相关申请的交叉引用[不适用]序列表该申请包含序列表,其通过引用而并入并且作为“28298us01_seqlist.txt”命名的文本文件与本申请一起提交。序列表文件创建于2014年12月22日,并且大小为3000字节。基金本文所公开的研究中至少一部分得到了日本政府机构的日本科技机构(jst)的资助。
背景技术:
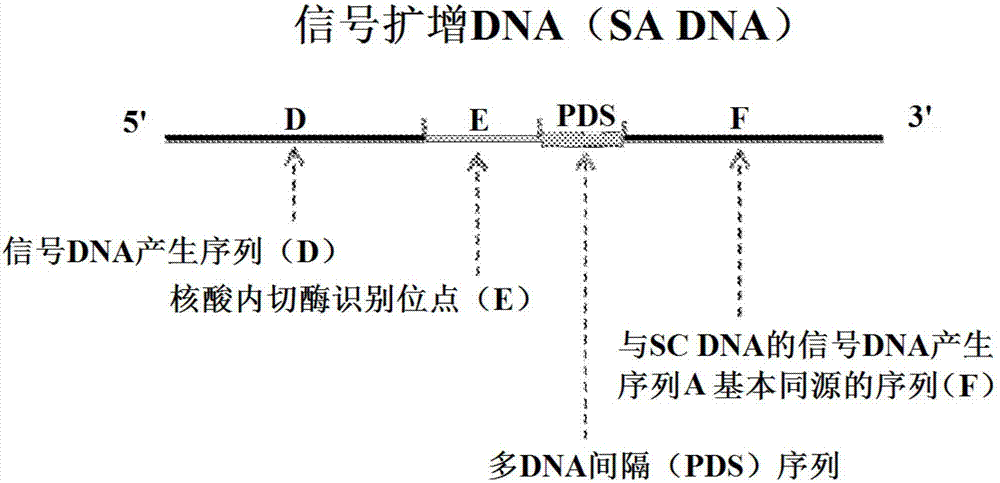
:检测试验样品中的靶核酸在各个领域中都重要,包括医学和生物学。许多组合物、测定平台和程序可用于检测特异性核酸分子。为了使检测在临床环境中有效,这些程序不仅应该是可重现和准确的,而且还必须是稳健和快速的。一种用于从混合核酸序列群扩增特定序列的常用方法是聚合酶链式反应(pcr)。由于典型的pcr在三个不同的温度下进行,所以反应可能与挑战相关,例如难以保持精确的温度,以及时间损失与扩增循环的数量成比例地增加。双链模板dna变性为单链(虽然在一定程度上依赖于特定序列)通常需要使用高“解链”温度,这限制了可用于那些高度耐热的dna聚合酶的类别。因此,已经开发了等温扩增平台技术以在比pcr中使用的反应条件更温和的反应条件下检测核酸。然而,这些等温扩增技术尚未解决与实现更快反应时间相关的挑战。以下公开内容提供了用于在比pcr中使用的反应条件更不严格的反应条件下检测核酸序列(例如dna或rna)的替代方法和组合物。该方法和组合物保持序列选择性和灵敏度,其允许检测样品中可能低浓度的核酸分子和/或短长度的核酸分子。该方法和组合物还减少反应时间。在其它方面,本发明提供了可以在低温、等温条件下提高样品中靶核酸的检测限的新方法和核酸分子,并且可以简化或改进样品制备和自动化检测方法,同时减少反应时间。技术实现要素:一方面,本发明涉及一种用于检测样品中靶核酸的方法,所述方法包括使所述样品与寡核苷酸、聚合酶和核酸内切酶接触,所述寡核苷酸(序列转换dna或scdna)在5'至3'方向上包含信号dna产生序列(a)、核酸内切酶识别位点(b)、多dna间隔序列(pds)和与靶核酸的3'末端互补的序列(c);所述核酸内切酶用于切口反应。在该方面的实施方式中,该方法还包括确定信号dna的存在或不存在,其中信号dna的存在表明样品中靶核酸的存在。一方面,本发明涉及一种用于检测样品中靶核酸的方法,所述方法包括使所述样品与第一寡核苷酸、第二寡核苷酸、聚合酶和核酸内切酶接触,所述第一寡核苷酸(序列转换dna或scdna)在5'至3'方向上包含信号dna产生序列(a)、核酸内切酶识别位点(b)、多dna间隔序列(pds)和与靶核酸的3'末端互补的序列(c);所述第二寡核苷酸(信号扩增dna或sadna)在5'至3'方向上包含与第一寡核苷酸的信号dna产生序列(a)同源的信号dna产生序列(d)、核酸内切酶识别位点(e)(其可以与scdna中的核酸内切酶识别位点(b)相同或不同)、多dna间隔序列(pds)(其可以与scdna中的pds相同或不同)和与第一寡核苷酸的信号dna产生序列(a)同源的序列(f);所述核酸内切酶用于切口反应。在该方面的实施方式中,该方法还包括确定信号dna的存在或不存在,其中信号dna的存在表明样品中靶核酸的存在。此外,在该方面的一些实施方式中,信号扩增dna(sadna)不具有多dna间隔序列(pds)。间隔是多“核酸”间隔,例如可以是多dna间隔(pds)、多rna间隔(prs)或其衍生物或类似物(例如人造核酸)。在某些实施方式中,多dna间隔(pds)序列可以包含g、a、t、c或其任何组合。多rna间隔(prs)序列可以包含g、a、u、c或其任何组合。此外,pds序列可以是1至20、1至15、1至10、1至8、1至6、或1至5个碱基长度。如下面更详细描述,本文中公开的方法对于检测样品中的靶rna特别有用。在核酸内切酶识别位点(b)和与靶标的3'末端互补的序列(c)之间的多dna间隔(pds)序列的scdna中的放置大大提高了产生信号dna的速率。在另一方面,本发明涉及加速或增强rna-dna混合双链体的下游、上游和/或相邻的核酸内切酶切口活性。在某些实施方式中,聚合酶可具有链置换活性。在另外的实施方式中,聚合酶可以是3'至5'核酸外切酶缺陷、5'至3'核酸外切酶缺陷、或3'至5'核酸外切酶缺陷和5'至3'核酸外切酶缺陷两者。在一些实施方式中,聚合酶包含dna聚合酶。在实施方式中,核酸内切酶可以包含可用于切割寡核苷酸的反应中的切口核酸内切酶或限制性核酸内切酶。虽然本文中公开的方法可以在典型的dna扩增条件(例如与标准pcr、反应物浓度、时间循环等相关的典型温度)下进行,但在一些实施方式中,该方法可以在等温条件或基本上恒温下进行。在另外的实施方式中,该方法可以在比标准pcr方法中使用的温度更低的温度下进行。作为一个示例,该方法的一些实施方式可以在等于或低于如下所述的scdna的靶核酸(t)和序列(c)或sadna的信号dna(s)和序列(f)的计算的最佳杂交或退火温度或实验测定的杂交或退火温度的温度下进行。在实施方式中,该方法可以在低于与scdna的序列(c)结合的靶核酸(t)或与sadna的序列(f)结合的信号dna(s)的解链温度的温度下进行。在其它实施方式中,该方法可以在允许聚合酶和/或核酸内切酶活性的温度下进行。在另外的实施方式中,该方法可以在处于或约为用于检测样品中靶核酸的反应混合物中存在的聚合酶和/或核酸内切酶的最佳反应温度的温度下进行。另一方面,本发明涉及一种寡核苷酸,其在本文中可称为“序列转换dna”(或“scdna”),其在5'至3'方向上包含信号dna产生序列(a)、核酸内切酶识别位点(b)、多dna间隔(pds)序列和与靶核酸的3'末端互补的序列(c)。另一方面,本发明涉及一种寡核苷酸,其可以在本文中称为“信号扩增dna”(或“sadna”),其在5'至3'方向上包含与序列转换dna(scdna)的信号dna产生序列(a)同源的信号dna产生序列(d)、核酸内切酶识别位点(e)、多dna间隔(pds)序列和与序列转换dna(scdna)的信号dna产生序列(a)同源的序列(f)。另一方面,本发明涉及信号扩增dna,其在5'至3'方向上包含与序列转换dna(scdna)的信号dna产生序列(a)同源的信号dna产生序列(d)、核酸内切酶识别位点(e)和与序列转换dna(scdna)的信号dna产生序列(a)同源的序列(f)。本发明的sc和/或sadna通常是线性的,然而这些dna也可以是圆形的(即微环dna(mc))。当靶核酸的3'末端与微环scdna结合时,或者当信号dna的3'末端与微环sadna结合时,可以引发滚环扩增(rca)。所得到的rca产物是含有数千拷贝的scdna或sadna的长单链dna片段。在一个实例中,scdna的信号dna产生序列(a)可以与靶核酸(t)的5'末端互补。在这方面,靶核酸(t)与scdna的信号dna产生序列(a)和序列(c)均结合。靶核酸(t)与scdna的结合(在dna连接酶存在下)导致形成微环scdna(mcscdna),并随后引发滚环扩增(rca)。所得到的rca产物是含有数千拷贝的scdna的长单链dna片段。在一个实施方式中,scdna的核酸内切酶识别位点在发夹结构的双链茎环区内,并因此在切口核酸内切酶存在下进行切割(在茎环的3'侧)。在切口核酸内切酶和聚合酶均存在的情况下,可以直接从rca产物产生信号dna。本发明的方法和寡核苷酸可以与其它扩增和/或检测方案组合使用。例如,根据本文公开的方法产生的信号dna中的任一种可以用作滚环扩增反应中的引物。在一个实施方式中,根据本发明的方法产生的信号dna的3'末端可以与微环dna模板互补,并且可以在结合信号dna时开始滚环扩增。靶核酸序列可以是感兴趣的任何核苷酸序列,并且在一些实施方式中可以包含源自感染剂或微rna的序列。在其它实施方式中,靶核酸可以包含可能与疾病或病症相关的基因的序列。在一些实施方式中,核酸内切酶识别位点包含与由核酸内切酶切割的序列互补的序列。在其它实施方式中,由核酸内切酶切割的序列与由核酸内切酶特异性识别的序列相邻(下游或上游)。在另一方面,本发明涉及一种用于检测样品中靶核酸的组合物,所述组合物包含:寡核苷酸(序列转换dna或scdna),其在5'至3'方向上包含信号dna产生序列(a)、核酸内切酶识别位点(b)、多dna间隔(pds)序列和与靶核酸的3'末端互补的序列(c);聚合酶;和用于切口反应的核酸内切酶。在另一方面,本发明涉及一种用于检测样品中靶核酸的组合物,所述组合物包含:第一寡核苷酸(序列转换dna或scdna),其在5'至3'方向上包含信号dna产生序列(a)、核酸内切酶识别位点(b)、多dna间隔(pds)序列和与靶核酸的3'末端互补的序列(c);第二寡核苷酸(信号扩增dna或sadna),其在5'至3'方向上包含与第一寡核苷酸的信号dna产生序列(a)同源的信号dna产生序列(d)、核酸内切酶识别位点(e)(其可以与sadna中的核酸内切酶识别位点(b)相同或不同)、多dna间隔(pds)序列(其可以与第一寡核苷酸的pds序列相同或不同)和与第一寡核苷酸的信号dna产生序列(a)同源的序列(f);聚合酶和用于切口反应的核酸内切酶。在该方面的一些实施方式中,第二寡核苷酸或信号扩增dna(sadna)不具有多dna间隔(pds)序列。当核酸内切酶识别位点是双链时,组合物还可以包含聚合酶和/或在第一和第二寡核苷酸的核酸内切酶识别位点处或邻近能够切割的核酸内切酶。组合物还可以包括用于新合成的dna的荧光检测的其它试剂,例如反应缓冲液、脱氧核糖核苷酸和报道分子,例如荧光团修饰的探针dna(例如分子信标探针)。可以使用本领域通常已知的报道分子,包括吖啶共轭的探针(即化学发光技术)。在另一方面,本发明涉及一种用于检测样品中靶核酸的试剂盒,所述试剂盒包含:寡核苷酸(序列转换dna或scdna),其在5'至3'方向上包含信号dna产生序列(a)、核酸内切酶识别位点(b)、多dna间隔(pds)序列和与靶核酸的3'末端互补的序列(c);聚合酶;和用于切口反应的核酸内切酶。在一些实施方式中,试剂盒可以还包含能够切割核酸内切酶识别位点或邻近核酸内切酶识别位点的位点的聚合酶和/或核酸内切酶。试剂盒还可以包括用于荧光检测新合成的dna例如信号dna的试剂,例如反应缓冲液、脱氧核糖核苷酸和报道分子,例如荧光团修饰的探针dna(例如分子信标探针)。试剂盒还可以包括用于实践本文公开的任何一种方法的说明书。在另一方面,本发明涉及一种用于检测样品中靶核酸的试剂盒,所述试剂盒包含:第一寡核苷酸(序列转换dna或scdna),其在5'至3'方向上包含信号dna产生序列(a)、核酸内切酶识别位点(b)、多dna间隔(pds)序列和与靶核酸的3'末端互补的序列(c);第二寡核苷酸(信号扩增dna或sadna),其在5'至3'方向上包含与第一寡核苷酸的信号dna产生序列(a)同源的信号dna产生序列(d)、核酸内切酶识别位点(e)(其可以与sadna中的核酸内切酶识别位点(b)相同或不同)、多dna间隔(pds)序列(其可以与第一寡核苷酸的pds序列相同或不同)和与第一寡核苷酸的信号dna产生序列(a)同源的序列(f)。在一些实施方式中,第二寡核苷酸或信号扩增dna(sadna)不具有多dna间隔(pds)序列。在一些实施方式中,试剂盒可以还包含能够切割核酸内切酶识别位点或邻近核酸内切酶识别位点的位点的聚合酶和/或核酸内切酶。试剂盒还可以包括用于荧光检测新合成的dna例如信号dna的试剂,例如反应缓冲液、脱氧核糖核苷酸和报道分子,例如荧光团修饰的探针dna(例如分子信标探针)。试剂盒还可以包括用于实践本文公开的任何一种方法的说明书。本文公开的方法、寡核苷酸、组合物和试剂盒可以与整体系统平台组合使用。例如,本发明的方法、寡核苷酸、组合物和试剂盒可以与abbott’sarchitect系统组合使用。本文公开的方法、寡核苷酸、组合物和试剂盒可以与样品制备系统平台一起使用,例如m2000sp样品制备系统(abbottdiagnostics,abbottpark,il)。类似地,本文公开的方法、寡核苷酸、组合物和试剂盒可以与检测系统平台一起使用,例如abbott’si-stat检测系统(abbottdiagnostics,abbottpark,il)。此外,本发明的方法、寡核苷酸、组合物和试剂盒可以与任何数量的其它装置、测定平台和仪器一起使用,例如手持式荧光检测器、微ph计、微流体装置、微阵列酶检测系统、免疫色谱条和侧流装置。本文公开的方法、寡核苷酸、组合物和试剂盒可用于分子诊断领域,包括非感染性和感染性疾病的诊断。例如,本发明的方法、寡核苷酸、组合物和试剂盒可以用于检测人体液例如血液、尿液、唾液、汗液和粪便中的微rna、信使rna、非编码rna和甲基化dna。类似地,本发明的方法、寡核苷酸、组合物和试剂盒可用于检测人体液例如血液、尿液、唾液、汗液和粪便中的源自感染性疾病的靶核酸,例如hbv、hcv、hiv、hpv、htlv-1、parvo病毒、结核病、梅毒、疟疾和溶组织内阿米巴。应当理解,在本发明的一些方面,本文公开的sc和sadna可以包含化学修饰的核苷酸。例如,本文公开的sc和sadna可以包含lna(锁定核酸)、bna(桥连核酸)、ena(乙烯桥连核酸)、gna(乙二醇核酸)、tna(苏糖核算)、pna(肽核酸)、吗啉代核酸、硫代磷酸酯核苷酸、或其组合和/或衍生物。考虑到下面的描述,由本发明提供的另外的方面、实施方式和优点将变得显而易见。附图说明图1a是示意性说明用于检测样品中靶核酸的序列转换dna(scdna)的非限制性实例的图。scdna在5'至3'方向上包含信号产生序列(a)、可用于切口反应的核酸内切酶识别位点(b)、多dna间隔(pds)序列和与靶核酸互补的序列(c)。图1b是示意性说明用于检测样品中靶核酸的信号扩增dna(sadna)的非限制性实例的图。sadna在5'至3'方向上包含与scdna的信号dna产生序列(a)同源的信号dna产生序列(d);核酸内切酶识别位点(e)(其可以与scdna的核酸内切酶识别位点(b)相同或不同)、多dna间隔(pds)序列(其可以与scdna的pds序列相同或不同)和与第一scdna的信号dna产生序列(a)同源的序列(f)。在一些实施方式中,信号扩增dna(sadna)不具有多dna间隔(pds)序列。图2a是示意性说明用于检测样品中靶核酸的靶(t)核酸与序列转换(sc)dna的示例性反应进展的图。序列(a)-(c)和pds如图1a中所述,序列(t)表示靶序列,序列(x)表示当与序列(c)结合的靶(t)通过聚合酶扩增时产生的序列,序列(x')表示切口扩增序列,序列(s)表示最终产生的信号dna序列。图2b是示意性说明用于检测样品中靶核酸的信号dna(s)与信号扩增(sa)dna的示例性反应进展的图。序列(d)-(f)和pds如图1b中所述,序列(s)是由target(t)核酸与scdna反应产生的信号dna,如图2a中所述,序列(y)表示与序列(d)结合的信号dna(s)通过聚合酶扩增时产生的序列,序列(y')表示切口扩增序列,序列(s)表示最终产生的信号dna序列。由于sa信号产生序列(d)与sc信号产生序列(a)同源,因此产生相同的信号dna(s)。图3显示了实施例1中进行的那些反应的结果,表明位于核酸内切酶识别位点(b)和scdna的序列(c)之间的pds序列的存在加速了信号dna的扩增。图4显示了实施例2中进行的这些反应的结果,表明位于核酸内切酶识别位点(b)和scdna的序列(c)之间的pds序列的存在增强了核酸内切酶识别位点(b)的切割。具体实施方式在一般意义上,本发明涉及核酸构建体,其在检测试验样品中的靶核酸中令人惊奇地有效。本文中公开的构建体包含允许在靶核酸存在下产生的信号dna产生的核酸序列,其伴随着反应速度的增加。本文公开的方法和核酸构建体提供了可有利地在低温和等温条件下进行靶核酸的选择、灵敏和快速的检测。在一方面,本发明涉及一种寡核苷酸,其在本文中可称为“信号扩增dna”(或“sadna”),其在5'至3'方向上包含与已知的信号dna序列互补的第一序列、核酸内切酶识别位点、多dna间隔(pds)序列和与第一序列的同一已知信号dna序列互补的第二序列。第一序列是图1b中的信号dna产生序列(d),其与scdna的已知信号dna产生序列(a)同源。第二序列是图1b中的序列(f),与同一scdna的同一已知信号dna产生序列(a)同源。在这方面,第一和第二序列的长度可以变化,但通常每个序列的长度与另一个序列大约相同。在实施方式中,序列的长度可以在约5至约100个核苷酸的范围内,但更通常为约5至约30、约10至约30、或约15至约30个核苷酸。核酸内切酶识别位点包含可被如本文所述的核酸内切酶识别、结合和切割的序列。这样的序列通常是本领域已知的。核酸内切酶识别位点可以包含核酸内切酶结合位点5'或3'(或5'和3'两者)的另外的核苷酸,但其长度通常不超过10个核苷酸。本发明提供了可用于检测样品中靶核酸的新型序列转换(sc)和信号扩增(sa)寡核苷酸构建体及其组合,其伴随着反应速度的增加。如图1a的说明性实施方式所示,用于检测样品中靶核酸的序列转换dna(scdna)寡核苷酸在5'至3'方向上包含信号dna产生序列(a)、核酸内切酶识别位点(b)、多dna间隔(pds)序列和与靶核酸的3'末端互补的序列(c)。如图1b的说明性实施方式所示,用于检测样品中靶核酸的信号扩增dna(sadna)在5'至3'方向上包含与scdna的信号dna产生序列(a)同源的信号dna产生序列(d);核酸内切酶识别位点(e)(其可以与scdna的核酸内切酶识别位点(b)相同或不同)、多dna间隔(pds)序列(其可以与scdna的pds相同或不同)和包含与scdna的信号dna产生序列(a)同源的序列的序列(f)。在一些实施方式中,信号扩增dna(sadna)不具有多dna间隔(pds)序列。如图1a中所示,本文公开的scdna包括信号产生序列(a)。scdna中的信号产生序列(a)可以包含任何所需的核酸序列,并且不受任何特定序列的限制。如下面更详细地讨论,信号产生序列(a)提供信号dna的模板的至少一部分(例如图2中的核酸(s)),其产生表明靶核酸的存在。scdna中的信号产生序列(a)不受长度限制。在一些实施方式中,scdna中的信号产生序列(a)为约5至约100个核酸碱基,以及5至100之间的所有整数。在实施方式中,scdna中的信号产生序列(a)为约5至约30个核酸碱基,以及5至30之间的所有整数。在一些实施方式中,scdna中的信号产生序列(a)为约10至约30个核酸碱基,以及10至30之间的所有整数。在另外的实施方式中,scdna中的信号产生序列(a)为约15至约30个核酸碱基,以及15至30之间的所有整数(例如约16、约17、约18、约19、约20、约21、约22、约23、约24、约25、约26、约27、约28、约29或约30个碱基)。如图1b中所示,本文公开的sadna包含与scdna的信号dna产生序列(a)同源的信号dna产生序列(d)和与同一scdna的同一信号dna产生序列(a)同源的序列(f)。在一些实施方式中,为了与scdna的信号dna产生序列(a)同源,序列(d)和(f)与相应的信号dna产生序列(a)完全相同。在其它实施方式中,序列(f)与scdna的相应信号dna产生序列(a)的序列相同,不同之处在于它为约1至约5、或约1至约4、或约1至约3个、或约1至约2个、或1个3'末端较短的碱基。当sadna的信号dna产生序列(d)与scdna的信号dna产生序列(a)同源时,产生相同的信号dna(s)并指数扩增。sc和sadna分别包含可以相同或不同的核酸内切酶识别位点(b)和(e)。在单链形式(例如图1a和1b的结构)中,核酸内切酶识别位点(b)和(e)可以包含与可由核酸内切酶切割的序列互补的序列。由核酸内切酶切割的序列可以在由核酸内切酶识别的序列的内部、下游或上游。适当地,当双链时,核酸内切酶识别位点(b)和(e)可被存在于反应中的一个或多个核酸内切酶识别,并且核酸内切酶识别位点(b)和(e)(或与核酸内切酶识别位点(b)和(e)相邻的序列)可以仅在双链dna的一条链上被切开(即切割)。如下面更详细描述,靶核酸与scdna的互补序列(c)的结合通过dna聚合酶引发复制以产生核酸内切酶识别位点(b)的活性双链形式,其现在能够作为核酸内切酶的识别位点(图2a)。核酸内切酶在新产生的双链核酸内切酶位点(b)或与新产生的双链核酸内切酶位点(b)相邻的位点处切割,然后通过dna聚合酶引发复制并产生信号dna(s)(参见例如图2a)。如图2a中所示,将核酸内切酶识别位点(b)取向以致使新复制的链被切割而不是scdna。也就是说,当产生新复制的链时,(b)中的核酸内切酶识别位点的取向指导新复制链的核酸内切酶活性(切开)。因此,核酸内切酶识别位点包含与由核酸内切酶切口的序列互补的序列,允许sc寡核苷酸在整个反应中保持完整(即scdna未被切割或切开)。sc和sadna还可以包含多dna间隔(pds)序列。在scdna中,pds序列位于核酸内切酶识别位点(b)和与靶核酸的3'末端互补的序列(c)之间。在sadna中,pds序列位于(e)中的核酸内切酶识别位点和与scdna的信号产生序列(a)基本同源的序列(f)之间。在一些实施方式中,信号扩增dna(sadna)不具有多dna间隔(pds)序列。sc和sadna的pds序列可以相同或不同。pds序列可以由天然碱基g、a、t或c中的任一个或多个组成,并且可以是3至20、3至18、3至16、3至14、3至14、3至12、3至10、3至8、3至6、或3至5个碱基长度。如下所述,本文公开的一个或多个sc或sadna中的pds序列的存在可以加速根据本文公开的方法进行的反应多达5至60、5至50、5至40、5至30、5至20、或5至10分钟。如下面更详细地描述,从scdna的信号产生序列(a)产生的信号dna(s)与sadna的序列(f)的结合通过dna聚合酶引发复制以产生sadna的核酸内切酶识别位点(e)的活性双链形式,其可以作为核酸内切酶识别位点(图2b)。核酸内切酶在sadna的新产生的双链核酸内切酶位点(e)或与新产生的双链核酸内切酶位点(e)邻近的位点处切割,然后通过dna聚合酶引发复制并产生信号dna(s),其与从scdna产生的信号dna(s)相同(图2b)。如图2b中所示,核酸内切酶识别位点(e)取向为使得新复制的链被切割,而不是sadna。也就是说,当产生新复制链时,e中的核酸内切酶识别位点的取向指导新复制链的核酸内切酶活性(切开)。因此,核酸内切酶识别位点包含与由核酸内切酶切口的序列互补的序列,允许sa寡核苷酸在整个反应中保持完整(即sadna未被切割或切开)。与靶dna互补的scdna的序列(c)不受长度的限制,并且可以是约5至约100个核酸碱基,以及5和100之间的所有整数。在一些实施方式中,scdna的序列(c)为约5至约30个核酸碱基,以及5至30之间的所有整数。在一些实施方式中,scdna中的序列(c)为约10至约30个核酸碱基,以及10和30之间的所有整数。在另外的实施方式中,scdna的序列(c)为约15至约30个核酸碱基,以及15至30之间的所有整数。互补序列能够形成氢键相互作用以形成双链核酸结构(例如核酸碱基对)。例如,与第一序列互补的序列包括能够与第一序列形成沃森-克里克碱基对的序列。如本文所用,术语“互补”不要求序列在其互补链的全长上是互补的,并且包含与另一序列的一部分互补的序列。因此,在一些实施方式中,互补序列包括在序列的整个长度上或其一部分上互补的序列(例如大于序列长度的约10%、约20%、约30%、约40%、约50%、约60%、约70%、约80%或约90%)。例如,两个序列可以在超过约2至约100范围内的连串(连续)核苷酸的长度上彼此互补,或2至100之间的任何整数。在一些实施方式中,两个序列可以在超过约15至约30范围内的连串(连续)核苷酸的长度上彼此互补,或15至30之间的任何整数。如本文所用,互补序列可以包含具有一些序列不匹配的序列。例如,互补序列可以包括与序列长度的至少约70%至100%,优选为大于95%互补的序列。尽管一些量的错配,互补序列通常具有在适当条件下选择性地彼此杂交的能力,例如严格和高度严格的条件,例如本文描述的那些或本领域普通技术人员通常已知的那些。sc和sadna可以通过已知方法合成。例如,sc和sadna可以使用亚磷酰胺法、磷酸三酯法、h-膦酸酯法或硫代膦酸酯法合成。在一些实施方式中,可以纯化sc和/或sadna,例如使用离子交换hplc。sc和sadna可以包括本领域通常已知的化学修饰。在一些实施方式中,例如sc和sadna可以包含化学修饰的核苷酸(例如2'-o甲基衍生物、硫代磷酸酯等)、3'末端修饰、5'末端修饰或其任何组合。在一些实施方式中,可以修饰sc和sadna的3'末端,使得扩增反应不会从sc或sadna的3'末端发生(例如当靶序列或另一非靶序列的结合,可能作为聚合酶扩增的引物)。如图2a中所示,它是靶核酸(t)的3'末端,而不是启动dna复制的scdna。从sc或sadna的3'末端发起的任何复制可能导致检测错误(例如假阳性)。此外,例如由scdna与非靶序列之间的结合、在scdna与靶序列之间的不正确位置的结合、sc和sadna之间的结合、或非模板的重新或从头的dna合成引起的scdna的未修饰的3'末端的非特异性扩增反应也可能导致检测错误。因此,在实施方式中,sc和sadna包含3'末端修饰,其可以减少或消除任何非期望的扩增反应的发生,例如上述讨论的那些。3'末端修饰的非限制性实例包括tamra、dabcyl和fam。修饰的其它非限制性实例包括例如生物素化、荧光色素沉淀、磷酸化、硫醇化、胺化、反转核苷酸或无碱基团。在另一方面,本发明包括用于检测样品中靶核酸(t)的方法。该方法通常包括使所述样品与第一寡核苷酸、第二寡核苷酸、聚合酶和核酸内切酶接触,所述第一寡核苷酸(或序列转换dna或scdna)在5'至3'方向上包含信号dna产生序列(a)、核酸内切酶识别位点(b)、多dna间隔(pds)序列和与所述靶核酸(t)的3'末端互补的序列(c);所述第二寡核苷酸(或信号扩增dna或sadna),其在5'至3'方向上包含与第一寡核苷酸的信号dna产生序列(a)同源的信号dna产生序列(d)、核酸内切酶识别位点(e)(与第一寡核苷酸的核酸内切酶识别位点(b)相同)、多dna间隔(pds)序列(与第一寡核苷酸的pds相同或不同)和与第一寡核苷酸的信号dna产生序列(a)同源的序列(f);所述核酸内切酶用于切口反应。在该方面的实施方式中,该方法还包括确定信号dna的存在或不存在,其中信号dna的存在表明样品中靶核酸的存在。在一些实施方式中,信号扩增dna(sadna)不具有多dna间隔(pds)序列。该方法包括使样品与核酸内切酶接触。核酸内切酶可以是切口核酸内切酶或限制性核酸内切酶,其能够或可用于切割与scdna内的核酸内切酶识别位点(b)互补的序列,或与sadna内的核酸内切酶识别位点(e)互补的序列。在一些实施方式中,核酸内切酶包含可催化或可用于催化双链dna切口反应的切口核酸内切酶或限制性核酸内切酶。在提供切口核酸内切酶的实施方式中,双链dna的一条链的磷酸二酯键可以被切割以在切割位点上产生5'侧的磷酸基和3'侧的羟基。切口核酸内切酶的非限制性实例包括nb.bbvci、ntalwi、nt.bbvci、nb.bsrdi、nb.bts1、nt.bspqi、nt.bstnbi、nb.bsmi、nt.cvipii和nt.bsmai。在一些实施方式中,核酸内切酶可以是限制性核酸内切酶。在这些实施方式中,可以修饰限制性核酸内切酶识别位点,使得限制性核酸内切酶仅在双链dna的一条链上切割磷酸二酯键,并在双链中产生切口。可以使用方法或策略来修饰限制性核酸内切酶的活性,例如包括未被限制酶切割的双链核酸的至少一条链中的化学修饰。该修饰的一个非限制性实例包括将一条链的磷酸二酯键的氧原子替换为硫原子。在提供限制性核酸内切酶的实施方式中,双链dna的一条链的磷酸二酯键可以被切割以在切割位点上产生5'侧的磷酸基和3'侧的羟基。限制性核酸内切酶的非限制性实例包括hincii、hindii、avai、fnu4hi、t1111和neil。该方法包括使样品与聚合酶接触。在一些实施方式中,聚合酶可以是具有链置换活性的dna聚合酶。在一些实施方式中,聚合酶可以是缺乏5'-3'核酸外切酶活性、缺乏3'-5'核酸外切酶活性或缺少5'-3'和3'-5'核酸外切酶活性的聚合酶。聚合酶可以来源于真核生物、原核生物或病毒,并且也可以进行遗传修饰。在一些实施方式中,聚合酶选自在较低温度(包括环境温度(例如室温))下起作用的那些。dna聚合酶的非限制性实例包括klenow片段、衍生自大肠杆菌的dna聚合酶i、衍生自嗜热脂肪芽孢杆菌(bacillusstearothermophilus)的5'至3'核酸外切酶缺陷型bstdna聚合酶和衍生自芽孢杆菌(bacilluscaldotenax)的5'至3'核酸外切酶缺陷型bcadna聚合酶。在图2a和2b中示出了本文公开的方法的一个非限制性实施方式。简言之,如图2a所示,样品在dna聚合酶和核酸内切酶存在下与scdna接触,所述核酸内切酶能够切割核酸内切酶识别位点(b)的双链形式(即互补序列)),或与核酸内切酶识别位点(b)的双链形式邻近的位点。如果样品中存在靶核酸(t),则靶核酸(t)的3'末端序列与scdna的序列(c)杂交,其与靶标互补并引发复制(通过反应混合物中存在的dna聚合酶),从而产生包含双链核酸内切酶识别位点(b)的双链扩增序列(x)。新产生的双链核酸内切酶识别位点(b)(通过反应混合物中存在的核酸内切酶)的识别和新产生的链(通过反应混合物中存在的核酸内切酶)随后的切割产生寡核苷酸信号序列(s)和扩增序列(x')。因为切口处的序列(x')的3'-oh作为后续的链置换复制的起始位点,所以寡核苷酸(s)通过在反应混合物中继续复制和扩增信号dna(s)的dna聚合酶从scdna中置换寡核苷酸(s)。如图2b中进一步所示,由信号扩增dna(或sadna)的存在可以进一步扩增由信号dna(s)的产生导致的信号。简言之,存在于反应中的信号dna(s)与引发或起始复制的sadna的序列(f)杂交(通过反应混合物中存在的dna聚合酶),从而产生双链扩增序列(y),其包括双链核酸内切酶识别位点(e)。新产生的双链核酸内切酶识别位点(e)(通过反应混合物中存在的核酸内切酶)的识别和新产生的链(通过反应混合物中存在的核酸内切酶)随后的切割产生寡核苷酸信号序列(s)和扩增序列(y')。因为切口处的序列(y')的3'-oh作为后续的链置换复制的起始位点,所以寡核苷酸(s)通过在反应混合物中继续复制和扩增信号dna(s)的dna聚合酶从sadna中置换寡核苷酸(s)。根据本发明的方法可用于检测样品中的靶核酸。特别地,本发明的方法可用于检测样品中的靶rna。如下所示,在核酸内切酶识别位点(b)和与靶核酸的3'末端互补的序列(c)之间的多dna间隔(pds)序列的scdna中的存在增强了信号dna(s)的产生。如下文所示,pds的存在增加了核酸内切酶切割核酸内切酶识别位点(b)的速率。根据本发明的方法可以在等温或基本上恒定的温度条件下进行。在涉及在基本上恒定的温度下进行该方法的实施方式中,允许温度的一些波动。例如,在一些实施方式中,基本上恒定的温度可以在期望的或识别的目标温度范围(例如约+/-2℃或约+/-5℃)内波动。在实施方式中,基本上恒定的温度可以包括不包括热循环的温度。在一些实施方式中,该方法可以在等温或基本上恒定的温度下进行,例如(1)等于或低于与scdna的序列(c)结合的靶核酸(t)的计算/预测或实验上确定的最佳杂交或退火温度的温度;(2)等于或低于与scdna结合的靶核酸(t)的解链温度(通常杂交或退火温度略低于解链温度)的温度;(3)低于与sadna结合的信号dna(s)的解链温度的温度;或(4)处于或约为反应混合物中存在的聚合酶和/或核酸内切酶的计算/预测或实验上确定的最佳反应温度的温度。该方法可以包括约20℃至约70℃的反应温度,包括落在约20℃至约42℃范围内的较低温度。在一些实施方式中,反应温度范围为35℃至40℃(例如35℃、36℃、37℃、38℃、39℃或40℃)。在其它实施方式中,反应温度低于65℃,包括低于约55℃、约50℃、约45℃、约40℃或约30℃的较低温度。在其它实施方式中,反应温度可为约20℃、约21℃、约22℃、约23℃、约24℃、约25℃、约26℃、约27℃、约28℃、约29℃、约30℃、约31℃、约32℃、约33℃、约34℃、约35℃、约36℃、约37℃、约38℃、约39℃、约40℃、约41℃、约42℃、约43℃、约44℃、约45℃、约46℃、约47℃、约49℃、约50℃、约51℃、约52℃、约53℃、约54℃、约55℃、约56℃、约57℃、约58℃、约59℃、约60℃、约61℃、约62℃、约63℃、约64℃、约65℃、约66℃、约67℃、约68℃、约69℃或约70℃。该方法可以进行一定时间,使得足以允许在靶核酸存在下扩增可检测量的信号序列。在一些实施方式中,反应时间可以为约5分钟至16小时,或约3分钟至16小时。在其它实施方式中,反应时间可以为约5至120分钟,或约15至60分钟。在整个说明书中,寡核苷酸(s)也称为信号dna(s)。因为信号dna仅在靶核酸(t)的存在下产生,所以根据本发明的方法通过检测信号dna的存在或不存在来检测样品中靶核酸(t)的存在或不存在。信号dna(s)不受序列限制,可以是可检测的任何序列。信号dna也不受长度限制。优选地,信号dna可以是约5至约100个碱基,以及5至100之间的任何整数。在一些实施方式中,信号dna可以是约5至约30个核酸碱基,以及5和30之间的所有整数。在一些实施方式中,信号dna的长度可以为约10至约30个碱基,以及10至30之间的所有整数。在另外的实施方式中,信号dna的长度可以为约15至约30个碱基,以及15和30之间的所有整数。根据本发明的方法可以在包含约4至约10或约7至约9的ph范围的缓冲条件下进行。缓冲液可以包含约10mm至约500mm或约50mm至150mm的盐浓度。在一些实施方式中,可以使用允许在靶核酸存在下扩增可检测量的信号序列的量的sc和/或sadna进行该方法。在一些实施方式中,sc和/或sadna浓度可以为约100pm至约100μm、约1nm至约1μm、约5nm至约50nm或约5nm至约25nm。信号dna(s)的存在可以通过本领域已知的任何方法检测。例如,可以使用凝胶电泳和用溴化乙锭染色。此外,信号dna的存在可以使用荧光偏振、免疫测定、荧光共振能量转移、酶标记(例如过氧化物酶或碱性磷酸酶)、荧光标记(例如荧光素或罗丹明)、化学发光、生物发光、表面等离子体共振(spr)或荧光团修饰的探针dna(例如taqman探针)来检测。扩增产物也可以通过使用例如用生物素标记的标记核苷酸来检测。在这种情况下,例如可以使用荧光标记的抗生物素蛋白或酶标记的抗生物素蛋白来检测扩增产物中的生物素。也可以通过使用本领域技术人员已知的氧化还原嵌入剂用电极检测扩增产物。也可以使用表面等离子体共振(spr)、夸克晶体微量天平(qcm)或电化学方法(包括使用纳米孔传感器的那些方法)来检测扩增产物。根据本发明的方法检测样品中靶核酸(t)的存在或不存在。根据本发明的方法还可以用于定量测量测试样品中靶核酸的浓度。例如,根据本发明的方法可以在靶核酸的不同已知浓度范围的存在下进行,然后可以如本领域通常实践的那样制备和使用校准曲线。靶核酸(图2中的(t))可以包含任何核酸序列,并且可以包括dna、rna、化学修饰的核酸、非天然核酸、核酸类似物或任何杂交体或其组合。因此,在一些实施方式中,dna可以包括cdna、基因组dna和合成的dna,并且rna可以包括总rna、mrna、rrna、sirna、hnrna、pirna、arna、mirna和合成的rna。虽然一些实施方式涉及特定靶核酸序列,但任何核酸序列(包括辅核酸序列)可以是待检测的靶核酸序列。本发明提供了即使当核酸是短链核酸时也具有选择性和灵敏性的检测靶核酸。因此,scdna的序列(c)与靶核酸(t)之间的互补度允许序列之间的特异性杂交(例如序列转换dna的序列(c)和靶核酸(t)序列中互补核苷酸数避免了在给定组的反应条件下的非特异性杂交)。在实施方式中,靶核酸序列可来自或衍生自任何数量的来源,包括例如基因组dna、表达mrna、来自病原体的核酸序列(微生物、病毒)或治疗性核酸。因此,sc和sadna以及本文公开的方法可用于疾病(例如由遗传和感染源产生)的诊断和预后,污染物(例如食源性疾病、设备污染)的鉴定和个体化用药(例如治疗的监测和/或预后)等。例如,可以针对以下感染性疾病进行分子诊断测试:乙型肝炎病毒(hbv);丙型肝炎(hcv);hcv(基因型1-6);人类免疫缺陷病毒1型(hiv-1);沙眼衣原体;淋病奈瑟氏球菌;甲型流感;乙型流感;呼吸道合胞病毒(rsv);和parvo病毒。在一些实施方式中,靶核酸可以包含微rna(mirna)。微rna包括约22个核苷酸的小型非编码rna分子。已知微rna在基因表达的转录和转录后调控中起作用。已知微rna通过与信使rna(mrna)的互补区域的碱基配对而起作用,通过翻译抑制或靶标降解导致基因沉默。可以包含靶核酸的任何类型的样品可用于本文公开的方法中。因此,含有或怀疑含有靶核酸的样品没有特别限制,例如包括来自生命体的生物样品,例如全血、血清、血沉棕黄层、尿液、粪便、脑脊髓液、精液、唾液、组织(例如癌组织或淋巴结)、细胞培养物(例如哺乳动物细胞培养物或细菌培养物);含有核酸的样品,例如类病毒、病毒、细菌、真菌、酵母、植物和动物;可能含有或被微生物例如病毒或细菌感染的样品(例如食品和生物制剂);以及可能含有生物物质的样品,例如土壤、工业过程和制造设备以及废水;和来自各种水源(例如饮用水)的样品。此外,可以通过任何已知方法处理样品以制备本文公开的方法中使用的含有核酸的组合物。这种制剂的实例可以包括细胞破裂(例如细胞裂解物和提取物)、样品馏分、样品中的核酸和特异性核酸分子基团例如富含mrna的样品。用于检测本发明的靶核酸的方法中的样品不限于如上所述的衍生自生物和天然产物的样品,并且可以是含有合成寡核苷酸的样品。根据本发明的方法可以与abbottm2000sp样品制备系统结合进行。m2000sp使用磁粉技术捕获核酸并洗涤颗粒以除去未结合的样品组分。将结合的核酸洗脱并转移到96深孔板中。abottm2000sp还可以将转移到96深孔板的洗涤后的核酸与根据本技术进行方法所需的任何试剂结合。例如,可以根据需要或期望添加sc和sadna、聚合酶、核酸内切酶、分子信标和任何其它试剂(例如dntp)。根据本发明的方法也可以与即时平台接合。例如,将三磷酸脱氧核糖核苷酸(dntp)并入生长的dna链中涉及共价键的形成和焦磷酸盐的释放以及影响反应ph的带正电荷的氢离子。因此,根据本发明的方法的信号dna的合成可以通过使用例如即时微量ph计跟踪ph的变化来检测。例如,abotti-stat即时系统可以提供一次性的一次性药筒,其中包含微型制造的传感器、校准溶液、流体系统和用于ph分析的废物室。本文公开的方法可以包含另外的试剂。可用于核酸扩增反应的其它试剂的一些非限制性实例包括金属盐例如氯化钠、氯化镁、乙酸镁和硫酸镁;底物例如dntp混合物;和缓冲液例如tris-hcl缓冲液、三氯缓冲液、磷酸钠缓冲液和磷酸钾缓冲液。同样,洗涤剂、氧化剂和还原剂也可用于本文公开的方法的实践中。此外,可以使用例如二甲基亚砜和甜菜碱(n,n,n-三甲基甘氨酸);国际公开号wo99/54455中描述的酸性物质;和阳离子络合物的试剂。本文提供的方法和核酸结构可以与其它方法组合使用,以在靶核酸存在下提供信号dna的指数扩增。例如,根据本发明的方法和组合物可以与覆盖序列转换dna组合使用,如名称为“覆盖序列转换dna和检测方法”的美国临时申请61/927710中所述,其通过引用并入本文。术语“约”通常是指本领域技术人员将认为等同于所引用的值(即具有相同的功能或结果)的数量范围。如本文所用,术语“约”旨在表示大于或小于参考值的约10-20%的范围。在某些情况下,本领域技术人员将认识到,由于参考值的性质,术语“约”可以意味着与该值的偏差大于或小于10-20%。以下实施例旨在说明上述方面和实施方式。上述公开和以下实施例都不应视为限制所附权利要求的范围。本领域技术人员将理解,本发明不受用于描述和示出本发明的各个方面的特定术语的限制。实施例1以下实施例证明位于scdna的内切核酸酶识别位点(b)和序列(c)之间的pds序列的存在加速了信号dna的扩增。使用scdna1:5'-agccctgtacaatgccctcagcctgttcctgctgaactgagcca-tamra-3'(序列号1),和在scdna的内切核酸酶识别位点(b)和序列(c)之间具有不同pds序列的多个其它scdna进行反应。这些scdna包括:在37℃下在含有终浓度为10mmtris-hcl、50mmnacl、10mmmgcl2、1mmdtt、ph7.9的纽英伦生物技术(neb)缓冲液2的25μl反应体积中进行反应。在反应中使用的切口内切核酸酶是nb.bbvci,其以0.1单位/μl的浓度存在。用于反应的聚合酶是bstdna聚合酶大片段,其以0.08单位/μl的浓度存在。dntp的终浓度各为200μm。scdna以200nm的终浓度存在于反应中。靶核酸是人微rna24-3p(序列号5),并且以1nm或100pm的浓度存在。以100nm的终浓度存在的分子信标探针用于检测信号dna的产生。使用bio-rad实时pcr系统cfx96进行荧光测量,结果示于图3中。如图所示,位于scdna的内切核酸酶识别位点(b)和序列(c)之间的pds序列存在加速了信号dna的扩增。实施例2以下实施例证明位于scdna内的内切核酸酶识别位点(b)和与靶核酸互补的序列(c)之间的pds序列的存在加速了内切核酸酶识别位点(b)的核酸内切酶切割。使用以下scdna:5'-agccctgtacaatgccctcagcctgttcctgctgaactgagcca-idt-idt'(序列号6)进行反应。使用许多不同的靶核酸,其包括以下:核酸序列号序列dna#1875'tggctcagttcagcaggaacagrna#655'uggcucaguucagcaggaacagrna#785'uggcucaguucagcaggaarna#895'uggcucaguucagcag在37℃下在含有终浓度为10mmtris-hcl、50mmnacl、10mmmgcl2、1mmdtt、ph7.9的纽英伦生物技术(neb)缓冲液2的50μl反应体积中进行第一聚合反应。用于反应的聚合酶是bstdna聚合酶大片段,其以0.08单位/μl的浓度存在。dntp的终浓度各为200μm。序列转换dna以200nm存在。靶核酸(dna#18、rna#6、7或8)以200nm存在。将聚合反应在37℃下保温10分钟,然后在80℃温育20分钟,然后将反应物移至4℃。然后将12.5μl上述聚合反应转移到内切核酸酶反应中。在37℃下在含有终浓度为10mmtris-hcl、50mmnacl、10mmmgcl2、1mmdtt、ph7.9的纽英伦生物技术(neb)缓冲液2的25μl反应体积中进行核酸内切酶反应。在反应中使用的切口内切核酸酶是nb.bbvci,其以0.1单位/μl的浓度存在。将反应物在37℃下培养0和5分钟。通过加入5μm的0.5medta终止反应,然后在95℃下培养。使用分子信标探针检测信号dna的产生,并在12%tbe-page上显现反应产物。(参见图4(箭头表示由nb.bbvci内切核酸切割产生的小片段的位置))。如图所示,当使用rna#6时,不产生短片段,表明在scdna的内切核酸酶识别位点(b)和与靶核酸互补的序列(c)中存在pds序列加速了核酸内切酶识别位点(b)的核酸内切酶切割。虽然已经参考某些方面和实施方式描述了应用,但是本领域技术人员将理解,可以对本文提供的公开内容进行改变,并且可以替换等同,而不脱离本发明的范围。因此,本申请不应该限于所公开的特定方面和实施方式,而应理解和意识为包括落在所附权利要求的范围内的所有方面和实施方式。对说明书的修改在第1页在标题之后,请插入以下段落:本申请要求于2014年10月14日提交的美国临时申请62/063,666的优先权以及于2014年12月30日提交的美国临时申请62/098,066的优先权。该美国临时申请均通过引用将其其全部并入本文。序列表<110>kenkomiyamakotokomoritoruyoshimura<120>具有多dna间隔序列的序列转换和信号扩增dna及使用其的检测方法<130>28298us01<160>9<170>patentin版本3.5<210>1<211>44<212>dna<213>人工序列<220><223>序列转换dna(scdna1)<220><221>尚未归类的特征<222>(44)..(44)<223>3'-末端修饰:tamra<400>1agccctgtacaatgccctcagcctgttcctgctgaactgagcca44<210>2<211>47<212>dna<213>人工序列<220><223>序列转换dna(具有t3间隔的scdna)<220><221>尚未归类的特征<222>(47)..(47)<223>3'-末端修饰:tamra<400>2agccctgtacaatgccctcagctttctgttcctgctgaactgagcca47<210>3<211>48<212>dna<213>人工序列<220><223>序列转换dna(具有t4间隔的scdna)<220><221>尚未归类的特征<223>3'-末端修饰:tamra<400>3agccctgtacaatgccctcagcttttctgttcctgctgaactgagcca48<210>4<211>49<212>dna<213>人工序列<220><223>序列转换dna(具有t5间隔的scdna)<220><221>尚未归类的特征<222>(49)..(49)<223>3'-末端修饰:tamra<400>4agccctgtacaatgccctcagctttttctgttcctgctgaactgagcca49<210>5<211>22<212>rna<213>人工序列<220><223>人微rna24-3p<400>5uggcucaguucagcaggaacag22<210>6<211>44<212>dna<213>人工序列<220><223>序列转换dna<220><221>尚未归类的特征<223>3'-末端修饰:两个倒置的胸腺嘧啶<400>6agccctgtacaatgccctcagcctgttcctgctgaactgagcca44<210>7<211>22<212>dna<213>人工序列<220><223>用于人微rna24-3p的dna序列<400>7tggctcagttcagcaggaacag22<210>8<211>19<212>rna<213>人工序列<220><223>人微rna24-3p<400>8uggcucaguucagcaggaa19<210>9<211>16<212>rna<213>人工序列<220><223>人微rna24-3p<400>9uggcucaguucagcag16当前第1页12
- 该技术已申请专利。仅供学习研究,如用于商业用途,请联系技术所有人。
- 技术研发人员:小宫健;小森诚;吉村徹
- 技术所有人:雅培日本有限公司;国立大学法人东京工业大学
- 我是此专利的发明人
- 上一篇:一种氧气盒子的制造方法与工艺
- 上一篇:一种节能环境干燥设备的制造方法与工艺
- 该领域下的技术专家
- 如您需求助技术专家,请点此查看客服电话进行咨询。
- 1、薛老师:1.CRISPR-Cas系统 2.基因编辑 3.基因修复 4.天然产物合成 5.单分子技术开发与应用
- 2、张老师:1.探索新型氧化还原酶结构-功能关系,电催化反应机制 2.酶电催化导向的酶分子改造 3.纳米材料、生物功能多肽对酶-电极体系的影响4. 生物电化学传感和生物电合成体系的设计与应用。
- 3、豆老师:1.环境纳米材料及挥发性有机化合物(VOCs) 2.CO污染物的催化氧化 3.低温等离子体 4.吸脱附等控制技术
- 4、赵老师:1.高分子材料改性及加工技术 2.微孔及过滤材料 3.环境友好高分子材料
- 5、邬老师:1.高分子材料的共混与复合 2.涉及材料功能化及结构与性能的研究; 高分子热稳定剂的研发
- 如您是高校老师,可以点此联系我们加入专家库。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
精彩留言,会给你点赞!