新型昆虫抑制性蛋白的制作方法
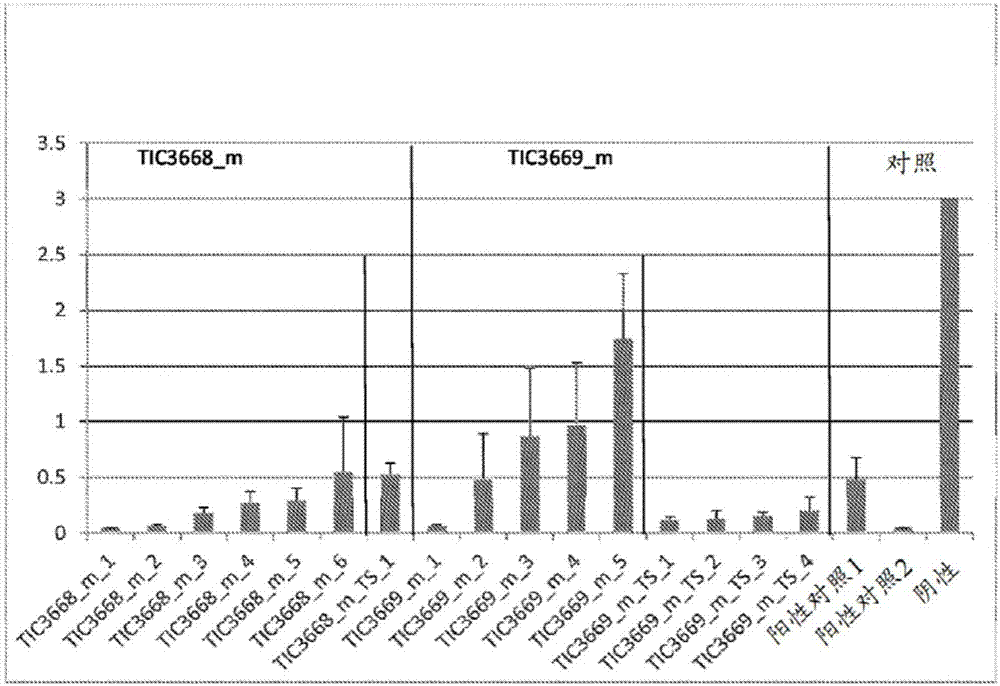
相关申请的引用本申请要求2014年11月20日提交的美国临时申请62/082,504的权益,所述美国临时申请以引用的方式整体并入本文。序列表的并入计算机可读形式的序列表通过电子提交稿随本文一起提交。序列表以引用的方式整体并入本文,包含在2015年11月13日创建的文件中,所述文件具有文件名称“mons387wo_st25.txt”并且大小是114kb(如ms-windows操作系统中所测量)。本发明一般涉及昆虫抑制性蛋白领域。公开一类展现针对作物植物和种子的农业相关害虫的昆虫抑制活性的新型蛋白质。具体来说,所公开类别的蛋白质针对作物植物和种子的农业相关害虫特别是鳞翅目(lepidopteran)和鞘翅目(coleopteran)物种的昆虫害虫具有杀虫活性。提供含有编码一种或多种所公开毒素蛋白质的重组多核苷酸构建体的植物、植物部分和种子。
背景技术:
:改进尤其包括玉米、大豆、甘蔗、稻、小麦、蔬菜和棉花的农业重要植物的作物产量已变得日益重要。除对用以为日益增长的人口提供食物、提供衣服和提供能量的农业产品的需要日益增长之外,预测气候相关影响和由日益增长的人口使用除用于农业实践的土地以外的土地所致的压力也会降低可用于农耕的可耕土地的数量。这些因素已导致对粮食保障的严峻预测,特别是在植物生物技术和农艺学实践方面不存在重大改进下。鉴于这些压力,在技术、农业技术和害虫治理方面的环境可持续改进是用以在可用于农耕的可耕土地的数量有限的情况下扩大作物产量的重要工具。昆虫,特别是属于鳞翅目和鞘翅目的昆虫,被视为是对田地作物的损害的主要原因,由此在受侵扰区域内降低作物产量。负面影响农业的鳞翅目害虫物种包括但不限于美洲棉铃虫(helicoverpazea)、欧洲玉米螟(ostrinianubilalis)、小蔗杆草螟(diatraeasaccharalis)、西南玉米杆草螟(diatraeagrandiosella)、黎豆夜蛾(anticarsiagemmatalis)、草地夜蛾(spodopterafrugiperda)、甜菜夜蛾(spodopteraexigua)、小地老虎(agrotisipsilon)、粉纹夜蛾(trichoplusiani)、大豆尺蠖(chrysodeixisincludens)、烟芽夜蛾(heliothisvirescens)、小菜蛾(plutellaxylostella)、棉红铃虫(pectinophoragossypiella)、棉铃虫(helicoverpaarmigera)、南美玉米苗斑螟(elasmopalpuslignosellus)、豆白缘切根虫(striacostaalbicosta)和柑桔潜叶蛾(phyllocnistiscitrella)。负面影响农业的鞘翅目害虫物种包括但不限于叩甲属某些种(agriotesspp.)、花象属某些种(anthonomusspp.)、甜菜隐食甲(atomarialinearis)、蚤凹胫跳甲(chaetocnematibialis)、根颈象属某些种(cosmopolitesspp.)、象虫属某些种(curculiospp.)、皮蠹属某些种(dermestesspp.)、根叶甲属某些种(diabroticaspp.)、食植瓢虫属某些种(epilachnaspp.)、eremnus属某些种、科罗拉多金花虫(leptinotarsadecemlineata)、稻水象属某些种(lissorhoptrusspp.)、鳃金龟属某些种(melolonthaspp.)、锯谷盗属某些种(orycaephilusspp.)、耳喙象属某些种(otiorhynchusspp.)、斑象属某些种(phlyctinusspp.)、弧丽金龟属某些种(popilliaspp.)、蚤跳甲属某些种(psylliodesspp.)、谷蠹属某些种(rhizoperthaspp.)、金龟子科(scarabeidae)、象虫属某些种(sitophilusspp.)、麦蛾属某些种(sitotrogaspp.)、拟步行虫属某些种(tenebriospp.)、拟谷盗属某些种(triboliumspp.)和斑皮蠹属某些种(trogodermaspp.),特别是当害虫是玉米根叶甲(diabroticavirgiferavirgifera)(西方玉米根虫,wcr)、巴氏根叶甲(diabroticabarberi)(北方玉米根虫,ncr)、墨西哥玉米根叶甲(diabroticavirgiferazeae)(墨西哥玉米根虫,mcr)、带斑黄瓜根叶甲(diabroticabalteata)(巴西玉米根虫(bzr))、霍华德十一星根叶甲(diabroticaundecimpunctatahowardii)(南方玉米根虫,scr)和由绿根叶甲(diabroticaviridula)和美丽根叶甲(diabroticaspeciosa)组成的巴西玉米根虫复合体(bcr)时。在历史上,作为农业中的害虫控制剂,合成化学杀虫剂的广泛应用得以信赖。除出现的抗性问题之外,对于环境和人类健康的担忧也刺激对生物杀虫剂的研究和开发。这个研究努力导致逐步发现和使用各种昆虫病原性微生物物种,包括细菌。当发现昆虫病原性细菌(尤其是属于芽孢杆菌属(bacillus)的细菌)的潜力并将其开发为生物害虫控制剂时,生物控制范例发生转变。细菌苏云金芽孢杆菌(bacillusthuringiensis(bt))的菌株已用作展现杀虫活性的蛋白质的来源,因为发现bt菌株显示针对特定昆虫的高毒性。bt的主要特征是产生含有一个或多个晶体的类芽孢体,所述晶体含有特定杀虫内毒素(cry蛋白),其通过对昆虫消化道上皮有害的成孔作用机理来作用于由易感昆虫进行的摄取。除bt之外,其它芽孢杆菌属物种,诸如球形芽孢杆菌(bacillussphaericus)和含有促进昆虫病原性表型的基因的其它细菌物种诸如侧孢短芽孢杆菌(brevibacilluslaterosporus),已显示害虫治理潜力。杀虫毒素蛋白质已在各种农业应用中用于保护农业重要植物和增加产量。杀虫毒素蛋白质用于通过机械方法,诸如喷洒以将含有各种细菌菌株的微生物制剂分散于植物表面上,和通过使用遗传转化技术以产生表达杀虫毒素蛋白质的转基因植物和种子,来控制作物植物的农业相关害虫。使用表达杀虫毒素蛋白质的转基因植物已在全球被采纳。举例来说,在2012年,种植了2610万公顷的表达bt毒素的转基因作物(james,c.,globalstatusofcommercializedbiotech/gmcrops:2012.第44期isaaa简报)。对转基因昆虫防护作物的扩大使用以及可商购获得的杀虫毒素蛋白质的数目有限正在产生对赋予对当前利用的杀虫蛋白质的抗性的等位基因的选择压力。在靶标害虫中产生对杀虫毒素蛋白质的抗性会损害这个技术的有效性和优势。此类优势包括作物产量增加、化学杀虫剂使用降低以及与化学杀虫剂使用相关的成本和劳动力降低。发现和开发新型杀虫毒素蛋白质对于治理昆虫对表达杀虫毒素蛋白质的转基因作物的抗性增加是重要的。具有改进的功效并且展现对更广泛谱系的易感昆虫物种的控制的新型蛋白质毒素将降低可产生抗性等位基因的存活昆虫的数目。此外,一种植物中的两种或更多种对相同昆虫害虫有毒并且显示不同作用模式的转基因毒素进一步降低在靶标昆虫物种中产生抗性的可能性。因此,关键是需要发现和开发相较于本领域中已知的蛋白质具有改进的杀虫性质的有效杀虫蛋白质,所述性质诸如针对更广泛谱系的靶标昆虫害虫物种的增加的功效和不同的作用模式。本申请中公开了来自侧孢短芽孢杆菌(b.laterosporus)的新型蛋白质毒素家族以及类似毒素蛋白质、变体蛋白和示例性重组蛋白质,这些蛋白质展现针对重要靶标鳞翅目和鞘翅目害虫物种,特别是针对西方玉米根虫的杀虫活性。技术实现要素:本文公开一组新型昆虫抑制性重组多核苷酸分子和由此编码的在本文中被称为tic3668型蛋白的多肽(毒素蛋白质),显示其展现针对作物植物的一种或多种害虫的抑制活性。各蛋白质可单独或彼此组合以及与其它杀虫蛋白质和毒性剂组合用于制剂中以及植物中(inplanta),由此提供当前用于农业系统中的杀虫蛋白质和杀虫剂化学物质的替代物。在一个方面,本发明提供一种编码昆虫抑制性多肽的重组多核苷酸分子,所述昆虫抑制性多肽包含选自由以下组成的组的氨基酸序列:seqidno:25、seqidno:23、seqidno:24、seqidno:26、seqidno:27、seqidno:28、seqidno:29、seqidno:30和seqidno:31。在一个实施方案中,重组多核苷酸分子编码包含与选自由以下组成的组的氨基酸序列具有至少35%同一性,例如至少40%、50%、60%、70%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一性的昆虫抑制性多肽:seqidno:25、seqidno:23、seqidno:24、seqidno:26、seqidno:27、seqidno:28、seqidno:29、seqidno:30和seqidno:31。在另一实施方案中,重组多核苷酸分子包含选自由以下组成的组的核苷酸序列:seqidno:37、seqidno:33、seqidno:35、seqidno:39、seqidno:41、seqidno:43、seqidno:45、seqidno:47、seqidno:49、seqidno:64、seqidno:65、seqidno:66、seqidno:67、seqidno:68、seqidno:69、seqidno:70、seqidno:71和seqidno:72。在另一实施方案中,重组多核苷酸分子包含与选自由以下组成的组的核苷酸序列具有至少35%同一性,例如至少40%、50%、60%、70%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一性:seqidno:37、seqidno:33、seqidno:35、seqidno:39、seqidno:41、seqidno:43、seqidno:45、seqidno:47、seqidno:49、seqidno:64、seqidno:65、seqidno:66、seqidno:67、seqidno:68、seqidno:69、seqidno:70、seqidno:71和seqidno:72。在另一实施方案中,重组多核苷酸分子包含与以下杂交的序列:(i)选自由以下组成的组的序列的位置4-885的核苷酸序列的反向互补序列:seqidno:37、seqidno:33、seqidno:35、seqidno:39、seqidno:41、seqidno:43、seqidno:45、seqidno:47、seqidno:49、seqidno:64、seqidno:65、seqidno:66、seqidno:67、seqidno:68、seqidno:69、seqidno:70、seqidno:71和seqidno:72;或(ii)选自由以下组成的组的序列的反向互补序列:seqidno:52、seqidno:53、seqidno:54、seqidno:55、seqidno:56、seqidno:57、seqidno:58、seqidno:59、seqidno:60和seqidno:61。在另一实施方案中,杂交条件是严格条件,举例来说,此类严格条件可包括在37c下在50%甲酰胺、1mnacl和1%sds中杂交4至12小时,以及在60c-65c下在0.1xssc中洗涤。在其它实施方案中,使重组多核苷酸分子可操作地连接至异源启动子。在另一方面,本发明提供一种由本文提供的重组多核苷酸分子编码的昆虫抑制性重组多肽。在一个实施方案中,昆虫抑制性重组多肽包含选自由以下组成的组的氨基酸序列:seqidno:25、seqidno:23、seqidno:24、seqidno:26、seqidno:27、seqidno:28、seqidno:29、seqidno:30和seqidno:31。在另一实施方案中,昆虫抑制性重组多肽包含与选自由以下组成的组的氨基酸序列具有至少35%同一性,例如至少40%、50%、60%、70%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一性:seqidno:25、seqidno:23、seqidno:24、seqidno:26、seqidno:27、seqidno:28、seqidno:29、seqidno:30和seqidno:31。在另一实施方案中,昆虫抑制性重组多肽展现针对鞘翅目的昆虫物种的抑制活性,所述物种例如包括西方玉米根虫、南方玉米根虫、北方玉米根虫、墨西哥玉米根虫、巴西玉米根虫或由绿根叶甲和美丽根叶甲组成的巴西玉米根虫复合体。在另一实施方案中,昆虫抑制性重组多肽展现针对鳞翅目的昆虫物种的抑制活性,所述物种例如包括欧洲玉米螟虫、西南玉米螟虫、黑切根虫、秋行军虫(fallarmyworm)、玉米耳虫(cornearworm)和大豆尺蠖。在另一方面,本发明提供一种包含本发明的重组多核苷酸分子的宿主细胞,其中所述宿主细胞选自由细菌宿主细胞和植物宿主细胞组成的组。在某些实施方案中,细菌宿主细胞包括土壤杆菌属(agrobacterium)、根瘤菌属(rhizobium)、苏云金芽孢杆菌、侧孢短芽孢杆菌、蜡样芽孢杆菌(bacilluscereus)、大肠杆菌(e.coli)、假单胞菌属(pseudomonas)、克雷伯氏菌属(klebsiella)和欧文氏菌属(erwinia)。在其它实施方案中,植物细胞包括苜蓿、香蕉、大麦、菜豆、西兰花、卷心菜、芸苔、胡萝卜、木薯、蓖麻、花椰菜、芹菜、鹰嘴豆、大白菜、柑桔、椰子、咖啡、玉米、三叶草、棉花、葫芦、黄瓜、花旗松、茄子、桉树、亚麻、大蒜、葡萄、蛇麻子、韭菜、莴苣、火炬松、粟、甜瓜、坚果、燕麦、橄榄、洋葱、观赏植物、棕榈、牧场草、豌豆、花生、胡椒、柿子、木豆、松树、石榴、白杨树、马铃薯、南瓜、辐射松、萝卜、油菜籽、稻、根茎、黑麦、红花、灌木、高粱、南方松、大豆、菠菜、南瓜小果、草莓、糖用甜菜、甘蔗、向日葵、甜玉米、枫香、甜薯、柳枝稷、茶树、烟草、番茄、黑小麦、草皮草、西瓜和小麦植物细胞。在另一方面,本发明提供一种可包含本发明的重组多核苷酸分子的昆虫抑制性组合物。在一个实施方案中,昆虫抑制性组合物可还包含编码至少一种其它杀虫剂的核苷酸序列。在某些实施方案中,至少一种其它杀虫剂不同于本发明的tic3668型昆虫抑制性多肽,并且可选自由昆虫抑制性蛋白、昆虫抑制性dsrna分子和辅助蛋白质组成的组。在其它实施方案中,其它杀虫剂展现针对鳞翅目、鞘翅目或半翅目(hemiptera)的一个或多个害虫物种的活性。在某些实施方案中,其它杀虫剂选自由以下组成的组:cry1a、cry1ab、cry1ac、cry1a.105、cry1b、cry1c、cry1d、cry1e、cry1f、cry1g、cry1h、cry1i、cry1j、cry1k、cry1l、cry2a、cry2ab、cry3a、cry3b、cry4b、cry6、cry7、cry8、cry9、cry15、cry34、cry35、cry43a、cry43b、cry51aa1、et29、et33、et34、et35、et66、et70、tic400、tic407、tic417、tic431、tic800、tic807、tic834、tic853、tic900、tic901、tic1201、tic1415、vip3a和vip3b蛋白。在另一方面,本发明提供一种包含昆虫抑制有效量的本发明的昆虫抑制性重组多肽诸如tic3668型昆虫抑制性多肽的昆虫抑制性组合物。在另一方面,本发明提供一种控制鞘翅目或鳞翅目物种害虫,以及控制鞘翅目或鳞翅目物种害虫侵扰植物例如作物植物的方法,其中所述方法包括使所述害虫与昆虫抑制量的本发明的昆虫抑制性重组多肽诸如tic3668型昆虫抑制性多肽接触。在另一方面,本发明提供一种包含本发明的重组多核苷酸分子或昆虫抑制性重组多肽诸如tic3668型昆虫抑制性多肽的种子。在另一方面,本发明提供一种包含可检测量的本发明的重组多核苷酸分子或昆虫抑制性多肽诸如tic3668型昆虫抑制性多肽的商品产品。在另一方面,本发明的商品产品可包含宿主细胞,所述宿主细胞包含本发明的重组多核苷酸分子,其中所述商品产品包含可检测量的所述重组多核苷酸分子或由所述重组多核苷酸编码的昆虫抑制性重组多肽。在某些实施方案中,商品产品可包括由谷物处理器装袋的商品玉米、玉米片、玉米饼、玉米粉、玉米面、玉米糖浆、玉米油、玉米青贮料、玉米淀粉、玉米谷物等、以及相应大豆、稻、小麦、高粱、木豆、花生、水果、甜瓜和蔬菜商品产品,当可适用时包括含有可检测量的本申请的此类多核苷酸和/或多肽的此类商品产品的浆汁、浓缩物、果酱、胶冻、柑桔酱和其它可食用形式。在另一方面,本发明提供一种产生包含本发明的重组多核苷酸的种子的方法,其中所述方法包括:(a)种植至少一个包含所述重组多核苷酸分子的种子;(b)由所述种子来使植物生长;和(c)从所述植物收获种子,其中收获的种子包含所述重组多核苷酸分子。在另一方面,本发明提供一种包含本发明的重组多核苷酸分子的重组载体。在一个实施方案中,重组载体选自由质粒、杆粒、噬菌粒和粘粒组成的组。在另一方面,本发明提供一种对昆虫侵扰具有抗性的植物,其中所述植物的细胞包含本发明的重组多核苷酸分子或昆虫抑制性重组多肽。附图说明图1说明拼贴蛋白(collageprotein)tic4260与5种示例性tic3668型蛋白的比对。在这个序列比对中,以灰色阴影将具有序列多样性的位置突出显示。图2说明在植物中示例性叶绿体靶向和非靶向的成熟长度tic3668型蛋白的西方玉米根虫(wcr)抑制活性。图3说明在植物中示例性叶绿体靶向和非靶向成熟长度tic-3668型蛋白的wcr抑制活性。序列简述seqidno:1是从侧孢短芽孢杆菌物种获得的由在核苷酸位置1-951处的开放阅读框和翻译终止密码子来编码tic3668蛋白的重组多核苷酸序列。seqidno:2是由如seqidno:1所示的开放阅读框获得的tic3668前体蛋白的氨基酸序列翻译物。seqidno:3是从侧孢短芽孢杆菌物种获得的由在核苷酸位置1-951处的开放阅读框和翻译终止密码子来编码tic3669蛋白的重组多核苷酸序列。seqidno:4是由seqidno:3所示的开放阅读框获得的tic3669蛋白的氨基酸序列翻译物。seqidno:5是从侧孢短芽孢杆菌物种获得的由在核苷酸位置1-951处的开放阅读框和翻译终止密码子来编码tic3670蛋白的重组多核苷酸序列。seqidno:6是由seqidno:5所示的开放阅读框获得的tic3670前体蛋白的氨基酸序列翻译物。seqidno:7是从侧孢短芽孢杆菌物种获得的由在核苷酸位置1-951处的开放阅读框和翻译终止密码子来编码tic4076蛋白的重组多核苷酸序列。seqidno:8是由seqidno:7所示的开放阅读框获得的tic4076前体蛋白的氨基酸序列翻译物。seqidno:9是从侧孢短芽孢杆菌物种获得的由在核苷酸位置1-951处的开放阅读框和翻译终止密码子来编码tic4078蛋白的重组多核苷酸序列。seqidno:10是由seqidno:9所示的开放阅读框获得的tic4078前体蛋白的氨基酸序列翻译物。seqidno:11是从侧孢短芽孢杆菌物种获得的由在核苷酸位置1-951处的开放阅读框和翻译终止密码子来编码拼贴tic4260蛋白的重组多核苷酸序列,其通过框内组合来自seqidno:1、seqidno:3、seqidno:5、seqidno:7和seqidno:9所示的编码序列中的每一个的dna区段以包括这5个不同开放阅读框的序列变化来产生。seqidno:12是由seqidno:11所示的开放阅读框获得的拼贴蛋白tic4260前体蛋白的氨基酸序列翻译物。seqidno:13是从侧孢短芽孢杆菌物种获得的由在核苷酸位置1-951处的开放阅读框和翻译终止密码子来编码tic4346蛋白的重组多核苷酸序列。seqidno:14是seqidno:13所示的开放阅读框的氨基酸序列翻译物。seqidno:15是从侧孢短芽孢杆菌物种获得的由在核苷酸位置1-951处的开放阅读框和翻译终止密码子来编码tic4826蛋白的重组多核苷酸序列。seqidno:16是seqidno:15所示的开放阅读框的氨基酸序列翻译物。seqidno:17是从侧孢短芽孢杆菌物种获得的由在核苷酸位置1-918处的开放阅读框和翻译终止密码子来编码tic4861蛋白的重组多核苷酸序列。seqidno:18是seqidno:17所示的开放阅读框的氨基酸序列翻译物。seqidno:19是从侧孢短芽孢杆菌物种获得的由在核苷酸位置1-945处的开放阅读框和翻译终止密码子来编码tic4862蛋白的重组多核苷酸序列。seqidno:20是seqidno:19所示的开放阅读框的氨基酸序列翻译物。seqidno:21是从侧孢短芽孢杆菌物种获得的由在核苷酸位置1-951处的开放阅读框和翻译终止密码子来编码tic4863蛋白的重组多核苷酸序列。seqidno:22是seqidno:21所示的开放阅读框的氨基酸序列翻译物。seqidno:23是成熟tic3668蛋白mtic3668的氨基酸序列。seqidno:24是成熟tic3669蛋白mtic3669的氨基酸序列。seqidno:25是成熟tic3670蛋白mtic3670的氨基酸序列。seqidno:26是成熟tic4076蛋白mtic4076的氨基酸序列。seqidno:27是成熟tic4078蛋白mtic4078的氨基酸序列。seqidno:28是成熟tic4260蛋白mtic4260的氨基酸序列。seqidno:29是成熟tic4346蛋白mtic4346的氨基酸序列。seqidno:30是成熟tic4826蛋白mtic4826的氨基酸序列。seqidno:31是成熟tic4861蛋白mtic4891的氨基酸序列。seqidno:32是经设计用于在植物中表达的编码tic3668蛋白的合成核苷酸序列。seqidno:33是经设计用于在植物中表达的编码成熟tic3668蛋白mtic3668的合成核苷酸序列。seqidno:34是经设计用于在植物中表达的编码tic3669蛋白的合成核苷酸序列。seqidno:35是经设计用于在植物中表达的编码成熟tic3669蛋白mtic3669的合成核苷酸序列。seqidno:36是经设计用于在植物中表达的编码tic3670蛋白的合成核苷酸序列。seqidno:37是经设计用于在植物中表达的编码成熟tic3670蛋白mtic3670的合成核苷酸序列。seqidno:38是经设计用于在植物中表达的编码tic4076蛋白的合成核苷酸序列。seqidno:39是经设计用于在植物中表达的编码成熟tic4076蛋白mtic4076的合成核苷酸序列。seqidno:40是经设计用于在植物中表达的编码tic4078蛋白的合成核苷酸序列。seqidno:41是经设计用于在植物中表达的编码成熟tic4078蛋白mtic4078的合成核苷酸序列。seqidno:42是经设计用于在植物中表达的编码tic4260蛋白的合成核苷酸序列。seqidno:43是经设计用于在植物中表达的编码成熟tic4260蛋白mtic4260的合成核苷酸序列。seqidno:44是经设计用于在植物中表达的编码tic4346蛋白的合成核苷酸序列。seqidno:45是经设计用于在植物中表达的编码成熟tic4346蛋白mtic4346的合成核苷酸序列。seqidno:46是经设计用于在植物中表达的编码tic4826蛋白的合成核苷酸序列。seqidno:47是经设计用于在植物中表达的编码成熟tic4826蛋白mtic4826的合成核苷酸序列。seqidno:48是经设计用于在植物中表达的编码tic4861蛋白的合成核苷酸序列。seqidno:49是经设计用于在植物中表达的编码成熟tic4861蛋白(mtic4861)、成熟tic4862蛋白(mtic4862)和成熟tic4863蛋白(mtic4863)的合成核苷酸序列。seqidno:50是经设计用于在植物中表达的编码tic4682蛋白的合成核苷酸序列。seqidno:51是经设计用于在植物中表达的编码tic4863蛋白的合成核苷酸序列。seqidno:52是表示用于与编码本申请中公开的蛋白质的dna的(-)链杂交的合成寡核苷酸的核苷酸序列,并且对应于seqidno:1的位置1至36(tic3668正向引物)。seqidno:53是表示用于与编码本申请中公开的蛋白质的dna的(+)链杂交的合成寡核苷酸的核苷酸序列,并且对应于seqidno:1的位置920至954(tic3668反向引物)。seqidno:54是表示用于与编码本申请中公开的蛋白质的dna的(-)链杂交的合成寡核苷酸的核苷酸序列,并且对应于seqidno:3的位置1至41(tic3669正向引物)。seqidno:55是表示用于与编码本申请中公开的蛋白质的dna的(+)链杂交的合成寡核苷酸的核苷酸序列,并且对应于seqidno:3的位置920至954(tic3669反向引物)。seqidno:56是表示用于与编码本申请中公开的蛋白质的dna的(-)链杂交的合成寡核苷酸的核苷酸序列,并且对应于seqidno:5的位置1至36(tic3670正向引物)。seqidno:57是表示用于与编码本申请中公开的蛋白质的dna的(+)链杂交的合成寡核苷酸的核苷酸序列,并且对应于seqidno:5的位置920至954(tic3670反向引物)。seqidno:58是表示用于与编码本申请中公开的蛋白质的dna的(-)链杂交的合成寡核苷酸的核苷酸序列,并且对应于seqidno:7的位置1至41(tic4076正向引物)。seqidno:59是表示用于与编码本申请中公开的蛋白质的dna的(+)链杂交的合成寡核苷酸的核苷酸序列,并且对应于seqidno:7的位置920至954(tic4076反向引物)。seqidno:60是表示用于与编码本申请中公开的蛋白质的dna的(-)链杂交的合成寡核苷酸的核苷酸序列,并且对应于seqidno:9的位置1至36(tic4078正向引物)。seqidno:61是表示用于与编码本申请中公开的蛋白质的dna的(+)链杂交的合成寡核苷酸的核苷酸序列,并且对应于seqidno:9的位置920至954(tic4078反向引物)。seqidno:62是从侧孢短芽孢杆菌物种获得的由在核苷酸位置1-951处的开放阅读框和翻译终止密码子来编码tic2462蛋白的重组多核苷酸序列。seqidno:63是seqidno:62所示的开放阅读框的氨基酸序列翻译物。seqidno:64是用于在细菌中表达的编码成熟tic3668蛋白mtic3668的合成核苷酸序列。seqidno:65是用于在细菌中表达的编码成熟tic3669蛋白mtic3669的合成核苷酸序列。seqidno:66是用于在细菌中表达的编码成熟tic3670蛋白mtic3670的合成核苷酸序列。seqidno:67是用于在细菌中表达的编码成熟tic4076蛋白mtic4076的合成核苷酸序列。seqidno:68是用于在细菌中表达的编码成熟tic4078蛋白mtic4078的合成核苷酸序列。seqidno:69是用于在细菌中表达的编码成熟tic4260蛋白mtic4260的合成核苷酸序列。seqidno:70是用于在细菌中表达的编码成熟tic4346蛋白mtic4346的合成核苷酸序列。seqidno:71是用于在细菌中表达的编码成熟tic4826蛋白mtic4826的合成核苷酸序列。seqidno:72是用于在细菌中表达的编码成熟tic4861(mtic4861)、tic4862(mtic4862)和tic4863(mtic4863)蛋白的合成核苷酸序列。具体实施方式可将农业害虫控制领域中的问题表征为需要针对靶标害虫有效力,展现针对靶标害虫物种的广谱毒性,能够在植物中表达而不导致不合需要的农艺学问题,并且相较于商业上用于植物中的当前毒素提供替代性作用模式的新型毒素蛋白质。由tic3668例示的新型杀虫蛋白质在本文中被公开,并且解决这些需要中的各个,特别是针对广谱鞘翅目和鳞翅目昆虫害虫,并且更特别是针对玉米根虫害虫物种。在本申请中提及“tic3668”、“tic3668蛋白”、“tic3668蛋白毒素”、“tic3668毒素蛋白”、“tic3668相关毒素”、“tic3668相关蛋白毒素类别或家族”、“tic3668相关毒素蛋白”、“tic3668型蛋白”、“tic3668样蛋白”、“tic3668相关毒素多肽”、“tic3668相关杀害虫蛋白”或“tic3668型昆虫抑制性多肽”等是指包含以下,由以下组成,与以下基本上同源,与以下类似,或源于以下的任何新型昆虫抑制性蛋白:tic3668(seqidno:2)的赋予针对鞘翅目害虫和鳞翅目害虫的活性的任何昆虫抑制性多肽序列及其昆虫抑制性区段或其组合,包括展现昆虫抑制活性的任何蛋白质,条件是此类蛋白质与tic3668(seqidno:2)、tic3669(seqidno:4)、tic3670(seqidno:6)、tic4076(seqidno:8)、tic4078(seqidno:10)、tic4346(seqidno:14)、tic4826(seqidno:16)、tic4861(seqidno:18)、tic4862(seqidno:20)和tic4863(seqidno:22)的比对产生约35%至约100%百分比的任何百分比分数的氨基酸序列同一性。本申请中公开的tic3668型蛋白毒素包括tic3668、tic3669、tic3670、tic4076、tic4078、tic4346、tic4826、tic4861、tic4862、tic4863和拼贴tic4260蛋白(seqidno:12)。tic3668型蛋白类别旨在包括前体形式以及成熟长度形式的蛋白质。术语“区段”或“片段”在本申请中用于描述相比于描述tic3668型蛋白的完全氨基酸或核酸序列,较短的连续氨基酸或核酸序列。本申请中也公开展现昆虫抑制活性的区段或片段,条件是此类区段或片段与seqidno:2所示的tic3668型蛋白的相应节段的比对产生在所述区段或片段与tic3668型蛋白的相应节段之间约35至约100百分比的任何百分比分数的氨基酸序列同一性。在本申请中提及术语“活性的”或“活性”、“杀虫活性”或“杀虫活性”、“昆虫抑制性的”或“杀虫性的”是指毒性剂诸如蛋白质毒素在抑制(抑制生长、摄食、繁殖力或活力)、遏制(遏制生长、摄食、繁殖力或活力)、控制(控制害虫侵扰,控制害虫对含有有效量的tic3668型蛋白的特定作物的摄食活动)或杀灭(导致致病、致死或繁殖力降低)害虫方面的功效。这些术语旨在包括向害虫提供杀害虫有效量的毒性蛋白质的结果,其中所述害虫暴露于所述毒性蛋白质导致致病、致死、繁殖力降低或阻碍生长。这些术语也包括由于在植物中或在植物上提供杀害虫有效量的毒性蛋白质而从植物、植物的组织、植物部分、种子、植物细胞或从植物可正在生长所处的特定地理位置排斥害虫。一般来说,杀虫活性是指毒性蛋白质能够有效抑制特定靶标害虫(包括但不限于鳞翅目或鞘翅目的昆虫)的生长、发育、活力、摄食行为、交配行为、繁殖力,或由昆虫以这个蛋白质、蛋白质片段、蛋白质区段或多核苷酸为食引起的使不利作用的任何可测量降低。毒性蛋白质可由植物产生,或可被施加于植物或植物所位于的位置内的环境。术语“生物活性”、“有效”、“有效力”或其变化形式也是在本申请中可互换用于描述本发明的蛋白质对靶标昆虫害虫的作用的术语。当提供在靶标害虫的膳食中时,杀害虫有效量的毒性剂在所述毒性剂接触所述害虫时展现杀虫活性。毒性剂可为杀害虫蛋白质或一种或多种本领域中已知的化学剂。杀虫化学剂和杀虫蛋白质试剂可单独或彼此组合使用。化学剂包括但不限于靶向特定基因以在靶标害虫中达成遏制的dsrna分子、有机氯化物、有机磷酸酯、氨基甲酸酯、拟除虫菊酯(pyrethroid)、新烟碱类(neonicotinoid)和莱恩碱类(ryanoid)。杀虫蛋白质试剂包括本申请中所示的蛋白质毒素,以及其它蛋白质毒性剂,包括靶向鳞翅目和鞘翅目的那些,以及用于控制其它植物害虫的蛋白质毒素诸如本领域中可用的用于控制半翅目和同翅目物种的cry蛋白。预期对害虫特别是作物植物的害虫的提及意指作物植物的昆虫害虫,特别是由tic3668相关蛋白毒素类别控制的那些。然而,当在以下情况下时提及害虫也可包括植物的半翅目和同翅目昆虫害虫以及线虫和真菌:靶向这些害虫的毒性剂连同tic3668相关蛋白毒素类别的一种或多种蛋白质一起共同定位或存在。构成tic3668相关蛋白类别的个别蛋白质由于具有共同功能而相关联,并且展现对来自鞘翅目和鳞翅目昆虫物种的昆虫害虫的杀虫活性,所述昆虫害虫包括成虫、蛹、幼虫和新生体(neonate)。鳞翅目的昆虫包括但不限于夜蛾科(noctuidae)中的行军虫、切根虫、尺蠖和棉铃虫,例如秋行军虫(草地夜蛾)、甜菜行军虫(甜菜夜蛾)、披肩行军虫(蓓带夜蛾(mamestraconfigurata))、黑切根虫(小地老虎)、卷心菜尺蠖(粉纹夜蛾)、大豆尺蠖(大豆夜蛾(pseudoplusiaincludens))、黎豆毛虫(黎豆夜蛾)、绿三叶草虫(苜蓿绿夜蛾(hypenascabra))、烟草蚜虫(烟芽夜蛾)、颗粒切根虫(粒肤地老虎(agrotissubterranea))、行军虫(一星粘虫(pseudaletiaunipuncta))、西方切根虫(西方灰地老虎(agrotisorthogonia));来自螟蛾科(pyralidae)的螟虫、鞘蛾幼虫、结网虫、锥虫、卷心菜虫和雕叶虫,例如欧洲玉米螟虫(欧洲玉米螟)、脐橙虫(脐橙螟(amyeloistransitella))、玉米根结网虫(玉米根草螟(crambuscaliginosellus))、草地结网虫(稻切叶野螟(herpetogrammalicarsisalis))、向日葵蛾(美洲葵螟(homoeosomaelectellum))、小玉米杆螟(南美玉米苗斑螟(elasmopalpuslignosellus));卷蛾科(tortricidae)中的卷叶虫、蚜虫、种子蠕虫和果实蠕虫,例如苹果蠹蛾(苹果小卷蛾(cydiapomonella))、葡萄浆果蛾(葡萄卷叶蛾(endopizaviteana))、东方果蛾(梨小食心虫(grapholitamolesta))、向日葵芽蛾(向日葵芽卷叶蛾(suleimahelianthana));以及许多其它在经济上重要的鳞翅目昆虫,例如菱纹背蛾(小菜蛾)、粉螟蛉虫(棉红铃虫)和吉卜赛蛾(舞毒蛾(lymantriadispar))。鳞翅目的其它昆虫害虫包括例如棉叶波纹夜蛾(alabamaargillacea)(棉叶蠕虫)、果树黄卷蛾(archipsargyrospila)(果树卷叶虫)、玫瑰黄卷蛾(archipsrosana)(欧洲卷叶虫)和其它黄卷蛾属物种、二化螟(chilosuppressalis)(亚洲稻螟或稻茎螟)、稻纵卷叶螟(cnaphalocrocismedinalis)(稻卷叶虫)、玉米根草螟(玉米根结网虫)、早熟禾草螟(crambusteterrellus)(早熟禾结网虫)、西南玉米杆草螟(西南玉米螟虫)、小蔗杆草螟(甘蔗螟虫)、埃及钻夜蛾(eariasinsulana)(多刺螟蛉虫)、翠纹钻夜蛾(eariasvittella)(斑点螟蛉虫)、棉铃虫(美洲螟蛉虫)、玉米穗蛾(helicoverpazea)(玉米耳虫或棉花螟蛉虫)、烟芽夜蛾(烟草蚜虫)、稻切叶野螟(草地结网虫)、葡萄花翅小卷蛾(lobesiabotrana)(欧洲葡萄藤蛾)、柑桔潜叶蛾(phyllocnistiscitrella)(柑桔潜叶虫)、欧洲粉蝶(pierisbrassicae)(大白蝴蝶)、菜粉蝶(pierisrapae)(外来卷心菜虫或小白蝴蝶)、小菜蛾(菱纹背蛾)、甜菜夜蛾(甜菜行军虫)、斜纹夜蛾(spodopteralitura)(烟草切根虫、群集毛虫)和番茄斑潜蝇(tutaabsoluta)(番茄潜叶虫)。鞘翅目的昆虫包括但不限于叩甲属某些种、花象属某些种、甜菜隐食甲、蚤凹胫跳甲、根颈象属某些种、象虫属某些种、皮蠹属某些种、根叶甲属某些种、食植瓢虫属某些种、eremnus属某些种、科罗拉多金花虫、稻水象属某些种、鳃金龟属某些种、锯谷盗属某些种、耳喙象属某些种、斑象属某些种、弧丽金龟属某些种、蚤跳甲属某些种、谷蠹属某些种、金龟子科、象虫属某些种、麦蛾属某些种、拟步行虫属某些种、拟谷盗属某些种和斑皮蠹属某些种,特别是当害虫是玉米根叶甲(西方玉米根虫wcr)、巴氏根叶甲(北方玉米根虫ncr)、墨西哥玉米根叶甲(墨西哥玉米根虫mcr)、带斑黄瓜根叶甲(巴西玉米根虫(bzr))、霍华德十一星根叶甲(南方玉米根虫scr)和由绿根叶甲和美丽根叶甲组成的巴西玉米根虫复合体(bcr)时。在本申请中提及“经分离的dna分子”、“经分离的多核苷酸分子”或等效术语或短语意指dna分子是单独或与其它组成成分组合存在,但不在它的天然环境内的dna分子。举例来说,天然见于生物体的基因组的dna内的核酸元件诸如编码序列、内含子序列、未翻译的前导序列、启动子序列、转录终止序列等不被视为是“经分离的”,只要所述元件在所述生物体的基因组内以及在它所天然见于其中的基因组内的位置处即可。然而,在本公开的范围内,这些元件中的各个以及这些元件的子部分将为“经分离的”,只要所述元件不在生物体的基因组内以及不在它所天然见于其中的基因组内的位置处即可。类似地,编码杀虫蛋白质或杀虫蛋白质的任何天然存在的杀虫变体的核苷酸序列将为经分离的核苷酸序列,只要所述核苷酸序列不在编码所述蛋白质的所述序列所天然见于的细菌的dna内即可。出于本公开的目的,编码天然存在的杀虫蛋白质的氨基酸序列的合成核苷酸序列将被视为是经分离的。出于本公开的目的,任何转基因核苷酸序列,即插入植物或细菌的细胞的基因组中或存在于染色体外载体中的dna的核苷酸序列,将被视为是经分离的核苷酸序列,无论它存在于用于转化细胞的质粒或类似结构内,植物或细菌的基因组内,抑或以可检测量存在于源于植物或细菌的组织、子代、生物样品或商品产品中。如本申请中进一步所述,在从侧孢短芽孢杆菌菌株eg5552获得的dna中发现编码tic3668(seqidno:2)的开放阅读框(orf)(seqidno:1)。接着筛选其它细菌基因组中编码tic3668相关蛋白的序列。在这些其它细菌基因组中鉴定了编码类似于eg5552tic3668蛋白的氨基酸序列的若干其它开放阅读框,包括以下tic3668样蛋白:在从侧孢短芽孢杆菌菌株eg5551获得的dna中发现的tic3669(seqidno:3编码seqidno:4)、在从侧孢短芽孢杆菌菌株eg5553获得的dna中发现的tic3670(seqidno:5编码seqidno:6)、在从侧孢短芽孢杆菌菌株atcc6456获得的dna中发现的tic4076(seqidno:7编码seqidno:8)、在从侧孢短芽孢杆菌菌株eg4227获得的dna中发现的tic4078(seqidno:9编码seqidno:10)、在从侧孢短芽孢杆菌菌株eg5551获得的dna中发现的tic4346(seqidno:13编码seqidno:14)、在从侧孢短芽孢杆菌菌株ag0021d10获得的dna中发现的tic4826(seqidno:15编码seqidno:16)、在从侧孢短芽孢杆菌菌株eg4227获得的dna中发现的tic4861(seqidno:17编码seqidno:18)、tic4862(seqidno:19编码seqidno:20)和tic4863(seqidno:21编码seqidno:22)。通过组合来自5种不同天然tic3668样蛋白的天然存在的氨基酸序列变体以产生拼贴蛋白来产生一种额外tic3668样蛋白tic4260(seqidno:11编码seqidno:12)。克隆相应编码序列,并且在微生物宿主细胞中表达以产生用于昆虫生物测定中的重组蛋白。如本申请中进一步所述,显示这些蛋白质展现针对根叶甲属物种的生物活性,所述物种包括西方玉米根虫(wcr,玉米根叶甲)、西方欧洲玉米螟虫(ecb,玉米螟)、西南玉米螟虫(swc,西南玉米杆草螟)和大豆尺蠖(soybeanlooper)(sbl,大豆尺蠖(chrysodeixisincludens))。tic3668型蛋白的一惊人特征是存在对应于氨基酸位置1至23(对于tic3668、tic3669、tic3670、tic4076、tic4078、tic4260、tic4346、tic4826、tic4863);1至12(对于tic4861);和1至21(对于tic4862)的n末端氨基酸区段。这些n末端氨基酸区段各自可从相应蛋白质省略,并且编码相应区段的多核苷酸序列也可被省略。当在植物中表达时,相较于表达含有省略区段的全长蛋白质毒素,省略这些相应区段惊人地导致针对玉米根虫物种的杀虫活性增加。缺乏以上提及的n末端氨基酸区段的蛋白质毒素区段在本文中被称为“成熟tic3668型毒素蛋白质”。一般来说,提及成熟形式的tic3668型蛋白在本文中用字母“m”在毒素的名称之前加以注解以区分提及来自全长天然序列的成熟序列。举例来说,tic3668的氨基酸序列(seqidno:2)的成熟形式是mtic3668(seqidno:23)。tic3669(seqidno:4)、tic3670(seqidno:6)、tic4076(seqidno:8)、tic4078(seqidno:10)、tic4260(seqidno:12)、tic4346(seqidno:14)和tic4826(seqidno:16)的成熟形式分别是mtic3669(seqidno:24)、mtic3670(seqidno:25)、mtic4076(seqidno:26)、mtic4078(seqidno:27)、mtic4260(seqidno:28)、mtic4346(seqidno:29)和mtic4826(seqidno:30)。全长蛋白质tic4861(seqidno:18)、tic4862(seqidno:20)和tic4863(seqidno:22)是彼此的序列长度变体,并且仅在它们的n末端氨基酸区段的长度方面不同。移除tic4861、tic4862和tic4863中的n末端氨基酸区段产生mtic4861、mtic4862和mtic4863的同一成熟氨基酸序列。因此,mtic4861、mtic4862和mtic4863的氨基酸序列由相同多核苷酸序列(mtic4861,seqidno:31)编码。成熟tic3668样蛋白序列由seqidno:64(编码mtic3668)、seqidno:65(编码mtic3669)、seqidno:66(编码mtic3670)、seqidno:67(编码mtic4076)、seqidno:68(编码mtic4078)、seqidno:69(编码mtic4260)、seqidno:70(编码mtic4346)、seqidno:71(编码mtic4826)和seqidno.72(编码mtic4861、mtic4862和mtic4863)编码以在细菌宿主中表达。可通过使用来自一些或所有家族成员的天然存在的氨基酸变化以产生具有较高氨基酸序列多样性水平和新型性质的新型蛋白质来产生tic3668型家族的额外成员。通过以下方式来产生tic3668型蛋白毒素类别的变体:比对tic3668型家族成员的氨基酸序列并在氨基酸序列水平上组合差异以获得新型氨基酸序列,以及对编码这些变体的多核苷酸进行适当变化。一个此类实例是tic4260。seqidno:11是编码tic4260蛋白(seqidno:12)的多核苷酸序列。成熟蛋白质(mtic4260,seqidno:28)由多核苷酸序列seqidno:43编码。tic3668型蛋白毒素的片段可为其中从蛋白质的n末端、c末端、中部或其组合缺失一个或多个氨基酸,具有昆虫抑制活性的截短形式。这些片段可为tic3668、tic3669、tic3670、tic4260、tic4076、tic4078、tic4346、tic4826、tic4861、tic4862或tic4863的天然存在的变体或合成变体,但应保留或改进tic3668、tic3669、tic3670、tic4260、tic4076、tic4078、tic4346、tic4826、tic4861、tic4862或tic4863的昆虫抑制活性。截短的n末端或c末端缺失变体包括但不限于缺乏来自n末端和/或c末端的氨基酸残基的tic3668、tic3669、tic3670、tic4260、tic4076、tic4078、tic4346、tic4826、tic4861、tic4862或tic4863蛋白。举例来说,tic3668蛋白的n末端氨基酸残基1至23可被缺失,从而产生具有seqidno:2的氨基酸24-317的毒素蛋白质。从tic3668蛋白的c末端氨基酸末端移除10或20个氨基酸导致杀虫活性损失,而移除单一氨基酸不影响活性。可通过使用本领域中已知的各种基于计算机的算法进行彼此比较来鉴定tic3668型蛋白类别的蛋白质以及类似于tic3668型蛋白类别的蛋白质的蛋白质(参见表1和2)。本文报道的氨基酸序列同一性是使用以下这些缺省参数进行clustalw比对的结果:权重矩阵:blosum,空位开放罚分:10.0,空位延伸罚分:0.05,亲水性空位:开启(on),亲水性残基:gpsndqerk,残基特异性空位罚分:开启(thompson等(1994)nucleicacidsresearch,22:4673-4680)。进一步通过100%乘以(同一氨基酸数目/主题蛋白质的长度)的乘积来计算氨基酸同一性百分比。其它比对算法在本领域中也是可用的,并且提供与使用clustalw比对获得的结果类似的结果。预期展现针对鳞翅目昆虫物种的昆虫抑制活性的蛋白质是tic3668型蛋白毒素类别的成员,条件是所述蛋白质用于例如在clustalw比对中的查询中,并且如阐述为mtic4260的至少一种本发明的蛋白质在此类比对中被鉴定为命中物(hit),其中查询蛋白质沿查询蛋白质的长度展现至少约85%至约100%氨基酸序列同一性,即85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、100%或在这个范围内的任何百分比分数;或如阐述为mtic3668的至少一种本发明的蛋白质在此类比对中被鉴定为命中物,其中查询蛋白质沿查询蛋白质的长度展现至少约89%至约100%氨基酸序列同一性,即89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、100%或在这个范围内的任何百分比分数;或如阐述为mtic3669和/或mtic3670的至少一种本发明的蛋白质在此类比对中被鉴定为命中物,其中查询蛋白质沿查询蛋白质的长度展现至少约90%至约100%氨基酸序列同一性,即90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、100%或在这个范围内的任何百分比分数;或如阐述为mtic4826的至少一种本发明的蛋白质在此类比对中被鉴定为命中物,其中查询蛋白质沿查询蛋白质的长度展现至少约91%至约100%氨基酸序列同一性,即91%、92%、93%、94%、95%、96%、97%、98%、99%、100%或在这个范围内的任何百分比分数。预期展现针对鞘翅目昆虫物种的昆虫抑制活性的蛋白质是tic3668型蛋白毒素类别的成员,条件是所述蛋白质用于例如在clustalw比对中的查询中,并且如阐述为mtic3668、mtic3669、mtic3670、mtic4076、mtic4078、mtic4260、mtic4346、mtic4826、mtic4861、mtic4862和mtic4863的至少一种本发明的蛋白质在此类比对中被鉴定为命中物,其中查询蛋白质沿查询蛋白质的长度展现至少约35%至约100%氨基酸同一性,即约35%、40%、50%、60%、70%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、85%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、100%或在这个范围内的任何百分比分数。使用clustalw算法将tic3668型蛋白毒素类别的示例性蛋白质彼此比对。创建各对全长蛋白质的氨基酸序列同一性百分比的成对矩阵,如表1中所报道。创建各对成熟长度蛋白质的氨基酸序列同一性百分比的成对矩阵,如表2中所报道。表1.示例性全长蛋白质的成对矩阵显示表格描述:在(x)相对于(y)之间的clustalw比对报道于成对矩阵中。在(n)下的各列是指seqidno。列(m)是指蛋白质名称(tic#)。计算在所有各对之间的氨基酸同一性百分比,并且由各框中的第一数字表示。各框中的第二数字(在括号中)表示这对之间的同一氨基酸数目。表2.示例性成熟蛋白质的成对矩阵显示表格描述:在(x)相对于(y)之间的clustalw比对报道于成对矩阵中。在(n)下的各列是指seqidno。列(m)是指蛋白质名称(tic#)。计算在所有各对之间的氨基酸同一性百分比,并且由各框中的第一数字表示。各框中的第二数字(在括号中)表示这对之间的同一氨基酸数目。tic3668型蛋白毒素类别的全长蛋白质和成熟蛋白质也可由于一级结构(保守氨基酸基序)、长度(成熟蛋白质约295个氨基酸,而全长蛋白质约317个氨基酸)和其它特征而相关联。当在变性条件下在蛋白质凝胶上跑胶时,本发明的全长蛋白质具有约35k道尔顿(dalton)的测量质量,而成熟蛋白质具有约32kda的测量质量。tic3668型蛋白毒素类别的全长形式和成熟形式的特征报道于表3和4中。表3.全长蛋白质的特征表4–成熟蛋白质的特征所公开tic3668型蛋白毒素类别的蛋白质表示一类新型杀虫蛋白质。参照表5,在对应于100%同一性的对角线以上的所有数字都表示在那个特定行和列的交叉点处所比较的相应蛋白质之间的氨基酸差异的数目。在对应于100%同一性的对角线以下的数字表示在那个特定行和列的交叉点处所比较的相应蛋白质的同一性百分比。这个蛋白质类别的成熟长度成员展现与本领域中已知的任何其它杀虫蛋白质的不大于90.54%氨基酸同一性,如表5中提供的比对中所显示。展现与本发明的任何成熟长度蛋白质的最接近同一性的杀虫蛋白质是美国专利申请公布号20110030093中的seqidno:50(axmi-209),与mtic4076、mtic4346、mtic4826和mtic4863具有90.5%序列同一性。这个公开仅教导针对鳞翅目的活性,而本发明的示例性蛋白质显示针对鞘翅目的活性。h0udd3_brela、f7tvp6_brela和u4wsu1_brela是根据基因组序列中的开放阅读框预测,被报道为已从侧孢短芽孢杆菌获得的未注释蛋白质序列。未报道这些蛋白质的杀虫活性。表5–成熟长度tic3886蛋白与先前技术蛋白质的比对本申请中公开的tic3668蛋白在膳食生物测定中展现针对包括wcr的鞘翅目的活性。在一些情况下,也观察到鳞翅目活性。如本申请的实施例中进一步所述,设计编码tic3668毒素蛋白的多核苷酸序列以用于植物中。经设计用于在植物中表达,并且编码全长昆虫抑制性tic3668、tic3669、tic3670、tic4260、tic4076、tic4078、tic4346、tic4826、tic4861、tic4862和tic4863蛋白的示例性多核苷酸以seqidno:32、seqidno:34、seqidno:36、seqidno:38、seqidno:40、seqidno:42、seqidno:44、seqidno:46、seqidno:48、seqidno:50和seqidno:51示出。经设计用于在植物中表达,并且编码成熟形式的昆虫抑制性mtic3668、mtic3669、mtic3670、mtic4260、mtic4076、mtic4078、mtic4346、mtic4826、mtic4861、mtic4862和mtic4863蛋白的示例性多核苷酸以seqidno:33、seqidno:35、seqidno:37、seqidno:39、seqidno:41、seqidno:43、seqidno:45、seqidno:47和seqidno:49示出。构建含有这些多核苷酸序列的表达盒和载体,并且根据本领域中已知的转化方法和技术引入玉米植物细胞中。使经转化的细胞再生成被观察到表达tic3668毒素蛋白的经转化的植物。为测试杀虫活性,使用从经转化的植物获得的植物叶盘,在鳞翅目或鞘翅目害虫幼虫存在下进行生物测定。tic3668型蛋白毒素类别的示例性成员的昆虫抑制活性更详细描述于实施例中。示例性蛋白质由于具有共同功能而相关联,并且展现对鞘翅目和鳞翅目昆虫物种的杀虫活性,所述物种包括成虫、蛹、幼虫和新生体。涵盖编码tic3668型蛋白的重组多核苷酸组合物。举例来说,tic3668型蛋白可用重组dna构建体来表达,其中使具有编码蛋白质的orf的多核苷酸分子可操作地连接至基因表达元件,诸如启动子和为在构建体所意图的系统中表达所必需的任何其它调控元件。非限制性实例包括植物功能性启动子可操作地连接至tic3668型蛋白编码序列以在植物中表达蛋白质,或bt功能性启动子可操作地连接至tic3668型蛋白编码序列以在bt细菌或其它芽孢杆菌属物种中表达蛋白质。可使其它元件可操作地连接至tic3668型蛋白编码序列,包括但不限于增强子、内含子、未翻译的前导物、编码蛋白质固定标签(his标签)、易位肽(即质体转运肽、信号肽)、翻译后修饰酶的多肽序列、核糖体结合位点和rnai靶标位点。随本文一起提供的示例性重组多核苷酸分子包括但不限于异源启动子可操作地连接至多核苷酸,诸如编码具有如以seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:12、seqidno:14、seqidno:16、seqidno:18、seqidno:20和seqidno:22示出的氨基酸序列的相应多肽或蛋白质的seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11、seqidno:13、seqidno:15、seqidno:17、seqidno:19和seqidno:21。编码本文公开的蛋白质的重组多核苷酸分子的密码子可由同义密码子取代(本领域中称为沉默取代)。编码本申请中公开的任何tic3668型蛋白的经修饰的多核苷酸的非限制性实例以seqidno:32、seqidno:34、seqidno:36、seqidno:38、seqidno:40、seqidno:42、seqidno:44、seqidno:46、seqidno:48、seqidno:50和seqidno:51(对于全长蛋白质序列)以及seqidno:33、seqidno:35、seqidno:37、seqidno:39、seqidno:41、seqidno:43、seqidno:45、seqidno:47和seqidno:49(对于成熟蛋白质序列)示出。包含tic3668型蛋白编码序列的重组dna构建体可还包含编码一种或多种昆虫抑制剂的dna区域,其可被构造来与编码tic3668型蛋白的dna序列并行表达或共同表达与tic3668型蛋白不同的蛋白质、昆虫抑制性dsrna分子或辅助蛋白质。辅助蛋白质包括但不限于辅因子、酶、结合配偶体或起有助于昆虫抑制剂的有效性的作用的其它试剂,例如通过帮助它的表达,影响它在植物中的稳定性,优化寡聚自由能,加强它的毒性,以及增大它的活性谱。辅助蛋白质可促进对一种或多种昆虫抑制剂的摄取(例如)或增强毒性剂的毒性作用。可装配重组dna构建体以使所有蛋白质或dsrna分子都根据一个启动子表达,或各蛋白质或dsrna分子处于单独启动子控制下,或处于其某一组合的情况下。本发明的蛋白质可由多基因表达系统表达,其中tic3668蛋白中的一种或多种蛋白质由共同核苷酸区段表达,所述区段视所选表达系统的类型而定也含有其它开放阅读框和启动子。举例来说,细菌多基因表达系统可利用单一启动子来驱动来自单一操纵子内的多重连接/串联开放阅读框的表达(即多顺反子表达)。另一实例中,植物多基因表达系统可利用未多重连接表达盒,其各自表达不同蛋白质或其它试剂诸如一种或多种dsrna分子。可通过载体来向宿主细胞递送包含tic3668型蛋白编码序列的重组多核苷酸或重组dna构建体,所述载体例如质粒、杆状病毒、合成染色体、病毒粒子、粘粒、噬菌粒、噬菌体或病毒载体。此类载体可用于实现在宿主细胞中稳定或瞬时表达tic3668型蛋白编码序列,或随后表达所编码多肽。包含tic3668型蛋白编码序列,并且引入宿主细胞中的外源性重组多核苷酸或重组dna构建体在本文中被称为“转基因”。本文提供含有表达任何一种或多种tic3668型蛋白编码序列的重组多核苷酸的转基因细菌、转基因植物细胞、转基因植物和转基因植物部分。术语“细菌细胞”或“细菌”可包括但不限于土壤杆菌属、芽孢杆菌属、埃希氏菌属(escherichia)、沙门氏菌属(salmonella)、假单胞菌属或根瘤菌属细胞。术语“植物细胞”或“植物”可包括但不限于单子叶植物、双子叶植物、苜蓿、香蕉、大麦、菜豆、西兰花、卷心菜、芸苔、胡萝卜、木薯、蓖麻、花椰菜、芹菜、鹰嘴豆、大白菜、柑桔、椰子、咖啡、玉米、三叶草、棉花、葫芦、黄瓜、花旗松、茄子、桉树、亚麻、大蒜、葡萄、蛇麻子、韭菜、莴苣、火炬松、粟、甜瓜、坚果、燕麦、橄榄、洋葱、观赏植物、棕榈、牧场草、豌豆、花生、胡椒、木豆、松树、马铃薯、白杨树、南瓜、辐射松、萝卜、油菜籽、稻、根茎、黑麦、红花、灌木、高粱、南方松、大豆、菠菜、南瓜小果、草莓、糖用甜菜、甘蔗、向日葵、甜玉米、枫香、甜薯、柳枝稷、茶树、烟草、番茄、黑小麦、草皮草、西瓜和小麦植物细胞或植物。在某些实施方案中,提供由转基因植物细胞再生的转基因植物和转基因植物部分。在某些实施方案中,转基因植物可通过切割、折断、研磨或另外解离来自植物的部分来从转基因种子获得。在某些实施方案中,植物部分可为种子、圆荚、叶、花、茎、根或其任何部分或转基因植物部分的非可再生部分。如在这个情形下所用,转基因植物部分的“非可再生”部分是不能被诱导以形成完整植物,或不能被诱导以形成能够有性和/或无性繁殖的完整植物的部分。在某些实施方案中,植物部分的非可再生部分是转基因种子、圆荚、叶、花、茎或根的部分。提供制备包含鞘翅目或鳞翅目昆虫抑制量的tic3668型蛋白的转基因植物的方法。此类植物可通过以下方式来制备:将编码本申请中提供的任何tic3668型蛋白的重组多核苷酸引入植物细胞中,以及选择源于所述植物细胞的表达鞘翅目或鳞翅目昆虫抑制量的tic3668型蛋白的植物。植物可通过再生、播种、授粉或分生组织转化技术来源于植物细胞。用于转化植物的方法在本领域中是已知的。本申请中也公开经加工的植物产品,其中所述经加工的产品包含可检测量的tic3668型蛋白、其昆虫抑制性区段或片段、或其任何特性部分。在某些实施方案中,经加工的产品选自由植物部分、植物生物质、油、餐食、糖、动物饲料、面粉、薄片、糠、棉绒、壳、经加工的种子和种子组成的组。在某些实施方案中,经加工的产品是不可再生的。植物产品可包括源于转基因植物或转基因植物部分的商品或其它商业产品,其中所述商品或其它产品可通过检测编码或包含tic3668型蛋白的特性部分的核苷酸区段或所表达的rna或蛋白质通过商业加以追踪。表达tic3668蛋白的植物可通过培育来与表达其它毒素蛋白质和/或表达其它转基因性状(诸如除草剂耐受基因、赋予产量或应激耐受性状的基因等)的转基因事件杂交,或此类性状可组合在单一载体中以使性状全都加以关联。可使用为本领域普通技术人员所知的方法诸如聚合酶链式反应(pcr)、热扩增和杂交来鉴定tic3668型蛋白编码序列和与tic3668型蛋白编码序列具有基本上百分比同一性的序列。举例来说,tic3668型蛋白毒素类别的蛋白质可用于产生特异性结合这个类别的蛋白质的抗体,并且可用于筛选和发现所述类别的其它成员。此外,编码tic3668型蛋白毒素类别的核苷酸序列(和反向互补序列)可用作筛选的探针和引物以使用热循环或等温扩增和杂交方法来鉴定所述类别的其它成员。具体来说,源于seqidno:52至61中的任一个所示的序列的寡核苷酸可用于确定在源于商品产品的脱氧核糖核酸样品中存在或不存在tic3668型转基因。鉴于采用寡核苷酸的某些核酸检测方法的灵敏性,预期源于seqidno:52、seqidno:53、seqidno:54、seqidno:55、seqidno:56、seqidno:57、seqidno:58、seqidno:59、seqidno:60和seqidno:61中的任一个所示的序列的寡核苷酸可用于检测源于汇合来源的商品产品中的tic3668、tic3669、tic3670、tic4076、tic4078或tic4260转基因,在所述商品产品中,仅一部分商品产品源于含有seqidno:52、seqidno:53、seqidno:54、seqidno:55、seqidno:56、seqidno:57、seqidno:58、seqidno:59、seqidno:60和seqidno:61中的任一个的转基因植物。应进一步认识到此类寡核苷酸可用于在seqidno:52、seqidno:53、seqidno:54、seqidno:55、seqidno:56、seqidno:57、seqidno:58、seqidno:59、seqidno:60和seqidno:61中的每一个中引入核苷酸序列变体。此类“诱变”寡核苷酸用于鉴定展现一定范围的昆虫抑制活性或展现在转基因植物宿主细胞中的表达有变化的tic3668、tic3669、tic3670、tic4076、tic4078或tic4260氨基酸序列变体。核苷酸序列同源物,例如由在严格杂交条件下与本申请中公开的各个或任何序列杂交的核苷酸序列编码的杀虫蛋白质,也是本发明的一实施方案。本发明也提供一种用于检测与第二核苷酸序列杂交的第一核苷酸序列的方法,其中所述第一核苷酸序列(或它的反向互补序列)编码杀虫蛋白质或其杀虫片段,并且在严格杂交条件下与所述第二核苷酸序列杂交。在此类情况下,第二核苷酸序列可为在严格杂交条件下以tic3668型蛋白毒素类别公开的任何核苷酸序列。核苷酸编码序列在适当杂交条件下彼此杂交,并且由这些核苷酸序列编码的蛋白质与针对任一其它蛋白质产生的抗血清交叉反应。严格杂交条件在本领域中是已知的并可根据所需应用和结果而变化,并且可涵盖多种试剂和条件。举例来说,在较高温度下洗涤构成更严格条件。在某些实施方案中,本发明的杂交条件可包括至少在42℃下杂交,随后在室温下用2xssc、0.1%sds洗涤2次,每次5分钟,接着在65℃下在0.5xssc、0.1%sds中洗涤2次,每次30分钟;或在68℃下杂交,随后在68℃下在含有0.1%sds的2xssc中洗涤;或在37c下在50%甲酰胺、1mnacl和1%sds中杂交4至12小时,并且在60c-65c下在0.1xssc中洗涤。本领域技术人员将认识到由于遗传密码冗余,许多其它序列能够编码此类相关蛋白质,并且那些序列就它们起在芽孢杆菌属菌株中或在植物细胞中表达杀虫蛋白质的作用而言是本发明的实施方案,当然应认识到许多此类冗余编码序列在这些条件下将不与编码tic3668的天然芽孢杆菌属序列杂交。本申请涵盖使用这些鉴定方法和为本领域普通技术人员所知的其它鉴定方法来鉴定tic3668型蛋白编码序列和与tic3668型蛋白编码序列具有基本上百分比同一性的序列。本公开也涵盖使用本领域中已知的分子方法来工程改造和克隆商业上有用的蛋白质,所述蛋白质包括来自杀害虫蛋白质的蛋白质的嵌合体;例如可由tic3668型蛋白的区段装配嵌合体以获得额外有用的实施方案,包括装配tic3668型蛋白区段与不同于tic3668和相关蛋白质的不同蛋白质的区段。可使tic3668型蛋白类别经受彼此比对以及与其它芽孢杆菌属杀害虫蛋白质(无论这些蛋白质是否在系统发生方面关系密切或疏远)比对,并且可鉴定各种此类蛋白质的用于在所比对蛋白质之间替换,从而导致构建嵌合蛋白的区段。可使此类嵌合蛋白经受害虫生物测定分析,并且表征相较于嵌合体中各个此类区段所源于的亲本蛋白质,存在或不存在生物活性增加和/或靶标害虫谱扩大。可通过与其它蛋白质调换结构域或区段或通过使用本领域中已知的定向进化方法来进一步工程改造多肽的杀虫活性以获得对特定害虫或对更广泛谱系的害虫的活性。本申请中也公开用来自tic3668毒素蛋白类别的蛋白质控制昆虫(特别是鳞翅目或鞘翅目昆虫)侵扰作物植物的方法。此类方法可包括使包含鞘翅目或鳞翅目昆虫抑制量的tic3668毒素蛋白类别的蛋白质的植物生长。在某些实施方案中,此类方法可还包括以下操作中的任何一个或多个:(i)将包含或编码tic3668型蛋白毒素类别的蛋白质的任何组合物施加于植物或产生植物的种子;和(ii)用编码tic3668型蛋白毒素类别的蛋白质的多核苷酸转化植物或产生植物的植物细胞。一般来说,预期tic3668型蛋白毒素类别中的任何蛋白质都可提供在组合物中,提供在微生物中,或提供在转基因植物中以赋予针对鳞翅目或鞘翅目昆虫的昆虫抑制活性。在某些实施方案中,tic3668型蛋白毒素类别的重组多肽是通过在适于表达和产生tic3668型蛋白毒素类别的蛋白质的条件下培养被转化以表达tic3668型蛋白毒素的重组芽孢杆菌属细胞或任何其它重组细菌细胞来制备的昆虫抑制性组合物的杀虫活性成分。此类组合物可通过干燥、冻干、均质化、提取、过滤、离心、沉降或浓缩表达/产生所述重组多肽的此类重组细胞的培养物来制备。此类方法可产生芽孢杆菌属细胞或其它昆虫病原性细菌细胞提取物、细胞混悬液、细胞匀浆、细胞溶解产物、细胞上清液、细胞滤液或细胞沉淀。通过获得如此产生的重组多肽,包括重组多肽的组合物可包括细菌细胞、细菌芽孢和伴孢包涵体,并且可被配制供各种用途,包括作为农业昆虫抑制性喷雾产品或作为膳食生物测定中的昆虫抑制性制剂。在一个实施方案中,为降低形成抗性的可能性,包含一种或多种来自tic3668型蛋白毒素类别的蛋白质的昆虫抑制性组合物可还包含至少一种展现针对同一鳞翅目或鞘翅目昆虫物种的昆虫抑制活性,但不同于tic3668型蛋白毒素的额外多肽。可能用于此类组合物的额外多肽包括昆虫抑制性蛋白和昆虫抑制性dsrna分子。使用此类核糖核苷酸序列来控制昆虫害虫的一个实例描述于baum等(美国专利公布2006/0021087a1)中。用于控制鳞翅目害虫的此类额外多肽可选自由以下组成的组:昆虫抑制性蛋白,诸如但不限于cry1a(美国专利号5,880,275)、cry1ab、cry1ac、cry1a.105、cry1ae、cry1b(美国专利公布号10/525,318)、cry1c(美国专利号6,033,874)、cry1d、cry1da及其变体、cry1e、cry1f、和cry1a/f嵌合体(美国专利号7,070,982;6,962,705;和6,713,063)、cry1g、cry1h、cry1i、cry1j、cry1k、cry1l、cry1型嵌合体(诸如但不限于tic836、tic860、tic867、tic869和tic1100)、cry2a、cry2ab(美国专利号7,064,249)、cry2ae、cry4b、cry6、cry7、cry8、cry9、cry15、cry43a、cry43b、cry51aa1、et66、tic400、tic400、tic800、tic834、tic1415、vip3a、vip3ab、vip3b、axmi-184、axmi-196、dig-3、dig-4、dig-5、dig-11、afip-1a及其衍生物(美国专利公布2014-0033361a1)、afip-1b及其衍生物(美国专利公布2014-0033361a1)、pip-1apip-1b(美国专利公布2014-0007292a1)、pseen3174(美国专利公布2014-0007292a1)、aecfg-592740(美国专利公布2014-0007292a1)、pput_1063(美国专利公布2014-0007292a1)、pput_1064(美国专利公布2014-0007292a1)、gs-135及其衍生物(美国专利公布2012-0233726a1)、gs153及其衍生物(美国专利公布2012-0192310a1)、gs154及其衍生物(美国专利公布2012-0192310a1)、gs155及其衍生物(美国专利公布2012-0192310a1)、如美国专利公布2012-0167259a1中所述的seqidno:2及其衍生物、如美国专利公布2012-0047606a1中所述的seqidno:2及其衍生物、如美国专利公布2011-0154536a1中所述的seqidno:2及其衍生物、如美国专利公布2011-0112013a1中所述的seqidno:2及其衍生物、如美国专利公布2010-0192256a1中所述的seqidno:2和4及其衍生物、如美国专利公布2010-0077507a1中所述的seqidno:2及其衍生物、如美国专利公布2010-0077508a1中所述的seqidno:2及其衍生物、如美国专利公布2009-0313721a1中所述的seqidno:2及其衍生物、如美国专利公布2010-0269221a1中所述的seqidno:2或4及其衍生物、如美国专利号7,772,465(b2)中所述的seqidno:2及其衍生物、如wo2014/008054a2中所述的cf161_0085及其衍生物、如美国专利公布us2008-0172762a1、us2011-0055968a1和us2012-0117690a1中所述的鳞翅目毒性蛋白质和它们的衍生物;如us7510878(b2)中所述的seqidno:2及其衍生物、如美国专利号7812129(b1)中所述的seqidno:2及其衍生物;以及为本领域普通技术人员所知的其它鳞翅目抑制性蛋白。用于控制鞘翅目害虫的此类额外多肽可选自由以下组成的组:昆虫抑制性蛋白,诸如但不限于cry3bb(美国专利号6,501,009)、cry1c变体、cry3a变体、cry3、cry3b、cry34/35、5307、axmi184、axmi205、axmir1、tic407、tic417、tic431、tic807、tic853、tic901、tic1201、tic3131、dig-10、ehips(美国专利申请公布号2010/0017914)以及为本领域普通技术人员所知的其它鞘翅目抑制性蛋白。.在其它实施方案中,此类组合物/制剂可还包含至少一种展现对不由本发明的另外昆虫抑制性蛋白抑制的昆虫的昆虫抑制活性的额外多肽以扩大所得昆虫抑制谱。举例来说,对于控制半翅目害虫,本发明的昆虫抑制性蛋白的组合可与半翅目活性蛋白质诸如tic1415(美国专利申请公布号2013/0097735)、tic807(美国专利号8609936)、tic834(美国专利申请公布号2013/0269060)和为本领域普通技术人员所知的其它半翅目活性蛋白质一起使用。用于控制鞘翅目、鳞翅目和半翅目昆虫害虫的额外多肽可见于由neilcrickmore维护的苏云金芽孢杆菌毒素命名网站上(在万维网上的btnomenclature.info处)。在本领域中已用文件记载昆虫对某些杀虫剂形成抗性的可能性。一种昆虫抗性管理策略在于采用表达两种通过不同作用模式来起作用的不同昆虫抑制剂的转基因作物。因此,对任一昆虫抑制剂具有抗性的任何昆虫都可由另一昆虫抑制剂控制。另一昆虫抗性管理策略采用对未针对所靶向鞘翅目或鳞翅目害虫物种加以防护的植物的使用以为所述未保护植物提供庇护。一个特定实例描述于以引用的方式整体并入本文的美国专利号6,551,962中。其它实施方案,诸如经设计用于控制也由本文公开的蛋白质控制的害虫的待与蛋白质一起用于种子处理的经局部施加的杀害虫化学物质、喷洒制剂、滴注制剂或擦拭制剂可直接施加于土壤(土壤浸润),施加于表达本文公开的蛋白质的生长植物,或被配制以施加于含有一种或多种编码一种或多种所公开蛋白质的转基因的种子。用于种子处理的此类制剂可与各种粘着剂和增粘剂一起施加。此类制剂可含有在作用模式方面与所公开蛋白质协同的杀虫剂,以使配制杀虫剂通过不同作用模式来起控制可由所公开蛋白质控制的相同或类似害虫的作用,或此类杀虫剂起控制在更广泛宿主范围内的害虫或不由tic3668型蛋白毒素类别有效控制的植物害虫物种的作用。以上提及的组合物/制剂可还包含农业上可接受的载体,所述组合物/制剂诸如诱饵、粉剂、粉尘剂、团粒、颗粒剂、喷雾剂、乳剂、胶体混悬剂、水溶液剂、芽孢杆菌属芽孢/晶体制剂、种子处理剂、被转化以表达一种或多种蛋白质的重组植物细胞/植物组织/种子/植物、或被转化以表达一种或多种蛋白质的细菌。视重组多肽固有的昆虫抑制或杀虫抑制水平和待施加于植物或应用于膳食测定的制剂水平而定,以重量计,组合物/制剂可包括各种量的重组多肽,例如以重量计,0.0001%至0.001%至0.01%至1%至99%的重组多肽。实施例鉴于前述内容,本领域技术人员应了解可在不脱离本发明的精神和范围下,在所公开的特定方面进行变化,并且仍然获得相似或类似结果。因此,本文公开的特定结构和功能细节不应解释为具有限制性。应了解本文引用的各参考文献的整个公开内容并入在本申请的公开内容内。实施例1发现tic3668相关蛋白毒素类别鉴定并制备在彼此比较时展现独特属性例如推断毒性、蛋白质组多样性和形态变异的细菌菌株以使用本领域中熟知的方法进行基因组测序。从侧孢短芽孢杆菌(b.laterosporus)菌株eg5552发现在体外生物测定中展现针对鞘翅目昆虫的抑制活性的蛋白质tic3668(seqidno:2)。也发现其它菌株含有类似于tic3668的蛋白质。克隆编码这些蛋白质的多核苷酸区段,并且将其插入重组宿主菌株中以测试表达。设计热扩增引物以从包括eg5552的不同侧孢短芽孢杆菌细菌菌株的总基因组dna扩增基因的全长拷贝。从各菌株产生单独热扩增产物(扩增子),并且分析这些产物中可编码tic3668相关蛋白的开放阅读框的存在。确定各扩增子具有单一开放阅读框,该开放阅读框含有翻译起始密码子,继之以框内连续开放阅读框,以框内翻译终止密码子终止。从这些额外不同细菌菌株的每一个获得的推导氨基酸序列分别以seqidno:2(tic3668)、seqidno:4(tic3669)、seqidno:6(tic3670)、seqidno:8(tic4076)、seqidno:10(tic4078)、seqidno:14(tic4346)、seqidno:16(tic4826)、seqidno:18(tic4861)、seqidno:20(tic4862)、seqidno:22(tic4863)示出。将这些扩增子克隆至重组苏云金芽孢杆菌(bt)质粒表达载体中的孢子形成特异性表达启动子的下游,并且转化至无晶体bt宿主细胞中。也将扩增子克隆至大肠杆菌表达系统中。观察到所得重组菌株表达重组蛋白。实施例2tic3668相关蛋白毒素类别的鞘翅目活性这个实施例说明由tic3668样蛋白针对鞘翅目展现的抑制活性。提交tic3668、tic3669、tic3670、tic4260、tic4076和tic2462全长蛋白的由如实施例1中所述的重组细菌产生的蛋白质制剂以进行针对科罗拉多马铃薯甲虫(科罗拉多金花虫,cpb)以及针对至少一个玉米根虫物种的昆虫膳食覆盖(diet-overlay)生物测定。玉米根虫物种的已知成员是玉米根叶甲(西方玉米根虫,wcr)、巴氏根叶甲(北方玉米根虫,ncr)、墨西哥玉米根叶甲(墨西哥玉米根虫,mcr)、带斑黄瓜根叶甲(巴西玉米根虫(bzr))、霍华德十一星根叶甲(南方玉米根虫,scr)和由绿根叶甲和美丽根叶甲组成的巴西玉米根虫复合体(bcr)。如表6中所显示,结果显示tic3668、tic3669、tic3670、tic4260和tic4076展现针对玉米根虫的致死性。tic2462(编码seqidno:63的seqidno:62)作为一种与axmi-209蛋白(相较于tic2462,在氨基酸水平上>99%同一,并且仅展现两个氨基酸差异)密切相关的蛋白质不展现针对玉米根虫的致死性,由此区分tic3668样蛋白毒素类别与类似于axmi-209的蛋白质的活性。惊人的是,未观察到任何测试蛋白质针对科罗拉多马铃薯甲虫的致死性,所述科罗拉多马铃薯甲虫是通常在生物测定中作为鞘翅目活性的指示物加以测试的物种。表6.观察到的示例性蛋白质针对鞘翅目昆虫害虫的致死性.毒素玉米根虫cpbtic2462--tic3668,tic3669,tic3670+-tic4260,tic4076+-tic4078nt-tic4346++tic4826,tic4861,tic4862,tic4863ntnt+=观察到致死性-=未观察到致死性nt=未测出实施例3成熟形式的tic3668蛋白毒素这个实施例说明膜转运肽在tic3668蛋白毒素类别内的天然蛋白质的氨基末端处存在以及对tic3668蛋白毒素类别的活性成熟毒素蛋白质的发现。使用signalp程序(petersen等(2011),naturemethods,8:785-786)对由tic3668编码序列(seqidno:1)获得的氨基酸序列翻译物的生物信息学分析预测存在对应于n末端前23个氨基酸的膜转运区段。设计实验以确认在tic3668样蛋白毒素类别的各成员内存在膜转运区段。将tic3668于非孢子形成特异性bt启动子之后克隆至bt宿主细胞中。测试所得培养上清液的杀虫活性。以混合物形式从上清液回收对应于tic3668的三种形式的蛋白质。稍后通过质谱分析和n末端序列分析来确定这些小于全长tic3668蛋白的不同片段在它们的相应氨基末端含有如seqidno:2所示的氨基酸16、19或24。在培养基中仅检测到少量这三种截短形式的tic3668。观察到所检测的蛋白质的最为丰富的形式在它的氨基末端具有在如seqidno:2所示的位置24处的丝氨酸。当在人工膳食生物测定中测试时,从培养上清液浓缩和纯化的蛋白质展现针对wcr的生物活性。产生不同表达构建体以鉴定各tic3668型蛋白的展现杀虫活性的最小肽区段。将这些构建体引入无晶体苏云金芽孢杆菌菌株或大肠杆菌菌株中。设计一种用于在无晶体bt菌株中表达如seqidno:2所示的从氨基酸1至317的全长tic3668蛋白的构建体。设计用于在具有羧基末端his标签序列(hhhhahhh)的大肠杆菌表达系统中表达全长tic3668蛋白和各种较短变异形式的tic3668蛋白的构建体。经设计用于在大肠杆菌中表达的构建体由以下组成:(1)经设计以表达如seqidno:2所示的从氨基酸位置1至317的全长tic3668蛋白的构建体;(2)经设计以表达具有如seqidno:2所示的氨基酸16至317的tic3668变体蛋白的构建体;(3)经设计以表达如seqidno:2所示的从氨基酸24至317的tic3668变体蛋白的构建体;(4)经设计以表达如seqidno:2所示的从氨基酸26至氨基酸317的tic3668变体蛋白的构建体;(5)经设计以表达如seqidno:2所示的从氨基酸28至317的tic3668变体蛋白的构建体。另外,从用以产生在如seqidno:2所示的残基no.24处起始的tic3668蛋白的大肠杆菌表达系统获得具有n末端10-his标签和tvmv(烟草叶脉斑点病毒)蛋白酶位点(mhhhhhhhhhhgtetvrfq)的tic3668蛋白。从bt表达系统的上清液获得蛋白质,并且使所述蛋白质经受质谱分析和n末端序列分析。bt表达系统产生从如seqidno:2所示的氨基酸24至317的预测的tic3668成熟毒素。未在大肠杆菌上清液中观察到蛋白质。通过渗透休克以使蛋白质从周质释放来从各相应大肠杆菌表达构建体获得蛋白质。确认由经设计以在小于全长蛋白质的氨基末端含有氨基酸16或24的构建体产生的蛋白质在它们的相应氨基末端含有这些氨基酸。由经设计以表达全长tic3668的构建体产生的蛋白质产生在氨基末端含有在如seqidno:2所示的位置24处的丝氨酸的成熟长度蛋白质。由经设计以含有如seqidno:2所示的氨基酸26或氨基酸28作为n末端氨基酸的构建体产生的蛋白质各自惊人地仅含有氨基酸28作为n末端氨基酸,从而表明在n末端维持如seqidno:2所示的氨基酸编号24的加工对于毒素稳定性可为重要的。提交从这些表达系统分析获得的蛋白质样品以在如实施例2中所述的昆虫膳食覆盖生物测定中针对西方玉米根虫幼虫进行测试。确定来自这个研究的某些n末端截短形式展现降低的生物活性。具体来说,观察到当氨基末端氨基酸是如seqidno:2所示的26或28时,杀虫活性显著降低。可外推的是n末端被截短以短于成熟蛋白质(对于tic3668、tic3669、tic3670、tic4076、tic4078、tic4260、tic4346、tic4826和tic4863,在氨基酸残基no.24处起始;对于tic4861,在氨基酸13处起始;以及对于tic4862,在氨基酸22处起始)的其它tic3668蛋白家族成员是所测试tic3668型蛋白的显示针对wcr的杀虫活性的最短形式。tic3668的相比于成熟蛋白质具有相等长度或更长的所有变体即使在相对较低浓度下也都显示针对wcr的高活性。数据也证明tic3668的大肠杆菌加工随构建体设计而变化。实施例4合成用于在植物中表达的编码tic3668型蛋白的基因设计编码全长和成熟形式的tic3668蛋白、tic3669蛋白、tic3670、tic4076、tic4078、tic4260蛋白、tic4346蛋白、tic4826蛋白、tic4861蛋白、tic4862蛋白和tic4863蛋白的核苷酸序列。根据一般描述于美国专利5,500,365中的方法合成编码tic3668、tic3669和tic3670的核苷酸序列,避免某些有害问题序列诸如富attta和a/t的植物多聚腺苷酸化序列,同时保留天然侧孢短芽孢杆菌蛋白质的氨基酸序列。这些核苷酸序列在本文中提供为seqidno:32、seqidno:34、seqidno:36、seqidno:38、seqidno:40、seqidno:42、seqidno:44、seqidno:46、seqidno:48、seqidno:50和seqidno:51(对于全长序列)以及seqidno:33、seqidno:35、seqidno:37、seqidno:39、seqidno:41、seqidno:43、seqidno:45、seqidno:47和seqidno:49(对于成熟序列)。实施例5用于在植物中表达tic3668型蛋白的表达盒设计具有如seqidno:32、seqidno:33、seqidno:34、seqidno:35、seqidno:36、seqidno:37、seqidno:38、seqidno:39、seqidno:40、seqidno:41、seqidno:42、seqidno:43、seqidno:44、seqidno:45、seqidno:46、seqidno:47、seqidno:48、seqidno:49、seqidno:50和seqidno:51所示的序列的多种植物表达盒。此类表达盒用于在植物原生质体中瞬时表达或转化植物细胞。就蛋白质在细胞内的最终放置来设计典型表达盒。以使蛋白质被翻译具有天然n末端区段的方式设计一组表达盒。设计另一组表达盒以使表达不具有n末端区段的蛋白质(即成熟长度蛋白质)。设计另一组表达盒以具有框内表达以及可操作地连接至成熟长度毒素蛋白质的转运肽,以允许靶向细胞的细胞器诸如叶绿体或质体。所有表达盒都经设计以在5’末端以启动子起始,所述启动子可由多个连续连接的启动子元件、增强子元件或为本领域普通技术人员所知用以加强转基因的表达的其它表达元件组成。启动子序列通常连续继之以一个或多个在启动子的3’的前导序列。在前导序列的3’提供内含子序列以改进转基因的表达。毒素或转运肽的编码序列以及毒素的编码序列位于启动子、前导序列和内含子构型的3’。在编码序列的3’提供3’utr序列以促进转录终止,并且提供对于所得转录物的多聚腺苷酸化重要的序列。所有上述元件都与被提供用于构建表达盒的通常额外的序列(诸如限制核酸内切酶位点或非连接依赖性克隆位点)连续排列。实施例6含有tic3668型蛋白表达盒的转化载体构建土壤杆菌属介导的转化载体以向植物基因组递送表达tic3668、mtic3668、tic3669、mtic3669、tic3670和mtic3670蛋白的dna。将表达盒克隆至适合载体中的土壤杆菌属边界序列之间以使它们将连同可选择标记基因一起由含有构建体载体的土壤杆菌属宿主转移至宿主植物细胞的基因组中。更具体来说,将含有编码以上提及的一种蛋白质的整个胞质表达盒的限制片段克隆至土壤杆菌属植物转化载体中。类似地,将含有整个质体靶向表达盒的限制片段克隆至土壤杆菌属植物转化载体中。将含有tic3668型蛋白表达盒(即未靶向盒或靶向盒)的载体通过电穿孔或通过三亲杂交(tri-parentalmating)来引入土壤杆菌属中。将含有编码tic4076、tic4078、tic4260、tic4346、tic4826、tic4861、tic4862和tic4863的人工基因(各自具有以及不具有编码存在于天然侧孢短芽孢杆菌开放阅读框中的n末端23个氨基酸(seqidno:2所示的氨基酸1-23)的序列)的表达盒克隆至适合载体中的土壤杆菌属边界序列之间,使得它们被转移至宿主细胞的基因组中,并且测试所编码蛋白质的表达和生物活性。实施例7植物中的tic3668型蛋白的鞘翅目活性这个实施例说明当在植物中表达并作为膳食向相应昆虫害虫提供时,由tic3668样蛋白针对鞘翅目诸如玉米根虫幼虫展现的抑制活性。使用含有实施例5中所述的表达盒的载体产生表达tic3668、mtic3668、tic3669、mtic3669、tic3670和mtic3670蛋白的r0转基因玉米植物由通过用来自r0转化体的花粉对非转化野生型商业种质植物的耳穗授粉所产生的种子来生长f1转基因玉米植物。在转移至笼形盆中的土壤之后,用新生玉米根虫昆虫侵扰f1植物,并且将其在控制条件下生长13天。使用oleson等人的0-3评级量表来确定根损害等级(rdr),其中0意指无损伤,并且3意指3个或更多个结节被剪裁至茎的1.5英寸内(j.d.oleson,y-l.park,t.m.nowatzki,j.j.tollefson,“node-injuryscaletoevaluaterootinjurybycornrootworms”,journalofeconomicentomology,98(1):1-8,2005)。通过在生长时期结束时对剩余的三龄幼虫的数目进行计数来评估昆虫致死性。在第一组实验中,针对wcr来测试表达全长tic3668、tic3669和tic3670蛋白的植物。相较于平均根损害等级(rdr)值在2与2.5之间的阴性对照,一些事件显示结节损伤在统计学上显著减轻,但未观察到全长蛋白质具有在商业上显著的活性。在第二组实验中,在玉米植物中表达具有或不具有叶绿体靶向肽的成熟蛋白质mtic3668(seqidno:23)、mtic3669(seqidno:24)和mtic3670(seqidno:25),并且针对wcr加以测试。观察到各成熟蛋白质的显著wcr致死性。相较于阴性对照,在存在以及不存在额外靶向序列下表达mtic3668、mtic3669和mtic3670的各植物显示结节损伤在统计学上显著减轻。图2描绘mtic3668和mtic3669蛋白的若干事件的平均rdr值,并且图3描绘mtic3670的若干事件的平均rdr值,此时的情况是无论蛋白质是否被靶向叶绿体都是在f1玉米植物中进行表达。图2和3的事件名称中的“ts”指示存在靶向序列。对于表达成熟蛋白质mtic3668、mtic3669和mtic3670的这些事件中的许多,观察到在商业上显著的活性。惊人的是,从tic3668样蛋白移除膜转运区段(如seqidno:2所示的氨基酸1-23)使在玉米植物中表达时针对玉米根虫的功效增加。当在植物中表达时,相比于全长蛋白质,成熟长度tic3668样蛋白显示针对鞘翅目害虫的更高水平的杀虫活性。实施例8在玉米中表达的tic3668相关蛋白针对cry3bb1抗性wcr的杀虫活性这个实施例说明由tic3668样蛋白针对已对bt毒素cry3bb1产生抗性的西方玉米根虫(wcr)品系展现的杀虫活性。用每个植物的2000个霍普金顿(hopkinton)品系的wcr卵侵扰使用如实施例7中所述的方法产生的表达mtic3668、mtic3669或mtic3670的f1转基因玉米植物。西方玉米根虫(玉米根叶甲leconte)的霍普金顿品系是对在玉米植物中表达的cry3bb1具有田地进化抗性的非滞育品系。所述品系源于从已连续7年种植cry3bb1玉米的田地获得的成体wcr样品。使种群与非滞育wcr品系回交三次,并且针对cry3bb1抗性选择三次(gassmann等(2011)plosone6(7):e22629;gassmann等(2012)gmcropsfood3(3):235-244)。从爱荷华州立大学(iowastateuniversity)的aarongassman博士的实验室获得集落,并且由位于密苏里州切斯特菲尔德(chesterfield,mo)的孟山都生物技术昆虫学组(monsantobiotechentomologygroup)维持。在侵扰之后,wcr-霍普金顿品系卵在48小时内孵化,并且新生体开始以根为食。在24天之后,将根从土壤移除,并且使用0-3量表,如实施例7中所述来评价玉米根损害。如表7中所示,相较于对照植物,在霍普金顿品系wcr新生体存在下,表达mtic3668、mtic3669和mtic3670的植物对保护玉米根免遭损害高度有效,由此克服wcr对cry3bb1毒素的抗性。表7.用cry3bb1抗性wcr侵扰的转基因玉米植物的平均rdr毒素n平均rdr(0-3)标准误差mtic3668180.060.004mtic3669150.051.82e-10mtic3670140.051.95e-10阴性对照62.140.24n:评价的植物数目实施例9在玉米中表达的tic3668相关蛋白针对田地测试地点的天然wcr侵扰的杀虫活性这个实施例说明由表达tic3668样蛋白的转基因玉米植物针对美国中西部农田中的天然wcr侵扰展现的减轻的根损害有效性。在4月底至5月初期间,在美国中西部的5个场地种植使用如实施例7中所述的方法产生的表达mtic3668、mtic3669或mtic3670的f1转基因玉米植物。在这些场地处的试验依赖于存在的天然玉米根虫侵扰压力。到7月末完成根挖掘以进行损害评估。如实施例7中所述,根据结节损伤量表确定根虫损害。来自根挖掘试验的结果指示在开放田地中的实际农耕条件下,在天然玉米根虫压力存在下,表达mtic3668、mtic3669和mtic3670的植物对保护玉米根免遭损害高度有效。表8显示测试植物在组合各场地时的评价的植物数目(n)、平均rdr和标准误差。表8.在具有天然wcr侵扰的农田中测试的转基因玉米植物的平均rdr毒素n平均rdr(0-3)标准误差mtic36687550.1440.009mtic366911080.1590.008mtic367013110.1200.007阴性对照3621.4260.047实施例10tic3668相关蛋白毒素类别的鳞翅目活性这个实施例说明由tic3668样蛋白针对鳞翅目展现的抑制活性。提交全长tic3668、tic3669、tic3670、tic4076和tic4078蛋白的如实施例1中所述的蛋白质制剂以进行针对黑切根虫(bcw,小地老虎)、西方菜豆切根虫(wbc,豆白缘切根虫)、玉米耳虫(cew,玉米穗蛾)、欧洲玉米螟虫(ecb,欧洲玉米螟)、甘蔗螟虫(scb,小蔗杆草螟)、西南玉米螟虫(swc,西南玉米杆草螟)、卷心菜尺蠖(clw,粉纹夜蛾)、大豆尺蠖(sbl,大豆尺蠖)和秋行军虫(faw,草地夜蛾)的昆虫膳食覆盖生物测定。制备和进行抑制性蛋白生物测定的方案和方法在本领域中是已知的。观察到某些tic3668型蛋白针对某些鳞翅目昆虫害虫的活性,如表9中所显示。表9.示例性蛋白质针对鳞翅目昆虫害虫的观察到的阻碍生长.+=观察到阻碍生长++=阻碍生长和致死性-=未观察到致死性nt=未测出实施例11植物中的tic3668型蛋白的鳞翅目活性这个实施例说明当在植物中表达并作为膳食向相应昆虫害虫提供时,tic3668型蛋白对ecb、swc、bcw、faw、cew、sbl的抑制活性。如美国专利8,344,207中所述以类似方式对表达tic3668、tic3669和tic3670的r0玉米植物进行使用植物叶盘达成的针对鳞翅目害虫的生物测定。基于叶盘由昆虫吞食的百分比,在0(0%被食用)至11(大于50%被食用)的标度上对叶损害等级(ldr)指定等级评分。等级评分步幅以5%以递增方式增加。不含有杀虫蛋白质的r0植物充当阴性对照。相对于未转化对照,全长tic3668型蛋白的胞质表达针对cew、faw和swc减轻摄食损害。相对于未转化对照,tic3670蛋白的胞质表达针对swc减轻摄食损害。实施例12产生拼贴蛋白tic4260这个实施例教导基于tic3668的家族成员来产生新型基因序列。组合来自5种天然tic3668型蛋白的氨基酸变化以产生新型拼贴蛋白tic4260(seqidno:12),其相较于天然存在的蛋白质展现不同氨基酸序列多样性。图1描绘5种天然tic3668型蛋白与tic4260的比对。在这个序列比对中,以灰色阴影将具有序列多样性的位置突出显示。构建编码tic4260蛋白的人工多核苷酸序列(seqidno:11)。成熟tic4260蛋白(mtic4260,seqidno:28)由seqidno:43所示的多核苷酸序列编码。可进行其它tic3668型蛋白的类似比对以产生展现鳞翅目和/或鞘翅目毒性活性的新型蛋白质。表达、纯化这些新型蛋白质,并且将其在膳食生物测定中针对鳞翅目和鞘翅目昆虫加以测试。产生这些新型蛋白质的表达盒,并且将其转化至植物中以表达这些蛋白质来控制植物的鳞翅目和鞘翅目害虫。本文公开和要求保护的所有组合物和方法都可鉴于本公开在不进行过度实验下制备和执行。尽管本发明的组合物和方法已用前述说明性实施方案描述,但将为本领域技术人员显而易知的是在不脱离本发明的真实构思、精神和范围下,变化、改变、修改和变动可应用于组合物、方法,以及应用在本文所述的方法的步骤方面或步骤的顺序方面。更具体来说,将显而易知的是在化学与生理两方面均相关的某些试剂可替代本文所述的试剂,同时将获得相同或类似结果。为本领域技术人员显而易知的所有所述类似替代物和修改都被视为在如由随附权利要求限定的本发明的精神、范围和构思内。应为本领域技术人员显而易知的是可组合这些不同的改进序列变化以产生也在本发明的范围内的变体。本说明书中引用的所有出版物和公布专利文件都以引用的方式并入本文,所述引用的程度就好像特定地和个别地指示将各个别出版物或专利申请以引用的方式并入一样。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1