使用灭活的修饰的痘苗病毒安卡拉作为实体肿瘤的单一免疫疗法或与免疫检查点阻断剂组合的制作方法
使用灭活的修饰的痘苗病毒安卡拉作为实体肿瘤的单一免疫疗法或与免疫检查点阻断剂组合
1.本技术是申请日为2016年2月25日、申请号为201680023964.5、发明名 称为“使用灭活的非复制型的修饰的痘苗病毒安卡拉(mva)作为实体肿瘤的单 一免疫疗法或与免疫检查点阻断剂的组合”的发明专利申请的分案申请。
2.相关应用
3.本技术要求于2015年2月25日提交的美国临时申请no.62/ 120,862的优先权。该临时申请的全部公开内容通过引用整体并入本文。
[0004][0005]
发明领域
[0006]
本发明一般涉及肿瘤学,病毒学和免疫治疗领域。更具体地说,它 涉及使用痘病毒,特别是具有灭活的修饰的痘苗病毒安卡拉病毒(“灭 活的mva”),其具有感染能力但是不能复制的,并且已经通过例如热 或紫外线(uv)照射进一步修饰。这种灭活的mva可以作为免疫治 疗剂用作治疗癌症,作为单一疗法或与免疫检查点阻断疗法的组合作为 联合疗法。
[0007]
背景
[0008]
免疫系统和癌症
[0009]
恶性肿瘤本身对常规治疗具有抵抗力,并呈现出显着的治疗挑战。 免疫治疗已成为不断发展的研究领域,也是治疗某些类型癌症的另一种 选择。免疫治疗方法取决于免疫系统可能被刺激以辨别肿瘤细胞,并将 其靶向用于破坏的原理。
[0010]
许多研究支持免疫系统成员在癌症进展中的不同存在的重要性[1]。 临床数据表明,肿瘤浸润淋巴细胞的高密度与临床结局改善有关[2]。 已经报道了各种类型的癌症,包括黑色素瘤,卵巢癌,头颈部癌,乳腺 癌,尿路上皮癌,结肠直肠癌,肺癌,肝细胞癌,胆囊癌和食管癌中的 淋巴细胞浸润和患者存活之间的相关性[3]。肿瘤免疫浸润包括巨噬细 胞,树突状细胞(dc),肥大细胞,天然杀伤(nk)细胞,初始和记 忆性淋巴细胞,b细胞和效应t细胞(t淋巴细胞),主要负责识别由 肿瘤细胞表达的抗原,随后由t细胞破坏肿瘤细胞。
[0011]
尽管由癌细胞呈递抗原和可潜在地与肿瘤细胞反应的免疫细胞的 存在,但在许多情况下,免疫系统不被激活。这种现象的关键是肿瘤通 过胁迫免疫系统的细胞来免疫免疫应答来抑制免疫系统的其他细胞。例 如,cd4
+
t细胞具有分化成具有抑制活化的t细胞的能力的t调节 (treg)细胞的能力。此外,癌细胞可能损害cd8
+
t细胞效应子功能, 导致逃避抗肿瘤免疫应答。最后,肿瘤微环境的局部免疫抑制性质以及 免疫编辑可导致不表达靶抗原的癌细胞亚群的逃逸。这样,找到可以保 护和/或恢复免疫系统的抗肿瘤活性的方法是至关重要的。
[0012]
已经确定i型ifn在宿主抗肿瘤免疫中起重要作用[4]。ifnar1 缺失小鼠在肿瘤细胞植入后更容易发生肿瘤。自发性肿瘤特异性t细 胞引发在ifnar1缺陷小鼠中也是缺失的
[5,6]。更多最近的研究表明, 细胞溶质dna感测途径(dna-sensing pathway)在先天免疫系统识别肿 瘤衍生的dna中是重要的。反过来,这导致抗肿瘤cd8
+
t细胞免疫的 发展[7]。该途径也在辐射诱导的抗肿瘤免疫中起重要作用[8]。
[0013]
黑色素瘤
[0014]
黑色素瘤是最致命癌症之一,是美国和全球增长最快的癌症。自 1980年以来,年轻白人妇女的发病率增加了50%,主要是由于阳光照 射过多和使用日晒床。根据美国癌症协会的数据,美国将有大约76,380 人被诊断患有黑色素瘤,预计2016年将有10,130人(或每小时1人) 死于黑色素瘤。在大多数情况下,晚期黑色素瘤对常规治疗有抗性,包 括化疗和放射。结果,转移性黑色素瘤患者的预后很差,预期寿命只有 6~10个月。发现大约50%的黑色素瘤在braf(一种关键的肿瘤促进 基因)中具有突变,为该疾病的靶向治疗打开了大门。braf抑制剂 的早期临床试验显示出显着的,但不幸的是,在具有braf突变的黑 色素瘤患者中不可持续的反应。因此,迫切需要这些患者的替代治疗策 略,以及无braf突变的黑色素瘤患者。
[0015]
人类病理资料表明,黑色素瘤病变中t细胞浸润的存在与患者生存 期较长有关[9]。免疫系统在免疫黑色素瘤保护方面的重要性进一步得 到免疫治疗部分成功的支持,如免疫激活剂ifn-α2b和il-2[10]以及转 移性黑色素瘤患者对免疫检查点封锁的前所未有的临床反应包括单独 或组合使用的抗ctla-4和抗pd-1/pd-l1[11-17]。然而,许多患者不 对免疫检查点阻断单一疗法有反应。添加病毒治疗可能会克服对免疫检 查点阻断的抵抗力,这得到动物肿瘤模型的支持[18]。
[0016]
痘病毒
[0017]
痘病毒,例如基因工程化痘苗病毒,在转移癌症的溶瘤治疗中处于 前列[19]。牛痘病毒是具有快速生命周期的大型dna病毒[20]。痘病毒 非常适合作为载体在癌细胞中表达多种转基因,从而提高治疗效果 [21]。临床前研究和临床试验已经证明使用溶瘤牛痘病毒和其他痘病毒 治疗常规治疗难治的晚期癌症的疗效[22-24]。基于痘病毒的溶瘤治疗具 有通过细胞裂解,细胞凋亡和坏死组合杀死癌细胞的优点。它还触发有 助于把免疫细胞召集到肿瘤和发展抗肿瘤适应性免疫应答的先天免疫 感测途径(sensing pathway)。目前在临床试验中的溶瘤牛痘菌株(例如 jx-594)使用具有胸苷激酶缺失的野生型牛痘来增强肿瘤选择性,并且 通过转基因如粒细胞巨噬细胞集落刺激因子(gm-csf)的表达来刺激 免疫反应[21]。然而许多研究表明,野生型牛痘对抗原呈递细胞(apc) 具有免疫抑制作用[25-28],因此增加了肿瘤本身的免疫抑制和免疫吸收 作用。
[0018]
痘病毒非常擅长通过编码阻断这些途径的细胞外和细胞内成分的 蛋白质来回避和拮抗多种先天免疫信号通路[29]。改良的痘苗病毒安卡 拉(mva)是通过在鸡胚胎成纤维细胞中连续传代而开发的减毒痘苗 病毒。mva具有31-kb的亲本痘苗基因组的缺失,并在世卫组织赞助 的天花根除运动中成功用作疫苗[30-32]。对mva作为针对艾滋病毒, 结核病,疟疾,流感和冠状病毒以及癌症的疫苗载体进行了深入研究 [33-38]。
[0019]
mva具有包括k1l,n1l和a52r在内的几种细胞内免疫调节基 因的缺失或截短,其涉及调节先天免疫应答[39-46]。另一方面,mva 保留编码双功能z-dna/dsrna结合蛋白的e3l基因,一种关键的痘 苗毒力因子[47-55]。已经表明,人单核细胞衍生的树突状细胞的mva 感染导致dc激活[56]。waibler等人[57]报道,大鼠flt3l-dc的mva 感染引发了与tlr
无关的i型ifn反应。此外,人类巨噬细胞的mva 感染通过tlr2/tlr6/myd88和mda5/mavs依赖性途径触发i型 ifn和促炎细胞因子和趋化因子[58]。
[0020]
感测(sensing)细胞溶质中的dna引发了一系列事件,导致产生 i型ifn和细胞因子以及细胞应激反应。sting(ifn基因的刺激物) 被鉴定为细胞溶质dna感测途径的重要适配器[59-61]。dna感测器 的性质仍然难以捉摸,直到发现循环gmp-amp合酶(cgas)为关键 的dna感测器,其产物环状gmp-amp在gpa步骤具有意外的2',5' 连接和在apg步骤具有标准的3',5'连接在[62-68]。随后的研究证实 sting是由cgamp激活的关键适配器,从而介导了导致干扰素反应的 涉及激酶和转录因子的下游事件的级联[66,68,69]。我们报道,大鼠常 规树突细胞的mva感染通过细胞溶质dna感测途径诱导i型ifn,该 途径由胞质dna传感器cgas,其适配器sting和转录因子irf3和 irf7介导。相比之下,野生型牛痘病毒不能激活该途径。通过尾静脉 注射静脉注射mva诱导wt小鼠中的i型ifn分泌,其在sting或irf3缺陷小鼠中减少[70]。此外,我们显示痘苗毒力因子e3和n1在 细胞溶质dna感测通路中起抑制作用[70]。
技术实现要素:
[0021]
一方面,本公开提供了治疗患有一种或多种实体恶性肿瘤的受试者 的方法,所述方法包括将灭活的修饰的牛痘安卡拉(灭活的mva)递送 至肿瘤细胞,从而治疗肿瘤。
[0022]
在另一方面,提供了一种治疗受试者中的实体恶性肿瘤的方法,所 述方法包括向受试者的肿瘤细胞递送有效诱导受试者的免疫系统发起抗 肿瘤免疫应答的量的灭活的mva。
[0023]
在另一方面,本公开提供了治疗受试者中的恶性肿瘤的方法,所述 方法包括以有效诱导受试者的免疫系统发起抗肿瘤免疫应答的量递送 灭活的mva到受试者的肿瘤细胞和以在肿瘤内有效阻断肿瘤细胞,基 质细胞,或肿瘤浸润性免疫细胞引起的免疫抑制机制的第二数量将免疫检 查点阻断剂联合给药受试者的组合,从而治疗肿瘤。如本文所用,“递 送”是指“给药”,前者主要用于灭活的mva,后者与免疫检查点阻断 剂相关。
[0024]
应当理解,除非另有明确说明,否则下文描述的实施例将涉及上述 各个方面,并且可以在特定方面单独呈现更多或更具体实施例的特征, 或者可以将它们中的一个或多个组合。
[0025]
在一些实施方案中,灭活的mva的量对于达成以下一个或多个是 有效的:
[0026]
a、诱导受试者的免疫系统发起抗肿瘤免疫应答;
[0027]
b、减少肿瘤的大小;
[0028]
c、根除肿瘤;
[0029]
d、抑制肿瘤生长;
[0030]
e、抑制肿瘤转移;和
[0031]
f、减少或根除转移性肿瘤。
[0032]
在一些实施方案中,经处理的肿瘤包括位于灭活的mva递送部位 的肿瘤,或同时位于受试者体内的所述部位和其它地方的肿瘤。换句话 说,mva递送的效果是全身性的,即使灭活的mva可以仅传递给受 试者的仅一个或仅多个实体瘤。
[0033]
在更具体的实施方案中,免疫应答包括以下一个或多个:
[0034]
a、在肿瘤和/或肿瘤引流淋巴结中增加细胞毒性cd8
+
t细胞;
[0035]
b、通过诱导i型ifn诱导浸润所述肿瘤的树突细胞的成熟;
[0036]
c、诱导在受试者中识别肿瘤中或全身的肿瘤细胞的活化的cd4
+
效应 t细胞;
[0037]
d、降低肿瘤内免疫抑制(调节)cd4
+
t细胞;和
[0038]
e、诱导肿瘤细胞在其表面上表达i型mhc,并产生i型ifn和其 它炎性细胞因子和趋化因子的一种或多种。
[0039]
在一些实施方案中,肿瘤是原发性或转移性黑色素瘤或原发性或转 移性结肠癌。
[0040]
在一些实施方案中,受试者是人。
[0041]
在一些实施方案中,以间隔开的时间间隔重复递送灭活的mva;在 更具体的实施方案中,只要获得利益持续或达到最大耐受剂量,重复递 送将持续数周,数月或数年或无限期;在进一步的实施方案中,以每周 一次至每周两次的频率重复灭活的mva的递送;在一些更具体的实施 方案中,每周重复递送一次。
[0042]
在一些实施方案中,灭活的mva的递送是通过肠胃外途径;在更具 体的实施方案中通过肿瘤内注射或静脉内注射。
[0043]
在一些实施方案中,灭活的mva以每次施用的剂量约10
5-10
10
个噬 菌斑形成单位(pfu)的范围内递送;在更具体的实施方案中,其以每次 施用的剂量约106至约109个噬菌斑形成单位(pfu)的范围内递送。
[0044]
在一些实施方案中,灭活的mva是uv灭活的mva;在其他实施 方案中,它是热灭活的mva;在其它实施方案中,是加热和uv灭活的 mva的组合。
[0045]
在一些实施方案中,效应t细胞的诱导和激活伴随着所述肿瘤中调 节性cd4
+
细胞的减少;在一些实施方案中,灭活的mva通过诱导i型ifn诱导浸润所述肿瘤的树突细胞的成熟;在一些实施方案中,灭活的 mva诱导i型mhc的表达和诱导感染的肿瘤细胞中的i型干扰素和其 它炎性细胞因子和趋化因子中的一种或多种。
[0046]
在一些实施方案中,诱导的免疫应答影响或有助于以下的一个或多 个:减少肿瘤大小,根除肿瘤,抑制肿瘤或转移生长。再次,肿瘤不局 限于注射灭活的mva的肿瘤。
[0047]
上述第三方面的具体实施方式包括前述及其另外的实施方案如下:
[0048]
在一些实施方案中,通过肿瘤内注射递送灭活的mva和通过静脉 内途径给药免疫检查点阻断剂;在其他实施方案中,递送和给药均通过 静脉内途径;在其它实施方案中,递送和给药都是通过肿瘤内注射。在 一些实施方案中,免疫检查点阻断剂选自抗pd-1抑制剂,pd-l1,抑 制剂和ctla4抑制剂,其在具体实施方案中为抗体。
[0049]
在一些实施方案中,所述灭活的mva的递送和所述免疫检查点阻断 剂的给药是根据其自身的间隔开的间隔的施用方案分别施用。在一些实施 方案中,递送和给药在相同的整个时间段期间并行发生。
[0050]
在一些实施方案中,第一剂量的灭活mva首先被递送,并且经过 一段时间,例如一周后,第一剂量的免疫检查点阻断剂被给药。在一些 实施方案中,灭活的mva的递送和免疫检查点阻断剂的给药的一个或同 时分别进行数周,数月或数年或在持续存在益处或达到最大耐受剂量的情 况下无限期。
[0051]
在一些实施方案中,同时施用免疫检查点阻断剂和灭活的mva;在 一些实施方案中,它们以相同的组合物施用;在一些实施方案中,它们 都是肿瘤内递送的。在一些实施方
案中,同时递送允许使用较低剂量的 免疫检查点阻断剂,并且两种活性剂的组合作用可以是协同的。
[0052]
可以排除被公开的任何实施例的特征或在多个实施例中被选择的 特征的组合。
附图说明
[0053]
图1是一系列数据的图形,显示了heat-mva比mvc诱导大鼠cdc 中i型ifn产生的水平较高。图1a是在mvi为10的mva或具有等 量的heat-mva感染的cdc(gm-csf培养的骨髓来源的dc)中,与 没有病毒的对照相比,相对的ifna4和ifnb mrna表达水平的条形 图。数据是平均值
±
sem(n=3)。显示一个代表性的实验,重复至少 两次。图1b是在mva或heat-mva感染后的cdc的培养基中分泌的 ifn-α和ifn-β的浓度的曲线图(***,p《0.001)。数据是平均值
±
sem (n=3)。显示一个代表性的实验,重复至少两次。图1c是显示痘苗 e3的p25,p-irf-3和β-肌动蛋白(作为加载对照)的蛋白质水平的蛋 白质印迹的扫描图像。“hpi”,感染后的小时。“m”,模拟感染对照。
[0054]
图2是一系列数据的图形,显示了heat-mva诱导的i型ifn产生 依赖于由cgas和sting介导的细胞溶质dna感测途径。图2a是 cgas
+/+
和cgas-/-小鼠产生并用heat-mva感染的cdc中,与无病毒 对照相比,ifna4和ifnb相对mrna表达的条形图(***,p《0.001)。 数据是平均值
±
sem(n=3)。显示一个代表性的实验,重复两次。图 2b是由cgas
+/+
和cgas-/-小鼠产生并用heat-mva感染的cdc的培养 基中分泌的ifn-α和ifn-β的浓度的柱状图(***,p《0.001)。数据是 平均值
±
sem(n=3)。显示一个代表性的实验,重复两次。图2c是 与sting
+/+
和sting
gt/gt
小鼠产生并用heat-mva感染的cdc中,与无 病毒对照相比,ifna4和ifnb相对mrna表达的条形图(***,p 《0.001)。数据是平均值
±
sem(n=3)。显示一个代表性的实验,重复 至少两次。图2d是从sting
+/+
和sting
gt/gt
小鼠产生,并且用heat-mva 感染的cdc的培养基中分泌的ifn-α和ifn-β的浓度的柱状图(***, p《0.001)。数据是平均值
±
sem(n=3)。显示一个代表性的实验,重 复至少两次。图2e是western印迹的扫描图像,显示heat-mva感染 后cgas
+/+
和cgas-/-cdc中p-irf3和β-肌动蛋白的蛋白水平。“hpi”, 感染后的小时。“m”,模拟感染对照。图2f是western印迹的扫描 图像,显示在加热mva感染后sting
+/+
和sting
gtgt cdc中的p-irf3 和β-肌动蛋白的蛋白水平。“hpi”,感染后的小时。“m”,模拟感 染控制。图2g是显示由sting
gtgt
和wt小鼠产生的受heat-mva感染 的cdc中的表面标志物mhc i(i型mhc),cd40,cd86和cd80的 表达的一系列图。显示一个代表性的实验,重复至少两次。
[0055]
图3是显示heat-mva诱导的i型ifn的产生依赖于转录因子irf3, irf7和ifnar1的一系列图形。图3a是描绘由wt,irf3-/-,irf5-/-, 和irf7-/-小鼠产生的cdc在heat-mva感染后ifna4和ifnb mrna 表达的倍数诱导的图。数据是平均值
±
sem(n=3)。显示一个代表性 的实验,重复两次。图3b是描绘由wt,irf3-/-,irf5-/-,和irf7-/-小 鼠产生的cdc在heat-mva感染后,培养液中分泌的ifn-α和ifn-β 的浓度的图。数据是平均值
±
sem(n=3)。显示一个代表性的实验, 重复两次。图3c是显示由ifnar
+/+
和ifnr-/-小鼠产生的cdc在heat
‑ꢀ
mva感染后ifna4和ifnb mrna的倍数诱导的条形图。数据是平均 值
±
sem(n=3)。显示一个代表性的实验,重复两次。图3d是显示 由ifnar
+/+
和ifnr-/-小鼠产生的cdc在heat-mva感染后的培养液 中分泌的ifn-α和ifn-β的浓度的柱状图。数据是平均值
±
sem(n= 3)。显示一个代
表性的实验,重复两次。
[0056]
图4是一系列散点图,显示heat-mva在体内诱导比mva高水平 的i型ifn,并且是以sting/irf3/irf7依赖性方式。图4a是通过 尾静脉注射接种mva(2
×
107pfu)或等量的heat-mva的wt小鼠的 血清中分泌的ifn-α和ifn-β的浓度的散点图。在接种后6小时收集 血清(***,p《0.001;n=5)。显示一个代表性的实验,重复两次。图 4b是用heat-mva接种的ifnar
+/+
或ifnr-/-小鼠血清中分泌的ifn
‑ꢀ
α和ifn-β的浓度的散点图(**,p《0.01;***,p《0.001;n=5)。显 示一个代表性的实验,重复两次。图4c是用heat-mva(n=5)接种 的来自wt,irf3-/-,irf7-/或sting
gt/gt
小鼠的血清中分泌的ifn
‑ꢀ
α和ifn-β的浓度的散点图。显示一个代表性的实验,重复两次。
[0057]
图5是显示b16-f10黑色素瘤细胞的heat-mva感染诱导产生i型 ifn和促炎细胞因子和趋化因子的数据的一系列图形。图5a是一系列 条形图,其显示了b16-f10黑色素瘤细胞的heat-mva或mva感染后 ifna4,ifnb,ccl5和il6的mrna水平的倍数诱导。数据是平均值
ꢀ±
sem(n=3)。显示一个代表性的实验,重复两次。图5b是一系列 条形图,其显示了在heat-mva或mva感染后b16-f10黑色素瘤细胞 培养基中分泌的ifn-α和ifn-β,ccl5和il-6的浓度。数据是平均 值
±
sem(n=3)。显示一个代表性的实验,重复两次。图5c是在用 heat-mva或mva感染的b16-f10细胞中显示p-irf,irf和gapdh 蛋白水平(作为加载对照)的western印迹的扫描图像。“hpi”,感染 后的小时。图5d是无病毒对照,感染mva或heat-mva的b16-f10 细胞中mhc i表达的图。显示一个代表性的实验,重复两次。
[0058]
图6是一系列数据的图形,显示在55℃下热灭活1小时的mva诱 导最高水平的来自cdc的ifn分泌。图6a和6b分别是在不同温度下 热处理1小时的mva感染的cdc的培养基中分泌的ifn-α(a)和ifn
‑ꢀ
β(b)的浓度的条形图。数据是平均值
±
sem(n=3)。显示一个代 表性的实验,重复两次。
[0059]
图7是显示heat-mva注射导致肿瘤根除和全身抗肿瘤免疫的数据 的一系列图形。图7a是pbs(空心圆圈;n=5)或heat-mva(实心圆 圈;n=10)注射后的肿瘤体积与时间(天)的曲线图。显示代表性实 验,重复至少五次。图7b是注射pbs(空心圆圈;n=5)或heat-mva (实心圆圈;n=10)(****,p《0.0001)的荷瘤小鼠的kaplan-meier存 活曲线。显示代表性实验,重复至少五次。图7c是初始小鼠(空心圆 圈;n=5)和heat-mva处理的小鼠(实心圆圈;n=10)的kaplan-meier 生存曲线,其在对侧用致死剂量的b16-f10黑色素瘤细胞(1
×
106个细 胞)再次攻击。显示代表性实验,重复至少五次。图7d是在静脉内输 送1
×
106个细胞(****,p《0.0001)后从初始小鼠(空心圆圈;n=9) 或heat-mva处理的小鼠(实心圆圈;n=10)在3周收集的肺表面上的 肿瘤灶数目的散点图。显示一个代表性的实验,重复至少两次。
[0060]
图8是显示肿瘤内注射heat-mva导致肿瘤微环境中的免疫学变化 的数据的一系列图形。图8a是cd3
+
cd45
+
t细胞的流式细胞分析。图 8b-c是表达颗粒酶b
+
(8b)或ki-67(8c)的cd8
+
细胞的流式细胞分 析的散点图。图8d-f是表达foxp3(8d),颗粒酶b(8e)或ki-67(8f) 的cd4
+
细胞的流式细胞分析的散点图。图8g-l是cd45
+
cd3
+
(8g), cd8
+
颗粒酶b
+
(8h),cd8
+
ki-67
+
(8i),cd4
+
foxp3
+
(8j),cd4
+
粒 酶b
+
(8k)和用pbs(n=5)或heat-mva(n=5;***,p《0.001;****, p《0.0001)处理的小鼠肿瘤内的cd4
+
ki67
+
(8l)细胞。显示一个代表 性的实验,重复两次。
[0061]
图9是显示了heat-mva诱导肿瘤引流淋巴结(tdln)的免疫学 变化数据的一系列图形。图9a-d是从pbs处理(n=5)或heat-mva 处理(n=5)小鼠的tdln分离的颗粒酶b
+
cd8
+
(9a),颗粒酶b
+
cd4
+ (9b),ki-67
+
cd8
+
(9c)和ki67
+
cd4
+
(9d)细胞的流式细胞术分析 的散点图。图9e-h是描述tdlns(n=5;***,p《0.001;****,p《0.0001) 中颗粒酶b
+
cd8
+
(9e),ki67
+
cd8
+
(9f),颗粒酶b
+
cd4
+
(9g)和 ki67
+
cd4
+
(9h)的比例的图形。显示一个代表性的实验,重复两次。
[0062]
图10是一系列数据的图形,显示与野生型对照相比,heat-mva在 消除sting缺陷小鼠或batf3缺陷小鼠中的b16-f10黑色素瘤的效果 较差。图10a是在携带肿瘤的wt,sting
gt/gt
和batf3-/-小鼠中注射 pbs或heat-mva后的肿瘤体积v时间(天)的图。图10b是用pbs 或heat-mva(不同组的n范围5-8;**,p《0.01;****,p《0.0001)处 理的肿瘤携带wt,sting
gt/gt
和batf3-/-小鼠的kaplan-meier存活曲线。 显示一个代表性的实验,重复两次。
[0063]
图11是显示heat-mva诱导的抗肿瘤作用主要由cd8
+
t细胞介导 和cd4
+
t细胞有助于发展针对肿瘤再次攻击的全身免疫的数据的一系 列图形。图11a是单侧b16-f10黑色素瘤植入模型中,存在或不存在 cd4
+
,cd8
+
和nk细胞耗竭抗体的情况下,肿瘤内注射heat-mva的 示意图。图11b是在同种型对照、cd4
+
,cd8
+
和nk细胞耗竭性抗体 存在下,用pbs或heat-mva处理的小鼠的kaplan-meier存活曲线(n =10;*,p《0.05;**,p《0.01;****,p《0.0001)。图11c-g是各种治 疗方案,包括pbs(11c),heat-mva+同种型对照(11d),heat-mva+ 抗-cd8(11e),heat-mva+抗cd4(11f)和heat-mva+抗nk(11g) 处理后的肿瘤体积对处理后天数做的图。图11h是在初始小鼠和用heat-mva处理,在存在或不存在cd4
+
和nk细胞耗竭的情况下,在右 侧植入原始肿瘤的存活小鼠的左侧真皮内植入致死剂量1
×
106个 b16-f10细胞的肿瘤再次攻击的示意图。图11i是初始小鼠(闭合圆圈, n=6),heat-mva处理的小鼠(实心圆圈,n=10),heat-mva处理的 带nk耗竭的小鼠(填充正方形,n=6),heat-mva处理的带cd4
+
t 细胞耗竭小鼠(实心三角形,n=6)在对侧用致死剂量的b16-f10黑 色素瘤细胞再次攻击的kaplan-meier存活曲线(*,p《0.05;**,p《0.01; ****,p《0.0001)。显示一个代表性的实验,重复两次。
[0064]
图12是显示肿瘤内注射依赖于sting和batf3的heat-mva诱导 的抗黑色素瘤抗体应答的数据的一系列图示。图12a是用heat-mva 或pbs(nt,无血清处理对照)处理的sting
gt/gt
,batf3-/和年龄匹 配的wt小鼠的血清中抗黑色素瘤抗体浓度(通过elisa测定)的散 点图。图12b是用heat-mva或pbs(nt,无血清处理对照)处理的 sting
gt/gt
,batf3-/和年龄匹配的wt小鼠血清中抗痘苗病毒抗体浓度 (通过elisa测定)的散点图。显示一个代表性的实验,重复两次。
[0065]
图13a-d是各种治疗方案(包括肿瘤内注射pbs加腹膜内递送同 种型抗体对照(13a,n=5),肿瘤内注射pbs加腹膜内递送抗ctla-4 抗体(13b,n=5),肿瘤内注射heat-mva加同种型对照(13c,n=10), 肿瘤内注射heat-mva加腹腔内递送抗ctla-4(13d,n=9))后的 肿瘤体积对时间(天数)的图。图13e是用pbs+同种型,heat-mva+ 同种型,pbs+抗ctla-4和heat-mva+抗-cctla4抗体处理的小鼠开 始病毒注射时的肿瘤体积的散点图。图13f是用pbs+同种型, heat-mva+同种型,pbs+抗ctla-4和heat-mva+抗ctla4抗体处理 的荷瘤小鼠的kaplan-meier存活曲线(*,p《0.05;**,p《0.01;****, p《0.0001)。显示一个代表性的实验,重复两次。
[0066]
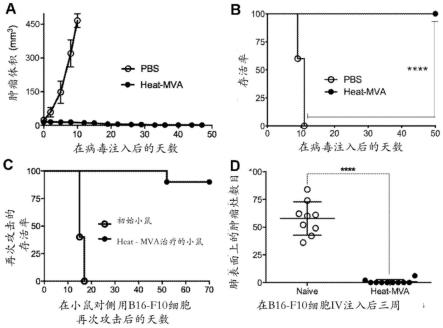
图14是一系列数据的图形,显示在根除注射的肿瘤以及控制非注 射的肿瘤的生长中,肿瘤内注射heat-mva比mva更有效。图14a是 其中在双侧真皮内肿瘤植入模型中b16-f10黑色素瘤用mva或 heat-mva肿瘤内处理的治疗方案的流程。图14b是moi为5(空心圆) 或0.05(实心圆)时mva在b16-f10细胞中的复制曲线图。图14c是 pbs(空心圆圈,n=7),mva(填充正方形,n=9)或heat-mva(实 心圆,n=9)处理的荷瘤小鼠的kaplan-meier存活曲线(**,p《0.01;***, p《0.001)。图14d-e是注射(d)和非注射(e)肿瘤体积相对于pbs 注射后的时间(天)的曲线图。图14f-g是注射(f)和非注射(g) 肿瘤体积对mva注射后的时间(天)的曲线图。图14h-i是注射(h) 和非注射(i)肿瘤体积对heat-mva注射后的时间(天)的曲线图。 显示一个代表性的实验,重复两次。
[0067]
图15是一系列数据的图示,其显示了在双侧b16-f10黑色素瘤模 型中,在非注射的肿瘤中,heat-mva的肿瘤内注射比mva或pbs更 有效召集和激活免疫细胞。用肿瘤内注射pbs,mva或heat-mva处 理肿瘤小鼠,如图14a所述。总共两次治疗,在第一次治疗后第7天 收集未注射的肿瘤。通过facs分析肿瘤浸润性免疫细胞。图15a是 肿瘤内注射pbs,mva或heat-mva至对侧肿瘤后,每克未注射肿瘤 的肿瘤浸润性cd45
+
,cd103
+
cd11c
+
,cd3
+
和cd8
+
的绝对数量的图。 图15b是肿瘤内注射pbs(n=5),mva(n=5)或heat-mva(n=5) 后每克未注射肿瘤的肿瘤浸润粒细胞颗粒酶b
+
cd8
+
,ki67
+
cd8
+
,颗粒 酶b
+
cd4
+
和ki67
+
cd4
+
的绝对数量的图。数据是平均值
±
sem(n=5)。 显示一个代表性的实验,重复两次。
[0068]
图16是数据的一系列图示,其显示与双侧肿瘤植入模型中的野生 型对照相比,heat-mva在sting缺陷小鼠或batf3缺陷小鼠中根除 b16-f10黑色素瘤的效果较差。图16a-b是注射(a)和非注射(b) 肿瘤体积对pbs注射后时间(天)的图(n=6)。图16c-d是注射(c) 和非注射(d)肿瘤体积对wt小鼠(n=10)中注射heat-mva后的 时间(天)的图。图16e-f是注射(e)和非注射(f)肿瘤体积对在 sting
gt/gt
小鼠(n=8)中注射heat-mva后的天数的图。图16g-h是 注射(g)和非注射(h)肿瘤体积对在batf3-/-小鼠(n=6)中注射 heat-mva后的天数的图。图16i是用pbs或heat-mva处理的携带肿 瘤的wt,sting
gt/gt
和batf3-/-小鼠的kaplan-meier存活曲线(**,p 《0.01;****,p《0.0001)。显示代表性实验,重复一次。
[0069]
图17是数据的一系列图示,显示在双侧b16-f10黑色素瘤模型中, 在wt小鼠中的肿瘤内注射heat-mva比在batf3-/-小鼠中的注射和非 注射的肿瘤中召集和激活免疫细胞更有效。用肿瘤内注射pbs或 heat-mva处理荷瘤小鼠,如图14a所述。总共两次治疗,第一次治疗 后第7天收集未注射的肿瘤。通过facs分析肿瘤浸润性免疫细胞。图 17a-b是在wt和batf3-/-老鼠的右侧肿瘤肿瘤内注射pbs或 heat-mva后每克注射(a)和非注射(b)肿瘤的肿瘤浸润cd3
+
和cd8
+
的绝对数量的图。图17c-d是在wt和batf3-/-小鼠右侧肿瘤肿瘤内注 射pbs或heat-mva后每克注射(c)和非注射(d)肿瘤的肿瘤浸润 性ki67
+
cd8
+
和ki67
+
cd4
+
细胞的绝对数量的图(**,p《0.01;***,p 《0.001;****,p《0.0001)。数据是平均值
±
sem(n=4)。显示一个代 表性的实验,重复两次。
[0070]
图18是一系列数据的图形,其显示heat-mva的肿瘤内注射与抗 ctla-4,抗pd-1或抗pd-l1抗体的全身递送的组合显着增加了总体 反应和肿瘤动物治愈率。图18a是治疗计划的方案,其中b16-f10黑 色素瘤在双侧皮内肿瘤植入模型中用肿瘤内递送pbs或heat-mva结 合或不结合全身递送免疫检查点阻断抗体进行处理。图18b是用pbs (n=5),heat-mva+同种型对照(n=10),heat-mva+抗-cctla4抗 体(n=10),heat-mva+抗pd1抗体(n=
10)或heat-mva+抗pd-l1 抗体处理的荷瘤小鼠的kaplan-meier存活曲线(n=10;*,p《0.05;**, p《0.01;****,p《0.0001)。图18c-d是pbs注射后几天的注射(c) 和非注射(d)肿瘤体积的图。图18e-f是肿瘤内注射heat-mva和腹 膜内递送同种型对照后几天的注射(e)和非注射(f)肿瘤体积的图。 图18g-h是肿瘤内注射heat-mva和腹膜内递送抗ctla-4抗体后的 几天中注射(g)和非注射(h)肿瘤体积的图。图18i-j是肿瘤内注射 heat-mva和腹膜内递送抗pd-1抗体后的几天内注射(i)和非注射(j) 肿瘤体积的图。图18k-l是肿瘤内注射heat-mva和腹膜内递送抗 pd-l1抗体后的几天内注射(k)和非注射(l)肿瘤体积的图。显示 一个代表性的实验,重复两次。
[0071]
图19是一系列数据的图形,显示了uv灭活的mva(uv-mva) 通过sting/irf3依赖的细胞溶质dna感测途径在cdc中诱导i型 ifn。图19a-b是mva(实心圆),heat-mva(空心正方形)或uv-mva (填充的正方形)感染cdc后培养基中分泌的ifn-α(a)和ifn-β (b)的浓度的曲线图(***,p《0.001)。数据是平均值
±
sem(n=3)。 显示一个代表性的实验,重复两次。图19c-d是从sting
gt/gt
,irf3-/-和 wt对照小鼠产生的并用uv-mva感染的cdc培养基中分泌的ifn
‑ꢀ
α(c)和ifn-β(d)浓度的柱状图。数据是平均值
±
sem(n=3)。 显示一个代表性的实验,重复两次。图19f是uv-mva感染后(hpi, 感染后小时;nt,无治疗)的sting
+/+
和sting
gtgt cdc中p-irf3,irf3, sting和β-肌动蛋白的蛋白质水平的western印迹的扫描图像。
[0072]
图20是一系列数据的图形,显示了uv灭活的mva(uv-mva) 诱导来自mc38结肠腺癌细胞系的炎性细胞因子和趋化因子,并且肿瘤 内注射uv-mva导致肿瘤根除和全身抗肿瘤免疫的发展具有与 heat-mva类似功效。图20a-c是感染mva,heat-mva,uv-mva或 模拟对照的mc38细胞培养基中分泌的il-6(a),ccl4(b)和ccl5 (c)的浓度的柱状图。数据是平均值
±
sem(n=3)。显示一个代表 性的实验,重复两次。图20d-f是肿瘤内注射pbs(d),heat-mva(e) 或uv-mva(f)后的肿瘤体积对时间(天)的图。图20g是用pbs (实心圆圈,n=5)或heat-mva(填充正方形,n=10)或uv-mva (实心三角形,n=7)注射的荷瘤小鼠的kaplan-meier存活曲线)。图 20h是初始小鼠(闭合圆圈,n=5),heat-mva处理的小鼠(填充正 方形,n=7)和uv-mva处理的小鼠(实心三角形,n=5),通过致死 剂量的mc38结肠腺癌细胞在对侧再次攻击的kaplan-meier存活曲线。 显示代表性实验,重复一次。
[0073]
图21是数据的一系列图形,显示了heat-mva的肿瘤内注射与抗 ctla-4或抗pd-l1抗体的全身递送的组合显着增加了携带肿瘤的动物 的总体反应和治愈率。图21a-b是pbs注射后,注射(a)和非注射 (b)肿瘤体积相对于时间(天)的图。图21c-d是肿瘤内注射pbs 和腹膜内递送抗ctla-4抗体后,注射(c)和非注射(d)肿瘤体积 对时间(天)的图。图21e-f是肿瘤内注射pbs和腹膜内递送抗-pd-l1 抗体后,注射(e)和非注射(f)肿瘤体积对时间(天)的图。图21g-h 是肿瘤内注射heat-mva和腹膜内递送同种型抗体对照后,注射(g) 和非注射(h)肿瘤体积与时间(天)的曲线图。图21i-j是肿瘤内注 射heat-mva和腹膜内递送抗ctla-4抗体后,注射(i)和非注射(j) 肿瘤体积与时间(天)的曲线图。图21k-l是肿瘤内注射heat-mva 和腹膜内递送抗pd-l1抗体后,注射(k)和非注射(l)肿瘤体积与 时间(天)的曲线图。图21m是用pbs(n=6),抗ctla4抗体(n= 7)或抗pd-l1抗体(n=7;***,p《0.001)处理的荷瘤小鼠的 kaplan-meier存活曲线。图21n是用pbs(n=6),heat-mva+同种型 对照(n=10),heat-mva+抗ctla4抗体(n=10),heat-mva+抗pd-l1 抗体(n=10;*,p《0.05;**,p《0.01;****,p《0.0001)处理的荷瘤小 鼠的kaplan-meier存活曲线。显示代表性实验,重复
一次。
[0074]
图22是数据的一系列图形,显示了在双侧b16-f10肿瘤植入模型 中,heat-mva和抗ctla-4的共同给药显着增加了总体反应和治愈率。 图22a-b是pbs(n=10)注射后,注射(a)和非注射(b)肿瘤体 积相对于时间(天)的图。图22c-d是肿瘤内注射heat-mva和同种 型对照抗体(n=10)后,注射(c)和非注射(d)肿瘤体积与时间(天) 的曲线图。图21e-f是在肿瘤内共同给予heat-mva和用于腹膜内输送 的剂量的十分之一的抗ctla-4抗体(n=10)后,注射(e)和非注 射(f)肿瘤体积的图表对时间(天)的曲线图。图22g-h是肿瘤内注 射heat-mva和腹膜内递送抗ctla-4抗体(n=10)后,注射(g) 和非注射(h)肿瘤体积对时间(天)的曲线图。
[0075]
详细说明
[0076]
定义:
[0077]
如本文所用,以下术语具有以下含义,除非上下文另有明确说明:
[0078]“癌症”是指一类以不受控制的细胞生长为特征的人类和动物疾 病。除非另有明确说明,否则术语“癌症”可以与术语“肿瘤”,(其又 包括原发性和转移性肿瘤)“恶性肿瘤”,“过度增生”和“肿瘤”互换 使用。术语“癌细胞”可以与术语“肿瘤细胞”,“恶性细胞”,“过度增 殖细胞”和“肿瘤细胞”互换。
[0079]“黑色素瘤”是指源于能够产生黑色素的细胞的恶性肿瘤。术语黑 色素瘤是“恶性黑色素瘤”的代名词。黑色素瘤广泛转移,涉及患者的 淋巴结,皮肤,肝,肺和脑组织。
[0080]“实体肿瘤”是指除如淋巴瘤,白血病和多发性骨髓瘤的血液性癌 症之外的所有赘生性细胞生长和增殖,原发性或转移性,以及所有癌前 和癌细胞和组织。实体肿瘤的实例包括但不限于:纤维肉瘤,粘液肉瘤, 脂肪肉瘤,软骨肉瘤,骨肉瘤,脊索瘤,血管肉瘤,内皮肉瘤,淋巴管 肉瘤,淋巴管内皮肉瘤,滑膜瘤,间皮瘤,尤因氏肿瘤,平滑肌肉瘤, 横纹肌肉瘤,结肠癌,胰腺癌,乳腺癌,卵巢癌,前列腺癌,鳞状细胞 癌,基底细胞癌,腺癌,汗腺癌,皮脂腺癌,乳头状癌,乳头状腺癌, 囊腺癌,髓样癌,支气管癌,肾细胞癌,肝癌,胆管癌,绒毛膜癌,精 原细胞瘤,胚胎癌,wilms肿瘤,宫颈癌,睾丸肿瘤,肺癌,小细胞肺 癌,膀胱癌,上皮癌,神经胶质瘤,星形细胞瘤,成神经管细胞瘤,颅 咽管瘤,室管膜瘤,松果体瘤,血管母细胞瘤,声神经瘤,少突胶质细 胞瘤,脑膜瘤,黑色素瘤,神经母细胞瘤和视网膜母细胞瘤。本公开的 组合物和方法可用的一些最常见的实体瘤包括:头颈癌,直肠腺癌,胶 质瘤,成神经管细胞瘤,尿路上皮癌,胰腺癌,子宫内膜癌,卵巢癌, 前列腺腺癌,非小细胞肺癌(鳞状细胞癌和腺癌),小细胞肺癌,黑色 素瘤,乳腺癌,肾细胞癌和肝细胞癌。
[0081]“转移”是指癌症从其主要部位扩散到身体中的相邻组织或远端位 置。癌细胞可以脱离原发性肿瘤,穿透淋巴管和血管,通过血液循环, 并在身体其他地方的正常组织中生长。转移是一个连续的过程,取决于 肿瘤细胞(或癌干细胞)从原发性肿瘤中脱离出来,穿过血流或淋巴管, 并停留在远处。一旦在另一个位点,癌细胞再次穿透血管或淋巴管,继 续扩展,最终形成新的肿瘤(转移性肿瘤)。在一些实施方案中,该新 肿瘤被称为转移性(或继发性)肿瘤。
[0082]“免疫应答”是指淋巴细胞,抗原呈递细胞,吞噬细胞,粒细胞和 由上述细胞或肝脏产生的可溶性大分子(包括抗体,细胞因子和补体) 中的一种或多种的作用,其导致选择性损伤,破坏或消除人体的癌细胞, 转移性肿瘤细胞等。免疫应答可以包括细胞反应,例如
75]。牛痘病毒感染 许多类型的细胞,但是优先在肿瘤细胞中复制,这是因为肿瘤细胞具有 有利于复制的代谢,表现出某些途径的激活,这些途径也有利于复制, 并创造了逃避先天免疫系统的环境,这也有利于病毒复制。热灭活的 mva不符合溶瘤病毒的定义。
[0092]“mva”是指“修饰的痘苗安卡拉”,是指来自安卡拉菌株的高度 减毒的牛痘菌株,开发用作疫苗和疫苗佐剂。通过连续通过鸡胚胎细胞 从野生型安卡拉菌株分离出原始的mva。由此,它损失了野生型牛痘 基因组的大约15%,包括其在灵长类动物(包括人)细胞中有效复制 的能力[76]。天花疫苗菌株mva:标记,遗传结构,肠胃外接种获得 的经验和具有虚弱防御机制的生物体内的行为。mva被认为是发展作 为针对传染病或肿瘤的基因或疫苗接种递送的重组载体的合适候选者 [77]。mva具有长度为178kb的基因组和首先在antoine,g等人[78] 中公开的序列。序列也在genbank u94848.1中公开。临床级mva是从 丹麦的巴伐利亚北欧a/s kvistgaard商业和公开获得的。此外,mva 可从atcc,rockville,md和cmcn(institut pasteur collection nationaledes microorganismes)paris,france获得。突变体mva e3l敲除(δ e3l-mva)及其制备已经在美国专利7,049,145中有所描述。
[0093]“热灭活的mva”或“加热mva(heat-mva)”是指在不破坏其 免疫原性或其进入靶细胞(肿瘤细胞)的能力但是除去病毒的残留复制 能力以及抑制宿主免疫应答的因子(例如,抑制感染细胞中ifn i诱导 的因子)的条件下通过暴露于热进一步处理的mva。这种条件的一个 例子是暴露在约50至约60℃范围内的温度约一小时的时间。其他时间 和温度可以通过常规实验确定,并且在感染的cdc的i型ifn诱导可 以与本文所述实验中使用的heat-mva进行比较,应高于mva。在本 发明人进行的一个实验中,通过用65℃和1小时暴露组合处理的mva 对cdc的感染不能诱导i型ifn。这种安全性和强免疫原性的组合使得 与wt牛痘甚至mva相比,heat-mva特别有吸引力。
[0094]“uv灭活的mva”或“uv-mva”是指在不破坏其免疫原性或其 进入靶细胞的能力(肿瘤细胞)但消除病毒的残留复制能力的条件下已 经通过暴露于uv灭活的mva。在本方法中有用的这种条件的实例是 使用例如365nm紫外灯泡暴露于uv约30分钟至约1小时的时间[56,79]。再次,如上所述的heat-mva所述,这些uv波长和暴露条件 的限制可以通过常规实验来确定,其通过确定接受给定暴露的uv-mva 诱导的i型ifn,并将其与i型ifn诱导通过以下实验中使用的uv-mva 和未处理的mva。uv-mva与heat-mva类似也是同样安全的,并且 也诱导显着的i型ifn。
[0095]
因此,“灭活的mva”应被用作包含热灭活的mva和uv灭活的 mva的通用术语,其是感染性的,非复制性的并且不抑制感染的dc 细胞中的i型ifn产生。通过热和紫外线辐射的组合灭活的mva也在 本公开的范围内。
[0096]“受试者”是指可能患有癌症的任何动物(哺乳动物,人类或其他) 患者。
[0097]“治疗有效量”或“有效量”是指以一种或多种剂量施用足量的药 剂,并且足以在减轻,治愈或缓解疾病方面提供所需生物学结果的时间。 在本公开内容中,有效量的灭活mva是(以适当的时间段和合适的频 率施用)减少癌细胞数量的量;或减少肿瘤大小或根除肿瘤;或抑制(即 减慢或停止)癌细胞浸润到周围器官中;抑制(即减慢或停止)转移性 生长;抑制(即稳定或阻止)肿瘤生长;允许治疗肿瘤,和/或诱导针对肿 瘤的免疫应答。根据本公开,本领域普通技术人员可以使用常规实验来 确定任何个别情况下的适当治疗量。这
样的测定将从在体外有效的量开 始,发现在动物中有效的量。治疗有效量将首先基于发现赋予培养细胞 益处的浓度或浓度来确定。有效量可以从细胞培养物中的数据外推,并 且可以基于诸如本文详细的因素来上调或下调。有效量范围的实例是每 次施用105个病毒颗粒至约10
12
个病毒颗粒。
[0098]
特别参考本文公开的基于病毒的免疫刺激剂,“治疗有效量”或“有 效量”是指包含足以减少,抑制或消除肿瘤细胞生长的灭活mva的组 合物的量,从而减少或消除肿瘤,或足以抑制,减少或消除体外或受试 者的转移性扩散,或引起针对肿瘤的免疫应答,根据情况可最终导致还 原,抑制和/或废除的一种或多种。肿瘤细胞生长的减少,抑制或根除 可能是坏死,细胞凋亡或免疫应答或两种或多种前述组合的结果。治疗 有效的量可以根据组合物中使用的特定灭活的mva,所治疗的受试者 的年龄和状况,肿瘤形成的程度,其它治疗方式的存在或不存在等因素 而变化。类似地,待施用的组合物的剂量和其施用的频率将取决于多种 因素,例如活性成分的效力,一旦施用其活性的持续时间,给药途径, 大小,年龄,受试者的性别和身体状况,不良反应的风险和医生的判断。 组合物以各种剂型施用,这些可注射溶液。
[0099]
特别参考与免疫检查点抑制剂的联合治疗,免疫检查点阻断剂的
ꢀ“
治疗有效量”是指足以阻断免疫检查点以避免被治疗的受试者的肿瘤 细胞中的细胞凋亡反应的免疫检查点封闭剂的量,在临床试验中还有待 开发的几种免疫检查点阻断剂,包括cd28抑制剂,如ctl4抑制剂(如 伊曲利单抗),pd-1抑制剂(如尼泊洛珠单抗,彭美罗珠单抗,哌替珠 单抗,lambrolizumab)pd-l1抑制剂mpdl3280a,bms-936559, medi4736,msb 00107180)icos和btla或其诱饵分子。前述的剂 量范围在本领域的技术人员中已知或容易掌握,因为几个给药临床试验 已经完成,从而使外推到其他试剂成为可能。
[0100]
优选地,肿瘤表达特定的检查点,但是这不是严格必要的,因为免 疫检查点阻断剂更一般地阻止肿瘤细胞,基质细胞和肿瘤浸润性免疫细 胞引起的免疫抑制机制。
[0101]
例如,ctla4抑制剂ipilimumab当在黑色素瘤手术后作为辅助治 疗作用时,以1-2mg/ml在90分钟内给药,总共输注量为3mg/kg每 3周共4次。这种治疗通常伴有严重的甚至危及生命的免疫介导的不良 反应,这限制了耐受剂量以及可以施用的累积量。预期当与灭活的mva 一起给药时,可以减少ipilimumab的剂量和/或累积量。特别是根据以 下实验结果,如果将ctla4抑制剂的剂量直接施用于同时或依次用灭 活的mva,则可以进一步降低ctla4抑制剂的剂量。因此,以上为 ipilimumab提供的量将是确定联合施用给予患者的特定剂量和累积量 的起点,但是需要进行剂量研究以确定最佳量。
[0102]
彭博拉珠单抗用于治疗黑色素瘤中的辅助治疗,稀释至25mg/ml, 以每3周在30分钟内2mg/kg的剂量施用。
[0103]
nivolumab用于治疗,以60分钟内静脉输注3mg/kg的剂量每两 周给药。
[0104]“药学上可接受的赋形剂”包括药学上可接受的载体或稀释剂,例 如任何和所有溶剂,分散介质,包衣,等渗和吸收延迟剂等,还包括防 腐剂和抗细菌剂和抗真菌剂。对于生物活性物质是本领域熟知的。赋形 剂的进一步细节在下面提供。
[0105]“递送”用于将本公开的灭活的mva放置在肿瘤微环境中,无论 这是通过局部施用于肿瘤还是通过全身给药,例如静脉内途径进行。该 术语重点是灭活mva到达肿瘤本身。
[0106]
本文中“联合给药”是指与灭活的mva组合给药第二治疗方式, 例如施用免疫检查
点阻断剂并且与灭活的mva紧密地临时接近。例如, pd-1/pdl-1抑制剂和/或ctla4抑制剂(在更具体的实施方案中,抗 体)可以与热灭活的mva同时施用(通过静脉内或肿瘤内注射,当给 予灭活的mva时肿瘤内或系统地如上所述)或在灭活的mva给药之 前或之后。如果灭活的mva递送和免疫检查点封闭剂给药分开1-7天 或甚至长达三周,则这将在本文所述的“紧密的临时接近”内。
[0107]
***
[0108]
在一个实施方案中,本公开内容涉及在具有肿瘤的受试者中引发抗 肿瘤免疫应答的方法,包括向肿瘤递送一定量的灭活的mva,以有效 地产生以下一种或多种:
[0109]
增加肿瘤内和/或肿瘤引流淋巴结中的细胞毒性cd8
+
t细胞;
[0110]
通过诱导i型ifn诱导浸润所述肿瘤的树突细胞的成熟;
[0111]
诱导受试者的效应t细胞识别肿瘤内和/或肿瘤引流淋巴结中的肿 瘤细胞;
[0112]
减少肿瘤内免疫抑制(调节)cd4
+
t细胞;和
[0113]
诱导肿瘤细胞在其表面上表达i型mhc并产生一种或多种i型ifn 或其它炎性细胞因子或趋化因子。
[0114]
本发明人探索了免疫应答的机制,并得出结论,其由介导1型ifn 生产的cgas/sting介导的细胞溶质dna感测途径引发。在实施例 中提供了引发的机制和召集的免疫细胞的进一步了解。其中提出的结论 并不局限于具体的实验环境,这些实验环境阐明这些机制。
[0115]
在一个实施方案中,本公开提供了治疗被诊断患有实体瘤的受试者 的方法,包括向肿瘤递送治疗有效量的本文所述的加热mva。
[0116]
在一个实施方案中,本公开提供了在诊断患有癌症的受试者中诱导 抗肿瘤免疫的方法,包括向受试者施用治疗有效量的灭活的mva。本 公开的方法包括可以减少肿瘤大小,根除肿瘤,抑制肿瘤生长或抑制肿 瘤转移或转移生长的抗肿瘤免疫的诱导。
[0117]
在另一个实施方案中,本公开提供了用于增强,刺激或引诱被诊断 患有实体恶性肿瘤的受试者的抗肿瘤免疫应答,其可包括先天免疫应答 和/或适应性免疫应答,例如通过将肿瘤暴露于治疗有效量的灭活mva 的t细胞应答。
[0118]
在具体实施方案中,本公开提供了引发介导同时针对肿瘤细胞的t 细胞细胞毒性和针对肿瘤细胞的t辅助细胞的适应性免疫应答的方法。 所述方法包括向肿瘤内或静脉内给予受试者包含非复制型热灭活或uv 灭活的mva的组合物,其中所述组合物的施用导致针对肿瘤的肿瘤特 异性免疫应答,并且最终导致肿瘤的减少,抑制或消除生长和/或抑制 转移生长。实际上,本发明人已经表明癌细胞正在被杀死,并且免疫应 答可以迁移到偏远的位置,如在转移的情况。
[0119]
在一些实施方案中,本公开提供了引发介导同时针对肿瘤细胞的t 细胞细胞毒性和针对肿瘤细胞的t辅助细胞的适应性免疫应答的方法。 所述方法包括向受试者胃肠外施用包含灭活的mva的组合物,其中所 述组合物的施用导致针对肿瘤的肿瘤特异性免疫应答,并且最终减少, 抑制或消除肿瘤生长和/或抑制转移性生长。实际上,本发明人已经表 明癌细胞正在被杀死,并且免疫应答可以迁移到偏远的位置,如在转移 的情况。
[0120]
因为灭活的mva不具有复制能力,它不会像复制型疫苗或载体一 样对免疫系统发挥其作用。因此,虽然认为免疫系统的刺激是对溶瘤疗 效的障碍[19],但是灭活的mva能够
利用先天免疫系统来刺激适应性 免疫,无论是在抗肿瘤的细胞毒性方面还是更广泛的t效应细胞激活。
[0121]
因此,本发明提供了一种治疗实体恶性肿瘤的方法,向受试者的肿 瘤递送在被诊断患有实体瘤的受试者中有效地使细胞毒性cd8
+
细胞增 加并减少肿瘤中调节性cd4
+
细胞并诱导免疫应答的量的灭活的mva。
[0122]
本公开还提供了通过治疗固体恶性肿瘤产生抗肿瘤全身性免疫的 方法,包括向受试者的肿瘤递送一定量的灭活的mva,以有效地引起 非注射的肿瘤中的免疫细胞的可观甚至是显着的增加,包括 cd103
+
dcs,细胞毒性cd8
+
细胞和cd4
+
效应细胞,从而引起所述受试 者中非注射的肿瘤的排斥反应和抗肿瘤转移的一种或两种(本发明人通 过肿瘤再刺激测试)。
[0123]
改良牛痘安卡拉(mva)
[0124]
改良牛痘安卡拉(mva)病毒是痘病毒科家族中的直链痘病毒属的 成员。mva是由安卡拉痘苗病毒株(cva)在鸡胚成纤维细胞(cef) 的大约570个连续传代产生的[80]。作为这些长期传代的结果,所得到 的mva病毒含有广泛的基因组缺失,并且是限制于禽类细胞的高度宿 主细胞[30]。在各种动物模型中显示所得mva显著无致病力[76]。
[0125]
mva的安全性和免疫原性已经在临床试验中进行了广泛的测试和 记录,特别是针对人类天花病。这些研究包括超过12万人,并且在人 类中表现出极好的疗效和安全性。此外,与其他痘苗疫苗相比,mva 已经削弱了毒力(感染性),同时触发了良好的特异性免疫应答。因此, 已经建立了mva作为安全的疫苗载体,具有诱导特异性免疫应答的能 力。
[0126]
由于上述特征,mva成为用于重组基因表达和疫苗的工程化mva 载体开发的有吸引力的目标。作为疫苗载体,已针对许多病理状况(包 括艾滋病毒,肺结核和疟疾以及癌症)对mva进行了研究[33,34]。
[0127]
已经证明,人单核细胞衍生的树突状细胞(dc)的mva感染导致 dc活化,其特征在于共刺激分子的上调和促炎细胞因子的分泌[56]。 在这方面,mva与标准野生型牛痘病毒(wt-vac)不同,其不能激 活dc。树突状细胞可以分为两个主要亚型:常规树突状细胞(cdcs) 和浆细胞样树突状细胞(pdc)。前者,特别是cd8
+
亚型,特别适用于 向t细胞呈递抗原;后者是i型ifn的强力生产者。
[0128]
人类细胞的病毒感染导致由i型干扰素(特别是干扰素-α(α)) 介导的先天免疫应答(第一道防线)的激活。这通常导致免疫学“级联
”ꢀ
的活化,其中活化的t细胞(ctl和辅助细胞)的召集和增殖以及最 终抗体的产生。然而,病毒表达抑制宿主免疫反应的因子。mva是比 wt-vac更好的免疫原,在哺乳动物细胞中复制不良[81]。
[0129]
然而,它并不完全是非复制性的,并且如本发明人所显示的,含有 一些免疫抑制活性。
[0130]
免疫反应
[0131]
除了通过上调特定免疫系统活性(例如抗体和/或细胞因子产生或 细胞介导的免疫的活化)来诱导免疫应答外,免疫应答还可以包括抑制, 衰减或任何其它下调可检测的免疫力,从而重建体内平衡,防止宿主自 身器官和组织的过度损伤。在一些实施方案中,根据本公开的方法诱导 的免疫应答产生细胞毒性cd8
+
t细胞或活化的t辅助细胞或同时两者, 可直接或间接地导致肿瘤细胞的死亡或散播能力的丧失。
[0132]
通过本公开的方法诱导的免疫应答可以通过检测各种众所周知的 免疫学参数中的任何一种来确定[82,83]。因此,免疫应答的诱导可以通 过许多众所周知的测定中的任何一种进行建立,包括免疫学测定。此类 测定包括但不限于体内,离体或体外测定可溶性免疫球蛋白或抗体;可 溶性介质如细胞因子,趋化因子,激素,生长因子等以及其他可溶性小 肽,碳水化合物,核苷酸和/或脂质介质;通过免疫系统的细胞的功能或 结构特性的改变,例如细胞增殖,改变的移动性,改变的细胞内阳离子 梯度或浓度(如钙)来确定细胞活化状态变化。细胞多肽的磷酸化或去 磷酸化;诱导特异性基因表达或细胞溶解行为等活动;免疫系统细胞的细 胞分化,包括改变的表面抗原表达谱,或凋亡的起动(程序性细胞死亡); 或可检测出免疫应答存在的任何其它标准。例如,区分免疫细胞类型的 细胞表面标志物可以通过结合cd4
+
,cd8
+
或nk细胞的特异性抗体来 检测。可以检测的其他标志物和细胞成分包括但不限于干扰素γ(ifn
‑ꢀ
γ),肿瘤坏死因子(tnf),ifn-α,ifn-β,il-6和ccl5。用于检 测免疫应答的常见方法包括但不限于流式细胞术,elisa,免疫组织化 学。用于进行这些和类似测定的程序是广为人知的,可以在letkovits (immunology methods manual:the comprehensive sourcebook oftechniques,current protocols in immunology,1998)中找到。
[0133]
药物组合物和制剂
[0134]
包含灭活的mva的药物组合物可以含有一种或多种药学上可接受 的赋形剂,例如载体或稀释剂。这些是不妨碍本公开的疫苗组分的活性 或有效性且不具有毒性的成分。载体或稀释剂可以是溶剂或分散介质, 其含有例如水,葡萄糖溶液,多元醇(例如,甘油,丙二醇和液体聚乙 二醇等),血清白蛋白,林格氏溶液,其合适的混合物和植物油。例如 通过使用诸如卵磷脂的涂层可以维持适当的流动性,通过在分散体的情 况下维持所需的粒径,并且通过使用表面活性剂和/或润湿剂如十二烷 基硫酸钠或乙醇。防止微生物的作用可以通过各种防腐剂,抗菌剂和抗 真菌剂,例如对羟基苯甲酸酯,氯丁醇,苯酚,山梨酸,硫柳汞,氯化 苄基氯化铵,苄索氯铵等进行。在许多情况下,优选包括等渗剂,例如 糖如甘露糖醇山梨糖醇,乳糖或氯化钠或氯化钾。可以通过在组合物中 使用延迟吸收的试剂,例如单硬脂酸铝和明胶来延长可注射组合物的吸 收。
[0135]
包含灭活的mva的药物组合物和制剂可以通过常规的混合,溶解, 乳化或冻干方法制备。药物病毒组合物可以使用一种或多种生理上可接 受的载体,稀释剂,赋形剂或辅助剂以常规方式配制,其有助于配制适 于体外,体内或离体使用的病毒制剂。组合物可以与一种或多种另外的 生物活性剂(例如平行施用gm-csf)组合,并且可与药学上可接受的 载体,稀释剂或赋形剂配制以产生本公开的药物(包括生物制剂)或兽 医组合物适合肠胃外或肿瘤内给药。
[0136]
许多类型的制剂是可能的并且是众所周知的。所选择的具体类型取 决于所选择的给药途径,如本领域公认的。例如,全身制剂通常设计用 于通过注射给药,例如静脉内给药,以及为肿瘤内给药设计的那些。优 选地,全身或肿瘤内制剂是无菌的。
[0137]
通过根据需要,将所需量的适当溶剂中的灭活的mva与本文列举 的各种其它成分并入合适的灭菌手段来制备无菌可注射溶液。通常,通 过将活性成分掺入含有碱性分散介质的无菌载体和从上述列举的所需 的其它成分制备分散体。在用于制备无菌注射溶液的无菌粉末的情况 下,优选的制备方法是真空干燥和冷冻干燥技术,其产生非活性mva 的粉
末加上来自先前无菌过滤的溶液中的任何另外的所需成分。
[0138]
在一些实施方案中,本公开的灭活的mva组合物可以配制在水溶 液中,或者在生理上相容的溶液或缓冲液如hanks溶液,林格氏溶液, 甘露醇溶液或生理盐水缓冲液中。在某些实施方案中,任何灭活的mva 组合物可含有配制剂,例如悬浮,稳定渗透剂或分散剂,缓冲液,冻干 保护剂或防腐剂,例如聚乙二醇,聚山梨酯80,1,1-十二烷基六氢-2h
‑ꢀ
氮杂-2-酮(laurocapran),油酸,柠檬酸钠,tris hcl,葡萄糖,丙二醇, 甘露糖醇,聚山梨醇酯聚乙烯脱水山梨醇单月桂酸酯(tween-20),肉 豆蔻酸异丙酯,苄醇,异丙醇,乙醇蔗糖,海藻糖等本技术可以用于本 发明的任何组合物[84]。
[0139]
本公开的生物或药物组合物可以配制成允许其中包含的病毒可用 于在向受试者施用组合物时感染肿瘤细胞。血清,肿瘤以及如果需要, 其他组织中的病毒水平可以通过各种公认的技术来监测,例如基于抗体 的测定(例如elisa,免疫组织化学等)。
[0140]
灭活mva的剂量
[0141]
通常,受试者以约105至约10
10
个噬菌斑形成单位(pfu)的范围施 用灭活的mva的剂量,尽管可以按照本领域普通技术人员确定施用较 低或较高剂量。在优选的实施方案中,剂量为约10
6-109pfu。该剂量可 以以约1至约10ml的单位剂型配制。pfu与病毒颗粒的等效性可根据 所用的特异性pfu滴定法而不同。通常,pfu等于约5至100个病毒颗 粒。灭活的mva的治疗有效量可以以一个或多个分开的剂量施用规定 的时间段并且以规定的施用频率施用。例如,根据本公开的治疗有效量 的灭活的mva可根据以下因素有所差异:诸如疾病状态,年龄,性别, 体重和受试者的一般状况,肿瘤大小,灭活
–
mva引发所需的足以抵 抗特定受试者的肿瘤的程度的免疫应答的能力,以及受试者的免疫系统 建立此类反应的能力。
[0142]
对于在癌症治疗领域工作的人员显而易见的是,剂量的变化必然会 发生,例如根据所治疗的受试者的状况,给药途径和受试者对治疗的反 应性以及受试者的最大耐受剂量。在向受试者递送灭活的mva时,剂 量也将根据诸如一般医学病症,先前病史,疾病进展,肿瘤负担等因素 而变化。
[0143]
以剂量单位形式配制本公开的组合物可能是有利的,以便于施用和 剂量的均匀性。本文所用的剂量单位形式是指适合作为待治疗的哺乳动 物受试者的单位剂量的物理上离散的单位;每个单位含有与所需药学上 可接受的载体结合产生所需治疗效果的预定量的活性物质。
[0144]
灭活mva的治疗和治疗方案
[0145]
可以使用不同途径的组合(包括肠胃外,例如肿瘤内,或静脉内施 用)来实现灭活的mva的施用。在一个实施方案中,灭活的mva直 接施用于肿瘤,例如,通过肿瘤内注射,其中需要直接的局部反应。此 外,灭活的mva的给药途径可以变化,例如,第一次给药使用肿瘤内 注射,以及随后通过静脉内注射给药,或其任何组合。治疗有效量的灭 活mva注射可以规定的时间段和给定的施用频率给予。在某些实施方 案中,灭活的mva可以与其它治疗性治疗结合使用。例如,灭活的 mva可以在用于大体积原发性肿瘤的受试者的新辅助疗法 (neoadjuvant)(术前)或辅助疗法(adjuvant)(术后)配置中施用。 预期这种优化的治疗方案将诱导针对肿瘤的免疫应答,并且降低主要治 疗之前和/或之后的肿瘤负担,例如手术。此外,灭活的mva可以与其 他治疗性治疗如化学疗法或辐射一起施用。
[0146]
在某些实施方案中,灭活的mva病毒以间隔的间隔重复施用,例 如每周或每月至
少一次,但是如果需要,可以更经常地施用,例如每周 两次,持续数周,数月,数年甚至在允许的情况下无限期长期坚持进行。 可以考虑更频繁的施用,如果它们导致持续或增加的收益。本发明方法 的优点包括但不限于以下:癌细胞数减少,肿瘤大小减少,肿瘤根除, 癌细胞侵入周围器官的抑制,转移生长的抑制或稳定,抑制或抑制稳定 肿瘤生长,稳定或改善生活质量。此外,益处可以包括诱导针对肿瘤的 免疫应答,t辅助细胞的活化,细胞毒性cd8
+
t细胞的增加或调节性 cd4
+
细胞的减少。例如,在黑色素瘤的情况下或,益处可以是在黑色 素瘤的初期诊断的1年,2年,4年,5年或更多年中,缺乏复发或转 移。可以对结肠癌和其他实体瘤进行类似的评估。
[0147]
在某些其他实施方案中,在体内,离体或体外用灭活的mva处理 肿瘤块或肿瘤细胞。
实施例
[0148]
材料和方法
[0149]
通常,本文使用的试剂来自商业来源,或者如果不是,其对应物可 商购或公开获得。
[0150]
病毒和细胞系
[0151]
mva病毒由gerd sutter(慕尼黑大学)提供,在bhk-21(幼仓鼠 肾细胞,atcc ccl-10)细胞中繁殖,但两种材料都是商业上和/或公 开可获得的。病毒通过36%的蔗糖垫提纯。bsc40细胞保持在补充有 5%胎牛血清(fbs),青霉素(100单位/ml)和链霉素(100μg/ml) 的dulbecco改良的eagle培养基(dmem,可购自life technologies, cat#11965-092)毫升)。将bhk-21在含有10%fbs,0.1mm非必需 氨基酸(neaa)和50mg/ml庆大霉素的eagle's minimal essentialmedium(eagle's mem,可购自life technologies,cat#11095-080)中 培养。鼠黑色素瘤细胞系b16-f10最初来自i.fidler(md安德森癌症 中心)。将b16-f10细胞保持在补充有10%fbs,100单位/ml青霉素,100μg/ml链霉素,0.1mm neaa,2mm l-谷氨酰胺,1mm丙酮酸钠 和10mm hepes缓冲液的rpmi 1640培养基中。
[0152]
所有细胞在37℃下在5%co2培养箱中生长。
[0153]
通过在55℃温育纯化的mva病毒1小时产生加热-mva。为了产 生uv-mva,使用365nm uv灯在stratalinker 1800 uv交联剂 (stratagene)中uva照射mva 15分钟。
[0154]
老鼠
[0155]
6至10周龄的雌性c57bl/6j小鼠购自jackson laboratory(库存 #000664),用于制备骨髓衍生的树突状细胞和体内实验。这些小鼠被 保存在sloan kettering研究所的动物设施中。所有程序均严格按照“国 家卫生研究所实验动物护理和使用指南”中的建议执行。该议定书由斯 隆-凯特林癌症研究所动物实验伦理委员会批准。在drs的实验室中 产生cgas-/-,irf3-/-,irf7-/-,irf5-/-,,batf3-/-和sting
gt/gt
小鼠。 陈志坚(德州西南医科大学;cgas-/-),谷口谷口(东京大学;irf3-/-和 irf7-/-),tak mak(多伦多大学;irf5-/-,);肯尼思
·
墨菲(华盛顿大 学;batf3-/-)和罗素
·
万斯(加州大学伯克利分校;sting
gt/gt
)。ifnar1 ‑
/-小鼠由eric pamer博士(sloan kettering institute)提供;小鼠购自b &k universal,与c57bl/6小鼠回交六代以上。在paula m.pitha博士 的实验室将irf5-/-,小鼠与c57bl/6j小鼠回交至少六代,然后转移到 sloan kettering研究所。
[0156]
上述动物的商业来源如下:
[0157][0158]
生成骨髓来源的树突状细胞
[0159]
通过首先从骨骼去除肌肉,然后用含有10%fcs的rpmi的0.5ccu-100胰岛素注射器(becton dickinson)冲洗细胞,收集来自胫骨和小鼠 股骨的骨髓细胞。离心后,通过将细胞在冰上温育1-3分钟,将细胞重悬 于ack裂解缓冲液(lonza)中以进行红细胞裂解。然后收集细胞,重新 悬浮于新鲜培养基中,并通过40-μm细胞过滤器(bd biosciences)过滤。 计数细胞数。对于gm-csf-bmdc的产生,在gm-csf(30ng/ml,由 斯隆-凯特林癌症研究所的单克隆抗体核心设施产生)的存在下,在cm 中培养骨髓细胞(每个15cm细胞培养皿中的500万个细胞)10-12天。cm 是补充有10%胎牛血清(fbs),100单位/ml青霉素,100μg/ml链霉素, 0.1mm必需和非必需氨基酸,2mm l-谷氨酰胺,1mm丙酮酸钠和10mm 的rpmi 1640培养基hepes缓冲液。通过每3天用新鲜培养基代替50% 的旧培养基喂养细胞,每3-4天再次接种以去除贴壁细胞。仅使用非贴壁 细胞进行实验。
[0160]
rna分离和实时pcr
[0161]
用rneasy mini试剂盒(qiagen)从全细胞裂解产物中提取rna, 并用first strand cdna合成试剂盒(fermentas)逆转录。使用基因特异性 引物,使用sybr green pcr mater mix(life technologies)和appliedbiosystems 7500 real-time pcr instrument(life technologies)进行一式三 份的定量实时pcr。相对表达量用甘油醛-3-磷酸脱氢酶(gapdh)的水 平进行标准化。
[0162]
以下引物用于实时pcr:ifna4正向: 5'-cctgtgtgatgcaggaacc-3',ifna4反向 5'-tcacctcccaggcacaga-3';ifnb前向: 5'-tggagatgacggagaagatg-3',ifnb反向: 5'-ttggatggcaaaggcagt-3';gapdh前向: 5'-atcaagaaggtggtgaagca-3',gapdh反向 5'-agacaacctggtcctcagtgt-3'。相对表达量用甘油醛-3-磷酸脱氢 酶(gadph)的水平进行标准化。
[0163]
细胞因子检测
[0164]
用moi为10的各种病毒感染细胞1小时或模拟感染。取出接种物, 细胞用pbs洗涤
两次,并与新鲜培养基一起培养。感染后不同时期收集上 清液。通过使用ifn-α/β(pbl biomedical laboratories),il-6,ccl4 和ccl5(r&d系统)的酶联免疫吸附实验(elisa)试剂盒测量细胞因 子水平。
[0165]
蛋白质印迹分析
[0166]
来自wt和ko小鼠的bmdc(1
×
106)用moi(感染复数)为 10或相当量的heat-mva或uv-mva感染mva。在感染后的不同时间, 移除培养基并收集细胞。制备全细胞裂解物。将等量的蛋白质进行十二烷 基硫酸钠-聚丙烯酰胺凝胶电泳,并将多肽转移到硝酸纤维素膜上。使 用特异于irf3(cell signaling)的磷酸丝氨酸396的兔多克隆抗体测定 irf3的磷酸化。使用针对irf3的兔多克隆抗体(cell signaling)测定irf3 的水平。抗sting抗体购自cell signaling。通过使用stuart n.easacs博士 (宾夕法尼亚大学)[85]善意提供的抗e3单克隆抗体(mab 3015b2)测 定牛痘e3蛋白水平。抗甘油醛-3-磷酸脱氢酶(gadph)或抗β-肌动蛋 白抗体(cell signaling)作为载体对照。
[0167]
big-f10黑色素瘤细胞用moi 10或mva等量的mva感染。在感 染后的不同时间收集细胞裂解物。使用如上所述的抗磷酸化irf3,抗irf3 和抗gapdh抗体进行蛋白质印迹分析。
[0168]
在有或无系统性给药免疫检查点封锁的情况下,单侧皮内肿瘤植入 和肿瘤内注射病毒
[0169]
将b16-f10黑色素瘤(体积为50μl的1x105细胞)皮内植入 sting
gt/gt
或batf3-/或年龄匹配的wt c57bl/6j小鼠右侧的剃光皮肤。 在植入后10至12天,测量肿瘤大小,直径3mm或更大的肿瘤将在小鼠 麻醉下注射加热-mva(相当于体积为50μl的2
×
107pfu的mva)或pbs。 根据实验具体情况每周或每周两次注射病毒。每天监测小鼠,每周测量两 次肿瘤大小。根据下列公式计算肿瘤体积:l(长)
×
w(宽)
×
h(高)/2。 将小鼠安乐死为遇难迹象,或肿瘤直径达到10毫米时。当小鼠安乐死时 收集血清。
[0170]
为了评估heat-mva与免疫检查点封闭的组合,我们在存在或不存 在抗ctla-4抗体(100μg,100μl体积为100μl)的情况下,用肿瘤内 注射heat-mva或pbs处理小鼠腹腔。小鼠每3-4天接受病毒和抗体(每 周两次)。每天监测动物,每3天测量肿瘤大小。根据下列公式计算肿瘤 体积:l(长)
×
w(宽)
×
h(高)/2。在存在痛苦迹象,或肿瘤直径达 到10毫米时将小鼠安乐死。
[0171]
在一些情况下,1x105mc38结肠腺癌细胞皮内植入剃光小鼠的右 侧。7天后,以与上述两次相同的剂量用pbs,heat-mva或uv-mva 注射肿瘤两周。每天监测动物,每3天测量肿瘤大小。根据下列公式计算 肿瘤体积:l(长)
×
w(宽)
×
h(高)/2。在存在痛苦迹象,或肿瘤直 径达到10毫米时将小鼠安乐死。
[0172]
评估系统性抗肿瘤免疫的发展的肿瘤免疫感染
[0173]
对于b16-f10鼠黑色素瘤模型,通过皮下注射1
×
105个细胞(体积 为50μl)并在肿瘤内递送pbs或heat-mva(相当于热灭活的2
×
107pfu 的mva,体积为50μl)。监测小鼠肿瘤生长和存活30-80天。将存活的 小鼠在对侧进行皮内递送致死剂量的b16-f10(1
×
105个细胞)的再次免 疫感染。监测小鼠肿瘤生长30-80天。或者,通过静脉内递送致死剂量的 b16-f10(1
×
105个细胞)来免疫感染它们,然后在再次免疫感染后3周 安乐死,以评估肺表面肿瘤的存在。
[0174]
对于mc38鼠结肠腺癌模型,通过皮内注射右侧1
×
105个细胞植入 肿瘤,并用肿瘤
内递送pbs,heat-mva或uv-mva(相当于加热或uv 灭活的2x107pfu的mva)。监测小鼠肿瘤生长和存活60天。将存活的 小鼠在对侧进行皮内递送致死剂量的b16-f10(1
×
105个细胞)的再次免 疫感染。监测小鼠肿瘤生长60天。
[0175]
t细胞耗竭实验
[0176]
将b16-f10鼠黑色素瘤细胞(体积为50μl的1
×
105个细胞)皮内植 入6-8周龄的剃光的wt c57b/6小鼠的右侧。在肿瘤植入后8天,肿瘤 每周注射两次heat-mva(等效剂量为2
×
107pfu的mva)或pbs。cd4
+
, cd8
+
细胞和nk细胞的耗竭抗体(200微克的gk1.5,2.43,和pk136) (单克隆抗体核心设施,mskcc)(参考文献,avogadri等人,plos one 2010)腹膜内注射每周两次,在病毒注射之前1天开始,使用它们直到 动物死亡或被安乐死或完全清除肿瘤。每天监测小鼠,并测量肿瘤大小。 目标免疫细胞的耗竭通过4剂量抗体后小鼠外周血的facs进行验证。
[0177]
在有或无全身或肿瘤内施用免疫检查点阻断的情况下,双侧肿瘤植 入模型和瘤内注射病毒
[0178]
简言之,将b16-f10黑色素瘤细胞皮内植入c57b/6小鼠的左侧和 右侧(向右侧5
×
105,左侧为1
×
105)。肿瘤植入后8天,我们将肿瘤内 注射2
×
107pfu的mva或等量的heat-mva给右侧较大的肿瘤。测量肿 瘤大小,肿瘤每周注射两次。监测小鼠的存活。
[0179]
在一些实验中,将mc38结肠腺癌细胞皮内植入c57b/6小鼠的左 侧和右侧(向右侧5
×
105,左侧为1
×
105)。
[0180]
在一些实验中,将sting
gt/gt
,batf3-/-小鼠和wt年龄匹配对照用 于双侧b16-f10黑色素瘤植入,并用pbs或heat-mva处理小鼠右侧较大 的肿瘤。
[0181]
在一些实验中,将具有双侧肿瘤的小鼠用肿瘤内注射heat-mva处 理右侧较大的肿瘤并腹膜内递送免疫检查点阻断抗体,包括抗ctla-4, 抗pd-1或抗-pd-l1。
[0182]
在一些实验中,用肿瘤内注射加热-mva和抗ctla-4抗体(用于 腹膜内递送的十分之一剂量)的双侧肿瘤小鼠治疗右侧较大的肿瘤。测量 注射和非注射肿瘤的大小,监测小鼠的存活。
[0183]
dc成熟的流式细胞术分析
[0184]
对于dc成熟分析,从wt和sting
gt/gt
小鼠产生bmdc,并以 mvi为10或以等量的heat-mva感染mva。在感染后14小时收集细胞, 然后在37℃用fix buffer i(bd biosciences)固定15分钟。洗涤细胞,用 permbuffer(bd biosciences)在冰上透化30分钟,并用针对i型mhc, cd40,cd86和cd80的抗体染色30分钟。使用lsrii流式细胞仪(bdbiosciences)分析细胞。数据用flowjo软件(treestar)进行分析。
[0185]
肿瘤浸润性免疫细胞的流式细胞术分析
[0186]
为了分析肿瘤或肿瘤引流淋巴结中的免疫细胞表型和特征,我们根 据以下方案在facs分析之前产生细胞悬浮液(zamarin等,2014)。首先, 我们使用镊子和手术剪刀在第二次治疗后三天和用pbs,mva或 heat-mva首次治疗后7天分离注射和/或未注射的肿瘤。然后称重肿瘤。 在37℃下与liberase(1.67w w
ü
nsch u/ml)和dnase(0.2mg/ml)温育30分钟之前,切碎肿瘤或肿瘤引流淋巴结。通过重复移液产生细胞悬浮液, 通过70-μm尼龙过滤器过滤,然后在ficoll纯化之前用完全rpmi洗涤 以除去死细胞。处理细胞用抗cd3,cd45,cd4和cd8抗体进行表面标 记。活细胞通过使用可固定染料efluor506(ebioscience)与死细胞
区分开 来。使用foxp3固定和透化试剂盒(ebioscience)进一步透化它们,并对 ki-67,foxp3和颗粒酶b染色。使用lsrii流式细胞仪(bd biosciences) 获得数据。数据用flowjo软件(treestar)进行分析。
[0187]
通过elisa测定抗黑色素瘤和抗病毒抗体
[0188]
为了确定小鼠血清中的抗b16黑色素瘤抗体浓度,将100μl培 养基/孔中的5
×
104个b16-f10细胞加入到96孔培养板中并在37℃下温 育过夜。将板用pbst洗涤两次。用10%缓冲福尔马林(125μl)处理细 胞,并在室温下固定15分钟。然后用pbs将板洗涤三次。用含有1%bsa (250μl)的pbs在室温下封闭1小时后,以100μl/孔加入用1%bsa (1:500)稀释在pbs中的小鼠血清。将板用pbst洗涤5次。在37℃温 育1小时。然后将辣根过氧化物酶(hrp)偶联的用1%bsa(1:2000) 的pbs稀释的抗小鼠igg加入到平板中并在37℃下温育1小时。用pbs 洗涤板5次,并在室温下与底物3,3',5,5'-四甲基联苯胺tmb(100μl/孔) 一起温育10分钟。通过加入硫酸(2n,50μl/孔)终止反应。通过使用设 置为450nm的酶标仪来确定每个孔的光密度。
[0189]
为了确定小鼠血清中的抗痘苗病毒抗体浓度,将100μlpbs/孔中 的heat-mva(10μg/ml)加入到96孔培养板中,并在37℃温育过夜。 将板用pbst洗涤两次。在室温下用1%bsa(250μl)的pbs封闭1小 时后,以100μl/孔加入用1%bsa(1:200)稀释在pbs中的小鼠血清。 检测方法的其余部分与上述相同。
[0190]
试剂
[0191]
试剂的商业来源如下:cpg寡脱氧核苷酸odn2216(invitrogen); 我们使用以下抗体。抗-ctla4(克隆9h10和9d9),抗pd1(克隆rmp1-14) 购自bioxcell;用于流式细胞术的抗体购自ebioscience(cd45.2 alexa fluor700,cd3 pe-cy7,cd4 apc-efluor780,cd8 percp-efluor710,foxp3 alexafluor 700,i型mhc apc,cd40 apc,cd80 apc,cd86 apc),invitrogen (cd4 qdot 605,granzyme b pe-texas red,granzyme b apc),bdpharmingen(ki-67-alexa fluor 488)。
[0192]
统计
[0193]
使用双尾不配对的student's t检验来比较研究中的两个组。通过对 数秩(mantel-cox)检验分析存活数据。被认为是显着的p值如下所示: *,p《0.05;**,p《0.01;***,p《0.001;****,p《0.0001。
[0194]
实施例1
[0195]
热灭活的mva在小鼠cdc中诱导比mva更高水平的i型ifn产 生
[0196]
为了测试热灭活的mva(加热mva,heat-mva)是否会导致比 mva更高水平的i型ifn诱导,mva在55℃下温育1小时,导致感染性 降低1000倍。在gm-csf(gm-csf-bmdc或cdc)存在下培养骨髓衍 生的树突状细胞,并以感染复数(moi)为10的mva或用等量的heat-mva 感染。在感染后6小时收获细胞,并对从感染细胞和模拟感染细胞分离的 rna进行定量实时pcr分析。发现与模拟感染细胞相比,cdc的mva 感染分别增加了ifna4和ifnb mrna水平4.8倍和148倍。相比之下, heat-mva的感染显着地将ifna4和ifnb mrna水平分别提高了22.4倍 和607倍(图1a)。这些结果表明,与mva相比,heat-mva是ifna4 和ifnb基因表达的更强诱导剂(***,p《0.001)。
[0197]
为了评估通过heat-mva或mva感染的cdc诱导i型ifn分泌的 动力学,在heat-mva
或mva感染后的不同时间(0,4,8,16和22小时) 收集上清液,并且通过elisa测定分泌的ifn-α和ifn-β水平。heat-mva 在感染后8小时强烈诱导ifn-α(1650pg/ml)和ifn-β(1975pg/ml), 这是相同时间点的mva诱导的10倍和6倍。而mva诱导的ifn-α和ifn
‑ꢀ
β在感染后8小时和22小时之间持续上升,在该时间段期间,heat-mva 诱导的ifn-α水平适度增加,而heat-mva诱导ifn-β在8小时达到峰 值后感染并随后平缓(图1b)。蛋白质印迹分析显示,减弱先天免疫反应 的e3蛋白在heat-mva感染的cdc中不产生,但在mva感染的细胞中 表达(图1c)。此外,heat-mva在感染后4和8h时触发了比mva更高 水平的irf3磷酸化(图1c)。总之,这些结果表明,在mvc中,heat-mva 是cdc中i型ifn产生的更强诱导剂。
[0198]
实施例2
[0199]
加热mva诱导的i型ifn产生依赖于由cgas/sting介导的细 胞溶质dna感测途径,以及转录因子irf3/irf7和ifnar1
[0200]
为了测试cdcs的heat-mva感染是否通过细胞溶质dna传感器 cgas(循环gmp-amp合酶)介导的细胞溶质dna感应途径触发i型ifn 诱导[62,63],及其适配器sting[59,69],cccs从cgas-/-小鼠和年龄匹 配的wt对照组产生,并用heat-mva感染。使用定量实时pcr分析, 发现在cgas缺陷细胞中,感染后6小时的heat-mva诱导的ifna4和 ifnb基因表达都减少(图2a,2b)。在感染后22小时收集的上清液的分 析也显示在cgas缺陷细胞中消除了heat-mva诱导的ifn-α/β分泌(图 2a,2b)。
[0201]
sting是细胞溶质dna感测途径的关键适配器[59,69,87,88]。cdc 也由sting
gt/gt
小鼠产生,其缺乏功能性sting[89]。发现heat-mva诱 导i型ifn基因表达,并且来自cdc的ifn-α/β分泌也依赖于sting(图 2c,d)。western印迹分析显示,在感染后4和8小时,heat-mva在ser-396 处诱导irf3的磷酸化,其在cgas或sting缺陷型细胞中被消除(图2e,f)。为了测试heat-mva感染是否通过细胞溶质dna感测途径触发dc 成熟,用heat-mva感染来自sting
gt/gt
小鼠的cdc和年龄匹配的wt对 照。在感染后14小时收集细胞,并染色dc激活标记,包括i型mhc(mhci)cd40,cd86和cd80。heat-mva感染显著诱导cd40和cd86的表 达,轻度增加wt细胞中mhc i和cd80的表达(图2g)。然而,sting 缺陷型细胞中激活标记物的表达显着降低(图2g)。这些结果表明,dc 成熟很大程度上是通过细胞溶质dna感测途径介导的。我们的研究结果 表明,来自mva和heat-mva的病毒dna被释放到受感染的cdc的胞 质溶胶中,并且被细胞溶质dna传感器cgas检测到,其又产生第二信 使cgamp,导致sting和下游信号通路的激活。
[0202]
为了测试i型ifn的heat-mva诱导是否需要irf3,irf5和irf7, 除了cgas和sting外,还从irf3-/-,irf5-/-,,irf7-/-和年龄匹配的 wt小鼠中产生cdc,并用heat-mva感染。heat-mva诱导的ifna4 基因表达和ifn-α蛋白的产生依赖于irf3和irf7,但与irf5无关(图 3a,b)。此外,heat-mva诱导的ifnb基因表达以及,与ifn-α分泌类 似,ifn-β蛋白分泌依赖于irf3和irf7,而不依赖于irf5(图3a,b)。 在irf7缺陷细胞中,heat-mva诱导的ifnb基因表达和ifn-β产生分别 降低了74%和67%(图3a,b)。heat-mva诱导的ifna4基因表达和ifn
‑ꢀ
α蛋白分泌依赖于ifnar1,而ifnar1缺乏型细胞中,heat-mva诱导 的ifnb基因表达和ifn-β分泌分别降低了82%和62%图3c,d)。这些 结果表明heat-mva诱导的i型ifn诱导需要转录因子irf3和irf7,以 及由ifnar1介导的i型ifn正反馈环。
[0203]
实施例3
[0204]
在体内,加热mva诱导比mva更高水平的i型ifn
[0205]
为了测试heat-mva在体内是否诱导比mva高水平的i型ifn, 通过尾静脉注射将heat-mva或mva接种到c57b/6小鼠中,并在感染 后6小时收集血清。在热mva治疗的小鼠中,血清中ifn-α和ifn-β的 水平显着高于在mva治疗的小鼠中(图4a)(***,p《0.001)。这些结果 表明,heat-mva不仅在培养的cdc中诱导比mva更高水平的i型ifn, 而且还在体内诱导比mva更高水平的i型ifn。
[0206]
实施例4
[0207]
加热mva以sting/irf3/irf7依赖的方式在体内触发i型ifn 产生
[0208]
为了测试heat-mva对i型ifn的体内诱导是否需要ifnar1,通 过尾静脉注射对ifnar1-/-和wt年龄匹配的对照小鼠进行静脉内(iv) 接种纯化的heat-mva。wt小鼠的heat-mva感染诱导ifn-α和ifn
‑ꢀ
β的产生为2256pg/ml和1901pg/ml的水平,其在ifnar1-/-小鼠中分 别降低了60%和35%(图4b)(**,p《0.01;***,p《0.001)。
[0209]
与wt对照相比,satgt/gt小鼠的heat-mva诱导的ifn-α分 泌降低了89%,而sting
gt/gt
小鼠中热mva诱导的ifn-β分泌被消除(图 4c),表明体内heat-mva诱导的i型ifn产生也依赖于sting。此外, 与wt对照相比,irm3-/-小鼠中的heat-mva诱导的ifn-α降低了74%, 而在irf3-/-小鼠中,heat-mva诱导的ifn-β降低了98%。在irf7-/-小 鼠中,heat-mva诱导的ifn-α和ifn-β分泌物减少(图4c)。这些结果 表明,heat-mva诱导的i型ifn在体内需要sting和irf3/irf7。
[0210]
实施例5
[0211]
b16-f10黑色素瘤细胞的加热mva感染诱导产生i型ifn和促炎 细胞因子和趋化因子
[0212]
为了检测肿瘤细胞的heat-mva感染是否触发先天免疫反应, bio-f10黑色素瘤细胞以moi为10的mva感染,或用等量的heat-mva 感染,并在感染后6小时收集细胞和在感染后22小时收集上清液。定量 实时pcr分析显示,b16-f10细胞的heat-mva感染诱导比mva更高水 平的ifna4,ifnb,ccl5和il6基因表达(图5a)。elisa分析显示,与 mva相比,heat-mva诱导b16-f10细胞中ifn-α,ifn-β,ccl5和il-6 蛋白分泌水平升高(图5b)。western印迹分析表明,heat-mva感染诱 导b16-f10黑色素瘤细胞中irf3的磷酸化水平高于mva(图5c)。此外, heat-mva感染诱导b16细胞上i型mhc分子表达的表达,而mva感染 则不能这样。这些结果表明,b16细胞的heat-mva感染不仅通过释放i 型ifn和促炎细胞因子和趋化因子而诱导针对肿瘤细胞的先天免疫应答, 而且还通过诱导肿瘤细胞上i型mhc分子来改变(增强)肿瘤的免疫原 性。
[0213]
实施例6
[0214]
55℃持续1小时是灭活mva的最佳条件
[0215]
为了评估55℃是否是灭活mva的最佳温度,我们在不同温度(包 括45℃,50℃,55℃,60℃和65℃)下温育mva1小时。用这些病毒制 剂感染来自wt小鼠的cdc,并在感染后22小时收集上清液。通过elisa 测量分泌的ifn-α和ifn-β的浓度。我们发现mva在55℃下灭活1小 时的感染诱导了来自cdc的ifn-α和ifn-β分泌的最高水平(图6a,b)。
[0216]
实施例7
[0217]
加热mva的肿瘤内注射导致大鼠移植b16-f10黑色素瘤模型中的 肿瘤消除和全身
抗肿瘤免疫
[0218]
可移植的体内b16-f10黑色素瘤模型涉及在c57b/6小鼠的一侧体 侧的皮内植入鼠b16-f10黑色素瘤细胞(1
×
105)。肿瘤植入后十天,当 肿瘤直径约3毫米时,每周向肿瘤注射heat-mva(相当于2
×
107pfu的 mva)或pbs。肿瘤内注射heat-mva导致肿瘤根除和小鼠的100%存活 (图7a,b),表现出优异的治疗功效。相比之下,肿瘤内注射pbs的所 有小鼠继续肿瘤生长,并在肿瘤植入后19和21天安乐死(图7a,b)。
[0219]
为了检测肿瘤内注射heat-mva后根除肿瘤的小鼠是否发生全身性 抗肿瘤免疫,初始肿瘤根除后8周,通过皮下注射致死剂量的b16黑色素 瘤细胞(1
×
105)给对侧来对动物进行攻毒。使用未暴露于b16黑色素瘤 细胞或heat-mva的空白小鼠(mice)作为对照。肿瘤攻毒后70天 内观察动物。90%的heat-mva
–
处理的小鼠在肿瘤攻击后存活,而所有 的空白小鼠发育成长的肿瘤并最终被安乐死(图7c)。为了测试heat-mva 处理的小鼠是否在与转移类似的不同器官产生全身抗肿瘤免疫,我们测试 了heat-mva处理的小鼠是否可以抵抗通过静脉内递送b16-f10黑色素瘤 细胞的肿瘤攻击。两只空白小鼠和heat-mva处理的小鼠通过静脉内输送 接受1
×
105个b16-f10细胞。肿瘤攻毒后3周将小鼠安乐死。收集小鼠的 肺并在含福尔马林的溶液中固定。将肺表面的肿瘤在解剖显微镜下观察并 计数。我们发现,所有的空白小鼠在肺表面平均发现了58个肿瘤,但在 10只heat-mva治疗的小鼠中只有1只在显微镜下可见2个肿瘤灶(图 7d,****,p《0.0001)。总之,这些结果表明,肿瘤内注射heat-mva导 致根除注射的肿瘤和发展全身抗肿瘤免疫。这些结果意味着肿瘤内注射 heat-mva可以通过增强肿瘤抗原呈递和肿瘤特异性t细胞的活化而引起 强烈的肿瘤疫苗效应。
[0220]
实施例8
[0221]
加热mva导致肿瘤微环境的免疫学变化
[0222]
为了研究肿瘤内注射heat-mva诱导的肿瘤内的免疫学变化,肿瘤 内注射heat-mva或pbs后3天收获肿瘤,并通过facs分析免疫细胞浸 润。肿瘤内活细胞的cd3
+
cd45
+
t细胞百分比从pbs处理肿瘤的6.5%增 加到heat-mva处理的肿瘤中的19.5%(p=0.0002;图8a,g)。在肿瘤内 也观察到表达颗粒酶b(即表达细胞毒性表型)的cd8
+
t细胞百分比的增 加,pbs-治疗肿瘤中的47.9%,在heat-mva处理的肿瘤中为92.8%(p 《0.0001;图8b,h)。ki-67
+
cd8
+
t细胞(即增殖性cd8
+
t细胞)的百分 比从51.2%增加到71.7%(p=0.0008;图8c,i)。与用pbs处理的那些相 比,用heat-mva处理的肿瘤中cd4
+
t细胞观察到相似的变化;颗粒酶 b
+
cd4
+
t细胞(即活化的t辅助细胞)的百分比从pbs处理的肿瘤中的3% 显着上升到heat-mva处理的肿瘤中的57%(p=0.0002;图8d,j)。另外, 在pbs处理的肿瘤中,ki-67
+
cd4
+
t细胞(即增殖的cd4
+
t细胞)的百 分比从37.5%增加到heat-mva处理的肿瘤中的79%(p《0.0001;图8e和 k)。相比之下,foxp3
+
cd4
+
t细胞(即调节性cd4
+
t细胞)的百分比从 pbs处理的肿瘤的34.7%下降到heat-mva处理的肿瘤中的9.1%(p 《0.0001;图8f,n)。这些结果表明,肿瘤内注射heat-mva显着上调肿瘤 微环境中的免疫应答,包括辅助cd4
+
,细胞毒性cd4
+
(统称为“效应t 细胞”)和细胞毒性cd8
+
t细胞的增殖和激活,以及肿瘤内cd4
+
调节t 细胞的伴随减少。结合实施例8的结果,这些结果表明,heat-mva的肿 瘤内注射深刻地改变肿瘤免疫抑制微环境,促进抗肿瘤免疫的发展。
[0223]
实施例9
[0224]
–
加热mva还诱导肿瘤引流淋巴结(tdln)的免疫学变化
[0225]
为了测试是否肿瘤内注射heat-mva导致tdlns的免疫学变化, 从heat-mva处理或pbs处理的小鼠中分离tdln,并通过facs进行分 析。在用heat-mva处理的小鼠中,tdln中粒细胞b
+
cd8
+
t细胞的百分 比从用pbs处理的小鼠的0.15%增加到3.04%(p《0.0001;图9a和e)。 另外,用heat-mva处理的小鼠,ki-67
+
cd8
+
t细胞的百分比从用pbs处 理的小鼠的7.2%增加到17%(p=0.0003;图9c,f)。这些结果表明,在 heat-mva处理小鼠的tdlns中比在pbs处理的小鼠中存在更多的活化 和复制的cd8
+
t细胞。与pbs处理的小鼠相比,在heat-mva处理的小 鼠中也观察到活化和复制的cd4
+
t细胞的类似增加。tdln中颗粒酶 b
+
cd4
+
t细胞的百分比从pbs处理的小鼠的0.25%增加到heat-mva处 理的小鼠中的0.77%(p=0.002;图9b,g),并且ki-67
+
cd4
+
tdln中的 t细胞从pbs处理的小鼠中的10.6%增加到heat-mva处理的小鼠中的 18.4%(p=0.002;图9d,h)。总之,这些结果表明,肿瘤内注射heat-mva 导致不仅在肿瘤内,而且在循环中的cd8
+
和cd4
+
t细胞的活化和增殖。
[0226]
实施例10
[0227]
与野生型对照相比,
–
加热mva的肿瘤内注射在sting缺陷小鼠 或batf3缺陷小鼠中根除b16黑色素瘤的效果较差
[0228]
最近的研究表明,sting介导的细胞溶质dna感测途径在自发性 t细胞对肿瘤的反应以及放射诱导的抗肿瘤免疫中起作用[7,8,90]。 batf3是一种转录因子,对于cd8α
+
系dcs的发育至关重要,其在病毒 和肿瘤抗原的交叉表达中起重要作用[91,92]。batf3缺陷小鼠不能排斥高 免疫原性肿瘤[91]。为了测试sting或batf3是否在heat-mva介导的肿 瘤清除中发挥作用,我们将b16-f10黑色素瘤细胞皮内植入wt c57b/6, sting
gt/gt
或batf3-/-小鼠的右侧。肿瘤植入后11天,按照指示每周一次注 射heat-mva(等效剂量为2
×
107pfu)或pbs(图10a和b)。我们发现, 100%的用heat-mva处理的wt小鼠存在很少肿瘤,如果有任何残留的 肿瘤,而用pbs处理的所有wt小鼠死亡(中位数存活24天)(图10a 和b)。我们还观察到,尽管只有7.7%的用heat-mva处理的sting缺陷 小鼠存活,但用pbs处理的所有sting缺陷小鼠都死亡。在heat-mva 处理后,wt和sting
gt/gt
小鼠的存活率差异具有统计学意义(p《0.0001) (图10a和b)。sting
gt/gt
小鼠的heat-mva处理将中位生存期从21天 延长至28天(p《0.0001)(图10a和b)。更令人惊讶的是,所有的batf3 ‑
/-小鼠都死亡,无论他们是用heat-mva还是pbs处理。然而,batf3-/-小 鼠中的heat-mva处理将中位生存期从21天延长至30天(p=0.0025)(图 10a和b)。这些结果表明,satt介导的细胞溶质dna感测途径和 cd8
+
dcs对于heat-mva诱导的抗肿瘤作用是必需的。
[0229]
实施例11
[0230]
–
加热mva诱导的抗肿瘤作用需要cd8
+
t细胞
[0231]
为了确定heat-mva的治疗效果需要哪种免疫细胞类型,我们进行 了抗体耗竭实验。简言之,我们将b16-f10黑色素瘤细胞(2
×
105)皮内 植入wt c57b/6小鼠的右侧。在肿瘤植入后8天,肿瘤按照指示每周一 次注射heat-mva(等效剂量为2
×
107pfu)或pbs(图10a)。在病毒注 射前一天开始腹膜内注射两次cd4
+
,cd8
+
和nk细胞的耗竭抗体(200 μg gk1.5,2.43和pk136)(图11a)。我们发现,虽然heat-mva的肿瘤 内递送导致有效的肿瘤根除,但cd8
+
t细胞的耗竭导致heat-mva的治疗 功效急剧下降(****,p《0.0001)(图11b,c,d,e)。cd4
+
和nk/nkt 细胞的耗竭导致heat-mva的治疗功效部分丧失(图11f,g)。这些结果 表
明cd8
+
t细胞是由heat-mva引起的抗肿瘤作用所必需的,而cd4
+
和 nk/nkt细胞也有助于抗肿瘤作用。cd4
+
t细胞在抗肿瘤作用中的作用 进一步证明了在cd4-耗竭性抗体存在下,用heat-mva成功处理的小鼠 缺乏对肿瘤攻击的保护作用(图11h-i)。相比之下,在存在或不存在 nk/nkt耗竭性抗体的情况下用heat-mva成功处理的小鼠有效地排除肿 瘤攻击(图11h-i)。我们得出结论,虽然cd4
+
t细胞不是根除加热mva 注射的肿瘤绝对需要的,但它们对抗肿瘤适应性免疫的发展至关重要,可 能是用于产生抗肿瘤抗体。与实施例8,9,10和11一起,我们推测 heat-mva的肿瘤内递送导致在免疫细胞和肿瘤细胞中诱导i型ifn,其 导致cd103
+
树突状细胞的活化,导致肿瘤内和循环中肿瘤抗原交叉呈递 给cd8
+
t细胞,以及产生适应性抗肿瘤免疫。
[0232]
实施例12
[0233]
sting介导的细胞溶质dna感测途径和cd103
+
dcs都是通过加 热mva诱导抗黑色素瘤抗体所必需的
[0234]
抗肿瘤抗体生产是适应性免疫的重要方面。为了测试heat-mva是 否诱导抗黑色素瘤抗体产生,我们进行了elisa,以确定用heat-mva或 模拟处理的小鼠中抗b16黑色素瘤抗体的血清浓度。我们发现只有 heat-mva治疗的小鼠产生抗黑色素瘤抗体(图12a)。在sting或batf3 缺陷小鼠中,该诱导被消除(图12a)。相比之下,抗病毒抗体的产生不 依赖于sting或batf3(图12b)。这些结果表明在该动物模型中促进肿 瘤和病毒抗原识别的过程可能是不同的。从实施例11,我们知道cd8
+
t 细胞对于在注射的肿瘤中的肿瘤杀伤是关键的,因此对于释放肿瘤抗原是 重要的,其可由b细胞加工以在辅助性cd4
+
t细胞存在下产生抗原特异 性抗体。因此,我们假设在缺乏batf3的小鼠中,抗肿瘤cd4
+
和cd8
+
t 细胞都缺乏,这导致抗黑色素瘤抗体的产生失败。
[0235]
实施例13
[0236]
肿瘤内注射加热mva与腹膜内递送抗ctla-4抗体的组合导致单 侧黑色素瘤植入模型中的协同抗肿瘤作用
[0237]
为研究肿瘤内注射heat-mva是否具有增强免疫治疗,如阻断免疫 检查点(例如抗ctla-4抗体),效果的能力,对患肿瘤小鼠进行肿瘤内 注射heat-mva与腹腔内递送抗ctla-4抗体相结合的处理。简言之,我 们将b16-f10黑色素瘤细胞(2
×
105)皮内植入wt c57b/6小鼠的右侧。 肿瘤植入后10天,当肿瘤生长大于实施例7,10或11的时,用以下组合 处理小鼠:pbs+同种型对照,pbs+抗-cctla-4抗体,heat-mva+同种 型对照,和heat-mva+抗ctla-4。如图11e所示,病毒注射开始时,测 试组的肿瘤体积一致。用pbs+同种型对照或用pbs+抗ctla-4处理的小 鼠由于肿瘤生长而迅速死亡(图13a,b)。然而,在heat-mva处理之后, 接受heat-mva注射的肿瘤显着减少或根除,在实验结束时(病毒注射后 第73天),30%的小鼠没有肿瘤(图13c)。用heat-mva和抗ctla-4 抗体处理,与单独的heat-mva处理相比,具有优异的治疗功效,实验结 束时78%的小鼠没有肿瘤(图13d)。我们观察到肿瘤内注射heat-mva 和腹膜内递送抗ctla-4抗体的协同效应,导致治愈率和存活率显着增加 (图13f,*,p《0.05;**,p《0.01;****,p《0.0001)。这些结果表明, heat-mva的肿瘤内递送导致肿瘤免疫抑制微环境的改变,产生抗肿瘤 cd8
+
和cd4
+
t细胞应答,其在抗ctla-4抗体存在下被增强或释放。
[0238]
实施例14
[0239]
加热mva是比mva更强的抗肿瘤免疫诱导剂
[0240]
mva是在大多数哺乳动物细胞中非复制的减毒痘苗病毒。我们发 现mva在b16黑色素瘤细胞中小幅复制(图12a)。heat-mva将感染 性降低了1000倍,并且不在b16黑色素瘤细胞中复制(数据未显示)。我 们假设heat-mva可能是比mva更强的抗肿瘤免疫活化剂,因为 heat-mva在感染的cdcs和肿瘤细胞在体外(实施例1和5)以及体内(实 施例4)诱导比mva高的i型ifn水平。我们进行了以下实验,以直接 比较双侧b16-f10黑色素瘤植入模型中肿瘤内注射heat-mva与mva之 间的肿瘤根除和产生全身免疫力的效力。简言之,将b16-f10黑色素瘤细 胞皮内植入c57b/6小鼠的左侧和右侧(向右侧5
×
105,左侧为1
×
105)。 肿瘤植入后8天,我们将肿瘤内注射2
×
107pfu的mva或等量的 heat-mva给右侧较大的肿瘤。测量肿瘤大小,每周注射两次肿瘤。也监 测小鼠的存活。我们发现在用pbs处理的小鼠中,肿瘤在右侧面迅速生长, 导致早期死亡(图12c,d和b)。heat-mva或mva的肿瘤内注射导致 与pbs相比延迟肿瘤生长和改善生存(图14b,***,mva对pbs,p《0.001, 对于热mva,p《0.0001与pbs)。对根除肿瘤,肿瘤内注射heat-mva比 mva更有效(heat-mva处理,9/9不含肿瘤;mva处理,6/9不含肿瘤), 并延迟或抑制对侧未注射肿瘤的生长(对于heat-mva处理而言为5/9不 含肿瘤,mva处理为1/9不含肿瘤)(图14e-h)。与mva处理的小鼠相 比,我们观察到heat-mva处理的小鼠的存活率提高(图14b,**,p《0.01)。 这些结果表明(i)病毒复制对于实现抗肿瘤作用是不必要的;和(ii) heat-mva的抗肿瘤作用与其强诱导i型ifn的能力相关。
[0241]
实施例15
[0242]
加热mva在非注射肿瘤中诱导比mva更多的免疫激活细胞
[0243]
为了了解heat-mva优于mva在诱导全身抗肿瘤免疫中的免疫机 制,我们研究了在heat-mva或mva处理的小鼠中未注射的肿瘤中的免 疫细胞浸润。简言之,我们将2.5
×
105个b16-f10黑色素瘤细胞皮内植入 左侧腹腔,将5
×
105个b16-f10黑色素瘤细胞植入小鼠的右侧。植入后7 天,我们将2
×
107pfu的mva或等量的heat-mva或pbs注射到右侧较 大的肿瘤中。三天后重复注射。收获未注射的肿瘤,产生细胞悬液。通过 facs分析肿瘤中浸润的活细胞免疫细胞。我们观察到与用mva或pbs 处理的小鼠相比,用heat-mva处理的小鼠的未注射的肿瘤中cd45
+
, cd103
+
,cd3
+
和cd8
+
免疫细胞显着增加。尽管与pbs治疗的小鼠相比, mva处理也导致非注射肿瘤中这些免疫细胞的增加,但mva处理在非注 射肿瘤中诱导免疫细胞方面比heat-mva比较不有效(图15a)。heat
‑–ꢀ
mva处理导致在非注射肿瘤中表达cd8
+
和cd4
+
t细胞的细胞毒性颗粒 酶b的召集和增殖(图15b)。mva在非注射肿瘤中诱导粒酶b
+
cd8
+
效果比heat-mva较差(图15b)。这些结果表明,在非注射的肿瘤中, heat-mva比mva更能够召集和激活多种免疫细胞,特别是颗粒酶 b
+
cd8
+
t细胞。这与其与mva相比,在消除或延迟非注射肿瘤的生长和 延长生存的增强的功效相关。
[0244]
实施例16
[0245]
加热mva的肿瘤内递送在sting或batf3缺陷小鼠的双侧肿瘤植 入模型中未能治愈b16-f10黑色素瘤
[0246]
在实施例10中,我们显示,在单侧植入模型中,热-mva的肿瘤 内递送对消除b16-f10黑色素瘤无效。为了进一步扩展这项研究,我们测 试了双侧肿瘤植入模型中heat-mva的肿瘤内递送的功效。在pbs治疗组 中,由于右侧较大肿瘤的快速生长,所有小鼠死亡,中位生存期为16天 (图16a,b,i)。heat-mva的肿瘤内注射导致根除所有注射的肿瘤,但 仅在
10只wt小鼠中的3只清除非注射肿瘤(图16c,d,i,****,p《0.0001 为heat-mva vs.pbs)。我们发现尽管heat-mva处理导致wt小鼠中黑 色素瘤治愈率达到30%,但在batf3 ko小鼠(图16g,h,i)中未见治 疗效果。在sting缺陷小鼠中,肿瘤内注射heat-mva导致肿瘤生长的 延迟和中位生存期的延长(图16e,f,i,**,p《0.01)。与实施例10一 起,我们得出结论,batf3依赖的cd103
+
dcs对于通过heat-mva的肿瘤 内递送诱导抗肿瘤免疫是关键的。由sting介导的细胞溶质dna感测途 径也在热mva诱导的适应性抗肿瘤免疫中起着重要作用。
[0247]
实施例17
[0248]
batf3 ko小鼠不利于发展抗肿瘤cd8
+
和cd4
+
t细胞,以响应加热 mva的肿瘤内递送
[0249]
鉴于cd103
+
dcs在实施例10和16中所示的heat-mva诱导的抗 肿瘤免疫中的重要性,以及cd8
+
和cd4
+
t细胞在heat-mva介导的抗肿 瘤作用中的关键作用,我们使用双侧肿瘤植入模型研究了在batf3 ko小 鼠响应肿瘤内注射heat-mva产生抗肿瘤cd4
+
和cd8
+
t细胞中是否存在 缺陷。简言之,我们将2.5
×
105个b16-f10黑色素瘤细胞皮内植入batf3-/-小鼠和wt年龄匹配对照组的小鼠的左侧腹腔和5
×
105个b16-f10黑色素 瘤细胞植入右侧腹腔。植入后7天,我们将右侧腹部的较大肿瘤中注射了 heat-mva或pbs。三天后重复注射。在第一次注射后第7天收获未注射 的肿瘤,并产生细胞悬液。通过facs分析注射和未注射的肿瘤中的活细 胞免疫细胞。与实施例15类似,我们观察到与用pbs处理的小鼠相比, 用heat-mva处理的小鼠的注射和未注射的肿瘤中的cd3
+
和cd8
+
免疫细 胞显着增加(图17ab,**,p《0.01;***,p《0.001)。我们还观察到注射 和非注射肿瘤中ki-67
+
cd8
+
和ki-67
+
cd4
+
t细胞显着增加(图17cd,**, p《0.01;***,p《0.001;****,p《0.0001)。相比之下,在batf3 ko小鼠 中,注射和未注射肿瘤的cd8
+
和cd4
+
t细胞的召集和增殖减少(图 17a-d)。这些结果表明cd103
+
dcs在交叉呈递肿瘤抗原和响应heat-mva 处理产生抗肿瘤cd8
+
t细胞是至关重要的。cd103
+
dc以外的许多细胞 类型能够在mhc ii型上呈递肿瘤抗原给初始cd4
+
t细胞。在这里,我们 发现batf3-/-小鼠中非注射肿瘤中肿瘤反应性cd4
+
t细胞的数量远低于 wt小鼠(图17d)。在batf3-/-小鼠的肿瘤中缺乏cd8
+
t细胞有可能导 致肿瘤杀伤不良和肿瘤抗原释放不良,影响肿瘤反应性cd4
+
t细胞的产 生。与实施例10,12和16一起,我们得出结论,batf3依赖的 cd103
+
/cd8-mva诱导的抗肿瘤作用中起重要作用,包括产 生肿瘤反应性cd8
+
,cd4
+
t细胞,肿瘤抗体。
[0250]
实施例18
[0251]
肿瘤内注射加热mva与腹膜内递送免疫检查点阻断的结合导致双 侧黑色素瘤植入模型中的协同治疗效果
[0252]
然后,我们在双侧b16-f10黑色素瘤模型中调查了heat-mva的肿 瘤内注射是否增强了免疫检查点阻断治疗如抗ctla-4,抗pd-1或抗 pd-l1抗体的治疗效果,该模型模仿具有转移性疾病的个体。简言之,将 b16-f10黑色素瘤细胞皮内植入c57b/6小鼠的左侧和右侧(向右侧5
×ꢀ
105,左侧为1
×
105)。肿瘤植入后8天,我们每周两次腹膜内注射heat-mva (热灭活的2
×
107pfu的mva)或pbs到较大的右侧肿瘤。使用heat-mva 处理四组小鼠,每组还分别接受同种型对照或抗ctla-4或抗pd-1或抗
ꢀ‑
pd-l1抗体的腹膜内递送(图18a)。
[0253]
pbs处理的小鼠在接下来的20天(图18b,c,d)中随着肿瘤生 长的增加而迅速死亡,用heat-mva+同种型对照治疗的小鼠根除了注射的 肿瘤并延迟了对侧的非注射肿瘤的生长(图18e,f)。结果,与pbs组相 比,heat-mva+同种型的治疗显着延长了存活率(图
18b,****,p 《0.0001)。肿瘤内注射heat-mva和抗ctla-4,抗pd-1和抗pd-l1抗体 的全身递送的组合进一步延迟或消除非注射肿瘤。结果,50%的用 heat-mva+抗ctla-4处理的小鼠,50%的用heat-mva+抗pd-1的小鼠 和70%用heat-mva+抗-pd-l1的小鼠在实验结束时(病毒注射后第57 天)无肿瘤,与此相比,10%的用heat-mva+同种型治疗的小鼠无肿瘤(图 18e-l)。
[0254]
与heat-mva+同种型对照相比,在heat-mva+免疫检查点阻断结 合组中,控制非注射远处肿瘤生长的能力与的改善生存率相关(图18b, **,p《0.0001)。b16-f10黑色素瘤模型中单独腹膜内递送抗ctla-4, 抗pd-1或抗pd-l1具有最小的治疗效果(图13b和数据未显示)。这些 结果表明,heat-mva的肿瘤内递送克服了对转移性b16黑色素瘤模型中 的免疫检查点阻断的治疗的抵抗,这意味着将该方法转移到人类治疗的有 利的结果。
[0255]
将重复该实验以评估联合免疫检查点阻断和灭活的mva治疗的长 期益处。
[0256]
实施例19
[0257]
uv-mva以sting依赖的方式在cdc中诱导i型干扰素
[0258]
我们假设,mva的紫外线灭活也可能通过激活sting介导的细 胞溶质dna感测途径导致免疫激活病毒。为了测试这个假设,感染了来 自sting
gt/gt
及其年龄匹配的wt对照小鼠的cdc。细胞(1
×
106)用 moi为10的mva或相当量的heat-mva或uv-mva感染。感染后22h 收集上清液,elisa法检测分泌ifn-α和ifn-β的浓度。与heat-mva 类似,uv灭活的mva在wt cdc中也诱导比mva更高水平的i型ifn (图19a,b)。uv-mva诱导的i型ifn在sting缺陷型细胞中完全消 除(图19a,b,***,p《0.001)。这些结果表明,heat-mva和uv-mva 介导的ifn-α和ifn-β的诱导都取决于sting途径,进一步证实 heat-mva和uv-mva通过类似的机制发挥其肿瘤抑制作用。
[0259]
实施例20
[0260]
在单侧结肠腺癌模型中,肿瘤内注射uv-mva和加热mva导致根 除注射的肿瘤和发展全身抗肿瘤免疫
[0261]
实施例7和14中公开的实验研究表明,肿瘤内注射heat-mva导 致鼠移植b16-f10黑色素瘤模型中的肿瘤消除和全身抗肿瘤免疫。为了测 试heat-mva或uv-mva是否能够消除其他实体瘤,我们测试了一种鼠 mc38结肠腺癌植入模型中heat-mva或uv-mva的抗肿瘤作用。结肠腺 癌是与黑色素瘤无关的实体肿瘤的代表,但不是任意选择。将5
×
105个 mc38结肠癌细胞皮内植入c57b/6小鼠的右侧。允许肿瘤生长7天,之 后,每周两次对肿瘤内注射heat-mva或uv-mva(通过加热或uv灭活 2
×
107pfu的mva)或pbs对照。每周测量两次肿瘤,根据下列公式计 算肿瘤体积:l(长度)
×
w(宽)
×
h(高)/2。我们发现用pbs处理的 所有小鼠都因肿瘤生长而死亡(图20d,g)。在实验结束时(病毒注射后 约60天),70%的heat-mva处理的小鼠和71%的uv-mva处理的小鼠 存活(图20e,f)。因此,与pbs对照相比,肿瘤内注射heat-或uv-mva 显着延长了小鼠的存活率(图20g,****,p《0.0001)。
[0262]
为了测试存活的小鼠是否已发展出全身抗肿瘤免疫,我们用致死剂 量的mc38细胞(1
×
105)攻击小鼠,并监测存活。我们发现,虽然所有 的初始小鼠发展成肿瘤并死亡,但100%的heat-或uv-mva治疗的小鼠 抵抗肿瘤攻击(图20h)。我们还测试了mc38细胞的感染,与heat-mva 或uv-mva一起是否比mva诱导更高水平的炎性细胞因子和趋化因子, 我们用moa为10的mva感染mc38细胞,或用等量的heat-mva或 uv-mva感染mc38细胞。感染后22小
时收集上清液。通过elisa测定 上清液中分泌的il-6,ccl4和ccl5的浓度。我们还发现heat-mva和 uv-mva比mva诱导mc38细胞更高水平的il-6,ccl4和ccl5(图20a, b,c)。总体上,在实施例7,14和实施例20中观察到的结果表明,heat-mva 和uv-mva在促进各种实体瘤中的抗肿瘤作用方面是有效的,并且本公 开中描述的发现不限于黑色素瘤,被推广到其他不同来源的实体肿瘤。
[0263]
实施例21
[0264]
双侧mc38结肠腺癌植入模型中,肿瘤内注射加热mva与腹膜内 递送免疫检查点阻断的组合导致协同治疗效果
[0265]
我们在其它双侧肿瘤植入模型中进一步调查了肿瘤内注射 heat-mva是否增强免疫检查点阻断疗法的治疗效果,如抗ctla-4,抗 或抗pd-l1抗体,该模型模拟具有转移性疾病的个体。简而言之,将mc38 结肠腺癌细胞皮内植入c57b/6小鼠的左侧和右侧(右侧5
×
105,左侧为 1
×
105)。肿瘤植入后8天,我们每周两次注射heat-mva(热灭活的2
×ꢀ
107pfu的mva)或pbs到右侧的较大肿瘤。有用pbs处理的三组小鼠, 每组接受pbs或抗ctla-4或抗-pd-l1抗体的腹膜内递送(图21a-f)。 有三组用heat-mva治疗的小鼠,每组接受同种型对照或抗ctla-4或抗
ꢀ‑
pd-l1抗体的腹膜内递送(图21g-l)。pbs处理的小鼠在随后的14天 内随着肿瘤生长的增加而迅速死亡(图21b,c,d),用pbs+抗ctla-4 或pbs+抗pd-l1处理的所有小鼠也全部死亡,尽管腹膜内递送抗ctla-4 或抗-pd-l1抗体导致与pbs处理的小鼠比较有较长的生存期(图21a-f,m, ***,p《0.001)。类似于我们在b16-f10双侧植入模型中观察到的(参见实 施例14),肿瘤内注射heat-mva导致根除注射的mc38肿瘤(图21g, 8/10无肿瘤),延迟了对侧非注射肿瘤的生长,然而,但仅根除其中1/10 (图21h,****,p《0.0001 heat-mva vs.pbs)。相比之下,heat-mva的 肿瘤内递送与抗ctla-4抗体或heat-mva+抗pd-l1的腹膜内递送的组 合导致比heat-mva更高的效率根除非注射的远端肿瘤(图21g-l),与 单独使用heat-mva相比,联合治疗与改善的存活相关(图21n,*,p《0.05, **,p《0.01)。这些结果对于使用灭活的mva和免疫检查点阻断的组合来 治疗转移性实体瘤具有意义。
[0266]
实施例22
[0267]
双侧b16-f10植入模型中,组合肿瘤内注射加热mva与肿瘤内递 送免疫检查点阻断抗ctla-4抗体导致协同治疗效果
[0268]
在实施例13,18和21中,我们显示heat-mva的肿瘤内注射与全身 (腹膜内)递送免疫检查点阻断的组合导致黑色素瘤和结肠腺癌模型中的 协同抗肿瘤作用。在这里,我们测试heat-mva和抗ctla-4抗体(用于 腹膜内递送的剂量的1/10)的共同给药是否将在严格的双侧肿瘤植入模型 中实现抗肿瘤作用。简言之,将b16-f10黑色素瘤细胞皮内植入c57b/6 小鼠的左侧和右侧(向右侧5
×
105,左侧为1
×
105)。肿瘤植入后8天, 我们每周两次腹膜内注射heat-mva(热灭活的2
×
107pfu的mva)或 pbs到右侧较大的肿瘤。三组小鼠用heat-mva治疗,每组接受:(i)腹 膜内递送抗ctla-4(100μg/小鼠)(ii)肿瘤内递送同种型抗体(10μg/ 小鼠)或(iii)肿瘤内递送抗ctla-4抗体(10μg/小鼠)(图22)。所有 pbs处理的小鼠由于注射和未注射的肿瘤的快速生长而早死亡(图 22a-b)。肿瘤内共注射heat-mva和同种型抗体消除了10个注射肿瘤中 的7个,但仅清除了10个非注射肿瘤中的1个(图22c-d)。相比之下, 肿瘤内共注射heat-mva和抗ctla-4抗体(10μg/小鼠)根除了10个注 射的肿瘤中的10个,并且清除了10个未注射的肿瘤中的7个(图22e-f), 其为相当于肿瘤内注射heat-mva和
腹膜内递送抗ctla-4抗体(100μ g/小鼠)的组合的治疗效果(图22g-h)。这些结果表明,以较低剂量共 同施用heat-mva和免疫检查点阻断,抗ctla-4抗体,可以实现与heat
‑ꢀ
mva的肿瘤内递送与系统递送较高剂量的抗ctla-4抗体的组合相似的 全身抗肿瘤效果。这种创新方法具有以下几个优点:(i)通过sting依赖 的细胞溶质dna感测机制激活先天免疫和通过batf3依赖性cd103
+
/cd8 交叉呈递dc激活适应性免疫,提供了“原位治疗疫苗”;(ii)这可以在 抗ctla-4抗体存在下进行cd8
+
和cd4
+
细胞毒性t细胞的强烈活化; (iii)这种组合导致cd4
+
调节性t细胞进一步耗尽;(iv)通过cd8
+
和 cd4
+
细胞毒性t细胞的作用,肿瘤抗原的释放和抗肿瘤适应性免疫的最 佳产生,包括抗肿瘤抗体,导致大量的肿瘤杀伤;和(v)该方法还通过以 腹膜内递送所用剂量的十分之一来肿瘤内递送,降低抗ctla-4抗体的全 身毒性。
[0269]
鉴于在iii期临床试验(larkin等,2015)中,抗ctla-4和抗pd-1 抗体的组合比单独的任一种在pd-l1阴性肿瘤中更有效,本发明人将递送 组合的灭活的mva和抗ctla-4和抗pd-1/抗-pd-l1的二者(阻断剂通 常以比单一疗法低的剂量递送,和比通过不同途径(例如肿瘤内静脉内) 的联合给药更低的剂量递送)将会在肿瘤内递送。预计这将导致抗肿瘤免 疫力的进一步增加和进一步改善的生存率,副作用发生率降低。此外,最 近开发的免疫检查点阻断抗体将被包括在这样的联合递送中,例如 lag-3,抗tim-3和抗tigit抗体,或在诸如上述那些临床前模型中的各 种实体瘤的治疗中。
[0270]
***
[0271]
前述实施例说明了本文所述的方法和特征,并不意图是限制。此 外,它们包含对本披露的一般适用性的陈述,并且这些陈述不仅限于它们 出现的特定实施例,而且构成本文所述实验结果的更广泛含义的结论描述 和表达。
[0272]
所有引用的参考文献的内容通过引用整体并入,如同为了所有目的 而完全转录的。
[0273]
更具体地,本技术提供下列各项:
[0274]
1.一种治疗患有一种或多种实体恶性肿瘤的受试者的方法,所述方法 包括将灭活的修饰的牛痘安卡拉(灭活的mva)递送至肿瘤细胞,从而 治疗肿瘤。
[0275]
2.项1的方法,其中所述量对于达成以下的一个或多个是有效的:
[0276]
g、诱导受试者的免疫系统发起抗肿瘤免疫应答;
[0277]
h、减少肿瘤的大小;
[0278]
i、根除肿瘤;
[0279]
j、抑制肿瘤生长;
[0280]
k、抑制肿瘤转移;和
[0281]
l、减少或根除转移性肿瘤。
[0282]
3.项2的方法,其中所述肿瘤包括位于灭活的mva递送位点的肿瘤, 或同时位于受试者体内的所述部位和其它部位的肿瘤。
[0283]
4.根据项1-3中任一项所述的方法,其中所述免疫应答包含以下一个 或多个:
[0284]
f、在肿瘤和/或肿瘤引流淋巴结中增加细胞毒性cd8
+ t细胞;
[0285]
g、通过诱导i型ifn诱导浸润所述肿瘤的树突细胞的成熟;
[0286]
h、诱导在受试者中识别肿瘤中或全身的肿瘤细胞的活化的cd4
+
效应 t细胞;
[0287]
i、减少肿瘤内免疫抑制(调节)cd4
+ t细胞;和
[0288]
j、诱导肿瘤细胞在其表面表达i型mhc并产生i型ifn。
[0289]
5.项1-4的方法,其中所述肿瘤是原发性或转移性黑色素瘤或原发性 或转移性结肠癌。
[0290]
6.根据项1-5中任一项所述的方法,其中以间隔开的时间间隔重复所 述灭活的mva的递送。
[0291]
7.根据项6所述的方法,其中所述重复递送持续数周,数月或数年或 在持续存在益处或达到最大耐受剂量的情况下无限期。
[0292]
8.根据项1-7中任一项所述的方法,其中所述灭活的mva的递送是 通过肠胃外途径。
[0293]
9.项8的方法,其中所述灭活的mva的递送是通过肿瘤内或静脉内 注射。
[0294]
10.根据项1-9中任一项所述的方法,其中所述受试者是人。
[0295]
11.根据项1-10中任一项所述的方法,其中所述灭活的mva以每次 施用的剂量约10
5-10
10
个噬菌斑形成单位(pfu)的范围内递送。
[0296]
12.项11的方法,其中所述灭活的mva以每次施用的剂量约106至 约109个噬菌斑形成单位(pfu)的范围内递送。
[0297]
13.根据项1-12中任一项所述的方法,其中所述递送以每月一次至每 周两次的频率重复。
[0298]
14.根据项13所述的方法,其中所述递送每周重复一次。
[0299]
15.根据项1-14中任一项或多项所述的方法,其中所述灭活的mva 是uv灭活的mva。
[0300]
16.根据项1-14中任一项所述的方法,其中所述灭活的mva是热灭 活的mva。
[0301]
17.项4的方法,其中效应t细胞的诱导和激活伴随着所述肿瘤中调 节性cd4
+
细胞的减少。
[0302]
18.项4的方法,其中所述灭活的mva通过诱导i型ifn诱导浸润 所述肿瘤的树突细胞的成熟。
[0303]
19.项4的方法,其中所述灭活的mva诱导感染的肿瘤细胞中i型 mhc的表达和i型干扰素的诱导。
[0304]
20.一种用于治疗受试者中的固体恶性肿瘤的方法,其包括向受试者 的肿瘤细胞递送有效诱导受试者的免疫系统以发起抗肿瘤的免疫应答的 量的灭活的mva。
[0305]
21.项20的方法,其中所述免疫应答是系统性的。
[0306]
22.项20的方法,其中所述免疫应答影响或有助于以下中的一个或多 个:减少肿瘤大小,根除肿瘤,抑制肿瘤或转移生长。
[0307]
23.根据项20或21所述的方法,其中所述灭活的mva有效地达成 以下的一个或多个:
[0308]
m、诱导受试者的免疫系统发起抗肿瘤免疫应答;
[0309]
n、减少肿瘤的大小;
[0310]
o、根除肿瘤;
[0311]
p、抑制肿瘤生长;
[0312]
q、抑制肿瘤转移;和
[0313]
r、减少或根除转移性肿瘤。
[0314]
24.根据项23所述的方法,其中所述肿瘤包括位于灭活的mva递送 位点的肿瘤,或同时位于受试者体内的所述部位和其它部位的肿瘤。
[0315]
25.根据项20-24中任一项所述的方法,其中所述免疫应答包含以下 一个或多个:
[0316]
k、在肿瘤和/或肿瘤引流淋巴结中增加细胞毒性cd8
+ t细胞;
[0317]
l、通过诱导i型ifn诱导浸润所述肿瘤的树突细胞的成熟;
[0318]
m、诱导在受试者中识别肿瘤中或全身的肿瘤细胞的活化的cd4
+
效应 t细胞;
[0319]
n、降低肿瘤内免疫抑制(调节)cd4
+ t细胞;和
[0320]
o、诱导肿瘤细胞在其表面表达i型mhc并产生i型ifn。
[0321]
26.项20-25的方法,其中所述肿瘤是原发性或转移性黑色素瘤或原 发性或转移性结肠癌。
[0322]
27.根据项20-25中任一项所述的方法,其中以间隔开的时间间隔重 复所述灭活的mva的递送。
[0323]
28.根据项27所述的方法,其中所述重复递送持续数周,数月或数年 或在持续存在益处或达到最大耐受剂量的情况下无限期。
[0324]
29.根据项20-28中任一项所述的方法,其中所述灭活的mva的递 送是通过肠胃外途径。
[0325]
30.根据项29所述的方法,其中所述灭活的mva的递送是通过肿瘤 内或静脉内注射。
[0326]
31.根据项20-30中任一项所述的方法,其中所述受试者是人。
[0327]
32.根据项31所述的方法,其中所述灭活的mva以每次施用的剂量 约10
5-10
10
个噬菌斑形成单位(pfu)的范围内递送。
[0328]
33.根据项32所述的方法,其中所述灭活的mva以每次施用的剂量 约106至约109个噬菌斑形成单位(pfu)的范围内递送,
[0329]
34.根据项20至33中任一项所述的方法,其中所述递送以每月一次 至每周两次的频率重复。
[0330]
35.根据项34所述的方法,其中所述递送每周重复一次。
[0331]
36.根据项20-35中任一项或多项所述的方法,其中所述灭活的mva 是uv灭活的mva。
[0332]
37.根据项20-35中任一项所述的方法,其中所述灭活的mva是热 灭活的mva。
[0333]
38.一种用于治疗受试者的恶性肿瘤的方法,所述方法包括以有效诱 导受试者的免疫系统发起抗肿瘤免疫应答的量递送灭活的mva至受试者 的肿瘤细胞并且以在肿瘤内有效阻断肿瘤细胞,基质细胞,或肿瘤浸润性 免疫细胞引起的免疫抑制机制的第二数量联合给药受试者免疫检查点阻 断剂。
[0334]
39.项38的方法,其中所述给药是通过肠胃外途径。
[0335]
40.根据项39所述的方法,其中所述递送是通过肿瘤内注射和所述给 药是通过静脉内途径。
[0336]
41.根据项39所述的方法,其中所述递送和给药均通过静脉内途径。
[0337]
42.项39的方法,其中递送和给药都是通过肿瘤内注射。
[0338]
43.项38的方法,其中所述免疫检查点阻断剂选自pd-1抑制剂, pd-l1抑制剂和ctla4抑制剂。
[0339]
44.项44的方法,其中每种所述抑制剂是抗体。
[0340]
45.项38的方法,其中所述肿瘤是原发性或转移性黑色素瘤或原发性 或转移性结肠癌。
[0341]
46.根据项38-46所述的方法,其中所述灭活的mva的递送和所述 免疫检查点阻断剂的给药是根据其自身的间隔开的间隔的施用方案分别 施用。
[0342]
47.项47的方法,其中首先递送第一剂量的灭活的mva,并且经过 一段时间之后给药第一剂量的免疫检查点阻断剂。
[0343]
48.根据项48或49所述的方法,其中所述递送和给药在相同的整个 时间段期间并行发生。
[0344]
49.根据项48所述的方法,其中灭活的mva的递送和免疫检查点阻 断剂的给药的一个或同时分别进行数周,数月或数年或在持续存在益处或 达到最大耐受剂量的情况下无限期。
[0345]
50.项38或48的方法,其中所述灭活的mva以每次施用剂量约 10
5-10
10
个噬菌斑形成单位(pfu)的范围内递送。
[0346]
51.项51的方法,其中所述灭活的mva以每次施用剂量约106至约 109个噬菌斑形成单位(pfu)的范围内递送。
[0347]
52.根据项47-52中任一项所述的方法,其中所述灭活的mva的递 送以每月一次至每周两次的频率重复灭活。
[0348]
53.项53的方法,其中所述灭活的mva的递送每周一次重复。
[0349]
54.根据项38-54中任一项或多项所述的方法,其中所述灭活灭活的 mva是uv灭活的mva。
[0350]
55.根据项38-54中任一项所述的方法,其中所述灭活的mva是热 灭活的mva。
[0351]
56.项38的方法,其中同时施用灭活的mva和免疫检查点阻断剂。
[0352]
57.项57的方法,其中所述灭活的mva和免疫检查点阻断剂在相同 的组合物施用。
[0353]
58.项57的方法,其中所述灭活的mva和免疫检查点阻断剂在肿瘤 内递送。
[0354]
59.项38的方法,其中顺序地给予灭活的mva和免疫检查点阻断剂。
[0355]
60.项60的方法,其中灭活的mva和免疫检查点阻断剂在肿瘤内递 送。
[0356]
参考文献:
[0357]
1.jochems,c.and j.schlom.(2011).tumor-infiltrating immune cells and prognosis:the potential link between conventional cancer therapy and immunity.exp biol med(maywood)236(5):567-579.
[0358]
2.mlecnik,b.,g.bindea,f.pages,and j.galon.(2011).tumor immunosurveillance in human cancers.cancer metastasis rev 30(1):5-12. 3.angell,h.and j.galon.(2013).from the immune contexture to the immunoscore:the role of prognostic and predictive immune markers in cancer.curr opin immunol 25(2):261-267.
activated protein kinase(pkr)-dependent pathway during the early phase of virus replication.virology 391(2):177-186.
[0399]
44.willis,k.l.,s.patel,y.xiang,and j.l.shisler.(2009).the effect of the vaccinia k1 protein on the pkr-eif2alpha pathway in rk13 and hela cells.virology 394(1):73-81.
[0400]
45.kotwal,g.j.,a.w.hugin,and b.moss.(1989).mapping and insertional mutagenesis of a vaccinia virus gene encoding a 13,800-da secreted protein. virology 171(2):579-587.
[0401]
46.bartlett,n.,j.a.symons,d.c.tscharke,and g.l.smith.(2002).the vaccinia virus n1l protein is an intracellular homodimer that promotes virulence.j gen virol 83(pt 8):1965-1976.
[0402]
47.brandt,t.,m.c.heck,s.vijaysri,g.m.jentarra,j.m.cameron,and b.l. jacobs.(2005).the n-terminal domain of the vaccinia virus e3l-protein is required for neurovirulence,but not induction of a protective immune response.virology 333(2):263-270.
[0403]
48.chang,h.w.,j.c.watson,and b.l.jacobs.(1992).the e3l gene of vaccinia virus encodes an inhibitor of the interferon-induced,double-stranded rna-dependent protein kinase.proc natl acad sci u s a 89(11): 4825-4829.
[0404]
49.xiang,y.,r.c.condit,s.vijaysri,b.jacobs,b.r.williams,and r.h.silverman. (2002).blockade of interferon induction and action by the e3l double-stranded rna binding proteins of vaccinia virus.j virol 76(10): 5251-5259.
[0405]
50.beattie,e.,e.b.kauffman,h.martinez,m.e.perkus,b.l.jacobs,e.paoletti, and j.tartaglia.(1996).host-range restriction of vaccinia virus e3l-specific deletion mutants.virus genes 12(1):89-94.
[0406]
51.brandt,t.a.and b.l.jacobs.(2001).both carboxy-and amino-terminal domains of the vaccinia virus interferon resistance gene,e3l,are required for pathogenesis in a mouse model.j virol 75(2):850-856.
[0407]
52.langland,j.o.,j.c.kash,v.carter,m.j.thomas,m.g.katze,and b.l.jacobs. (2006).suppression of proinflammatory signal transduction and gene expression by the dual nucleic acid binding domains of the vaccinia virus e3l proteins.j virol 80(20):10083-10095.
[0408]
53.smith,e.j.,i.marie,a.prakash,a.garcia-sastre,and d.e.levy.(2001).irf3 and irf7 phosphorylation in virus-infected cells does not require double-stranded rna-dependent protein kinase r or ikappa b kinase but is blocked by vaccinia virus e3l protein.j biol chem 276(12):8951-8957.
[0409]
54.guerra,s.,a.caceres,k.p.knobeloch,i.horak,and m.esteban.(2008). vaccinia virus e3 protein prevents the antiviral action of isg15.plos pathog 4(7):e1000096.
cd8alpha+dendritic cells in cytotoxic t cell immunity.science 322(5904):1097-1100.
[0447]
92.edelson,b.t.,w.kc,r.juang,m.kohyama,l.a.benoit,p.a.klekotka,c. moon,j.c.albring,w.ise,d.g.michael,d.bhattacharya,t.s.stappenbeck, m.j.holtzman,s.s.sung,t.l.murphy,k.hildner,and k.m.murphy.(2010). peripheral cd103+dendritic cells form a unified subset developmentally related to cd8alpha+conventional dendritic cells.j exp med 207(4): 823-836.
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1