用于治疗髓系恶性肿瘤的嵌合抗原受体
hct。在一些实施例中,已经对受试者进行造血干细胞移植治疗。在其他实施例中,未对受试者进行造血干细胞移植治疗。在一些实施例中,受试者不符合allohct的资格。
11.在一些实施例中,抗cd83结合剂是特异性结合cd83的抗体片段。例如,抗原结合结构域可以是特异性结合cd83的抗体的fab或单链可变片段(scfv)。在一些实施例中,抗cd83结合剂是特异性结合cd83的适体。例如,抗cd83结合剂可以是基于其结合cd83的能力从随机序列库中选择的肽适体。抗cd83结合剂也可以是cd83的天然配体,或其能够结合cd83的变体和/或片段。
12.在一些实施例中,抗cd83 scfv可以包含具有cdr1、cdr2和cdr3序列的可变重(vh)结构域和具有cdr1、cdr2和cdr3序列的可变轻(v
l
)结构域。
13.例如,在一些实施例中,vh结构域的cdr1序列包含氨基酸序列gfsittggywwt(seq id no:1)、sdgis(seq id no:7)、或snami(seq id no:13);vh结构域的cdr2序列包含氨基酸序列gyifssgntnynpsiks(seq id no:2)、iissggntyyaswakg(seq id no:8)、或amdsnsrtyyatwakg(seq id no:14);vh结构域的cdr3序列包含氨基酸序列caraygklgfdy(seq id no:3)、vvggtysi(seq id no:9)、或gdggssdytem(seq id no:15);v
l
的cdr1序列包含氨基酸序列tlssqhstytig(seq id no:4)、qssqsvynndfls(seq id no:10)、或qssqsvygnnels(seq id no:16);v
l
结构域的cdr2序列包含氨基酸序列vnsdgshskgd(seq id no:5)、yastlas(seq id no:11)、或qasslas(seq id no:17);并且v
l
结构域的cdr3序列包含氨基酸序列gssdssgyv(seq id no:6)、tgtygnsawyeda(seq id no:12)、或lgeysisadnh(seq id no:18)。
14.例如,在一些实施例中,vh结构域的cdr1序列包含氨基酸序列gfsittggywwt(seq id no:1),vh结构域的cdr2序列包含氨基酸序列gyifssgntnynpsiks(seq id no:2),vh结构域的cdr3序列包含氨基酸序列caraygklgfdy(seq id no:3),v
l
结构域的cdr1序列包含氨基酸序列tlssqhstytig(seq id no:4),v
l
结构域的cdr2序列包含氨基酸序列vnsdgshskgd(seq id no:5),并且v
l
结构域的cdr3序列包含氨基酸序列gssdssgyv(seq id no:6)。
15.例如,在一些实施例中,vh结构域的cdr1序列包含氨基酸序列sdgis(seq id no:7),vh结构域的cdr2序列包含氨基酸序列iissggntyyaswakg(seq id no:8),vh结构域的cdr3序列包含氨基酸序列vvggtysi(seq id no:9),v
l
结构域的cdr1序列包含氨基酸序列qssqs vynndfls(seq id no:10),v
l
结构域的cdr2序列包含氨基酸序列yastlas(seq id no:11),并且v
l
结构域的cdr3序列包含氨基酸序列tgtygnsawyeda(seq id no:12)。
16.例如,在一些实施例中,vh结构域的cdr1序列包含氨基酸序列snami(seq id no:13),vh结构域的cdr2序列包含氨基酸序列amdsnsrtyyatwakg(seq id no:14),vh结构域的cdr3序列包含氨基酸序列gdggssdytem(seq id no:15),v
l
结构域的cdr1序列包含氨基酸序列qssqsvygnnels(seq id no:16),v
l
结构域的cdr2序列包含氨基酸序列qasslas(seq id no:17),并且v
l
结构域的cdr3序列包含氨基酸序列lgeysisadnh(seq id no:18)。
17.在一些实施例中,抗cd83 scfv vh结构域包含以下氨基酸序列:qvqlkesgpglvkpsqslsltcsvtgfsittggywwtwirqfpgqklewmgyifssgntnynpsiksrisitrdtsknqfflqlnsvttegdtaryycaraygklgfdywgqgtlvtvss(seq id no:19,vh-gbm00)。
18.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:qpvltqspsasaslgnsvkitctlssqhstytigwyqqhpdkapkyvmyvnsdgshskgdgipdrfsgsssgahrylsisniqpedeady
id no:26,14c12)。
25.在一些实施例中,抗cd83 scfv vh结构域包含以下氨基酸序列:metglrwlllvavlkgvqcqsveesggrlvtpgtpltltctvsgfslssydmtwvrqapgkglewigiiyasgttyyanwakgrftisktsttvdlkvtsptigdtatyfcaregagvsmtlwgpgtlvtvssgqpkapsvfplapccgdtpsstvtlgclvkgylpepvtvtwnsgtltngvrtfpsvrqssglyslssvvsvtsssqpvtcnvahpatntkvdktvapstcskptcpppellggpsvfifppkpkdtlmisrtpevtcvvvdvsqddpevqftwyinneqvrtarpplreqqfnstirvvstlpiahqdwlrgkefkckvhnkalpapiektiskargqplepkvytmgppreelssrsvsltcmingfypsdisvewekngkaednykttpavldsdgsyflynklsvptsewqrgdvftcsvmhealhnhytqksisrspgk(seq id no:27,020b08)。
26.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:mdmraptqllgllllwlpgarcaydmtqtpasvevavggtvtikcqasqsistyldwyqqkpgqppklliydasdlasgvpsrfkgsgsgtqftltisdlecadaatyycqqgythsnvdnvfgggtevvvkgdpvaptvllfppssdevatgtvtivcvankyfpdvtvtwevdgttqttgiensktpqnsadctynlsstltltstqynshkeytckvtqgttsvvqsfsrknc(seq id no:28,020b08)。
27.在一些实施例中,抗cd83 scfv vh结构域包含以下氨基酸序列:metglrwlllvavlkgvqcqsveesggrlvspgtpltltctasgfslssydmswvrqapgkgleyigiisssgstyyaswakgrftisktsttvdlevtslttedtatyfcsrehagysgdtghlwgpgtlvtvssgqpkapsvfplapccgdtpsstvtlgclvkgylpepvtvtwnsgtltngvrtfpsvrqssglyslssvvsvtsssqpvtcnvahpatntkvdktvapstcskptcpppellggpsvgigppkpkdtlmisrtpevtcvvvdvsqddpevqftwyinneqvrtarpplreqqfnstirvvstlpiahqdwlrgkefkckvhnkalpapiektiskargqplepkvytmgppreelssrsvsltcmingfypsdisvewekngkaednykttpavldsdgsyflynklsvptsewqrgdvftcsvmhealhnhytqksisrspgk(seq id no:29,006g05)。
28.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:mdmraptqllgllllwlpgarcaydmtqtpasvevavggtvaikcqasqsvssylawyqqkpgqppkpliyeasmlaagvssrfkgsgsgtdftltisdlecddaatyycqqgysisdidnafgggtevvvkgdpvaptvllfppssdevatgtvtivcvankyfpdvtvtwevdgttqttgiensktpqnsadctynlsstltltstqynshkeytckvtqgttsvvqsfsrknc(seq id no:30,006g05)。
29.在一些实施例中,抗cd83 scfv vh结构域包含以下氨基酸序列:metglrwlllvavlkgvqcqsveesggrlvtpgtpltltctvsgidlssdgiswvrqapgkglewigiissggntyyaswakgrftisrtsttvdlkmtslttedtatyfcarvvggtysiwgqgtlvtvssastkgpsvyplapgsaaqtnsmvtlgclvkgyfpepvtvtwnsgslssgvhtfpavlqsdlytlsssvtvpsstwpsetvtcnvahpasstkvdkkivprdcgckpcictvpevssvfifppkpdvltitltpkvtcvvvdiskddpevqfswfvddvevhtaqtqpreeqfnstfrsvselpimhqdwlngkefkcrvnsaafpapiektisktkgrpkapqvytipppkeqmakdkvsltcmitdffpeditvewqwngqpaenykntqpimdtdgsyfvysklnvqksnweagntftcsvlheglhnhhtekslshspgk(seq id no:31,96g08)。
30.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:mdtraptqllgllllwlpgatfaqvltqtaspvsapvggtvtincqssqsvynndflswyqqkpgqppklliyyastlasgvpsrfkgsgsgtqftltisdlecddaatyyctgtygnsawyedafgggtevvvkrtpvaptvllfppssaelatgtativcvankyfpdgtvtwkvdgitqssginnsrtpqnsadctynlsstltlssdeynshdeytcqvaqdsgspvvqsfsrksc
(seq id no:32,96g08)
31.在一些实施例中,抗cd83 scfv vh结构域包含以下氨基酸序列:metglrwlllvavlkgvqcqsveesggrlvtpgtpltltctvsgidlssnamiwvrqapreglewigamdsnsrtyyatwakgrftisrtssitvdlkitspttedtatyfcargdggssdytemwgpgtlvtvssastkgpsvyplapgsaaqtnsmvtlgclvkgyfpepvtvtwnsgslssgvhtfpavlqsdlytlsssvtvpsstwpsetvtcnvahpasstkvdkkivprdcgckpcictvpevssvfifppkpkdvltitltpkvtcvvvdiskddpevqfswfvddvevhtaqtqpreeqfnstfrsvselpimhqdwlngkefkcrvnsaafpapiektisktkgrpkapqvytipppkeqmakdkvsltcmitdffpeditvewqwngqpaenykntqpimdtdgsyfvysklnvqksnweagntftcsvlheglhnhhtekslshspgk(seq id no:33,95f04)。
32.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:mdtraptqllgllllwlpgatfaqavvtqttspvsapvggtvtincqssqsvygnnelswyqqkpgqppklliyqasslasgvpsrfkgsgsgtqftltisdlecddaatyyclgeysisadnhfgggtevvvkrtpvaptvllfppssaelatgtativcvankyfpdgtvtwkvdgitqssginnsrtpqnsadctynlsstltlssdeynshdeytcqvaqdsgspvvqsfsrksc(seq id no:34,95f04)
33.在一些实施例中,抗cd83 scfv vh结构域包含以下氨基酸序列:qvqlvqsggavvqpgrslrlscaasgftfstygmhwvrqapgkglewvaavsydgsnkyyadfvkgrftisrdnpkntlylqmnslraddtavyycarrggldiwgqgttvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscaaa(seq id no:35)。
34.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:ltqpppasgtpgqqrvtiscsgsssnigsntvnwyqqlpgtapklliyygndqrpsgvpdrfsasksgtsaslaisglqsedeahyycaawdgslnggvifgggtkvtlg(seq id no:36)。
35.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:vtqppsasgtpgqrvtiscsgsssnigtnpvnwyqqlpgtapklliyttdqrpsgvpdrfsgsksgtsaslaisglqsedeadyycaawddslsglyvfgtgtkvtvlg(seq id no:37)。
36.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:mthtplslsvtpgqpasisckssqsllhsdgktylywylqrpgqspqpliyevsnrfsgvpdrfsgsgsgtdftlkisrvqaedvgvyycmqslqlwtfgqgtkveikr(seq id no:38)。
37.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:mtqsplslpvtlgqpasiscrssqslihsdgntyldwfqqrpgqsprrliykvsnrdsgvpdrfsgsgsgtdftlrisrveaedigvyycmqathwprtfgqgtkveikr(seq id no:39)。
38.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:mtqsplslpvtlgqpasiscrssqslvdsagntflhwfhqrpgqsprrliykvsnrdsgvpdrfsgsgsgtdftlkisrveaedvgvyycmqgthwprtfgqgtkveikr(seq id no:40)。
39.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:ltqsplslpvtlgqpasisckssqslvdsdgntylnwfqqrpgqsprrliykvsnrdsgvpdrfsgsgsgtdftlkisrveaedvgvyycmqgthwprtfgqgtkveikr(seq id no:41)。
40.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:mtqsplslpvtlgqpasiscrssqslvhsdgnmylnwfqqrpgqsprrliykvsnrdsgvpdrfsgsgsgtdftlkisrveaedvgvyycmqatqptwtfgqgtkleikr(seq id no:42)。
41.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:mtqspsslsasvgdrvtitcqasqdisnylnwyqqkpgkapklliydasnletgvpsrfsgsgsgtdftftissatyycqqtyqgtkleikr(seq id no:43)。
42.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:mtqspsslsasvghpvtitcrasqslisylnwyhqkpgkapklliyaasilqsgvpsrfsgsgsgtdftltisslqpenfasyycqhtdsfprtfghgtkveikr(seq id no:44)。
43.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:ltqppsasgtpgqgvtiscrgstsnignnvvnwyqhvpgsapklliwsniqrpsgipdrfsgsksgtsaslaisglqsedqavyycavwddglagwvfgggttvtvls(seq id no:45)。
44.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:mtqapvvsvaleqtvritcqgdslaiyydfwyqhkpgqapvlviygknnrpsgiphrfsgsssntdsltitgaqaedeadyycnsrdssgnhwvfgggtnltvlg(seq id no:46)。
45.在一些实施例中,抗cd83 scfv v
l
结构域包含以下氨基酸序列:ltqsplslpvtlgqpasiscksnqslvhsdgntylnwfqqrpgqsprrliykvsnrdsgvpdrfsgsgsgtdftlkinrveaedvgvyycmqgtqwprtfggqgtkldikr(seq id no:47)。
46.在一些实施例中,抗cd83 scfv vh结构域已人源化并且包含以下氨基酸序列:
47.qvqlqesgpglvkpsetlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrvtisvdtsknqfslklssvtaadtavyycaraygklgfdywgqgtlvtvss(seq id no:48,vh-gbm01)。
48.在一些实施例中,抗cd83 scfv vh结构域已人源化并且包含以下氨基酸序列:
49.qvqlqesgpglvkpsqtlsltctvsgfsittggywwtwirqhpgkglewigyifssgntnynpsikslvtisvdtsknqfslklssvtaadtavyycaraygklgfdywgqgtlvtvss(seq id no:49,vh-gbm02)。
50.在一些实施例中,抗cd83 scfv vh结构域已人源化并且包含以下氨基酸序列:
51.qvqlqesgpglvkpsqtlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrvtisvdtsknqfslklssvtaadtavyycaraygklgfdywgqgtlvtvss(seq id no:50,vh-gbm03)。
52.在一些实施例中,抗cd83 scfv vh结构域已人源化并且包含以下氨基酸序列:
53.qvqlqesgpglvkpsetlsltctvsgfsittggywwtwirqppgkglewi gyifssgntnynpsiksrvtisrdtsknqfslklssvtaadtavyycaraygklgfdywgqgtlvtvss(seq id no:51,vh-gbm04)。
54.在一些实施例中,抗cd83 scfv vh结构域已人源化并且包含以下氨基酸序列:
55.qvqlqesgpglvkpsetlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrvtisvdtsknqfslklssvtaadtaryycaraygklgfdywgqgtlvtvss(seq id no:52,vh-gbm05)。
56.在一些实施例中,抗cd83 scfv vh结构域已人源化并且包含以下氨基酸序列:
57.qvqlqesgpglvkpsetlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrisitrdtsknqfflqlnsvttegdtaryycaraygklgfdywgqgtlvtvss(seq id no:53,vh-gbm06)。
58.在一些实施例中,抗cd83 scfv v
l
结构域已人源化并且包含以下氨基酸序列:
59.qlvltqspsasaslgasvkltctlssqhstytigwhqqqpekgprylmkvnsdgshskgdgipdrfsgsssgaeryltisslqsedeadyycgssdssgyvfgsgtkvtvl(seq id no:54,vl-gbm01)。
60.在一些实施例中,抗cd83 scfv v
l
结构域已人源化并且包含以下氨基酸序列:
61.lpvltqppsasallgasikltctlssqhstytigwyqqrpgrspqyimkvn sdgshskgdgipdrfmgsssgadryltfsnlqsddeaeyhcgssdssgyvfgsgtkvtvl(seq id no:55,vl-gbm02)。
62.重链和轻链优选由接头分开。scfv抗体的合适接头是本领域已知的。在一些实施例中,接头包含氨基酸序列ggggsggggsggggs(seq id no:56)。
63.在一些实施例中,抗cd83 scfv包含氨基酸序列:qpvltqspsasaslgnsvkitctlssqhstytigwyqqhpdkapkyvmyvnsdgshskgdgipdrfsgsssgahrylsisniqpedeadyfcgssdssgyvfgsgtqltvlraaassggggsggggsggggsqpvltqspsasaslgnsvkitctlssqhstytigwyqqhpdkapkyvmyvnsdgshskgdgipdrfsgsssgahrylsisniqpedeadyfcgssdssgyvfgsgtqltvlraaa(seq id no:57)。
64.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlkesgpglvkpsqslsltcsvtgfsittggywwtwirqfpgqklewmgyifssgntnynpsiksrisitrdtsknqfflqlnsvttegdtaryycaraygklgfdywgqgtlvtvssggggsggggsggggsqvqlkesgpglvkpsqslsltcsvtgfsittggywwtwirqfpgqklewmgyifssgntnynpsiksrisitrdtsknqfflqlnsvttegdtaryycaraygklgfdywgqgtlvtv(seq id no:58)。
65.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlqesgpglvkpsetlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrvtisvdtsknqfslklssvtaadtavyycaraygklgfdywgqgtlvtvssggggsggggsggggsqlvltqspsasaslgasvkltctlssqhstytigwhqqqpekgprylmkvnsdgshskgdgipdrfsgsssgaeryltisslqsedeadyycgssdssgyvfgsgtkvtvl(seq id no:59)。
66.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlqesgpglvkpsqtlsltctvsgfsittggywwtwirqhpgkglewigyifssgntnynpsikslvtisvdtsknqfslklssvtaadtavyycaraygklgfdywgqgtlvtvssggggsggggsggggsqlvltqspsasaslgasvkltctlssqhstytigwhqqqpekgprylmkvnsdgshskgdgipdrfsgsssgaeryltisslqsedeadyycgssdssgyvfgsgtkvtvl(seq id no:60)。
67.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlqesgpglvkpsqtlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrvtisvdtsknqfslklssvtaadtavyycaraygklgfdywgqgtlvtvssggggsggggsggggsqlvltqspsasaslgasvkltctlssqhstytigwhqqqpekgprylmkvnsdgshskgdgipdrfsgsssgaeryltisslqsedeadyycgssdssgyvfgsgtkvtvl(seq id no:61)。
68.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlqesgpglvkpsetlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrvtisrdtsknqfslklssvtaadtavyycaraygklgfdywgqgtlvtvssggggsggggsggggsqlvltqspsasaslgasvkltctlssqhstytigwhqqqpekgprylmkvnsdgshskgdgipdrfsgsssgaeryltisslqsedeadyycgssdssgyvfgsgtkvtvl(seq id no:62)。
69.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlqesgpglvkpsetlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrvtisvdtsknqfslklssvtaadtaryycaraygklgfdywgqgtlvtvssggggsggggsggggsqlvltqspsasaslgasvkltctlssqhstytigwhqqqpekgprylmkvnsdgshskgdgipdrfsgsssgaeryltisslqsedeadyycgssdssgyvfgsgtkvtvl(seq id no:63)。
70.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlqesgpglvkpsetlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrisitrdtsknqfflqlnsvttegdtaryycarayg
klgfdywgqgtlvtvssggggsggggsggggsqlvltqspsasaslgasvkltctlssqhstytigwhqqqpekgprylmkvnsdgshskgdgipdrfsgsssgaeryltisslqsedeadyycgssdssgyvfgsgtkvtvl(seq id no:64)。
71.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlqesgpglvkpsetlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrvtisvdtsknqfslklssvtaadtavyycaraygklgfdywgqgtlvtvssggggsggggsggggslpvltqppsasallgasikltctlssqhstytigwyqqrpgrspqyimkvnsdgshskgdgipdrfmgsssgadryltfsnlqsddeaeyhcgssdssgyvfgsgtkvtvl(seq id no:65)。
72.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlqesgpglvkpsqtlsltctvsgfsittggywwtwirqhpgkglewigyifssgntnynpsikslvtisvdtsknqfslklssvtaadtavyycaraygklgfdywgqgtlvtvssggggsggggsggggslpvltqppsasallgasikltctlssqhstytigwyqqrpgrspqyimkvnsdgshskgdgipdrfmgsssgadryltfsnlqsddeaeyhcgssdssgyvfgsgtkvtvl(seq id no:66)。
73.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlqesgpglvkpsqtlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrvtisvdtsknqfslklssvtaadtavyycaraygklgfdywgqgtlvtvssggggsggggsggggslpvltqppsasallgasikltctlssqhstytigwyqqrpgrspqyimkvnsdgshskgdgipdrfmgsssgadryltfsnlqsddeaeyhcgssdssgyvfgsgtkvtvl(seq id no:67)。
74.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlqesgpglvkpsetlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrvtisrdtsknqfslklssvtaadtavyycaraygklgfdywgqgtlvtvssggggsggggsggggslpvltqppsasallgasikltctlssqhstytigwyqqrpgrspqyimkvnsdgshskgdgipdrfmgsssgadryltfsnlqsddeaeyhcgssdssgyvfgsgtkvtvl(seq id no:68)。
75.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlqesgpglvkpsetlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrvtisvdtsknqfslklssvtaadtaryycaraygklgfdywgqgtlvtvssggggsggggsggggslpvltqppsasallgasikltctlssqhstytigwyqqrpgrspqyimkvnsdgshskgdgipdrfmgsssgadryltfsnlqsddeaeyhcgssdssgyvfgsgtkvtvl(seq id no:69)。
76.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlqesgpglvkpsetlsltctvsgfsittggywwtwirqppgkglewigyifssgntnynpsiksrisitrdtsknqfflqlnsvttegdtaryycaraygklgfdywgqgtlvtvssggggsggggsggggslpvltqppsasallgasikltctlssqhstytigwyqqrpgrspqyimkvnsdgshskgdgipdrfmgsssgadryltfsnlqsddeaeyhcgssdssgyvfgsgtkvtvl(seq id no:70)。
77.在一些实施例中,抗cd83 scfv包含氨基酸序列:qvqlkesgpglvkpsqslsltcsvtgfsittggywwtwirqfpgqklewmgyifssgntnynpsiksrisitrdtsknqfflqlnsvttegdtaryycaraygklgfdywgqgtlvtvssggggsggggsggggsqpvltqspsasaslgnsvkitctlssqhstytigwyqqhpdkapkyvmyvnsdgshskgdgipdrfsgsssgahrylsisniqpedeadyfcgssdssgyvfgsgtqltvl(seq id no:71)。
78.与其他car一样,所披露的多肽也可以包含能够激活免疫效应细胞的跨膜结构域
和胞内结构域。例如,胞内结构域可以包含信号传导结构域和一个或多个共刺激信号传导区。
79.在一些实施例中,细胞内信号传导结构域是cd3ζ(cd3 zeta)信号传导结构域。在一些实施例中,共刺激信号传导区包含cd28、4-1bb或其组合的细胞质结构域。在一些情况下,共刺激信号传导区包含一个或多个细胞内信号传导和/或共刺激分子的1、2、3或4个细胞质结构域。在一些实施例中,共刺激信号传导区包含增强信号传导的cd28和/或4-1bb的细胞质结构域中的一个或多个突变。
80.在一些实施例中,car多肽包含不完整的胞内结构域。例如,car多肽可以仅包含细胞内信号传导结构域或共刺激结构域,而不是包含两者。在这些实施例中,除非免疫效应细胞和包含缺失结构域的第二car多肽(或内源性t细胞受体)均结合它们各自的抗原,否则免疫效应细胞不被激活。因此,在一些实施例中,car多肽包含cd3ζ(cd3 zeta)信号传导结构域,但不包含共刺激信号传导区(csr)。在其他实施例中,car多肽包含cd28、4-1bb或其组合的细胞质结构域,但不包含cd3ζ(cd3 zeta)信号传导结构域(sd)。
81.还披露了编码所披露的car多肽的分离的核酸序列、包含这些分离的核酸的载体以及包含这些载体的细胞。例如,细胞可以是选自由以下组成的组的免疫效应细胞:α-βt细胞、γ-δt细胞、自然杀伤(nk)细胞、自然杀伤(nkt)细胞、b细胞、先天性淋巴细胞(ilc)、细胞因子诱导的杀伤(cik)细胞、细胞毒性t淋巴细胞(ctl)、淋巴因子激活的杀伤(lak)细胞和调节性t细胞。
82.在附图和下面的描述中阐述了本发明的一个或多个实施例的详情。从说明书和附图以及权利要求书看,本发明的其他特征、目的和优点将显而易见。
附图说明
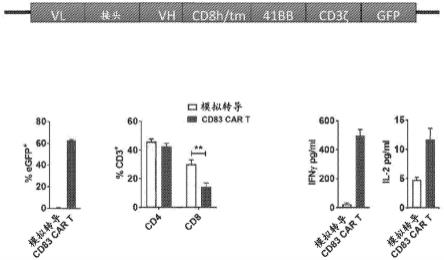
83.图1a至图1g显示了人cd83靶向cart构建体和功能特征。图1a显示了抗cd83单链可变片段之后是cd8铰链和跨膜结构域,以及41bb共刺激结构域和cd3s激活结构域。将car在3'端用荧光报告子加标签。将car报告基因克隆到sfg逆转录病毒载体中。图1b是显示在模拟转导t细胞(egfp阴性)或cd83 car t细胞(egfp阳性)中产生后表达egfp报告子的t细胞的量(平均值
±
sem)的条形图。图1c是展示了模拟转导t细胞或cd83 cart细胞中cd4或cd8表达的相对量(平均值
±
sem)的条形图,sidak检验。图1d和图1e显示了模拟转导t细胞或cd83 cart细胞在cd83+dc刺激后释放的ifny和il-2的量。图1f显示了cd83 cart细胞或模拟转导t细胞与cd83+dc共培养,并在实时细胞分析系统上测量细胞毒性。数据呈现为重复孔随时间变化的平均归一化细胞指数。归一化细胞指数计算为给定时间点的细胞指数除以归一化时间点(即添加t细胞后的第1天)的细胞指数。显示了2个代表性实验中的1个,dunnett检验。图1g显示了cd83+dc刺激cd83 cart细胞或模拟转导t细胞,并在14天时间段内每周计算t细胞的绝对数量。显示了2个代表性实验中的1个,sidak检验。**p=.001-.01,***p=.0001-.001,并且****p《.0001。
84.图2显示了人cd83嵌合抗原受体t细胞降低同种异体反应性。将人t细胞与同种异体的细胞因子成熟的单核细胞衍生的树突状细胞(modc)以1:30的dc:t细胞比率(即100000个t细胞和3333个modc)培养。将cd83 cart(对于培养的t细胞为自体的)以特定比率添加到modc中(3:1至1:10,其中添加的cart最低量为333个细胞)。在第+5天,通过ki-67表达测量t
细胞增殖。car t因gfp的表达而被门控在外。对照包括单独的t细胞(即没有增殖)、模拟转导t细胞和cd19 cart细胞。这些模拟转导t细胞不表达嵌合抗原受体,但以与转导cd83细胞相同的方式处理。cd19 cart细胞使用41bb共刺激结构域,并靶向该系统中的无关抗原。显示了2个代表性实验中的1个。
85.图3a至图3d显示了与调节性t细胞(treg)相比,cd83在激活的人常规cd4+ t细胞(tcon)上差异表达。用同种异体modc(dc:t细胞比率1:30)或cd3/cd28珠(珠:t细胞比率1:30)刺激人t细胞。在基线,刺激后4小时、8小时、24小时和48小时,测量激活的tconv(cd4+、cd127+、cd25+)或treg(cd4+、cd127-、cd25+、foxp3+)上的cd83表达。条形图显示了同种异体dc(图3a)或cd3/cd28珠(图3b)刺激后cd83+tconv或treg(平均值
±
sem)的量。n=5个独立实验,sidak检验。将人cd83 car或模拟t细胞与dc同种异体刺激的pbmc以1:10的比率培养48小时。代表性等高线图显示了随时间变化的cd83+、cd3-和cd3+靶细胞的频率(图3c)以及egfp+cart细胞中cd83的表达(图3d)。显示了2个代表性实验中的1个。****p《.0001。
86.图4a至图4j显示了人cd83 cart细胞预防异种gvhd。图4a显示了接受25x106个人pbmc并接种低剂量(1x106)或高剂量(10x106)cd83 car或(1-10x106)模拟转导t细胞的nsg小鼠。car对于pbmc供体是自体的。另一个对照组小鼠单独接受pbmc。图4a和图4b显示了存活率(图4a)和gvhd(图4b)临床评分。临床评分包括对活动、皮毛和皮肤状况、体重减轻和姿势的综合评估。汇集了来自3个独立实验的数据,每个实验组最多9只小鼠。对数秩检验。在单独的实验中,受体小鼠在+21天时被人道地安乐死,并且组织gvhd严重程度由专家、致盲病理学家评估。显示了受体肺(图4c-4f)和肝脏(图4g-4j)的异种gvhd路径评分,代表性h&e图像,ki-67+、cd3+ t细胞/hpf的量,和代表性ihc图像(cd3=红色,ki-67=棕色)。汇集了来自2个独立实验的数据,每个实验组最多6只小鼠。dunnett检验(组比较)或mann-whitney。**p=.001-.01并且***p=.0001-.001。
87.图5a至图5d显示了人cd83靶向car t细胞显著降低cd83+dc。nsg小鼠如所述接受25x106个人pbmc加1x106个cd83 car或模拟转导t细胞。在+21天时人道地对小鼠实施安乐死并收获脾脏。图5a包含代表性等高线图,显示了+21天时小鼠脾脏中人cd83+、cdlc+dc的频率。图5b是显示了在+21天时小鼠脾脏中人cd83+、cd1c+dc的绝对数量(平均值
±
sem)的条形图,dunn检验。图5c包含代表性等高线图,显示了在+21天时受体脾脏中mhc ii+类、cdlc+dc的百分比。图5d是描绘这些细胞的绝对数量(平均值
±
sem)的条形图,dunn检验。汇集了来自2个独立实验的数据,每个实验组最多6只小鼠。**p=.001-.01。
88.图6:人cd83靶向cart细胞体内显著降低cd4+、cd83+ t细胞,同时增加treg:激活的tconv比率。nsg小鼠如所述接受25x l06个人pbmc加1x106个cd83 car或模拟转导t细胞。在+21天时人道地对小鼠实施安乐死并收获脾脏。a)代表性等高线图显示了与接受模拟转导t细胞的小鼠相比,在+21天时接种的小鼠中egfp+cd83 car t细胞的量。b)代表性等高线图显示了受体脾脏中人cd4+ t细胞的频率。条形图显示在+21天时小鼠脾脏中c)cd4+和d)cd4+、cd83+ t细胞的绝对数量(平均值
±
sem),dunn检验。e)等高线图描绘了+21天时小鼠脾脏中cd4+、cd12t、cd25+、foxp3+treg的百分比。条形图显示了在+21天时受体小鼠中f)treg和g)treg:激活的tconv的量(平均值
±
sem),dunnett检验。h)等高线图描绘了在+21天时小鼠脾脏中cd4+、ifny+thl细胞和cd4+、il-4+th2细胞的频率。条形图展示了受体脾脏中l)thl和j)th2细胞的绝对数量(平均值
±
sem),dunn检验。汇集了来自2个独立实验的数据,
每个实验组最多6只小鼠。*p《.05,**p=.001-.01。
89.图7:人cd83 cart细胞杀伤急性髓系白血病细胞系。直方图显示了在增殖中的(a)k562和(b)thp-1细胞中的cd83表达,右下角注有mfi。人cd83 car或模拟转导的t细胞与新鲜的k562或thp-1细胞以10:1的e/t比率共培养。使用xcelligence rtca系统监测靶细胞杀伤,dunnett检验。显示了每个的代表性实验。****p《.0001.
90.图8:人cd83 cart细胞表现出可忽略不计的中靶脱瘤毒性(on-target,off-tumor toxicity)。从正常人骨髓中分离出的cd34+细胞与cart细胞、模拟t细胞或单独的培养基以10:1的效应子与靶标的比率共同孵育4小时。将细胞一式两份铺板在methocult培养基中并培养14天,然后进行集落计数。条形图显示以下项的量:a)总集落、b)集落形成单位(cfu)-粒细胞/巨噬细胞(gm)、c)cfu-粒细胞/红细胞/单核细胞/巨核细胞(gemm)和d)成红细胞形成单位(bfu)。结果代表3个独立实验,dunnett检验。ns=不显著。
91.图9:当暴露于他克莫司时,人cd83 cart细胞仍然可以响应于cd83+靶细胞而杀伤和增殖。a)将来自同一供体的人cd83 cart细胞或未转导的t细胞与同种异体cd83+细胞因子成熟的modc以不同的t细胞与dc比率培养24小时。将培养物暴露于临床相关剂量的他克莫司(10ng/ml)或dmso对照(《0.01%)。条形图显示每次比色ldh测定在24小时时的dc裂解。b)将来自同一供体的人cd83 car t细胞或未转导的t细胞与同种异体cd83+细胞因子成熟的modc以1:30的t:dc比率培养。在第0天加入一次他克莫司或dmso对照,并在3天后通过比色测定评估增殖。显示每个的2个代表性实验中的1个,sidak检验。***p=0.0001-.001并且****p《.0001。
92.图10:人cd83 cart细胞降低体内供体细胞的扩增。nsg小鼠被移植了25x106个人pbmc加1x106个cd83 car或模拟转导t细胞。对照组由未接受pbmc的小鼠(阴性对照)和接受具有未修饰t细胞的pbmc的小鼠(二级阳性对照)组成。在+21天时人道地对受体小鼠实施安乐死,并取出它们的脾脏进行总体评估。代表性图像显示,接受pbmc和cd83 car t细胞的小鼠脾脏体积减小,支持体内供体t细胞扩增的抑制。2个代表性实验中的1个。
93.图11:人cd83 cart细胞在+21天时消除cd83+靶标。nsg小鼠被移植了25x106个人pbmc加1x106个cd83 car或模拟转导t细胞。在+21天时人道地对受体小鼠实施安乐死,并通过流式细胞术分析egfp+car、cd83+、cdlc+dc和cd83+、cd4+ t细胞的量。a)条形图显示了在+21天时受体脾脏中egfp+cart细胞的量,以及脾脏中cd83+靶标的减少百分比(通过注射模拟t细胞的小鼠归一化)。b、c)图显示了根据+21天时恢复的egfp+cart细胞的量的cd83+靶标的线性回归(虚线)。显示了斯皮尔曼级序相关系数。汇集了来自2个独立实验的数据,每个实验组最多6只小鼠。
94.图12:dc消耗不会阻止由人t细胞介导的异种gvhd。nsg小鼠单独接受7.5x106个纯化的人t细胞或与1.87x105个自体树突状细胞一起接受。通过磁珠纯化(miltenyi)分离树突状细胞,并且包括浆细胞样dc、cd1c+1型髓系dc和cd1c-,cd141
亮
2型髓系dc。显示了(a)存活率和(b)gvhd临床评分。显示了代表性实验,每个实验组4只小鼠。
95.图13:人cd83 car t细胞不会降低供体thl 7细胞的量。nsg小鼠如所述接受25x106个人pbmc加l x 106个cd83 car或模拟转导t细胞。在+21天时人道地对小鼠实施安乐死并收获脾脏。a)代表性等高线图显示了在+21天时小鼠脾脏中人cd4+、il-17+thl 7细胞的频率。b)条形图显示了在+21天时小鼠脾脏中人thl 7细胞的绝对数量(平均值
±
sem)。
汇集了来自2个独立实验的数据,每个实验组最多6只小鼠。
96.图14:人cd83 car t细胞呈现在+100天时。nsg小鼠接受25x106个人pbmc加1-10x106个cd83 car或10x106个模拟转导t细胞。等高线图显示了存活到+100天终点的代表性小鼠的脾脏中的cd83+靶细胞与egfp+cd83 cart细胞的量。显示了来自3个代表性实验中的1个的数据。
97.图15:cd83在u937和molm-13细胞上的表达。直方图显示了在增殖中的a)u937和b)molm-13细胞中的cd83表达,右下角注有mfi。
98.图16:人cd83 car t细胞降低体内供体cd8+ t细胞的量。nsg小鼠如所述接受25x106个人pbmc加1x106个cd83 car或模拟转导t细胞。a)在+21天时对供体人cd8+ t细胞的量进行计数,dunn检验。汇集了来自2个独立实验的数据,每个实验组最多6只小鼠。
具体实施方式
99.在更详细地描述本披露内容之前,应当理解,本披露内容不限于所描述的特定实施例,因此当然可以变化。还应当理解的是,因为本披露的范围将仅由所附权利要求限制,所以本文所用的术语仅是出于描述具体实施方案的目的,而不旨在是限制性的。
100.在提供了一系列值时,应当理解的是每个中间值,到下限的第十个单位(除非上下文清晰地另外指示),该范围的上限与下限之间以及任何其他陈述的或在该陈述范围内的中间值均被涵盖在本披露之内。这些更小范围的上限和下限可以独立地被包括在更小范围之内,并且也被涵盖在本披露之内,服从于在所陈述范围内任何确切排除的限制。在所陈述的范围包括一个或两个限制时,排除了那些被包括的限制的任一个或两者的范围也被包括在本披露之内。
101.除非另外定义,否则本文使用的所有技术和科学术语均具有与本披露所属领域中的普通技术人员通常所理解的相同含义。尽管类似于或等同于在此所描述的方法和材料的任何方法和材料也可以用于对本披露的实践或测试,但是现在将对优选方法和材料进行描述。
102.本说明书中引用的所有出版物和专利都通过引用并入本文,就好像每个单独的出版物或专利被具体地和单独地指示通过引用并入并且通过引用并入本文以披露和描述与引用出版物相关的方法和/或材料。对任何出版物的引用是针对其在申请日之前的披露内容,并且不应被解释为承认本披露因为先前的披露而不能获得比这种出版物更早的申请日。另外,所提供的出版物的日期可以不同于实际公开日期,所述实际公开日期可能需要被独立确认。
103.在阅读本披露之后,如对于本领域的普通技术人员将清楚的是,在此所描述和说明的单独实施例中的每个实施例具有不连续的组成部分和特征,所述组成部分和特征可以在不偏离本披露的范围或精神的情况下易于与任何其他若干实施方案的特征分离或组合。可以按所叙述的事件的顺序或按逻辑上可能的任何其他顺序来执行所叙述的任何方法。
104.除非另有指示,否则本披露内容的实施例将采用本领域技术范围内的化学、生物学等技术。
105.提出以下实例以向本领域普通技术人员提供关于如何执行本文披露和要求保护的方法和使用本文披露和要求保护的探针的完整披露内容和描述。已经努力确保关于数字
(例如,量、温度等)的准确性,但是应该考虑到一些误差和偏差。除非另有指示,份数为重量份数,温度按℃计,压力为大气压或接近大气压。标准温度和压力定义为20℃和1个大气压。
106.在详细描述本披露的实施例之前,应当理解,除非另有指示,本披露不限于特定的材料、试剂、反应材料、制造工艺等,因为这些可以变化。还应理解,本文使用的术语仅用于描述特定实施例的目的,而非旨在限制。在本披露中也有可能的是,步骤也可以在逻辑上可能的情况下以不同的顺序执行。
107.必须注意,如说明书和所附权利要求书中所用,除非上下文另外明确指出,否则单数形式“一个/种”和“该”包括复数指示物。
108.本文披露了靶向抗原呈递细胞上的cd83的嵌合抗原受体(car)。还披露了经工程化以表达这些car的免疫效应细胞,如t细胞或自然杀伤(nk)细胞。表达这些car的car t细胞可以抑制同种异体反应供体细胞,如t细胞。因此,还披露了用于预防受试者中的gvhd的方法,该方法涉及过继转移披露的免疫效应细胞,这些免疫效应细胞被工程化以表达披露的cd83特异性car。
109.cd83特异性嵌合抗原受体(car)
110.car通常包含来自单克隆抗体(mab)的单链可变片段(scfv)的抗原识别结构域,该抗体具有参与淋巴细胞激活的跨膜信号传导基序(sadelain m,等人nat rev cancer[癌症自然评论]2003 3:35-45)。本文披露了cd83特异性嵌合抗原受体(car),它可以在免疫效应细胞中表达以抑制同种异体反应供体细胞。
[0111]
披露的car通常由三个结构域组成:胞外结构域、跨膜结构域和胞内结构域。胞外结构域包含cd83结合区并负责抗原识别。它还任选地包含信号肽(sp),使得car可以被糖基化并锚定在免疫效应细胞的细胞膜中。顾名思义,跨膜结构域(td)将胞外结构域与胞内结构域连接起来,并在由细胞表达时位于细胞膜内。胞内结构域是car的业务端,在抗原识别后向免疫效应细胞传递激活信号。例如,胞内结构域可以包含细胞内信号传导结构域(isd)和任选的共刺激信号传导区(csr)。
[0112]“信号传导结构域(sd)”通常包含基于免疫受体酪氨酸的激活基序(itam),当itam被磷酸化时,这些激活基序会激活信号传导级联。术语“共刺激信号传导区(csr)”是指来自共刺激蛋白受体的细胞内信号传导结构域,如cd28、41bb和icos,它们能够增强t细胞受体对t细胞的激活。
[0113]
在一些实施例中,胞内结构域包含sd或csr,而不是包含两者。在这些实施例中,含有所披露的car的免疫效应细胞仅在含有缺失结构域的另一个car(或t细胞受体)也结合其各自的抗原时才被激活。
[0114]
在一些实施例中,所披露的car由下式定义:
[0115]
sp-cd83-hg-tm-csr-sd;或
[0116]
sp-cd83-hg-tm-sd-csr;
[0117]
其中“sp”代表任选的信号肽,
[0118]
其中“cd83”代表cd83结合区,
[0119]
其中“hg”代表任选的铰链结构域,
[0120]
其中“tm”代表跨膜结构域,
[0121]
其中“csr”代表一个或多个共刺激信号传导区,
[0122]
其中“sd”代表信号传导结构域,并且
[0123]
其中
“‑”
代表肽键或接头。
[0124]
描述了另外的car构建体,例如,在fresnak ad,等人engineered t cells:the promise and challenges of cancer immunotherapy[工程化t细胞:癌症免疫疗法的前景和挑战].nat rev cancer.[癌症自然评论]2016年8月23日;16(9):566-81,针对这些car模型的传授内容将其通过引用以其全文并入。
[0125]
例如,car可以是truck、通用car、自驱动car、装甲car、自毁car、条件car、标记car、tencar、双car或scar。
[0126]
经工程化以抵抗免疫抑制的car t细胞(装甲car)可能经过基因修饰不再表达各种免疫检查点分子(例如,细胞毒性t淋巴细胞相关抗原4(ctla4)或程序性细胞死亡蛋白1(pd1)),具有免疫检查点开关受体,或者可以与阻断免疫检查点信号传导的单克隆抗体一起施用。
[0127]
可以使用通过电穿孔递送以编码car的rna来设计自毁car。可替代地,可基于基因修饰淋巴细胞或最近描述的通过小分子二聚体激活人半胱天冬酶9的系统中更昔洛韦与胸苷激酶的结合来实现t细胞的可诱导凋亡。
[0128]
默认情况下,条件car t细胞是无反应的或开关“关闭”,直到添加小分子来完成电路,从而实现信号1和信号2的完全转导,从而激活car t细胞。可替代地,可以对t细胞工程化以表达对随后施用的针对靶抗原的二抗具有亲和力的衔接子特异性受体。
[0129]
串联car(tancar)t细胞表达由两个连接的单链可变片段(scfvs)组成的单个car,这些片段具有与一个或多个细胞内共刺激结构域和cd3ζ结构域融合的不同亲和力。只有当靶细胞同时表达两个靶标时,才能实现tancar t细胞激活。
[0130]
双car t细胞表达两个具有不同配体结合靶标的单独的car;一个car仅包含cd3ζ结构域,而另一个car仅包含该一个或多个共刺激结构域。双car t细胞激活需要两个靶标的共表达。
[0131]
安全car(scar)由融合到细胞内抑制结构域的细胞外scfv组成。共表达标准car的scar t细胞只有在遇到具有标准car靶标但缺乏scar靶标的靶细胞时才会被激活。
[0132]
所披露的car的抗原识别结构域通常是scfv。然而,有很多替代方案。已经描述了来自天然t细胞受体(tcr)α和β单链的抗原识别结构域,以及简单的胞外结构域(例如,识别hiv感染细胞的cd4胞外结构域)和更奇特的识别成分,如连接的细胞因子(其导致识别带有细胞因子受体的细胞)。事实上,几乎任何以高亲和力结合给定靶标的东西都可以用作抗原识别区。
[0133]
胞内结构域是car的业务端,在抗原识别后向免疫效应细胞传递信号,激活免疫效应细胞的至少一项正常效应功能。例如,t细胞的效应子功能可以是细胞溶解活性或辅助活性,包括细胞因子的分泌。因此,胞内结构域可包含t细胞受体(tcr)的“细胞内信号传导结构域”和任选的共受体。虽然通常可以使用整个细胞内信号传导结构域,但在许多情况下不需要使用整个链。就使用细胞内信号传导结构域的截短部分而言,此类截短部分可用于代替完整链,只要它转导效应子功能信号即可。
[0134]
以刺激方式起作用的调节tcr复合物初级激活的细胞质信号传导序列可能包含称为基于免疫受体酪氨酸的激活基序(itam)的信号传导基序。含有itam的细胞质信号传导序
列的实例包括衍生自cd8、cd3ζ、cd3δ、cd3γ、cd3ε、cd32(fcγriia)、dap10、dap12、cd79a、cd79b、fcγriγ、fcγriiiγ、fcεriβ(fcerib)和fcεriγ(fcerig)的那些。
[0135]
在特定实施例中,细胞内信号传导结构域衍生自cd3ζ(cd3 zeta)(tcrζ,genbank accno.bag36664.1)。t细胞表面糖蛋白cd3ζ(cd3 zeta)链,也称为t细胞受体t3ζ链或cd247(分化簇247),是一种在人中由cd247基因编码的蛋白。
[0136]
第一代car通常具有来自cd3ζ链的细胞内结构域,其为内源性tcr信号的主要传递者。第二代car将来自各种共刺激蛋白受体(例如,cd28、41bb、icos)的细胞内信号传导结构域添加到car的胞内结构域,以向t细胞提供另外的信号。最近,第三代car组合了多个信号传导结构域以进一步增强效力。移植有这些car的t细胞已证明了独立于共刺激受体/配体相互作用的增强的扩增、激活、持久性和肿瘤根除效率(imai c,等人leukemia[白血病]2004 18:676-84;maher j,等人nat biotechnol[自然
·
生物技术]2002 20:70-5)。
[0137]
例如,car的胞内结构域可以设计为包含cd3ζ信号传导结构域本身或与在本发明的car上下文中有用的一种或多种任何其他所需细胞质结构域组合。例如,car的细胞质结构域可以包含cd3ζ链部分和共刺激信号传导区。共刺激信号传导区是指包含共刺激分子的细胞内结构域的car的一部分。共刺激分子是淋巴细胞对抗原的有效反应所需的抗原受体或其配体以外的细胞表面分子。此类分子的实例包括cd27、cd28、4-1bb(cd137)、ox40、cd30、cd40、icos、淋巴细胞功能相关抗原-1(lfa-1)、cd2、cd7、light、nkg2c、b7-h3和与cd123、cd8、cd4、b2c、cd80、cd86、dap10、dap12、myd88、btnl3和nkg2d特异性结合的配体。因此,虽然car主要以cd28作为共刺激信号传导元件为例,但其他共刺激元件可以单独使用或与其他共刺激信号传导元件组合使用。
[0138]
在一些实施例中,car包含铰链序列。铰链序列是促进抗体灵活性的氨基酸短序列(参见,例如,woof等人,nat.rev.immunol.[自然评论:免疫学],4(2):89-99(2004))。铰链序列可以位于抗原识别部分(例如,抗cd83scfv)和跨膜结构域之间。铰链序列可以是衍生自或获得自任何适合的分子的任何适合的序列。在一些实施例中,例如,铰链序列衍生自cd8a分子或cd28分子。
[0139]
跨膜结构域可以衍生自天然来源或合成来源。在来源是天然的情况下,该结构域可以衍生自任何膜结合或跨膜蛋白。例如,跨膜区可以衍生自t细胞受体、cd28、cd3ε、cd45、cd4、cd5、cd8(例如,cd8α、cd8β)、cd9、cd16、cd22、cd33、cd37、cd64、cd80、cd86、cd134、cd137、或cd154、kirds2、ox40、cd2、cd27、lfa-1(cd11a、cd18)、icos(cd278)、4-1bb(cd137)、gitr、cd40、baffr、hvem(lightr)、slamf7、nkp80(klrf1)、cd160、cd19、il2rβ、il2rγ、il7rα、itga1、vla1、cd49a、itga4、ia4、cd49d、itga6、vla-6、cd49f、itgad、cd11d、itgae、cd103、itgal、cd11a、lfa-1、itgam、cd11b、itgax、cd11c、itgb1、cd29、itgb2、cd18、lfa-1、itgb7、tnfr2、dnam1(cd226)、slamf4(cd244、2b4)、cd84、cd96(tactile)、ceacam1、crtam、ly9(cd229)、cd160(by55)、psgl1、cd100(sema4d)、slamf6(ntb-a、ly108)、slam(slamf1、cd150、ipo-3)、blame(slamf8)、selplg(cd162)、ltbr和pag/cbp的α、β或ζ链(即,至少包含其一个或多个跨膜区)。可替代地,跨膜结构域可以是合成的,在这种情况下,它将主要包含疏水残基,如亮氨酸和缬氨酸。在某些情况下,苯丙氨酸、色氨酸和缬氨酸的三联体会出现在合成跨膜结构域的每一端。短的寡核苷酸或多肽接头,如长度在2到10个氨基酸之间,可以形成跨膜结构域和car内质结构域之间的连接。
[0140]
在一些实施例中,car具有多于一个跨膜结构域,其可以是相同跨膜结构域的重复,或者可以是不同的跨膜结构域。
[0141]
在一些实施例中,car是多链car,如wo 2015/039523中所述,针对本传授内容将其通过引用并入。多链car可以在不同的跨膜多肽中包含单独的细胞外配体结合和信号传导结构域。信号传导结构域可以设计为在近膜位置组装,形成更接近天然受体的灵活结构,从而提供最佳的信号转导。例如,多链car可以包含fceriα链的一部分和fceriβ链的一部分,使得fceri链自发地二聚在一起形成car。
[0142]
下面的表1、2和3提供了可以在所披露的car中出现的cd83结合区、共刺激信号传导区和细胞内信号传导结构域的一些示例组合。
[0143][0144]
[0145]
[0146]
[0147]
[0148]
[0149][0150]
[0151]
[0152]
[0153]
[0154]
[0155]
[0156]
[0157]
[0158]
[0159]
[0160]
[0161]
[0162]
[0163]
[0164]
[0165]
[0166]
[0167]
[0168]
[0169]
[0170]
[0171]
[0172]
[0173]
[0174]
[0175]
[0176]
[0177]
[0178]
[0179]
[0180]
[0181]
[0182]
[0183]
[0184]
[0185]
[0186]
[0187]
[0188]
[0189]
[0190]
[0191]
[0192]
[0193]
[0194]
[0195]
[0196]
[0197]
[0198]
[0199]
[0200]
[0201]
[0202]
[0203]
[0204]
[0205]
[0206]
[0207]
[0208]
[0209]
[0210]
[0211]
[0212]
[0213]
[0214]
[0215]
[0216]
[0217]
[0218]
[0219]
[0220]
[0221]
[0222]
[0223]
[0224]
[0225]
[0226]
[0227]
[0228]
[0229]
[0230]
[0231]
[0232]
[0233]
[0234]
[0235]
[0236]
[0237]
[0238]
[0239]
[0240]
[0241]
[0242]
[0243]
[0244]
[0245]
[0246]
[0247]
[0248]
[0249]
[0250]
[0251]
[0252]
[0253]
[0254]
[0255]
[0256]
[0257]
[0258]
[0259]
[0260]
[0261]
[0262]
[0263]
[0264]
[0265]
[0266]
[0267]
[0268]
[0269]
[0270]
[0271]
[0272]
[0273]
[0274]
[0275]
[0276]
[0277]
[0278]
[0279]
[0280]
[0281]
[0282]
[0283]
[0284]
[0285][0286]
[0287][0288][0289]
[0290]
[0291]
[0292]
[0293]
[0294]
[0295]
[0296]
[0297]
[0298]
[0299]
[0300][0301]
在一些实施例中,抗cd83结合剂是单链可变片段(scfv)抗体。抗cd83scfv的亲和力/特异性在很大程度上由重(vh)和轻(v
l
)链中的互补决定区(cdr)内的特定序列驱动。每个vh和v
l
序列将具有三个cdr(cdr1、cdr2、cdr3)。
[0302]
在一些实施例中,抗cd83结合剂衍生自天然抗体,如单克隆抗体。在某些情况下,抗体是人的。在某些情况下,抗体发生了改变,使其在施用于人时免疫原性降低。例如,改变包含从选自由嵌合、人源化、cdr移植、去免疫化和框架氨基酸突变组成的组中的一种或多种技术,以对应于最接近的人生殖系序列。
[0303]
还披露了靶向cd83和至少一种另外的抗原的双特异性car。还披露了设计为仅与另一种结合不同抗原的car联合工作的car。例如,在这些实施例中,所披露的car的胞内结构域可以仅包含信号传导结构域(sd)或共刺激信号传导区(csr),而不是包含两者。如果第二个car(或内源性t细胞)被激活,它会提供缺失信号。例如,如果披露的car包含sd但不包含csr,则包含该car的免疫效应细胞只有在另一个包含csr的car(或t细胞)结合其各自的抗原时才被激活。同样,如果披露的car包含csr但不包含sd,则包含该car的免疫效应细胞只有在另一个包含sd的car(或t细胞)结合其各自的抗原时才被激活。
[0304]
核酸和载体
[0305]
还披露了编码所披露的cd83特异性car的多核苷酸和多核苷酸载体,其允许cd83特异性car在所披露的免疫效应细胞中表达。
[0306]
编码所披露的car及其区域的核酸序列可以使用本领域已知的重组方法获得,诸如像通过从表达该基因的细胞中筛选文库,通过从已知包含该基因的载体衍生该基因,或通过使用标准技术直接从含有该基因的细胞和组织中分离。可替代地,感兴趣的基因可以合成产生,而不是克隆。
[0307]
编码car的核酸的表达通常通过将编码car多肽的核酸可操作地连接至启动子,并将构建体掺入到表达载体中来实现。典型的克隆载体含有转录和翻译终止子、起始序列和可用于调节所需核酸序列表达的启动子。
[0308]
披露的核酸可以克隆到多种类型的载体中。例如,可以将核酸克隆到载体中,包括但不限于质粒、噬菌粒、噬菌体衍生物、动物病毒和粘粒。特别感兴趣的载体包括表达载体、复制载体、探针生成载体和测序载体。
[0309]
此外,表达载体可以以病毒载体的形式提供给细胞。病毒载体技术在本领域中是众所周知的并且例如在sambrook等人(2001,molecular cloning:a laboratory manual[分子克隆:实验室手册],冷泉港实验室,纽约)以及其他病毒学和分子生物学手册中有所描述。可用作载体的病毒包括但不限于逆转录病毒、腺病毒、腺相关病毒、疱疹病毒和慢病毒。一般而言,合适的载体包含在至少一种生物体中具有功能的复制起点、启动子序列、方
便的限制性内切核酸酶位点和一种或多种可选择标记。在一些实施例中,多核苷酸载体是慢病毒或逆转录病毒载体。
[0310]
已经开发了许多基于病毒的系统用于将基因转移到哺乳动物细胞中。例如,逆转录病毒为基因递送系统提供了一个方便的平台。可以使用本领域已知的技术将选定的基因插入载体中并包装在逆转录病毒颗粒中。然后可以在体内或离体分离重组病毒并将其递送至受试者的细胞。
[0311]
适合的启动子的一个实例是立即早期巨细胞病毒(cmv)启动子序列。该启动子序列是强组成型启动子序列,能够驱动与其操作性连接的任何多核苷酸序列的高水平表达。适合的启动子的另一个实例是延伸生长因子-1α(ef-1α)。然而,也可以使用其他组成型启动子序列,包括但不限于猿猴病毒40(sv40)早期启动子、mnd(骨髓增殖性肉瘤病毒)启动子、小鼠乳腺肿瘤病毒(mmtv)、人免疫缺陷病毒(hiv)长末端重复(ltr)启动子、momulv启动子、禽白血病病毒启动子、爱泼斯坦-巴尔病毒立即早期启动子、劳斯肉瘤病毒启动子,以及人基因启动子,诸如但不限于肌动蛋白启动子、肌球蛋白启动子、血红蛋白启动子和肌酸激酶启动子。可替代地,启动子可以是诱导型启动子。诱导型启动子的实例包括但不限于金属硫氨酸启动子、糖皮质激素启动子、孕酮启动子和四环素启动子。
[0312]
另外的启动子元件,例如增强子,调节转录起始的频率。通常,这些启动子位于起始位点上游30-110bp的区域,尽管最近已显示许多启动子也包含起始位点下游的功能元件。启动子元件之间的间距通常是灵活的,因此当元件相对于彼此倒置或移动时,启动子功能得以保留。
[0313]
为了评估car多肽或其部分的表达,待引入细胞中的表达载体还可以包含可选择标记基因或报告基因或两者,以促进从试图通过病毒载体转染或感染的细胞群中鉴定和选择表达细胞。在其他方面,可选择标记可携带在单独的dna片段上并用于共转染程序。可选择标记和报告基因的两侧都可以有适当的调节序列,以使其能够在宿主细胞中表达。有用的可选择标记包括,例如,抗生素抗性基因。
[0314]
报告基因用于鉴定可能转染的细胞和评估调节序列的功能。一般而言,报告基因是不存在于受体生物体或组织中或不由受体生物体或组织表达的基因,并且其编码多肽,该多肽的表达通过一些容易检测的特性(例如,酶活性)来证明。在将dna引入受体细胞后的适合的时间测定报告基因的表达。适合的报告基因可以包括编码荧光素酶、β-半乳糖苷酶、氯霉素乙酰转移酶、分泌型碱性磷酸酶或绿色荧光蛋白基因的基因。合适的表达系统是众所周知的并且可以使用已知技术制备或商业获得。通常,具有显示最高水平的报告基因表达的最小5'侧翼区的构建体被鉴定为启动子。此类启动子区可与报告基因连接并用于评估试剂调节启动子驱动的转录的能力。
[0315]
将基因引入和表达到细胞中的方法是本领域已知的。在表达载体的情况下,可以通过本领域中的任何方法将载体容易地引入宿主细胞,例如,哺乳动物、细菌、酵母或昆虫细胞。例如,表达载体可以通过物理、化学或生物学方式转移到宿主细胞中。
[0316]
将多核苷酸引入宿主细胞的物理方法包括磷酸钙沉淀、脂质转染、粒子轰击、显微注射、电穿孔等。用于产生包含载体和/或外源核酸的细胞的方法是本领域众所周知的。参见,例如,sambrook等人(2001,molecular cloning:a laboratory manual[分子克隆:实验室手册],冷泉港实验室,纽约)。
[0317]
将感兴趣的多核苷酸引入宿主细胞的生物学方法包括使用dna和rna载体。病毒载体,尤其是逆转录病毒载体,已成为将基因插入哺乳动物(例如,人细胞)中最广泛使用的方法。
[0318]
用于将多核苷酸引入宿主细胞的化学方法包括胶体分散系统,如大分子复合物、纳米胶囊、微球、珠和基于脂质的系统(包括水包油乳液、胶束、混合胶束和脂质体)。用作体外和体内递送媒介物的示例性胶体系统是脂质体(例如,人工膜囊泡)。
[0319]
在使用非病毒递送系统的情况下,示例性递送媒介物是脂质体。在另一个方面,核酸可以与脂质缔合。与脂质缔合的核酸可以包封在脂质体的水性内部,散布在脂质体的脂质双层中,通过与脂质体和寡核苷酸两者缔合的连接分子附接至脂质体,包埋在脂质体中,与脂质体复合,分散在含有脂质的溶液中,与脂质混合,与脂质组合,作为悬浮液包含在脂质中,包含或与胶束复合,或以其他方式与脂质缔合。脂质、脂质/dna或脂质/表达载体相关组合物不限于溶液中的任何特定结构。例如,它们可以以双层结构、胶束形式或具有“塌陷”结构的形式存在。它们也可以简单地散布在溶液中,可能形成大小或形状不均匀的聚集体。脂质是脂肪物质,可以是天然存在的或合成的脂质。例如,脂质包括天然存在于细胞质中的脂肪滴以及含有长链脂肪烃及其衍生物的化合物类别,如脂肪酸、醇、胺、氨基醇和醛。适合使用的脂质可以从商业来源获得。例如,二聚豆蔻基卵磷脂(“dmpc”)可以从密苏里州圣路易斯的西格玛公司(sigma)获得;二鲸蜡醇磷酸酯(“dcp”)可以k&k实验室(纽约州普莱恩维尤)获得;胆固醇(“choi”)可以从calbiochem-behring获得;二肉豆蔻酰磷脂酰甘油(“dmpg”)和其他脂质可以从阿凡提极性脂质公司(avanti polar lipids)(阿拉巴马州伯明翰)获得。
[0320]
免疫效应细胞
[0321]
还披露了经工程化以表达所披露的car的免疫效应细胞(在本文中也称为“car-t细胞”)。这些细胞优选从待治疗的受试者获得(即,是自体的)。然而,在一些实施例中,使用免疫效应细胞系或供体效应细胞(同种异体)。然而,在其他实施例中,免疫效应细胞不是hla匹配的。免疫效应细胞可从多种来源获得,包括外周血单核细胞、骨髓、淋巴结组织、脐带血、胸腺组织、感染部位的组织、腹水、胸腔积液、脾脏组织、和肿瘤。可以使用本领域技术人员已知的许多技术,如ficoll
tm
分离,由从受试者收集的血液中获得免疫效应细胞。例如,来自个体循环血液的细胞可以通过单采术获得。在一些实施例中,免疫效应细胞通过裂解红细胞和消耗单核细胞从外周血淋巴细胞中分离,例如通过percoll
tm
梯度离心或通过逆流离心淘析。可以通过阳性或阴性选择技术进一步分离免疫效应细胞的特定亚群。例如,可以使用针对阳性选择的细胞特有的表面标记的抗体组合来分离免疫效应细胞,例如,通过与抗体缀合珠孵育足够时间以阳性选择所需免疫效应细胞。可替代地,免疫效应细胞群的富集可以通过使用针对负选择细胞特有的表面标记的抗体组合进行负选择来完成。
[0322]
在一些实施例中,免疫效应细胞包含参与保护身体免受感染性疾病和外来物质的任何白细胞。例如,免疫效应细胞可以包含淋巴细胞、单核细胞、巨噬细胞、树突状细胞、肥大细胞、嗜中性粒细胞、嗜碱性粒细胞、嗜酸性粒细胞或其任何组合。例如,免疫效应细胞可以包含t淋巴细胞。
[0323]
通过在细胞表面存在t细胞受体(tcr),可以将t细胞或t淋巴细胞与其他淋巴细胞,如b细胞和自然杀伤细胞(nk细胞)区分开来。它们被称为t细胞,因为它们在胸腺中成熟
(尽管有些也在扁桃体中成熟)。有几个t细胞亚群,每个亚群都有不同的功能。
[0324]
t辅助细胞(th细胞)在免疫过程中协助其他白细胞,包括b细胞成熟为浆细胞和记忆b细胞,以及细胞毒性t细胞和巨噬细胞的激活。这些细胞也被称为cd4+ t细胞,因为它们在其表面表达cd4糖蛋白。辅助t细胞在被mhc ii类分子呈递肽抗原时被激活,这些分子在抗原呈递细胞(apc)的表面上表达。一旦被激活,它们会迅速分裂并分泌称为细胞因子的小蛋白,调节或协助主动免疫反应。这些细胞可以分化为几种亚型之一,包括th1、th2、th3、th17、th9、或t
fh
,它们分泌不同的细胞因子以促进不同类型的免疫反应。
[0325]
细胞毒性t细胞(tc细胞、或ctl)破坏病毒感染的细胞和肿瘤细胞,并且还与移植排斥有关。这些细胞也被称为cd8
+ t细胞,因为它们在其表面表达cd8糖蛋白。这些细胞通过与存在于所有有核细胞表面的mhc i类分子相关的抗原缔合来识别它们的靶标。通过调节性t细胞分泌的il-10、腺苷和其他分子,cd8+细胞可以失活到无能状态,从而预防自身免疫性疾病。
[0326]
记忆t细胞是在感染消退后会长期存在的抗原特异性t细胞亚群。当再次暴露于同源抗原时,它们迅速扩展到大量效应t细胞,从而为免疫系统提供针对过去感染的“记忆”。记忆细胞可以是cd4
+
或cd8
+
。记忆t细胞通常表达细胞表面蛋白cd45ro。
[0327]
调节性t细胞(t
reg
细胞),以前被称为抑制性t细胞,对维持免疫耐受至关重要。它们的主要作用是在免疫反应结束时关闭t细胞介导的免疫,并抑制在胸腺中逃脱负选择过程的自身反应性t细胞。已经描述了两大类cd4
+
t
reg
细胞——天然存在的t
reg
细胞和适应性t
reg
细胞。
[0328]
自然杀伤t(nkt)细胞(不要与自然杀伤(nk)细胞混淆)连接适应性免疫系统和先天免疫系统。与识别主要组织相容性复合物(mhc)分子呈递的肽抗原的常规t细胞不同,nkt细胞识别由称为cd1d的分子呈递的糖脂抗原。
[0329]
在一些实施例中,t细胞包含cd4+细胞的混合物。在其他实施例中,t细胞基于细胞表面表达富集一个或多个亚群。例如,在一些情况下,t包含细胞毒性cd8
+ t淋巴细胞。在一些实施例中,t细胞包含γδt细胞,其具有独特的t细胞受体(tcr),其具有一条γ链和一条δ链而不是α和β链。
[0330]
自然杀伤(nk)细胞是cd56
+
cd3-大颗粒淋巴细胞,其可以杀伤病毒感染和转化的细胞,并构成先天免疫系统的关键细胞亚群(godfrey j,等人leuk lymphoma 2012 53:1666-1676)。与细胞毒性cd8
+ t淋巴细胞不同,nk细胞对肿瘤细胞具有细胞毒性,无需预先致敏,还可以根除mhc-i阴性细胞(narni-mancinelli e,等人int immunol[国际免疫学]2011 23:427-431)。nk细胞是更安全的效应细胞,因为它们可以避免细胞因子风暴(morgan ra,等人mol ther[分子治疗]2010 18:843-851)、肿瘤溶解综合征(porter dl,等人n engl j med[新英格兰医学杂志]2011 365:725-733)和中靶脱瘤效应的潜在致命并发症。
[0331]
治疗方法
[0332]
表达所披露的car的免疫效应细胞抑制同种异体反应性供体细胞(如t细胞),并预防gvhd。因此,所披露的car可施用于有gvhd风险的任何受试者。在一些实施例中,受试者接受骨髓移植并且所披露的car修饰的免疫效应细胞抑制供体t细胞或树突状细胞的同种异体反应性。
[0333]
所披露的car修饰的免疫效应细胞可以单独施用,也可以作为药物组合物与稀释
剂和/或其他组分(如il-2、il-15或其他细胞因子)或细胞群组合施用。
[0334]
在一些实施例中,所披露的car修饰的免疫效应细胞与er应激阻断(靶向ire-1/xbp-1途径的化合物(例如,b-i09))组合施用。在一些实施例中,所披露的car修饰的免疫效应细胞与以下项组合施用:jak2抑制剂、stat3抑制剂、极光激酶抑制剂、mtor抑制剂或其任何组合。
[0335]
简而言之,药物组合物可包含如本文所述的靶细胞群,以及一种或多种药学或生理学可接受的载体、稀释剂或赋形剂。此类组合物可包含缓冲剂(如中性缓冲盐水、磷酸盐缓冲盐水等);碳水化合物(如葡萄糖、甘露糖、蔗糖或葡聚糖、甘露醇);蛋白质;多肽或氨基酸(如甘氨酸);抗氧化剂;螯合剂(如edta或谷胱甘肽);佐剂(例如,氢氧化铝);和防腐剂。在一些实施例中,用于所披露的方法的组合物被配制用于静脉内施用。可以以任何适合治疗mm的方式施用药物组合物。施用的数量和频率将由如患者的病症和患者疾病的严重程度等因素决定,尽管适当的剂量可以通过临床试验来确定。
[0336]
当指示“治疗量”时,本发明组合物的准确施用量可由医师考虑患者(受试者)的年龄、体重、移植程度和病症的个体差异来确定。通常可以说包含本文所述的t细胞的药物组合物可以104至109个细胞/kg体重,如105至106个细胞/kg体重的剂量施用,包括那些范围内的所有整数值。t细胞组合物也可以这些剂量多次施用。可以通过使用免疫疗法中通常已知的输注技术来施用细胞(参见,例如,rosenberg等人,new eng.j.of med.[新英格兰医学杂志]319:1676,1988)。医学领域的技术人员可以通过监测患者的疾病迹象并相应地调整治疗来容易地确定特定患者的最佳剂量和治疗方案。
[0337]
在某些实施例中,可能需要将激活的t细胞施用于受试者,然后再抽血(或进行单采术),根据所披露的方法从其中激活t细胞,并将这些激活的和扩增的t细胞重新注入患者。该过程可以每隔几周进行多次。在某些实施例中,t细胞可以从10cc至400cc的抽血中被激活。在某些实施例中,t细胞从20cc、30cc、40cc、50cc、60cc、70cc、80cc、90cc、或100cc的抽血中被激活。使用该多次抽血/多次回输方案可用于选择某些t细胞群。
[0338]
披露的组合物的施用可以任何方便的方式进行,包括通过注射、输血或植入。本文所述的组合物可以皮下、皮内、结内、髓内、肌内、通过静脉内(i.v.)注射或腹膜内施用于患者。在一些实施例中,所披露的组合物通过皮内或皮下注射施用于患者。在一些实施例中,所披露的组合物通过静脉内注射施用。也可以将组合物直接注射到移植部位。
[0339]
在某些实施例中,将所披露的car修饰的免疫效应细胞与任何数量的相关治疗方式(包括但不限于沙利度胺、地塞米松、硼替佐米和来那度胺)联合(例如,之前、同时或之后)施用于患者。在进一步的实施例中,car修饰的免疫效应细胞可以与以下项组合:化学疗法、放射、免疫抑制剂(如环孢菌素、硫唑嘌呤、甲氨蝶呤、霉酚酸酯和fk506)、抗体、或其他免疫消融剂(如cam path、抗cd3抗体或其他抗体疗法)、细胞毒素、氟达利滨、环孢菌素、fk506、雷帕霉素、霉酚酸、类固醇、fr901228、细胞因子和辐射。在一些实施例中,将car修饰的免疫效应细胞与骨髓移植、使用任一种化疗剂(如氟达拉滨)的t细胞消融疗法、外束放射疗法(xrt)、环磷酰胺、或抗体(如okt3或campath)联合(例如,之前、同时或之后)施用于患者。在另一个实施例中,本发明的细胞组合物在b细胞消融疗法(如与cd20反应的试剂,例如,美罗华)之后施用。例如,在一些实施例中,受试者可以接受高剂量化学疗法的标准治疗,然后进行外周血干细胞移植。在某些实施例中,移植后,受试者接受本发明扩增的免疫
细胞的输注。在另一个实施例中,在手术之前或之后施用扩增的细胞。
[0340]
将car-t细胞作为一种形式的“活体治疗剂”的一个主要问题是它们在体内的可操作性及其潜在的免疫刺激副作用。为了更好地控制car-t疗法并预防不必要的副作用,已经工程化了多种特征,包括关闭开关、安全机制和条件控制机制。例如,自毁和标记/加标签的car-t细胞都被工程化以具有促进表达car的t细胞清除的“关闭开关”。自毁car-t包含car,但也被工程化以表达在施用外源分子后可诱导的促凋亡自杀基因或“消除基因”。多种自杀基因可用于此目的,包括hsv-tk(单纯疱疹病毒胸苷激酶)、fas、icasp9(诱导型半胱天冬酶9)、cd20、myc tag和截短的egfr(内皮生长因子受体)。例如,hsk会将前药更昔洛韦(gcv)转化为gcv-三磷酸盐,后者将自身掺入到复制的dna中,最终导致细胞死亡。icasp9是一种嵌合蛋白,含有可结合小分子ap1903的fk506结合蛋白的组分,导致半胱天冬酶9二聚化和细胞凋亡。然而,标记/加标签的car-t细胞是一种具有car但也被工程化以表达选择标记的细胞。针对该选择标记施用mab将促进car-t细胞的清除。截短的egfr是抗egfr mab的一种此类可靶向抗原,西妥昔单抗的施用可促进car-t细胞的消除。为具有这些特征而建立的car也称为
‘
可切换car’的scar,以及
‘
可调节car’的rcar。“安全car”,也称为“抑制性car”(icar),被工程化以表达两个抗原结合结构域。这些细胞外结构域之一针对第一抗原并与细胞内共刺激和刺激结构域结合。然而,第二个细胞外抗原结合结构域对正常组织具有特异性,并与细胞内检查点结构域(如ctla4、pd1或cd45)结合。将多个细胞内抑制结构域掺入到icar中也是可能的。一些可能提供这些抑制结构域的抑制分子包括b7-h1、b7-1、cd160、pih、2b4、ceacam(ceacam-1.ceacam-3、和/或ceacam-5)、lag-3、tigit、btla、lair1和tgfβ-r。在正常组织存在的情况下,该第二个抗原结合结构域的刺激将起到抑制car的作用。需要注意的是,由于该双重抗原特异性,icar也是一种双特异性car-t细胞。安全car-t工程化增强了car-t细胞对组织的特异性,并且在某些正常组织可能表达极低水平的抗原的情况下是有利的,这将导致标准car的脱靶效应(morgan 2010)。条件car-t细胞表达与细胞内共刺激结构域和单独的细胞内共刺激物相连的细胞外抗原结合结构域。将共刺激和刺激结构域序列以这样一种方式工程化,即在施用外源分子后,所得蛋白质将在细胞内聚集在一起以完成car回路。通过这种方式,可以调节car-t激活,甚至可能针对特定患者进行“微调”或个性化。与双car设计类似,刺激结构域和共刺激结构域在条件car中不活动时在物理上是分开的;出于该原因,这些也被称为“分裂氏car”。
[0341]
通常,car-t细胞是使用α-βt细胞建立的,但也可以使用γ-δt细胞。在一些实施例中,用于产生car-t细胞的所述car构建体、结构域和工程化特征可以类似地用于产生其他类型的表达car的免疫细胞,包括nk(自然杀伤)细胞、b细胞、肥大细胞、髓源吞噬细胞和nkt细胞。可替代地,可以建立表达car的细胞以具有t细胞和nk细胞的特性。在另一个实施例中,用car转导的可以是自体的或同种异体的。
[0342]
可以使用几种不同的car表达方法,包括逆转录病毒转导(包括γ-逆转录病毒)、慢病毒转导、转座子/转座酶(sleeping beauty和piggybac系统)和信使rna转移介导的基因表达。基因编辑(基因插入或基因缺失/破坏)对于工程化car-t细胞的可能性也变得越来越重要。crispr-cas9、zfn(锌指核酸酶)和talen(转录激活因子样效应核酸酶)系统是生成car-t细胞的三种潜在方法。
[0343]
定义
[0344]
术语“氨基酸序列”是指代表氨基酸残基的缩写、字母、字符或单词的列表。此处使用的氨基酸缩写是氨基酸的常规单字母代码,表示如下:a,丙氨酸;b,天冬酰胺或天冬氨酸;c,半胱氨酸;d天冬氨酸;e,谷氨酰胺,谷氨酸;f,苯丙氨酸;g,甘氨酸;h,组氨酸;i,异亮氨酸;k,赖氨酸;l,亮氨酸;m,蛋氨酸;n,天冬酰胺;p,脯氨酸;q,谷氨酰胺;r,精氨酸;s,丝氨酸;t,苏氨酸;v,缬氨酸;w,色氨酸;y,酪氨酸;z,谷氨酰胺或谷氨酸。
[0345]
术语“抗体”是指免疫球蛋白、其保持特异性结合能力的衍生物、以及具有与免疫球蛋白结合结构域同源或很大程度上同源的结合结构域的蛋白。这些蛋白可以衍生自天然来源,或部分或全部合成产生。抗体可以是单克隆或多克隆的。抗体可以是来自任何物种的任何免疫球蛋白类别的成员,包括任何人类类别:igg、igm、iga、igd和ige。在示例性实施例中,与本文所述的方法和组合物一起使用的抗体是igg类的衍生物。除了完整的免疫球蛋白分子之外,术语“抗体”还包括那些免疫球蛋白分子的片段或聚合物,以及选择性结合靶抗原的免疫球蛋白分子的人或人源化形式。
[0346]
术语“抗体片段”是指小于全长的抗体的任何衍生物。在示例性实施例中,抗体片段保留了全长抗体特异性结合能力的至少重要部分。抗体片段的实例包括但不限于fab、fab'、f(ab')2、scfv、fv、dsfv双抗体、fc和fd片段。抗体片段可以通过任何方式产生。例如,抗体片段可以通过完整抗体的片段化酶促或化学产生,它可以从编码部分抗体序列的基因重组产生,或者它可以完全或部分合成产生。抗体片段可以任选地是单链抗体片段。可替代地,该片段可包含连接在一起的多条链,例如,通过二硫键。该片段还可以任选地是多分子复合物。功能性抗体片段通常包含至少约50个氨基酸,并且更通常包含至少约200个氨基酸。
[0347]
术语“抗原结合位点”是指特异性结合抗原表位的抗体区。
[0348]
术语“适体”是指与特定靶分子结合的寡核酸或肽分子。这些分子通常选自随机序列库。选择的适体能够适应独特的三级结构并以高亲和力和特异性识别靶分子。“核酸适体”是dna或rna寡核酸,其通过其构象与靶分子结合,从而抑制该分子的功能。核酸适体可由dna、rna或其组合构成。“肽适体”是一种组合蛋白分子,其可变肽序列插入恒定支架蛋白中。肽适体的鉴定通常在严格的酵母双杂交条件下进行,这提高了所选肽适体在细胞内环境中稳定表达和正确折叠的可能性。
[0349]
术语“载体”是指化合物、组合物、物质或结构,当与化合物或组合物组合时,有助于或促进化合物或组合物的制备、储存、施用、递送、有效性、选择性或任何其他特征用于其预期用途或目的。例如,可以选择载体以使活性成分的任何降解最小化并且使受试者中的任何不利副作用最小化。
[0350]
术语“嵌合分子”是指通过连接两个或多个以其天然状态分别存在的分子而建立的单个分子。单个嵌合分子具有其所有组成分子所需的功能。一种类型的嵌合分子是融合蛋白。
[0351]
术语“工程化抗体”是指至少包含抗体片段的重组分子,该抗体片段包含衍生自抗体重链和/或轻链可变结构域的抗原结合位点,并且可以任选地包含来自任何ig类别(例如iga、igd、ige、igg、igm和igy)的抗体的可变和/或恒定结构域的全部或部分。
[0352]
术语“表位”是指抗体优先和特异性结合的抗原区。单克隆抗体优先结合可以在分子上进行定义的分子的单个特定表位。在本发明中,多个表位可以被多特异性抗体识别。
[0353]
术语“融合蛋白”是指两个或多个多肽通过一个多肽的氨基末端和另一个多肽的羧基末端之间形成的肽键连接形成的多肽。融合蛋白可以通过组成多肽的化学偶联形成,或者它可以表达为来自编码单个连续融合蛋白的核酸序列的单个多肽。单链融合蛋白是具有单个连续多肽骨架的融合蛋白。可以使用分子生物学中的常规技术将两个框内基因连接成单个核酸,然后在产生融合蛋白的条件下在适合的宿主细胞中表达该核酸,可以制备融合蛋白。
[0354]
术语“fab片段”是指包含通过用木瓜蛋白酶切割抗体产生的抗原结合位点的抗体片段,其在h链间二硫键n末端的铰链区切割并产生来自一个抗体分子的两个fab片段。
[0355]
术语“f(ab')2片段”是指包含两个抗原结合位点的抗体片段,其通过用胃蛋白酶切割抗体分子产生,该酶在铰链区c末端切割至h链间二硫键。
[0356]
术语“fc片段”是指包含其重链恒定结构域的抗体片段。
[0357]
术语“fv片段”是指包含其重链和轻链可变结构域的抗体片段。
[0358]“基因构建体”是指核酸,如载体、质粒、病毒基因组等,其包括多肽的“编码序列”或以其他方式可转录成生物活性rna(例如,反义、诱饵、核酶)等),可以转染到细胞中,例如在某些实施例中,哺乳动物细胞,并且可以引起编码序列在用构建体转染的细胞中表达。基因构建体可以包括与编码序列以及内含子序列、多聚腺苷酸化位点、复制起点、标记基因等可操作地连接的一种或多种调节元件。
[0359]
术语“同一性”是指两个核酸分子或多肽之间的序列同一性。同一性可以通过比较每个序列中的位置来确定,为了比较的目的,这些位置可以对齐。当比较序列中的一个位置被相同的碱基占据时,则该位置的分子是相同的。核酸或氨基酸序列之间的相似性或同一性程度是核酸序列共有的位置处相同或匹配的核苷酸数量的函数。可以使用各种比对算法和/或程序来计算两个序列之间的同一性,包括fasta或blast,它们可作为gcg序列分析包(威斯康星州麦迪逊威斯康星大学)的一部分获得,并可以以例如默认设置使用。例如,考虑了与本文所述的特定多肽具有至少70%、85%、90%、95%、98%或99%同一性并且优选地表现出基本上相同的功能的多肽,以及编码此类多肽的多核苷酸。除非另有指示,否则相似性评分将基于blosum62的使用。当使用blastp时,相似性百分比基于blastp阳性评分,而序列同一性百分比基于blastp同一性评分。blastp“同一性”显示了相同的高评分序列对中总残基的数量和分数;并且blastp“阳性”显示比对评分具有阳性值且彼此相似的残基的数量和分数。本披露考虑并涵盖了与本文所披露的氨基酸序列具有这些程度的同一性或相似性或者任何中间程度的同一性或相似性的氨基酸序列。相似多肽的多核苷酸序列是使用遗传密码推断的,并且可以通过常规方法获得,特别是通过使用遗传密码反向翻译其氨基酸序列来获得。
[0360]
术语“接头”是本领域公认的并且是指连接两种化合物(如两种多肽)的分子或分子组。接头可以由单个连接分子组成或可以包含连接分子和间隔子分子,旨在将连接分子和化合物分开特定距离。
[0361]
术语“多价抗体”是指包含多于一个抗原识别位点的抗体或工程化抗体。例如,“二价”抗体具有两个抗原识别位点,而“四价”抗体具有四个抗原识别位点。术语“单特异性”、“双特异性”、“三特异性”、“四特异性”等是指存在于多价抗体中的不同抗原识别位点特异性的数量(与抗原识别位点的数量形成对照)。例如,“单特异性”抗体的抗原识别位点都结
合相同的表位。“双特异性”抗体具有至少一个结合第一表位的抗原识别位点和至少一个结合不同于第一表位的第二表位的抗原识别位点。“多价单特异性”抗体具有多个抗原识别位点,它们都结合相同的表位。“多价双特异性”抗体具有多个抗原识别位点,其中一些数量结合第一表位,而一些数量结合与第一表位不同的第二表位。
[0362]
术语“核酸”是指天然或合成分子,其包含单个核苷酸或者通过一个核苷酸的3'位置处的磷酸基团连接到另一个核苷酸的5'端的两个或更多个核苷酸。核酸不受长度限制,因此核酸可以包括脱氧核糖核酸(dna)或核糖核酸(rna)。
[0363]
术语“可操作地连接到”是指核酸与另一核酸序列的功能关系。启动子、增强子、转录和翻译终止位点以及其他信号序列是与其他序列可操作连接的核酸序列的实例。例如,dna与转录控制元件的可操作连接是指dna和启动子之间的物理和功能关系,使得这种dna的转录由特异性识别、结合和转录dna的rna聚合酶从启动子开始。
[0364]
术语“肽”、“蛋白”和“多肽”可互换使用,是指包含两个或多个氨基酸的天然或合成分子,这些氨基酸通过一个氨基酸的羧基连接到另一个氨基酸的α氨基。
[0365]
术语“药学上可接受的”是指在合理医学判断的范围内适合于与人类和动物的组织接触使用而无过多毒性、刺激、过敏反应或其他问题或并发症,与合理的效益/风险比相称的那些化合物、材料、组合物、和/或剂型。
[0366]
术语“多肽片段”或“片段”,当用于提及特定多肽时,是指与参考多肽本身相比氨基酸残基缺失但剩余氨基酸序列通常与参考多肽相同的多肽。此类缺失可发生在参考多肽的氨基末端或羧基末端,或可替代地两者兼而有之。片段通常至少约5、6、8或10个氨基酸长,至少约14个氨基酸长,至少约20、30、40或50个氨基酸长,至少约75个氨基酸长,或至少约100、150、200、300、500或更多个氨基酸长。片段可以保留参考多肽的一种或多种生物活性。在各种实施例中,片段可以包含参考多肽的酶活性和/或相互作用位点。在另一个实施例中,片段可以具有免疫原性。
[0367]
术语“蛋白质结构域”是指蛋白的一部分、蛋白的多个部分或显示结构完整性的整个蛋白;该确定可以基于蛋白的一部分、蛋白的多部分或整个蛋白的氨基酸组成。
[0368]
术语“单链可变片段或scfv”是指其中重链结构域和轻链结构域连接的fv片段。一个或多个scfv片段可以连接到其他抗体片段(如重链或轻链的恒定结构域)以形成具有一个或多个抗原识别位点的抗体构建体。
[0369]
如本文所用,“间隔子”是指连接包括融合蛋白在内的蛋白的肽。通常,间隔子除了连接蛋白或保持它们之间的一些最小距离或其他空间关系之外没有特定生物活性。然而,可以选择间隔子的组成氨基酸以影响分子的某些特性,如分子的折叠、净电荷、或疏水性。
[0370]
如本文所用,术语“特异性结合”,当指多肽(包括抗体)或受体时,指的是确定蛋白或多肽或受体在蛋白和其他生物制品的异质群体中的存在的结合反应。因此,在指定条件下(例如,在抗体情况下的免疫测定条件),当特定配体或抗体未与样品中存在的其他蛋白或者在生物体中配体或抗体可能接触的其他蛋白大量结合时,该特定配体或抗体“特异性结合”到其特定“靶标”(例如抗体特异性结合到内皮抗原)。通常,“特异性结合”第二分子的第一分子与第二分子具有大于约105m-1
(例如,106m-1
、107m-1
、108m-1
、109m-1
、10
10
m-1
、10
11
m-1
、和10
12
m-1
或更多)的亲和常数(ka)。
[0371]
如本文所用,术语“特异性递送”是指分子与携带特定靶分子或标记的细胞或组织
的优先缔合,而不是与缺乏该靶分子的细胞或组织的优先结合。当然,人们认识到分子和非靶细胞或组织之间可能发生某种程度的非特异性相互作用。然而,特异性递送可以区分为通过对靶分子的特异性识别来介导。通常,特异性递送导致所递送的分子与携带靶分子的细胞之间的缔合比递送的分子与缺乏靶分子的细胞之间的缔合强得多。
[0372]
术语“受试者”是指作为施用或治疗靶标的任何个体。受试者可以是脊椎动物,例如,哺乳动物。因此,受试者可以是人或兽医患者。术语“患者”是指正在接受临床医生(例如,医师)治疗的受试者。
[0373]
术语“治疗有效”是指所用组合物的量足以改善疾病或障碍的一种或多种原因或症状。该改善只需要降低或改变,不一定要消除。
[0374]
术语“转化”和“转染”是指将核酸例如表达载体引入受体细胞,包括将核酸引入所述细胞的染色体dna。
[0375]
术语“治疗”是指以治愈、改善、稳定或预防疾病、病理病症或障碍为目的对患者进行的医学管理。该术语包括积极治疗,即专门针对改善疾病、病理病症或障碍的治疗,还包括因果治疗,即针对消除相关疾病、病理病症或障碍的原因的治疗。此外,该术语还包括姑息治疗,即旨在缓解症状而不是治愈疾病、病理病症或障碍的治疗;预防性治疗,即旨在最小化或部分或完全抑制相关疾病、病理病症或障碍的发展的治疗;和支持治疗,即用于补充另一种针对改善相关疾病、病理病症或障碍的特定疗法的治疗。
[0376]
术语“变体”是指具有保守氨基酸取代、非保守氨基酸取代(即简并变体)、编码氨基酸的每个密码子(即dna和rna)的摆动位置内的取代、添加到肽的c末端的氨基酸的氨基酸或肽序列,或与参考序列具有60%、70%、80%、90%、95%、96%、97%、98%、99%序列同一性的肽。
[0377]
术语“载体”是指能够将与载体序列相连的另一种核酸转运到细胞中的核酸序列。术语“表达载体”包括任何含有适合细胞表达的形式(例如,连接到转录控制元件)的基因构建体的载体(例如,质粒、粘粒或噬菌体染色体)。
[0378]
已经描述了本发明的多个实施例。然而,应当理解,在不背离本发明的精神和范围的情况下可以进行各种修改。因此,其他实施例在以下权利要求的范围内。
[0379]
实例
[0380]
实例1:cd83靶向嵌合抗原受体t细胞预防gvhd并杀伤髓系白血病
[0381]
材料和方法
[0382]
研究设计:这是一项关于用于gvhd预防的人cd83 car t细胞的设计、生产和功效的临床前研究。该研究的第一部分描述了car构建体以及cd83 car t细胞响应于cd83+靶标在表型、细胞因子产生、靶向杀伤和增殖方面的体外活性。随后在体外用标准allomlr证明cd83 car t细胞的免疫抑制作用。此外,在人t细胞中测量cd83表达,显示cd83在tconv和treg细胞上的差异表达。在人t细胞介导的异种gvhd模型中(betts b.c.等人,science translational medicine[科学转化医学]9:eaai8269(2017)),证明了cd83 car在gvhd预防中的临床前功效。这包括对cd83+树突状细胞和tconv的体内靶向杀伤的全面评估。还显示了cd83 car t细胞对体内各种t细胞亚群的影响。已证明cd83在人恶性髓系细胞系上表达,并使用xcelligence rtca(实时细胞分析)系统通过cd83 car t细胞有效杀伤(li g.等人,jci insight 3(2018))。对于gvhd实验,使用人道的临终终点。经常监测小鼠的gvhd临
床评分。gvhd组织病理学由致盲的专家病理学家进行评估和评分(betts b.c.等人,science translational medicine[科学转化医学]9:eaai8269(2017);betts b.c.等人,proc natl acad sci u s a.[美国科学院院报],201712452(2018);betts b.c.等人,front immunol[免疫学前沿]9:2887(2018))。鼠体内数据由至少两个独立实验汇集,每个实验组有6-9只小鼠。
[0383]
cd83 car t细胞构建体和产生:cd83 car由genewiz合成并克隆到sfg逆转录病毒构建体中(li,g.等人,methods mol biol[分子生物学方法]1514:111-118(2017);li g.等人,jci insight 3(2018))。然后使用磷酸钙将cd83 sfg克隆构建体转染到h29细胞中,并使用来自转染的h29细胞的逆转录病毒上清液转导rd114。将rd114细胞的逆转录病毒上清液通过0.45μm过滤器(milliporesigma)过滤以纯化γ逆转录病毒。具体地,通过如所述转导人t细胞来产生cd83 car t细胞(li g.等人,jci insight 3(2018))。简而言之,通过密度梯度离心分离来自健康人供体(all细胞)的单采术中获得的白细胞。使用磁珠(干细胞公司(stem cells inc.))分离t细胞,并在具有重组人il-2的rpmi中用人dynabeads cd3和cd28(赛默飞世尔公司(thermo fisher))进行刺激。用cd83γ逆转录病毒在retronectin(塔卡拉公司(takara bio inc.))涂层板上转导激活的t细胞。在激活7-8天后,cd83 car t细胞被脱珠。如通过流式细胞术检测的,通过gfp+细胞估计基因转移或转导效率。
[0384]
单克隆抗体和流式细胞术:荧光染料缀合小鼠抗人单克隆抗体包括抗cd3、cd4、cd8、cd25、cd83、cd1c、cd127、mhcii、foxp3、ki-67、ifn-γ、il-17a和il-4(bd生物科学公司(bd biosciences,加利福尼亚州圣何塞市,美国;e生物科学公司(ebioscience),加利福尼亚州圣何塞市,美国;细胞信号传导技术公司(cell signaling technology),马萨诸塞州波士顿,美国)。活/死可固定黄色或水性死细胞染色(生命技术公司(life technologies),纽约州格兰德岛)用于测定存活率。在bd facscanto ii或lsrii流式细胞仪(flowjo软件,7.6.4版;treestar,美国俄勒冈州阿什兰)上采集活事件。
[0385]
细胞因子免疫测定:将cd83 car和模拟转导t细胞(1x105)用cd83+modc(1x104)共培养24小时。收获上清液并在luminex 100系统(luminex)上使用人luminex测定试剂盒(r&d系统公司(r&d systems))和在ella仪器(biotechne)上使用simple plex测定试剂盒(biotechne)分析。遵循制造商的说明(li g.等人,jci insight 3(2018))。
[0386]
人cd83 car t细胞的细胞毒性和体外增殖:将归一化的cd83 cart细胞(1x105个细胞)与cd83+modc、k562、或thp-1细胞以10:1的et比率在e-plate 96中重复培养。根据制造商的说明,在xcelligence rtca(实时细胞分析)仪器(acea生物科学公司(acea biosciences))上进行细胞毒性测定。类似地,将人cd83 car t细胞与modc一起以1:1的et比率在非组织培养物治疗的6孔板中共培养,一式三份。将细胞在补充有60iu/ml il-2的人t细胞完全培养基中生长。在+1天、+7天和+14天时,用台盼蓝染色法在细胞计数器(伯乐公司(bio-rad))上测量每个孔中的细胞活力和总细胞数量。
[0387]
体外allomlr:如所述,人单核细胞衍生的树突状细胞(modc)是细胞因子产生的、分化的和成熟的(betts b.c.等人,science translational medicine[科学转化医学]9:eaai8269(2017))。将从白细胞浓缩物(oneblood或memorial blood center)纯化的t细胞(105个)与同种异体modc(t细胞:dc比率30:1)在100μl补充10%热灭活的混合人血清的完全rpmi中培养(betts b.c.等人,science translational medicine[科学转化医学]9:
eaai8269(2017);betts b.c.等人,proc natl acad sci u s a.[美国科学院院报],201712452(2018);betts b.c.等人,front immunol[免疫学前沿]9:2887(2018))。将cd83 car、cd19 car或模拟转导t细胞(对t细胞供体为自体的)以一定的car与dc比率范围添加到allomlr中。5天后通过ki-67表达测量t细胞增殖。
[0388]
cd83表达时间过程:用同种异体modc(t细胞:dc比率30:1)或cd3/cd28珠(t细胞:珠比率30:1)刺激纯化的人t细胞。在培养4、8、24和48小时后,从96孔板中的一式三份孔中收获t细胞。对t细胞进行cd3、cd4、cd127、cd25和cd83染色,然后固定。在激活的tconv(cd3+、cd4+、cd127+、cd25+)(betts b.c.等人,science translational medicine[科学转化医学]9:eaai8269(2017))、treg(cd3+、cd4+、cd127-、cd25+)(betts b.c.等人,science translational medicine[科学转化医学]9:eaai8269(2017))和cd8 t细胞(cd3+、cd4-)中评估cd83表达。在指示的情况下,将cd83car或模拟t细胞与dc同种异体刺激的pbmc一起培养,并在48小时内评估cd3-和cd3+靶细胞中的cd83表达。
[0389]
集落形成单位:从正常人骨髓中分离的cd34+细胞购自allcells。将103个细胞与cd83病毒转导的car t细胞、模拟t细胞或单独培养基共培养。将细胞以10:1的e:t比率孵育4小时。培养后,根据制造商的说明,将细胞铺板在6孔smartdish板(干细胞公司(stemcell))中的methocult培养基(干细胞公司)中,并培养14天。在培养期结束时,使用stemvision软件对集落进行成像、分析和计数。
[0390]
异种gvhd模型:将nod scidγ(nsg)小鼠(雄性或雌性,6-24周龄)在moffitt/usf饲养场维持的iacu-c批准聚居地内饲养。受体小鼠在移植的第0天接受一次25x106个新鲜的人pbmc(oneblood)。如所示的,小鼠要么接受单独pbmc,要么接受pbmc加cd83 car t细胞(低剂量:1x106或高剂量:10x106),或pbmc加模拟转导t细胞(10x106)。每个独立的实验都是用不同的人pbmc供体进行的,其中car t细胞和模拟转导t细胞衍生自pbmc供体。监测小鼠的gvhd临床评分和临终状态。在指示的情况下,通过人道安乐死在+21天时完成短期实验,以评估盲法gvhd靶器官病理学、组织驻留淋巴细胞以及鼠脾脏中人dc和t细胞亚群的含量(betts b.c.等人,science translational medicine[科学转化医学]9:eaai8269(2017);betts b.c.等人,proc natl acad sci u s a.[美国科学院院报],201712452(2018);betts b.c.等人,front immunol[免疫学前沿]9:2887(2018))。如前所述,对组织样品进行制备、染色(维塔纳医学系统公司(ventana medical systems))和成像(vista)以鉴定人ki67+ t细胞(betts b.c.等人,science translational medicine[科学转化医学]9:eaai8269(2017))。这些小鼠被移植了带有或不带有cd83 car(1x106)或模拟转导t细胞(1x106)的pbmc(25x106)。所有脊椎动物工作均在aicuc批准的方案下进行。
[0391]
统计分析:数据报告为平均值
±
sem。anova用于组比较,包括dunnett或sidak的事后检验,其中对多重比较进行了校正。mann-whitney被用于所有其他项。为了比较存活曲线,使用了对数秩检验。使用prism软件5.04版(graphpad)进行统计分析。统计显著性由双尾p《0.05(双尾)定义。
[0392]
结果
[0393]
人cd83 car构建体的示意图:抗cd83单链可变片段(scfv)与cd8铰链和跨膜结构域配对,然后是细胞内41bb共刺激结构域和cd3ζ激活结构域(图1a)。为了便于跟踪car t细胞,该构建体包含egfp标签,其可用于鉴定正常非car t细胞中的car t细胞(图1a)。如我们
已经公布的,逆转录病毒转导并产生cd83靶向car t细胞(图1a)(li,g.等人,methods mol biol[分子生物学方法]1514:111-118(2017);li g.等人,jci insight 3(2018))。
[0394]
人cd83 car t细胞的表征:cd83 car构建体表现出高度的转导效率,超过60%的t细胞表达egfp(图1b)。虽然两组的cd4表达相似,但与模拟转导t细胞相比,cd83 car t细胞中观察到cd8表达显著降低(图1c)。然而,当与cd83+靶细胞一起培养时,cd83 car t细胞展示出稳健的ifnγ和il-2产生;如细胞因子成熟的人单核细胞衍生的dc(modc)(图1d、e)。此外,与模拟转导t细胞相比,cd83 car t细胞展示出对cd83+modc的有效杀伤和增殖(图1f、1g)。这些实验中的靶modc对t细胞是同种异体的,因此模拟转导t细胞的裂解和增殖代表了基线同种异体反应性(图1f、1g)。
[0395]
人cd83 car t细胞降低同种异体反应性:为了测试人cd83 car t细胞是否在体外降低同种异体反应性,研究了它们在同种异体混合白细胞反应(allomlr)中的抑制功能。cd83和模拟转导car t细胞由健康供体人t细胞产生。cd19 car t细胞靶向b细胞(一种在allomlr中不相关的细胞类型),并被用作另一个对照。此外,cd19和cd83 car t细胞的相似之处在于它们都通过41bb接受共刺激。将car t细胞添加到由自体t细胞(1x105)和同种异体的细胞因子成熟的cd83+modc(3.33x103)组成的5天allomlr中。car t细胞:modc比率范围为3:1至1:10。cd83 car t细胞有效地降低了同种异体反应性t细胞增殖(图2,上图)。相反,模拟转导t细胞和cd19靶向car t细胞对同种异体反应性t细胞没有抑制作用(图2,中图和下图)。
[0396]
与treg相比,cd83在激活的人tconv上差异表达:cd83是人树突状细胞成熟的既定标记并且也在激活的人b细胞上表达(szabolcs p.等人,blood[血液]87:4520-4530(1996);krzyzak l.等人,j immunol[免疫学杂志]196:3581-3594(2016))。使用cd83报告子小鼠系统,先前显示激活的鼠t细胞也表达cd83(lechmann,m.等人,proc natl acad sci u s a[美国科学院院报]105:11887-11892(2008))。已知在刺激后cd83在人t细胞上表达,并且可在来自急性gvhd患者的循环t细胞上检测到(ju x.等人,j immunol[免疫学杂志]197:4613-4625(2016))。然而,cd83在cd4+treg与cd4+tconv或cd8+ t细胞上的精确表达尚不清楚。实验证实,人t细胞表达cd83会伴随发生刺激,包括同种异体树突状细胞或cd3/cd28珠(图3a、3b)。重要的是,已证明与免疫抑制性cd4+treg(cd127-、cd25+)或溶细胞性cd8+ t细胞相比,cd83在人cd4+tconv(cd127+、cd25+)上差异表达,以响应于dc同种异体激活(图3a)。cd83的cd4+tconv表达在dc同种异体刺激4-8小时时达到峰值,并在48小时下降至基线水平,在treg或cd8+ t细胞上观察到的量最少(图3a)。cd83的表达在超生理cd3/cd28珠刺激下更加丰富,这也导致treg和cd8+ t细胞上cd83表达在激活48小时后的后期增加(图3b)。鉴于cd83表达在促炎成熟dc以及同种异体反应性tconv之间共享,研究了cd83 car t细胞是否可以消耗培养物中的任一靶细胞。将人cd83 car或模拟t细胞与通过同种异体modc刺激的自体外周血单核细胞(pbmc)一起培养,并在培养4、8、24和48小时评估cd83+靶细胞的量。我们在8小时观察到cd3-和cd3+靶细胞的cd83表达出现类似的峰形(图3c)。然而,cd83+靶细胞在培养48小时时基本被cd83 car t细胞消除,并且远低于培养后8小时的基线量(图3c)。此外,cd83-t细胞仍然存在于所有实验组中(图3c),支持t细胞没有被不加选择地破坏。接下来,评估了48小时内egfp+car t细胞上cd83的表达。car t细胞上的cd83表达适中,并且在培养48小时后仍观察到egfp+car t细胞比例增加(图3d),提供证据表明
cd83 car t细胞不会明显屈服于cd83介导的自相残杀。为了平行临床实践,测试了cd83 car t细胞在存在临床相关剂量的他克莫司(5-10ng/ml)的情况下的功能能力。有趣的是,尽管暴露于他克莫司,cd83 car t细胞仍然可以杀伤和增殖以响应于cd83+靶细胞(图9a、9b)。
[0397]
人cd83靶向car t细胞预防异种gvhd:使用异种gvhd模型评估人cd83 car t细胞在体内的功效。使用了已建立的nsg小鼠模型(betts b.c.等人,science translational medicine[科学转化医学]9:eaai8269(2017)),其中在第0天,受体接种了25x106个人pbmc加1-10x106个自体cd83或模拟转导car t细胞。每天监测移植小鼠的异种gvhd临床症状直至+100天。与单独的pbmc相比,输注cd83或模拟转导car t的nsg小鼠没有早期gvhd或毒性的证据(图4a、4b)。然而,与单独的pbmc或模拟转导car t细胞相比,cd83 car t细胞在移植后显著提高了异种gvhd的存活率(图4a)。此外,cd83靶向car t细胞降低了异种gvhd临床严重程度(图4b)。值得注意的是,cd83靶向car t细胞的两个剂量组的小鼠展示出90%或更高的3个月存活率(图4a)。在单独的实验中,移植的nsg小鼠接受单独的pbmc或与模拟转导t细胞(1x106)或cd83靶向car t细胞(1x106)一起接受,并在+21天时人道地安乐死以评估靶器官gvhd的严重程度。gvhd路径评分由致盲的专家病理学家确定(betts b.c.等人,science translational medicine[科学转化医学]9:eaai8269(2017);betts b.c.等人,proc natl acad sci u s a.[美国科学院院报],201712452(2018);betts b.c.等人,front immunol[免疫学前沿]9:2887(2018))。与单独pbmc或模拟转导t细胞相比,cd83 car t细胞消除了人t细胞在受体肺(图4c-4e)和肝脏(图4g-j)中对异种gvhd靶器官组织的损伤。此外,很少有人t细胞直接浸润鼠靶器官,并且根据ki-67染色,它们没有增殖(图4e、4f、4i、4j)。
[0398]
人cd83靶向car t细胞在体内显著降低cd83+dc:成熟的cd83+树突状细胞与同种异体反应性供体t细胞的致敏有关。因此,确定了cd83car t细胞对移植小鼠中人cd1c+dc免疫恢复的影响。在+21天对移植了人pbmc加cd83 car或模拟转导t细胞的nsg小鼠实施安乐死。在收获受体脾脏后,确定cd83靶向car t细胞降低了供体细胞在体内的扩增,正如该治疗组中小得多的脾脏所指示的那样(图10)。cd83靶向car t细胞显著降低了受体小鼠中人cd1c+、cd83+dc的量(图5a、5b)。虽然在实验组中,表达mhc ii类的cd1c+dc的比例相似,但移植了cd83 car t细胞的小鼠总体表现出显著更少的dc总数(图5c、5d)。
[0399]
人cd83靶向car t细胞体内显著降低cd4+、cd83+ t细胞,同时增加treg:激活的tconv比率:将egfp标签用于证实在+21天时可在鼠脾脏中检测到的输注的人cd83 car t细胞(图6a)。在+21天时,经cd83靶向car t细胞治疗的小鼠的脾脏中人cd4+ t细胞总量显著降低(图6b、6c)。由于在体外观察到dc同种异体刺激后有大量cd83+cd4+tconv,因此进行了实验以证实在+21天时在使用单独的pbmc或模拟转导t细胞治疗的小鼠中cd83+tconv增加(图6d)。此外,在体内接受cd83 car t细胞的受体中,cd83+tconv的量显著减少(图6d)。总体而言,与模拟t细胞相比,cd83 car t细胞在+21天时能稳健清除cd83+靶细胞(图11a)。虽然在+21天时,循环egfp+car t细胞的数量较多与cd83+dc的数量较少有关,但cd83+ t细胞的降低在体内car t细胞数量上是一致的(图11b、11c)。
[0400]
在单独的实验中,nsg小鼠被移植单独人t细胞或t细胞加树突状细胞。虽然缺乏树突状细胞略微延迟了gvhd的发病,但两组的中位gvhd存活率相似(图12a、12b)。这与其他项的工作一致,表明纯化的人t细胞足以诱导异种gvhd(li w.等人,jci insight 1(2016))。
[0401]
据推测,cd83靶向car t细胞主要通过消除与gvhd有关的同种异体反应性tconv,同时增强treg与同种异体反应性tconv的比率来保护受体免受gvhd的影响(图6e-6g)。在+21天时,所有实验组鼠脾脏中人treg的频率相似(图6e)。与总cd4+ t细胞的降低类似,在用cd83靶向car t细胞治疗的小鼠中,treg的绝对数量显著减少(图6f)。然而,treg(cd4+、cd127-、cd25+、foxp3+)与激活的tconv(cd4+、cd127+、cd25+)的比率(betts b.c.等人,science translational medicine[科学转化医学]9:eaai8269(2017)))在接受cd83靶向car t细胞的小鼠中显著增加(图6g)。th1细胞促进了gvhd发病机制。重要的是,用cd83 car t细胞治疗的小鼠表现出人cd4+、ifnγ+th1细胞的显著降低(图6h、6i)。此外,在注射cd83 car t细胞的小鼠中,脾脏驻留的人th2细胞(cd4+、il-4+)的量也显著减少(图6h、6j)。相反,与单独的pbmc或模拟转导car t细胞相比,cd83靶向car t细胞不会抑制受体脾脏中人th17细胞的量(图13a、13b)。有趣的是,在长期实验中,在存活至+100天终点的小鼠的脾脏中也检测到egfp+cd83 car t细胞(图14)。移植后超过3个月,在低(1x106)或高(10x106)剂量的cd83 car t细胞治疗的小鼠中,观察到循环cd83+靶细胞的剂量依赖性减少(图14)。
[0402]
人cd83 car t细胞杀伤急性髓系白血病细胞系:根据国际血液和骨髓移植研究中心(cibmtr)的纵向数据,每年有1000多名患者因高风险aml接受allo-hct(gupta,v.等人,blood[血液]117:2307-2318(2011))。即使患者能够耐受清髓预备方案,无复发生存率也限于67.8%,而降低强度调理后为47.3%(scott b.l.等人,j clin oncol[临床肿瘤学杂志]35:1154-1161(2017))。因此,迫切需要预防aml复发的策略。鉴于cd83 car t细胞在异种gvhd预防中具有强大的裂解活性,且移植小鼠对其耐受性良好,因此进行了实验,以研究人髓系白血病是否潜在地表达cd83。发现cd83确实在恶性髓系k562、thp-1、u937和molm-13细胞系上表达(图7a、7b,图15a、15b)。此外,使用xcelligence平台,cd83 car t细胞对k562和thp-1细胞展示出显著的抗肿瘤活性(图7c、7d)。因此,人cd83 car t细胞具有预防gvhd并提供直接杀伤aml的能力。
[0403]
人cd83 car t细胞表现出可忽略不计的中靶脱瘤毒性:人aml抗原通常与祖干细胞共享。虽然cd83 car t细胞明显杀伤aml靶标,但证实它们允许造血干细胞在集落形成单位(cfu)中的生长和分化(图8a-8d)。总体而言,模拟t细胞、cd83 car t细胞和培养基治疗组的集落总数相似。虽然对cd83 car t细胞观察到粒细胞/巨噬细胞cfu减少,但与单独培养基相比没有显著差异(图8b)。此外,在治疗组中,来自粒细胞/红细胞/单核细胞/巨核细胞cfu和红系爆式形成单位的集落基本相同(图8c、8d)。这些实验提供了人cd83 car t细胞选择性杀伤aml同时保留正常的造血功能的证据。
[0404]
讨论
[0405]
使用car t细胞作为细胞免疫疗法来预防gvhd是一种创新策略,不同于供体treg的药物免疫抑制或过继转移。靶向表达cd83的细胞有效地消耗移植受体的炎症性成熟dc以及同种异体反应性cd4+tcovnv。供体cd8+t细胞也可以介导gvhd(okiyama n.等人,j invest dermatol[皮肤病学研究杂志]134:992-1000(2014);shindo t.等人,blood[血液]121:4617-4626(2013))。虽然很少有人cd8+ t细胞表达cd83,但cd83 car t细胞也显著降低了供体cd8+ t细胞的量(图16)。从机制上讲,据推测,同种异体反应性t细胞的体内消除驱动了这些car t细胞的功效,因为树突状细胞的消耗并没有减少异种gvhd。cd83 car t细胞对同种异体反应性t效应子的体内消耗也介导了treg:激活的tconv比率的显著升高,这
是控制gvhd的临床相关指标(koreth j.等人,n engl j med[新英格兰医学杂志]365:2055-2066(2011))。
[0406]
cd83 car t细胞在体内显著降低了致病性人th1和th2细胞。使用stat4和stat6敲除供体t细胞的实验表明,th1和th2细胞独立介导小鼠中的致死性gvhd(nikolic,b.等人,j clin invest[临床研究杂志]105:1289-1298(2000))。此外,体内th1和th2细胞的组合协同恶化了鼠gvhd(nikolic,b.等人,j clin invest[临床研究杂志]105:1289-1298(2000))。在某种程度上,th1和th2细胞分别对肠道和肺部造成组织特异性损伤(yi t.等人,blood[血液]114:3101-3112(2009))。目前存在靶向供体th1反应的策略,并且主要由p40细胞因子中和或相关下游受体信号转导的抑制驱动(betts b.c.等人,science translational medicine[科学转化医学]9:eaai8269(2017);betts b.c.等人,proc natl acad sci u s a.[美国科学院院报],201712452(2018);betts b.c.等人,front immunol[免疫学前沿]9:2887(2018);pidala j.等人,haematologica[血液学]2017.171199(2017);yu y.等人,blood[血液]118:5011-5020(2011))。然而,很少有方法同时靶向致病性th1和th2细胞。因此,人cd83 car t细胞代表了一种细胞产物,可在allo-hct后同时抑制供体th1/th2反应。人th17细胞在很大程度上不受cd83 car t细胞的影响,尽管经治疗的小鼠明显受到针对gvhd的保护。虽然供体th17细胞有可能促进gvhd(iclozan c.等人,biol blood marrow transplant 16:170-178(2010)),但缺乏可用的th1细胞可能会减轻存活th17细胞的致病性(yu y.等人,blood[血液]118:5011-5020(2011))。
[0407]
所披露的数据支持人cd83 car t细胞提供持久的保护以防止激活的tconv和gvhd死亡。虽然cd83在人treg上没有显著表达,但用人cd83car t细胞治疗的小鼠表现出treg的量减少。这可能是由于用于treg分化的cd4+ t细胞前体的可用性有限,或者由于循环供体t细胞的整体减少而导致的il-2浓度降低。在啮齿类动物中,cd83参与体内treg的稳定性,而携带cd83缺陷型treg的小鼠易患自身免疫综合征(doebbeler m.等人,jci insight 3(2018))。然而,在异种移植实验中,与对照相比,用cd83 car t细胞治疗的小鼠中人treg与激活的tconv的比率显著增加。treg与tconv的比率增加是临床相关的免疫指标,并且甚至与对treg导向的gvhd疗法(如低剂量il-2)的反应相关(koreth j.等人,n engl j med[新英格兰医学杂志]365:2055-2066(2011);koreth j.等人,blood[血液]128:130-137(2016))。此外,人cd83 car t细胞在体内具有良好的耐受性,并消除免疫介导的器官损伤。因此,cd83在鼠和人treg中的作用可能不同。
[0408]
cd83是一种独特的免疫调节分子。在小鼠中,可溶性cd83通过吲哚胺2,3-双加氧酶和tgfβ机制增强treg反应来介导免疫抑制作用(bock f.等人,j immunol[免疫学杂志]191:1965-1975(2013))。人cd83的细胞外结构域也显示在体外损害同种异体反应性t细胞增殖(lechmann m.等人,j exp med[实验医学期刊]194:1813-1821(2001))。相反,用单克隆抗体3c12c直接中和cd83显著降低了体内人t细胞介导的异种gvhd(wilson j.等人,j exp med[实验医学期刊]206:387-398(2009))。cd83抗体还保留了供体人cd8+ t细胞的treg和抗病毒反应(seldon t.a.等人,leukemia[白血病]30:692-700(2016))。这表明虽然可溶性cd83可能具有免疫抑制特性,但靶向cd83的细胞表面表达可以预防gvhd,同时保留关键效应子和treg功能。与单克隆抗体不同的是,cd83 car t细胞单独引起稳健的靶细胞杀伤;不需要nk细胞介导的抗体依赖性细胞毒性(seldon t.a.等人,leukemia[白血病]30:
692-700(2016))。当需要快速、有效地消除同种异体反应性t细胞以预防gvhd时,这是一个优势。事实上,人cd83靶向car t细胞提供了持久的gvhd预防,即使在单次输注后,在+100天的小鼠中也能检测到。
[0409]
除了在gvhd预防中消除同种异体反应性t细胞外,cd83显现是靶向髓系恶性肿瘤的有希望的候选者。在恶性髓系k562、thp-1、u937和molm-13细胞上观察到cd83表达。此外,cd83 car t细胞有效地杀伤了aml细胞系。许多aml抗原在祖干细胞上表达。因此,在人cfu测定中进行实验来评估干细胞的杀伤作用,证明中靶/脱瘤毒性可以忽略不计。尽管复发仍然是移植后失败和死亡的重要原因,但allo-hct通常是治疗高风险aml所必需的。与hla介导的经典gvl不同,cd83 car t细胞选择性地破坏表达cd83的恶性细胞。此外,最近发现cd83也在霍奇金淋巴瘤上表达(li z.等人,haematologica[血液学]103:655-665(2018))。因此,cd83car t细胞在治疗aml或hl方面可能具有功效,而不依赖于allo-hct。鉴于cd19 car t细胞在all和弥漫性大b细胞淋巴瘤中的临床成功,这在转化上是强效的(neelapu s.s.等人,n engl j med[新英格兰医学杂志]377:2531-2544(2017);schuster s.j.等人,engl j med[新英格兰医学杂志]380:45-56(2019);maude s.l.等人,n engl j med[新英格兰医学杂志]378:439-448(2018);davila m.l.等人,sci transl med[科学转化医学]6:224ra225(2014))。
[0410]
总之,cd83 car t细胞代表了第一种旨在预防gvhd的人程序化溶细胞效应细胞。cd83 car t细胞的转化潜力已在gvhd预防中得到证明,尽管预计它在实体器官或血管化的复合同种异体移植物移植后也具有预防排斥反应的优点。此外,即使暴露于钙调神经磷酸酶抑制剂,cd83 car t细胞仍保持其杀伤活性。cd83 car t细胞可以克服在造血细胞hla差异和实体器官供体选择中的障碍,并大大扩展了治愈性移植手术在有需要的患者中的应用。重要的是,cd83 car t细胞提供了一个平台来消除同种异体反应性t细胞,而无需广泛抑制、非选择性的钙调神经磷酸酶抑制剂或糖皮质激素。此外,cd83 car t细胞杀伤髓系白血病细胞的能力进一步扩大了其临床影响。因此,cd83 car t细胞极有可能降低移植相关死亡率并改善allo-hct后的结果。
[0411]
除非另有定义,否则本文使用的所有技术和科学术语具有与所披露的发明所属领域中的技术人员通常所理解的相同含义。本文引用的出版物和引用它们的材料通过引用具体并入。
[0412]
本领域的技术人员仅仅使用常规实验将认识到或能够确认在此描述的发明的具体实施例的许多等同物。这些等同物旨在被以下权利要求所涵盖。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1