异硫氰酸苄酯靶向结合蛋白及其应用
1.本发明属于生物与新医药技术领域,具体属于异硫氰酸苄酯靶向结合蛋白及其应用。
背景技术:
2.异硫氰酸苄酯bitc,分子量为149.21g/mol,分子式c8h7ns,是亲油性的,难溶于水,是研究最多的异硫氰酸酯类分子之一,存在于十字花科蔬菜中,如豆瓣菜、卷心菜、花椰菜、羽衣甘蓝和西兰花。前期研究发现异硫氰酸苄酯对标准金黄色葡萄球菌菌株atcc25923和临床上两株mrsa菌株表现出相同的杀菌效果,mic均为10μg/ml。在哺乳动物中,bitc在肝脏中分别通过谷胱甘肽-s-转移酶和谷氨酰转肽酶的降解和结合进行代谢。据报道,62%的bitc以巯基尿酸的形式从尿中排出。除了抗菌、抗真菌和抗炎特性外,bitc在临床上还具有抗癌作用。大量动物实验和流行性病学研究表明,异硫氰酸苄酯具有很强的抑制肿瘤细胞生长作用,对肺癌细胞、肝癌细胞、前列腺癌细胞、食管癌细胞和白血病细胞等均有抑制其生长的作用。美国明尼苏达大学癌症中心开展的一项关于异硫氰酸苄酯对健康吸烟人群肺癌预防作用的ⅱ期临床研究发现异硫氰酸苄酯能降低烟草代谢后的致癌物活性。植物及其天然产物具有抑制癌症的潜力,可以降低患癌症的风险。之前的几项研究表明,每天摄入十字花科蔬菜有助于避免癌症的发生。十字花科蔬菜中存在的itcs具有更高的抗癌特性,可以抑制细胞增殖。
3.金黄色葡萄球菌是一种食源性致病菌,其引起的食品污染可能发生在不同种类的食物资源中,其产生的肠毒素可导致中毒性休克和各种自身免疫性疾病。金黄色葡萄球菌对食品安全和公共健康产生的危害都不容忽视。多重耐药金黄色葡萄球菌污染已成为全球关注的食品安全及公共健康问题,研发新型抗菌剂和探索新型控制技术已刻不容缓。
4.杀菌剂的活性范围比抗生素更广。因此,对革兰氏阴性菌具有显著抗菌活性的抗菌产品可以成为控制该菌特异性感染的有价值的治疗药物来源。其中一些水解产物比传统抗生素更能有效地抑制病原微生物的生长,如异硫氰酸苄酯(benzyl isothiocyanate,bitc)。bitc是异硫氰酸酯(isothiocyanates,itcs)最重要的植物源生物活性成分之一,具有抗菌性。据报道,bitc可以抑制病原体的生长。前期研究发现异硫氰酸苄酯对标准金黄色葡萄球菌菌株atcc25923和临床上两株mrsa菌株(耐甲氧西林菌株)表现出相同的杀菌效果,但靶点作用蛋白不清晰。
技术实现要素:
5.为了解决现有技术中存在的问题,本发明提供了53个异硫氰酸苄酯靶向结合蛋白及其应用,在蛋白组学数据基础上借鉴修饰蛋白组学思维,通过筛选与药物分子具有相同偏移量的蛋白位组点来鉴定bitc处理独有的修饰位点,全面解析出金黄色葡萄球菌中53个异硫氰酸苄酯靶向结合蛋白,共涉及99个作用修饰位点,为bitc靶向抗癌药物或靶向抗菌剂开发应用提供了分子基础。
6.为实现上述目的,本发明提供如下技术方案:
7.定稿后补充
8.与现有技术相比,本发明至少具有以下有益效果:
9.本发明提供了异硫氰酸苄酯靶向结合蛋白及应用,基于蛋白质组学解析bitc对金黄色葡萄球菌的抑制作用,共鉴定出88个差异蛋白,其中下调76个、上调12个。生物信息学分析及蛋白质组学鉴定等揭示多靶点协同作用促进了bitc对金黄色葡萄球菌的杀菌效果。全面解析出金黄色葡萄球菌中53个异硫氰酸苄酯的共价结合蛋白,共涉及99个作用修饰位点,为bitc靶向开发应用提供了分子基础;基于蛋白组学的作用靶点鉴定方法能在复杂生物系统中无差别地鉴定化合物的未知作用靶点,具有高复现性、高准确性等优点,逐渐成为目前发现化合物作用靶点的重要手段,极大促进了活性分子的靶点识别;本发明采用的异硫氰酸苄酯(benzyl isothiocyanate,bitc)是来源于十字花科植物的活性精油,常用于食品防腐保鲜,具有抑菌抗肿瘤活性,具有极大的药物开发价值,符合环保要求。
附图说明
10.图1是csha质谱图;图2是cshb质谱图;图3是lgt质谱图;图4是translation initiation factor if-2质谱图;图5是capg质谱图;图6是cyclic-di-amp质谱图;图7是ldh质谱图;图8是pyk质谱图;图9是ezra质谱图;图10是abc transporter质谱图;图11是aminoglycoside 6'-n-acetyltransferase质谱图;图12是udk质谱图;图13是aur质谱图;图14是hslu质谱图;图15是thre质谱图;图16是phislt orf401-like protein质谱图;图17是phi pv83 orf 19-like protein质谱图;图18是yyci质谱图;图19是aldehyde dehydrogenase b质谱图;图20是lysr substrate binding domain protein质谱图;图21是agcs_2质谱图;图22是dihydrolipoamide acetyltransferase component质谱图;图23是exotoxin质谱图;图24是bshc质谱图;图25是ribosomal protein l7ae质谱图;图26是hemn质谱图;图27是infc质谱图;图28是mene_1质谱图;图29是toxin,beta-grasp domain protein质谱图;图30是cobb质谱图;图31是nade质谱图;图32是dape质谱图;图33是cbix质谱图;图34是ysdc质谱图;图35是gltb_1质谱图;图36是srmb质谱图;图37是argf质谱图;图38是n-acetyltransferase domain-containing protein质谱图;图39是dna-3-methyladenine glycosylase质谱图;图40是mfd质谱图;图41是lipoprotein质谱图;图42是gyra_1质谱图;图43是nadh dehydrogenase质谱图;图44是formate dehydrogenase,alpha subunit质谱图;图45是lysr family transcriptional regulator质谱图;图46是putative membrane protein质谱图;图47是entc质谱图;图48是aroc_3质谱图;图49是uvra_3质谱图;图50是glcb_2质谱图;图51是irta质谱图;图52是superfamily i dna/rna helicase protein质谱图;图53是collagen adhesin质谱图。
具体实施方式
11.下面结合附图和具体实施方式对本发明作进一步的说明。
12.本发明获取bitc抗金黄色葡萄球菌靶点蛋白的方法,具体如下:
13.s1在培养至对数生长期的金黄色葡萄球菌培养液(1x107 cfu/ml)中加入bitc溶液,使其终浓度达到40μg/ml,以不添加bitc的一组作为对照组;
19-like蛋白的ncbi检索码为phx24470.1;18yyci蛋白的ncbi检索码为uks35359.1;19.aldehyde dehydrogenase b蛋白的ncbi检索码为scs45302.1;20.lysr substrate binding domain蛋白的ncbi检索码为kxa39781.1;21.agcs_2蛋白的ncbi检索码为kxa40614.1;22.dihydrolipoamide acetyltransferase component蛋白的ncbi检索码为cag42804.1;23.exotoxin蛋白的ncbi检索码为azl91819.1;24.bshc蛋白的ncbi检索码为azl91176.1;25.ribosomal protein l7ae蛋白的ncbi检索码为oby01910.1;26.hemn蛋白的ncbi检索码为uks36683.1;27.infc蛋白的ncbi检索码为oby01036.1;28.mene_1蛋白的ncbi检索码为eik32941.1;29.toxin,beta-grasp domain蛋白的ncbi检索码为efw31165.1;30.cobb蛋白的ncbi检索码为eju82514.1;31.nade蛋白的ncbi检索码为bba24545.1;32.dape蛋白的ncbi检索码为cpm20293.1;33.cbix蛋白的ncbi检索码为cpl97512.1;34.ysdc蛋白的ncbi检索码为cpl70611.1;35.gltb_1蛋白的ncbi检索码为bar12687.1;36.srmb蛋白的ncbi检索码为kfd31251.1;37.argf蛋白的ncbi检索码为cpl88181.1;38.n-acetyltransferase domain-containing蛋白的ncbi检索码为txo06693.1;39.dna-3-methyladenine glycosylase蛋白的ncbi检索码为efw31974.1;40.mfd蛋白的ncbi检索码为eik25642.1;41lipoprotein蛋白的ncbi检索码为bba23099.1;42.gyra_1蛋白的ncbi检索码为kki64673.1;43.nadh dehydrogenase蛋白的ncbi检索码为oby02024.1;44.formate dehydrogenase,alpha subunit蛋白的ncbi检索码为kxa35290.1;45.lysr family transcriptional regulator蛋白的ncbi检索码为oby02041.1;46.putative membrane蛋白的ncbi检索码为aew65823.1;47.entc蛋白的ncbi检索码为kfd32374.1;48.aroc_3蛋白的ncbi检索码为oby00605.1;49.uvra_3蛋白的ncbi检索码为eik35378.1;50.glcb_2蛋白的ncbi检索码为scs40645.1;51.irta蛋白的ncbi检索码为kfd31171.1;52.superfamily i dna/rnahelicase蛋白的ncbi检索码为ccw20441.1;53.collagen adhesin蛋白的ncbi检索码为aaa20874.1;
21.实施例1
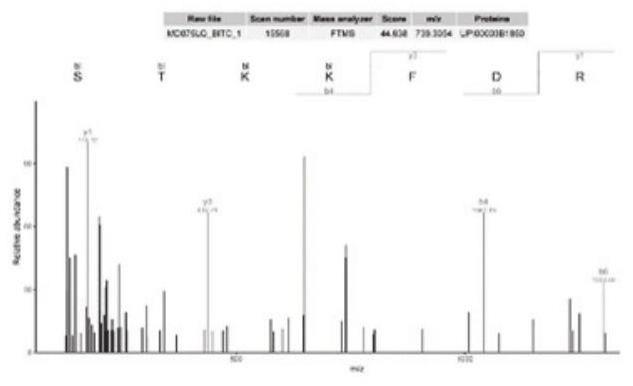
22.csha蛋白可以保护特定的mrna或srnas不被降解,提高金黄色葡萄球菌在毒素诱导下的存活率;cshb是金黄色葡萄球菌脂肪酸稳态所必需的。由图1可知csha蛋白第479位的s(丝氨酸)或480位的t(苏氨酸)或481位的k(赖氨酸)或482位的k(赖氨酸)存在一个bitc分子。图2可知cshb蛋白第395位的t(苏氨酸)存在一个bitc分子。rna分子参与了大量的细胞过程。由于它们的相对简单性(只由4个不同的核酸构建块组成)和在细胞胞浆中发现的分子拥挤性,rna容易形成不必要的分子间和分子内相互作用。为了防止这些相互作用,每个细胞都表达有助于rna分子正常运转的蛋白质。其中一类主要的蛋白质是dead-box rna解旋酶。dead-box rna解旋酶是一种普遍存在的酶,由高度保守的解旋酶核心组成,该核心包含两个reca样结构域。在这两个结构域内,已鉴定出12个特征序列基序。这些基序涉及atp和rna结合、atp水解以及不同位点之间的通信。除了保守的解旋酶核心外,大多数dead-box蛋白还包含可变的n端或c端延伸。csha是四种rna解旋酶中最大的一种,在其c末端具有特定结构域。csha以及rna解旋酶cshb参与冷适应。这两种蛋白质都位于类核周围,这种模式让人联想到核糖体或冷休克蛋白(csp)。进一步利用cshb和冷休克蛋白b(cold shock protein b,cspb)进行荧光共振能量转移分析,证明了这两种蛋白之间存在相互作用,支持了cshb在冷适应中的作用。在研究单个突变体的生长时,已经表明了dead-box解旋酶的不
同功能。解旋酶编码基因csha、cshb中任何一个的缺失都会导致冷敏感表型。显然,解旋酶不能相互替换。用革兰氏阳性病原体单核细胞增生李斯特菌和蜡状芽孢杆菌获得了类似的结果。在这两种生物体中,缺乏枯草芽孢杆菌csha、cshb的同源物会导致冷敏感表型。相比之下,之前的一项研究发现csha在低温下是非必要的,即使在环境温度下也具有cshb突变的综合致死性。差异可能是由于使用不同的媒介或不同的遗传背景造成的。
23.实施例2
24.lgt细菌脂蛋白具有广泛而重要的生物学功能,例如维持细胞包膜结构、插入和稳定外膜蛋白、营养吸收、运输、粘附、侵袭和毒力。由图3可知prolipoprotein diacylglyceryl transferase蛋白第276位的t(苏氨酸)或277位的r(精氨酸)存在一个bitc分子。尽管脂蛋白的结构、功能和来源各不相同,但这些脂蛋白中的大多数都含有n-酰基二酰基甘油基-半胱氨酸作为它们的n末端氨基酸。脂蛋白的生物合成途径在革兰氏阳性菌和革兰氏阴性菌中包括三个反应。由于该途径已被证明能够影响g-的生存,因此这些酶为未来具有广谱作用的抗生素的研发提供了良好的潜在靶标。
25.实施例3
26.translation initiation factor if-2由图4可知translation initiation factor if-2蛋白第41位的t(苏氨酸)或42位的s(丝氨酸)存在一个bitc分子。细菌蛋白质合成或翻译的起始沿着多步途径进行,30s起始复合物(ic)的组装为该途径的开端位置。30s ic的组装可能通过多个路径发生,但在速度上优选的路径已被确定,三个if耦合到30s子基,并协调地调节trna耦合的速度。if-2在整个引通路中起着核心作用,它确保了fmet-trnafmet的正确选择。在30s ic组装过程中,起始因子(if)2通过选择性地增加50s核糖体亚基与携带n-甲酰基-甲硫氨酰-trna(fmet-trnafmet)的30s起始复合物(ic)的结合率来控制翻译起始的保真度。研究发现if2的结构域iii在if2的激活中起着关键的变构作用,这表明这个结构域可以作为开发新型抗生素的靶点。
27.实施例4
28.capg细菌荚膜多糖是保护细菌细胞免受环境胁迫的生物膜的主要组成部分。它们也在致病性中发挥重要作用。由图5可知capg蛋白第8位的t(苏氨酸)或12位的t或13位的r(精氨酸)存在一个bitc分子。金黄色葡萄球菌的致病性取决于微生物对宿主的成功适应和毒力因子的协调表达。金黄色葡萄球菌通常被薄的荚膜包围,其中5型(cp5)和8型(cp8)荚膜多糖(cp)是最常见的临床分离株。荚膜多糖在细胞表面形成一层厚厚的碳水化合物,赋予其抗吞噬作用,并帮助金黄色葡萄球菌在感染宿主的血液中持续存在。而合成细菌荚膜多糖的酶是很有吸引力的抗菌靶标。
29.实施例5
30.cyclic-di-ampcyclic-di-amp(c-di-amp)是一种由多种人类病原体合成的细菌信号核苷酸。这种广泛而特定的细菌产物被受感染的宿主细胞识别以触发先天免疫反应。由图6可知cyclic-di-amp蛋白第337位的k(赖氨酸)存在一个bitc分子。二级信使分子c-di-gmp在控制基因表达方面的明确功能已在多种细菌物种中确立。现在,有强有力的证据表明,环状二核苷酸在包括绿脓杆菌等重要人类病原体在内的各种细菌中,可以控制生物膜的形成和致病性基因表达。也有研究表明,该信号分子广泛存在于细菌中,但在高等真核生物中未发现,可作为真核细胞中的危险信号,促进了对c-di-gmp免疫调节和免疫刺激特
性的研究。金黄色葡萄球菌细胞质中c-di-amp的明确鉴定和调节其水平的能力为鉴定靶蛋白或其他化合物提供了可能性,这种环状二核苷酸通过这些化合物发挥其功能并调节细胞过程。c-di-amp在宿主细胞质中的检测主要通过sting-cgas信号转轴导致i型干扰素的诱导,但也与nf-kb通路的激活有关。在它们长期的相互作用中,宿主和病原体共同进化以控制先天免疫的c-di-amp激活。在细菌方面,细胞内释放的c-di-amp的数量允许通过避免免疫识别或相反地通过sting-cgas途径超载来操纵宿主反应以加剧感染。
31.实施例6
32.ldh金黄色葡萄球菌厌氧糖代谢的主要终产物是乳酸,乳酸可以抑制tca循环,增加乳酸水平会引起丁二醇途径的上调,在硝化胁迫下维持氧化还原平衡,这对细菌毒力和生物膜的形成起到关键作用。由图7可知ldh蛋白第45位的r(精氨酸)存在一个bitc分子。金黄色葡萄球菌的发病机制涉及多种代谢途径,该病原体具有完整的tca循环,在细菌的生长和定植中发挥重要作用。在高葡萄糖浓度的情况下,磷酸戊糖(pp)途径被上调,三羧酸(tca)循环被抑制;此外,当金黄色葡萄球菌在高糖条件下生长时,丙酮酸的氧化和细胞色素含量降低。tca循环酶的活性降低很可能是由于它们的生物合成受到抑制;这种现象被称为葡萄糖效应或碳分解代谢物抑制。根据d-或l-乳酸的特异性,乳酸脱氢酶可分为两类。第i类包括l-乳酸铁细胞色素c氧化还原酶(l-lcr)和nad依赖性l-乳酸脱氢酶,而第ii类包括三部分。在哺乳动物中,乳酸不仅是细胞氧化还原稳态的调节剂,而且还是人类肝细胞癌g2细胞线粒体中l-乳酸代谢的穿梭物。在稻瘟病菌中,d-乳酸是一种穿过线粒体膜间隙的主要代谢物,被d-乳酸脱氢酶1(modld1)氧化成丙酮酸,同时形成fadh2,为真菌呼吸链提供燃料。
33.实施例7
34.丙酮酸激酶pyk可以催化磷酸烯醇丙酮酸的不可逆转变,从而控制有机体中丙酮酸的水平。因此,厌氧条件下的高丙酮酸形成无助于能量产生,但有利于上调参与生物膜形成的生物合成途径,这是关键的致病因素之一。由图8可知pyk蛋白第264位的r(精氨酸)或266位的c(半胱氨酸)存在一个bitc分子。金黄色葡萄球菌主要通过糖酵解和克雷布斯循环从葡萄糖分解代谢中获取能量。糖酵解丙酮酸的终产物进入tca循环并调节与生物体致病性相关的能量水平。丙酮酸激酶(pyk)属于一组与pep水解的自由能耦合的转移酶,使用k+和mg2+作为辅助因子,生成atp和丙酮酸。
35.实施例8
36.ezra由图9可知ezra蛋白第256位的s(丝氨酸)或257位的t(苏氨酸)或260位的s存在一个bitc分子。细菌的细胞分裂是一个高度调节的过程,在这个过程中,细胞经历一系列时间和空间控制的事件,从而产生两个相同的子细胞。在几乎所有的细菌中,细胞分裂是随着一种类似微管蛋白的蛋白ftsz的聚合而开始的,形成位于未来分裂位点的环状结构。这种z-环作为支架招募参与细胞分裂的其他蛋白,导致多蛋白复合体的组装。ezra是分裂体复合物中的完整膜蛋白,在低gc含量的革兰氏阳性细菌中是保守的。有文献表明ezra在金黄色葡萄球菌中的主要作用是维持适当的ftsz在细胞中期的组装动力学,有助于协调细胞生长和细胞分裂,而不是阻止z环在不适当的位置组装。ezra在枯草杆菌中期细胞中发挥作用的一个假设是,随着细胞因子水平的提高,ezra通过加速z环的拆解而促进z环的重塑。因此,缺乏ezra会导致稳定的z-环,这种z-环可能在细菌细胞周期中存在较长时间。在金黄色
葡萄球菌中,隔膜是细胞壁合成的唯一场所。因此,可想而知,在ezra缺失的情况下,在隔膜处合成细胞壁的时间更多,因此可能会有额外的细胞壁材料掺入到隔膜中。含有较多细胞壁物质的隔膜有可能导致子细胞内半球较大,从而导致子细胞较大。这些细胞可能由于在ezra突变细胞周围分散的细胞壁合成而进一步增大尺寸。这些机制可以解释在金黄色葡萄球菌ezra突变体中观察到的较大的细胞尺寸。
37.实施例9
38.abc transporter,atp-binding protein在细菌、古菌和真核生物中都可以找到atp结合盒(abc)转运体,因为它是最大的超级家族之一。它们的功能作用非常多样化,从将必需营养素导入细菌细胞到赋予癌细胞多药耐药性。由图10可知abc transporter蛋白第15位的k(赖氨酸)或16位的t(苏氨酸)存在一个bitc分子。abc转运体分为两类:在细菌中,abc转运蛋白可能输出底物,包括药物和抗生素,或介导必需营养素的吸收;然而,在真菌和寄生虫中发现的大多数abc转运蛋白几乎都发挥着作用仅作为输出者,介导底物从富含atp的胞质溶胶转移出细胞或进入细胞内细胞器。值得注意的是,在细菌、病原真菌和寄生虫中发现的某些abc转运蛋白与抗微生物药物的耐药性有关。在哺乳动物中,abc转运蛋白具有重要的临床意义。对于abc输入一类,在没有底物的情况下可能会发生无效的atp水解,尽管速度比底物结合时的速度要慢。对于多特异性转运蛋白(包括多药转运蛋白abcb1),尚不清楚是只有一个结合位点还是存在多个位点。
39.实施例10
40.aminoglycoside 6'-n-acetyltransferase氨基糖苷类是一种杀菌抗生素,由图11可知aminoglycoside 6'-n-acetyltra-nsferase蛋白第135位的y(酪氨酸)或138位的s(丝氨酸)存在一个bitc分子。多药耐药(mdr)细菌引起的严重感染的治疗正变得更加复杂和昂贵得令人望而却步。因此迫切需要开发针对这些病原体的新疗法,不仅需要设计新型抗生素,还需要寻找与现有药物相结合规避耐药性的佐剂。后一种策略延长了已经在使用的抗生素的使用寿命,但由于耐药性特征的传播,这些抗生素正在变得无效。氨基糖苷类会干扰翻译保真度,产生不正确的初级序列蛋白质,从而导致多种毒性生理效应,并最终导致细胞死亡。这些抗生素在治疗由革兰氏阴性菌引起的危及生命的感染以及与其他抗菌素联合治疗革兰氏阳性菌引起的感染方面发挥了重要作用。尽管细菌已经发展出多种机制来抵抗氨基糖苷类,但在临床环境中酶失活是最普遍的。
41.实施例11
42.udk已有报道确定了t7噬菌体与其宿主大肠杆菌之间的遗传关系,鉴定出了四种宿主基因,其中有基因udk编码尿苷/胞苷激酶,这是一种将尿苷和胞苷转化为它们各自的核糖核苷单磷酸、ump和cmp的酶。由图12可知udk蛋白第102位的k(赖氨酸)存在一个bitc分子。该反应最有效的磷酸盐供体是gtp和dgtp。udk基因对大肠杆菌来说不是必需的,它是嘧啶核糖核苷酸补救途径的一部分。udk(一种编码尿苷/胞苷激酶的大肠杆菌基因)的过表达会限制t7噬菌体的生长。
43.实施例12
44.aursabat等人克隆并测序了编码金黄色葡萄球菌金属蛋白酶(aureys)的结构基因(aur),此外,他们还证实了aur基因以两种等位基因形式出现(i型和ii型),在人类金黄色葡萄球菌分离株中具有很强的保守性,表明该蛋白酶可能具有重要的管家功能。由图13
可知aur蛋白第131位的s(丝氨酸)存在一个bitc分子。然而,目前还不清楚是否所有来自各种家畜的分离株都保存着带有两种等位基因形式的aur基因。此外,ⅰ型和ⅱ型分离株金属蛋白酶的酶学性质未知。
45.实施例13
46.hslu由图14可知hslu蛋白第91位的t(苏氨酸)或95位的y(酪氨酸)存在一个bitc分子。atp依赖的蛋白酶是一类重要的细胞机器,在调控蛋白质的周转和清除受损蛋白质中发挥重要作用。它们利用atp水解产生的化学能,将其转化为机械力来展开蛋白质底物,并将其转移到蛋白水解室中进行降解。这些atp依赖的蛋白酶小室将蛋白水解活性位点从胞浆中分离出来,从而阻止胞浆蛋白不受控制地进入活性位点。hslvu是细菌中atp依赖的双组分蛋白酶之一,由hslv蛋白酶和hslu atp酶组成。hslv是20s蛋白酶体β亚基的同源物。它通过堆叠两个相同的hslv亚基的六聚环形成桶状十二聚体ic复合物,每个hslv亚基都含有一个n端的thr(thr1)蛋白水解活性位点。属于aaa+atp酶家族的六聚体hslu atp酶与hslv十二聚体的一端或两端结合形成hslvu复合物。在hslvu复合物中,hslu和hslv的中心孔排列,使hslu通过孔道将底物多肽链转移到hslv的内部蛋白水解室。
47.实施例15
48.如图15所示,thre蛋白第40位的t(苏氨酸)或41位的r(精氨酸)或44位的k(赖氨酸)存在一个bitc分子。
49.实施例16
50.如图16所示,phislt orf401-like蛋白105位的k(赖氨酸)存在一个bitc分子。
51.实施例17
52.如图17所示,phi pv83 orf 19-like蛋白第55位的r(精氨酸)存在一个bitc分子。
53.实施例18
54.如图18所示,yyci蛋白第77位的s(丝氨酸)或78位的s或79位的k(赖氨酸)存在一个bitc分子。
55.实施例19
56.如图19所示,aldehyde dehydrogenase b蛋白第487位的t(苏氨酸)或488位的s(丝氨酸)或492位的k(赖氨酸)存在一个bitc分子。
57.实施例20
58.如图20所示,lysr substrate binding domain protein第170位的r(精氨酸)或171位的r存在一个bitc分子。
59.实施例21
60.如图21所示,agcs_2蛋白第39位的r(精氨酸)存在一个bitc分子。
61.实施例22
62.如图22所示,dihydrolipoamide acetyltransferase component蛋白第149位的k(赖氨酸)或152位的r(精氨酸)或154位的t(苏氨酸)存在一个bitc分子。
63.实施例23
64.如图23所示,exotoxin蛋白第149位的y(酪氨酸)或148位的t(苏氨酸)存在一个bitc分子。
65.实施例24
66.如图24所示,bshc蛋白第305位的k(赖氨酸)或306位的t(苏氨酸)或308位的k或309位的k存在一个bitc分子。
67.实施例25
68.如图25所示,ribosomal protein l7ae蛋白第78位的k(赖氨酸)存在一个bitc分子。
69.实施例26
70.如图26所示,hemn蛋白第149位的s(丝氨酸)或151位的s(丝氨酸)存在一个bitc分子。
71.实施例27
72.如图27所示,infc蛋白第81位的k(赖氨酸)或82位的k或83位的k存在一个bitc分子。
73.实施例28
74.如图28所示,mene_1蛋白第490位的y(酪氨酸)存在一个bitc分子。
75.实施例29
76.如图29所示,toxin,beta-grasp domain蛋白第93位的c(半胱氨酸)存在一个bitc分子。
77.实施例30
78.如图30所示,cobb蛋白第180位的y(酪氨酸)存在一个bitc分子。
79.实施例31
80.如图31所示,nade蛋白第17位的r(精氨酸)或20位的s(丝氨酸)存在一个bitc分子。
81.实施例32
82.如图32所示,dape蛋白第392位的y(酪氨酸)存在一个bitc分子。
83.实施例33
84.如图33所示,cbix蛋白第245位的k(赖氨酸)或236位的s(丝氨酸)存在一个bitc分子。
85.实施例34
86.如图34所示,ysdc蛋白第278位的k(赖氨酸)或282位的k存在一个bitc分子。
87.实施例35
88.如图35所示,gltb_1蛋白第515位的t(苏氨酸)或519位的y(酪氨酸)或520位的k(赖氨酸)存在一个bitc分子。
89.实施例36
90.如图36所示,srmb蛋白第22位的s(丝氨酸)存在一个bitc分子。
91.实施例37
92.如图37所示,argf蛋白第2位的t(苏氨酸)存在一个bitc分子。
93.实施例38
94.如图38所示,n-acetyltransferase domain-containing蛋白第132位的k(赖氨酸)或133位的c(半胱氨酸)存在一个bitc分子。
95.实施例39
96.如图39所示,dna-3-methyladenine glycosylase蛋白第57位的k(赖氨酸)或59位的t(苏氨酸)或60位的s(丝氨酸)存在一个bitc分子。
97.实施例40
98.如图40所示,mfd蛋白第711位的k(赖氨酸)或713位的t(苏氨酸)或714位的k存在一个bitc分子。
99.实施例41
100.如图41所示,lipoprotein蛋白第362位的k(赖氨酸)或366位的k或364位的k存在一个bitc分子。
101.实施例42
102.如图42所示,gyra_1蛋白第372位的r(精氨酸)存在一个bitc分子。
103.实施例43
104.如图43所示,nadh dehydrogenase蛋白第8位的t(苏氨酸)或6位的k(赖氨酸)存在一个bitc分子。
105.实施例44
106.如图44所示,formate dehydrogenase,alpha subunit蛋白第803位的k(赖氨酸)或808位的s(丝氨酸)或第812位的y(酪氨酸)存在一个bitc分子。
107.实施例45
108.如图45所示,lysr family transcriptional regulator蛋白第257位的t(苏氨酸)存在一个bitc分子。
109.实施例46
110.如图46所示,putative membrane蛋白第568位的t(苏氨酸)存在一个bitc分子。
111.实施例47
112.如图47所示,entc蛋白第225位的y(酪氨酸)或228位的k(赖氨酸)或231位的y或232位的s(丝氨酸)或233位的k存在一个bitc分子。
113.实施例48
114.如图48所示,aroc_3蛋白第47位的r(精氨酸)存在一个bitc分子。
115.实施例49
116.如图49所示,uvra_3蛋白第942位的k(赖氨酸)或945位的t(苏氨酸)存在一个bitc分子。
117.实施例50
118.如图50所示,glcb_2蛋白第112位的t(苏氨酸)或118位的s(丝氨酸)或119位的y(酪氨酸)或121位的k(赖氨酸)存在一个bitc分子。
119.实施例51
120.如图51所示,irta蛋白第298位的t(苏氨酸)或300位的y(酪氨酸)存在一个bitc分子。
121.实施例52
122.如图52所示,superfamily i dna/rnahelicase蛋白第951位的r(精氨酸)或950位的k(赖氨酸)存在一个bitc分子。
123.实施例53
124.如图53所示,collagen adhesin蛋白第962位的k(赖氨酸)或第963位的y(酪氨酸)存在一个bitc分子。
125.上述53个异硫氰酸苄酯靶向结合蛋白中的一个或多个均可以用于制备bitc靶向抗菌剂或bitc靶向抗肿瘤药物;含有上述异硫氰酸苄酯靶向结合蛋白的多价靶向融合蛋白也可以用于制备bitc靶向抗菌剂或bitc靶向抗肿瘤药物;编码上述53个异硫氰酸苄酯靶向结合蛋白的核酸和/或编码上述多价靶向结合蛋白的核酸也可以用于制备bitc靶向抗菌剂或bitc靶向抗肿瘤药物。
126.本发明采用生物信息学分析及蛋白质组学鉴定等揭示多靶点协同作用促进了bitc对金黄色葡萄球菌的杀菌效果,全面解析出金黄色葡萄球菌中53个异硫氰酸苄酯的共价结合蛋白,共涉及99个作用修饰位点,以上述53个异硫氰酸苄酯的共价结合蛋白中的一个或多个为靶点,针对其上对应的结合靶点,通过生物信息学结构模拟及分子对接,开发靶向性金黄色葡萄球菌抗菌剂,可以极大提升抗菌剂的抗菌效果,减少耐药菌产生,治疗临床或食源性金黄色葡萄球菌污染。
127.针对常见致病菌包括幽门螺杆菌(helicobacter pylori)、银白色葡萄球菌(staphylococcus argenteus)、铜绿假单胞菌(pseudomonas aeruginosa)、溶血性链球菌(streptococcus hemolyticus)、单核细胞增生李斯特氏菌(listeria monocytogenes)、大肠杆菌(escherichia coli)、克罗诺杆菌(cronobacter spp)、小肠结肠炎耶尔森氏菌(yersinia enterocolitica)、蜡样芽孢杆菌(bacillus cereus)、肺炎链球菌(streptococcus pneumoniae)、肺炎克雷伯菌(klebsiella pneumoniae)、摩根菌属(morganella spp)、普罗威登斯菌属(providencia spp)以及淋病奈瑟菌(neisseria gonorrhoeae)、肠道沙门氏菌(salmonella enterica)、伤寒沙门氏菌(salmonella enterica serovar typhi)、鲍曼不动杆菌(acinetobacter baumannii)、粪肠球菌(enterococcus faecalis)、屎肠球菌(enterococcus faecium)、脑膜炎奈瑟菌(neisseria meningitidis)、流感嗜血杆菌(haemophilus influenzae)、霍乱弧菌(vibrio cholerae)、艰难梭菌(clostridioidesdifficile)、奈瑟氏淋球菌(neisseria gonorrhoeae)等,可以通过blast分别查找53个靶点蛋白的同源蛋白及结合位点,做靶向药物和靶向抗菌剂开发。
128.针对常见癌细胞,诸如肺癌细胞、肝癌细胞、前列腺癌细胞、食管癌细胞和白血病细胞等,针对53个靶点蛋白中的一种或几种,通过blast查找肺癌细胞、肝癌细胞、前列腺癌细胞、食管癌细胞和白血病细胞等细胞中对应的同源蛋白,针对同源蛋白和bitc结合位点,做靶向药物筛选和靶向药物设计,开发针对某一癌细胞单个或多个靶点的特异性抗癌药物。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1