增强抗凋亡途径以预防DOX毒性的方法与流程
本申请涉及增强抗凋亡途径以预防dox毒性的方法。
背景技术:
蒽环类药物,包括阿霉素(dox),被广泛用作有效的化学治疗剂,但会引起严重的剂量依赖性心脏毒性。蒽环类药物引起的心力衰竭很大程度上归因于心肌细胞凋亡。
gata4是gata转录因子家族的成员,其具有结合共有的dna基序“gata”的能力,在心肌分化和功能中起着至关重要的作用。gata4激活抗凋亡基因bcl-2和bcl-xl等的转录,从而防止dox诱导的心肌细胞死亡。在dox处理后,gata4在转录本和蛋白质水平上均迅速下调。有趣的是,gata4的过度表达会导致心脏肥大。这些发现表明,gata4可能还进行更多层次的微调,这需要在应用gata4修复作为防止dox引起的心脏毒性的临床策略之前进行进一步的检查。
sirt6属于高度保守的nad+依赖性沉默调节蛋白家族,其使组蛋白和非组蛋白底物去乙酰化,从而调节染色质稳定性并限制转录。通过这些功能,sirt6可以维持机体健康,并防止衰老和包括癌症和代谢紊乱在内的各种疾病。sirt6通过使h3k9去乙酰化以抑制igf-akt和nf-κb信号传导而参与预防心肌肥大和心力衰竭。心脏sirt6对应激刺激,即血管紧张素ii、异丙肾上腺素和缺血/再灌注诱导的活性氧(ros)和dox,敏感。怀孕期间进行运动可保护新生儿心肌细胞免受dox毒性,并伴有sirt6表达的增加。尽管取得了这些进展,但sirt6如何保护心肌细胞免受dox的影响尚不清楚。
技术方案
鉴于上述技术问题,本申请的目的之一是提供用于促进哺乳动物细胞中的抗凋亡途径的方法,其防止dox诱导的毒性,从而增强dox化学疗法的安全性。
为了实现上述目的,根据本申请的一实施例,提供了一种通过增强转录因子gata4的乙酰化来促进哺乳动物细胞中抗凋亡途径的方法。该方法包括:使所述哺乳动物细胞与有效增加gata4乙酰化的作用剂接触。所述作用剂被用于增强gata4在其羧基末端的锌指区域的第328以及330位的赖氨酸残基(k328/330)的乙酰化。
根据本申请的另一实施例,提供了调节gata4乙酰化的作用剂在制备用于治疗其中促进gata4乙酰化是有益的疾病或病况的药物中的用途。所述作用剂用于增强gata4在其羧基末端的锌指区域的k328以及k330的乙酰化。
根据本申请的又一实施例,提供了一种抑制哺乳动物细胞中sirt6(具有由seqidno:4表示的氨基酸序列)的脱乙酰酶活性的方法。该方法包括:使哺乳动物细胞与有效抑制sirt6的脱乙酰酶活性的量的作用剂接触。所述作用剂为包括由seqidno:2表示的氨基酸序列的肽段,或者其类似物。
根据本申请的又一实施例,提供了抑制sirt6的脱乙酰基酶活性的作用剂在制备用于治疗其中抑制sirt6的脱乙酰基酶活性是有益的疾病或病况的药物中的用途。所述作用剂是包括由seqidno:2表示的氨基酸序列的肽段,或其类似物。
根据本申请的又一实施例,提供了一种筛选抑制sirt6的脱乙酰酶活性的作用剂的方法。所述方法包括:a)使候选分子与测试样品中表达sirt6的细胞接触;b)确定所述测试样品中的脱乙酰酶活性;如果所述候选分子抑所述制测试样品中sirt6的脱乙酰酶活性水平,则选择所述候选分子作为抑制sirt6的脱乙酰酶活性的作用剂。候选分子是包含由seqidno:2表示的氨基酸序列的肽或其类似物。
有益效果
根据本申请的实施方案的用于在哺乳动物细胞中促进抗凋亡途径的方法能够预防dox诱导的毒性,从而增强dox化学疗法的安全性。从原理上讲,sirt6募集tip60乙酰转移酶以在k328/330处乙酰化gata4,从而增强其与染色质结合能力。反过来,gata4抑制sirt6的脱乙酰基酶活性,从而通过tip60促进的h3k9乙酰化确保局部染色质的开放性,并促进抗凋亡基因的转录。重要的是,抗癌化学疗法阿霉素(dox)的治疗损害了sirt6-tip60-gata4三聚体,阻断了gata4的乙酰化作用,并导致心肌细胞凋亡。虽然gata4超乙酰化模拟物保留了对dox的保护作用,但低乙酰化模拟物却失去了这种能力。因此,本申请基于新的sirt6-tip60-gata4轴,该轴促进了抗凋亡途径以防止dox毒性,并靶向三聚体,以提高dox化疗的安全性。
附图说明
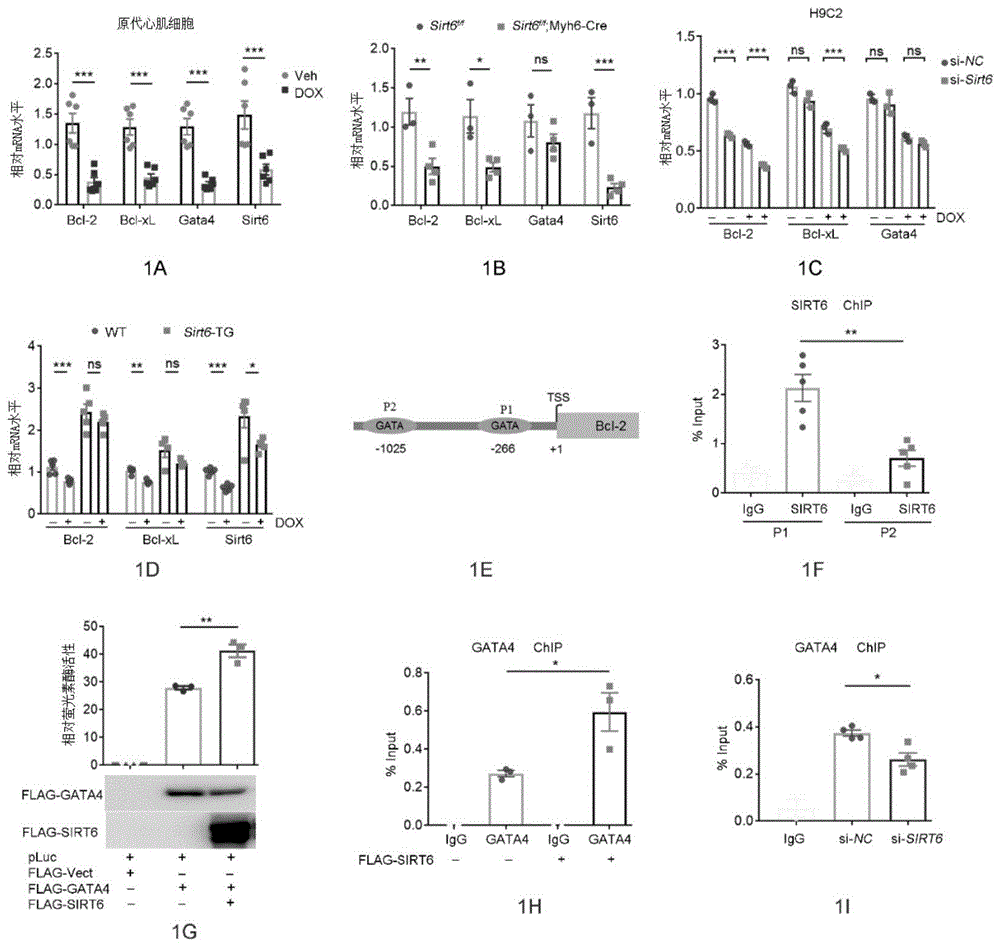
图1a-1i显示sirt6通过gata4拮抗dox诱导的心肌细胞凋亡途径。(图1a)定量pcr(qpcr),显示在dox(1μm)处理6小时后,原代小鼠心肌细胞中bcl-2、bcl-xl、gata4和sirt6的mrna水平。***p<0.001。(图1b)qpcr结果显示了从用他莫昔芬(tamoxifen)治疗的sirt6f/f和sirt6f/f;myh6-cre分离的心脏组织中bcl-2、bcl-xl、sirt6和gata4的mrna水平。*p<0.05,**p<0.01,***p<0.001,”ns”表示无意义。(图1c)将具有si-nc(阴性对照)或si-sirt6小干扰(si)rna的h9c2细胞暴露于dox(1μm,6h)。通过定量pcr测定bcl-2、bcl-xl和gata4的mrna水平。***p<0.001,“ns”表示无意义。(图1d)qpcr结果显示在腹膜内注射dox后,野生型(wt)和sirt6转基因(sirt6-tg)小鼠中bcl-2、bcl-xl和sirt6的mrna水平(n=5/组)。*p<0.05,**p<0.01,***p<0.001,“ns”表示无意义。(图1e)在bcl-2启动子区域上的gata共有基序的示意图。p1和p2分别表示–266gata基序和–1025gata基序。tss代表bcl-2基因的转录起始位点。(图1f)使用抗sirt6抗体和igg进行chip。通过qpcr用靶向p1和p2基序的引物分析bcl-2启动子区域上sirt6的富集。**p<0.01。(图1g)用所示质粒转染hek293细胞,并测量荧光素酶活性并将其标准化为β-gal活性。**p<0.01。蛋白质印迹表明flag-gata4和flag-sirt6的表达水平。(图1h)在具有或不具有flag-sirt6过表达的hek293细胞中使用抗gata4抗体进行chip以测试bcl-2启动子上的gata4富集。*p<0.05。(图1i)在具有si-nc或si-sirt6的hek293细胞中使用抗gata4抗体进行chip以测试bcl-2启动子上的gata4富集。*p<0.05。
图2a-2g显示sirt6与gata4物理相互作用。(图2a)在用所示构建体转染的hek293细胞中进行了使用抗ha琼脂糖的共免疫沉淀。在纯化的ha-sirt6库中通过蛋白质印迹法检测了flag-gata4的水平。(图2b)在用所示构建体转染的hek293细胞中进行了使用抗flag琼脂糖的共免疫沉淀。ha-sirt6的水平通过蛋白印迹法在纯化的flag-gata4库中进行检测。(图2c-2d)通过共免疫沉淀(co-ip)并用抗gata4和抗sirt6抗体进行免疫染色来评估h9c2细胞中gata4和sirt6的内源性相互作用。比例尺,50μm。(图2e)进行gst拉下实验法以测试纯化的his-gata4和来自大肠杆菌的gst融合的sirt6的体外结合。考马斯亮蓝染色指示gst和gst-sirt6的负载水平。(图2f)具有一个或两个锌指dna结合结构域的全长(f-l)和片段化的人gata4的图。(图2g)在hek293细胞中ha-gata4的基于域的截短与flag-sirt6共表达。沉淀ha-gata4蛋白,并通过蛋白质印迹分析flag-sirt6。
图3a-3f显示sirt6增强gata4乙酰化并保护dox诱导的心肌细胞凋亡,而与其脱酰基酶活性无关。(图3a)蛋白质印迹显示了从过量表达flag-sirt6和ha-gata4的hek293细胞中免疫沉淀的gata4的乙酰化水平。用抗乙酰赖氨酸(ack)抗体测定乙酰化水平。(图3a’)相对于仅ha-gata4对照,gata4的乙酰化水平的倍数变化,由imagej确定。n=6。***p<0.001。(图3b)将共表达flag-sirt6和ha-gata4的hek293细胞用10μm烟酰胺(nam)处理4小时。免疫沉淀后,使用抗ack抗体分析gata4的乙酰化水平。(图3b’)相对于仅ha-gata4对照,gata4的乙酰化水平的倍数变化,由imagej确定。n=3。***p<0.001。“ns”表示没有意义。(图3c)用抗ha珠粒免疫沉淀分别在hek293细胞中与不同形式的flag-sirt6(wt、d63y、d116n和h133y突变体)共表达的ha-gata4。ips中的flag-sirt6印迹表明gata4和sirt6的关联。(图3c’)相对于仅ha-gata4对照,gata4的乙酰化水平的倍数变化,由imagej确定。n=3。**p<0.01,*p<0.05,”ns”表示没有意义。(图3d)wt、重新表达flag-sirt6的sirt6–/–mefs或酶失活突变体中bcl-2的mrna水平的实时pcr分析。**p<0.01;***p<0.001。18srrna用作参考基因。(图3e-3f)代表性的流程图和定量显示了在荧光激活的细胞分选测定法中,用dox处理(1μm,12小时)过表达指示基因的凋亡h9c2细胞的百分比。*p<0.05;**p<0.01;***p<0.001。
图4a-4h显示sirt6募集tip60用于gata4在赖氨酸328/330处的乙酰化。(图4a)抗ha免疫沉淀物的免疫印迹显示ha-gata4在过量表达的异位ha-gata4的hek293细胞中的乙酰化水平,用指定的组蛋白乙酰转移酶(hat)抑制剂(即cpth2(20μm)、mg149(20μm)和c646(10μm),持续6小时。(图4a’)相对于仅ha-gata4对照,ha-gata4的乙酰化水平的倍数变化,由imagej确定。n=3。***p<0.001。“ns”表示没有意义。(图4b)抗ha免疫沉淀物的免疫印迹显示ha-gata4在过量表达ha-gata4以及flag-tip60或空载体ha-gata4蛋白的hek293细胞(从si-nc-或si-sirt6处理的共表达ha-gata4的hek293细胞纯化)中的乙酰化水平。gata4的乙酰化状态通过抗ack抗体的蛋白质印迹法确定。(图4b’)相对于仅ha-gata4对照,ha-gata4的乙酰化水平的倍数变化,由imagej确定。n=4。***p<0.001。(图4c)蛋白质印迹显示来自原代小鼠心肌细胞的抗gata4ip中内源性tip60和sirt6的水平。igg用作抗体对照。(图4d)在野生型(wt)或sirt6敲除(ko)hek293细胞中,在有或没有ha-gata4的情况下共表达flag-tip60。进行了ha-gata4的co-ip分析,并评估了flag-tip60和gata4乙酰化的水平。(图4d’)相对于wthek293细胞对照,与ha-gata4结合的flag-tip60蛋白的倍数变化。n=4。**p<0.01。(图4e)将截短的ha-gata4构建体与或不与flag-sirt6一起共转染。用所示抗体进行蛋白质印迹。(图4e')相对于仅ha-gata4(f-l)对照的ha-gata4乙酰化水平的倍数变化,由imagej确定。n=3。***p<0.001。“ns”表示没有意义。(图4f)在小鼠、大鼠和非洲爪蟾中人gata4的c末端锌指结构域及其直向同源物的蛋白质序列比对。突出显示了该区域内所有被乙酰化的k残基。(图4g)将wt和位点突变的(k328/330r)ha-gata4的构建体与flag-tip60共表达在hek293细胞中。测定了gata4的乙酰化水平。(图4g’)相对于仅ha-gata4wt对照,ha-gata4乙酰化水平的倍数变化,由imagej确定。n=4。***p<0.001。“ns”表示没有意义。(图4h)蛋白质印迹显示了在用flag-tip60和ha-sirt6或空载体共转染的hek293细胞中ha-sirt6蛋白和tip60乙酰化的水平。(图4h’)相对于仅flag-tip60对照,flag-tip60乙酰化水平的倍数变化,由imagej确定。n=3,“ns”表示无意义。
图5a-5h显示gata4的低乙酰化减弱了针对dox的心脏保护。(图5a)用0、1、2μmdox进行6小时处理后,ha-gata4沉淀池中flag-sirt6蛋白和ha-gata4乙酰化水平的蛋白质印迹分析。(图5a’)相对于溶媒/无flag-sirt6对照的乙酰化gata4的百分比水平。(图5a')相对于空白对照,绑定到ha-gata4的flag-sirt6的百分比(右)。定量由imagej进行。n=3。*p<0.05。**p<0.01。(图5b)暴露于dox(1μm,6h)后,过表达gata4wt、k328/330r(2kr)和k328/330q(2kq)的原代小鼠心肌细胞中bcl-2mrna水平的qpcr分析。***p<0.001,”ns”表示无意义。(图5c)用dox(1μm)处理24小时后,tunel阳性染色的原代心肌细胞百分比。**p<0.01,***p<0.001,”ns”表示无意义。(图5d)将稳定表达gata4wt、k328/330r(2kr)和k328/330q(2kq)的h9c2细胞暴露于dox(1μm,6h)。通过qpcr评估bcl-2的mrna转录物。**p<0.01,”ns”表示无意义。18srrna用作参考基因。(图5e)在存在/不存在dox的情况下,通过使用抗gata4抗体在稳定转染的h9c2细胞中进行chip分析。通过qpcr测量在bcl-2启动子区域上的gata4富集。***p<0.001,**p<0.01,”ns”表示无意义。(图5f-5g)将稳定转染的h9c2细胞暴露于dox(1μm)24小时,并用tunel或tritc-鬼笔环肽染色。捕获染色细胞的图像。tunel染色阳性的h9c2细胞数量的量化(图5f)。测量并定量细胞面积(图5g)。比例尺,20μm。***p<0.001,”ns”表示无意义。(图5h-5h’)显示了指定剂量的dox1小时后h9c2细胞的存活分数的细胞集落形成。将板(n=3/组)用结晶紫染色并计数菌落数。**p<0.01,***p<0.001,”ns”表示无意义。
图6a-6i显示sirt6-tip60-gata4轴有助于染色质的开放性。(图6a-6b)在具有或不具有flag-tip60的过表达的wt和sirt6kohek293细胞中,使用抗tip60或抗h3k9ac抗体进行chip测定。通过qpcr分析tip60和h3k9ac在bcl-2启动子上的富集。**p<0.01,***p<0.001。(图6c)在具有或不具有flag-sirt6过表达的hek293细胞中,使用抗h3k9ac和h3k56ac抗体进行chip分析,显示出h3k9ac和h3k56ac在bcl-2启动子区域的p1基序上富集。”ns”表示没有意义。(图6d-6f)通过使用抗h3k9ac和带有si-nc或si-gata4的hek293细胞中的抗体,进行chip分析以测试bcl-2启动子区域上h3k9ac、sirt6和tip60的富集。*p<0.05。”ns”表示没有意义。(图6g)在单独表达ha-gata4或与野生型flag-sirt6或flag-sirt6h133y突变体表达ha-gata4的hek293细胞中进行使用抗flag琼脂糖的co-ip。通过蛋白质印迹分析了flag-sirt6沉淀物中的ha-gata4水平。(图6h)在体外实验中与或不与纯化的his-gata4一起预温育后,重组sirt6的脱乙酰基活性。***p<0.001。(图6i)在体外实验中与指定剂量的g4-13肽(0、0.2、2、20、200μm)预孵育后,重组sirt6wt和sirt6h133y的脱乙酰基活性。***p<0.001。”ns”表示没有意义。
图7显示了示意性模型:sirt6-tip60-gata4轴将基因转录和表观遗传激活耦合在一起。sirt6最初通过核小体结合特性占据启动子区域,并募集tip60来平衡局部组蛋白的动态乙酰化。当gata4通过识别gata序列而同时相互作用并阻断sirt6脱乙酰基酶活性时,tip60将gata4乙酰化以增强其转录活性。sirt6的抑制作用会破坏局部组蛋白乙酰化平衡,从而确保转录的开放染色质结构。
发明的实施方式
为了进一步说明本申请,下面描述了详述增强抗凋亡途径以防止dox毒性的方法的实验。应当注意,以下实施例旨在描述而非限制本申请。
gata4的乙酰化的增强
根据一实施例,本申请提供了一种通过增强转录因子gata4的乙酰化来促进哺乳动物细胞中抗凋亡途径的方法。该方法包括:使哺乳动物细胞与有效增加gata4乙酰化的作用剂接触。所述作用剂用于增强gata4在羧基末端锌指区域内的k328和k330处的乙酰化。
在一实施例中,所述作用剂是乙酰转移酶或其类似物。
在一实施例中,所述作用剂是tip60或被用于激活所述哺乳动物细胞内的tip60的激活剂。
在一实施例中,tip60包含由seqidno:1表示的氨基酸序列或其类似物。
在一实施例中,该作用剂是一种被用于使所述哺乳动物细胞过表达sirt6或其类似物的作用剂。sirt6及其类似物能够在所述哺乳动物细胞中募集tip60以对gata4在k328和k330处进行乙酰化。
sirt6脱乙酰酶活性的抑制
在一实施例中,本申请提供了一种在哺乳动物细胞中抑制sirt6的脱乙酰基酶活性的方法,该方法包括:使所述哺乳动物细胞与有效抑制sirt6的脱乙酰基酶活性的量的作用剂接触,其中,所述作用剂是包含由seqidno:2表示的氨基酸序列的肽段,或其类似物。
在一实施例中,所述作用剂是包括gata4的羧基末端的锌指区域(a.a.251-350)的肽段或者其类似物,其中,gata4包括由seqidno:3表示的氨基酸序列。
在一实施例中,所述作用剂是gata4或其类似物。
在一实施例中,用于抑制哺乳动物细胞中sirt6的脱乙酰酶活性的方法被应用于促进哺乳动物细胞中的抗凋亡途径或抑制肿瘤细胞的存活。
药物筛选测定法
在一实施例中,本申请涉及用于筛选抑制sirt6的脱乙酰酶活性的治疗剂的方法。治疗剂可以是药物、化学物质、化合物、蛋白质或肽,或核酸分子(例如dna、rna,例如sirna)。
在一实施例中,本申请提供了一种筛选抑制sirt6的脱乙酰基酶活性的作用剂的方法,包括:a)使候选分子与测试样品中表达sirt6的细胞接触;b)确定测试样品中的脱乙酰酶活性;如果候选分子抑制测试样品中sirt6的脱乙酰酶活性水平,则选择候选分子作为抑制sirt6的脱乙酰酶活性的作用剂。
在抑制sirt6的脱乙酰基酶活性的作用剂的筛选测定法的一实施例中,候选分子是包含由seqidno:2表示的氨基酸序列的肽段或其类似物。在另一实施例中,候选分子是包括gata4的羧基末端的锌指区域(a.a.251-350)的肽段或者其类似物。在另一实施例中,候选分子是gata4或其类似物。
在抑制sirt6的脱乙酰基酶活性的作用剂的筛选测定的一实施例中,候选分子可以选自增加gata4活性的作用剂以及增加gata4表达的作用剂,诸如增强gata4的转录、翻译和/或处理的作用剂。
脱乙酰活性可以通过以下方法测定:包括但不限于共免疫沉淀、蛋白质印迹、elisa、免疫荧光、放射免疫测定、免疫细胞化学及其组合。
疾病治疗
在一实施例中,本申请提供了一种用于治疗其中促进gata4乙酰化是有益的疾病或病况的方法,所述方法包括给予需要所述治疗的患者或受试者有效量的作用剂,以增强gata4在羧基末端的锌指区域内的k328和k330处的乙酰化。在一实施例中,所述作用剂是tip60或被用于激活哺乳动物细胞内的tip60的激活剂。在一实施例中,该作用剂是一种被用于使哺乳动物细胞过表达sirt6或其类似物的作用剂。sirt6和sirt6及其类似物能够在哺乳动物细胞中募集tip60以在gata4的k328和k330处进行乙酰化。
在各种实施例中,其中促进gata4乙酰化将是有益的并且可以根据本申请进行治疗的疾病或病况包括但不限于dox诱导的心脏毒性或其他相关疾病。
在一实施例中,本申请提供了一种用于治疗其中对sirt6的脱乙酰酶活性的抑制是有益的疾病或病况的方法,包括向需要这种治疗的患者或受试者施用有效量的包含seqidno:2表示的氨基酸序列的肽段,或其类似物。在一实施例中,该作用剂是包括gata4的羧基末端的锌指区域(a.a.251-350)的肽段或者其类似物。在一实施例中,所述作用剂是gata4或其类似物。在一实施例中,其中抑制sirt6的脱乙酰基酶活性将是有益的疾病或病况包括但不限于dox诱导的心脏毒性、癌症或其他相关疾病。
如本文所用,术语“受试者”或“患者”描述了可以向其提供用根据本申请的组合物进行治疗的生物,包括哺乳动物,例如灵长类。可以从所公开的治疗方法中受益的哺乳动物物种包括但不限于猿、黑猩猩、猩猩、人、猴子;驯养动物,例如狗和猫;家畜,例如马、牛、猪、绵羊、山羊和鸡等;以及其他动物,例如小鼠、大鼠、豚鼠和仓鼠。
本文所用的术语“治疗”或其任何语法变化(例如,treat、treating和treatment等)包括但不限于减轻疾病或病况的症状;和/或减少、压制、抑制、减缓或影响疾病或病况的进展、严重程度和/或范围。如本文所用,术语“有效量”是指能够治疗、预防或改善疾病或病况或能够产生预期治疗效果的量。
治疗组合物和制剂
在一实施例中,本申请提供了调节gata4乙酰化的作用剂在制备用于治疗其中促进gata4乙酰化是有益的疾病或病况的药物中的用途。所述作用剂用于增强gata4在其羧基末端的锌指区域的k328以及k330的乙酰化。在一实施例中,所述作用剂是tip60或被用于激活哺乳动物细胞内的tip60的激活剂。所述作用剂是用于使哺乳动物细胞过表达sirt6或其类似物的作用剂;sirt6和sirt6及其类似物能够在哺乳动物细胞中募集tip60以在gata4的k328和k330处进行乙酰化。在一实施例中,要治疗的疾病或病况与dox诱导的心脏毒性有关。
在一实施例中,本申请提供了抑制sirt6的脱乙酰基酶活性的作用剂在制备用于治疗其中抑制sirt6的脱乙酰基酶活性是有益的疾病或病况的药物中的用途。所述作用剂是包括由seqidno:2表示的氨基酸序列的肽段,或其类似物。在一实施例中,该作用剂是包括gata4的羧基末端的锌指区域(a.a.251-350)的肽段或者其类似物。在一实施例中,所述作用剂是gata4或其类似物。在一实施例中,待治疗的疾病或病况与dox诱导的心脏毒性或癌症有关。
本申请还提供了治疗或药物组合物,其包括可以与药学上可接受的载体组合的形式的本申请的化合物。在这种情况下,该化合物可以是例如分离的或基本上纯的。
术语“载体”是指与化合物一起施用的稀释剂、佐剂、赋形剂或溶媒。这样的药物载体可以是无菌液体,例如水和油,包括石油(例如矿物油)、植物油(例如花生油、大豆油和芝麻油)、动物油或合成来源的油的那些。盐溶液以及葡萄糖水溶液和甘油溶液也可以用作液体载体,特别是用于注射溶液。用于治疗或减轻中枢神经系统中的炎症的特别优选的药物载体是可以穿透血/脑屏障的载体。如本文所用,载体不包括天然存在的天然植物材料。
合适的药物赋形剂包括淀粉、葡萄糖、乳糖、蔗糖、明胶、麦芽、大米、面粉、白垩、硅胶、硬脂酸钠、单硬脂酸甘油酯、滑石粉、氯化钠、脱脂奶粉、甘油、丙烯、乙二醇、水、乙醇和类似。如果需要,治疗组合物还可包含少量的湿润剂或乳化剂或ph缓冲剂。这些组合物可以采取溶液剂、混悬剂、乳剂、片剂、胶囊剂、散剂、缓释制剂等形式。该组合物可以与传统的粘合剂和载体(例如甘油三酸酯)一起配制。合适的药物载体的实例在e.w.martin的“remington'spharmaceuticalsciences”中有所描述。这样的组合物包含治疗有效量的治疗组合物以及适当量的载体,以提供用于向患者适当给药的形式。该制剂应适合于给药方式。
在一实施例中,本申请提供了适于向人类局部注射给药的药物组合物。通常,用于局部注射给药的组合物是在无菌等渗水性缓冲液中的溶液。必要时,该组合物还可包含增溶剂和局部麻醉剂,例如利多卡因,以减轻注射部位的疼痛。通常,成分以单位剂型单独或混合在一起提供,例如,作为干燥的冻干粉末或无水浓缩物,放在指示活性剂含量的密闭容器(例如安瓿或小药囊)中。在通过注射施用组合物的情况下,可以提供安瓿瓶的无菌注射用水或盐水,从而可以在施用之前将成分混合。
本申请的治疗或药物组合物可以配制成中性或盐形式。药学上可接受的盐包括与游离氨基形成的盐,例如来源于盐酸、磷酸、乙酸、草酸、酒石酸等的盐,以及与游离羧基形成的盐,例如源自钠、钾、铵、钙、异丙胺、三乙胺、2-乙基氨基乙醇、组氨酸、普鲁卡因等的盐,
本申请还提供了一种药物包装或药盒,其包括一个或多个装有本申请的药物组合物的一种或多种成分(例如化合物、载体)的容器。
给药途径
可以通过标准途径将本发明的化合物和组合物施用于正在治疗的受试者,包括口服、吸入或肠胃外给药,包括静脉内、皮下、局部、经皮、皮内、跨粘膜、腹膜内、肌内、囊内、眼眶内、心内、气管内、皮下、皮下、表皮内、关节内、囊下、蛛网膜下、脊柱内、硬膜外和胸骨内注射、输注和电穿孔,以及作为(暂时或永久地)插入受试者中的任何医疗设备或物体的组分共同给药。
在治疗特定疾病、病况或障碍中有效的本申请的治疗或药物组合物的量将取决于给药途径以及疾病、病况或障碍的严重性,并且应根据取决于从业医师和每个患者的情况。通常,剂量为约0.001mg/kg至约3g/kg。例如,合适的单位剂量可以在约0.01至约3g、约0.01至约1g、约0.01至约500mg、约0.01至约400mg、约0.01至约300mg、约0.01至约200mg之间。约0.01至约100mg、约0.01至约50mg、约0.01至约30mg、约0.01至约20mg、约0.01至约10mg、约0.01至约5mg、约0.01至约3mg0.01至约1mg、或约0.01至约0.5mg。这样的单位剂量可以每天多次给药,例如一天两次或三次。
可与载体材料组合以产生单一剂型的活性成分的量将根据病况的类型和待治疗的受试者而变化。一般来说,治疗组合物包含约5%至约95%的活性成分(w/w)。更具体地说,治疗组合物包含约20%(w/w)至约80%或约30%至约70%的活性成分(w/w)。
一旦患者状况得到改善,必要时可给予维持剂量。随后,可以根据症状将剂量或给药频率或两者降低至保持改善的状况的水平。当症状减轻至所需水平时,应停止治疗。但是,一旦疾病况状复发,患者可能需要长期间歇治疗。另外,可任选地采用体外实验法以帮助鉴定最佳剂量范围。制剂中要使用的精确剂量还取决于给药途径以及疾病、病况或障碍的严重性,并且应根据从业医师的判断和每个患者的情况来决定。有效剂量可以从源自体外或动物模型测试系统的剂量反应曲线中推断出来。
材料与方法
细胞培养和处理
hek293(crl-1573)和h9c2(gnr-5)细胞购自atcc。如先前所述,获得野生型(wt)和sirt6-/-小鼠胚胎成纤维细胞(mef)。如先前所述,使用crispr/cas9系统产生sirt6敲除(ko)hek293细胞系。用标准方法制备原代新生小鼠心肌细胞。简而言之,分离1至3日龄的c57bl/6小鼠的心脏,并与消化培养基一起孵育。离心和铺板后,存活的心肌细胞在培养两天内形成同步跳动的单层细胞。所有细胞系都在37℃的5%co2和大气氧气中在dulbecco改良的eagle培养基(dmem,lifetechnologies,美国)中培养,该培养基中添加了15%的胎牛血清、100u/ml青霉素和链霉素。用指定剂量的dox处理细胞以进行特异性分析。
小鼠和dox给药
将sirt6f/f小鼠与myh6-cre/esr1小鼠杂交,以产生sirt6f/f;myh6-cre小鼠。为了产生心脏特异性的sirt6ko小鼠,连续三天每天在sirt6f/f;myh6-cre小鼠的腹膜内(i.p.)注射4-羟基他莫昔芬(4-hydroxytamoxifen)。1个月后,收集心脏组织并通过基于pcr的基因分型和定量pcr(qpcr)进行分析。所有小鼠均在深圳大学实验动物研究中心饲养和处理。所有实验均根据机构动物护理和使用委员会(iacuc)的指导进行。
质粒和rna干扰
人flag-gata4和flag-tip60构建体购自vigenebiosciences(中国)。人flag-sirt1、2、6和7构建体购自addgene(usa)。使用kod-plus试剂盒(日本toboyo),通过基于pcr的诱变产生具有氨基酸取代的flag-gata4和flag-sirt6。基于ha-sirt6和ha-gata4模板,通过基于pcr的缺失构建了截短的sirt6和gata4质粒。
对于rna干扰,根据制造商的说明,使用
细胞转染和集落形成测定
为了建立稳定转染的h9c2细胞系,将pcdna3.1、gata4、gata42kr(k328/330r)和gata42kq(k328/330q)质粒用pvui核酸酶消化30分钟以线性化,然后用
对于集落形成测定,将细胞一式三份接种在6孔板中,并在正常生长条件下在有或没有dox的情况下以所示剂量培养。再培养10-14天后,用0.5%结晶紫溶液将细胞集落染色。定量每个孔中的菌落数量并计算存活分数。
染色质结合级分测定
小心地将细胞从培养容器中的1ml冷pbs缓冲液中取出,然后通过以3,000×g离心1分钟沉淀。用500μl缓冲液a(10mmhepes、10mmkcl、1.5mmmgcl2、0.34m蔗糖、10%甘油、1mmdtt、0.1%tritonx-100、10mmna3vo4和蛋白酶抑制剂混合物)重悬细胞沉淀。并在冰上孵育10分钟。然后将细胞裂解物以1300×g离心5分钟,并收集含有胞浆蛋白的上清液。沉淀用缓冲液a洗涤一次,然后在250μl缓冲液b(3mmedta、0.2mmegta、1mmdtt、10mmna3vo4和蛋白酶抑制剂混合物)中裂解。孵育和离心(1700×g,5分钟)后,获得沉淀(染色质结合级分),并用1×sds上样缓冲液变性。蛋白质提取、蛋白质印迹和抗体
将小鼠组织在1ml冰冷的组织裂解缓冲液(25mmtris-hcl、ph7.5、10mmna3vo4,100mmnaf,50mmna4p2o7、5mmegta、5mmedta、0.5%sds,1%np-40和蛋白酶抑制剂混合物)。超声处理后,将裂解物以16,000×g离心10分钟,然后仔细收集。使用比辛可宁酸(bicinchoninicacid)试剂盒(bca;thermo)测定蛋白质浓度。按照标准方案进行蛋白质印迹。抗乙酰赖氨酸(cst,ac-k-100,#6952)、抗聚/单adp核糖(cst,#83732)和抗tip60(cst,#12058)抗体购自cellsignalingtechnology(美国)。抗gata4(ab84593)、抗sirt6(ab62739)和抗h3(ab1791)抗体购自abcam(英国)。抗p300(sc48343)和抗gst(sc138)抗体购自santacruzbiotechnology(美国)。抗-h3k9ac(abe18)购自emdmillipore(美国),抗flagm2购自sigma-aldrich(美国)。
免疫共沉淀(co-ip)和拉下实验(pull-downassays)
用冷pbs洗涤细胞并收集。然后将细胞沉淀重悬于500μlip裂解缓冲液(20mmtris-hclph8.0、250mmnacl、5m焦磷酸钠、1mm氟化钠、10%甘油、2mmedta、10mmna3vo4、蛋白酶抑制剂混合物以及1mm烟酰胺)。超声和离心后,收集上清液,并在旋转平台上于4℃与抗ha或抗flagm2琼脂糖(sigma,美国)孵育过夜。然后小珠用1ml冷裂解缓冲液洗涤3次。为了变性,将1×sds上样缓冲液添加到样品中,并在98℃下煮沸6分钟。然后通过蛋白质印迹分析样品(包括input(5%))。
对于体外拉下实验,将gst或gst-sirt6(1μg)偶联的gsh珠悬浮在netn缓冲液(20mmtris-hclph8、100mmnacl,1mmedta、0.5%np40和磷酸酶以及蛋白酶抑制剂混合物)。然后加入纯化的his-gata4蛋白(0.2μg),并在4℃下孵育过夜,然后用netn缓冲液洗涤3次。之后将煮沸的洗脱液分离并通过考马斯染色和蛋白质印迹法检测。
萤光素酶报告实验
用pgl3-bcl-2-启动子报告基因,表达质粒和对照pcmxβgal质粒转染hek293细胞。根据制造商的说明,使用荧光素酶报告试剂盒(promega,#e1501)测量荧光素酶活性。荧光素酶值根据β-gal活性标准化。
rna制备和实时qpcr分析
按照苯酚-氯仿提取方法,使用试剂rnaisoplus(takara,日本)从细胞或小鼠组织中提取总rna。使用primescriptrtmastermix(takara,日本)和以下热循环条件,从纯化的rna构建cdna文库:37℃持续40分钟;85℃持续5s。使用cfxconnectedreal-timepcr检测系统(美国bio-rad)和sybrextaqpremixes(日本takara)分析基因表达。每个实验至少进行三次。数值标准化为18srrna。
染色质免疫沉淀(chip)分析
将细胞交联并用裂解缓冲液(50mmtris·hclph8.0、2mmedta,15mmnacl,1%sds、0.5%脱氧胆酸盐、蛋白酶抑制剂混合物和1mmpmsf裂解)。超声和离心后,收集上清液,并用蛋白a/gsepharose预先纯化。将预清除的样品与指定的抗体(抗h3k9ac、抗sirt6、抗gata4抗体(2μg/样品)和适当的对照igg)孵育过夜。通过补充50%的蛋白a/gsepharose悬浮液沉淀结合的dna片段。用一系列缓冲液顺序洗涤后,将珠粒加热至65℃以使交联反向。纯化dna片段并通过qpcr分析。
tunel染色法
根据制造商的说明,使用deadendtmtunel荧光法检测系统(promega,g3250)评估了dox诱导的心肌细胞凋亡。简而言之,将细胞固定并与含有末端脱氧核苷酸转移酶和荧光素-12-dutp的反应混合物在37℃孵育1小时。dapi用于对细胞核进行复染。随机选择五个视野,并在显微镜下捕获图像以计算每个切片的tunel阳性染色率。
荧光激活细胞分选(facs)分析
将h9c2细胞用1μmdox处理12h,然后用fitc-annexinv(beyotime,c1062l)和7-aad(bdpharmingen)染色以区分活/死细胞。使用lsr-ii流式细胞仪进行流式细胞仪分析,并使用flowjox7.5进行分析。
体外sirt6去乙酰化实验
按照制造商的说明,使用sirt6活性测定试剂盒(fluorometric)(ab156068,abcam,uk)进行sirt6去乙酰化实验。简而言之,将细菌纯化的gst-sirt6或gst-sirt6h133y与纯化的his-gata4,具有由seqidno:2表示的氨基酸序列的g4-13肽段或具有由seqidno:5表示的氨基酸序列的g4-24肽段(glbiochemltd,中国上海)在4℃的反应缓冲液中预孵育30分钟。一旦添加荧光团偶联的合成ac-肽,移动板以测量荧光强度。
统计分析
所有值均表示为平均值±s.e.m.。使用student的t检验或使用graphpadprism软件的双向anova确定组之间的统计差异。使用对数秩(mantel-cox)检验分析生存曲线数据。
示例1:sirt6通过gata4增强针对dox的抗凋亡基因表达
为了确定sirt6如何保护细胞免受dox侵害,我们评估了来自两个独立的sirt6基因敲除细胞系的rna-seq数据集:小鼠胚胎干细胞(mescs)和小鼠胚胎成纤维细胞(mefs)。众所周知,sirt6的缺失会激活凋亡途径,并显著降低抗凋亡基因的表达,例如bcl-2和bcl-xl。通过直接靶向抗凋亡基因的启动子区域,gata4转录因子对于出生后抗dox的心肌细胞存活至关重要。实际上,经dox处理后,h9c2细胞和原代小鼠心肌细胞中gata4、bcl-2和bcl-xl的mrna水平都被迅速下调(图1a)。为了进一步了解sirt6和gata4/bcl-2途径之间的因果关系,我们使用他莫昔芬诱导的α-肌球蛋白重链(myh6)cre系,即sirt6f/f;myh6-cre(其会自发性发展心力衰竭),在成年小鼠中产生了心脏特异性sirt6敲除。qpcr分析表明,小鼠心脏中sirt6的缺失显著降低了bcl-2和bcl-xl的水平,而对gata4mrna或蛋白质表达的影响很小(图1b)。通过sirna对sirt6的敲低进一步加剧了dox诱导的h9c2细胞中bcl-2和bcl-xlmrna表达的降低(图1c)。相比之下,sirt6的过表达增强了bcl-2和bcl-xl的表达水平,并保护了小鼠免受dox诱导的下降(图1d)。
然后,我们质疑sirt6是否通过gata4调节抗凋亡途径。为此,我们首先比较了sirt6基因敲除mescs中下调的基因和由mescs中胚层中染色质免疫沉淀和测序(chip-seq)获得的gata4结合的基因。有趣的是,由sirt6缺乏引起的下调基因中几乎有50%带有gata4结合位点,而基因本体分析表明调节细胞凋亡的途径显著富集。因此,我们推测sirt6和gata4可能在调节抗凋亡基因表达中协同作用。如所描述的,gata4通过与近端-266gata基序(p1)而非远端-1025gata基序(p2)结合来调节心脏bcl-2转录。有趣的是,chip分析表明sirt6在p1位点也高度富集(图1e-1f)。然后,我们确定了在含有p1基序的bcl-2启动子的转录控制下的荧光素酶活性。我们发现过表达的sirt6显著提高了bcl-2启动子(p1)驱动的萤光素酶活性(图1g)。此外,sirt6过表达增强了p1位点的gata4占有率(图1h),但是sirt6kd则使这种现象有所折中(图1i)。总之,这些数据表明sirt6增强了gata4染色质结合能力,从而促进了抗凋亡基因的表达。
示例2:gata4与sirt6物理关联
如上所示,sirt6和gata4都在p1启动子区域富集并协调bcl-2的转录,这表明这两种蛋白可能直接相互作用。实际上,我们在表达带有或不带有ha-sirt6的flag-gata4的hek293细胞中的抗ha-sirt6免疫沉淀物中检测到了flag-gata4(图2a)。相应地,ha-sirt6存在于来自表达带有或不带有flag-gata4的ha-sirt6的hek293细胞的flag-gata4的纯化池中(图2b)。为了确认在生理条件下的这种相互作用,我们在h9c2心肌细胞中进行了内源性免疫共沉淀(coip)和免疫荧光显微镜检查。再次,在抗gata4免疫沉淀物中检测到sirt6,并将其与细胞核中的gata共染色(图2c-2d)。接下来,我们使用纯化的表达his-gata4和gst-sirt6的大肠杆菌进行了体外拉下实验。如图所示,结果表明了gata4和sirt6之间的直接相互作用(图2e)。gata4具有两个锌指(zn-finger)域;c末端的锌指对于dna和辅因子的结合是必不可少的(图2f)。域图谱显示sirt6在c末端锌指区(a.a.251-350)处特异性结合gata4(图2g)。
示例3:sirt6促进gata4的乙酰化,与其脱乙酰酶活性无关
接下来我们试图弄清楚sirt6如何调控gata4靶向的基因表达。值得注意的是,翻译后修饰,特别是磷酸化和乙酰化,对gata4dna结合能力和激活至关重要。为了研究sirt6是否调节gata4的乙酰化水平,我们在共过量表达gata4和sirt6的hek293细胞中对flag-gata4进行了免疫共沉淀,并使用泛抗乙酰赖氨酸抗体检查了其乙酰化水平。出乎意料的是,在过sirt6存在下,gata4的乙酰化水平显著增加(图3a-3a’)。在缺失sirt6的细胞中检测到低水平的gata4乙酰化。除脱乙酰基酶活性外,sirt6还具有单adp-核糖基酶(mar)活性,并且可以自身核糖基化(mar)。但是,未观察到gata4的mar,表明gata4不太可能成为sirt6的mar修饰目标。
接下来,我们研究了gata4乙酰化的增加是否需要sirt6的脱乙酰基酶活性。烟酰胺(nam)与邻近nad+的保守口袋结合,以阻断sirt6的脱乙酰基酶和单adp-核糖基酶活性。我们观察到nam消除了沉默调节蛋白(sirtuin)介导的h3k9脱乙酰基作用,但不能抑制sirt6过表达诱导的gata4乙酰化水平升高(图3b-3b')。此外,无催化活性的sirt6突变体,d63y和d116n,仍然能够结合并增强gata4的乙酰化作用(图3c-3c’)。更重要的是,sirt6wt、d63y和d116n的过表达恢复了sirt6缺失的mef中的bcl-2和bcl-xl表达(图3d),并显著抑制了暴露于dox的h9c2细胞的凋亡(图3e-3f)。这些数据因此暗示sirt6促进gata4乙酰化,并因此转录活性独立于其脱乙酰酶活性。
示例4:sirt6诱导的gata4乙酰化需要tip60
sirt6与gata4相互作用并增强其乙酰化水平的数据表明,还涉及其他因素,最可能是组蛋白乙酰转移酶(hat)。为了确定哪个hat负责增强gata4的乙酰化作用,我们研究了一系列hat抑制剂对sirt6调节的gata4乙酰化作用的影响,包括抑制gcn5的cpth2、抑制tip60的mg149以及抑制p300的c646。有趣的是,尽管通过所有hat抑制剂的处理进一步降低了h3k56ac的水平,但是只有mg149处理显著抑制了sirt6诱导的gata4乙酰化的增加(图4a-4a’)。小鼠心脏中特异性的tip60缺失会缩短小鼠的寿命,并伴有增加的心肌细胞密度和凋亡。tip60-p400hat复合物的缺失严重影响了esc中gata4调控的原始内胚层分化。但是,是否gata4是否是tip60的直接目标仍然未知。我们发现,tip60的过表达增加了gata4的乙酰化水平,并且在应用mg149后消除了这种影响。sirna的tip60kd也显著降低了gata4的乙酰化水平。这些结果表明tip60是新型的gata4的乙酰转移酶。
为了确认tip60在sirt6增强的gata4乙酰化中的作用,我们研究了sirt6过表达的tip60kd细胞中gata4的乙酰化水平。我们发现,在tip60kd细胞中,gata4的sirt6增强乙酰化作用被显著消除。换句话说,在sirt6缺失的细胞中tip60的过表达不能完全乙酰化gata4(图4b-4b')。重要的是,内源性sirt6和tip60存在于来自原代小鼠心肌细胞的抗gata4免疫沉淀物中(图4c),这表明这些蛋白质形成了三聚体复合物。此外,sirt6ko细胞中低水平的gata4乙酰化与不可检测的tip60和gata4之间相互作用有关(图4d-4d')。这些数据表明sirt6充当促进tip60介导的gata4乙酰化的骨架。
示例5:tip60在k328/330处乙酰化gata4
如上所示,gata4的c端锌指经常被乙酰化,这就是sirt6的结合位点。如通过抗gata4免疫沉淀中的抗ack和抗组蛋白h3免疫印迹法和染色质分级测定法所确定的,c端锌指结构域的缺失消除了gata4的乙酰化和染色质结合。在该区域内有六个进化保守的赖氨酸(k)残基(图4f)。据报道,p300使gata4在k313/320/322/324处乙酰化,从而增强了dna结合活性,导致应激引起的心脏肥大。但是,通过k-to-r突变,我们注意到k313/320/322/324不太可能被tip60乙酰化。相反,k328/330r突变体(2kr)几乎完全消除了异位sirt6或tip60诱导的gata4乙酰化增加(图4g-4g')。总之,这些数据表明,tip60介导了gata4在k328/330处的乙酰化。
自我乙酰化会激活tip60,其受sirt1的负调控。因此,我们研究了sirt6是否调节tip60的乙酰化状态。如图所示,sirt6的过表达对tip60的乙酰化水平无效用(图4h-4h’)。进一步的co-ip分析表明,wt和酶失活的sirt6均与tip60结合。我们还分析了sirt6过表达时tip60的mar水平,但几乎没有观察到tip60的mar修饰,这表明tip60的核糖基化的可能性较小。数据支持sirt6在gata4乙酰化中的脱乙酰基酶活性非依赖作用和结构作用。
示例6:gata4在k328/330处的乙酰化对于dox抗性至关重要
恢复gata4转录活性已被公认为是预防dox引起的心力衰竭的有效的方法。接下来,我们研究了tip60介导的gata4乙酰化的心脏保护作用。我们共表达了ha-gata4和flag-sirt6,然后用dox处理细胞。重要的是,dox处理破坏了gata4和sirt6的相互作用,并同时以剂量依赖的方式降低了gata4乙酰化的水平(图5a-5a')。在原代心肌细胞中,dox处理显著降低了bcl-2表达,这在很大程度上由gata4wt和k328/330q(2kq),而不是2kr,的过表达所弥补(图5b)。因此,如通过tunel测定所确定的,gata4wt或2kq的过表达能够减轻dox诱导的心肌细胞凋亡,但是2kr突变体却不能(图5c)。我们进一步生成了稳定表达gata4wt、2kr突变体或2kq突变体的h9c2细胞系。尽管异位gata4wt和2kq突变体显著减弱了暴露于dox的h9c2细胞中bcl-2和bcl-xl水平的下降,但是2kr突变体的作用很小(图5d)。chip数据一致性地显示在存在或不存在dox的情况下,2kr突变体,而不是2kq突变体,损害了gata4的dna结合能力(图5e)。gata4的过表达减轻了dox诱导的心肌细胞凋亡和导致的心肌萎缩。然而,该效果被2kr突变所消除(图5f-5g)。进一步的集群形成试验表明,gata42kq突变体在响应dox的情况下促进了与wt相似的心肌细胞存活,而2kr突变体却不能(图5h-5h')。这些数据支持以下观点:tip60介导的乙酰化对于gata4的心脏保护作用至关重要。
示例7:sirt6-tip60-gata4轴表观遗传确保基因表达
sirt6和tip60是h3k9和/或h3k56乙酰化的主要hdac和hat,它们决定了开放的染色质结构以促进转录。因此,我们想了解sirt6-tip60-gata4轴是否/如何将染色质重塑整合到基因转录中。再次,以bcl2的启动子为例,chip数据显示tip60在p1区域高度富集,当sirt6被敲低时,tip60被显著抑制(图6a)。一致性地,tip60的过表达显著增强了p1处h3k9ac的水平(图6b),该现象被sirt6缺失所减弱(图6c)。有趣的是,尽管在过表达sirt6时,h3k9ac和h3k56ac的总体水平显著降低,但是chip测定显示在p1基序处的h3k9ac和h3k56ac的富集并没有改变(图6c)。同样,sirt6敲低(kd)并不影响h3k9ac在p1基序上的富集。相比之下,gata4kd在启动子位点显著下调了h3k9ac的水平,但不影响sirt6和tip60的富集(图6d-6f)
我们注意到,酶失活突变体h133y完全消除了sirt6和gata4之间的相互作用(图6g),以及消除了dox诱导的心肌细胞凋亡的能力。因此,我们认为gata4的关联可能会抑制sirt6在靶启动子上的脱乙酰基酶活性。为了获得更多的实验证据,我们进行了体外去乙酰化测定,发现纯化的带有his标签的gata4显著阻断了重组sirt6的脱乙酰酶活性(图6h)。考虑到sirt6/tip60复合物介导gata4在k328/330处的乙酰化,并且gata4的c末端锌指对于sirt6结合至关重要,我们合成了在gata4的该区域(aa295-331)的两个肽段:具有由seqidno:5表示的氨基酸序列的g4-24(aa295-318),以及具有由seqidno:2表示的氨基酸序列的g4-13(aa319-331)。体外去乙酰化测定表明g4-13,而非g4-24,显著抑制了sirt6的脱乙酰基酶活性(14μm,50%的抑制)(图6i)。
考虑到sirt6的核小体缔合也需要h133位点,结合我们的发现,我们提出sirt6通过一般的核小体结合特性占据启动子区域。sirt6总体地或在特定基因启动子处募集tip60,以平衡组蛋白的动态乙酰化;当募集gata4并阻断sirt6脱乙酰基酶活性时,tip60将gata4乙酰化以增强其转录活性,并使组蛋白(例如h3k9ac)乙酰化,从而确保了用于转录的开放的染色质结构(图7)。
除非另有说明,否则本申请中涉及的数值范围包括上限值和下限值。尽管已经示出并描述了本申请的特定实施例,但是对于本领域技术人员显而易见的是,可以在不脱离本申请的更广泛方面的情况下进行改变和修改,因此,所附权利要求书的目的是涵盖落入本申请的真实精神和范围内的所有这样的改变和修改。
序列表
<110>深圳大学
<120>增强抗凋亡途径以预防dox毒性的方法
<130>pct1921316-1
<160>5
<170>patentinversion3.5
<210>1
<211>546
<212>prt
<213>智人(homosapiens)
<400>1
metalagluvalvalserprovalproglyalaglyargargglupro
151015
glygluvalglyargalaargglyproprovalalaaspproglyval
202530
alaleuserproglnglygluileilegluglycysargleuproval
354045
leuargargasnglnaspasngluaspglutrpproleualagluile
505560
leuservallysaspileserglyarglysleuphetyrvalhistyr
65707580
ileasppheasnlysargleuaspglutrpvalthrhisgluargleu
859095
aspleulyslysileglnpheprolyslysglualalysthrprothr
100105110
lysasnglyleuproglyserargproglyserprogluarggluval
115120125
proalaseralaglnalaserglylysthrleuproileprovalgln
130135140
ilethrleuargpheasnleuprolysgluargglualaileprogly
145150155160
glygluproaspglnproleusersersersercysleuglnproasn
165170175
hisargserthrlysarglysvalgluvalvalserproalathrpro
180185190
valprosergluthralaproalaservalpheproglnasnglyala
195200205
alaargargalavalalaalaglnproglyarglysarglysserasn
210215220
cysleuglythraspgluaspserglnaspserseraspglyilepro
225230235240
seralaproargmetthrglyserleuvalseraspargserhisasp
245250255
aspilevalthrargmetlysasnileglucysilegluleuglyarg
260265270
hisargleulysprotrptyrpheserprotyrproglngluleuthr
275280285
thrleuprovalleutyrleucysgluphecysleulystyrglyarg
290295300
serleulyscysleuglnarghisleuthrlyscysaspleuarghis
305310315320
proproglyasngluiletyrarglysglythrileserphepheglu
325330335
ileaspglyarglysasnlyssertyrserglnasnleucysleuleu
340345350
alalyscyspheleuasphislysthrleutyrtyraspthrasppro
355360365
pheleuphetyrvalmetthrglutyraspcyslysglyphehisile
370375380
valglytyrpheserlysglulysgluserthrgluasptyrasnval
385390395400
alacysileleuthrleuproprotyrglnargargglytyrglylys
405410415
leuleuilegluphesertyrgluleuserlysvalgluglylysthr
420425430
glythrproglulysproleuseraspleuglyleuleusertyrarg
435440445
sertyrtrpserglnthrileleugluileleumetglyleulysser
450455460
gluserglygluargproglnilethrileasngluilesergluile
465470475480
thrserilelyslysgluaspvalileserthrleuglntyrleuasn
485490495
leuileasntyrtyrlysglyglntyrileleuthrleusergluasp
500505510
ilevalaspglyhisgluargalametleulysargleuleuargile
515520525
aspserlyscysleuhisphethrprolysasptrpserlysarggly
530535540
lystrp
545
<210>2
<211>13
<212>prt
<213>智人(homosapiens)
<400>2
arglysarglysprolysasnleuasnlysserlysthr
1510
<210>3
<211>443
<212>prt
<213>智人(homosapiens)
<400>3
mettyrglnserleualametalaalaasnhisglyproproprogly
151015
alatyrglualaglyglyproglyalaphemethisglyalaglyala
202530
alaserserprovaltyrvalprothrproargvalproserserval
354045
leuglyleusertyrleuglnglyglyglyalaglyseralasergly
505560
glyalaserglyglyserserglyglyalaalaserglyalaglypro
65707580
glythrglnglnglyserproglytrpserglnalaglyalaaspgly
859095
alaalatyrthrproproprovalserproargpheserpheprogly
100105110
thrthrglyserleualaalaalaalaalaalaalaalaalaargglu
115120125
alaalaalatyrserserglyglyglyalaalaglyalaglyleuala
130135140
glyarggluglntyrglyargalaglyphealaglysertyrserser
145150155160
protyrproalatyrmetalaaspvalglyalasertrpalaalaala
165170175
alaalaalaseralaglypropheaspserprovalleuhisserleu
180185190
proglyargalaasnproalaalaarghisproasnleuvalaspmet
195200205
pheaspaspphesergluglyargglucysvalasncysglyalamet
210215220
serthrproleutrpargargaspglythrglyhistyrleucysasn
225230235240
alacysglyleutyrhislysmetasnglyileasnargproleuile
245250255
lysproglnargargleuseralaserargargvalglyleusercys
260265270
alaasncysglnthrthrthrthrthrleutrpargargasnalaglu
275280285
glygluprovalcysasnalacysglyleutyrmetlysleuhisgly
290295300
valproargproleualametarglysgluglyileglnthrarglys
305310315320
arglysprolysasnleuasnlysserlysthrproalaalaproser
325330335
glysergluserleuproproalaserglyalaserserasnserser
340345350
asnalathrthrsersersergluglumetargproilelysthrglu
355360365
proglyleuserserhistyrglyhisserserservalserglnthr
370375380
pheservalseralametserglyhisglyproserilehisproval
385390395400
leuseralaleulysleuserproglnglytyralaserprovalser
405410415
glnserproglnthrserserlysglnaspsertrpasnserleuval
420425430
leualaaspserhisglyaspileilethrala
435440
<210>4
<211>355
<212>prt
<213>智人(homosapiens)
<400>4
metservalasntyralaalaglyleuserprotyralaasplysgly
151015
lyscysglyleuprogluilepheaspproproglugluleugluarg
202530
lysvaltrpgluleualaargleuvalargglnserserservalval
354045
phehisthrglyalaglyileserthralaserglyileproaspphe
505560
argglyprohisglyvaltrpthrmetglugluargglyleualapro
65707580
lyspheaspthrthrphegluseralaargprothrglnthrhismet
859095
alaleuvalglnleugluargvalglyleuleuargpheleuvalser
100105110
glnasnvalaspglyleuhisvalargserglypheproargasplys
115120125
leualagluleuhisglyasnmetphevalgluglucysalalyscys
130135140
lysthrglntyrvalargaspthrvalvalglythrmetglyleulys
145150155160
alathrglyargleucysthrvalalalysalaargglyleuargala
165170175
cysargglygluleuargaspthrileleuasptrpgluaspserleu
180185190
proaspargaspleualaleualaaspglualaserargasnalaasp
195200205
leuserilethrleuglythrserleuglnileargproserglyasn
210215220
leuproleualathrlysargargglyglyargleuvalilevalasn
225230235240
leuglnprothrlyshisasparghisalaaspleuargilehisgly
245250255
tyrvalaspgluvalmetthrargleumetlyshisleuglyleuglu
260265270
ileproalatrpaspglyproargvalleugluargalaleupropro
275280285
leuproargproprothrprolysleugluprolysglugluserpro
290295300
thrargileasnglyserileproalaglyprolysglngluprocys
305310315320
alaglnhisasnglysergluproalaserprolysarggluargpro
325330335
thrserproalaprohisargproprolysargvallysalalysala
340345350
valproser
355
<210>5
<211>24
<212>prt
<213>智人(homosapiens)
<400>5
alacysglyleutyrmetlysleuhisglyvalproargproleuala
151015
metarglysgluglyileglnthr
20
- 还没有人留言评论。精彩留言会获得点赞!