重组李斯特菌疫苗菌株及其用于癌症免疫疗法的方法与流程
本发明提供治疗、防御人乳头瘤病毒相关肿瘤或癌症以及诱导对人乳头瘤病毒相关肿瘤或癌症的免疫应答的方法,该方法包括向受试者施用表达人乳头瘤病毒抗原的重组李斯特菌的步骤。
背景技术:
人乳头瘤病毒(hpv)相关口咽癌(hpvopc)在美国日益流行(从1988年到2004年增加225%)。hpvopc患者倾向于年轻化并且预后良好,与hpv阴性患者相比,死亡风险低69%。然而,大多数hpvopc患者都已是晚期,而标准的化疗方案可导致严重的毒性。因此,具有良好预后的患者自相矛盾地处于更高的治疗相关长期不良生活质量结局的风险中。免疫疗法通过逐渐减弱化疗方案而具有降低毒性的潜力,并可能增强长期疾病控制。
hr-hpve6和e7蛋白在发育异常和癌中一致表达,分别破坏细胞周期调控蛋白p53和prb。发育异常和浸润性恶性病变对e6和e7的强制性表达以及这些蛋白的病毒性来源使得它们成为hpv治疗性疫苗的优异靶标。
单核细胞增多性李斯特菌(listeriamonocytogenes)(lm)是一种食源性革兰氏阳性菌,偶尔会在人类尤其是老年人、新生儿、孕妇和免疫低下个体中导致疾病。除了强烈激活先天免疫并诱导增强抗原呈递细胞(apc)功能的细胞因子应答外,lm还能够在从吞噬溶酶体逃离后在apc的胞质溶胶中复制,这主要是通过李斯特菌溶血素o(llo)蛋白的作用。这种独特的胞内生命周期使得由lm分泌的抗原可在mhci类和ii类分子的背景下加工和呈递,从而导致强效的细胞毒性cd8+和th1cd4+t细胞介导的免疫应答。
本发明通过提供综合疗法来解决具有人乳头瘤病毒(hpv)相关口咽癌的患者中与治疗相关的长期不良生活质量结局这一问题,该疗法包括单核细胞增多性李斯特菌免疫疗法,从而通过逐渐减弱化疗方案而降低毒性,并可能增强长期疾病控制。
技术实现要素:
在一个方面,本发明涉及一种诱导人受试者中的抗肿瘤或抗癌症免疫应答的方法,该方法包括向所述受试者施用包含重组李斯特菌菌株的组合物的步骤,所述重组李斯特菌菌株包含重组核酸,所述核酸包含第一开放阅读框,所述第一开放阅读框编码重组多肽,所述重组多肽包含融合至异源性抗原或其片段的llo蛋白的n末端片段,其中所述重组核酸还包含第二开放阅读框,所述第二开放阅读框编码突变体prfa基因或代谢酶,从而诱导对肿瘤或癌症的免疫应答。在另一个实施例中,该免疫应答降低所述受试者接受化疗或放疗的需要。在另一个实施例中,所述免疫应答消除所述受试者接受化疗或放疗的需要。在另一个实施例中,所述免疫应答通过允许用较低剂量的化疗或放疗治疗患者而降低与向所述受试者施用化疗或放疗相关的副作用的严重性。
在另一方面,所述免疫应答使得疾病的分期能够下降,以使得更保守的治疗选择变得可用。
本发明的其他特征和优点将由于以下详细描述的实例和附图而变得显而易见。但是,应当理解,该详细描述和具体实例尽管指出了本发明的优选实施例,但是仅仅通过例证给出,因为本发明的精神和范围内的各种变化和修改对阅读了该详细描述的本领域技术人员而言将是显而易见的。
附图说明
以下附图形成了本说明书的一部分,将它们包括进来以进一步说明本公开的某些方面,通过参考一个或多个这些附图并结合本文提供的对具体实施例的描述可更好地理解本发明。本专利或申请文件包含至少一个用彩色表现的附图。根据请求并支付必要的费用后,专利局将提供带有彩色附图的本专利或专利申请公开的副本。
图1.lm-e7和lm-llo-e7使用不同的表达系统来表达和分泌e7。通过将基因盒引入单核细胞增多性李斯特菌基因组的orfz域中来产生lm-e7(a)。hly启动子驱动hly信号序列和llo的前五个氨基酸(aa)以及随后的hpv-16e7的表达。b)通过以质粒pgg-55转化prfa-菌株xfl-7来产生lm-llo-e7。pgg-55具有驱动llo-e7非溶血性融合体的表达的hly启动子。pgg-55也含有prfa基因,以选择xfl-7在体内对质粒的保留。
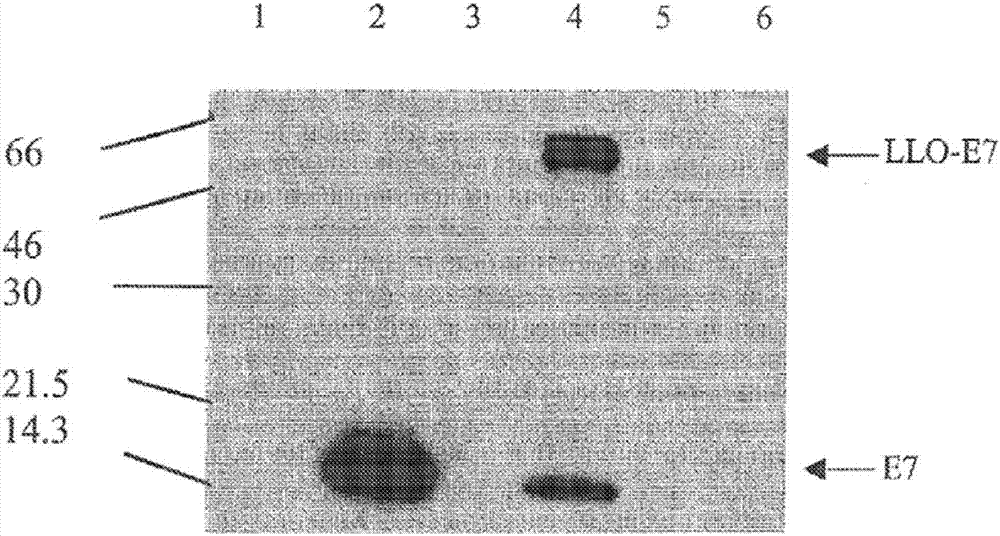
图2.lm-e7和lm-llo-e7分泌e7。使lm-gag(泳道1)、lm-e7(泳道2)、lm-llo-np(泳道3)、lm-llo-e7(泳道4)、xfl-7(泳道5)和10403s(泳道6)在luria-bertoni液体培养基中于37℃生长过夜。沉淀相等数目的细菌(通过600nm处吸光度od确定),并将18ml的每种上清液进行tca沉淀。通过western印迹分析e7表达。用抗e7mab、随后用hrp-偶联的抗小鼠(amersham)探测该印迹,接着使用ecl检测试剂显影。
图3.llo-e7融合体的肿瘤免疫抑制功效。显示了肿瘤接种后第7、第14、第21、第28和第56天小鼠中以毫米计的肿瘤大小。未暴露的小鼠:空心圆圈;lm-llo-e7:实心圆圈;lm-e7:正方形;lm-gag:空心菱形;和lm-llo-np:实心三角形。
图4.来自lm-llo-e7-免疫的小鼠的脾细胞在暴露于tc-1细胞时增殖。将c57bl/6小鼠用lm-llo-e7、lm-e7或对照rlm菌株免疫和强化。在强化后6天收获脾细胞,并以显示的比率与经照射的tc-1细胞一起铺板。将细胞用3h胸苷脉冲处理并收获。将cpm定义为(实验cpm)-(无tc-1对照)。
图5.a.在施用tc-1肿瘤细胞并随后施用lm-e7、lm-llo-e7、lm-acta-e7或无疫苗(未暴露的)的小鼠中,脾中e7特异性的分泌ifn-γ的cd8+t细胞的诱导和浸润肿瘤的数目。b.针对(a)描述的小鼠的脾和肿瘤中e7特异性的cd8+细胞的诱导和浸润。
图6.含有pest区域的李斯特菌构建体在肿瘤中诱导更高百分比的e7特异性淋巴细胞。a.来自1个实验的代表性数据。b.来自所有3个实验的数据的平均值和se。
图7a.传代对重组李斯特菌疫苗载体的细菌负荷(毒力)的作用。上图:lm-gag下图:lm-llo-e7图7b.传代对脾脏中重组lm-e7的细菌负荷的作用。描绘了来自四只小鼠的每毫升脾匀浆的平均活细菌cfu。
图8显示了在施用经传代的lm-gag与未传代的lm-gag后对于hiv-gag和llo而言的抗原特异性cd8+t细胞的诱导。将小鼠用103个(a、b、e、f)或105个(c、d、g、h)cfu的经传代的李斯特菌疫苗载体免疫,并分析抗原特异性t细胞。b、d、f、h:未传代的李斯特菌疫苗载体。a-d:对mhci类hiv-gag肽的免疫应答。e-h:对llo肽的免疫应答。i:来自用105个cfu经传代的lm-gag免疫的小鼠的脾细胞用来自hpve7的对照肽刺激。
图9a显示了在整个lb稳定性研究中的质粒分离。图9b显示了在整个tb稳定性研究中的质粒分离。图9c显示了tb稳定性研究的定量。
图10显示了来自在lb中生长的细菌的活细菌氯霉素(cap)抗性和cap敏感性集落形成单位(cfu)的数量。深色条:cap+;白色条:cap-。每个时间点的两个深色条和两个白色条代表重复样。
图11显示了来自在tb中生长的细菌的活细菌cap抗性和cap敏感性cfu的数量。深色条:cap+;白色条:cap。每个时间点的两个深色条和两个白色条代表重复样。
图12.显示了d133v突变区(箭头)的实际色谱图。混合物比率在括号中示出。
图13.adv451、452和453引物以及在反应中扩增的prfa基因片段的位置的表示。
图14.使用引物adv451和adv453的pcr反应的特异性。
图15.使用引物adv452和adv453的pcr反应的特异性。
图16.使用引物adv452和1ng初始量的dna进行pcr反应来检测野生型prfa序列的灵敏度。
图17.使用引物adv452和5ng初始量的dna进行pcr反应来检测野生型prfa序列的灵敏度。
图18.来自图16中描绘的pcr的条带的平均密度。
图19.来自图17中描绘的pcr的条带的平均密度。
图20.对使用引物adv452进行pcr反应来检测野生型prfa序列进行的验证。
图21.来自图16中描绘的pcr的条带的平均密度。
图22.对lm-llo-e7中的d133vprfa突变的分析。a,用于密度测量技术的原始图像;b,对图像进行数字增强以有利于观察低密度条带。
图23.显示了用于施用lm-llo-e7疫苗(adxs-hpv)、用于样品(肿瘤组织或血液)采集以及用于执行各种测定的试验方案。
图24.对肿瘤样品的苏木精和伊红(h&e)染色,显示了嗜碱性淋巴细胞样浸润巢(白色箭头)。
图25.adxs-hpv疫苗接种后的肿瘤样品的多重免疫荧光,显示了致密的瘤内cd8浸润和基质cd4浸润。绿色=cd4、粉色=cd8、紫色=cd68、黄色=cd20。
图26.elispot–直接分析(无再刺激和培养),显示了adxs-hpv疫苗接种后hpv-e7应答的增加>3倍。
图27.elispot–直接分析(无再刺激和培养),显示了采用非疫苗抗原hpv-e6和hpv-e2的情况下不存在疫苗接种后的应答增加。
图28a-b.ii期临床试验设计。图28a显示了研究中的各流程和样品采集的总体时间轴(mo,月;pbmc,外周血单核细胞;tors,经口腔机器人手术;wk,周)。图28b显示了各个hpvopc患者的流程和样品采集计划。第一列中的数字表示各个患者。v1和v2样品在疫苗接种前抽取,v3在第一与第二疫苗施用之间抽取,而v4样品在手术当天抽取。样品v5-v9在指定的术后时间的随访从患者抽取。
图29.elispot-直接分析,显示了adxs-hpv疫苗接种后系统性hpv-e6和hpve7应答增加。第一列指定elispot目标。空:无肽刺激,e2:hpve2肽刺激(证实总体而言对hpv的t细胞应答),e6和e7:hpve6和e7肽分别刺激(与hpv发育异常相关的早期肽基因);p/i:阳性对照。
图30a-d.ifn-γ细胞内细胞因子染色(ics)测定的概要,显示了在adxs-hpv疫苗接种后e6和e7特异性ifn-γ分泌性cd4+和cd8+t细胞的诱导。图30a显示了e7特异性ifn-γ分泌性cd4+t细胞的诱导。图30b显示了e7特异性ifn-γ分泌性cd8+t细胞的诱导。图30c显示了e6特异性ifn-γ分泌性cd4+t细胞的诱导。图30d显示了e6特异性ifn-γ分泌性cd8+t细胞的诱导。
图31a-d.ifn-α细胞内细胞因子染色(ics)测定的概要,显示了在adxs-hpv疫苗接种后e6-和e7特异性ifn-α分泌性cd4+和cd8+t细胞的诱导。图31a显示了e7特异性tnf-α分泌性cd4+t细胞的诱导。图31b显示了e7特异性tnf-α分泌性cd8+t细胞的诱导。图31c显示了e6特异性tnf-α分泌性cd4+t细胞的诱导。图31d显示了e6特异性tnf-α分泌性cd8+t细胞的诱导。
图32a-c.hpv特异性应答的概要。各表格第一列中的数字表示各个患者。图32a显示了在adxs-hpv疫苗接种后对e6或e7有响应的患者。图32b显示了显示出e7应答的患者。图32c显示了显示出e6应答的患者。使用以下测试来测量应答:exvivoeli、ifngeli、ifn-γelispot、ifngics、ifn-γ细胞内细胞因子染色、tnfaics、tnf-α细胞内细胞因子染色。
图33.患者肿瘤的多重免疫荧光。图33a显示了疫苗接种前的肿瘤环境(tumormilieu)。图33b显示了疫苗接种前的肿瘤。图33c显示了adxs-hpv疫苗接种后肿瘤环境的致密的瘤内cd8浸润。图33d显示了adxs-hpv疫苗接种后减小的肿瘤大小。红色=cd8、黄色=cd3、品红色=pd-1、绿色=pd-l1。
图34a-f.患者肿瘤的多重免疫荧光。图34a-c显示了疫苗接种前肿瘤环境的三个独立样品。图34d-f显示了在adxs-hpv疫苗接种前后在肿瘤环境中pd-l1表达的抑制和pd-1表达性细胞的浸润。红色=cd8、黄色=cd4、品红色=pd-1、绿色=pd-l1。
图35a-h.免疫荧光结果的初步定量,显示了adxs-hpv疫苗接种前后cd4、cd8、pd-1和pd-l1的水平。图35a显示了在七个单独的样品中观测到的cd4水平的变化。图35b显示了在七个单独的样品中观测到的cd8水平的变化。图35c显示了在七个单独的样品中观测到的pd-1水平的变化。图35c显示了在七个单独的样品中观测到的pd-l1水平的变化。图35e显示了图35a中的样品队列的cd4的平均变化。图35f显示了图35b中的样品队列的cd8的平均变化。图35g显示了图35c中的样品队列的pd-1的平均变化。图35h显示了图35d中的样品队列的pd-l1的平均变化。
图36.疫苗接种后在adxs-hpv疫苗接种患者的样品中pd-1和pd-l1的相对水平。pd-1的水平沿x轴绘图。pd-l1的水平沿y轴绘图。
图37.疫苗接种前后在adxs-hpv疫苗接种患者的样品中cd8+细胞水平的相对变化。疫苗接种后的cd8+细胞水平沿x轴绘图。疫苗接种前的cd8+细胞水平沿y轴绘图。
应当理解,为了阐述的简洁和清楚,显示在图中的要素未必是按比率绘制的。例如,为了清楚,一些要素的尺寸可以是相对其他要素而放大的。另外,在认为适合时,附图标记可以在附图之间重复,以指出对应的或类似的要素。
具体实施方式
本发明提供治疗疾病、防御疾病以及诱导对疾病的免疫应答的方法,该方法包括向受试者施用重组李斯特菌菌株的步骤,所述菌株包含融合肽,所述融合肽包含李斯特菌溶血素o(llo)片段和所述疾病表达的异源性抗原或其片断。
在一个实施例中,本发明提供一种诱导人受试者中的抗肿瘤或抗癌症免疫应答的方法,该方法包括向所述受试者施用包含重组李斯特菌菌株的组合物的步骤,所述重组李斯特菌菌株包含重组核酸,所述核酸包含第一开放阅读框,所述第一开放阅读框编码重组多肽,所述重组多肽包含融合至异源性抗原或其片段的llo蛋白的n末端片段,其中所述重组核酸还包含第二开放阅读框,所述第二开放阅读框编码突变体prfa基因或代谢酶,从而诱导对肿瘤或癌症的免疫应答。在另一个实施例中,所述免疫应答降低所述受试者接受化疗或放疗的需要。在另一个实施例中,所述免疫应答消除所述受试者接受化疗或放疗的需要。在另一个实施例中,所述免疫应答通过允许用较低剂量的化疗或放疗治疗患者而降低与向所述受试者施用化疗或放疗相关的副作用的严重性。
在一个实施例中,本发明提供一种诱导人受试者中的抗肿瘤或抗癌症免疫应答的方法,该方法包括向所述受试者施用包含重组李斯特菌菌株的组合物的步骤,所述重组李斯特菌菌株包含重组核酸,所述核酸包含第一开放阅读框,所述第一开放阅读框编码重组多肽,所述重组多肽包含融合至异源性抗原或其片段的llo蛋白的n末端片段,其中所述重组核酸还包含第二开放阅读框,所述第二开放阅读框编码代谢酶,其中所述免疫应答降低所述受试者接受化疗或放疗的需要,从而诱导对肿瘤或癌症的免疫应答。在另一个实施例中,所述李斯特菌在内源性dal/dat和acta基因中包含突变。
在一个实施例中,本文所公开的核酸分子包含编码重组多肽的第一开放阅读框,该重组多肽包含异源性抗原或其片段。在另一个实施例中,该重组多肽还包含融合至异源性抗原的n末端llo。在另一个实施例中,本文所公开的核酸分子还包含编码代谢酶的第二开放阅读框。在另一个实施例中,该代谢酶补充重组李斯特菌菌株的染色体中缺乏的内源性基因。在另一个实施例中,由该第二开放阅读框编码的代谢酶为丙氨酸消旋酶(dal)。在另一个实施例中,由该第二开放阅读框编码的代谢酶为d-氨基酸转移酶(dat)。在另一个实施例中,本文所公开的李斯特菌菌株在基因组dal、dat或acta基因中包含突变、缺失或失活。在另一个实施例中,本文所公开的李斯特菌菌株在基因组dal、dat和acta基因中包含突变、缺失或失活。在另一个实施例中,该李斯特菌缺乏基因组dal、dat或acta基因。在另一个实施例中,该李斯特菌缺乏基因组dal、dat和acta基因。
在另一个实施例中,施用本文所公开的李斯特菌或本文所公开的基于李斯特菌的免疫疗法能够降低具有肿瘤或癌症的受试者接受化疗或放疗的需要。在另一个实施例中,施用本文所公开的李斯特菌或本文所公开的基于李斯特菌的免疫疗法能够消除具有肿瘤或癌症的受试者接受化疗或放疗的需要。在另一个实施例中,施用本文所公开的李斯特菌或本文所公开的基于李斯特菌的免疫疗法能够减轻具有肿瘤或癌症的受试者中与放疗或放疗相关的副作用的严重性。本发明还提供诱导人受试者中的抗疾病细胞毒性t细胞(ctl)应答并治疗与所述疾病相关的障碍和症状的方法,该方法包括施用重组李斯特菌菌株。在一个实施例中,本文公开一种重组李斯特菌菌株,所述重组李斯特菌菌株包含重组核酸,所述核酸包含第一开放阅读框,所述第一开放阅读框编码重组多肽,所述重组多肽包含融合至异源性抗原或其片段的llo蛋白的第一n末端片段,并且其中所述重组核酸还包含第二开放阅读框,所述第二开放阅读框编码突变体prfa基因。在一个实施例中,突变体prfa基因是编码氨基酸位置133处从氨基酸d(也称为“asp”、“天冬氨酸盐”或“天冬氨酸”)到氨基酸v(也称为“val”或“缬氨酸”)的点突变的基因。在一个实施例中,本文所公开的重组李斯特菌菌株在内源性prfa基因中包含突变或缺失。在另一个实施例中,在本文所公开的李斯特菌的prfa基因中的染色体突变或缺失经由质粒来补充,所述质粒包含编码突变体prfa基因的核酸序列,所述基因编码包含d133v氨基酸置换的突变体prfa蛋白。在另一个实施例中,包含d133v氨基酸置换的突变体prfa蛋白补充本文所公开的李斯特菌中的内源性prfa突变。
在另一个实施例中,该重组李斯特菌是减毒的李斯特菌。将认识到,术语“减毒”或“减毒的”可涵盖经过修饰以降低对宿主毒性的细菌、病毒、寄生虫、感染性生物体、朊病毒、肿瘤细胞、感染性生物体中的基因等。宿主可以是人或动物,或器官、组织或细胞。以非限制性方式举例,细菌可被减毒以减少与宿主细胞的结合,减少从一个宿主细胞向另一宿主细胞的传播,减少胞外生长,或减少在宿主细胞中的细胞内生长。在一个实施例中,减毒可通过测量例如毒性的一个或多个指标、ld50、从器官中的清除率或竞争指数而评估(参见例如auerbuch等人(2001)infect.immunity69:5953-5957)。通常,减毒导致ld50增加和/或清除率增加至少25%;更通常增加至少50%;最通常增加至少100%(2倍);正常增加至少5倍;更正常增加至少10倍;最正常增加至少50倍;经常增加至少100倍;更经常增加至少500倍;并且最经常增加至少1000倍;一般增加至少5000倍;更一般增加至少10,000倍;并且最一般增加至少50,000倍;并且最经常增加至少100,000倍。在另一个实施例中,减毒导致ld50增加和/或清除率增加至少25%。在另一个实施例中,减毒导致ld50增加和/或清除率增加3-5倍。在其他实施例中,减毒导致ld50增加和/或清除率增加5-10倍、11-20倍、21-30倍、31-40倍、41-50倍、51-100倍、101-500倍、501-1,000倍、1001-10,000倍或10,001-100,000倍。
技术人员将容易认识到,术语“减毒基因”可涵盖介导对宿主的毒性、病理或毒力、在宿主内生长或在宿主内存活的基因,其中该基因以减轻、降低或消除毒性、病理或毒力的方式突变。可通过比较由突变基因介导的毒力或毒性与由非突变(或亲本)基因介导的毒力或毒性来评估所述降低或消除。“突变基因”涵盖基因调控区、基因编码区、基因非编码区或其任意组合中的缺失、点突变、倒位、截短和移码突变。
在一个实施例中,本文公开一种诱导人受试者中对肿瘤或癌症的免疫应答的方法,该方法包括向所述受试者施用包含重组核酸的重组李斯特菌菌株的步骤,所述核酸包含第一开放阅读框,所述第一开放阅读框编码重组多肽,所述重组多肽包含融合至异源性抗原或其片段的llo蛋白的n末端片段,其中所述重组核酸还包含第二开放阅读框,所述第二开放阅读框编码突变体prfa蛋白,从而诱导对肿瘤或癌症的免疫应答。在一个实施例中,本发明提供一种治疗人受试者中的癌症的方法,该方法包括向所述受试者施用本文所公开的重组李斯特菌菌株的步骤。在另一个实施例中,本发明提供一种避免人受试者罹患宫颈癌的方法,该方法包括向所述受试者施用本文所公开的重组李斯特菌菌株的步骤。在另一个实施例中,该重组李斯特菌菌株表达重组多肽。在另一个实施例中,该重组李斯特菌菌株包含编码重组多肽的质粒。在另一个实施例中,该方法还包括用本发明的重组李斯特菌菌株强化人受试者的步骤。在另一个实施例中,该方法还包括用包含本文所公开的异源性抗原或其片段的免疫原性组合物强化人受试者的步骤。在另一个实施例中,该方法还包括用免疫原性组合物强化人受试者的步骤,该组合物引导受试者的细胞表达该异源性抗原。在另一个实施例中,细胞是肿瘤细胞。在另一个实施例中,该方法还包括用本发明的疫苗强化人受试者的步骤。
在一个实施例中,在本文所公开的llo蛋白和acta蛋白的背景下的片段是指包含llo或acta蛋白的至少5个连续氨基酸残基的氨基酸序列的肽或多肽。在另一个实施例中,该术语是指包含llo或acta蛋白或多肽的至少10个连续氨基酸残基、至少15个连续氨基酸残基、至少20个连续氨基酸残基、至少25个连续氨基酸残基、至少40个连续氨基酸残基、至少50个连续氨基酸残基、至少60个连续氨基酸残基、至少70个连续氨基酸残基、至少80个连续氨基酸残基、至少90个连续氨基酸残基、至少100个连续氨基酸残基、至少125个连续氨基酸残基、至少150个连续氨基酸残基、至少175个连续氨基酸残基、至少200个连续氨基酸残基、至少250个连续氨基酸残基、至少300个连续氨基酸残基、至少350个连续氨基酸残基、至少400个连续氨基酸残基或至少450个连续氨基酸残基的氨基酸序列的肽或多肽。
在另一个实施例中,该片段是如本发明的预期而发挥作用的功能性片段(例如,当呈n末端llo/异源性抗原融合蛋白或n末端acta/异源性抗原融合蛋白形式时引起对疾病相关抗原的免疫应答)。在另一个实施例中,该片段为功能性的,形式为非融合的。在另一个实施例中,片段为免疫原性片段。
本发明在某些实施例中提供李斯特菌异源性核酸或李斯特菌内源性核酸的密码子优化。针对每种氨基酸由单核细胞增多性李斯特菌使用的最佳密码子在美国专利公开2007/0207170中示出,该专利据此以引用方式并入本文。如果核酸中的至少一个密码子被下述密码子替换,则核酸为密码子优化的:单核细胞增多性李斯特菌将该密码子比原始序列中的密码子更频繁地用于所述氨基酸。
n末端llo蛋白片段和异源性抗原在另一个实施例中直接互相融合。在另一个实施例中,编码n末端llo蛋白片段和异源性抗原的基因直接互相融合。在另一个实施例中,n末端llo蛋白片段和异源性抗原通过接头肽连接。在另一个实施例中,n末端llo蛋白片段和异源性抗原通过异源性肽连接。在另一个实施例中,n末端llo蛋白片段是异源性抗原的n末端。在另一个实施例中,n末端llo蛋白片段是融合蛋白的最n末端部分。
如本文所公开,表达llo-抗原融合体的重组李斯特菌菌株诱导抗肿瘤免疫(实例1)、引起抗原特异性t细胞增殖(实例2)、产生抗原特异性和肿瘤浸润性t细胞(实例3)。
在另一个实施例中,本发明提供一种治疗人受试者中的宫颈癌的方法,该方法包括向受试者施用重组李斯特菌菌株的步骤,该重组李斯特菌菌株包含含有llo蛋白的n末端片段和hpve7抗原的重组多肽,由此重组李斯特菌菌株诱导对e7抗原的免疫应答,从而治疗人受试者中的宫颈癌。在另一个实施例中,该重组李斯特菌菌株表达重组多肽。在另一个实施例中,该重组李斯特菌菌株包含编码重组多肽的质粒。
在另一个实施例中,本发明提供一种避免人受试者罹患宫颈癌的方法,该方法包括向受试者施用重组李斯特菌菌株的步骤,该重组李斯特菌菌株包含含有llo蛋白的n末端片段和hpve7抗原的重组多肽,由此重组李斯特菌菌株诱导对e7抗原的免疫应答,从而避免人受试者罹患宫颈癌。在另一个实施例中,该重组李斯特菌菌株表达重组多肽。在另一个实施例中,该重组李斯特菌菌株包含编码重组多肽的质粒。
在另一个实施例中,本发明提供一种诱导人受试者中对宫颈癌的免疫应答的方法,该方法包括向受试者施用重组李斯特菌菌株的步骤,该重组李斯特菌菌株包含含有llo蛋白的n末端片段和hpve7抗原的重组多肽,从而诱导人受试者中对宫颈癌的免疫应答。在另一个实施例中,该重组李斯特菌菌株表达重组多肽。在另一个实施例中,该重组李斯特菌菌株包含编码重组多肽的质粒。
在另一个实施例中,本发明提供一种治疗人受试者中的宫颈癌的方法,该方法包括向受试者施用重组李斯特菌菌株的步骤,该重组李斯特菌菌株包含含有acta蛋白的n末端片段和异源性抗原的重组多肽,由此重组李斯特菌菌株诱导对异源性抗原的免疫应答,从而治疗人受试者中的宫颈癌。在另一个实施例中,该重组李斯特菌菌株表达重组多肽。在另一个实施例中,该重组李斯特菌菌株包含编码重组多肽的质粒。
在另一个实施例中,本发明提供一种避免人受试者罹患宫颈癌的方法,该方法包括向受试者施用重组李斯特菌菌株的步骤,该重组李斯特菌菌株包含含有acta蛋白的n末端片段和异源性抗原的重组多肽,由此重组李斯特菌菌株诱导对异源性抗原的免疫应答,从而避免人受试者罹患宫颈癌。在另一个实施例中,该重组李斯特菌菌株表达重组多肽。在另一个实施例中,该重组李斯特菌菌株包含编码重组多肽的质粒。
在另一个实施例中,本发明提供一种诱导人受试者中对宫颈癌的免疫应答的方法,该方法包括向受试者施用重组李斯特菌菌株的步骤,该重组李斯特菌菌株包含含有acta蛋白的n末端片段和异源性抗原的重组多肽,从而诱导人受试者中对宫颈癌的免疫应答。在另一个实施例中,该重组李斯特菌菌株表达重组多肽。在另一个实施例中,该重组李斯特菌菌株包含编码重组多肽的质粒。
n末端acta蛋白片段和异源性抗原在另一个实施例中直接互相融合。在另一个实施例中,编码n末端acta蛋白片段和异源性抗原的基因直接互相融合。在另一个实施例中,n末端acta蛋白片段和异源性抗原通过接头肽连接。在另一个实施例中,n末端acta蛋白片段和异源性抗原通过异源性肽连接。在另一个实施例中,n末端acta蛋白片段是异源性抗原的n末端。在另一个实施例中,n末端acta蛋白片段是融合蛋白的最n末端部分。
在另一个实施例中,本发明提供一种诱导人受试者中对宫颈癌的免疫应答的方法,该方法包括向受试者施用重组李斯特菌菌株的步骤,该重组李斯特菌菌株包含重组多肽,该重组多肽包含含pest氨基酸序列的肽和异源性抗原,由此重组李斯特菌菌株诱导对异源性抗原的免疫应答,从而治疗人受试者中的宫颈癌。在另一个实施例中,该重组李斯特菌菌株表达重组多肽。在另一个实施例中,该重组李斯特菌菌株包含编码重组多肽的质粒。在另一个实施例中,该方法避免人受试者罹患宫颈癌。在另一个实施例中,该方法治疗所述人受试者中的宫颈癌。
含pest氨基酸序列的肽和异源性抗原在另一个实施例中直接互相融合。在另一个实施例中,编码含pest氨基酸序列的肽和异源性抗原的基因直接互相融合。在另一个实施例中,含pest氨基酸序列的肽和异源性抗原通过接头肽连接。在另一个实施例中,含pest氨基酸序列的肽和异源性抗原通过异源性肽连接。在另一个实施例中,含pest氨基酸序列的肽是异源性抗原的n末端。在另一个实施例中,含pest氨基酸序列的肽是融合蛋白的最n末端部分。
在另一个实施例中,本发明提供一种对人受试者进行疫苗接种以防hpv的方法,该方法包括向受试者施用本文所公开的重组李斯特菌菌株的步骤,其中该李斯特菌表达hpve7抗原并且其中该李斯特菌表达突变体prfa蛋白。在另一个实施例中,该突变体prfa基因编码prfa蛋白中的d133v突变。在另一个实施例中,该突变体prfa基因在所述重组李斯特菌中的质粒中。在另一个实施例中,该重组李斯特菌菌株表达重组多肽。在另一个实施例中,该重组李斯特菌菌株包含编码重组多肽的质粒。
在一个实施例中,如本文所用的术语“可操作地连接”意味着将转录和翻译调控核酸相对于任何编码序列以使得引发转录的方式定位。一般来讲,这意味着将启动子和转录引发或起始序列定位在编码区的5'。在另一个实施例中,如本文所用的术语“可操作地连接”意味着若干开放阅读框的融合方式使得形成单个连续阅读框,其导致包括接连地布置的原始蛋白的序列的蛋白得以表达。
在一个实施例中,“融合”是指通过共价键合可操作地连接。在一个实施例中,该术语包括(核酸序列或其开放读框的)重组融合。在另一个实施例中,该术语包括化学偶联。
在一个实施例中,本文所公开的融合蛋白由本文所公开的重组李斯特菌表达和分泌。
在另一个实施例中,受试者处于发生hpv介导的癌变(例如,宫颈癌)风险中。在另一个实施例中,受试者为hpv阳性的。在另一个实施例中,受试者表现出宫颈上皮内瘤样病变。在另一个实施例中,受试者表现出鳞状上皮内病变。在另一个实施例中,受试者表现出宫颈发育异常。
在一个实施例中,该异源性抗原是本领域已知的且本文所公开的任何肿瘤相关抗原。在另一个实施例中,该异源性抗原是自身免疫抗原。在另一个实施例中,该异源性抗原是感染性疾病抗原。在另一个实施例中,该异源性抗原是hpv相关抗原。
为本发明的方法的靶标的hpv在另一个实施例中为hpv16。在另一个实施例中,该hpv为hpv-18。在另一个实施例中,该hpv选自hpv-16和hpv-18。在另一个实施例中,该hpv为hpv-31。在另一个实施例中,该hpv为hpv-35。在另一个实施例中,该hpv为hpv-39。在另一个实施例中,该hpv为hpv-45。在另一个实施例中,该hpv为hpv-51。在另一个实施例中,该hpv为hpv-52。在另一个实施例中,该hpv为hpv-58。在另一个实施例中,该hpv为高风险hpv型。在另一个实施例中,该hpv为粘膜hpv型。
在另一个实施例中,本发明提供一种针对目的抗原对人受试者进行疫苗接种的方法,该方法包括向人受试者静脉内施用包含或表达该目的抗原的重组李斯特菌菌株的步骤,其中第一肽选自(a)llo蛋白的n末端片段、(b)acta蛋白或其n末端片段和(c)含pest氨基酸序列的肽,从而针对目的抗原对人受试者进行疫苗接种。
在另一个实施例中,本发明提供一种针对目的抗原对人受试者进行疫苗接种的方法,该方法包括向人受试者静脉内施用免疫原性组合物的步骤,该组合物包含第一肽与目的抗原的融合体,其中该第一肽选自(a)llo蛋白的n末端片段、(b)acta蛋白或其n末端片段和(c)含pest氨基酸序列的肽,从而针对目的抗原对人受试者进行疫苗接种。
在另一个实施例中,本发明提供一种针对目的抗原对人受试者进行疫苗接种的方法,该方法包括向人受试者静脉内施用包含重组多肽的重组李斯特菌菌株的步骤,该重组多肽包含融合至目的抗原的第一肽,其中该第一肽选自(a)llo蛋白的n末端片段、(b)acta蛋白或其n末端片段和(c)含pest氨基酸序列的肽,从而针对目的抗原对人受试者进行疫苗接种。
在另一个实施例中,本发明提供一种诱导人受试者中针对目的抗原的ctl应答的方法,该方法包括向人受试者施用包含或表达目的抗原的重组李斯特菌菌株的步骤,从而诱导人受试者中针对目的抗原的ctl应答。在另一个实施例中,施用步骤为静脉内施用。
如本文所公开,表达llo-抗原融合体的重组李斯特菌菌株诱导抗肿瘤免疫(实例1)、引起抗原特异性t细胞增殖(实例2)、产生抗原特异性和肿瘤浸润性t细胞(实例3)。因此,本发明的疫苗有效诱导对e7和e6的免疫应答。
在另一个实施例中,本发明提供一种诱导受试者中的癌症消退的方法,该方法包括向受试者施用本文所公开的重组李斯特菌菌株的步骤。
在另一个实施例中,本发明提供一种降低受试者中的癌症的发生率或复发率的方法,该方法包括向受试者施用本文所公开的重组李斯特菌菌株的步骤。
在另一个实施例中,本发明提供一种抑制受试者中的肿瘤形成的方法,该方法包括向受试者施用本文所公开的重组李斯特菌菌株的步骤。
在另一个实施例中,本发明提供一种诱导受试者中的癌症缓解的方法,该方法包括向受试者施用本文所公开的重组李斯特菌菌株的步骤。
在另一个实施例中,本发明提供一种阻遏人受试者中的肿瘤生长的方法,该方法包括向受试者施用本文所公开的重组李斯特菌菌株的步骤。
在另一个实施例中,本发明提供一种减小受试者中的肿瘤大小的方法,该方法包括向受试者施用本文所公开的重组李斯特菌菌株的步骤。
在一个实施例中,疾病为感染性疾病、自身免疫疾病、呼吸疾病、癌前病症或癌症。
技术人员将容易认识到,术语“癌前病症”可涵盖发育异常、肿瘤发生前结节;大再生结节(mrn);低级别异型增生结节(lg-dn);高级别异型增生结节(hg-dn);胆管上皮发育异常;变异肝细胞病灶(fah);变异肝细胞结节(nah);染色体失衡;端粒酶异常活化;端粒酶催化亚单位再表达;内皮细胞标记物诸如cd31、cd34和bnh9的表达(参见例如terracciano和tomillo(2003)pathologica95:71-82;su和bannasch(2003)toxicol.pathol.31:126-133;rocken和carl-mcgrath(2001)dig.dis.19:269-278;kotoula等人(2002)liver22:57-69;frachon等人(2001)j.hepatol.34:850-857;shimonishi等人(2000)j.hepatobiliarypancreat.surg.7:542-550;nakanuma等人(2003)j.hepatobiliarypancreat.surg.10:265-281)。诊断癌症和发育异常的方法已有公开(参见例如riegler(1996)semin.gastrointest.dis.7:74-87;benvegnu等人(1992)liver12:80-83;giannini等人(1987)hepatogastroenterol.34:95-97;anthony(1976)cancerres.36:2579-2583)。
在一个实施例中,感染性疾病为由以下病原体的任一种但并不限于它们所引起的感染性疾病:bcg/结核病、疟疾、恶性疟原虫、三日疟原虫、间日疟原虫、轮状病毒、霍乱、白喉-破伤风、百日咳、流感嗜血杆菌、乙型肝炎、人乳头瘤病毒、季节性流感)、a型流感(h1n1)流行病、麻疹和风疹、腮腺炎、脑膜炎球菌a+c、口服脊髓灰质炎疫苗(单价、二价和三价)、肺炎球菌、狂犬病、破伤风类毒素、黄热病、炭疽芽孢杆菌(炭疽)、肉毒棱状芽孢杆菌毒素(肉毒中毒)、鼠疫耶尔森菌(瘟疫)、重型天花(天花)和其他有关的痘病毒、土拉热弗朗西丝菌(土拉菌病)、病毒性出血热、沙粒病毒(lcm、胡宁病毒、马丘波病毒、瓜纳瑞托病毒、拉沙热)、布尼亚病毒(汉坦病毒属、裂谷热)、黄病毒属(登革热)、线状病毒(埃博拉病毒、马尔堡病毒)、假鼻疽伯克霍尔德菌、伯内特考克斯体(q热)、布鲁杆菌属种(布鲁杆菌病)、鼻疽伯克霍尔德菌(鼻疽)、鹦鹉衣原体(鹦鹉热)、蓖麻蛋白毒素(来自蓖麻)、产气荚膜梭菌的ε毒素、葡萄球菌肠毒素b、斑疹伤寒热(普氏立克次体)、其他立克次体、食品和水传播的病原体、细菌(致腹泻性的大肠杆菌、致病性的弧菌属、志贺氏菌属种、沙门氏菌属bcg/、空肠弯曲杆菌、小肠结肠炎耶尔森菌)、病毒(杯状病毒、甲型肝炎、西尼罗病毒、lacrosse、加利福尼亚脑炎、vee、eee、wee、日本脑炎病毒、科萨努尔森林病毒、尼帕病毒、汉坦病毒属、蜱传出血热病毒、切昆贡亚病毒、克里米亚-刚果出血热病毒、蜱传脑炎病毒、乙型肝炎病毒、丙型肝炎病毒、单纯疱疹病毒(hsv)、人免疫缺陷病毒(hiv)、人乳头状瘤病毒(hpv)、原生动物(小隐孢子虫、卡晏环孢子虫、兰伯贾第虫、溶组织内阿米巴、弓形虫属)、真菌(微孢子目)、黄热病、结核病(包括耐药的tb)、狂犬病、朊病毒、严重急性呼吸综合征相关的冠状病毒(sars-cov)、coccidioidesposadasii、粗球孢子菌、细菌性阴道病、沙眼衣原体、巨细胞病毒、腹股沟肉芽肿、杜克雷嗜血杆菌、淋病奈瑟球菌、苍白密螺旋体、阴道毛滴虫或本领域已知的在本文中没有列出的任何其他感染性疾病。
在另一个实施例中,该感染性疾病是家畜感染性疾病。在另一个实施例中,家畜疾病可传播给人,并被称作“人畜共患病”。在另一个实施例中,这些疾病包括但不限于口蹄疫、西尼罗病毒、狂犬病、犬细小病毒、猫白血病病毒、马流感病毒、传染性牛鼻气管炎(ibr)、伪狂犬病、古典猪瘟(csf)、由牛1型牛疱疹病毒(bhv-1)感染造成的ibr以及猪伪狂犬病(奥尔斯基病)、弓形虫病、炭疽、水疱性口炎病毒、马红球菌、土拉菌病、瘟疫(鼠疫耶尔森菌)、毛滴虫。
在另一个实施例中,本文所公开的疾病为呼吸道疾病或炎症性疾病。在另一个实施例中,该呼吸道疾病或炎症性疾病为慢性阻塞性肺病(copd)。在另一个实施例中,该疾病为哮喘。
在一个实施例中,活的减毒李斯特菌菌株能够缓解哮喘症状,无需其他治疗剂的共施用,例如抗炎剂或支气管扩张剂。在另一个实施例中,本文所公开的方法还包括向受试者共同施用活的减毒李斯特菌菌株和一种或多种治疗剂的步骤。在另一个实施例中,该治疗剂为平喘药。在另一个实施例中,该治疗剂为抗炎剂、非甾体抗炎剂、抗生素、抗胆碱能药、支气管扩张药、皮质类固醇、短效β受体激动剂、长效β受体激动剂、组合吸入剂、抗组织胺药或它们的组合。
在一个实施例中,本文所公开的疾病为癌症或肿瘤。在一个实施例中,该肿瘤为癌性的。在另一个实施例中,该癌症为乳腺癌。在另一个实施例中,该癌症为宫颈癌。在另一个实施例中,该癌症为含her2癌症。在另一个实施例中,该癌症为黑素瘤。在另一个实施例中,该癌症为胰腺癌。在另一个实施例中,该癌症为卵巢癌。在另一个实施例中,该癌症为胃癌。在另一个实施例中,该癌症为胰腺的癌性病变。在另一个实施例中,该癌症为肺腺癌。在另一个实施例中,其为多形性胶质母细胞瘤。在另一个实施例中,该癌症为结直肠腺癌。在另一个实施例中,该癌症为肺鳞癌。在另一个实施例中,该癌症为胃腺癌。在另一个实施例中,该癌症为卵巢表面上皮肿瘤(例如其良性的、增生性的或恶性的种类)。在另一个实施例中,该癌症为口腔鳞状细胞癌。在另一个实施例中,该癌症为非小细胞肺癌。在另一个实施例中,该癌症为子宫内膜癌。在另一个实施例中,该癌症为膀胱癌。在另一个实施例中,该癌症为头颈癌。在另一个实施例中,该癌症为前列腺癌。在另一个实施例中,该癌症为口咽癌。在另一个实施例中,该癌症为肺癌。在另一个实施例中,该癌症为肛门癌。在另一个实施例中,该癌症为结肠直肠癌。在另一个实施例中,该癌症为食道癌。本发明的方法靶向的宫颈肿瘤在另一个实施例中为鳞状细胞癌。在另一个实施例中,该宫颈肿瘤为腺癌。在另一个实施例中,该宫颈肿瘤为腺鳞癌。在另一个实施例中,该宫颈肿瘤为小细胞癌。在另一个实施例中,该宫颈肿瘤为本领域已知的任何其他类型的宫颈肿瘤。
本发明的方法靶向的宫颈肿瘤在另一个实施例中为鳞状细胞癌。在另一个实施例中,该宫颈肿瘤为腺癌。在另一个实施例中,该宫颈肿瘤为腺鳞癌。在另一个实施例中,该宫颈肿瘤为小细胞癌。在另一个实施例中,该宫颈肿瘤为本领域已知的任何其他类型的宫颈肿瘤。
在一个实施例中,本文所公开的抗原为异源性肿瘤抗原,其在本文也称为“肿瘤抗原、“抗原性多肽”或“外源抗原”。在另一个实施例中,该抗原为人类乳头瘤病毒e7(hpv-e7)抗原,其在一个实施例中来自hpv16(在一个实施例中,genbank登录号aad33253),在另一个实施例中来自hpv18(在一个实施例中,genbank登录号p06788)。在另一个实施例中,该抗原性多肽为hpv-e6,其在一个实施例中来自hpv16(在一个实施例中,genbank登录号aad33252、aam51854、aam51853或aab67615),在另一个实施例中来自hpv18(在一个实施例中,genbank登录号p06463)。在另一个实施例中,该抗原性多肽为her/2-neu抗原。在另一个实施例中,该抗原性多肽为前列腺特异性抗原(psa)(在一个实施例中genbank登录号cad30844、cad54617、aaa58802或np—001639)。在另一个实施例中,该抗原性多肽为角质层胰凝乳蛋白酶(scce)抗原(在一个实施例中,genbank登录号aak69652、aak69624、aag33360、aaf01139或aac37551)。在另一个实施例中,该抗原性多肽为wilms肿瘤抗原1,其在另一个实施例中为wt-1端粒酶(genbank登录号p49952、p22561、np—659032、cac39220.2或eaw68222.1)。在另一个实施例中,该抗原性多肽为htert或端粒酶(genbank登录号nm003219(变体1)、nm198255(变体2)、nm198253(变体3)或nm198254(变体4)。在另一个实施例中,该抗原性多肽为蛋白酶3(在一个实施例中,genbank登录号m29142、m75154、m96839、x55668、nm00277、m96628或x56606)。在另一个实施例中,该抗原性多肽为酪氨酸酶相关蛋白2(trp2)(在一个实施例中,genbank登录号np—001913、abi73976、aap33051或q95119)。在另一个实施例中,该抗原性多肽为高分子量黑素瘤相关抗原(hmw-maa)(在一个实施例中,genbank登录号np—001888、aai28111或aaq62842)。在另一个实施例中,该抗原性多肽为testisin(在一个实施例中,genbank登录号aaf79020、aaf79019、aag02255、aak29360、aad41588或np—659206)。在另一个实施例中,该抗原性多肽为ny-eso-1抗原(在一个实施例中,genbank登录号caa05908、p78358、aab49693或np—640343)。在另一个实施例中,该抗原性多肽为psca(在一个实施例中,genbank登录号aah65183、np—005663、np—082492、o43653或cab97347)。在另一个实施例中,该抗原性多肽为白介素(il)13受体α(在一个实施例中,genbank登录号np—000631、np—001551、np—032382、np—598751、np—001003075或np—999506)。在另一个实施例中,该抗原性多肽为碳酸酐酶ix(caix)(在一个实施例中,genbank登录号cai13455、cai10985、eaw58359、np—001207、np—647466或np—001101426)。在另一个实施例中,该抗原性多肽为癌胚抗原(cea)(在一个实施例中,genbank登录号aaa66186、caa79884、caa66955、aaa51966、aad15250或aaa51970)。在另一个实施例中,该抗原性多肽为mage-a(在一个实施例中,genbank登录号np—786885、np—786884、np—005352、np—004979、np—005358或np—005353)。在另一个实施例中,该抗原性多肽为存活素(在一个实施例中,genbank登录号aac51660、aay15202、abf60110、np—001003019或np—001082350)。在另一个实施例中,该抗原性多肽为gp100(在一个实施例中,genbank登录号aac60634、yp—655861或aab31176)。在另一个实施例中,该抗原性多肽为本领域已知的任何其他抗原性多肽。在另一个实施例中,本发明的组合物和方法的抗原性肽包含所述抗原性多肽的免疫原性部分。
在另一个实施例中,该抗原为hpv-e6。在另一个实施例中,该抗原为hpv16-e6。在另一个实施例中,该抗原为hpv18-e6。在另一个实施例中,抗原为hpv-e7。在另一个实施例中,该抗原为hpv16-e7。在另一个实施例中,该抗原为hpv18-e7。在另一个实施例中,该抗原为端粒酶(tert)。在另一个实施例中,该抗原为lmp-1。在另一个实施例中,该抗原为p53。在另一个实施例中,该抗原为间皮素。在另一个实施例中,该抗原为egfrviii。在另一个实施例中,该抗原为碳酸酐酶ix(caix)。在另一个实施例中,该抗原为psma。在另一个实施例中,该抗原为hmw-maa。在另一个实施例中,该抗原为hiv-1gag。在另一个实施例中,该抗原为酪氨酸酶相关蛋白2。在另一个实施例中,该抗原选自hpv-e7、hpv-e6、her-2、hiv-1gag、lmp-1、p53、psma、癌胚抗原(cea)、lmp-1、激肽释放酶相关肽酶3(klk3)、klk9、muc、酪氨酸酶相关蛋白2、muc1、fap、il-13rα2、psa(前列腺特异性抗原)、gp-100、热休克蛋白70(hsp-70)、β-hcg、egfr-iii、粒细胞集落刺激因子(g-csf)、血管生成素、促血管生成素-1、del-1、成纤维细胞生长因子:酸性(afgf)或碱性(bfgf)、卵泡抑素、粒细胞集落刺激因子(g-csf)、肝细胞生长因子(hgf)/分散因子(sf)、白细胞介素-8(il-8)、痩素、中期因子、胎盘生长因子、血小板衍生内皮细胞生长因子(pd-ecgf)、血小板衍生生长因子-bb(pdgf-bb)、多效生长因子(ptn)、颗粒蛋白前体、增殖素、转化生长因子-α(tgf-α)、转化生长因子-β(tgf-β)、肿瘤坏死因子-α(tnf-α)、血管内皮生长因子(vegf)/血管渗透因子(vpf)、vegfr、vegfr2(kdr/flk-1)或其片段、flk-1或其表位、flk-e1、flk-e2、flk-i1、内皮因子或其片段、神经菌毛素1(nrp-1)、促血管生成素-1(ang1)、tie2、血小板衍生生长因子(pdgf)、血小板衍生生长因子受体(pdgfr)、转化生长因子-β(tgf-β)、内皮因子、tgf-β受体、单核细胞趋化蛋白-1(mcp-1)、ve-钙粘蛋白、cd31、ephrin、icam-1、v-cam-1、vap-1、e-选择素、纤溶酶原激活物、纤溶酶原激活物抑制剂-1、一氧化氮合酶(nos)、cox-2、ac133或id1/id3、促血管生成素3、促血管生成素4、促血管生成素6、cd105、edg、hht1、orw、orw1或tgfβ共受体或它们的组合。在另一个实施例中,所述抗原为美国专利申请公开no.2011/0142791中公开的嵌合her2抗原,通过引用将该专利全文并入本文。对本文所公开的抗原的片段的使用也涵盖在本发明中。
在另一个实施例中,本文所公开的异源性肿瘤抗原为肿瘤相关抗原,其在一个实施例中为以下肿瘤抗原之一:mage(黑素瘤相关抗原e)蛋白,例如mage1、mage2、mage3、mage4、酪氨酸酶;突变体ras蛋白;突变体p53蛋白;p97黑素瘤抗原、与晚期癌症相关的ras肽或p53肽;与宫颈癌相关的hpv16/18抗原、与乳腺癌相关的klh抗原、与结肠直肠癌相关的cea(癌胚抗原)、与黑素瘤相关的mart1抗原或与前列腺癌相关的psa抗原。在另一个实施例中,用于本文所公开的组合物和方法的抗原为黑素瘤相关抗原,其在一个实施例中为trp-2、mage-1、mage-3、gp-100、酪氨酸酶、hsp-70、β-hcg或它们的组合。应当理解,技术人员能够将本文未提到但本领域已知的任何异源抗原用在本文所公开的方法和组合物中。还应当理解的是,本发明提供但不限于编码本文所公开的至少一种抗原的核酸的减毒李斯特菌。本发明涵盖编码所公开的抗原的突变体、突变蛋白、剪接变体、片段、截短的变体、可溶性变体、胞外域、胞内域、成熟序列等的核酸。公开了编码这些抗原的表位、寡肽和多肽的核酸。还公开了密码子优化的实施例,这些实施例针对在李斯特菌中的表达而优化。所引用的参考文献、genbank登录号以及本文所公开的核酸、肽和多肽均整体以引用方式并入本文。在另一个实施例中,所选的核酸序列可编码全长或截短的基因、融合或标记的基因,并且可以是cdna、基因组dna或dna片段,优选cdna。其根据需要可以突变或以其他方式修饰。这些修饰包括密码子优化以优化密码子在所选宿主细胞或细菌即李斯特菌中的使用。所选的序列也可以编码分泌的、细胞质的、细胞核的、膜结合的或细胞表面的多肽。
在一个实施例中,血管内皮生长因子(vegf)为重要的参与血管发生(胚胎循环系统的形成)和血管生成(从现存脉管系统生长血管)的信号蛋白。在一个实施例中,vegf活性主要限于血管内皮的细胞,尽管其对有限数量的其他细胞类型有作用(例如刺激单核细胞/巨噬细胞迁移)。vegf在体外已显示出刺激内皮细胞有丝分裂和细胞迁移。vegf还增强微血管渗透性,并且有时也被称为血管通透性因子。
在一个实施例中,vegf家族的所有成员都刺激细胞应答,这通过结合细胞表面上的酪氨酸激酶受体(vegfr),导致它们二聚化并经由转磷酸作用而被活化来实现。该vegf受体具有由7个免疫球蛋白样域组成的胞外部分、单个跨膜区和含有分开的酪氨酸激酶域的胞内部分。
在一个实施例中,vegf-a为vegfr-2(kdr/flk-1)配体以及vegfr-1(flt-1)配体。在一个实施例中,vegfr介导几乎全部已知的对vegf的细胞应答。vegfr-1的功能不太明确,尽管在一个实施例中认为其通过隔离vegf与vegfr-2结合而调节vegfr-2信号转导,在一个实施例中,这在胚胎的血管发生期间是尤其重要的。在一个实施例中,vegf-c和vegf-d为vegfr-3受体的配体,该vegfr-3受体在一个实施例中介导淋巴管生成。
在一个实施例中,本发明的组合物包含vegf受体或其片段,其在一个实施例中为vegfr-2,在另一个实施例中为vegfr-1,以及在另一个实施例中为vegfr-3。
在一个实施例中,血管内皮生长因子受体2(vegfr2)在活化的内皮细胞(ec)上高表达并参与新血管的形成。在一个实施例中,vegfr2结合vegf的全部5个同种型。在一个实施例中,vegf经由vegfr2在ec上的信号转导诱导增殖、迁移和最终的分化。在一个实施例中,vegfr2的小鼠同源物为胎儿肝激酶基因-1(flk-1),其为强治疗靶标,并在肿瘤生长、入侵和转移中起重要作用。在一个实施例中,vegfr2也称为激酶插入区受体(iii型受体酪氨酸激酶)(kdr)、分化群309(cd309)、flk1、ly73、krd-1、vegfr、vegfr-2或6130401c07。
在一个实施例中,抗原源自真菌病原体、细菌、寄生虫、蠕虫或病毒。在另一个实施例中,该抗原选自破伤风类毒素、来自流感病毒的血凝素分子、白喉类毒素、hivgp120、hivgag蛋白、iga蛋白酶、胰岛素肽b、马铃薯粉痂病菌(spongosporasubterranea)抗原、弧菌抗原、沙门氏菌(salmonella)抗原、肺炎球菌抗原、呼吸道合胞体病毒抗原、流感嗜血杆菌(haemophilusinfluenza)外膜蛋白、幽门螺杆菌(helicobacterpylori)脲酶、脑膜炎奈瑟球菌(neisseriameningitidis)菌毛蛋白、淋病奈瑟菌(n.gonorrhoeae)菌毛蛋白、黑素瘤相关抗原(trp-2、mage-1、mage-3、gp-100、酪氨酸酶、mart-1、hsp-70、β-hcg)、来自hpv-16、hpv-18、hpv-31、hpv-33、hpv-35或hpv-45型人类乳头瘤病毒的人类乳头瘤病毒抗原e1和e2、肿瘤抗原cea、突变或者其他形式的ras蛋白、突变或者其他形式的p53蛋白、muc1或psa。
在其他实施例中,抗原与以下疾病之一相关:霍乱、白喉、嗜血杆菌、甲型肝炎、乙型肝炎、流感、麻疹、脑膜炎、腮腺炎、百日咳、天花、肺炎球菌肺炎、脑灰质炎、狂犬病、风疹、破伤风、肺结核、伤寒、水痘-带状疱疹、百日咳、黄热病、来自爱迪生氏病的免疫原和抗原、过敏症、过敏反应、布鲁顿综合征、癌症、包括实体和血源性肿瘤、湿疹、桥本氏甲状腺炎、多肌炎、皮肌炎、1型糖尿病、获得性免疫缺陷综合征、移植排斥、例如肾、心脏、胰腺、肺、骨骼和肝脏移植物、格雷夫斯病、多内分泌自身免疫病、肝炎、显微镜下多动脉炎、结节性多动脉炎、天疱疮、原发性胆汁性肝硬化、恶性贫血、乳糜泻、抗体介导的肾炎、血管球性肾炎、风湿病、全身性红斑狼疮、类风湿性关节炎、血清阴性脊椎关节病、鼻炎、sjogren综合征、全身性硬化症、硬化性胆管炎、韦格纳氏肉芽肿、疱疹样皮炎、牛皮癣、白癜风、多发性硬化症、脑脊髓炎、吉兰-巴雷综合征、重症肌无力、兰伯特-伊顿综合征、巩膜、巩膜外层、色素层炎、慢性粘膜皮肤念珠菌病、风疹、婴儿短暂性低丙种球蛋白血症、骨髓瘤、x-连锁高igm综合征、斯科特-奥尔德里奇综合征、共济失调性毛细血管扩张症、自身免疫性溶血性贫血、自身免疫性血小板减少症、自身免疫性中性粒细胞减少症、瓦尔登斯特伦巨球蛋白血、淀粉样变性、慢性淋巴细胞白血病、非霍奇金淋巴瘤、恶性疟原虫环子孢子蛋白、微生物抗原、病毒抗原、自身抗原以及李斯特菌病。
在另一个实施例中,在用于治疗宫颈癌、防御宫颈癌或诱导对宫颈癌的免疫应答的本发明的方法中,代替e7抗原或作为其补充,使用hpve6抗原。
在另一个实施例中,在用于治疗宫颈癌、防御宫颈癌或诱导对宫颈癌的免疫应答的本发明的方法中,代替llo片段或作为其补充,使用acta蛋白片段。
在另一个实施例中,在用于治疗宫颈癌、防御宫颈癌或诱导对宫颈癌的免疫应答的本发明的方法中,代替llo片段或作为其补充,使用含pest氨基酸序列的蛋白片段。
在另一个实施例中,本文提供了包含本发明的重组李斯特菌的免疫原性组合物。在另一个实施例中,本发明的方法和组合物的免疫原性组合物包含本发明的重组疫苗载体。在另一个实施例中,免疫原性组合物包含本发明的质粒。在另一个实施例中,免疫原性组合物包含佐剂。在一个实施例中,本发明的载体可作为疫苗组合物的一部分施用。
在另一个实施例中,本发明的疫苗与佐剂一起递送。在一个实施例中,佐剂有利于主要由th1介导的免疫应答。在另一个实施例中,佐剂有利于th1型免疫应答。在另一个实施例中,佐剂有利于th1介导的免疫应答。在另一个实施例中,佐剂有利于细胞介导的免疫应答超过抗体介导的应答。在另一个实施例中,佐剂是本领域已知的任何其他类型的佐剂。在另一个实施例中,免疫原性组合物诱导针对靶蛋白的t细胞免疫应答的形成。
在另一个实施例中,本发明提供一种诱导诱导人受试者中的抗e7细胞毒性t细胞(ctl)应答的方法,该方法包括向受试者施用重组李斯特菌菌株的步骤,该重组李斯特菌菌株包含含有llo蛋白的n末端片段和hpve7抗原的重组多肽,从而诱导人受试者中的抗e7ctl应答。在另一个实施例中,该重组李斯特菌菌株包含编码重组多肽的质粒。在另一个实施例中,该方法还包括用本发明的重组李斯特菌菌株强化受试者的步骤。在另一个实施例中,该方法还包括用包含e7抗原的免疫原性组合物强化受试者的步骤。在另一个实施例中,该方法还包括用免疫原性组合物强化受试者的步骤,该组合物引导受试者的细胞表达e7抗原。在另一个实施例中,ctl应答能够对hpv介导的疾病、障碍或症状产生治疗性功效。在另一个实施例中,ctl应答能够对hpv介导的疾病、障碍或症状产生预防性功效。
在另一个实施例中,本发明提供治疗或改善受试者中的hpv介导的疾病、障碍或症状的方法,该方法包括向受试者施用重组李斯特菌菌株的步骤,该重组李斯特菌菌株包含含有llo蛋白的n末端片段和hpve7抗原的重组多肽,由此重组李斯特菌菌株诱导对e7抗原的免疫应答,从而治疗或改善受试者中的hpv介导的疾病、障碍或症状。在另一个实施例中,受试者是人受试者。在另一个实施例中,受试者是非人受试者。在另一个实施例中,受试者是本领域已知的任何其他类型的受试者。
导致疾病、障碍或症状的hpv在另一个实施例中为hpv16。在另一个实施例中,该hpv为hpv-18。在另一个实施例中,该hpv为hpv-31。在另一个实施例中,该hpv为hpv-35。在另一个实施例中,该hpv为hpv-39。在另一个实施例中,该hpv为hpv-45。在另一个实施例中,该hpv为hpv-51。在另一个实施例中,该hpv为hpv-52。在另一个实施例中,该hpv为hpv-58。在另一个实施例中,该hpv为高风险hpv型。在另一个实施例中,该hpv为粘膜hpv型。
在另一个实施例中,hpv介导的疾病、障碍或症状为生殖器疣。在另一个实施例中,hpv介导的疾病、障碍或症状为非生殖器疣。在另一个实施例中,hpv介导的疾病、障碍或症状为呼吸道乳头瘤。在另一个实施例中,hpv介导的疾病、障碍或症状为本领域已知的任何其他hpv介导的疾病、障碍或症状。
在另一个实施例中,在用于治疗或改善hpv介导的疾病、障碍或症状的本发明的方法中,代替e7抗原或作为其补充,使用hpve6抗原。
在另一个实施例中,在用于治疗或改善hpv介导的疾病、障碍或症状的本发明的方法中,代替llo片段或作为其补充,使用acta蛋白片段。
在另一个实施例中,在用于治疗或改善hpv介导的疾病、障碍或症状的本发明的方法中,代替llo片段或作为其补充,使用含pest氨基酸序列的蛋白片段。
在另一个实施例中,在用于治疗或改善hpv介导的疾病、障碍或症状的本发明的方法中,代替e7抗原或作为其补充,使用hpve6抗原。
在另一个实施例中,本发明的方法和组合物的抗原是hpve7蛋白。在另一个实施例中,该抗原是hpve6蛋白。在另一个实施例中,该抗原是本领域已知的任何其他hpv蛋白。
“e7抗原”在另一个实施例中是指e7蛋白。在另一个实施例中,该术语是指e7片段。在另一个实施例中,该术语是指e7肽。在另一个实施例中,该术语是指本领域已知的任何其他类型的e7抗原。
本发明的方法和组合物的e7蛋白在另一个实施例中为hpv16e7蛋白。在另一个实施例中,该e7蛋白为hpv-18e7蛋白。在另一个实施例中,该e7蛋白为hpv-31e7蛋白。在另一个实施例中,该e7蛋白为hpv-35e7蛋白。在另一个实施例中,该e7蛋白为hpv-39e7蛋白。在另一个实施例中,该e7蛋白为hpv-45e7蛋白。在另一个实施例中,该e7蛋白为hpv-51e7蛋白。在另一个实施例中,该e7蛋白为hpv-52e7蛋白。在另一个实施例中,该e7蛋白为hpv-58e7蛋白。在另一个实施例中,该e7蛋白为高风险hpv型e7蛋白。在另一个实施例中,该e7蛋白为粘膜hpv型e7蛋白。
“e6抗原”在另一个实施例中是指e6蛋白。在另一个实施例中,该术语是指e6片段。在另一个实施例中,该术语是指e6肽。在另一个实施例中,该术语是指本领域已知的任何其他类型的e6抗原。
本发明的方法和组合物的e6蛋白在另一个实施例中为hpv16e6蛋白。在另一个实施例中,该e6蛋白为hpv-18e6蛋白。在另一个实施例中,该e6蛋白为hpv-31e6蛋白。在另一个实施例中,该e6蛋白为hpv-35e6蛋白。在另一个实施例中,该e6蛋白为hpv-39e6蛋白。在另一个实施例中,该e6蛋白为hpv-45e6蛋白。在另一个实施例中,该e6蛋白为hpv-51e6蛋白。在另一个实施例中,该e6蛋白为hpv-52e6蛋白。在另一个实施例中,该e6蛋白为hpv-58e6蛋白。在另一个实施例中,该e6蛋白为高风险hpv型e6蛋白。在另一个实施例中,该e6蛋白为粘膜hpv型e6蛋白。
本发明的方法和组合物诱导的免疫应答在另一个实施例中为t细胞应答。在另一个实施例中,免疫应答包括t细胞应答。在另一个实施例中,该应答为cd8+t细胞应答。在另一个实施例中,该应答包括cd8+t细胞应答。
在一个实施例中,本发明的组合物诱导强烈的干扰素-γ先天刺激,该干扰素-γ在一个实施例中具有抗血管新生性质。在一个实施例中,本发明的李斯特菌诱导强烈的干扰素-γ先天刺激,该干扰素-γ在一个实施例中具有抗血管新生性质(dominiecki等人,cancerimmunolimmunother.2005年5月;54(5):477-88.2004年10月6日电子版,以引用方式全文并入本文;beatty和paterson,jimmunol.2001年2月15日;166(4):2276-82,以引用方式全文并入本文)。在另一个实施例中,本发明的方法增加产干扰素γ细胞的水平。在一个实施例中,李斯特菌的抗血管新生性质通过cd4+t细胞介导(beatty和paterson,2001)。在另一个实施例中,李斯特菌的抗血管新生性质通过cd8+t细胞介导。在另一个实施例中,因李斯特菌疫苗接种导致的ifn-γ分泌由nk细胞、nkt细胞、th1cd4+t细胞、tc1cd8+t细胞或它们的组合介导。
在另一个实施例中,本发明的组合物诱导一种或多种抗血管新生蛋白或因子的生成。在一个实施例中,该抗血管新生蛋白为ifn-γ。在另一个实施例中,该抗血管新生蛋白为色素上皮衍生因子(pedf)、血管新生抑制素、内皮抑素、fms样酪氨酸激酶(sflt)-1或可溶性内皮因子(seng)。在一个实施例中,本发明的李斯特菌参与抗血管新生因子的释放,由此,在一个实施例中,除了作为用于将抗原引入受试者的载体之外还具有治疗作用。每个李斯特菌菌株及其类型代表了本发明的独立实施例。
在另一个实施例中,由本发明的方法和组合物诱导的免疫应答是抑制目标肿瘤细胞中程序性细胞死亡受体1-配体1(pd-l1)表达。在另一个实施例中,免疫应答包括肿瘤内表达程序性细胞死亡受体-1(pd-1)的免疫细胞的水平升高。在另一个实施例中,免疫应答包括pd-1表达水平与pd-l1表达水平的比率升高。在另一个实施例中,免疫应答包括抑制肿瘤pd-l1介导的免疫抑制。
在另一个实施例中,施用本发明的组合物诱导稳健的系统性抗原特异性免疫。在另一个实施例中,施用本发明的组合物诱导表位扩展。在另一个实施例中,施用本发明的组合物诱导对自身来源的肿瘤抗原的广泛应答。在另一个实施例中,由本发明的方法和组合物诱导的免疫应答包括改善肿瘤微环境(tme)中抑制性与效应免疫细胞的总体平衡。在另一个实施例中,由本发明的方法和组合物诱导的免疫应答包括改善抑制性与效应免疫细胞的系统平衡。
在一个实施例中,如本文所公开的组合物及其使用方法生成能够浸润肿瘤、破坏肿瘤细胞并根除疾病的效应t细胞。在另一个实施例中,本发明的使用方法增加效应t细胞产生的浸润。在另一个实施例中,效应t细胞包括cd8+t细胞。在另一个实施例中,效应t细胞包括cd4+t细胞。
在一个实施例中,肿瘤浸润性淋巴细胞(til)与多种肿瘤诸如结肠癌、卵巢癌和黑素瘤中更好的预后相关。在结肠癌中,不具有微转移信号的肿瘤具有增加的免疫细胞浸润和th1表达谱,这与改善的患者存活相关。此外,t细胞对肿瘤的浸润在临床前和临床试验中都已与免疫疗法的成功相关。在一个实施例中,淋巴细胞浸润到肿瘤部位中依赖于肿瘤血管的内皮细胞中粘附分子的上调,这种上调通常通过促炎性细胞因子诸如ifn-γ、tnf-α和il-1进行。若干粘附分子已在淋巴细胞浸润到肿瘤中的过程中牵涉,包括细胞间粘附分子1(icam-1)、血管内皮细胞粘附分子1(v-cam-1)、血管粘附蛋白1(vap-1)和e-选择素。然而,这些细胞粘附分子通常在肿瘤血管中下调。因此,在一个实施例中,如本文所公开的癌症疫苗增加til,上调粘附分子(在一个实施例中,icam-1、v-cam-1、vap-1、e-选择素或它们的组合),上调促炎性细胞因子(在一个实施例中,ifn-γ、tnf-α、il-1或它们的组合)或它们的组合。
本发明的方法和组合物的n末端llo蛋白片段在另一个实施例中包含seqidno:2。在另一个实施例中,该片段包含llo信号肽。在另一个实施例中,该片段包含seqidno:2。在另一个实施例中,该片段大约由seqidno:2组成。在另一个实施例中,该片段基本上由seqidno:2组成。在另一个实施例中,该片段对应于seqidno:2。在另一个实施例中,该片段与seqidno:2同源。在另一个实施例中,该片段与seqidno:2的片段同源。用于一些实例的δllo的长度为416个aa(不包括信号序列),因为包括含有半胱氨酸484的激活域在内的氨基末端的88个残基被截短。本领域的技术人员将会知道,无激活域具体地讲无半胱氨酸484的任何δllo均适用于本发明的方法和组合物。在另一个实施例中,e7或e6抗原与任何δllo(包括pest氨基酸序列,seqidno:1)的融合增强抗原的细胞介导的抗肿瘤免疫性。
在另一个实施例中,用于构建本发明的疫苗的llo蛋白具有以下序列:
mkkimlvfitlilvslpiaqqteakdasafnkensissmappasppaspktpiekkhadeidkyiqgldynknnvlvyhgdavtnvpprkgykdgneyivvekkkksinqnnadiqvvnaissltypgalvkanselvenqpdvlpvkrdsltlsidlpgmtnqdnkivvknatksnvnnavntlverwnekyaqaypnvsakidyddemaysesqliakfgtafkavnnslnvnfgaisegkmqeevisfkqiyynvnvneptrpsrffgkavtkeqlqalgvnaenppayissvaygrqvylklstnshstkvkaafdaavsgksvsgdveltniiknssfkaviyggsakdevqiidgnlgdlrdilkkgatfnretpgvpiayttnflkdnelaviknnseyiettskaytdgkinidhsggyvaqfniswdevnydpegneivqhknwsennksklahftssiylpgnarninvyakectglawewwrtviddrnlplvknrnisiwgttlypkysnkvdnpie(genbank登录号p13128;seqidno:3;核酸序列在genbank登录号x15127中示出)。对应于该序列的前蛋白的前25个aa为信号序列,并在由细菌分泌时从llo切除。因此,在本实施例中,全长的活性llo蛋白为504个残基长。在另一个实施例中,将以上llo片段用作并入本发明的疫苗中的llo片段的来源。
在另一个实施例中,用于本发明的组合物和方法的llo蛋白的n末端片段具有以下序列:
mkkimlvfitlilvslpiaqqteakdasafnkensissvappasppaspktpiekkhadeidkyiqgldynknnvlvyhgdavtnvpprkgykdgneyivvekkkksinqnnadiqvvnaissltypgalvkanselvenqpdvlpvkrdsltlsidlpgmtnqdnkivvknatksnvnnavntlverwnekyaqaysnvsakidyddemaysesqliakfgtafkavnnslnvnfgaisegkmqeevisfkqiyynvnvneptrpsrffgkavtkeqlqalgvnaenppayissvaygrqvylklstnshstkvkaafdaavsgksvsgdveltniiknssfkaviyggsakdevqiidgnlgdlrdilkkgatfnretpgvpiayttnflkdnelaviknnseyiettskaytdgkinidhsggyvaqfniswdevnyd(seqidno:2)。
在另一个实施例中,该llo片段对应于本文所用的llo蛋白的约aa20-442。
在另一个实施例中,llo片段具有以下序列:
mkkimlvfitlilvslpiaqqteakdasafnkensissvappasppaspktpiekkhadeidkyiqgldynknnvlvyhgdavtnvpprkgykdgneyivvekkkksinqnnadiqvvnaissltypgalvkanselvenqpdvlpvkrdsltlsidlpgmtnqdnkivvknatksnvnnavntlverwnekyaqaysnvsakidyddemaysesqliakfgtafkavnnslnvnfgaisegkmqeevisfkqiyynvnvneptrpsrffgkavtkeqlqalgvnaenppayissvaygrqvylklstnshstkvkaafdaavsgksvsgdveltniiknssfkaviyggsakdevqiidgnlgdlrdilkkgatfnretpgvpiayttnflkdnelaviknnseyiettskaytd(seqidno:4)。
在另一个实施例中,“截短的llo”或“δllo”是指包含pest氨基酸域的llo片段。在另一个实施例中,该术语是指包含pest序列的llo片段。
在另一个实施例中,该术语是指不包含氨基末端的活化域并且不包含胱氨酸484的llo片段。在另一个实施例中,该术语是指非溶血性llo片段。在另一个实施例中,llo片段因活化域的缺失或突变而成为非溶血的。在另一个实施例中,llo片段因半胱氨酸484的缺失或突变而成为非溶血的。在另一个实施例中,llo片段因另一位置的缺失或突变而成为非溶血的。
在另一个实施例中,llo片段由llo蛋白的大约前441个aa构成。在另一个实施例中,llo片段由llo的大约前420个aa构成。在另一个实施例中,该llo片段为llo蛋白的非溶血形式。
在另一个实施例中,llo片段包含对应于以上aa范围之一的同源llo蛋白的残基。残基编号在另一个实施例中不必精确对应以上列举的残基编号;例如,如果该同源llo蛋白相对于本文所用的llo蛋白具有插入或缺失,则可因此而调整该残基编号。
在另一个实施例中,该llo片段为本领域已知的任何其他llo片段。
在另一个实施例中,将重组李斯特菌菌株以1×109-3.31×1010个cfu的剂量施用给人受试者。在另一个实施例中,剂量为5-500×108个cfu。在另一个实施例中,剂量为7-500×108个cfu。在另一个实施例中,剂量为10-500×108个cfu。在另一个实施例中,剂量为20-500×108个cfu。在另一个实施例中,剂量为30-500×108个cfu。在另一个实施例中,剂量为50-500×108个cfu。在另一个实施例中,剂量为70-500×108个cfu。在另一个实施例中,剂量为100-500×108个cfu。在另一个实施例中,剂量为150-500×108个cfu。在另一个实施例中,剂量为5-300×108个cfu。在另一个实施例中,剂量为5-200×108个cfu。在另一个实施例中,剂量为5-150×108个cfu。在另一个实施例中,剂量为5-100×108个cfu。在另一个实施例中,剂量为5-70×108个cfu。在另一个实施例中,剂量为5-50×108个cfu。在另一个实施例中,剂量为5-30×108个cfu。在另一个实施例中,剂量为5-20×108个cfu。在另一个实施例中,剂量为1-30×109个cfu。在另一个实施例中,剂量为1-20×109个cfu。在另一个实施例中,剂量为2-30×109个cfu。在另一个实施例中,剂量为1-10×109个cfu。在另一个实施例中,剂量为2-10×109个cfu。在另一个实施例中,剂量为3-10×109个cfu。在另一个实施例中,剂量为2-7×109个cfu。在另一个实施例中,剂量为2-5×109个cfu。在另一个实施例中,剂量为3-5×109个cfu。
在另一个实施例中,剂量为1×109个生物体。在另一个实施例中,剂量为1.5×109个生物体。在另一个实施例中,剂量为2×109个生物体。在另一个实施例中,剂量为3×109个生物体。在另一个实施例中,剂量为4×109个生物体。在另一个实施例中,剂量为5×109个生物体。在另一个实施例中,剂量为6×109个生物体。在另一个实施例中,剂量为7×109个生物体。在另一个实施例中,剂量为8×109个生物体。在另一个实施例中,剂量为10×109个生物体。在另一个实施例中,剂量为1.5×1010个生物体。在另一个实施例中,剂量为2×1010个生物体。在另一个实施例中,剂量为2.5×1010个生物体。在另一个实施例中,剂量为3×1010个生物体。在另一个实施例中,剂量为3.3×1010个生物体。在另一个实施例中,剂量为4×1010个生物体。在另一个实施例中,剂量为5×1010个生物体。
在另一个实施例中,本发明的方法的重组多肽由重组李斯特菌菌株表达。在另一个实施例中,该表达由重组李斯特菌菌株携带的核苷酸分子介导。
在另一个实施例中,该重组李斯特菌菌株通过编码重组多肽的质粒表达重组多肽。在另一个实施例中,该质粒包含编码细菌转录因子的基因。在另一个实施例中,该质粒编码李斯特菌转录因子。在另一个实施例中,该转录因子为prfa。在另一个实施例中,该prfa为突变体prfa。在另一个实施例中,该prfa包含d133v氨基酸突变。在另一个实施例中,转录因子是本领域已知的任何其他转录因子。
在另一个实施例中,该质粒包含编码代谢酶的基因。在另一个实施例中,该代谢酶为细菌代谢酶。在另一个实施例中,该代谢酶为李斯特菌代谢酶。在另一个实施例中,该代谢酶为氨基酸代谢酶。在另一个实施例中,该氨基酸代谢酶涉及细胞壁合成途径。在另一个实施例中,该代谢酶是d-氨基酸氨基转移酶基因(dat)的产物。在另一个实施例中,该代谢酶是丙氨酸消旋酶基因(dal)的产物。在另一个实施例中,该代谢酶是本领域已知的任何其他代谢酶。
在另一个实施例中,本发明的方法还包括用本发明的重组李斯特菌菌株强化人受试者的步骤。在另一个实施例中,用于强化接种的重组李斯特菌菌株与用于初始的“初免”接种的菌株相同。在另一个实施例中,强化菌株不同于初免菌株。在另一个实施例中,将相同的剂量用于初免和强化接种。在另一个实施例中,将大剂量用于强化。在另一个实施例中,将小剂量用于强化。
在另一个实施例中,本发明的方法还包括用包含e7抗原的免疫原性组合物接种人受试者的步骤。在另一个实施例中,免疫原性组合物包含重组e7蛋白或其片段。在另一个实施例中,免疫原性组合物包含表达重组e7蛋白或其片段的核苷酸分子。在另一个实施例中,在李斯特菌接种之后施用非李斯特菌接种。在另一个实施例中,在李斯特菌接种之前施用非李斯特菌接种。
在另一个实施例中,“强化”是指给受试者施用另外的疫苗剂量。在本发明方法的另一个实施例中,施用2次强化(或总共3次接种)。在另一个实施例中,施用3次强化。在另一个实施例中,施用4次强化。在另一个实施例中,施用5次强化。在另一个实施例中,施用6次强化。在另一个实施例中,施用超过6次强化。
本发明的方法和组合物的重组李斯特菌菌株在另一个实施例中是重组单核细胞增多性李斯特菌菌株。在另一个实施例中,李斯特菌菌株是重组西尔李斯特菌(listeriaseeligeri)菌株。在另一个实施例中,李斯特菌菌株是重组格雷李斯特菌(listeriagrayi)菌株。在另一个实施例中,李斯特菌菌株是重组伊万诺夫李斯特菌(listeriaivanovii)菌株。在另一个实施例中,李斯特菌菌株是重组默里李斯特菌(listeriamurrayi)菌株。在另一个实施例中,李斯特菌菌株是重组威耳逊李斯特菌(listeriawelshimeri)菌株。在另一个实施例中,该李斯特菌菌株是本领域已知的任何其他李斯特菌属菌种的重组菌株。
本发明提供多种用于制备或工程化本发明的减毒李斯特菌的李斯特菌属菌种和菌株。在一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌10403s野生型(参见bishop和hinrichs(1987)j.immunol.139:2005–2009;lauer等人(2002)j.bact.184:4177–4186)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌dp-l4056(噬菌体解离的)(参见lauer等人(2002)j.bact.184:4177–4186)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌dp-l4027,其为噬菌体解离的并为hly基因缺失的(参见lauer等人(2002)j.bact.184:4177–4186;jones和portnoy(1994)infect.immunity65:5608–5613)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌dp-l4029,其为噬菌体解离的、acta缺失的(参见lauer等人(2002)j.bact.184:4177–4186;skoble等人(2000)j.cellbiol.150:527–538)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌dp-l4042(δpest)(参见brockstedt等人(2004)proc.natl.acad.sci.usa101:13832–13837;支持信息)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌dp-l4097(llo-s44a)(参见brockstedt等人(2004)proc.natl.acad.sci.usa101:13832–13837;支持信息)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌dp-l4364(δlpla;脂蛋白连接酶(lipoateproteinligase))(参见brockstedt等人(2004)proc.natl.acad.sci.usa101:13832–13837;支持信息)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌dp-l4405(δinla)(参见brockstedt等人(2004)proc.natl.acad.sci.usa101:13832–13837;支持信息)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌dp-l4406(δinlb)(参见brockstedt等人(2004)proc.natl.acad.sci.usa101:13832–13837;支持信息)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌cs-l0001(δacta-deltainlb)(参见brockstedt等人(2004)proc.natl.acad.sci.usa101:13832–13837;支持信息)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌cs-l0002(δacta-deltalpla)(参见brockstedt等人(2004)proc.natl.acad.sci.usa101:13832–13837;支持信息)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌cs-l0003(l461t-δlpla)(参见brockstedt等人(2004)proc.natl.acad.sci.usa101:13832–13837;支持信息)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌dp-l4038(δacta-llol461t)(参见brockstedt等人(2004)proc.natl.acad.sci.usa101:13832–13837;支持信息)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌dp-l4384(s44a-llol461t)(参见brockstedt等人(2004)proc.natl.acad.sci.usa101:13832–13837;支持信息)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌。脂蛋白中的突变(参见o'riordan等人(2003)science302:462–464)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌dp-l4017(10403shly(l461t),其在溶血素基因中具有点突变(参见美国临时专利申请no.60/490,089,2003年7月24日提交)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌egd(参见genbank登录号al591824)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌egd-e(参见genbank登录号nc_003210,atcc登录号baa-679)。在另一个实施例中,李斯特菌菌株为uvrab缺失的单核细胞增多性李斯特菌dp-l4029(参见2004年2月2日提交的美国临时专利申请no.60/541,515、2003年7月24日提交的美国临时专利申请no.60/490,080)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌acta-/inlb-双突变体(参见atcc登录号pta-5562)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌lpla突变体或hly突变体(参见授予portnoy等人的美国专利申请no.20040013690)。在另一个实施例中,李斯特菌菌株为单核细胞增多性李斯特菌dal/dat双突变体。(参见授予frankel和portnoy的美国专利申请no.20050048081)。本发明涵盖包含上述李斯特菌菌株的试剂和方法,以及例如通过质粒和/或通过基因组整合进行修饰以包含以下核酸的这些菌株,即编码以下基因之一或其任意组合的核酸:hly(llo,李斯特菌溶血素)、iap(p60)、inla、inlb、inlc、dal(丙氨酸消旋酶)、dat(d-氨基酸氨基转移酶)、plca、plcb、acta;或介导以下方面的核酸:单壁囊泡的生长、传播、破裂,双壁囊泡的破裂,与宿主细胞的结合,由宿主细胞摄取。本发明不受上文公开的特定菌株的限制。
在另一个实施例中,本发明的重组李斯特菌菌株已在动物宿主中传代。在另一个实施例中,该传代让该菌株作为疫苗载体的效能最佳化。在另一个实施例中,该传代稳定李斯特菌菌株的免疫原性。在另一个实施例中,该传代稳定李斯特菌菌株的毒力。在另一个实施例中,该传代增强李斯特菌菌株的免疫原性。在另一个实施例中,该传代增强李斯特菌菌株的毒力。在另一个实施例中,该传代除去李斯特菌菌株的不稳定亚株。在另一个实施例中,该传代降低李斯特菌菌株的不稳定亚株的普遍性。在另一个实施例中,李斯特菌菌株包含编码含抗原重组肽的基因的基因组插入。在另一个实施例中,李斯特菌菌株携带包含编码含抗原重组肽的基因的质粒。在另一个实施例中,该传代如本文所述进行(例如,在实例12中)。在另一个实施例中,该传代通过本领域已知的任何其他方法进行。
在另一个实施例中,用于本发明的方法中的重组李斯特菌菌株已储存在冷冻细胞库中。在另一个实施例中,重组李斯特菌菌株已储存在冻干细胞库中。
在另一个实施例中,本发明的方法和组合物的细胞库为主细胞库。在另一个实施例中,细胞库为工作细胞库。在另一个实施例中,细胞库为良好生产规范(gmp)细胞库。在另一个实施例中,细胞库旨在用于制备临床级材料。在另一个实施例中,细胞库符合人用监管规范。在另一个实施例中,细胞库是本领域已知的任何其他类型的细胞库。
“良好生产规范”在另一个实施例中由美国联邦法规(unitedstatescodeoffederalregulations)的(21cfr210–211)定义。在另一个实施例中,“良好生产规范”由生产临床级材料或供人类消耗的其他标准定义,例如美国之外的国家/地区的标准。
在另一个实施例中,本发明的方法中所用的重组李斯特菌菌株来自一批疫苗剂量。
在另一个实施例中,本发明的方法中所用的重组李斯特菌菌株来自由美国专利no.8,114,414中所公开的方法制备的冷冻或冻干储备品,该专利以引用方式并入本文。
在另一个实施例中,本发明的肽为融合肽。在另一个实施例中,“融合肽”是指包含2个或更多个通过肽键或其他化学键连接在一起的蛋白的肽或多肽。在另一个实施例中,蛋白通过肽或其他化学键直接连接在一起。在另一个实施例中,蛋白通过2个或更多个蛋白之间的1个或更多个氨基酸(例如,“间隔物”)连接在一起。
在另一个实施例中,本发明的疫苗还包含佐剂。在另一个实施例中,本发明的方法和组合物所用的佐剂是粒细胞/巨噬细胞集落刺激因子(gm-csf)蛋白。在另一个实施例中,该佐剂包含gm-csf蛋白。在另一个实施例中,该佐剂为编码gm-csf的核苷酸分子。在另一个实施例中,该佐剂为包含编码gm-csf的核苷酸分子。在另一个实施例中,该佐剂为皂苷qs21。在另一个实施例中,该佐剂包含皂苷qs21。在另一个实施例中,该佐剂为单磷酰脂质a。在另一个实施例中,该佐剂包含单磷酰脂质a。在另一个实施例中,该佐剂为sbas2。在另一个实施例中,该佐剂包含sbas2。在另一个实施例中,该佐剂为含有未甲基化的cpg的寡核苷酸。在另一个实施例中,该佐剂包含含有未甲基化的cpg的寡核苷酸。在另一个实施例中,该佐剂为免疫刺激性细胞因子。在另一个实施例中,该佐剂包含免疫刺激性细胞因子。在另一个实施例中,该佐剂为编码免疫刺激性细胞因子的核苷酸分子。在另一个实施例中,该佐剂包含编码免疫刺激性细胞因子的核苷酸分子。在另一个实施例中,该佐剂为或包含刺糖苷(quillglycoside)。在另一个实施例中,该佐剂为或包含细菌有丝分裂原。在另一个实施例中,该佐剂为或包含细菌毒素。在另一个实施例中,该佐剂为或包含本领域已知的任何其他佐剂。
在另一个实施例中,本发明的核苷酸可操作地连接至驱动编码肽在李斯特菌菌株中表达的启动子/调控序列。可用于驱动基因的组成型表达的启动子/调控序列是本领域熟知的,并且包括但不限于例如李斯特菌的phlya、pacta和p60启动子,链球菌(streptococcus)bac启动子,灰色链霉菌(streptomycesgriseus)sgia启动子以及苏云金芽孢杆菌(b.thuringiensis)phaz启动子。在另一个实施例中,编码本发明的肽的核酸的诱导型和组织特异性表达通过将编码该肽的核酸置于诱导型或组织特异性启动子/调控序列的调控下实现。可用于此目的的组织特异性或诱导型启动子/调控序列的例子包括但不限于mmtvltr诱导型启动子和sv40晚期增强子/启动子。在另一个实施例中,采用响应于诱导剂(诸如金属、糖皮质素等等)而诱导的启动子。因此,应当理解,本发明包括使用任何已知或未知的并且能够驱动与其可操作地连接的所需蛋白的表达的启动子/调控序列。
本发明的方法和组合物中使用的acta蛋白的n末端片段在另一个实施例中具有seqidno:5:
mrammvvfitancitinpdiifaatdsedsslntdeweeekteeqpsevntgpryetarevssrdikeleksnkvrntnkadliamlkekaekgpninnnnseqtenaaineeasgadrpaiqverrhpglpsdsaaeikkrrkaiassdselesltypdkptkvnkkkvakesvadasesdldssmqsadesspqplkanqqpffpkvfkkikdagkwvrdkidenpevkkaivdksaglidqlltkkkseevnasdfpppptdeelrlalpetpmllgfnapatsepssfefpppptdeelrlalpetpmllgfnapatsepssfefpppptedeleiiretassldssftrgdlaslrnainrhsqnfsdfppipteeelngrggrp。在另一个实施例中,acta片段包含seqidno:5.在另一个实施例中,acta片段为本领域已知的任何其他acta片段。
在另一个实施例中,编码acta蛋白的片段的重组核苷酸包含seqidno:6:
atgcgtgcgatgatggtggttttcattactgccaattgcattacgattaaccccgacataatatttgcagcgacagatagcgaagattctagtctaaacacagatgaatgggaagaagaaaaaacagaagagcaaccaagcgaggtaaatacgggaccaagatacgaaactgcacgtgaagtaagttcacgtgatattaaagaactagaaaaatcgaataaagtgagaaatacgaacaaagcagacctaatagcaatgttgaaagaaaaagcagaaaaaggtccaaatatcaataataacaacagtgaacaaactgagaatgcggctataaatgaagaggcttcaggagccgaccgaccagctatacaagtggagcgtcgtcatccaggattgccatcggatagcgcagcggaaattaaaaaaagaaggaaagccatagcatcatcggatagtgagcttgaaagccttacttatccggataaaccaacaaaagtaaataagaaaaaagtggcgaaagagtcagttgcggatgcttctgaaagtgacttagattctagcatgcagtcagcagatgagtcttcaccacaacctttaaaagcaaaccaacaaccatttttccctaaagtatttaaaaaaataaaagatgcggggaaatgggtacgtgataaaatcgacgaaaatcctgaagtaaagaaagcgattgttgataaaagtgcagggttaattgaccaattattaaccaaaaagaaaagtgaagaggtaaatgcttcggacttcccgccaccacctacggatgaagagttaagacttgctttgccagagacaccaatgcttcttggttttaatgctcctgctacatcagaaccgagctcattcgaatttccaccaccacctacggatgaagagttaagacttgctttgccagagacgccaatgcttcttggttttaatgctcctgctacatcggaaccgagctcgttcgaatttccaccgcctccaacagaagatgaactagaaatcatccgggaaacagcatcctcgctagattctagttttacaagaggggatttagctagtttgagaaatgctattaatcgccatagtcaaaatttctctgatttcccaccaatcccaacagaagaagagttgaacgggagaggcggtagacca中示出的序列。在另一个实施例中,重组核苷酸具有seqidno:6中示出的序列。在另一个实施例中,重组核苷酸包含任何其他的编码acta蛋白的片段的序列。
在本发明的方法和组合物的另一个实施例中,pest氨基酸序列融合至e7或e6抗原。如本文所公开,表达pest氨基酸序列-抗原融合体的重组李斯特菌菌株诱导抗肿瘤免疫(实例3)并产生抗原特异性、肿瘤浸润性t细胞(实例4)。另外,证实了包含抗原和含pest氨基酸序列kensissmappasppaspktpiekkhadeidk(seqidno:1)的llo的融合蛋白增强的细胞介导的免疫。
因此,抗原融合至来源于其他原核生物体的其他lmpest氨基酸序列和pest氨基酸序列也将增强抗原的免疫原性。在另一个实施例中,pest氨基酸序列具有选自seqidno:7-12的序列。在另一个实施例中,pest氨基酸序列为来自lmacta蛋白的pest氨基酸序列。在另一个实施例中,pest氨基酸序列为kteeqpsevntgpr(seqidno:7)、kasvtdtsegdldssmqsadestpqplk(seqidno:8)、kneevnasdfpppptdeelr(seqidno:9)或rggiptseefsslnsgdftddensetteeeidr(seqidno:10)。在另一个实施例中,该pest氨基酸序列来自链球菌属菌种(streptococcussp)的链球菌溶血素o蛋白。在另一个实施例中,pest氨基酸序列来自酿脓链球菌(streptococcuspyogenes)的链球菌溶血素o,例如aa35-51处的kqntastettttneqpk(seqidno:11)。在另一个实施例中,pest氨基酸序列来自类马链球菌(streptococcusequisimilis)的链球菌溶血素o,例如aa38-54处的kqntantettttneqpk(seqidno:12)。在另一个实施例中,该pest氨基酸序列为源自原核生物体的另一pest氨基酸序列。在另一个实施例中,该pest氨基酸序列为本领域已知的任何其他pest氨基酸序列。
可根据诸如由例如rechsteiner和rogers(1996,trendsbiochem.sci.21:267-271)针对lm所述的方法来鉴定其他原核生物体的pest氨基酸序列。或者,来自其他原核生物体的pest氨基酸序列也可基于这种方法来鉴定。可预期其中pest氨基酸序列的其他原核生物体包括但不限于其他的李斯特菌菌种。在另一个实施例中,pest氨基酸序列嵌入抗原性蛋白内。因此,在另一个实施例中,“融合体”是指既包含抗原也包含连接在抗原的一端或嵌入抗原内的pest氨基酸序列的抗原性蛋白。
在另一个实施例中,使用本领域已知的任何其他方法或算法,例如caspredictor(garay-malpartidahm,occhiuccijm,alvesj,belizarioje.bioinformatics.2005年6月;21增刊1:i169-76)来鉴定pest氨基酸序列。在另一个实施例中,采用以下方法:
通过将1的值分配给氨基酸ser、thr、pro、glu、asp、asn或gln,计算每30-35个氨基酸的段的pest指数。每个pest残基的系数值(cv)为1,每个其他氨基酸(非-pest)的系数值为0。
在另一个实施例中,本发明的llo蛋白、acta蛋白或其片段不需要精确地如本文所示的序列那样,而是可以作出其他保持如本文其他地方所示的融合至抗原的llo或acta蛋白的功能特性的改变、修饰或变化。在另一个实施例中本发明利用llo蛋白、acta蛋白或其片段的类似物。类似物在另一个实施例中与天然存在的蛋白或肽的不同之处在于保守氨基酸序列差异或不对序列产生影响的修饰或以上两者。
在另一个实施例中,整个e7蛋白或其片段融合至llo蛋白、acta蛋白或含pest氨基酸序列的肽以生成本发明的方法的重组肽。所用的e7蛋白(作为整体或作为片段的来源)在另一个实施例中具有序列
mhgdtptlheymldlqpettdlycyeqlndsseeedeidgpagqaepdrahynivtfcckcdstlrlcvqsthvdirtledllmgtlgivcpicsqkp(seqidno:13)。在另一个实施例中,该e7蛋白为seqidno:13的同源物。在另一个实施例中,该e7蛋白为seqidno:13的变体。在另一个实施例中,该e7蛋白为seqidno:13的异构体。在另一个实施例中,该e7蛋白为seqidno:13的片段。在另一个实施例中,该e7蛋白为seqidno:13的同源物的片段。在另一个实施例中,该e7蛋白为seqidno:13的变体的片段。在另一个实施例中,该e7蛋白为seqidno:13的异构体的片段。
在另一个实施例中,e7蛋白的序列为:
mhgpkatlqdivlhlepqneipvdllcheqlsdseeendeidgvnhqhlparraepqrhtmlcmcckcearielvvessaddlrafqqlflntlsfvcpwcasqq(seqidno:14)。在另一个实施例中,该e7蛋白为seqidno:14的同源物。在另一个实施例中,该e7蛋白为seqidno:14的变体。在另一个实施例中,该e7蛋白为seqidno:14的异构体。在另一个实施例中,该e7蛋白为seqidno:14的片段。在另一个实施例中,该e7蛋白为seqidno:14的同源物的片段。在另一个实施例中,该e7蛋白为seqidno:14的变体的片段。在另一个实施例中,该e7蛋白为seqidno:14的异构体的片段。
在另一个实施例中,该e7蛋白具有以下genbank条目之一中示出的序列:m24215、nc_004500、v01116、x62843或m14119。在另一个实施例中,该e7蛋白为来自以上genbank条目之一的序列的同源物。在另一个实施例中,该e7蛋白为来自以上genbank条目之一的序列的变体。在另一个实施例中,该e7蛋白为来自以上genbank条目之一的序列的异构体。在另一个实施例中,该e7蛋白为来自以上genbank条目之一的序列的片段。在另一个实施例中,该e7蛋白为来自以上genbank条目之一的序列的同源物的片段。在另一个实施例中,该e7蛋白为来自以上genbank条目之一的序列的变体的片段。在另一个实施例中,该e7蛋白为来自以上genbank条目之一的序列的异构体的片段。
在另一个实施例中,整个e6蛋白或其片段融合至llo蛋白、acta蛋白或含pest氨基酸序列的肽以生成本发明的方法的重组肽。所用的e6蛋白(作为整体或作为片段的来源)在另一个实施例中具有序列
mhqkrtamfqdpqerprklpqlctelqttihdiilecvyckqqllrrevydfafrdlcivyrdgnpyavcdkclkfyskiseyrhycyslygttleqqynkplcdllircincqkplcpeekqrhldkkqrfhnirgrwtgrcmsccrssrtrretql(seqidno:15)。在另一个实施例中,该e6蛋白为seqidno:15的同源物。在另一个实施例中,该e6蛋白为seqidno:15的变体。在另一个实施例中,该e6蛋白为seqidno:15的异构体。在另一个实施例中,该e6蛋白为seqidno:15的片段。在另一个实施例中,该e6蛋白为seqidno:15的同源物的片段。在另一个实施例中,该e6蛋白为seqidno:15的变体的片段。在另一个实施例中,该e6蛋白为seqidno:15的异构体的片段。
在另一个实施例中,e6蛋白的序列为:
marfedptrrpyklpdlctelntslqdieitcvycktvleltevfefafkdlfvvyrdsiphaachkcidfysrirelrhysdsvygdtlekltntglynllirclrcqkplnpaeklrhlnekrrfhniaghyrgqchsccnrarqerlqrrretqv(seqidno:16)。在另一个实施例中,该e6蛋白为seqidno:16的同源物。在另一个实施例中,该e6蛋白为seqidno:16的变体。在另一个实施例中,该e6蛋白为seqidno:16的异构体。在另一个实施例中,该e6蛋白为seqidno:16的片段。在另一个实施例中,该e6蛋白为seqidno:16的同源物的片段。在另一个实施例中,该e6蛋白为seqidno:16的变体的片段。在另一个实施例中,该e6蛋白为seqidno:16的异构体的片段。
在另一个实施例中,该e6蛋白具有以下genbank条目之一中示出的序列:m24215、m14119、nc_004500、v01116、x62843或m14119。在另一个实施例中,该e6蛋白为来自以上genbank条目之一的序列的同源物。在另一个实施例中,该e6蛋白为来自以上genbank条目之一的序列的变体。在另一个实施例中,该e6蛋白为来自以上genbank条目之一的序列的异构体。在另一个实施例中,该e6蛋白为来自以上genbank条目之一的序列的片段。在另一个实施例中,该e6蛋白为来自以上genbank条目之一的序列的同源物的片段。在另一个实施例中,该e6蛋白为来自以上genbank条目之一的序列的变体的片段。在另一个实施例中,该e6蛋白为来自以上genbank条目之一的序列的异构体的片段。
在另一个实施例中,“同源性”是指与llo序列(如,与seqidno:2-4之一)的同一性大于70%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于64%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于68%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于72%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于75%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于78%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于80%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于82%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于83%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于85%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于87%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于88%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于90%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于92%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于93%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于95%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于96%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于97%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于98%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性大于99%。在另一个实施例中,“同源性”是指与seqidno:2-4之一的同一性为100%。
在另一个实施例中,“同源性”是指与e7序列(如,与seqidno:13-14之一)的同一性大于70%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于62%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于64%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于68%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于72%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于75%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于78%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于80%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于82%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于83%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于85%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于87%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于88%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于90%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于92%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于93%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于95%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于96%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于97%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于98%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性大于99%。在另一个实施例中,“同源性”是指与seqidno:13-14之一的同一性为100%。
在另一个实施例中,“同源性”是指与e6序列(如,与seqidno:15-16之一)的同一性大于70%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于64%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于68%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于72%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于75%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于78%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于80%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于82%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于83%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于85%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于87%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于88%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于90%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于92%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于93%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于95%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于96%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于97%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于98%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性大于99%。在另一个实施例中,“同源性”是指与seqidno:15-16之一的同一性为100%。
在另一个实施例中,“同源性”是指与pest氨基酸序列(如,与seqidno:1和7-12之一)或与acta序列(如,与seqidno:5-6之一)的同一性大于70%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于60%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于64%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于68%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于72%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于75%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于78%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于80%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于82%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于83%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于85%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于87%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于88%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于90%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于92%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于93%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于95%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于96%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于97%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于98%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性大于99%。在另一个实施例中,“同源性”是指与seqidno:1和7-12或seqidno:5-6之一的同一性为100%。
在一个实施例中,通过本领域充分描述的方法确定蛋白和/或肽对本文列出的任何氨基酸序列的同源性,包括免疫印迹分析,或通过建立的方法采用多种可用软件包中的任一种通过氨基酸序列的计算机算法分析进行。这些软件包中的一些例如包括fasta、blast、mpsrch或scanps软件包,并且在其他实施例中采用smith和waterman算法,和/或整体/局部或blocks比对用于分析。
在另一个实施例中,llo蛋白、acta蛋白或其片段通过化学偶联连接到抗原。在另一个实施例中,将戊二醛用于偶联。在另一个实施例中,偶联采用本领域已知的任何合适的方法进行。
在另一个实施例中,本发明的融合蛋白通过任何合适的方法制备,这些方法包括例如克隆和适当序列的限制或通过下文讨论的方法直接化学合成。在另一个实施例中,克隆子序列,并使用适当的限制性酶切割适当的子序列。在另一个实施例中,然后连接片段生成所需的dna序列。在另一个实施例中,编码融合蛋白的dna使用dna扩增法例如聚合酶链式反应(pcr)制备。首先,在天然dna片段的新末端的任一侧单独扩增。一个扩增的序列的5'末端编码肽接头,而另一个扩增的序列的3'末端也编码肽接头。由于第一片段的5'末端与第二片段的3'末端互补,两个片段(在部分纯化后,如在lmp琼脂糖上)可在第三pcr反应中用作重叠模板。扩增的序列将包含密码子、开口位点羧基侧上的片段(现在形成氨基序列)、接头和开口位点氨基侧上的序列(现在形成羧基序列)。然后将插入片段连接进质粒。
在另一个实施例中,llo蛋白、acta蛋白或其片段以及抗原或其片段通过本领域技术人员已知的方式偶联。在另一个实施例中,抗原或其片段直接地或通过接头(间隔物)偶联到acta蛋白或llo蛋白。在另一个实施例中,嵌合分子作为单链融合蛋白以重组方式表达。
在另一个实施例中,本发明的融合肽使用标准化学肽合成技术而合成。在另一个实施例中,嵌合分子作为单个连续多肽合成。在另一个实施例中,llo蛋白、acta蛋白或其片段以及抗原或其片段单独地合成,然后通过一个分子的氨基末端与另一个分子的羧基末端的缩合而融合,从而形成肽键。在另一个实施例中,acta蛋白或llo蛋白和抗原各自与肽间隔分子的一端缩合,从而形成连续的融合蛋白。
在另一个实施例中,本发明的肽和蛋白通过固相肽合成(spps)而制备,如由stewart等人在solidphasepeptidesynthesis,第2版,1984,piercechemicalcompany,rockford,ill.中所述,或由bodanszky和bodanszky(thepracticeofpeptidesynthesis,1984,springer-verlag,newyork)所述。在另一个实施例中,适当保护的氨基酸残基通过其羧基基团连接到衍生化的不溶性聚合物支持物,诸如交联的聚苯乙烯或聚酰胺树脂。“适当保护的”是指在氨基酸的α-氨基基团和任何侧链官能团上均存在保护基团。侧链保护基团通常是对整个合成过程中所用的溶剂、试剂和反应条件稳定的,并且可在不会影响最终肽产物的条件下移除。寡肽的逐步合成通过以下方式进行:从初始氨基酸移除n保护基团,然后将所需肽的序列中的下一个氨基酸的羧基端偶合到其上。该氨基酸也加以适当保护。可使新来的氨基酸的羧基失活以与支持物结合的氨基酸的n末端反应,反应方式是通过形成反应性基团,诸如形成碳二亚胺、对称酸酐或“活性酯”基团,诸如羟基苯并三唑或五氟苯基酯。
在另一个实施例中,本发明提供一种试剂盒,该试剂盒包括本发明的疫苗、施用装置和描述本发明的使用方法的说明性材料。虽然下文描述了模型试剂盒,但是其他可用试剂盒的内容物根据本公开对技术人员而言将是显而易见的。这些试剂盒中的每一个代表了本发明的独立实施例。
如本文所用,术语“约”是数量术语,意指加或减5%,或在另一个实施例中,加或减10%,或在另一个实施例中,加或减15%,或在另一个实施例中,加或减20%。
在一个实施例中,术语“受试者”是指需要治疗病症或其后遗症或易受该病症或其后遗症影响的哺乳动物(包括人)。受试者可包括狗、猫、猪、牛、绵羊、山羊、马、大鼠和宠物小鼠以及人。受试者还可包括家畜。在一个实施例中,术语“受试者”不排除在所有方面都健康并且不具有或显示出疾病或病症的体征的个体。
为了更充分地举例说明本发明的优选实施例,提供了以下实例。然而,它们绝不应解释为限制本发明的广泛范围。
实验细节章节
(a)实例1:llo-抗原融合蛋白诱导抗肿瘤免疫
材料和实验方法(实例1-2)
细胞系
c57bl/6同基因tc-1肿瘤经hpv-16e6和e7永生化并以c-ha-ras癌基因转化。t.c.wu(johnshopkinsuniversityschoolofmedicine,baltimore,md)所公开的tc-1为高度致瘤的肺上皮细胞,该细胞表达低水平的hpv-16e6和e7,并被c-ha-ras癌基因转化。让tc-1于37℃及10%co2下生长在rpmi1640、10%fcs、2mml-谷氨酰胺、100u/ml青霉素、100μg/ml链霉素、100μm非必需氨基酸、1mm丙酮酸钠、50微摩尔(mcm)2-me、400微克(mcg)/mlg418以及10%国家典型菌种保藏中心-109培养基(nationalcollectiontypeculture-109medium)中。c3为来自c57bl/6小鼠的小鼠胚胎细胞,该细胞经完整hpv16基因组永生化并经pej-ras转化。el-4/e7为经e7逆转录病毒转导的胸腺瘤el-4。
单核细胞增多性李斯特菌菌株和繁殖
所用的李斯特菌菌株为lm-llo-e7(在游离型表达系统中的hly-e7融合基因;图1a)、lm-e7(整合进李斯特菌基因组中的单拷贝e7基因盒)、lm-llo-np(“dp-l2028”;在游离型表达系统中的hly-np融合基因)和lm-gag(“zy-18”;整合进染色体中的单拷贝hiv-1gag基因盒)。将e7用引物5'-ggctcgagcatggagatacacc-3'(seqidno:17;xhoi位点标有下划线)和5'-ggggactagtttatggtttctgagaaca-3'(seqidno:18;spei位点标有下划线)通过pcr扩增并连接进pcr2.1(invitrogen,sandiego,ca)。通过xhoi/spei消化从pcr2.1切离e7,并将其连接进pgg-55。将hly-e7融合基因和多能转录因子prfa克隆进pam401,即一种多拷贝穿梭质粒(wirthr等人,jbacteriol,165:831,1986),从而生成pgg-55。hly启动子驱动hly基因产物的前441个氨基酸(缺乏溶血性c-端,在下文中被称作“δllo”,并具有seqidno:25中示出的序列)的表达,其通过xhoi位点连接至e7基因,从而产生hly-e7融合基因,该基因被转录和分泌成llo-e7。用为了在体内保留质粒而选择的pgg-55转化prfa阴性李斯特菌菌株xfl-7(由haoshen博士所公开,universityofpennsylvania)(图1a-b)。使用引物5'-gggggctagccctcctttgattagtatattc-3'(seqidno:19;nhei位点标有下划线)和5'-ctccctcgagatcataatttacttcatc-3'(seqidno:20;xhoi位点标有下划线)生成hly启动子和基因片段。将prfa基因用引物5'-gactacaaggacgatgaccgacaagtgataacccgggatctaaataaatccgttt-3'(seqidno:27;xbai位点标有下划线)和5'-cccgtcgaccagctcttcttggtgaag-3'(seqidno:21;sali位点标有下划线)进行pcr扩增。通过将含有驱动e7的表达和分泌的hly启动子和信号序列的表达盒引入lm基因组的orfz域中,产生lm-e7。将e7用引物5'-gcggatcccatggagatacacctac-3'(seqidno:22;bamhi位点标有下划线)和5'-gctctagattatggtttctgag-3'(seqidno:23;xbai位点标有下划线)通过pcr扩增。然后将e7连接进pzy-21穿梭载体。用得到的质粒pzy-21-e7转化lm菌株10403s,该质粒包括插入在与lm基因组的orfx、y、z域对应的1.6-kb序列的中央的表达盒。该同源域允许e7基因盒通过同源重组插入orfz域中。针对e7基因盒在orfz域中的整合,筛选克隆。在含有(lm-llo-e7和lm-llo-np)或不含(lm-e7和zy-18)氯霉素(20μg/ml)的脑心浸液培养基中培养细菌。在-80℃以等分试样冷冻细菌。通过western印迹法验证表达(图2)。
western印迹法
使李斯特菌菌株在luria-bertoni培养基中于37℃生长,并在600nm处测得的相同光密度下收获。将上清液tca沉淀,并重悬于补充了0.1nnaoh的1x样品缓冲液中。将相同量的每种细胞沉淀物或每种经tca沉淀的上清液上样至4-20%tris-甘氨酸sds-page凝胶(novex,sandiego,ca)。将凝胶转移至聚偏二氟乙烯,并用抗-e7单克隆抗体(mab)(zymedlaboratories,southsanfrancisco,ca)探测,然后与hrp-偶联的抗小鼠二抗(amershampharmaciabiotech,littlechalfont,u.k.)一起温育,用amershamecl检测试剂显影,并暴露于hyperfilm(amershampharmaciabiotech)。
肿瘤生长的测量
每隔一天用卡尺跨最短和最长表面直径测量肿瘤。将这两个测量结果的平均值作为平均肿瘤直径(以毫米计)相对于不同时间点作图。当肿瘤直径达到20mm时,处死小鼠。仅显示存活小鼠在每个时间点的肿瘤测量结果。
李斯特菌重组体对建立的肿瘤生长的影响
6-8周龄c57bl/6小鼠(charlesriver)在左胁腹皮下接受2×105个tc-1细胞。肿瘤接种后一周,肿瘤已经达到直径为4-5mm的可触知大小。然后在第7和14天用0.1ld50腹膜内lm-llo-e7(107个cfu)、lm-e7(106个cfu)、lm-llo-np(107个cfu)或lm-gag(5×105个cfu)处理8只小鼠的组。
51cr释放测定
以0.1ld50lm-llo-e7、lm-e7、lm-llo-np或lm-gag腹膜内免疫6-8周龄c57bl/6小鼠。免疫接种后十天,收获脾。将经照射的tc-1细胞作为饲养细胞,在培养物中建立脾细胞(100:1,脾细胞:tc-1);在体外刺激5天,然后用在标准51cr释放测定中,其中使用下述靶标:el-4、el-4/e7或经e7h-2b肽(rahynivtf)脉冲处理的el-4。一式三份地进行的e:t细胞比率为:80:1、40:1、20:1、10:1、5:1和2.5:1。在37℃温育4h以后,将细胞沉淀,并从每个孔取出50μl上清液。以wallac1450闪烁计数器(gaithersburg,md)测定样品。将特异性裂解百分比确定为[(每分钟的实验计数(cpm)-自发cpm)/(总cpm-自发cpm)]×100。
tc-1特异性增殖
将c57bl/6小鼠用0.1ld50免疫,并在20天后通过腹膜内注射1ld50lm-llo-e7、lm-e7、lm-llo-np或lm-gag进行强化。强化后六天,从经免疫的小鼠和未暴露的小鼠收获脾。以2.5×104、1.25×104、6×103或3×103个经照射的tc-1细胞/孔作为e7ag的来源,或不用tc-1细胞或用10μg/mlcona,在平底96孔平板中以5×105个/孔在培养物中建立脾细胞。45h后以0.5μci[3h]胸苷/孔脉冲处理细胞。18h以后,采用tomtec采集器96(orange,ct)收获平板,并用wallac1450闪烁计数器评估增殖。将以cpm计的变化计算为实验cpm–无agcpm。
流式细胞术分析
将c57bl/6小鼠用0.1ld50lm-llo-e7或lm-e7静脉内(i.v.)免疫,并在30天后强化。采用带
b16f0-ova实验
给24只c57bl/6小鼠接种5×105个b16f0-ova细胞。在第3、10和17天,将8只小鼠的组用0.1ld50lm-ova(106个cfu)、lm-llo-ova(108个cfu)免疫,另八只动物保持不处理。
统计
为了对比肿瘤直径,确定每个组的肿瘤大小的平均值和sd,并通过学生t检验确定统计显著性。p≤0.05视作显著的。
结果
对比了lm-e7和lm-llo-e7影响tc-1生长的能力。在c57bl/6小鼠的左胁腹上建立皮下肿瘤。七天后,肿瘤已经达到可触知的大小(4-5mm)。在第7和14天,给小鼠接种0.1ld50lm-e7、lm-llo-e7或作为对照的lm-gag和lm-llo-np。lm-llo-e7诱导75%的建立的tc-1肿瘤的完全消退,而在该组中的其他2只小鼠中控制了肿瘤生长(图3)。相反,使用lm-e7和lm-gag的免疫接种没有诱导肿瘤消退。该实验重复多次,总是得到非常类似的结果。另外,在不同的免疫方案下,lm-llo-e7也得到了类似的结果。在另一实验中,单次免疫能够治愈小鼠建立的5mmtc-1肿瘤。
在其他实验中,用另外2个表达e7的肿瘤细胞系得到了类似的结果:c3和el-4/e7。为了确认以lm-llo-e7接种的效力,分别在第60天或第40天用tc-1或el-4/e7肿瘤细胞重新挑战已经消除了它们的肿瘤的动物。经lm-llo-e7免疫的动物保持无肿瘤直到实验终止(就tc-1而言,第124天;就el-4/e7而言,第54天)。
因此,抗原作为与δllo的融合蛋白的表达会增强所述抗原的免疫原性。
实例2:lm-llo-e7处理引起tc-1特异性脾细胞增殖
为了用lm-llo-e7测量lm-e7对t细胞的诱导,在经免疫的小鼠中测量了tc-1特异性增殖性应答(抗原特异性免疫活性的度量)。来自经lm-llo-e7免疫的小鼠的脾细胞当以20:1、40:1、80:1和160:1的脾细胞:tc-1比率暴露于经照射的tc-1细胞(作为e7的来源)时发生增殖(图4)。反之,来自经lm-e7和rlm对照免疫的小鼠的脾细胞仅表现出背景水平的增殖。
实例3:e7与llo、acta或pest氨基酸序列的融合增强e7特异性免疫并产生浸润肿瘤的e7特异性cd8+细胞
材料和实验方法
在12只c57bl/6小鼠(n=3)的左胁腹皮下植入500mcl(微升)
在存在布雷菲德菌素a的情况下以107个细胞/ml将脾细胞和肿瘤细胞与1微摩尔(mcm)e7肽一起温育5小时。将细胞洗涤两次,并在50微升的抗小鼠fc受体上清液(2.4g2)中4℃温育1小时或过夜。针对表面分子cd8和cd62l将细胞染色,渗透化,使用渗透化试剂盒
对于四聚体染色,让h-2db四聚体负载藻红蛋白(pe)偶联的e7肽(rahynivtf,seqidno:24),在室温染色1小时,并用抗别藻蓝蛋白(apc)偶联的mel-14(cd62l)和fitc偶联的cd8
结果
为了分析lm-acta-e7增强抗原特异性免疫的能力,给小鼠植入tc-1肿瘤细胞,并用lm-llo-e7(1×107个cfu)、lm-e7(1×106个cfu)或lm-acta-e7(2×108个cfu)免疫,或者不处理(未暴露的)。来自lm-llo-e7和lm-acta-e7组的小鼠肿瘤含有比在lm-e7或未暴露的小鼠中更高百分比的分泌ifn-γ的cd8+细胞(图5a)和四聚体特异性cd8+细胞(图5b)。
在另一个实验中,给荷瘤小鼠施用lm-llo-e7、lm-pest-e7、lm-δpest-e7或lm-e7epi,并测量肿瘤内的e7特异性淋巴细胞的水平。在第7和14天用0.1ld50的4种疫苗处理小鼠。在第21天收获肿瘤,并用针对cd62l、cd8的抗体和用e7/db四聚体染色。在接种lm-llo-e7和lm-pest-e7的小鼠中观察到四聚体阳性的淋巴细胞在肿瘤内增加的百分比(图6a)。该结果在三个实验中是可再现的(图6b)。
因而,lm-llo-e7、lm-acta-e7和lm-pest-e7各自有效地诱导浸润肿瘤的cd8+t细胞和肿瘤消退。
实例4:李斯特菌疫苗载体通过小鼠传代引起对异源性和内源性抗原的增强的免疫应答
材料和实验方法
细菌菌株
单核细胞增多性李斯特菌菌株10403s血清型1(atcc,manassas,va.)是用于这些研究的野生型生物体和下述构建体的亲本菌株。菌株10403s在腹膜内注入balb/c小鼠时具有大约5×104个cfu的ld50。“lm-gag”是包含hiv-1菌株hxb(具有形成合胞体的表型的b亚型实验室菌株)gag基因的拷贝的重组lm菌株,该基因使用经修饰的穿梭载体pksv7稳定整合进李斯特菌染色体。gag蛋白由该菌株表达和分泌,如通过western印迹所测定。所有菌株均在脑心浸液(bhi)肉汤或琼脂板(difcolabs,detroit,mich)中生长。
细菌培养
选择来自表达过客抗原(passengerantigen)和/或融合蛋白的单个克隆的细菌并在bhi肉汤中培养过夜。将该培养物的等分试样在-70℃下冷冻,不添加添加剂。通过该储备液,使培养物生长到600nm下0.1-0.2的o.d.,并将等分试样在-70℃下再次冷冻,不添加添加剂。为了制备克隆的细菌库,使用以上程序,但在每次传代后,选择多个细菌克隆,并检查靶抗原的表达,如本文所述。将在其中确认了异源抗原的表达的克隆用于下一次传代。
细菌在小鼠中传代
6-8周龄雌性balb/c(h-2d)小鼠购自jacksonlaboratories(barharbor,me)并维持在无病原体的微隔离环境中。冷冻储存在-70℃下的储备培养物的等分试样中的活细菌滴度通过在解冻后且在使用前在bhi琼脂板上铺板而测定。总计,将5×105个细菌静脉内注射进balb/c小鼠。3天后,收获脾脏,匀化,并将脾脏匀浆的系列稀释样在bhi肉汤中温育过夜,然后在bhi琼脂板上铺板。对于进一步传代,再次使等分试样生长到0.1-0.2o.d.,在-70℃下冷冻,并再次通过系列稀释来测定细菌滴度。在初始传代(0次传代)后,将该过程总共重复4次。
针对ifn-γ的胞内细胞因子染色
将淋巴细胞在补充了50u/ml人重组il-2和1微升/mlbrefeldina(golgistoptm;pharmingen,sandiego,ca)的完全rpmi-10培养基中在存在或不存在hiv-gag的细胞毒性t细胞培(ctl)表位(amqmlketi;seqidno:25)、李斯特菌llo(gykdgneyi;seqidno:26)或hpv病毒基因e7(rahynivtf;seqidno:24)的情况下以1微摩尔的浓度培养5小时。首先对细胞进行表面染色,然后洗涤并接受使用cytofix/cytoperm试剂盒根据制造商建议(pharmingen,sandiego,ca)进行的胞内细胞因子染色。对于胞内ifn-γ染色,使用了fitc偶联的大鼠抗小鼠ifn-γ单克隆抗体(克隆xmg1.2)及其同种型对照ab(大鼠igg1;均得自pharmingen)。总计,在4℃下将106个细胞在含有1%牛血清白蛋白和0.02%叠氮化钠的pbs(facs缓冲液)中染色30分钟,然后在facs缓冲液中洗涤3次。在facscantm流式细胞仪或facscaliburtm仪器(bectondickinson,sanjose,ca)上采集样品数据。使用facscaliburtm流式细胞仪进行cd8(percp偶联的,大鼠抗小鼠,克隆53-6.7pharmingen,sandiego,calif.)、cd62l(apc偶联的,大鼠抗小鼠,克隆mel-14)和胞内ifn-γ的三色流式细胞术,并通过cellquestsoftware(bectondickinson,mountainview,ca)对数据进行进一步分析。将细胞进行cd8高和cd62l低设门,然后分析它们的cd8+和胞内ifn-γ染色。
结果
在小鼠中传代增强重组单核细胞增多性李斯特菌的毒力
将三个不同的构建体用于确定传代对重组李斯特菌疫苗载体的影响。这些构建体中的两个携带过客抗原的基因组插入:第一个包含hivgag基因(lm-gag),而第二个包含hpve7基因(lm-e7)。第三个(lm-llo-e7)包含的质粒具有与截短形式的llo融合的过客抗原(hpve7)的融合基因和编码prfa的基因,prfa是控制李斯特菌毒力因子的正调控因子。该质粒用于补充prfa阴性突变体,以使得在活的宿主中,选择压力将有利于质粒保种,因为没有质粒细菌则无毒性。所有3个构建体均已在体外在细菌中广泛繁殖了许多代。
细菌传代导致了细菌毒力的增加,如通过脾脏中的存活细菌数所测量,每一者都是前2代。对于lm-gag和lm-llo-e7,毒力随着每次传代而增加,直至第2代(图7a)。含质粒的构建体lm-llo-e7展现出最显著的毒力增加。在传代前,lm-llo-e7的初始免疫剂量必须增至107个细菌,并且脾脏必须在第2天收获以便回收细菌(而对于lm-gag的105个细菌的初始剂量则在第3天收获)。在初始传代后,lm-llo-e7的标准剂量足以使得可以在第3天收获。对于lm-e7,经未传代的细菌,毒力增加1.5个数量级(图7b)。
因此,通过小鼠传代增加了李斯特菌疫苗菌株的毒力。
传代增加单核细胞增多性李斯特菌诱导cd8+t细胞的能力
接下来,使用hiv-gag肽amqmlketi(seqidno:25)和llo91-99(gykdgneyi;seqidno:26),通过用mhci类特异性免疫显性表位进行细胞内细胞因子染色,而确定传代对抗原特异性cd8+t细胞的作用。将103个cfu的传代细菌(lm-gag)注射进小鼠产生了大量的hiv-gag特异性cd8+t细胞,而相同剂量的未传代lm-gag未诱导可检测的gag特异性cd8+t细胞。甚至将未传代细菌的剂量增加100倍也未补偿它们的相对毒力;事实上,甚至在更高的剂量下也未引起可检测的gag特异性cd8+t细胞。采用传代细菌的相同剂量将gag特异性t细胞诱导增加了50%(图8)。采用llo特异性cd8+t细胞,观察到了相同的抗原特异性cd8+t细胞诱导模式,表明这些结果不是由过客抗原的性质导致的,因为它们在用内源性李斯特菌抗原llo时观察到。
因此,通过小鼠传代增加了李斯特菌疫苗菌株的免疫原性。
实例5:含prfa的质粒在不存在抗生素的情况下在具有prfa缺失的lm菌株中是稳定的
材料和实验方法
细菌
单核细胞增多性李斯特菌菌株xfl7在prfa基因中包含300个碱基对缺失。xfl7携带部分地恢复毒力并赋予cap抗性的pgg55,并在美国专利申请公开no.200500118184中有所描述。
从李斯特菌提取质粒的方案开发
将1ml单核细胞增多性李斯特菌lm-llo-e7研究工作细胞库病毒接种到含有34μg/mlcap的27mlbh1培养基中,并在37℃和200rpm下生长24小时。
使七个2.5ml的培养物样品沉淀(15000rpm持续5分钟),并将沉淀物在37℃下与50μl溶菌酶溶液温育从0-60分钟的不同时长。
溶菌酶溶液:
-29μl1m磷酸氢二钾
-21μl1m磷酸二氢钾
-500μl40%蔗糖(通过0.45μm过滤器除菌过滤)
-450μl水
-60μl溶菌酶(50mg/ml)
在与溶菌酶温育后,将悬浮液如前所述离心,并丢弃上清液。然后使各沉淀接受通过经修改版本的qiaprepspinminiprep
1.缓冲液pi、p2和n3的体积均提高三倍,以使得增加的生物质可被完全裂解。
2.将2mg/ml的溶菌酶加入重悬的细胞中,然后添加p2。然后将裂解溶液在37℃下温育15分钟,再进行中和。
3.将质粒dna重悬在30μl而不是50μl中以提高浓度。
在其他实验中,将细胞在p1缓冲液+溶菌酶中温育15分钟,然后在室温下用p2(裂解缓冲液)和p3(中和缓冲液)温育。
将相等体积的得自各继代培养的分离的质粒dna在0.8%琼脂糖凝胶上运行,用溴化乙锭染色并观察结构或分离不稳定性的迹象。
结果表明从单核细胞增多性李斯特菌lm-llo-e7的质粒提取效率随着溶菌酶温育时间的延长而提高,在大约50分钟温育时达到最佳水平。
这些结果提供从李斯特菌疫苗菌株提取质粒的有效方法。
影印培养法
将原始培养物的稀释样在不存在或存在34μg/mlcap的情况下在含lb或tb琼脂的板上铺板。将选择性与非选择性琼脂之间的计数差异用于确定是否存在质粒任何明显的分离不稳定性。
结果
在不存在抗生素的情况下pgg55质粒在单核细胞增多性李斯特菌菌株xfl7中的遗传稳定性(即,在不存在选择压力例如抗生素选择压力的情况下质粒被细菌保持的程度或质粒保持与细菌稳定相关联的程度)通过在luria-bertani培养基(lb:5g/lnacl、10g/ml大豆蛋白胨、5g/l酵母提取物)和terrificbroth培养基(tb:10g/l葡萄糖、11.8g/l大豆蛋白胨、23.6g/l酵母提取物、2.2g/lkh2po4、9.4g/lk2hpo4)两者中以一式两份培养的方式通过系列传代培养而评估。将250ml带挡板的摇瓶中的50ml新鲜培养基用固定数量的细胞(1odml)接种,然后以24小时的间隔传代培养。将培养物在定轨摇床中在37℃和200rpm下温育。在每次传代培养时,测量od600,并将其用于计算已经过的细胞倍增时间(或代数),直到在lb和tb中分别达到30代和42代。通过离心法沉淀每个传代培养阶段(大约每4代)的已知数量的细胞(15odml),然后使用上述qiagenqiaprepspin
还通过在每个传代培养阶段在琼脂板上的影印培养而在稳定性研究期间监测质粒稳定性。与得自琼脂糖凝胶电泳的结果一致,在lb或tb液体培养(分别为图10和图11)中在整个研究期间含质粒的细胞的数量均无整体变化。
这些发现证明编码prfa的质粒在不存在抗生素的情况下在含prfa突变的李斯特菌菌株中表现出稳定性。
材料与方法(实例6-10)
pcr试剂:
用于扩增prfa基因及辨别d133v突变的引物在表1中示出。通过将引物在te缓冲液中稀释到400μm而制备了引物adv451、452和453的储备溶液。将储备溶液的等分试样在水(pcr级)中进一步稀释到20μm以制备工作溶液。将引物储存在-20℃。pcr中所用的试剂在表2中示出。
表1.引物adv451、452和453。
表2.pcr试剂。
质粒dna制备
使用purelinktmhipure质粒中量提取试剂盒(invitrogen,k2100-05)根据制造商的说明从大肠杆菌或单核细胞增多性李斯特菌通过中量制备而提取并纯化具有(pgg55d133v)和不具有(pgg55wt)prfa突变的pgg55质粒。对于从李斯特菌的质粒纯化,将携带pgg55d133v或wt质粒的细菌菌株从冷冻的储备品划线接种到补充了氯霉素(25μg/ml)的bhi琼脂板中。使来自每个菌株的单个集落在5ml选择性培养基(具有25μg/ml氯霉素的bhi肉汤)中在37℃和剧烈摇晃下生长6小时,然后以1:500传代接种在100ml选择性培养基中在相似的条件下生长过夜。将来自过夜培养的细菌通过在4,000xg下离心10分钟而收获,然后重悬在含有2mg/ml溶菌酶(sigma,l7001)的缓冲液r3(重悬缓冲液)中。将细菌悬液在37℃下温育至少1小时,然后转到常规方案。在分光光度计中在260nm和280nm下测量经稀释的质粒的浓度和纯度。为了制备模板dna,从中量制备储备溶液将pgg55d133v和wt质粒重悬在水中达到1ng/μl的最终浓度。对于pgg55wt质粒,从1ng/μl溶液制备了系列10倍稀释样,对应于从10-1至10-7的稀释。
测试临床级材料的prfa特异性pcr方案
反应混合物包含1xpcr缓冲液、1.5mmmgcl2、0.8mmdntp、0.4μm的各引物、0.05u/μltaqdna聚合酶和0.04ng/μlpgg55d133v模板质粒。对于各测试,需要10根管,在25μl反应中每根管的关键组分在表3中示出。对于pcr反应,用如表4所示的足够11次反应的试剂制备主混合物,然后将24μl该pcr混合物加入各管中。随后,将总共1μl系列稀释的pgg55wt质粒加入对应的管中:管3中1ng;管4中100pg;管5中10pg;管6中1pg;管7中100fg;管8中10fg;管9中1fg;管10中0.1fg。将该系列稀释样用于校准标准曲线以确定方法灵敏度。另外,将0.5μl水和0.5μl引物adv451(20μm储备液)加入管1中,并将1μl水加入管2中,达到25μl的最终体积。对于25μl反应,每根管的每种试剂的量在表5中示出。用在反应中的pcr循环条件在表6中示出。
在pcr反应结束后,将5μl凝胶上样缓冲液(6x,具有溴酚蓝)加到每个样品中,并通过在tbe缓冲液中的1.2%琼脂糖凝胶上电泳分析10μl。凝胶尺寸为7cm×7cm×1cm,具有15个样品孔(1mm×2mm)的梳。凝胶在100v下运行约30分钟,直到溴酚蓝染料到达凝胶的中间。将凝胶在溴化乙锭(0.5μg/ml)中染色20分钟,在水中脱色10分钟。通过用紫外光照射而观察凝胶,并拍照。使用条带密度测量软件(quantityone4.5.1版,biorad)分析图像。
表3.一套单独的pcr反应对方法进行验证以检测野生型prfa序列在lm-llo-e7样品中的存在性。
表4.主pcr混合物制备。
表5.使用引物adv451、452和453对方法进行验证以检测野生型prfa序列的存在性的pcr方案。
apgg55wt(管3中1ng;管4中100pg;管5中10pg;管6中1pg;管7中100fg;管8中10fg;管9中1fg;管10中0.1fg)。
b在管1中,添加0.5μl水和0.5μl引物adv451(20μm储备液);在管2中添加1μl水。
表6.使用引物adv451、452和453检测野生型prfa序列的存在性的pcr循环条件。
测序:
使用双脱氧测序法进行了质粒的测序。将质粒pgg55d133v和pgg55wt以不同的比率(1:1、1:10、1:100、1:1,000和1:10,000)混合。将混合物中质粒的总量保持恒定(500μg),并将包含野生型序列的质粒关于d133v质粒进行10倍系列稀释以确定方法的灵敏度。
结果
实例6:测序不是检测d133v突变的回复的灵敏方法。
为了估计测序在检测野生型prfa序列中的灵敏度,将pgg55d133v和wt质粒以不同的比率混合并测序。结果在图12中示出,并揭示测序在辨别prfad133v突变中具有高度特异性(图12)。在另一方面,灵敏度较低,并且在序列中具有可检测峰的野生型prfapgg55质粒的最大稀释度是1/10(图12)。总之,虽然测序具有非常高的特异性,但是该方法的灵敏度较低,且不适于筛选罕见事件(诸如prfad133v突变在lm-llo-e7样品中的回复体)的存在性。
实例7:开发高度特异性且灵敏的pcr方法来检测d133v突变的回复。
鉴于用测序来检测罕见事件的低灵敏度,有必要开发具有类似特异性的更灵敏的方法来检测野生型d133v突变的回复。为了实现该目标,我们设计了一种基于pcr的方法,该方法特异性扩增野生型序列并且足够灵敏检测10,000,000个d133v突变序列拷贝中的至少1个野生型prfa拷贝。我们设计了3个引物用于该方法:adv451、adv452和adv453(表1)。adv451和adv452均为正向引物,且差异在于3’位置的最后一个核苷酸以区分prfa基因的位置398处的a→t(d133v)突变。adv453引物是位于adv451和adv452引物的退火位点下游大约300bp处的反向引物(图13)。引物adv451或adv452和adv453所得的预期pcr条带为326bp。在严格条件下,adv451引物不应扩增pgg55d133v质粒,而adv452将为野生型prfa序列特异性的。
实例8:pcr方法的特异性。
使用引物adv451的反应具有非常高的特异性,并扩增了突变的d133vprfa序列(泳道1至3),但未扩增野生型序列(泳道4至6)。然而,当使用5ng的模板dna而不是使用1ng时,可在泳道4中检测到非常弱的条带(图14)。
如图15中所示,使用adv452引物的反应仅扩增了野生型prfa序列(泳道4、5和6),当将携带d133vprfa突变的pgg55用作模板时未检测到条带(泳道1、2和3),甚至当在反应中使用5ng质粒时也是如此(图16)。总之,使用引物adv451和adv452的pcr反应具有非常高的特异性,并且能够辨别在pgg55质粒中的prfa基因的位置398处的
实例9:pcr方法的灵敏度。
使用1ng的模板dna测试了反应的灵敏度。对于携带野生型prfa序列的质粒,还将降低的dna量(对应于从10-1至10-7的10倍稀释)包括在反应中以估计灵敏度。在这些反应中,仅使用了引物adv452和adv453。在具有30个循环(10个循环的退火温度为53℃,另20个循环的退火温度为50℃)的pcr反应中,方法的灵敏度为1/100,000(数据未示出)。如图5中所示,对于d133v回复体的检测,将pcr循环数增至37将方法的视觉灵敏度提高至10-6,而不明显损害特异性。当将1ng质粒用作初始dna量时,在10-6稀释下可见明显的条带,这对应于一百万个d133v突变体中1个野生型序列拷贝的检测水平。在较长的暴露后在泳道1和9中仅可观察到非常弱的条带,再次确保了方法可靠的特异性。在另一方面,当以5ngdna开始时,在10-7稀释下可容易地检测到条带,从而提高了pcr的灵敏度。然而,使用pgg55d133v质粒也可检测到类似强度的条带,从而表明方法的特异性限度(图17)。通过pgg55d133v质粒观察到的该条带可能是由于用引物adv452对d133v突变的非特异性扩增,通过增加的循环数,该突变可以明显蓄积。这些结果表明在不明显损害特异性的情况下该方法的灵敏度限度位于1:1,000,000与1:10,000,000之间。
实例10:表达llo与e7的融合蛋白(lm-llo-e7)的重组李斯特菌
该菌株的减毒程度比野生型亲本菌株10403s高大约4-5个数量级,并分泌融合蛋白tllo-e7。该免疫疗法基于骨架xfl7,其通过毒力基因转录激活因子prfa中的不可逆缺失而衍生自10403s。prfa调控若干毒力基因诸如李斯特菌溶血素o(llo)、acta、plca(磷脂酶a)、plcb(磷脂酶b)等的转录,这些基因是单核细胞增多性李斯特菌的体内胞内生长和存活所需的。质粒pgg55借助“氯霉素”的选择而由lm-llo-e7在体外保持。然而,对于lm-llo-e7对质粒的体内保持而言,其携带突变的prfa(d133v)的拷贝,已证实其在dna结合和激活毒力基因的转录方面的活性不如野生型prfa。我们已观察到,与突变的prfa的互补作用导致与野生型菌株10403s相比从lm-llo-e7分泌的llo的量降低大约40倍。这暗示:菌株lm-llo-e7可能表现出降低的受prfa调控的毒力基因诸如acta、inla、inlb、inlc、plcb等的表达。在lm-llo-e7中,与突变的prfa拷贝的互补作用可能导致受prfa调控的不同毒力基因的表达降低,从而导致总体减毒大约4-5个数量级。
实例11:抗e7肿瘤应答
招募
–6名具有ii–iv期hpvopc的患者入选
–5名患者用adxs-hpv治疗
–5名患者采集疫苗前和初始疫苗后肿瘤及血液样品。
测定
在疫苗施用前后执行组织测定
–h&e组织病理学
–多重免疫荧光
–基因表达签名的nanostring分析
血液
–通过流式细胞术进行的免疫表型分型
–hpv抗原特异性t细胞应答
–hpv血清学
–癌-睾丸抗原血清学
结果
在用lm-llo-e7(adxs-hpv)进行疫苗接种前后,从5名患者获取人肿瘤组织样品。结果显示出嗜碱性淋巴细胞样浸润巢(参见图24)和致密的瘤内cd8浸润和基质cd4浸润(图25)。
在用lm-llo-e7(adxs-hpv)进行疫苗接种前后,从5名患者获取人血液样品,并执行elispot-直接分析测定(无再刺激和培养)。在疫苗接种后观测到hpv-e7应答的平均增加>3倍,并在5名患者的3名中观测到应答增加(图26)。相比之下,相比非疫苗抗原(hpv-e6和e2),不存在疫苗治疗后的应答增加,证实了hpv-e7应答是抗原特异性的(图27)。
实例12:adxs-hpv肿瘤应答的ii期临床研究
招募和治疗
-11名具有ii–iv期hpvopc的患者入选
–8名患者在第1和15天用1×109个集落形成单位的lm-llo-e7(adxs-hpv)治疗
-9名患者采集疫苗前和初始疫苗后肿瘤及血液样品。
–之前未进行adxs-hpv治疗的接受了经口腔机器人手术(tors)的3名患者的观察组也入选。
关键纳入标准
–具有新诊断的、经活检证实的ii–iv期hpvopc的成年患者(-18岁)。
–能进行tors,接受或未接受颈清扫术。
–美国东部肿瘤协作组(easterncooperativeoncologygroup)体能状态≤2。
–能够理解并作出知情同意
关键排除标准
–另一部位有活动癌,或在过去3年内有癌症史。
–之前接受过全身化疗或放疗。
–处于免疫抑制状态或在使用免疫抑制药物。
–患有肝病或其他对研究药物的医疗禁忌症。
测定
在疫苗施用前后执行组织测定
–在研究开始时以及在手术时采集肿瘤活检样品
–h&e组织病理学
–多重免疫荧光
血液
–hpv抗原特异性t细胞应答
–hpv血清学
–癌-睾丸抗原血清学
分析
–将基于组织的变化与对外周血中免疫变化的综合分析相关联。
表7.实验室研究
elispot,酶联免疫斑点技术;hnscca,人头颈鳞状细胞癌抗原;hpv,人乳头瘤病毒;
if,免疫荧光;ihc,免疫组织化学;tcr,t细胞受体;tme,肿瘤微环境。
结果
研究的总体设计在图28a中示出。总共11名具有ii–iv期hpvopc的患者入选试验。在开始研究前从每名患者采集肿瘤活检样品。八名患者用1×109个集落形成单位的lm-llo-e7(adxs-hpv)进行疫苗接种,在第一次注射后14天接受加强注射。未接受疫苗接种的具有ii–iv期hpvopc的3名患者的观察组也入选。在输注adxs-hpv前给予布洛芬、苯海拉明和止吐剂,在输注后还施用布洛芬;在每次adxs-hpv给药后72小时施用一个疗程的阿莫西林(或替代性抗生素)。所有患者均在加强注射后10-14天接受医护标准的经口腔机器人手术(tors)以取出肿瘤样品。根据医护标准向所有患者施用辅助放疗/放化疗(tors后4–6周)。在疫苗接种前1-10天,从除了接受疫苗的两名患者外的所有患者采集外周血单核细胞(pbmc)样品。在每次疫苗注射前以及在手术前,从所有患者采集另外的pbmc样品。在不同的时间点直至初次疫苗接种后12个月,抽取另外的pbmc样品(图28b)。样品的elispot测定显示在一些患者中在疫苗接种后e6和e7应答均增加(图29),以及e6和e7特异性ifn-γ分泌性cd4+和cd8+t细胞水平增加(图30)。此外,一些患者在手术时表现出甚至更稳健的e6和e7特异性ifn-γ分泌性cd4+和cd8+t细胞诱导(图31)。总体而言,8名患者中的5名对e7或e6具有ifn-γ应答,并且8名患者中的7名对e7或e6具有tnf-α应答(图32)。多重免疫荧光测定显示出在adxs-hpv疫苗接种后肿瘤环境的致密的瘤内cd8浸润(对比图33a和33c)和减小的肿瘤大小(对比图33b和33d),以及疫苗接种后肿瘤中cd8和pd-1的增加(对比图34a-c和34d-f)。这些观测结果在定量分析中得以证实(图35),定量分析另外显示出相对于pd-l1而言pd1表达水平升高(图36),以及在adxs-hpv疫苗接种后cd8表达水平升高(图37),从而证明了在癌症患者中adxs-hpv成功激活了抗e6和e7应答。
结论:
在研究结束时,外周血中的hpv特异性cd8+ctl应答从手术时的基线发生变化。这种变化可在手术后的各个时间点观测到。此外,肿瘤浸润性效应(自然杀伤[nk]细胞、cd4+和cd8+t细胞)和抑制性(treg和mdsc)免疫细胞的概况在研究过程中变化。通过美国国立癌症研究院常见不良事件评价标准4.0版(nationalcancerinstitutecommonterminologycriteriaforadverseeventsversion4.0)评估了不良事件。
已参考附图对本发明的优选实施例进行了描述,应当理解,本发明不限于精确的实施例,本领域技术人员可在不脱离所附权利要求书中定义的本发明范围和精神的情况下对其进行各种改动和修饰。
序列表
<110>宾夕法尼亚大学理事会
y·佩特森
<120>重组李斯特菌疫苗菌株及其用于癌症免疫疗法的方法
<130>p-78581-pc
<140>pct/us15/999999
<141>2015-10-14
<150>62/065,973
<151>2014-10-20
<150>62/063,828
<151>2014-10-14
<160>30
<170>patentin3.5版
<210>1
<211>32
<212>prt
<213>单核细胞增多性李斯特菌
<400>1
lysgluasnserilesersermetalaproproalaserproproala
151015
serprolysthrproileglulyslyshisalaaspgluileasplys
202530
<210>2
<211>441
<212>prt
<213>单核细胞增多性李斯特菌
<400>2
metlyslysilemetleuvalpheilethrleuileleuvalserleu
151015
proilealaglnglnthrglualalysaspalaseralapheasnlys
202530
gluasnserileserservalalaproproalaserproproalaser
354045
prolysthrproileglulyslyshisalaaspgluileasplystyr
505560
ileglnglyleuasptyrasnlysasnasnvalleuvaltyrhisgly
65707580
aspalavalthrasnvalproproarglysglytyrlysaspglyasn
859095
glutyrilevalvalglulyslyslyslysserileasnglnasnasn
100105110
alaaspileglnvalvalasnalaileserserleuthrtyrprogly
115120125
alaleuvallysalaasnsergluleuvalgluasnglnproaspval
130135140
leuprovallysargaspserleuthrleuserileaspleuprogly
145150155160
metthrasnglnaspasnlysilevalvallysasnalathrlysser
165170175
asnvalasnasnalavalasnthrleuvalgluargtrpasnglulys
180185190
tyralaglnalatyrserasnvalseralalysileasptyraspasp
195200205
glumetalatyrsergluserglnleuilealalyspheglythrala
210215220
phelysalavalasnasnserleuasnvalasnpheglyalaileser
225230235240
gluglylysmetglnglugluvalileserphelysglniletyrtyr
245250255
asnvalasnvalasngluprothrargproserargphepheglylys
260265270
alavalthrlysgluglnleuglnalaleuglyvalasnalagluasn
275280285
proproalatyrileserservalalatyrglyargglnvaltyrleu
290295300
lysleuserthrasnserhisserthrlysvallysalaalapheasp
305310315320
alaalavalserglylysservalserglyaspvalgluleuthrasn
325330335
ileilelysasnserserphelysalavaliletyrglyglyserala
340345350
lysaspgluvalglnileileaspglyasnleuglyaspleuargasp
355360365
ileleulyslysglyalathrpheasnarggluthrproglyvalpro
370375380
ilealatyrthrthrasnpheleulysaspasngluleualavalile
385390395400
lysasnasnserglutyrilegluthrthrserlysalatyrthrasp
405410415
glylysileasnileasphisserglyglytyrvalalaglnpheasn
420425430
ilesertrpaspgluvalasntyrasp
435440
<210>3
<211>529
<212>prt
<213>单核细胞增多性李斯特菌
<400>3
metlyslysilemetleuvalpheilethrleuileleuvalserleu
151015
proilealaglnglnthrglualalysaspalaseralapheasnlys
202530
gluasnserilesersermetalaproproalaserproproalaser
354045
prolysthrproileglulyslyshisalaaspgluileasplystyr
505560
ileglnglyleuasptyrasnlysasnasnvalleuvaltyrhisgly
65707580
aspalavalthrasnvalproproarglysglytyrlysaspglyasn
859095
glutyrilevalvalglulyslyslyslysserileasnglnasnasn
100105110
alaaspileglnvalvalasnalaileserserleuthrtyrprogly
115120125
alaleuvallysalaasnsergluleuvalgluasnglnproaspval
130135140
leuprovallysargaspserleuthrleuserileaspleuprogly
145150155160
metthrasnglnaspasnlysilevalvallysasnalathrlysser
165170175
asnvalasnasnalavalasnthrleuvalgluargtrpasnglulys
180185190
tyralaglnalatyrproasnvalseralalysileasptyraspasp
195200205
glumetalatyrsergluserglnleuilealalyspheglythrala
210215220
phelysalavalasnasnserleuasnvalasnpheglyalaileser
225230235240
gluglylysmetglnglugluvalileserphelysglniletyrtyr
245250255
asnvalasnvalasngluprothrargproserargphepheglylys
260265270
alavalthrlysgluglnleuglnalaleuglyvalasnalagluasn
275280285
proproalatyrileserservalalatyrglyargglnvaltyrleu
290295300
lysleuserthrasnserhisserthrlysvallysalaalapheasp
305310315320
alaalavalserglylysservalserglyaspvalgluleuthrasn
325330335
ileilelysasnserserphelysalavaliletyrglyglyserala
340345350
lysaspgluvalglnileileaspglyasnleuglyaspleuargasp
355360365
ileleulyslysglyalathrpheasnarggluthrproglyvalpro
370375380
ilealatyrthrthrasnpheleulysaspasngluleualavalile
385390395400
lysasnasnserglutyrilegluthrthrserlysalatyrthrasp
405410415
glylysileasnileasphisserglyglytyrvalalaglnpheasn
420425430
ilesertrpaspgluvalasntyraspprogluglyasngluileval
435440445
glnhislysasntrpsergluasnasnlysserlysleualahisphe
450455460
thrserseriletyrleuproglyasnalaargasnileasnvaltyr
465470475480
alalysglucysthrglyleualatrpglutrptrpargthrvalile
485490495
aspaspargasnleuproleuvallysasnargasnileseriletrp
500505510
glythrthrleutyrprolystyrserasnlysvalaspasnproile
515520525
glu
<210>4
<211>416
<212>prt
<213>单核细胞增多性李斯特菌
<400>4
metlyslysilemetleuvalpheilethrleuileleuvalserleu
151015
proilealaglnglnthrglualalysaspalaseralapheasnlys
202530
gluasnserileserservalalaproproalaserproproalaser
354045
prolysthrproileglulyslyshisalaaspgluileasplystyr
505560
ileglnglyleuasptyrasnlysasnasnvalleuvaltyrhisgly
65707580
aspalavalthrasnvalproproarglysglytyrlysaspglyasn
859095
glutyrilevalvalglulyslyslyslysserileasnglnasnasn
100105110
alaaspileglnvalvalasnalaileserserleuthrtyrprogly
115120125
alaleuvallysalaasnsergluleuvalgluasnglnproaspval
130135140
leuprovallysargaspserleuthrleuserileaspleuprogly
145150155160
metthrasnglnaspasnlysilevalvallysasnalathrlysser
165170175
asnvalasnasnalavalasnthrleuvalgluargtrpasnglulys
180185190
tyralaglnalatyrserasnvalseralalysileasptyraspasp
195200205
glumetalatyrsergluserglnleuilealalyspheglythrala
210215220
phelysalavalasnasnserleuasnvalasnpheglyalaileser
225230235240
gluglylysmetglnglugluvalileserphelysglniletyrtyr
245250255
asnvalasnvalasngluprothrargproserargphepheglylys
260265270
alavalthrlysgluglnleuglnalaleuglyvalasnalagluasn
275280285
proproalatyrileserservalalatyrglyargglnvaltyrleu
290295300
lysleuserthrasnserhisserthrlysvallysalaalapheasp
305310315320
alaalavalserglylysservalserglyaspvalgluleuthrasn
325330335
ileilelysasnserserphelysalavaliletyrglyglyserala
340345350
lysaspgluvalglnileileaspglyasnleuglyaspleuargasp
355360365
ileleulyslysglyalathrpheasnarggluthrproglyvalpro
370375380
ilealatyrthrthrasnpheleulysaspasngluleualavalile
385390395400
lysasnasnserglutyrilegluthrthrserlysalatyrthrasp
405410415
<210>5
<211>390
<212>prt
<213>单核细胞增多性李斯特菌
<400>5
metargalametmetvalvalpheilethralaasncysilethrile
151015
asnproaspileilephealaalathraspsergluaspserserleu
202530
asnthraspglutrpglugluglulysthrglugluglnproserglu
354045
valasnthrglyproargtyrgluthralaarggluvalserserarg
505560
aspilelysgluleuglulysserasnlysvalargasnthrasnlys
65707580
alaaspleuilealametleulysglulysalaglulysglyproasn
859095
ileasnasnasnasnsergluglnthrgluasnalaalaileasnglu
100105110
glualaserglyalaaspargproalaileglnvalgluargarghis
115120125
proglyleuproseraspseralaalagluilelyslysargarglys
130135140
alailealaserseraspsergluleugluserleuthrtyrproasp
145150155160
lysprothrlysvalasnlyslyslysvalalalysgluservalala
165170175
aspalasergluseraspleuaspsersermetglnseralaaspglu
180185190
serserproglnproleulysalaasnglnglnprophepheprolys
195200205
valphelyslysilelysaspalaglylystrpvalargasplysile
210215220
aspgluasnprogluvallyslysalailevalasplysseralagly
225230235240
leuileaspglnleuleuthrlyslyslysserglugluvalasnala
245250255
serasppheproproproprothraspglugluleuargleualaleu
260265270
progluthrprometleuleuglypheasnalaproalathrserglu
275280285
proserserpheglupheproproproprothraspglugluleuarg
290295300
leualaleuprogluthrprometleuleuglypheasnalaproala
305310315320
thrsergluproserserpheglupheproproproprothrgluasp
325330335
gluleugluileilearggluthralaserserleuaspserserphe
340345350
thrargglyaspleualaserleuargasnalaileasnarghisser
355360365
glnasnpheserasppheproproileprothrgluglugluleuasn
370375380
glyargglyglyargpro
385390
<210>6
<211>1170
<212>dna
<213>单核细胞增多性李斯特菌
<400>6
atgcgtgcgatgatggtggttttcattactgccaattgcattacgattaaccccgacata60
atatttgcagcgacagatagcgaagattctagtctaaacacagatgaatgggaagaagaa120
aaaacagaagagcaaccaagcgaggtaaatacgggaccaagatacgaaactgcacgtgaa180
gtaagttcacgtgatattaaagaactagaaaaatcgaataaagtgagaaatacgaacaaa240
gcagacctaatagcaatgttgaaagaaaaagcagaaaaaggtccaaatatcaataataac300
aacagtgaacaaactgagaatgcggctataaatgaagaggcttcaggagccgaccgacca360
gctatacaagtggagcgtcgtcatccaggattgccatcggatagcgcagcggaaattaaa420
aaaagaaggaaagccatagcatcatcggatagtgagcttgaaagccttacttatccggat480
aaaccaacaaaagtaaataagaaaaaagtggcgaaagagtcagttgcggatgcttctgaa540
agtgacttagattctagcatgcagtcagcagatgagtcttcaccacaacctttaaaagca600
aaccaacaaccatttttccctaaagtatttaaaaaaataaaagatgcggggaaatgggta660
cgtgataaaatcgacgaaaatcctgaagtaaagaaagcgattgttgataaaagtgcaggg720
ttaattgaccaattattaaccaaaaagaaaagtgaagaggtaaatgcttcggacttcccg780
ccaccacctacggatgaagagttaagacttgctttgccagagacaccaatgcttcttggt840
tttaatgctcctgctacatcagaaccgagctcattcgaatttccaccaccacctacggat900
gaagagttaagacttgctttgccagagacgccaatgcttcttggttttaatgctcctgct960
acatcggaaccgagctcgttcgaatttccaccgcctccaacagaagatgaactagaaatc1020
atccgggaaacagcatcctcgctagattctagttttacaagaggggatttagctagtttg1080
agaaatgctattaatcgccatagtcaaaatttctctgatttcccaccaatcccaacagaa1140
gaagagttgaacgggagaggcggtagacca1170
<210>7
<211>14
<212>prt
<213>单核细胞增多性李斯特菌
<400>7
lysthrglugluglnprosergluvalasnthrglyproarg
1510
<210>8
<211>28
<212>prt
<213>单核细胞增多性李斯特菌
<400>8
lysalaservalthraspthrsergluglyaspleuaspsersermet
151015
glnseralaaspgluserthrproglnproleulys
2025
<210>9
<211>20
<212>prt
<213>单核细胞增多性李斯特菌
<400>9
lysasnglugluvalasnalaserasppheproproproprothrasp
151015
glugluleuarg
20
<210>10
<211>33
<212>prt
<213>单核细胞增多性李斯特菌
<400>10
argglyglyileprothrsergluglupheserserleuasnsergly
151015
aspphethraspaspgluasnsergluthrthrgluglugluileasp
202530
arg
<210>11
<211>17
<212>prt
<213>链球菌属菌种g148
<400>11
lysglnasnthralaserthrgluthrthrthrthrasngluglnpro
151015
lys
<210>12
<211>17
<212>prt
<213>类马链球菌
<400>12
lysglnasnthralaasnthrgluthrthrthrthrasngluglnpro
151015
lys
<210>13
<211>98
<212>prt
<213>人乳头瘤病毒16型
<400>13
methisglyaspthrprothrleuhisglutyrmetleuaspleugln
151015
progluthrthraspleutyrcystyrgluglnleuasnaspserser
202530
gluglugluaspgluileaspglyproalaglyglnalagluproasp
354045
argalahistyrasnilevalthrphecyscyslyscysaspserthr
505560
leuargleucysvalglnserthrhisvalaspileargthrleuglu
65707580
aspleuleumetglythrleuglyilevalcysproilecyssergln
859095
lyspro
<210>14
<211>105
<212>prt
<213>人乳头瘤病毒16型
<400>14
methisglyprolysalathrleuglnaspilevalleuhisleuglu
151015
proglnasngluileprovalaspleuleucyshisgluglnleuser
202530
aspsergluglugluasnaspgluileaspglyvalasnhisglnhis
354045
leuproalaargargalagluproglnarghisthrmetleucysmet
505560
cyscyslyscysglualaargilegluleuvalvalgluserserala
65707580
aspaspleuargalapheglnglnleupheleuasnthrleuserphe
859095
valcysprotrpcysalaserglngln
100105
<210>15
<211>158
<212>prt
<213>人乳头瘤病毒16型
<400>15
methisglnlysargthralametpheglnaspproglngluargpro
151015
arglysleuproglnleucysthrgluleuglnthrthrilehisasp
202530
ileileleuglucysvaltyrcyslysglnglnleuleuargargglu
354045
valtyraspphealapheargaspleucysilevaltyrargaspgly
505560
asnprotyralavalcysasplyscysleulysphetyrserlysile
65707580
serglutyrarghistyrcystyrserleutyrglythrthrleuglu
859095
glnglntyrasnlysproleucysaspleuleuileargcysileasn
100105110
cysglnlysproleucysprogluglulysglnarghisleuasplys
115120125
lysglnargphehisasnileargglyargtrpthrglyargcysmet
130135140
sercyscysargserserargthrargarggluthrglnleu
145150155
<210>16
<211>158
<212>prt
<213>人乳头瘤病毒16型
<400>16
metalaargphegluaspprothrargargprotyrlysleuproasp
151015
leucysthrgluleuasnthrserleuglnaspilegluilethrcys
202530
valtyrcyslysthrvalleugluleuthrgluvalpheglupheala
354045
phelysaspleuphevalvaltyrargaspserileprohisalaala
505560
cyshislyscysileaspphetyrserargilearggluleuarghis
65707580
tyrseraspservaltyrglyaspthrleuglulysleuthrasnthr
859095
glyleutyrasnleuleuileargcysleuargcysglnlysproleu
100105110
asnproalaglulysleuarghisleuasnglulysargargphehis
115120125
asnilealaglyhistyrargglyglncyshissercyscysasnarg
130135140
alaargglngluargleuglnargargarggluthrglnval
145150155
<210>17
<211>22
<212>dna
<213>人工序列
<220>
<223>引物
<400>17
ggctcgagcatggagatacacc22
<210>18
<211>28
<212>dna
<213>人工序列
<220>
<223>引物
<400>18
ggggactagtttatggtttctgagaaca28
<210>19
<211>31
<212>dna
<213>人工序列
<220>
<223>引物
<400>19
gggggctagccctcctttgattagtatattc31
<210>20
<211>28
<212>dna
<213>人工序列
<220>
<223>引物
<400>20
ctccctcgagatcataatttacttcatc28
<210>21
<211>27
<212>dna
<213>人工序列
<220>
<223>引物
<400>21
cccgtcgaccagctcttcttggtgaag27
<210>22
<211>25
<212>dna
<213>人工序列
<220>
<223>引物
<400>22
gcggatcccatggagatacacctac25
<210>23
<211>22
<212>dna
<213>人工序列
<220>
<223>引物
<400>23
gctctagattatggtttctgag22
<210>24
<211>9
<212>prt
<213>人乳头瘤病毒16型
<400>24
argalahistyrasnilevalthrphe
15
<210>25
<211>9
<212>prt
<213>人免疫缺陷病毒
<400>25
alametglnmetleulysgluthrile
15
<210>26
<211>9
<212>prt
<213>单核细胞增多性李斯特菌
<400>26
glytyrlysaspglyasnglutyrile
15
<210>27
<211>55
<212>prt
<213>人工序列
<220>
<223>引物
<400>27
glyalacysthralacysalaalaglyglyalacysglyalathrgly
151015
alacyscysglyalacysalaalaglythrglyalathralaalacys
202530
cyscysglyglyglyalathrcysthralaalaalathralaalaala
354045
thrcyscysglythrthrthr
5055
<210>28
<211>18
<212>dna
<213>人工序列
<220>
<223>adv451正向引物
<400>28
cctagctaaatttaatgt18
<210>29
<211>18
<212>dna
<213>人工序列
<220>
<223>adv452正向引物
<400>29
cctagctaaatttaatga18
<210>30
<211>21
<212>dna
<213>人工序列
<220>
<223>adv453反向引物
<400>30
taattttccccaagtagcagg21
- 还没有人留言评论。精彩留言会获得点赞!