或primescriptrt试剂盒(タカラ社)生成cdna。qrt-pcr在viia7实时pcr系统(appliedbiosystems社)上使用powersybrgreen(appliedbiosystems社),或者在cfx96实时pcr检测系统(bio-rad社)上使用kapasybrfastqpcr试剂盒(kapabiosystems)进行。每个反应进行4次技术重复。使用ama-1(蠕虫)、rpl32(苍蝇)、18srrna(小鼠)和gapdh(人)作为内部对照。
(8)多蠕虫追踪分析
将13cm×10cm的琼脂填充板分成4个具有相等面积的区域。为了防止动物在区域之间移动,将各区域都用作为秀丽隐杆线虫(c.elegans)的厌恶性刺激的甘油包围。将来自某实验组的蠕虫放置在四个区域中的一个,将多个实验组同时进行试验。使用多蠕虫追踪器的适当版本(swierczek,n.a.etal.,natmethods8,592-598(2011))来记录秀丽隐杆线虫(c.elegans)的琼脂板上的运动。记录运动10分钟,使用choreography(多蠕虫追踪器软件的一部分)和自定义制成脚本对记录进行分析,然后整理和概括数据。动物的轨迹在记录中作为最后2分钟中各蠕虫的重心位置的时间序列收集。为了稳定追踪器对动物的识别,忽略最初的8分钟。使用choreography滤镜(无影和-t10)避免图像的伪影。个体的速度作为一系列重心之间的距离的合计除以轨迹的持续时间而得的值计算出。实验组使用由动物的轨迹长度加权而得的标本平均和标准误差来概括。
(9)爬升试验
爬升试验根据已发表的方案(suzuki,m.etal.,humanmoleculargenetics24,6675-6686(2015))稍加改变而进行。将10~20只蝇放入无麻醉的圆锥形玻璃管(长15cm;直径2.5cm)中。从蝇落到管的底部起10秒后,数出各铅直区域的蝇数并如下分数:分数0(0-2cm)、1(2-3.9cm)、2(4-5.9cm)、3(6-7.9cm)、4(8-9.9cm)、5(10-15cm)。以20秒的间隔对各组进行三次试验,并如下计算出爬升的分数。将各分数乘以蝇的数,然后所得的值除以蝇总数,计算出三次试验的平均值。结果作为3~9次独立实验获得的分数的标本平均±标准误差表示。
(10)显微镜使用和定量
为了监测自噬活性,用0.1%叠氮化钠麻醉l4阶段的gfp::lgg-1和mcherry::gfp::lgg-1动物,并使用olympusfv1000共聚焦显微镜获得图像。使用这些图像来计数和定量前肠区域或咽区域的gfp和/或mcherrypuncta。对于cerubicon::egfp和q35::yfp蠕虫,用配备有dp80ccd相机的体视显微镜szx16(olympus)拍摄排列在冰冷的空板上的蠕虫整体的荧光图像,以计算每蠕虫的polyq凝集物的数。
(11)蝇的眼的摄像
使用配备ccd相机(dp22,olympus)的szx16型体视显微镜(olympus)拍摄1日龄的成体雌蝇的光学显微镜图像。针对各基因型计数相对于1个复眼的死的小眼的数。
(12)蝇的自噬流
为了使摄食状态一致,在da-gal4驱动基因下、并且在drubicon-ir存在或不存在的状况下,用0.75%琼脂使表达gfp-mcherry-atg8a的4日龄的蝇饥饿15小时,然后再摄取4小时。将脑解剖,在pbs中的4%福尔马林中固定,用80%甘油孵育过夜,然后用dapifluoromount-g(southernbiotech)封装。通过共聚焦显微镜(ライカ社tcssp8)拍摄kc层的荧光图像,计数含有150~200个细胞的25×50μm的感兴趣区域中的gfp和/或mcherrypuncta的数。使用三个不重叠的图像来定量各kc层。
(13)应激耐受性测定
将第1天的成体蠕虫分别暴露于氧化应激(空板中4.4mm的h2o2)或热应激(35℃)3小时或7小时,并计数存活的蠕虫。对于各条件使用30只蠕虫,将实验重复3次。
(14)western印迹(蛋白质印迹)法
使用匀浆器在裂解缓冲液(50mmtris/hcl[ph7.4]、150mmnacl、1mmedta、0.1%np-40、蛋白酶和磷酸酶抑制剂合剂(roche社))中裂解蠕虫或小鼠的组织。离心分离后,将所得上清液进行蛋白质定量和western印迹(蛋白质印迹)。通过sds-聚丙烯酰胺凝胶电泳分离蠕虫和小鼠组织的蛋白质裂解物,并转移至pvdf膜,然后将其封闭并与特异性一抗一起孵育。用于小鼠组织的western印迹(蛋白质印迹)的一抗和稀释液如下。
rubicon(cellsignalingtechnology社、#8465,1:500)、lc3(cellsignalingtechnology社、#2755,1:1000)、p62(株式会社医学生物学研究所,pm045,1:1000)、磷酸化p70s6激酶(thr398)(cellsignalingtechnology社、#9209,1:500)、和β-肌动蛋白(sigmaaldrich社、a5316,1,8000)。
(15)免疫组织化学
对于蝇,将成体雌性的脑(自羽化起3天后)解剖,在pbs中的4%福尔马林内固定,用pbs/t(pbs中0.5%tritonx-100)中的50%blockace(大日本住友制药株式会社)封闭,与抗ha抗体(3f10,1:500,sigma社)一起孵育,然后用ha标签染色mjdtrq78蛋白质。用alexa488标记抗大鼠抗体使免疫染色可视化,用dapi染色核。用共聚焦显微镜(lsm780,zeiss社)拍摄kenyon(凯尼恩)细胞层(kc层)的图像。用imagej1.5s软件定量ha染色的信号强度。对于各组分析来自5只动物的10枚双侧kc层。在小鼠中,在石蜡包埋的切片上进行i型胶原的免疫组织化学染色。通过在0.01mmol/l柠檬酸盐缓冲液(ph6.0)中于120℃高压灭菌10分钟进行抗原回收后,将切片用pbs中的3%bsa封闭60分钟。将封闭的切片与作为一抗的抗i型胶原抗体(abcam社、ab34710,1:400)一起于4℃孵育过夜,于室温孵育30分钟,然后用hrp-二氨基联苯胺化合物(株式会社ニチレイ)进行检测。将切片用苏木精进行对比染色。
(16)重组α-syn原纤维的注射和检测
原纤维的注入和检测目前已有报道(luk,k.c.etal.,science338,949-953(2012);watanabe,y.etal.,autophagy13,133-148(2017))。
具体地,在脑内注射之前,将制备好的原纤维用无菌pbs稀释并进行超声波处理。将8~10周龄的小鼠用水合氯醛(250mg/kg,腹膜内)麻醉,并将重组α-syn原纤维(1mg/ml的1μl)定位地注射到两个半球中(坐标:相对于前囟+0.2mm,距离中线+2.0mm)。提供了出生后10个月的动物。使用cm3050s低温恒温器(leicabiosystems社)获得包含原纤维注射区域的冷冻脑切片。将切片与小鼠单克隆抗体psyn#64(稀释,1:1000,和光纯药工业株式会社)一起孵育,然后与alexafluor488标记二抗(稀释,1:1000,invitrogen社)一起孵育,然后进行检测。用biorevobz-9000(株式会社キーエンス)获得图像并定量。
(17)细胞培养
以每6孔5.0×104个的密度接种rpe-1细胞。一天后,针对萤光素酶和mondoa(santacruz社),用20nm的sirna处理细胞。经过48小时的sirna处理后,将rpe1细胞在样品缓冲液中裂解,进行sds-聚丙烯酰胺凝胶电泳和western印迹(蛋白质印迹)。通过western印迹(蛋白质印迹)定量rubicon的蛋白质水平。在sirna处理的一天前,以每6孔5.0×104个的密度接种rpe-1细胞。
(18)统计分析
结果作为标本平均±标准误差显示。统计学检验使用graphpadprism(graphpadsoftware社)或excel(microsoftoffice2011)的通过利用tukey检验或t检验的单因素方差分析或双因素方差分析进行。
trap染色
将骨髓细胞在24孔板中培养,通过加入m-csf和rankl,从而向破骨细胞诱导。用4%多聚甲醛固定5分钟,用蒸馏水洗涤,然后加入含有50mm酒石酸的缓冲液(ph5.0)和生色底物,在37℃下反应45分钟。利用trap活性的染色用bz-x700(keyence)进行显微镜检查。
(20)窝形成测定(pitformationassay)
在osteoassayplate(corning#3988)中培养骨髓细胞,并且添加m-csf和rankl从而向破骨细胞诱导。破骨细胞通过溶解骨基质而形成吸收窝(pit),用bz-x700(keyence)进行显微镜检查。
(21)alp染色
对于原代培养系统在12孔板中,对于培养细胞系统在96孔板中,在骨分化培养基中培养5天。骨分化培养基由补充有10%fbs的αmem、50μg/mll-抗坏血酸磷酸酯(shigma)、10mmβ-甘油磷酸(shigma)制成。用4%多聚甲醛(在0.1m二甲胂酸缓冲液中,ph7.3)中固定30分钟,添加溴-氯-吲哚基-磷酸盐/氯化硝基四氮唑兰,在37℃染色20分钟,用bz-x700(keyence)进行显微镜观察。
(22)茜素染色
对于原代培养系统在12孔板中,对于培养细胞系统在96孔板中,在骨分化培养基中培养5天。骨分化培养基由补充有10%fbs的αmem、50μg/mll-抗坏血酸磷酸酯(shigma)、10mmβ-甘油磷酸(shigma)制成。骨分化培养基每2天进行培养基更换,用4%多聚甲醛(在0.1m二甲胂酸缓冲液中,ph7.3)中固定30分钟。用0.1m二甲胂酸缓冲液(ph7.3)洗涤,并用茜素红s(ph4.0)染色5分钟,然后用蒸馏水再次洗涤。使孔干燥,用bz-x700(keyencne)进行显微镜检查。
(23)骨分析
使用c57bl/6北京的12周龄的雄性小鼠进行分析。但是在骨质疏松症模型中,从9周龄的雌性小鼠摘除卵巢,4周后测量骨形态。骨形态通过ct扫描(小中型实验动物用3d显微x射线ct/cosmoscanax;rigaku)小鼠的股骨远端获得。然后,用tri/3d-bon软件(ratocsystems)分析3d显微ct数据,进行骨形态分析。
实施例
(1)介由rubicon产生的自噬的调节来控制寿命
探索秀丽隐杆线虫(c.elegans)的rubicon同源物,发现了y56a3a.16与人rubicon(kiaa0226)、人kiaa0226l(最近被鉴定为pacer)(cheng,x.etal.,molcell65,1029-1043(2017))、和果蝇rubicon(drubicon,cg12772)(banreti,a.etal.,devcell28,56-69(2014))具有序列类似性(图5a)。
在哺乳类中,rubicon阻碍自噬,但pacer是正常进行自噬所必须的。因此,接下来,进行了这两种蛋白质中的哪一者在功能上更类似于y56a3a.16的研究。如到目前为止在rubicon敲低哺乳类细胞中所观察到那样(非专利文献6),通过利用rnai的y56a3a.16的敲低,从而作为自噬体标志物的gfp::lgg-1点的数在肠细胞中大幅增加(图1a(i),(b))。该增加通过同时敲低在自噬囊泡和形成中起作用的bec-1/beclin1而完全消失。这个事实表明y56a3a.16的敲低导致自噬囊泡的增加(图1a(i),(ii))。
为了进一步判别y56a3a.16的敲低是否增强或阻断自噬活性,使用最近开发的表达串联荧光lgg-1(mcherry::gfp::lgg-1)的转基因蠕虫监测自噬流(非专利文献1)。观察到通过y56a3a.16的敲低,肠和咽肌中的自噬体(ap)和自噬溶酶体(al)的数大幅增加(图1b(i)、(ii),图5b(i)、(ii)),这显示自噬流通过敲低而提高。结合斑马鱼和果蝇中仅有一个rubicon同源物的事实(图5a)(banreti,a.etal.,devcell28,56-69(2014)),该结果意味着y56a3a.16表示rubicon的祖先同源物(因此将y56a3a.16称为“cerubicon”)。令人感兴趣的是,通过使用cerubicon::egfp转基因蠕虫和qrt-pcr分析的基因表达实验判明了,与年轻蠕虫(第1天)相比,cerubicon在年老蠕虫(第7天)中被上调(图1c(i)、(ii))。由这些结果导入如下假说:作为自噬的负的调节因子的rubicon的年龄依赖性增加,可能成为自噬年龄依赖性地减弱的原因。并且,如该观点所述,利用rnai的cerubicon的敲低大幅延长了野生型蠕虫的寿命(图1d(i)、(ii),图5c(i))。cerubicon的敲低不改变咽部抽吸速率,显示包括rnai的细菌被适当地摄入(图6b(i))。rubicon转录水平在这些动物中降低至正常水平的约50%(图5c(ii))。
在哺乳类细胞中,显示rubicon抑制自噬和胞吞作用,但是如果通过rnai将作为自噬控制因子的bec-1/beclin1、unc-51/ulk1、和atg-18/atg18与cerubicon一起敲低,则寿命的增加完全消失(图1d(i)、(ii),图5c(i))。这显示,由cerubicon敲低付与的寿命提高依赖于自噬活性。相反,通过cerubicon的过表达,野生型蠕虫的寿命变短(图1e(i)、(ii))。
上述结果提示,cerubicon通过调节自噬活性来控制寿命。
(2)由全身rubicon敲低产生的年龄依赖性表型的改善
研究了rubicon的敲低是否影响其它年龄发病型。在体壁肌中表达多聚谷氨酰胺(polyq)-yfp的蠕虫显示出polyq融合蛋白质的年龄发病凝集(morley,j.f.等,pnatlacadsciusa99,10417-104229(2002))。cerubicon的敲低使凝集的水平降低,但bec-1/beclin1的同时敲低则使降低恢复。这显示cerubicon的敲低自噬依赖性地使polyq融合蛋白质的凝集降低(图2a,图2b(i))。
运动量随年龄下降,并且速度与寿命良好相关(hahm,j.h.etal.,naturecommunications6,doi:artn8919)。通过多蠕虫追踪系统,能够监测各个蠕虫的总计,并计算运动速度(swierczek,n.a.etal.,natmethods8,592-598(2011))。该分析表明,cerubicon的敲低自噬依赖性地减缓运动量的年龄依赖性下降(图2b(ii),图6a)。
大量长寿动物经常表现出对包括氧化应激和热应激在内的多种应激的耐受性。我们发现,通过cerubicon的敲低,氧化应激耐受性自噬依赖性地增加了(图2b(iii))。另一方面,热应激耐受性不变(图6b(ii))。这显示,cerubicon的敲低提供了关于应激耐受性的不同下游输出。
为了拓展秀丽隐杆线虫(c.elegans)中的发现,研究了rubicon敲低是否也延长蝇的寿命。起结果是,果蝇rubicon(drubicon)的整体敲低大幅增加了脑内的自噬流,雌性特异性地延长了寿命(图2c、图6c(i)、(ii)、d(i))。特别是通过drubicon的敲低,mjdtr-q78(包含膨胀谷氨酰胺管的mjd/sca3的缩短型)polyq疾病模型雌蝇中的复眼的变性改善了(图2d(i)、(ii))。作为mtor激酶处理的抑制剂的雷帕霉素,在雌性小鼠中比在雄性小鼠中增加得更多。因此,该观察可能反映了自噬对动物寿命的性二态的贡献。
与秀丽隐杆线虫(c.elegans)同样地,发现了肾脏和肝脏中的rubicon蛋白质水平在老年小鼠(20个月龄)中高于年轻小鼠(2个月)(图2e(i)、(ii),图6d(ii))。为了理解rubicon在小鼠中的动物衰老中的作用,开发了高lc3-ii的水平高、且自噬底物p62的水平低,即基础自噬激活了的rubicon整体敲除小鼠(图2f(i))。进行性纤维化症是肾脏衰老的组织学特征(yamamoto,t.etal.,autophagy12,801-813(2016))。如在rubicon整体敲除小鼠中通过i型胶原的免疫组织化学所确定的那样,年龄相关性纤维化症减少(图2f(ii),图2g(i))。该表型通过编码纤维化标记物的mrna的qrt-pcr检测来确认(图2g(ii))。简言之,通过rubicon的敲低或缺陷,蠕虫、蝇和小鼠中的年龄相关性表型改善了。
(3)由神经元中rubicon水平的降低带来的衰老的延迟
研究了哪些组织cerubicon承担秀丽隐杆线虫(c.elegans)的寿命控制。对于cerubicon的敲低,使用包括tu3401(神经元特异、sid-1系统)(calixto,a.etal.,naturemethods7,554-u102(2010))、nr350(肌肉)、vp303(肠)和nr222(皮下组织)在内的组织特异性的rnai敏感株(图3a(i)、7a(i)~(iii))(qadota,h.etal.,gene400,166-173(2007))。神经细胞中的cerubicon的敲低最有效地延长寿命(图3a(i))。另一方面,cerubicon的皮下组织和肠敲低虽然程度比神经细胞中的cerubicon的敲低低,但也发福延长寿命(图7a(i)~(iii))。由于神经细胞通常对rnai敲低有耐受性,因此由野生型蠕虫的cerubicon的敲低付与的寿命延长可能反映了来自皮下组织、肠和其他未鉴定组织的组合效应(图1d(i)、(ii)、5c(i))。
接着,研究了由cerubicon的敲低带来的神经元的自噬作用的增加是否足以延长动物的寿命。其结果是,神经元细胞中的通过cerubicon的敲低所付与的寿命提高,通过atg-18/wipi基因的同时敲低而完全消失(图7b(i))。这显示自噬的神经元激活足以延长动物的寿命。同样地,果蝇的rubicon的神经元特异性敲低也延长了雌蝇的寿命(图3a(ii),图7b(ii))。进而发现,rubicon敲低在雌性的脑神经元中使polyq内含体减少了(图3b(i),(ii)),缓解了被polyq细胞毒性加速的运动功能的年龄相关性下降(图3b(iii))。
自噬涉及包括帕金森病在内的几种与衰老相伴的神经变性疾病。在散发性和家族性的帕金森病中,受影响的神经元表达包含α-突触核蛋白(α-syn)的包含体。上升的该蛋白质的量足以引起帕金森病。α-syn过表达、且错误折叠的易于凝集的形态被自噬分解。因此,研究了由神经元中的rubicon的缺失引起的自噬的强制激活,是否足以防止脑中的α-syn的凝集。为了探索该可能性,如目前已经报告的那样(luk,k.c.etal.,science338,949-953(2012);watanabe,y.etal.,autophagy13,133-148(2017)),将预先形成的α-syn原纤维注射到对照和rubicon神经元(巢蛋白-cre)敲除小鼠的纹状体中,10个月后比较了lewy小体和lewy神经突状α-syn包含体的形成。值得注意的是,磷酸化α-syn阳性信号的水平在神经元特异性rubicon敲除小鼠中大幅降低(图3c(i),(ii)),这提示神经元rubicon缺失抑制了α-突触核蛋白病变的扩大。这些结果提示,神经元的敲低足以改善许多生物体中的衰老表型。
(4)多个寿命延长条件下的rubicon的下调
研究了rubicon表达是否可通过多种寿命延长条件来改变。在蠕虫中,多种不同的长寿途径和相应的长寿变异体已经被充分鉴定出来。其中包括胰岛素/igf-1信号传导(daf-2)、生殖销细胞系列的去除(glp-1)、热量限制(eat-2)和线粒体功能障碍(isp-1)的减少(kenyon,c.j.nature464,504-512(2010))。与野生型相比,cerubicon表达在所有这些长寿蠕虫中均被抑制(图4a(i)、图8b(ii))。此外,受到9个月的热量限制的小鼠显示肝脏和肾脏中rubicon表达的减少(图4a(ii)、图8b(i))。还发现cerubicon的敲低不会进一步延长daf-2、glp-1、eat-2和isp-1的寿命(图8a(i)~(iv))。该结果显示,rubicon在多个寿命范式的下游被控制,这些动物的寿命部分由rubicon表达的减少来付与。
在蠕虫中,mml-1/mondo被鉴定为多个长寿途径所需的趋同转录因子之一(nakamura,s.etal.,natcommun7,10944(2016);johnson,d.w.etal.,plosgenet10(2014))。特别是,mml-1的敲低使生殖细胞系列缺陷的长寿glp-1动物的自噬活性和寿命消失。然而,尚未完全表征mml-1如何控制自噬。因此,研究了mml-1是否参与cerubicon的控制。令人感兴趣的是,尽管cerubicon在其他长寿动物中仍被大幅抑制(图8b(ii)),但glp-1动物中的cerubicon的抑制在mml-1或作为其异二聚体伴侣的mxl-2的敲低之后恢复完全(图4b(i))。此外,带来缓慢的寿命延长的mml-1过表达(上述,nakamura,s.etal.),与野生型相比,使cerubicon表达减少了(图4b(ii))。此外,尽管rubicon的转录水平未变化,但通过作为mml-1的哺乳动物同源物的mondoa的敲低,哺乳类细胞中的rubicon蛋白质水平上升(图4c(i),(ii))。这些结果显示,rubicon特别是在生殖细胞系列寿命的体系中,在mml-1/mondoa的下游被控制,认为该控制在哺乳动物中是保守的。
营养素传感器mtor响应营养素缺乏而失活。另外mtor的抑制还诱导自噬,延长多个物种的寿命(kapahi,p.etal.,cellmetab11,453-465(2010))。此外,mml-1通过抑制亮氨酸传感器lars-1来控制tor活性(nakamura,s.etal.,同上)。因此,研究了rubicon与mtor路径之间的可能的交互作用。其结果令人惊讶地判明了,cerubicon和let-363/mtor的双重敲低与仅敲低任一方相比,进一步延长了寿命(图4d)。cerubicon的敲低不影响tor活性,但tor敲低不改变cerubicon表达(图8c(i)、(ii))。这些结果显示,作为自噬的两个主要的负的控制因子的tor和rubicon通过部分独立的下游机制而有助于寿命。
(5)介由rubicon带来的自噬的骨代谢的调节
用ct拍摄全身的rubicon缺陷小鼠的股骨,结果在rubicon缺陷小鼠中,股骨的骨密度(bmd)、骨体积/骨组织体积比(bv/tv)、骨矿物质含量(bmc)显著升高(图9a)。因此,制作破骨细胞和成骨细胞的原代培养系统,评价了各自的分化能力和功能。破骨细胞的分化用trap染色评价,在rubicon缺陷破骨细胞中分化得到增强(图9b)。另外,用窝形成测定(pittformulationassay)评价破骨细胞的骨吸收功能,结果在rubicon缺陷成骨细胞中骨吸收功能得到增强(图9c)。接着,在原代培养系统和培养细胞系统中研究成骨细胞的分化能力和矿化功能。培养细胞系统中的rubicon缺陷细胞通过crisper-cas9系统制成。用alp染色评价分化能力,结果在rubicon缺陷成骨细胞中分化能力显著上升(图9d)。另外,用茜素染色评价矿化功能,结果在rubicon缺陷成骨细胞中矿化功能得到增强(图9e)。在全身的rubiconko小鼠中,破骨细胞和成骨细胞均分化能力和功能增强,但在rubicon缺陷小鼠中,股骨的骨形态得到增强,由此推测,成骨细胞的功能更为重要。另外,用lc3flux测定确认在rubicon缺陷成骨细胞中自噬是否增强。其结果是,rubicon缺陷成骨细胞在营养条件下、饥饿条件下双方均lc3flux增强(图10)。
成骨细胞特异性小鼠制作中,将osterix-cre小鼠和rubiconflox/flox小鼠交配,制作rubiconflox/flox;osterix-cre小鼠。同时,将osterix-cre小鼠与atg5flox/flox小鼠和rubiconflox/flox;atg5flox/flox小鼠交配,制作成骨细胞特异性atg5缺陷小鼠和rubicon/atg5缺陷小鼠,对骨形态进行ct分析。其结果是,成骨细胞特异性rubicon缺陷小鼠中,骨密度显著增加。另一方面,有报告称成骨细胞特异性atg7缺陷小鼠中骨密度降低(li,h.etal.,autophagy14(10),1726-1741(2018)),在本实验中atg5缺陷小鼠中骨密度也显著降低。此外,在rubicon/atg5缺陷小鼠中,骨密度与atg5缺陷小鼠同等地降低(图11a)。该事实显示,成骨细胞特异性rubicon缺陷小鼠中的骨形成依赖于自噬而发生。
在本实施例中,成骨细胞特异性rubicon缺陷小鼠显示成骨性的变化,制作了骨质疏松症模型研究了这是否有助于骨质疏松症的改善。作为骨质疏松症模型,使用通过摘除两侧卵巢、在四周内使雌激素耗尽而制成的绝经后骨质疏松症模型。其结果是,在连体小鼠中,骨质疏松症被诱导,骨密度降低,骨体积/骨组织体积比也降低,与此相对,在rubicon缺陷小鼠中,骨质疏松症模型中骨密度和骨体积/骨组织体积比显著改善(图11b)。
在rubicon缺陷小鼠中,确认到自噬依赖性地发生成骨性变化,在rubicon缺陷成骨细胞中,自噬、分化及成骨性功能增强。该事实提示,自噬与成骨细胞的分化相关。迄今为止,已知wnt、bmp或notch的信号通路与成骨细胞的分化相关(zanotti,s.&canalis,e.molecularandcellularbiology,30(4),886-896(2010)、muguruma,y.etal.,journalofcellularphysiology,232(9),2569-2580(2017)),在本实施例中,着眼于notch的信号通路。这是因为,已经报告了成骨细胞特异性notch2缺陷小鼠(notch具有1~4个异形体,在成骨细胞中notch2高表达)显示成骨性的变化,成骨细胞的分化和功能也增强,notch受体自噬依赖性地减少(yorgan,t.etal.,bone87(c),136-146(2016)、wu,x.etal.,naturecommunications7,1-17(2016)、cao,y.etal.,stemcellsanddevelopment24(22),2660-2673(2015))。在已有的报告中,成骨细胞的notch2缺陷小鼠显示成骨性变化的机制被报告如下。即,通过notch2受体缺陷而转移至核内的nicd降低,notch下游的转录因子受到抑制。notch下游的hes、hey负地控制作为成骨性的转录因子的runx2、osterix,因此通过hes、hey减少,从而runx2、osterix等成骨性转录因子增加,成骨细胞的分化能力和骨形成功能增强。因此,本发明者预测,在rubicon缺陷成骨细胞中,通过增强的自噬而促进notch受体的分解,从而notch下游的hes、hey减少,进而runx2、osterix增加。因此,首先,确认了notch下游的转录因子和成骨性转录因子的mrna的表达,结果判明,notch下游的hey1、heyl、hes1、hes3、hes7在rubicon缺陷成骨细胞中的mrna的表达降低,runx2、osteocalsin等成骨性转录因子的mrna的表达增加(图12a)。进而,通过western印迹法观察在rubicon缺陷成骨细胞中notch2和nicd是否被分解,结果由溶酶体抑制所致的notch2和nicd的流(flux)在rubicon缺陷成骨细胞中增加,提示了在rubicon缺陷细胞中,notch2和nicd进一步被分解(图12b)。
在rubicon缺陷成骨细胞中,notch受体和nicd的分解增强,但不清楚nicd是否在rubicon的下游发挥功能。因此,使nicd在rubicon缺陷的成骨细胞中过表达,用alp染色评价其分化能力,结果与野生型成骨细胞同等地抑制了分化。由该事实判明,在rubicon的下游nicd被控制(图13a)。由于在nicd中有两处lir基序,因此认为其有可能成为自噬的底物。因此,使用附加有flag标签的nicd,通过免疫沉淀法确认了nicd与lc3的结合。用flag进行免疫沉淀,通过免疫印迹对lc3进行检测,结果判明了lc3与nicd直接结合(图13b)。
产业可利用性
如以上所说明的那样,根据本发明,通过以rubicon作为靶标,能够抑制与自噬的经时性减弱相伴的衰老,广泛地预防、改善或治疗衰老相关疾病或症状,延长寿命。另外,通过以rubicon作为靶标,还能够检查衰老的程度、这些疾病或症状的发病风险、或缩短寿命的风险。因此,本发明特别能够对医疗领域有很大贡献。
- 还没有人留言评论。精彩留言会获得点赞!