用于淋巴系统病症的诊断和治疗的组合物和方法与流程
本申请要求于2018年9月7日提交的美国临时申请号62/728,444的优先权,其全部内容通过引用的方式并入本文,如同完整阐述一样。
本发明涉及遗传学,个性化医疗和淋巴系统畸形的领域。更具体地,本发明提供了新的遗传靶标和治疗性处理方案,用于改善与淋巴管瘤病和其他广泛性淋巴管异常(gla)有关的症状。
背景技术:
在整个说明书中引用了若干出版物和专利文献,以描述本发明所属领域的现有技术水平。这些引用中的每一个都通过引用的方式并入本文,如同完整阐述一样。
淋巴系统在维持体液循环、保护身体免受疾病侵害以及吸收小肠中的膳食脂肪方面起着关键作用(1)。复杂淋巴管异常的特征是淋巴管的异常形成和组织过度生长。患者经常出现可能导致严重的肺部疾病的重叠症状(2、3)。淋巴管异常的例子包括广泛性淋巴管异常(gla)、淋巴管扩张和乳糜性积液(心包、胸膜或腹膜)。由于诊断上的重大挑战,分类和命名的不一致阻碍了对于复杂淋巴管异常的研究(3-6)。尽管对复杂淋巴管异常的分子遗传病因了解甚少,但先天性淋巴系统畸形似乎具有相关的潜在遗传病因(7-9)。实际上,已经在集中于pi3k/mtor和ras/mapk途径的基因中鉴定出种系和体细胞突变(1、8)。
pi3k/mtor和ras/mapk信号通路的破坏或畸变已被证明会损害成熟淋巴网络构建过程中的正常扩张和重塑,其中这种破坏与淋巴疾病有关。akt1和pik3ca功能突变的获得(导致雷帕霉素复合物1(mtorc1)活性的哺乳动物靶标升高)在淋巴畸形患者中被确认,其包含部分综合征,例如proteus综合征(omim176920)、cloves综合征(omim612918)以及klippel-trenaunay-weber综合征(omim149000)(9-11)。kras、hras、raf1、ptpn11、sos1和rasa1的突变(其导致ras通路活性失调),导致在以下综合征中的淋巴水肿或淋巴管扩张:noonan综合征(omim163950)、costello综合征(omim218040)、心脸皮肤综合征(omim115150)和毛细血管畸形-动静脉畸形(cm-avm)综合征(omim608354)(12-17)。
尽管有这些理解,但是缺乏用于确认患有淋巴病症和淋巴管异常(例如淋巴管瘤病/淋巴管扩张(lam)、广泛性淋巴管异常(gla)和乳糜性积液)的患者的遗传生物标志物,也缺乏靶向与这些病症相关的遗传标志物的治疗剂。
技术实现要素:
因此,在本发明的一个实施方案中,提供了一种用于在人类患者中诊断淋巴管异常的方法。示例性方法包括:从所述患者获得包含核酸的生物样品;分析核酸以确定:i)在ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1、ephb4、nf1、cbl和araf的一个或多个中是否存在单核苷酸变异(snv),或ii)是否存在与ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1、ephb4、nf1、cbl和araf的一个或多个中的snv处于连锁不平衡的snv;以及淋巴管异常,如果存在i)或ii)的snv,则诊断患者患有淋巴管异常。在另一方面,用于在人类患者中诊断淋巴管异常的方法需要从人类患者获得基因型序列信息,根据所述序列信息确定:i)在ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1、ephb4、nf1、cbl和araf的一个或多个中是否存在单核苷酸变异(snv),或ii)是否存在与ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1、ephb4、nf1、cbl和araf的一个或多个中的snv处于连锁不平衡的snv;以及淋巴管异常,如果存在i)或ii)的snv,则诊断患者患有淋巴管异常。
本发明还提供了一种用于在人类患者中治疗淋巴管异常的方法。示例性方法包括:从所述患者获得包含核酸的生物样品;分析核酸以确定i)在ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1、ephb4、nf1和cbl的一个或多个中是否存在单核苷酸变异(sanv)或ii)是否存在与ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1、ephb4、nf1和cbl的一个或多个中的snv处于连锁不平衡的snv;以及向确认具有i)或ii)的一种或多种snv的患者施用适于治疗所述淋巴管异常的一种或多种药剂,从而治疗所述淋巴管异常。在该方法的替代实施方案中,从患者获得基因型信息并进行分析以确定:i)在ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1、ephb4、nf1和cbl的一个或多个中是否存在单核苷酸变异(snv),或ii)是否存在与ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1、ephb4、nf1和cbl的一个或多个中的snv处于连锁不平衡的snv;以及向确认具有i)或ii)的一种或多种snv的患者施用适于治疗所述淋巴管异常的一种或多种药剂,从而治疗所述淋巴管异常。在该方法的替代实施方案中,从患者获得基因型信息。
在某些实施方案中,所述淋巴管异常的特征是淋巴管的异常形成和/组织过度生长。在其他实施方案中,所述淋巴管异常是淋巴管瘤病(lam)。在另一个实施方案中,所述淋巴管异常是广泛性淋巴管异常(gla)。所述淋巴管异常的特征是乳糜性积液,包括心包、胸膜或腹膜积液。
所述诊断方法可以进一步包括在生物样品中检测后产生确认snv的报告。上述治疗方法可以进一步包括基于在该方法中确认的snv,生成确认针对淋巴管异常的建议治疗的报告。
在一些实施方案中,施用给snv阳性受试者的药剂是1)mek/erk抑制剂;2)表1和2中列出的药剂/抑制剂;3)mek/erk抑制剂和表1和2中所列出的一种或多种药剂/抑制剂的组合;和/或4)1)mtor抑制剂和/或pik3k抑制剂;和2)一种或多种mek/erk抑制剂的组合。在又一个实施方案中,本文所述的诊断方法可以进一步包括向经诊断的患者施用有效量的一种或多种适于治疗所述淋巴管异常的药剂。
在所述治疗方法的某些实施方案中,所施用的药剂(例如施用于携带一种或多种淋巴管异常相关的snv的患者)选自一种或多种mek/erk抑制剂,和任意所述抑制剂的一种或多种的组合。在一些实施方案中,所施用的药剂是mek/erk抑制剂。在一些实施方案中,所施用的药剂是表1和表2中列出的一种或多种药剂。在一些实施方案中,所施用的药剂是mek/erk抑制剂以及表1和表2中列出的一种或多种药剂。在一些实施方案中,所施用的药剂是1)mtor抑制剂和/或pik3k抑制剂;和2)一种或多种mek/erk抑制剂的组合。在一些实施方案中,当所述药剂是mtor抑制剂时,施用雷帕霉素和/或bez-235(达克利司(dactolisib))。在某些实施方案中,所述一种或多种mtor抑制剂、一种或多种pik3k抑制剂和/或一种或多种mek/erk抑制剂具有小于100μm、小于10μm、小于1μm、小于100nm、小于10nm、小于1nm的ic50。
在一些实施方案中,所述患者在ptpn11中没有snv。在一些实施方案中,所述患者在braf没有snv。在一些实施方案中,所述患者在kras没有snv。在一些实施方案中,所述患者在sos1中没有snv。在一些实施方案中,所述患者在itga9中没有snv。
在一些实施方案中,将表1和表2中列出的药剂组合使用。这些组合包括但不限于,a)地磷莫司(ridaforolimus)和曲美替尼(trametinib);b)地磷莫司和司美替尼(selumetinib)或考比替尼(cobimetinib);c)bez235和司美替尼;d)奥米利塞(omipalisib)和司美替尼或曲美替尼;e)依维莫司(everolimus)和曲美替尼或司美替尼;f)西罗莫司(sirolimus)、地磷莫司和司美替尼;g)西罗莫司、地磷莫司和曲美替尼;h)torkinib和曲美替尼;i)bez235、torkinib和曲美替尼;以及j)西罗莫司和gedatolisib和曲美替尼。在其他实施方案中,所述治疗还包括施用全身化疗、干扰素α、放疗和/或手术。
在一些实施方案中,所述snv选自选自以下的snv:ptpn11基因中的c.1504t>g:ps502a、c.1510a>g:pm504v和/或c.1507g>c:pg503r、kras中的c.35g>a:pg12d、braf基因中的c.1403t>c:pf468s、sos1基因中的c.2536g>a:pe846k,以及包含itga9基因中的c.1236+4a>g和c.289t>g:p.c97g的复合突变,其中“c.”指定编码dna序列,“p.”指定蛋白质序列。
在一些实施方案中,所述诊断方法包括检测一个或多个上述snv。在一些实施方案中,所述诊断方法还包括向受试者施用一种或多种已知可治疗淋巴管异常的药剂。在一些实施方案中,所述药剂是mek/erk抑制剂。在一些实施方案中,所施用的药剂是表1和表2中列出的一种或多种药剂。在一些实施方案中,所施用的药剂是mek/erk抑制剂以及表1和表2中列出的一种或多种药剂。在一些实施方案中,所施用的药剂是1)mtor抑制剂和/或pik3k抑制剂;和2)一种或多种mek/erk抑制剂的组合。所述药剂的施用改善了一种或多种淋巴结构、减少了乳糜性胸膜积液、改善了呼吸功能、允许逐渐减少伴随用药、和/或增加了存活率。
在一些实施方案中,所述诊断方法包括检测以下的一种或多种:ptpn11基因中的c.1504t>g:ps502a、c.1510a>g:pm504v和/或c.1507g>c:pg503r,以及单独或组合施用至少一种或多种mek/erk抑制剂。在其他实施方案中,所述药剂选自表1-2,从而改善了一种或多种淋巴结构、减少了乳糜性胸膜积液、改善了呼吸功能、允许逐渐减少伴随用药、或增加了存活率。
在一些实施方案中,所述诊断方法包括检测braf基因中的c.1403t>c:pf468s。在一些实施方案中,所述诊断方法还包括单独或组合使用一种或多种mek/erk抑制剂治疗所述患者。在其他实施方案中,药剂选自表1-2,从而改善了一种或多种淋巴结构、减少了乳糜性胸膜积液、改善了呼吸功能、允许逐渐减少伴随用药、或增加了存活率。
在一些实施方案中,所述诊断方法包括检测kras中的c.35g>a:pg12d,以及单独或组合施用一种或多种mtor抑制剂和一种或多种mek/erk抑制剂。在其他实施方案中,至少一种药剂选自表1-2用于施用,从而改善了一种或多种淋巴结构、减少了乳糜性胸膜积液、改善了呼吸功能、允许逐渐减少伴随用药、或增加了存活率。
在一些实施方案中,所述诊断方法包括检测sos1基因中的c.2536g>a:pe846k。在一些实施方案中,所述诊断方法还包括单独或组合使用一种或多种mek/erk抑制剂治疗所述患者。在其他实施方案中,选择表1-2的药剂,从而改善了一种或多种淋巴结构、减少了乳糜性胸膜积液、改善了呼吸功能、允许逐渐减少伴随用药、或增加了存活率。
在一些实施方案中,所述诊断方法包括检测itga9基因中的c.1236+4a>g和/或c.289t>g:p.c97g,以及单独或组合施用一种或多种mek/erk抑制剂。在一些实施方案中,施用表1-2的至少一种药剂,从而改善了一种或多种淋巴结构、减少了乳糜性胸膜积液、改善了呼吸功能、允许逐渐减少伴随用药、或增加了存活率。
适用于治疗的erk/mek抑制剂包括但不限于司美替尼(azd6244)、pd0325901,曲美替尼(gsk1120212)、pd184352(ci-1040)、pimasertib(as-703026)、tak-733,azd8330、比美替尼(binimetinib)(mek162、arry-162、arry-438162)、sl-327、瑞法替尼(refametinib)(rdea119,bay86-9766)以及考比替尼(gdc-0973、rg7420)。
在一些实施方案中,分析所述核酸以确定在ptpn11基因中的c.1504t>g:ps502a、c.1510a>g:pm504v和/或c.1507g>c:pg503r、kras中的c.35g>a:pg12d、braf基因中的c.1403t>c:pf468s、sos1基因中的c.2536g>a:pe846k,以及包含itga9基因中的c.1236+4a>g和c.289t>g:p.c97g的复合突变的一个或多个中是否存在单核苷酸变异(snv)的步骤进一步包括分析多核苷酸样品的步骤,以通过执行选自以下的过程来确定所述snv的存在:特异性杂交的检测、等位基因大小的测量、限制性片段长度多态性分析、等位基因特异性杂交分析、单碱基引物延伸反应和扩增的多核苷酸测序。
在一些实施方案中,所述生物样品包含dna。
在一些实施方案中,所述生物样品包含rna。
在一些实施方案中,包含所述snv的核酸获自人类患者的分离的细胞。
在一些实施方案中,分离的载体编码带有snv的核酸,其中所述snv选自:ptpn11基因中的c.1504t>g:ps502a、c.1510a>g:pm504v和/或c.1507g>c:pg503r、kras中的c.35g>a:pg12d、braf基因中的c.1403t>c:pf468s、sos1基因中的c.2536g>a:pe846k,以及包含itga9基因中的c.1236+4a>g和c.289t>g:p.c97g的复合突变。
在一些实施方案中,宿主细胞包含编码带有snv的核酸的分离的载体。在一些实施方案中,转基因动物包含宿主细胞。在一些实施方案中,所述转基因动物是小鼠或斑马鱼。
在一些实施方案中,筛选药剂作用的方法包括使宿主细胞或转基因动物与单独或组合的一种或多种本文所述的抑制剂接触,或涵盖表1-2的试剂。在一些实施方案中,被筛选的药剂的作用是在斑马鱼中的尾部挽救(caudalrescue)或分支挽救(branchingrescue)。
在一些实施方案中,用于鉴定改变细胞信号转导的试剂的方法包括:提供表达至少一种包含如上所述的至少一种snv的核酸的细胞、提供表达缺少所述snv的同源野生型序列的细胞;使两种细胞群都与测试试剂接触;以及分析所述试剂是否改变具有含所述snv核酸的细胞(相对于缺少所述snv的细胞)的细胞信号转导。
附图说明
图1:ea.hy926细胞系中braf和ptpn11突变体的过表达激活erk信号转导。
图2:ea.hy926细胞系中braff486s的过表达改变了细胞的形态和肌动蛋白的组织。
图3:在过表达brafwt和f486s突变体的细胞中,用mek抑制剂曲美替尼治疗可增加ve-钙粘蛋白表面染色和丝状肌动蛋白。
图4a-4b:mek抑制剂可降低过度表达brafwt和f486s突变体的细胞中erk的激活/磷酸化。图4b-mek抑制剂可降低过度表达brafwt和f486s突变体的细胞中erk的激活/磷酸化。
图5a-5b:mek抑制剂可降低过度表达ptpn11突变体的细胞中erk的激活/磷酸化(图5a)。mek抑制剂可降低过度表达ptpn11突变体的细胞中erk的激活/磷酸化(图5b)。
图6:pd0325901对表达araf突变s214p的hdlec的作用。pd0325901挽救了araf-s214p诱导的细胞形态差异,如通过在细胞膜处的降低的perk水平和增加的ve-钙粘蛋白积累所示。绿色表示ha染色,它标记表达araf的细胞;红色表示ve-钙粘蛋白染色。
图7:ci1040对表达araf突变s214p的hdlec的作用。ci1040挽救了araf-s214p诱导的细胞形态差异,如通过在细胞膜处的降低的perk水平和增加的ve-钙粘蛋白积累所示。绿色表示ha染色,它标记表达araf的细胞;红色表示ve-钙粘蛋白染色。
图8:pimasertib对表达araf突变s214p的hdlec的作用。pimasertib挽救了araf-s214p诱导的细胞形态差异,如通过在细胞膜处的降低的perk水平和增加的ve-钙粘蛋白积累所示。绿色表示ha染色,它标记表达araf的细胞;红色表示ve-钙粘蛋白染色。
图9:tak-733对表达araf突变s214p的hdlec的作用。tak-733挽救了araf-s214p诱导的细胞形态差异,如通过在细胞膜处的降低的perk水平和增加的ve-钙粘蛋白积累所示。绿色表示ha染色,它标记表达araf的细胞;红色表示ve-钙粘蛋白染色。
图10:azd8330对表达araf突变s214p的hdlec的作用。azd8330挽救了araf-s214p诱导的细胞形态差异,如通过在细胞膜处的降低的perk水平和增加的ve-钙粘蛋白积累所示。绿色表示ha染色,它标记表达araf的细胞;红色表示ve-钙粘蛋白染色。
图11:瑞法替尼对表达araf突变s214p的hdlec的作用。瑞法替尼挽救了araf-s214p诱导的细胞形态差异,如通过在细胞膜处的降低的perk水平和增加的ve-钙粘蛋白积累所示。绿色表示ha染色,它标记表达araf的细胞;红色表示ve-钙粘蛋白染色。
图12:乌利替尼(ulixertinib)对表达araf突变s214p的hdlec的作用。乌利替尼挽救了araf-s214p诱导的细胞形态差异,如通过在细胞膜处的降低的erk底物rsk3的磷酸化水平和增加的ve-钙粘蛋白积累所示。绿色表示ha染色,它标记表达araf的细胞;红色表示ve-钙粘蛋白染色;白色表示细胞核的dapi染色。
图13:乌利替尼对表达braf突变f486s的hdlec的作用。乌利替尼挽救了braf-f486s诱导的细胞形态差异,如通过在细胞膜处的增加的ve-钙粘蛋白积累所示。绿色表示ha染色,它标记表达braf的细胞;红色表示ve-钙粘蛋白染色;白色表示细胞核的dapi染色。
图14:曲美替尼对表达braf突变f486s的hdlec的作用。曲美替尼挽救了braf-f486s诱导的细胞形态差异,如通过在细胞膜处的降低的erk的磷酸化水平和增加的ve-钙粘蛋白积累所示。绿色表示ha染色,它标记表达braf的细胞;红色表示ve-钙粘蛋白染色;白色表示细胞核的dapi染色。
图15:曲美替尼对表达raf1突变t145p的hdlec的作用。曲美替尼挽救了raf1-t145p诱导的细胞形态差异,如通过在细胞膜处的降低的erk的磷酸化水平和增加的ve-钙粘蛋白积累所示。
图16:乌利替尼和曲美替尼对表达kras突变g12d的hdlec的作用。乌利替尼和曲美替尼挽救了kras-g12d诱导的细胞形态差异,如通过在细胞膜处的降低的erk的磷酸化水平和增加的ve-钙粘蛋白积累所示。
图17:由rit1突变诱导的p-erk的弱激活。
图18:三维淋巴球体发芽试验显示,如通过底部发芽长度衡量的,与ephb4-wt相比,表达三种不同ephb4突变的hdlec中发芽活性升高。雷帕霉素和osi-027均可挽救由突变l778_g779inslmgs诱导的发芽增加。
图19:用mek抑制剂和bez235的治疗可显著改善kras突变g12d引起的水肿。
图20:ptpn11s502a或g503r突变的嵌合体表达(mosaicexpression)导致轻度淋巴管异常表型。
图21:crispr敲除残基749周围的rasa1a和rasa1b上的靶标导致斑马鱼中形成大水肿,单一靶向(rasa1a或rasa1b)或atg靶向不会导致任何表型。
图22a-22g:淋巴管异常先证者的临床图像及分子分析。图22a)t2加权非对比淋巴管造影照片的冠状切片,显示出较大的心包积液(箭头)。图22b)在健康对照者中的动态对比增强的磁共振淋巴管造影照片的最大强度投影,显示出正常的td向左无名静脉移动。图22c)p1中的动态对比增强的磁共振淋巴管造影照片的最大强度投影,显示出扩张的腰椎淋巴管网具有逆行肝门血流(箭头),并且扩张和弯曲的td(箭头)向左侧的无名静脉移动,并且还向纵隔和心包提供逆行灌注(方框)。图22d)c中框内区域的对比淋巴管造影照片,显示扩张和弯曲的远端td具有通过起源于远端td的扩张的淋巴管网朝向纵隔、心包和肺的逆行血流(箭头)。图22e)骨盆和生殖器的冠状最大强度投影,显示起源于双侧腹股沟淋巴结的多个扩张的导管(箭头),并向阴茎和阴囊(箭头)提供逆行血流。图22f)在不相关的亲属中确认出的复发突变(araf中的c.640t>c(p.s214p))的谱系和基因型。图22g)araf蛋白的示意拓扑,其中星号表示cr2中p.s214p突变的位置。ser214残基在脊椎动物物种和所有raf同种型中高度保守。
图23a-23j:araf-s214p突变在hdlec中增加了erk1/2活性,增强了淋巴管生成能力,并改变了肌动蛋白骨架和ve-钙粘蛋白连接,并在斑马鱼中导致胸导管(td)扩张,其可被考比替尼逆转。图23a),heaf293t细胞中的araf突变体转染削弱了与14-3-3蛋白的结合并增加了p-erk。右图示出了标准化的14-3-3/flag比,显示了突变体中14-3-3蛋白的免疫共沉淀减少。数据显示为三个独立实验的平均值±s.e.m.。双尾不成对t检验(自由度(df)为4),****p=8.6×10-6。将图像裁剪以更好地呈现。图23b),与表达wt的细胞相比,hek293t细胞中的araf突变体转染诱导增加的p-erk1/2表达(**p=0.0026;双尾不成对t检验;df=8)。arat-s214p不会改变akt、p70s6k\mtor和p38的磷酸化。标准化的比率由右侧的箱线图(最小到最大值,显示所有点)示出,其中中心线表示中位数,方框范围表示四分位距,而晶须表示最小到最大数据范围。进行了六个独立的实验。将图像裁剪以更好地呈现。图23c)用araf-wt或araf-s214p转导的原代hdlec在浓度逐渐增加的曲美替尼下培养。量化来自三个独立转导的细胞的结果,并将其绘制在散点图上,其中每个单独的值作为一个点叠加。数据显示为三个独立实验的平均值±s.e.m.(误差条)。araf-s214p的转导显著增加了p-erk的水平(*p=0.03;双尾不成对t检验,df为4)。曲美替尼治疗导致p-erk显著减少(对于100nm的曲美替尼治疗和300nm的曲美替尼治疗,*p=0.02;对于1000nm的曲美替尼治疗和3000nm的曲美替尼治疗,*p=0.01;双尾不成对t检验,df为4);ns,不显著。将图像裁剪以更好地呈现。图23d)三维淋巴球体发芽试验显示,如通过在底部的发芽数量(***p=0.0002)和发芽长度(***p=0.0005)所衡量的,与araf-wt相比,表达araf-s214p的hdlec的发芽活性升高。双尾不成对t检验,df为22。还在浓度逐渐增加的曲美替尼下培养球体,这在30nm(****p=4.68×10-5;df=25)、100nm(***p=9.5×10-4;df=25)和300nm(***p=4.4×10-4;df=24)的浓度下显著减少了发芽数量以及在30nm(***p=1.8×10-4;df=25)100nm(**p=0.001;df=23)和300nm(***p=3.2×10-4;df=24)的浓度下显著减少了发芽长度。双尾不成对t检验。量化了使用独立转导的hdlec进行的三个实验,并在交错箱线图上绘制了所有这三个实验的点(每种条件15点)(每个实验3至6个球体),其中中心线表示中位数,方框范围表示四分位距,而晶须表示最小到最大数据范围。图23e)araf突变体影响ve-钙粘蛋白的定位(****p=1.88×10-26),而用曲美替尼的治疗导致ve-钙粘蛋白的细胞表面定位增加(****p=1.63×10-19)。红色箭头表示被称为质膜染色的染色,而黄色箭头表示细胞内染色。量化了使用独立转导的hdlec进行的三个实验用于细胞内和质膜染色,并在左边箱线图上绘制了所有这三个实验的点(每种条件75点)(最小到最大),其中中心线表示中位数,方框范围表示四分位距,而晶须表示最小到最大数据范围。双尾不成对t检验,df为148;ns,不显著。测量了来自图23e中实验的细胞的最大长度和宽度,并计算了长宽比,并将其绘制在右侧箱线图上(最小值至最大值;右下方),其中中心线表示中位数,方框范围表示四分位距,而晶须表示最小到最大数据范围。araf-s214p表达导致长宽比显著增加(****p=9.57×10-15),而用曲美替尼的治疗可使所述比值正常化(****p=7.51×10-17;双尾不成对t检验,df为148;ns,不显著)。图23f)mtt(3-(4,5-二甲基噻唑-2-基)-2,5-二苯基溴化四唑)试验显示,araf-s214p不会导致转导的hdlec的增殖增加。在两个波长(550和700nm)处测量代谢活性。如方法中所述,在指定的时间收集一式三份孔(n=3个独立孔)的内容。所述趋势线连接每个时间点每个转导体的平均值,这些点显示所有数据点的测量值。该实验代表了两个独立的逆转录病毒转导的结果。图23g-图23i)上:鱼淋巴系统总览;白色框表示在图23g-23i中研究的区域,其显示了共聚焦扫描的叠加图(最大强度投影)。td和后主静脉(pcv)被标记为绿色(tg(mrc1a:egfp)),并且将td在图23g–图23i中用虚线标出。将araf转基因(mrc1a:araf)细胞标记为红色(绿色+红色→黄色)。图23g)araf-s214p表达导致td严重扩张。图23h)araf-wt(红色)的表达对td和pcv(均为绿色)的形态没有影响。图23i)考比替尼(1um)部分逆转了由突变诱导的扩张。重复三个独立的实验,在图23g-23i中,具有相似的发现。图23j),针对使用和不使用考比替尼治疗的扩张的身体部分表型评分类别:正常、中度扩张(td已扩展,但可以与pcv分开)以及重度扩张(td与pcv重叠)。考比替尼治疗可显著减少重度扩张(****p=1.33×10-5;单尾不成对t检验;蓝色直方图)并挽救至正常形态(***p=0.00051;单尾不成对t检验;绿色直方图)。在三个独立的实验中,总共分析了40个幼体和120个身体部分。数据显示为三个独立实验的平均值±s.e.m.。未经处理的点图像可用作源数据。
图24a-24f:mek抑制剂治疗前、后,先证者的肺功能测试和临床图像。图24a)mek抑制剂治疗之前和之后的肺功能测试结果。请注意,所有肺量测定度量均得到了显著改善,其中fev1的预测值从23%提高到42%,tlc的预测值从29%提高到56%,最大吸气压力(mip)的预测值从71%提高到115%(标记为红色)。fvc,用力肺活量;fef25-75,中度用力呼气流速;rv,余气量;rv/tlc,rv与tlc之比;dlco[hb],用于血红蛋白的校正的肺一氧化碳弥散量;dlco/va,dlco除以肺泡体积(va);mep,最大呼气压力;o2sat,氧饱和度。图24b)开始药物治疗之前的(左)和mek抑制剂治疗开始后12个月的(右)的t2加权非对比淋巴管造影照片的冠状最大强度投影,证实了大面积扩张和串珠状皮下淋巴管的几乎完全吸收。图24c)治疗前骨盆和胸部的对比淋巴管造影照片的冠状最大强度投影(左),证实了中央淋巴管稀少,隔膜上方缺少中央淋巴流,大量扩张和串珠状的双侧皮下导管沿腹壁移动。开始治疗后十二个月(右),扩张的皮下导管被吸收,形成新的、外观更正常的淋巴网,其沿着腹壁延伸到胸部内。图24d)治疗前骨盆和大腿的对比淋巴管造影照片的冠状最大强度投影(左),还证实了大腿中的导管稀少,淋巴管大量扩张和成串珠状。治疗后(右),再次出现异常扩张的导管吸收,并形成新的和外观更正常的淋巴网。图24e)治疗前(左)和治疗后12个月(右)的胸部x光片显示积液减少,肺活量明显改善。图24f)患者的生长图(左)。曲美替尼治疗始于13岁(*)之前,治疗开始后约3–6个月开始观察到淋巴水肿和临床状况的改善。右上方显示了所述患者在其峰值体重时去除压力袜后下肢的照片。右下方的图像显示了下肢的相应图片。
具体实施方式
ras/有丝分裂原活化蛋白激酶(mapk)通路在细胞增殖、迁移、分化和凋亡中起着至关重要的作用,所有这些对于正常发育都是必不可少的。中枢传导淋巴管异常(ccla)是复杂的淋巴管异常,其特征是淋巴通道扩张、淋巴通道运动障碍和影响淋巴引流的远端阻塞。首先由trenoriii和chaudry以及clemens等人描述,ccla在2015年被国际血管异常研究协会(issva)归类为通道型淋巴畸形,其表现出临床症状与其密切相关的诊断-广泛性淋巴管异常(gla)-的明显的重叠模式,包括但不限于乳糜胸、乳糜性腹水、淋巴液渗漏或倒流以及四肢肿胀。我们最近发现,araf中功能突变的获得是导致淋巴管异常(包括lam、gla和ccla)的原因。在这里,我们将kras、biraf和pnpn11中功能突变的获得描述为淋巴管异常(包括lam、gla和ccla)的致病变异。还描述了sos1、itag9、rasa1、raf1、rit1、piezo1、ephb4、nf1、cbl和araf中的其他突变,这些突变有助于该病症的发病机制。
淋巴管异常是一种罕见的毁灭性疾病,病因大多未知,其治疗取决于患者的表现。鉴定致病基因将允许开发与精准医疗实施保持一致的负担得起的疗法。
在实施例ii中,我们在一名患有晚期异常淋巴疾病的12岁男孩中以及在另一无关系的成年患者中表征了复发性功能获得性araf突变(c.640t>c:p.s214p),所述疾病对常规西罗莫司疗法无反应。所述突变导致保守的磷酸化位点的丢失。用araf-s214p转导的细胞表现出升高的erk1/2活性,增强的淋巴管生成能力以及肌动蛋白骨架的分解和ve-钙粘蛋白连接,这些可使用mek抑制剂曲美替尼进行挽救。通过在斑马鱼模型中重建淋巴表型,也验证了突变的功能相关性,其中使用mek抑制剂挽救异常表型。用mek抑制剂对先证者的后续治疗导致了显著的临床改善,其中患者的淋巴系统重塑、淋巴水肿消退、肺功能测试显著改善、补充氧需求停止、日常活动接近正常。我们的结果提供了一个有代表性的证明,说明遗传分类和机械理解的知识如何指导基于生物学的医学治疗,在我们看来,这是挽救生命的。
现在将详细参考本发明的某些实施方案,其示例在附图中示出。尽管将结合图示的实施方案描述本发明,但是应当理解,它们并不旨在将本发明限制于那些实施方案。相反,本发明旨在覆盖所有替换、修改和等同形式,其可以包括在由所附权利要求限定的本发明之内。
在详细描述本教导之前,应当理解,本公开不限于特定的组成或处理步骤,因为它们可以变化。应当指出的是,如本说明书和所附权利要求书中所使用的,单数形式“一”,“一个”和“该”包括复数对象,除非上下文另外明确指出。因此,例如,对“缀合物”的引用包括多个缀合物,而对“一个细胞”的引用包括多个细胞等。
要理解的是,在本公开中讨论的温度、浓度、时间等之前存在隐含的“约”,使得轻微和无实质的偏差在本文的本教导的范围内。而且,“包含(comprise)”、“包含(comprises)”、“包含(comprising)”、“含有(contain)”、“含有(contains)”、“含有(containing)”、“包括(include)”、“包含(includes)”以及“包括(including)”的使用不意图是限制性的。应当理解,前面的一般描述和详细描述都仅是示例性和说明性的,并且不限制本教导。
除非在以上说明书中特别指出,说明书中陈述“包括”各种组件的实施方案也被认为是“由所述组件组成”或“基本上由所述组件组成”;说明书中陈述“由多种组件组成”的实施方案也被认为是“包括所述组件”或“基本上由所述组件组成”;以及说明书中陈述“基本上由多种组件组成”的实施方案也被认为是“由所述组件组成”或“包括所述组件”(这种互换性不适用于权利要求中这些术语的使用)。
本文使用的章节标题仅用于组织目的,而不应以任何方式被解释为限制所期望的主题。如果通过引用的方式并入的任何文献与本说明书中限定的任何术语相抵触,则以本说明书为准。尽管结合多种实施方案描述了本教导,但是并不意图将本教导限于这样的实施方案。相反,本领域技术人员将理解,本教导涵盖各种替代、修改和等同形式。
此外,化合物“选自以下组成的组”是指以下列表中的一种或多种化合物,其包括两种或更多种所述化合物的混合物(即,组合)。根据本发明,分离的或生物学上纯的分子是已经与其天然环境脱离的化合物。这样,“分离的”和“生物纯的”不一定反映化合物已纯化的程度。本发明的分离的化合物可以从其天然来源获得、可以使用实验室合成技术来生产、或者可以通过任何这样的化学合成途径来生产。
“淋巴管异常”是指以淋巴管的异常形成和组织过度生长为特征的疾病或病症。淋巴管异常的非限制性实例包括“淋巴管瘤病”或“淋巴管扩张”(在本文中统称为lam)、淋巴管瘤、广泛性淋巴管异常(gla)和乳糜性积液、广泛性淋巴管瘤,全身性囊性血管瘤病、多发性淋巴管扩张、广泛性淋巴管畸形、ccla、弥漫性淋巴管畸形、kaposi型lam和gorham-stout疾病(gsd)、罕见的淋巴管源性血管病症(其特征是进行性骨溶解)。
临床上,淋巴管瘤分为几种类型。其中包括(1)单纯型(simplex),它由毛细管尺寸的薄壁淋巴管组成。这种类型通常会影响皮肤(局限性淋巴管瘤);(2)囊性淋巴管瘤(或囊性水瘤):其尺寸范围从几毫米到几厘米,在年轻时候就可见,通常在颈部或腋窝;(3)海绵体型(cavernosum):这种类型由扩张的淋巴管组成,通常带有纤维状外膜层。这是通常影响胸部、腹部和骨骼中的器官的类型。这些淋巴管瘤的每一种都被包括在本发明中。
“单核苷酸变异(snv)”是指基因组dna中的一个位置,其中存在与该位置的通常碱基不同的单个碱基。snv与snp相似,不同之处在于snp通常是指以某个频率出现的snv(例如,发生在超过一定比例的人口中),而snv不提供频率信息。在人类基因组中已经编录有数百万的snv。一些snv导致疾病,而其他snv是基因组中的正常变异。
“淋巴管异常-相关的snv或-特异性标志物”是与发生淋巴管异常的风险增加相关的snv,在没有这种疾病的患者中未发现。这样的标志物可以包括但不限于核酸、由其编码的蛋白质或其他小分子。
如本文所用,术语“遗传改变”是指从一个或多个核酸分子的野生型或参考序列的改变。遗传改变包括但不限于snv和snp、拷贝数变异(cnv)、已知序列的核酸分子的至少一个核苷酸的碱基对取代、添加和缺失。
“连锁”描述了基因、等位基因、基因座或遗传标志物由于其位于同一染色体上而导致一起遗传的趋势,并通过两个基因、等位基因、基因座或遗传标志物之间的重组百分比(也称为重组率,或θ)来衡量。两个基因座在染色体上的物理距离越近,重组率就越低。通常,当测试致病基因中的多态性位点与疾病的连锁时,重组率将为零,这表明所述疾病和致病基因始终是共同遗传的。在极少数情况下,当一个基因跨越基因组的很大一部分时,有可能观察到该基因一端上的多态性位点与另一端上的致病突变之间的重组。但是,如果所述致病突变是正在测试的与疾病连锁的多态性,则不会观察到重组。
“厘摩(centimorgan)”是遗传距离的单位,表示两个遗传标志物、等位基因、基因或基因座之间的连锁,其对应于任何减数分裂事件的两个标志物或基因座之间重组的概率为1%。
“连锁不平衡”或“等位基因关联”是指,特定等位基因、基因座、基因或遗传标志物与附近染色体位置上的特定等位基因、基因座、基因或遗传标志物的优先关联的频率高于所述种群中偶然预期的任何特定等位基因频率。
如本文所用,术语“固体基质”是指任何形式,例如珠、微粒、微阵列、微量滴定孔或试管的表面、浸棒(dipstick)或过滤器。所述基质的材料可以是聚苯乙烯、纤维素、乳胶、硝化纤维、尼龙、聚丙烯酰胺、葡聚糖或琼脂糖。固体基质可以包含固定在其上的核酸,使得它们不能从溶液中的基质上除去。
如本文所用,“靶核酸”是指存在于复杂核酸混合物中的核酸的先前限定的区域,其中所限定的野生型区域包含至少一个已知的核苷酸变异,其可能与淋巴管异常相关或不相关。可以通过cdna克隆或消减杂交或手动合成从天然来源分离所述核酸分子。可以通过三酯合成方法或通过使用自动dna合成仪来人工合成所述核酸分子。
关于本发明中使用的核酸,术语“分离的核酸”应用于dna时,是指从与其起源的生物的天然存在的基因组中紧接连续(在5'和3'方向上)的序列中分离的dna分子。例如,“分离的核酸”可包含插入载体(例如质粒或病毒载体)中的,或整合到原核生物或真核生物的基因组dna中的dna分子。“分离的核酸分子”也可以包含cdna分子。插入载体中的分离的核酸分子在本文中有时也称为重组核酸分子。
关于rna分子,术语“分离的核酸”主要是指由如上定义的分离的dna分子编码的rna分子。或者,该术语可以指从与其天然状态(即,在细胞或组织中)相关联的rna分子中充分分离的rna分子,使得其以“基本上纯的”形式存在。
关于核酸的术语“富集”,是指特定的dna或rna序列在目的细胞或溶液中的总dna或rna中所占的比例明显高于(2-5倍)其在正常细胞中或在提取该序列的细胞中所占的比例。这可能是由于人们通过优先减少存在的其他dna或rna的量,或通过优先增加特定dna或rna序列的量,或通过两者的结合而引起的。但是,应该注意的是,“富集”并不意味着不存在其他dna或rna序列,而只是目的序列的相对数量已显著增加。
对于某些目的,核苷酸序列为纯化形式也是有利的。涉及核酸的术语“纯化的”不要求绝对纯度(例如均质的制备物);相反,它表明该序列比自然环境中的序列相对更纯。
术语“互补的”描述了可以彼此形成多种有利相互作用的两个核苷酸。例如,腺嘌呤与胸腺嘧啶互补,因为它们可以形成两个氢键。同样,鸟嘌呤和胞嘧啶是互补的,因为它们可以形成三个氢键。因此,如果核酸序列包含以下碱基序列:胸腺嘧啶、腺嘌呤、鸟嘌呤和胞嘧啶,该核酸分子的“补体”是包含替代胸腺嘧啶的腺嘌呤、替代腺嘌呤的胸腺嘧啶、替代鸟嘌呤的胞嘧啶和替代胞嘧啶的鸟嘌呤的分子。因为补体可以包含与亲本核酸分子形成最佳相互作用的核酸序列,所以这样的补体可以与其亲本分子以高亲和力结合。
关于单链核酸,特别是寡核苷酸,术语“特异性杂交”是指具有足够互补序列的两个单链核苷酸分子之间的关联,以允许在本领域通常使用的预定条件下进行这种杂交(有时称为“基本上互补”)。具体地,该术语指寡核苷酸与本发明的单链dna或rna分子中包含的基本上互补的序列的杂交,基本排除寡核苷酸与非互补序列的单链核酸的杂交。例如,特异性杂交可以指与任何淋巴管异常特异性标志物核酸杂交但不与其他核苷酸杂交的序列。这样的标志物包括,例如,本文包含的表中所示的淋巴管异常特异性标志物。能够使具有不同互补性的单链核酸分子特异性杂交的合适条件是本领域众所周知的。
例如,下面示出了一种通用公式用于计算实现具有特定序列同源性的核酸分子之间的杂交所需的严格条件(sambrook等人,molecularcloning,coldspringharborlaboratory(1989):
tm=81.5"c+16.6log[na+]+0.41(%g+c)-0.63(%甲酰胺)-600/#双链体中的碱基对(bpinduplex)
作为上述公式的说明,使用[na+]=[0.368]和50%的甲酰胺,gc含量为42%,平均探针大小为200个碱基,tm为57℃。同源性每降低1%,dna双链体的tm就会降低1-1.5℃。因此,使用42℃的杂交温度将观察到具有大于约75%序列同一性的靶标。杂交和洗涤的严格性主要取决于溶液的盐浓度和温度。通常,为了使探针与其靶标的退火速率最大化,杂交通常在比计算的杂交tm低20-25℃的盐和温度条件下进行。洗涤条件应尽可能严格,以确保针对靶标的探针的同一性程度。通常,将洗涤条件选择为低于杂交的tm约12-20℃。在某些方面,本发明的核酸,中等严格杂交定义为在42℃下在6xssc、5xdenhardt溶液、0.5%sds和100μg/ml变性鲑鱼精子dna中的杂交,并在55℃下于2xssc和0.5%sds中洗涤15分钟。高度严格杂交定义为在42℃下在6xssc、5xdenhardt溶液、0.5%sds和100μg/ml变性鲑鱼精子dna中的杂交,并在65℃下于1xssc和0.5%sds中洗涤15分钟。非常高度严格杂交定义为在42℃下在6xssc、5xdenhardt溶液、0.5%sds和100μg/ml变性鲑鱼精子dna中的杂交,并在65℃下于0.1xssc和0.5%sds中洗涤15分钟。
如本文所用,术语“寡核苷酸”定义为在由两个或更多个,优选三个以上的核糖核苷酸或脱氧核糖核苷酸组成的核酸分子。寡核苷酸的确切大小将取决于多种因素以及寡核苷酸的特定应用和用途。包括探针和引物的寡核苷酸可以是从3个核苷酸到核酸分子全长的任何长度,并且明确地包括从3个核苷酸到多核苷酸全长的每个可能数目的连续核酸。优选地,寡核苷酸的长度为至少约10个核苷酸,更优选为至少15个核苷酸,更优选为至少约20个核苷酸。
如本文所用,术语“探针”是指寡核苷酸、多核苷酸或核酸,rna或是dna,无论是天然存在于纯化的限制酶消化物中还是合成产生的,其能够与具有与探针互补的序列的核酸退火或特异性杂交。探针可以是单链的也可以是双链的。探针的确切长度将取决于许多因素,包括温度、探针来源和方法的使用。例如,对于诊断应用,取决于靶序列的复杂性,所述寡核苷酸探针(在某些情况下,与指定rs号相关的核酸与dbsnp数据库中可用的单核苷酸多态性相关)通常包含15-25、15-35、20-50或100个或更多核苷酸,尽管它可能包含更少的核苷酸,但前提是snv的位点包含在探针中。选择本文的探针以与特定靶核酸序列的不同链互补。这意味着探针必须足够互补,以便能够在一组预定条件下与其各自的靶链“特异性杂交”或退火。因此,所述探针序列不必反映靶标的确切互补序列。例如,非互补核苷酸片段可以连接到探针的5’或3’端,探针序列的剩余部分与靶链互补。或者,可以将非互补碱基或更长的序列散布到探针中,条件是所述探针序列与靶标核酸的序列具有足够的互补性,以与其进行特异性退火。
如本文所用,术语“引物”是指寡核苷酸,其可以是rna或dna,可以是单链的或双链的,可以源自生物系统,通过限制性酶消化产生,或合成产生,当置于适当的环境中时,其能够在功能上充当模板依赖性核酸合成的引发剂。当提供合适的核酸模板、合适的核酸三磷酸核苷前体、聚合酶、合适的辅因子和条件(如合适的温度和ph)时,所述引物可以在其3'末端通过添加核苷酸(通过聚合酶作用或类似活性)来延伸,以产生引物延伸产物。引物的长度可以根据特定条件和应用要求而变化。例如,在诊断应用中,寡核苷酸引物的长度通常为15-25、15-40、20-50个等或更多个核苷酸。所述引物必须与所需模板具有足够的互补性,以引发所需延伸产物的合成,也就是说,能够以足以提供适当的并列的引物的3’羟基部分的方式与所需模板链退火,以用于由聚合酶或类似酶起始合成。不需要所述引物序列代表所需模板的精确互补。例如,非互补核苷酸序列可以连接到另外的互补引物的5'末端。或者,非互补碱基可以散布在寡核苷酸引物序列中,前提是所述引物序列与所需模板链的序列具有足够的互补性,从而在功能上为延伸产物的合成提供模板-引物复合物。
聚合酶链式反应(pcr)已经在美国专利4,683,195、4,800,195和4,965,188中进行了描述,其全部公开内容通过引用的方式并入本文。
“sirna”是指通过提供与靶基因的序列具有同源性的小干扰rna(sirna)而参与序列特异性转录后基因沉默或基因敲落的rna干扰过程中的分子。小干扰rna(sirna)可以在体外合成,也可以通过核糖核酸酶iii从更长的dsrna切割而产生,并且是序列特异性mrna降解的介体。优选地,使用适当保护的核糖核苷亚磷酰胺和常规dna/rna合成仪化学合成本发明的sirna。所述sirna可以合成为两个单独的互补rna分子,也可以合成为具有两个互补区域的单个rna分子。合成rna分子或合成试剂的商业供应商包括appliedbiosystems(fostercity,ca,美国)、proligo(hamburg,德国)、dharmaconresearch(lafayette,colo.,美国)、piercechemical(partofperbioscience,rockford,ill.,美国)、glenresearch(sterling,va.,美国)、chemgenes(ashland,mass.,美国)以及cruachem(glasgow,英国)。例如,用于抑制淋巴管瘤病mrna的特异性sirna构建体的长度可以在15-35个核苷酸之间,更典型地,其长度可以为约21个核苷酸。
术语“载体”涉及可以被感染、转染或转化到细胞中并独立地或在宿主细胞基因组内复制的单链或双链环状核酸分子。可以切割环状双链核酸分子,从而在用限制酶处理后线性化。载体、限制酶的种类以及限制酶所靶向的核苷酸序列的知识对于本领域技术人员来说是容易获得的,并包括任何复制子(例如质粒、粘粒、杆粒、噬菌体或病毒),在其上可以连接另一个遗传序列或元件(dna或rna),从而复制所连接的序列或元件。通过用限制酶切割载体并将两个片段连接在一起,可以将本发明的核酸分子插入载体。当克隆包含重复或缺失的遗传区域时,本领域技术人员充分意识到,在克隆过程中将采用合适长度(例如,50-100个或更多个核苷酸之间)的受影响区域上游和下游的侧翼序列。这样的载体例如在细胞系中将具有实用性,用于研究这种改变对编码的蛋白质的影响。
本领域技术人员可以使用许多技术来促进所述表达构建体转化、转染或转导进入原核或真核生物。术语“转化”、“转染”和“转导”是指将核酸和/或表达构建体插入细胞或宿主生物中的方法。这些方法涉及多种技术,例如用高浓度的盐、电场或去污剂处理细胞,使宿主细胞的外膜或壁可渗透目的核酸分子、显微注射、peg融合等。
术语“启动子元件”描述了掺入载体中的核苷酸序列,该核苷酸序列一旦进入适当的细胞,便可以促进转录因子和/或聚合酶的结合以及随后将载体dna的部分转录成mrna。在一个实施方案中,本发明的启动子元件在淋巴管异常特异性标志物核酸分子的5'端之前,使得后者被转录成mrna。然后宿主细胞机制将mrna翻译成多肽。启动子元件可以驱动目的编码区的组成型或诱导型表达。
本领域技术人员将认识到,核酸载体可以包含除启动子元件和编码淋巴管异常特异性标志物的核酸以外的核酸元件。这些其他核酸元件包括但不限于复制起点、核糖体结合位点、编码抗药性酶或氨基酸代谢酶的核酸序列以及编码分泌信号、定位信号或对多肽纯化有用的信号的核酸序列。
“复制子”是能够主要在其自身控制下复制的任何遗传元件,例如质粒、粘粒、杆粒、质体、噬菌体或病毒。复制子可以是rna或dna,并且可以是单链或双链。
“表达操纵子”是指可以具有转录和翻译控制序列的核酸片段,所述转录和翻译控制序列例如启动子、增强子、翻译起始信号(例如atg或aug密码子)、聚腺苷酸化信号,终止子等,并且其促进多肽编码序列在宿主细胞或生物中的表达。
如本文所用,术语“报道子”、“报道子系统”、“报道子基因”或“报道子基因产物”应表示可操作的遗传系统,其中核酸包含编码产物的基因,所述产物表达后可产生(例如通过生物测定、免疫测定、放射免疫测定、或通过比色、荧光、化学发光或其他方法)易于测量的报道子信号。所述核酸可以是rna或dna、线性的或环状的、单链的或双链的、反义的或有义极性的,并且可操作地连接至必要的控制元件以表达报道子基因产物。所需的控制元件将根据报道子系统的性质以及所述报道子基因是dna还是rna形式而有所不同,但是其可以包括但不限于,如启动子、增强子、翻译控制序列、聚a添加信号、转录终止信号等的元件。
所引入的核酸可以或可以不整合(共价连接)到受体细胞或生物的核酸中。例如,在细菌、酵母、植物和哺乳动物细胞中,所引入的核酸可以保持为游离基因元件或独立的复制子,例如质粒。或者,所引入的核酸可以整合到受体细胞或生物的核酸中,并稳定地维持在该细胞或生物中,并进一步传递或遗传给受体细胞或生物的后代细胞或生物。最后,所引入的核酸可能仅短暂地存在于受体细胞或宿主生物中。
术语“可选择标志基因”是指当表达时赋予转化细胞以可选择表型(例如抗生素抗性)的基因。
术语“可操作地连接”是指将编码序列表达所必需的调节序列置于dna分子中相对于编码序列的适当位置,以实现编码序列的表达。这一定义有时适用于表达载体中转录单元和其他转录控制元件(例如增强子)的布置。
术语“重组生物”或“转基因生物”是指具有基因或核酸分子的新组合的生物。可以使用本领域技术人员可以使用的多种核酸操纵技术将基因或核酸分子的新组合引入生物。术语“生物”涉及由至少一个细胞组成的任何生命。生物可以像一个真核细胞一样简单,也可以像哺乳动物一样复杂。因此,短语“重组生物”涵盖重组细胞以及真核和原核生物。示例性的转基因生物包括斑马鱼或小鼠。
本文有时使用术语“分离的蛋白质”或“分离和纯化的蛋白质”。该术语主要指通过本发明的分离的核酸分子的表达产生的蛋白质。或者,该术语可以指与其天然相关的其他蛋白质充分分离,使得以“基本上纯的”形式存在的蛋白质。“分离的”并不意味着排除与其他化合物或材料的人工或合成混合物,或排除不干扰基本活性的杂质的存在,并且所述杂质可能是存在的,例如由于不完全纯化、添加稳定剂或混入例如免疫原性制剂或药学上可接受的制剂中。
“特异性结合对”包括特异性结合成员(sbm)和结合伴侣(bp),它们彼此具有特定的特异性,并且在正常条件下优先于其他分子彼此结合。特异性结合对的实例是抗原和抗体、配体和受体以及互补核苷酸序列。技术人员知道许多其他实例。此外,术语“特异性结合对”也适用于特异性结合成员和结合伴侣之一或两者包含一部分大分子的情况。在其中特异性结合对包含核酸序列的实施方案中,它们将具有在试验条件下彼此杂交的长度,优选大于10个核苷酸长,更优选大于15或20个核苷酸长。
“样品”或“患者样品”或“生物样品”通常是指可以测试特定分子的样品,所述分子优选为淋巴管异常特异性标志物分子,例如下表中所示的标志物。样品可以包括但不限于细胞、体液,所述体液包括血液、血清、血浆、尿液、淋巴液、唾液、眼泪、胸膜液等。
“基因型序列信息”通常是指与受试者的dna或rna序列有关的任何信息。基因型序列信息包括受试者基因组内的目的区域的完整基因组、完整外显子组测序、外显子组测序或靶向测序。基因型序列信息还可包括关于特定snv存在与否的数据的产生,例如在本文中发现与淋巴管异常相关的snv。另外,基因型序列信息将包括使用探针检测一种或多种与淋巴管异常相关的snv的存在和/或表达。如何将探针用于获取基因型序列信息的实例包括但不限于:(1)原位杂交;(2)southern杂交(3)northern杂交;(4)各种扩增反应,例如聚合酶链式反应(pcr)。
术语“试剂”和“测试化合物”在本文可互换使用,表示化学化合物、化学化合物的混合物、生物大分子或由生物材料制成的提取物,所述生物材料例如细菌、植物、真菌或动物(尤其是哺乳动物)细胞或组织。生物大分子包括sirna、shrna、反义寡核苷酸、肽,肽/dna复合物、以及任何基于核酸的分子,其能够调节本文所述的含snv的核酸或其编码的蛋白质的活性。示例性试剂包括但不限于至少一种mek抑制剂。其他试剂还包括表1-2中列出的试剂。通过包括在下文所述的筛选试验中来评估试剂的潜在生物活性。
如本文所用,“治疗”涵盖用于哺乳动物(包括人类)的疾病的治疗剂的任何施用或应用,其包括抑制疾病或疾病的进展、抑制或减缓疾病或其进展、阻止其发展、部分或完全缓解疾病、预防疾病发作或预防疾病症状复发。示例性治疗包括以有效剂量施用至少一种mek抑制剂和/或表1-2中所列的药剂的至少一种。
术语“抑制(inhibition)”或“抑制(inhibit)”是指减少或停止任何事件(例如蛋白质配体结合),或减少或停止任何表型特征,或减少或停止该特征的发生率、程度或可能性。“减少”或“抑制”是指与参照相比减少、降低或阻止活性、功能和/或数量。不需要完全抑制或降低。例如,在某些实施方案中,“降低”或“抑制”是指引起总体减少20%或更多的能力。在另一实施方案中,“降低”或“抑制”是指引起总体减少50%或更多的能力。在另一实施方案中,“降低”或“抑制”是指引起总体减少75%、85%、90%、95%或更多的能力。
术语“抑制剂”是指减慢或阻止特定化学反应、信号转导途径或其他过程的试剂,或降低特定反应物、催化剂或酶的活性的试剂。
术语“患者”和“受试者”可互换使用,是指哺乳动物(包括人类)。
术语“mek”是指mapk/erk通路(也称为ras-raf-mek-erk通路),其包含细胞中的一系列蛋白质,所述一系列蛋白质将信号从细胞表面上的受体传达至细胞核中的dna。当信号分子与细胞表面的受体结合时,信号开始,而当细胞核中的dna表达蛋白质并在细胞中产生某些变化(例如细胞分裂)时,信号终止。所述途径包括许多蛋白质,包括mapk(促有丝分裂原激活蛋白激酶,原来称为erk,细胞外信号调节激酶),它们通过在邻近的蛋白质上添加磷酸基团进行通讯,从而充当“开”或“关”开关。
术语“mek抑制剂”或“mek/erk抑制剂”是指抑制促有丝分裂原激活蛋白激酶mek1、mek2和/或erk的试剂。它们可用于影响mapk/erk途径,所述mapk/erk途径在某些癌症中常常过度活跃。术语“细胞信号转导”将包括mtor信号转导以及控制细胞稳态或活性的任何其他信号转导途径过程。
诊断患有淋巴管异常的患者
在一些实施方案中,在从获自患者的生物样品中获得基因型序列信息之后,基于snv的存在,来诊断具有淋巴管异常的患者。在一些实施方案中,基于在选自ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1,ephb4、nf1、araf和cbl的基因中检测到一种或多种snv的存在,或检测到与选自ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1,ephb4、nf1、araf和cbl的基因中的snv处于连锁不平衡的snv的存在来诊断患有淋巴管异常的患者,上述基因与淋巴管异常相关。在一些实施方案中,所述一种或多种snv是ptpn11基因中的c.1504t>g:ps502a、c.1510a>g:pm504v和/或c.1507g>c:pg503r,kras中的c.35g>a:pg12d,braf基因中的c.1403t>c:pf468s,sos1基因中的c.2536g>a:pe846k,以及包含itga9基因中的c.1236+4a>g和c.289t>g:p.c97g的复合突变或表3中列出的任何突变。
在一些实施方案中,可以从实验数据中生成确认存在于特定受试者中的snv的报告。在一些实施方案中,确认针对淋巴管异常的建议治疗的报告可以基于使用基因型序列信息确认的关于snv的数据来生成。
在一些实施方案中,在从获自患者的生物样品中获得基因型序列信息后,基于以下的诊断可以指导针对患者的治疗的选择:检测到在选自ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1、ephb4、nf1、araf和cbl的基因中一种或多种snv的存在,或检测到与选自ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1,ephb4、nf1、araf和cbl的基因中的snv处于连锁不平衡的snv的存在。在一些实施方案中,在从获自患者的生物样品中获得基因型序列信息后,基于以下的诊断不会指导或影响针对患者的治疗的选择:检测到在选自ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1、ephb4、nf1、araf和cbl的基因中一种或多种snv的存在,或检测到与选自ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1,ephb4、nf1、araf和cbl的基因中的snv处于连锁不平衡的snv的存在。
在一些实施方案中,仅基于临床表现、扫描结果和/或家族史来进行淋巴管异常的诊断。在一些实施方案中,无需测试遗传序列信息就可以进行淋巴管异常的诊断。在一些实施方案中,基于临床表现以及遗传序列信息来进行淋巴管异常的诊断。
本发明中公开的与淋巴管异常有关的snv可以以多种方式用于诊断淋巴管异常。
例如,包含与淋巴管异常相关的snv的核酸可以被用作探针来检测淋巴管异常特异性标志物的存在和/或表达。可以将与淋巴管异常相关的标志物核酸用作探针以用于此类测定的方法包括但不限于:(1)原位杂交;(2)southern杂交;(3)northern杂交;(4)各种扩增反应,例如聚合酶链式反应(pcr)。
此外,用于检测与淋巴管异常相关的snv或由其编码的蛋白质的试验,可以对任意类型的生物样品进行,包括但不限于体液(包括血液、尿液、血清、胃灌洗液)、任意类型的细胞(例如脑细胞、白细胞、单核细胞)或身体组织。
含与淋巴管异常相关的snv的核酸、表达其的载体、含与淋巴管异常相关的snv的标志物蛋白以及抗淋巴管异常特异性标志物抗体可被用于在体组织、细胞或体液中检测与淋巴管异常相关的snv,并改变含与淋巴管异常相关的snv的标志物蛋白表达,以检测和诊断淋巴管异常。
包括了基于与淋巴管异常相关的snv的检测和/或诊断淋巴管异常的方法。所述方法可以包括检测与淋巴管异常相关的snv,样本中包含与淋巴管异常相关的snv的核酸将首先被扩增(例如使用pcr),以与样品中存在的其他序列相比增加模板的数量。这允许以高度灵敏度检测靶标序列,如果它们存在于样品中。可以通过使用在本领域中变得越来越重要的高度灵敏的阵列技术来避免该初始步骤。
或者,新的检测技术可以克服这一局限性,并能够分析包含低至1μg总rna的小样本。与传统的荧光技术相比,使用共振光散射(rls)技术,多次读取可以使用生物素标记的杂交靶标和抗生物素抗体检测少量的mrna。pcr扩增的另一种替代涉及平面波导技术(pwg),以增加信噪比并减少背景干扰。两种技术都可以从qiageninc.(美国)商购获得。
因此,任何前述技术可用于检测或定量与淋巴管异常相关的snv标志物表达,并因此并用于诊断淋巴管异常或其发展的风险。
治疗患有淋巴管异常的患者
阐明本文所述的与淋巴管异常相关的snv在调节淋巴管异常表型中所起的作用有助于重新利用现有疗法,并开发出可用于治疗淋巴管异常的新疗法。在一些实施方案中,本发明包括向患有淋巴管异常的患者施用一种或多种mtor抑制剂、一种或多种pik3k抑制剂和/或一种或多种mek抑制剂(例如,表1-2中的一种或多种剂)。
在一些实施方案中,已经基于淋巴管异常的症状和阳性家族史诊断了待治疗的患有淋巴管异常的患者。在一些实施方案中,多种扫描技术,例如平片放射线照相、骨扫描、计算机断层扫描、磁共振成像和淋巴显像与临床表现一起被用于诊断淋巴管异常。在一些实施方案中,进行活检以诊断淋巴管异常。在一些实施方案中,基于淋巴管过度生长来诊断淋巴管异常。在一些实施方案中,基于淋巴管的异常形成来诊断淋巴管异常。在一些实施方案中,基于乳糜性积液(包括心包、胸膜或腹膜积液)来诊断淋巴管异常。
在一些实施方案中,已经根据本文所述的诊断方法诊断待治疗的患有淋巴管异常的患者。
在一些实施方案中,一种或多种mek抑制剂(例如,表1-2的一种或多种剂;实施例ii)可用于制备治疗淋巴管异常的药物。可以将所述一种或多种剂与药学上可接受的赋形剂、载体、缓冲剂、稳定剂或本领域技术人员所熟知的其他材料一起配制。此类材料应是无毒的,并且不应干扰活性成分的功效。载体或其他材料的确切性质可以取决于施用途径,例如口服、静脉内、皮肤或皮下、鼻、雾化、肌内和腹膜内途径。包含淋巴管异常相关突变的体外系统或转基因生物可用于选择特定药剂用于治疗人类。
可用于治疗的药剂包括但不限于表1和表2的药剂。表1和表2都列出了某些药剂,并且它们未在这两个表中都列出的事实不应被赋予任何含义。
表1
表2提供了可单独使用或与表1或表2中的任何药剂组合使用的药剂,以治疗淋巴管异常。
表2
*iso-27的结构(mateo等人(2016)br.j.cancer114(8):889-96*
为了治疗患有淋巴管异常的个体,或减轻该疾病的病征或症状,可以组合施用靶向本文公开的基因的合适药剂,以便为患者提供治疗益处。此类药剂应以有效剂量施用。
一旦确认遗传改变,就可以设计疗法来调节被改变了的基因影响的生物学途径和信号转导途径。例如,在需要抑制mapk(mek1/mek2)和erk途径的情况下,mek抑制剂可单独使用或与其他mek/erk抑制剂组合使用。在某些实施方案中,治疗需要将表1或2中列出的药剂(如mtor抑制剂)与pik3k抑制剂一起施用。在其他实施方案中,将mtor抑制剂和mek/erk抑制剂组合以为患者提供治疗益处。在另一种方法中,将pik3k和mek/erk抑制剂组合以改善疾病症状。对于本文所述的特异性araf功能突变的获得,有效的疗法包括施用mek/erk抑制剂。上述的组合疗法可以以加和的方式起作用。在其他实施方案中,所组合的药剂协同作用以减轻症状。
首先,可以从患者获得生物学样品和/或基因分型信息。然后评估从样本中存在的核酸中收集的遗传信息,例如是否存在与淋巴管异常相关的snv。表明淋巴管异常风险或疾病存在的这些突变的存在,以及同时确认出受影响的基因,为临床医生提供了有关哪些治疗剂是合适的指导。一个或多个总治疗剂量(当要调节两个或更多个靶标时)可以以单次剂量施用于受试者,也可以使用分级治疗方案施用,其中在更长的时间内施用多次/单独的剂量,例如,在一天的时间内允许以每日剂量施用,或在更长的时间段内,在需要的时间段内施用剂量。本领域技术人员将知道,在受试者中获得有效剂量所需的淋巴管异常药剂的量取决于许多因素,包括受试者的年龄、体重和总体健康状况,以及施用途径和要施用的治疗的数量。鉴于这些因素,本领域技术人员将调整特定剂量,以获得用于治疗患有淋巴管异常的个体的有效剂量。
淋巴管异常治疗剂的有效剂量将取决于施用方式和所治疗个体的体重。本文所述的剂量通常是普通成年人的剂量,但是可以进行调整以用于儿童的治疗。所述剂量通常为约0.001mg至约1000mg。
在患有淋巴管异常,特别是患有较严重形式的所述疾病的个体中,当与例如用于治疗这种疾病的常规药物组合施用时,淋巴管异常治疗剂的施用是特别有用的。技术人员将单独或组合施用淋巴管异常治疗剂,并将使用常规方法监测此类治疗的有效性,所述方法例如,肺、肠、甲状腺或炎性功能测定、放射或免疫学试验、或在指明的情况下,采用组织病理学方法。其他用于治疗淋巴管异常的药剂包括全身化疗、α干扰素疗法、放疗或手术,以减轻该疾病的症状。
药物制剂的施用优选为“有效量”,其足以显示出对个体的益处。该量防止、缓解、减缓或以其他方式降低患者的淋巴管异常症状的严重性。用有效量的mek抑制剂和/或表1-2)的药剂治疗淋巴管异常的患者,可能会改善淋巴结构、减少乳糜性胸膜积液、改善呼吸功能、逐渐减少伴随用药或提高生存率。
所述药物制剂可以被配制成剂量单位形式,以便于给药和剂量的一致性。如本文所用,剂量单位形式是指适合于接受治疗的患者的药物制剂的物理上离散的单位。每个剂量应包含一定量的活性成分,所述成分经计算可与选定的药物载体一起产生所需的效果。确定合适剂量单位的规程是本领域技术人员众所周知的。
剂量单位可以基于患者的体重成比例地增加或减少。如本领域已知的,用于缓解特定病理状况的适当浓度可以通过剂量浓度曲线计算来确定。
可用于本发明方法的药物组合物可以肠胃外、口服固体和液体制剂的形式全身施用,或以皮下、皮内、肌肉内、舌下、局部、耳廓(otic)、颊、结膜、皮肤、牙齿、经电渗透、子宫颈内、窦道或气管、肠内、硬膜外、经渗透、间质、腹腔内、动脉内、关节内、胆内、支气管内、囊内、心脏内、软骨内、尾内、海绵窦内、腔内、脑内、皮内、淋巴管内、心包内、腹膜内、鼻腔、经皮、呼吸道、眼睛、栓剂、气雾剂、局部或其他已知的施用途径施用。除了可用于治疗淋巴管异常的药剂外,所述药物组合物还可包含药学上可接受的载体和其他已知能增强和促进药物施用的成分。因此,这样的组合物可以任选地包含其他组分,例如佐剂,例如氢氧化铝和氢氧化镁的水性悬浮液,和/或其他药学上可接受的载体,例如盐水。其他可能的制剂,例如纳米颗粒、脂质体、再封红细胞和基于免疫的系统,也可以用于根据本发明的方法向患者递送/施用适当的药剂。使用纳米颗粒来递送此类试剂以及可以使用的细胞膜可渗透肽载体描述于crombez等人,biochemicalsocietytransactionsv35:p44(2007)。
在成功检测或定量与淋巴管异常相关的snv标志物表达并因此诊断淋巴管异常或其发展的风险之后,可完成可用于治疗淋巴管异常的药剂的施用。检测或定量与淋巴管异常相关的snv标志物表达可以指导用于治疗的特定药物的选择。检测或定量与淋巴管异常相关的snv标志物表达可以表明特定治疗不适用于给定的受试者。
在其他实施方案中,可以基于疾病的临床诊断来进行淋巴管异常的治疗,并且可以在不检测或量化遗传序列信息的情况下开始治疗。在其他实施方案中,可以基于疾病的临床诊断来进行淋巴管异常的治疗,并且可以在不检测或量化与淋巴管异常相关的snv标志物表达的情况下开始治疗。
在其他实施方案中,可以基于疾病的临床诊断来进行淋巴管异常的治疗,并且当与淋巴管异常相关的snv标志物表达与对照组无差异时,可以开始治疗。
在一些实施方案中,在ptpn11、kras、braf、sos1、itga9、rasa1、raf1、rit1、peizo1、ephb4、nf1、araf和cbl中没有snv的患者中施用治疗。
在一些实施方案中,所述抑制剂是mek1/2抑制剂,其抑制促有丝分裂原激活蛋白激酶mek1和/或mek2。它们可用于影响mapk/erk途径,所述mapk/erk途径在某些癌症和其他病症中常常过度活跃。
在一些实施方案中,所共同施用的药剂是雷帕霉素或bez-235(dactolisib)。雷帕霉素,一种mtor抑制剂,也被称为西罗莫司。bez-235,也称为dactolisib或nvp-bez235,是对p110、pi3k和mtor具有已知活性的化合物。
在一些实施方案中,在治疗方法中所施用的药剂选自:雷帕霉素(西罗莫司)、依维莫司(rad001)、azd8055、temsirolimus(cci-779、nsc683864)、ku-0063794、mhy1485、bez235(nvp-bez235、dactolisib)、pi-103、torkinib(pp242)、他克莫司(fk506)、地磷莫司(deforolimus、mk-8669)、ink128(mln0128)、voxtalisib(sar245409、xl765)、torin1、奥米利塞(gsk2126458、gsk458)、osi-027、pf-04691502、apitolisib(gdc-0980、rg7422)、gsk1059615、gedatolisib(pf-05212384、pki-587)、wye-354、azd2014、torin2、wye-125132(wye-132)、pp121、wye-687、ch5132799、way-600、etp-46464、gdc-0349、xl388、唑他莫司(abt-578)、他克莫司(fk506)、bgt226(nvp-bgt226)、palomid529(p529)以及大黄根酸。
在一些实施方案中,待施用的药剂是选自以下的mek抑制剂:司美替尼(azd6244)、pd0325901、曲美替尼(gsk1120212)、pd184352(ci-1040)、pimasertib(as-703026)、tak-733,azd8330、比美替尼(mek162、arry-162、arry-438162)、sl-327、瑞法替尼(rdea119,bay86-9766)以及考比替尼(gdc-0973、rg7420)。
上述药剂的组合也可以用于本文所述的治疗方法中以治疗淋巴管异常。在一些实施方案中,以下组合可以以加和或协同方式起作用,以治疗淋巴管异常(包括gla和lam)。在某些实施方案中,用于施用的组合选自1)地磷莫司和曲美替尼;2)地磷莫司和司美替尼或考比替尼;3)bez235和司美替尼;4)奥米利塞和司美替尼或曲美替尼;5)依维莫司和曲美替尼或司美替尼;6)西罗莫司、地磷莫司和司美替尼;7)西罗莫司、地磷莫司和曲美替尼;8)torkinib和曲美替尼;9)bez235、torkinib和曲美替尼;以及10)西罗莫司和gedatolisib和曲美替尼。
在一些实施方案中,用本文列出的药剂进行的治疗与全身化疗、干扰素α、放疗和/或手术中的一种或多种组合使用。
确认其他有用治疗剂的方法
由于本文确认的snv已与淋巴管异常的病因相关,因此确认调节基因活性的试剂及其包含此类snv的编码产物的方法应导致产生用于治疗该病况的有效治疗剂。
本文所述的染色体区域包含蛋白质编码区,其提供合适的靶标用于合理设计调节其活性的治疗剂。可以使用对应于这些区域的小肽分子,以有利于设计可有效调节编码蛋白活性的治疗剂。
分子建模应有助于根据功能所需的构象或关键氨基酸残基,确认具有结合至由含snv核酸所编码的蛋白质活性位点的能力的特定有机分子。将使用组合化学方法来确认具有最大活性的分子,然后将开发这些分子的迭代以用于进一步的筛选循环。在某些实施方案中,可以从合成或天然化合物的大型文库中筛选候选药物。一个例子是fda批准的人类可以使用的化合物文库。另外,化合物文库可从许多公司商购获得,其包括但不限于maybridgechemicalco.(trevillet,康沃尔,英国)、comgenex(普林斯顿,nj)、microsource(新米尔福,ct)、aldrich(密尔沃基,wi)、akosconsultingandsolutionsgmbh(巴塞尔,苏格兰)、ambinter(巴黎,法国)、asinex(莫斯科,俄罗斯)、aurora(格拉茨,澳大利亚)、biofocusdpi,瑞士,bionet(卡姆尔福德,英国)、chembridge,(圣地亚哥,ca)、chemdiv,(圣地亚哥,ca)、chemicalblocklt,(莫斯科,俄罗斯)、chemstar(莫斯科,俄罗斯)、exclusivechemistry,ltd(奥布宁斯克,俄罗斯)、enamine(基辅,乌克兰)、evotec(汉堡,德国)、indofine(希斯堡,nj)、interbioscreen(莫斯科,俄罗斯)、interchim(蒙吕松,法国)、lifechemicals,inc.(奥兰治,ct)、microchemistryltd.(莫斯科,俄罗斯)、otava,(多伦多,on)、pharmexltd.(莫斯科,俄罗斯)、princetonbiomolecular(蒙茅斯章克申,nj)、scientificexchange(森特奥西皮,nh)、specs(代尔夫特,荷兰)、timtec(纽瓦克,de)、torontoresearchcorp.(北约克on)、ukrorgsynthesis(基辅,乌克兰)、vitas-m,(莫斯科,俄罗斯)、zelinskyinstitute,(莫斯科,俄罗斯)和bicoll(上海,中国)。
细菌、真菌、植物和动物提取物形式的天然化合物文库是可商购的,或者可以通过本领域众所周知的方法容易地制备。建议可以将从天然来源(例如动物、细菌、真菌、植物来源(包括叶和树皮))以及海洋样品中分离的化合物作为候选物进行测定,以检测潜在有用药物试剂的存在。应当理解,待筛选的药物试剂也可以从化学组合物或人造化合物衍生或合成。在筛选中可以使用数种商业文库。
用于药物筛选试验的多肽或片段可以是溶液中游离的、固定至固体支持物上或在细胞内的。一种药物筛选方法利用真核或原核宿主细胞,其优选地在竞争性结合试验中用表达所述多肽或片段的重组多核苷酸稳定转化。活的或固定形式的此类细胞可用于标准结合试验。例如,可以确定多肽或片段与被测试剂之间的复合物的形成,或者检查多肽或片段与已知底物之间的复合物的形成受被测试剂干扰的程度。
用于药物筛选的另一种技术涉及使用宿主真核细胞系、细胞(例如内皮细胞)或完整动物模型(例如转基因小鼠或斑马鱼),其具有无功能或改变的淋巴管异常相关基因。在某些情况下,转基因生物包含具有以下突变的细胞:ptpn11基因中的c.1504t>g:ps502a、c.1510a>g:pm504v和/或c.1507g>c:pg503r,kras中的c.35g>a:pg12d,braf基因中的c.1403t>c:pf468s,sos1基因中的c.2536g>a:pe846kin,以及包含itga9基因中的c.1236+4a>g和c.289t>g:p.c97g的复合突变或表3中所示的一种或多种突变。这些宿主细胞系、细胞或转基因动物在多肽水平上是有缺陷的。所述宿主细胞系或细胞在药物化合物存在下生长。例如,在斑马鱼模型中,可以评估尾血管和/或d/v血管结构的挽救。另外,可以评估在宿主细胞系中由mtor引起的磷酸化诱导。
药物筛选的示例方法是用于确认改变细胞信号转导的试剂(例如表1-2中列出的试剂)的方法。该方法将包括提供表达至少一种核酸的细胞,所述核酸包含至少一种与淋巴管异常相关的snv;提供表达对应于所述与淋巴管异常相关的snv的同源野生型序列的细胞;使表达至少一种与淋巴管异常相关snv的细胞和表达同源野生型序列的细胞与测试试剂接触;以及分析所述试剂是否改变细胞信号转导。
表达本发明的与淋巴管异常相关的snv或其功能片段的宿主细胞提供了一种系统,在该系统中针对调节淋巴管异常发生的能力筛选潜在的化合物或试剂。因此,在一个实施方案中,本发明的核酸分子可用于产生重组细胞系,用于试验,以确认调节与淋巴管异常和异常血管形成有关的异常mapk信号转导方面的试剂。本文还提供了筛选化合物的方法,所述化合物能够调节由含snv的核酸编码的蛋白质的功能。
另一方法需要使用噬菌体展示文库,其经工程改造以在噬菌体表面上表达由含snv的核酸编码的多肽片段。然后在可检测到表达的肽与化学文库组分之间的结合亲和力的条件下,使此类文库与组合化学文库接触。美国专利6,057,098和5,965,456提供了进行这种试验的方法和设备。
在另一个实施方案中,与淋巴管异常相关的改变的核酸的可用性使得能够产生携带本发明的改变的核酸的实验室小鼠品系。这些与淋巴管异常相关的改变的核酸可能是ptpn11基因中的c.1504t>g:ps502a、c.1510a>g:pm504v和/或c.1507g>c:pg503r,kras中的c.35g>a:pg12d,braf基因中的c.1403t>c:pf468s,sos1基因中的c.2536g>a:pe846k,以及包含itga9基因中的c.1236+4a>g和c.289t>g:p.c97g的复合突变或表3中所示的任何其他突变。表达本发明的与淋巴管异常相关的突变的转基因小鼠提供了一种模型系统,其中可以检查由突变的核酸编码的蛋白质在淋巴管异常发生和进展中的作用。在实验室小鼠中引入转基因的方法是本领域技术人员已知的。三种常见方法包括:1.将编码目的外源基因的逆转录病毒载体整合到早期胚胎中;2.将dna注入新受精卵的原核;以及3.将经过基因处理的胚胎干细胞整合到早期胚胎中。上述转基因小鼠的产生将有助于分子阐明靶蛋白在与淋巴管异常表型有关的各种过程中所起的作用。这样的小鼠提供了在整个动物模型中研究推定的治疗药物的体内筛选工具,并且被本发明涵盖。
本文使用的术语“动物”包括人类以外的所有脊椎动物。它还包括处于各个发育阶段(包括胚胎和胎儿阶段)的个体动物。“转基因动物”是指任何含有一个或多个带有遗传信息的细胞的动物,这些遗传信息通过在亚细胞水平上的有意遗传操作(例如通过靶向重组、显微注射或重组病毒感染)而直接或间接地被改变或接收。术语“转基因动物”并非旨在涵盖经典的杂交或体外受精,而是旨在涵盖其中一个或多个细胞被重组dna分子改变或接受重组dna分子的动物。该分子可以专门针对特定的遗传基因座,被随机整合在染色体中,或者它也可以是染色体外复制的dna。术语“生殖细胞系转基因动物”是指这样的转基因动物,其中将遗传改变或遗传信息引入生殖细胞,从而赋予将遗传信息转移给后代的能力。如果这些后代实际上拥有这种改变或遗传信息的一些或全部,那么它们也是转基因动物。
遗传信息的改变可能对接收者所属的动物物种而言是外来的,或者仅对特定的个体接收者而言是外来的,或者可能是接收者已经拥有的遗传信息。在最后一种情况下,改变或引入的基因可能与天然基因表达不同。这种改变的或外来的遗传信息将包括改变的与淋巴管异常相关的核苷酸序列的引入。
用于改变靶基因的dna可以通过多种技术获得,包括但不限于从基因组来源分离、从分离的mrna模板制备cdna、直接合成或其组合。
用于转基因导入的靶细胞的一种类型是胚胎干细胞(es)。es细胞可以从体外培养的植入前胚胎中获得(evans等人,(1981)nature292:154-156;bradley等人,(1984)nature309:255-258;gossler等人,(1986)proc.natl.acad.sci.83:9065-9069)。可以通过标准技术(例如dna转染)或通过逆转录病毒介导的转导,将转基因有效地引入es细胞。此后,可以将所得的转化的es细胞与来自非人类动物的胚泡合并。此后,引入的es细胞定植在胚胎中并有助于所得嵌合体动物的种系。
确定个体基因及其表达产物的贡献的一种方法是使用分离的、含有突变的与淋巴管异常相关的基因作为插入盒,以选择性失活全能es细胞中的野生型基因(例如如上所述的那些),然后产生转基因小鼠。描述了基因靶向的es细胞在产生基因靶向转基因小鼠中的用途,并在其他地方进行了综述(frohman等人,(1989)cell56:145-147;bradley等人,(1992)bio/technology10:534-539)。
通过使用靶向的同源重组将特定的变化插入染色体等位基因中,可以使用灭活或改变任何遗传区域至所需突变的技术。但是,与在接近100%频率发生的同源染色体外重组相比,最初报道同源质粒-染色体重组仅在10-6和10-3之间的频率被检测到。非同源质粒-染色体相互作用在比同源插入物高105-102倍的水平上更频繁发生。
为了克服鼠es细胞中靶向重组的低比例,已经开发出多种策略来检测或选择罕见的同源重组体。一种检测同源改变事件的方法是使用聚合酶链反应(pcr)筛选转化细胞池中的同源插入片段,然后筛选单个克隆。或者,已经开发出一种积极的遗传选择方法,其中构建了一种标记基因,所述标记基因仅在发生同源插入时才有活性,从而可以直接选择这些重组体。为选择同源重组体而开发的最强大的方法之一是为不存在直接选择改变的基因开发的正负选择(pns)方法。由于标记基因具有自己的启动子,所述pns方法更有效地用于靶向未高水平表达的基因。通过使用单纯疱疹病毒胸苷激酶(hsv-tk)基因选择非同源重组体,并使用有效的疱疹药物选择非同源重组体,所述药物例如丙氧鸟苷(gancyclovir)(ganc)或(1-(2-脱氧-2-氟-b-d阿拉伯氟烷糖基)-5-碘尿嘧啶(fiau)。通过这种反选择,可以增加存活的转化体中同源重组体的数量。利用突变的与淋巴管异常相关的核酸作为靶向插入盒,可提供检测成功插入的方法,例如,通过获得与对由ephb4核酸编码的多肽具有免疫学特异性的抗体的免疫反应性是可见的,因此有助于筛选/选择具有所需基因型的es细胞。
如本文所用,敲入动物是其中例如内源鼠基因已被本发明的人类淋巴管异常相关基因替代的动物。这种敲入动物为研究淋巴管异常的发展提供了理想的模型系统。
如本文所用,可以使用载体以“组织特异性方式”或“细胞类型特异性方式”靶向突变的淋巴管异常相关核酸,其片段或淋巴管异常相关融合蛋白的表达,其中编码全部或部分与淋巴管异常相关的核酸的核酸序列可操纵地连接至调节序列(例如,启动子和/或增强子),所述调节序列指导特定组织或细胞类型中所编码的蛋白的表达。这样的调节元件可以有利地用于体外和体内应用。指导组织特异性蛋白的启动子是本领域众所周知的,并在本文中进行了描述。替代地,所述转基因可以在诱导型启动子的控制下,所述启动子可以以组织特异性或“全身”的方式起作用。
可以将编码本发明的与淋巴管异常相关的突变体的核酸序列可操作地连接至多种不同的启动子序列,以在转基因动物中表达。这样的启动子包括但不限于朊病毒基因启动子,例如用于在所需细胞类型中表达转基因的仓鼠或小鼠thy-1启动子;pgk启动子;或cmv启动子。
本文还提供了本发明的转基因小鼠的使用方法。例如,已经向其中引入了包含与突变的淋巴管异常相关的核酸的核酸或其编码的蛋白质的转基因小鼠,可用于开发筛选方法以筛选治疗剂以确认能够调节淋巴管异常发生的那些。
检测产品和试剂盒
涵盖了可用于检测淋巴管异常snv的组合物或产品。例如,含淋巴管异常相关的snv的核酸、表达其的载体、含淋巴管异常相关的snv的标志物蛋白以及抗淋巴管异常特异性标志物的抗体是能够检测snv的产品,所述snv为ptpn11基因中的c.1504t>g:ps502a、c.1510a>g:pm504v和/或c.1507g>c:pg503r,kras中的c.35g:pg12d,braf基因中的c.1403t>c:pf468s,sos1基因中的c.2536g>a:pe846k,以及包含itga9基因中的c.1236+4a>g和c.289t>g:p.c97g的复合突变。还包括具有足够的长度和特征以检测snv的核酸探针,所述snv是ptpn11基因中的c.1504t>g:ps502a、c.1510a>g:pm504v和/或c.1507g>c:pg503r,kras中的c.35g>a:pg12d,braf基因中的c.1403t>c:pf468s,sos1基因中的c.2536g>a:pe846k,以及包含itga9基因中的c.1236+4a>g和c.289t>g:p.c97g的复合突变。可以标记检测产物,以便可以对其进行检测。
任何可用于检测与淋巴管异常相关的snv的产品都可以被整合到试剂盒中。任何可用于治疗淋巴管异常的产品都可以被整合到试剂盒中。包括了包含此类检测和治疗产品的试剂盒。所述试剂盒可以包含下列的一种或多种:淋巴管异常相关的snv特异性标志物多核苷酸或固定在固体支持物上的一种或多种此类标志物、基因芯片、寡核苷酸、多肽、肽、抗体、标记、标志物、报道子、药学上可接受的载体、生理上可接受的载体、使用说明、容器、施用器皿、试验底物或其任意组合。
提供以下实施例以说明本发明的某些实施方案。它们不意图以任何方式限制本发明。
实施例i
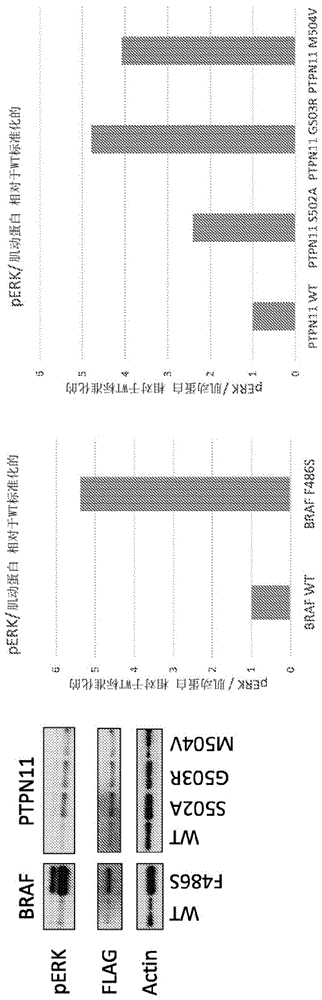
如对于其他淋巴畸形所述的,ccla是先天性淋巴发育错误的结果。为了确定ccla的遗传基础,我们对来自7名患者的dna样本进行了全外显子组测序,所有患者均具有通过动态对比磁共振淋巴管造影(dcmrl)成像的畸形淋巴管,这是一种新的先进成像技术,可以更好地诊断这组患者。我们检查了五名患者的错义、无义、剪接改变和编码插入缺失标记(indel)突变,这些突变可能解释了这些表型。过滤结果以排除同义变体、次要等位基因频率(maf)大于0.5%的变体以及通过我们内部外显子组变体数据库先前在对照中确认的变体。相关候选者已被推荐进行人工操作(manualcuration)。结果,我们在四个不同的基因(ptpn11、kras、braf和sos1)中鉴定出一个体细胞突变和五个新生的(denovo)错义突变,它们均涉及ras/mapk信号转导途径(表3)。另外,在一名患有原发性淋巴水肿和逆行淋巴流的患者中发现了两个杂合变体,即itga9中的c.1236+4a>g和c.289t>g:p.c97g。itga9编码整合素α-9,这是一种介导细胞-细胞粘附和细胞-基质相互作用的细胞表面糖蛋白。整合素α-9结合vegf-c,并且itga9的失活在小鼠中导致乳糜胸并破坏淋巴管瓣的形成。
这些蛋白质的每一种的序列可以在ncbi数据库中找到,并在下面列出。a)ptpn11中的c.1504t>g:ps502a;
ncbi参考序列中存在的s502:np_002825.3
b)ptpn11中的c.1510a>g:pm504v;
ncbi参考序列中存在的m504:np_002825.3
c)ptpn11中的c.1507g>c:pg503r;
ncbi参考序列中存在的g503:np_002825.3
d)kras中的c.35g>a:pg12d;
ncbi参考序列中存在的g12:np_203524.1。
e)braf中的c.1403t>c:pf468s;
ncbi参考序列中存在的f468:np_004324.2
f)sos1中的c.2536g>a:pe846k;
ncbi参考序列中存在的e846:np_005624.2
g)itga9中的c.1236+4a*>g和c.289t>g:p.c97g;
ncbi参考序列中存在的pc97:np_002198.2。
*变体距离外显子-内含子连接点4bp,并存在于内含子中
表3.导致淋巴病症的基因的突变
nm_002834.3:381-2162智人蛋白质酪氨酸磷酸酶、11型非受体(ptpn11)、转录物变体1、mrna
atgacatcgcggagatggtttcacccaaatatcactggtgtggaggcagaaaacctactgttgacaagaggagttgatggcagttttttggcaaggcctagtaaaagtaaccctggagacttcacactttccgttagaagaaatggagctgtcacccacatcaagattcagaacactggtgattactatgacctgtatggaggggagaaatttgccactttggctgagttggtccagtattacatggaacatcacgggcaattaaaagagaagaatggagatgtcattgagcttaaatatcctctgaactgtgcagatcctacctctgaaaggtggtttcatggacatctctctgggaaagaagcagagaaattattaactgaaaaaggaaaacatggtagttttcttgtacgagagagccagagccaccctggagattttgttctttctgtgcgcactggtgatgacaaaggggagagcaatgacggcaagtctaaagtgacccatgttatgattcgctgtcaggaactgaaatacgacgttggtggaggagaacggtttgattctttgacagatcttgtggaacattataagaagaatcctatggtggaaacattgggtacagtactacaactcaagcagccccttaacacgactcgtataaatgctgctgaaatagaaagcagagttcgagaactaagcaaattagctgagaccacagataaagtcaaacaaggcttttgggaagaatttgagacactacaacaacaggagtgcaaacttctctacagccgaaaagagggtcaaaggcaagaaaacaaaaacaaaaatagatataaaaacatcctgccctttgatcataccagggttgtcctacacgatggtgatcccaatgagcctgtttcagattacatcaatgcaaatatcatcatgcctgaatttgaaaccaagtgcaacaattcaaagcccaaaaagagttacattgccacacaaggctgcctgcaaaacacggtgaatgacttttggcggatggtgttccaagaaaactcccgagtgattgtcatgacaacgaaagaagtggagagaggaaagagtaaatgtgtcaaatactggcctgatgagtatgctctaaaagaatatggcgtcatgcgtgttaggaacgtcaaagaaagcgccgctcatgactatacgctaagagaacttaaactttcaaaggttggacaagggaatacggagagaacggtctggcaataccactttcggacctggccggaccacggcgtgcccagcgaccctgggggcgtgctggacttcctggaggaggtgcaccataagcaggagagcatcatggatgcagggccggtcgtggtgcactgcagtgctggaattggccggacagggacgttcattgtgattgatattcttattgacatcatcagagagaaaggtgttgactgcgatattgacgttcccaaaaccatccagatggtgcggtctcagaggtcagggatggtccagacagaagcacagtaccgatttatctatatggcggtccagcattatattgaaacactacagcgcaggattgaagaagagcagaaaagcaagaggaaagggcacgaatatacaaatattaagtattctctagcggaccagacgagtggagatcagagccctctcccgccttgtactccaacgccaccctgtgcagaaatgagagaagacagtgctagagtctatgaaaacgtgggcctgatgcaacagcagaaaagtttcagatga(seqidno:1)
>nm_004985.4:193-759智人kras原癌基因、gtp酶(kras)、转录物变体b、mrna
atgactgaatataaacttgtggtagttggagctggtggcgtaggcaagagtgccttgacgatacagctaattcagaatcattttgtggacgaatatgatccaacaatagaggattcctacaggaagcaagtagtaattgatggagaaacctgtctcttggatattctcgacacagcaggtcaagaggagtacagtgcaatgagggaccagtacatgaggactggggagggctttctttgtgtatttgccataaataatactaaatcatttgaagatattcaccattatagagaacaaattaaaagagttaaggactctgaagatgtacctatggtcctagtaggaaataaatgtgatttgccttctagaacagtagacacaaaacaggctcaggacttagcaagaagttatggaattccttttattgaaacatcagcaaagacaagacagggtgttgatgatgccttctatacattagttcgagaaattcgaaaacataaagaaaagatgagcaaagatggtaaaaagaagaaaaagaagtcaaagacaaagtgtgtaattatgtaa(seqidno:2)
>nm_004333.4:62-2362智人b-raf原癌基因、丝氨酸/苏氨酸激酶(braf)、mrna
atggcggcgctgagcggtggcggtggtggcggcgcggagccgggccaggctctgttcaacggggacatggagcccgaggccggcgccggcgccggcgccgcggcctcttcggctgcggaccctgccattccggaggaggtgtggaatatcaaacaaatgattaagttgacacaggaacatatagaggccctattggacaaatttggtggggagcataatccaccatcaatatatctggaggcctatgaagaatacaccagcaagctagatgcactccaacaaagagaacaacagttattggaatctctggggaacggaactgatttttctgtttctagctctgcatcaatggataccgttacatcttcttcctcttctagcctttcagtgctaccttcatctctttcagtttttcaaaatcccacagatgtggcacggagcaaccccaagtcaccacaaaaacctatcgttagagtcttcctgcccaacaaacagaggacagtggtacctgcaaggtgtggagttacagtccgagacagtctaaagaaagcactgatgatgagaggtctaatcccagagtgctgtgctgtttacagaattcaggatggagagaagaaaccaattggttgggacactgatatttcctggcttactggagaagaattgcatgtggaagtgttggagaatgttccacttacaacacacaactttgtacgaaaaacgtttttcaccttagcattttgtgacttttgtcgaaagctgcttttccagggtttccgctgtcaaacatgtggttataaatttcaccagcgttgtagtacagaagttccactgatgtgtgttaattatgaccaacttgatttgctgtttgtctccaagttctttgaacaccacccaataccacaggaagaggcgtccttagcagagactgccctaacatctggatcatccccttccgcacccgcctcggactctattgggccccaaattctcaccagtccgtctccttcaaaatccattccaattccacagcccttccgaccagcagatgaagatcatcgaaatcaatttgggcaacgagaccgatcctcatcagctcccaatgtgcatataaacacaatagaacctgtcaatattgatgacttgattagagaccaaggatttcgtggtgatggaggatcaaccacaggtttgtctgctaccccccctgcctcattacctggctcactaactaacgtgaaagccttacagaaatctccaggacctcagcgagaaaggaagtcatcttcatcctcagaagacaggaatcgaatgaaaacacttggtagacgggactcgagtgatgattgggagattcctgatgggcagattacagtgggacaaagaattggatctggatcatttggaacagtctacaagggaaagtggcatggtgatgtggcagtgaaaatgttgaatgtgacagcacctacacctcagcagttacaagccttcaaaaatgaagtaggagtactcaggaaaacacgacatgtgaatatcctactcttcatgggctattccacaaagccacaactggctattgttacccagtggtgtgagggctccagcttgtatcaccatctccatatcattgagaccaaatttgagatgatcaaacttatagatattgcacgacagactgcacagggcatggattacttacacgccaagtcaatcatccacagagacctcaagagtaataatatatttcttcatgaagacctcacagtaaaaataggtgattttggtctagctacagtgaaatctcgatggagtgggtcccatcagtttgaacagttgtctggatccattttgtggatggcaccagaagtcatcagaatgcaagataaaaatccatacagctttcagtcagatgtatatgcatttggaattgttctgtatgaattgatgactggacagttaccttattcaaacatcaacaacagggaccagataatttttatggtgggacgaggatacctgtctccagatctcagtaaggtacggagtaactgtccaaaagccatgaagagattaatggcagagtgcctcaaaaagaaaagagatgagagaccactctttccccaaattctcgcctctattgagctgctggcccgctcattgccaaaaattcaccgcagtgcatcagaaccctccttgaatcgggctggtttccaaacagaggattttagtctatatgcttgtgcttctccaaaaacacccatccaggcagggggatatggtgcgtttcctgtccactga(seqidno:3)
>nm_005633.3:42-4043智人sosras/rac鸟嘌呤核苷酸交换因子1(sos1)、mrna
atgcaggcgcagcagctgccctacgagtttttcagcgaagagaacgcgcccaagtggcggggactactggtgcctgcgctgaaaaaggtccaggggcaagttcatcctactctcgagtctaatgatgatgctcttcagtatgttgaagaattaattttgcaattattaaatatgctatgccaagctcagccccgaagtgcttcagatgtagaggaacgtgttcaaaaaagtttccctcatccaattgataaatgggcaatagctgatgcccaatcagctattgaaaagaggaagcgaagaaaccctttatctctcccagtagaaaaaattcatcctttattaaaggaggtcctaggttataaaattgaccaccaggtttctgtttacatagtagcagtcttagaatacatttctgcagacattttaaagctggttgggaattatgtaagaaatatacggcattatgaaattacaaaacaagatattaaagtggcaatgtgtgctgacaaggtattgatggatatgtttcatcaagatgtagaagatattaatatattatctttaactgacgaagagccttccacctcaggagaacaaacttactatgatttggtaaaagcatttatggcagaaattcgacaatatataagggaactaaatctaattataaaagtttttagagagccctttgtctccaattcaaaattgttttcagctaatgatgtagaaaatatatttagtcgcatagtagatatacatgaacttagtgtaaagttactgggccatatagaagatacagtagaaatgacagatgaaggcagtccccatccactagtaggaagctgctttgaagacttagcagaggaactggcatttgatccatatgaatcgtatgctcgagatattttgcgacctggttttcatgatcgtttccttagtcagttatcaaagcctggggcagcactttatttgcagtcaataggcgaaggtttcaaagaagctgttcaatatgttttacccaggctgcttctggcccctgtttaccactgtctccattactttgaacttttgaagcagttagaagaaaaaagtgaagatcaagaagacaaggaatgtttaaaacaagcaataacagctttgcttaatgttcagagtggtatggaaaaaatatgttctaaaagtcttgcaaaacgaagactgagtgaatctgcatgtcggttttatagtcagcaaatgaaggggaaacaactagcaatcaagaagatgaacgagattcagaagaatattgatggttgggagggaaaagacattggacagtgttgtaatgaatttataatggaaggaactcttacacgtgtaggagccaaacatgagagacacatatttctctttgatggcttaatgatttgctgtaaatcaaatcatgggcagccaagacttcctggtgctagcaatgcagaatatcgtcttaaagaaaagttttttatgcgaaaggtacaaattaatgataaagatgacaccaatgaatacaagcatgcttttgaaataattttaaaagatgaaaatagtgttatattttctgccaagtcagctgaagagaaaaacaattggatggcagcattgatatctttacagtaccggagtacactggaaaggatgcttgatgtaacaatgctacaggaagagaaagaggagcagatgaggctgcctagtgctgatgtttatagatttgcagagcctgactctgaagagaatattatatttgaagagaacatgcagcccaaggctggaattccaattatcaaagcaggaactgttattaaacttatagagaggcttacgtaccatatgtacgcagatcccaattttgttcggacatttcttacaacatacagatccttttgcaaacctcaagaactactgagtcttataatagaaaggtttgaaattccagagcctgagccaacagaagctgatcgcatagctatagagaatggagatcaacccttgagtgcagaactgaaaagatttagaaaagaatatatacagcctgtgcaactgcgagtattaaatgtatgtcggcactgggtagagcaccacttctatgattttgaaagagatgcatatcttttgcaacgaatggaagaatttattggaacagtaagaggtaaagcaatgaaaaaatgggttgaatccatcactaaaataatccaaaggaaaaaaattgcaagagacaatggaccaggtcataatattacatttcagagttcacctcccacagttgagtggcatataagcagacctgggcacatagagacttttgacctgctcaccttacacccaatagaaattgctcgacaactcactttacttgaatcagatctataccgagctgtacagccatcagaattagttggaagtgtgtggacaaaagaagacaaagaaattaactctcctaatcttctgaaaatgattcgacataccaccaacctcactctgtggtttgagaaatgtattgtagaaactgaaaatttagaagaaagagtagctgtggtgagtcgaattattgagattctacaagtctttcaagagttgaacaactttaatggtgtccttgaggttgtcagtgctatgaattcatcacctgtttacagactagaccacacatttgagcaaataccaagtcgccagaagaaaattttagaagaagctcatgaattgagtgaagatcactataagaaatatttggcaaaactcaggtctattaatccaccatgtgtgcctttctttggaatttatctcactaatatcttgaaaacagaagaaggcaaccctgaggtcctaaaaagacatggaaaagagcttataaactttagcaaaaggaggaaagtagcagaaataacaggagagatccagcagtaccaaaatcagccttactgtttacgagtagaatcagatatcaaaaggttctttgaaaacttgaatccgatgggaaatagcatggagaaggaatttacagattatcttttcaacaaatccctagaaatagaaccacgaaaccctaagcctctcccaagatttccaaaaaaatatagctatcccctaaaatctcctggtgttcgtccatcaaacccaagaccaggtaccatgaggcatcccacacctctgcagcaggagccaaggaaaattagttatagtaggatccctgaaagtgaaacagaaagtacagcatctgcaccaaattctccaagaacaccgttaacacctccgcctgcttctggtgcttccagtaccacagatgtttgcagtgtatttgattccgatcattcgagcccttttcactcaagcaatgataccgtctttatccaagttactctgccccatggcccaagatctgcttctgtatcatctataagtttaaccaaaggcactgatgaagtgcctgtccctcctcctgttcctccacgaagacgaccagaatctgccccagcagaatcttcaccatctaagattatgtctaagcatttggacagtcccccagccattcctcctaggcaacccacatcaaaagcctattcaccacgatattcaatatcagaccggacctctatctcagaccctcctgaaagccctcccttattaccaccacgagaacctgtgaggacacctgatgttttctcaagctcaccactacatctccaacctccccctttgggcaaaaaaagtgaccatggcaatgccttcttcccaaacagcccttccccctttacaccacctcctcctcaaacaccttctcctcacggcacaagaaggcatctgccatcaccaccattgacacaagaagtggaccttcattccattgctgggccgcctgttcctccacgacaaagcacttctcaacatatccctaaactccctccaaaaacttacaaaagggagcacacacacccatccatgcacagagatggaccaccactgttggagaatgcccattcttcctga(seqidno:4)
>nm_002207.2:54-3161智人整合素亚基α9(itga9)、mrna
atgggcggcccggctgcgccgaggggcgccgggaggctccgcgcgctgctgctggcgctggtggtcgcggggatccccgcgggcgcctacaacctcgacccgcagcgccccgtgcacttccagggccccgctgactcgttcttcggctacgcagttctggagcatttccacgacaacacgcgctgggtccttgtgggcgcaccaaaggcagattccaaatacagcccttcagtgaagtctcctggggctgtgtttaagtgccgtgttcacaccaaccctgaccggagatgcaccgaactggacatggctcgagggaagaatcggggcacgtcctgcggaaagacctgccgggaagaccgcgatgatgagtggatgggggtgagcctggcccgacagcccaaggctgatggccgtgtgttggcctgtgctcatcgctggaagaacatctactatgaagccgaccacatcctaccccatggcttctgctacatcatcccctccaacctccaggccaaaggcaggacgctgatcccttgctatgaagagtataagaagaagtacggagaggaacacggctcctgccaggctgggatagcgggcttcttcaccgaggagctggtggtgatgggtgctccagggtcattttattgggctggaaccatcaaagtgctgaaccttacggacaacacctatttaaaactgaacgacgaagtgatcatgaacaggcggtacacctacctgggctacgcagtgaccgctggccacttctctcacccgtccaccattgatgtggtaggaggtgccccacaggacaaaggcatcggcaaggtttatattttcagagctgaccgaagatcaggcaccttaattaagatctttcaagcatcaggtaaaaagatgggctcttacttcggctcctccttgtgcgcagttgacctgaatggggacggcctctctgacctgctggtgggggcccccatgttttctgagatcagggatgagggacaggtcactgtctacatcaacagaggaaatggagccctcgaggagcagctggctctgactggggatggtgcctacaatgcgcactttggagagagcattgccagcctggacgatctggacaatgatgggttcccagatgtggccattggtgcacccaaggaggatgacttcgcaggggcggtctatatctatcatggtgatgccggtgggatagtccctcagtactcaatgaaactgtctgggcagaagataaatccagtgctccggatgtttggtcagtccatatcgggaggcattgatatggatggaaatggctatcctgatgtcactgttggagccttcatgtccgacagcgtggttcttctcagagcaaggcctgtcattacggtggatgtctccatcttcctcccgggctccatcaacatcacagcgcctcagtgtcacgacggacagcagcctgtgaactgcctgaacgtcaccacctgcttcagcttccatggcaaacacgttccaggagagattggcctgaattatgttctgatggctgacgtggccaaaaaggagaagggccagatgcccagggtctactttgtgctgctgggagagaccatgggtcaggtcacagagaagctgcagctgacttacatggaggagacgtgtcgtcactatgtggcccatgtgaagcggagggtgcaggacgtcatcagcccgatcgtgtttgaagcagcctacagcctcagtgagcatgtgactggagaggaggagagggaactgccgcctctgacaccagttctccgctggaaaaagggacaaaagattgcccaaaagaatcagactgtttttgaaaggaattgccgttcagaggactgtgccgcagacctgcagcttcagggtaaactgctgctctccagtatggatgagaaaaccctgtatctagctttgggggctgtgaagaacatctccctaaacatctctatctccaacctcggagatgatgcctatgatgccaacgtgtccttcaatgtttcccgggagctcttcttcatcaacatgtggcagaaggaggagatgggcatctcctgtgagctgctggaatcggacttcctcaaatgcagcgtgggatttcctttcatgaggtcaaagtcaaagtatgaattcagcgtgatctttgatacaagccacctgtctggggaagaggaagttctcagcttcattgttactgctcagagtggcaacacggagcgctctgaatccctgcatgacaacaccctcgtgctgatggtgccactgatgcacgaggtggacacgtccatcaccggaatcatgtctccaacctcctttgtatatggcgagtccgtggacgcagccaacttcattcagctggatgacctggagtgtcactttcagcccatcaatatcacccttcaggtctacaacactggcccaagcacccttccagggtcatctgtcagcatctctttccctaatcgactctcatctggtggtgcagagatgtttcatgtccaggaaatggtggtgggccaagagaagggaaactgctctttccagaaaaacccaactccctgcatcatccctcaagaacaagaaaatatcttccacacaatatttgcttttttcacaaagtctggaagaaaagtcttggactgtgaaaaaccaggaatttcttgcctaacagcacactgtaactttagtgctcttgctaaagaagaaagtcgtactatagacatttacatgctgctgaacacagaaatactgaaaaaggacagttcgtctgtcatccagttcatgtcccgcgccaaggtgaaggtggatcctgccctaagggtggtggaaatagctcatgggaacccagaagaggtgacggtggtcttcgaggccctgcacaatctggagccccgtggctacgtcgtggggtggatcatcgccatcagtttgttggtgggaatcctcatcttcctgctgctggccgtgctgctctggaagatgggcttctttcgccgaaggtacaaagaaattatcgaagctgagaagaaccggaaagagaatgaagacagttgggactgggtccagaaaaaccagtga(seqidno:5)
nm_002890.24402bpmrna智人rasp21蛋白激活剂1(rasa1)、转录物
1cgtaacccaggcagctggggagcctgggctgtggccctaggagggggcgcggcggcgggc
61tctctccttttgttgttgtttcctcagcctggggagctgaaggggagacgcgtctgggtg
121gggctgctcggagcccgggcctggtggcccctggggctcccgggcgggcagggtagggca
181gagtagagcgggcttcaacatgatggcggccgaggccggcagtgaggagggcggcccggt
241aacagccggagctggaggaggcggcgcggcagcgggctccagtgcctatcccgcagtgtg
301tcgggtgaagatacccgcggccctgcctgtggcagccgccccctatcctgggctggtgga
361gaccggagtggctggaactctgggtggcggagccgctttggggtcagagttcctaggagc
421cgggtctgtggcaggggcactggggggagctggactgacagggggaggtactgctgctgg
481cgtagctggtgctgctgctggcgtggccggtgctgctgttgctggacctagtggagacat
541ggctctcaccaaactgcccacttcgttgcttgctgagactctcgggccaggcggcggttt
601tccccctctgccccctcccccttacctgccccctttgggggcgggcctcgggacagtgga
661cgaaggtgactctctggatggaccagaatacgaggaggaagaggtggccataccgttgac
721cgctcctccaactaaccagtggtatcacggaaaacttgacagaacgatagcagaagaacg
781cctcaggcaggcagggaagtctggcagttatcttataagagagagtgatcggaggccagg
841gtcctttgtactttcatttcttagccagatgaatgttgtcaaccattttaggattattgc
901tatgtgtggagattactacattggtggaagacgtttttcttcactgtcagacctaatagg
961ttattacagtcatgtttcttgtttgcttaaaggagaaaaattactttacccagttgcacc
1021accagagccagtagaagatagaaggcgtgtacgagctattctaccttacacaaaagtacc
1081agacactgatgaaataagtttcttaaaaggagatatgttcattgttcataatgaattaga
1141agatggatggatgtgggttacaaatttaagaacagatgaacaaggccttattgttgaaga
1201cctagtagaagaggtgggccgggaagaagatccacatgaaggaaaaatatggttccatgg
1261gaagatttccaaacaggaagcttataatttactaatgacagttggtcaagtctgcagttt
1321tcttgtgaggccctcagataatactcctggcgattattcactttatttccggaccaatga
1381aaatattcagcgatttaaaatatgtccaacgccaaacaatcagtttatgatgggaggccg
1441gtattataacagcattggggacatcatagatcactatcgaaaagaacagattgttgaagg
1501atattatcttaaggaacctgtaccaatgcaggatcaagaacaagtactcaatgacacagt
1561ggatggcaaggaaatctataataccatccgtcgtaaaacaaaggatgccttttataaaaa
1621cattgttaagaaaggttatcttctgaaaaagggcaaaggaaaacgttggaaaaatttata
1681ttttatcttagagggtagtgatgcccaacttatttattttgaaagcgaaaaacgagctac
1741caaaccaaaaggattaatagatctcagtgtatgttctgtctatgtcgttcatgatagtct
1801ctttggcaggccaaactgttttcagatagtagttcagcactttagtgaagaacattacat
1861cttttactttgcaggagaaactccagaacaagcagaggattggatgaaaggtctgcaggc
1921attttgcaatttacggaaaagtagtccagggacatccaataaacgccttcgtcaggtcag
1981cagccttgttttacatattgaagaagcccataaactcccagtaaaacattttactaatcc
2041atattgtaacatctacctgaatagtgtccaagtagcaaaaactcatgcaagggaagggca
2101aaacccagtatggtcagaagagtttgtctttgatgatcttcctcctgacatcaatagatt
2161tgaaataactcttagtaataaaacaaagaaaagcaaagatcctgatatcttatttatgcg
2221ctgccagttgagccgattacagaaagggcatgccacagatgaatggtttctgctcagctc
2281ccatataccattaaaaggtattgaaccagggtccctgcgtgttcgagcacgatactctat
2341ggaaaaaatcatgccagaagaagagtacagtgaatttaaagagcttatactgcaaaagga
2401acttcatgtagtctatgctttatcacatgtatgtggacaagaccgaacactactggccag
2461catcctactgaggatttttcttcacgaaaagcttgaatcgttgttgttatgcacactaaa
2521tgacagagaaataagcatggaagatgaagccactaccctatttcgagccacaacacttgc
2581aagcaccttgatggagcagtatatgaaagccactgctacacagtttgttcatcatgcttt
2641gaaagactctattttaaagataatggaaagcaagcagtcttgtgagttaagtccatcaaa
2701gttagaaaaaaatgaagatgtgaacactaatttaacacacctattgaacatactttcaga
2761gcttgtggagaaaatattcatggcttcagaaatacttccaccgacattgagatatattta
2821tgggtgtttacagaaatctgttcagcataagtggcctacaaataccaccatgagaacaag
2881agttgttagtggttttgtttttcttcgactcatctgtcctgccatcctgaatccacggat
2941gttcaatatcatctcagattctccatctcctattgctgcaagaacactgatattagtggc
3001taaatctgtgcagaacttagcaaatcttgtggaatttggagctaaggagccctacatgga
3061aggtgtcaatccattcatcaaaagcaacaaacatcgtatgatcatgtttttagatgaact
3121tgggaatgtacctgaacttccggacactacagagcattctagaacggacctgtcccgtga
3181tttagcagcattgcatgagatttgcgtggctcattcagatgaacttcgaacgctcagtaa
3241tgagcgtggtgcacagcagcacgtattgaaaaagcttctggctataacagaactgcttca
3301acaaaaacaaaaccagtatacaaaaaccaatgatgtcaggtagcagccttcgccccagtg
3361ttctgcatggattcagcatgtccaacatggtaattcacttcagtttaatgtctcctttgc
3421tcttgccaaaaaatagcacacttttccacattccagtgatgtgtgagctatgcaaacaaa
3481atccaagattctgctggtgaataactatgccagcaaccttgtaagctatctgtgcaggat
3541atttgcactatttccacatggaatcaatctttaacaacctctgagccttggtgtacagac
3601cacctttcacaaaacgaaatgctatgactgtatcttgatatctcgaactttcaaaatata
3661ttttcagtacacccagttgccaaagttttgctgtctcttagagaaagaactatgaaatca
3721actgacaagaaacacattcttattgacaattgtgtataactggattgcagactgttctta
3781ctgtaactacttcctgattaggaatatgaccatttgactgttcaatgattatttgtattt
3841acagtttccagagtttgtcattataataggaacaatctttgctgtatacttttaaaaaat
3901actctgctatttctcttgctggaactgttgaaagaaaatatatagaatgatctattgctc
3961atcagctttattttttaaacatacgacttattttgttgaaattgtcaaagactgtattta
4021gatctcataatgctttgttaaatgtttacaagtaaatagtttgaattcagtaaatattat
4081tggttgttgtattgatcaatgcatgttacccattcaaccattttatagactaccaatttc
4141ttttatgttaactagaatgcttttgttaaaagttatttgttcattatttgtgctacccct
4201ttgattatgcagacaacctcatcagctgcctaacttatccatctttgaacttctgactac
4261ttgttgtatctgctggatatttagttcaactgtatagttttatttacttctgtatgtgta
4321tttttgtgaagtattcacaaaggttaagttaaaataaaaccaagggatatcttgcataat
4381tgattaaaaaaaaaaaaaaaaa(seqidno:6)
nm_002880.23291bpmrna智人raf-1原癌基因、丝氨酸/苏氨酸激酶(raf1)、转录物变体2、mrna。
1agaatcggagagccggtggcgtcgcaggtcgggaggacgagcaccgagtcgagggctcgc
61tcgtctgggccgcccgagagtcttaatcgcgggcgcttgggccgccatcttagatggcgg
121gagtaagaggaaaacgattgtgaggcgggaacggctttctgctgccttttttgggccccg
181aaaagggtcagctggccgggctttggggcgcgtgccctgaggcgcggagcgcgtttgcta
241cgatgcgggggctgctcggggctccgtcccctgggctggggacgcgccgaatgtgaccgc
301ctcccgctccctcacccgccgcggggaggaggagcgggcgagaagctgccgccgaacgac
361aggacgttggggcggcctggctccctcaggtttaagaattgtttaagctgcatcaatgga
421gcacatacagggagcttggaagacgatcagcaatggttttggattcaaagatgccgtgtt
481tgatggctccagctgcatctctcctacaatagttcagcagtttggctatcagcgccgggc
541atcagatgatggcaaactcacagatccttctaagacaagcaacactatccgtgttttctt
601gccgaacaagcaaagaacagtggtcaatgtgcgaaatggaatgagcttgcatgactgcct
661tatgaaagcactcaaggtgaggggcctgcaaccagagtgctgtgcagtgttcagacttct
721ccacgaacacaaaggtaaaaaagcacgcttagattggaatactgatgctgcgtctttgat
781tggagaagaacttcaagtagatttcctggatcatgttcccctcacaacacacaactttgc
841tcggaagacgttcctgaagcttgccttctgtgacatctgtcagaaattcctgctcaatgg
901atttcgatgtcagacttgtggctacaaatttcatgagcactgtagcaccaaagtacctac
961tatgtgtgtggactggagtaacatcagacaactcttattgtttccaaattccactattgg
1021tgatagtggagtcccagcactaccttctttgactatgcgtcgtatgcgagagtctgtttc
1081caggatgcctgttagttctcagcacagatattctacacctcacgccttcacctttaacac
1141ctccagtccctcatctgaaggttccctctcccagaggcagaggtcgacatccacacctaa
1201tgtccacatggtcagcaccaccctgcctgtggacagcaggatgattgaggatgcaattcg
1261aagtcacagcgaatcagcctcaccttcagccctgtccagtagccccaacaatctgagccc
1321aacaggctggtcacagccgaaaacccccgtgccagcacaaagagagcgggcaccagtatc
1381tgggacccaggagaaaaacaaaattaggcctcgtggacagagagattcaagctattattg
1441ggaaatagaagccagtgaagtgatgctgtccactcggattgggtcaggctcttttggaac
1501tgtttataagggtaaatggcacggagatgttgcagtaaagatcctaaaggttgtcgaccc
1561aaccccagagcaattccaggccttcaggaatgaggtggctgttctgcgcaaaacacggca
1621tgtgaacattctgcttttcatggggtacatgacaaaggacaacctggcaattgtgaccca
1681gtggtgcgagggcagcagcctctacaaacacctgcatgtccaggagaccaagtttcagat
1741gttccagctaattgacattgcccggcagacggctcagggaatggactatttgcatgcaaa
1801gaacatcatccatagagacatgaaatccaacaatatatttctccatgaaggcttaacagt
1861gaaaattggagattttggtttggcaacagtaaagtcacgctggagtggttctcagcaggt
1921tgaacaacctactggctctgtcctctggatggccccagaggtgatccgaatgcaggataa
1981caacccattcagtttccagtcggatgtctactcctatggcatcgtattgtatgaactgat
2041gacgggggagcttccttattctcacatcaacaaccgagatcagatcatcttcatggtggg
2101ccgaggatatgcctccccagatcttagtaagctatataagaactgccccaaagcaatgaa
2161gaggctggtagctgactgtgtgaagaaagtaaaggaagagaggcctctttttccccagat
2221cctgtcttccattgagctgctccaacactctctaccgaagatcaaccggagcgcttccga
2281gccatccttgcatcgggcagcccacactgaggatatcaatgcttgcacgctgaccacgtc
2341cccgaggctgcctgtcttctagttgactttgcacctgtcttcaggctgccaggggaggag
2401gagaagccagcaggcaccacttttctgctccctttctccagaggcagaacacatgttttc
2461agagaagctgctgctaaggaccttctagactgctcacagggccttaacttcatgttgcct
2521tcttttctatccctttgggccctgggagaaggaagccatttgcagtgctggtgtgtcctg
2581ctccctccccacattccccatgctcaaggcccagccttctgtagatgcgcaagtggatgt
2641tgatggtagtacaaaaagcaggggcccagccccagctgttggctacatgagtatttagag
2701gaagtaaggtagcaggcagtccagccctgatgtggagacacatgggattttggaaatcag
2761cttctggaggaatgcatgtcacaggcgggactttcttcagagagtggtgcagcgccagac
2821attttgcacataaggcaccaaacagcccaggactgccgagactctggccgcccgaaggag
2881cctgctttggtactatggaacttttcttaggggacacgtcctcctttcacagcttctaag
2941gtgtccagtgcattgggatggttttccaggcaaggcactcggccaatccgcatctcagcc
3001ctctcagggagcagtcttccatcatgctgaattttgtcttccaggagctgcccctatggg
3061gcggggccgcagggccagccttgtttctctaacaaacaaacaaacaaacagccttgtttc
3121tctagtcacatcatgtgtatacaaggaagccaggaatacaggttttcttgatgatttggg
3181ttttaattttgtttttattgcacctgacaaaatacagttatctgatggtccctcaattat
3241gttattttaataaaataaattaaatttaggtgtaaaaaaaaaaaaaaaaaa
seqidno:7)
nm_0011428647833bpmrna智人压电型机械敏感离子通道组件1(piezo1)、mrna。
1atggagccgcacgtgctcggcgcggtcctgtactggctgctgctgccctgcgcgctgctg
61gctgcctgcctgctccgcttcagcggactctcgctggtctacctgctcttcctgctgctg
121ctgccctggttccccggccccacccgatgcggcctccaaggtcacacaggccgcctcctg
181cgggcattgctgggcctcagcctgctcttcctggtggcccatctcgccctccagatctgc
241ctgcatattgtgccccgcctggaccagctcctgggacccagctgcagccgctgggagacc
301ctctcgcgacacataggggtcacaaggctggacctgaaggacatccccaacgccatccgg
361ctggtggcccctgacctgggcatcttggtggtctcctctgtctgcctcggcatctgcggg
421cgccttgcaaggaacacccggcagagcccacatccacgggagctggatgatgatgagagg
481gatgtggatgccagcccgacggcagggctgcaggaagcagcaacgctggcccctacacgg
541aggtcacggctggccgctcgtttccgagtcacggcccactggctgctggtggcggctggg
601cgggtcctggccgtaacactgcttgcactggcaggcatcgcccacccctcggccctctcc
661agtgtctacctgctgctcttcctggccctctgcacctggtgggcctgccactttcccatc
721agcactcggggcttcagcagactctgcgtcgcggtggggtgcttcggcgccggccatctc
781atctgcctctactgctaccagatgcccttggcacaggctctgctcccgcctgccggcatc
841tgggctagggtgctgggtctcaaggacttcgtgggtcccaccaactgctccagcccccac
901gcgctggtcctcaacaccggcctggactggcctgtgtatgccagccccggcgtcctcctg
961ctgctgtgctacgccacggcctctctgcgcaagctccgcgcgtaccgcccctccggccag
1021aggaaggaggcggcaaaggggtatgaggctcgggagctggagctagcagagctggaccag
1081tggccccaggaacgggagtctgaccagcacgtggtgcccacagcacccgacaccgaggct
1141gataactgcatcgtgcacgagctgaccggccagagctccgtcctgcggcggcctgtgcgg
1201cccaagcgggctgagcccagggaggcgtctccgctccacagcctgggccacctcatcatg
1261gaccagagctatgtgtgcgcgctcattgccatgatggtatggagcatcacctaccacagc
1321tggctgaccttcgtactgctgctctgggcctgcctcatctggacggtgcgcagccgccac
1381caactggccatgctgtgctcgccctgcatcctgctgtatgggatgacgctgtgctgccta
1441cgctacgtgtgggccatggacctgcgccctgagctgcccaccaccctgggccccgtcagc
1501ctgcgccagctggggctggagcacacccgctacccctgtctggaccttggtgccatgttg
1561ctctacaccctgaccttctggctcctgctgcgccagtttgtgaaagagaagctgctgaag
1621tgggcagagtctccagctgcgctgacggaggtcaccgtggcagacacagagcccacgcgg
1681acgcagacgctgttgcagagcctgggggagctggtgaagggcgtgtacgccaagtactgg
1741atctatgtgtgtgctggcatgttcatcgtggtcagcttcgccggccgcctcgtggtctac
1801aagattgtctacatgttcctcttcctgctctgcctcaccctcttccaggtctactacagc
1861ctgtggcggaagctgctcaaggccttctggtggctcgtggtggcctacaccatgctggtc
1921ctcatcgccgtctacaccttccagttccaggacttccctgcctactggcgcaacctcact
1981ggcttcaccgacgagcagctgggggacctgggcctggagcagttcagcgtgtccgagctc
2041ttctccagcatcctggtgcccggcttcttcctcctggcctgcatcctgcagctgcactac
2101ttccacaggcccttcatgcagctcaccgacatggagcacgtgtccctgcctggcacgcgc
2161ctcccgcgctgggctcacaggcaggatgcagtgagtgggaccccactgctgcgggaggag
2221cagcaggagcatcagcagcagcagcaggaggaggaggaggaggaggaggactccagggac
2281gaggggctgggcgtggccactccccaccaggccacgcaggtgcctgaaggggcagccaag
2341tggggcctggtggctgagcggctgctggagctggcagccggcttctcggacgtcctctca
2401cgcgtgcaggtgttcctgcggcggctgctggagcttcacgttttcaagctggtggccctg
2461tacaccgtctgggtggccctgaaggaggtgtcggtgatgaacctgctgctggtggtgctg
2521tgggccttcgccctgccctacccacgcttccggcccatggcctcctgcctgtccaccgtg
2581tggacctgcgtcatcatcgtgtgtaagatgctgtaccagctcaaggttgtcaacccccag
2641gagtattccagcaactgcaccgagcccttccccaacagcaccaacttgctgcccacggag
2701atcagccagtccctgctgtaccgggggcccgtggaccctgccaactggtttggggtgcgg
2761aaagggttccccaacctgggctacatccagaaccacctgcaagtgctgctgctgctggta
2821ttcgaggccatcgtgtaccggcgccaggagcactaccgccggcagcaccagctggccccg
2881ctgcctgcccaggccgtgtttgccagcggcacccgccagcagctggaccaggatctgctc
2941ggctgcctcaagtacttcatcaacttcttcttctacaaattcgggctggagatctgcttc
3001ctgatggccgtgaacgtgatcgggcagcgcatgaactttctggtgaccctgcacggttgc
3061tggctggtggccatcctcacccgcaggcaccgccaggccattgcccgcctctggcccaac
3121tactgcctcttcctggcgctgttcctgctgtaccagtacctgctgtgcctggggatgccc
3181ccggccctgtgcattgattatccctggcgctggagccgggccgtccccatgaactccgca
3241ctcatcaagtggctgtacctgcctgatttcttccgggcccccaactccaccaacctcatc
3301agcgactttctcctgctgctgtgcgcctcccagcagtggcaggtgttctcagctgagcgc
3361acagaggagtggcagcgcatggctggcgtcaacaccgaccgcctggagccgctgcggggg
3421gagcccaaccccgtgcccaactttatccactgcaggtcctaccttgacatgctgaaggtg
3481gccgtcttccgatacctgttctggctggtgctggtggtggtgtttgtcacgggggccacc
3541cgcatcagcatcttcgggctgggctacctgctggcctgcttctacctgctgctcttcggc
3601acggccctgctgcagagggacacacgggcccgcctcgtgctgtgggactgcctcattctg
3661tacaacgtcaccgtcatcatctccaagaacatgctgtcgctcctggcctgcgtcttcgtg
3721gagcagatgcagaccggcttctgctgggtcatccagctcttcagccttgtatgcaccgtc
3781aagggctactatgaccccaaggagatgatggacagagaccaggactgcctgctgcctgtg
3841gaggaggctggcatcatctgggacagcgtctgcttcttcttcctgctgctgcagcgccgc
3901gtcttccttagccattactacctgcacgtcagggccgacctccaggccaccgccctgcta
3961gcctccaggggcttcgccctctacaacgctgccaacctcaagagcattgactttcaccgc
4021aggatagaggagaagtccctggcccagctgaaaagacagatggagcgtatccgtgccaag
4081caggagaagcacaggcagggccgggtggaccgcagtcgcccccaggacaccctgggcccc
4141aaggaccccggcctggagccagggcccgacagtccagggggctcctccccgccacggagg
4201cagtggtggcggccctggctggaccacgccacagtcatccactccggggactacttcctg
4261tttgagtccgacagtgaggaagaggaggaggctgttcctgaagacccgaggccgtcggca
4321cagagtgccttccagctggcgtaccaggcatgggtgaccaacgcccaggcggtgctgagg
4381cggcggcagcaggagcaggagcaggcaaggcaggaacaggcaggacagctacccacagga
4441ggtggtcccagccaggaggtggagccagcagagggccccgaggaggcagcggcaggccgg
4501agccatgtggtgcagagggtgctgagcacggcgcagttcctgtggatgctggggcaggcg
4561ctagtggatgagctgacacgctggctgcaggagttcacccggcaccacggcaccatgagc
4621gacgtgctgcgggcagagcgctacctcctcacacaggagctcctgcagggcggcgaagtg
4681cacaggggcgtgctggatcagctgtacacaagccaggccgaggccacgctgccaggcccc
4741accgaggcccccaatgccccaagcaccgtgtccagtgggctgggcgcggaggagccactc
4801agcagcatgacagacgacatgggcagccccctgagcaccggctaccacacgcgcagtggc
4861agtgaggaggcagtcaccgaccccggggagcgtgaggctggtgcctctctgtaccaggga
4921ctgatgcggacggccagcgagctgctcctggacaggcgcctgcgcatcccagagctggag
4981gaggcagagctgtttgcggaggggcagggccgggcgctgcggctgctgcgggccgtgtac
5041cagtgtgtggccgcccactcggagctgctctgctacttcatcatcatcctcaaccacatg
5101gtcacggcctccgccggctcgctggtgctgcccgtgctcgtcttcctgtgggccatgctg
5161tcgatcccgaggcccagcaagcgcttctggatgacggccatcgtcttcaccgagatcgcg
5221gtggtcgtcaagtacctgttccagtttgggttcttcccctggaacagccacgtggtgctg
5281cggcgctacgagaacaagccctacttcccgccccgcatcctgggcctggagaagactgac
5341ggctacatcaagtacgacctggtgcagctcatggcccttttcttccaccgctcccagctg
5401ctgtgctatggcctctgggaccatgaggaggactcaccatccaaggagcatgacaagagc
5461ggcgaggaggagcagggagccgaggaggggccaggggtgcctgcggccaccaccgaagac
5521cacattcaggtggaagccagggtcggacccacggacgggaccccagaaccccaagtggag
5581ctcaggccccgtgatacgaggcgcatcagtctacgttttagaagaaggaagaaggagggc
5641ccagcacggaaaggagcggcagccatcgaagctgaggacagggaggaagaagagggggag
5701gaagagaaagaggcccccacggggagagagaagaggccaagccgctctggaggaagagta
5761agggcggccgggcggcggctgcagggcttctgcctgtccctggcccagggcacatatcgg
5821ccgctacggcgcttcttccacgacatcctgcacaccaagtaccgcgcagccaccgacgtc
5881tatgccctcatgttcctggctgatgttgtcgacttcatcatcatcatttttggcttctgg
5941gcctttgggaagcactcggcggccacagacatcacgtcctccctatcagacgaccaggta
6001cccgaggctttcctggtcatgctgctgatccagttcagtaccatggtggttgaccgcgcc
6061ctctacctgcgcaagaccgtgctgggcaagctggccttccaggtggcgctggtgctggcc
6121atccacctatggatgttcttcatcctgcccgccgtcactgagaggatgttcaaccagaat
6181gtggtggcccagctctggtacttcgtgaagtgcatctacttcgccctgtccgcctaccag
6241atccgctgcggctaccccacccgcatcctcggcaacttcctcaccaagaagtacaatcat
6301ctcaacctcttcctcttccaggggttccggctggtgccgttcctggtggagctgcgggca
6361gtgatggactgggtgtggacggacaccacgctgtccctgtccagctggatgtgtgtggag
6421gacatctatgccaacatcttcatcatcaaatgcagccgagagacagagaagaaatacccg
6481cagcccaaagggcagaagaagaagaagatcgtcaagtacggcatgggtggcctcatcatc
6541ctcttcctcatcgccatcatctggttcccactgctcttcatgtcgctggtgcgctccgtg
6601gttggggttgtcaaccagcccatcgatgtcaccgtcaccctgaagctgggcggctatgag
6661ccgctgttcaccatgagcgcccagcagccgtccatcatccccttcacggcccaggcctat
6721gaggagctgtcccggcagtttgacccccagccgctggccatgcagttcatcagccagtac
6781agccctgaggacatcgtcacggcgcagattgagggcagctccggggcgctgtggcgcatc
6841agtccccccagccgtgcccagatgaagcgggagctctacaacggcacggccgacatcacc
6901ctgcgcttcacctggaacttccagagggacctggcgaagggaggcactgtggagtatgcc
6961aacgagaagcacatgctggccctggcccccaacagcactgcacggcggcagctggccagc
7021ctgctcgagggcacctcggaccagtctgtggtcatccctaatctcttccccaagtacatc
7081cgtgcccccaacgggcccgaagccaaccctgtgaagcagctgcagcccaatgaggaggcc
7141gactacctcggcgtgcgtatccagctgcggagggagcagggtgcgggggccaccggcttc
7201ctcgaatggtgggtcatcgagctgcaggagtgccggaccgactgcaacctgctgcccatg
7261gtcattttcagtgacaaggtcagcccaccgagcctcggcttcctggctggctacggcatc
7321atggggctgtacgtgtccatcgtgctggtcatcggcaagttcgtgcgcggattcttcagc
7381gagatctcgcactccattatgttcgaggagctgccgtgcgtggaccgcatcctcaagctc
7441tgccaggacatcttcctggtgcgggagactcgggagctggagctggaggaggagttgtac
7501gccaagctcatcttcctctaccgctcaccggagaccatgatcaagtggactcgtgagaag
7561gagtaggagctgctgctggcgcccgagagggaaggagccggcctgctgggcagcgtggcc
7621acaaggggcggcactcctcaggccgggggagccactgccccgtccaaggccgccagctgt
7681gatgcatcctcccggcctgcctgagccctgatgctgctgtcagagaaggacactgcgtcc
7741ccacggcctgcgtggcgctgccgtcccccacgtgtactgtagagttttttttttaattaa
7801aaaatgttttatttatacaaatggacaatcaga(seqidno:8)
nm_0044444369bpmrna智人eph受体b4(ephb4)、mrna。
1ttccagcgcagctcagcccctgcccggcccggcccgcccggctccgcgccgcagtctccc
61tccctcccgctccgtccccgctcgggctcccaccatccccgcccgcgaggagagcactcg
121gcccggcggcgcgagcagagccactccagggagggggggagaccgcgagcggccggctca
181gcccccgccacccggggcgggaccccgaggccccggagggaccccaactccagccacgtc
241ttgctgcgcgcccgcccggcgcggccactgccagcacgctccgggcccgccgcccgcgcg
301cgcggcacagacgcggggccacacttggcgccgccgcccggtgccccgcacgctcgcatg
361ggcccgcgctgagggccccgacgaggagtcccgcgcggagtatcggcgtccacccgccca
421gggagagtcagacctgggggggcgagggccccccaaactcagttcggatcctacccgagt
481gaggcggcgccatggagctccgggtgctgctctgctgggcttcgttggccgcagctttgg
541aagagaccctgctgaacacaaaattggaaactgctgatctgaagtgggtgacattccctc
601aggtggacgggcagtgggaggaactgagcggcctggatgaggaacagcacagcgtgcgca
661cctacgaagtgtgtgacgtgcagcgtgccccgggccaggcccactggcttcgcacaggtt
721gggtcccacggcggggcgccgtccacgtgtacgccacgctgcgcttcaccatgctcgagt
781gcctgtccctgcctcgggctgggcgctcctgcaaggagaccttcaccgtcttctactatg
841agagcgatgcggacacggccacggccctcacgccagcctggatggagaacccctacatca
901aggtggacacggtggccgcggagcatctcacccggaagcgccctggggccgaggccaccg
961ggaaggtgaatgtcaagacgctgcgtctgggaccgctcagcaaggctggcttctacctgg
1021ccttccaggaccagggtgcctgcatggccctgctatccctgcacctcttctacaaaaagt
1081gcgcccagctgactgtgaacctgactcgattcccggagactgtgcctcgggagctggttg
1141tgcccgtggccggtagctgcgtggtggatgccgtccccgcccctggccccagccccagcc
1201tctactgccgtgaggatggccagtgggccgaacagccggtcacgggctgcagctgtgctc
1261cggggttcgaggcagctgaggggaacaccaagtgccgagcctgtgcccagggcaccttca
1321agcccctgtcaggagaagggtcctgccagccatgcccagccaatagccactctaacacca
1381ttggatcagccgtctgccagtgccgcgtcgggtacttccgggcacgcacagacccccggg
1441gtgcaccctgcaccacccctccttcggctccgcggagcgtggtttcccgcctgaacggct
1501cctccctgcacctggaatggagtgcccccctggagtctggtggccgagaggacctcacct
1561acgccctccgctgccgggagtgccgacccggaggctcctgtgcgccctgcgggggagacc
1621tgacttttgaccccggcccccgggacctggtggagccctgggtggtggttcgagggctac
1681gtcctgacttcacctatacctttgaggtcactgcattgaacggggtatcctccttagcca
1741cggggcccgtcccatttgagcctgtcaatgtcaccactgaccgagaggtacctcctgcag
1801tgtctgacatccgggtgacgcggtcctcacccagcagcttgagcctggcctgggctgttc
1861cccgggcacccagtggggctgtgctggactacgaggtcaaataccatgagaagggcgccg
1921agggtcccagcagcgtgcggttcctgaagacgtcagaaaaccgggcagagctgcgggggc
1981tgaagcggggagccagctacctggtgcaggtacgggcgcgctctgaggccggctacgggc
2041ccttcggccaggaacatcacagccagacccaactggatgagagcgagggctggcgggagc
2101agctggccctgattgcgggcacggcagtcgtgggtgtggtcctggtcctggtggtcattg
2161tggtcgcagttctctgcctcaggaagcagagcaatgggagagaagcagaatattcggaca
2221aacacggacagtatctcatcggacatggtactaaggtctacatcgaccccttcacttatg
2281aagaccctaatgaggctgtgagggaatttgcaaaagagatcgatgtctcctacgtcaaga
2341ttgaagaggtgattggtgcaggtgagtttggcgaggtgtgccgggggcggctcaaggccc
2401cagggaagaaggagagctgtgtggcaatcaagaccctgaagggtggctacacggagcggc
2461agcggcgtgagtttctgagcgaggcctccatcatgggccagttcgagcaccccaatatca
2521tccgcctggagggcgtggtcaccaacagcatgcccgtcatgattctcacagagttcatgg
2581agaacggcgccctggactccttcctgcggctaaacgacggacagttcacagtcatccagc
2641tcgtgggcatgctgcggggcatcgcctcgggcatgcggtaccttgccgagatgagctacg
2701tccaccgagacctggctgctcgcaacatcctagtcaacagcaacctcgtctgcaaagtgt
2761ctgactttggcctttcccgattcctggaggagaactcttccgatcccacctacacgagct
2821ccctgggaggaaagattcccatccgatggactgccccggaggccattgccttccggaagt
2881tcacttccgccagtgatgcctggagttacgggattgtgatgtgggaggtgatgtcatttg
2941gggagaggccgtactgggacatgagcaatcaggacgtgatcaatgccattgaacaggact
3001accggctgcccccgcccccagactgtcccacctccctccaccagctcatgctggactgtt
3061ggcagaaagaccggaatgcccggccccgcttcccccaggtggtcagcgccctggacaaga
3121tgatccggaaccccgccagcctcaaaatcgtggcccgggagaatggcggggcctcacacc
3181ctctcctggaccagcggcagcctcactactcagcttttggctctgtgggcgagtggcttc
3241gggccatcaaaatgggaagatacgaagaaagtttcgcagccgctggctttggctccttcg
3301agctggtcagccagatctctgctgaggacctgctccgaatcggagtcactctggcgggac
3361accagaagaaaatcttggccagtgtccagcacatgaagtcccaggccaagccgggaaccc
3421cgggtgggacaggaggaccggccccgcagtactgacctgcaggaactccccaccccaggg
3481acaccgcctccccattttccggggcagagtggggactcacagaggcccccagccctgtgc
3541cccgctggattgcactttgagcccgtggggtgaggagttggcaatttggagagacaggat
3601ttgggggttctgccataataggaggggaaaatcaccccccagccacctcggggaactcca
3661gaccaagggtgagggcgcctttccctcaggactgggtgtgaccagaggaaaaggaagtgc
3721ccaacatctcccagcctccccaggtgcccccctcaccttgatgggtgcgttcccgcagac
3781caaagagagtgtgactcccttgccagctccagagtgggggggctgtcccagggggcaaga
3841aggggtgtcagggcccagtgacaaaatcattggggtttgtagtcccaacttgctgctgtc
3901accaccaaactcaatcatttttttcccttgtaaatgcccctcccccagctgctgccttca
3961tattgaaggtttttgagttttgtttttggtcttaatttttctccccgttccctttttgtt
4021tcttcgttttgtttttctaccgtccttgtcataactttgtgttggagggaacctgtttca
4081ctatggcctcctttgcccaagttgaaacaggggcccatcatcatgtctgtttccagaaca
4141gtgccttggtcatcccacatccccggaccccgcctgggacccccaagctgtgtcctatga
4201aggggtgtggggtgaggtagtgaaaagggcggtagttggtggtggaacccagaaacggac
4261gccggtgcttggaggggttcttaaattatatttaaaaaagtaactttttgtataaataaa
4321agaaaatgggacgtgtcccagctccaggggtaaaaaaaaaaaaaaaaaa
(seqidno:9)
nm_00026712381bpmrna智人神经纤维蛋白1(nf1)、转录物变体2、mrna。
1aatctctagctcgctcgcgctccctctccccgggccgtggaaaggatcccacttccggtg
61gggtgtcatggcggcgtctcggactgtgatggctgtggggagacggcgctagtggggaga
121gcgaccaagaggccccctcccctccccgggtccccttcccctatccccctccccccagcc
181tccttgccaacgccccctttccctctccccctcccgctcggcgctgaccccccatcccca
241cccccgtgggaacactgggagcctgcactccacagaccctctccttgcctcttccctcac
301ctcagcctccgctccccgccctcttcccggcccagggcgccggcccacccttccctccgc
361cgccccccggccgcggggaggacatggccgcgcacaggccggtggaatgggtccaggccg
421tggtcagccgcttcgacgagcagcttccaataaaaacaggacagcagaacacacatacca
481aagtcagtactgagcacaacaaggaatgtctaatcaatatttccaaatacaagttttctt
541tggttataagcggcctcactactattttaaagaatgttaacaatatgagaatatttggag
601aagctgctgaaaaaaatttatatctctctcagttgattatattggatacactggaaaaat
661gtcttgctgggcaaccaaaggacacaatgagattagatgaaacgatgctggtcaaacagt
721tgctgccagaaatctgccattttcttcacacctgtcgtgaaggaaaccagcatgcagctg
781aacttcggaattctgcctctggggttttattttctctcagctgcaacaacttcaatgcag
841tctttagtcgcatttctaccaggttacaggaattaactgtttgttcagaagacaatgttg
901atgttcatgatatagaattgttacagtatatcaatgtggattgtgcaaaattaaaacgac
961tcctgaaggaaacagcatttaaatttaaagccctaaagaaggttgcgcagttagcagtta
1021taaatagcctggaaaaggcattttggaactgggtagaaaattatccagatgaatttacaa
1081aactgtaccagatcccacagactgatatggctgaatgtgcagaaaagctatttgacttgg
1141tggatggttttgctgaaagcaccaaacgtaaagcagcagtttggccactacaaatcattc
1201tccttatcttgtgtccagaaataatccaggatatatccaaagacgtggttgatgaaaaca
1261acatgaataagaagttatttctggacagtctacgaaaagctcttgctggccatggaggaa
1321gtaggcagctgacagaaagtgctgcaattgcctgtgtcaaactgtgtaaagcaagtactt
1381acatcaattgggaagataactctgtcattttcctacttgttcagtccatggtggttgatc
1441ttaagaacctgctttttaatccaagtaagccattctcaagaggcagtcagcctgcagatg
1501tggatctaatgattgactgccttgtttcttgctttcgtataagccctcacaacaaccaac
1561actttaagatctgcctggctcagaattcaccttctacatttcactatgtgctggtaaatt
1621cactccatcgaatcatcaccaattccgcattggattggtggcctaagattgatgctgtgt
1681attgtcactcggttgaacttcgaaatatgtttggtgaaacacttcataaagcagtgcaag
1741gttgtggagcacacccagcaatacgaatggcaccgagtcttacatttaaagaaaaagtaa
1801caagccttaaatttaaagaaaaacctacagacctggagacaagaagctataagtatcttc
1861tcttgtccatggtgaaactaattcatgcagatccaaagctcttgctttgtaatccaagaa
1921aacaggggcccgaaacccaaggcagtacagcagaattaattacagggctcgtccaactgg
1981tccctcagtcacacatgccagagattgctcaggaagcaatggaggctctgctggttcttc
2041atcagttagatagcattgatttgtggaatcctgatgctcctgtagaaacattttgggaga
2101ttagctcacaaatgcttttttacatctgcaagaaattaactagtcatcaaatgcttagta
2161gcacagaaattctcaagtggttgcgggaaatattgatctgcaggaataaatttcttctta
2221aaaataagcaggcagatagaagttcctgtcactttctccttttttacggggtaggatgtg
2281atattccttctagtggaaataccagtcaaatgtccatggatcatgaagaattactacgta
2341ctcctggagcctctctccggaagggaaaagggaactcctctatggatagtgcagcaggat
2401gcagcggaacccccccgatttgccgacaagcccagaccaaactagaagtggccctgtaca
2461tgtttctgtggaaccctgacactgaagctgttctggttgccatgtcctgtttccgccacc
2521tctgtgaggaagcagatatccggtgtggggtggatgaagtgtcagtgcataacctcttgc
2581ccaactataacacattcatggagtttgcctctgtcagcaatatgatgtcaacaggaagag
2641cagcacttcagaaaagagtgatggcactgctgaggcgcattgagcatcccactgcaggaa
2701acactgaggcttgggaagatacacatgcaaaatgggaacaagcaacaaagctaatcctta
2761actatccaaaagccaaaatggaagatggccaggctgctgaaagccttcacaagaccattg
2821ttaagaggcgaatgtcccatgtgagtggaggaggatccatagatttgtctgacacagact
2881ccctacaggaatggatcaacatgactggcttcctttgtgcccttgggggagtgtgcctcc
2941agcagagaagcaattctggcctggcaacctatagcccacccatgggtccagtcagtgaac
3001gtaagggttctatgatttcagtgatgtcttcagagggaaacgcagatacacctgtcagca
3061aatttatggatcggctgttgtccttaatggtgtgtaaccatgagaaagtgggacttcaaa
3121tacggaccaatgttaaggatctggtgggtctagaattgagtcctgctctgtatccaatgc
3181tatttaacaaattgaagaataccatcagcaagttttttgactcccaaggacaggttttat
3241tgactgataccaatactcaatttgtagaacaaaccatagctataatgaagaacttgctag
3301ataatcatactgaaggcagctctgaacatctagggcaagctagcattgaaacaatgatgt
3361taaatctggtcaggtatgttcgtgtgcttgggaatatggtccatgcaattcaaataaaaa
3421cgaaactgtgtcaattagttgaagtaatgatggcaaggagagatgacctctcattttgcc
3481aagagatgaaatttaggaataagatggtagaatacctgacagactgggttatgggaacat
3541caaaccaagcagcagatgatgatgtaaaatgtcttacaagagatttggaccaggcaagca
3601tggaagcagtagtttcacttctagctggtctccctctgcagcctgaagaaggagatggtg
3661tggaattgatggaagccaaatcacagttatttcttaaatacttcacattatttatgaacc
3721ttttgaatgactgcagtgaagttgaagatgaaagtgcgcaaacaggtggcaggaaacgtg
3781gcatgtctcggaggctggcatcactgaggcactgtacggtccttgcaatgtcaaacttac
3841tcaatgccaacgtagacagtggtctcatgcactccataggcttaggttaccacaaggatc
3901tccagacaagagctacatttatggaagttctgacaaaaatccttcaacaaggcacagaat
3961ttgacacacttgcagaaacagtattggctgatcggtttgagagattggtggaactggtca
4021caatgatgggtgatcaaggagaactccctatagcgatggctctggccaatgtggttcctt
4081gttctcagtgggatgaactagctcgagttctggttactctgtttgattctcggcatttac
4141tctaccaactgctctggaacatgttttctaaagaagtagaattggcagactccatgcaga
4201ctctcttccgaggcaacagcttggccagtaaaataatgacattctgtttcaaggtatatg
4261gtgctacctatctacaaaaactcctggatcctttattacgaattgtgatcacatcctctg
4321attggcaacatgttagctttgaagtggatcctaccaggttagaaccatcagagagccttg
4381aggaaaaccagcggaacctccttcagatgactgaaaagttcttccatgccatcatcagtt
4441cctcctcagaattcccccctcaacttcgaagtgtgtgccactgtttataccaggtggtta
4501gccagcgtttccctcagaacagcatcggtgcagtaggaagtgccatgttcctcagattta
4561tcaatcctgccattgtctcaccgtatgaagcagggattttagataaaaagccaccaccta
4621gaatcgaaaggggcttgaagttaatgtcaaagatacttcagagtattgccaatcatgttc
4681tcttcacaaaagaagaacatatgcggcctttcaatgattttgtgaaaagcaactttgatg
4741cagcacgcaggtttttccttgatatagcatctgattgtcctacaagtgatgcagtaaatc
4801atagtctttccttcataagtgacggcaatgtgcttgctttacatcgtctactctggaaca
4861atcaggagaaaattgggcagtatctttccagcaacagggatcataaagctgttggaagac
4921gaccttttgataagatggcaacacttcttgcatacctgggtcctccagagcacaaacctg
4981tggcagatacacactggtccagccttaaccttaccagttcaaagtttgaggaatttatga
5041ctaggcatcaggtacatgaaaaagaagaattcaaggctttgaaaacgttaagtattttct
5101accaagctgggacttccaaagctgggaatcctattttttattatgttgcacggaggttca
5161aaactggtcaaatcaatggtgatttgctgatataccatgtcttactgactttaaagccat
5221attatgcaaagccatatgaaattgtagtggaccttacccataccgggcctagcaatcgct
5281ttaaaacagactttctctctaagtggtttgttgtttttcctggctttgcttacgacaacg
5341tctccgcagtctatatctataactgtaactcctgggtcagggagtacaccaagtatcatg
5401agcggctgctgactggcctcaaaggtagcaaaaggcttgttttcatagactgtcctggga
5461aactggctgagcacatagagcatgaacaacagaaactacctgctgccaccttggctttag
5521aagaggacctgaaggtattccacaatgctctcaagctagctcacaaagacaccaaagttt
5581ctattaaagttggttctactgctgtccaagtaacttcagcagagcgaacaaaagtcctag
5641ggcaatcagtctttctaaatgacatttattatgcttcggaaattgaagaaatctgcctag
5701tagatgagaaccagttcaccttaaccattgcaaaccagggcacgccgctcaccttcatgc
5761accaggagtgtgaagccattgtccagtctatcattcatatccggacccgctgggaactgt
5821cacagcccgactctatcccccaacacaccaagattcggccaaaagatgtccctgggacac
5881tgctcaatatcgcattacttaatttaggcagttctgacccgagtttacggtcagctgcct
5941ataatcttctgtgtgccttaacttgtacctttaatttaaaaatcgagggccagttactag
6001agacatcaggtttatgtatccctgccaacaacaccctctttattgtctctattagtaaga
6061cactggcagccaatgagccacacctcacgttagaatttttggaagagtgtatttctggat
6121ttagcaaatctagtattgaattgaaacacctttgtttggaatacatgactccatggctgt
6181caaatctagttcgtttttgcaagcataatgatgatgccaaacgacaaagagttactgcta
6241ttcttgacaagctgataacaatgaccatcaatgaaaaacagatgtacccatctattcaag
6301caaaaatatggggaagccttgggcagattacagatctgcttgatgttgtactagacagtt
6361tcatcaaaaccagtgcaacaggtggcttgggatcaataaaagctgaggtgatggcagata
6421ctgctgtagctttggcttctggaaatgtgaaattggtttcaagcaaggttattggaagga
6481tgtgcaaaataattgacaagacatgcttatctccaactcctactttagaacaacatctta
6541tgtgggatgatattgctattttagcacgctacatgctgatgctgtccttcaacaattccc
6601ttgatgtggcagctcatcttccctacctcttccacgttgttactttcttagtagccacag
6661gtccgctctcccttagagcttccacacatggactggtcattaatatcattcactctctgt
6721gtacttgttcacagcttcattttagtgaagagaccaagcaagttttgagactcagtctga
6781cagagttctcattacccaaattttacttgctgtttggcattagcaaagtcaagtcagctg
6841ctgtcattgccttccgttccagttaccgggacaggtcattctctcctggctcctatgaga
6901gagagacttttgctttgacatccttggaaacagtcacagaagctttgttggagatcatgg
6961aggcatgcatgagagatattccaacgtgcaagtggctggaccagtggacagaactagctc
7021aaagatttgcattccaatataatccatccctgcaaccaagagctcttgttgtctttgggt
7081gtattagcaaacgagtgtctcatgggcagataaagcagataatccgtattcttagcaagg
7141cacttgagagttgcttaaaaggacctgacacttacaacagtcaagttctgatagaagcta
7201cagtaatagcactaaccaaattacagccacttcttaataaggactcgcctctgcacaaag
7261ccctcttttgggtagctgtggctgtgctgcagcttgatgaggtcaacttgtattcagcag
7321gtaccgcacttcttgaacaaaacctgcatactttagatagtctccgtatattcaatgaca
7381agagtccagaggaagtatttatggcaatccggaatcctctggagtggcactgcaagcaaa
7441tggatcattttgttggactcaatttcaactctaactttaactttgcattggttggacacc
7501ttttaaaagggtacaggcatccttcacctgctattgttgcaagaacagtcagaattttac
7561atacactactaactctggttaacaaacacagaaattgtgacaaatttgaagtgaatacac
7621agagcgtggcctacttagcagctttacttacagtgtctgaagaagttcgaagtcgctgca
7681gcctaaaacatagaaagtcacttcttcttactgatatttcaatggaaaatgttcctatgg
7741atacatatcccattcatcatggtgacccttcctataggacactaaaggagactcagccat
7801ggtcctctcccaaaggttctgaaggataccttgcagccacctatccaactgtcggccaga
7861ccagtccccgagccaggaaatccatgagcctggacatggggcaaccttctcaggccaaca
7921ctaagaagttgcttggaacaaggaaaagttttgatcacttgatatcagacacaaaggctc
7981ctaaaaggcaagaaatggaatcagggatcacaacaccccccaaaatgaggagagtagcag
8041aaactgattatgaaatggaaactcagaggatttcctcatcacaacagcacccacatttac
8101gtaaagtttcagtgtctgaatcaaatgttctcttggatgaagaagtacttactgatccga
8161agatccaggcgctgcttcttactgttctagctacactggtaaaatataccacagatgagt
8221ttgatcaacgaattctttatgaatacttagcagaggccagtgttgtgtttcccaaagtct
8281ttcctgttgtgcataatttgttggactctaagatcaacaccctgttatcattgtgccaag
8341atccaaatttgttaaatccaatccatggaattgtgcagagtgtggtgtaccatgaagaat
8401ccccaccacaataccaaacatcttacctgcaaagttttggttttaatggcttgtggcggt
8461ttgcaggaccgttttcaaagcaaacacaaattccagactatgctgagcttattgttaagt
8521ttcttgatgccttgattgacacgtacctgcctggaattgatgaagaaaccagtgaagaat
8581ccctcctgactcccacatctccttaccctcctgcactgcagagccagcttagtatcactg
8641ccaaccttaacctttctaattccatgacctcacttgcaacttcccagcattccccaggaa
8701tcgacaaggagaacgttgaactctcccctaccactggccactgtaacagtggacgaactc
8761gccacggatccgcaagccaagtgcagaagcaaagaagcgctggcagtttcaaacgtaata
8821gcattaagaagatcgtgtgaagcttgcttgctttcttttttaaaatcaacttaacatggg
8881ctcttcactagtgaccccttccctgtccttgccctttccccccatgttgtaatgctgcac
8941ttcctgttttataatgaacccatccggtttgccatgttgccagatgatcaactcttcgaa
9001gccttgcctaaatttaatgctgccttttctttaactttttttcttctacttttggcgtgt
9061atctggtatatgtaagtgttcagaacaactgcaaagaaagtgggaggtcaggaaactttt
9121aactgagaaatctcaattgtaagagaggatgaattcttgaatactgctactactggccag
9181tgatgaaagccatttgcacagagctctgccttctgtggttttcccttcttcatcctacag
9241agtaaagtgttagtcctatttatacatttttcaagatacaagtttatgagagaaatagta
9301ttataaccccagtatgtttaatcttttagctgtggactttttttttaaccgtacaaaact
9361gaaagaaccatagaggtcaagcctcagtgacttgacaccataaagccacagacaaggtac
9421ttgggggggagggcagggaaatttcatattttatagtggattcttaagaaatactaacac
9481ttgagtattagcaataattacaggaaaataagtgcgaccacatatatcttaacattactg
9541aattaaaactatggcttctaagtccttatccaaactcagtcatccaaactagtttatttt
9601tttctccagttgattatcttttaatttttaattttgctaaaggtggtttttttgtgtttt
9661gttttttgtaaaccaaaactatactaagtatagtaattatatatatatatatattttttc
9721ccctccccctcttctttcctaactaattctgagcagggtaatcagtgaacaaagtgttga
9781aaattgttcccagaaggtaattttcatagatgtttgcattagctccatagcaaaatggaa
9841tggtacgtgacatttagggtagctgatatttttattttgttaaataatttccaagaatag
9901agtatggtgtatattataaatttctttgataagatgtattttgaatgtcttttaatcttc
9961ctcctcctctccaaaaaaatcagaaacctctttaagaaaacatgtaggttatatatgcta
10021gaattgcatttaatcactgtgaaaagactggtcagcctgcattagtatgacagtaggggg
10081gctgttagaattgctgctatactggtggtatggattatcatggcattggaattttcatag
10141taatgcagatccaatttctttgtggtacctgcagtttacaaaataatttgacttcagtga
10201gcatattggtatctggatgttccaatttagaactaaaccatatttattacaaaaagatat
10261taatccctctactcccaggttccctttatatgttaagatataatggctttgaggggggaa
10321aaaataaacctaggggagaggggagtttcctgtagtgctgtttcattagaggatttcagt
10381aaattaaattccacagctaattcaataaataatggtacatttaagtgttctgattttaat
10441aatatatttcacatttatccacacagtaacaatgtaatatgttaatgtaaataaaattgg
10501ttttgatactcagaaataacaagaatttaattttttaaatttgtttacagtcctgggaaa
10561agtaagaattatttgccaaaataagaggaaagaaaaccttagtattattaatgagtttac
10621catagaattgttggaaatactgaagacaggtgcaatttactaaacttttgtttttaaact
10681attgtagaggctgcattagaagaaaatgtttataatgacagagcaactatgactatataa
10741aaaagctgaaattagaactgtgtttagaaatagatcagtaacccagtgccaaggatgcca
10801agctgccaccatggtcttggctctcccacaacccagtgtttctggggtaagtttcacagt
10861ttctaggccctggaatagcaggcagtgtaagcctttgataactttagttcgatgtttttc
10921ttgtttttgtttgttggtttggtgcatatgatagtgggtgttatgctattttgctcttcc
10981catcaaaataaagaaacttccagaggtttactgttaaaaatactgatatttccataaacg
11041ggtttaccaagggtgtagtatttcataccgcctgaaatgatcagcattggcacaaatcaa
11101aattcagccgcctttgaaatgcaaaaatacctttgactagtaagtacatcctaggagttt
11161gaaaacttaactaaggtttaaaatttaccttgtttaaagaacttctgacttttgaggaaa
11221atctagctttccaagtaactaaaatgtacatgagataaacctctcaccactatgtgtccc
11281ttgagaaatgcaacacttttttagtcttcatacttgtaatctataaaagaaattctgaag
11341tttagaccaagttgcccatttctgcgtaattgacataagttctgttaaaaatattataag
11401taattcgtttcggtttgtagatgtttcccctgacttgttaaagaggaaaccaggaactca
11461gtcatgtttttgtcctggataatctacctgttatgccagtactcccatccgaggggcatg
11521cccttagttgcccagatggagatgcagttcagtagatttggggcaaagtggctacagctc
11581tgtcttccattcactcaacacctgttcatgactgagccaggtgcccaggacacatcctaa
11641acagtcagcttctatcctgtgtcctagttggggagacagagtgccagccagcaaccctcc
11701caggtttgtaggttttaggggttttcagttttgtttgggttttttgttttttgtttttgt
11761ttctacatccttccccgactcccaggcataatgaggcatgtcttactcaatgttatgcaa
11821tggatttaggcaaaaattcattcttagtgtcagccacacaattttttttaatgcagtata
11881ttcacctgtaaatagtttgtgtaaaatttgacaaaaaaagtatatttactatactgtaaa
11941tatatgtgatgatatattgtattattttgcttttttgtaaagcagttagttgctgcacat
12001ggataacaacaaaaatttgattattctcgtgttagtattgttaacttctttttgcgactg
12061cgttacatcatttaaagaaaatgctgtgtattgtaaacttaaattgtatatgataactta
12121ctgtcctttccatccgggcctaaactttggcagttcctttgtctacaaccttgttaatac
12181tgtaaacagttgtacgccagcaggaaaaatactgcccaacagacaaaatcgatcattgta
12241ggggaaaatcatagaaatccatttcagatctttattgttcctcaccccattttcctcctt
12301gtgtatgtacttcccccacccccctttttttaagtaaaatgtaaattcaatctgctctaa
12361gaaaaaaaaaaaaaaaaaaaa(seqidno:10)
nm_00518811241bpmrna智人cbl原癌基因(cbl)、mrna。
1tccgcccggatagccggcggcggcggcggcggcggcggcggcggcggccgggagaggccc
61ctccttcacgccctgcttctctccctcgctcgcagtcgagccgagccggcggacccgcct
121gggctccgaccctgcccaggccatggccggcaacgtgaagaagagctctggggccggggg
181cggcagcggctccgggggctcgggttcgggtggcctgattgggctcatgaaggacgcctt
241ccagccgcaccaccaccaccaccaccacctcagcccccacccgccggggacggtggacaa
301gaagatggtggagaagtgctggaagctcatggacaaggtggtgcggttgtgtcagaaccc
361aaagctggcgctaaagaatagcccaccttatatcttagacctgctaccagatacctacca
421gcatctccgtactatcttgtcaagatatgaggggaagatggagacacttggagaaaatga
481gtattttagggtgtttatggagaatttgatgaagaaaactaagcaaaccataagcctctt
541caaggagggaaaagaaagaatgtatgaggagaattctcagcctaggcgaaacctaaccaa
601actgtccctcatcttcagccacatgctggcagaactaaaaggaatctttccaagtggact
661ctttcagggagacacatttcggattactaaagcagatgctgcggaattttggagaaaagc
721ttttggggaaaagacaatagtcccttggaagagctttcgacaggctctacatgaagtgca
781tcccatcagttctgggctggaggccatggctctgaaatccactattgatctgacctgcaa
841tgattatatttcggtttttgaatttgacatctttacccgactctttcagccctggtcctc
901tttgctcaggaattggaacagccttgctgtaactcatcctggctacatggcttttttgac
961gtatgacgaagtgaaagctcggctccagaaattcattcacaaacctggcagttatatctt
1021ccggctgagctgtactcgtctgggtcagtgggctattgggtatgttactgctgatgggaa
1081cattctccagacaatccctcacaataaacctctcttccaagcactgattgatggcttcag
1141ggaaggcttctatttgtttcctgatggacgaaatcagaatcctgatctgactggcttatg
1201tgaaccaactccccaagaccatatcaaagtgacccaggaacaatatgaattatactgtga
1261gatgggctccacattccaactatgtaaaatatgtgctgaaaatgataaggatgtaaagat
1321tgagccctgtggacacctcatgtgcacatcctgtcttacatcctggcaggaatcagaagg
1381tcagggctgtcctttctgccgatgtgaaattaaaggtactgaacccatcgtggtagatcc
1441gtttgatcctagagggagtggcagcctgttgaggcaaggagcagagggagctccctcccc
1501aaattatgatgatgatgatgatgaacgagctgatgatactctcttcatgatgaaggaatt
1561ggctggtgccaaggtggaacggccgccttctccattctccatggccccacaagcttccct
1621tcccccggtgccaccacgacttgaccttctgccgcagcgagtatgtgttccctcaagtgc
1681ttctgctcttggaactgcttctaaggctgcttctggctcccttcataaagacaaaccatt
1741gccagtacctcccacacttcgagatcttccaccaccaccgcctccagaccggccatattc
1801tgttggagcagaatcccgacctcaaagacgccccttgccttgtacaccaggcgactgtcc
1861ctccagagacaaactgccccctgtcccctctagccgccttggagactcatggctgccccg
1921gccaatccccaaagtaccagtatctgccccaagttccagtgatccctggacaggaagaga
1981attaaccaaccggcactcacttccattttcattgccctcacaaatggagcccagaccaga
2041tgtgcctaggctcggaagcacgttcagtctggatacctccatgagtatgaatagcagccc
2101attagtaggtccagagtgtgaccaccccaaaatcaaaccttcctcatctgccaatgccat
2161ttattctctggctgccagacctcttcctgtgccaaaactgccacctggggagcaatgtga
2221gggtgaagaggacacagagtacatgactccctcttccaggcctctacggcctttggatac
2281atcccagagttcacgagcatgtgattgcgaccagcagattgatagctgtacgtatgaagc
2341aatgtataatattcagtcccaggcgccatctatcaccgagagcagcacctttggtgaagg
2401gaatttggccgcagcccatgccaacactggtcccgaggagtcagaaaatgaggatgatgg
2461gtatgatgtcccaaagccacctgtgccggccgtgctggcccgccgaactctctcagatat
2521ctctaatgccagctcctcctttggctggttgtctctggatggtgatcctacaacaaatgt
2581cactgaaggttcccaagttcccgagaggcctccaaaaccattcccgcggagaatcaactc
2641tgaacggaaagctggcagctgtcagcaaggtagtggtcctgccgcctctgctgccaccgc
2701ctcacctcagctctccagtgagatcgagaacctcatgagtcaggggtactcctaccagga
2761catccagaaagctttggtcattgcccagaacaacatcgagatggccaaaaacatcctccg
2821ggaatttgtttccatttcttctcctgcccatgtagctacctagcacaccatctccctgct
2881gcaggtttagaggaccagtgagttgggagttattactcaagtggcacctagaagggcagg
2941agttcctttggtgacttcacagtgaagtcttgccctctctgtgggatatcacatcagtgg
3001ttccaagatttcaaagtggtgaaatgaaaatggagcagctagtatgttttattattttat
3061gggtcttgagtgcatttgaaggtgtccttcagttcccacgtagagagagtgtggattata
3121ttacatgataacctacctggggaacagtccagaaagctatagaacaagtattttgctgga
3181aatcctaattgaggacttaagacttcctgggttaaggatgtggccgtgtgtgtgtgtgtc
3241tgcctgtggttgtatgtgtccttgtgattataagattaacctgctgtgtgtgttaattcc
3301aggcagggaattagcacaaaaggtttaggaaggaatctttttttaaagacttccatctac
3361tgtggtattatacccaagcctagtgtgtattacaacttcaacactcccctttggcttata
3421ttaccatgtgcatagctaaagtcttctatttttagaacaccttctgtctgttctttcccc
3481atcaactccttcctcatccttcttggtgttctgtcatgggccatgggcttgctatggcca
3541gccttactgaggccaagcagcttatgggatgttctttattgtgtgtgatggtattggttt
3601gtttggtagataagtgggaggaaaagtactgttgctacactattataggcatgtttgata
3661ctagcagctaacactggtcactccaaagcactgtttctataggaacattgaagctattaa
3721gatgttttgattatcctaattacataatgaccgatttgagatagaggcctttaaatacat
3781tccatgccctccccagaaaatagtctgtgggagtcagttgccttggtgccaggtatgtgt
3841tctgatgtaggtcatgagtctttctacttaatgggaagggaagaacatttgtttccagga
3901tgactttctggccagaataccggaaagcttttaggaagcttcgttcacatgctatttaaa
3961tgcacaaaatagacagtaaggatttatctgttcagtttttcttcccagtgaattaatttc
4021agcttatatgggtgtcttcatttgaacatgaggaatattaggttatattttcagcagtgg
4081ttttttcctttgccctttaaggagtggggataatgtccacggtggcccagcctcttgctg
4141atggcaccttccctgcattgctgcctcccgatgatgtggttcttttcttgtgcctgtggc
4201tttgggaatgtaacatctctttcctcctttccttcccttttcctcttcacctgaggtcct
4261aaatactctctgtaattactgtgttcttcacggtaattagacatcattcagtgaataaat
4321tactgtagtcaaagacagtatgggctggcagtttgtgtaattgcaagttcataaagagaa
4381ttgagggtccagttgggagaactattagtcagttcttttatatgctgataaatgatccct
4441cgagttcagttagtattctgtccagagtgtttagctcactttcttagcagtgtgtaagct
4501ttctccatgtcagaagcaagcctgctctttgataaatctgtcttcctgaaaatctaaatc
4561atgcttttgtctttagatctacacagaaatgaccctccttggatcagttttctttccagt
4621ctaatcatctttggaactaaaacttgttctaactcgtctcttggcattcagctactccta
4681gatcttttggttttatcccctggcctcagagccatttatattcccagagtaggcagtaca
4741ggatctcgtgttgatttgctgtggttacccagtgtcttctctacatggcataaagcggca
4801aagcccaccattaggtgaggcggtcccgagttgaggtagagtggggcagaggaagatggc
4861agtgaatatcaaacagtagaccgccatcaacttctaacagccagtacacacactgtttca
4921ttttgaggtaacgttcagttttgcattttgtttaaatattgaaggcctagacaaagaact
4981agaaaaaaaaaagcagtttccaggcccatccatattgtaatttttctttatctgcagata
5041ttgcctgtagtctaaagatctctttggaagacaaagcattggctatatatcttttgcctt
5101ttccatgcatctaaatcttctctggagattatctccctactgtgtaggttaagggcagtc
5161tcgacttttccttttttgagtcctgtgtggctctttgaatcagcgtgaaactgaggctcc
5221agctccctgtgttgtgtgtgtgtgccatccatgggcttgggtgtcagtttgtcacaggta
5281tctgccagcattcaaggttttggatcatttcatgaggatctttcctttgactgggtgctg
5341tgaggacacacctgggtctgtgcctgagattgccaggcaagattaaggaaagttttcatg
5401tggcttttgttttgaggttattctcaaaaccttaatttcttatattttctgttgactaag
5461gcaccagtaacccattcttcaccctccatttgtatggcaatttaaaagtctttggctttg
5521ctctgaatttaattaaaactgccttttatgaacagacttcgagttttgccattttgggca
5581agcccttccgcttgtcccttcctagtggctaataaagtaaaaaaacccacactactttgt
5641tctctttttctcatattcattgggctgttgtattcagccagtctcatgctttccctgggt
5701cttcacggattgctttccaagctgccttgttgcggggttgctgcagagcagcaactggac
5761ctttccagctgtcgccatgttccttccactaaagtagagggttcttaaaatggaaaaacc
5821tgtgggctcttcatatacctccctttagttaagtaatagaccaggcagcttctcatctca
5881gcatttacctgttaatatttttgtgaatagtgctctctacctgtgggtggccgttctctt
5941ccacttgctcgtctccccccagccccattctgcataatctaccattcttctcctctcttt
6001ctcttcttatacagaccctcattactggggcccaagatgtgggatactactgttagtatt
6061atttaactattttgtagatttaaaagatttctggttaagggaggtgggggtcactgttca
6121tcactcttaaaatatgtgttttctctatagaaaagtaaaatgtgtttatggtcccaaaca
6181gtcaactcacaaatttttataacaaaatttccttgtaaaaactagggaccatctatatat
6241tccctttaagatctagttctttttgtaggtgttcagcaatggtgataaagcagaatattc
6301tcctacctcacgtcattaaagtcagaagattatagaccttctcaaactataagtccctct
6361tcttgccgttggcctttctgactctggaatgaccactgttcattgaaaaatagttttctg
6421actattggtctggctctaacagtttgtttgttcatccagcaaatgtttatgagtgatgac
6481catgtgccagaaatgtcaggtatgtgtccttcccttggcgccacatagtagtttactaat
6541gtttgggggattgtacttggactgtcatagcctctgcgtttgaccttaaaatagctcttc
6601ccagtaagattgtgcaatttttattcacagctcttccatgtagacttacctttcctcata
6661gagctatcctggttaataacaggccaagattctcccattatcccctgttgtctcctgtag
6721ctttgataatgcctgggagattccttggtgtaagtgtcatggataccgactgtttttatg
6781ttggaatttgttccaacataattagaatctgtttggtgagttgaaaggtaagttggctca
6841gagttgcacagtagggcattaaatgtttaagcaaagcatctgcccacactcccctttcca
6901atctagtgccttccttgaactttttcctgagctgctacgtccctaatcccccttgttggg
6961aggattttcgtatcacccttatgggacctgtcaccatgtcctgtactatttggaattggt
7021tttccagtctttcaacaaccgttgtggctaactatgttttagaagggctggaggtgtggg
7081ccctgtcttcgggtctcaggacccaaagatcctttagtcagttgttgggtcttccaagag
7141ccagacattaatacagattgaactccatcagtcccctaattgtcagcctttacctccctc
7201ccagagcaaggagtttagggattctaaagcttagtgtccacacatcattctaccagacct
7261tagagctttagaagctcaatctaaaatactgtaactcagcataaactattactatcactc
7321ctttgaactcagtctccatgagcagtgttttgttggaaatacatagaacggcttaatgcc
7381tagagggtggtggatagtgaaggacggtcaaggttatatttttgactgcttagggattct
7441ttggatccaagaaacagaaatgttcaagcggaataaaggagggagtggagttgtggtaag
7501gatgcagggtatttcgcagaacccaggacgggaagtgcctttggttcttgggtggagctg
7561gaactgcagagctttgcacctagtcctttctcccgcttcacagtctgcttatggtatatg
7621tggcccccaaataggcactctagtcctcaagtctacaccaccttccaactctggggatca
7681ccatgaacaaattctcaatttcccatacttaattttttttttttttgagatggagtctcg
7741ctgtgtcgcccaggctggagtgcagtggtgcagtctcaactcaccacaacctctgcctcc
7801caggttcaagcagttctctgcctcaacctcccgagtagctgggattacaggcgcctgcca
7861ccatgcccagctaatgttcatatttttagtagagacagggtttcaccgtcttggctaggc
7921tggtcttgaactcctgaccctcatgatccacccacctcggcctcccaaagtgctaagatt
7981acaggcgtgagccaccgcgcccggcccatacttcgtattcttaaaaaaaactacactcag
8041cccagcacattgatcaagtatctatctctgagcagttggccttgccagggagagcagaga
8101tgtggcaggctccttcagctggagacagggagcttctcagagaagtgagcagagactcca
8161cagacaccctaaaaaggcttctactcaagaagtaaagccactactcctgcctttttgctt
8221agtggacaggaaggcacaggagtttgtctgggacatcatagaaattcttaggtttaactt
8281aattctggtcattgtcttctttatttcctgtttttcttccctttgtcagtcttcgcatcc
8341aagatttcttccctccctcttgtgggccagcctgtcctgttccagagctagcctgttcct
8401gggtagccttccttagcctccattcagcctcaggtcttttgccttcttccgtgtttattt
8461agagagcagaatctaataacgggttccactgtagccactatccatggacttctgggtcct
8521cttcaggtttgagtgcttgaaaatgttcattctctgggcttgtggcctgtctcctccact
8581ctcctcctcaccctctcgctccttcctgtgtgagggccgctctgcagtaatgttctcagg
8641caagccttcctaggcacctcagaaactactttgccagagccagtaagaatatataatatt
8701ggagcagttgccaggatagaaattaaatatagattccagtttaggatagagtttttaccg
8761agagctctttagacagtatacctgtgtcttctctggcaattgctttcattttagtcctat
8821ataaaagctttccttttctgtttttttttaaaactatgcttttgcttgcctaaatctttt
8881gatcttatatttctctcatctcagagcctgtcctgagttgtaaggtatttcatactgcct
8941tacttaaaagttttttaaactactagagtcatttgatacacacagaagttacctaataat
9001ccaaagatgtccatcaagggaggaagggtgggtcatcagactttgcctttgatgttgtag
9061actaggctcctgagttaagcagcagagggacagcagtgccatgtgccttcactgtgtccc
9121aggaaatctgggttggttccagtgggaaataccagtatttcttggttctggaaagtagca
9181aaagagtaggagatggggaaatagggatggggagagcaagccccgcatgtccatggcgag
9241tcaggtggggagcacgggtggaagggccggctgttgacagacagactaagctgtgtggtg
9301ctcttgccgccccttcctgggtacagagcttgagaaaaatgcagccgaccactccctgtg
9361tttgtacagagcaaagcccaaaagccaacctcagatctcctgatttggcagctgaagaaa
9421tcagcagagtcctgattgcctgattcagtcccaaaaatgaatgtcaggccccgccccctc
9481cccaccaacattgcctctcctacattctccttctgcccctaaatcagacaggaggccaga
9541gaggagtattgctcaatgcgtgctatgtgcaactcctcaggccttgtgccacctccatgc
9601tgagccctgaagcagggtgtcctgggtgcctgtgtgtcagctccctcctctctacctacc
9661tctgaccttcttgtgggtgagggtggccatgcttatggccatcttaaaactggagaggca
9721gagaactacttatgagtctgtagaccacgtgttgtcttccatggcctgtttctcctgctg
9781tctgggtgagtgagcctgcaacgcaatgcccatgagagtaaatgcctcctgacctaccct
9841gctcagcactgttctagtgtcttggccttgaaagaaaagcctgacttcctgctgacacat
9901gtggtaggggcatggcagctatgaggcacctcctacgtctgttttctggctgtggtgact
9961tgggatttttaaccttatatatctttttcctttactcaaaacaaaacaatttttagcaca
10021ctgaaaaaaaaaaaaagccaaatgttttgtgcctttctaaggcagcactgtatcccaggc
10081tgcattttaggacttaatatggaaataccagagtctgagctcctctaccttgagtttcat
10141tagtccttagtgtctaggagacaggaaagaatgctctctgtgactggagaggtgacatgc
10201aggtgcagtgtgtctggagtccctttcccctgctgtgagacttcagtggaggagagaagc
10261attgtaccctgggatcatttggttggttccaatcacaagcttagttatcaggttgcatgc
10321cttgtctcctgcaaaagacagaatgtttcacaattcccaggtaaactctggaccattcca
10381agtgtcctagccttctgatgacattaattacctagttgtgtcgaggagtataggatggac
10441tctcctgagaaggggaggttggtggctttgtcttttctttttgctggatcctgaactggt
10501ctagacctcctgcccccaccccccagcccccatcagatgtggctggcctttcatttgaag
10561gcttcagacttaaagcattaagcagctagtgccctctgcagggcctggtttccccaggga
10621agggcagcaaggaacatgggaccagaagcctgtcctcagtaatgtgactatagtgagctt
10681tagcaaaagtttttctatataatgacatcttacttatcttttaccctttcctcagttttc
10741ccctgcctttaactaataaagaattgggagacagaaattttaaagtcctccttattcaag
10801attttgaaattcttagcctgggagtgctggagagaacctgatgctttctccagaatgaag
10861agtcccaatttgtatatcagtgttaagaagaaaacaaaacaaacacataggtgagatttt
10921cgtggactattttaaaaatgtgtcattaatataaaaaatttatattagcagtatttaatc
10981attctcacctgtaaagaataagaaaaacagaaggtaaatattcttacagagaatagcaga
11041gctttaagattcattttcattttaagtccattttattttgccagtgtattaatgtttaga
11101agtctgttttactaatgttatttattaattttttttcatttccatacacagttagttaac
11161taaagagctttttcaagcacccatgtctgtaaaaaaatatttttaaataaagtttctttt
11221gttgtagcagaaaaaaaaaaa
ras病(rasopathies)是一组遗传异质性病症,包括但不限于noonan综合征(ns)、心血管筋膜综合症(cfc)、1型神经纤维瘤病(nf1)、noonan综合征伴多发痣(nsml)、costello综合征(cs)和legius综合征,其具有ras/mapk途径突变,所述突变涉及ptpn11、sos1、raf1、kras、hras、map2k1、map2k2、nras、cbl、shoc2、braf、rit1、a2ml1、spred1和nf1。ns中的淋巴缺陷已在部分ns患者中进行了描述,但其严重程度、位置和发作时间有所不同。回顾文献,我们总共鉴定了52名表现出noonan或noonan样综合征和淋巴缺陷的临床特征的产前和产后患者,所述临床特征包括胸膜积液、心包积液、乳糜胸、积水、淋巴管扩张和淋巴水肿。我们鉴定了可解释其疾病的ptpn11中的突变。因此,对ras/mapkerk途径的治疗性干预应逆转这些患者的疾病表型,类似于我们先前对araf突变所示出的。因此,我们的中枢传导淋巴管异常患者具有ras/mapk途径中的生殖系或体细胞突变的发现是新颖的,并确定了治疗剂开发的新靶标。支持这些ras/mapk途径的细胞和模型数据描述如下。
开发了用于研究潜在的erk激活突变影响的细胞模型。我们已经在细胞系ea.hy926中逆转录表达了野生型和突变型的蛋白质。所述ea.hy926细胞系是人类脐静脉内皮细胞与黑色素瘤细胞系的融合体。它是永生化的,并保留了内皮细胞的某些特征。我们已经成功地使用该细胞系来证明araf基因突变的细胞后果,这种突变会导致某种形式的中枢传导淋巴管异常。我们还示出了使用曲美替尼(一种mek抑制剂)在同一细胞系中突变型araf的作用的逆转。
图1示出了表达wt型或突变型braf或ptpn11蛋白的ea.hy926细胞中激活的erk的水平。当相对于蛋白质加载标准化时,在该实验中,表达突变braf的细胞所含的磷酸化erk的量是表达野生型braf的细胞的5倍以上。如先前用araf所观察到的,突变braf比wt表达更高。
在图2中,可以在表达f486sbraf突变体的细胞中观察到细胞形态变化。如通过鬼笔环肽染色所揭示的,丝状肌动蛋白存在于表达野生型braf的细胞的整个细胞体中。然而,肌动蛋白在很大程度上限于表达braf突变体的细胞的外周。此行为与先前对于表达araf突变的细胞的观察结果一致。在表达wt型或突变型braf的细胞中ve-钙粘蛋白染色主要在细胞内。我们先前关于araf的结果表明,即使是wtbraf的表达,也在ea.hy926细胞系中诱导erk活化。尽管wtbraf不会显著改变细胞形态,但这会导致内化的ve-钙粘蛋白。图3突出显示了erk活化对ve-钙粘蛋白定位和细胞形态的影响。表达wt型或突变型braf的细胞的处理导致ve-钙粘蛋白的细胞表面染色显著增加,并且细胞体中的丝状肌动蛋白增加。表达wt型与突变型的细胞之间的形态学差异在本实验中并不那么明显,这可能表示实验与实验之间的变异性,或者是由于与图2相比本实验中培养时间和细胞密度的增加。在表达ptpn11突变体的细胞中未观察到细胞形态变化和ve-钙粘蛋白的分布。
为了鉴定具有更高治疗功效的抑制剂,我们用多种mek抑制剂处理了表达wt型和突变型braf或ptpn11的细胞。参见图4和图5。我们使用曲美替尼作为比较点,已对pd0352971、pimasertib和瑞法替尼建立了滴定曲线。如所期望的,每种抑制剂能够以剂量依赖性方式降低细胞中的perk水平。请注意,在这组实验中,ptpn11的突变体似乎未激活上述erk也未超越用wtptpn11所示的水平。尽管具体的倍数增加因凝胶而异,但braff486s始终诱导erk活化超过wt水平。
淋巴管疾病的斑马鱼模型
使用gateway系统(invitrogen,kwan2007,villefranc200717937395,17948311)将克隆组装在带有侧翼tol2转座酶位点的载体中以整合到基因组中(kawakami1999,10564832)。将大约10pl的25ng/ultol2mrna和载体dna注射到新鲜受精卵的第一个细胞中。将构建体注射到表达mrc1a:gfp的卡斯珀鱼(casperfish)中以使淋巴管可视化。使用zeisslsm710共聚焦显微镜和zen软件进行共聚焦扫描。使用zeisszen软件的最大强度投影功能叠加了图像的共焦z堆栈。图像在imagej(fiji){schindelin2012}和powerpoint(microsoft)中进行编辑。
为了评估突变的人类krasg12d是否可以影响斑马鱼的淋巴管发育,我们在淋巴特异性mrc1a启动子后克隆了突变型和wt型基因(jung2017,pmid:28506987),并将其与具有v2a自催化切割位点的mcherry荧光团连接(provost2007,17941043)。我们使用侧翼的tol2转座酶位点进行基因组整合。当在7dpf(受精后的天数)对表型进行分析时,注射具有tol2mrna的构建体会在大多数被注射的鱼中产生表达转基因和mcherry标志物的淋巴管细胞小斑块。
我们研究了主干胸导管中的表型,这是现阶段最大的血管之一,其显示了转基因克隆的最一致表达。wtkras表达未显示对淋巴管发育的影响,但是krasg12d导致了胸导管扩展和扩张,其似乎与腹主静脉融合。
在其他研究中,我们鉴定了ras/mapk途径中更多的基因/突变,这些基因/突变可以解释淋巴管异常,并为新的治疗干预提供了机会。如表3所示,我们鉴定出在rasa1、raf1、rit1、nf1、cbl1和braf中的生殖系或体细胞突变,它们均涉及ras/mapk信号转导通路,进一步支持了这些疾病实体之间共有的遗传病因学,以及淋巴管异常中ras/mapk通路突变的重要性。此外,在一名淋巴水肿和淋巴传导病症患者中发现了一种纯合的错义变体。因此,我们提供的数据表明,线性ras/mapk途径的治疗性干预可以在斑马鱼中逆转由突变引起的erk1/2活性的升高以及淋巴表型,类似于我们先前对araf突变所示出的。
我们开发了一种细胞模型,以更好地理解和研究潜在的erk激活突变的影响,方法是使用原发性淋巴管内皮细胞代替已建立的细胞系来捕获疾病相关细胞类型的突变特征。我们已经在人类皮肤淋巴管内皮细胞(hdlec)中逆转录表达了野生型和突变型的蛋白质。我们已经成功地使用这些细胞来证明araf基因突变的细胞后果,这种突变导致某种形式的中枢传导淋巴管异常。我们还示出了使用曲美替尼(一种mek抑制剂)可以在细胞中逆转突变araf的作用,这是我们最近在naturemedicine中发表的。此外,我们在该模型中测试了另外7种mek/erk抑制剂,并示出了由araf突变诱导的效应的生化和形态逆转,其与曲马替尼相当,所述7种mek/erk抑制剂包括pd0325901(图6)、ci1040/pd184352(图7)、pimasertib(图8)、tak-733(图9)、azd8330(图10)、瑞法替尼(图11)以及乌利替尼(图12)。
图6-11显示了每种不同药物(如图所示)对表达araf突变s214p的hdlec的作用。所有测试的药物足以逆转由araf-s214p诱导的形态差异,表明更多的ve-钙粘蛋白在细胞膜上积累。绿色表示ha染色,它标记表达araf的细胞;红色表示ve-钙粘蛋白染色。
乌利替尼是erk1/2的口服有效抑制剂。先前的研究表明,在乌利替尼治疗后,多种癌细胞系中的p-erk1/2水平增加。但是,rsk(erk1/2蛋白底物)的磷酸化降低,这与持续的erk1/2抑制作用是一致的。如图12的上图所示,乌利替尼治疗的确减少了erk底物rsk3的磷酸化。在下图中,乌利替尼可以挽救表达araf-s214p的hdlec中细胞-细胞连接处ve-钙粘蛋白的损失,并在300nm的浓度下几乎完全恢复(图12:绿色表示ha染色,它标记表达araf的细胞;红色表示ve-钙粘蛋白染色;白色表示细胞核的dapi染色)。类似地,在hdlec中表达braf-f486s会导致相邻细胞之间ve-钙粘蛋白积累的明显缺乏(图13-14),这基本上产生了与araf突变相似的表型。不出所料,乌利替尼(图13)或曲美替尼(图14)治疗可以挽救该表型,而且曲美替尼(图14上图)可以抑制erk磷酸化升高。此外,表达raf1或kras突变的hdlec表现出相似的形态变化和相似的p-erk升高,这可以通过曲美替尼和乌利替尼进行标准化(图15-16)。另一方面,rit1突变诱导了p-erk的弱激活(图17)。
我们利用以hdlec进行的球状发芽试验(3d淋巴管生成试验)来深入了解突变对淋巴管生成的影响。与表达ephb4-wt的hdlec相比,表达ephb4-ins、ephb4-r763q或ephb4-k885的hdlec表现出增强的淋巴管生成能力,如在具有或没有肝配蛋白(ephrin)b2的血管内皮生长因子c(vegfc)存在下进行的三维淋巴球体发芽试验中通过发芽长度所测量的(图18,上图和左下图)。雷帕霉素和osi-027(一种有效的和选择性的mtorc1和mtorc2抑制剂)均可挽救突变体中发芽的增加(图18,右下图)。
在斑马鱼模型中,kras-g12d突变体(而非野生型kras)在淋巴管中的过度表达导致胸导管扩张和明显的水肿。我们进一步研究了mtor或mek/erk抑制剂的任一种是否可以挽救所述表型。与雷帕霉素、ci1040和sl-327(没有明显作用)不同,使用mek抑制剂的治疗可显著改善表型。具体而言,用考比替尼、pimasertib、tak-733、azd8330和pd0325901进行治疗可导致水肿减少和改善淋巴支化紊乱(图19)。此外,用nvp-bez235(pi3k和mtor的双重抑制剂)进行治疗,显示水肿幼体显著减少(图19)。
我们还在斑马鱼模型中过表达ptpn11突变。图20示出了ptpn11s502a或g503r突变的嵌合表达导致轻度淋巴管异常表型。存在淋巴组织的某些扩张和误导,或后主静脉和胸导管的融合。
斑马鱼中有两种rasa1同源物,rasa1a和rasa1b。我们设计了靶向rasa1a和rasa1b基因两者的grna,并将其注入cas9转基因胚胎中,这导致了大水肿的形成(图21)。
实施例i的参考文献
1.alitalok,tammelat,petrovatv2005lymphangiogenesisindevelopmentandhumandisease.nature438:946-953
2.trenorcc,3rd,chaudryg2014complexlymphaticanomalies.seminpediatrsurg23:186-190
3.levinec1989primarydisordersofthelymphaticvessels--aunifiedconcept.jpediatrsurg24:233-240
4.wassefm,bleif,adamsd,alomaria,baselgae,berensteina,burrowsp,friedenij,garzonmc,lopez-gutierrezjc,lorddj,mitchels,powellj,prendivillej,vikkulam2015vascularanomaliesclassification:recommendationsfromtheinternationalsocietyforthestudyofvascularanomalies.pediatrics136:e203-214
5.hilliardri,mckendryjb,phillipsmj1990congenitalabnormalitiesofthelymphaticsystem:anewclinicalclassification.pediatrics86:988-994
6.smeltzerdm,sticklergb,flemingre1986primarylymphaticdysplasiainchildren:chylothorax,chylousascites,andgeneralizedlymphaticdysplasia.eurjpediatr145:286-292
7.fauljl,berrygj,colbytv,ruosssj,waltermb,rosengd,raffinta2000thoraciclymphangiomas,lymphangiectasis,lymphangiomatosis,andlymphaticdysplasiasyndrome.amjrespircritcaremed161:1037-1046
8.brouillardp,boonl,vikkulam2014geneticsoflymphaticanomalies.jclininvest124:898-904
9.luksvl,kamitakin,viveromp,ullerw,rabr,boveejv,rialonkl,guevaracj,alomariai,greeneak,fishmansj,kozakewichhp,maclellanra,mullikenjb,rahbarr,spencersa,trenorcc,3rd,uptonj,zurakowskid,perkinsja,kirsha,bennettjt,dobynswb,kurekkc,warmanml,mccarrollsa,murillor2015lymphaticandothervascularmalformative/overgrowthdisordersarecausedbysomaticmutationsinpik3ca.jpediatr166:1048-1054e1041-1045
10.kurekkc,luksvl,ayturkum,alomariai,fishmansj,spencersa,mullikenjb,bowenme,yamamotogl,kozakewichhp,warmanml2012somaticmosaicactivatingmutationsinpik3cacauseclovessyndrome.amjhumgenet90:1108-1115
11.lindhurstmj,sappjc,teerjk,johnstonjj,finnem,petersk,turnerj,cannonsjl,bickd,blakemorel,blumhorstc,brockmannk,calderp,chermann,deardorffma,evermandb,golasg,greensteinrm,katobm,keppler-noreuilkm,kuznetsovsa,miyamotort,newmank,ngd,o'brienk,rothenbergs,schwartzentruberdj,singhalv,tiraboscor,uptonj,wientroubs,zackaieh,hoagk,whitewood-nealt,robeypg,schwartzbergpl,darlingtn,tosill,mullikinjc,bieseckerlg2011amosaicactivatingmutationinakt1associatedwiththeproteussyndrome.nengljmed365:611-619
12.revencun,boonlm,mendolaa,cordiscomr,duboisj,clapuytp,hammerf,amordj,irvinead,baselgae,dompmartina,syeds,martin-santiagoa,adesl,collinsf,smithj,sandaraduras,barriovr,burrowspe,bleif,cozzolinom,brunetti-pierrin,vicentea,abramowiczm,desirj,vilainc,chungwk,wilsona,gardinerca,dwighty,lorddj,fishmanl,cytrynbaumc,chamlins,ghalif,gilabertey,josss,boentemdelc,leaute-labrezec,delruema,baylisss,martorelll,gonzalez-ensenatma,mazereeuw-hautierj,o'donnellb,bessisd,pyeritzre,salhia,tanot,wargono,mullikenjb,vikkulam2013rasa1mutationsandassociatedphenotypesin68familieswithcapillarymalformation-arteriovenousmalformation.hummutat34:1632-1641
13.burrowspe,gonzalez-garayml,rasmussenjc,aldrichmb,guilliodr,mausea,fifece,kwons,lapinskipe,kingpd,sevick-muracaem2013lymphaticabnormalitiesareassociatedwithrasa1genemutationsinmouseandman.procnatlacadsciusa110:8621-8626
14.loif,brewerc,shannonn,shortoj,tangb,blackg,soomt,ngdk,lamst,kerrb2008severeneonatalmanifestationsofcostellosyndrome.jmedgenet45:167-171
15.fabrettoa,kutschek,harmsenmb,demarinis,gasparinip,fertzmc,zenkerm2010twocasesofnoonansyndromewithsevererespiratoryandgastroenteralinvolvementandthesos1mutationf623i.eurjmedgenet53:322-32416.joyces,gordonk,briceg,ostergaardp,nagarajar,shortj,moores,mortimerp,mansours2016thelymphaticphenotypeinnoonanandcardiofaciocutaneoussyndrome.eurjhumgenet24:690-696
17.morcaldig,bellinit,rossic,maghniem,boccardof,boniolie,bellinic2015lymphodysplasiaandkrasmutation:acasereportandliteraturereview.lymphology48:121-127
18.makinent,adamsrh,baileyj,luq,ziemieckia,alitalok,kleinr,wilkinsonga2005pdzinteractionsiteinephrinb2isrequiredfortheremodelingoflymphaticvasculature.genesdev19:397-410
19.kumet2010specificationofarterial,venous,andlymphaticendothelialcellsduringembryonicdevelopment.histolhistopathol25:637-646
20.hashimotot,tsunekim,fostertr,santanajm,baih,wangm,huh,hanischjj,dardika2016membrane-mediatedregulationofvascularidentity.birthdefectsrescembryotoday108:65-84
21.martin-almedinas,martinez-corrali,holdhusr,vicentea,fotioue,lins,petersenk,simpsonma,hoischena,gilissenc,jefferyh,attong,karapouliouc,briceg,gordonk,wisemanjw,wedinm,rocksonsg,jefferys,mortimerps,snydermp,berlands,mansours,makinent,ostergaardp2016ephb4kinase-inactivatingmutationscauseautosomaldominantlymphatic-relatedhydropsfetalis.jclininvest126:3080-3088
22.kettleboroughrn,busch-nentwichem,harveysa,dooleycm,debruijne,vaneedenf,sealyi,whiterj,herdc,nijmanij,fenyesf,mehrokes,scahillc,gibbonsr,walin,carrutherss,halla,yenj,cuppene,stempledl2013asystematicgenome-wideanalysisofzebrafishprotein-codinggenefunction.nature496:494-497
23.suns,chens,liuf,wuh,mchughj,berginil,guptaa,adamsd,guanjl2015constitutiveactivationofmtorc1inendothelialcellsleadstothedevelopmentandprogressionoflymphangiosarcomathroughvegfautocrinesignaling.cancercell28:758-772.
实施例ii
araf复发突变导致可通过mek抑制剂治疗的中枢传导淋巴管异常
尽管最近的研究表明西罗莫司在广泛性淋巴管异常(gla)和中枢传导淋巴管异常(ccla)的治疗中有益3–5,但由于它们的稀有性和诊断标准的重叠,这些实体之间缺乏明确的临床区别,这阻碍了创新疗法的发展6–9。gla被定义为多灶性淋巴管异常,其具有多个微/大囊性淋巴畸形的区域,并通常涉及骨破坏9–11。另一方面,ccla描述了胸导管(td)或乳糜池的功能障碍,这导致淋巴液逆行流动或淋巴液引流异常1,12,13。两种病况均可表现为乳糜胸、积液、乳糜性腹水或淋巴水肿。这些明显不同的疾病的重叠表明,多种临床症状是由共同途径而非共同基因引起的,并意味着实体之间的区分可能是人为的。在这里,我们报道了使用全外显子组测序(wes)来鉴定araf中的复发性错义突变,其在两名无关患者中是以复杂淋巴管异常为特征的严重晚期淋巴疾病的基础。我们的结果提供了一个有代表性的证明,说明遗传分类如何提供一种对复杂医学疾病进行分类的方法,从而指导基于生物学的医学治疗,在我们看来,这是挽救生命的。
提供以下材料和方法以帮助进行实施例ii。
患者
获得费城儿童医院(chop)的机构审查委员会的批准并获得知情同意书后,从先证者(p1)和他的父母那里获得了血液样本用于测序分析。所述先证者在其胸部、心包、腹部、下肢和生殖器中有严重的淋巴液积聚,并在chop的淋巴影像和干预中心跟进和治疗。通过淋巴管瘤病和gorham疾病联盟(lgda)的患者登记处招募了一名不相关的第二成年患者(p2),以及可用的家庭成员。除了腹部左侧的毛细血管畸形外,p1的出生和家族史无异常,并且他的童年生长和发育历程是正常的。10岁时,他出现了下腹部、大腿、阴囊和阴茎肿胀。两个月后,他因呼吸急促和运动不耐症到当地一家医院就诊。胸部x光片显示心脏肥大,超声心动图显示大的心包积液。心包穿刺术是在引流1升乳糜性液体的情况下进行的。尽管实行了全胃肠外营养,引流仍在继续,他被转移到chop进行进一步治疗。在chop,他的初步评估包括动态对比增强磁共振淋巴管造影照片,其显示出大的心包积液和在扩张的腰椎和腹膜后网络中的顺行血流,进入扩张且弯曲的td,所述td向左侧的无名静脉移动(图1a、c、d)。未受影响的人的图像在图1b中示出作为参考。此外,淋巴液逆行流入肝脏、肠系膜、阴茎和阴囊,并且从远端td逆行流入纵隔和心包(图1c–e)。他在远端td接受了支架植入和碘油栓塞,目的是停止异常的纵隔和心包淋巴积液。他在一个月后因心包积液稳定出院,但此后不久因大量液体再积聚而出现呼吸窘迫,这使得对补充氧气的需求增加(至多5升通过鼻插管)。他开始使用西罗莫司,并根据每天2.5mg的耐受性良好的谷值水平进行给药,这导致2016年5月至2016年11月的中谷水平为11.8(范围6.8–16.2μgdl-1)。在1.5年的时间里,他经历了多次经皮的干预和外科淋巴手术,包括反复的胸腔穿刺术和胸膜引流、多次经皮淋巴栓塞、两次双侧外科胸膜固定术、大腿、腹部和腹膜后腔外科淋巴静脉吻合术,以及由于阴茎和阴囊水肿恶化,外科手术结扎和腹股沟淋巴管栓塞术。尽管多次尝试控制他的心包积液,但他的阴茎、阴囊、下肢和下腹淋巴水肿恶化,他的病况持续恶化,以至于讨论了姑息治疗的考虑。在使用曲美替尼前五个月进行了最后一个手术,并停用了西罗莫司。
患者p2是一名无关的成年女性,在31岁时被诊断患有淋巴管瘤病。她在诊断前多年有广泛的症状,伴有明显的肺部受累,在胸膜固定术前需要多次胸膜穿刺术。她胃肠道广泛受累,需要专门的脂肪限制性饮食和中链甘油三酯油补充以及间歇性全胃肠外营养。持续出现无法解释的症状后,她接受了计算机断层扫描和磁共振成像,这些症状与影响其肾脏、肝脏、脾脏和肺部的淋巴管瘤病一致。
肝活检证实了淋巴管瘤病的诊断。她还接受了沙丁胺醇和利尿剂的治疗,并由于疲劳和呼吸困难使用了机动踏板车(motorizedscooter)。从未证实她有骨受累。由于该患者是从淋巴管瘤病和gorham疾病联盟招募的,而且不是本地人,因此她失去了随访机会,无法进行其他疗法的试验,因为她死于与她的潜在的淋巴疾病相关的并发症。
wes和生物信息学分析。
我们检查了与外显子组数据中的显性或隐性遗传模型匹配的错义、无义、剪接改变和编码插入缺失标记。过滤结果以排除具有以下因素的变体:同义变体;已知假基因的变体;在1000基因组项目中的或在国家心肺血液研究所外显子组测序项目(esp6500si)的6503个外显子组中的次要等位基因频率(maf)大于0.5%的变体;通过我们内部的外显子组变体数据库先前在对照中鉴定的变体。通过参考在线人类孟德尔遗传数据库,在有害预测和生物学相关性的基础上进行随后的基因优先排序。
araf突变在哺乳动物细胞系中的表达和表征。
hek293t和hela细胞得自美国典型培养物保藏中心,并在杜氏改良的eagle培养基(添加10%胎牛血清)中于37℃下生长。从promocell获得原代成人hdlec,并根据制造商的说明在内皮细胞生长培养基mv2(promocell)中进行培养。从原始载体扩增从addgene获得的全长arafcdna(质粒编号23725)41,并将其作为bamhi/xhoi片段克隆到pcdna3.1载体中,所述载体包含flag标签(dykddddk)的两个拷贝,然后是两个strep标签(wshpqfek)。按照制造商的说明,使用neb的q5诱变试剂盒通过定点诱变引入s214p突变。根据制造商的方案,使用fugenehd(promega),以及3μgdna(空载体,wtaraf(araf-wt)或araf突变体(araf-s214p))和9μl转染试剂在hek293t和hela中进行转染,转染后36–48小时,将细胞用冰冷的磷酸盐缓冲盐水(pbs)洗涤两次,并使用新鲜制备的冰冷细胞裂解缓冲液在冰上裂解,所述裂解缓冲液含有50mmtris-hcl、ph为7.4、100mmnacl、50mmβ-甘油磷酸酯、10%甘油(w/v)、1%np-40(w/v)、1mmedta、2mmnavo4和完整的不含edta的蛋白酶抑制剂混合物(rocheappliedscience)(20μl/毫升裂解缓冲液)。通过离心清除细胞裂解物后,收集上清液,用于western印迹,或使用anti-flagm2亲和凝胶(目录号a2220,sigma)进行免疫沉淀,然后进行western印迹。免疫沉淀物和裂解物在nupage4–12%bis-tris凝胶(thermofisherscientific)上电泳,并用包括以下抗体的一抗进行印迹:抗磷酸化p70s6k-thr389(目录号9205s,cellsignalingtechnology;1:1000)、抗磷酸化mtorser2448(目录号5536p,cellsignalingtechnology;1:1000)、抗flag(目录号f3165,sigma;1:4000)、抗磷酸化p38thr180/tyr182(目录号4511,cellsignalingtechnology;1:1000)、抗pan-14-3-3(目录号sc-629,santacruzbiotechnology;1:500)、抗磷酸化akt-ser473(目录号4060,cellsignalingtechnology;1:1000)、抗磷酸44/42-(erk1/2)-thr202/tyr204(目录号4376,cellsignalingtechnology;1:1000)或抗β肌动蛋白(目录号sc-69879,santacruzbiotechnology;1:1000)抗体。
用bamhi/xhoi切割araf序列(如前所述,来自pcdna3.1-f2s2-araf-wt或-s214p构建体),并将其引入包含氨基末端flag和ha标签的pmscv质粒的修饰形式的bglii/xhoi位点。使用fugene,以及8μg的总dna(pmscv-araf-wt或-s214p以及包膜和包装质粒)和18μl的转染试剂,在hek293t中进行病毒生产。72小时后,收集病毒上清液并过滤。通过用病毒上清液(补充有8μgml-1聚凝胺)替换细胞培养基,感染hdlec,然后通过0.45μm过滤器过滤。将细胞以650g离心90分钟,然后培养6小时,此时将所述病毒上清液替换为标准培养基。将转导的hdlec培养48小时,然后用于实验。通过ha染色观察到的转导效率在40%至60%之间。
hdlec的免疫荧光染色和western印迹。
将圆形(12mm)盖玻片(vwr)在24孔板(corning)中用0.1%明胶水溶液涂覆10分钟,然后风干15分钟。在存在或不存在曲美替尼的情况下,将转导的hdlec以每孔100,000个细胞的浓度在0.5ml培养基中铺板48h。将细胞在温暖的无血清杜氏改良eagle培养基中洗涤,并在4%多聚甲醛中固定。将固定的细胞用pbs洗涤两次,并用在pbs中的0.1%bsa洗涤两次。通过与pbs中的10%正常驴血清(jacksonimmunoresearch)和0.3%tritonx-100(sigmaaldrich)一起培养来渗透和封闭细胞。将ve-钙粘蛋白抗体(thermofisherscientific)在pbs中的0.01%正常驴血清、0.1%bsa和0.3%tritonx-100中稀释(终浓度:2μgml-1),并染色1小时。将盖玻片用pbs中的0.1%bsa洗涤两次。将山羊抗兔alexa546(thermofisherscientific;终浓度:8μgml-1)和鬼笔环肽(thermofisherscientific;终浓度:5单位ml-1)在pbs中的0.01%正常驴血清、0.1%bsa和0.3%tritonx-100中稀释,并染色1小时。使用时,将ha标签(6e2)小鼠抗体(目录号2367,cellsignalingtechnology)在pbs中的0.1%bsa和0.3%tritonx-100中以1:100稀释,然后染色1小时。将盖玻片用pbs中的0.1%bsa洗涤两次并用pbs洗涤两次。将盖玻片浸入水中以去除残留的盐,并使用prolonggold抗淬灭试剂(thermofisherscientific)固定在载玻片上。使用lasaf软件(leicamicrosystems),在带有photometricshq2高分辨率单色ccd(电荷耦合器件)相机的leicadm6000电动立式显微镜上进行图像采集。z堆栈以10倍的放大倍数获取。在fiji软件包中对图像进行了进一步处理42。进行了亮度和对比度调整。相同的亮度和对比度设置应用于所有图像。在绘制为包含整个个体细胞的目标区域(roi)或绘制为包含整个细胞体但不包括细胞与细胞连接的roi中测量荧光值。从这些测量值中,推导出质膜的值(总细胞-细胞内),并推导并绘制质膜与细胞内值的比率。此外,使用线条工具和roi管理器测量细胞的长度和宽度。对于这两种分析,每×10视野分析了5个通过ha染色确定的明显表达araf的细胞。每个实验每个条件采集5个×10视野。对于每种条件下总共75个细胞,使用来自3个独立解冻和hdlec的转导的细胞进行了实验。为了用曲美替尼对hdlec进行western印迹,在存在越来越多的曲美替尼的情况下,将20000个转导的hdlec细胞接种到96孔板中。在药物存在下将细胞培养24小时,然后用40mmhepesph7.5、120mmnacl、0.3%chaps、50mmnaf、1.5mmnavo3和蛋白酶抑制剂混合物裂解。通过在4℃下以20000g离心5分钟来清除裂解物。将蛋白质在4–12%nupagebis-tris凝胶上分离。使用上述抗体进行印迹。
三维淋巴球体发芽试验
通过将7,500个表达araf-wt或araf-s214p的hdlec接种到预先用1.5%琼脂糖包被的96孔板的孔中,启动用于淋巴发芽试验的多细胞球体。在这些条件下,所有hdlec都将在24小时内聚集为单个球体。形成后,将每个球状体转移到由指定的浓度的i型胶原蛋白(目录号354236,corning;最终浓度=1.5mgml-1;ph用naoh中和)和曲美替尼组成的凝胶溶液中,然后使其在37℃下聚合。固化后,将含有适当浓度曲美替尼的内皮细胞生长培养基mv2(不含vegfc)添加到胶原蛋白凝胶上。培养2天后,使用evosfl自动成像系统(thermofisherscientific)对嵌入的球体进行拍摄,得到步长约为8.5μm的z堆栈图像。使用imagej软件测量从每个球状体长出的毛细血管状芽的数量和长度(https://imagej.nih.gov/ij/)。
用转导的hdlec进行mtt增殖试验。
使用rocheappliedscience的细胞增殖试剂盒i(mtt)测量了转导的hdlec的增殖。简而言之,在逆转录病毒转导后第2天,收集表达araf-wt-和-s214p的hdlec,计数并以每孔10,000个细胞的浓度在100μl培养基中重新接种到平底96孔板中。在接种后的指定时间,将10μlmtt加入适当的孔中,并在37℃下孵育4小时。加入100μl体积的增溶试剂,随后在37℃下孵育过夜。在spectramaxi3多模式读板器(moleculardevices)上在550nm和700nm处测量吸光度,并计算出a550nm–a700nm。将接种后4小时的时间点包括在内,作为以最小增殖加载到实验中的细胞的近似量度。
人类araf在斑马鱼中的转基因表达。
所有使用斑马鱼的程序均已获得chop机构动物护理和使用委员会(iac001154)的批准,并符合美国国立卫生研究院《实验室动物的护理和使用指南》的规定。将人类突变型和wt型arafcdna在没有终止密码子的情况下克隆到pdonr221载体中;使用了适合斑马鱼的kozak序列(gcaaacatgg)43。使用包括gateway克隆盒的tol2主链载体组装表达构建体(44、45)。将构建体与tol2信使rna共注射(46)。
使用斑马鱼mrc1a启动子在血管和淋巴管中表达araf,并通过mcherry可视化表达,所述mcherry通过自动催化v2a蛋白切割位点连接到araf。对于成像,将幼体固定在低熔点琼脂糖中,并使用zeisslsm710共焦显微镜使用×20镜头拍摄多个z图像。使用zeisszen软件的最大强度投影功能叠加了图像的共焦z堆栈。
为了分析td的扩张,选择了td中具有转基因表达的、由节间淋巴管分隔的身体部分。形态学分为正常(wt)、中度扩张(td扩展但与pcv分开)或重度扩张(td和pcv在z投影中无法区分)。图像在imagej(fiji)中编辑。每个实验进行3次,总共分析了40只动物。
斑马鱼中的抑制性药物处理。
药物处理在6孔板中进行,每组多达20个幼体。将考比替尼在含有0.01mtrisph7.2和0.1%dmso的胚胎培养基中稀释。使用的考比替尼的浓度为1μm。
斑马鱼中的p-erk抗体染色。
如上所述对鱼进行注射,并选择具有显著wt型或突变型araf/mcherry表达的幼体进行分析。将幼体在含有tween-20的pbs(pbst)中的4%多聚甲醛溶液中固定过夜。将幼体用pbst洗涤,并在2%tritonx-100中于4℃孵育24小时。然后,将幼体在10%的牛血清中封闭,并在4℃下用磷酸化erkt202/y204抗体(目录号9101,cellsignalingtechnology,1:200)染色过夜,用pbst洗涤并用alexafluor488山羊抗兔二抗(目录号a11008,thermofisherscientific,1:400)染色。
统计。
对于所有基于细胞的试验,均通过不成对的双尾学生t检验评估其显著性,用于两组的比较。使用graphpadprism7.0d软件进行统计分析。如所示,数据以箱线图表示,其中框的范围从25%到75%,晶须从最小到最大,并且以中位数为中心,或以带有条形图的点状图表示,表示平均值±s.e.m.。对于在hdlec上进行的所有测定,除了未进行统计学分析的增殖研究外,均使用hdlec的独立转导进行了三个独立的实验。对于14-3-3蛋白结合试验,用hek293t细胞的独立转染进行了三个独立的实验,而hek293t细胞的其他结果则代表了六个独立的实验。所有与斑马鱼相关的试验都是在三个独立的实验中进行的,并通过不成对的单尾学生t检验进行了检验,以比较两组。
结果
对已知的淋巴管异常相关基因的第一层wes分析未披露,所述分析包括akt1、pik3ca、kras、hras、nras、braf、raf1、ptpn11、shoc2、cbl、rit1和sos1的突变分析。随后的基因优先排序在两名患者中均揭示了新的x染色体araf突变,c.640t>c:p.s214p,其中所述患者p1为男性,其患有ccla(图22a,22c-22e;有关详细的临床说明,请参见“方法”;genbank登录号ng_016339),患者p2为女性,其先前被诊断患有淋巴管瘤病。所述突变影响保守的磷酸化位点,推测导致功能获得(gof)效应,因为残基ser214是其同源蛋白c-raf(也称为raf1)中的旁系同源调节位点,可被14-3-3蛋白抑制。该错义突变在1000基因组项目、esp6500si、exacv0.3、gnomadv2.1或我们在内部数据库中拥有的5,000多个样本中的其他外显子组测序数据中不存在。如在男性患者中所示,对来自p1和父母双方的血液dna进行的sanger测序证实,这种x连锁的araf突变是作为体细胞杂合事件发生的(图22f)。对p2中的araf突变进行sanger测序,她未受影响的女儿和母亲证实了所述突变仅在p2中存在(图22f)。对父亲无法进行测序;但是,由于她的父亲没有被报道有呼吸道症状,因此很可能所述araf突变是p2中的新生突变或体细胞突变引起的。患者p2失去了随访,而后来我们被告知她随后在诊断后五年死于她的淋巴疾病的并发症。
ser214残基,其是araf中保守区2(cr2)(14)中14-3-3结合位点之一,在整个脊椎动物物种以及raf蛋白中都是高度保守的,这表明它可能在这些激酶的功能中起重要作用(图22g)。14-3-3蛋白与araf的磷酸化ser214的结合将阻止激活的ras将araf蛋白招募到质膜上(15)。先前的研究表明,c-raf中的araf-s214旁系同源的残基ser259的突变会损害14-3-3蛋白的结合,这导致质膜定位并诱导erk/mek信号转导(16)。如图23a所示,与表达野生型(wt)araf的hek293t细胞相比,用araf-s214p转染的hek293t细胞显示14-3-3蛋白的免疫共沉淀减少,并且进而erk1/2的活化显著增强,如通过增加的磷酸化来衡量的(图23a、23b)。arat-s214p不会改变akt、p70s6k、mtor和p38(另一个map激酶家族)的磷酸化(图23b)。在hela细胞和原代人类皮肤淋巴管内皮细胞(hdlec)中获得了相似的结果(图23c)。即使在不存在细胞因子或生长因子的情况下,这种明显的过度活化也存在。
与表达araf-wt的hdlec相比,表达araf-s214p的hdlec表现出增强的淋巴管生成能力,如通过在不存在血管内皮生长因子c(vegfc)的情况下进行的三维淋巴球体发芽试验中的发芽数量和发芽长度所衡量的(图23d)。mek抑制剂曲美替尼挽救了突变体中发芽的增加(图23d)。然后,我们对表达araf-s214p的原代hdlec的内皮粘附连接处进行了形态学分析。如免疫荧光显微镜检查所示,araf-s214p表达引起相邻细胞之间ve-钙粘蛋白积累的大量缺乏,表明ve-钙粘蛋白的内化增加(图23e,黄色箭头)。此外,araf-s214p的表达改变了肌动蛋白的组织,其中表达突变型的细胞在细胞体内具有较少的离散f-肌动蛋白丝。
然后,我们检查了mek1/2抑制剂逆转这些异常的能力。mek抑制剂曲美替尼以100nm的浓度,挽救了在表达araf-s214p的hdlec中观察到的细胞间连接中ve-钙粘蛋白的损失,几乎完全恢复了细胞单层完整性,并恢复了在连接处和肌动蛋白丝处的ve-钙粘蛋白的正常外观(图23e)。尽管araf-s214p明显激活了hdlec中的erk,并且erk激活通常与细胞增殖有关,但我们在两次独立的逆转录病毒转导中,都没有观察到表达araf-wt-和-s214p的hdlec之间在增殖方面存在任何可测量的差异(图23f)。
在tg(mrc1a:egfp)y251转基因株系17中进行了斑马鱼淋巴发育的分析,其中所有淋巴管内皮细胞都标记有egfp。使用mrc1a启动子将araf表达靶向淋巴管,并通过mcherry表达标记表达araf的细胞。araf-s214p表达诱导不同位置的淋巴管扩张,最一致的是,我们观察到躯干td扩张(图23g)。相比之下,araf-wt的表达对淋巴形态无影响(图23h)。araf-s214p的表达诱导斑马鱼中的p-erk。
为了确定mek信号转导抑制剂是否可以逆转异常,当淋巴祖细胞发芽形成td(17)时,我们在受精后3天(dpf)用考比替尼处理了mrc1a:arafs214p幼体。我们分析了在7dpf时td中具有araf表达的身体部分(体节),并发现通过考比替尼可以显著挽救导管形态(图23i、23j)。同时,我们用考比替尼处理了wttg(mrc1a:egfp)y251幼体,并发现它们对药物的耐受性良好。
鉴于我们的研究表明,araf突变在对西罗莫司无反应的p1中导致了功能增加作用,并且mek抑制剂可以挽救转导的内皮细胞和转基因斑马鱼模型中的淋巴表型,我们寻求机构审查委员会的批准,以便在p1中使用mek抑制剂疗法。曲美替尼(mekinist)是美国食品药品监督管理局(fda)批准的mek抑制剂,经过全面的基线评估后,随后在该12岁患者中进行适应症外(off-label)使用。
我们使用了1mgd-1的曲美替尼起始剂量,并在治疗后2个月内开始观察到肺功能测试的改善(图24a)。此外,治疗三个月后,淋巴液滞留和补充氧气需求量显著减少,并且随着身体活动水平的提高,他能够适应室内空气,并且没有观察到曲马替尼的任何不良事件。在治疗12个月时,他的肺功能测试表明他的总肺容量(tlc)几乎翻了一番,并且他在1s内的用力呼气量(fev1)从23%提高到预计的42%(图24a)。电解质(低na和k)正常化和他的磁共振成像扫描显示他的淋巴系统的淋巴重塑和重构(图24b-24f)、肺总容量(tlc)翻倍的显著恢复以及他在1s内的用力呼气量(fev1)从23%提高到预计的42%(图24a)。电解质(低na和k)正常化和他的磁共振成像扫描显示他的淋巴系统的淋巴重塑和重构(图24b-24f),在开始这种基因指导治疗之前经常住院的个体的显著恢复(图24f)。
总之,我们对两名无关的淋巴管异常患者进行了wes,并鉴定了araf基因中的反复发生的功能获得性突变,包括一名对西罗莫司治疗无反应的患有晚期淋巴疾病的12岁男性。用突变araf转导的hdlec表现出升高的erk1/2活性,增强的淋巴管生成能力以及肌动蛋白骨架和ve-钙粘蛋白连接的分解,这可使用mek抑制剂曲美替尼进行挽救。在不存在vegfc(有效的淋巴管生成因子)的情况下,在表达araf-s214p的hdlecs中观察到发芽(18)。在相同条件下,表达araf-wt的细胞中没有发芽。这表明araf突变体正在模仿vegfc的刺激行为或通过hdlec诱导vegfc的表达,这是内皮细胞发芽所必需的,如在许多基质细胞类型中所见(19-21)。我们在斑马鱼模型中复制了由araf的gof突变造成的异常淋巴表型,在所述斑马鱼模型中可以观察到使用mek抑制剂治疗挽救了所述表型。值得注意的是,使用曲马替尼对具有araf突变的先证者进行治疗,导致显著改善了患者的症状、重塑了其扩张且疼痛的淋巴管、消除了淋巴水肿,并在治疗的12个月内恢复了日常活动。
从正在进行的患者招募中,我们研究了其他淋巴管异常患者,包括患有noonan(或与noonan相关的)综合征、gorham–stout病、kaposi型淋巴管瘤病(kla)、淋巴管扩张和ccla的患者。在对另外43位患者进行测序时,我们在kras、braf、rasa1、ptpn11和sos1中鉴定出了7个其他突变(表3),这表明ras-mapk信号转导是导致各种淋巴疾病临床表现的共同通路。的确,人们越来越认识到,ras-mapk通路在淋巴管生成的信号转导中起着关键作用(21-23)。回顾文献,我们鉴定了50多名患者,所述患者具有kras、hras、braf、raf1、ptpn11、shoc2、cbl、rit1和sos1中的突变,并且表现出noonan或noonan样综合征与淋巴缺陷的临床特征,所述临床特征包括胸膜积液、心包积液、乳糜胸、积水、淋巴管扩张和淋巴水肿(21-23)。在我们的研究进行期间,一个复发的nras变体涉及到gla(37)和kla(38),这进一步支持了这些疾病实体之间共有的遗传病因学和淋巴管异常中ras-mapk通路突变的重要性。
数十年来,已经认识到ras病中的突变在人类癌症中非常普遍。我们使用cbioportal(39)数据库(n=71,857个受试者,于2019年2月6日查询)仔细检查了我们发现的araf突变,发现有2名患者的araf具有相同的确切突变。有趣的是,他们都具有并发的tp53突变,其被认为是致癌的驱动因素。在十名患有不同类型癌症的患者中也观察到了这个残基的不同突变(s214t、s214a、s214y、s214c和s214f),其中三个已显示导致mek/erk磷酸化升高(40)。但是,十分之九的患者在tp53、gnas、akt2、apc、egfr、atm、chek2、kit或u2af1中同时发生致癌突变,从而增加了这些致癌驱动因素可能导致癌细胞过度增殖的可能性。具有araf突变的先证者具有扩张的淋巴管,但在多年的随访中,病变的大小未见增加。因此,这些数据与我们的观察结果一致,即我们发现的araf突变可能不会在体外驱动淋巴管内皮细胞增殖的增加。
关于突变阳性淋巴管异常的患病率,在美国的11个中心中形成了淋巴管异常联盟,以促进该组淋巴管异常的多中心临床试验,所述淋巴管异常包括但不限于gla、gorham–stout病、ccla、kla、klippel–trenaunay综合征和kaposi型血管内皮瘤,招募了3000多名患有中度至重度疾病病程的患者,每年新增患者的总数约为300。根据目前的分子诊断率(20%),我们预计他们中约20%在ras-mapk通路中存在缺陷,这表明美国数千名患者可能会受益于mek抑制剂治疗。因此,我们的工作例证了遗传发现如何影响疾病分类并发现新的生物学和拯救生命的治疗方法,如本文患有先前未知病因的淋巴管异常的患者所示,这是一种精准医疗方法的实现。
参考文献
1.trenor,c.c.3rd&chaudry,g.complexlymphaticanomalies.semin.pediatr.surg.23,186–190(2014).
2.collins,f.s.&varmus,h.anewinitiativeonprecisionmedicine.n.engl.j.med.372,793–795(2015).
3.adams,d.m.等人efficacyandsafetyofsirolimusinthetreatmentofcomplicatedvascularanomalies.pediatrics137,e20153257(2016).
4.hammill,a.m.等人sirolimusforthetreatmentofcomplicatedvascularanomaliesinchildren.pediatr.bloodcancer57,1018–1024(2011).
5.mccormick,a.,rosenberg,s.,trier,k.&balest,a.acaseofacentralconductinglymphaticanomalyresponsivetosirolimus.pediatrics137,e20152694(2016).
6.hilliard,r.i.,mckendry,j.b.&phillips,m.j.congenitalabnormalitiesofthelymphaticsystem:anewclinicalclassification.pediatrics86,988–994(1990).7.levine,c.primarydisordersofthelymphaticvessels—aunifiedconcept.j.pediatr.surg.24,233–240(1989).
8.smeltzer,d.m.,stickler,g.b.&fleming,r.e.primarylymphaticdysplasiainchildren:chylothorax,chylousascites,andgeneralizedlymphaticdysplasia.eur.j.pediatr.145,286–292(1986).
9.wassef,m.等人vascularanomaliesclassification:recommendationsfromtheinternationalsocietyforthestudyofvascularanomalies.pediatrics136,e203–e214(2015).
10.chen,w.,adams,d.,patel,m.,gupta,a.&dasgupta,r.generalizedlymphaticmalformationwithchylothorax:long-termmanagementofahighlymorbidconditioninapediatricpatient.j.pediatr.surg.48,e9–e12(2013).
11.lala,s.等人gorham–stoutdiseaseandgeneralizedlymphaticanomaly—clinical,radiologic,andhistologicdifferentiation.skeletalradiol.42,917–924(2013).12.clemens,r.k.,pfammatter,t.,meier,t.o.,alomari,a.i.&amann-vesti,b.r.combinedandcomplexvascularmalformations.vasa44,92–105(2015).
13.li,d.等人pathogenicvariantinephb4resultsincentralconductinglymphaticanomaly.hum.mol.genet.27,3233–3245(2018).
14.wellbrock,c.,karasarides,m.&marais,r.therafproteinstakecentrestage.nat.rev.mol.cellbiol.5,875–885(2004).
15.lavoie,h.&therrien,m.regulationofrafproteinkinasesinerksignalling.nat.rev.mol.cellbiol.16,281–298(2015).
16.molzan,m.等人impairedbindingof14-3-3toc-rafinnoonansyndromesuggestsnewapproachesindiseaseswithincreasedrassignaling.mol.cellbiol.30,4698–4711(2010).
17.jung,h.m.等人developmentofthelarvallymphaticsysteminzebrafish.development144,2070–2081(2017).
18.karkkainen,m.j.等人vascularendothelialgrowthfactorcisrequiredforsproutingofthefirstlymphaticvesselsfromembryonicveins.nat.immunol.5,74–80(2004).
19.carmeliet,p.&jain,r.k.molecularmechanismsandclinicalapplicationsofangiogenesis.nature473,298–307(2011).
20.karaman,s.,leppanen,v.m.&alitalo,k.vascularendothelialgrowthfactorsignalingindevelopmentanddisease.development145,dev151019(2018).
21.potente,m.&makinen,t.vascularheterogeneityandspecializationindevelopmentanddisease.nat.rev.mol.cellbiol.18,477–494(2017).
22.coso,s.,bovay,e.&petrova,t.v.pressingtherightbuttons:signalinginlymphangiogenesis.blood123,2614–2624(2014).
23.brouillard,p.,boon,l.&vikkula,m.geneticsoflymphaticanomalies.j.clin.invest.124,898–904(2014).
24.bulow,l.等人hydrops,fetalpleuraleffusionsandchylothoraxinthreepatientswithcblmutations.am.j.med.genet.a167a,394–399(2015).
25.gargano,g.等人hydropsfetalisinapretermnewbornheterozygousforthec.4a>gshoc2mutation.am.j.med.genet.a164a,1015–1020(2014).
26.gos,m.等人contributionofrit1mutationstothepathogenesisofnoonansyndrome:fournewcasesandfurtherevidenceofheterogeneity.am.j.med.genet.a164a,2310–2316(2014).
27.hanson,h.l.等人germlinecblmutationassociatedwithanoonan-likesyndromewithprimarylymphedemaandteratomaassociatedwithacquireduniparentalisodisomyofchromosome11q23.am.j.med.genet.a164a,1003–1009(2014).
28.milosavljevic,d.等人twocasesofrit1associatednoonansyndrome:furtherdelineationoftheclinicalphenotypeandreviewoftheliterature.am.j.med.genet.a170,1874–1880(2016).
29.koenighofer,m.等人mutationsinrit1causenoonansyndrome-additionalfunctionalevidenceandexpandingtheclinicalphenotype.clin.genet.89,359–366(2016).
30.lee,k.a.等人ptpn11analysisfortheprenataldiagnosisofnoonansyndromeinfetuseswithabnormalultrasoundfindings.clin.genet.75,190–194(2009).
31.croonen,e.a.等人prenataldiagnostictestingofthenoonansyndromegenesinfetuseswithabnormalultrasoundfindings.eur.j.hum.genet.21,936–942(2013).
32.joyce,s.等人thelymphaticphenotypeinnoonanandcardiofaciocutaneoussyndrome.eur.j.hum.genet.24,690–696(2016).
33.yaoita,m.等人spectrumofmutationsandgenotype–phenotypeanalysisinnoonansyndromepatientswithrit1mutations.hum.genet.135,209–222(2016).
34.lo,i.f.等人severeneonatalmanifestationsofcostellosyndrome.j.med.genet.45,167–171(2008).
35.ebrahimi-fakhari,d.等人congenitalchylothoraxastheinitialpresentationofptpn11-associatednoonansyndrome.j.pediatr.185,248–248.e1(2017).
36.morcaldi,g.等人lymphodysplasiaandkrasmutation:acasereportandliteraturereview.lymphology48,121–127(2015).
37.manevitz-mendelson,e.等人somaticnrasmutationinpatientwithgeneralizedlymphaticanomaly.angiogenesis21,287–298(2018).
38.barclays.f.等人asomaticactivatingnrasvariantassociatedwithkaposiformlymphangiomatosis.genet.med.https://doi.org/10.1038/s41436-018-0390-0(2018).
39.gao,j.等人integrativeanalysisofcomplexcancergenomicsandclinicalprofilesusingthecbioportal.sci.signal.6,pl1(2013).
40.imielinski,m.等人oncogenicandsorafenib-sensitivearafmutationsinlungadenocarcinoma.j.clin.invest.124,1582–1586(2014).
尽管上面已经描述并具体举例说明了本发明的某些优选实施方案,但是这并不意味着本发明限于这些实施方案。如所附权利要求书所述,在不脱离本发明的范围和精神的情况下可以对其进行多种修改。
- 还没有人留言评论。精彩留言会获得点赞!