活化T细胞与阻断性抗体联合制备抗肿瘤药物的用途及抗肿瘤药物的制作方法
活化t细胞与阻断性抗体联合制备抗肿瘤药物的用途及抗肿瘤药物
技术领域
1.本发明涉及肿瘤免疫治疗技术领域,具体涉及活化t细胞与阻断性抗体联合制备抗肿瘤药物的用途及抗肿瘤药物。
背景技术:
2.使用t细胞进行肿瘤免疫治疗在临床上已取得突破性进展,主要治疗思路是通过基因工程技术,将t细胞激活,并装上定位导航装置car(肿瘤嵌合抗原受体),成为car-t细胞,这种t细胞利用其“定位导航装置”car,专门识别体内肿瘤细胞,并通过直接免疫杀伤及释放大量的多种效应因子,进而杀灭肿瘤细胞,达到治疗恶性肿瘤的目的。但该技术难度大、实施成本高,且目前只对b细胞白血病有效。
技术实现要素:
3.本发明的目的是针对现有技术中的上述问题,提供一种活化t细胞与阻断性抗体联合制备抗肿瘤药物的用途及抗肿瘤药物。
4.本发明技术方案详述如下:第一方面,本发明提供了活化t细胞与阻断性抗体联合制备抗肿瘤药物的用途,所述活化t细胞,是从患者自身组织或体液中分离细胞,先进行抗人cd3抗体分选得到cd3
+
t细胞,再经抗人cd3抗体、抗人cd28抗体和人il-2刺激活化获得;所述阻断性抗体是抗人癌细胞产生的唾液酸化igg(sia-igg,或sig)的抗体、抗人siglec-7抗体、抗人siglec-9抗体、抗人siglec-10抗体及抗人pd-1抗体中的任意一种或几种。
5.可选或优选的,上述用途中,所述组织为癌组织,所述体液为外周血、胸水或腹水。其中由于胸水和腹水相较于癌组织和外周血量更大、对患者来说属于废物获取更容易,因此是更为适合临床应用的体液材料。
6.可选或优选的,上述用途中,所述体液为胸水或腹水时,活化t细胞的制备方法如下:患者胸水或腹水经离心得细胞沉淀,细胞沉淀经pbs清洗后,用密度梯度离心法分离获得初级体液细胞,初级体液细胞用人淋巴细胞分离液进行离心分离获得体液细胞;体液细胞用抗人cd3抗体避光孵育,然后用pbs洗涤一次并重悬于含1%(v/v)fbs的pbs中,用流式分选仪分选出cd3
+
t细胞;cd3
+
t细胞培养体系中加入抗人cd3抗体、抗人cd28抗体和人il-2刺激诱导培养,获得活化t细胞。
7.其中,所述的初级体液细胞、体液细胞仅用于描述目的,代表经过密度梯度离心后获得的细胞、用人淋巴细胞分离液分离后获得的细胞,并非具有医学或者生物学特定含义的词。pbs即磷酸盐缓冲液(phosphate buffered saline),fbs即胎牛血清(fetal bovine serum)。
8.可选或优选的,上述用途中,所述分选得到的cd3
+
t细胞,是cd3
+
7aad-的t细胞。7aad是一种经典的核酸标记染料,在流式细胞术中能够用于标记死细胞,上述cd3
+
7aad-的t细胞,是经7aad染色后呈阴性(未被染色)的活t细胞。
9.可选或优选的,上述用途中,所述抗人癌细胞产生的sia-igg的抗体为rp215或抗sia-igg人源化抗体,所述抗sia-igg人源化抗体的重链可变区的氨基酸序列如seq id no.1所示,轻链可变区的氨基酸序列如seq id no.2所示。
10.rp215是gregory lee教授从卵巢癌细胞株oc-3-vgh中筛选出来的小鼠抗人单克隆抗体,以往认为其对应的抗原是ca215,该抗原包含了人癌细胞表达的igg,还包含了其他糖蛋白。本发明的发明人前期研究证实rp215的识别抗原实际为人癌细胞表达的ch1区域162位点具有独特n糖基化修饰且糖链末端发生高唾液酸化修饰的sia-igg。rp215可通过请求赠与或购买的方式获得。
11.第二方面,本发明还提供了一种抗肿瘤药物,其包括活化t细胞和阻断性抗体,所述活化t细胞,是从患者自身组织或体液中分离细胞,先进行抗人cd3抗体分选得到cd3
+
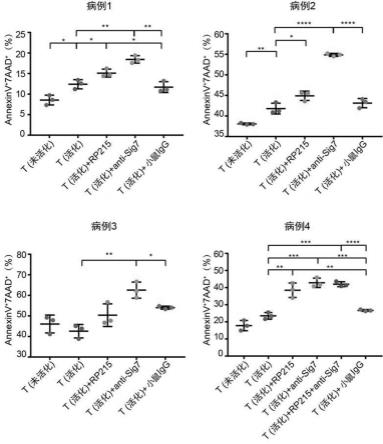
t细胞,再经抗人cd3抗体、抗cd28抗体和人il-2刺激活化获得;所述阻断性抗体是抗人癌细胞产生的唾液酸化igg(sia-igg)的抗体、抗人siglec-7抗体、抗人siglec-9抗体、抗人siglec-10抗体及抗人pd-1抗体中的任意一种或几种。
12.可选或优选的,上述药物中,所述组织为癌组织,所述体液为外周血、胸水或腹水。
13.可选或优选的,上述药物中,所述组织为胸水或腹水时,活化t细胞的制备方法如下:患者胸水或腹水经离心得细胞沉淀,细胞沉淀经pbs清洗后,用密度梯度离心法分离获得初级体液细胞,初级体液细胞用人淋巴细胞分离液进行离心分离获得体液细胞;体液细胞用抗人cd3抗体避光孵育,然后用pbs洗涤一次并重悬于含1%(v/v)fbs的pbs中,用流式分选仪分选出cd3
+
t细胞;cd3
+
t细胞中加入抗人cd3抗体、抗cd28抗体和人il-2刺激诱导培养,获得活化t细胞。
14.可选或优选的,上述药物中,所述抗人癌细胞产生的唾液酸化igg的抗体为rp215或抗sia-igg人源化抗体,所述抗sia-igg人源化抗体的重链可变区的氨基酸序列如seq id no.1所示,轻链可变区的氨基酸序列如seq id no.2所示。
15.术语解释:cd3:分化簇3,为一种跨膜蛋白,其跨膜区通过盐桥与tcr(t cell receptor,t细胞抗原受体)两条肽链的跨膜区连接,形成tcr-cd3复合体,共同参与t细胞对抗原的识别。抗人cd3抗体能够特异性识别并结合cd3,是一种t细胞抗原受体。
16.cd28:是白细胞表面分化抗原28。抗人cd28抗体能够特异性识别并结合cd28,是一种t细胞抗原受体。
17.人癌细胞产生的唾液酸化igg,即sia-igg、sialylated-igg,是恒定区ch1区域162位点具有独特n糖基化修饰且糖链末端发生高唾液酸化修饰的igg,表达于癌细胞表面。我们研究发现,该igg可与肿瘤浸润的t细胞表面singles结合,导致t细胞活化及增殖受到抑制,t细胞失去杀伤肿瘤能力。抗sia-igg抗体可阻断sia-igg对siglec下游信号的增殖抑制作用。
18.siglec:sialic acid binding ig-like lectins,唾液酸结合型ig样凝集素,在肿瘤浸润的t细胞表面高表达,其配体是包含唾液酸的蛋白。siglec是单次跨膜蛋白,由包含唾液酸结合位点的胞外段、跨膜区和传递信号的胞内段组成。目前发现 15 种人源siglec分子,siglec家族分为两类:一类是序列保守的siglecs,包括唾液酸黏附素(siglec-1)、cd22(siglec-2)、cd33(siglec-3)、mag(siglec-4)和 siglec-15;另一类是与 cd33 相关的序列可变的 siglecs。siglec-7、siglec-9及siglec-10常常组成性表达在肿瘤患者的t细胞表面,与肿瘤细胞表面的sia-igg上唾液酸结合后获得增殖抑制信号,导致其无法杀伤肿瘤细胞。单用抗siglec-7抗体、抗siglec-9抗体及抗siglec-10抗体或上述3种抗siglec联用能够阻断sia-igg与siglec结合,从而阻断sia-igg对siglec下游信号的抑制作用,激活t细胞的杀伤肿瘤效应。
19.pd-1:programmed death 1,程序性死亡受体1,也是肿瘤患者t细胞表面的抑制分子(免疫检查点分子),诱导t细胞增殖抑制效应,抗pd-1抗体能够阻断pd-1的增殖抑制信号,增强t细胞杀伤肿瘤能力。
20.与现有技术相比,本发明具有如下有益效果:cd3分子是t细胞特有的分子标志物,本发明中使用抗人cd3抗体标记t细胞,进而用流式细胞术从血液、胸水、腹水等中分选纯化t细胞,由于t细胞杀伤肿瘤时需要t细胞由非活化状态转为活化状态,而抗人cd3抗体可以诱导t细胞的第一活化信号,抗人cd28抗体可以诱导t细胞的第二活化信号,il-2可以通过结合t细胞表面其特异性受体使t细胞获得活化的第三信号。当然,活化t细胞并非只能使用上述三种物质,其他抗原和细胞因子例如il-4、il-7、il-15、il-21、il-18、ifn-γ、tnf(肿瘤坏死因子)、gm-csf(粒细胞巨噬细胞集落刺激因子)、cd70、glycam-1、cd34、madcam-1、psgl-1等等虽然也能够启动t细胞,从而促进t细胞的活化、成熟和/或增殖,本发明经过分析及大量实验发现,使用抗人cd3抗体、抗人cd28抗体和il-2三者进行组合刺激诱导是使t细胞活化并具备抗肿瘤功能且成本最低适合临床应用的一种组合方式。
21.本发明通过将特定方式刺激活化获得的活化t细胞与特定阻断性抗体进行联合,二者能够产生协同作用,更加高效地促进t细胞杀伤效应,诱导肿瘤细胞凋亡,提升治疗效果。且活化t细胞能够从患者自身的血液及胸水、腹水等废物中获得,分离和活化过程操作简单、对操作人员的技术要求低、成本低廉,非常适合临床上大范围推广。
附图说明
22.图1为实施例1中用淋巴细胞分离液分离单个核细胞的分层示意图。
23.图2为实施例1中用正常外周血分离的t细胞进行诱导活化后不同时间使用流式分别检测cd4
+
t细胞,cd8
+
t细胞表达表达cd25、siglec-7和tim-3(免疫检查点分子)的比例。
24.图3为实施例1中从结肠癌分选的t细胞用抗cd3及抗cd28抗体和il-2活化后cd4
+
t细胞及cd8
+
t细胞表达cd25(t细胞活化标志分子)、siglec-7及t细胞免疫检查点分子pd-1、lag-3及tim-3的比例。
25.图4为实施例1从卵巢癌腹水分选的t细胞用抗cd3及抗cd28抗体和il-2活化后t细胞表达siglec-7、siglec-10及t细胞免疫检查点分子pd-1 及tim-3表达频率及水平。a为腹水中分离出的t细胞加入不同试剂进行96小时孵育后t细胞增殖状态的照片,左图加入pbs,
右图加入抗cd3及抗cd28抗体和il-2;b为96小时后 cd4
+
t细胞和cd8
+
t细胞中cd25(t细胞活化的标志分子)、siglec-7、siglec-10、pd-1、tim-3的表达百分比,pbs表示未活化t细胞,αcd3/cn28+il-2表示活化t细胞。计量结果以平均值
±
标准差表示。**p<0.01,***p<0.001,**** p<0.0001;ns,not significant (p>0.05)。
26.图5为实施例1从卵巢癌腹水中分选得到siglec-7
+
与siglec-7-t细胞后细胞活化能力及表达siglec-9、siglec-10、pd-1及lag-3的差异,a为每孔细胞增长的倍数,b、c、d、e分别为流式检测siglec-7
+
与siglec-7-t细胞活化后表达pd-1、lag-3、siglec-9、以及siglec-10的比例。计量结果以平均值
±
标准差表示。**p<0.01,**p<0.001。
27.图6为实施例2检测卵巢癌腹水中肿瘤细胞表达sia-igg情况的实验结果,其中a为使用rp215对卵巢癌腹水肿瘤细胞进行流式染色,圈门显示rp215阳性的细胞群及比例;b为使用rp215对卵巢癌腹水甩片进行免疫荧光染色的结果(绿光)。
28.图7为实施例2流式分析活化的t细胞与同一个体来源的卵巢癌细胞共培养体系中加入活化或未活化t细胞后epcam
+
肿瘤细胞的凋亡情况。凋亡的肿瘤细胞为epcam
+ annexinv
+ 7aad
+
染色。
29.图8为实施例2从病例1-4卵巢癌腹水纯化的t细胞经抗cd3、抗cd28抗体和il-2活化后,与肿瘤细胞的共培养检测肿瘤细胞凋亡的比例。分别在活化t细胞培养体系中加入同一个体来源的卵巢癌细胞,共培养24小时后流式检测epcam
+
肿瘤细胞中annexinv
+
7aad
+
晚期凋亡细胞的比例数据统计结果;计量结果以平均值
±
标准差表示,*p《0.05,**p<0.01,***p《0.001,****p《0.0001。
30.图9为实施例2从病例5-6卵巢癌腹水纯化的t细胞经抗cd3、抗cd28抗体和il-2活化后与同一个体来源的肿瘤细胞共培养,24小时后流式检测epcam
+
肿瘤细胞中annexinv
+
7aad
+
晚期凋亡细胞的比例数据统计结果;计量结果以平均值
±
标准差表示,*p《0.05,**p<0.01,***p《0.001,****p《0.0001。
31.图10为实施例2的2.4从卵巢癌腹水纯化的肿瘤细胞与未活化t细胞共培养体系中,分别加入不同的抗体后,检测7aad
+
的晚期凋亡肿瘤细胞比例统计结果,其中t细胞为未活化t细胞,ab包括抗人cd3抗体、抗人cd28抗体和重组人il-2,αsig-7为抗siglec-7抗体,αpd-1为抗pd-1抗体,migg为小鼠igg;计量结果以平均值
±
标准差表示,*p《0.05,**p<0.01,***p《0.001,****p《0.0001。
32.图11为实施例3在裸鼠皮下构建卵巢癌pdx模型,成瘤后分别用卵巢癌腹水纯化并活化的t细胞、及该活化的t细胞加阻断siglec-7处理,分析活化t细胞的抗肿瘤作用的实验过程及结果图。a为在裸鼠中建立oac-e卵巢癌肿瘤模型的示意图;b为第26天各组小鼠卵巢癌肿瘤体积比较;c为各组小鼠卵巢癌肿瘤的生长曲线;d为第26天各组小鼠卵巢癌肿瘤的体积与质量的统计比较;e为第26天各组小鼠卵巢癌肿瘤中cd4
+
t细胞、cd8
+
t细胞的百分比。计量结果以平均值
±
标准差表示,*p《0.05,**p<0.01,ns,not significant (p>0.05)。f是对各组小鼠卵巢癌肿瘤进行he染色的结果,标尺:50μm。
33.图12为实施例3 在nod-scid小鼠皮下构建卵巢癌pdx模型,分析阻断sia-igg/siglec-7增强t细胞的抗肿瘤作用实验过程及结果图。小鼠成瘤后分为5组,分别是pbs组(对照组)、单纯活化t细胞组、单加rp215组、活化t细胞加sia-igg及活化t细胞加抗siglec-7抗体组。a为在nod-scid小鼠中建立oac-e卵巢癌肿瘤模型的示意图;b为第29天各组小鼠
卵巢癌肿瘤体积比较;c为各组小鼠卵巢癌肿瘤的生长曲线;d为第29天各组小鼠卵巢癌肿瘤的体积与质量的统计比较;e为第29天各组小鼠卵巢癌肿瘤中cd4
+
t细胞、cd8
+
t细胞的百分比。计量结果以平均值
±
标准差表示,*p《0.05,**p<0.01,ns,not significant (p>0.05);f为对各组小鼠卵巢癌肿瘤进行he染色的结果。标尺:50μm。
具体实施方式
34.下面结合具体实施例对本发明的技术方案进行详细解释和说明,以使本领域技术人员能够更好地理解本发明并予以实施。除特殊说明外,实施例中所用的仪器、试剂、生物材料等均为本领域常规,可通过商业途径购买或根据现有知识配置。
35.实施例1 制备活化t细胞1.1淋巴细胞的分离与培养1.1.1 人外周血单个核细胞的分离采用密度梯度离心法从正常人外周血中分离单个核细胞(peripheral blood mononuclear cell,pbmc),提前将人淋巴细胞分离液平衡至室温,铺4 ml淋巴细胞分离液至15 ml离心管底部;将4 ml血样离心后弃去血浆,血细胞沉淀用pbs(磷酸盐缓冲液,ph 7.2)稀释至8 ml;沿离心管壁将稀释后的血细胞缓慢加到平衡至室温的淋巴细胞分离液上,此步的关键在于不能破坏分离界面;室温,2000 rpm,升速和降速刹车设为0,离心20分钟。离心后,离心管内液体将分为三层,上层为pbs,中层为淋巴细胞分层液,上层和中层之间的白色云雾状细胞层即为pbmc,下层为粒细胞和红细胞,如图1所示。小心吸取pbmc层细胞,用大体积pbs洗一遍之后计数。
36.1.1.2 结直肠癌组织中细胞的分离取得结直肠癌标本后,将组织剪至1mm3大小,使用5ml含hepes buffer的1640培养基,加入100μg/ml胶原酶iv和150u/ml dna酶i,37℃ 180rpm消化30分钟后,用5ml pbs终止消化,将得到的细胞悬液经150目筛网过滤,收集滤过的液体,1600rpm离心5分钟,用pbs洗一遍之后计数。
37.1.1.3 卵巢癌腹水中细胞的分离获得的卵巢癌肿瘤患者的腹水经2000rpm离心,得到细胞沉淀。大体积pbs洗2次后,使用密度梯度离心法分离腹水中的细胞,后续操作步骤与1.1.1中人外周血单个核细胞的分离一致。
38.1.2 t淋巴细胞的分选与活化1.2.1 t淋巴细胞的分选外周血pbmc、结肠癌组织细胞悬液或卵巢癌腹水细胞悬液使用含5%fbs的pbs重悬,在冰上静置30分钟封闭。离心弃上清,加入合适剂量的荧光标记的抗人cd3抗体的抗体,用500 μl 1% fbs 的pbs稀释抗体(5 μl/107细胞),4℃避光孵育30 min;1 ml pbs洗涤一次,重悬于 200~400 μl 1% fbs的pbs中置于冰上避光待用。使用bd facsmelody流式分选仪进行分选,分选前5分钟加入7aad(5 μl/107细胞),分选cd3
+
7aad-的t细胞。
39.1.2.2 t淋巴细胞的活化用无菌4℃预冷的pbs(ph 7.2)将leaf
™ꢀ
purified anti-human cd3(okt3)抗体稀释为3 μg/ml,加入至96 孔板(50 μl/孔)或 48孔板(200 μl/孔)中,4℃静置过夜或37℃
孵育2小时以上。取上述包被好的孔板,弃去孔内液体,用无菌4℃预冷pbs洗涤1次(200 μl/孔)后,将计数好的1.2.2分选的cd3
+
7aad-的t细胞用含有1μg/ml leaf
™ꢀ
purified anti-human cd28(抗人cd28抗体)、1μg/ml抗人cd3抗体和30u/ml 重组人il-2的 rpmi 1640培养基重悬,铺入孔中。96孔板细胞密度为2
×
105/孔,培养基体积为200 μl;48孔板密度为4
×
105/孔,培养基体积为400 μl,培养96小时,获得活化t细胞。
40.同时采用上述相同的方法,将抗人cd3抗体、抗人cd28抗体、il-2对应替换为等浓度等剂量的pbs,获得的t细胞作为未活化t细胞阴性对照。
41.1.2.3 刺激活化检测对上述活化t细胞和未活化t细胞,进行直接免疫荧光标记抗体的流式细胞术检测。
42.流式细胞术检测方法:将待检测的细胞制备为单细胞悬液,pbs(ph 7.2)洗涤一次后,用含有5%(v/v)fbs的pbs,冰上静置30 min进行封闭。离心弃上清,加入合适剂量的荧光标记的不同膜分子的抗体,用50 μl 1% (v/v)fbs 的pbs稀释抗体(0.1~0.3 μl/样品),4℃避光孵育30 min;1 ml pbs洗涤一次,重悬于 200~400 μl 1%(v/v)fbs的pbs中置于冰上避光待用。用bd facscantoplus流式仪或 bd facsverse流式仪检测荧光信号。对于淋巴细胞,每组细胞不超过5
×
106/ml,离心转速为5,000 rpm;肿瘤细胞每组不超过5
×
105/ml,离心转速为3,500 rpm。
43.(1)正常外周血制备的活化t细胞加入抗人cd3抗体、抗人cd28抗体和人il-2进行刺激诱导活化后的不同时间,检测cd4
+
t细胞,cd8
+
t细胞表达cd25、siglec-7和tim-3的比例。
44.结果显示,在未刺激的情况下,外周血中的t细胞表达cd25、siglec-7和tim-3这几种分子的水平都较低。在接受抗体刺激活化后,t细胞表达cd25明显上调,说明刺激活化是成功的,参见图2。在cd4
+
t细胞中,siglec-7和tim-3都出现了幅度较小的上调。而在cd8
+
t细胞中,siglec-7和tim-3上调更加明显,siglec-7在48小时阳性比例可以达到40%以上。
45.(2)结直肠癌组织制备的活化t细胞加入抗人cd3抗体、抗人cd28抗体和人il-2进行刺激诱导活化后的不同时间,检测cd4
+
t细胞,cd8
+
t细胞表达cd25、pd-1、siglec-7、tim-3以及lag-3的比例。
46.结果显示,在结肠癌的cd4
+
t细胞中,未刺激时lag-3、siglec-7有20%左右的阳性表达,其他分子的阳性率较低;而随着细胞被刺激活化,cd25、siglec-7以及 pd-1、lag-3等分子都出现明显上调,但tim-3则上调不明显,参见图3。cd8
+
t细胞则在未刺激的情况下表达高达pd-1(60%)及低频率表达siglec-7(10%左右),其他检测的分子则低于5%;在t细胞活化后,pd-1虽然在24小时出现了轻微下调,但在72小时又恢复到了较高的表达水平,cd25、lag-3和siglec-7均出现明显上调,tim-3也出现轻微上调。肿瘤微环境中的t细胞在静息状态下表达分子的水平与外周血存在很大差异,但在接受抗人cd3抗体、抗人cd28抗体和人il-2刺激并活化后,其siglec-7和其他免疫检查点分子都出现同向的上调。
47.(3)卵巢癌腹水制备的活化t细胞加入抗人cd3抗体、抗人cd28抗体和人il-2进行刺激诱导活化后的不同时间,用流式细胞术检测cd4
+
t细胞,cd8
+
t细胞中cd25、pd-1、siglec-7、siglec-10以及tim-3的百分比。
48.流式细胞术检测过程中加入的荧光标记的抗体为:抗cd25抗体(作为t细胞活化标志)、抗pd-1抗体(检测免疫检查点分子)及抗tim-3抗体(检测免疫检查点分子)、抗siglec-7抗体、抗siglec-10抗体及抗cd4抗体(检测cd4
+
t细胞)和抗cd8抗体(检测cd8
+
t细胞)。
49.诱导活化后96小时的检测结果如图4所示,相比于未活化t细胞,经过诱导活化后,cd4
+
t细胞中活化相关分子cd25,免疫检查点分子pd-1、tim-3以及siglec-7、siglec-10等分子都出现明显上调,其中以siglec-7表达最为明显。在cd8
+
t细胞中,pd-1、siglec-7、siglec-10表达水平略低于cd4
+
t细胞,但相比于未活化t细胞,在活化后也出现类似的上调,tim-3则没有显著的变化。
50.以上实验结果表明,在不同样本来源的t细胞中,siglec-7都在细胞接受刺激活化后出现上调,提示siglec-7与pd-1、lag-3等免疫检查点分子类似,是与t细胞活化相关的分子。
51.1.2.4 siglec-7
+
的t细胞表达高水平的免疫抑制分子为了进一步探究表达siglec-7的t细胞有哪些特点,我们从卵巢癌腹水中分选得到了siglec-7
+
和siglec-7-的t细胞,按每孔5000个铺板,并使用抗人cd3/cd28抗体+人il-2进行活化,48小时后检测两种细胞增殖倍数的区别,以及表达分子的差异。
52.结果参见图5,siglec-7
+
t细胞表现出明显的增殖抑制,其刺激活化后细胞数的增加倍数只有2-3倍,而sigecle-7-的t细胞可以增殖10倍以上(图5中 a)。在活化之后,siglec-7
+
cd8
+
t细胞表达更高水平的免疫检查点分子pd-1和lag-3,提示其活化后的功能处于更加受到抑制的状态(图5的b,c)。同时siglec-7
+
cd8
+
t细胞也表达更多的抑制性siglec分子如siglec-9和siglec-10(图5中d,e)。
53.以上结果说明,表达siglec-7的t细胞处于更加抑制的状态,表现为增殖能力更低、表达免疫抑制分子的水平更高。
54.实施例2 活化t细胞联合阻断性抗体抗肿瘤效果的验证(体外实验)2.1使用卵巢癌腹水细胞建立共培养模型显示活化t细胞可杀伤同一个体的肿瘤细胞根据前期的实验结果,卵巢癌腹水中的t细胞表达较高水平的siglec分子。并且其中含有大量的肿瘤细胞和免疫细胞,使我们能够获得足以用于实验的细胞。因此,我们首先选择卵巢癌腹水,分别分选其中的t细胞及肿瘤细胞,构建体外共培养模型。
55.用抗人epcam抗体从卵巢癌腹水中分选肿瘤细胞,对分选的卵巢癌中的肿瘤细胞进行分析,使用特异性识别sia-igg的抗体-rp215进行流式染色;此外,对腹水肿瘤细胞甩片固定后,使用rp215进行免疫荧光染色,显微镜观察。结果如图6,圈门显示rp215阳性的细胞群及比例,表明腹水中的肿瘤细胞膜上表达高水平的sia-igg(图6中a),免疫荧光染色也证实了sia-igg在卵巢癌腹水肿瘤细胞的细胞膜和细胞质内均有表达(图6中b)。
56.通过流式分选得到卵巢癌腹水中cd3
+
7aad-的t细胞后,在体外加入抗人cd3抗体和抗人cd28抗体以及重组人白细胞介素-2(il-2)进行活化培养,培养进行刺激诱导活化48小时后获得活化t细胞;以不加入两种抗体及il-2刺激诱导的t细胞进行相同时间的培养获得未活化t细胞(作为对照)。
57.再分选得到同一例卵巢癌腹水来源的epcam
+
上皮来源的肿瘤细胞,以t细胞:肿瘤细胞=5:1~3:1的比例向肿瘤细胞中加入t细胞(由于不同比例个体差异较大,每个病例加入
的比例不完全一致)。共培养34小时后,使用annexinv、7aad染色,流式检测epcam
+
肿瘤细胞中annexinv
+
7aad
+
的晚期凋亡细胞比例。
58.检测结果如图7所示,我们发现,相比未活化t细胞,在使用活化t细胞与肿瘤细胞进行共培养后,晚期凋亡的肿瘤细胞群明显增加,说明我们构建的体外共培养模型能够反映t细胞杀伤肿瘤细胞的功能。
59.2.2抗sia-igg抗体、抗siglec-7抗体可以增强活化t细胞对肿瘤细胞的杀伤作用接下来,我们在成功建立的体外共培养模型中检测阻断性抗体:抗sia-igg抗体、抗siglec-7抗体是否能够增强活化t细胞对肿瘤细胞的杀伤作用。
60.使用不同病例的卵巢癌腹水经epcam
+
分选获得的肿瘤细胞,进行活化t细胞与肿瘤细胞的共培养实验,肿瘤细胞与t细胞数量比为4:1。同一病例的共培养细胞分为五组:t(未活化)组:肿瘤细胞加入未活化t细胞,作为阴性对照;t(活化)组:肿瘤细胞加入活化t细胞;t(活化)+ rp215组:肿瘤细胞加入活化t细胞和rp215;t(活化)+anti-sig7组:肿瘤细胞加入活化t细胞和小鼠抗siglec-7抗体;t(活化)+小鼠igg组:肿瘤细胞加入活化t细胞和小鼠igg,作为对照migg组。
61.病例4增加一组:t(活化)+ rp215+ anti-sig7组:肿瘤细胞加入活化t细胞和rp215、小鼠抗人siglec-7抗体。
62.病例1-4未活化t细胞和活化t细胞加入剂量相同,病例1-3中rp215、小鼠抗人siglec-7抗体、小鼠igg三者加入剂量相同,其在整个培养体系中的终浓度均为10μg/ml,病例4中rp215和小鼠抗siglec-7抗体的终浓度各5μg/ml,总计10μg/ml)。共培养24小时后流式检测epcam
+
肿瘤细胞中annexinv
+
7aad
+
晚期凋亡细胞的比例,反映t细胞的杀伤作用。
63.结果如图8,在四例不同的卵巢癌腹水中,相比只加t细胞活化组或加入对照migg组,阻断siglec-7都能更加显著地增强肿瘤细胞的凋亡。同时,使用rp215阻断sia-igg活性也能够增强活化t细胞的杀伤作用,但在不同病例中的效果存在一定差异。同时加入rp215和抗siglec-7的阻断抗体,相比单独阻断并没有更强的作用,推测是由于单独使用一种抗体的阻断作用已经饱和(图8中病例4)。这样的结果初步证明了使用抗sia-igg抗体阻断sia-igg,或使用抗siglec-7抗体阻断siglec-7都能够增强卵巢癌腹水活化t细胞对同一个体肿瘤细胞的杀伤作用。
64.2.3 抗siglec-7抗体、抗siglec-9抗体、抗siglec-10抗体可以增强活化t细胞对肿瘤细胞的杀伤作用在之前的结果中,我们发现了卵巢癌腹水的t细胞也共表达其它几种siglec分子,如siglec-9、siglec-10。因此我们也在共培养实验中探究了阻断其他的siglec分子能否增强t细胞杀伤肿瘤细胞的功能。
65.使用不同病例的卵巢癌腹水经epcam
+
分选获得的肿瘤细胞,进行活化t细胞与肿瘤细胞的共培养实验。同一病例的共培养细胞分为六组:t(未活化)组:肿瘤细胞加入未活化t细胞;t(活化)组:肿瘤细胞加入活化t细胞;t(活化)+anti-sig7组:肿瘤细胞加入活化t细胞和山羊抗siglec-7抗体;
t(活化)+anti-sig9组:肿瘤细胞加入活化t细胞和山羊抗siglec-9抗体;t(活化)+anti-sig10组:肿瘤细胞加入活化t细胞和山羊抗siglec-10抗体;t(活化)+山羊igg组:肿瘤细胞加入活化t细胞和山羊igg。
66.未活化t细胞和活化t细胞加入剂量相同,山羊抗siglec-7抗体、山羊抗siglec-9抗体、山羊抗siglec-10抗体、山羊 igg加入剂量相同(在培养体系中的终浓度均为10μg/ml)。共培养24小时后流式检测epcam
+
肿瘤细胞中annexinv
+
7aad
+
晚期凋亡细胞的比例。
67.实验结果见图9,首先,使用与前文中不同来源的抗siglec-7阻断抗体仍然能够增强活化t细胞的杀伤作用。并且在两例不同的卵巢癌腹水中,我们观察到阻断siglec-9、-10也都能够增强活化t细胞对肿瘤细胞的杀伤作用。以上结果表明,在卵巢癌腹水这一模型中,阻断sia-igg和/或siglec能够增强活化t细胞的抗肿瘤免疫功能。
68.2.4不同阻断性抗体组合与活化t细胞之间协同作用不同用抗人cd3抗体和7aad从卵巢癌腹水中分选得到cd3
+
7aad-的t细胞,为未活化t细胞。
69.用抗人epcam抗体从卵巢癌腹水中分选得到肿瘤细胞。
70.以肿瘤细胞与t细胞4:1的比例进行体外共培养。
71.共培养体系分为八组:肿瘤细胞+t细胞(pbs)组:肿瘤细胞与未活化t细胞共培养,加入pbs(与以下几组中用于稀释加抗体及il-2的pbs体积一致)作为阴性对照;肿瘤细胞+t细胞(ab)组:肿瘤细胞与未活化t细胞共培养,加入含有抗人cd3抗体、抗人cd28抗体,重组人il-2 的pbs;肿瘤细胞+t细胞(ab+rp215)组:肿瘤细胞与未活化t细胞共培养,加入含有抗人cd3抗体、抗人cd28抗体、重组人il-2和rp215的pbs;肿瘤细胞+t细胞(ab+αsig-7)组:肿瘤细胞与未活化t细胞共培养,加入含有抗人cd3抗体、抗人cd28抗体、重组人il-2和抗siglec-7抗体的pbs;肿瘤细胞+t细胞(ab+αpd-1)组:肿瘤细胞与未活化t细胞共培养,加入含有抗人cd3抗体、抗人cd28抗体、重组人il-2和抗pd-1抗体的pbs;肿瘤细胞+t细胞(ab+rp215+αsig-7)组:肿瘤细胞与未活化t细胞共培养,加入含有抗人cd3抗体、抗人cd28抗体、重组人il-2和rp215(终浓度5
ꢀµ
g/ml)抗siglec-7抗体 (终浓度5
ꢀµ
g/ml)的pbs;肿瘤细胞+t细胞(ab+αpd-1+αsig-7)组:肿瘤细胞与未活化t细胞共培养,加入含有抗人cd3抗体、抗人cd28抗体、重组人il-2和抗siglec-7抗体(终浓度5
ꢀµ
g/ml)、抗pd-1抗体(终浓度5
ꢀµ
g/ml)的pbs;肿瘤细胞+t细胞(ab+migg)组:肿瘤细胞与未活化t细胞共培养,加入含有抗人cd3抗体、抗人cd28抗体、重组人il-2和小鼠igg (终浓度10
ꢀµ
g/ml)的pbs,作为同型对照。
72.各组中pbs体系的体积相同,肿瘤细胞数量相同,未活化t细胞数量相同。各组中加入抗人cd3抗体、抗人cd28抗体、重组人il-2的对应终浓度分别为:3
µ
g/ml、1
µ
g/ml、30 u/ml。
73.结果如图10,肿瘤细胞与未活化t细胞共培养体系中单独加入ab(ab即抗人cd3抗体、抗人cd28抗体和重组人il-2)与阴性对照组无显著差别,但在加入ab的情况下再单独加
入rp215或抗siglec-7或抗pd-1抗体都显示明显杀伤效果;在加入ab的情况下再同时加入rp215及siglec-7单抗组显示了更强的杀伤效果;但在加入ab的情况下再同时加入抗siglec-7及抗pd-1单抗相较于同时加rp215和siglec-7单抗未见明显协同作用,但依然显著区别于肿瘤细胞+t细胞(ab)组、肿瘤细胞+t细胞(ab+migg)组和肿瘤细胞+t细胞(pbs)组。
74.实施例3 活化t细胞联合阻断性抗体抗肿瘤效果验证(体内实验)3.1抗siglec-7抗体与活化t细胞联合在小鼠体内能够增强t细胞抗肿瘤作用自卵巢癌患者腹水中分选得到单个核细胞,并单克隆化建系获得卵巢癌细胞系oac-e,进行动物体内实验。
75.选取6~8周龄的nu/nu雌性鼠,提前一周复苏小鼠卵巢癌细胞系oac-e细胞,常规培养传代,在生长对数期将其用胰酶消化,用无菌4℃预冷的pbs(ph 7.2)洗两遍,600 rpm离心5 min,计数。最终以 5
×
107/ml 的细胞浓度重悬于pbs中,每管1 ml分装于数个1.5 ml无菌ep管中待用。在第0天时在nu/nu鼠腋窝皮下接种5
×
106细胞。观察肿瘤生长情况,约第7天可肉眼见到肿瘤块,完成小鼠肿瘤模型建立。
76.使用卵巢癌腹水中分选得到的t细胞与人il-2、抗人cd3抗体、抗人cd28抗体进行体外培养活化,获得活化t细胞。
77.将成瘤的小鼠随机分三组,每组6只:pbs组:注射pbs,每个肿瘤瘤周40μl;t细胞+山羊igg:注射加入了终浓度20μg/ml、剂量为每个肿瘤瘤周40μl的山羊igg的活化t细胞;每个肿瘤瘤周每次注射5
×
105个t细胞;t细胞+anti-siglec-7:注射加入了终浓度20μg/ml、剂量为每个肿瘤瘤周40μl的山羊抗siglec-7抗体的活化t细胞,每个肿瘤瘤周每次注射5
×
105个t细胞。
78.三组小鼠瘤周注射上述药物进行肿瘤治疗,每隔3天瘤周皮下注射1次,共注射8次。每天在体测量小鼠肿瘤生长情况并计算瘤体积,用游标卡尺量取肿瘤长径(a)和短径(b),计算肿瘤体积v=a
×b×
b/2。第26天处死小鼠,取出肿瘤进行测量统计,检测各组小鼠卵巢癌肿瘤中cd4
+
t细胞、cd8
+
t细胞的百分比。
79.结果显示,相比不给予t细胞治疗组(pbs组),活化t细胞治疗能够显著抑制肿瘤生长(图11中b和c)。并且与给予对照igg培养的活化t细胞(t细胞+山羊igg组)相比,使用抗体阻断siglec-7的活化t细胞治疗组(t细胞+抗siglec-7抗体组)肿瘤大小和重量的降低更为显著(图11中d)。
80.接下来,我们对荷瘤小鼠肿瘤微环境中人cd4
+
t和cd8
+
t细胞浸润的比例进行分析,反映了治疗注射的人活化t细胞向肿瘤中浸润的水平。结果显示,加入抗siglec-7抗体阻断后,肿瘤浸润的cd8
+
t细胞比例明显增加,而cd4
+
t细胞比例无明显改变(图11中e)。对肿瘤组织的he染色显示,相比pbs组,使用活化t细胞治疗组肿瘤中坏死更为明显。并且阻断siglec-7治疗组具有更明显的液化灶。以上结果说明阻断性抗体抗siglec-7抗体能够增强活化t细胞的抗肿瘤功能,并主要是通过增加cd8
+
t细胞的浸润水平,阻止肿瘤免疫逃逸。
81.3.2 抗sia-igg抗体与活化t细胞联合在小鼠体内增强t细胞抗肿瘤作用我们在免疫缺陷程度更严重的nod-scid小鼠中进行oac-e成瘤模型构建,构建流程与nu/nu小鼠基本一致,不同之处在于在接种细胞前一天对nod-scid小鼠腋下进行脱毛
处理,nu/nu鼠无需处理。在第0天时在nod-scid鼠腋窝皮下接种3
×
10
6 细胞(注意进针深一些,退针时用镊子夹住10 s左右防止细胞悬液流出)。
82.分别使用rp215阻断sia-igg、抗siglec-7抗体阻断siglec-7培养活化t细胞进行实验。
83.使用卵巢癌腹水中分选得到的t细胞与人il-2、抗人cd3抗体、抗人cd28抗体进行体外培养活化,获得活化t细胞。
84.将成瘤的小鼠随机分五组,每组6只:pbs组:注射pbs,每个肿瘤瘤周40μl;rp215组:注射阻断sia-igg的抗体rp215,终浓度20μg/ml,每个肿瘤瘤周40μl;t细胞+小鼠igg组:注射加入了终浓度20μg/ml、剂量为每个肿瘤瘤周40μl的小鼠igg的活化t细胞,每个肿瘤瘤周每次注射3
×
105个t细胞;t细胞+ rp215组:注射加入了终浓度20μg/ml、剂量为每个肿瘤瘤周40μl 的rp215的活化t细胞,每个肿瘤瘤周每次注射3
×
105个t细胞;t细胞+anti-siglec-7组:注射加入了终浓度20μg/ml、剂量为每个肿瘤瘤周40μl 小鼠抗siglec-7抗体的活化t细胞,每个肿瘤瘤周每次注射3
×
105个t细胞。
85.3.1的裸鼠成瘤的实验结果提示,瘤周注射活化t细胞本身就有抗肿瘤的作用,因此本次实验中减少了活化t细胞的用量和频率,共注射7次,每次每侧3
×
105个细胞(图12中a)。每天在体测量小鼠肿瘤生长情况并计算瘤体积,用游标卡尺量取肿瘤长径(a)和短径(b),计算肿瘤体积v=a
×b×
b/2。第29天处死小鼠,取出肿瘤进行测量统计,检测各组小鼠卵巢癌肿瘤中cd4
+
t细胞、cd8
+
t细胞的百分比。
86.此外,由于本实验室的前期研究表明,阻断sia-igg可以通过阻止其与肿瘤细胞表面的整合素结合,从而起到直接的抗肿瘤作用(参见公开号cn108610414a、cn114031688a的专利)。为了排除rp215直接抑制肿瘤的作用,我们在体内实验时使用相对低浓度(20μg/ml)的rp215进行实验,并设置了只使用rp215不使用活化t细胞的治疗组。
87.同时,本次实验中使用小鼠抗siglec-7的阻断型单克隆抗体进行实验,以小鼠igg培养t细胞作为对照。
88.结果发现,低浓度的rp215并不能显著抑制肿瘤生长,然而同一浓度rp215培养的活化t细胞治疗能够显著抑制肿瘤生长(图12中b、c)。在降低瘤周注射活化t细胞数目与频率后,给予对照igg培养的活化t细胞不再具有显著的抗肿瘤效果。而使用抗体阻断sia-igg或siglec-7的活化t细胞治疗组肿瘤大小和重量都显著降低(图12中d)。
89.我们也对荷瘤小鼠肿瘤中人cd4
+
t和cd8
+
t细胞的浸润水平进行了检测。由于本次活化t细胞用量很少,各组组内差异较大,只有rp215培养活化t细胞治疗组的cd4
+
t细胞浸润水平出现显著上升,而阻断siglec-7培养活化t细胞治疗组的cd8
+
t细胞浸润水平则有增加的趋势。
90.对肿瘤组织的he染色显示,相比pbs组或rp215组,加入t细胞治疗组都有明显的坏死,其中rp215培养t细胞组出现细胞溶解空洞,阻断siglec-7培养t细胞组出现更加明显的肿瘤坏死灶。
91.以上结果说明,使用抗体阻断sia-igg和/或siglec能够增强活化t细胞的抗肿瘤功能,且可以用量更少,具有临床应用前景。
92.本文中应用了具体个例对发明构思进行了详细阐述,以上实施例的说明只是用于帮助理解本发明的核心思想。应当指出,对于本技术领域的普通技术人员来说,在不脱离该发明构思的前提下,所做的任何显而易见的修改、等同替换或其他改进,均应包含在本发明的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1