尖叶假龙胆来源的Gen-miR-1抑制心肌纤维化的用途
尖叶假龙胆来源的gen-mir-1抑制心肌纤维化的用途
技术领域
1.本发明属于医药技术领域,涉及尖叶假龙胆来源的gen-mir-1抑制心肌纤维化的用途。
背景技术:
2.尖叶假龙胆(gentianella acuta,g.acuta)为龙胆科假龙胆属一年生草本植物,广泛分布于我国河北、东北、内蒙古、山西、山东等地区。在蒙药中全草入药,味微苦、性凉,具有清热解毒之效。民间药用植物调查发现,鄂温克族猎民用其泡水代茶饮治疗心绞痛等疾病,疗效极为显著,是该属唯一一种具有治疗心脏病作用的植物
[1,2]
。基于中医心系疾病的“热毒学说”,认为心肌梗死后心肌纤维化(myocardial fibrosis,mf)是由于浊瘀蓄积,蕴结成热毒,导致热毒瘀并存的病理演变结果。现代临床和实验研究表明,g.acuta及其有效成分防治mf的作用
[3]
,其水提物可通过抑制tlr4/nf-κb炎症信号通路减轻大鼠急性心肌损伤;也可通过抗炎、抗氧化、抑制tgf-β1/smads信号通路改善异丙肾上腺素(isoproterenol,iso)诱导的大鼠mf
[4-6]
。近年来有研究报道,g.acuta 的酮类有效成分可通过抗氧化和抗凋亡作用抑制离体大鼠缺血/再灌注损伤;减轻h2o2所致心肌细胞毒性和损伤;其含有的四种主要酮类化合物均能通过激活nrf2/are通路,抑制h2o2所致的h9c2细胞氧化损伤
[7,8]
。我们最新实验研究证实,g.acuta有效成分雏菊叶龙胆酮(bellidifolin,bel)可抑制小鼠mf。 bel通过抑制tgf-β1/smads和p38mapk信号通路,阻断smad2/3和p38/mapk 的磷酸化活化,阻止nr4a1磷酸化及其胞核到胞浆转位,进而抑制tgf-β1诱导的心脏成纤维细胞(cardiac fibroblasts,cfs)活化(该研究已发表在frontpharmacol.2021)
[9]
。此外,我们研究证实g.acuta中另一种酮成分芒果苷可通过调控gsh合成和线粒体能量代谢发挥抗mf作用
[10]
。上述研究表明,g. acuta及酮类有效成分确有抗mf的功效。但由于g.acuta中有效成分复杂,作用机制尚未完全明确等因素,极大制约了g.acuta及其活性成分在心血管疾病中的应用。因此寻找g.acuta中更高效、更稳定、更安全的新型活性成分并明确其作用机制,成为心血管疾病治疗药物开发过程中亟待解决的问题。
[0003]
植物来源mirnas可以通过依赖sdit1蛋白介导胃部吸收,进入体内发挥靶向调控作用
[11]
,不仅可以调节人体脂肪代谢
[12]
,甚至能够影响肿瘤的发生
[13]
。研究还发现,中药金银花中mir2911可以抑制甲流病毒在动物体内的复制
[14]
,自此开启了中药来源mirnas跨物种调节治疗疾病的研究热点。有趣的是,我们发现了丹参中特异性存在的sal-mir-1和sal-mir-3可以跨物种调控 otud7b/klf4/nmhcⅱa通路影响vsmc表型转换,从而抑制细胞的迁移和粘附,有效改善了血管内膜增生(研究成果发表在:theranostics.2020)
[15]
;此外,丹参来源sal-mir-58可以通过调控klf3/nedd4l/pfkp轴诱导自噬并减轻vsmc 炎症反应(研究成果发表在:mol ther nucleic acids.2020)
[16]
。但是并非所有的中药中都存在mirna,也并非存在于中药中的mirna必然与该味中药或其有效成分具有相同的药理作用。因此,g.acuta中是否存在mirna,他们是否能在体内稳定存在并发挥作用,以及具备何种药效尚无法预料。
[0004]
参考文献:
[0005]
[1]乌尼尔,春亮,哈斯巴根.鄂温克族民间药用植物及其与蒙古医药的比较[j].中国民族民间医药.2009;18(17):156-158.
[0006]
[2]庞宇航,管伟,胡晓阳,高彦宇,李冀.尖叶假龙胆的化学成分及药理作用研究进展[j].中草药.2018;49(22):5468-5476.
[0007]
[3]朱骏宏,金政,褚庆民,都治伊,吴伟.清热解毒法辅助pci术对冠心病病人临床疗效及mace影响的meta分析[j].中西医结合心脑血管病杂志.2021;19(07):1070-1078.
[0008]
[4]sunjh,yanghx,yaott,liy,ruanl,xugr,zhangc,guogx,liay.gentianellaacutapreventsacutemyocardialinfarctioninducedbyisoproterenolinratsviainhibitionofgalectin-3/tlr4/myd88/nf-κbinflammatorysignalling[j].inflammopharmacology.2021;29:205-219.
[0009]
[5]liay,wangjj,yangsc,zhaoys,lijr,liuy,sunjh,anlp,guanp,jies.protectiveroleofgentianellaacutaonisoprenalineinducedmyocardialfibrosisinratsviainhibitionofnf-κbpathway[j].biomedpharmacother.2019;110:733-741.
[0010]
[6]yanghx,xugr,zhangc,sunjh,zhangy,songjn,liyf,liuy,liay.theaqueousextractofgentianellaacutaimprovesisoproterenol-inducedmyocardialfibrosisviainhibitionofthetgf-β1/smadssignalingpathway[j].intjmolmed.2020;45(1):223-233.
[0011]
[7]wangz,wug,yuy,liuh,yangb,kuangh,wangq.xanthonesisolatedfromgentianellaacutaandtheirprotectiveeffectsagainsth2o
2-inducedmyocardialcellinjury[j].natprodres.2018;32(18):2171-2177.
[0012]
[8]renk,suh,lvlj,yilt,gongx,dangls,zhangrf,limh.effectsoffourcompoundsfromgentianellaacuta(michx.)hultenonhydrogenperoxide-inducedinjuryinh9c2cells[j].biomedresint.2019;2692970.
[0013]
[9]yanghx,sunjh,yaott,liy,xugr,zhangc,liuxc,zhouww,songqh,zhangy,liay.bellidifolinamelioratesisoprenaline-inducedmyocardialfibrosisbyregulatingtgf-beta1/smadsandp38signalingandpreventingnr4a1cytoplasmiclocalization[j].frontpharmacol.2021;12:644886.
[0014]
[10]songj,mengy,wangm,lil,liuz,zhengk,wul,liub,houf,lia.mangiferinactivatesnrf2toattenuatecardiacfibrosisviaredistributingglutaminolysis-derivedglutamate[j].pharmacolres.2020;157:104845.
[0015]
[11]zhouz,lix,liuj,dongl,chenq,liuj,kongh,zhangq,qix,houd,zhangl,zhangg,liuy,zhangy,lij,wangj,chenx,wangh,zhangj,chenh,zenk,zhangcy.honeysuckle-encodedatypicalmicrorna2911directlytargetsinfluenzaaviruses[j].cellres.2015;25(1):39-49.
[0016]
[12]chinar,fongmy,somlog,wuj,swiderskip,wux,wangse.cross-kingdominhibitionofbreastcancergrowthbyplantmir159[j].cellres.2016;026(002):217-228.
[0017]
[13]chenq,zhangf,dongl,wuh,xuj,lih,wangj,zhouz,liuc,wangy,liuy,lul,wangc,lium,chenx,wangc,zhangc,lid,zenk,wangf,zhangq,zhangcy.sidt1-dependentabsorptioninthestomachmediateshostuptakeofdietaryandorallyadministeredmicrornas[j].cellres.2020;0:1-12.
[0018]
[14]yanggs,zhengb,qiny,zhouj,yangz,zhangxh,zhaohy,yanghj,wenjk.salviamiltiorrhiza-derivedmirnassuppressvascularremodelingthroughregulatingotud7b/klf4/nmhcⅱaaxis[j].theranostics.2020;10(17):7787-7811.
[0019]
[15]qiny,zhengb,yanggs,yanghj,zhouj,yangz,zhangxh,zhaohy,shijh,wenjk.salviamiltiorrhiza-derivedsal-mir-58inducesautophagyandattenuatesinflammationinvascularsmoothmusclecells[j].molthernucleicacids.2020;21:492-511.
[0020]
[16]murphymp.howmitochondriaproducereactiveoxygenspecies[j].biochemj.2009;417(1):1-13.
技术实现要素:
[0021]
本发明的目的是提供gen-mir-1在制备治疗抑制心肌纤维化的药物中的应用。
[0022]
本发明的另一目的是提供含有gen-mir-1的组合物。
[0023]
本发明的目的可通过以下技术方案实现:
[0024]
seqidno.1所示的gen-mir-1在制备治疗抑制心肌纤维化的药物中的应用。
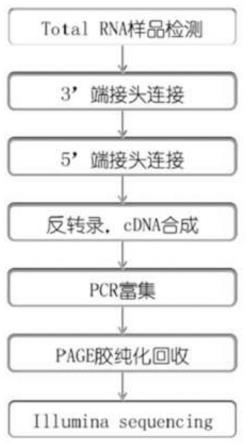
[0025]
gen-mir-1的前体序列如seqidno.2所示,成熟体序列如seqidno.1所示,二级建构如图2所示。
[0026]
作为本发明的一种优选,所述的gen-mir-1在其3'末端核苷酸上存在2
’‑
o-methylated结构的修饰。
[0027]
作为本发明的一种优选,所述的gen-mir-1来源于尖叶假龙胆(gentianellaacuta)或直接化学合成。
[0028]
作为本发明的一种优选,,所述的gen-mir-1在制备抑制心肌纤维化过程中氧化应激和炎性反应的药物中的应用。
[0029]
含有seqidno.1所示的gen-mir-1的组合物,所述的gen-mir-1在其3'末端核苷酸上存在2
’‑
o-methylated结构的修饰。
[0030]
作为本发明的一种优选,所述的组合物还含有药用辅料。
[0031]
作为本发明的一种优选,所述的组合物被制备成片剂、胶囊剂或注射剂。
[0032]
有益效果:
[0033]
本发明结合对心肌纤维化历经数十年的实验研究,经对尖叶假龙胆进行二代高通量测序、将尖叶假龙胆的有效成分单一化并合成、体外实验抗氧化应激、抑制炎症、细胞迁移机制的研究、动物模型的建立等,发现尖叶假龙胆来源的gen-mir-1在抑制心肌纤维化过程中发挥抑制氧化应激和炎性反应的作用。这种发现突破了现有技术的教导与启示,是本领域技术人员不经过创造性劳动就得不出来的,具有突出的实质性特点和显著的进步。为了进一步研究尖叶假龙胆来源的gen-mir-1在抑制氧化应激和炎性反应过程中发挥抑制心肌纤维化的作用机制。本发明采用了生物化学、分子生物学、组织病理学等方法,研制出
具有抑制氧化应激和炎性反应作用的gen-mir-1,为临床治疗心血管重塑提供一种高效、低毒的新型药物奠定基础,为研发中药治疗心血管重塑提供体内药效学及作用机制及理论实验依据,为科技成果转化创造条件。
附图说明
[0034]
图1.尖叶假龙胆rna建库测序流程
[0035]
图2.尖叶假龙胆来源gen-mir-1二级结构
[0036]
图3.尖叶假龙胆mirna生物信息分析流程
[0037]
图4.尖叶假龙胆mirna kegg pathway富集分析(a)和kog富集分析(b)
[0038]
图5.尖叶假龙胆mirnas分类
[0039]
图6.尖叶假龙胆特有的mirnas表达丰度
[0040]
图7.gen-mir-1在小鼠组织中的表达水平
[0041]
图8.gen-mir-1在小鼠心肌纤维化细胞模型中的实验结果
[0042]
a.western blot检测gen-mir-1对tgf-β1诱导的心肌纤维化细胞模型中 collagen i、collagenⅲ、α-sma、il-1β、tnf-α、il-6蛋白表达的影响
[0043]
b.划痕实验检测gen-mir-1对tgf-β1诱导的成纤维细胞模型迁移的影响
[0044]
图9.gen-mir-1小鼠体内实验结果
[0045]
a.gen-mir-1对iso诱导的心肌纤维化小鼠心电图变化的影响
[0046]
b.gen-mir-1对iso诱导的心肌纤维化小鼠不同组左心室组织masson和he染色的代表性图像
[0047]
c.western blot检测gen-mir-1对iso诱导的心肌纤维化小鼠不同组心脏组织中tβr i、tβrⅱ、collagen iii、il-1β、il-6的蛋白表达的影响
[0048]
d.免疫组化检测gen-mir-1对iso诱导的心肌纤维化小鼠不同组心脏组织中collagen i、collagen iii、il-1β、il-6、tnf-α表达的影响
[0049]
e.荧光观察gen-mir-1对iso诱导的心肌纤维化小鼠不同组心脏组织中ros的产生情况
[0050]
图10尖叶假龙胆来源的gen-mir-1在cf中稳定表达
具体实施方式
[0051]
实施例1
[0052]
为了鉴定尖叶假龙胆中的mirnas,我们利用高通量二代测序手段,对尖叶假龙胆中可能存在的mirnas进行序列分析,在建立文库前,我们首先对中药尖叶假龙胆的rna进行提取,质量合格后开始建立文库并测序(图1)。测序结果显示,共筛选出了111种mirnas(如图5),其中既有与其它植物同源的mirnas,又存在6种在尖叶假龙胆中特异存在的mirnas。
[0053]
6种mirna,具有如下序列:
[0054]
name
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
sequence
[0055]
gen-mir-1
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
uggaaucgaaaaucucucuug
[0056]
gen-mir-2
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
gcuuugucagagauccugagc
[0057]
gen-mir-3
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
uuaugcagccaaucauguaaag
[0058]
gen-mir-4
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
uuggacugaagggagcucccuuc
[0059]
gen-mir-5
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
guagaucguaugguagaaaga
[0060]
gen-mir-8
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
cggcaagucgucuuuggcu
[0061]
上述所提供mirna均在其3'末端核苷酸上存在2
’‑
o-methylated结构的修饰。
[0062]
进一步通过对尖叶假龙胆中mirnas的功能进行了kegg pathway和kog富集分析,kegg和kgo信号通路富集分析显示g.acuta来源的mirna在内质网应激、自噬调节以及糖脂代谢过程中发挥作用(如图4)。从中选取了与心血管功能相关的几种mirnas进行后续验证,我们提取尖叶假龙胆rna进行qrt-pcr检测,结果显示在尖叶假龙胆中能检测到尖叶假龙胆来源的mirnas的表达,且以尖叶假龙胆特异性的gen-mir-1表达相对较高(图6)。我们决定将这种mirnas作为研究对象。
[0063]
由于植物来源的mirnas在其3’末端核苷酸上存在2
′‑
o-methylated结构的修饰,这使得它们对高碘酸盐具有抗性。相反,具有游离2'和3'羟基的哺乳动物mirnas对高碘酸盐敏感。我们进一步还提取了尖叶假龙胆中的mirna,并给予高碘酸钠(氧化剂)处理,进行qrt-pcr检测,结果显示尖叶假龙胆特异性的gen-mir-1仍有表达。我们进一步验证,尖叶假龙胆来源的mirnas能否在cf 中稳定表达,我们对gen-mir-1进行了制备,并在各自的3’末端核苷酸上添加了2
′‑
o-methylated结构的修饰,分别转染cf,提取rna并给予高碘酸钠处理后,进行qrt-pcr检测。结果表明,尖叶假龙胆来源的gen-mir-1不被高碘酸钠氧化降解,可以在小鼠cf中稳定存在(图10)。
[0064]
涉及的实验步骤如下:
[0065]
1、植物总rna的提取
[0066]
1)将干燥后的尖叶假龙胆的全植株进行液氮研磨,保证无块状组织,取100mg 粉末置于2ml无菌离心管中,添加500μl buffer rcl/β-巯基乙醇,(样品解冻之前加入β-巯基乙醇),快速混匀;
[0067]
2)55℃水浴1-3min,室温,14,000g离心5min;
[0068]
3)吸取上清液(大概可获得450μl),加入至含有2ml收集管的gdna filter colum中,室温14,000g离心2min;
[0069]
4)加入等倍体积的buffer rcb于收集管中,并上下颠倒混匀5-10次;
[0070]
5)将从(4)中得到的全部混合液包括沉淀置于hibind rna mini colum,加入一个新的2ml收集管,室温10,000离心1min,除去流动相,并把柱子放回收集管中;
[0071]
6)加入400μl rwc wash buffer置于柱子中,室温10,000g离心1min除去流动相,柱子放回收集管中;此时,可选择用dnaaseⅰ处理;
[0072]
7)把柱子放在一个新的2ml收集管中,加入500μl rna wash bufferⅱ,室温10,000g离心1min,除去流动相,并把柱子放回收集管中;
[0073]
8)重复(7),把柱子放回收集管中,10,000g离心2min,离心干燥;
[0074]
9)将离心柱置于新的1.5ml离心管中,在离心柱中加入30-50μl depc水,室温静置2min后,全速(≥13,000g)离心1min,将流出液收集,-80℃保存;
[0075]
2、尖叶假龙胆rna建库测序流程
[0076]
从rna样品到最终数据获得,样品检测、建库、测序每一个环节都会对数据质量和数量产生影响,而数据质量又会直接影响后续信息分析的结果;为了从源头上保证测序数
据的准确性、可靠性,对样品检测、建库、测序每一个步骤都严格把控,确保高质量数据的产出;流程图如图1。
[0077]
2.1 total rna样品检测
[0078]
对rna样品的检测主要包括4种方法:
[0079]
(1)琼脂糖凝胶电泳分析rna降解程度以及是否有污染;
[0080]
(2)nanodrop检测rna的纯度(od260/280比值);
[0081]
(3)qubit对rna浓度进行精确定量;
[0082]
(4)agilent 2100精确检测rna的完整性;
[0083]
2.2文库构建
[0084]
样品检测合格后,使用small rna sample pre kit构建文库,利用small rna 的3’及5’端特殊结构(5’端有完整的磷酸基团,3’端有羟基),以total rna 为起始样品,直接将small rna两端加上接头,然后反转录合成cdna;随后经过pcr扩增,page胶电泳分离目标dna片段,切胶回收得到的即为cdna文库;构建原理图如附图1。
[0085]
2.3库检
[0086]
文库构建完成后,先使用qubit2.0进行初步定量,稀释文库至1ng/μl,随后使用agilent 2100对文库的insert size进行检测,insert size符合预期后,使用q-pcr方法对文库的有效浓度进行准确定量(文库有效浓度>2nm),以保证文库质量;
[0087]
2.4上机测序
[0088]
库检合格后,把不同文库按照有效浓度及目标下机数据量的需求pooling 后进行hiseq/miseq测序获取尖叶假龙胆中存在的植物源性的mirna;
[0089]
3、植物mirna的提取
[0090]
1)将干燥的尖叶假龙胆植株组织进行液氮研磨,保证无块状组织,取50-100 mg粉末置于2ml无菌离心管中,添加700μl lysis mixture,最高速度涡旋 30s,充分混匀样品;
[0091]
2)55℃水浴3min,室温,12,000g离心5min;
[0092]
3)吸取上清液,加入至含有2ml收集管的gdna removel colum中,室温12,000 g离心2min;
[0093]
4)将收集管液体转移至新的2ml离心管中,加入1.1倍体积的无水乙醇涡旋 20s,充分混匀;12,000g离心1min,弃掉流动相;
[0094]
5)将从(4)中得到的700μl混合液置于microelute rna mini colum,加入一个新的2ml收集管,室温12,000离心1min,除去流动相,并把柱子放回收集管中;
[0095]
6)重复(5)直至所有液体均被转移;
[0096]
7)加入500μl无水乙醇至microelute rna mini colum,室温12,000g离心 1min,除去流动相;
[0097]
8)加入500μl xd binding buffer到microelute rna mini colum,室温12,000 g离心1min,除去流动相;
[0098]
9)加入750μl rna wash bufferⅱ,室温10,000g离心1min,除去流动相,并把柱子放回收集管中;重复一次;
[0099]
10)最大转速离心(≥12,000g)2min,离心干燥
[0100]
11)将离心柱置于新的1.5ml离心管中,在离心柱中加入30-50μl depc水,室温静
置5min后,全速(≥12,000g)离心1min,将流出液收集,-80℃保存;
[0101]
4、高碘酸钠消化处理rna
[0102]
(1)将提取的rna取5μl加入95μl 10mm naio3在0℃避光孵育40min;
[0103]
(2)加入1ml无水乙醇及1μl糖原在冰上静置20min;
[0104]
(3)12,000g,15min,4℃离心后弃上清;
[0105]
(4)加入1ml无水乙醇12,000g,15min,4℃离心弃上清;
[0106]
(5)加入1ml 75%乙醇12,000g,15min,4℃离心弃上清;
[0107]
(6)室温静置5min,用ddh2o溶解后进行定量;
[0108]
5、mirna的逆转录
[0109]
5.1连接反应体系:
[0110][0111]
将上述反应体系在pcr仪中按照如下条件进行反转录反应:
[0112][0113]
5.2反转录反应体系:
[0114][0115]
4μl连接产物加入反转录体系中(加入前在室温平衡2min)
[0116][0117]
6、mirna的半定量检测
[0118]
pcr反应体系:
[0119]
20μl的cdna加入200μlrnase-free h2o
[0120][0121][0122][0123]
name
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
primers
[0124]
gen-mir-1
ꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀꢀ
cgtggaatcgaaaatctctcttg
[0125]
7、mirna的测序
[0126]
将扩增的pcr产物进行测序确定为gen-mir-1的序列一致。
[0127]
gen-mir-1序列信息:
[0128]
precursor(前体):
[0129]
uggaaucgaaaaucucucuugcauugucuuuguauauuaccuuccugagcccuggugaaaauaucugaa cuaaucgaugcccucuacaaggaugggaggauagaagaugcaaacuauuuaguuuagaccaugacccag gaaggu
guuaguuaagauauuuucacuuuuaguuguuuugauggguguguauguguugcguggucaaaa gacaaagcaagagagauuucugauuccaug(seq id no.2)
[0130]
mature(成熟体):
[0131]
uggaaucgaaaaucucucuug(seq id no.1)
[0132]
二级结构:
[0133]
如图2。
[0134]
实施例2
[0135]
我们在体外培养小鼠心肌成纤维细胞(cf),制备了3'末端加有2'-o-甲基化的gen-mir-1agomir转染cf,明确gen-mir-1可以抑制引起的心肌纤维化和转化生长因子-β1(transforming growth factor-β1,tgf-β1)诱导的对cf 氧化应激和炎症反应的分子机制。
[0136]
心肌梗死后引起心肌组织无菌性炎性和氧化应激反应,激活cfs进而诱导mf。大量研究证实,氧化应激和炎症反应是mf的核心事件。为了明确gen-mir-1对cf 活化的过程中的作用,我们将体外合成的g.acuta来源gen-mir-1转染cfs,实验结果显示,gen-mir-1能够抑制tgf-β1诱导的cfs活化标志物collagen i、 collagenⅲ、α-sma的表达水平(图8a)以及il-1β、tnf-α、il-6等炎症因子的表达水平和cfs的迁移(如图8b)。研究结果表明,gen-mir-1通过靶抑制了 tgf-β1诱导的cfs活化与炎症反应。
[0137]
实施例3体内试验
[0138]
为了明确g.acuta来源的gen-mir-1是否可以跨物种进入体内发挥调控作用,c57小鼠灌胃给药gen-mir-1agomir,每天1次,1od/次,连续七天。摘眼球取血、处死后取心脏和血管标本,用试剂盒提取血液、组织中的mirna, rt-pcr检测gen-mir-1在血和组织中的水平。结果显示,给小鼠灌胃gen-mir-1,在小鼠血液、血管和心脏组织中均可检测到gen-mir-1(图7),表明gen-mir-1 可以跨物种进入小鼠体内。
[0139]
选用c57bl/6小鼠40只(20g左右的雄性小鼠),适应性喂养一周后,随机分为4组即:
①
mir-ctl+生理盐水组、
②
mir-ctl+iso组、
③
生理盐水 +gen-mir-1组、
④
iso+gen-mir-1组,每组10只。组
②
和组
④
每24h皮下注射 iso(5mg/kg),同时采用灌胃的方式给与小鼠mir-ctl agomir或gen-mir-1agomir(1od/只/天),持续7d。组
①
和组
③
每24h皮下注射生理盐水(5mg/kg),同时采用灌胃的方式给与小鼠mir-ctl agomir或gen-mir-1 agomir(1od/只/ 天),持续7d。心电图检测观察心脏生物学的变化。动物处死后取心脏标本,he和masson染色观察心脏组织形态学、胶原沉积的变化,western blot检测心脏组织中tgf-β受体i(tβr i)、tgf-β受体ⅱ(tβrⅱ)、collagen iii、白介素1β(il-1β)和白介素6(il-6)的表达水平,免疫组化检测心脏组织中collagen i、collagen iii、il-1β、il-6、tnf-α的表达水平。
[0140]
心电图结果显示,gen-mir-1可明显改善iso诱导的st段降低,改善小鼠心肌缺血(图9a);he和masson染色结果表明,gen-mir-1可显著减低心肌梗死的范围,减少梗死区胶原纤维沉积(图9b);western blot检测显示,gen-mir-1 可抑制tβr i、tβrⅱ、collagen iii、il-1β、il-6的蛋白表达水平(图 9c)。免疫组化检测显示,gen-mir-1可抑制collagen i、collagen iii、il-1β、 il-6、tnf-α的表达水平(图9d)上述结果表明,g.acuta来源的gen-mir-1 可以跨物种抑制iso诱导的小鼠心肌纤维化和炎症反应。
[0141]
tgf-β1与氧化应激密切相关:tgf-β1可增加ros的产生,导致氧化还原失衡;反过来,ros又可激活并介导tgf-β1的许多促纤维化作用,二者相互调节形成恶性循环,参与纤维化疾病的病理过程。利用二氢乙锭(dihydroethidium, dhe)对组织中产生的ros进行检测,结果显示,gen-mir-1可抑制ros的产生(图 9e)。据此提示,gen-mir-1在调控tgf-β1信号的同时也影响了ros的产生。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1