用锌转运蛋白7抑制神经退行性变
背景技术:
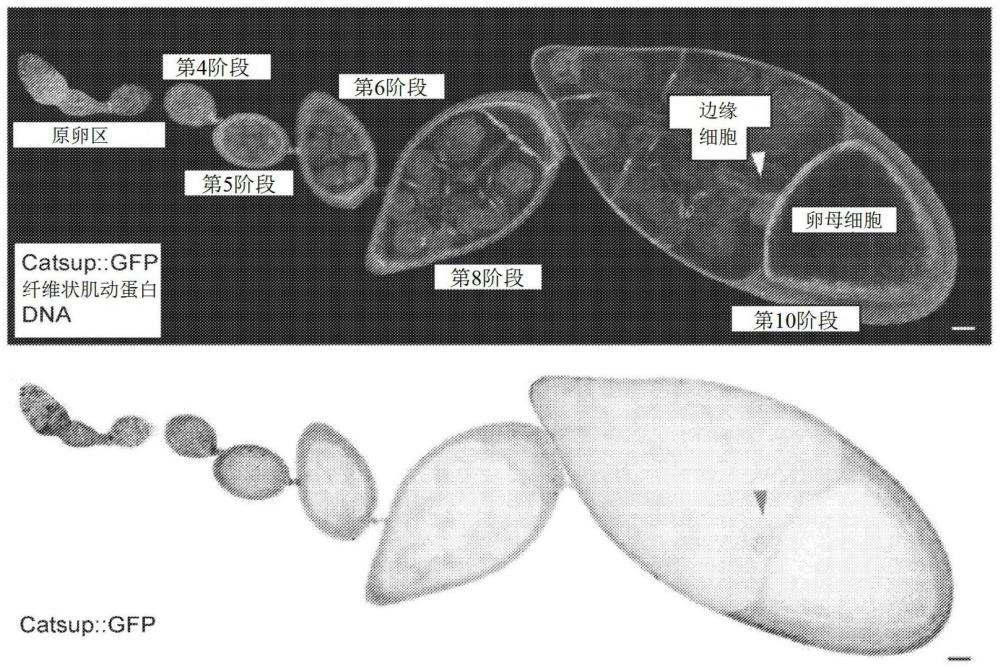
1、细胞集体迁移已成为正常器官发育、伤口修复和肿瘤转移的关键驱动因素(mishra等人,《发展(development)》146,(2019);friedl和mayor,《冷泉港生物学观点(cold spring harb.perspect.biol.)》9,(2017);friedl和gilmour,《自然评论-分子细胞生物学(nat.rev.mol.cell biol.)》10,445-457(2009);friedl等人,《自然细胞生物学(nat.cell biol.)》14,777-783(2012))。果蝇卵巢中的边缘细胞迁移提供了一个强大的细胞集体迁移的体内模型,该模型适合无偏遗传筛选。果蝇卵巢由卵巢管组成,卵巢管是一串串卵室,经过14个发育阶段发育成成熟的卵子(图1a)。每个卵室由16个生殖细胞组成,包括15个哺育细胞和一个卵母细胞,它们被上皮卵泡细胞包围。在第9阶段(图1b),4至8个边缘细胞在卵室的前端被指定,从卵泡上皮层离,并在第9发育阶段期间向后迁移,在第10阶段到达卵母细胞的前边缘。
2、遗传筛选已经深入了解了分子机制,这些分子机制指定了约850个卵泡细胞中有哪些卵泡细胞获得迁移的能力(silver和montell,《细胞(cell)》107,831-841(2001);bai和montell,《发展》129,5377-5388(2002))、其迁移的发展时机(jang等人,《自然细胞生物学》11,569-579(2009);bai和montell,《细胞》103,1047-1058(2000))、方向感应(mcdonald等人,《发展》130,3469-3478(2003);mcdonald等人,《发展生物学(dev.biol.)》296,94-103(2006);dai等人,《科学(science)》370(6519),987-990(2020))和细胞骨架动力学(mcdonald等人(2003),同上;murphy和montell,《细胞生物学杂志(j.cell biol.)》133,617-630(1996);kim等人,《基因与发育(genes dev.)》25,730-741(2011);lee等人,《发展》122,409-418(1996);cai等人,《细胞》157,1146-1159(2014);mcdonald,《当前生物学(curr.biol.)》18,1659-1667(2008);wang等人,《自然细胞生物学》12,591-597(2010);duchek和《科学》291,131-133(2001);fulga和rorth,《自然细胞生物学》4,715-719(2002);ramel等人,《自然细胞生物学》15,317-324(2013);assaker等人,《美国国家科学院院刊(proc natl acad sci usa)》107,22558-22563(2010))。边缘细胞研究继续提供新的生物学见解(dai等人,同上;miao等人,《发育细胞(dev.cell)》54,501-515.e9(2020);colombie等人,《发育生物学(dev.biol.)》423,12-18(2017);chen等人,《伊莱夫(elife)》9,e52979(2020);laflamme等人,《细胞生物学杂志》198,57-67(2012);ogienko等人,《国际分子科学杂志(int.j.mol.sci.)》21,(2020);berez等人,《生理学前沿(front.physiol.)》11,803(2020);wang等人,《交叉科学(iscience)》23,101335(2020);wang,h.、guo,x.、wang,x.、wang、x.和chen,j.“超细胞肌动球蛋白介导细胞间通讯并塑造集体迁移形态(supracellular actomyosin mediates cell-cell communication and shapescollective migratory morphology)”。《交叉科学》23,101204(2020);fox等人,《细胞分子生物学(mol.biol.cell)》31,1584-1594(2020);plutoni等人,《自然通讯(nat.commun.)》10,3940(2019);zeledon等人,《细胞报告(cell rep.)》28,3238-3248.e3(2019);lamb等人,《发育动力学(dev.dyn.)》249,961-982(2020);ghiglione,c.、jouandin,p.、cérézo,d.和noselli,s.“果蝇胰岛素途径在细胞集体迁移期间控制前纤维蛋白的表达和富含肌动蛋白的动态突起(the drosophila insulin pathway controls profilin expression anddynamic actin-rich protrusions during collective cell migration)”。《发展》145,dev161117(2018);sharma等人,《发展》145,(2018))。在用于嵌合克隆中的边缘细胞迁移缺陷的大规模甲磺酸乙酯诱变筛选(liu和montell,《发展》126,1869-1878(1999)和边缘细胞的全基因组表达谱(wang等人,《发育细胞》10,483-495(2006))中鉴定出基因catsup。
3、名称catsup是“儿茶酚胺up(catecholamines up)”的缩写,其丧失会增加芳香胺的合成,包括肾上腺素和多巴胺等神经递质(stathakis等人,《遗传学(genetics)》153,361-382(1999))。catsup是果蝇气管形态发生所必需的,在这种情况下,它直接结合并抑制酪氨酸羟化酶ple的果蝇同源物,以限制多巴胺合成(hsouna等人,《发育生物学》308,30-43(2007))。相反,在翅成虫盘细胞中,catsup调控notch和egfr的丰度和定位(groth等人,《发展》140,3018-3027(2013))。
4、catsup与其哺乳动物同源物锌转运蛋白7(zip7),也称为slc39a7或hke4(groth等人,同上),具有62%的相似性和53%的同一性,该蛋白是zn2+转运蛋白的两个主要家族之一的成员(kambe,“锌转运:调控,无机化学和生物无机化学百科全书(zinc transport:regulation.in encyclopedia of inorganic and bioinorganic chemistry)”(编辑scott,r.a.)1-9(约翰·威利父子出版公司(john wiley&sons,ltd),2011)。zip7位于包括er的内膜系统内,在那里它将zn2+转运到胞质溶胶(taylor等人,《生物化学杂志(biochem.j.)》377,131-139(2004))。zn2+是许多蛋白发挥作用所必需的微量元素,并且zn2+稳态由人体内24种zn2+转运蛋白精心维持,其中14种是zip43。zip7在整个真核生物中是保守的,其丧失会导致酵母、植物和动物等多种生物体的er应激(nguyen等人,《生物科学生物技术与生物化学(biosci.biotechnol.biochem.)》77,1337-1339(2013);tan等人,《转基因研究(transgenic res.)》24,109-122(2015);zhang等人,《国际分子科学杂志》15,20413-20433(2014);adulcikas等人,《生物学和医学中的计算机(comput.biol.med.)》100,196-202(2018);tuncay等人,《糖尿病(diabetes)》66,1346-1358(2017);fauster等人,《细胞死亡和分化(cell death differ.)》26,1138-1155(2019))。然而,er应激、zn2+转运、notch和egfr定位和活性以及细胞运动之间的关系仍有待阐明。
技术实现思路
1、本发明提供了用于通过增加zip7的表达或活性来增强内质网相关降解(erad)以及抑制错误折叠的蛋白在细胞中的病理性积累的方法和组合物。还提供了治疗受试者与蛋白错误折叠相关的病症的方法,包括用于在体内以足以抑制错误折叠的蛋白的病理性积累的有效量表达zip7的基因治疗方法。
2、在一个方面,提供了一种治疗受试者与蛋白错误折叠相关的病症的方法,该方法包括向受试者施用治疗有效量的锌转运蛋白7(zip7)。
3、可以通过本文所描述的方法治疗的与蛋白错误折叠相关的病症包括导致蛋白错误折叠和/或蛋白聚集的任何病况,其中错误折叠的蛋白和/或蛋白聚集体在内质网(er)内积累,在内质网中它们引起er应激并抑制er相关降解(erad)。与蛋白错误折叠相关的病症包括但不限于视网膜色素变性和神经退行性疾病,诸如亨廷顿氏症、阿尔茨海默氏症、帕金森氏症、额颞叶痴呆、肌萎缩性侧索硬化症(als)、齿状核红核苍白球丘脑下部核萎缩(drpla)、脊髓延髓肌肉萎缩症(sbma),也称为肯尼迪病、多系统萎缩症和脊髓小脑性共济失调(sca)。
4、在某些实施例中,zip7蛋白包含与seq id no:5的序列具有至少约80%至100%序列同一性的氨基酸序列,包括这个范围内的任何同一性百分比,诸如与该序列具有的81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%序列同一性。
5、在某些实施例中,将zip7蛋白局部施用到受试者的眼睛或大脑中。在一些实施例中,将zip7蛋白局部施用到眼睛的光感受器中。
6、在某些实施例中,zip7蛋白根据每日给药方案施用或间歇施用。
7、在某些实施例中,zip7蛋白由表达载体诸如病毒载体提供。
8、在某些实施例中,错误折叠的蛋白是变体视紫红质(例如,rh1g69d)、vap33或淀粉样蛋白β42。
9、在另一方面,提供了一种向受试者提供zip7以抑制错误折叠的蛋白的病理性积累的方法,该方法包括向细胞中引入表达载体,该表达载体包含可操作地连接到编码zip7的编码序列的启动子,其中该细胞在受试者中以足以抑制错误折叠的蛋白在细胞中的病理性积累的有效量体内表达zip。
10、在某些实施例中,细胞是视网膜细胞或大脑细胞。在一些实施例中,视网膜细胞是光感受器细胞。
11、在某些实施例中,错误折叠的蛋白是错误折叠的视紫红质蛋白。
12、在某些实施例中,将表达载体在体外或在体内引入细胞中。
13、在另一方面,提供了一种治疗受试者与蛋白错误折叠相关的病症的方法,该方法包括向受试者施用表达载体,该表达载体包含可操作地连接到编码zip7的核苷酸序列的启动子,其中zip7在受试者中以足以抑制错误折叠的蛋白的病理性积累的治疗有效量体内表达。
14、在另一方面,提供了一种增强细胞中内质网(er)相关降解(erad)或蛋白体相关降解的方法,该方法包括增加zip7在细胞中的表达或活性。
15、在某些实施例中,细胞是视网膜细胞。在一些实施例中,视网膜细胞是光感受器细胞。
16、在某些实施例中,zip7的表达被充分增加以增加错误折叠的视紫红质蛋白的降解并抑制错误折叠的视紫红质蛋白在视网膜细胞中的积累。
17、在某些实施例中,增加zip7的表达包括用包含编码zip7的编码序列的重组多核苷酸转染细胞。在一些实施例中,重组多核苷酸还包含病毒载体,该病毒载体包含可操作地连接到编码zip7的编码序列的启动子。在一些实施例中,编码zip7的编码序列被整合到经转染的细胞的染色体座位中,其中内源性启动子在染色体座位处可操作地连接到编码zip7的经整合的编码序列。
18、在另一方面,提供了一种抑制错误折叠的蛋白在受试者的器官、细胞或组织中积累的方法,该方法包括增加zip7在器官、细胞或组织中的表达或活性。
19、在某些实施例中,器官是眼睛或大脑。
20、在某些实施例中,组织是神经组织。
21、在某些实施例中,组织是视网膜组织。
22、在某些实施例中,细胞是视网膜细胞。在一些实施例中,视网膜细胞是光感受器细胞。
23、在某些实施例中,错误折叠的蛋白是视紫红质蛋白。
24、在某些实施例中,受试者患有视网膜色素变性。
25、在某些实施例中,与蛋白错误折叠相关的病症是视网膜色素变性、亨廷顿氏症、阿尔茨海默氏症、帕金森氏症、肌萎缩性侧索硬化症或额颞叶痴呆。
26、在某些实施例中,zip7的表达被充分增加以减少错误折叠的蛋白的病理性积累并提高细胞存活。
27、在某些实施例中,通过向器官、细胞或组织提供包含编码zip7的编码序列的重组多核苷酸使zip7表达增加,其中zip7在器官、细胞或组织中表达。
28、在某些实施例中,重组多核苷酸还包含病毒载体,该病毒载体包含可操作地连接到编码zip7的编码序列的启动子。
29、在某些实施例中,编码zip7的编码序列被整合到染色体座位中。在一些实施例中,内源性启动子在染色体座位处可操作地连接到编码zip7的经整合的编码序列。
30、在另一方面,提供了一种用于治疗与蛋白错误折叠相关的病症的方法的组合物,该组合物包含zip7或表达载体,该表达载体包含可操作地连接到编码zip7的编码序列的启动子。在一些实施例中,该组合物还包含药学上可接受的赋形剂。
- 还没有人留言评论。精彩留言会获得点赞!