去铁胺在制备治疗帕金森病的药物中的应用
1.本发明属于帕金森防治新机制技术领域,具体涉及去铁胺在制备治疗帕金森病的药物中的应用。
技术背景
2.①
帕金森病的现状
3.随着人口的老龄化,神经退行性疾病的发病率及患病率逐年升高,其中帕金森病(parkinson’s disease,pd)已经成为严重影响人类生活的第二大神经退行性疾病,临床表现为肌强直、运动迟缓、静止性震颤等症状,严重影响患者及其家庭的生活质量。遗传因素、环境因素、衰老、脑血管病、中毒、感染等多种因素均会引发帕金森病,其主要病理特点是黑质多巴胺能神经元选择性死亡和α-突触核蛋白(路易小体的主要成分)的大量聚集。
4.研究表明帕金森病的发生与氧化压力、线粒体功能异常、泛素化蛋白酶降解系统的缺陷、神经细胞凋亡、突触传递障碍、胞内钙稳态失衡等有关。尽管帕金森病的临床表现、病理改变以及致病相关基因都已经开展了深入研究,但是目前的治疗手段包括药物治疗、手术治疗,干细胞和基因治疗都只能暂时改善症状,并不能阻止或延缓疾病的进程,主要原因是帕金森病的发生机制仍然没有完全清楚,因此进一步开展帕金森病发生发展的分子机制研究是治疗和缓解帕金森病的基础。
5.②
铁过载与帕金森病密切相关
6.在神经退行性疾病的研究中发现,金属离子稳态失衡会加剧氧化压力从而导致神经元死亡。铁作为机体代谢过程中必须的微量金属元素,维持着机体的多种生物学功能。在脑中铁元素不仅参与神经组织的新陈代谢、能量供给、dna 复制、细胞周期等过程,还涉及髓磷脂的合成、神经递质的合成与代谢。然而铁元素的过度累积会引发多种疾病,例如:血浆铜蓝蛋白缺乏症和神经铁蛋白病变,就是由于铁元素的过度沉积导致的神经退行性病变。临床研究发现,帕金森病人脑中铁含量明显高于正常人,而且铁在黑质神经元和胶质细胞中的含量与帕金森病的严重程度呈正相关。值得注意的是,动物的脑中注射铁剂也会引发行为学的改变以及帕金森病的症状。铁过载可能引发帕金森病:
①
黑质中铁水平的升高会增加活性氧(reactive oxygen species,ros),造成氧化压力,引发神经退行性病变、肌张力障碍和震颤等运动障碍。
②
fe
3+
在过氧化氢的作用下将多巴胺氧化为神经毒素六羟基多巴胺,六羟基多巴胺会进一步地释放更多的fe
2+
,加重帕金森病。
③
铁作为酪氨酸羟化酶限速酶的辅因子,影响多巴胺合成。过量的铁导致多巴胺合成过度,首先抑制多巴胺的正常功能,其次释放到细胞质产生大量ros,促使六羟基多巴胺的合成增加,损伤多巴胺神经元。
④
铁还会诱导α-突触核蛋白从α-螺旋错误折叠形成病理状态的β-折叠,而β-折叠的α-突触核蛋白是路易小体的主要成分。
⑤
在帕金森病中铁水平的增加会产生大量丙二醛,降低谷胱甘肽,进而导致线粒体呼吸链酶复合体i和 iv活性降低,引起线粒体功能异常。以上研究指出,铁大量累积会产生ros、增加氧化压力、引起线粒体功能障碍、α-突触核蛋白错误折叠,其中氧化压力被认为是铁过载导致帕金森病的主要诱因之一。但是抗氧化剂并不能治
疗帕金森病,提示铁过载可能还存在其他机制影响帕金森病的发生和发展。因此,铁介导帕金森病的分子机制有待进一步的阐明。
7.③
自噬缓解帕金森病
8.引发帕金森病的氧化压力、线粒体功能异常以及α-突触核蛋白的聚集都与自噬(autophagy)有着密切的联系。自噬在真核生物中高度保守,作为细胞内的“清道夫”,将大分子物质、长寿命蛋白以及受损细胞器包裹形成自噬小体,通过溶酶体降解,维持机体的内稳态。自噬过程发生的关键蛋白 atg5-atg12-atg16和lc3/atg8类泛素连接系统、atg1/ulk1蛋白激酶复合体、 vps34-beclin1/atg6复合体、atg9等,控制着自噬小体的膜融合、形成、运输和识别等功能。越来越多的研究表明,自噬在神经元的存活、神经退行性变性中起着至关重要的作用,而自噬的功能异常导致错误折叠的蛋白、损伤的细胞器的聚集,引起亨廷顿病、阿尔兹海默、帕金森病等多种神经退行性疾病。最新研究发现遗传性帕金森病人常染色体的基因突变都与自噬的相关基因有关。 sala等报道帕金森病中的自噬水平明显低于正常人,在过表达α-突触核蛋白的pd细胞模型中,自噬基因beclin1的表达显著抑制。此外由dj-1基因功能缺失造成的遗传性帕金森模型中,lc3-ii的表达显著低于正常小鼠,并伴有明显的线粒体功能障碍。因此,提高自噬水平有望成为治疗帕金森病的策略。激活自噬增加组蛋白乙酰化,可以减少pd小鼠多巴胺神经元的死亡。spencer等报道过表达自噬相关基因beclin1也能够减轻小鼠pd的症状。同样地,用1
‑ꢀ
甲基-4-苯基-1,2,3,6-四氢吡啶(1-methyl-4-phenyl-1,2,3,6
‑ꢀ
tetrahydropyridine,mptp)诱导的小鼠pd模型中,使用自噬激活剂-雷帕霉素处理后,显著增加多巴胺能神经元,减少α-突触核蛋白的聚集。诱导pd模型的神经毒素
‑‑
鱼藤酮就是通过抑制自噬功能实现的。
技术实现要素:
9.自噬功能降低参与帕金森病的发生与发展。且有文献报道,在神经退行性疾病中直接激活自噬可能是有害的,当自噬被抑制时,最佳处理方法是通过相应的不同的途径去活化自噬。
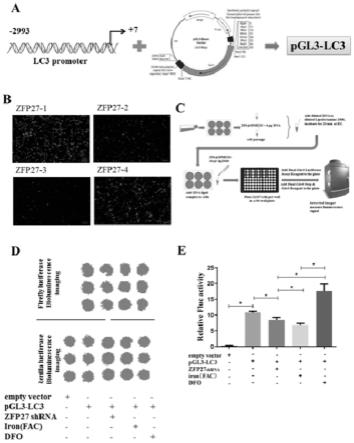
10.为实现上述目的,本发明提供了去铁胺在制备治疗帕金森病的药物中的应用。
11.进一步地,所述药物中的去铁胺通过激活转录因子zfp27促进自噬基因lc3 启动子增加达到激活自噬作用,通过自噬治疗帕金森病。
12.所述去铁胺为去铁胺甲磺酸酯,分子式为:c
25h48
n6o8.ch4o3s,分子结构式为:
13.本发明的机理介绍:小鼠中的zfp27相当于人的znf585,在帕金森病模型中,铁过度沉积降低转录因子zfp27和igf-2的活性抑制了自噬,进而促进多巴胺神经元的死亡和增加α-syn的积累,通过去铁胺(dfo)减少帕金森病模型中铁沉积,促进激酶激活剂igf-2增加,活化zfp27活化lc3启动子增强自噬,以达到治疗帕金森疾病的目的。
附图说明
14.图1:a为转棒行为学检测铁沉积对pd小鼠运动功能的影响;
15.b为western blot检测pd小鼠中转铁蛋白受体1(tfr1)的表达;
16.c为perl’s染色显示pd小鼠铁含量;
17.d为铁沉积对小鼠脑黑质中的酪氨酸羟化酶和α-突触核蛋白的聚集的影响;
18.e为image j软件分析酪氨酸羟化酶和α-突触核蛋白的灰度值
19.f为免疫荧光染色显示:铁对黑质多巴胺神经元的存活影响;
20.图2:为帕金森病小鼠黑质中铁沉积和铁剥夺(去铁胺处理)调控自噬的变化;
21.其中,a为通过western blot实验检测对照组、mptp模型组、氯喹组、铁组和去铁胺组中自噬相关基因lc3,p62和beclin1的变化;
22.b为image j软件分析各组中自噬相关基因的蛋白水平变化;
23.c为透射电镜呈现小鼠脑黑质中自噬小体的变化(用黑色箭头标注);
24.d为免疫双染实验观察小鼠脑黑质多巴胺神经元中lc3的变化;
25.图3:a为转录组测序的热图结果显示图;
26.b为转录组测序的韦恩图;
27.c为qpcr验证差异表达基因中的4个激酶相关基因和2个转录因子的变化;
28.d为western blot实验检测铁组与pd模型组的脑黑质zfp27的表达;
29.e:image j软件分析zfp27的灰度值;
30.图4:为pd细胞模型中,铁与zfp27和igf-2的表达关系图;
31.其中,a为cck8实验检测mpp
+
、柠檬酸铁铵(fac)、去铁胺(df0)和氯喹(cq) 在shsy5y细胞中的最适作用浓度;
32.b,c为d细胞模型不同处理组中通过western blot检测转铁蛋白tfr1的蛋白水平,并通过image j进行灰度分析;
33.d,e为pd细胞模型不同处理组中通过western blot检测自噬相关基因lc3、p62、beclin1的蛋白水平变化,并通过image j进行灰度分析;
34.f,g为pd细胞模型不同处理组中通过western blot检测zfp27和igf-2 的蛋白水平变化,并通过image j进行灰度分析;
35.图5:为帕金森病小鼠模型中去铁胺活化转录因子zfp27激活自噬
36.其中,a:lc3启动子克隆到pgl3 basic载体上获得融合质粒pgl3-lc3;
37.b:通过荧光显微镜观察筛选出的转染效率最高的zfp27 shrna质粒;
38.c:质粒转染后通过活体成像系统检测空载组、pgl3-lc3、zfp27 shrna(敲低zfp27)、铁处理组和去铁胺处理组萤火虫荧光素酶和海肾荧光素酶的实验流程;
39.d:双荧光素酶报告基因实验成像结果图;
40.e:相对荧光素酶分析敲减zfp27对lc3启动子的活性的影响。
具体实施方式
41.为使本发明的目的、技术方案和优点更加清楚明了,下面结合具体实施方式并参照附图,对本发明进一步详细发明。应该理解,这些描述只是示例性的,而并非要限制本发明的范围。此外,在以下发明中,省略了对公知结构和技术的描述,以避免不必要地混淆本
发明的概念。
42.实施例1:铁沉积加重帕金森病
43.为了评估铁对pd小鼠的影响。通过转棒行为学检测,mptp组的潜伏期与对照组相比显著降低(图1a)。与mptp组相比,铁处理组(iron)小鼠的潜伏期也明显降低,而铁螯合剂去铁胺dfo处理组小鼠的潜伏期增强,但没有显著性差异(图1a)。转铁蛋白受体1(tfr1)维持细胞内铁稳态,tfr1的表达与铁含量呈正相关。如图1b所示,mptp组tfr1表达高于对照组,铁组tfr1表达高于对照组,dfo组tfr1表达降低。进一步经perl’s染色发现(如图1c所示), mptp诱导的pd小鼠铁含量明显高于对照组。经dfo处理后,铁呈现减少趋势。为了验证铁的积累是否伴随着pd的发展,如图1d和1e,相对于pd模型小鼠,铁诱导后酪氨酸羟化酶(th)蛋白水平明显降低。此外,随着dfo减少铁的累积后,th蛋白水平得到部分恢复(图1d和1e)。与mptp动物相比,铁处理小鼠的α-synuclein(α-syn)积累明显增加,但dfo处理的α-syn表达无显著性(图 1d和1e)。此外,免疫荧光染色结果也显示(图1f):与mptp组相比,铁能减少黑质sn中th阳性细胞的数量;dfo组中th阳性神经元较mptp组增多;
44.综上所述,这些数据表明铁的积累加重pd小鼠运动缺陷和病理变化。
45.实施例2:自噬能够缓解帕金森病
46.在mptp诱导的pd模型中,检测了自噬基因lc3,western blot结果显示:相比正常对照组,sn中lc3ii明显降低,而beclin 1没有变化(图2a和2b)。那么铁是否调控自噬,如图2a所示,铁降低lc3ii和增加p62的表达,而dfo 处理后显著激活自噬(图2a和2b)。此外,透射电子显微镜(tem)观察双层膜结构的自噬小体(用黑色标记),与图2c中的pd模型相比,经铁处理后,自噬小体减少;相反,dfo处理组自噬小体数量增加(图2c)。此外,免疫荧光双染(lc3/th) 也显示,铁处理组自噬减少,铁处理后th阳性细胞数量也减少(图2d)。与pd 模型相比,dfo组多巴胺神经元自噬部分恢复(图2d)。
47.以上数据表明,在pd模型中,铁是通过lc3而不是beclin 1抑制自噬,而自噬的减少参与pd的发生发展。
48.实施例3:zfp27和igf-2参与帕金森病中铁抑制自噬的作用
49.为了研究铁是如何调节自噬加速帕金森病的进展,通过转录组测序分析,通过热图(图3a)和韦恩图(图3b)中发现pd小鼠和铁处理组小鼠相比有37 个差异表达基因(degs),包括20个上调基因和17个下调基因(degs)(图3a和 3b)。在所有37个degs中包括2个转录因子和4个激酶相关基因(gm4724、zfp27、 wdcp、myo5a、igf-2、fgfrlop)。通过qpcr检测,在小鼠脑黑质(sn)中6个基因中有gm4724、igf-2、fgfrlop 3个基因表达升高或降低(图3c)。又从蛋白水平上筛选两个转录因子(zfp27、gm4724)和两个激酶相关基因包括胰岛素样生长因子2(igf-2)和fgfrlop。结果发现:铁处理组的小鼠sn中转录因子zfp27 明显下降,而dfo处理后zfp27表达水平高于mptp模型,但铁组和去铁胺组的 gm4724无变化,(图3d和3e);sn中igf-2抗体检测不到任何无信号。因此,为了确定是哪些基因介导pd中铁抑制自噬的作用,用sh-sy5y细胞建立pd细胞模型。如图4a所示,通过cck-8检测,不同浓度柠檬酸铁铵(fac)、dfo、cq 分别处理sh-sy5y细胞系12h、24h(图4a)。如图4a所示,当10μm cq、50μg/ml fac、100μm dfo分别处理sh-sy5y细胞后,细胞活力与对照组无差异。因此,使用10μm cq、50μg/ml fac和100μm dfo进行以下检测。在pd细胞模型中,mpp+/mptp组tfr1蛋白水平显著高于对照组,dfo组tfr1蛋白水平显著低于mptp模型组(图4b、4c)。同时发现:铁处理组
lc3ii降低,dfo组恢复了mpp+/mptp模型组被降低的lc3ii蛋白表达水平(图4d和4e)。pd细胞模型中beclin 1蛋白表达降低,但铁处理后细胞的beclin 1蛋白水平升高,因此 pd细胞模型中铁通过降低lc3ii抑制自噬。进一步检测:pd模型组zfp27/znf585 表现出低于正常组的趋势,dfo处理组zfp27的表达高于模型组(图4f和4g)。同样,较正常对照组,pd模型组igf-2明显受到抑制。此外,与pd模型组相比,铁处理组的igf-2蛋白水平下降,但dfo处理增加igf-2的表达(图4f和4g)。而小鼠中的zfp27相当于人的znf585
50.综上所述,小鼠和细胞pd模型的数据均表明,铁通过转录因子zfp27和激酶激活剂igf-2抑制自噬在pd发生发展中发挥作用。
51.实验结果4:去铁胺活化转录因子zfp27激活自噬
52.为了进一步了解zfp27如何调控自噬,通过双荧光素酶报告基因实验检测 zfp27与自噬基因启动子的结合能力。细胞和动物模型中均显示铁抑制自噬是通过lc3ii和p62而不是beclin 1(图4d和4e)。因此,获得lc3启动子并与双荧光素酶报告基因pgl3-basic载体融合,构建了包含lc3与pgl3-basic载体的报告基因质粒pgl3-lc3(图5a)。同时,评价四个zfp27 shrna的转染效率,最终使用zfp27-1质粒进行后续实验(图5b)。最后,使用活体成像系统,通过荧光素酶报告基因检测lc3的启动子活性(图5c)。如图5d和e所示,hek293t 细胞中转染质粒pgl3-lc3后荧光显著升高,而用shrna敲减zfp27的表达后,荧光素酶活性明显降低(图5d和5e)。铁处理后细胞的荧光素酶活性也低于敲减 zfp27组,而dfo处理后即活化zfp27后确实增强lc3的启动子活性(图5d和 5e)。综上所述,这些结果表明zfp27直接结合lc3启动子增强自噬,而dfo激活zfp27激活自噬。
53.综上,在帕金森病模型中,铁过度沉积降低转录因子zfp27和igf-2的活性抑制了自噬;而dfo处理后激活igf-2和zfp27促进自噬进而减少多巴胺神经元的死亡,减少α-syn的积累
54.实施例5:上述使用的去铁胺dfo为去铁胺甲磺酸酯,动物实验用量是 100mg/kg,一天两次,共计10天,细胞实验是100μm,12h;去铁胺甲磺酸酯分子式为c
25h48
n6o8.ch4o3s,分子量:656.79g/mol,分子结构式如下:
[0055][0056]
应当理解的是,本发明的上述具体实施方式仅仅用于示例性发明或解释本发明的原理,而不构成对本发明的限制。因此,在不偏离本发明的精神和范围的情况下所做的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。此外,本发明所附权利要求旨在涵盖落入所附权利要求范围和边界、或者这种范围和边界的等同形式内的全部变化和修改例。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1